Local inflammatory events induced by Bothrops atrox snake venom and the release of distinct classes of inflammatory mediators Vanessa Moreira a,1 , Maria Cristina Dos-Santos b,1 , Neide Galvão Nascimento a , Henrique Borges da Silva c , Cristina Maria Fernandes a , Maria Regina D’Império Lima c , Catarina Teixeira a, * a Laboratório de Farmacologia, Instituto Butantan, Av. Vital Brazil,1500, CEP 05503-900, Sao Paulo, SP, Brazil b Laboratório de Imunologia, Departamento de Parasitologia, ICB, Universidade Federal do Amazonas, Manaus, AM, Brazil c Departamento de Imunologia, Instituto de Ciências Biomédicas, Universidade de Sao Paulo, Sao Paulo, SP, Brazil article info Article history: Received 21 October 2011 Received in revised form 27 February 2012 Accepted 6 March 2012 Available online 20 March 2012 Keywords: Bothrops atrox Inflammation Leukocytes Cytokines Eicosanoids Cyclooxygenases abstract Bothrops atrox is responsible for most accidents involving snakes in the Brazilian Amazon and its venom induces serious systemic and local effects. The local effects are not neutralized effectively by commercial antivenoms, resulting in serious sequelae in indi- viduals bitten by this species. This study investigates the local inflammatory events induced in mice by B. atrox venom (BaV), such as vascular permeability, leukocyte influx and the release of important inflammatory mediators such as cytokines, eicosanoids and the chemokine CCL-2, at the injection site. The effect of BaV on cyclooxygenase (COX-1 and COX-2) expression was also investigated. The results showed that intraperitoneal (i.p.) injection of BaV promoted a rapid and significant increase in vascular permeability, which reached a peak 1 h after venom administration. Furthermore, BaV caused leukocyte infiltration into the peritoneal cavity between 1 and 8 h after i.p. injection, with mono- nuclear leukocytes (MNs) predominating in the first 4 h, and polymorphonuclear leuko- cytes (PMNs) in the last 4 h. Increased protein expression of COX-2, but notof COX-1, was detected in leukocytes recruited in the first and fourth hours after injection of BaV. The venom caused the release of eicosanoids PGD 2 , PGE 2 , TXA 2 and LTB 4 , cytokines TNF-a, IL-6, IL-10 and IL-12p70, but not IFN-g, and chemokine CCL-2 at different times. The results show that BaV is able to induce an early increase in vascular permeability and a leukocyte influx to the injection site consisting mainly of MNs initially and PMNs during the later stages. These phenomena are associated with the production of cytokines, the chemokine CCL-2 and eicosanoids derived from COX-1 and COX-2. Ó 2012 Elsevier Ltd. All rights reserved. 1. Introduction Bothrops atrox snake, commonly known in Brazil as jararaca, jararaca do norte, surucucurana or jararaca do rabo branco, has a widespread distribution in the Northern region of Brazil, where it is responsible for most ophidian accidents (Waldez and Vogt, 2009). Poisoning by B. atrox, as with other species, triggers local and systemic effects in victims (Borges et al., 1999). Local lesions caused by this species are considered to be of greater clinical importance and are characterized by edema, pain, erythema, cyanosis and necrosis (Barros et al., 1998; Petretski et al., 2000; Otero et al., 2000). Based on comparative in vivo studies of the venom of nine species of snakes from genus Bothrops, it was shown that B. atrox venom (BaV) has the most intense local effects among the species investigated (Furtado et al., * Corresponding author. Tel.: þ55 11 37267222; fax: þ55 11 37261505. E-mail address: [email protected] (C. Teixeira). 1 These authors contributed equally to this work. Contents lists available at SciVerse ScienceDirect Toxicon journal homepage: www.elsevier.com/locate/toxicon 0041-0101/$ – see front matter Ó 2012 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2012.03.004 Toxicon 60 (2012) 12–20

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

e at SciVerse ScienceDirect
Toxicon 60 (2012) 12–20
Contents lists availabl
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
Local inflammatory events induced by Bothrops atrox snake venomand the release of distinct classes of inflammatory mediators
Vanessa Moreira a,1, Maria Cristina Dos-Santos b,1, Neide Galvão Nascimento a,Henrique Borges da Silva c, Cristina Maria Fernandes a, Maria Regina D’Império Lima c,Catarina Teixeira a,*
a Laboratório de Farmacologia, Instituto Butantan, Av. Vital Brazil, 1500, CEP 05503-900, Sao Paulo, SP, Brazilb Laboratório de Imunologia, Departamento de Parasitologia, ICB, Universidade Federal do Amazonas, Manaus, AM, BrazilcDepartamento de Imunologia, Instituto de Ciências Biomédicas, Universidade de Sao Paulo, Sao Paulo, SP, Brazil
a r t i c l e i n f o
Article history:Received 21 October 2011Received in revised form 27 February 2012Accepted 6 March 2012Available online 20 March 2012
Keywords:Bothrops atroxInflammationLeukocytesCytokinesEicosanoidsCyclooxygenases
* Corresponding author. Tel.: þ55 11 37267222; faE-mail address: [email protected] (C. Teixeira).
1 These authors contributed equally to this work
0041-0101/$ – see front matter � 2012 Elsevier Ltddoi:10.1016/j.toxicon.2012.03.004
a b s t r a c t
Bothrops atrox is responsible for most accidents involving snakes in the Brazilian Amazonand its venom induces serious systemic and local effects. The local effects are notneutralized effectively by commercial antivenoms, resulting in serious sequelae in indi-viduals bitten by this species. This study investigates the local inflammatory eventsinduced in mice by B. atrox venom (BaV), such as vascular permeability, leukocyte influxand the release of important inflammatory mediators such as cytokines, eicosanoids andthe chemokine CCL-2, at the injection site. The effect of BaV on cyclooxygenase (COX-1 andCOX-2) expression was also investigated. The results showed that intraperitoneal (i.p.)injection of BaV promoted a rapid and significant increase in vascular permeability, whichreached a peak 1 h after venom administration. Furthermore, BaV caused leukocyteinfiltration into the peritoneal cavity between 1 and 8 h after i.p. injection, with mono-nuclear leukocytes (MNs) predominating in the first 4 h, and polymorphonuclear leuko-cytes (PMNs) in the last 4 h. Increased protein expression of COX-2, but not of COX-1, wasdetected in leukocytes recruited in the first and fourth hours after injection of BaV. Thevenom caused the release of eicosanoids PGD2, PGE2, TXA2 and LTB4, cytokines TNF-a, IL-6,IL-10 and IL-12p70, but not IFN-g, and chemokine CCL-2 at different times. The resultsshow that BaV is able to induce an early increase in vascular permeability and a leukocyteinflux to the injection site consisting mainly of MNs initially and PMNs during the laterstages. These phenomena are associated with the production of cytokines, the chemokineCCL-2 and eicosanoids derived from COX-1 and COX-2.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Bothrops atrox snake, commonly known in Brazil asjararaca, jararaca do norte, surucucurana or jararaca do rabobranco, has a widespread distribution in the Northernregion of Brazil, where it is responsible for most ophidian
x: þ55 11 37261505.
.
. All rights reserved.
accidents (Waldez and Vogt, 2009). Poisoning by B. atrox, aswith other species, triggers local and systemic effects invictims (Borges et al., 1999). Local lesions caused by thisspecies are considered to be of greater clinical importanceand are characterized by edema, pain, erythema, cyanosisand necrosis (Barros et al., 1998; Petretski et al., 2000;Otero et al., 2000). Based on comparative in vivo studies ofthe venom of nine species of snakes from genus Bothrops, itwas shown that B. atrox venom (BaV) has the most intenselocal effects among the species investigated (Furtado et al.,

V. Moreira et al. / Toxicon 60 (2012) 12–20 13
1991). As with snakebites involving other species fromgenus Bothrops, these local reactions are not neutralizedeffectively by conventional antivenom serum therapy(Avila-Aguero et al., 2001). Furthermore, there have beenfew studies of the mechanisms involved in these effects,and the literature on this subject is scarce.
Using in vivo experimental models, it was shown thatBaV induced an increase in serum levels of nitric oxide andproinflammatory cytokines IL-6 and IFN-g (Barros et al.,1998). In addition, leukocytes were found at the sitewhere BaV had been injected. In light of this, it was sug-gested that neutrophils play an important role in thesynthesis of inflammatory cytokines IL-1b and IL-6 andanti-inflammatory cytokine IL-10 by increasing mRNAexpression of these molecules and the release of nitricoxide by adjacent cells (Escocard et al., 2006). However, theinduction of other inflammatory events and secretion ofmediators of the acute-phase inflammatory response at thesite of the injection of BaV was not investigated. Proteomicanalysis revealed that the main components of BaV aremetalloproteinases, phospholipases A2 (PLA2), serineproteinases and L-amino acid oxidases (Núñez et al., 2009;Calvete et al., 2011). It was found that one of the PLA2s inthe venom, BaPLA2M-I, induced edema, myonecrosis andthe release of IL-6 at the injection site (Núñez et al., 2004).Phospholipases BaPLA2I and BaPLA2III, which are also iso-lated from BaV, were able to degranulate mast cells in vitroand to cause edema, myonecrosis and the release ofcreatine kinase in vivo (Kanashiro et al., 2002). In addition,the metalloproteinase HI-5 exhibited proteolytic andcomplement-inactivating activity in vitro, and in vivo,in complement-sufficient mice, it induced local inflam-mation characterized by edema, accumulation of poly-morphonuclear leukocytes (PMNs) and hemorrhage(Rodrigues et al., 2004).
Various studies have shown that the inflammatoryeffects induced by bothropic venoms include the release ofeicosanoids such as prostaglandins (PGs), thromboxaneand leukotrienes (Farsky et al., 1997; Zamuner and Teixeira,2002; Moreira et al., 2007, 2009), which are involved in thedevelopment of inflammatory events such as pain, edema,chemotaxis and leukocyte activation at the inflammatorysite (Olivo et al., 2007; Chaves et al., 1995; Zamuner andTeixeira, 2002; Teixeira et al., 2009). PGs in turn areproduced from arachidonic acid by the catalytic activity ofcyclooxygenases (COX-1, COX-2 and COX-3) (Warner andMitchell, 2004), COX-1 and COX-2 being the main targetof nonsteroidal anti-inflammatory drugs (Simon, 1996).However, despite the importance of lipid mediators in thedevelopment and amplification of the inflammatory reac-tion, the release of these mediators and of cytokines at thesite of the injection of BaV has not been fully investigated todate.
In the present study, a comprehensive analysis of theinflammatory reaction elicited by B. atrox venom in theperitoneal cavity of mice was performed, with a specialfocus on acute events such as vascular permeability,leukocyte recruitment, the release of selected groups ofinflammatory mediators and COX-1 and COX-2 expressionby leukocytes. Elucidation of these mechanisms is criticalfor the development of novel treatment strategies based on
a more in-depth understanding of the pathophysiology ofB. atrox venom-induced acute inflammatory response.
2. Materials and methods
2.1. Animals
Male Swiss mice (20 g) were housed in temperature-controlled rooms with a 12 h dark–light period andreceived water and food ad libitum. The research protocolsand animals used were in accordance with the guidelines ofthe Committee for Ethics in the Use of Animals at theButantan Institute, SP, Brazil (CEUAIB, referencenumber729/10) and international law and policies. All efforts weremadetominimize the number of animals used and their suffering.
2.2. Venom
BaV was supplied by the Herpetology Laboratory at theButantan Institute, Sao Paulo, Brazil, where it was obtainedby manual extraction from adult specimens. The venomwas lyophilized, maintained at �20 �C and dissolved insterile saline at the time of use. Prior to the experiments,the venom solutions were filtered through 0.22 mm dura-pore PVDF membranes (Millipore, Brazil). The absence ofendotoxin contamination in the B. atrox venom batchesused was demonstrated by performing the limulusamebocyte lysate (LAL) test (Takayama et al., 1994) on thevenom samples, which revealed undetectable levels ofendotoxin (<0.125 EU/mL).
2.3. Induction of an inflammatory reaction
Animals were injected i.p. with BaV (0.5 mg/animal)dissolved in 1 mL of sterile saline or with 1 mL of sterilesaline alone (Control). Mice were killed 1, 4 and 8 h afterinjection of venom by overexposure to CO2, and the peri-toneal exudates were harvested by washing the cavitieswith 2 mL of saline solution. Aliquots of the washes werecentrifuged at 500g for 6 min at 22 �C, and total anddifferential cells counts were carried out with the exudatecells. Supernatants were stored at �80 �C and later used todetermine the concentrations of eicosanoids (PGD2, PGE2,TXA2 and LTB4), cytokines (IL-12p70, IL-6, IL-10, IFN-g andTNF-a) and the chemokine CCL-2 also named MCP-1. Theleukocytes were used to determine COX-1 and COX-2protein expression by western blotting.
2.4. Leukocyte harvesting and counting
Leukocytes were harvested after i.p. injection of BaV orsterile saline by washing peritoneal cavities with 2 mL ofsaline. Aliquots of the washes were used for total cellcounts in a Neubauer chamber after dilution (1:20, v/v) inTurk solution (0.2% crystal violet dye in 30% acetic acid). Fordifferential cell counts, cytospin preparations were stainedwith Hema3 stain (Renylab Química e Farmacêutica, Cai-çaras, Barbacena, MG). Differential cell counts were per-formed by counting at least 100 cells, which were classifiedas either polymorphonuclear or mononuclear based onconventional morphological criteria.

0.0
0.5
1.0
1.5
2.0
2.5ControlBaV
15 30 60 120
*
*
Time (min)
Con
c. E
vans
blu
e dy
e(μ
g/m
L)
Fig. 1. Plasma extravasation induced by BaV in mouse peritoneal cavity.Animals were injected i.p. with BaV (0.5 mg/animal) or sterile saline (control)in a final volume of 1 mL. After the above indicated time intervals, VP wasdetermined by measuring extravasation of Evans blue dye as described inMaterials and methods. Values are mean � SEM of 4–6 animals. *p < 0.05compared with the control group.
V. Moreira et al. / Toxicon 60 (2012) 12–2014
2.5. Evaluation of vascular permeability (VP)
Changes in VP were evaluated by quantifying extrava-sation of Evans blue dye into the mouse peritoneal cavity atseveral time intervals (15, 30, 60 and 120 min) after i.p.injection of BaV (0.5 mg/animal) or sterile saline (control). EB(20 mg/kg, i.v.) was injected 20 min before each timeinterval, when the animals were killed by overexposure toCO2 andexsanguinated. Theperitoneal cavityof each animalwas then washed with 1 mL of saline solution pH 7.2 andcentrifuged at 500g for 5min. The concentration of EB in thesupernatant was determined by recording the absorbanceat 620 nm (Sirois et al., 1988). Results were expressed asmg EB/mL, using a standard curve (0.5–25 mg/mL).
2.6. Western blotting analysis
Aliquots of 2�106 cellswere lysedwith 100 mL of samplebuffer (0.5 M, Tris–HCl, pH 6.8, 20% SDS, 1% glycerol, 1 Mb-mercaptoethanol, 0.1% bromophenol blue) and boiled for10 min at 100 �C. The samples were subjected to SDS-polyacrylamide gel electrophoresis (SDS-PAGE) on 10% bis-acrylamide gels overlaid with a 5% stacking gel. Theproteins were then transferred to a nitrocellulosemembrane (GE Healthcare, Buckinghamshire, UK) usinga Mini Trans-Blot� (Bio-Rad Laboratories, Richmond, CA,USA). Themembranewas blocked for 1 hwith 5% nonfat drymilk in Tris-buffered saline with Tween-20 (TBST) (20 mMTris, 100 mM NaCl and 0.5% Tween-20) and incubated withprimary polyclonal antibody against COX-2 (1:500 v/v)(Cayman Company, Ann Arbor, MI) for 12 h and againstCOX-1 (1:1000 v/v) (Santa Cruz Biotechnology, Santa Cruz,CA, USA) and b-actin (1:3000 v/v) (Sigma, St Louis, MO) for1h. Themembranewas thenwashedand incubatedwith theappropriate secondary antibody conjugated to horseradishperoxidase. Detection was accomplished using theenhanced chemiluminescence method according to themanufacturer’s instructions (GE Healthcare, Buck-inghamshire, UK). Band densities were measured witha GS 800 Densitometer� (Bio-Rad Laboratories, Richmond,CA, USA) using Molecular Analyst� image analysis software(Bio-Rad Laboratories, Richmond, CA, USA).
2.7. Quantification of eicosanoids
The concentration of PGD2, PGE2, TXB2 (a stablemetabolite of TXA2) and LTB4 was measured by specificenzyme immunoassay (Pradelles et al., 1985), usingcommercial kits (Cayman Chemical Company. Ann Arbor,MI), following the manufacturer’s instructions. To thispurpose, the eicosanoids were extracted from samples onSep-Pak C18 columns (Waters Corporations, Milford, MA)and eluted with ethanol.
2.8. Quantification of cytokines and CCL-2
To measure the levels of TNF-a, IL-6, IL-10, IL-12p70,IFN-g and the chemokine CCL-2 present in the peritonealcavities of the mice, we used the BD Cytometric Bead Array(CBA) Inflammation Kit (BD Biosciences, NJ, USA), followingthe manufacturer’s instructions.
2.9. Data analysis
Results are expressed as mean � SEM. Differencesbetween groups were analyzed by one-way analysis ofvariance (ANOVA), followed by the Tukey test. Differenceswith an associated probability (p value) of less than 5%(p < 0.05) were considered significant.
3. Results
3.1. Effects of BaV on VP
Changes in VP were determined at selected intervalsafter i.p. injection of BaV (0.5 mg/animal). Fig. 1 shows thetime course of the increase in VP induced by the venom inthe peritoneal cavity. There was a significant increase inEvans blue extravasation from 30 to 60 min, with thehighest values at 60 min, decreasing rapidly thereafter toreach basal levels at 120 min.
3.2. Cellular influx into the peritoneal cavity induced by BaV
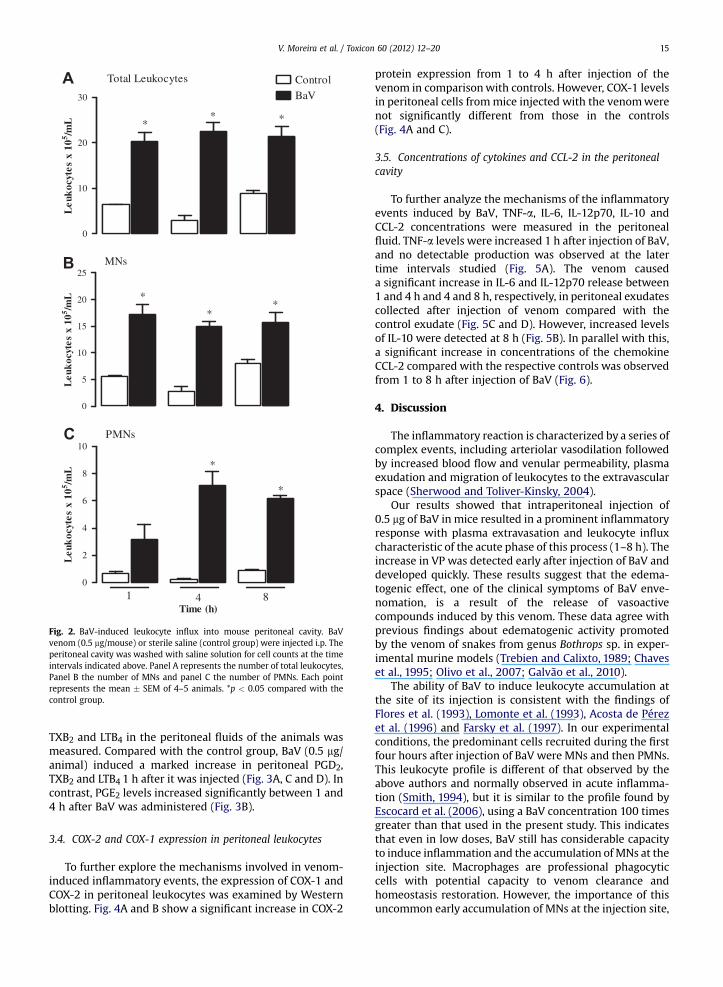
Fig. 2A shows the time course of leukocyte influx intothe peritoneal cavity after injection of BaV (0.5 mg/animal)or saline solution (control group). BaV led to a significantaccumulation of leukocytes at the injection site (Fig. 2A).Compared with the saline group, the number of these cellsremained increased from the 1st to the 8th hour followingBaV injection. Differential cell counts showed that cellspresent in the peritoneal cavity after BaV injection werepredominantly MNs or PMNs, the former being present ingreater quantities. Maximal MNs migration was observedfrom 1 up to 8 h (Fig. 2B). As shown in Fig. 2C, there wassignificant accumulation of PMNs 4 and 8 h after injectionof BaV compared with the respective control groups.
3.3. Eicosanoid concentration in the peritoneal cavity
To investigate the ability of BaV to release lipid media-tors in the injection site, the concentration of PGD2, PGE2,
0
10
20
30
Total Leukocytes
**
BaVControl
*
Leu
koc
ytes
x 1
05 /mL
0
5
10
15
20
25MNs
*
**
Leu
koc
ytes
x 1
05 /mL
0
2
4
6
8
10
*
*
PMNs
1 4 8Time (h)
Leu
koc
ytes
x 1
05 /mL
A
B
C
Fig. 2. BaV-induced leukocyte influx into mouse peritoneal cavity. BaVvenom (0.5 mg/mouse) or sterile saline (control group) were injected i.p. Theperitoneal cavity was washed with saline solution for cell counts at the timeintervals indicated above. Panel A represents the number of total leukocytes,Panel B the number of MNs and panel C the number of PMNs. Each pointrepresents the mean � SEM of 4–5 animals. *p < 0.05 compared with thecontrol group.
V. Moreira et al. / Toxicon 60 (2012) 12–20 15
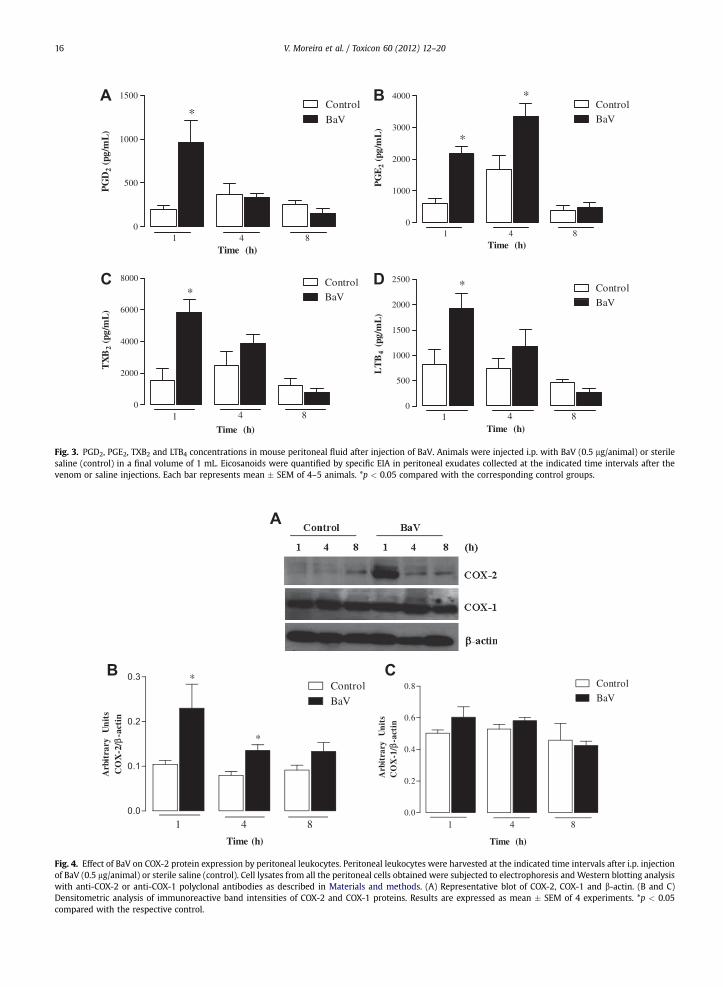
TXB2 and LTB4 in the peritoneal fluids of the animals wasmeasured. Compared with the control group, BaV (0.5 mg/animal) induced a marked increase in peritoneal PGD2,TXB2 and LTB4 1 h after it was injected (Fig. 3A, C and D). Incontrast, PGE2 levels increased significantly between 1 and4 h after BaV was administered (Fig. 3B).
3.4. COX-2 and COX-1 expression in peritoneal leukocytes
To further explore the mechanisms involved in venom-induced inflammatory events, the expression of COX-1 andCOX-2 in peritoneal leukocytes was examined by Westernblotting. Fig. 4A and B show a significant increase in COX-2
protein expression from 1 to 4 h after injection of thevenom in comparisonwith controls. However, COX-1 levelsin peritoneal cells frommice injected with the venomwerenot significantly different from those in the controls(Fig. 4A and C).
3.5. Concentrations of cytokines and CCL-2 in the peritonealcavity
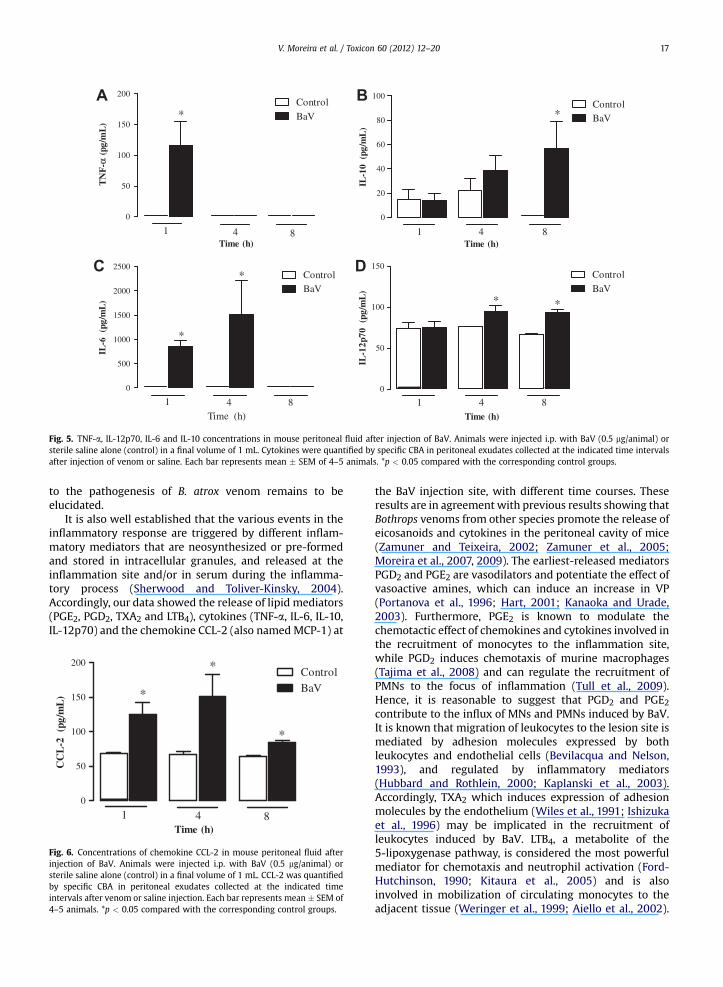
To further analyze the mechanisms of the inflammatoryevents induced by BaV, TNF-a, IL-6, IL-12p70, IL-10 andCCL-2 concentrations were measured in the peritonealfluid. TNF-a levels were increased 1 h after injection of BaV,and no detectable production was observed at the latertime intervals studied (Fig. 5A). The venom causeda significant increase in IL-6 and IL-12p70 release between1 and 4 h and 4 and 8 h, respectively, in peritoneal exudatescollected after injection of venom compared with thecontrol exudate (Fig. 5C and D). However, increased levelsof IL-10 were detected at 8 h (Fig. 5B). In parallel with this,a significant increase in concentrations of the chemokineCCL-2 compared with the respective controls was observedfrom 1 to 8 h after injection of BaV (Fig. 6).
4. Discussion
The inflammatory reaction is characterized by a series ofcomplex events, including arteriolar vasodilation followedby increased blood flow and venular permeability, plasmaexudation and migration of leukocytes to the extravascularspace (Sherwood and Toliver-Kinsky, 2004).
Our results showed that intraperitoneal injection of0.5 mg of BaV in mice resulted in a prominent inflammatoryresponse with plasma extravasation and leukocyte influxcharacteristic of the acute phase of this process (1–8 h). Theincrease in VP was detected early after injection of BaV anddeveloped quickly. These results suggest that the edema-togenic effect, one of the clinical symptoms of BaV enve-nomation, is a result of the release of vasoactivecompounds induced by this venom. These data agree withprevious findings about edematogenic activity promotedby the venom of snakes from genus Bothrops sp. in exper-imental murine models (Trebien and Calixto, 1989; Chaveset al., 1995; Olivo et al., 2007; Galvão et al., 2010).
The ability of BaV to induce leukocyte accumulation atthe site of its injection is consistent with the findings ofFlores et al. (1993), Lomonte et al. (1993), Acosta de Pérezet al. (1996) and Farsky et al. (1997). In our experimentalconditions, the predominant cells recruited during the firstfour hours after injection of BaV were MNs and then PMNs.This leukocyte profile is different of that observed by theabove authors and normally observed in acute inflamma-tion (Smith, 1994), but it is similar to the profile found byEscocard et al. (2006), using a BaV concentration 100 timesgreater than that used in the present study. This indicatesthat even in low doses, BaV still has considerable capacityto induce inflammation and the accumulation ofMNs at theinjection site. Macrophages are professional phagocyticcells with potential capacity to venom clearance andhomeostasis restoration. However, the importance of thisuncommon early accumulation of MNs at the injection site,
0
500
1000
1500
2000
2500 *
BaVControl
1 4 8Time (h)
LT
B4
(pg/
mL
)
0
2000
4000
6000
8000
* BaVControl
1 4 8
Time (h)
TX
B2
(pg/
mL
)
0
1000
2000
3000
4000 *ControlBaV
*
1 4 8Time (h)
PG
E2
(pg/
mL
)
0
500
1000
1500ControlBaV*
1 4 8Time (h)
PG
D2
(pg/
mL
)A B
C D
Fig. 3. PGD2, PGE2, TXB2 and LTB4 concentrations in mouse peritoneal fluid after injection of BaV. Animals were injected i.p. with BaV (0.5 mg/animal) or sterilesaline (control) in a final volume of 1 mL. Eicosanoids were quantified by specific EIA in peritoneal exudates collected at the indicated time intervals after thevenom or saline injections. Each bar represents mean � SEM of 4–5 animals. *p < 0.05 compared with the corresponding control groups.
0.0
0.1
0.2
0.3 *
1 4 8
Time (h)
*
ControlBaV
Arb
itra
ry
Un
its
CO
X-2
/ββ-a
ctin
0.0
0.2
0.4
0.6
0.8
Arb
itra
ry U
nits
CO
X-1
/β-a
ctin
1 4 8
Time (h)
BaVControl
A
B C
Fig. 4. Effect of BaV on COX-2 protein expression by peritoneal leukocytes. Peritoneal leukocytes were harvested at the indicated time intervals after i.p. injectionof BaV (0.5 mg/animal) or sterile saline (control). Cell lysates from all the peritoneal cells obtained were subjected to electrophoresis and Western blotting analysiswith anti-COX-2 or anti-COX-1 polyclonal antibodies as described in Materials and methods. (A) Representative blot of COX-2, COX-1 and b-actin. (B and C)Densitometric analysis of immunoreactive band intensities of COX-2 and COX-1 proteins. Results are expressed as mean � SEM of 4 experiments. *p < 0.05compared with the respective control.
V. Moreira et al. / Toxicon 60 (2012) 12–2016
0
50
100
150
200
* BaVControl
1 4 8Time (h)
TN
F-α
(pg
/mL
)
0
50
100
150
*BaVControl
1 4 8Time (h)
*
IL-1
2p70
(pg
/mL
)
0
500
1000
1500
2000
2500*
BaVControl
*
1 4 8Time (h)
IL-6
(pg
/mL
)
0
20
40
60
80
100
* BaVControl
1 4 8Time (h)
IL-1
0 (
pg/m
L)
A B
C D
Fig. 5. TNF-a, IL-12p70, IL-6 and IL-10 concentrations in mouse peritoneal fluid after injection of BaV. Animals were injected i.p. with BaV (0.5 mg/animal) orsterile saline alone (control) in a final volume of 1 mL. Cytokines were quantified by specific CBA in peritoneal exudates collected at the indicated time intervalsafter injection of venom or saline. Each bar represents mean � SEM of 4–5 animals. *p < 0.05 compared with the corresponding control groups.
V. Moreira et al. / Toxicon 60 (2012) 12–20 17
to the pathogenesis of B. atrox venom remains to beelucidated.
It is also well established that the various events in theinflammatory response are triggered by different inflam-matory mediators that are neosynthesized or pre-formedand stored in intracellular granules, and released at theinflammation site and/or in serum during the inflamma-tory process (Sherwood and Toliver-Kinsky, 2004).Accordingly, our data showed the release of lipid mediators(PGE2, PGD2, TXA2 and LTB4), cytokines (TNF-a, IL-6, IL-10,IL-12p70) and the chemokine CCL-2 (also namedMCP-1) at
0
50
100
150
200 *
BaVControl
*
1 4 8Time (h)
*
CC
L-2
(pg
/mL
)
Fig. 6. Concentrations of chemokine CCL-2 in mouse peritoneal fluid afterinjection of BaV. Animals were injected i.p. with BaV (0.5 mg/animal) orsterile saline alone (control) in a final volume of 1 mL. CCL-2 was quantifiedby specific CBA in peritoneal exudates collected at the indicated timeintervals after venom or saline injection. Each bar represents mean � SEM of4–5 animals. *p < 0.05 compared with the corresponding control groups.
the BaV injection site, with different time courses. Theseresults are in agreement with previous results showing thatBothrops venoms from other species promote the release ofeicosanoids and cytokines in the peritoneal cavity of mice(Zamuner and Teixeira, 2002; Zamuner et al., 2005;Moreira et al., 2007, 2009). The earliest-released mediatorsPGD2 and PGE2 are vasodilators and potentiate the effect ofvasoactive amines, which can induce an increase in VP(Portanova et al., 1996; Hart, 2001; Kanaoka and Urade,2003). Furthermore, PGE2 is known to modulate thechemotactic effect of chemokines and cytokines involved inthe recruitment of monocytes to the inflammation site,while PGD2 induces chemotaxis of murine macrophages(Tajima et al., 2008) and can regulate the recruitment ofPMNs to the focus of inflammation (Tull et al., 2009).Hence, it is reasonable to suggest that PGD2 and PGE2contribute to the influx of MNs and PMNs induced by BaV.It is known that migration of leukocytes to the lesion site ismediated by adhesion molecules expressed by bothleukocytes and endothelial cells (Bevilacqua and Nelson,1993), and regulated by inflammatory mediators(Hubbard and Rothlein, 2000; Kaplanski et al., 2003).Accordingly, TXA2 which induces expression of adhesionmolecules by the endothelium (Wiles et al., 1991; Ishizukaet al., 1996) may be implicated in the recruitment ofleukocytes induced by BaV. LTB4, a metabolite of the5-lipoxygenase pathway, is considered the most powerfulmediator for chemotaxis and neutrophil activation (Ford-Hutchinson, 1990; Kitaura et al., 2005) and is alsoinvolved in mobilization of circulating monocytes to theadjacent tissue (Weringer et al., 1999; Aiello et al., 2002).

V. Moreira et al. / Toxicon 60 (2012) 12–2018
In light of this, LTB4 may be an important mediator for BaV-induced leukocyte influx. It should be stressed that this isthe first time the release of these lipid mediators at theinjection site in response to the action of BaV has beendemonstrated. The cell populations responsible forproducing and releasing prostanoids under the stimuluswith BaV were not characterized in this study. This matterdeserves further investigation.
To better understand the mechanisms involved in thesynthesis of PGs and thromboxane induced by BaV, theeffect of this venom on protein expression of COX-1 andCOX-2 was investigated. Our data show that COX-2, anisoform expressed during a variety of inflammatory events,was up-regulated in leukocytes recruited by BaV in theearly stages, between 1 and 4 h after the injection. Asexpected, the venom did not change protein levels ofCOX-1, a constitutively expressed isoform. These resultsagree with studies showing that Bothrops jararaca andBothrops asper venom are also able to induce proteinexpression of COX-2 (Moreira et al., 2007, 2009). Thereforeour data suggest that COX-2 is involved in generation of PGsand TXA2 production in peritoneal leukocytes recruitedfollowing the injection of BaV. On the other hand, the factthat BaV does not change COX-1 protein expression inleukocytes does not exclude the possibility that it stimu-lates the enzymatic activity of this isoform. In this context,it has been shown that the constitutive COX-1 is mainlyrecruited in the immediate biosynthesis of prostanoids,which occurs within several minutes after stimuli whereasthe inducible COX-2 is required for delayed prostanoidbiosynthesis, which lasts for several hours followingvarious stimuli (James et al., 2001; Naraba et al., 1998).Depending on cell type or different pathological stimuli,the formation of PGE2 is linked mainly to COX-2(Matsumoto et al., 1997). Therefore, the production ofprostanoids detected at the early periods after B. atroxvenom injection may be at least in part due to an initialstimulatory effect of this venom on the enzymatic activityof COX-1 followed by induction of COX-2 expression, owingto differential conversion of the common precursor PGH2into the distinct prostanoids at 1 or 4 h, respectively.
Cytokines are believed to be mediators that regulate allthe phases of the inflammatory response (Arend andGabay, 2004). The kinetics of BaV-induced cytokinerelease observed here confirms that this venom induces animmediate inflammatory response. Because of the ability ofTNF-a to induce expression of distinct classes of adhesionmolecules (Miyata et al., 1999; Sedgwick et al., 2000), thiscytokine may contribute to infiltration of leukocytes to thelocal of BaV injection. Moreover, considering that TNF-a positively regulates COX-2 expression (Di Rosa et al.,1996; Smith et al., 2000), it is reasonable to suggest thatthis cytokine is an important mediator in the induction ofCOX-2 expression by BaV and, consequently, in increasedPG production. Concomitantly, the increase in IL-6 levelsseen in the first four hours after injection of BaV suggeststhat IL-6 is involved in BaV-induced leukocyte infiltrationas this cytokine mediates the maturation of bone marrowPMNs and the increase in levels of these cells in the bloodcirculation (Ulich et al., 1989; Suwa et al., 2000). Further,the increase in IL-10 seen 8 h after injection of BaV implies
this cytokine is a mediator in the resolution phase of theinflammatory process induced by the venom since IL-10 isknown to be an anti-inflammatory and immune regulatorcytokine (Vieira and O’Garra, 2007; Sabat, 2010). It inhibitsthe release of growth factors and inflammatory mediatorsTNF-a, IL-1b, IL-6 and CXCL-8 (DeWaal Malefyt et al., 1991),as well as COX-2 synthesis and the resulting production ofPGE2 (Niiro et al., 1997). Different cell populations in theinnate and adaptive immune system secrete IL-10(Mehrotra et al., 1998; Wendy et al., 2003; Speiran et al.,2009), and its production can be stimulated by IL-12(Gerosa et al., 1996; Garcia et al., 2009; Durrant andMetzger, 2010). Interestingly, injection of BaV led to IL-12p70 production from 4 up to 8 h after the injection,suggesting that IL-12 acts as a stimulatory factor for IL-10production. Moreover, IL-12p70 is known to favor IFN-gproduction by activating natural killer (NK) cells andinducing later differentiation of T-helper 1 cells (TH1)(Trinchieri, 2003; Wendy et al., 2003). However, IFN-g wasnot detected at the BaV injection site at the time intervalsevaluated (data not shown), suggesting that NK cells werenot activated by BaV during this time period. Of note wasthe fact that MN recruitment took place as soon as the firsthour after injection of BaV. CCL-2 is one of the mostpowerful chemotactic mediators for monocyte recruitment(Jiang et al., 1992; Luster, 1998; Henderson et al., 2003).Therefore, the elevated levels of CCL-2 observed soon afterinjection of BaVmay explain the early accumulation of MNsat the injection site. In addition, since PGD2 and PGE2 areinvolved in positive modulation of the synthesis of CCL-2 inthe presence of TNF-a, as well as in expression of its mainreceptor, CCR2 (Panzer and Uguccioni, 2004; Hirano et al.,2007), it is reasonable to suggest that PGD2, PGE2 andTNF-a released by BaV act in synergy to induce productionof the chemokine CCL-2.
In conclusion, our data show that BaV induces an acuteinflammatory reaction at the site of its injection charac-terized by increased VP and leukocyte influx. The infiltratedcells consist mainly of macrophages and neutrophils, whichaccumulate soon after the venom injection. In parallel, BaV-induced production of the eicosanoids PGD2, PGE2, TXA2and LTB4, the cytokines TNF-a, IL-6, IL-10 and IL-12 and thechemokine CCL-2, but not INF-g. Furthermore, BaV up-regulates COX-2 expression in inflammatory cells. Thismay be an important mechanism in the increased synthesisof PGs and TXA2 induced by BaV. Taken together, theunderstanding of the inflammatory events and theinvolvement of inflammatory mediators released at theBaV injection site may help provide a more effective adju-vant to serum therapy to fight the acute local effectsinduced by this venom.
Acknowledgments
The authors thank to Renata Hage Amaral for technicalassistance. This investigation was supported by researchgrants from FAPESP, Brazil (grants 07/03336-9, 09/50896and 10/51150-4). C.T. is recipient of CNPq-PQ (grant306099/2008-0); V.M. is recipient of Pos doctoral fellow-ship from FAPESP-Brazil (grant 07/03337-5); M.C.S. isrecipient of CNPq-PQ (grant 302615/2010-5); N.G.N. is

V. Moreira et al. / Toxicon 60 (2012) 12–20 19
recipient of PhD fellowship from CAPES, H.B.S. is recipientof PhD fellowship from FAPESP (grants 09/08559-1) andM.R.D.L. is recipient of CNPq-PQ (grant 302476/2010-5).
Ethical statement
The animals and research protocols used in this studyfollowed the guidelines of the Ethical Committee for use ofanimals of Butantan Institute, SP, Brazil (protocol number729/10) and international law and policies. All efforts weremade to minimize the number of animals used and theirsuffering.
Conflict of interest
None declared.
References
Acosta de Pérez, O., Teibler, P., Koscinczuk, P., Sánchez Negrette, M.,Trulls, H., Maruñak, S., 1996. Edema and myonecrosis induced byBothrops jararaca venom of Argentina in mice. Acta Physiol. Phar-macol. Ther. Latinoam. 46, 233–238.
Aiello, R.J., Bourassa, P.A., Lindsey, S., Weng, W., Freeman, A., Showell, H.J.,2002. Leukotriene B4 receptor antagonism reduces monocytic foamcells in mice. Arterioscler. Thromb. Vasc. Biol. 22, 443–449.
Arend, W.P., Gabay, C., 2004. Cytokines in the rheumatic diseases. Rheum.Dis. Clin. North Am. 30, 41–67.
Avila-Agüero, M.L., París, M.M., Hu, S., Peterson, P.K., Gutiérrez, J.M.,Lomonte, B., Faingezicht, I; Snakebite Study Group, 2001. Systemiccytokine response in children bitten by snakes in Costa Rica. Pediatr.Emerg. Care 17, 425–429.
Barros, S.F., Friedlanskaia, I., Petricevich, V.L., Kipnis, T.L., 1998. Localinflammation, lethality and cytokine release in mice injected withBothrops atrox venom. Mediators Inflamm. 7, 339–346.
Bevilacqua, M.P., Nelson, R.M., 1993. Selectins. J. Clin. Invest. 91, 379–387.Borges, C.C., Sadahiro, M., Dos-Santos, M.C., 1999. Aspectos epi-
demiológicos e clínicos dos acidentes ocorridos nos municípios doEstado do Amazonas. Rev. Soc. Bras. Med. Trop. 32, 637–646.
Calvete, J.J., Sanz, L., Pérez, A., Borges, A., Vargas, A.M., Lomonte, B.,Ângulo, Y., Gutiérrez, J.M., Chalkidis, H.M., Mourão, R.H., Furtado, M.F.,Moura-Da-Silva, A.M., 2011. Snake population venomics and anti-venomics of Bothrops atrox: paedomorphism along its trans-amazonian dispersal and implications of geographic venomvariability on snakebite management. J. Proteomics 74, 510–527.
Chaves, F., Barbosa, M., Gutierrez, J.M., 1995. Pharmacological study ofedema induced by venom of the snake Bothrops asper (terciopelo) inmice. Toxicon 33, 31–39.
De Waal Malefyt, R., Abrams, J., Bennett, B., Figdor, C.G., de Vries, J.E., 1991.Interleukin 10 (IL-10) inhibits cytokine synthesis by human mono-cytes: an autoregulatory role of IL-10 produced by monocytes. J. Exp.Med. 174, 1209–1220.
Di Rosa, M., Ialenti, A., Ianaro, A., Sautebin, L., 1996. Interaction betweennitric oxide and cyclooxygenase pathways. Prostaglandins Leukot.Essent. Fatty Acids 54, 229–238.
Durrant, D.M., Metzger, D.W., 2010. IL-12 can alleviate Th17-mediatedallergic lung inflammation through induction of pulmonary IL-10expression. Mucosal. Immunol. 3, 301–311.
Escocard, R.C., Kanashiro, M.M., Petretski, J.H., Azevedo-Silva, J., Queiroz-de-Carvalho, E.C., Dias-da-Silva, W., Kipnis, T.L., 2006. Neutrophilsregulate the expression of cytokines, chemokines and nitric oxidesynthase/nitric oxide in mice injected with Bothrops atrox venom.Immunobiology 211, 37–43.
Farsky, S.H., Walber, J., Costa-Cruz, M., Cury, Y., Teixeira, C.F.P., 1997.Leukocyte response induced by Bothrops jararaca venom: in vivo andin vitro studies. Toxicon 35, 185–193.
Flores, C.A., Zappellini, A., Prado-Franceschi, J., 1993. Lypoxygenase-derived mediators may be involved in in vivo neutrophil migrationinduced by Bothrops erytromelas and Bothrops alternatus venom.Toxicon 31, 1551–1559.
Ford-Hutchinson, A.W., 1990. Leukotriene B4 in inflammation. Crit. Rev.Immunol. 10, 1–12.
Furtado, M.F., Colletto, G.M.D.D., Silva, W.D., 1991. Controle de qualidadedos venenos animais e dos correspondentes antivenenos: I. Padro-nização dos métodos de ensaio das atividades bioquímicas e farm-acológicas dos venenos de algumas espécies do gênero Bothrops eCrotalus. Mem. Inst. Butantan 53, 149–159.
Galvão, N.N., Sampaio, M.C., Amaral, R.O., Teixeira, C., 2010. Contributionof mast cells to the oedema induced by Bothrops moojeni snakevenom and a pharmacological assessment of the inflammatorymediators involved. Toxicon 55, 343–352.
Garcia,C.A.,Wang,H., Benakanakere,M.R.,Barrett, E.,Kinane,D.F.,Martin,M., 2009. C-jun controls the ability of IL-12 to induce IL-10production fromhumanmemory CD4þ T cells. J. Immunol. 183, 4475–4482.
Gerosa, F., Paganin, C., Perilt, D., Paiola, F., Seupoli, M.T., Asteanezaga, M.,Frank, I., Trinchieri, G., 1996. Inteleukin-12 prime human CD4 e CD8 Tcell clones for high production of both Interferon-gamma and IL-10. J.Exp. Med. 183, 2559–2569.
Hart, P.H., 2001. Regulation of the inflammatory response in asthma bymast cell products. Immunol. Cell Biol. 79, 149–153.
Henderson, R.B., Hobbs, J.A., Mathies, M., Hogg, N., 2003. Rapid recruit-ment of inflammatory monocytes is independent of neutrophilmigration. Blood 102, 328–335.
Hirano, Y., Shichijo, M., Deguchi, M., Nagira, M., Suzuki, N., Nishitani, Y.,Hattori, M., Arimura, A., 2007. Synergistic effect of PGD2 via prosta-noid DP receptor on TNF-alpha-induced production of MCP-1 and IL-8in human monocytic THP-1 cells. Eur. J. Pharmacol. 560, 81–88.
Hubbard, A.K., Rothlein, R., 2000. Intercellular adhesion molecule-1(ICAM-1) expression and cell signaling cascades. Free Radic. Biol.Med. 28, 1379–1386.
Ishizuka, T., Suzuki, K., Kawakami, M., Hidaka, T., Matsuki, Y.,Nakamura, H., 1996. Thromboxane A2 receptor blockade suppressesintercellular adhesion molecule-1 expression by stimulated vascularendothelial cells. Eur. J. Pharmacol. 312, 367–377.
James, M.J., Penglis, P.S., Caughey, G.E., Demasi, M., Clenland, L.G., 2001.Eicosanoids production by human monocytes: does COX-2 contributeto a self-limiying inflammatory response? Inflamm. Res. 50, 249–253.
Jiang, Y., Beller, D.I., Frendl, G., Graves, D.T.,1992.Monocyte chemoattractantprotein-1 regulates adhesion molecule expression and cytokineproduction in humanmonocytes. J. Immunol. 148, 2423–2428.
Kanaoka, Y., Urade, Y., 2003. Hematopoietic prostaglandin D synthase.Prostaglandins Leukot. Essent. Fatty Acids 69, 163–167.
Kanashiro, M.M., Escocard, R.C.M., Petretski, J.H., Prates, M.V., Alves, E.W.,Machado, O.L., Dias-da-Silva, W.D., Kipnis, T.L., 2002. Biochemical andbiological properties of phospholipases A(2) from Bothrops atroxsnake venom. Biochem. Pharmacol. 64, 1179–1186.
Kaplanski, G., Marin, V., Montero-Julian, F., Mantovani, A., Farnarier, C.,2003. IL-6: a regulator of the transition from neutrophil to monocyterecruitment during inflammation. Trends Immunol. 24, 25–29.
Kitaura, J., Kinoshita, T., Matsumoto, M., Chung, S., Kawakami, Y.,Leitges, M., Wu, D., Lowell, C.A., Kawakami, T., 2005. IgE- and IgEþAg-mediated mast cell migration in an autocrine/paracrine fashion.Blood 105, 3222–3229.
Lomonte, B., Tarkowski, A., Hanson, L.A., 1993. Host response to Bothropsasper snake venom. Analysis of edema formation, inflammatory cells,and cytokine release in a mouse model. Inflammation 17, 93–105.
Luster, A.D., 1998. Chemokines – chemotactic cytokines that mediateinflammation. N. Engl. J. Med. 338, 436–445.
Matsumoto, H., Naraba, H., Murakami, I., Kudo, I., Yakami, K., Ueno, A., Oh-Ishi, S., 1997. Concordant induction of prostaglandin E2 synthase withcyclooxygenase-2 leads to preferred production of prostaglandin E2over thromboxane and prostaglandin D2 in lipopolysaccharide-stimulated rat peritoneal macrophages. Biochem. Biophys. Res.Commun. 230, 110–114.
Mehrotra, P.T., Donnelly, R.P., Wong, S., Kanegane, H., Geremew, A.,Mostowski, H.S., Furuke, K., Siegel, J.P., Bloom, E.T., 1998. Production ofIL-10 by human natural killer cells stimulated with IL-2 and/or IL-12.J. Immunol. 160, 2637–2644.
Miyata, R., Iwabuchi, K., Watanabe, S., Sato, N., Nagaoka, I., 1999. Shortexposure of intestinal epithelial cells to TNF-alpha and histamineinduces Mac-1-mediated neutrophil adhesion independent of proteinsynthesis. J. Leukoc. Biol. 66, 437–446.
Moreira, V., Zamuner, S.R., Wallace, J.L., Teixeira, C.de F., 2007. Bothropsjararaca and Crotalus durissus terrificus venoms elicit distinctresponses regarding to production of prostaglandins E2 and D2, andexpression of cyclooxygenases. Toxicon 49, 615–624.
Moreira, V., Gutiérrez, J.M., Amaral, R.B., Zamunér, S.R., Teixeira, C.F., 2009.Effects of Bothrops asper snake venom on the expression of cyclo-oxygenases and production of prostaglandins by peritoneal leuko-cytes in vivo, and by isolated neutrophils and macrophages in vitro.Prostaglandins Leukot. Essent. Fatty Acids 80, 107–114.

V. Moreira et al. / Toxicon 60 (2012) 12–2020
Naraba, H., Murakami, M., Matsumoto, H., Shimbara, S., Ueno, A., Kudo, I.,Oh-Ishi, S., 1998. Segregated coupling of phospholipases A2, cyclo-oxygenases, and terminal prostanoid synthases in different phases ofprostanoid biosynthesis in rat peritoneal macrophages. J. Immunol.160, 2974–2982.
Niiro, H., Otsuka, T., Izuhara, K., Yamaoka, K., Ohshima, K., Tanabe, T.,et al., 1997. Regulation by interleukin-10 and interleukin-4 ofcyclooxygenase-2 expression in human neutrophils. Blood 89,1621–1628.
Núñez, V., Arce, V., Gutiérrez, J.M., Lomonte, B., 2004. Structural andfunctional characterization of myotoxin I, a Lys49 phospholipase A2homologue from the venom of the snake Bothrops atrox. Toxicon 44,91–101.
Núñez, V., Cid, P., Sanz, L., De La Torre, P., Angulo, Y., Lomonte, B.,Gutiérrez, J.M., Calvete, J.J., 2009. Snake venomics and antivenomicsof Bothrops atrox venoms from Colombia and the Amazon regions ofBrazil, Perú and Ecuador suggest the occurrence of geographic vari-ation of venom phenotype by a trend towards paedomorphism. J.Proteomics 73, 57–78.
Olivo, R.A., Teixeira, C.F.P., Wallace, J.L., Gutierrez, J.M., Zamuner, S.R.,2007. Role of cyclooxygenases in oedema-forming activity ofbothropic venoms. Toxicon 49, 670–677.
Otero, R., Núñes, V., Barona, J., Fonnegra, R., Jiménez, S.L., Osorio, R.G.,Saldarriaga, M., Díaz, A., 2000. Snakebites and ethnobotany in thenorthwest region of Colombia. Part III: neutralization of the hae-morrhagic effect of Bothrops atrox venom. J. Ethnopharmacol. 73,233–241.
Panzer, U., Uguccioni, M., 2004. Prostaglandin E2 modulates the functionalresponsiveness of human monocytes to chemokines. Prostaglandin E2modulates the functional responsiveness of human monocytes tochemokines. Eur. J. Immunol. 34, 3682–3689.
Petretski, J.H., Kanashiro, M.M., Rodrigues, F.R., Alves, E.W., Machado, O.L.T., Kipnis, T.L., 2000. Edema induction by the disintegrin-like/cysteine-rich domains from a Bothrops atrox hemorrhagin. Biochem.Biophys. Res. Commun. 276, 29–34.
Portanova, J.P., Zhang, Y., Anderson, G.D., Hauser, S.D., Masferrer, J.L.,Seibert, K., Gregory, S.A., Isakson, P.C., 1996. Selective neutralization ofprostaglandin E2 blocks inflammation, hyperalgesia, and interleukin 6production in vivo. J. Exp. Med. 184, 883–891.
Pradelles, P., Grassi, J., Maclouf, J., 1985. Enzyme immunoassays ofeicosanoids using acetylcholine esterase as label: an alternative toradioimmunoassay. Anal. Chem. 57, 1170–1173.
Rodrigues, F.G., Petretski, J.H., Kanashiro, M.M., Lemos, L., da Silva, W.D.,Kipnis, T.L., 2004. The complement system is involved in acuteinflammation but not in the hemorrhage produced by a Bothropsatrox snake venom lowmolecular mass proteinase. Mol. Immunol. 40,1149–1156.
Sabat, R., 2010. IL-10 family of cytokines. Cytokine Growth Factor Rev. 21,315–324.
Sedgwick, J.D., Riminton, D.S., Cyster, J.G., Körner, H., 2000. Tumornecrosis factor: a master-regulator of leukocyte movement. Immunol.Today 21, 110–113.
Sherwood, E.R., Toliver-Kinsky, T., 2004. Mechanisms of the inflammatoryresponse. Best. Pract. Res. Clin. Anaesthesiol. 18, 385–405.
Simon, L.S., 1996. Nonsteroidal antiinflammatory drugs and their effects. J.Clin. Rheumatol. 2, 135–140.
Sirois, M.G., Jancar, S., Braquet, P., Plante, G.E., Sirois, P., 1988. PAFincreases vascular permeability in selected tissues: effect of BN-52021 and L-655,240. Prostaglandins 36, 631–644.
Smith, W.L., DeWitt, D.L., Garavito, R.M., 2000. Cyclooxygenases:structural, cellular, and molecular biology. Annu. Rev. Biochem. 69,145–182.
Smith, J.A., 1994. Neutrophils, host defense, and inflammation: a double-edged sword. J. Leukoc. Biol. 56, 672–686.
Speiran, K., Bailey, D.P., Fernando, J., Macey, M., Barnstein, B., Kolawole, M.,Curley, D., Watowich, S.S., Murray, P.J., Oskeritzian, C., Ryan, J.J., 2009.Endogenous suppression of mast cell development and survival by IL-4 and IL-10. J. Leukoc. Biol. 85, 826–836.
Suwa, T., Hogg, J.C., English, D., Van Eeden, S.F., 2000. Interleukin-6induces demargination of intravascular neutrophils and shortenstheir transit in marrow. Am. J. Physiol. Heart Circ. Physiol. 279,H2954–H2960.
Tajima, T., Murata, T., Aritake, K., Urade, Y., Hirai, H., Nakamura, M.,Ozaki, H., Hori, M., 2008. Lipopolysaccharide induces macrophagemigration via prostaglandin D(2) and prostaglandin E(2). J. Pharma-col. Exp. Ther. 326, 493–501.
Takayama, K., Mitchell, D.H., Din, Z.Z., Mukerjee, P., Li, C., Coleman, D.L.,1994. Monomeric Re lipopolysaccharide from Escherichia coli is moreactive than the aggregated form in the Limulus amebocyte lysateassay and in inducing Egr-1 mRNA in murine peritoneal macro-phages. J. Biol. Chem. 269, 2241–2244.
Teixeira, C., Cury, Y., Moreira, V., Picolo, G., Chaves, F., 2009. Inflammationinduced by Bothrops asper venom. Toxicon 54, 988–997.
Trebien, H.A., Calixto, J.B., 1989. Pharmacological evaluation of rat pawoedema induced by Bothrops jararaca venom. Agents Actions 26,292–300.
Trinchieri, G., 2003. Interleukin-12 and the regulation of innate resistanceand adaptative immunity. Nat. Rev. Immunol. 3, 133–146.
Tull, S.P., Yates, C.M., Maskrey, B.H., O’Donnell, V.B., Madden, J., Grimble, R.F., Calder, P.C., Nash, G.B., Rainger, G.E.D., 2009. Omega-3 fatty acidsand inflammation: novel interactions reveal a new step in neutrophilrecruitment. PLoS Biol. 7, 1–11.
Ulich, T.R., Del Castillo, J., Busser, K., Guo, K.Z., Yin, S.M., 1989. Acutein vivo effects of IL-3 alone and in combination with IL-6 on the bloodcells of the circulation and bone marrow. Am. J. Pathol. 135, 663–670.
Vieira, P., O’Garra, A., 2007. Regula’ten’ the gut. Nat. Immunol. 8, 905–907.Waldez, F., Vogt, R.C., 2009. Aspectos ecológicos e epidemiológicos de
acidentes ofídicos em comunidades ribeirinhas do Baixo Rio Purus,Amazonas, Brasil. Acta Amaz. 39, 681–692.
Warner, T.D., Mitchell, J.A., 2004. Cyclooxygenases: new forms, newinhibitors, and lessons from the clinic. FASEB J. 18, 790–804.
Wendy, T., Moriguchi, W.M., Morinobu, A., O’Shea, J.J., 2003. The biologyof IL-12: coordinating innate and adaptive immune responses. Cyto-kine Growth Factor Rev. 14, 361–368.
Weringer, E.J., Perry, B.D., Sawyer, P.S., Gilman, S.C., Showell, H.J., 1999.Antagonizing leukotriene B4 receptors delays cardiac allograft rejec-tion in mice. Transplantation 67, 808–815.
Wiles, M.E., Welbourn, R., Goldman, G., Hechtman, H.B., Shepro, D., 1991.Thromboxane-induced neutrophil adhesion to pulmonary microvas-cular and aortic endothelium is regulated by CD18. Inflammation 15,181–199.
Zamuner, S.R., Teixeira, C.F., 2002. Cell adhesion molecules involved in theleukocyte recruitment induced by venom of the snake Bothrops jar-araca. Mediators Inflamm. 11, 351–357.
Zamuner, S.R., Zuliani, J.P., Fernandes, C.M., Gutiérrez, J.M., Teixeira, C.F.P.,2005. Inflammation induced by Bothrops asper venom: release ofproinflammatory cytokines and eicosanoids, and role of adhesionmolecules in leukocyte infiltration. Toxicon 46, 806–813.
Related Documents











![SUEROS ANTIOFÍDICOS EN COLOMBIA: ANÁLISIS DE LA … · (Bothrops asper y B. atrox), entre otras [son 18 las víboras reportadas para Colombia] (13). Los anticuerpos producidos neutralizan](https://static.cupdf.com/doc/110x72/5ec825cb80505d7a8a78edbd/sueros-antiofdicos-en-colombia-anlisis-de-la-bothrops-asper-y-b-atrox-entre.jpg)