Please cite this article in press as: Partyka, A., et al., Lipid peroxidation and antioxidant enzymes activity in avian semen. Anim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anireprosci.2012.07.007 ARTICLE IN PRESS G Model ANIREP-4584; No. of Pages 7 Animal Reproduction Science xxx (2012) xxx–xxx Contents lists available at SciVerse ScienceDirect Animal Reproduction Science jou rn al h om epa ge: www .elsevier.com/locate/anireprosci Lipid peroxidation and antioxidant enzymes activity in avian semen Agnieszka Partyka a,∗ , Ewa Łukaszewicz b , Wojciech Ni ˙ za ´ nski a a Wrocław University of Environmental and Life Sciences, Faculty of Veterinary Medicine, Department of Reproduction and Clinic of Farm Animals, pl. Grunwaldzki 49, 50-366 Wrocław, Poland b Wrocław University of Environmental and Life Sciences, Faculty of Biology and Animal Breeding, Department of Poultry Breeding, ul. Chełmo´ nskiego 38c, 51-630 Wrocław, Poland a r t i c l e i n f o Article history: Received 29 June 2011 Received in revised form 16 July 2012 Accepted 20 July 2012 Available online xxx Keywords: Avian semen Lipid peroxidation Antioxidant enzymes Superoxide dismutase Glutathione peroxidase Catalase a b s t r a c t The present study compared the antioxidant system and lipid peroxidation in semen of two avian species: chicken and goose. The experiment was conducted on Greenleg Partridge roosters and White Koluda ® ganders, each represented by 10 mature males. Malondialde- hyde (MDA) concentration, catalase (CAT), glutathione peroxidase (GPx) and superoxide dismutase (SOD) activities were determined in sperm cells and seminal plasma. In gander spermatozoa, the amount of MDA was 10 times greater (P < 0.01) than in rooster spermatozoa. Each of the investigated antioxidant enzymes had greater (P < 0.01) activity in goose than chicken sperm. Catalase activity was detected in seminal plasma and sperma- tozoa from both studied species for the first time. In seminal plasma, the activity of GPx was two times greater (P < 0.01) in the White Koluda ® than in chickens, whereas SOD activity was less (P < 0.01) than in chickens. This is the first study describing the presence of CAT in avian semen and the occurrence of indicator of lipid peroxidation (LPO) in geese. Data from the present study clearly show the species-specific differences in the activity of antioxidant defense and LPO. The greater amount of lipid peroxidation and greater activity of antioxidant enzymes in goose semen might suggest that spermatozoa were under greater oxidative stress and the enzymes were not utilized for the protection of functionally and structurally impaired cells. In turn, in fresh chicken semen a lesser activity of antioxidant enzymes accompanied with a lesser lipid peroxidation amount and good semen quality could indicate that fowl spermatozoa were under oxidative stress, but the enzymes were employed to protect and maintain sperm quality. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Avian spermatozoa membranes are rich in polyunsat- urated fatty acids (PUFAs) and can easily undergo lipid peroxidation (LPO) in the presence of reactive oxygen species (ROS) (Cerolini et al., 2006; Fujihara and Howarth, 1978; Surai et al., 1998). ROS are reactive molecules produced during oxygen reduction that affect the deteri- oration of the function and viability of the spermatozoa, if ∗ Corresponding author. Tel.: +48 71 32 05 300; fax: +48 71 32 01 006. E-mail address: [email protected] (A. Partyka). produced at greater than optimal concentrations (Aitken et al., 1989; De Lamirande and Gagnon, 1993). Peroxidation of PUFAs in the sperm cell membranes is an autocatalytic, self-propagating reaction, which can cause cell dysfunc- tion associated with the loss of the membrane function, integrity and finally leads to decreased fertilizing ability of spermatozoa (Alvarez and Storey, 1982). As a consequence of the destructive potential of ROS, spermatozoa membranes must be protected by a highly effective antioxidant system to prevent the peroxida- tive damage. The antioxidant system in semen includes enzymes such as glutathione peroxidase (GPx), superox- ide dismutase (SOD) and catalase (CAT) and also natural 0378-4320/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.anireprosci.2012.07.007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

GA
L
Aa
Gb
5
a
ARRAA
KALASGC
1
ups1po
0h
ARTICLE IN PRESS ModelNIREP-4584; No. of Pages 7
Animal Reproduction Science xxx (2012) xxx– xxx
Contents lists available at SciVerse ScienceDirect
Animal Reproduction Science
jou rn al h om epa ge: www .e lsev ier .com/ locate /an i reprosc i
ipid peroxidation and antioxidant enzymes activity in avian semen
gnieszka Partykaa,∗, Ewa Łukaszewiczb, Wojciech Nizanskia
Wrocław University of Environmental and Life Sciences, Faculty of Veterinary Medicine, Department of Reproduction and Clinic of Farm Animals, pl.runwaldzki 49, 50-366 Wrocław, PolandWrocław University of Environmental and Life Sciences, Faculty of Biology and Animal Breeding, Department of Poultry Breeding, ul. Chełmonskiego 38c,1-630 Wrocław, Poland
r t i c l e i n f o
rticle history:eceived 29 June 2011eceived in revised form 16 July 2012ccepted 20 July 2012vailable online xxx
eywords:vian semenipid peroxidationntioxidant enzymesuperoxide dismutaselutathione peroxidaseatalase
a b s t r a c t
The present study compared the antioxidant system and lipid peroxidation in semen of twoavian species: chicken and goose. The experiment was conducted on Greenleg Partridgeroosters and White Koluda® ganders, each represented by 10 mature males. Malondialde-hyde (MDA) concentration, catalase (CAT), glutathione peroxidase (GPx) and superoxidedismutase (SOD) activities were determined in sperm cells and seminal plasma.
In gander spermatozoa, the amount of MDA was 10 times greater (P < 0.01) than in roosterspermatozoa. Each of the investigated antioxidant enzymes had greater (P < 0.01) activityin goose than chicken sperm. Catalase activity was detected in seminal plasma and sperma-tozoa from both studied species for the first time. In seminal plasma, the activity of GPx wastwo times greater (P < 0.01) in the White Koluda® than in chickens, whereas SOD activitywas less (P < 0.01) than in chickens.
This is the first study describing the presence of CAT in avian semen and the occurrenceof indicator of lipid peroxidation (LPO) in geese. Data from the present study clearly showthe species-specific differences in the activity of antioxidant defense and LPO. The greateramount of lipid peroxidation and greater activity of antioxidant enzymes in goose semenmight suggest that spermatozoa were under greater oxidative stress and the enzymes were
not utilized for the protection of functionally and structurally impaired cells. In turn, in freshchicken semen a lesser activity of antioxidant enzymes accompanied with a lesser lipidperoxidation amount and good semen quality could indicate that fowl spermatozoa wereunder oxidative stress, but the enzymes were employed to protect and maintain spermquality.. Introduction
Avian spermatozoa membranes are rich in polyunsat-rated fatty acids (PUFAs) and can easily undergo lipideroxidation (LPO) in the presence of reactive oxygenpecies (ROS) (Cerolini et al., 2006; Fujihara and Howarth,
Please cite this article in press as: Partyka, A., et al., Lipid peroxiAnim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anirepros
978; Surai et al., 1998). ROS are reactive moleculesroduced during oxygen reduction that affect the deteri-ration of the function and viability of the spermatozoa, if
∗ Corresponding author. Tel.: +48 71 32 05 300; fax: +48 71 32 01 006.E-mail address: [email protected] (A. Partyka).
378-4320/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.anireprosci.2012.07.007
© 2012 Elsevier B.V. All rights reserved.
produced at greater than optimal concentrations (Aitkenet al., 1989; De Lamirande and Gagnon, 1993). Peroxidationof PUFAs in the sperm cell membranes is an autocatalytic,self-propagating reaction, which can cause cell dysfunc-tion associated with the loss of the membrane function,integrity and finally leads to decreased fertilizing ability ofspermatozoa (Alvarez and Storey, 1982).
As a consequence of the destructive potential of ROS,spermatozoa membranes must be protected by a highly
dation and antioxidant enzymes activity in avian semen.ci.2012.07.007
effective antioxidant system to prevent the peroxida-tive damage. The antioxidant system in semen includesenzymes such as glutathione peroxidase (GPx), superox-ide dismutase (SOD) and catalase (CAT) and also natural

ING Model
oductio
ARTICLEANIREP-4584; No. of Pages 7
2 A. Partyka et al. / Animal Repr
antioxidants: vitamins A, C, E, uric acid, glutathione,carotenoids (Breque et al., 2003; Cerolini et al., 2006;Partyka et al., 2012).
The superoxide anion (O2•−) is one of the major free rad-
icals produced in cells in the physiological state, therefore,SOD is considered to be the main element of initial antiox-idant defense in the semen (Surai et al., 2001). Superoxidedismutase catalyzes dismutation of the superoxide anionin the reaction: O2
•− + O2− + 2H+ → H2O2 + O2. This creates
another highly toxic reactive oxygen species – hydrogenperoxide (H2O2), which in turn is removed by glutathioneperoxidase and catalase (De Lamirande and Gagnon, 1993).
Glutathione peroxidase is a selenoprotein that catalyzesthe reduction reaction, not only of H2O2, but also of organicperoxides, especially lipid peroxides with reduced glu-tathione (GSH): 2GPx + H2O2 → GSSG + 2H2O. Glutathionedisulfide (GSSG), which is formed in this reaction is danger-ous for a cell, but is reduced by glutathione reductase viaoxidation of NADPH: GSSG + NADPH + H+ → NADP+ + 2GSH.
As noted above, the primary function of catalase in thecells is the participation in the reaction of disproportion-ation of hydrogen peroxide: 2H2O2 → 2H2O + O2. Catalasecontains a heme group and the binding site for NADPH. Thisenzyme is effective in the cells, protecting them againstH2O2 toxicity. Depending on the concentration of H2O2,CAT removes it, converting into H2O and O2, or removesH2O2, subsequently causing oxidation of suitable hydro-gen donors (such as ethanol, methanol; Scibor and Czeczot,2006).
Despite such a significant role of the antioxidant system,information regarding antioxidant enzymes in the semenof birds is scarce. Surai et al. (1998) have demonstrated thepresence of SOD and GPx in the spermatozoa and seminalplasma of chickens, turkeys, geese, ducks and guinea fowls.These enzymes, however, showed species-specific differ-ences in activity and distribution in the semen. Recently,we have reported for the first time the activity of CATin chicken semen (Partyka et al., 2012). However, to theauthors’ knowledge, the available data are lacking on theactivity of catalase and LPO in goose semen.
Therefore, the aim of this study was to compare theactivity of antioxidant enzymes in relation to lipid per-oxidation in the semen of two different avian species –chicken and geese. The relationship between sperm plasmamembrane integrity, lipid peroxidation and antioxidantenzymatic activities (CAT, GPx, SOD) in seminal plasma andspermatozoa was also determined.
2. Materials and methods
2.1. Animals
The experiment was conducted on Greenleg Par-tridge roosters and White Koluda® geese, each rep-resented by 10 mature males (2-year-old) kept indi-vidually in cages (60 cm × 50 cm × 75 cm – roosters)
Please cite this article in press as: Partyka, A., et al., Lipid peroxiAnim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anirepros
and (70 cm × 95 cm × 85 cm – geese) at 18–20 ◦C, under14L:10D photoperiod. Birds were fed commercial feed forbreeding flocks, without additional antioxidant supple-mentation. Water was provided ad libitum.
PRESSn Science xxx (2012) xxx– xxx
Protocols for working with the animals were in accor-dance with the Polish national guidelines for the careand use of experimental animals. All procedures wereperformed with the agreement of the IInd Local EthicalCommittee in Wroclaw.
2.2. Semen collection
The pooled semen from 10 individuals of each specieswas collected two times a week, by the dorso-abdominalmassage method (Burrows and Quinn, 1937). To maximizethe semen quality and quantity, the collection was alwaysperformed by the same people, under the same conditions,time, and massage method. Only clean ejaculates were usedfor further analysis. Fifteen semen collections from eachspecies were performed during 8 consecutive weeks.
2.3. Experimental design
In pooled samples the concentration of spermatozoawas determined by hemocytometer counts. Each pooledsample was divided into five aliquots. One of the dropletswas diluted in EK diluent (Partyka et al., 2011a) to obtain50 × 106 spermatozoa per mL for flow cytometric evalua-tion of sperm plasma membrane integrity.
The rest of semen aliquots (500 �L each) were cen-trifuged at 4000 × g for 10 min (MiniSpin® Eppendorf,Hamburg, Germany). The samples of seminal plasma werestored at −80 ◦C until further analysis. The sperm pel-lets were washed in PBS and centrifuged 3×. After thelast centrifugation, 1 mL of deionized water was added tothe spermatozoa and they were snap-frozen and stored at−80 ◦C until further analysis.
To determine LPO and the antioxidant enzymes inthe spermatozoa the cells were frozen-thawed threetimes and centrifuged (8000 × g, 5 min). After centrifu-gation the resultant supernatant was used for the assay(Kasimanickam et al., 2006). The lipid peroxidation andantioxidant enzymatic activity in sperm cells were calcu-lated in appropriate units per 109 spermatozoa (spz).
2.4. Plasma membrane integrity
The percentage of viable (plasma membrane intact;PMI) sperm was determined by flow cytometry using40,000 cells per sample, as we described before (Partykaet al., 2010). Five �L of SYBR-14 working solution (commer-cial solution diluted 50-fold; Live/Dead Sperm Viability Kit,InvitrogenTM, Eugene, OR, USA) was added to the dilutedsemen (300 �L) and incubated at room temperature for10 min and then the cells were counterstained with 5 �Lpropidium iodide (PI) 5 min before analysis. The sampleswere analyzed using FACSCalibur (Becton Dickinson, SanJose, CA, USA) equipped with an argon laser (488 nm), the
dation and antioxidant enzymes activity in avian semen.ci.2012.07.007
FL-2 detector to detect SYBR-14 positive PI-negative sperm(PMI), and the FL-1 detector to detect PI-positive sperm. Fif-teen plasma membrane integrity evaluations in semen ofeach species were performed.

ING Model
A
oductio
2e
wU
2
tb24oa55po1bs
ppMa
2
s(etawiwt3pors5Cs
2
ma(doptrpw3
ARTICLENIREP-4584; No. of Pages 7
A. Partyka et al. / Animal Repr
.5. Determination of lipid peroxidation and antioxidantnzymes activity
Lipid peroxidation and antioxidant enzyme activitiesere determined using a spectrophotometer Carry ConcV–vis (Varian Inc. Palo Alto, USA).
.5.1. Lipid peroxidationFor determination of the quantity of lipid peroxida-
ion in the seminal plasma and spermatozoa, the methodased on the reaction of a chromogenic reagent, N-methyl--phenylindole (NMPI) with malondialdehyde (MDA) at5 ◦C was used (Kasimanickam et al., 2006). One moleculef MDA reacted with two molecules of NMPI to yield
stable carbocyanine dye with maximum absorption at86 nm. According to the protocol of commercial kit (MDA-86, OxisResearchTM, OR, USA), 200 �L of samples (seminallasma or supernatant from sperm) were mixed with 10 �Lf probucol and 640 �L of NMPI and briefly vortexed. Then50 �L of hydrochloric acid was added, mixed and incu-ated at 45 ◦C for 60 min. To obtain a clear supernatant,amples were centrifuged 10,000 × g for 10 min.
The MDA in the sample was determined from the sam-le’s absorbance at 586 nm and the standard curve wasrepared using the MDA standard provided in the kit. TheDA concentration in the plasma was expressed as �M/mL
nd in spermatozoa as �M/109 spz.
.5.2. Catalase (CAT)Catalase (EC 1.11.1.6) activity was assessed by a two-
tep procedure using a commercial kit Catalase-520 AssayOxisResearchTM, Portland, OR, USA) as described by C oyant al. (2011). The samples (30 �L) were incubated inhe presence of 10 mM hydrogen peroxide (500 �L) andfter the incubation for precisely 1 min, the reactionas quenched with sodium azide (500 �L). The remain-
ng amount of H2O2 in 20 �L of the reaction mixtureas then determined by the oxidative coupling reac-
ion of 4-aminophenazone (4-aminoantipyrene, AAP) and,5-dichloro-2-hydroxybenzenesulfonic acid (DHBS) in theresence of H2O2 and catalyzed by horseradish per-xidase (HRP) (total volume of reagents – 2 mL). Theesulting quinoneimine dye (N-(4-antipyrl)-3-chloro-5-ulfonate-p-benzoquinonemonoimine) was measured at20 nm after a 10 min incubation at room temperature. TheAT activity in the plasma was expressed as U/mL, and inpermatozoa as U/109 spz.
.5.3. Glutathione peroxidase (GPx)GPx (EC 1.11.1.9) activity was determined using com-
ercial kit GPx-340 (OxisResearchTM, Portland, OR, USA)s described Kasimanickam et al. (2006) and C oyan et al.2011). The assay was based on a nicotinamide adenineinucleotide phosphate (NADPH) coupled reaction wherexidized glutathione, produced upon reduction of organiceroxide (tert-butyl hydroperoxide) by GPx, was recycledo its reduced state by utilizing the enzyme glutathione
Please cite this article in press as: Partyka, A., et al., Lipid peroxiAnim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anirepros
eductase and NADPH. Immediately prior to assay, the sam-les were diluted in an adequate ratio in buffer (providedith the commercial kit). To 350 �L of assay buffer and
50 �L of NADPH, 70 �L of the sample was added. Then
PRESSn Science xxx (2012) xxx– xxx 3
350 �L of tert-butyl hydroperoxide was added at 25 ◦C.The oxidation of NADPH to NADP+ was associated witha decrease in absorbance at 340 nm, and was measuredfor 3 min providing a spectrophotometric means for mon-itoring GPx activity. The rate of decrease in absorbance at340 nm was directly proportional to the GPx activity. TheGPx activity in the plasma was expressed as mU/mL, and inspermatozoa as mU/109 spz.
2.5.4. Superoxide dismutase (SOD)The SOD (EC 1.15.1.1) assay using commercial
SOD Assay Kit (Sigma–Aldrich®, St. Louis, Missouri,USA) was based on utilization of Dojindo’s highlywater-soluble tetrazolium salt, WST-1 (2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium,monosodium salt) that produced a water-soluble for-mazan dye upon reduction with a superoxide anion. Therate of the reduction with O2 was linearly related to thexanthine oxidase (XO) activity, and was inhibited by SOD.According to the protocol of kit, 40 �L of sample in 900 �Lof buffer (provided in commercial kit) were incubatedwith 30 �L of 1-methyl-2-vinylpyridinium trifluoro-methanesulfonate in HCl at 37 ◦C for 1 min. Then 30 mL of5,6,6a,11b-tetrahydro-3,9,10-trihydroxybenzo[c]fluorenein HCl containing diethyl-enetriamine-pentaacetic acid(DTPA) and ethanol were added, vortexed briefly andimmediately measured.
The SOD activity, as an inhibition activity, was quanti-fied by measuring the decrease in the color developmentat 450 nm. The SOD activity in the plasma was expressedas U/mL and in spermatozoa as U/109 spz. Samples withenzymes activities outside the kit’s assay range were fur-ther diluted or evaluated again in smaller volume
2.6. Data analysis
Statistical analyses were performed using STATISTICA(StatSoft, Inc. (2001), version 6). The data obtained wereanalyzed by ANOVA and Duncan’s multiple range test.Pearson correlation coefficients were calculated to deter-mine the associations between sperm plasma membraneintegrity, lipid peroxidation and antioxidant enzymaticactivities (CAT, GPx, SOD) in seminal plasma and sper-matozoa. The results are presented as mean ± SD ofmeasurements on samples from 15 replicate determina-tions.
3. Results
By creating two dimensional dot plots of PI comparedto SYBR-14 fluorescence, the subpopulation of sperm PInegative and stained by SYBR-14, indicating that thesecells were plasma membrane intact (PMI) were noted.Chicken sperm with intact plasma membrane (PMI) was89.1 ± 1.9% which was greater (P < 0.01) than for goosesperm (75.2 ± 8.6%) (Table 1).
dation and antioxidant enzymes activity in avian semen.ci.2012.07.007
3.1. Lipid peroxidation in sperm cells and seminal plasma
The amount of lipid peroxidation differed greatlyin both species. The concentration of MDA in chicken

ARTICLE ING Model
ANIREP-4584; No. of Pages 7
4 A. Partyka et al. / Animal Reproductio
Table 1Mean (±SD) of plasma membrane integrity, malondialdehyde concen-tration and antioxidant enzymes activity in chicken and goose seminalplasma and spermatozoa (n = 15).
Parameters Chicken semen Goose semen
PMI (%) 89.1 ± 1.9 A 75.2 ± 8.6 B
MDAp [�M/ml] 1.8 ± 0.4 A 0.9 ± 0.1 B
MDAspz [�M/109 spz] 5.0 ± 0.9 A 50.4 ± 15.8 B
CATp [U/mL] 46.8 ± 14.5 A 76.3 ± 8.7 B
CATspz [U/109 spz] 9.7 ± 1.5 A 239.2 ± 50.7 B
GPxp [mU/mL] 12.7 ± 3.0 A 28.7 ± 7.8 B
GPxspz [mU/109 spz] 7.6 ± 1.2 A 61.5 ± 25.4 B
SODp [U/mL] 91.7 ± 3.9 A 73.7 ± 13.0 B
SODspz [U/109 spz] 44.4 ± 4.0 A 308.5 ± 116.4 B
Different superscripts indicate differences for seminal plasma and sper-matozoa between species: A,B P < 0.01. PMI: plasma membrane integrity(%); MDAp: malondialdehyde concentration as amount of lipid perox-idation in seminal plasma [�M/ml]; CATp: catalase activity in seminalplasma [U/mL]; GPxp: glutathione peroxidase activity in seminal plasma[mU/mL]; SODp: superoxide dismutase activity in seminal plasma [U/mL];MDAspz: malondialdehyde concentration as amount of lipid peroxidationin spermatozoa [�M/109 spz]; CATspz: catalase activity in spermato-
zoa [U/109 spz]; GPxspz: glutathione peroxidase activity in spermatozoa[mU/109 spz]; SODspz: superoxide dismutase activity in spermatozoa[U/109 spz].seminal plasma was greater (P < 0.01) than in geese, but10× greater in goose than chicken sperm (P < 0.01; Table 1).
3.2. Activity of antioxidant enzymes
The activity of the antioxidant enzymes differed greatlyin both species. Catalase activity was detected in seminalplasma and spermatozoa obtained from the both species.The activity of this enzyme in spermatozoa and seminalplasma was greater (P < 0.01) in geese than in chickens(Table 1).
The activity of other antioxidants in seminal plasma alsoshowed species-specific differences. In White Koluda®, GPxactivity was two times greater (P < 0.01) than in chickenseminal plasma, whereas SOD activity was less (P < 0.01)than in chicken seminal plasma (Table 1). Each antioxidantenzyme had greater (P < 0.01) activity in goose than chickensperm (Table 1).
3.3. Correlation of sperm plasma membrane integrity,lipid peroxidation and antioxidant enzyme activity
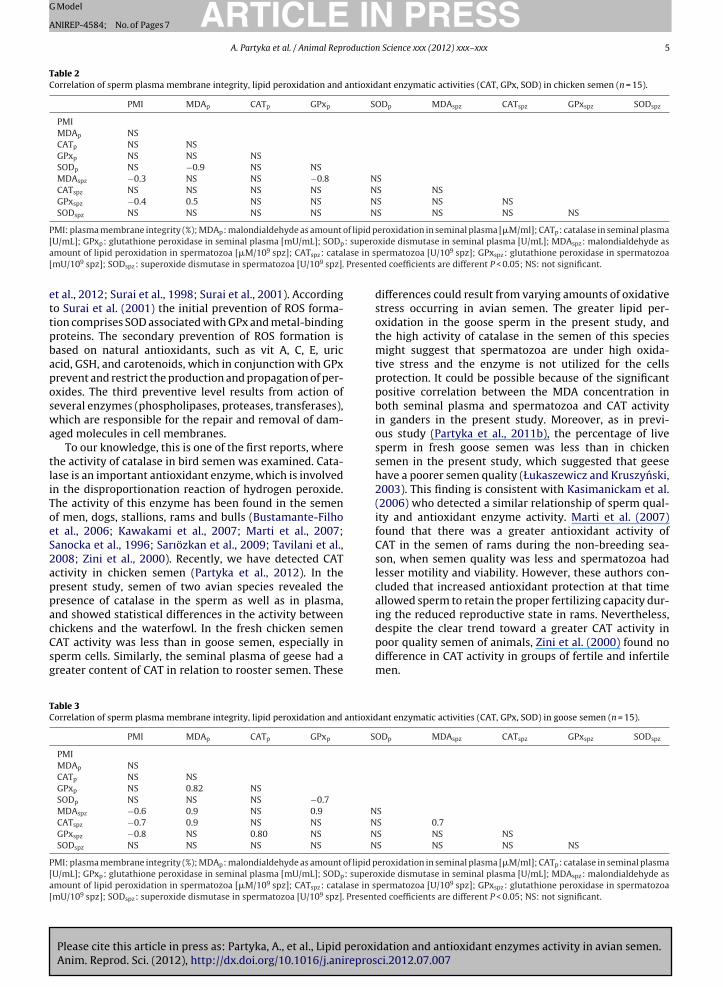
Correlations between PMI, MDA concentration andantioxidant enzymatic activity in Greenleg Partridge andWhite Koluda® semen are presented in Tables 2 and 3,respectively. In chicken semen, sperm lipid peroxidationand sperm GPx activity were negatively correlated with thePMI (P < 0.05). The MDA concentration was negatively cor-related with seminal plasma SOD activity, and positivelycorrelated with sperm GPx activity (P < 0.05). The amountof MDA in spermatozoa was negatively correlated withplasma GPx activity (P < 0.05) (Table 2).
In goose semen, the PMI was negatively correlated withspermatozoal MDA concentration and CAT, GPx activities
Please cite this article in press as: Partyka, A., et al., Lipid peroxiAnim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anirepros
(P < 0.05). The concentration of MDA in seminal plasma waspositively correlated with spermatozoal MDA concentra-tion and intracellular CAT activity, as well as with seminalplasma GPx activity (P < 0.05). The amount of MDA in
PRESSn Science xxx (2012) xxx– xxx
spermatozoa was positively correlated with CAT activityin cells and GPx activity in seminal plasma (P < 0.05). TheGPx activity was negatively correlated with SOD activityin plasma (P < 0.05). The activity of CAT in seminal plasmaand GPx activity in spermatozoa was positively correlated(P < 0.05; Table 3).
4. Discussion
This is the initial study performed to investigate theactivity of antioxidant enzymes, catalase and lipid per-oxidation in semen of two avian species, particularly ingeese semen. The antioxidant system in the semen con-taining enzymes and natural antioxidants acts to preventor restrict the formation and propagation of peroxides.Lesser amounts of antioxidants, or inhibition of antioxi-dant enzymes cause the oxidative stress and may damageor kill the germ cells. In mammals the effect of ROS on sper-matozoa and semen quality is well documented (Aitkenand Baker, 2004; Alvarez and Storey, 1982; Sanocka et al.,1997), while in avian semen such data are scarce.
One of the by-products of lipid peroxide decompo-sition is malondialdehyde, which is commonly used inbiochemical assay to monitor the degree of peroxida-tive damage sustained by spermatozoa. The results of thepresent study showed that the goose spermatozoa hadsignificantly greater MDA concentrations than chickencells. It is consistent with our previous study (Partykaet al., 2011b), where with the use of C11-BODIPY581/591, wedetected a greater percentage of spermatozoa with LPO infresh goose than chicken semen. Therefore, based on theaccessible reports and investigations in the present study,indicate there are species-specific differences in amountsof LPO in the semen of birds. However, the method toassess these by-products might have also had a consider-able influence. In the present study the concentration ofMDA in goose seminal plasma was two times less thanin chickens, and was due to the lesser sperm concentra-tion in gander semen. Whereas in goose spermatozoa 10times greater concentrations of the intracellular MDA wasdetected as compared to chicken cells. Previous research,using a similar commercial kit obtained a similar value ofMDA in whole fresh semen of turkeys (Long and Kramer,2003), but Blesbois et al. (1993) reported a small amount ofMDA in chicken semen. Lesser amounts of lipid peroxida-tion in fresh chicken semen, assessed by the same method,were also obtained in our recent study (Partyka et al., 2012)and as well as by thiobarbituric acid reaction (TBARs) meth-ods (Partyka et al., 2007; Partyka and Łukaszewicz, 2008).Nevertheless, it is noteworthy that comparisons among dif-ferent studies are often difficult to interpret, due to thedifferences in the method of analysis and sperm processingprotocols. Therefore these apparent discrepancies might bemainly due to the more sensitive test used in the presentstudy, which allowed the measurement of both, free andprotein-bound MDA.
While the antioxidant system in semen of mammals has
dation and antioxidant enzymes activity in avian semen.ci.2012.07.007
been well characterized in several experiments (Kankoferet al., 2005; Kawakami et al., 2007; Marti et al., 2007),only few studies are available about the antioxidant sys-tem in avian semen (Froman and Thurston, 1981; Partyka
ARTICLE IN PRESSG Model
ANIREP-4584; No. of Pages 7
A. Partyka et al. / Animal Reproduction Science xxx (2012) xxx– xxx 5
Table 2Correlation of sperm plasma membrane integrity, lipid peroxidation and antioxidant enzymatic activities (CAT, GPx, SOD) in chicken semen (n = 15).
PMI MDAp CATp GPxp SODp MDAspz CATspz GPxspz SODspz
PMIMDAp NSCATp NS NSGPxp NS NS NSSODp NS −0.9 NS NSMDAspz −0.3 NS NS −0.8 NSCATspz NS NS NS NS NS NSGPxspz −0.4 0.5 NS NS NS NS NSSODspz NS NS NS NS NS NS NS NS
PMI: plasma membrane integrity (%); MDAp: malondialdehyde as amount of lipid peroxidation in seminal plasma [�M/ml]; CATp: catalase in seminal plasma[ : superoa lase in s[ Present
ettpbaposwa
tliToeS2appacCsg
TC
P[a[
U/mL]; GPxp: glutathione peroxidase in seminal plasma [mU/mL]; SODp
mount of lipid peroxidation in spermatozoa [�M/109 spz]; CATspz: catamU/109 spz]; SODspz: superoxide dismutase in spermatozoa [U/109 spz].
t al., 2012; Surai et al., 1998; Surai et al., 2001). Accordingo Surai et al. (2001) the initial prevention of ROS forma-ion comprises SOD associated with GPx and metal-bindingroteins. The secondary prevention of ROS formation isased on natural antioxidants, such as vit A, C, E, uriccid, GSH, and carotenoids, which in conjunction with GPxrevent and restrict the production and propagation of per-xides. The third preventive level results from action ofeveral enzymes (phospholipases, proteases, transferases),hich are responsible for the repair and removal of dam-
ged molecules in cell membranes.To our knowledge, this is one of the first reports, where
he activity of catalase in bird semen was examined. Cata-ase is an important antioxidant enzyme, which is involvedn the disproportionation reaction of hydrogen peroxide.he activity of this enzyme has been found in the semenf men, dogs, stallions, rams and bulls (Bustamante-Filhot al., 2006; Kawakami et al., 2007; Marti et al., 2007;anocka et al., 1996; Sarıözkan et al., 2009; Tavilani et al.,008; Zini et al., 2000). Recently, we have detected CATctivity in chicken semen (Partyka et al., 2012). In theresent study, semen of two avian species revealed theresence of catalase in the sperm as well as in plasma,nd showed statistical differences in the activity between
Please cite this article in press as: Partyka, A., et al., Lipid peroxiAnim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anirepros
hickens and the waterfowl. In the fresh chicken semenAT activity was less than in goose semen, especially inperm cells. Similarly, the seminal plasma of geese had areater content of CAT in relation to rooster semen. These
able 3orrelation of sperm plasma membrane integrity, lipid peroxidation and antioxid
PMI MDAp CATp GPxp S
PMIMDAp NSCATp NS NSGPxp NS 0.82 NSSODp NS NS NS −0.7MDAspz −0.6 0.9 NS 0.9 NCATspz −0.7 0.9 NS NS NGPxspz −0.8 NS 0.80 NS NSODspz NS NS NS NS N
MI: plasma membrane integrity (%); MDAp: malondialdehyde as amount of lipid pU/mL]; GPxp: glutathione peroxidase in seminal plasma [mU/mL]; SODp: superomount of lipid peroxidation in spermatozoa [�M/109 spz]; CATspz: catalase in smU/109 spz]; SODspz: superoxide dismutase in spermatozoa [U/109 spz]. Present
xide dismutase in seminal plasma [U/mL]; MDAspz: malondialdehyde aspermatozoa [U/109 spz]; GPxspz: glutathione peroxidase in spermatozoaed coefficients are different P < 0.05; NS: not significant.
differences could result from varying amounts of oxidativestress occurring in avian semen. The greater lipid per-oxidation in the goose sperm in the present study, andthe high activity of catalase in the semen of this speciesmight suggest that spermatozoa are under high oxida-tive stress and the enzyme is not utilized for the cellsprotection. It could be possible because of the significantpositive correlation between the MDA concentration inboth seminal plasma and spermatozoa and CAT activityin ganders in the present study. Moreover, as in previ-ous study (Partyka et al., 2011b), the percentage of livesperm in fresh goose semen was less than in chickensemen in the present study, which suggested that geesehave a poorer semen quality (Łukaszewicz and Kruszynski,2003). This finding is consistent with Kasimanickam et al.(2006) who detected a similar relationship of sperm qual-ity and antioxidant enzyme activity. Marti et al. (2007)found that there was a greater antioxidant activity ofCAT in the semen of rams during the non-breeding sea-son, when semen quality was less and spermatozoa hadlesser motility and viability. However, these authors con-cluded that increased antioxidant protection at that timeallowed sperm to retain the proper fertilizing capacity dur-ing the reduced reproductive state in rams. Nevertheless,
dation and antioxidant enzymes activity in avian semen.ci.2012.07.007
despite the clear trend toward a greater CAT activity inpoor quality semen of animals, Zini et al. (2000) found nodifference in CAT activity in groups of fertile and infertilemen.
ant enzymatic activities (CAT, GPx, SOD) in goose semen (n = 15).
ODp MDAspz CATspz GPxspz SODspz
SS 0.7S NS NSS NS NS NS
eroxidation in seminal plasma [�M/ml]; CATp: catalase in seminal plasmaxide dismutase in seminal plasma [U/mL]; MDAspz: malondialdehyde aspermatozoa [U/109 spz]; GPxspz: glutathione peroxidase in spermatozoaed coefficients are different P < 0.05; NS: not significant.

ING Model
oductio
ARTICLEANIREP-4584; No. of Pages 7
6 A. Partyka et al. / Animal Repr
Glutathione peroxidase plays a major role in main-taining the balance of pro- and antioxidation, because itremoves lipid peroxides from the cells, converting theminto non-reactive products (Fraczek and Kurpisz, 2005).Kasimanickam et al. (2006) showed the negative correla-tion between GPx activity in spermatozoa of young rams,and the percentage of normal spermatozoa with progres-sive movement. In addition, the group of animals withreduced semen quality had a greater activity of glutathioneperoxidase. The present study also confirmed this find-ing. We noted the negative relation between GPx activityin spermatozoa and sperm plasma membrane integrityin both species. The greater GPx activity in gander semi-nal plasma and spermatozoa, supports the hypothesis thatthere is distinctly lesser semen quality in geese derivedfrom wild species Anser anser L., as reported previously(Łukaszewicz and Kruszynski, 2003; Partyka et al., 2011b)may result in the functional and structural impairment ofsperm metabolic mechanisms and finally the decreasedutilization of antioxidant enzymes. Previous studies havealso shown species-specific differences in the distributionof Se-dependent glutathione peroxidase (Surai et al., 1998).Greater GPx activity was detected in goose than chickensperm, but in chicken seminal plasma the activity of thisenzyme was relatively greater as compared to goose sperm(Surai et al., 1998).
In agreement with previous observations, the currentstudy also indicated species-specific differences in super-oxide dismutase activity. Surai et al. (1998) reported thegreatest intracellular antioxidant protection from SODactivity in geese and the least in turkey spermatozoawhereas SOD activity in seminal plasma was less in waterfowl and greater in turkeys. In the present experiment therewas a similar trend. Moreover, there was a negative rela-tionship between seminal plasma activity of SOD and GPxin geese, which could explain why in ganders only semi-nal plasma SOD activity was less as compared with otherenzymes. Nevertheless, the concurrent increase in MDAproduction and SOD activity in spermatozoa observed inthe present study might indicate that SOD also was notengaged for sperm supporting against oxidative stress andwas not able to maintain sperm quality in fresh semen.Results of the present study are consistent with previ-ous studies indicating that greater amounts of MDA wereaccompanied by greater SOD activity in human sperma-tozoa (Aitken et al., 1993, 2006). However, Cerolini et al.(2001) showed that boar semen samples with the great-est viability after freezing-thawing also had greater SODactivity. This finding may be reflected in previous studies(Partyka et al., 2011b), where goose semen had a greaterpercentage of live spermatozoa after thawing than chickensemen.
In conclusion, the comparative studies described here,emphasize the profound implications of species on activ-ity of antioxidant defense and amount of lipid peroxidation.This is one of the first reports, where the activity of catalasein bird semen was found. We demonstrated that the activ-
Please cite this article in press as: Partyka, A., et al., Lipid peroxiAnim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anirepros
ity of CAT, GPx and SOD remained greater in goose than inchicken sperm. The greater lipid peroxidation and greateractivity of antioxidant enzymes in goose semen might sug-gest that spermatozoa were under high oxidative stress and
PRESSn Science xxx (2012) xxx– xxx
the enzymes were not utilized for the protection of func-tionally and structurally impaired cells. In turn, in freshchicken semen we detected a lesser activity of antioxidantenzymes accompanied with lesser lipid peroxidation andgood semen quality that could indicate that fowl sperma-tozoa were under oxidative stress, but the enzymes wereemployed to protect and maintain sperm quality. How-ever, the excessive usage of antioxidant defense systemin chicken semen, would probably lead to its insufficientreserve, necessary for the spermatozoa protection againstLPO during cryopreservation, as has been recently demon-strated (Partyka et al., 2012). Therefore, such data wouldbe of particular importance to gain a greater understand-ing of sperm biology in avian species and to understandthe species-specific differences in the manifestation andeffectiveness of the semen antioxidant system.
Acknowledgments
This study was supported by the Polish Ministry of Sci-ence and Higher Education, grant no. N N311 2217 33. Theauthors wish to thank Dr Maria Chrzanowska, Dr AnnaJerysz for the technical support and Dr Małgorzata Ochotafor improving language.
References
Aitken, R.J., Baker, M.A., 2004. Oxidative stress and male reproductivebiology. Reprod. Fertil. Dev. 16, 581–588.
Aitken, R.J., Clarkson, J.S., Fishel, S., 1989. Generation of reactive oxygenspecies, lipid peroxidation and human sperm function. Biol. Reprod.40, 183–197.
Aitken, R.J., Harikss, D., Buckingham, D.W., 1993. Analysis of lipid per-oxidation mechanisms in human spermatozoa. Mol. Reprod. Dev. 35,302–315.
Aitken, R.J., Wingate, J.K., De Iuliis, G.N., Koppers, A.J., McLaughlin, E.A.,2006. Cis-unsaturated fatty acids stimulate reactive oxygen speciesgeneration and lipid peroxidation in human spermatozoa. J. Clin.Endocrinol. Metab. 91, 4154–4163.
Alvarez, J.G., Storey, B.T., 1982. Spontaneous lipid peroxidation in rabbitepididymal spermatozoa: its effect on sperm motility. Biol. Reprod.27, 1102–1108.
Blesbois, E., Grasseau, I., Blum, J.C., 1993. Effect of vitamin E on fowl semenstorage at 4 ◦C. Theriogenology 39, 771–779.
Breque, C., Surai, P., Brillard, J.P., 2003. Roles of antioxidants on prolongedstorage of avian spermatozoa in vivo and in vitro. Mol. Reprod. Dev.66, 314–323.
Burrows, W.H., Quinn, J.P., 1937. The collection of spermatozoa from thedomestic fowl and turkey. Poult. Sci. 16, 19–24.
Bustamante-Filho, I.C., Pederzolli, C.D., Sgaravatti, A.M., Mattos, R.C.,Dutra-Filho, C.S., Jobim, M.I.M., 2006. Activity of glutathione perox-idase and catalase in stallion semen during cryopreservation. Anim.Reprod. Sci. 94, 70–73.
Cerolini, S., Maldjian, A., Pizzi, F., Gliozzi, T.M., 2001. Changes in spermquality and lipid composition during cryopreservation of boar semen.Reproduction 121, 395–401.
Cerolini, S., Zainiboni, L., Maldjian, A., Gliozzi, T., 2006. Effect of docosahex-aenoic acid and �-tocopherol enrichment in chicken sperm on semenquality, sperm lipid composition and susceptibility to peroxidation.Theriogenology 66, 877–886.
C oyan, K., Bas pınar, N., Bucak, M.N., Akalın, P.P., 2011. Effects of cysteineand ergothioneine on post-thawed Merino ram sperm and biochem-ical parameters. Cryobiology 63, 1–6.
De Lamirande, E., Gagnon, C., 1993. A positive role for superoxide anionsin triggering hyperactivation and capacitation of human spermatozoa.Int. J. Androl. 16, 21–25.
dation and antioxidant enzymes activity in avian semen.ci.2012.07.007
Fraczek, M., Kurpisz, M., 2005. Redox system in the male semen andsperm peroxidative damage. Postepy Hig. Med. Dosw. 59, 523–534(in Polish).
Froman, D.P., Thurston, R.J., 1981. Chicken and turkey superoxide dismu-tase: a comparative study. Biol. Reprod. 24, 193–200.

ING Model
A
oductio
F
K
K
K
L
Ł
M
P
P
P
P
2008. Activity of antioxidant enzymes in seminal plasma and their
ARTICLENIREP-4584; No. of Pages 7
A. Partyka et al. / Animal Repr
ujihara, N., Howarth, B., 1978. Lipid peroxidation in fowl spermatozoa.Poult. Sci. 57, 1766–1768.
ankofer, M., Kolm, G., Aurich, J., Aurich, C., 2005. Activity of glutathioneperoxidase, superoxide dismutase and catalase and lipid peroxidationintensity in stallion semen during storage at 5 ◦C. Theriogenology 63,1354–1365.
asimanickam, R., Pelzer, K.D., Kasimanickam, V., Swecker, W.S., Thatcher,C.D., 2006. Association of classical semen parameters, sperm DNAfragmentation index, lipid peroxidation and antioxidant enzymaticactivity of semen in ram-lambs. Theriogenology 65, 1407–1421.
awakami, E., Takemura, A., Sakuma, M., Takano, M., Hirano, T., Hori, T.,Tsutsui, T., 2007. Superoxide dismutase and catalase activities in theseminal plasma of normozoospermic and asthenozoospermic Beagles.J. Vet. Med. Sci. 69, 133–136.
ong, J.A., Kramer, M., 2003. Effect of vitamin E on lipid peroxidation andfertility after artificial insemination with liquid-stored turkey semen.Poult. Sci. 82, 1802–1807.
ukaszewicz, E., Kruszynski, W., 2003. Evaluation of fresh and frozen-thawed semen of individual ganders by assessment of spermatozoamotility and morphology. Theriogenology 59, 1627–1640.
arti, E., Mara, L., Marti, J.I., Muino-Blanco, T., Cebrián-Pérez, J.A., 2007.Seasonal variations in antioxidant enzyme activity in ram seminalplasma. Theriogenology 67, 1446–1454.
artyka, A., Jerysz, A., Pokorny, P., 2007. Lipid peroxidation in fresh andstored semen of Green-legged Partridge. E.J.P.A.U. 10 (2) (on-line).
artyka, A., Łukaszewicz, E., 2008. The effect of storage time and tempera-ture on lipid peroxidation and semen quality of various chicken breedsand lines. “Na pograniczu chemii i biologii”. Wydawnictwo NaukowePoznan XXI, 223–230 (in Polish).
artyka, A., Łukaszewicz, E., Nizanski, W., 2011a. Flow cytometric assess-
Please cite this article in press as: Partyka, A., et al., Lipid peroxiAnim. Reprod. Sci. (2012), http://dx.doi.org/10.1016/j.anirepros
ment of fresh and frozen-thawed Canada goose (Branta canadensis)semen. Theriogenology 76, 843–850.
artyka, A., Łukaszewicz, E., Nizanski, W., Twardon, J., 2011b. Detectionof lipid peroxidation in frozen-thawed avian spermatozoa using C11-BODIPY581/591. Theriogenology 75, 1623–1629.
PRESSn Science xxx (2012) xxx– xxx 7
Partyka, A., Nizanski, W., Łukaszewicz, E., 2010. Evaluation of fresh andfrozen-thawed fowl semen by flow cytometry. Theriogenology 74,1019–1027.
Partyka, A., Łukaszewicz, E., Nizanski, W., 2012. Effect of cry-opreservation on sperm parameters, lipid peroxidation andantioxidant enzymes activity in fowl semen. Theriogenology 77,1497–1504.
Sanocka, D., Miesel, R., Jedrzejczak, P., Chełmonska-Soyta, A., Kurpisz, M.K.,1997. Effect of reactive oxygen species and the activity of antioxi-dant systems on human semen; association with male infertility. Int.J. Androl. 20, 255–264.
Sanocka, D., Miesel, R., Jedrzejczak, P., Kurpisz, M.K., 1996. Oxidative stressand male infertility. J. Androl. 17, 449–454.
Sarıözkan, S., Bucak, M.N., Tuncer, P.B., Ulutas , P.A., Bilgen, A., 2009. Theinfluence of cysteine and taurine on microscopic-oxidative stressparameters and fertilizing ability of bull semen following cryopreser-vation. Cryobiology 58, 134–138.
Scibor, D., Czeczot, H., 2006. Catalase – structure, properties, functions.Postepy Hig. Med. Dosw. 60, 170–180 (in Polish).
Surai, P.F., Blesbois, E., Grasseau, I., Chalah, T., Brillard, J.P., Wishart,G.J., Cerolini, S., Sparks, N.H.C., 1998. Fatty acid composition, glu-tathione peroxidase and superoxide dismutase activity and totalantioxidant activity of avian semen. Comp. Biochem. Physiol. B 120,527–533.
Surai, P.F., Fujihara, N., Speake, B.K., Brillard, J.P., Wishart, G.J., Sparks,N.H.C., 2001. Polyunsaturated fatty acids, lipid peroxidation andantioxidant protection in avian semen. Asian-Aust. J. Anim. Sci. 14,1024–1050.
Tavilani, H., Goodarzi, M.T., Vaisi-Raygani, A., Salimi, S., Hassanzadeh, T.,
dation and antioxidant enzymes activity in avian semen.ci.2012.07.007
relationship with lipid peroxidation of spermatozoa. Int. Braz. J. Urol.34, 485–491.
Zini, A., Garrels, K., Phang, D., 2000. Antioxidant activity in the semen offertile and infertile men. Urology 55, 922–926.
Related Documents











