ELSEVIER Chemistry and Physics of Lipids 73 (1994) 139-158 Chemistry and Physics of LIPID$ Lipid domains and lipid/protein interactions in biological membranes Jean-Franqois Tocanne*, Laurence C6zanne, Andr6 Lopez, Barbora Piknova, Vincent Schram, Jean-Francois Tournier, M. Welby Laboratoire de Pharmacologieet ToxicologieFondamentales du CNRS, Dpt III: Gtvcoconjugu~set Biomembranes, 118, Route de Narbonne, F-31062 Toulouse Cedex, France Abstract In the fluid mosaic model of membranes, lipids are organized in the form of a bilayer supporting peripheral and integral proteins. This model considers the lipid bilayer as a two-dimensional fluid in which lipids and proteins are free to diffuse. As a direct consequence, both types of molecules would be expected tobe randomly distributed within the membrane. In fact, evidences are accumulating to indicate the occurrence of both a transverse and lateral regionalization of membranes which can be described in terms of micro- and macrodomains, including the two leaflets of the lipid bilayer. The nature of the interactions responsible for the formation of domains, the way they develop and the time- and space-scale over which they exist represent today as many challenging problems in membranology. In this report, we will first consider some of the basic observations which point to the role of proteins in the transverse and lateral regionalization of membranes. Then, we will discuss some of the possible mechanisms which, in particular in terms of lipid/protein interactions, can explain lateral heterogenities in membranes and which have the merit of providing a thermodynamic support to the existence of lipid domains in membranes. Keywords: Membranes; Lipid microdomains and macrodomains; Protein-lipid interactions; Hydrophobic mismatch; Lateral diffusion; Bacteriorhodopsin 1. Introduction Biological membranes are complex assemblies of lipids and proteins. One of the central features of these highly organized systems is the absence of covalent linkages between lipid and protein molecules which makes them normally free to undergo conformational, rotational and transla- * Corresponding author. Abbret~ations: ANoc, 8-(2-anthroyl)-octanoic acid; ANno, 9-(2-anthryl)-nonanoic acid; ASte, 12-(9-anthroyloxy)-stearic acid; DL-, DM-, DO-, DP-, DSPC, dilauroyl, dimyristoyl, dioleoyl, dipalmitoyl, distearoylphosphatidylcholine; DC13PC, ditridecanoylphos- phatidylcholine; DC15PC, dipentadecanoy!phosphatidylcholine; egg-PC, egg yolk phosphatidylcholine; C16-, Cts-diI, 1,1'-dihexade- cyl, 1,1'-dioctadecyl-3,3,3',13'-tetramethylindocarbocyanine; ClsdiO , 1,1'-dioctadecyl-3,3,3',3'-tetramethylindooxatricarbocyaninc; NBD-PC, 1-acyl-2-(N-4-nitrobenzo-2-oxa-l,3-diazol)- aminododecanoyl-phosphatidylcholine; NBD-PE, N-4-nitrobenzo-2-oxa-l,3- diazol-4-yl-phosphatidylethanolamine; C6-NBD-PE, 1-acyl-2-(N-4-nitrobenzo-2-oxa-l,3-diazol)- aminohexanoyl-phosphatidy- iethanolamine; HEDAF, 5-(N-hexadecanoyl)-aminofluorescein;R-PE, tetramethyl rodamine phosphatidylethanolamine; BR, bacte- riorhodopsin; TMA-DPH, 1-(4-trimethylammonium)-6-phenyl-l,3,5-hexatriene, p-toluene sulfonate. 0009-3084/94/$07.00 © 1994 Elsevier Science Ireland Ltd. All rights reserved. SSDI 0009-3084(94)02364-B

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ELSEVIER Chemistry and Physics of Lipids 73 (1994) 139-158
Chemistry and Physics of
LIPID$
Lipid domains and lipid/protein interactions in biological membranes
Jean-Franqois Tocanne*, Laurence C6zanne, Andr6 Lopez, Barbora Piknova, Vincent Schram, Jean-Francois Tournier, M. Welby
Laboratoire de Pharmacologie et Toxicologie Fondamentales du CNRS, Dpt III: Gtvcoconjugu~s et Biomembranes, 118, Route de Narbonne, F-31062 Toulouse Cedex, France
Abstract
In the fluid mosaic model of membranes, lipids are organized in the form of a bilayer supporting peripheral and integral proteins. This model considers the lipid bilayer as a two-dimensional fluid in which lipids and proteins are free to diffuse. As a direct consequence, both types of molecules would be expected tobe randomly distributed within the membrane. In fact, evidences are accumulating to indicate the occurrence of both a transverse and lateral regionalization of membranes which can be described in terms of micro- and macrodomains, including the two leaflets of the lipid bilayer. The nature of the interactions responsible for the formation of domains, the way they develop and the time- and space-scale over which they exist represent today as many challenging problems in membranology. In this report, we will first consider some of the basic observations which point to the role of proteins in the transverse and lateral regionalization of membranes. Then, we will discuss some of the possible mechanisms which, in particular in terms of lipid/protein interactions, can explain lateral heterogenities in membranes and which have the merit of providing a thermodynamic support to the existence of lipid domains in membranes.
Keywords: Membranes; Lipid microdomains and macrodomains; Protein-lipid interactions; Hydrophobic mismatch; Lateral diffusion; Bacteriorhodopsin
1. Introduction
Biological membranes are complex assemblies of lipids and proteins. One of the central features
of these highly organized systems is the absence of covalent linkages between lipid and protein molecules which makes them normally free to undergo conformational, rotational and transla-
* Corresponding author. Abbret~ations: ANoc, 8-(2-anthroyl)-octanoic acid; ANno, 9-(2-anthryl)-nonanoic acid; ASte, 12-(9-anthroyloxy)-stearic acid; DL-, DM-, DO-, DP-, DSPC, dilauroyl, dimyristoyl, dioleoyl, dipalmitoyl, distearoylphosphatidylcholine; DC13PC, ditridecanoylphos- phatidylcholine; DC15PC, dipentadecanoy!phosphatidylcholine; egg-PC, egg yolk phosphatidylcholine; C16-, Cts-diI, 1,1'-dihexade- cyl, 1,1'-dioctadecyl-3,3,3',13'-tetramethylindocarbocyanine; ClsdiO , 1,1'-dioctadecyl-3,3,3',3'-tetramethylindooxatricarbocyaninc; NBD-PC, 1-acyl-2-(N-4-nitrobenzo-2-oxa-l,3-diazol)- aminododecanoyl-phosphatidylcholine; NBD-PE, N-4-nitrobenzo-2-oxa-l,3- diazol-4-yl-phosphatidylethanolamine; C6-NBD-PE, 1-acyl-2-(N-4-nitrobenzo-2-oxa-l,3-diazol)- aminohexanoyl-phosphatidy- iethanolamine; HEDAF, 5-(N-hexadecanoyl)-aminofluorescein; R-PE, tetramethyl rodamine phosphatidylethanolamine; BR, bacte- riorhodopsin; TMA-DPH, 1-(4-trimethylammonium)-6-phenyl-l,3,5-hexatriene, p-toluene sulfonate.
0009-3084/94/$07.00 © 1994 Elsevier Science Ireland Ltd. All rights reserved. SSDI 0009-3084(94)02364-B

140 J.-F. Tocanne et al. /Chem. Phys. Lipids 73 (1994) 139-158
tional motions. In the fluid mosaic model of mem- branes, lipids are organized in the form of a bilayer supporting peripheral and integral pro- teins. This model, which stems from the above remarks, considers the lipid bilayer as a two-di- mensional fluid in which lipids and proteins are free to diffuse. As a direct consequence, both types of molecules would be expected to be ran- domly distributed within the membrane.
In fact, although the concept of the lipid bi- layer is currently accepted, membrane organiza- tion is certainly much more complex. Biological membranes contain a large variety of lipids and proteins. Such a diversity in lipid structure is not required to maintain the bilayer assembly and fluidity. Despite extensive studies on the structure of biological membranes during the last decade, we still have no direct information on the poten- tial specificity of lipid distribution around mem- brane proteins nor on the molecular organization of those lipids which are not in contact with proteins. Nevertheless, evidences are accumulat- ing to indicate the occurrence of both a trans- verse [1-3] and lateral [4-6] regionalization of membranes which can be described in terms of macro- and microdomains.
Macrodomains, which correspond to large membrane areas of at least a few tenths /xm 2 in surface, have been recognized in the plasma membrane of certain specialized cells with highly regionalized functions, such as spermatozoa, eggs and epithelia [4] and quite recently by fluores- cence digital imaging microscopy in the mem- brane of rabbit erythrocytes [7,8]. As we will see below, the two leaflets of the lipid bilayer can also be considered as two large and nearly indepen- dent membrane domains but with the possibility of transbilayer lipid redistribution.
Lipid microdomains extend from the submicron to the molecular scale. They have been recog- nized in the plasma membrane of many cell types and by means of a rather large variety of ap- proaches [5,6] among which the fluorescence re- covery after photobleaching technique is the most widely used [4,20].
In terms of structure, lipid and protein micro- domains are still poorly understood and it is clear that with a few exceptions, lipid domains detected
in native membranes have been operationally de- fined by heterogeneities in the behaviour of probe molecules [9]. The nature of the interactions re- sponsible for the formation of domains, the way they develop and the time- and space-scale over which they exist represent today as many chal- lenging problems in membranology. It is often suggested that membrane lateral heterogeneities might originate from intrinsic properties of the lipids, for example in terms of gel-phase/liquid- crystalline-phase separation [6]. Such'a possibility, which can occur in some specific cases, like in ram sperm plasma membrane [10,11], is not of general relevance to natural membranes. More probably, membrane proteins are responsible for the existence of these macro- and microdomains since in most cases, the lateral heterogeneities detected in a natural membrane are no longer observed in the corresponding protein-free lipid bilayers. Various hypotheses have been proposed to explain membrane compartmentation includ- ing the existence of a lattice of membrane pro- teins or the existence of specific and non-specific lipid/protein interactions. In this report, we will first consider some of the basic observations which point to the role of proteins in the transverse and lateral regionalization of membranes. Then, we will discuss some of the possible mechanisms which, in particular in terms of lipid/protein in- teractions, can explain lateral heterogenities in membranes and which have the merit of provid- ing a thermodynamic support to the existence of lipid domains in membranes.
2. Role of proteins on the transverse and lateral regionalization of membranes
2.1. Transverse regionalization of membranes
It is now well established that intrinsic and extrinsic membrane proteins are asymmetrically oriented and distributed with respect to the mem- brane plane [1,2]. Most of them are not free to diffuse laterally, presumably due to their attach- ment to cytoskeleton components [12]. In many cellular systems, lipids have been shown to be asymmetrically distributed between the two leaflets of the lipid bilayer [2,13]. In synthetic

J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158 141
lipid bllayers, spontaneous flip-flop of lipids from one leaflet to the other occurs at a very slow rate [14]. In contrast, in biological membranes, tran- sloeation of the two aminophospholipids phos- phatidylserine and phosphatidylethanolamine can occur very rapidly [3,14,15,19] and there is in- creasing evidence that the lipid transverse asymmetry is maintained by an energy- and pro- tein-dependent specific 'aminophospholipid tran- slocase' activity [3,14,15].
In terms of molecular interactions, the two halves of the bilayer are weakly coupled. Thus, in lipid vesicles, the gel-to-liquid phase transition can occur in one leaflet independently of the other [16]. As shown in the plasma membrane of human red blood cells [17], Chinese hamster lung fibroblasts [18] and in the apical plasma mem- brane of bovine aortic endothelial cells [19], lipids diffuse laterally more rapidly in the intraeyto- plasmic than in the exoeytoplasmic leaflet.
In these conditions, the two leaflets of the lipid bilayer in biological membranes can be con- sidered as two large and nearly independent membrane domains but with the possibility of transbilayer lipid redistribution, via a protein-de- pendent mechanism controlled by the physiologi- cal state of the cell [14].
2.2 Lateral regionalization of membranes
2.2 1. Lipid macrodomains The fact that a protein network can exist in
membranes and act as a fence preventing lipid and protein molecules to diffuse laterally from one membrane region to the other is clearly illus- trated by epithelial cells with the role of tight junctions. Epithelia are composed of highly polar- ized cells having a surface membrane which is separated by tight junctions into apical and ba- solateral domains. These two membrane domains differ in terms of morphology, transport processes, ionic permeability, distribution of glycolipids and proteins and sensitivity to hormones and drugs ([21] and references cited therein). FRAP data [22,23] shows that for probe molecules located in the exoplasmic leaflet of the plasma membrane, confluent cell monolayers can be described as a mosaic of closed diffusion areas. In these macro-
domains, which are clearly restricted to the apical and basolateral areas, lipids can diffuse but they do not flow from one cell to the next or from the apical to the basolateral membrane. In contrast, FRAP [24] and lipid transfer [25] data suggest that lipids can exchange between the apical and basal domains of the plasma membrane in the cytoplasmic leaflet. Differences in fluidity also exist between the apical and basolateral domains, the highest lipid D values being found in the apical pole in bovine aortic [22] and pulmonary artery [23] endothelial cells but in the basal pole in rat proximal tubule epithelial cells [26]. Tight junctions are believed to be the barrier responsi- ble for such membrane macrocompartmentation [22,24,25,27-29]. After disruption of cell contacts, probes located in the exoplasmic leaflet can move from the apical to the basolateral pole of the membrane [22]. In vascular endothelium, cell junctions have also been shown to reversibly af- fect overall plasma membrane organization [22]. When cell junctions are disrupted, the lateral diffusion coefficient of HEDAF is the same in both apical and basal domains (2 10 -9 cm2/s). It regains its initial higher value (2.7 10 -9 cm2/s) in the apical pole when cell contacts are restored [22]. In renal epithelium, exposure of the cells to calcium-free medium which provokes microfile- ment retraction and reversible opening of the tight junctions, brought about a 4-fold increase in the diffusion coefficient of the probe C6-NBD-PE in the apical plasma membrane [26], indicating that tight junctions and also the cytoskeleton con- trol the membrane organization and therefore the lipid D.
A tight junction-like barrier to movements of lipids has recently been demonstrated to exist in polarized hippocampal neurons in culture. This functional barrier, which is located at the axonal hillock/initial segments and concerns both the outer and inner leaflets of the plasma membrane, is probably involved in the maintenance of the compositional differences which exist between the axonal and somatodentritic domains in these cells [30].
Highly polarized and differentiated cells like spermatozoa and early embryo are able to over- come the randomizing effect of lateral diffusion

142 J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158
and to localize certain membrane components in specific regions of the cell surface [31-33].
Thus, spermatozoa exhibit membrane regional- ization, with different lipid and protein composi- tion between the anterior and posterior head, midpiece and tail regions [32-34]. Correlatively, the lateral diffusion coefficient D and the mobile fraction M of lipid probes (C16-diI) inserted into the plasma membrane of ejaculated sperm vary from one cell surface region to the other [35]. D and M also differ between testicular and ejacu- lated cells [35] and they vary during cell capacita- tion [36] and sperm maturation [34]. A progres- sive surface regionalization with changes in the diffusibility properties of lipids is observed during spermatogenesis which reflects changes in the plasma membrane organization. This can pre- sumably be related to the membrane protein re- gionalization which can be inferred from ultra- structural studies carried out on mature cells [31-33]. Nevertheless, this large membrane regio- nalization between the head, midpiece and tail in spermatozoa is not due to the existence of protein belts acting as fences against the lateral diffusion of molecules since bleaching experiments carried out on large areas of these various domains indi- cate that the lipid probes are free to exchange between head and midpiece and midpiece and tail [35].
As another example, D and M values mea- sured for lipid probes in the membrane of em- bryos of the mollusc Nassarius reticulatus indicate the existence of an animal-vegetal membrane re- gionalization which increases with the number of cleavage cycles [37]. In addition, lipid D in the polar lobe membrane area shows a cell-cycle de- pendent modulation, with maximum and mini- mum values during the S phase and early mitosis respectively. These observations are in agreement with ultrastructural studies indicating hetero- geneities in the Nassarius egg plasma membrane [37].
Xenopus eggs also exhibit an animal-vegetal plasma membrane polarity in terms of lipid mobility which is strongly enhanced upon fertil- ization [38]. Two distinct macrodomains can now be distinguished: the animal half in which the probe molecules are nearly completely im-
mobilized ( D < < 10 -1° cm2/s, M < < 5 % ) ; the vegetal half in which D and M are only slightly affected with values of 4.4 10 -8 cm2/s and 66%, respectively. The transition between the animal and vegetal domains is sharp and seems to coin- cide with the boundary between the presumptive ecto- and endoderm. In this case, the immobiliza- tion of lipids in the animal plasma membrane might arise from the fusion of cortical granules which are more numerous in this part of the cell [381.
Lipid macrodomains have also been identified in the plasma membrane of red blood cells using fluorescence microscopy coupled with digital image processing [6-8]. No significant lipid do- mains were observed in vesicles made of a total lipid extract thus suggesting that membrane pro- teins are responsible for the formation of these lipid domains.
2.2.2. Lipid microdomains As recently reviewed, a large body of experi-
mental evidence suggests the occurrence of lipid microdomains in biomembranes [4-6,20]. How- ever, only a few techniques can be considered as being really pertinent for detecting lateral hetero- geneities in membranes [4,5,20]. We will restrict our presentation to the FRAP techniques [4,5,20] which provide a macroscopic view of the lateral organization of membranes and to a photo-cross- linking approach we have developed in our laboratory and which provides specific informa- tion on the lateral distribution of lipids at a molecular level [47,55].
FRAP techniques afford an interesting way to study the lateral motion and organization of lipids in membranes [4,5,20]. This method has the ad- vantage of being non-invasive and applicable to individual living intact cells. Two parameters can be evaluated: the lateral diffusion coefficient, D, and the mobile fraction, M, of the probe. If all the labelled lipids are free to diffuse, then M must be equal to 100%. In contrast, and provided that labelling is restricted to the plasma mem- brane (no probe internalization into the intracel- lular membranes), partial fluorescence recovery indicates the existence of microdomains corre- sponding to immobile lipid molecules, at least

J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158 143
within the time scale of FRAP experiments. This was observed to occur in the plasma membrane of many cellular systems [4,20]. More than likely, membrane proteins are responsible for these par- tial immobile fractions since M is close to 100% in protein-free lipid bilayers in the fluid state.
Measuring D and M for lipids and proteins in membranes while varying the radius r i of the illuminated area, can provide useful information on the membrane lateral organization at a macroscopic [39] and a microscopic [40] scale.
In cases when the radius r i and R of the illuminated and diffusion areas tend to be similar ( R / r i < 2, conditions of a finite reservoir), an 'apparent' immobile fraction can be observed and we have proposed a numerical approach enabling D, M and the ratio R / r i to be calculated [39]. As shown for the apical/basolateral plasma mem- branes of vascular endothelial cells [22], this ap- proach enables membranes to be described in terms of macrodomains.
In cases when R / r i > 5 (condition of an infinite reservoir), changes in r i can reveal the existence of lipid microdomains. Thus, for the probe NBD-PC in the plasma membrane of hu- man fibroblasts, increasing r i resulted in an in- crease in D and a decrease in M. This was interpreted as reflecting the occurrence of pro- tein-rich domains about 1 /xm in diameter char- acterized by a low lipid diffusion coefficient, em- bedded in a relatively protein-poor and more mobile lipid continuum [40]. These changes in D and M were an intrinsic property of the plasma membrane and not an artifact of the measuring system since the effects were not seen if diffusion of NBD-PC was measured in liposomes of egg-PC. An increase in D and a decrease in M with increasing beam radius were also observed in the outer bilayer of the envelope of the parasite Schistosoma mansoni, suggesting the existence of /zm-scale lipid microdomains in this membrane [411.
Recently, abnormally low diffusion coefficients and large immobile fractions have been reported for the two lipid probes Cls-diI and R-PE in the plasma membrane of spheroplasts of the yeast Saccharomyces cerevisiae [42]. Removal of the sur- face proteins by cell trypsinization revealed a
considerable increase in D and the appearance of a large mobile fraction of 96%. Lipid immobility was interpreted as being due to the existence of sub-micron size microdomains in the form of a cage of proteins configured in arrays that trap the lipids within [42].
As for any probe approach, the validity of the FRAP techniques relies on the ability of the lipophilic probes to report correctly on the host membrane lipids. A rather large variety of probe molecules has been used and, irrespective of their molecular structures, all share the characteristic of being extrinsic probes which are supposed to randomly distribute within the constitutive mem- brane lipids. This is a crucial point which is not necessarily met. These probes can label different parts of the membranes depending on both their chemical structure and the physical state (gel/liquid) of the host lipids [4,20]. Furthermore, extrinsic probes can be excluded from given parts of the membrane with, as a consequence, a loss of information. To circumvent this drawback, a very attractive and promising way is to work with con- stitutive lipids after the fluorophore has been introduced into the membrane lipids through the regular lipid metabolic pathway. Such an ap- proach has been developed in our laboratory, using anthracene-labelled lipids [43-47].
Anthracene was selected as fluorophore be- cause of the very interesting fluorescence and photochemical properties it displays. Under il- lumination at 360 nm, this group fluoresces and can also form 9-9', 10-10' covalently bound dimers which are not fluorescent. Thus, after incorpora- tion into membrane lipids, anthracene can be used (i) as an indicator of membrane fluidity [48-50] and micropolarity [50-52], (ii) to measure the lateral diffusion rate of the labelled molecules using fluorescence recovery after photobleaching (photodimerization) techniques [53,54] and (iii) to study the topological distribution of lipids in membranes by identification of the photodimers formed between adjacent anthracene-labelled molecules [47,55].
8-(2-Anthroyl)-octanoic acid (ANoc) [50] and 9-(2-anthryl)-nonanoic acid (ANno) [48] and cor- responding phospholipids (Fig. 1) have been syn- thesized and characterized for their physical
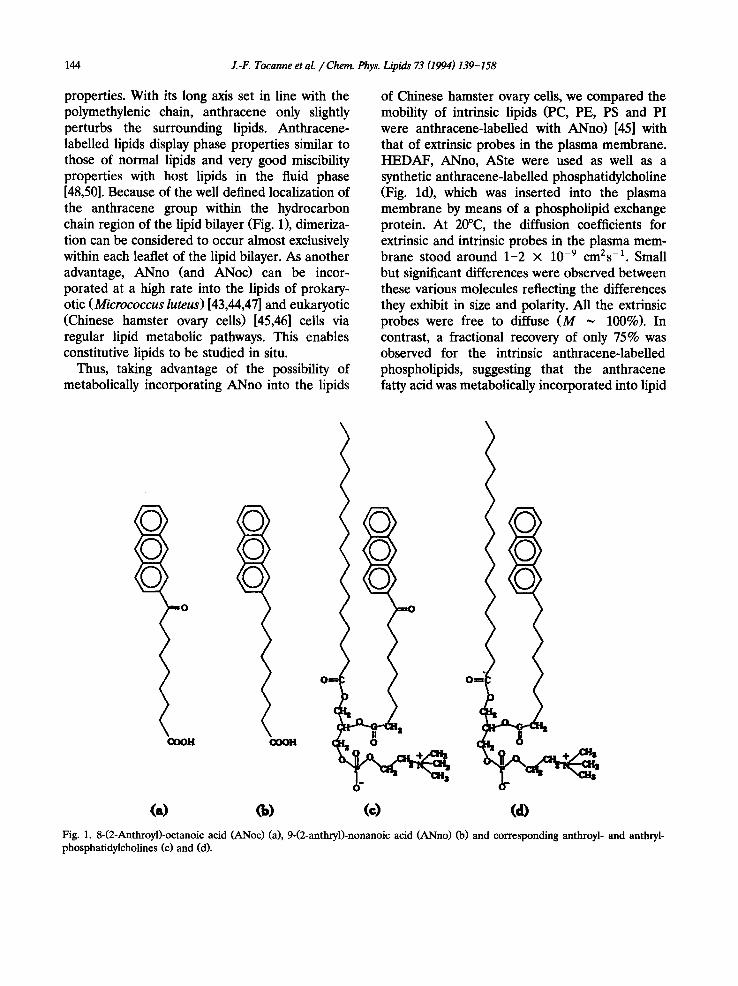
144 J.-F. Tocanne et al. / Cherr~ Phys. Lipids 73 (1994) 139-158
properties. With its long axis set in line with the polymethylenic chain, anthracene only slightly perturbs the surrounding lipids. Anthracene- labelled lipids display phase properties similar to those of normal lipids and very good miscibility properties with host lipids in the fluid phase [48,50]. Because of the well defined localization of the anthracene group within the hydrocarbon chain region of the lipid bilayer (Fig. 1), dimeriza- tion can be considered to occur almost exclusively within each leaflet of the lipid bilayer. As another advantage, ANno (and ANoc) can be incor- porated at a high rate into the lipids of prokary- otic (Micrococcus luteus) [43,44,47] and eukaryotic (Chinese hamster ovary cells) [45,46] cells via regular lipid metabolic pathways. This enables constitutive lipids to be studied in situ.
Thus, taking advantage of the possibility of metabolically incorporating ANno into the lipids
of Chinese hamster ovary cells, we compared the mobility of intrinsic lipids (PC, PE, PS and PI were anthracene-labelled with ANno) [45] with that of extrinsic probes in the plasma membrane. HEDAF, ANno, ASte were used as well as a synthetic anthracene-labelled phosphatidylcholine (Fig. ld), which was inserted into the plasma membrane by means of a phospholipid exchange protein. At 20°C, the diffusion coefficients for extrinsic and intrinsic probes in the plasma mem- brane stood around 1-2 × 10 -9 cnl2s -1. Small but significant differences were observed between these various molecules reflecting the differences they exhibit in size and polarity. All the extrinsic probes were free to diffuse (M ~ 100%). In contrast, a fractional recovery of only 75% was observed for the intrinsic anthracene-labelled phospholipids, suggesting that the anthracene fatty acid was metabolically incorporated into lipid
0
I:~0011 COOH
o I o o~ /
Fig. 1. 8-(2-Anthroyl)-octanoic acid (ANoc) (a), 9-(2-anthryl)-nonanoic acid (ANno) (b) and corresponding anthroyl- and anthryl- phosphatidylcholines (c) and (d).
J.-F. Tocanne et at / Chem. Phys. Lipids 73 (1994) 139-158 145
domains which were inaccessible to the extrinsic probes [46].
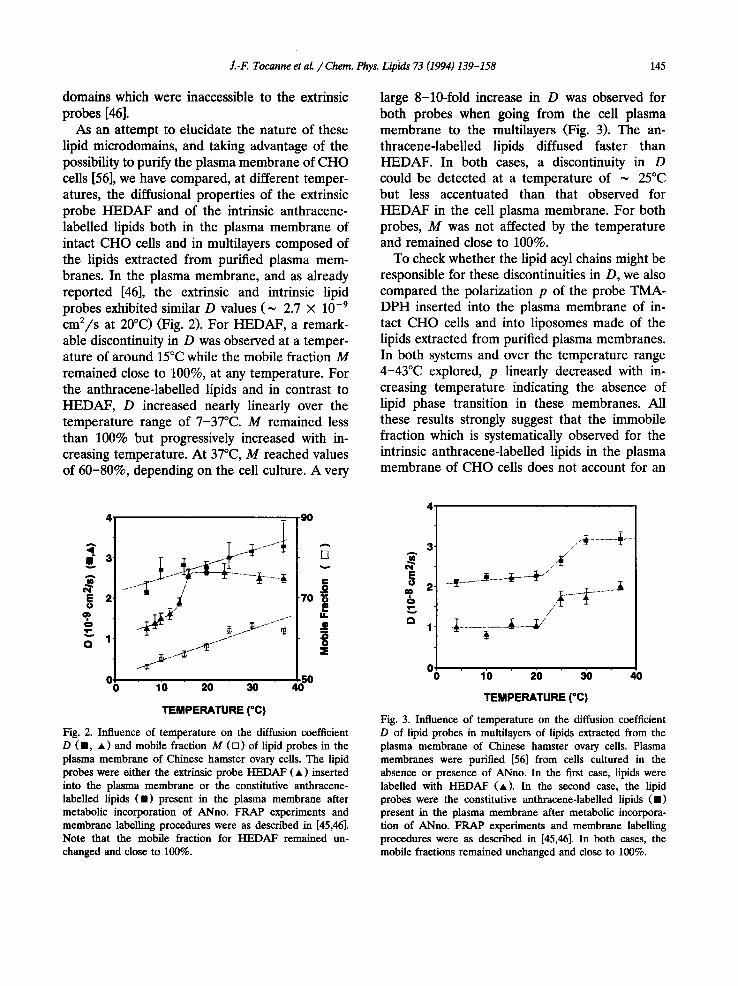
As an attempt to elucidate the nature of these lipid microdomains, and taking advantage of the possibility to purify the plasma membrane of CHO cells [56], we have compared, at different temper- atures, the diffusional properties of the extrinsic probe HEDAF and of the intrinsic anthracene- labelled lipids both in the plasma membrane of intact CHO cells and in multilayers composed of the lipids extracted from purified plasma mem- branes. In the plasma membrane, and as already reported [46], the extrinsic and intrinsic lipid probes exhibited similar D values (~ 2.7 × 10 -9
cm2/s at 20°C) (Fig. 2). For HEDAF, a remark- able discontinuity in D was observed at a temper- ature of around 15°C while the mobile fraction M remained close to 100%, at any temperature. For the anthracene-labelled lipids and in contrast to HEDAF, D increased nearly linearly over the temperature range of 7-37°C. M remained less than 100% but progressively increased with in- creasing temperature. At 37°C, M reached values of 60-80%, depending on the cell culture. A very
large 8-10-fold increase in D was observed for both probes when going from the cell plasma membrane to the multilayers (Fig. 3). The an- thracene-labelled lipids diffused faster than HEDAF. In both cases, a discontinuity in D could be detected at a temperature of ~ 25°C but less accentuated than that observed for HEDAF in the cell plasma membrane. For both probes, M was not affected by the temperature and remained close to 100%.
To check whether the lipid acyl chains might be responsible for these discontinuities in D, we also compared the polarization p of the probe TMA- DPH inserted into the plasma membrane of in- tact CHO cells and into liposomes made of the lipids extracted from purified plasma membranes. In both systems and over the temperature range 4-43°C explored, p linearly decreased with in- creasing temperature indicating the absence of lipid phase transition in these membranes. All these results strongly suggest that the immobile fraction which is systematically observed for the intrinsic anthracene-labelled lipids in the plasma membrane of CHO cells does not account for an
3
2 U
...~rj~.....~ -"'~I'I-" 10 20 30
TEMPERATURE {°C)
.90
5O
Fig. 2. Influence of temperature on the diffusion coefficient D ( • , • ) and mobile fraction M ([]) of lipid probes in the plasma membrane of Chinese hamster ovary ceils. The lipid probes were either the extrinsic probe HEDAF ( • ) inserted into the plasma membrane or the constitutive anthracene- labelled lipids ( • ) present in the plasma membrane after metabolic incorporation of ANno. FRAP experiments and membrane labelling procedures were as described in [45,46]. Note that the mobile fraction for HEDAF remained un- changed and close to 100%.
3 A
E 2
O
r~ 1
.." /
o 1~o 20 3o 40
TEMPERATURE (°C)
Fig. 3. Influence of temperature on the diffusion coefficient D of lipid probes in multilayers of lipids extracted from the plasma membrane of Chinese hamster ovary cells. Plasma membranes were purified [56] from cells cultured in the absence or presence of ANno. In the first case, lipids were labelled with HEDAF (A). In the second case, the lipid probes were the constitutive anthracene-labelled lipids ( • ) present in the plasma membrane after metabolic incorpora- tion of ANno. FRAP experiments and membrane labelling procedures were as described in [45,46]. In both cases, the mobile fractions remained unchanged and close to 100%.

146 J.-F. Tocanne et al. /Chem. Phys. Lipids 73 (1994) 139-158
intrinsic property of these lipids but originates rather from the presence of the membrane pro- teins. These observations reinforce our first con- clusion that the lipid microdomains so detected might correspond to the existence of a protein lattice trapping lipid molecules and acting like a fence preventing lipids from laterally diffusing [46]. These microdomalns would be accessible, not to the extrinsic probes, but only to the intrin- sic ones, in the course of membrane biogenesis.
To investigate the lateral distribution of lipids in membranes and still using the photochemical properties of anthracene derivatives, we have de- vised a new photo-cross-linking method [47,55]. It is based on the possibility of identifying and counting quantitatively the various anthracene- lipid dimers which can form upon illumination of membranes in which the lipids have been metabolically anthracene-labelled. This method, which has the advantage of being non-invasive and applicable to living intact cells, gives informa- tion on the lateral distribution of the various lipid molecular species present in a membrane [47,55]. Anthracene has a short lifetime of about 4 ns [49]. In other words, since the dimerization reac- tion proceeds on a nanosecond time scale while lipids exchange lattice positions only every 100 ns or so, photodimerization corresponds to an inte- gration in time of a series of instantaneous pic- tures of the lipid distribution within the mem- brane. If lipid domains existed in a membrane with functional significance, they would present a certain character of permanency and would be detected with our method. Accordingly, in the membrane of the bacterium Micrococcus luteus, this approach clearly demonstrated that PG and DMDG, the two major lipids present in the bac- terial membrane (80% of total lipids), were not homogeneously distributed in the membrane plane [47]. Indeed, the ratio of PG-DMDG het- ero-dimer to the PG-PG and DMDG-DMDG homo-dimers was found to be around half of that expected for an homogeneous mixture of the two lipids. The possibility for this to arise from an asymmetric distribution of the two lipids in the inner and outer leaflets could be discarded since PG and DMDG were found to be evenly dis-
tributed between the two membrane leaflets [47]. This non-random lateral distribution of the two anthracene-labelled lipids in the bacterial mem- brane is to be linked to the presence of the membrane proteins and does not correspond to a property of the lipid phase itself. Indeed, natural PG and DMDG (and anthracene-labelled molecules) are mainly substituted by methyl- branched tetra- and hexa-decanoic acids and re- main in a fluid phase over a wide temperature range. In addition, the photodimerization ap- proach revealed a random lateral distribution of PG and DMDG in mixed liposomes [47], in agree- ment with previous studies demonstrating very good miscilbility properties for the two lipids [57].
3. Influence of proteins on lipid D in membranes
As already mentioned in the Introduction, most membrane proteins are not free to diffuse, pre- sumably due to their attachment to cytoskeleton components, and de facto, act as many obstacles to the diffusion of lipids. Such an influence of membrane proteins on the lateral mobility of lipids can be inferred from the comparison of the diffusion coefficients measured for lipids in natu- ral membranes and in multilayers composed of the corresponding membrane lipid extracts. Thus, from the data obtained from erythrocytes [58,59], human fibroblastes [60] and CHO cells (Figs. 2 and 3 and Table 1), the lateral diffusion rate of lipids is three- to ten-times faster in protein-free lipid bilayer than in the corresponding natural membrane. As shown by Cribier et al. [59] for the membrane of human erythrocytes, these differ- ences concern both the outer and inner leaflets of the membrane lipid bilayer. Influence of proteins on the lateral motion of lipids in membranes is also supported by the FRAP data reported above (see Section 2.2.2) for the yeast Saccharomyces cerevisiae [42] and rat proximal tubule epithelial cells [26] and by the fact that the lipid immobile fractions which are very often detected in natural membranes are no longer observed in the corre- sponding protein-free lipid bilayers.
Influence of membrane proteins was also de- monstrated in a very elegant way by Chazotte et
Z-F. Tocanne et al. /Chem. Phys. Lipids 73 (1994) 139-158 147
Table 1 Diffusion coefficients of lipophilic probes in the plasma membrane of different cells and in multilayers obtained from the corresponding lipid extracts
Cellular system Probe Temperature (°C) D (10 -s em2/s) D Ref.
Plasma Lipid membrane multilayers
Human fibroblasts
Human erythrocytes
Human erythrocytes
inner leaflet outer leaflet Chinese hamster
ovary cells
C16-diI 37 2 6 60
NBD-PE 37 0.2 0.85 58
NBD-PC 59
Metabolically anthracene-labelled lipids
37 0.8 1.8 37 0.14 0.35 20 0.27 2
37 0.32 3.2 this paper
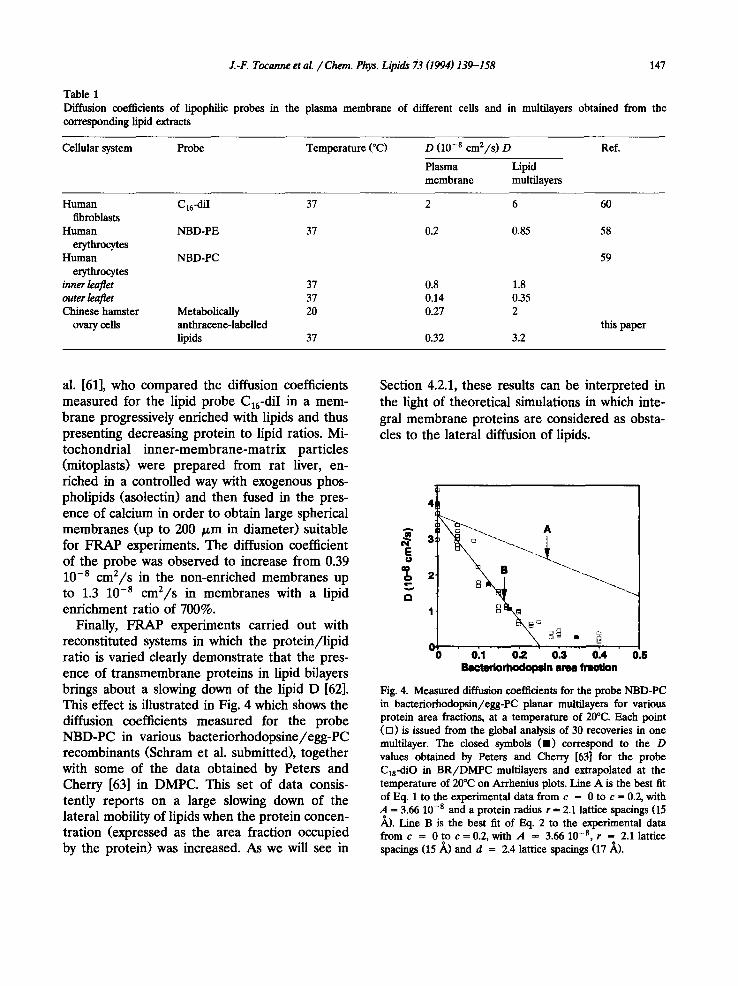
al. [61], who compared the diffusion coefficients measured for the lipid probe C16-diI in a mem- brane progressively enriched with lipids and thus presenting decreasing protein to lipid ratios. Mi- tochondrial inner-membrane-matrix particles (mitoplasts) were prepared from rat liver, en- riched in a controlled way with exogenous phos- pholipids (asolectin) and then fused in the pres- ence of calcium in order to obtain large spherical membranes (up to 200 /~m in diameter) suitable for FRAP experiments. The diffusion coefficient of the probe was observed to increase from 0.39 10 .8 cm2/s in the non-enriched membranes up to 1.3 10 .8 c m 2 / s in membranes with a lipid enrichment ratio of 700%.
Finally, FRAP experiments carried out with reconstituted systems in which the protein/lipid ratio is varied clearly demonstrate that the pres- ence of transmembrane proteins in lipid bilayers brings about a slowing down of the lipid D [62]. This effect is illustrated in Fig. 4 which shows the diffusion coefficients measured for the probe NBD-PC in various bacteriorhodopsine/egg-PC recombinants (Schram et al. submitted), together with some of the data obtained by Peters and Cherry [63] in DMPC. This set of data consis- tently reports on a large slowing down of the lateral mobility of lipids when the protein concen- tration (expressed as the area fraction occupied by the protein) was increased. As we will see in
Section 4.2.1, these results can be interpreted in the light of theoretical simulations in which inte- gral membrane proteins are considered as obsta- cles to the lateral diffusion of lipids.
c~
4
, ' o21 ' o'~ o23 ' o24 ' o.s I ~ t m ' i o r l ~ l o ~ n area fmotlon
Fig. 4. Measured diffusion coefficients for the probe NBD-PC in bacteriorhodopsin/egg-PC planar multilayers for various protein area fractions, at a temperature of 20°C. Each point ([]) is issued from the global analysis of 30 recoveries in one multilayer. The closed symbols (11) correspond to the D values obtained by Peters and Cherry [63] for the probe Cls-diO in BR/DMPC multilayers and extrapolated at the temperature of 20°C on Arrhenius plots. Line A is the best fit of Eq. 1 to the experimental data from c = 0 to c = 0.2, with A ffi 3.66 10 -s and a protein radius r ffi 2.1 lattice spacings (15 ~,). Line B is the best fit of Eq. 2 to the experimental data f romc = 0 t o c = 0 . 2 , with A = 3.6610 - s , r = 2.1 lattice spacings (15/~) and d = 2.4 lattice spacings (17/~).

148 Z-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158
4. Protein-lipid interactions
4.1. Specific interactions
Specific interactions between lipids and pro- teins have long been recognized to occur in mem- brane model systems in which peripheral and integral membrane proteins were recombined with a variety of lipids. It is out of the scope of this article to present a detailed review of the avail- able data which was obtained using a wide variety of approaches (NMR, ESR, fluorescence, protein purification, binding measurements....). Well documented review articles exist on this topic and the interested reader is to be referred to [6,64-71]. With regard to membrane lateral organization, the most interesting points which concern the specificity, the stoichiometry and the time scale of these interactions and which were obtained prin- cipally by means of ESR experiments, can be summarized as follows: i. The stoiehiometry of lipid association with a number of different integral membrane proteins has been determined and the motionally re- stricted lipid component observed by ESR spec- troscopy may be identified with the first layer of lipid molecules in contact with the intramembra- nous hydrophobic surface of the protein. This conclusion stems from the observation that, with a few exceptions, the stoichiometries scale with the square root of the protein molecular weight, as would be expected for proteins of roughly cylindrical cross-section which protrude to equiva- lent extents from the membrane surface. The structure of most of the proteins tested is still unknown, at least at a high degree of resolution. However, the numbers of motionally restricted lipids found associated with these proteins agree reasonably well with the number of lipids which would form the first shell around the intramem- branous perimeter of these proteins, and esti- mated on the basis of available dimensional data [66,68]. ii. A rather large variety of both peripheral and integral membrane proteins exhibit selectivity for different phospholipid headgroups. Results are expressed in the form of the ratio of the average relative association constant K~ for a particular
spin-labelled lipid to that for spin-labelled phos- phatidylcholine [66]. The selectivity patterns re- ported differ from one protein to another both in intensity and with respect to the relative order of the various lipids tested. Thus, a Kay of 5-6 (relative to phosphatidylcholine) was observed for cardiolipin interacting with cytochrome c oxydase [70,72]. In contrast, a Kay value < 2 was mea- sured for cardiolipin interacting with Ca 2÷- ATPase while rhodopsin exhibited no selectivity for lipids [66]. In general, preferential selectivities are found for acidic lipids, especially cardiolipin, phosphatidic acid and free fatty acids, depending on the particular protein. The more specific inter- actions are observed with peripheral proteins, in particular /3-hydroxybutyrate dehydrogenase and protein kinase c which exhibit high specificity for phosphatidylcholine and phosphatidylserine, re- spectively [71].
It should be stressed that these average relative association constants Kay can be misleading since it is not known whether the lipid specificity arises from a uniform increase in association constant for all the sites or is due to a much higher specificity of just a few sites [66]. Kay values of 5-10 correspond to relatively weak specificity and are extremely low as compared to the binding constants usually found for a ligand interacting with its protein (105 to 107 M -1) [67]. However and as previously discussed by Griffith et al. [64], conditions in a membrane are quite different from those in solution, due to reduction in dimen- sion and to the increase in the concentration of the interacting species in the highly confined re- gion of space a membrane is. Thus, to achieve about 90% occupancy of one specific lipid binding site on a protein embedded in a bilayer contain- ing 10 mol % of a specific lipid requires a relative binding constant of only about 100. A K~ value of 5 averaged over the 40-60 lipid molecules currently found in the first shell around mem- brane proteins [66] might correspond to a value of 100 if only a few sites are concerned by a highly specific protein-lipid interaction. iii. The perturbation of the lipids by the protein is of short range and practically concerns only the first shell of lipids around the protein.

J.-F.. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158 149
iv. In most cases, lipids in contact with the protein exchange rapidly with the others. Off-rate con- stants stand around 2 × 107 S- 1 [65-67,70] which is comparable to the frequency of jump for lipids diffusing freely (D = 10 -8 cm2/s) in a fluid bilayer. However, in a few cases, the relative exchange rates of lipids can be found to follow the relative selectivities. This is the case for myelin proteolipid apoprotein which displays selectivity for phosphatidic acid relative to phosphatidyl- choline. In myelin proteolipid/dimyristoyl- phosphatidylcholine recombinants, off-rate con- stants of 4.5 × 106 S -1 and 1.6 × 107 S -1 were measured for spin-labelled phosphatidic acid and phosphatidylcholine respectively, indicating a longer residence time near the protein for the former than for the latter, in agreement with the selectivity order.
In summary, one can be confident about the fact that some peripheral and integral membrane proteins exhibit selectivity for lipid headgroups. Nevertheless, there is no general model to de- scribe these interactions; different proteins will show different lipid selectivities, stoichiometries and strength of association [68]. These interac- tions are weak and in the case of integral pro- teins, concern only the first layer of lipids around the protein.
4.2. Non specific interactions
4.2.1. Long-range lipid/protein interactions in the absence of hydrophobic mismatch
Interactions between lipids and integral pro- teins have been the object of many investigations which have been extensively described in some recent review articles [67,69,73]. It is now well recognized that lipids in contact with the tran- smembrane hydrophobic segments of integral membrane proteins are partly immobilized and that this can extend beyond the first layer of the lipid shell [69,73]. It should be emphasized that such an effect is general in nature and therefore an important question arises as to whether these partially immobilized lipids can constitute lipid microdomains in a membrane and whether they can be detected as such. These two questions are intimately related and, as illustrated by the dif-
ferent views fluorescence, ESR and NMR spec- troscopies give on the way intrinsic proteins per- turb the lipid dynamics, the answer clearly de- pends on the time scale of the method used for analysing membrane organization [69,73-75]. From comparison of ESR and NMR data, and as discussed in the above section, these interactions do not yield permanent structures. They are dy- namic and in many systems, the perturbed and unperturbed lipid molecules exchange at a rate close to that of the lateral diffusion of lipids in a fluid phase [67,69,73-75]. In these conditions, the next problem is to know whether FRAP experi- ments, which relate the lateral diffusion and dis- tribution of lipids in membranes, can reflect the presence of these partially immobilized lipids. As shown in Section 3, intrinsic membrane proteins bring about a considerable slowing down in the lateral diffusion rate of lipids. This can be under- stood if one considers the relative size of lipids and proteins and the fact that many membrane proteins are immobilized due to interactions with cytoskeleton elements. This slowing down is also well accounted for by theoretical models devel- oped by Saxton [76-79], Eisinger et al. [80], Almeida et al. [81] and quite recently in our laboratory (Schram et al., submitted), which all describe the diffusion of particles in the presence of obstacles. As reviewed in [20], these models are based on the free volume theory of diffusion [82,83] which seems to be the most appropriate for describing the lateral motions of lipids in membranes [84,85]. In short, it is necessary to create a void large enough for diffusive lipid displacement to occur, implying that the vacant site for the translational diffusion and the lipid tracer have similar structural parameters. In the computer simulations of diffusion [76-79,80] (Schram et al., submitted), a lipid leaflet assembly is represented by a triangular lattice of points arranged in the form of a square matrix. Like- wise, to simulate complex assemblies of lipid and proteins molecules, each element of the matrix can be randomly occupied either by an immobile or less mobile point obstacle (protein), or by a diffusing particle (lipids or tracers). In Saxton's approach [76-79], D obtained after random walks on the lattice was calculated by the computation

150 J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158
of the mean square displacement. Eisinger et al. [80] proposed another approach which evaluated D by a double travelling, involving the probe and the simulated 'monomeric non-excited probe'. These two approaches do not consider the mobile fraction of lipids and for that reason, we have developed a computer simulation of FRAP exper- iments (Schram et al., submitted). A membrane proteolipid leaflet is represented again by lipid sites arranged into triangular lattice, in which each matrix site can be occupied randomly either by an immobile point obstacle (the proteins), or by a diffusing particle (the lipid tracers). In order to simulate FRAP experiments, an area of obser- vation is introduced at the centre of the matrix. The initial fluorescence intensity is first evaluated by averaging the number of tracers found in the observation area. After, and in order to mimic a 'bleaching' step in the condition of 'uniform disk illumination', all the tracers present in the obser- vation area are deleted. During the 'recovery' step, the fluorescence intensity at time t after the bleaching procedure is estimated by counting the number of tracers present in the observation area. In this new approach, it is now possible to analyse the effect of the size and fractional area of im- mobile obstacles on the diffusion coefficient D but also on the mobile fraction M of the diffusing particles. Despite differences in their conceptual formulations, these various computational models give identical results as to the changes in D for various obstacle area fractions. Clearly, the greater the obstacle area fraction, the lower D is, and for a given obstacle area fraction, the smaller the obstacle size, the greater its influence on the translational diffusion coefficient of the mobile species. Furthermore, large decreases of D [77,78,80,81] (Schram et al., submitted) and of M (Schram et al., submitted) are noted when the obstacle area fraction reaches a value of 50%, which corresponds to the value of a percolation threshold noted by Stauffer [86] for a triangular two dimensional lattice.
From the simulations of FRAP experiments, a phenomenological equation has been determined which relates the diffusion coefficient D of the mobile particles to the radius r and area fraction
c of the obstacles:
D =A {1 - [1.574 - 0.475. ln(r)] .c} (1)
If one considers the proteins as being surrounded by a shell of immobilized lipids of thickness d, then this equation takes the form:
r
/ 1 - [1.574 - 0.475. ln(r + d)] D ~ a "
L
(r + d) 2 1 "¢prot" r------T'--- (2)
d
In both equations, A is a constant which charac- terizes the numerical translation between simu- lated and measured D coefficients. This approach has been applied to the analysis of the diffusion coefficients measured experimentally by a FRAP technique for the probe NBD-PC incorporated in bacteriorhodopsin/egg-PC multilayers, at a tem- perature of 20°C and for different protein-to-lipid ratios (Schram et al., submitted). As can be seen in Fig. 4, the diffusion coefficient of the probe decreased by a factor of 10 when bacteriorho- dopsin area fractions varied from 0 to 0.3, above which value D remained nearly constant. In the full protein concentration range explored, the mobile fraction did not vary and was close to 100% (not shown). The diffusion coefficients measured by Peters and Cherry [63] for the probe C18-diO in B R / D M P C multilayers and extrap- olated in Arrhenius plots at 20°C are in excellent agreement with our own values (Fig. 4, full squares). This suggests that these diffusion coef- ficients (obtained for different experimental con- ditions of fluorescent probe structure, reconstitu- tion procedure and FRAP technique) can be con- sidered as characteristic of the system studied.
Fitting Eq. 1 to the experimental data in Fig. 4, with a radius of 15 A for the protein (in egg-PC bilayers, BR can be assumed to stay in the monomeric form up to an area fraction of about 0.3 [87,88]) and a molecular area of 50 ~2 for egg-PC, lead to line A which strongly deviated from the measured diffusion coefficients. In agreement with the arguments presented in Sec- tion 4.1, one possibility for fitting the equation to the data in Fig. 4 was to consider that the effec-

J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158 151
tive size of the obstacles was greater than that normally expected from the structure of monomeric bacteriorhodopsin or in other words, to consider that the protein was surrounded by a few layers of partially immobilized lipids. From the point of view of translational diffusion, the existence of such an annular domain of partly rigidified lipids would further decrease the over-all translational diffusion. Eq. 2 was used and good fitting to the experimental data (line B in Fig. 4) was achieved with a d value of 17 + 3 ~,, which corresponds to about two layers of partly im- mobilized lipids around the protein. This d value of 17 .~ is not the coherence length of the actual lipid annular domain surrounding each bacteri- orhodopsin molecule. It must simply be con- sidered as an estimation of the distance over which the influence of the protein extends in the lipid phase when viewed by the FRAP method- ology. In this respect, Almeida and co-workers [81] have recently proposed a theoretical treat- ment of the effects of immobile and rigid circular obstacles on translational diffusion, which is based on the free-volume theory of diffusion and which takes into account long-range lipid/protein inter- actions and their exponential decrease away from the protein surface. By appreciating the diffusion coefficient from a two-dimensional integration of the relative free area around randomly dis- tributed obstacles, an analytical expression relat- ing the relative diffusion coefficient < D > to the obstacle size and area fractions was obtained [81]:
< D > -- 1 +ac +bc2 (3)
with
a = - 1.208 - 24.3 exp( - 1.763R/~ )
- 2.408 e x p ( - 0.3138R/~ )
b = 185 exp(2.587R/~)
and in which R is the radius of the solid-phase obstacle, ~ is the coherence length and c is the obstacle area fraction. Best fitting of Eq. 3 to the data in Fig. 4, over the protein concentration ran.ge 0-0.2, leads to a coherence length g = 11 4 A. This value is compatible with the 17 + 3 A protein radius increase detected with our ap-
proach. These values are consistent with the coherence length proposed to account for long- range protein/lipid interactions and which has been estimated at around 15 ~, [89]. As we will see in the next section, they are also consistent with those inferred from the analysis of phase transition data obtained for lipids perturbed by the reaction center proteins of Rhodopseudomo- nas spaheroides [90,91] and bacteriorhodopsin from Halobacterium halobium [92,93].
It is clear from this analysis that the diffusion coefficient of lipids in membranes integrate not only the presence of transmembrane proteins as obstacles to diffusion but also the presence of an annular domain of partly immobilized lipids sur- rounding the protein. The above results indicate that this kind of lipid microdomain is susceptible to detection in FRAP experiments provided a quantitative analysis of the diffusion data can be achieved. This also demonstrates that the detec- tion of lipid microdomains is not necessarily to be linked to the detection of a lipid immobile frac- tion. This last point will be discussed in the final section together with the problem of the time- scale of the method used for detecting lipid do- mains in membranes.
4. 2. 2. Long-range lipid ~protein interactions in con- ditions of hydrophobic mismatch
As already mentioned, membranes contain a large variety of transmembrane proteins and of lipids with different acyl chain lengths. Therefore, and in addition to the interactions described above, a very attractive possibility is to consider the consequences of a mismatch between the hydrophobic thicknesses of transmembrane pro- teins and the surrounding lipid bilayer. Intro- duced first by Mouritsen and Bloom [94], this concept of hydrophobic matching has received much theoretical attention and its basic state- ments and consequences on lipid organization are extensively described in many papers [94-98] and review articles [73,99-101].
The consequences of the hydrophobic matching condition on the acyl chain order of lipids in a bilayer of hydrophobic thickness d L, supporting a protein with a hydrophobic length dp are shown

152 J.-F. Tocanne et al. /Chem. Phys. Lipids 73 (1994) 139-158
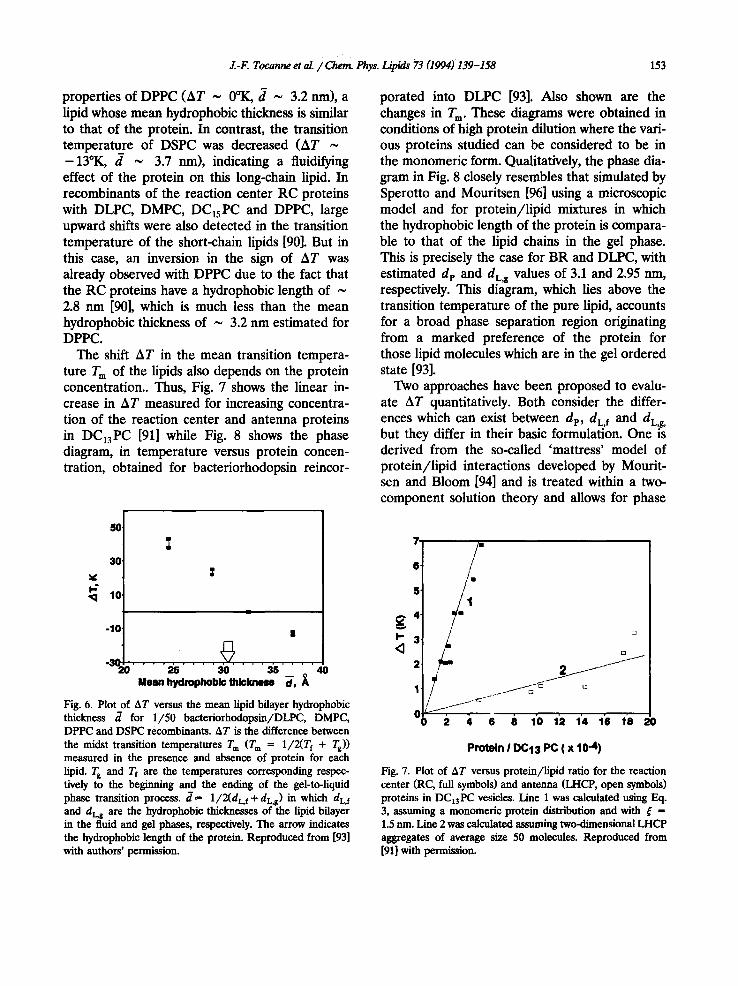
in Fig. 5. Lipids in contact with that protein would be expected to be rigidified (Fig. 5A) or fluiditied (Fig. 5B) depending on whether d L is respectively smaller or larger than dp. In both cases, the perturbation would extend over a few lipid layers around the protein in order either to taper or to enlarge the bilayer to its typical unperturbed thickness [96]. The decay of the lipid bilayer hy- drophobic thickness, which is exponential in na- ture, is characterized by a coherence length [97]. It has long been recognized that hydrophobic mismatch generates thermodynamic forces which might be responsible for lateral segregation of proteins in membranes [94,102]. Conversely, and in absence of protein segregation, hydrophobic mismatch might also contribute to the formation of lipid microdomains around intrinsic membrane proteins with fluidity different to that of the bulk of the lipid phase [96,100]. These local perturba- tions in the lipid acyl chain order would result in an overall upward or downward shift in the phase transition temperature of the host lipids, a ther- modynamic prediction which is easily accessible to experimental verification by studying the phase behaviour of protein/lipid recombinants.
Today, only the reaction center (RC) and the light harvesting antenna (LHCP), two transmem- brane proteins f rom Rhodopseudomonas Sphaeroides [90,91] and bacteriorhodopsin from Halobacterium halobium [93], have been reconsti- tuted into vesicles of phosphatidylcholines with different chain lengths and shown to modify the gel-to-liquid phase transition of the lipid bilayer. Both the reaction center proteins and bacteri- orhodopsin produced positive or negative shifts in the phase transition temperature of the host lipids depending on the strength and sign of the mis- match between the mean hydrophobic thickness d of the lipid bilayer and the protein hydrophobic length dp. Data obtained with BR [93], in the region of high protein concentration (BR/PC molar ratios ~ 1/50), are shown in Fig. 6. De- spite the presence of the endogenous lipids, bac- teriorhodopsin ( d p ~ 3.0-3.1 nm) brought about a large upward shift in the phase transition tem- perature of DLPC (AT ~ 40°K, tl ~ 2.4 nm), and to a lesser extent of DMPC (AT ~ 23°K, d ~ 2.8 nm), accounting for a strong rigidifying effect of the protein on these short-chain lipids. Bacteriorhodopsin had no influence on the phase
A
Fig. 5. Schematic representation of an intrinsic membrane protein of hydrophobic length dp embedded in a lipid bilayer in which the unperturbed'hydrophobic thickness d L is smaller (A) or larger (B) than dp. The influence of the protein, characterized by the coherence length ~ extends over a certain distance from the protein surface and progressively vanishes so that the bilayer recovers its t~pical unperturbed thickness d L.
J..-F. Tocanne et al. /Chem. Phys. Lipids 73 (1994) 139"158 153
properties of DPPC (AT ~ 0°K, d ~ 3.2 nm), a lipid whose mean hydrophobic thickness is similar to that of the protein. In contrast, the transition temperature of DSPC was decreased (AT ~ -13°K, d ~ 3.7 nm), indicating a fluidifying effect of the protein on this long-chain lipid. In recombinants of the reaction center RC proteins with DLPC, DMPC, D C 1 5 P C and DPPC, large upward shifts were also detected in the transition temperature of the short-chain lipids [90]. But in this case, an inversion in the sign of AT was already observed with DPPC due to the fact that the RC proteins have a hydrophobic length of ~ 2.8 nm [90], which is much less than the mean hydrophobic thickness of ~ 3.2 nm estimated for DPPC.
The shift AT in the mean transition tempera- ture T m of the lipids also depends on the protein concentration.. Thus, Fig. 7 shows the linear in- crease in AT measured for increasing concentra- tion of the reaction center and antenna proteins in DC13PC [91] while Fig. 8 shows the phase diagram, in temperature versus protein concen- tration, obtained for bacteriorhodopsin reincor-
porated into DLPC [93]. Also shown are the changes in T m. These diagrams were obtained in conditions of high protein dilution where the vari- ous proteins studied can be considered to be in the monomeric form. Qualitatively, the phase dia- gram in Fig. 8 closely resembles that simulated by Sperotto and Mouritsen [96] using a microscopic model and for protein/lipid mixtures in which the hydrophobic length of the protein is compara- ble to that of the lipid chains in the gel phase. This is precisely the case for BR and DLPC, with estimated dp and dL¢ values of 3.1 and 2.95 nm, respectively. This diagram, which lies above the transition temperature of the pure lipid, accounts for a broad phase separation region originating from a marked preference of the protein for those lipid molecules which are in the gel ordered state [93].
Two approaches have been proposed to evalu- ate AT quantitatively. Both consider the differ- ences which can exist between dp, dL, f a n d dL,g ' but they differ in their basic formulation. One is derived from the so-called 'mattress' model of protein/lipid interactions developed by Mourit- sen and Bloom [94] and is treated within a two- component solution theory and allows for phase
50 ̧
30,
I.- 10'
-10.
-3(
|
25 30 3 5 - - o Moan hydrophoblo thickness d , A
40
Fig. 6. Plot of AT versus the mean lipid bilayer hydrophobic thickness d for 1/50 baeteriorhodopsin/DLPC, DMPC, DPPC and DSPC recombinants. AT is the difference between the midst transition temperatures T m (T m ffi 1/2(Tf + Ts)) measured in the presence and absence of protein for each lipid. T s and Tf are the temperatures corresponding respec- tively to the be~nning and the ending of the gel-to-liquid phase transition process, d = 1/2(dL, f + dL~) in which dL, f and dLg are the hydrophobic thicknesses of the lipid bilayer in the fluid and gel phases, respectively. The arrow indicates the hydrophobic length of the protein. Reproduced from [93] with authors' permission.
7
6"
5'
g4. ~ 3' <1
6 2 4 6 8 10 12 14 16 18 20
Protein I DC13 PC ( x 10 "4)
Fig. 7. Plot of AT versus protein/lipid ratio for the reaction center (RC, full symbols) and antenna (LHCP, open symbols) proteins in DC13PC vesicles. Line 1 was calculated using Eq. 3, assuming a monomerie protein distribution and with ~ = 1.5 nm. Line 2 was calculated assuming two-dimensional LHCP aggregates of average size 50 molecules. Reproduced from [91] with permission.
154 J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158
20. f
15
O° 10
-5
BR/DLPC, tool/tool
Fig. 8. Phase diagrams, in temperature versus protein concen- tration, for recombinants of bacteriorhodopsin with dilau- roylpliospliatidylcholine. The full symbols correspond to the measured Tf and Tg values and the open symbols to the midst transition temperatures T m = 1/2(Tf + Ts). Line T m shows the best fitting of F_x 1. 3 to the experimental data ( ~ = 1.2 rim, r = 0.989). Reproduced from [93] with authors ' permission.
separation. The other [91] is based upon elastic models within the Landau-de Gennes theory, which attribute the full excess free energy to an elastic distorsion of the lipid phase. In the case of protein/lipid hydrophobic mismatch, the main elastic force results from an expansion or a com- pression of the lipid bilayer by the protein in normal direction and changes in the hydrophobic thickness of the lipids in contact with the protein are derived from changes in the order parameter of their acyl chains. Note that phase separation is not allowed and a sharp phase transition remains in the presence of the protein within this descrip- tion [89]. In this approach, it is possible to evalu- ate AT as a function of d e, d and ~, the coher- ence length of the lipid membrane using the following expression [95]:
AT= 16~2[~e/ (1r -~) + 1]
-dL,g)] X e (Eq. 4)
in which ¢ is the characteristic coherence length of the lipid phase, (1)P is the perimeter of the protein and Xp is the protein mole fraction.
Very good agreement between theory and ex-
periments was achieved for the reaction center [91] and bacteriorhodopsin [93] data using coher- ence length values of 1.5 and 1.2 nm, respectively. These values are consistent with that of 1.5 nm predicted by J~ihnig on the grounds of theoretical arguments [89]. In contrast, the antenna protein data could be fitted only after considering protein aggregation within the lipid bilayer [91].
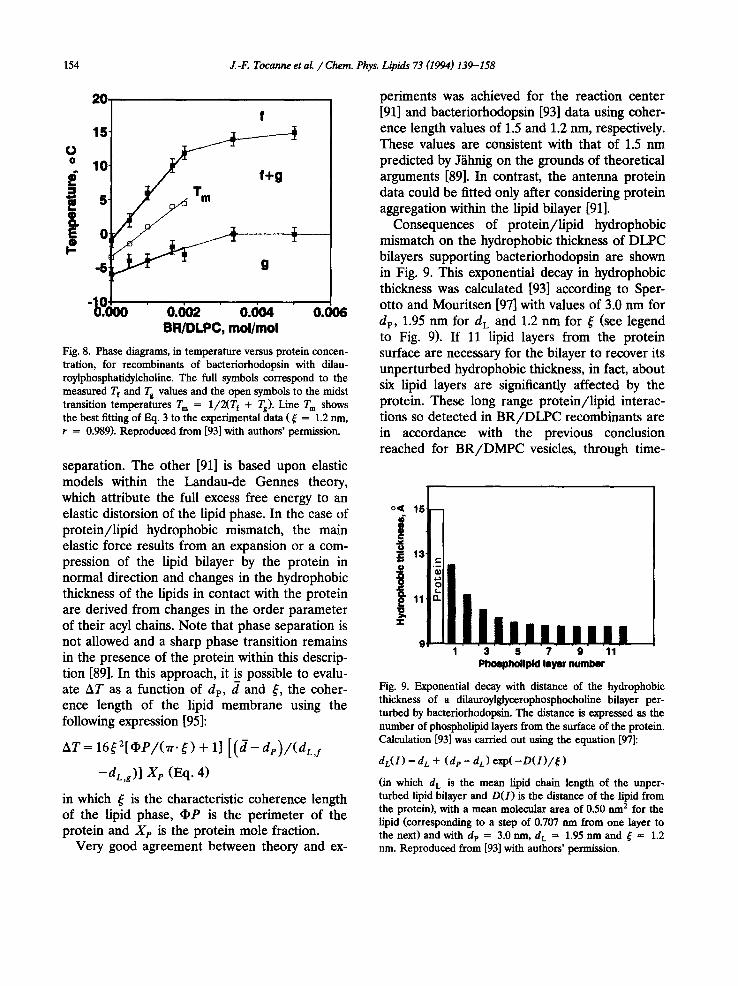
Consequences of protein/lipid hydrophobic mismatch on the hydrophobic thickness of DLPC bilayers supporting bacteriorhodopsin are shown in Fig. 9. This exponential decay in hydrophobic thickness was calculated [93] according to Sper- otto and Mouritsen [97] with values of 3.0 nm for dp, 1.95 nm for d L and 1.2 nm for ~ (see legend to Fig. 9). If 11 lipid layers from the protein surface are necessary for the bilayer to recover its unperturbed hydrophobic thickness, in fact, about six lipid layers are significantly affected by the protein. These long range protein/lipid interac- tions so detected in BR/DLPC recombinants are in accordance with the previous conclusion reached for B R / D M P C vesicles, through time-
o< 16
i /:! r -
~,~11 n
$: iiln,,,,,,,, 1 3 5 7 9 11
Phoephollpld layer number
Fig. 9. Exponential decay with distance of the hydrophobic thickness of a dilauroylglycerophosphocholine bilayer per- turbed by bactedorhodopsin. The distance is expressed as the number of phospholipid layers from the surface of the protein. Calculation [93] was carried out using the equation [97]:
dL(1) = d L + (de - d L) exp( - D ( 1 ) / ~ )
(in which d L is the mean lipid chain length of the unper- turbed lipid bilayer and D(1) is the distance of the lipid from the protein), with a mean molecular area of 0.50 urn2 for the lipid (corresponding to a step of 0.707 nm from one layer to the next) and with dp = 3.0 nm, d L = 1.95 nm and ~ = 1.2 nm. Reproduced from [93] with authors ' permission.

J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158 155
resolved fluorescence depolarization and energy- transfer experiments, that the influence of BR on the order of the lipid phase extends beyond a distance of 4.5 nm from the protein, which corre- sponds to about six to seven lipid layers [92].
To conclude this section, both the experimental results obtained with the reaction center proteins and bacteriorhodopsin and their theoretical inter- pretation strongly support the concept that a transmembrane protein can increase or decrease the chain order of the surrounding lipid phase with an intensity and over a distance which will depend on the sign and strength of the lipid/pro- tein hydrophobic mismatch. In other Words, lipid microdomains can exist around a membrane pro- tein with an acyl chain order different from that of the other lipids. Quite interestingly and as recently shown by Sperotto and Mouritsen [103], the concept of hydrophobic mismatch can also lead to lipid sorting in lipid mixtures with differ- ent acyl chain lengths. Through computer simula- tions based on a microscopic model of lipid/pro- tein interactions in conditions of hydrophobic mismatch, these authors show that lateral hetero- geneities in lipid composition might occur be- cause of selection by the protein of those lipid species whose chain length is such that they will most easily wet its hydrophobic surface [103].
5. Conclusion
It is clear from the set of data presented above that proteins play a predominant role in the lat- eral organization of membranes and therefore in the formation of lipid macro- and microdomains. Many of the results obtained with protein recom- binants in membrane model systems clearly show that, through both short-range or long-range spe- cific and non-specific interactions, proteins can modulate the lateral organization of lipids. When acting in synergy, these various interactions might explain the formation of lipid microdomains in membranes. However, it remains difficult today to provide a general and precise description of the various lipid microdomains which can exist in cell membranes. As a matter of fact, this is due to the great structural and compositional diversity of lipids and proteins in membranes which makes
any investigation at a molecular level extremely difficult to achieve. This is also due to the fact that the detection of these lipid microdomains and their characterization in nature (closed or open domains), composition (lipid and protein species present in the domain) and size (one protein surrounded by a few layers of perturbed lipids or larger domains involving a great number of lipid and protein molecules) depend on the time-scale and nature of the technique used, with many ambiguities in the interpretation of the experimental data still remaining. Typically, short-range specific lipid/protein interactions can be detected via ESR spectrocopy but not by NMR while long-range non-specific lipid/protein inter- actions can be detected when the thermodynamic and diffusion approaches described for the reac- tion center proteins and bacteriorhodopsin are used. Our photodimerization approach can lead to the detection of an irregular lateral distribu- tion of anthracene-labelled lipid homo- and het- ero-dimers indicating the presence of different lipid pools in the membrane. These pools can account for the existence of closed protein do- mains with different lipid compositions. However, the occurrence of specific lipid/protein interac- tions cannot be excluded, even if the perturbed and unperturbed lipids exchange at a rate close to that of the lateral diffusion of lipids. This is due to the fact that anthracene has a life-time of ~ 4 ns much shorter than the characteristic time of ~ 100 ns (or more) for the lateral diffusion of lipids. The D and M values measured for lipids in FRAP experiments contain information on the lateral organization of lipids in membranes. How- ever, interpretation of these parameters will de- pend on the relative time and space scales over which the lateral motion of lipids is considered to occur. Thus, the FRAP data obtained with bacte- riorhodopsin recombinants (see Section 4.2.1), which shows a decrease in the lipid D with in- creasing protein concentration but without alter- ation of the mobile fraction, illustrates the first concept that in FRAP experiments, the absence of an immobile lipid fraction does not necessarily mean the absence of lipid microdomains. Indeed, interpretation of this FRAP data leads to the conclusion that bacteriorhodopsin is surrounded

156 J.-F. Tocanne et al. / Chem. Phys. Lipids 73 (1994) 139-158
by about two layers of partly immobilized lipids in contact with freely diffusing lipids. In such a configuration, which corresponds to long-range protein/lipid interactions in conditions of rela- tively low protein concentrations, an immobile lipid fraction is not to be expected even assuming that the protein molecules are completely im- mobile and surrounded by immobilized lipids in a gel-like state. Indeed, and although these per- turbed lipids would diffuse laterally with a rela- tively high characteristic time of about 1 /~s (D ~ 10 -1° c m 2 / s for lipids in the gel state [20]), exchange between perturbed and unperturbed lipids would occur within less than 0.5 ms, be- cause the perturbed lipids have to cover a small area of only two lipid layers around the protein; 0.5 ms is a very short time as compared to the 10-15 s duration of a fluorescence recovery and exchange between perturbed and unperturbed lipids which can occur, thus explaining the absence of an immobile fraction. On the contrary, it is clear that the detection of an immobile lipid fraction in FRAP experiments accounts for the existence of lipid microdomains. Without really convincing arguments, these domains are sometimes postulated as being closed membrane regions, in the form of cages of proteins in which lipids are trapped. More simply, the possibility also exists of considering open domains in the form of extended protein-rich regions. On the grounds of the data obtained with bacteriorho- dopsin, simple calculation shows that for protein/lipid molar ratios of 1/100 and above, all the lipids in such domains would be perturbed by the proteins and therefore have a restricted dif- fusion. As discussed in Section 2.2.2, this type of domains would be detected by varying the radius r i of the illuminated area. With a size of around 1 /~m, approaching that of r i (1-2/xm), and enough surface concentration, they would make a signifi- cant contribution to the immobile lipid fraction in FRAP experiments.
In connection with the more general problem of how a living cell spatially controls the biogene- sis of its various membrane fractions and tem- porally maintains their spatial organization in re- lation to their functions, investigating the lateral
and transverse distribution and dynamics of mem- brane components remains a challenge of prime interest in cell biology. Development of new methodologies enabling the structure and compo- sition of protein/lipid domains to be described at a submicroscopic and even molecular level, mod- elization of membrane structure and dynamics and comparison with experimental data obtained from natural and model membranes, should be of great help.
Acknowledgements
We are grateful to J. Robb for rereading the English manuscript.
References
[1] A.H. Etemadi (1980) Biochim. Biophys. Acta 604, 347-422.
[2] G. Benga and R.P. Holmes (1984) Progr. Biophys. Mol. Biol. 43, 195-257.
[3] P.F. Devaux (1993) Curr. Opin. Struct. Biol. 3, 489-494. [4] J.F. Tocarme, L. Dupou-C6zanne, A. Lopez and J.F.
Tournier (1989) FEBS Lett. 257, 10-16. [5] J.F. Toeanne (1992) Comm. Mol. Cell. Biophys. 8,
53-72. [6] M. Giaser (1993) Curr. Opin. Struct. Biol. 3, 475-481. [7] W. Rodgers and M. Glaser (1991) Proc. Natl. Acad. Sci.
USA 88, 1364-1368. [8] M. Glaser (1992) Comm. Mol. Cell. Biophys. 8, 37-51. [9] M. Edidin (1992) Comm. Mol. Cell. Biophys. 8, 73-82.
[10] D.E. Wolf, V.M. Maynard, C.A. McKinnon and D.L. Melchior (1990) Proc. Natl. Acad. Sci. USA 87, 6893-6896.
[11] D.E. Wolf (1992) Comm. Mol. Cell. Biophys. 8, 83-95. [12] R.J. Cherry (1979) Biochim. Biophys. Acta 559,
289-327. [13] A.H. Etemadi (1980) Bioehim. Biophys. Aeta 604,
423-475. [14] P.F. Devaux (1988) FEBS Lett. 234, 8-12. [15] P.F. Devaux (1991) Biochemistry 30, 1163-1171. [16] L.O. Sillerud and R.E. Barnett (1982) Biochemistry 21,
1756-1760. [17] G. Morrot, S. Cribier, P.F. Devaux, D. Geldwerth, J.
Davoust, J.F. Bureau, P. Felmann, P. Herr6 and B. Friley (1986) Proc. Natl. Acad. Sci. USA 83, 6863-6867.
[18] J.M. El Hage Chahine, S. Cribier and P.F. Devaux (1993) Proc. Natl. Acad. Sei. USA 90, 447-451.
[19] M. Julien, J.F. Tournier and J.F. Tocanne (1993) Exp. Cell Res. 208, 387-397.
[20] J.F. Tocanne, L. Dupou-C6zanne and A. Lopez (1994) Prog. Lipid Res., 33, 203-237.

J.-F. Tocanne et aL / Chent Phys. Lipids 73 (1994) 139-158 157
[21] S.S. Spiegel, G.R. Matyas, L Cheng and B. Saektor (1988) Biechim. Biophys. Acta 938, 270-278.
[22] J.F. Tournier, A. Lopez, N. Gas and J.F. Tecanne (1989) Exp. Cell Res. 181, 375-384.
[23] M. Nakaehe, A.B. Schreiber, H. Gaub and H.M. Me- Connell (1985) Nature 317, 75-77.
[24] P.R. Dragsten, R. Blumentlaal and J.S. Handler (1981) Nature 294, 718-722.
[25] G. Van Meer and K. Simons (1986) EMBO J. 5, 1455-1464.
[26] M.S. PaUer (1993) Am. J. Physiol. 264, C201-C208. [27] S. Spiegel, R. Blumenthal, P.M. Fishman and J.S. Han-
dler (1985) Biechim. Biophys. Acta 821, 310-318. [28] G. Van Meer, B. Gumbiner and K. Simons (1986)
Nature 322, 639-641. [29] G.E. Nichols, C.A. Borgman and W.W. Young (1986)
Biochem. Biophys. Res. Commun. 138, 1163-1169. [30] T. Kobayashi, B. Storrie, K. Simons and C.G. Dotti
(1992) Nature 359, 647-650. [31] B. Gumbiner and D. Louvard (1985) Trends Biechem.
Sci. 435-438. [32] D.S. Friend (1982) J. Cell. Biol. 93, 243-249. [33] D.E. Wolf (1986) BioEssays 6, 116-121. [34] D.E. Wolf, B.K. Scott and C.F. Millette (1986) J. Cell.
Biol. 103, 1745-1750. [35] D.E. Wolf and J.K. Voglmayr (1984) J. Cell. Biol. 98,
1678-1684. [36] D.E. Wolf, S.S. Hagopian and S. Ishijima (1986) J. Cell.
Biol. 102, 1372-1377. [37] J.E. Speksnijder, M.R. Dohmen, L.G.J. Tertoolen and
S.W. de Laat (1985) Dev. Biol. 110, 207-216. [38] W.J.A.G. Dictus, E.J.J. Van Zoelen, P.A.T. Tetteroo,
L.G.J. Tertoolen, S.W. de Laat and J.G. Bluemink (1984) Dev. Biol. 101, 201-211.
[39] A. Lopez, L. Dupou, A. Altibelli, J. Trotard and J.F. Tecanne (1988) Biophys. J. 53, 963-970.
[40] E. Yechiel and M. Edidin (1987) J. Cell. Biol. 105, 755-760.
[41] J.P. Caulfield, C.P. Chiang, P.W. Yacono, L.A. Smith and D.E. Golan (1991) J. Cell Sci. 99, 167-173.
[42] M.L. Greenberg and D. Axelrod (1993) J. Membr. Biol. 131, 115-127.
[43] M. Welby and J.F. Tocanne (1982) Biechim. Biophys. Acta 689, 173-176.
[44] J. de Bony, G. Martin, M. Welby and J.F. Tecarme (1984) FEBS Lett. 174, 1-6.
[45] L. Dupou, J. Teissi6 and J.F. Tocanne (1986) Eur. J. Biechem. 154, 171-177.
[46] L. Dupou, A. Lopez and J.F. Toearme (1988) Eur. J. Biechem. 171, 669-674.
[47] J. de Bony, A. Lopez, M. Gilleron, M. Welby, G. Lan6elle, B. Rousseau, J.P. Beancourt and J.F. To- canne (1989) Biochemistry 28, 3728-3737.
[48] J. de Bony and J.F. Tocanne (1983) Chem. Phys. Lipids 32, 105-121.
[49] M. Vincent, J. Gallay, J. de Bony and J.F. Tecanne (1985) Eur. J. Biechem. 150, 341-347.
[50] E. P~rochon and J.F. Tecanne (1991) Chem. Phys. Lipids 58, 7-17.
[51] E. P~rechon, A. Lopez and J.F. Tecanne (1991) Chem. Phys. Lipids 59, 17-28.
[52] E. P~rechon, A. Lopez and J.F. Tocanne (1992) Biochemistry 31, 7672-7682.
[53] A. Lopez, L. Dupou, A. Altibelli, J. Trotard and J.F. Tecanne (1988) Biophys. J. 53, 963-970.
[54] X. Ferri~res, A. lopez, A. Altibelli, L. Dupou-C~zaune, J.L. Lagouanelle and J.F. Tocanne (1989) Biophys. J. 55, 1081-1091.
[55] J. de Bony and J.F. Tecanne (1984) Eur. J. Biechem. 143, 373-379.
[56] L. CEzanne, L. Navarro and J.F. Tocanne (1992) Biechim. Biophys. Acta 1112, 205-214.
[57] F. Lakhdar-Ghazal and J.F. Tecanne, (1981) Biochim. Biophys. Acta 644, 284-294.
[58] D.E. Golan, M.R. Alecio, W.R. Veatch and R.R. Rando (1984) Biochemistry 23, 332-339.
[59] S. Cribier, G. Morrot, J.M. Neumarm and P.F. Devaux (1990) Eur. Biophys. J. 18, 33-41.
[60] K. Jacobson, Y. Hou, Z. Derzko, J. Wojcieszyn and D. Organisciak (1982) Biophys. J. 37, 8-9.
[61] B. Chazotte, E.S. Wu and R. Hackenbreck (1984) Biechem. Sec. Trans. 12, 463-464.
[62] W.L.C. Vaz, F. Goodsaid-Zalduendo and K. Jacobson (1984) FEBS Lett. 174, 199-207.
[63] R. Peters and R.J. Cherry (1982) Prec. Natl. Acad. Sci USA 79, 4317-4321.
[64] O.H. Griffith, J.R. Brotherus and P. Jost (1982) in: P. Jost and O.H. Griffith (Eds.), Lipid-Protein Interac- tions, Vol. 2: John Wiley, New-York, pp. 225-237.
[65] D. Marsh (1983) Trends Biechem. Sci. 8, 330-333. [66] D. Marsh (1985) in: A. Watts and J.J. De Pont (Eds.),
Progress in Protein-Lipid Interactions, Vol. 1: Elsevier, Amsterdam, pp. 143-172.
[67] P.F. Devaux and M. Seigneuret (1985) Biochim. Bio- phys. Acta 822, 63-125.
[68] D. Marsh and A. Watts (1988) in: R.C. Aloia, C.C. Curtain and L.M. Gordon (Eds.), Advances in Mem- brane Fluidity, Vol. 2: Alan R. Liss, Inc., New York, pp. 163-200.
[69] B.R. Lentz (1988) in: R.C. Aloia, C.C. Curtain and L.M. Gordon (Eds.), Advances in Membrane Fluidity, Vol. 2, Alan R. Liss, New York, pp. 141-161.
[70] D. Marsh (1990) FEBS Lett. 2, 371-375. [71] A.C. Newton (1993) Annu. Rev. Biophys. Biomol.
Struct. 22, 1-25. [72] P.F. Knowles, A. Watts and D. Marsh (1981) Biochem-
istry 20, 5888-5894. [73] M. Bloom, E. Evans and O.G. Mouritsen (1991) O.
Rev. Biophys. 24, 293-397. [74] D. Chapman, J.C. Gomez-Fernadez and F.M. Goni
(1982) TIBS 67-70. [75] M.R. Paddy and F.W. Dahlquist (1982) Biophys. J. 37,
110-112.

158 J.-F. Tocanne et aL / Chem. Phys. Lipids 73 (1994) 139-158
[76] M.J. Saxton (1987) Biophys. J. 52, 989-997. [77] M.J. Saxton (1989) Biophys. J. 56, 615-622. [78] M.J. Saxton (1992) Biophys. J. 61, 119-128. [79] M.J. Saxton (1993) Biophys. J. 64, 1766-1780. [80] J. Einsenger, J. Flores and W.P. Petersen (1986) Bin-
phys. J. 49, 987-1001. [81] P.F.F. Almeida, W.L.C. Vaz and T.E. Thompson (1992)
Biochemistry 31, 7198-7210. [82] M.H. Cohen and D.J. Turnbull (1959) Chem. Phys. 31,
1164-1169. [83] H.J. Galla, W. Hartmann, U. Theilen and E.U. Sack-
mann (1979) J. Membr. Biol. 48, 215-236. [84] W.LC. Vaz and D. HaUmann (1983) FEBS Lett. 152,
287-290. [85] W.L.C. Vaz, R.M. Clegg and D. Hallmann (1985)
Biochemistry 24, 781-786. [86] D. Stauffer and A. Aharony (1992) in Introduction to
Percolation Theory, Taylor and Francis, London, p. 17. [87] M.P. Heyn, A~ Blurae, M. Rehorek and N.A. Dencher
(1981) Biochemistry 20, 7109-7115. [88] T. Gulik-Kxzywicki, M. Seigneuret and J.L. Rigaud
(1987) J. Biol. Chem. 262, 15580-15588. [89] F. J~anig (1981) Biophys. J. 36, 329-345. [90] J. Riegler and H. M6hwald (1986) Biophys. J., 49,
1111-1118. [91] J. Peschke, J. Riegler and H. M6hwald (1987) Eur.
Biophys. J. 14, 385-391.
[92] M. Rehorek, N.A. Dencher and M.P. Heyn (1985) Biochemistry 24, 5980-5988.
[93] B. Piknova, E. P~rochon and J.F. Tocanne (1993) Eur. J. Biochem. 218, 385-396.
[94] O.G. Mouritsen and M. Bloom (1984) Biophys. J. 46, 141-153.
[95] M.M. Sperotto and O.G. Mouritsen (1988) Eur. Bio- phys. J. 16, 1-10.
[96] M.M. Sperotto and O.G. Mouritsen (1991) Eur. Bio- phys. J. 19, 157-168.
[97] M.M. Sperotto and O.G. Mouritsen (1991) Biophys. J. 59, 261-270.
[98] D.R. Fattal and A. Ben-Shaal (1993) Biophys. J. 65, 1795-1809.
[99] O.G. Mouritsen and M.M. Sperotto (1992) in: M. Jack- son (Ed.), Thermodynamics of Cell Surface Receptors, CRC Press, Boca Raton, FL, pp. 127-181.
[100] O.G. Mouritsen and R.L. Biltonen (1993) in: A. Watts (Ed.), Protein-Lipid Interactions, Elsevier, Amsterdam, pp. 1-39.
[101] O.G. Mouritsen and M. Bloom (1993) Annu. Rev. Biophys. Biomol. Struct. 22, 145-171.
[102] M.M. Sperotto, J.H. Ipsen and O.G. Mouritsan (1989) Cell Biophys. 14, 79-95.
[103] M.M. Sperotto and O.G. Mouritsen (1993) Eur. Bio- phys. J. 22, 323-328.
Related Documents











