Littoral chironomid communities of Alpine lakes in relation to environmental factors A. Boggero 1, *, L. Fu¨reder 2 , V. Lencioni 3 , T. Simcic 4 , B. Thaler 5 , U. Ferrarese 6 , A.F. Lotter 7 & R. Ettinger 2 1 C.N.R.-Institute for Ecosystem Study (ISE), Largo Tonolli 50/52, 28922, Verbania Pallanza, Italy 2 Institute of Zoology and Limnology, University of Innsbruck, Technikerstasse 25, 6020, Innsbruck, Austria 3 Department of Invertebrate Zoology and Hydrobiology, Museum of Natural Science, Via Calepina 14, 38100, Trento, Italy 4 National Institute of Biology, Vecna pot 111, P.O. Box 141, 1001, Ljubljana, Slovenia 5 Laboratorio Biologico, Agenzia Provinciale per l’Ambiente, Via Sottomonte 2, 39055, Laives, Italy 6 Via Lucca 38, 35143, Padova, Italy 7 Department of Palaeoecology, Laboratory of Palaeobotany and Palynology, Utrecht University, Budapestlaan 4, NL-3584 CD, Utrecht, The Netherlands (*Author for correspondence: E-mail: [email protected]) Key words: lake-littoral, chironomids, distribution, high altitude, Alps, pollution Abstract The results of a study of littoral chironomid communities from 89 lakes in the Alps are presented. The lakes are located on the northern and southern sides of the Alps from the western Bernese to the eastern Julian Alps. Due to the different origins of the data set here considered, different sampling years and sampling methodologies were adopted. All of the lakes were sampled during the ice-free season, but 79/89 lakes were sampled mainly in autumn 2000, while for a few of them the sample dated back to 1996 or the early 90s. Samples were taken by kicking the different substrata or a stretch of lakeshore (10–30 m) for at least 2–5 min with a standard net and were then preserved in alcohol. Larvae and pupal exuviae were sampled by skimming the lake surface with the same net, to extend identification to species level. Notwithstanding the large altitudinal gradient, almost all of the lakes are situated above the timberline, and show a wide range of morphological (depth, watershed and lake area) and chemical characteristics. Chironomids were the most abundant insects represented in these lakes, with highest abundances at pH between 6.0 and 7.5. In par- ticular, Orthocladiinae and Chironominae were the two subfamilies with the widest distribution. Altitude and temperature played an important role in determining their distribution, the higher altitudes were dominated by the former, whereas the latter were more numerous in lower lakes. The prevalence of Orthocladiinae and Tanytarsini at high altitudes was regarded as a general pattern of taxonomic compo- sition in harsh climatic areas. A reverse condition occurred only in the Julian Alps, where Tanytarsini dominate, followed by Tanypodinae and rare Orthocladiinae. Although our results demonstrated that the composition of chironomid assemblages was also determined by pH, the pattern was unbalanced. Due to the fact that no cases of strong water acidification were present, taxa restricted to very low pH were poorly represented (only three taxa—Limnophyes, Endochironomus and Orthocladius (O.) spp.) compared with circumneutral (8) and alkaline (14) taxa, which occurred in slightly acid and non-acidified soft water lakes. As regard conductivity and altitude, we found that almost all the taxa present at higher pH were con- temporary present at conductivity higher than 100 leq l )1 and altitude lower than 2100 m a.s.l. At species level, Pseudodiamesa branickii, Corynoneura arctica, Heterotrissocladius marcidus and Paratanytarsus austriacus resulted to be the most ubiquitous and abundant species in the Alps. When the west to east distribution of taxa was taken into account, Chironomini (particularly the genera Chironomus and Hydrobiologia (2006) 562:145–165 Ó Springer 2006 A. Lami & A. Boggero (eds), Ecology of High Altitude Aquatic Systems in the Alps DOI 10.1007/s10750-005-1809-6

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Littoral chironomid communities of Alpine lakes in relation
to environmental factors
A. Boggero1,*, L. Fureder2, V. Lencioni3, T. Simcic4, B. Thaler5, U. Ferrarese6, A.F. Lotter7
& R. Ettinger21C.N.R.-Institute for Ecosystem Study (ISE), Largo Tonolli 50/52, 28922, Verbania Pallanza, Italy2Institute of Zoology and Limnology, University of Innsbruck, Technikerstasse 25, 6020, Innsbruck, Austria3Department of Invertebrate Zoology and Hydrobiology, Museum of Natural Science, Via Calepina 14, 38100, Trento,Italy4National Institute of Biology, Vecna pot 111, P.O. Box 141, 1001, Ljubljana, Slovenia5Laboratorio Biologico, Agenzia Provinciale per l’Ambiente, Via Sottomonte 2, 39055, Laives, Italy6Via Lucca 38, 35143, Padova, Italy7Department of Palaeoecology, Laboratory of Palaeobotany and Palynology, Utrecht University, Budapestlaan 4,NL-3584 CD, Utrecht, The Netherlands
(*Author for correspondence: E-mail: [email protected])
Key words: lake-littoral, chironomids, distribution, high altitude, Alps, pollution
Abstract
The results of a study of littoral chironomid communities from 89 lakes in the Alps are presented. The lakesare located on the northern and southern sides of the Alps from the western Bernese to the eastern JulianAlps. Due to the different origins of the data set here considered, different sampling years and samplingmethodologies were adopted. All of the lakes were sampled during the ice-free season, but 79/89 lakes weresampled mainly in autumn 2000, while for a few of them the sample dated back to 1996 or the early 90s.Samples were taken by kicking the different substrata or a stretch of lakeshore (10–30 m) for at least2–5 min with a standard net and were then preserved in alcohol. Larvae and pupal exuviae were sampled byskimming the lake surface with the same net, to extend identification to species level. Notwithstanding thelarge altitudinal gradient, almost all of the lakes are situated above the timberline, and show a wide range ofmorphological (depth, watershed and lake area) and chemical characteristics. Chironomids were the mostabundant insects represented in these lakes, with highest abundances at pH between 6.0 and 7.5. In par-ticular, Orthocladiinae and Chironominae were the two subfamilies with the widest distribution. Altitudeand temperature played an important role in determining their distribution, the higher altitudes weredominated by the former, whereas the latter were more numerous in lower lakes. The prevalence ofOrthocladiinae and Tanytarsini at high altitudes was regarded as a general pattern of taxonomic compo-sition in harsh climatic areas. A reverse condition occurred only in the Julian Alps, where Tanytarsinidominate, followed by Tanypodinae and rare Orthocladiinae. Although our results demonstrated that thecomposition of chironomid assemblages was also determined by pH, the pattern was unbalanced. Due tothe fact that no cases of strong water acidification were present, taxa restricted to very low pH were poorlyrepresented (only three taxa—Limnophyes, Endochironomus and Orthocladius (O.) spp.) compared withcircumneutral (8) and alkaline (14) taxa, which occurred in slightly acid and non-acidified soft water lakes.As regard conductivity and altitude, we found that almost all the taxa present at higher pH were con-temporary present at conductivity higher than 100 leq l)1 and altitude lower than 2100 m a.s.l. At specieslevel, Pseudodiamesa branickii, Corynoneura arctica, Heterotrissocladius marcidus and Paratanytarsusaustriacus resulted to be the most ubiquitous and abundant species in the Alps. When the west to eastdistribution of taxa was taken into account, Chironomini (particularly the genera Chironomus and
Hydrobiologia (2006) 562:145–165 � Springer 2006A. Lami & A. Boggero (eds), Ecology of High Altitude Aquatic Systems in the AlpsDOI 10.1007/s10750-005-1809-6

Dicrotendipes) and Tanypodinae (Larsia and Paramerina) were more typical of the eastern lakes (JA). Theymay be better indicators of more alkaline waters. Assembling all the existing data and presenting a com-prehensive panorama of lake-littoral chironomid taxa of the Alps was one of the aims of this paper. Furthermonitoring, but also harmonization of the sampling methodologies and taxonomical identificationincluding inter-calibration exercises among laboratories are recommended, to provide reliable basis andmore knowledge for future studies of comparative biogeography, and to preserve these environments whereglobal climatic changes may cause abrupt shifts in the faunal assemblages with a loss of taxa typical of thesemountainous areas.
Introduction
Despite their importance and abundance at highaltitude, chironomid communities in mountainlakes in the Alps were neither extensively norintensively studied until the 20th century. Not-withstanding the rich literature produced in thefirst years of the 20th century, no papers dealingwith chironomids of mountain lakes in the Alpsabove the treeline were written.
Published work only existed from sporadicsampling and for faunistic purposes, e.g. thestudy of lakes on the Northern slope of thePrealps (Lundbeck, 1936), while, at the sametime, Thienemann (1936) in his monograph onhigh altitude chironomids referred to lakeslocated below the treeline, with the sole exceptionof the Alpine Stuibensee in Upper Bavaria.
Papers covering the chironomids of mountainlakes sensu stricto are available only on Norwegianlakes (Lenz, 1927), and somewhat later on lakes onthe French slopes of the Pyrenees (Laville, 1972),the Sierra Nevada (Spain) (Laville & Vilchez-Quero, 1986), and the French Central Massif(Laville & Serra-Tosio, 1987). More recentlyanother paper has been written on Pyrenees andSierra Nevada (Spain) and Serra da Estrela(Portugal) (Rieradevall & Prat, 1999).
Reiss (1968) was the first author to study thechironomid community of 14 Alpine lakes at alti-tudes above 1000 m a.s.l., as a threshold betweenhigh and low elevations in the Alps. He tried todefine chironomid species characteristic for theseecosystems, explaining their exclusive presence inthem and their absence in the prealpine lakesthrough the cold-stenothermy of the larval stagerather than through biogeography. He believed
that the existence of a very small number of alpineendemic and boreoalpine species among chirono-mids strengthened his assertion.
A few years later, sporadic records of chirono-mids from Alpine mountain pools and small lakeswere only present in taxonomic, faunistic or eco-logical papers (Reiss & Fittkau, 1971; Serra-Tosio,1989; Ferrarese, 1990; Serra-Tosio & Laville, 1991;Rossaro, 1992; Orendt, 2000; Ferrarese, 2003). Inthe 1990s chironomids were studied in the SwissAlps to establish empirical relationships betweentheir occurrence and environmental parameterssuch as climate or the trophic state of lakes (Lotteret al., 1997, 1998; Heiri et al., 2003). Only recently,after a first tentative attempt by Garibaldi et al.(1987) has there been a spate of samplingcampaigns due to pan-European projects (i.e.AL:PE, Acidification of mountain Lakes:Palaeo-limnology and Ecology; MOLAR, MOuntainLAke Research; EMERGE, European Mountainlake Ecosystems: Regionalisation, diaGnostics andsocio-economic Evaluation) which include studiesof the composition and the structure of thechironomid community, and indeed of the entirebenthos, of Alpine lakes at high altitudes. With twolakes included in the first project and a total of 70 inthe last one, a wide spectrum of the Alps from westto east was covered. Research was devoted to thestudy of physical, chemical, morphologicaland biological characteristics of the same lakes(Mosello et al., 1993; Pugnetti et al., 1993; Boggeroet al., 1996a; Boggero & Nobili, 1998). Wathneet al. (1995) and Fjellheim et al. (2000) alreadydiscussed the distribution of chironomids inrelation to environmental factors (trophy, lake andbasin morphology, hydro-chemistry, and airpollution level).
146

At the same time, Mietto et al. (1997) compareda few Alpine lakes to lowland lakes with the aim offinding a relationship between trophic status andspecies composition. Recently, Franceschini &Lencioni (2002) followed the approach of theEU-projects by studying high altitude lakes with asecond survey considering 10 lakes in the RhaetianAlps, describing them and their inlets and outletsfrom the benthological point of view.
The paper is designed to assemble all existingprevious and recent data to provide a compre-hensive (though not exhaustive) panorama of thelittoral chironomid taxa of high altitude Alpinelakes and to evaluate their distribution among 89mountain lakes in relation to environmentalfactors along a longitudinal gradient of the Alpinechain. Biogeographical notes are also providedregarding a N–S and an E–W gradient.
Study area
One of the major mountain systems of Europe, theAlps, stretch from the Gulf of Genoa to Vienna,and rise between the plains of Northern Italy and ofSouthern Germany (Fig. 1). They do not present acontinuous barrier, being made up of numerousranges divided by comparatively deep valleys.
Following the Italian partitioning of the Alpscreated in 1926 by the Italian National GeographicCommittee and unchanged since then, three maindivisions are generally distinguished:
� WesternAlps—from the Col di Tenda to theSimplon Pass
� Central Alps—from the Simplon Pass to theReschen Scheideck Pass
� Eastern Alps—from the Reschen Scheideck Passto the Radstadter Tauern route
Following this division, the Alpine mountainsystem was subdivided into 18 smaller groups.Seven out of these 18 subgroups constitute the studyarea reported here: Bernese, Pennine, Lepontine,Rhaetian, Dolomites (Central Alps), andNoric andJulian (Eastern Alps). More detailed informationon watersheds and lakes can be found in papers byGuthruf et al. (1999) (Bernese Alps), Mosello et al.(1994) and Boggero et al. (1996b) (Pennine andLepontine Alps), Cantonati et al. (2002) (RhaetianAlps—Province of Trento), Tolotti et al. (2003),Hansen et al. (2002) and Thies et al. (2000)(Rhaetian Alps—Provice of Bolzano and Noric),and Dobravec and Sisko (2002) (Julian Alps).
Table 1 shows the number of sampled lakesper valley and alpine sector and the institutionsinvolved in the present study, while Table 2
Figure 1. Orographic partitioning of the Alps and Alpine sectors considered in the study.
147

summarizes themorphological characteristics of thesame lakes and their watersheds subdivided peralpine sector. All lakes are situated within an areabounded by 46� and 47� north and 7� and 13� east,and, with the exception of one Bernese and sevenJulian lakes, above the modern timberline (around1500–2000 m a.s.l., depending on the region and on
the orientation). Consequently, ice-cover lasts fromNovember to June, at least for most of the lakes.
Different forms of direct anthropogenic impactare observable at all lakes (e.g. tourism and land-use, fish stocking, cattle grazing and waterabstraction), apart from six Trentino-Rhaetianlakes for which information is lacking.
Table 1. Orographic division of the Alps, code, side, states, main valleys, number of sampled lakes and longitude range per each group
of lakes
Alpine sector Alps Code Side State Valley Sampled
lakes
Longitude Institution
involved
Central Bernese, 10 lakes BE North Switzerland Aare 10 7.40–8.42 UB-GBI
Pennine, 9 lakes PE South Italy Strona 1 8.07–8.22 CNR-ISE
Anzasca 2
Bognanco 6
Lepontine, 15 lakes LE South Italy Devero 1 8.33–8.97 CNR-ISE
Formazza 2
Onsernone 3
Isorno 1
Vigezzo 3
South Switzerland Bavona 1 8.48–8.57 CNR-ISE
Lavizzara 4
Central-eastern Rhaetian, 27 lakes RH_N North Austria Inntal 5 10.48–11.12 UI-IZL
Otztal 5
Stubaital 1
RH_S South Italy Nambrone 1 10.50–10.80 MNS-DIZH
Nambino 2
Meledrio 3
Di Genova 2
Siniciaga 1
Canton 1
d’Adige 1 LB-APPA
Passiria 2
d’Ultimo 2
Sarentino 1
Dolomites, 1 lake DO Italy Badia 1 12.01 LB-APPA
Eastern Noric, 13 lakes NO_N North Austria Zillertal 5 11.70–12.77 UI-IZL
Unterinntal 1
Pustertal 5
NO_S South Italy Aurina 2 12.01–12.13 LB-APPA
Julian, 14 lakes JA South Slovenia Bohinj 8 13.68–13.81 NIB
Trenta 6
UB-GBI: University of Bern-Geobotanical Institute; CNR-ISE: National Research Council-Institute for Ecosystem Study; UI-IZL:
University of Innsbruck-Institute of Zoology and Limnology; MNS-DIZH: Museum of Natural Science-Department of Invertebrate
Zoology and Hydrobiology; LB-APPA: Laboratorio Biologico-Agenzia Provinciale per l’Ambiente; NIB: National Institute of
Biology.
148

North side
On the northern side 34 high altitude lakes weresampled (Table 1), 10 of them in the Bernese(Switzerland), 11 in the Rhaetian (Austria) and 13in the Noric Alps (North Tyrol, Austria). Twentylakes are located in a limestone area with some ofthem in a contact zone with siliceous bedrock(granite), and the remaining with catchmentsdominated by siliceous rocks. The Swiss lakes draininto the River Aare, which is a major tributary ofthe Rhine; the Austrian ones feed tributaries to theRiver Danube. For details on watersheds and lake
morphometry see Table 2. The catchments of theSwiss lakes consist predominantly of alpinemeadows and pastures with a changing amount ofscrees and bare rock surfaces, while a second groupconsists of bare ground and scarce vegetationcover. The shores and littoral regions of all thelakes are mainly sandy to rocky.
South side
Most of the lakes in this study lie on the south sideof the Alps. Six sectors and 57 lakes were consid-ered from the Pennine, Lepontine, Rhaetian,
Table 2. Selected statistics describing the total lakes and each group of Alpine lakes studied
Total (89 lakes) Min 25% 50% 75% Max Mean
Altitude (m a.s.l.) 1325 1993 2232 2398 2796 2175
Watershed area (ha) 3.8 26.0 59.0 103.0 783.4 86.5
Lake area (ha) 0.1 0.9 1.6 3.0 45.9 2.8
Maximum depth (m) 1.0 5.0 8.0 14.0 49.4 10.6
BE (10 lakes)
Altitude (m a.s.l.) 1592 1868 1989 2210 2365 2018
Watershed area (ha) 19.8 40.3 106.9 186.2 783.4 186.3
Lake area (ha) 0.7 2.8 6.6 8.2 45.9 9.4
Maximum depth (m) 3.0 8.7 15.6 18.6 49.4 16.8
PE+LE (24 lakes)
Altitude (m a.s.l.) 1910 2081 2210 2336 2450 2196
Watershed area (ha) 3.8 17.3 55.0 89.5 140.0 59.7
Lake area (ha) 0.1 0.7 0.9 1.3 5.9 1.4
Maximum depth (m) 1.0 3.0 5.5 8.5 16.0 6.4
RH (28 lakes)
Altitude (m a.s.l.) 1874 2208 2358 2472 2796 2329
Watershed area (ha) 12.0 36.0 61.5 103.0 275.0 82.4
Lake area (ha) 0.3 1.4 2.2 3.1 9.2 2.7
Maximum depth (m) 2.2 6.2 8.3 14.6 33.7 12.1
NO (13 lakes)
Altitude (m a.s.l.) 1840 2290 2438 2532 2660 2387
Watershed area (ha) 17.0 22.0 38.0 65.0 144.0 52.2
Lake area (ha) 0.9 1.5 2.0 2.5 7.2 2.4
Maximum depth (m) 2.7 6.5 7.4 15.4 44.6 12.7
JA (14 lakes)
Altitude (m a.s.l.) 1325 1490 1815 1975 2150 1743
Watershed area (ha) 13.0 28.5 60.0 93.8 550.0 101.5
Lake area (ha) 0.2 0.6 0.8 1.3 4.5 1.1
Maximum depth (m) 2.5 5.7 8.2 9.9 17.0 8.4
For Alpine sector code see Table 1.
149

Noric, Dolomites to the Julian Alps (Table 1).Only one lake in the Dolomites was studied, but itis described and considered with the southernRhaetian for statistics to avoid a separate chapter.
The origin of most of the lakes is glacial, apartfrom 14 belonging in the Julian Alps which originis glacial-karstic or simply karstic.
The Piedmont lakes feed tributaries to the RiverToce and the Swiss-Lepontine lakes to the RiverTicino, the two main inlets of Lake Maggiore.The Trentino–Rhaetian lakes feed tributaries to theSarca River, the main inlet of Lake Garda, and theAlto Adige–Rhaetian, Noric and Dolomites lakesfeed those to the River Adige. The Julian lakes canbe divided into three groups: the Triglav lakes arelocated in the Bohinj area, and the Kriz and theKrn lakes distributed in the Trenta Valley.
In 35 cases the lithology is mainly acidic, in 8cases there is a prevalence of calcareous rocks, orthey mainly consist of limestone (14 Julian lakes).The watersheds are mainly covered by bare rocks,pastures, shrubs and pioneer-vegetation, but rareconifers are present in the catchments of four lakeslocated at the timberline.
Substrate in the lake littoral is generally dom-inated by larger grain size from gravel to stonesand boulders; silt, mud and sand were prevalentonly in rare cases. In particular, most of the Julianlakeshores consisted of screes or rocks fallen intothe lakes. The bottoms of almost all the Julianlakes were covered with variously coloured thin siltstrictly in relation to oxygen depletion, but onlytwo lakes showed incomplete transparency. Themajority of these lakes were subject to strongfluctuations in their level (from 1 to 4–5 m peryear) depending on precipitation and evaporation,and due to the absence of inlets and outlets.
Chemical characteristics
A summary of the chemical characteristics of thelakes is presented in Figure 2, considering onlysurface samples or samples taken at the lake-outletstations. Unfortunately, there is not a completechemical spectrum for the Bernese Alps, so thisgroup was not considered in the followingdescription.
The chemical composition of lake-water ismainly related to geo-lithology and the watershed
area. In fact, all the lakes lying on siliceous bedrockhad extremely low conductivity (<50 lS cm)1 in68% of the lakes), while those with carbonaticlithology had values up to 228 lS cm)1 (JA andDO lakes). Alkalinity was also low (58% of thelakes with values lower than 200 leq l)1), buthigher in areas dominated by calcareous rocks witha spotty distribution along the Alps and very high(>1000 leq l)1) for one lake in the DO, six lakes inthe NO (1 southern and 5 northern) and all of thosein the JA. While pH was generally in the rangebetween 4.9 and 9.4, it was lower than 6 in 15% ofthe sampled lakes, and <5.0 in one lake.
The main cations were calcium and magne-sium, with sodium and potassium showing similarand lower concentrations. In most of the lakes,ammonium was measured in low concentrations,with only one case in the LE equal of147 lg N l)1. Bicarbonate, sulphate and nitratewere the most important anions. One lake had aparticularly high sulphate concentration of130 mg l)1 due to the high release of this anionfrom the watershed (RH_S, province of Bolzano).Chloride concentrations were below 0.3 mg l)1 in89% of the lakes, while only the lakes in the JAshowed higher figures due to their proximity to thesea. Total phosphorus concentrations were belowor equal to the detection limit of 3 lg P l)1 in 47%of the lakes, while values >9 lg P l)1 were mea-sured in one lake in RH_S (province of Trento)and in JA, respectively, indicating a higher trophiclevel. Notwithstanding the long ice-cover period,due to low nutrient levels there is not usually anyoxygen limitation, except in lakes with highernutrient concentrations.
Secchi disk values reached the bottom layer inmost lakes, with some exceptions were transpar-ency was no more than half of the maximum depthor reduced to one third, or even to 30% or lessindicating a relatively high primary production.
Methods
Sampling procedure generally followed a commonprotocol, developed within the EU-Project‘‘EMERGE’’ (Fjellheim et al., 2000), using thekicking method with a standard net size (250 lm).
Most of the lakes (79/89) were sampled mainlyin autumn 2000, but for a few of them the sample
150

4
5
6
7
8
9
10
TOT PE+LE RH NO JA
pH
0
500
1000
1500
2000
TOT PE+LE RH NO JA
Alkalinity (µeq l-1)
0
50
100
150
200
250
TOT PE+LE RH NO JA
Conductivity (µS cm-1)
0
10
20
30
40
TOT PE+LE RH NO JA
Base cations (µeq l-1)
0
5
10
15
20
TOT PE+LE RH NO JA
Sulphate (mg l-1)
0
100
200
300
400
500
600
TOT PE+LE RH NO JA
Nitrate (µg Nl-1)
0.0
0.5
1.0
1.5
TOT PE+LE RH NO JA
Chloride (mg l-1)
0
20
40
60
80
TOT PE+LE RH NO JA
Total Phosphorus (µg P l-1)
Figure 2. Selected water chemistry variables for the entire set of lakes studied (TOT) and grouped by sector (see codes in Table 1).
Box plots show minimum, maximum values, and 25–50–75% quartiles of the data distribution.
151

dated back to the early 90s. Samples were takenwith a hand-net by kicking the different substrataor a stretch of lakeshore (10–30 m) for at least2 min. The sampled material was immediatelypreserved in 85% alcohol. Larval and pupal exu-viae were captured by skimming the lake surfacewith the same net, in order to facilitate lateridentification to species level.
Ten RH lakes were sampled once during sum-mer 1996, from June to August, when all the lakeswere free from ice. Samples were collected in theselakes using a standard pond net (225 lm) with5 min investigation of different microhabitats inthe littoral zone of each lake. Animals were pre-served in the field in 70% ethanol and sorted andidentified later in the laboratory under a stereo-microscope (50–100�).
Larvae and pupal exuviae of chironomids wereidentified as far as possible following the currenttaxonomic literature.
For analytical purposes, the lakes were selectedon the basis of their respective numbers of indi-viduals; an arbitrary minimum of 200 individualswas regarded as giving an adequate representationof the littoral community as commonly used fordiatoms and pollen grains (Battarbee, 1978, 1984;Rull, 1987; Madanes & Dadon, 1998). The lakeswith lower numbers of individuals were not con-sidered because the number of species was too lowfor a statistical evaluation. Due to differencesamong the participating laboratories regardingidentification level, the complete list of taxaobtained (116) was harmonised to produce a sub-set of 95 homogeneous taxa, excluding unidenti-fied juvenile specimens, which is used in thesubsequent presentation.
To evaluate the zoogeographical distributionalong the Alpine chain, the lakes were divided intofour classes representing rare, common, verycommon and widespread taxa. Taxa were consid-ered rare when present in a maximum of 5 out of89 lakes, common and very common when presentin 6–10, and 11–20 lakes, respectively, and wide-spread when found in more than 20 lakes. Meanfrequency distributions of the species were alsoconsidered, calculating the means among the lakesin which the species was present, and not on theentire set of lakes. As before, four classes weredefined to represent infrequent, frequent, veryfrequent and abundant taxa.
Canonical Correspondence Analysis (CCA) wasused to verify the trend of the abiotic componentswith regard to the groups of lakes and taxa; elabo-ration was performed by the CANOCO 4.5programme (ter Braak & Smilauer, 2002). Abioticvariables considered were chemical (pH, conduc-tivity, main cations and anions, total inorganicnitrogen, representing the sum of ammonium andnitrate, total phosphorus and reactive silica),physical (temperature), and geo-morphological(altitude, longitude, lake and watershed areas, andlake depth) measurements and values. Only surfacelog transformed chemical data were used, as theywere thought to represent the littoral conditionsbetter than deeper samples. The Bernese lakes werenot taken into account in the statistical analysisbecause no complete chemical data set is availablefor them; as a consequence, in this case only thetotal number of lakes considered was 79. The bioticcomponent (list of taxa) abundances were trans-formed in percentages of the respective chironomidcommunity. Taxa rarely represented in the entire setof lakes, i.e. taxa present in less than three lakes withpercentages equal to or lower than 1%, wereexcluded from the analysis (23 rare taxa), and theremaining data (36 taxa) were square-root trans-formed. Four lakes were excluded, 3 from the RHgroup and 1 from the LEAlps, and the remaining 75lakes were analysed, but after a first run two morelakes were excluded, both from JA, as outliers.
Finally, we calculated the respective weightedmean values and the optimum for the 36 taxataken into consideration (ter Braak, 1987) for thetwo variables showing the higher loads in the CCA(conductivity and altitude) and for pH. The latterloaded strongly to the second axis of the CCA thatrepresents a geological gradient from lakes lyingon acidic bedrocks to those on calcareous catch-ments. This gradient is described by several relatedvariables: pH, alkalinity, conductivity and theconcentrations of calcium and magnesium.
Results
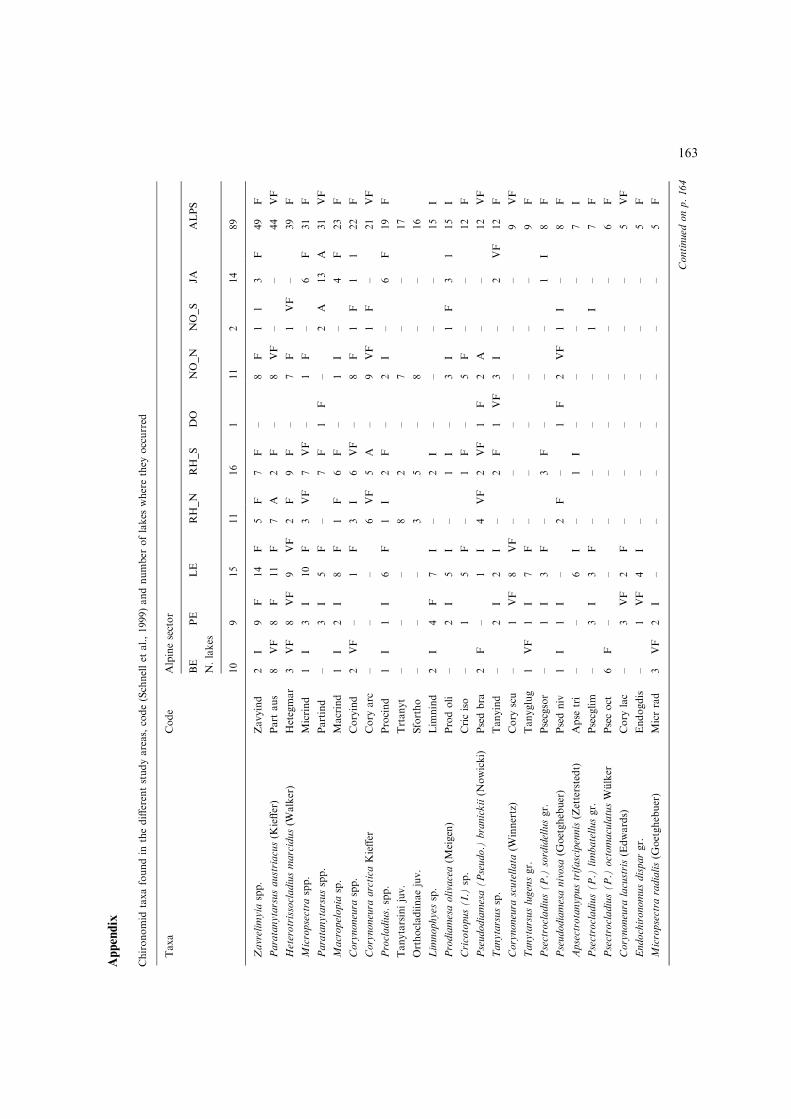
A total of 116 taxa were found and identified in thestudy lakes, taking into consideration subfamilies,tribus, groups, subgroups, genus and species.Appendix A shows the complete list of chironomidtaxa found for the total number of lakes and per
152
each sector of the Alps (specifying the number oflakes in which the considered taxa were present).After harmonisation of the data set only 95 taxaremained for the analysis.
Of the 95 taxa, 6 belonged to the subfamilyDiamesinae, 1 to Prodiamesinae, 16 to Tanypo-dinae, 48 to Orthocladiinae and 24 to Chiro-nominae, 10 of which belonged to the Tanytarsinitribe and 14 to the Chironomini tribe. In partic-ular, Tanypodinae prevailed in RH_S (10 taxa),Orthocladiinae in PE, LE and RH_S (each with18 taxa), Chironominae in LE and RH_S (9 and 8taxa, respectively) and Diamesinae in LE (5 taxa).The number of taxa found varied between 1 in JAand 19 in PE, 75% of the lakes had a maximumof 9 taxa. The lakes with the highest numbers arein PE, LE, RH_S and NO_N. Thirty-five taxawere found in one area only and the remaining 60taxa (64%) were observed more than once.RH_N and NO_S were the only two sectorswithout exclusive taxa, while the others had 10(BE and RH_S), 6 (LE), 4 (JA), 3 (PE) and 2(NO_N), respectively.
Orthocladiinae dominated the littoral fauna inhigh altitude lakes both in taxa number andabundance (here values refer to 75% of the lakesin each sector), with values ranging between 43and 86%, followed by Chironominae, in particularTanytarsini, with values of >30% in each sector.The third important subfamily, Tanypodinae, hadrelative abundances of between 4 and 33% in thesampled lakes, while Diamesinae and Prodiame-sinae represented only a small proportion withvalues lower than 3%. In contrast, in JA Tany-tarsini prevailed (91%), followed by Tanypodinae(15%) and rare Orthocladiinae (3%).
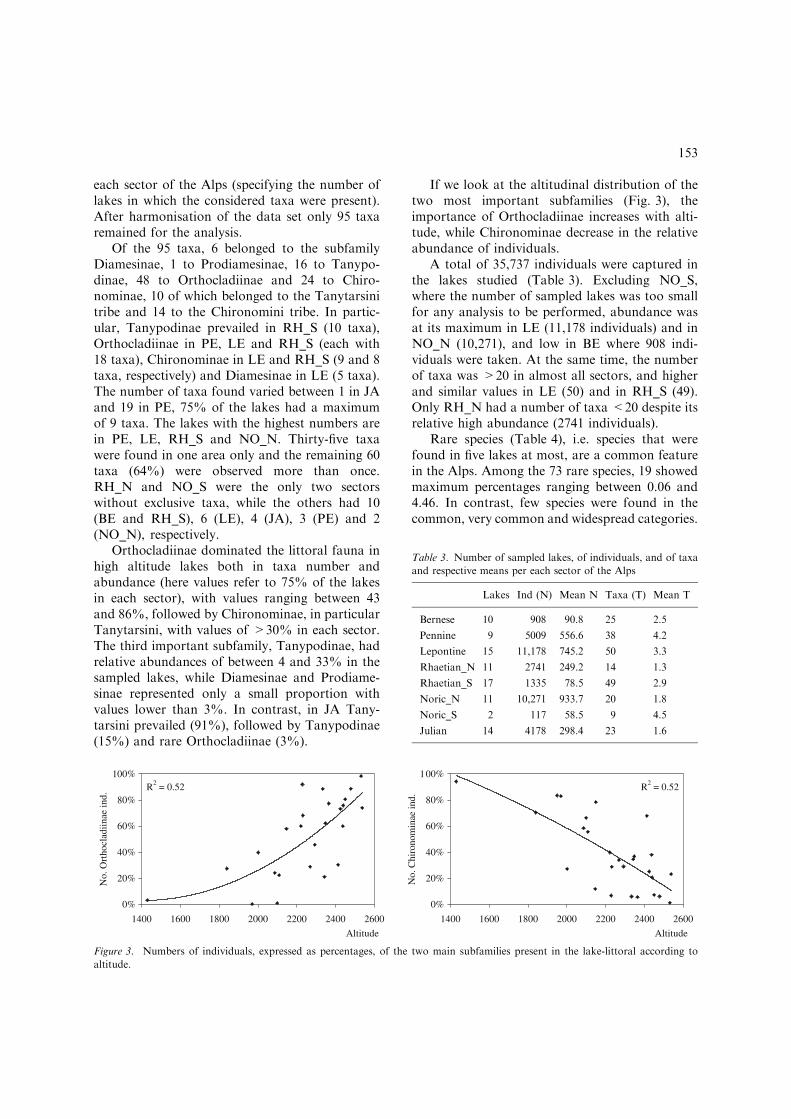
If we look at the altitudinal distribution of thetwo most important subfamilies (Fig. 3), theimportance of Orthocladiinae increases with alti-tude, while Chironominae decrease in the relativeabundance of individuals.
A total of 35,737 individuals were captured inthe lakes studied (Table 3). Excluding NO_S,where the number of sampled lakes was too smallfor any analysis to be performed, abundance wasat its maximum in LE (11,178 individuals) and inNO_N (10,271), and low in BE where 908 indi-viduals were taken. At the same time, the numberof taxa was >20 in almost all sectors, and higherand similar values in LE (50) and in RH_S (49).Only RH_N had a number of taxa <20 despite itsrelative high abundance (2741 individuals).
Rare species (Table 4), i.e. species that werefound in five lakes at most, are a common featurein the Alps. Among the 73 rare species, 19 showedmaximum percentages ranging between 0.06 and4.46. In contrast, few species were found in thecommon, very common and widespread categories.
R2 = 0.52
0%
20%
40%
60%
80%
100%
1400 1600 1800 2000 2200 2400 2600
Altitude
No.
Ort
hocl
adiin
ae in
d.
R2 = 0.52
0%
20%
40%
60%
80%
100%
1400 1600 1800 2000 2200 2400 2600
Altitude
No.
Chi
rono
min
ae in
d.
Figure 3. Numbers of individuals, expressed as percentages, of the two main subfamilies present in the lake-littoral according to
altitude.
Table 3. Number of sampled lakes, of individuals, and of taxa
and respective means per each sector of the Alps
Lakes Ind (N) Mean N Taxa (T) Mean T
Bernese 10 908 90.8 25 2.5
Pennine 9 5009 556.6 38 4.2
Lepontine 15 11,178 745.2 50 3.3
Rhaetian_N 11 2741 249.2 14 1.3
Rhaetian_S 17 1335 78.5 49 2.9
Noric_N 11 10,271 933.7 20 1.8
Noric_S 2 117 58.5 9 4.5
Julian 14 4178 298.4 23 1.6
153

Although some taxa were found to be presentin most of the lakes (Appendix A) they frequentlyoccurred in different proportions, and it was notalways the most widespread species that were alsothe most abundant (Table 4).
Considering the species level and the entirealpine chain, Pseudodiamesa branickii, Corynone-ura arctica, Heterotrissocladius marcidus andParatanytarsus austriacus were not only verycommon or widespread species in the Alps, butthey were also abundant in some cases (maximumvalues between 81 and 98%). Pseudodiamesabranickii and Paratanytarsus austriacus alsoreached high mean values (about 30% of the pop-ulation). At genus level, Cricotopus (I.) sp., Proc-ladius spp.,Micropsectra spp. and Zavrelimyia spp.were widespread and abundant in so far as morethan one species was present, but due to identifi-cation difficulties this was not analysed further.
In contrast, Paratanytarsus lauterborni, Para-cladius conversus, Psectrocladius (P.) octomacula-tus, Ablabesmyia monilis and Dicrotendipes lobigerwere found in high abundances but only in a fewlakes (from 1 up to 6 out of 89 lakes) where theyrepresented more than half of the sampled indi-viduals.
Maximum relative abundances per sector (avalue equal to or higher than 70% was consideredmaximum) revealed that BE lakes were rich inMicropsectra radialis, Paracladius conversus, Abla-besmyia monilis,Psectrocladius (P.) octomaculatus,Paratanytarsus austriacus, PE in Corynoneuralacustris, LE in Paratanytarsus laccophilus gr. andHeterotrissocladius marcidus, RH_S in Corynone-ura arctica and Micropsectra spp., RH_N inPseudodiamesa branickii, Paratanytarsus austriacusand Corynoneura arctica, NO_N in Paratanytarsusaustriacus, Pseudodiamesa nivosa and Corynone-ura spp., NO_S in Paratanytarsus spp., and JAin Dicrotendipes lobiger, Tanytarsus sp. and
Paratanytarsus spp. (mean values are shown inAppendix A). Despite these findings, only a few ofthem are also widespread; in fact, the most diffusespecies, i.e. species present in more than the half ofthe lakes in each group of lakes, are:Paratanytarsusaustriacus in BE, PE, LE, RH_N and NO_N,Heterotrissocladius marcidus in PE, LE, RH_S andNO_N, Psectrocladius (P.) octomaculatus in BE,Corynoneura scutellata in LE, Corynoneura arcticain RH_N and NO_N. At genus level: Zavrelimyiain PE and LE, Micropsectra in LE andParatanytarsus in JA.
Two genera, Corynoneura and Zavrelimyia,were observed in all regions, and one species(Heterotrissocladius marcidus) and two more gen-era, Macropelopia and Micropsectra, were absentthe former in JA and the latter in NO_S Alps.
The presence of some rare or less frequentgenera might be justified by the restricted habitatsthey characterise, like terrestrial or semi-terrestrial(Bryophenocladius and Limnophyes), cow dung(Camptocladius), dead wood (Orthocladius (Sympo-siocladius)), macrophytes living tissues (Endo-chironomus), plant held waters (Metriocnemus) andmeadow soil (Parasmittia). Two species not previ-ously seen in the Alps were found: Acamptocladiusreissi Cranston & Sæther (PE and RH_S) andOrthocladius (Pogonocladius) consobrinus (Holmgren)(JA).
The number of chironomid taxa (Fig. 4) alonga gradient of pH (from 5.0 to 8.7) of the studiedlakes shows its maximum values for intermediatepH units (6.0–7.0), in presence of circum-neutraland alkaline soft waters, while the more acidic andthe more alkaline lakes show lower numbers oftaxa.
CCA (Fig. 5) highlighted, in a two-dimensionaltriplot, the correlation between the groups oflakes, the selected environmental characteristics,the morphological features and the distribution
Table 4. Classification of the taxa and of species on the basis of their distribution and abundance
Distribution Abundances
No. lakes Taxa (95) Species (48) Mean % Taxa (95) Species (48)
Rare 1–5 73 35 Infrequent 0–5 56 25
Common 6–10 8 8 Frequent 5–25 28 14
Very common 11–20 6 2 Very frequent 25–50 9 8
Widespread >20 8 3 Abundant >50 2 1
154

patterns of taxa. The first axis, explaining 6.7% ofthe total variance, loads heavily with total phos-phorous and chloride and is negatively relatedwith altitude, lake area, and to a lesser extentsulphate, reactive silica and sodium. The secondaxis, explains 4.4% of the total variance. It ispositively related with calcium, magnesium, alka-linity, and conductivity and negatively with total
inorganic nitrogen. As regards species distribution,there was a distinction between the northern(black symbol) and southern (white symbol) sidesof the Alps on the basis of morphology andchemistry and consequently of taxa distribution.Chironominae prevailed in lower lakes with highertotal phosphorous content and alkalinity, whileOrthocladiinae dominated in higher siliceous andTIN enriched lakes.
Table 5 shows the distribution of taxa in rela-tion to increasing altitude (left), conductivity(middle) and to pH values (right), based on therelative weighted mean values and respective tol-erance. Each lake presents higher abundances of ataxon whose optimal values are nearest to that ofthe lake. The taxa, which show lower tolerances,are better indicators of a specific location, of solutecontent of a lake or of acid–alkaline situation.
In applying an analysis to evaluate the potentialeffects of pH on the chironomid assemblage (Ta-ble 5), we found that Chironomini (in particular,
0
4
8
12
16
20
4.0 4.5 5.0 5.5 6.0 6.5 7.0 7.5 8.0 8.5 9.0
pH
No.
of
chir
onom
id's
taxa
Figure 4. Number of taxa found in the studied lakes and their
distribution with pH.
Figure 5. CCA of environmental features, chemical variables and taxa in the surveyed lakes. (: PE; n:LE; r: RH_N; }: RH_S;
}: DO; d: NO_N; �: NO_S; : JA Alps.
155

Chironomus sp. and Dicrotendipes sp.), more typi-cal of the eastern lakes (JA) and the Tanypodinae(Ablabesmyia sp.) characteristic taxon of the cen-tral eastern RH_S, were better indicators of morealkaline waters. Taxa confined to lower pH (from5.8 to 6.4), and thus regarded as tolerant, i.e.Limnophyes sp., Endochironomus sp. and Ortho-cladius (O.) spp. were primarily found the formerin the central western Alps (PE and LE), and thelatter in the central eastern part of the Alps(RH_S). In contrast, 14 taxa were found to besensitive to pH and could be encountered in theAlps only at a pH>7.0, while eight circumneutralspecies were observed in lakes with pH rangingbetween 6.5 and 7.0.
As regard conductivity and altitude, we foundthat almost all the taxa present at higher pH werecontemporary present at conductivity higher than100 leq l)1 and altitude lower than 2100 m a.s.l.In particular, the Chironomini (Chironomus sp.and Dicrotendipes sp.), and the Tanypodinae(Larsia sp. and Ablabesmyia sp.) were the taxapresent at lower altitude in our lakes.
Obviously, no general conclusions can bedrawn from these results; it does not necessarilyfollow that the first-mentioned taxa in Table 5 willnot be found in other sectors of the Alps, but it isvery likely that they will be found in lakes withmorphological and chemical characteristics similarto those in the JA.
Table 5. Distribution of taxa in relation to increasing altitude (left), conductivity (middle) and pH values (right)
Altitude Conductivity pH
TAXA O T TAXA O T TAXA O T
Dicrotendipes sp. 1375 73 Orthocladius (O.) spp. 10.4 1.4 Limnophyes sp. 5.85 0.63
Chironomus sp. 1455 232 Endochironomus sp. 13.2 3.0 Endochironomus sp. 5.90 0.39
Larsia sp. 1701 331 Limnophyes sp. 19.1 38.0 Orthocladius (O.) spp. 6.20 0.19
Ablabesmyia sp. 1838 137 Heterotrissocladius
marcidus
23.5 27.4 Heterotrissocladius
marcidus
6.60 0.77
Tanytarsus sp. 1882 311 Corynoneura spp. 25.9 29.4 Corynoneura spp. 6.79 0.65
Psectrocladius sp. 1905 229 Zavrelimyia spp. 33.1 39.6 Zavrelimyia spp. 6.80 0.82
Endochironomus sp. 1959 89 Chaetocladius sp. 33.4 22.5 Tanytarsini indet. 6.83 1.16
Procladius spp. 2051 267 Orthocladiinae indet. 35.0 33.3 Procladius spp. 6.87 0.95
Paratanytarsus spp. 2066 254 Procladius spp. 50.8 47.1 Chaetocladius sp. 6.90 0.48
Paracladius sp. 2080 131 Tanypodinae indet. 53.7 44.8 Micropsectra spp. 6.91 1.01
Prodiamesa olivacea 2102 238 Cricotopus sp. 60.9 51.1 Orthocladiinae indet. 6.98 0.60
Limnophyes sp. 2120 179 Macropelopia sp. 68.3 56.2 Cricotopus sp. 7.22 1.11
Parakiefferiella sp. 2124 163 Tanytarsini indet. 72.2 74.4 Tanypodinae indet. 7.23 0.66
Macropelopia sp. 2149 242 Pseudodiamesa sp. 79.7 71.4 Macropelopia sp. 7.35 0.78
Zavrelimyia spp. 2202 290 Psectrocladius sp. 87.7 63.5 Tanytarsus sp. 7.39 0.66
Cricotopus sp. 2217 245 Paratanytarsus spp. 89.4 63.9 Paratanytarsus spp. 7.40 0.86
Heterotrissocladius
marcidus
2264 177 Chironomus sp. 106.1 31.6 Psectrocladius sp. 7.66 1.16
Chaetocladius sp. 2281 109 Prodiamesa olivacea 109.2 43.6 Pseudodiamesa sp. 7.67 0.82
Tanypodinae indet. 2320 243 Larsia sp. 109.8 57.1 Prodiamesa olivacea 7.80 0.45
Corynoneura spp. 2350 167 Micropsectra spp. 111.4 90.3 Larsia sp. 7.82 0.63
Pseudodiamesa sp. 2364 166 Tanytarsus sp. 117.6 79.7 Parakiefferiella sp. 7.90 0.70
Orthocladiinae indet. 2393 229 Dicrotendipes sp. 119.8 14.6 Paracladius sp. 7.96 0.28
Micropsectra spp. 2402 262 Paracladius sp. 126.1 38.6 Dicrotendipes sp. 8.09 1.13
Orthcladius (O.) spp. 2446 113 Ablabesmyia sp. 127.6 42.2 Ablabesmyia sp. 8.25 0.82
Tanytarsini indet. 2536 233 Parakiefferiella sp. 149.7 69.2 Chironomus sp. 8.48 1.23
The values shown are weighted mean altitude, conductivity and pH, and their respective tolerance. O: Optima; T: Tolerance.
156

Discussion
The species collected in the present study are all ofPalaearctic origin (Ashe et al., 1987). Orthocla-diinae and Chironominae, of the Tanytarsini tribe,were the most frequent subfamilies observed in thelittoral area of high altitude lakes in the Alps, aspreviously recorded by Bretschko (1974), Wathneet al. (1995), Boggero et al. (1996a) and by Fran-ceschini and Lencioni (2002); this is considered tobe a general pattern of taxonomic composition inharsh climatic areas (Armitage et al., 1995). JA,where Chironominae were found to prevail overOrthocladiinae, was in a different category fromthe other Alpine areas, not only because of thevery different chemical and morphological featuresof these lakes, but also on the basis of their chi-ronomid communities. In fact, they should prob-ably be compared to lowland lakes.
The general rule, the number of chironomidtaxa increase with decreasing altitude (Ashe et al.,1987; Wathne et al., 1995; Lotter et al., 1997), andis higher for lentic systems, could not be found inthe present study. There was not any clear relationbetween number of taxa and altitude. In fact, whenthe faunistic results from the different Alpineregions are compared, in the RH_S and BE Alpinesector, lakes had the highest ratio between taxafound and individuals collected, as well as thehighest number of exclusive taxa (10). This couldbe explained partly by the presence of lakeslocated at the timberline (i.e. lakes with a morediversified and vegetated lake-shore) in the firstarea and by the higher alkalinity and buffercapacity in the second area.
The taxa found generally confirmed the distri-bution known from current literature. Most ofthem were cold-stenothermic and typical of highaltitude or high latitude lakes, like Apsectrotany-pus trifascipennis, Heterotrissocladius marcidus,Zavrelimyia melanura, Zavrelimyia punctatissima,Micropsectra radialis, and the genera Macropel-opia, Diamesa, Chaetocladius, Krenosmittia, Para-cladius, Neozavrelia and Paracladopelma. A fewother genera (Cardiocladius, Eukiefferiella, Heleni-ella, Parametriocnemus, Rheocricotopus andTvetenia) were more typical of fast flowing waters,evidently due to the presence of inlets.
A lake classification based on littoral/sub-lit-toral fauna along a trophic gradient was proposed
by Saether (1975, 1979), and then tested byKansanen et al. (1984) in some Finnish lakes, butit was quite difficult to create a lake typology basedon the morphological features characterising thelake shore to a depth of 1 m. Usually the littoralhabitat is not as uniform as the profundal one, sothat the littoral fauna was also more diverse, withvery few common and typical species, in fact thesubstratum was more varied due to erosion andthe presence of vegetation. In contrast, the markedsimilarity among high altitude lakes, which are inmost instances shallow, not stratified, at oxygensaturation level, and in watersheds with a thin soillayer and low production of allochthonous organicmatter, a fairly uniform littoral habitat developedfrequently covered with scarce vegetation. Conse-quently, a classification may be possible consider-ing littoral chironomid communities and theirwide similarity.
Our research suggested that there are taxarestricted to oligotrophic systems (Protanypus,Diamesa,Heterotrissocladius marcidus, Paracladiusalpicola, Micropsectra radialis and Tanytarsuslugens), and some others inhabiting more eutrophiclakes (Natarsia, Paramerina divisa, Cricotopus (C.)tremulus, Cricotopus (I.) trifasciatus, Orthocladiusconsobrinus, Psectrocladius (Allop.) obvius, Dicro-tendipes gr. tritomus). It is obvious, as stated also byKansanen et al. (1984), that for genera unidentifiedto species level (like Macropelopia, Procladius,Zavrelimyia, Micropsectra, Tanytarsus and Para-tanytarsus) which are widespread throughout theAlps, it might be useful to have a more detailedidentification for distinguishing lakes.
Many studies highlight the fact that the overallfactors controlling the chironomid distribution aretemperature/altitude and chemical features, whichalso to a large extent control the availability offood and suitable substrate (Thienemann, 1954;Lotter et al., 1997, 1998; Heiri et al., 2003).
The recorded Orthocladiinae were in generalcold adapted and most of them defined asscrapers preferring periphyton incrusting rocksand debris. Consequently, they became morefrequent with increasing altitude, and were poorlyrepresented in lower eastern lakes. Chironomini,merely considered as gathering collectors ofdetritus and algae, in our study increased withdecreasing altitude and dominated in lowlandstanding waters, where temperatures are warmer
157

and oxygen concentrations are likely to be low.The presence of macrophytes seemed to be acondition for the presence of Paratanytarsus,Cricotopus and Corynoneura scutellata, and alsoEndochironomus as Buskens (1987) found thesetaxa in presence of water plants. Dicrotendipeswas only found in more eutrophic waters (JA).As a result, we observed a substitution of Chi-ronominae with Orthocladiinae with increasingaltitude. Also within the Chironominae, Chiro-nomini prevailed at lower heights, while Tany-tarsini, with the genera Micropsectra spp., andthe species Paratanytarsus austriacus andTanytarsus lugens gr. were more abundant athigher elevations.
The application of CCA corroborated thehypothesis that altitude and temperature were themain key factors determining the prevalence ofone subfamily over another (Rossaro, 1991a,b) athigh altitudes and high latitudes. In fact, the firstaxis represented an altitudinal gradient, wheresiliceous weathering was higher at higher altitude.JA lakes were thus the lowest in altitude and hadthe highest total phosphorous, chloride concen-trations and temperatures, while NO and RHlakes were the highest and the largest of thedistribution, and had high concentrations of sul-phate–sodium-reactive silica due to the contribu-tion from the watershed. The second axis might beregarded as an acidity axis, where lakes withhigher longitude values (eastern) showed highercalcium concentrations and were better bufferedthan western lakes with lower areas and high totalinorganic nitrogen content.
CCA demonstrated that in addition to altitudeand temperature, pH too might determine thecomposition of chironomid assemblages, as foundby Leuven et al. (1987). Although pH had nodirect impact on the presence/absence of a taxon,it affected mainly the vegetal growth and theaccumulation of organic matter, and these factorsmodified the bottom substrate, one of the mostimportant variables for the distribution ofchironomid larvae.
There are no cases of strong water acidificationin the Alps, consequently very few (3) taxarestricted to very low pH were found compared tothe circum-neutral (8) and alkaline (14) taxaoccurring in slightly acid and non-acidified softwater lakes. In contrast with the work of Buskens
(1987) we found Ablabesmyia and Psectrocladiusonly in alkaline waters, and while the first of thesetaxa was restricted to five lakes, of which three hadpHP7.0, the second was present in a wide spec-trum of lakes (26) with only one case below 6.0.Again in contrast with Raddum and Sæther (1981)we found the following taxa more frequently inalkaline conditions, but also each in one case atlower pH values: Prodiamesa olivacea was at aminimum pH of 6.06, Eukiefferiella at 5.75,Orthocladius at 5.76 and Pseudosmittia at 5.35. Allof them in very low percentages (<1.0%).
At these altitudes/latitudes, in fact, food avail-ability had little effect on the benthic fauna andwas replaced by temperature and ice-cover dura-tion (Welch, 1976), so that Orthocladiinae pre-vailed where temperatures were lower and siliceousweathering processes occurred, and both were themajor features. On the other hand, becauseChironominae are more resistant to stress andpollution and are ubiquitous, they were mainlydistributed at lower altitudes, where higher trophicconditions and nutrient levels were typical, andfluctuations in temperature were more frequentand might cause short periods of partial de-oxygenation and fluctuation in the water level, twomajor causes of habitat disturbance (Jonasson,1972; Prat et al., 1992).
Environmental parameters thus influence thepresence/absence of species, but also populationdynamics and community variability. In particular,one of these habitat descriptors, the chemistry oflake water, at high altitude is considerably affectedby the chemical composition of atmospheric depo-sition and by climate characteristics (Sommaruga-Wograth et al., 1997; Koinig et al., 1998; Rogoraet al., 2003). These last two factors determine aseries of cascade effects on lakes such as: changesin temperature and precipitation regime, in thehydrological cycle and the extent of ice and snowcover, in solute content and the biological activityand finally in primary production (Schindler et al.,1990, Henriksen & Hessen, 1997; Tait & Thaler,2000). In response to these variations, some moun-tain lakes in the Alps have shown abrupt shifts intheir faunal assemblages with the loss of taxa typicalof these areas. This did in fact occur in Lake PaioneSuperiore (PE; Guilizzoni et al., 1996) where at thebeginning of the XX century Micropsectra radialisdominated the chironomid population in the
158

lake-bottom with a few individuals of Tanytarsussp., and was displaced by the latter species about100 years ago due to climate changes.
The role played by taxonomy in a research onthe geographical distribution of species has beenshown several fold (Orendt, 2000; Stribling et al.,2003; Rossaro et al. present issue; Maiolini et al.present issue). In fact the taxonomical level reachedcould add a degree of uncertainty to the resultsobtained and could become crucial if qualitativeassessments were based on the presence/absence ofparticular taxa. This was one of the first causes ofthe ‘‘reduction’’ of the original Alpine data set.Sampling strategy and sorting also played a keyrole in the list of species obtained. Although due tooverall study design only semi-quantitative and notreplicated samples were available from each lake,we found interesting results when those were con-sidered on a geographical (or altitudinal) compar-ison. The high ratio between number of taxa andnumber of individuals shown by RH_S and BEmight be further explained, despite the samplingstrategy adopted, by the improved methodologyused by the respective labs, which considered notonly larvae, but also pupal exuviae, probably to agreater extent than in other labs, providing a highertaxonomic determination level, as defined byOrendt (2000), Rossaro et al. (present issue) andMaiolini et al. (present issue). Moreover, generaoccurring in very specific habitats were not usuallytaken into account by general collecting methods,which might explain some differences in theirzoogeographical distribution or their spotty pres-ence throughout the Alps. The finding of newspecies corroborate these results, and must beconsidered a stimulus to future research in thissector.
Conclusions
As was generally recognised, chironomids repre-sent the most abundant group of benthic macro-invertebrates both in lakes and in running watersof the Alps, able to exploit a wide range of trophic,temporal and spatial habitats. Notwithstandingthese features and the improvement in studiesconducted in the Alpine area, the knowledgeavailable on this community in high altitudeecosystems was still relatively scanty.
As might be expected of a fauna typical of highaltitude and cold areas, chironomids becomeincreasingly dominate the colder and harsher theclimate grows, accounting for more than half of allinvertebrates. At the same time, however, due tothe structurally simple habitats of Alpine lakes,there is a lower variety of species in these lakesthan in lowland lakes of similar size and incomparable lakes in different mountain chainsacross Europe.
This work can be regarded as a stimulus tocontinuing and extending research leading to asounder basis for future studies on comparativebiogeography. There is a pressing need to pre-serve these mountain environments where globalclimatic changes may cause abrupt shifts in thefaunal assemblages with the loss of taxa typicalof these areas. A few points are relevant to thisgoal:
1. As a general rule, Orthocladiinae and Chiro-nominae, of the Tanytarsini tribe, were themost frequent subfamilies observed in thelittoral area of high altitude lakes of the Alps.Pseudodiamesa branickii, Corynoneura arctica,Heterotrissocladius marcidus and Paratany-tarsus austriacus were the most widespreadspecies, in some cases also representing themost abundant species in a lake. There wasnot any clear relation between number of taxaand altitude.
2. The distribution of taxa based on relativeweighted mean values of pH, conductivityand altitude and respective tolerance showedthat Chironomini (in particular, Chironomussp. and Dicrotendipes sp.) more typical ofthe eastern lakes (JA) and Tanypodinae(Ablabesmyia sp.) present in the central RH_Swere to be considered better indicators of morealkaline waters. Of the taxa restricted to lowerpH (from 5.8 to 6.4), and thought up to now tobe tolerant, Limnophyes sp., Endochironomussp. andOrthocladius (O.) spp. were commonlyfound the former two in the central westernAlps (PE and LE) and the latter in the centraleastern part of the Alps (RH_S). Moreover,almost all the taxa present at higher pH werecontemporary present at conductivity higherthan 100 leq l)1 and altitude lower than2100 m a.s.l.
159

3. The present study identified altitude andtemperature as the main key factors influenc-ing patterns of chironomid community struc-ture, but pH and substratum were asimportant as the first two factors.
4. A strong need emerged to harmonize sam-pling methodologies used at high altitudes andto fix a minimum sample size of Chironomi-dae individuals to obtain sufficient and preciseenough information on the composition of thecommunity. Any deficiency in this step-by-step work makes it difficult to interpret thezoogeographical distribution of taxa, andidentification limited to genus level does notallow the establishment of optimal ecologicalconditions for each species.
5. As a practical result of the international, multi-lab projects, we experienced the deficienciesin chironomid taxonomy. Meetings are highlyrecommended to fine-tune the precision ofidentification, and inter-calibration exercisesamong laboratories for verifying anddiscussingexisting discrepancies in the identification ofspecies and for harmonising larval taxonomy.
Acknowledgements
The research described here was partly fundedby the European Commission EnvironmentProgramme EMERGE project (CE/CNR EVK1-CT-1999-00032), by the Val Grande NationalPark, in the framework of the EU Inter-Reg IIProgramme ‘‘Concetto di paesaggio transfrontali-ero quale sostegno per la creazione di una rete diaree protette tra il Parco Nazionale Val Grande(Italia) e l’area Centovalli-Collina di Maia-LagoVerbano (Confederazione Elvetica)’’, and by theNatural Science Museum of Trento and theAdamello-Brenta Regional Park. Thanks aredue to Øyvind Schnell (Department of Zoology-University of Bergen, Norway) as responsiblefor chironomid validation of the data set ofEMERGE programme and to Gunnar Raddum(same University), Rosario Mosello (CNR-Insti-tute for Ecosystem Study-Verbania Pallanza, Ita-ly) and to Martin Kernan (Environmental ChangeResearch Centre, University College London,England) for providing us with biological, chemi-cal and geographical data.
References
Ashe, P., D. A. Murray, F. Reiss, 1987. The zoogeographical
distribution of Chironomidae (Insecta: Diptera) Annales de
Limnologie 23: 27–60.
Armitage, P., P. S. Cranston & L. C. V. Pinder (eds), 1995. The
Chironomidae: the Biology and Ecology of Non-biting
Midges. Chapmann & Hall, London, 572 pp.
Battarbee, R. W., 1978. Observations on the recent history of
Lough Neagh and its drainage basin. Philosophical Trans-
actions of the Royal Society B 281: 303–345.
Battarbee, R. W., 1984. Spatial variations in the water qua-
lita of Lough Erne, Northern Ireland, on the basis of
surface sediment diatom analysis. Freshwater Biology 14:
539–545.
Boggero, A. & M. Nobili, 1998. Macrobenthic community and
chemical characteristics of four Alpine lakes in Canton
Ticino. Bollettino della Societa Ticinese di Scienze Naturali
86: 17–23.
Boggero, A., & A. M. Nocentini, 1994. Macrozoobentos di un
lago alpino d’alta quota (Lago di Paione Superiore, Val
Bognanco). Proceedings of the X Congress of the ‘‘Associ-
azione Italiana Oceanologia e Limnologia’’ (A.I.O.L.),
Alassio (Italy), November 4–6, 1992: 177–187.
Boggero, A., A. M. Nocentini, M. Nobili & M. Gianatti, 1996a.
Ricerche sulla fauna macrobentonica litorale in laghi d’alta
quota nel bacino imbrifero del Lago Maggiore. Proceedings
of the VII National Congress of the ‘‘Societa Italiana di
Ecologia’’ (S.It.E.) Napoli (Italy), September 11–14, 1996:
83–86.
Boggero, A., A. Marchetto, A. Barbieri, A. Sassi, M. Conedera,
G. A. Tartari & R. Mosello, 1996b. Idrochimica dei laghi
alpini del Canton Ticino (Alpi Centrali) in relazione con la
chimica delle precipitazioni. Documenta dell’Istituto italiano
di Idrobiologia 57: 273 pp.
Bretschko, G., 1974. The chironomid fauna of a high alpine
lake (Vorderer Finstertaler See) Tyrol, Austria, 2237 m asl.
Entomologisk Tidskrift Supplement 95: 22–33.
Buskens, R. F. M., 1987. The chironomid assemblages in
shallow lentic waters differing in acidity, buffering capacity
and trophic level in the Netherlands. Entomologica Scandi-
navica Supplement 29: 217–224.
Cantonati, M., M. Tolotti & M. Lazzara, 2002. I laghi del
Parco Naturale Adamello-Brenta. Ricerche limnologiche su
laghi d’alta quota del settore siliceo del Parco. Documenti
del Parco 14: 285.
Dobravec, J. & M. Sisko, 2002. Geographical location and
description of the lakes. In A. Brancelj (ed.), High-mountain
Lakes in the Eastern Part of the Julian Alps. ZRC Pub-
lishing, Ljubljana, 49–76.
Ferrarese, U., 1990. Contributo alla conoscenza dei Chirono-
midi del Trentino (Diptera, Chironomidae). Studi Trentini di
Scienze Naturali-Acta Biologica 66: 171–185.
Ferrarese, U., 2003. Chironomidi (Diptera, Chironomidae) di
alcuni torrenti, laghi e zone umide del Trentino, con segna-
lazione di specie nuove per l’Italia. Studi Trentini di Scienze
Naturali-Acta Biologica 79: 203–211.
Fjellheim, A., A. Boggero, G. A. Halvorsen, A. M. Nocen-
tini, M. Rieradevall, G. G. Raddum & Ø. A. Schnell,
160

2000. Distribution of benthic invertebrates in relation to
environmental factors. A study of European remote alpine
lake ecosystems. Verhandlugen der Internationale Vereini-
gung fur Theoretische und Angewandte Limnologie 27:
484–488.
Franceschini, A., & V. Lencioni, 2002. Lo zoobenthos. In
Cantonati M., M. Tolotti & M. Lazzara (eds), I laghi del
Parco Adamello-Brenta. Ricerche limnologiche su laghi
d’alta quota del settore siliceo del Parco. Parco Adamello-
Brenta, Strembo (Trento), Documenti del Parco 14: 245–
266 pp.
Garibaldi, L., R. Mosello, B. Rossaro & F. Setti, 1987.
Caratteristiche chimiche e biologiche di due laghi d’alta
quota suscettibili ad acidificazione. In Mosello, R. & B. De
Margaritis (eds), Deposizioni acide: Un Problema per Acque
e Foreste. Documenta dell’Istituto italiano di Idrobiologia
14: 165–180.
Guilizzoni, P., A. Marchetto, A. Lami, N. G. Cameron, P. G.
Appleby, N. L. Rose, Ø. A. Schnell, C. A. Belis, A. Giorgis &
L.Guzzi, 1996. The environmental hystory of amountain lake
(Lago Paione Superiore, Central Alps, Italy) for the last c.
100 years: A multidisciplinary, palaeolimnological study.
Journal of Paleolimnology 15: 245–264.
Guthruf, J., K. Guthruf-Seiler & M. Zeh, 1999. Kleinsee im
Kanton Bern. Bern, Paul Haupt AG, 229 pp.
Hansen, C. M. E., M. Tolotti, R. Ettinger, H. Thies, D. Tait &
R. Psenner, 2002. The application of geographic information
science (GIS) for data synthesis and evaluation of remote
high mountain lakes in the lake district Tyrol (Austria &
Italy). Informatics for Environmental Protection 1: 597–604.
Heiri, O., A. F. Lotter, S. Hausmann & F. Kienast, 2003. A
chironomid-based Holocene summer air temperature recon-
struction from the Swiss Alps. The Holocene 13: 477–484.
Henriksen, A. & D. O. Hessen, 1997. Whole catchment studies
on nitrogen cycling: nitrogen from mountains to fjords.
Ambio 26: 254–257.
Jonasson, P. M., 1972. Ecology and production of the
profundal benthos. Oikos Supplement 14: 1–148.
Kansanen, P. H., J. Aho & L. Paasivirta, 1984. Testing the
benthic lake type concept based on chironomid associations
in some Finnish lakes using multivariate statistical methods.
Annales Zoologici Fennici 21: 55–76.
Koinig, K. A., R. Schmidt, S. Sommaruga-Wograth, R. Som-
maruga, R. Tessadri & R. Psenner, 1998. Climate change as
the primary cause for pH shifts in a high alpine lake. Water
Air and Soil Pollution 104: 167–180.
Laville, H., 1972. Recherches ecologiques sur les Chironomides
(Diptera) des lacs de montagne. Doctoral Thesis. Universite
P. Sabatier, Toulouse, 414 pp.
Laville, H. & B. Serra-Tosio, 1987. Chironomides (Diptera) du
Massif Central et des basses regions avoisinantes. Annales de
Limnologie 23(2): 135–145.
Laville, H. & A. Vilchez-Quero, 1986. Les Chironomides
(Diptera) de quelques ‘‘lagunas de haute altitude de la Sierra
Nevada (Granada, Espagne). Annales de Limnologie 22(1):
53–63.
Lenz, F., 1927. Chironomiden aus norwegischen Hochgebirgs-
seen. Zugleich ein Beitrag zur Seetypenlehre. Nytt Magazin
for Naturvidenskabene 66: 111–192.
Leuven, R. S. E. W., J. A. Velden, J. A. M. Vanhemelrijk &
G. Velde, 1987. Impact of acidification on chironomid
communities in poorly buffered waters in the Netherlands.
Entomologica scandinavica Supplement 29: 269–280.
Lotter, A. F., H. J. B. Birks, W. Hofmann & A. Marchetto,
1997. Modern diatom, cladocera, chironomid, and chryso-
phyte cyst assemblages as quantitative indicators for the
reconstruction of past environmental conditions in the Alps.
I. Climate. Journal of Paleolimnology 18: 395–420.
Lotter, A. F., H. J. B. Birks, W. Hofmann & A. Marchetto,
1998. Modern diatom, cladocera, chironomid, and chryso-
phyte cyst assemblages as quantitative indicators for the
reconstruction of past environmental conditions in the Alps.
II. Nutrient. Journal of Paleolimnology 19: 443–463.
Lundbeck, J., 1936. Untersuchungen uber die Bodenbesiedlung
der Alpenrandseen. Archiv fur Hydrobiologie Supplement
10: 207–358.
Madanes, N. & J. R. Dadon, 1998. Assessment of the minimum
sample size required to characterize site-scale airborne pol-
len. Grana 37: 239–245.
Mietto, S., B. Rossaro & V. Lencioni, 1997. The Chironomid
species of the Italian lakes: A review. In O. Hoffrichter (ed.),
Late 20th Century Research on Chironomidae: An Anthol-
ogy from the 13th International Symposium on Chironom-
idae. Schaker Verlag, Aachen, 549–564.
Mosello, R., A. Boggero, M. Carmine, A. Marchetto, A. Sassi
& G. A. Tartari, 1994. Ricerche idrochimiche sui laghi alpini
delle valli Ossola e Sesia (Alpi Pennine e Lepontine). Doc-
umenta dell’Istituto italiano di Idrobiologia 46: 436 pp.
Mosello,R.,A.Lami, P.Guilizzoni,M.Manca,A.M.Nocentini,
A. Pugnetti, A. Boggero, A. Marchetto, G. A. Tartari,
R. Bettinetti, M. Bonardi & P. Cammarano, 1993. Limno-
logical studies on two acid sensitive lakes in the south western
Alps (lakes Paione Superiore and Paione Inferiore, Italy).
Memorie dell’Istituto italiano di Idrobiologia 51: 127–146.
Orendt, C., 2000. Chironomids of small Alpine water bodies
(springs, spring brooks, pools, small lakes) of the northern
Calcareous Alps (Insecta, Diptera, Chironomidae). In:
Baehr M. & M. Spies, (eds), Contributions to Chironomid
Research in Memory of Dr. Friedrich Reiss. Spixiana, 23(2):
121–128.
Prat, N., M. Real & M. Rieradevall, 1992. Benthos of Spanish
lakes and reservoirs. Limnetica 8: 221–229.
Pugnetti, A., M. Manca, A. M. Nocentini, A. Boggero,
R. Bettinetti, M. Bonardi & P. Cammarano, 1993. Ecologi-
cal aspects of two Alpine lakes (Lakes Paione, Italy). Pro-
ceedings of the 5th International Conference, Conservation
and management of lakes: Strategies for Lake Ecosystems
Beyond 2000. Stresa (VB, Italy), May 17–21, 1993: 440–443.
Raddum, G. G. & O. A. Sæther, 1981. Chironomid commu-
nities in Norwegian lakes with different degrees of acidifi-
cation. Verhandlugen der Internationale Vereinigung fur
Theoretische und Angewandte Limnologie 21: 399–405.
Reiss, F., 1968. Verbreitung lakustrischer Chironomiden
(Diptera) des Alpengebietes. Annales Zoologici Fennici 5:
119–125.
Reiss, F. & E. J. Fittkau, 1971. Taxonomie und Okologie eu-
ropaisch verbreiteter Tanytarsus-Arten (Chironomidae,
Diptera). Archiv fur Hydrobiologie Supplement 40: 75–200.
161

Rieradevall, M. & N. Prat, 1999. Chironomidae from high
mountain lakes in Spain and Portugal. In O. Hoffrichter (ed.),
Late 20th Century Research on Chironomidae: An Anthol-
ogy from the 13th International Symposium on Chironomi-
dae. Shaker Verlag, Aachen, 605–613. ISBN 3–8265–7431–1.
Rogora, M., R. Mosello & S. Arisci, 2003. The effect of climate
warming on the hydrochemistry of alpine lakes. Water Air
and Soil Pollution 148: 347–361.
Rossaro, B., 1991a. Chironomids and water temperature.
Aquatic Insects 13: 87–98.
Rossaro, B., 1991b. Factors that determine Chironomidae
species distribution in fresh waters. Bollettino di Zoologia
58: 281–286.
Rossaro, B., 1992. Chironomids collected in the Western and
Central Alps (Diptera, Chironomidae). Studi Trentini di
Scienze Naturali-Acta Biologica 67: 181–195.
Rull, V., 1987. A note on pollen counting in Palaeoecology.
Pollen et Spores XXIX: 471–480.
Sæther, O. A., 1975. Nearctic chironomids as indicators of lake
typology. Verhandlugen der Internationale Vereinigung fur
Theoretische und Angewandte Limnologie 19: 3127–3133.
Sæther, O. A., 1979. Chironomid communities as water quality
indicators. Holarctic Ecology 2: 65–74.
Schindler, D. W., K. G. Beaty, E. J. Fee, D. R. Cruikshnak,
E. R. DeBruyn, D. L. Findlay, G. A. Linsey, J. A. Shearer,
M. P. Stainton & M. A. Turner, 1990. Effects of climatic
warming on lakes of the Central Boreal Forest. Science 250:
967–970.
Schnell, O. A., M. Rieradevall, I. Granados & O. Hanssen,
1999. A Chironomid taxa coding system for use in ecological
and palaeoecological databases. NIVA Report No. 3710–97,
Molar Project Manual, Annex B, 19 pp.
Serra-Tosio, B. & H. Laville, 1991. Liste annotee des Dipteres
Chironomides de France continentale et de Corse. Annales
de Limnologie 27: 37–74.
Serra-Tosio, B., 1989. Chironomides (Diptera) des Alpes
francaises et des bases regions avoisinantes. Annales de
Limnologie 25: 159–175.
Sommaruga-Wograth, S., K. A. Koinig, R. Schmidt, R. Som-
maruga, R. Tessadri & R. Psenner, 1997. Temperature effects
on the acidity of remote alpine lakes. Nature 387: 64–67.
Stribling, J. B., S. R. Moulton, II & G. T. Lester, 2003.
Determining the quality of taxonomic data. Journal of
North American Benthological Society 22: 621–631.
Tait, D. & B. Thaler, 2000. Atmospheric deposition and lake
chemistry trends at a high mountain site in the Eastern Alps.
Journal of Limnology 59: 61–71.
ter Braak, C. J. F., 1987. Calibration. In Jongman R. H. G.,
C. J. F. ter Braak & O. F. R. Tongeren (eds.), Data Analysis
in Community and Landscape Ecology. Pudoc, Wagenin-
gen, The Netherlands, 79–90.
ter Braak, C. J. F. & P. Smilauer, 2002. Canoco reference
manual and canodraw for Windows. User’s guide (version
4.5). Biometris, Wageningen and Ceske Budejovice: 499 pp.
Thienemann, A., 1936. Alpine Chironomiden (Ergebnisse von
Untersuchungen in der Gegend von Garmisch-Partenkir-
chen, Oberbayern). Archiv fur Hydrobiologie 30: 167–262.
Thienemann, A., 1954..
Thies, H., U. Nickus, C. Arnold, R. Schnegg, A. Wille &
R. Psenner, 2000. Biogeochemistry of a high mountain lake
in the Austrian Alps. Verhandlugen der Internationale
Vereinigung fur Theoretische und Angewandte Limnologie
27: 517–520.
Tolotti, M., H. Thies, M. Cantonati, C. M. E. Hansen &
B. Thaler, 2003. Flagellate algae (Chrysophyceae, Dino-
phyceae, Cryptophyceae) in 48 high mountain lakes of the
Northern and Southern slope of the Eastern Alps: Biodi-
versity, taxa distribution and their driving variables. Hyd-
robiologia 502: 331–348.
Wathne, B. M., S. T. Patrick, D. Monteith & H. Barth, 1995.
AL:PE Project. Part 1. April 1991–April 1993. Report
EUR16129EN. EuropeanCommission, Luxembourg, 296 pp.
Welch, H. E., 1976. Ecology of Chironomidae (Diptera) in a
polar lake. Journal of the Fisheries Research Board of
Canada 33: 227–247.
162
Chironomid
taxafoundin
thedifferentstudyareas,code(Schnellet
al.,1999)andnumber
oflakes
wherethey
occurred
Taxa
Code
Alpinesector
BE
PE
LE
RH_N
RH_S
DO
NO_N
NO_S
JAALPS
N.lakes
10
915
11
16
111
214
89
Zavrelim
yia
spp.
Zavyind
2I
9F
14
F5
F7
F–
8F
11
3F
49
F
Paratanytarsusaustriacus(K
ieffer)
Part
aus
8VF
8F
11
F7
A2
F–
8VF
––
44
VF
Heterotrissocladiusmarcidus(W
alker)
Hetegmar
3VF
8VF
9VF
2F
9F
–7
F1
VF
–39
F
Micropsectra
spp.
Micrind
1I
3I
10
F3
VF
7VF
–1
F–
6F
31
F
Paratanytarsusspp.
Partind
–3
I5
F–
7F
1F
–2
A13
A31
VF
Macropelopia
sp.
Macrind
1I
2I
8F
1F
6F
–1
I–
4F
23
F
Corynoneura
spp.
Coryind
2VF
–1
F3
I6
VF
–8
F1
F1
122
F
Corynoneura
arctica
Kieffer
Cory
arc
––
–6
VF
5A
–9
VF
1F
–21
VF
Procladius.spp.
Procind
1I
1I
6F
1I
2F
–2
I–
6F
19
F
Tanytarsinijuv.
Trtanyt
––
–8
2–
7–
–17
Orthocladiinaejuv.
Sfortho
––
–3
5–
8–
–16
Lim
nophyes
sp.
Lim
nind
2I
4F
7I
–2
I–
––
–15
I
Prodiamesaolivacea(M
eigen)
Prodoli
–2
I5
I–
1I
–3
I1
F3
115
I
Cricotopus(I.)sp.
Criciso
–1
5F
–1
F–
5F
––
12
F
Pseudodiamesa(Pseudo.)
branickii(N
owicki)
Psedbra
2F
–1
I4
VF
2VF
1F
2A
––
12
VF
Tanytarsussp.
Tanyind
–2
I2
I–
2F
1VF
3I
–2
VF
12
F
Corynoneura
scutellata
(Winnertz)
Cory
scu
–1
VF
8VF
––
––
––
9VF
Tanytarsuslugensgr.
Tanyglug
1VF
1I
7F
––
––
––
9F
Psectrocladius(P.)
sordidellusgr.
Psecgsor
–1
I3
F–
3F
––
–1
I8
F
Pseudodiamesanivosa
(Goetghebuer)
Psedniv
1I
1I
–2
F–
1F
2VF
1I
–8
F
Apsectrotanypustrifascipennis(Zetterstedt)
Apse
tri
––
6I
–1
I–
––
–7
I
Psectrocladius(P.)
limbatellusgr.
Psecglim
–3
I3
F–
––
–1
I–
7F
Psectrocladius(P.)
octomaculatusWulker
Psecoct
6F
––
––
––
––
6F
Corynoneura
lacustris(Edwards)
Cory
lac
–3
VF
2F
––
––
––
5VF
Endochironomusdispargr.
Endogdis
–1
VF
4I
––
––
––
5F
Micropsectra
radialis(G
oetghebuer)
Micrrad
3VF
2I
––
––
––
–5
F
Appendix
Continued
onp.164
163

Paracladiuscf.alpicola
Parl?alp
–1
F–
2F
––
2I
––
5F
Tanypodinaejuv.
sftanyp
––
–3
2–
––
–5
Zavrelim
yia
punctatissim
a(G
oetghebuer)
Zavypun
––
––
5I
––
––
5I
Ablabesmyia
monilis(Linnaeus)
Abla
mon
2VF
––
–2
F–
––
–4
F
Chaetocladiussp.
Chaeind
–1
I–
1F
1F
––
–1
I4
F
Chironomusthummigr.
Chirgthu
1I
–1
I–
––
––
1F
4F
Dicrotendipes
lobiger
(Kieffer)
Dicrlob
2I
––
––
––
–2
A4
VF
Larsia
sp.
Larsind
––
––
1F
––
–3
F4
F
Paracladiussp.
Parlind
––
3I
––
–1
I–
–4
I
Pentaneurinijuv.
Trpenta
––
––
3–
–1
–4
Smittiasp.
Smitind
––
3I
1I
––
––
–4
I
Cardiocladiussp.
Cardind
––
3I
––
––
––
3I
Chironominaejuv.
Sfchiro
1I
––
–2
––
––
3
Cricotopussp.
Cricind
–1
I2
I–
––
––
–3
I
Orthocladius(O.)
s.str.
Orthort
––
––
3F
––
––
3F
Paracladopelmacamptolabis/laminata
Pard?cam
–2
I–
–1
I–
––
–3
I
Parorthocladiusnudipennis(K
ieffer)
Paornud
–1
I1
I–
––
–1
I–
3I
Protanypussp.
Protind
–1
I2
I–
––
––
–3
I
Pseudodiamesasp.
Psepind
––
3I
––
––
––
3I
Tvetenia
calvescens(Edwards)
Tvet
cal
–2
I1
I–
––
––
–3
I
Acamptocladiusreissi(C
ranston&
Sæther)
Acam
rei
––
1I
–1
F–
––
–2
I
Bryophaenocladiussubvernalis(Edwards)
Bryosub
–2
I–
––
––
––
2I
Corynoneura
lobata
(Edwards)
Cory
lob
––
2I
––
––
––
2I
Cricotopus(C.)
trem
ulus(Linnaeus)
Crictre
––
––
––
––
2I
2I
Appendix
(Continued)
Taxa
Code
Alpinesector
BE
PE
LE
RH_N
RH_S
DO
NO_N
NO_S
JAALPS
N.lakes
10
915
11
16
111
214
89
164

Cricotopus(I.)laricomalis(Edwards)
Criclar
––
–1
F1
I–
––
–2
F
Diamesaspp.
Diamind
–1
I1
I–
––
––
–2
I
Eukiefferiellaminor(Edwards)
Eukimin
––
––
2F
––
––
2F
Metriocnem
ushygropetricusgr.
Metrghyg
––
––
2F
––
––
2F
Natarsia
sp.
Nataind
––
––
––
––
2I
2I
Orthocladius(Eud.)
sp.
Ortheud
––
2I
––
––
––
2I
Paracricotopussp.
Parrind
–1
I–
––
–1
I–
–2
I
Paramerinadivisa(W
alker)
Parm
div
––
––
––
––
2I
2I
Parametriocnem
ussp.
Pareind
––
––
2I
––
––
2I
Parakiefferiellasp.
Parkind
––
––
1I
––
––
1I
Parakiefferiellatriquetra
(Pankratova)
Park
tri
––
––
1I
––
––
1I
Paramerinasp.
Parm
ind
––
––
1I
––
––
1I
Parasm
ittiasp.
Parsind
–1
I–
––
––
––
1I
Paratrichocladiusnivalis(G
oetghebuer)
Patr
niv
––
––
––
1I
––
1I
Polypedilum
convictum
gr.
Polygcon
––
––
––
––
1I
1I
ProtanypuscaudatusEdwards
Protcau
––
––
––
1I
––
1I
Psectroclacdius(Allop.)
obvius(W
alker)
Psecobv
––
––
––
––
1I
1I
Psectrocladius(Allo.)
sp.
PsecA
ind
––
1I
––
––
––
1I
Psectrocladius(Monop.)
sp.
PseeM
ind
–1
I–
––
––
––
1I
Thienem
annim
y,iaspp.
Srthiy
––
––
1I
––
––
1I
Stictochironomussp.
Sticind
––
1I
––
––
––
1I
StilocladiusmontanusRossaro
Stilmon
––
––
–1
F–
––
1F
Symposiocladiussp.
Sympind
1F
––
––
––
––
1F
Tanytarsuspallidicornisgr.
Tanygpal
–1
F–
––
––
––
1F
Trissopelopia
sp.
Trisind
––
––
1I
––
––
1I
Tvetenia
sp.
Tvetind
––
1I
––
––
––
1I
Zavrelim
yia
melanura
(Meigen)
Zavymel
––
––
1I
––
––
1I
ForAlpinesectorcodeseeTable
1.Samedistributionfortheentire
alpinechain.Letters
refers
tothemeanfrequencies
distributionexperiencedbyeach
taxon.I,infrequent(0–
5%
);F,frequent(5–10%
);VF,veryfrequent(10–20%
);A,abundant(>
20%
).
165
Related Documents











