Lipopeptide particles as the immunologically active component of CTL inducing vaccines Ikuo Tsunoda a , Alessandro Sette b , Robert S. Fujinami a , Carla Osero b , Jorg Ruppert b , Carol Dahlberg b , Scott Southwood b , Thomas Arrhenius c , Li-Qing Kuang a , Ralph T. Kubo c , Robert W. Chesnut b , Glenn Y. Ishioka b, * a Department of Neurology, University of Utah School of Medicine, 50 North Medical Dr., Salt Lake City, UT 84132, USA b Epimmune Inc., 6555 Nancy Ridge Dr. #200, San Diego, CA 92121, USA c Cytel Corporation, 3525 John Hopkins Ct., San Diego, CA 92121, USA Received 30 January 1998; received in revised form 4 June 1998; accepted 2 July 1998 Abstract Using a bipalmitoylated lipopeptide consisting of an ovalbumin helper T-cell epitope covalently linked to an influenza virus cytotoxic T-lymphocyte (CTL) epitope, we addressed possible factors that may be critical for CTL induction. Antigen processing of lipopeptide appears to be required for T-cell induction since there was virtually no in vitro binding of lipopeptide to purified MHC molecules. A major portion of lipopeptide immunogenicity was due to its particulate nature inasmuch as CTL induction in mice correlated with insoluble lipopeptide constructs, whereas more soluble analogs were significantly less immunogenic. Immunohistological analysis of tissue from immunized animals revealed that lipopeptide migration from the s.c. injection site to the spleen could be detected as early as 1 h after immunization and cell-associated lipopeptide was observed on macrophages and dendritic cells, implicating both cell populations in the processing and presentation of lipopeptide particles to CTLs. # 1999 Elsevier Science Ltd. All rights reserved. Keywords: Lipopeptide particles; Cytotoxic T-lymphocytes; Antigen processing 1. Introduction The importance of class I MHC-restricted cytotoxic T-lymphocytes (CTLs) in conferring protection in the host against viral infection is well established [1–4]. Since CTLs recognize short peptide fragments of viral antigen (Ag) presented in association with class I MHC molecules, a possible strategy for the development of CTL-inducing vaccines has been to use peptide epitopes as the immunogen. A particularly successful strategy for enhancing the immunogenicity of CTL epitopes uti- lizes covalent attachment of long-chain lipids to the peptide moiety. In vivo studies in various animal species have shown that lipidation of T- and B-cell epitopes result in induction of strong antibody (Ab), helper T- lymphocyte (Th), and CTL responses [5–9]. Our labora- tory has developed a modular lipopeptide vaccine con- struct containing two palmitic acid (PAM) residues and a peptide unit composed of a Th and CTL peptide epi- tope covalently linked by a trialanine spacer. Bipalmitoylated Th–CTL constructs containing selected immunodominant epitopes gave vigorous and durable CTL responses in mice [10]. Similarly, we observed strong CTL induction in normal human subjects who were vaccinated with a lipopeptide containing a domi- nant hepatitis B virus (HBV), HLA-A2.1-restricted CTL peptide and a universal tetanus toxin Th epitope [10], the magnitude of which was comparable to that induced during acute HBV infection [11]. These findings help to validate the potential use of lipopep- tides for therapeutic and prophylactic treatment of dis- eases involving CTL immunity. Although lipopeptides are potent T-cell immunosti- mulators, the mechanism underlying their in vivo immunogenicity has yet to be adequately explained. In the following report we examine potential factors that Vaccine 17 (1999) 675–685 0264-410X/99/$19.00 # 1999 Elsevier Science Ltd. All rights reserved. PII: S0264-410X(98)00250-3 * Corresponding author. Tel.: +1-619-404-7171; fax: +1-619-404- 7177.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Lipopeptide particles as the immunologically active component ofCTL inducing vaccines
Ikuo Tsunodaa, Alessandro Sette b, Robert S. Fujinami a, Carla Osero�b,Jorg Ruppert b, Carol Dahlberg b, Scott Southwoodb, Thomas Arrhenius c,
Li-Qing Kuanga, Ralph T. Kuboc, Robert W. Chesnutb, Glenn Y. Ishiokab, *aDepartment of Neurology, University of Utah School of Medicine, 50 North Medical Dr., Salt Lake City, UT 84132, USA
bEpimmune Inc., 6555 Nancy Ridge Dr. #200, San Diego, CA 92121, USAcCytel Corporation, 3525 John Hopkins Ct., San Diego, CA 92121, USA
Received 30 January 1998; received in revised form 4 June 1998; accepted 2 July 1998
Abstract
Using a bipalmitoylated lipopeptide consisting of an ovalbumin helper T-cell epitope covalently linked to an in¯uenza viruscytotoxic T-lymphocyte (CTL) epitope, we addressed possible factors that may be critical for CTL induction. Antigen processing
of lipopeptide appears to be required for T-cell induction since there was virtually no in vitro binding of lipopeptide to puri®edMHC molecules. A major portion of lipopeptide immunogenicity was due to its particulate nature inasmuch as CTL inductionin mice correlated with insoluble lipopeptide constructs, whereas more soluble analogs were signi®cantly less immunogenic.
Immunohistological analysis of tissue from immunized animals revealed that lipopeptide migration from the s.c. injection site tothe spleen could be detected as early as 1 h after immunization and cell-associated lipopeptide was observed on macrophagesand dendritic cells, implicating both cell populations in the processing and presentation of lipopeptide particles to CTLs. # 1999
Elsevier Science Ltd. All rights reserved.
Keywords: Lipopeptide particles; Cytotoxic T-lymphocytes; Antigen processing
1. Introduction
The importance of class I MHC-restricted cytotoxic
T-lymphocytes (CTLs) in conferring protection in the
host against viral infection is well established [1±4].
Since CTLs recognize short peptide fragments of viral
antigen (Ag) presented in association with class I MHC
molecules, a possible strategy for the development of
CTL-inducing vaccines has been to use peptide epitopes
as the immunogen. A particularly successful strategy
for enhancing the immunogenicity of CTL epitopes uti-
lizes covalent attachment of long-chain lipids to the
peptide moiety. In vivo studies in various animal species
have shown that lipidation of T- and B-cell epitopes
result in induction of strong antibody (Ab), helper T-
lymphocyte (Th), and CTL responses [5±9]. Our labora-
tory has developed a modular lipopeptide vaccine con-
struct containing two palmitic acid (PAM) residues and
a peptide unit composed of a Th and CTL peptide epi-
tope covalently linked by a trialanine spacer.
Bipalmitoylated Th±CTL constructs containing selected
immunodominant epitopes gave vigorous and durable
CTL responses in mice [10]. Similarly, we observed
strong CTL induction in normal human subjects who
were vaccinated with a lipopeptide containing a domi-
nant hepatitis B virus (HBV), HLA-A2.1-restricted
CTL peptide and a universal tetanus toxin Th
epitope [10], the magnitude of which was comparable to
that induced during acute HBV infection [11]. These
®ndings help to validate the potential use of lipopep-
tides for therapeutic and prophylactic treatment of dis-
eases involving CTL immunity.
Although lipopeptides are potent T-cell immunosti-
mulators, the mechanism underlying their in vivo
immunogenicity has yet to be adequately explained. In
the following report we examine potential factors that
Vaccine 17 (1999) 675±685
0264-410X/99/$19.00 # 1999 Elsevier Science Ltd. All rights reserved.
PII: S0264-410X(98 )00250 -3
* Corresponding author. Tel.: +1-619-404-7171; fax: +1-619-404-
7177.

may contribute to the immunogenicity of a model lipo-peptide containing the well characterized H-2Kd-restricted in¯uenza nucleoprotein 147±155 (FLU) pep-tide and the I-Ad-restricted ovalbumin 323±336 (OVA)Th epitope, both covalently linked and lipidated at theN-terminus by addition of two PAM residues.Elucidation of the mechanisms of lipopeptide immuno-genicity was explored by examining the capacity of thelipidated OVA±FLU peptide to bind puri®ed MHCmolecules, the immunogenicity of particulate and sol-uble lipopeptide constructs, and the in vivo tra�ckingof lipopeptide and their association with antigen pre-senting cells (APCs).
2. Materials and methods
2.1. Peptides and lipopeptides
Peptides were synthesized according to standard F-moc solid phase synthesis methods, which have beenpreviously described [12]. The peptides used in thisstudy include OVA (ISQAVHAAHAEINE), FLU(TYQRTRALV), and the linked OVA±FLU peptide(ISQAVHAAHAEINEAAATYQRTRALV).
All lipopeptides used in this study contain theOVA±FLU linked construct. The standard lipopeptideconstruct (PAM)2±OVA±FLU ((PAM)2KSSISQA-VHAAHAEINEAAATYQRTRALV) contains twoPAM residues attached via the a- and e-amino groupsof a lysine that is part of a lysyl-serine±serine spacer atthe amino terminus of the OVA±FLU construct.Analogs containing lipid tails of varying carbon chainlengths were also used in some experiments. All lipo-peptides were prepared by coupling the preformedsymmetrical anhydride of octanoic, decanoic, dodeca-noic, tetradecanoic, or PAM, to the a- and e-aminogroups of the lysine residue located at the amino-ter-minus of the resin-bound elongated peptide.Monopalmitoylated lipopeptide was synthesized bycoupling PAM to resin-bound peptide, bearing amono-Boc protected lysine residue at the amino termi-nus. In some experiments, analogs of the (PAM)2±OVA±FLU lipopeptide containing a DDDDD orKKKKK spacer instead of the standard SS betweenthe OVA epitope and the palmitoylated lysine residuewere synthesized.
The biotinylated (PAM)2±OVA±FLU analog wassynthesized by incorporating an e-allyloxycarbonylprotected lysine residue at position 147 in place ofthreonine in the FLU peptide. Prior to cleavage of thepeptide from the resin, the allyloxycarbonyl protectinggroup was removed by exposure to tetrakis(triphenyl-phosphine) palladium (o) in the presence of morpho-line. Biotin was then coupled to the free e-amino
group of the lysine residue by treatment with diisopro-pylcarbodiimide and hydroxybenzotriazole.
For puri®cation, lipopeptides were subjected to re-petitive washes in 50% acetic acid, followed by twowashes in water and the washed material was sub-sequently lyophilized. Puri®cation of unlipidated pep-tides was accomplished by conventional HPLCmethods. Peptide purity was determined by analyticalreverse-phase HPLC and their composition con®rmedby amino acid analysis, sequencing, and/or mass spec-trometry.
2.2. MHC-peptide binding assay
The mouse mastocytoma P815 (Kd, Dd) and lym-phomas EL-4 (Kb, Db), A20-1.11 (I-Ad/Ed), andLB27.4 (I-Ab), all obtained from ATCC (Rockville,MD), were used as sources for H-2 molecules. MHCisolation was performed by a�nity chromatographyusing the MKD6 (anti-I-Ad), 14.4.4 (anti-I-Ed), Y3JP(anti-I-Ab), 34-5-8S (anti-Dd) and M1/42 (pan anti-class I) Abs [12, 13].
Assays that measure binding of peptides to detergentsolubilized MHC class I and II molecules on the basisof the inhibition of the binding of a radiolabeled stan-dard probe peptide, were performed as describedpreviously [12, 13]. Brie¯y, puri®ed MHC class I and IImolecules were incubated with 1 to 10 nM 125I-radio-labeled peptide probes and varying concentrations ofunlabeled (test) peptide inhibitors for 48 h in PBS con-taining 5% DMSO and 0.05% NP-40. A protease in-hibitor cocktail consisting of PMSF (1.3 nM ®nal),phenanthroline (1:10), pepstatin A (73 mM), EDTA (8mM), N-ethylmaleimide (6 mM), and N-a-p-tosyl-L-lysine chloromethyl ketone (200 mM) was also addedto the peptide mixture. For MHC class I bindingassays, 1 mg/ml of human b-2-microglobulin (ScrippsLaboratories, San Diego, CA) was also included. Atthe end of the incubation period, the percent of MHCbound radioactivity was calculated by size-exclusiongel chromatography [14]. For competitive assays, theconcentration of test peptide yielding 50% inhibitionof the binding of the radiolabeled probe (IC50%) wasalso calculated as described [12, 13].
The peptides that served as standards in the compe-tition binding assay are as follows (sequence, IC50%for restricting element): the adenovirus E1A analog(SGPSNTYPEI, 4.1 nM for Db); HIV-IIIB envelopeglycoprotein analog (RGPYRAFVTI, 3.7 nM for Dd);vesicular stomatitis virus nucleoprotein 52±59(RGYVFQGL, 3.0 nM for Kb); a Kd pool sequencingconsensus peptide (KFNPMKTYI, 1.1 nM for Kd);the ROIV reiterative peptide (YAHAAHAA-HAAHAAHAA, 28 nM for I-Ab); OVA (110 nM forI-Ad); and lambda repressor 12±26 (LEDARRLK-AIYEKKK, 150 nM for I-Ed).
I. Tsunoda et al. / Vaccine 17 (1999) 675±685676

2.3. Lipopeptide formulation
Lyophilized lipopeptide was ®rst solubilized at 10mg/ml in DMSO (Sigma) containing 0.1% tri¯uoroace-tic acid (TFA) with warming to 458C then aliquotedinto vials that were stored at ÿ208C. Two di�erent for-mulation methods were used to generate a particulatesuspension. In the ®rst method, lipopeptide was thawedat 458C for 10 min then diluted 1:10 (v/v) in room tem-perature PBS (Gibco BRL) and mixed immediately byvortexing for 30±60 s. In the second method, PBS con-taining 10% DMSO and 0.01% TFA was added to lyo-philized powder to achieve a 1 mg/ml suspension (w/v)and mixed by vortexing for 2 min, then brie¯y sonicatedin a bath sonicator (Branson Ultrasonics, Danbury,CT). Lipopeptide suspensions were diluted further inPBS containing 10% DMSO and 0.01% TFA toachieve concentrations required for injecting mice.
2.4. In vivo CTL immunization and measurement ofCTL activity
Eight±twelve week old female BALB/cByJ or (C57BL/6�DBA/2)F1 mice (Jackson Laboratory, Bar Harbor,ME)were immunized with lipopeptide s.c. at the tail base.Eleven±fourteen days later, 3� 107 splenocytes fromeach immunized animal were cultured with 10 mg/ml ofthe FLU peptide in an upright 25 cm2 ¯ask (Costar).Culture medium consisted of RPMI 1640 supplementedwith 10%FBS, 4mMglutamine, 50 mM2-ME, 100 mg/mlstreptomycin, and 100U/ml penicillin. Six days later, cul-tures were harvested and cells assayed for cytotoxic ac-tivity in a 51Cr release assay [10]. The level of cytotoxicactivity was determined by calculating the percentagelysis of target cells as described [10]. Tomore readily com-pare responses, the data were converted into lytic units(LU) per 106 e�ector cells with one LU de®ned as the lyticactivity required to achieve 30% lysis of target cells at a100:1 E:T ratio. Speci®c CTL activity (DLU) was calcu-lated by subtracting the LU value obtained in the absenceof peptide from the LUvalue obtainedwith peptide.
2.5. Analysis of lipopeptide solubility and particle size
A 1 mg/ml suspension of lipopeptide was divided intotwo aliquots. One aliquot was centrifuged for 10 min at10,000 rpm and the supernate collected. After furtherdiluting 1:2 (v/v) with DMSO, the protein concentrationof the supernate and the untreated suspension wasmeasured using the BCA protein assay (Pierce,Rockford, IL). The percentage solubility of the samplewas then calculated by dividing its supernatant concen-tration by the total sample protein concentration.
Particle size analysis was performed by Aveka(Woodbury, MN) with a Horiba LA-910 laser scatter-ing particle size distribution analyzer.
2.6. Immunohistological analysis of tissue sections
At various times after immunization, mice were per-fused under halothane anesthesia and 5 mm thick par-a�n or frozen sections were made of skin, spleen, anddraining lymph nodes. Biotinylated lipopeptide wasdetected in situ by avidin±biotin±peroxidase complex(ABC), ¯uorescein streptoavidin, or rhodamine avidinDCS (Vector Laboratories, Burlingame, CA). Adjacentsections were stained with hematoxylin and eosin.
To de®ne the lipopeptide-bearing cell type observedin situ, sections were double-labeled with mononuclearcell markers and ¯uorochrome-avidin for biotin-lipopeptide [15, 16]. Monocyte/macrophages wereidenti®ed by using the F4/80 monoclonal Ab(ATCC) [17] while DCs were detected by the N418(anti-CD11c) and NLDC 145 Abs (Serotec USA,Washington, DC) [18, 19]. The NLDC 145 Ab recog-nizes DEC-205 found on DCs, an endocytic receptorthat is involved in antigen uptake [20]. To block non-speci®c staining, 5% nonfat milk was added in bothprimary and secondary Ab solutions [21].
3. Results
3.1. MHC class I and class II binding capacity of CTLinducing lipopeptides
To assess the processing requirements of a lipidatedTh±CTL epitope constructs, we compared the capacityof the OVA±FLU lipopeptide to bind puri®ed class I andII MHC molecules in vitro with that of the unlinkedOVA and FLU peptides. As shown in Table 1, the OVAand FLU peptides were each capable of binding to its re-spective restriction element when tested in an unlinkedcon®guration. The OVA peptide bound in the 100±500nM a�nity range to puri®ed I-Ad and I-Ab moleculesand the FLU CTL epitope bound with even higher a�-nity to Kd (IC50% value of 28 nM). The constructformed by the covalent linkage of both peptides stillshowed good class II MHC binding capacity (IC50% of136 and 233 nM against I-Ad and I-Ab, respectively).However, in keeping with the stringent length require-ments for peptide binding to class I molecules, the linkedOVA±FLU peptide failed to bind Kd. Finally, the ad-dition of two PAM residues to the OVA±FLU linkedconstruct virtually abolished all the class II MHC bind-ing capacity. These ®ndings suggest that intracellular orextracellular processing of lipidated constructs is likely tobe required in order to generate peptide fragmentscapable of binding to class I and II molecules, and ofbeing recognized by FLU and OVA-speci®c T-cells, re-spectively.
I. Tsunoda et al. / Vaccine 17 (1999) 675±685 677

3.2. Particulate material is the biologically active formof lipopeptide
During preliminary studies of the physico-chemical
characteristics of the (PAM)2±OVA±FLU lipopeptide,
it was observed that although the lyophilized lipopep-
tide was soluble in DMSO containing 0.1% TFA, di-
lution of the DMSO-solubilized material in PBS,
nonbu�ered saline, water, or other aqueous solvents
promptly resulted in the formation of a ®ne suspen-
sion. Since the particulate nature of this (and other
lipopeptides) complicated its puri®cation and analysis,
we substituted the diserine spacer positioned between
the OVA epitope and the PAM residues with hydro-
philic penta-K or penta-D spacers in order to increase
the solubility of the resulting lipopeptide construct.
The solubility of the modi®ed lipopeptides in 10%
DMSO, 0.01% TFA in PBS, a standard diluent for
immunizing animals was assessed as described in
Section 2. The results indicated that addition of ®ve
lysine or aspartic acids greatly increased the solubility
of the constructs in the range of 75±95% as opposed
to 11% solubility observed with the unmodi®ed(PAM)2±OVA±FLU construct (Table 2).
When the CTL inducing activity of these relativelysoluble constructs was analyzed, it was found to bereduced in comparison to that of the less soluble unmo-di®ed construct, particularly at the highest primingdose tested where the (PAM)2±OVA±FLU constructinduced 90.1 DLU of CTL activity compared to 18.3and 24.6 DLU for the poly-D and poly-K constructs,respectively (Table 2). Furthermore, most strikingly,the capacity of the modi®ed constructs to induce CTLswas further reduced after removal of residual aggre-gates by ®ltration through a 0.2 mm membrane. TheCTL-inducing activity of ®ltered poly-K constructdropped from 24.6 DLU to 13.5 DLU at a priming doseof 27 nmol (Table 2). Likewise, ®ltration of the poly-Dconstruct dropped the activity from 18.3 to 1 DLU atthe highest priming dose and from 12.8 to 0.7 DLU at a20-fold lower dose. A ®ltrate of the diserine constructcould not be obtained due to its highly particulatenature. These results suggest that particulate material isthe biological active form of the (PAM)2±OVA±FLUconstruct. Similar results have been obtained with a
Table 1
Lipidated Th±CTL peptide binds ine�ciently to class I and II MHC molecules
MHC binding a�nity (IC50%, nM)a
Peptide Kd Dd I-Ad I-Ed I-Ab
OVA ±b NDc 150 ± 352
FLU 28 ± ± ± ±
OVA±FLU ± ND 136 ± 233
(PAM)2±OVA±FLU ± ND 4688 ± ±
a50% inhibitory concentration (IC50%) based on the mean of a minimum of three independent experi-
ments.b±, indicates IC50%>10,000 nM.cND, not done.
Table 2
Solubility and immunogenicity of lipopeptide constructs containing di�erent spacer residues
Immunizing dose (nmol) (geometric mean DLU (X/6S.D.)d)
Spacer residuesa % Solubilityb Filtrationc 27 1.35 0.07
SS 11 ± 90.1 (1.9) 14.4 (2.4) 3.3 (4.0)
DDDDD 78 ± 18.3 (1.7) 12.8 (1.7) 0.4 (2.9)
+ 1.0 (3.9) 0.7 (6.7) 0.6 (3.2)
KKKKK 94 ± 24.6 (2.9) 4.3 (3.6) 0.7 (7.9)
+ 13.5 (1.7) 1.1 (5.2) 0.2 (6.0)
aAmino acid spacer residues linking the bipalmitoylated lysine to the OVA Th epitope in the (PAM)2±OVA±FLU lipopeptide
construct.bLipopeptide solubilized in DMSO containing 0.1% TFA (10 mg/ml) was heated at 458C. PBS at room temperature was then added to
bring the concentration to 1 mg/ml followed by immediate vortexing. The solubility and immunogenicity of this material was then analyzed as
described in Section 2.cAn aliquot of lipopeptide material prepared in PBS was passed through a 0.2 mm ®lter and the ®ltrate was injected into
animals.dThe values shown indicate the geometric mean of net (D) LU per 106 e�ector cells (background subtracted), calculated as described in
Section 2. In parenthesis, the X/6SD value indicates the error factor by which the geometric mean value should be divided and multiplied to
obtain the con®dence interval corresponding to one standard deviation. The mean CTL response was calculated from six individual animals per
group, in two independent experiments.
I. Tsunoda et al. / Vaccine 17 (1999) 675±685678

lipopeptide construct consisting of a tetanus toxin Thpeptide and a HBV CTL peptide [10] when tested inHLA±A2.1/Kb transgenic mice, suggesting that thisphenomenon might be of general biological andmechanistic signi®cance (data not shown).
3.3. Immunogenicity of constructs containing di�erenttypes of lipid
The solubility of lipopeptides can also be changedby varying the length of the carbon chain lipid tail, orattaching a single instead of double lipid tail to thepeptide. Accordingly, constructs containing mono- orbilipid tails with varying number of carbon atomswere synthesized and studied. Fig. 1 shows the strikinginverse correlation that was observed between solubi-lity of lipopeptide constructs having lipids of varyingcarbon chain lengths and their ability to induce CTLsin animals (Pearson correlation coe�cient,R= ÿ0.92). Constructs with lipid moieties composedof two fatty acid chains with 10 to 16 carbon atomswere shown to be the least soluble, with solubility ran-ging between 11 and 25%, and these constructs werethe most immunogenic, inducing over 16 DLU of CTLactivity at a 1 nmol priming dose. By contrast, con-structs carrying either a single PAM chain, or a doubleoctyl acid chain were more soluble (30 and 42% solu-bility values, respectively) and virtually inactive interms of CTL induction, achieving less than 2 DLU ofCTL activity. These results con®rm, by an independentapproach, that the insolubility of lipopeptides is a
major contributor of its ability to induce CTLs, asnoted in the preceding section.
3.4. Correlation between particle size andimmunogenicity
As previously mentioned, lipopeptide constructs aregenerally soluble in DMSO containing 0.1% TFA butmixing with aqueous solutions results in precipitation ofthe lipopeptide and formation of a particulate suspen-sion. Furthermore, it was observed that depending onhow the lipopeptide was formulated prior to dilution,particles of di�erent appearances were formed. Morespeci®cally, if room temperature PBS was mixed vigor-ously with DMSO-solubilized (PAM)2±OVA±FLU, thatwas heated to 458C, an opalescent suspension character-ized by ®ne particles resulted (hereafter referred to as the458C DMSO formulation). However, by directly resus-pending lyophilized lipopeptide powder in a solution of10% DMSO in PBS, a suspension consisting of larger,rapidly settling particles was generated (referred to asthe powder formulation). When the size distribution ofparticles contained in these two visually di�erent suspen-sions was quantitated by laser di�raction technology,signi®cant di�erences were observed. The powder for-mulation contained a broad distribution of larger par-ticles with diameters ranging from 10 to 200 mm and witha mean of 67 mm whereas the 458C DMSO formulationcontained a bimodal size distribution of smaller particleswith a mean diameter of 0.3 mm (range, 0.15±0.6 mm)and 7.7 mm (range, 2±20 mm) (Fig. 2).
When the in vivo immunogenicity of the very samesamples subjected to particle size analysis was investi-gated, strong CTL responses were elicited by the sus-pension generated from 458C DMSO formulation,while very little activity was observed with the powderformulation (Table 3). Similar results have beenobserved with all other lipidated constructs tested inour laboratory, suggesting that a general correlationexists between particle size and immunogenicity of lipi-dated Th±CTL constructs (data not shown).
3.5. Immunohistological analysis of lipopeptideimmunized animals
To further clarify the mechanism involved in the vig-orous CTL induction by lipopeptide, we performed animmunohistological analysis of tissues from immunizedanimals to examine the phenotype of APCs that as-sociate with lipopeptide following immunization, andto determine the tra�cking of lipopeptide in vivo.With respect to peptide immunizations, especially lipo-peptide, very little is known about the tra�cking ofAg and the involvement of APCs. Since macrophagesand DCs have been implicated in presenting particu-late Ags to CTLs [22±25], we addressed whether both
Fig. 1. Correlation between solubility and potency of OVA±FLU
constructs with lipid tails of varying carbon chain lengths. Lipidated
OVA±FLU constructs containing bilipid tails of 8 (w), 10 (.), 12
(Q), 14 (r), and 16 (R) carbon residues or a monolipid tail of 16
carbon residues (q) were synthesized and tested for solubility and in
vivo immunogenicity at a 1 nmol priming dose as described in
Section 2.
I. Tsunoda et al. / Vaccine 17 (1999) 675±685 679

professional APC populations were involved in thelocal and peripheral immune response.
In order to facilitate immunohistological analysis,mice were immunized with a biotinylated analog of the(PAM)2±OVA±FLU lipopeptide, thereby allowing fordetection of lipopeptide in tissue sections by stainingwith peroxidase- or ¯uorochrome-conjugated avidin.The biotin-conjugated lipopeptide analog, when for-mulated either from 458C DMSO-solubilized materialor lyophilized powder, behaved similarly to the non-biotinlyated lipopeptide in terms of particle size distri-bution and in vivo CTL induction (data not shown).
Following s.c. immunization at the tail base with thebiologically active 458C DMSO formulation, theimmunohistological analysis of the skin site of immu-nization revealed an early in®ltration of polymorpho-nuclear leukocytes at 3 h post injection (PI) (Fig. 3a).Mononuclear cell in®ltration, consisting largely of
macrophages, was seen initially at 24 h PI and waspredominant in the in¯amed hypodermis after 3±4days PI. Cell free lipopeptide was visualized mainly inthe hypodermis with both hematoxylin and eosin stainand the ABC method 1 h PI. Interestingly, cell-associ-ated lipopeptide was also observed and the cell popu-lations staining peroxidase-positive coincided with thein®ltrating populations, that is, being mainly polymor-phonuclear cells during the ®rst 24 h PI (Fig. 3b) andmacrophages thereafter. Free lipopeptide in the hypo-dermis became encapsulated 3 days PI and was seque-strated by collagen ®ber 1 week PI. Eventually thesequestrated lipopeptide was partially engulfed bymacrophages beginning 1 week PI and was completelyreplaced by lipopeptide-engulfed macrophages at 2week PI (Fig. 3c and d).
Surprisingly, when tra�cking of lipopeptide to per-ipheral organs was analyzed, the spleen from animalsinjected 1 h previously showed intense staining with bio-tin-conjugated lipopeptide, mainly in the red pulp and tosome extent in the white pulp (Fig. 4a). The intensity ofstaining diminished with time, such that only occasionallipopeptide-positive cells could be seen in the red andwhite pulp after 24 h PI (Fig. 4b). In the regional lymphnode of some mice, a few lipopeptide-bearing cells weredetected as early as 1 h PI (data not shown).
In repeated experiments using the same and ad-ditional injection sites, such as the footpad and skin inthe hindcalf, virtually identical histopathologicalchanges were observed and, overall, the intensity oflipopeptide staining was the greatest with the tail baseimmunization (data not shown). We could not detectthe lipopeptide in the thymus or the liver. As negativecontrols, mice were immunized with the unconjugatedlipopeptide, vehicle only (PBS/10% DMSO/0.01%TFA), or PBS. While an in¯ammatory response could
Fig. 2. Suspensions of (PAM)2±OVA±FLU generated by di�erent methods vary in particle size. One mg/ml suspensions of (PAM)2±OVA±FLU
were generated by solubilizing lyophilized powder in DMSO containing 0.1% TFA, heating this material to 458C, then mixing in PBS (solid line)
or by mixing lyophilized powder directly in 10% DMSO, 0.01% TFA in PBS (dotted line). Suspensions were then analyzed for particle size with
a laser di�raction particle size analyzer. Data is represented as the percentage of the total particles having a given diameter. The numbers shown
above each particle distribution curve represents its mean diameter.
Table 3
Di�erent formulations of (PAM)2±OVA±FLU lipopeptide have
di�erent in vivo potency
Experiment Lipopeptide
formulation
Geometric mean
DLU (X/6SD)a
1 powderb 0.3 (8.7)
458C DMSOc 22.9 (1.7)
2 powder 0.7 (9.2)
458C DMSO 20.6 (1.4)
aSee footnote d, Table 2. Mean CTL responses were calculated from
three individual animals per group.bLyophilized (PAM)2±OVA±FLU
powder was mixed by vigorous vortexing with 10% DMSO in PBS
to achieve a 20 nmol/mouse priming dose.c(PAM)2±OVA±FLU (10
mg/ml in DMSO with 0.1% TFA) was heated for 10 min at 458C,then mixed by vigorous vortexing with PBS to achieve a 20 nmol/
mouse priming dose.
I. Tsunoda et al. / Vaccine 17 (1999) 675±685680
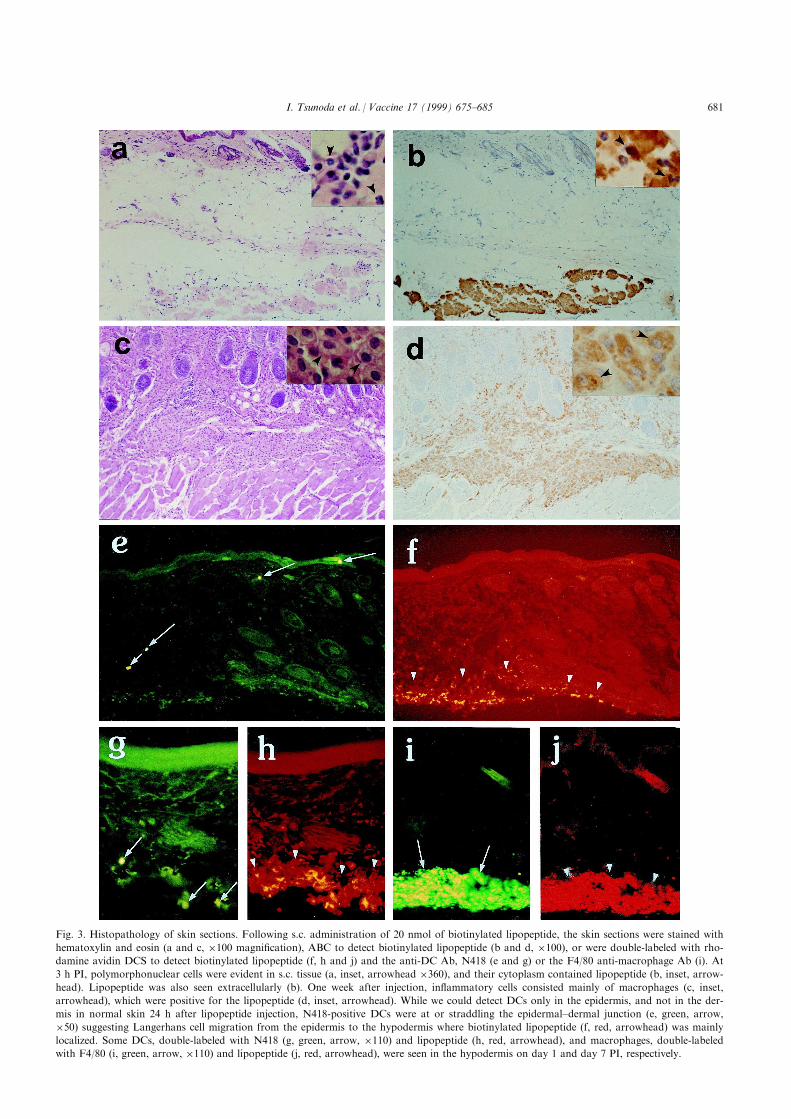
Fig. 3. Histopathology of skin sections. Following s.c. administration of 20 nmol of biotinylated lipopeptide, the skin sections were stained with
hematoxylin and eosin (a and c, �100 magni®cation), ABC to detect biotinylated lipopeptide (b and d, �100), or were double-labeled with rho-
damine avidin DCS to detect biotinylated lipopeptide (f, h and j) and the anti-DC Ab, N418 (e and g) or the F4/80 anti-macrophage Ab (i). At
3 h PI, polymorphonuclear cells were evident in s.c. tissue (a, inset, arrowhead �360), and their cytoplasm contained lipopeptide (b, inset, arrow-
head). Lipopeptide was also seen extracellularly (b). One week after injection, in¯ammatory cells consisted mainly of macrophages (c, inset,
arrowhead), which were positive for the lipopeptide (d, inset, arrowhead). While we could detect DCs only in the epidermis, and not in the der-
mis in normal skin 24 h after lipopeptide injection, N418-positive DCs were at or straddling the epidermal±dermal junction (e, green, arrow,
�50) suggesting Langerhans cell migration from the epidermis to the hypodermis where biotinylated lipopeptide (f, red, arrowhead) was mainly
localized. Some DCs, double-labeled with N418 (g, green, arrow, �110) and lipopeptide (h, red, arrowhead), and macrophages, double-labeled
with F4/80 (i, green, arrow, �110) and lipopeptide (j, red, arrowhead), were seen in the hypodermis on day 1 and day 7 PI, respectively.
I. Tsunoda et al. / Vaccine 17 (1999) 675±685 681

Fig. 4. Representative histopathology in the spleen. Spleen sections were stained with ABC (a and b), the N418 (c), or the F4/80 Ab (d), or were
double-stained with N418 (e) and rhodamine avidin DCS (f), or F4/80 (g) and ¯uorescein streptoavidin (h). Each panel (a±d) are displayed so
that the marginal zone and the beginning of the red pulp are on the right, while the white pulp is on the left. (a) At 1 h PI, the lipopeptide was
most easily seen in the red pulp (arrowhead), but was also evident in the white pulp (arrow), most likely in the T-cell zone surrounding a central
artery (*) (a, �180 magni®cation). (b) Biotinylated lipopeptide could also be detected 24 h PI, although its intensity had decreased. The N418
Ab stained DCs not only in the red pulp (arrowhead), but also in the T-cell zone (c, arrow, �180) with an evident central artery (*), while the
F4/80 Ab labeled only red pulp macrophages d, (arrowhead). Some DCs double-labeled with N418 (e, green, arrow, �220) and lipopeptide
(f, red, arrow), and macrophages were also seen on day 1 PI labeled with both F4/80 (g, red, �110) and lipopeptide (h, green, arrow).
I. Tsunoda et al. / Vaccine 17 (1999) 675±685682

be seen in skin, injected with both the unconjugatedlipopeptide and vehicle, biotinylated lipopeptide-posi-tive cells were absent in the skin, lymph node, andspleen by the ABC method. PBS injection did notcause in¯ammation (data not shown).
To de®ne the lipopeptide-bearing cell type, wedouble-stained sections with monoclonal Abs speci®cfor DC and macrophage cell markers and ¯uoro-chrome-avidin for biotinylated lipopeptide. To con®rmthe speci®city of the Abs, they were tested on tissuesections from unimmunized animals. In the spleen,N418- and NLDC 145-positive DCs, having thecharacteristic DC morphology [26], were detected inboth areas of the spleen (Fig. 4c) whereas macrophagesstained with F4/80 were detected in the red pulp, butnot in the white pulp (Fig. 4d). In the epidermis,N418- and NLDC 145-positive Langerhans cells weredetected easily in the footpad but rarely in the tailbase, probably because the epidermis at the tail basewas too thin, and the numbers of Langerhans cellswere few and di�cult to ®nd in cross-sections.
Examination of double-stained tissue sections fromlipopeptide-immunized animals revealed that both DCsand macrophages appeared to be associating with Ag atthe site of injection. Whereas the number of N418- andNLDC 145-positive DCs are normally low in the epider-mis and undetectable in the dermis and hypodermis, cellsstaining with both Abs were present in increasing num-bers 24 h PI in all three layers of the skin (Fig. 3e)suggesting Langerhans cell migration from the epidermisto the hypodermis, where the lipopeptide was mainlylocalized (Fig. 3f). N418- and NDLC 145-positive DCswere also found directly in the hypodermis at the site ofinjection where lipopeptide was deposited (Fig. 3g andh). F4/80-positive macrophages were also detected in theinjected lipopeptide depot as early as 24 h PI. On day 4and thereafter, large numbers of double-positive macro-phages that had engulfed lipopeptide were detected inthe hypodermis (Fig. 3i and j). In peripheral lymphoidorgans, lipopeptide-positive DCs (Fig. 4e and f) andmacrophages (Fig. 4g and h) were observed in the spleen24 h PI and in the lymph node (data not shown).
4. Discussion
Despite numerous studies demonstrating the aug-mented immunogenicity of lipopeptide molecules invivo, very little is known regarding its mechanisms ofaction. In our current study, we address the contri-bution of the following factors towards the potentimmunogenicity of lipopeptides for CTL induction: (1)the MHC binding capacity of lipopeptide molecules,(2) the correlation between lipopeptide (in)solubilityand immunogenicity in vivo, and (3) the tra�cking of
Ag in vivo and identity of APCs that associate withlipopeptide following immunization.
The major ®ndings of our study are three-fold.First, marginal binding activity of a model lipopeptide,(PAM)2±OVA±FLU, was detected with puri®ed class Iand II MHC molecules of the appropriate restrictingelement, indicating that processing of this lipopeptideis a requirement for induction of epitope-speci®cCD4+ and CD8+ cells. Secondly, an inverse corre-lation between the solubility of (PAM)2±OVA±FLUand in vivo immunogenicity was demonstrated. Theparticulate nature of the lipopeptide is crucial forimmunogenicity, since particulate (PAM)2±OVA±FLUsuspensions were more immunogenic than partially sol-uble analogs of this lipopeptide and removal of Agaggregates by ®ltration eliminated immunogenicity.Finally, based on immunohistological studies, cell-as-sociated lipopeptide could be detected on macrophagesand DCs found both at the site of injection and in theperipheral lymphoid organs. More remarkably, whenin vivo lipopeptide tra�cking was examined, thereappeared to be rapid movement of Ag from the injec-tion site at the tail base to the spleen within 1 h fol-lowing immunization. Collectively, these ®ndings pointto insoluble lipopeptide particles and macrophages/DCs as the biologically active form and the relevantAPCs, respectively, that are involved in CTL inductionfollowing lipopeptide immunization.
Macrophages and DCs have been shown to beinvolved in CTL induction following uptake of Agparticles such as protein-coated latex beads andbacteria [22±25, 27±29], thus both APC populationsmay contribute signi®cantly to the strong in vivoimmunogenicity observed with particulate lipopeptidesuspensions. Another contributing factor of lipopep-tide immunogenicity may be its tra�cking propertiesin vivo. In this regard, the rapid egress of lipopeptidefrom the site of injection is reminiscent of a similar®nding recently described by Torres et al. [30]. Theseinvestigators found that the magnitude and longevityof Ab responses following i.m. immunization withcDNA was not a�ected by surgical ablation of themuscle receiving cDNA when it was performed asearly as 1±10 min following immunization. The rapiddissemination of cDNA or cDNA-transfected cells alsosuggested that initiation of the Ab response occurredat a distal site. Since DCs and macrophages are impli-cated in cDNA-induced immune responses in vivo,either through transfection or cross-priming [31±35],the ®ndings of Torres et al. with cDNA and the cur-rent study with lipopeptide are consistent with a cru-cial common pathway shared by many of themechanisms leading to immunogenicity in general, andto CTL induction in vivo, in particular. This pathwayinvolves the rapid capture of Ag by macrophages andDCs shortly following immunization and the dissemi-
I. Tsunoda et al. / Vaccine 17 (1999) 675±685 683

nation of Ag or Ag-sensitized APCs into peripheralorgans where lymphocyte induction occurs.
Immunohistological analysis of tissue sections of ani-mals immunized with a biotinylated analog of (PAM)2±OVA±FLU, revealed that macrophages and DCs wereassociating with the lipopeptide depot at the site of injec-tion as early as 24 h PI. In addition, at the same timepoint, lipopeptide-positive macrophages and DCs werealso detected in peripheral lymphoid organs, mainly inthe spleen and to a lesser extent in the draining lymphnodes. Prior to the appearance of lipopeptide-coatedAPCs in the spleen, we observed lipopeptide in the redand white pulp of the spleen within 1 h following s.c. tailbase immunization. Whereas the involvement of splenicDCs as APCs after i.v. administration of soluble proteinhas been reported previously [36], as well as the mi-gration of haptenated Langerhans cells into draininglymph nodes after topical administration of contactallergens [37], the rapid accumulation of Ag and Ag-bearing APCs in the spleen shortly after s.c. immuniz-ation has, to our knowledge, not been reported pre-viously. In addition to rapid tra�cking of lipopeptide tothe spleen, the depot of lipopeptide detected at the site ofinjection up to 2 week PI may further contribute toimmunogenicity, since it serves as a source of Ag that iscontinually being engaged by in®ltrating DCs andmacrophages.
From a more practical viewpoint, our current studyalso addresses some of the challenges encountered withdeveloping CTL-inducing lipopeptide vaccines.Although lipopeptide suspensions are highly immuno-genic in vivo, their poor solubility may complicatepuri®cation and analysis. To circumvent these con-cerns, two structural modi®cations, aimed at increasingthe solubility of the (PAM)2±OVA±FLU lipopeptide,were investigated. Increased lipopeptide solubility wasachieved either by the insertion of hydrophilic polyly-sine or polyaspartic acid spacer residues between thelipid moiety and the OVA±FLU peptide itself, or bythe attachment of smaller lipids instead of the doublePAM chain to the OVA±FLU peptide. Although thesemodi®cations did enhance solubility, in all instancesthe more soluble lipopeptide analogs were found to besigni®cantly less immunogenic than the particulatenative compound. With respect to the lipid moiety,although the monopalmitoylated construct and the(Octyl)2±OVA±FLU construct showed increased solu-bility compared to (PAM)2±OVA±FLU, they still con-tained a considerable amount of insoluble material, onthe order of 60±70%, which apparently did not con-tribute to its immunogenicity. Thus, the lipid moietymight serve multiple functions, on one hand promotinginsolubility, and on the other hand, playing some asyet unspeci®ed role(s), such as intercalating with bio-logical membranes [6, 38] or in¯uencing the type and/or size of insoluble particle formed.
In the current study we also found that the formu-lation process profoundly in¯uences the biophysicalproperties of particulate lipopeptide suspensions andits level of in vivo potency. The most immunogenicsuspensions, generated from 458C DMSO-solubilizedlipopeptide, contained particles of two predominantsizes, one with a mean diameter in the range of 0.3 mmand the other with a diameter of 7.7 mm. In contrast,inactive lipopeptide suspension generated by mixinglyophilized powder with PBS contained larger particleswith a mean distribution of 67 mm. The decreasedimmunogenicity of the latter formulation may be dueto the inability of macrophages and DCs to internalizelarge particles, as evidenced by studies showing thatDCs can internalize particles within vesicles formed bycell surface ru�es that are up to 5 mm indiameter [39, 40]. In addition, Kovacsovics-Bankowskiet al. [23] and Harding and Song [24] observed thatparticle size is a key parameter for Ag uptake and pro-cessing by macrophages through the alternate class Ipathway, with e�cient processing occurring with par-ticles ranging in diameter from 59 nm to 3 mm.
Based on the ®ndings of our current study, the ma-nipulation of lipopeptide structure and formulation tooptimize particle size and spleen tra�cking, may pro-vide the basis for improving the potency of epitope-based lipopeptide vaccines that raise protective andtherapeutic CTL responses. Such studies should alsocontribute signi®cantly to our understanding of theprocessing pathway of lipopeptides used by macro-phages and DCs.
Acknowledgements
The authors would like to thank Neal D. Tolley,Dr. Ingeborg J. McCright, Kornelia Edes and Dr. CarlPerez for many helpful discussions and to AjeshMaewal and Anthony Chiem for technical assistancein peptide synthesis. We also thank Dr. Michael B.A.Oldstone for his generous gift of the N418 Ab. We aregrateful to Ms. Kathleen Borick for preparation of themanuscript. This work was supported in part byGrants NS34497, AI42525, and AI35198 (to R.S.F.)from the National Institutes of Health and also withFederal funds from the NIH under Contract N01-AI-45241 (to A.S.).
References
[1] Byrne JA, Oldstone MBA. Biology of cloned cytotoxic T-lym-
phocytes speci®c for lymphocytic choriomeningitis virus: clear-
ance of virus in vivo. J. Virol. 1984;51:682±6.
[2] Yap KL, Ada GL, McKenzie IFC. Transfer of speci®c cyto-
toxic T-lymphocytes protects mice inoculated with in¯uenza
virus. Nature 1978;273:238±9.
I. Tsunoda et al. / Vaccine 17 (1999) 675±685684

[3] McMichael AJ, Gotch FM, Noble GR, Beare PAS. Cytotoxic
T-cell immunity to in¯uenza. N. Engl. J. Med. 1983;309:13±17.
[4] Quinnan GV, Kirmeni N, Rook AH, Manishevitz JV, Jackson
L, Moreschi G, Santos GW, Saral R, Burns WH. Cytotoxic T-
cells in cytomegalovirus infection: HLA-restricted T-lymphocyte
and non-T-lymphocyte cytotoxic response correlate with recov-
ery from cytomegalovirus infection in bone-marrow-transplant
recipients. N. Engl. J. Med. 1982;307:6±13.
[5] Deres K, Schild H, WeismuÈ ller K-H, Jung G, Rammensee H-G.
In vivo priming of virus-speci®c cytotoxic T-lymphocytes with
synthetic lipopeptide vaccine. Nature 1989;342:561±4.
[6] WeismuÈ ller K-H, Bessler WG, Jung G. Solid phase peptide syn-
thesis of lipopeptide vaccines eliciting epitope-speci®c B-, T-
helper, and T-killer cell response. Int. J. Peptide Protein Res.
1992;40:255±60.
[7] Allsopp CE, Plebanski M, Gilbert S, Harris RE, Frankel G,
Dougan G, Hioe C, Nixon D, Paoletti E, Layton G, Hill AV.
Comparison of numerous delivery systems for the induction of
cytotoxic T-lymphocytes by immunization. Eur. J. Immunol.
1996;26:1951±9.
[8] Hioe CE, Qiu H, Chend PD, Bian Z, Li ML, Li J, Singh M,
Kuebler P, McGee P, O'Hagan D, Zamb T, Ko� W, Allsopp
C, Yang CY, Nixon DF. Comparison of adjuvant formulations
for cytotoxic T-cell induction using synthetic peptides. Vaccine
1996;14:412±8.
[9] BenMohamed L, Gras-Masse H, Tartar A, Daubersies P,
Brahimi K, Bossus M, Thomas A, Druilhe P. Lipopeptide im-
munization without adjuvant induces potent and long-lasting B,
T helper and cytotoxic T-lymphocyte responses against a ma-
laria liver stage antigen in mice and chimpanzees. Eur. J.
Immunol. 1997;27:1242±53.
[10] Vitiello A, Ishioka G, Grey HM, Rose R, Farness P, LaFond
R, Yuan L, Chisari FV, Furze J, Bartholomeuz R, Chesnut
RW. Development of a lipopeptide-based therapeutic vaccine to
treat chronic HBV infection. I. Induction of a primary cytotoxic
T-lymphocyte response in man. J. Clin. Invest. 1995;95:341±9.
[11] Livingston BD, Crimi C, Grey H, Ishioka G, Chisari FV, Grey
H, Chesnut RW, Sette A. The hepatitis B virus-speci®c CTL re-
sponses induced in humans by lipopeptide vaccination are com-
parable to those elicited by acute viral infection. J. Immunol.
1997;159:1383±92.
[12] Sette A, Southwood S, O'Sullivan D, Gaeta FCA, Sidney J,
Grey HM. E�ect of pH on MHC class II-peptide interactions.
J. Immunol. 1992;148:844±51.
[13] Vitiello A, Yuan L, Chesnut RW, Sidney J, Southwood S,
Farness P, Jackson MR, Peterson PA, Sette A.
Immunodominance analysis of CTL responses to the in¯uenza
PR8 virus reveals two new dominant and subdominant Kb-
restricted epitopes. J. Immunol. 1996;157:5555±62.
[14] Sette A, Buus S, Colon S, Miles C, Grey HM. Structural analy-
sis of peptides capable of binding to more than one Ia antigen.
J. Immunol. 1989;142:35±40.
[15] Tsunoda I, Kurtz CI, Fujinami RS. Apoptosis in acute and
chronic central nervous system disease induced by Theiler's
murine encephalomyelitis virus. Virology 1997;228:388±93.
[16] Borrow P, Evans CF, Oldstone MBA. Virus-induced immuno-
suppression: immune system-mediated destruction of virus-
infected dendritic cells results in generalized immune suppres-
sion. J. Virol. 1995;69:1059±70.
[17] Austyn JM, Gordon S. F4/80, a monoclonal antibody directed
against the mouse macrophage. Eur. J. Immunol. 1981;11:805±
15.
[18] Metlay JP, Witmer-Pack MD, Agger R, Crowley MT, Lawless
D, Steinman RM. The distinct leukocyte integrins of mouse
spleen dendritic cells as identi®ed with new hamster monoclonal
antibodies. J. Exp. Med. 1990;171:1753±71.
[19] Kraal G, Breel M, Janse M, Bruin G. Langerhans' cells, veiled
cells, and interdigitating cells in the mouse recognized by a
monoclonal antibody. J. Exp. Med. 1986;163:981±7.
[20] Jiang W, Swiggard WJ, Heu¯er C, Peng M, Mirza A, Steinman
RM, Nussenzweig MC. The receptor DEC-205 expressed by
dendritic cells and thymic epithelial cells is involved in antigen
processing. Nature 1995;375:151±5.
[21] Duhamel RC, Johnson DA. Use of nonfat dry milk to block
nonspeci®c nuclear and membrane staining by avidin conju-
gates. J. Histochem. Cytochem. 1985;33:711±4.
[22] Rock KL, Gamble S, Rothstein L. Presentation of exogenous
antigen with class I major histocompatibility complex molecules.
Science 1990;249:918±21.
[23] Kovacsovics-Bankowski M, Clark K, Benacerraf B, Rock KL.
E�cient major histocompatibility complex class I presentation
of exogenous antigen upon phagocytosis by macrophages. Proc.
Natl. Acad. Sci. USA 1993;90:4942±6.
[24] Harding CV, Song R. Phagocytic processing of exogenous par-
ticulate antigens by macrophages for presentation by class I
MHC molecules. J. Immunol. 1994;153:4925±33.
[25] Svensson M, Stockinger B, Wick MJ. Bone marrow-derived
dendritic cells can process bacteria for MHC-I and MHC-II
presentation to T-cells. J. Immunol. 1997;158:4229±36.
[26] Steinman RM, Cohn ZA. Identi®cation of a novel cell type in
peripheral lymphoid organs of mice. I. Morphology, quanti-
tation, tissue distribution. J. Exp. Med. 1973;137:1142±62.
[27] Steinman RM. The dendritic cell system and its role in immuno-
genicity. Ann. Rev. Immunol. 1991;9:271±96.
[28] Inaba K, Inaba M, Naito M, Steinman RM. Dendritic cell pro-
genitors phagocytose particulates, including Bacillus Calmette±
Guerin organisms, and sensitize mice to mycobacterial antigens
in vivo. J. Exp. Med. 1993;178:479±88.
[29] Lanzavecchia A. Mechanisms of antigen uptake for presen-
tation. Curr. Opin. Immunol. 1996;8:348±53.
[30] Torres CAT, Iwasaki A, Barber BH, Robinson HL. Di�erential
dependence on target site tissue for gene gun and intramuscular
DNA immunizations. J. Immunol. 1997;158:4529±32.
[31] Doe B, Selby M, Barnett S, Baenziger J, Walker CM. Induction
of cytotoxic T-lymphocytes by intramuscular immunization with
plasmid DNA is facilitated by bone marrow-derived cells. Proc.
Natl. Acad. Sci. USA 1996;93:8578±83.
[32] Condon C, Watkins SC, Celluzzi CM, Thompson K, Falo L.
DNA-based immunization by in vivo transfection of dendritic
cells. Nat. Med. 1996;2:1122±8.
[33] Corr M, Lee DJ, Carson DA, Tighe H. Gene vaccination with
naked plasmid DNA: mechanism of CTL priming. J. Exp. Med.
1996;184:1555±60.
[34] Ulmer JB, Deck RR, Dewitt CM, Donnhly JI, Liu MA.
Generation of MHC class I-restricted cytotoxic T-lymphocytes
by expression of a viral protein in muscle cells: antigen presen-
tation by nonmuscle cells. Immunology 1996;89:59±67.
[35] Song ES, Lee V, Surh CD, Lynn A, Brumm D, Jolly DJ,
Warner JF, Chada S. Antigen presentation in retroviral vector-
mediated gene transfer in vivo. Proc. Natl. Acad. Sci. USA
1997;94:1943±8.
[36] Crowley M, Inaba K, Steinman RM. Dendritic cells are the
principal cells in mouse spleen bearing immunogenic fragments
of foreign proteins. J. Exp. Med. 1990;172:383±6.
[37] Macatonia SE, Knight SC, Edwards AJ, Gri�ths S, Fryer P.
Localization of antigen on lymph node dendritic cells after ex-
posure to the contact sensitizer ¯uorescein isothiocyanate.
Functional and morphological studies. J. Exp. Med.
1987;166:1654±67.
[38] Wolf B, Hauschildt S, Uhl B, Metzger J, Jung G, Bessler WG.
Localization of the cell activator lipopeptide in bone marrow-
derived macrophages by electron loss spectroscopy (EELS).
Immunol. Lett. 1989;20:121±6.
I. Tsunoda et al. / Vaccine 17 (1999) 675±685 685
Related Documents





![Inducing Patent Infringement - Law Review...2005] Inducing Patent Infringement 229 understanding, inducing infringement is a natural outgrowth of the common law principle of respondeat](https://static.cupdf.com/doc/110x72/5f9608d795a783197246401f/inducing-patent-infringement-law-review-2005-inducing-patent-infringement.jpg)





