1 3 J Ind Microbiol Biotechnol (2016) 43:1467–1480 DOI 10.1007/s10295-016-1820-0 GENETICS AND MOLECULAR BIOLOGY OF INDUSTRIAL ORGANISMS - ORIGINAL PAPER Lipase genes in Mucor circinelloides: identification, sub‑cellular location, phylogenetic analysis and expression profiling during growth and lipid accumulation Xinyi Zan 1 · Xin Tang 1 · Linfang Chu 1 · Lina Zhao 1 · Haiqin Chen 1,3 · Yong Q. Chen 1,3 · Wei Chen 1,3 · Yuanda Song 1,2 Received: 24 May 2016 / Accepted: 30 July 2016 / Published online: 17 August 2016 © Society for Industrial Microbiology and Biotechnology 2016 lipases also contained a typical acyltransferase motif of H-(X) 4 -D, and these lipases may play a dual role in lipid metabolism, catalyzing both lipid hydrolysis and transacyla- tion reactions. The differential expression of all lipase genes were confirmed by quantitative real-time PCR, and the expression profiling were analyzed to predict the possible biological roles of these lipase genes in lipid metabolism in M. circinelloides. We preliminarily hypothesized that lipases may be involved in triacylglycerol degradation, phospho- lipid synthesis and beta-oxidation. Moreover, the results of sub-cellular localization, the presence of signal peptide and transcriptional analyses of lipase genes indicated that four lipase in WJ11 most likely belong to extracellular lipases with a signal peptide. These findings provide a platform for the selection of candidate lipase genes for further detailed functional study. Introduction Lipases or triacylglycerol hydrolases (EC 3.1.1.3) are enzymes which hydrolyze ester bonds between fatty acids and glycerol [4]. They hydrolyze triglycerides (TAG) into diglycerides, monoglycerides, fatty aicds and glycerol [15]. Lipases can also catalyze ester synthesis via esterification and transesterification (alcoholysis and acidolysis) and interesterification reactions. Additionally, they can be used in numerous bioconversion reactions such as aminolysis, amide and thioester synthesis and hydrolysis [24]. Due to these diverse properties, lipases are widely used in waste treatment [13], fine chemicals refinery [14], traditional food making [25, 29], citric acid production [17] and phar- maceutical industries [35]. Lipases belong to the structural super family of α/β- hydrolases [10]. Their activities rely on a catalytic triad Abstract Lipases or triacylglycerol hydrolases are widely spread in nature and are particularly common in the micro- bial world. The filamentous fungus Mucor circinelloides is a potential lipase producer, as it grows well in triacylglyc- erol-contained culture media. So far only one lipase from M. circinelloides has been characterized, while the major- ity of lipases remain unknown in this fungus. In the present study, 47 potential lipase genes in M. circinelloides WJ11 and 30 potential lipase genes in M. circinelloides CBS 277.49 were identified by extensive bioinformatics analysis. An overview of these lipases is presented, including several characteristics, sub-cellular location, phylogenetic analysis and expression profiling of the lipase genes during growth and lipid accumulation. All of these proteins contained the consensus sequence for a classical lipase (GXSXG motif) and were divided into four types including α/β-hydrolase_1, α/β-hydrolase_3, class_3 and GDSL lipase (GDSL) based on gene annotations. Phylogenetic analyses revealed that class_3 family and α/β-hydrolase_3 family were the con- served lipase family in M. circinelloides. Additionally, some Electronic supplementary material The online version of this article (doi:10.1007/s10295-016-1820-0) contains supplementary material, which is available to authorized users. * Yuanda Song [email protected] 1 State Key Laboratory of Food Science and Technology, School of Food Science and Technology, Jiangnan University, Wuxi, People’s Republic of China 2 Colin Ratledge Center for Microbial Lipids, School of Agriculture Engineering and Food Science, Shandong University of Technology, Zibo, People’s Republic of China 3 Synergistic Innovation Center for Food Safety and Nutrition, Wuxi, People’s Republic of China Downloaded from https://academic.oup.com/jimb/article/43/10/1467/5996015 by guest on 11 January 2022

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 3
J Ind Microbiol Biotechnol (2016) 43:1467–1480DOI 10.1007/s10295-016-1820-0
GENETICS AND MOLECULAR BIOLOGY OF INDUSTRIAL ORGANISMS - ORIGINAL PAPER
Lipase genes in Mucor circinelloides: identification, sub‑cellular location, phylogenetic analysis and expression profiling during growth and lipid accumulation
Xinyi Zan1 · Xin Tang1 · Linfang Chu1 · Lina Zhao1 · Haiqin Chen1,3 · Yong Q. Chen1,3 · Wei Chen1,3 · Yuanda Song1,2
Received: 24 May 2016 / Accepted: 30 July 2016 / Published online: 17 August 2016 © Society for Industrial Microbiology and Biotechnology 2016
lipases also contained a typical acyltransferase motif of H-(X) 4-D, and these lipases may play a dual role in lipid metabolism, catalyzing both lipid hydrolysis and transacyla-tion reactions. The differential expression of all lipase genes were confirmed by quantitative real-time PCR, and the expression profiling were analyzed to predict the possible biological roles of these lipase genes in lipid metabolism in M. circinelloides. We preliminarily hypothesized that lipases may be involved in triacylglycerol degradation, phospho-lipid synthesis and beta-oxidation. Moreover, the results of sub-cellular localization, the presence of signal peptide and transcriptional analyses of lipase genes indicated that four lipase in WJ11 most likely belong to extracellular lipases with a signal peptide. These findings provide a platform for the selection of candidate lipase genes for further detailed functional study.
Introduction
Lipases or triacylglycerol hydrolases (EC 3.1.1.3) are enzymes which hydrolyze ester bonds between fatty acids and glycerol [4]. They hydrolyze triglycerides (TAG) into diglycerides, monoglycerides, fatty aicds and glycerol [15]. Lipases can also catalyze ester synthesis via esterification and transesterification (alcoholysis and acidolysis) and interesterification reactions. Additionally, they can be used in numerous bioconversion reactions such as aminolysis, amide and thioester synthesis and hydrolysis [24]. Due to these diverse properties, lipases are widely used in waste treatment [13], fine chemicals refinery [14], traditional food making [25, 29], citric acid production [17] and phar-maceutical industries [35].
Lipases belong to the structural super family of α/β-hydrolases [10]. Their activities rely on a catalytic triad
Abstract Lipases or triacylglycerol hydrolases are widely spread in nature and are particularly common in the micro-bial world. The filamentous fungus Mucor circinelloides is a potential lipase producer, as it grows well in triacylglyc-erol-contained culture media. So far only one lipase from M. circinelloides has been characterized, while the major-ity of lipases remain unknown in this fungus. In the present study, 47 potential lipase genes in M. circinelloides WJ11 and 30 potential lipase genes in M. circinelloides CBS 277.49 were identified by extensive bioinformatics analysis. An overview of these lipases is presented, including several characteristics, sub-cellular location, phylogenetic analysis and expression profiling of the lipase genes during growth and lipid accumulation. All of these proteins contained the consensus sequence for a classical lipase (GXSXG motif) and were divided into four types including α/β-hydrolase_1, α/β-hydrolase_3, class_3 and GDSL lipase (GDSL) based on gene annotations. Phylogenetic analyses revealed that class_3 family and α/β-hydrolase_3 family were the con-served lipase family in M. circinelloides. Additionally, some
Electronic supplementary material The online version of this article (doi:10.1007/s10295-016-1820-0) contains supplementary material, which is available to authorized users.
* Yuanda Song [email protected]
1 State Key Laboratory of Food Science and Technology, School of Food Science and Technology, Jiangnan University, Wuxi, People’s Republic of China
2 Colin Ratledge Center for Microbial Lipids, School of Agriculture Engineering and Food Science, Shandong University of Technology, Zibo, People’s Republic of China
3 Synergistic Innovation Center for Food Safety and Nutrition, Wuxi, People’s Republic of China
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022

1468 J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
formed by Ser, His and Asp residues. Serine is especially essential for lipase activities and usually appears in a highly conserved motif (G/A) XSXG, which is characteris-tic of α/β-hydrolases [16]. The sequence change within the motif (G/A) XSXG is one of the factors to distinguish vari-ous lipolytic families [10]. For example, the true lipolytic enzymes including triacylglycerol lipase usually contain the G (H/F) SQG sequence, while the GDSLG sequence is a conserved characteristic of GDSL family. Recent studies found that some lipases contain not only the lipase motif GXSXG but also an H-(X)4-D acyltransferase motif such as lipases Tgl3p, Tgl4p, and Tgl5p in the yeast Saccharo-myces cerevisiae [24, 28] and ptl1, ptl2 and ptl3 in the fis-sion yeast Schizosaccharomyces pombe [37]. Furthermore, site-directed mutagenesis experiments clearly demon-strated that the histidine residue is indispensable for acyl-transferase activity of Tgl3p [24].
In general, lipases derived from microorganism can be divided into extracellular and intracellular enzymes. Extra-cellular lipases can be excreted into the culture medium such as CdLIP1 in yeast Candida deformans [4] and LIP2, LIP7 and LIP8 in yeast Yarrowia lipolytica [10]. Interest-ingly, CdLIP1, LIP7 and LIP8 are mainly associated to the cell wall to hydrolyze triglycerides, whereas YlLIP2 is the main lipase released in the culture medium at the end of the growth phase [11]. The syntheses and secretion of extracellular lipases cannot be triggered by glucose, glyc-erol or mineral nitrogen compounds (NH4Cl, (NH4)2SO4), but they are highly increased in the presence of hydropho-bic substrates such as fatty acid, methyl esters and oils [8]. Moreover, the expression of extracellular lipase genes such as YlLIP2 also requires the regulation of SOA genes (spe-cific for oleic acid) [7]. In contrast, intracellular lipases are mainly located on the surface of lipid bodies (LBs) and a few of them are located in mitochondria. Distinct sub-cel-lular location could affect the role of intracellular lipases in cell metabolism. For example, lipases Tgl3, Tgl4 and Tgl5 in yeast S. cerevisiae [24, 28] and TGL3 and TGL4 in yeast Y. lipolytica [9] are located in LBs. These enzymes usually participate in the LB-associated lipid metabolism including TAG homeostasis, regulation of fatty acid com-position, and spore formation of cells. However, lipase Tgl2 in yeast S. cerevisiae is located in mitochondria and may be essential for cell survival under stress-inducing conditions [15, 19].
Fungal cells have been considered as important sources of lipase [2, 22]. The filamentous fungus Mucor circinel-loides grows well in triacylglycerol-containing culture media, which indicates that the fungus is a potential extra-cellular lipase producer [2]. Although a few studies have also investigated the potential of immobilized M. circinel-loides whole cells as a catalyst for ethyl esters synthesis, transesterificantion and ethanolysis [1, 3, 5, 23, 30, 31],
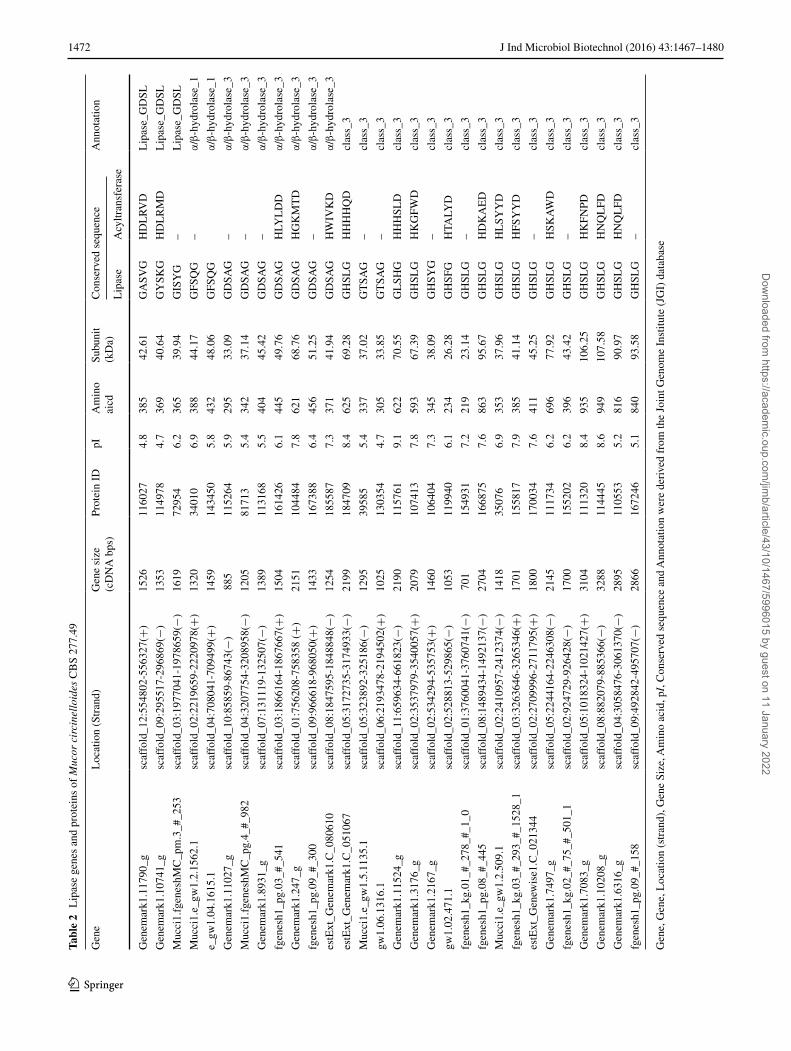
only one mycelium-bound lipase has been reported in Mucor javanicus, which has been reclassified as Mucor circinelloide [12]. Previous studies from genetic perspec-tive have revealed that a large number of lipase genes exist in the fungus Mucor circinelloide [33, 36, 40]. Therefore, it is necessary to study the roles and specificity of the remaining lipases in M. circinelloides. Moreover, a lot of enzymes and/or genes related to TAG synthesis such as malic enzyme, glucose 6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase have been investi-gated to explore the mechanisms of lipid accumulation in M. circinelloides [26, 34, 38, 41]. However, the enzymes involved in TAG degradation such as lipase in this fungus have not been studied at all. We have previously reported several characteristics of the 30 potential lipases in a low lipid-producing strain M. circinelloides CBS 277.49 (15 % w/w lipid, cell dry weight) [40]. Lipase Lip6 or Lip10 from CBS 277.49 may control acyltransferase activity and play a role in fatty acid reconstruction for TAG, while Lip19 or Lip24 was involved in TAG degradation as a TAG lipase. Recently, a new strain M. circinelloides WJ11 isolated in our laboratory is able to produce up to 36 % (w/w) lipid of cell dry weight (CDW) [32]. The differ-ence on lipases genes involved in lipid metabolism might provide an insight for the different lipid content between strain CBS 277.49 and strain WJ11. Thus, in the present study we searched all possible genes encoding lipase in M. circinelloides WJ11 based on its genome database and compared these genes with that of CBS 277.49. The characteristics, sub-cellular location of these lipase genes were analyzed and phylogenetic analysis and expression profiling of the lipase genes during growth and lipid accu-mulation were performed. To our knowledge, this is the first and comprehensive report of the characterizations of lipases in M. circinelloides.
Materials and methods
Identification of lipase genes in M. circinelloides
Based on genomes of M. circinelloides CBS 277.49 and WJ11, related lipase genes were retrieved. Crite-ria for the identification of lipase genes in M. circinel-loides include gene annotations and the presence of con-served domains. The sequence of gene and protein, gene location and size, protein ID, conserved sequence and annotations of lipase genes in CBS 277.49 and WJ11 were acquired from the Joint Genome Institute (JGI) (http://genome.jgi.doe.gov/pages/search-for-genes.jsf?organism=Mucci2) and the National Center for Bio-technology Information (NCBI) (http://www.ncbi.nlm.nih.gov/genome/?term=LGTF00000000), respectively.
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022

1469J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
The numbers of amino acid and molecular weight were analyzed on the website (http://www.bio-soft.net/sms/prot_mw.html). The protein isoelectric point was analyzed by using isoeletric point calculator (IPC) (http://isoelectric.ovh.org/).
Analysis of sub‑cellular location, signal peptide and transmembrane domains
TargetP1.1 Server (http://www.cbs.dtu.dk/services/Tar-getP/), SignalP 4.1 Server program (http://www.cbs.dtu.dk/services/SignalP/) and TMHMM (http://www.cbs.dtu.dk/service/TMHMM) were used to predict the sub-cellular location, the presence and location of signal peptide and transmembrane spanning region of lipase proteins from M. circinelloides CBS 277.49 and WJ11, respectively.
Phylogenetic and sequence homology analysis of lipase proteins
The phylogenetic tree was constructed based on the full-length sequences of these lipases. A neighbor-joining tree was built using MEGA 6.06, adopting the interior-branch test. Support for the tree obtained was assessed using the bootstrap method with 1000 replicates. Sequence homol-ogy analyses were performed using pairwise sequence alignment.
RNA isolation and transcriptional analyses of lipase genes by qRT‑PCR
100 μL spore suspension (approx. 107 spores/mL) of M. circinelloides CBS 277.49 or WJ11 was inoculated into 150 mL K&R medium [18] held in a 1 L flask equipped with baffles to increase aeration. Cultures were incubated at 30 °C for 24 h with shaking at 150 rpm and then 10 % of the culture were used as the seed culture to inoculate the 2-L fermentor containing 1.5 L modified K&R medium (2 g diammonium tartrate, 80 g glucose per liter plus inor-ganic salts), which was incubated with aeration at 0.5 v/v min−1 and stirring at 700 rpm. pH was maintained at 6.0 by auto-addition of sterilized 4 M KOH or 2 M H2SO4.
Based on the characteristic lipid accumulation of M. circinelloides WJ11 and CBS 277.49, cells at 6, 24 and 72 h were harvested and used for transcriptional analysis of lipase genes. Total RNA was extracted by an RNAiso Plus kit after grinding under liquid N2 and reverse-tran-scribed using the Prime ScriptRT reagent kit (Takara, Japan) according to the manufacturer’s instructions. Real-Time quantitative PCR was performed in BioRad CFX96 (BioRad, CA, USA) using the iTaq™ Universal SYBR® Green Supermix (BioRad, CA, USA). Relative quantifica-tion was based on the 2−△△Ct method using 18S rRNA of
M. circinelloides as a housekeeping gene [21]. The ther-mal cycling conditions for the amplification reaction were as follows: 95 °C 30 s, 51/53/55/58 °C 30 s (40 cycles). Three replicates, prepared from independent biological samples, were analyzed. The primer sequences used for amplification of lipase genes are listed in Tables S1 and S2. We applied a fold-change cutoff of ≥1.5 for up-regulation, and ≤0.5 for down-regulation.
Results and discussion
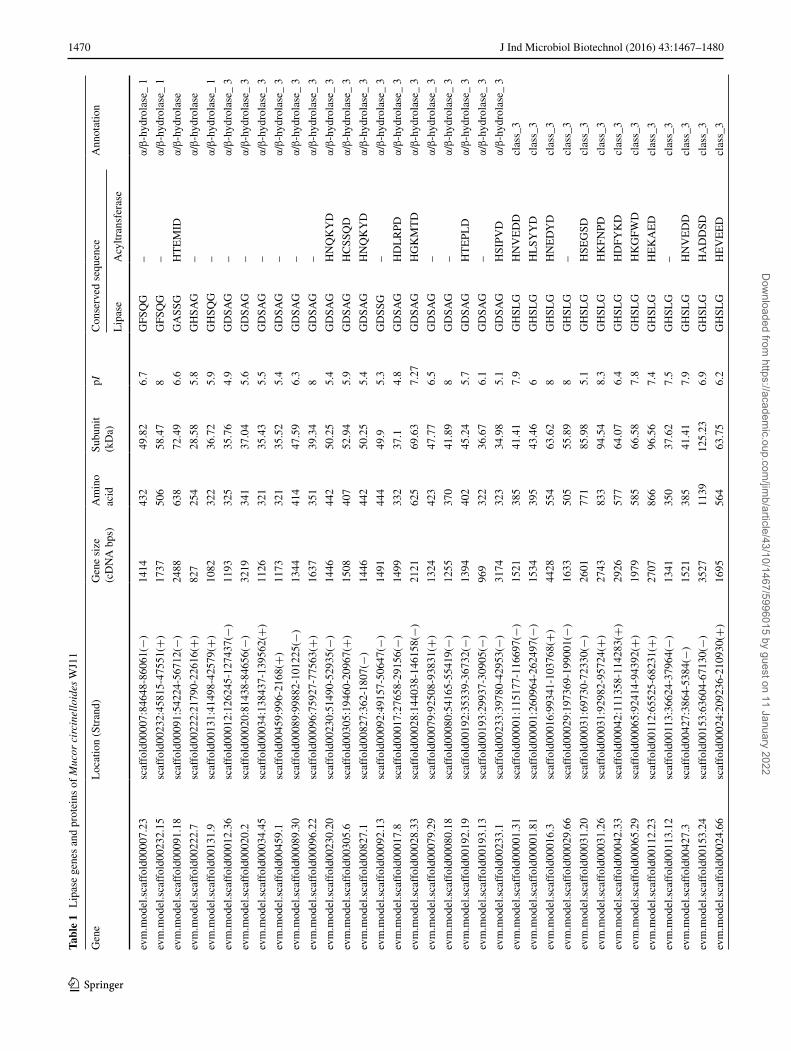
Identification of lipase genes in M. circinelloides WJ11
Based on the gene annotations and conserved motif sequences, we are surprised to find 47 potential genes of lipase in high lipid-producing strain M. circinelloides WJ11 (Table 1), which is more than the number of poten-tial lipase genes in low lipid-producing strain M. circinel-loides CBS 277.49 (30, Table 2). However, strain WJ11 has similar average lipase gene and protein length, and the average isoelectric point with strain CBS 277.49. All of these proteins contained the consensus sequence for a classical lipase (GXSXG motif) with a classical catalytic triad containing a nucleophile serine. Gene annotations showed that lipases in strain WJ11 are mainly divided into four types including α/β-hydrolase_1, α/β-hydrolase_3, class_3 and GDSL. The sequence variation within the motif (G/A) XSXG may be a factor to distinguish various lipase families. For example, the α/β-hydrolase_1 fam-ily, α/β-hydrolase_3 family and class_3 family in strain WJ11 mainly conserve the GFSQG, GDSAG and GHSLG sequence, respectively, whereas the motif (G/A) XSXG of GDSL family is diverse in strain WJ11. Most lipases from the filamentous fungi including the fungus WJ11 and CBS 277.49 contain the GHSLG motif in class_3 family, while some unusual motifs (GLSVG, GHSFG, GHSYG, and GTSAG) were also observed in class_3 family in strain WJ11. Moreover, 25 lipases in WJ11 not only contain the typical lipase motif (G/A) XSXG, but also the con-sensus sequence motif H-(X) 4-D (a typical of acyltrans-ferase motif), while only 16 similar lipases are observed in strain CBS 277.49. These lipases may play a dual role in lipid metabolism, catalyzing both lipid hydrolysis and transacylation reactions. Similar results have been reported that lipases Tgl3 and Tgl5 in S. cerevisiae not only exhib-ited lipase activity but also catalyzed acylation of lysophos-phatidylethanolamine and lysophosphatidic acid, respec-tively [27, 28]. The microalga phospholipid:diacylglycerol acyltransferase (PDAT) also exerts multiple functions as a typical PDAT, galactolipid:DAG acyltransferase and lipase [39]. We speculate that synthesis of these multi-functional enzymes allows better resource utilization and management
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022
1470 J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
Tabl
e 1
Lip
ase
gene
s an
d pr
otei
ns o
f M
ucor
cir
cine
lloi
des
WJ1
1
Gen
eL
ocat
ion
(Str
and)
Gen
e si
ze
(cD
NA
bps
)A
min
o ac
idSu
buni
t (k
Da)
pIC
onse
rved
seq
uenc
eA
nnot
atio
n
Lip
ase
Acy
ltran
sfer
ase
evm
.mod
el.s
caff
old0
0007
.23
scaf
fold
0000
7:84
648-
8606
1(−
)14
1443
249
.82
6.7
GFS
QG
–α
/β-h
ydro
lase
_ 1
evm
.mod
el.s
caff
old0
0232
.15
scaf
fold
0023
2:45
815-
4755
1(+
)17
3750
658
.47
8G
FSQ
G–
α/β
-hyd
rola
se_
1
evm
.mod
el.s
caff
old0
0091
.18
scaf
fold
0009
1:54
224-
5671
2(−
)24
8863
872
.49
6.6
GA
SSG
HT
EM
IDα
/β-h
ydro
lase
evm
.mod
el.s
caff
old0
0222
.7sc
affo
ld00
222:
2179
0-22
616(+
)82
725
428
.58
5.8
GH
SAG
–α
/β-h
ydro
lase
evm
.mod
el.s
caff
old0
0131
.9sc
affo
ld00
131:
4149
8-42
579(+
)10
8232
236
.72
5.9
GH
SQG
–α
/β-h
ydro
lase
_ 1
evm
.mod
el.s
caff
old0
0012
.36
scaf
fold
0001
2:12
6245
-127
437(−
)11
9332
535
.76
4.9
GD
SAG
–α
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0020
.2sc
affo
ld00
020:
8143
8-84
656(−
)32
1934
137
.04
5.6
GD
SAG
–α
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0034
.45
scaf
fold
0003
4:13
8437
-139
562(+
)11
2632
135
.43
5.5
GD
SAG
–α
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0459
.1sc
affo
ld00
459:
996-
2168
(+)
1173
321
35.5
25.
4G
DSA
G–
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0089
.30
scaf
fold
0008
9:99
882-
1012
25(−
)13
4441
447
.59
6.3
GD
SAG
–α
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0096
.22
scaf
fold
0009
6:75
927-
7756
3(+
)16
3735
139
.34
8G
DSA
G–
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0230
.20
scaf
fold
0023
0:51
490-
5293
5(−
)14
4644
250
.25
5.4
GD
SAG
HN
QK
YD
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0305
.6sc
affo
ld00
305:
1946
0-20
967(+
)15
0840
752
.94
5.9
GD
SAG
HC
SSQ
Dα
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0827
.1sc
affo
ld00
827:
362-
1807
(−)
1446
442
50.2
55.
4G
DSA
GH
NQ
KY
Dα
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0092
.13
scaf
fold
0009
2:49
157-
5064
7(−
)14
9144
449
.95.
3G
DSS
G–
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0017
.8sc
affo
ld00
017:
2765
8-29
156(−
)14
9933
237
.14.
8G
DSA
GH
DL
RPD
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0028
.33
scaf
fold
0002
8:14
4038
-146
158(−
)21
2162
569
.63
7.27
GD
SAG
HG
KM
TD
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0079
.29
scaf
fold
0007
9:92
508-
9383
1(+
)13
2442
347
.77
6.5
GD
SAG
–α
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0080
.18
scaf
fold
0008
0:54
165-
5541
9(−
)12
5537
041
.89
8G
DSA
G–
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0192
.19
scaf
fold
0019
2:35
339-
3673
2(−
)13
9440
245
.24
5.7
GD
SAG
HT
EPL
Dα
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0193
.13
scaf
fold
0019
3:29
937-
3090
5(−
)96
932
236
.67
6.1
GD
SAG
–α
/β-h
ydro
lase
_ 3
evm
.mod
el.s
caff
old0
0233
.1sc
affo
ld00
233:
3978
0-42
953(−
)31
7432
334
.98
5.1
GD
SAG
HSI
PVD
α/β
-hyd
rola
se_
3
evm
.mod
el.s
caff
old0
0001
.31
scaf
fold
0000
1:11
5177
-116
697(−
)15
2138
541
.41
7.9
GH
SLG
HN
VE
DD
clas
s_3
evm
.mod
el.s
caff
old0
0001
.81
scaf
fold
0000
1:26
0964
-262
497(−
)15
3439
543
.46
6G
HSL
GH
LSY
YD
clas
s_3
evm
.mod
el.s
caff
old0
0016
.3sc
affo
ld00
016:
9934
1-10
3768
(+)
4428
554
63.6
28
GH
SLG
HN
ED
YD
clas
s_3
evm
.mod
el.s
caff
old0
0029
.66
scaf
fold
0002
9:19
7369
-199
001(−
)16
3350
555
.89
8G
HSL
G–
clas
s_3
evm
.mod
el.s
caff
old0
0031
.20
scaf
fold
0003
1:69
730-
7233
0(−
)26
0177
185
.98
5.1
GH
SLG
HSE
GSD
clas
s_3
evm
.mod
el.s
caff
old0
0031
.26
scaf
fold
0003
1:92
982-
9572
4(+
)27
4383
394
.54
8.3
GH
SLG
HK
FNPD
clas
s_3
evm
.mod
el.s
caff
old0
0042
.33
scaf
fold
0004
2:11
1358
-114
283(+
)29
2657
764
.07
6.4
GH
SLG
HD
FYK
Dcl
ass_
3
evm
.mod
el.s
caff
old0
0065
.29
scaf
fold
0006
5:92
414-
9439
2(+
)19
7958
566
.58
7.8
GH
SLG
HK
GFW
Dcl
ass_
3
evm
.mod
el.s
caff
old0
0112
.23
scaf
fold
0011
2:65
525-
6823
1(+
)27
0786
696
.56
7.4
GH
SLG
HE
KA
ED
clas
s_3
evm
.mod
el.s
caff
old0
0113
.12
scaf
fold
0011
3:36
624-
3796
4(−
)13
4135
037
.62
7.5
GH
SLG
–cl
ass_
3
evm
.mod
el.s
caff
old0
0427
.3sc
affo
ld00
427:
3864
-538
4(−
)15
2138
541
.41
7.9
GH
SLG
HN
VE
DD
clas
s_3
evm
.mod
el.s
caff
old0
0153
.24
scaf
fold
0015
3:63
604-
6713
0(−
)35
2711
3912
5.23
6.9
GH
SLG
HA
DD
SDcl
ass_
3
evm
.mod
el.s
caff
old0
0024
.66
scaf
fold
0002
4:20
9236
-210
930(+
)16
9556
463
.75
6.2
GH
SLG
HE
VE
ED
clas
s_3
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022
1471J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
for maintaining lipid homeostasis necessary for growth and reproduction, as well as for adapting to changing environment.
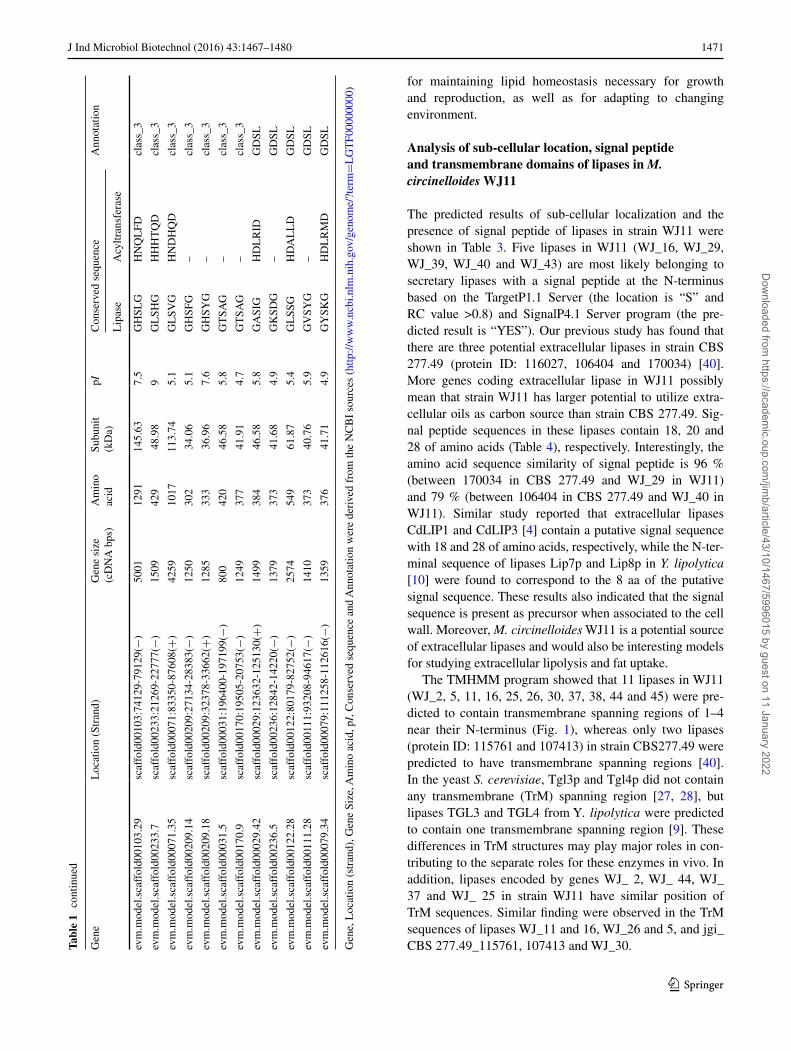
Analysis of sub‑cellular location, signal peptide and transmembrane domains of lipases in M. circinelloides WJ11
The predicted results of sub-cellular localization and the presence of signal peptide of lipases in strain WJ11 were shown in Table 3. Five lipases in WJ11 (WJ_16, WJ_29, WJ_39, WJ_40 and WJ_43) are most likely belonging to secretary lipases with a signal peptide at the N-terminus based on the TargetP1.1 Server (the location is “S” and RC value >0.8) and SignalP4.1 Server program (the pre-dicted result is “YES”). Our previous study has found that there are three potential extracellular lipases in strain CBS 277.49 (protein ID: 116027, 106404 and 170034) [40]. More genes coding extracellular lipase in WJ11 possibly mean that strain WJ11 has larger potential to utilize extra-cellular oils as carbon source than strain CBS 277.49. Sig-nal peptide sequences in these lipases contain 18, 20 and 28 of amino acids (Table 4), respectively. Interestingly, the amino acid sequence similarity of signal peptide is 96 % (between 170034 in CBS 277.49 and WJ_29 in WJ11) and 79 % (between 106404 in CBS 277.49 and WJ_40 in WJ11). Similar study reported that extracellular lipases CdLIP1 and CdLIP3 [4] contain a putative signal sequence with 18 and 28 of amino acids, respectively, while the N-ter-minal sequence of lipases Lip7p and Lip8p in Y. lipolytica [10] were found to correspond to the 8 aa of the putative signal sequence. These results also indicated that the signal sequence is present as precursor when associated to the cell wall. Moreover, M. circinelloides WJ11 is a potential source of extracellular lipases and would also be interesting models for studying extracellular lipolysis and fat uptake.
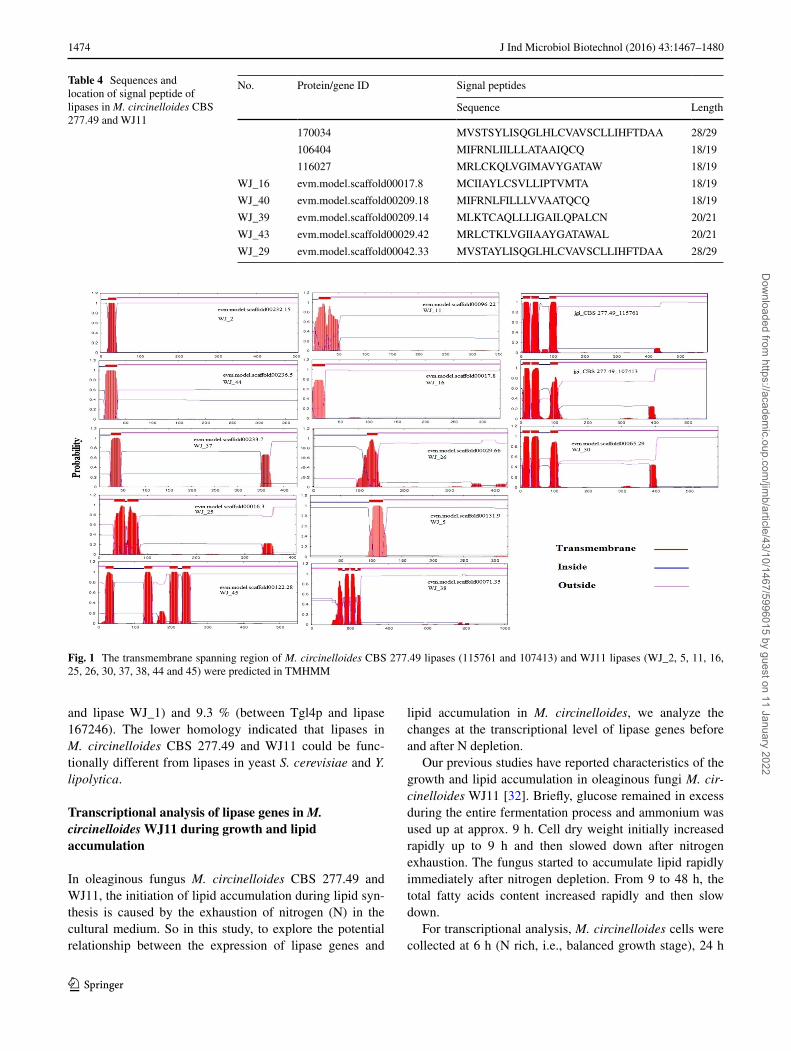
The TMHMM program showed that 11 lipases in WJ11 (WJ_2, 5, 11, 16, 25, 26, 30, 37, 38, 44 and 45) were pre-dicted to contain transmembrane spanning regions of 1–4 near their N-terminus (Fig. 1), whereas only two lipases (protein ID: 115761 and 107413) in strain CBS277.49 were predicted to have transmembrane spanning regions [40]. In the yeast S. cerevisiae, Tgl3p and Tgl4p did not contain any transmembrane (TrM) spanning region [27, 28], but lipases TGL3 and TGL4 from Y. lipolytica were predicted to contain one transmembrane spanning region [9]. These differences in TrM structures may play major roles in con-tributing to the separate roles for these enzymes in vivo. In addition, lipases encoded by genes WJ_ 2, WJ_ 44, WJ_ 37 and WJ_ 25 in strain WJ11 have similar position of TrM sequences. Similar finding were observed in the TrM sequences of lipases WJ_11 and 16, WJ_26 and 5, and jgi_CBS 277.49_115761, 107413 and WJ_30.Ta
ble
1 c
ontin
ued
Gen
eL
ocat
ion
(Str
and)
Gen
e si
ze
(cD
NA
bps
)A
min
o ac
idSu
buni
t (k
Da)
pIC
onse
rved
seq
uenc
eA
nnot
atio
n
Lip
ase
Acy
ltran
sfer
ase
evm
.mod
el.s
caff
old0
0103
.29
scaf
fold
0010
3:74
129-
7912
9(−
)50
0112
9114
5.63
7.5
GH
SLG
HN
QL
FDcl
ass_
3
evm
.mod
el.s
caff
old0
0233
.7sc
affo
ld00
233:
2126
9-22
777(−
)15
0942
948
.98
9G
LSH
GH
HH
TQ
Dcl
ass_
3
evm
.mod
el.s
caff
old0
0071
.35
scaf
fold
0007
1:83
350-
8760
8(+
)42
5910
1711
3.74
5.1
GL
SVG
HN
DH
QD
clas
s_3
evm
.mod
el.s
caff
old0
0209
.14
scaf
fold
0020
9:27
134-
2838
3(−
)12
5030
234
.06
5.1
GH
SFG
–cl
ass_
3
evm
.mod
el.s
caff
old0
0209
.18
scaf
fold
0020
9:32
378-
3366
2(+
)12
8533
336
.96
7.6
GH
SYG
–cl
ass_
3
evm
.mod
el.s
caff
old0
0031
.5sc
affo
ld00
031:
1964
00-1
9719
9(−
)80
042
046
.58
5.8
GT
SAG
–cl
ass_
3
evm
.mod
el.s
caff
old0
0170
.9sc
affo
ld00
170:
1950
5-20
753(−
)12
4937
741
.91
4.7
GT
SAG
–cl
ass_
3
evm
.mod
el.s
caff
old0
0029
.42
scaf
fold
0002
9:12
3632
-125
130(+
)14
9938
446
.58
5.8
GA
SIG
HD
LR
IDG
DSL
evm
.mod
el.s
caff
old0
0236
.5sc
affo
ld00
236:
1284
2-14
220(−
)13
7937
341
.68
4.9
GK
SDG
–G
DSL
evm
.mod
el.s
caff
old0
0122
.28
scaf
fold
0012
2:80
179-
8275
2(−
)25
7454
961
.87
5.4
GL
SSG
HD
AL
LD
GD
SL
evm
.mod
el.s
caff
old0
0111
.28
scaf
fold
0011
1:93
208-
9461
7(−
)14
1037
340
.76
5.9
GV
SYG
–G
DSL
evm
.mod
el.s
caff
old0
0079
.34
scaf
fold
0007
9:11
1258
-112
616(−
)13
5937
641
.71
4.9
GY
SKG
HD
LR
MD
GD
SL
Gen
e, L
ocat
ion
(str
and)
, Gen
e Si
ze, A
min
o ac
id, p
I, C
onse
rved
seq
uenc
e an
d A
nnot
atio
n w
ere
deri
ved
from
the
NC
BI
sour
ces
(http
://w
ww
.ncb
i.nlm
.nih
.gov
/gen
ome/
?ter
m=
LG
TF0
0000
000)
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022
1472 J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
Tabl
e 2
Lip
ase
gene
s an
d pr
otei
ns o
f M
ucor
cir
cine
lloi
des
CB
S 27
7.49
Gen
e, G
ene,
Loc
atio
n (s
tran
d), G
ene
Size
, Am
ino
acid
, pI,
Con
serv
ed s
eque
nce
and
Ann
otat
ion
wer
e de
rive
d fr
om th
e Jo
int G
enom
e In
stitu
te (
JGI)
dat
abas
e
Gen
eL
ocat
ion
(Str
and)
Gen
e si
ze
(cD
NA
bps
)Pr
otei
n ID
pIA
min
o ai
cdSu
buni
t (k
Da)
Con
serv
ed s
eque
nce
Ann
otat
ion
Lip
ase
Acy
ltran
sfer
ase
Gen
emar
k1.1
1790
_gsc
affo
ld_1
2:55
4802
-556
327(+
)15
2611
6027
4.8
385
42.6
1G
ASV
GH
DL
RV
DL
ipas
e_G
DSL
Gen
emar
k1.1
0741
_gsc
affo
ld_0
9:29
5517
-296
869(−
)13
5311
4978
4.7
369
40.6
4G
YSK
GH
DL
RM
DL
ipas
e_G
DSL
Muc
ci1.
fgen
eshM
C_p
m.3
_#_2
53sc
affo
ld_0
3:19
7704
1-19
7865
9(−
)16
1972
954
6.2
365
39.9
4G
ISY
G–
Lip
ase_
GD
SL
Muc
ci1.
e_gw
1.2.
1562
.1sc
affo
ld_0
2:22
1965
9-22
2097
8(+
)13
2034
010
6.9
388
44.1
7G
FSQ
G–
α/β
-hyd
rola
se_1
e_gw
1.04
.161
5.1
scaf
fold
_04:
7080
41-7
0949
9(+
)14
5914
3450
5.8
432
48.0
6G
FSQ
Gα
/β-h
ydro
lase
_1
Gen
emar
k1.1
1027
_gsc
affo
ld_1
0:85
859-
8674
3(−
)88
511
5264
5.9
295
33.0
9G
DSA
G–
α/β
-hyd
rola
se_3
Muc
ci1.
fgen
eshM
C_p
g.4_
#_98
2sc
affo
ld_0
4:32
0775
4-32
0895
8(−
)12
0581
713
5.4
342
37.1
4G
DSA
G–
α/β
-hyd
rola
se_3
Gen
emar
k1.8
931_
gsc
affo
ld_0
7:13
1119
-132
507(−
)13
8911
3168
5.5
404
45.4
2G
DSA
G–
α/β
-hyd
rola
se_3
fgen
esh1
_pg.
03_#
_541
scaf
fold
_03:
1866
164-
1867
667(+
)15
0416
1426
6.1
445
49.7
6G
DSA
GH
LYL
DD
α/β
-hyd
rola
se_3
Gen
emar
k1.2
47_g
scaf
fold
_01:
7562
08-7
5835
8 (+
)21
5110
4484
7.8
621
68.7
6G
DSA
GH
GK
MT
Dα
/β-h
ydro
lase
_3
fgen
esh1
_pg.
09_#
_300
scaf
fold
_09:
9666
18-9
6805
0(+
)14
3316
7388
6.4
456
51.2
5G
DSA
G–
α/β
-hyd
rola
se_3
estE
xt_G
enem
ark1
.C_0
8061
0sc
affo
ld_0
8:18
4759
5-18
4884
8(−
)12
5418
5587
7.3
371
41.9
4G
DSA
GH
WIV
KD
α/β
-hyd
rola
se_3
estE
xt_G
enem
ark1
.C_0
5106
7sc
affo
ld_0
5:31
7273
5-31
7493
3(−
)21
9918
4709
8.4
625
69.2
8G
HSL
GH
HH
HQ
Dcl
ass_
3
Muc
ci1.
e_gw
1.5.
1135
.1sc
affo
ld_0
5:32
3892
-325
186(−
)12
9539
585
5.4
337
37.0
2G
TSA
G–
clas
s_3
gw1.
06.1
316.
1sc
affo
ld_0
6:21
9347
8-21
9450
2(+
)10
2513
0354
4.7
305
33.8
5G
TSA
G–
clas
s_3
Gen
emar
k1.1
1524
_gsc
affo
ld_1
1:65
9634
-661
823(−
)21
9011
5761
9.1
622
70.5
5G
LSH
GH
HH
SLD
clas
s_3
Gen
emar
k1.3
176_
gsc
affo
ld_0
2:35
3797
9-35
4005
7(+
)20
7910
7413
7.8
593
67.3
9G
HSL
GH
KG
FWD
clas
s_3
Gen
emar
k1.2
167_
gsc
affo
ld_0
2:53
4294
-535
753(+
)14
6010
6404
7.3
345
38.0
9G
HSY
G–
clas
s_3
gw1.
02.4
71.1
scaf
fold
_02:
5288
13-5
2986
5(−
)10
5311
9940
6.1
234
26.2
8G
HSF
GH
TALY
Dcl
ass_
3
fgen
esh1
_kg.
01_#
_278
_#_1
_0sc
affo
ld_0
1:37
6004
1-37
6074
1(−
)70
115
4931
7.2
219
23.1
4G
HSL
G–
clas
s_3
fgen
esh1
_pg.
08_#
_445
scaf
fold
_08:
1489
434-
1492
137(−
)27
0416
6875
7.6
863
95.6
7G
HSL
GH
DK
AE
Dcl
ass_
3
Muc
ci1.
e_gw
1.2.
509.
1sc
affo
ld_0
2:24
1095
7-24
1237
4(−
)14
1835
076
6.9
353
37.9
6G
HSL
GH
LSY
YD
clas
s_3
fgen
esh1
_kg.
03_#
_293
_#_1
528_
1sc
affo
ld_0
3:32
6364
6-32
6534
6(+
)17
0115
5817
7.9
385
41.1
4G
HSL
GH
FSY
YD
clas
s_3
estE
xt_G
enew
ise1
.C_0
2134
4sc
affo
ld_0
2:27
0999
6-27
1179
5(+
)18
0017
0034
7.6
411
45.2
5G
HSL
G–
clas
s_3
Gen
emar
k1.7
497_
gsc
affo
ld_0
5:22
4416
4-22
4630
8(−
)21
4511
1734
6.2
696
77.9
2G
HSL
GH
SKA
WD
clas
s_3
fgen
esh1
_kg.
02_#
_75_
#_50
1_1
scaf
fold
_02:
9247
29-9
2642
8(−
)17
0015
5202
6.2
396
43.4
2G
HSL
G–
clas
s_3
Gen
emar
k1.7
083_
gsc
affo
ld_0
5:10
1832
4-10
2142
7(+
)31
0411
1320
8.4
935
106.
25G
HSL
GH
KFN
PDcl
ass_
3
Gen
emar
k1.1
0208
_gsc
affo
ld_0
8:88
2079
-885
366(−
)32
8811
4445
8.6
949
107.
58G
HSL
GH
NQ
LFD
clas
s_3
Gen
emar
k1.6
316_
gsc
affo
ld_0
4:30
5847
6-30
6137
0(−
)28
9511
0553
5.2
816
90.9
7G
HSL
GH
NQ
LFD
clas
s_3
fgen
esh1
_pg.
09_#
_158
scaf
fold
_09:
4928
42-4
9570
7(−
)28
6616
7246
5.1
840
93.5
8G
HSL
G–
clas
s_3
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022
1473J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
Evolution of lipases in M. circinelloides CBS 277.49 and WJ11
To investigate the evolutionary relationship of lipase genes, a neighbor-joining tree was generated by align-ing protein sequences of lipases in M. circinelloides CBS 277.49 and WJ11, Tgl3p, Tgl4p and Tgl5p in the yeast S. cerevisiae, and TGL3 and TGL4 in the yeast Y. lipolytica. The phylogram was clustered into three major groups (I, II and III), while the group I appeared earlier by 800 million years than group II and III (Fig. 2). The lipases in group I contain nine sequence variation within the motif GXSXG including GHSLG, G (D/T) SAG, GAS (V/I) G, GLS (S/V) G, GFSQG and GKSDG, whereas these sequence variations appear partly in group II or III. Interestingly, 76 % percentage of lipases in class_3 family and 78 % percentage of lipases in α/β-hydrolase_3 fam-ily fall into group I. The class_3 family showed a closer relationship to the α/β-hydrolase_3 family. This result indicates that class_3 family and α/β-hydrolase_3 family are the conserved lipase family in the fungus M. circinel-loides. The sequence GHSLG is a typical conserved motif in the class_3 family as same as the motif GDSAG in the α/β-hydrolase_3 family. Eighteen pairs of lipases between strain WJ11 and CBS 277.49 show closer evolutionary relationships on the phylogram tree, but further homology analyses based on their amino acid sequences reveal that only ten pairs of lipases have higher identify from 60.8 to 95.7 % with the sequence similarity from 63.1 to 98.5 % (Table 5). This result suggests that most of lipases between strain WJ11 and CBS 277.49 have different amino acid sequences, which may result in different functions of these lipases and associated to the different lipid accumulation. The sequence homology analyses between TGL4 and WJ_1, WJ_25, WJ_26 and Tgl3p, and Tgl4p and 167246 were also performed. TGL4 displayed 9.5 % sequence identify to lipase (WJ_1) and 15 % to lipase (WJ_25). The amino acid sequence identity was 3.7 % (between Tgl3p
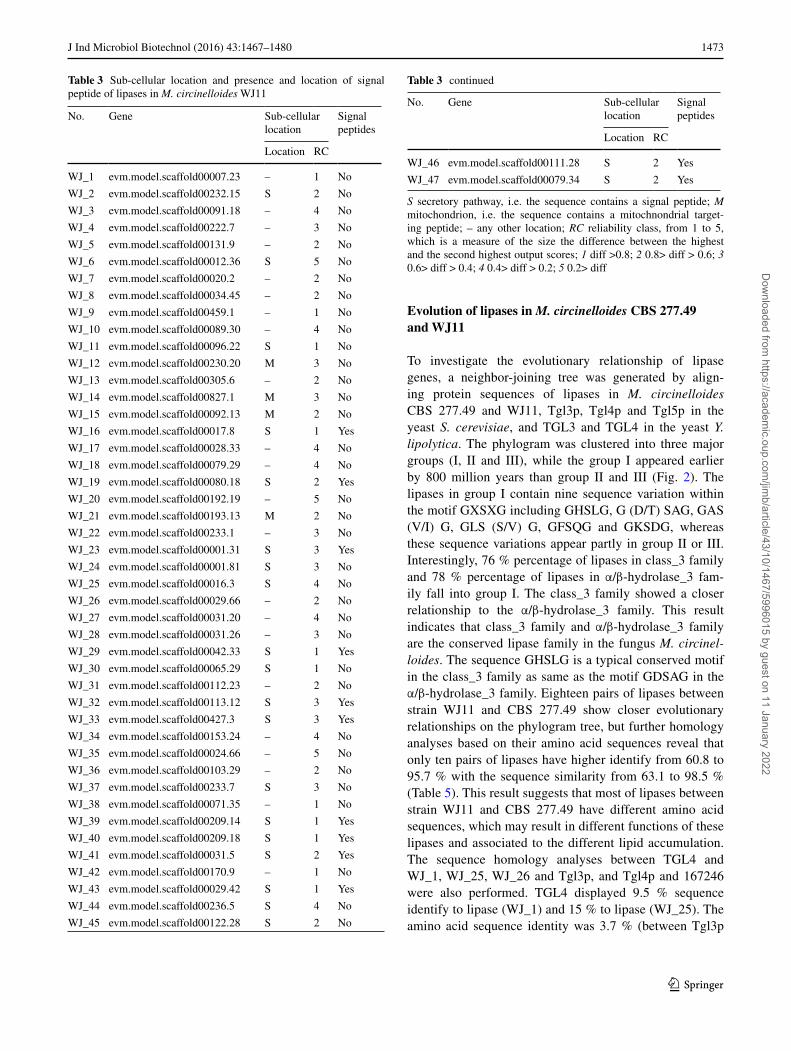
Table 3 Sub-cellular location and presence and location of signal peptide of lipases in M. circinelloides WJ11
No. Gene Sub-cellular location
Signal peptides
Location RC
WJ_1 evm.model.scaffold00007.23 – 1 No
WJ_2 evm.model.scaffold00232.15 S 2 No
WJ_3 evm.model.scaffold00091.18 – 4 No
WJ_4 evm.model.scaffold00222.7 – 3 No
WJ_5 evm.model.scaffold00131.9 – 2 No
WJ_6 evm.model.scaffold00012.36 S 5 No
WJ_7 evm.model.scaffold00020.2 – 2 No
WJ_8 evm.model.scaffold00034.45 – 2 No
WJ_9 evm.model.scaffold00459.1 – 1 No
WJ_10 evm.model.scaffold00089.30 – 4 No
WJ_11 evm.model.scaffold00096.22 S 1 No
WJ_12 evm.model.scaffold00230.20 M 3 No
WJ_13 evm.model.scaffold00305.6 – 2 No
WJ_14 evm.model.scaffold00827.1 M 3 No
WJ_15 evm.model.scaffold00092.13 M 2 No
WJ_16 evm.model.scaffold00017.8 S 1 Yes
WJ_17 evm.model.scaffold00028.33 – 4 No
WJ_18 evm.model.scaffold00079.29 – 4 No
WJ_19 evm.model.scaffold00080.18 S 2 Yes
WJ_20 evm.model.scaffold00192.19 – 5 No
WJ_21 evm.model.scaffold00193.13 M 2 No
WJ_22 evm.model.scaffold00233.1 – 3 No
WJ_23 evm.model.scaffold00001.31 S 3 Yes
WJ_24 evm.model.scaffold00001.81 S 3 No
WJ_25 evm.model.scaffold00016.3 S 4 No
WJ_26 evm.model.scaffold00029.66 – 2 No
WJ_27 evm.model.scaffold00031.20 – 4 No
WJ_28 evm.model.scaffold00031.26 – 3 No
WJ_29 evm.model.scaffold00042.33 S 1 Yes
WJ_30 evm.model.scaffold00065.29 S 1 No
WJ_31 evm.model.scaffold00112.23 – 2 No
WJ_32 evm.model.scaffold00113.12 S 3 Yes
WJ_33 evm.model.scaffold00427.3 S 3 Yes
WJ_34 evm.model.scaffold00153.24 – 4 No
WJ_35 evm.model.scaffold00024.66 – 5 No
WJ_36 evm.model.scaffold00103.29 – 2 No
WJ_37 evm.model.scaffold00233.7 S 3 No
WJ_38 evm.model.scaffold00071.35 – 1 No
WJ_39 evm.model.scaffold00209.14 S 1 Yes
WJ_40 evm.model.scaffold00209.18 S 1 Yes
WJ_41 evm.model.scaffold00031.5 S 2 Yes
WJ_42 evm.model.scaffold00170.9 – 1 No
WJ_43 evm.model.scaffold00029.42 S 1 Yes
WJ_44 evm.model.scaffold00236.5 S 4 No
WJ_45 evm.model.scaffold00122.28 S 2 No
S secretory pathway, i.e. the sequence contains a signal peptide; M mitochondrion, i.e. the sequence contains a mitochnondrial target-ing peptide; – any other location; RC reliability class, from 1 to 5, which is a measure of the size the difference between the highest and the second highest output scores; 1 diff >0.8; 2 0.8> diff > 0.6; 3 0.6> diff > 0.4; 4 0.4> diff > 0.2; 5 0.2> diff
Table 3 continued
No. Gene Sub-cellular location
Signal peptides
Location RC
WJ_46 evm.model.scaffold00111.28 S 2 Yes
WJ_47 evm.model.scaffold00079.34 S 2 Yes
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022
1474 J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
and lipase WJ_1) and 9.3 % (between Tgl4p and lipase 167246). The lower homology indicated that lipases in M. circinelloides CBS 277.49 and WJ11 could be func-tionally different from lipases in yeast S. cerevisiae and Y. lipolytica.
Transcriptional analysis of lipase genes in M. circinelloides WJ11 during growth and lipid accumulation
In oleaginous fungus M. circinelloides CBS 277.49 and WJ11, the initiation of lipid accumulation during lipid syn-thesis is caused by the exhaustion of nitrogen (N) in the cultural medium. So in this study, to explore the potential relationship between the expression of lipase genes and
lipid accumulation in M. circinelloides, we analyze the changes at the transcriptional level of lipase genes before and after N depletion.
Our previous studies have reported characteristics of the growth and lipid accumulation in oleaginous fungi M. cir-cinelloides WJ11 [32]. Briefly, glucose remained in excess during the entire fermentation process and ammonium was used up at approx. 9 h. Cell dry weight initially increased rapidly up to 9 h and then slowed down after nitrogen exhaustion. The fungus started to accumulate lipid rapidly immediately after nitrogen depletion. From 9 to 48 h, the total fatty acids content increased rapidly and then slow down.
For transcriptional analysis, M. circinelloides cells were collected at 6 h (N rich, i.e., balanced growth stage), 24 h
Table 4 Sequences and location of signal peptide of lipases in M. circinelloides CBS 277.49 and WJ11
No. Protein/gene ID Signal peptides
Sequence Length
170034 MVSTSYLISQGLHLCVAVSCLLIHFTDAA 28/29
106404 MIFRNLIILLLATAAIQCQ 18/19
116027 MRLCKQLVGIMAVYGATAW 18/19
WJ_16 evm.model.scaffold00017.8 MCIIAYLCSVLLIPTVMTA 18/19
WJ_40 evm.model.scaffold00209.18 MIFRNLFILLLVVAATQCQ 18/19
WJ_39 evm.model.scaffold00209.14 MLKTCAQLLLIGAILQPALCN 20/21
WJ_43 evm.model.scaffold00029.42 MRLCTKLVGIIAAYGATAWAL 20/21
WJ_29 evm.model.scaffold00042.33 MVSTAYLISQGLHLCVAVSCLLIHFTDAA 28/29
Fig. 1 The transmembrane spanning region of M. circinelloides CBS 277.49 lipases (115761 and 107413) and WJ11 lipases (WJ_2, 5, 11, 16, 25, 26, 30, 37, 38, 44 and 45) were predicted in TMHMM
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022

1475J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
Fig. 2 Phylogenetic analyses of lipases from M. circinelloides CBS 277.49 and WJ11, Tgl3p, Tgl4p and Tgl5p from yeast S. cerevisiae, and TGL3 and TGL4 from yeast Y. lipolytica
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022

1476 J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
(after N depletion, i.e., fast lipid and accumulation stage) and 72 h (after N depletion, i.e., slow lipid accumulation stage), and qRT-PCR was carried out to analyze the tran-scriptional level of lipase genes (Table 6). We applied a fold-change cutoff of ≥1.5 for up-regulation, and ≤0.5 for down-regulation.
The results showed that twenty-one lipase genes in WJ11 were intensely up-regulated at the fast lipid accumu-lation stage (24 h) but slightly up-regulated at the slow lipid accumulation stage (72 h) compared to the balanced growth stage (6 h). The glucose at the fast lipid accumulation stage (24 h) was in excess and enough to provide energy for cell growth, but the question remained why lipase genes were up-regulated at this stage? One possibility might be that fatty acids from TAG degradation induced by lipases may serve primarily for phospholipid synthesis which is vital to maintaining cell growth. Many lipases with acyltransferase activity are prominent contributor to TAG synthesis in the logarithmic growth stage or the exponential growth phase but less crucial under stress conditions or in the stationary phase, such as PDAT in microalga Chlamydomonas rein-hardtii [39] or Tgl3p, Tgl4p and Tgl5p in the yeast S. cer-evisiae [24, 27, 28]. Moreover, Daum et al. demonstrated that the presence of intracellular TAG was important for the acyltransferase activity of these multi-functional lipases [27]. In vitro Tgl3p in S. cerevisiae utilizes acyl-CoAs for efficient acylation of lysophosphatidylethanolamine, indi-cating that fatty acyl activation is required for the phospho-lipid biosynthetic route. Fewer these lipase genes (six) in strain CBS 277.49 possible resulted in the lower content of phospholipid compared with strain WJ11 [40]. Inter-estingly, seven lipase genes in WJ11 were down-regulated at the fast lipid accumulation stage, but were enhanced sharply at the last fermentation stage. These results sug-gested that these lipases were activated under a lower concentration of glucose in the cultural medium, and may be involved in β-oxidation at the later fermentation stage
(72 h). Similarly, Mycobacteriaceae tuberculosis strain lipase gene (F11 TBFG_11087) was predicted to be part of the operon (mtf_TBFG_11089) together with two other genes, TBFG_11089 and TBFG_11088, encoding enoyl-CoA hydrotases, which are involved in β-oxidation of FFA [20]. Fourteen lipase genes in strain WJ11 but five lipase genes in strain CBS 277.49 were down-regulated during the lipid accumulation. The remaining lipase genes in strains WJ11 and CBS 277.49 have no significant changes in the transcriptional level during the entire bioprocess. Similarly, transcriptions of some triacylglycerol lipase genes in the fungus Mortierella alpina were also inhibited during the lipid accumulation [6]. More lipase genes in strain WJ11 were inhibited compared to strain CBS 277.49, which may be associated with high lipid accumulation in WJ11. More-over, this result also supported our previous prediction that lipases (WJ_16, 29, 39 and 40) in WJ11 belong to the fam-ily of secretary lipases except the lipase (WJ_43), because transcriptional levels of extracellular lipase genes should not be triggered by glucose in the culture medium.
Conclusion
In this work we searched all genes encoding lipase in a high lipid-producing strain M. circinelloides WJ11 based on its genome database and compared these genes to a low lipid-producing strain M. circinelloides CBS 277.49, and ana-lyzed several characteristic, sub-cellular location, phyloge-netic analysis and expression profiling of the lipase genes during growth and lipid accumulation. Our results reveal that all of these proteins contain the typical lipase motif of GXSXG and were divided into four types including α/β-hydrolase_1, α/β-hydrolase_3, class_3 and GDSL. Inter-estingly, some lipases also contain a typical acyltransferase motif of H-(X) 4-D, and these lipases may play a dual role in lipid metabolism, catalyzing both lipid hydrolysis and
Table 5 Homology analyses of lipases between strain WJ11 and CBS 277.49 based on their amino acid sequences
WJ11 CBS 277.49 Amino sequence
No. Gene ID Protein ID Identify (%) Similarity (%)
WJ_30 evm.model.scaffold00065.29 107413 92.7 95.4
WJ_43 evm.model.scaffold00029.42 116027 91.1 97.1
WJ_35 evm.model.scaffold00024.66 111734 72 76.6
WJ_31 evm.model.scaffold00112.23 166875 92.4 96.2
WJ_19 evm.model.scaffold00080.18 185587 88.9 94.3
WJ_20 evm.model.scaffold00192.19 113168 89.8 95.5
WJ_7 evm.model.scaffold00020.2 81713 93 96.5
WJ_29 evm.model.scaffold00042.33 170034 60.8 63.1
WJ_24 evm.model.scaffold00001.81 155202 95.7 98.5
WJ_40 evm.model.scaffold00209.18 106404 79.7 85.7
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022

1477J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
Table 6 The transcriptional levels of lipase genes in strains WJ11 and CBS 277.49 were analyzed by qRT-PCR at 6, 24 and 72 h
Strain No. Gene/protein ID The transcriptional level (h)
6 24 72
WJ11 WJ_1 evm.model.scaffold00007.23 1.00 3.07 ± 0.06a 1.72 ± 0.23b
WJ_2 evm.model.scaffold00232.15 1.00 2.66 ± 0.13a 0.99 ± 0.48b
WJ_3 evm.model.scaffold00091.18 1.00 0.74 ± 0.13b 3.59 ± 0.48a
WJ_4 evm.model.scaffold00222.7 1.00 0.51 ± 0.06b 0.07 ± 0.01a
WJ_6 evm.model.scaffold00012.36 1.00 2.28 ± 0.42b 1.35 ± 0.25b
WJ_7 evm.model.scaffold00020.2 1.00 0.16 ± 0.03a 0.03 ± 0.01a
WJ_8 evm.model.scaffold00034.45 1.00 0.05 ± 0.01a 0.22 ± 0.04a
WJ_9 evm.model.scaffold00459.1 1.00 1.82 ± 0.01b 0.42 ± 0.04a
WJ_10 evm.model.scaffold00089.30 1.00 0.19 ± 0.07b 2.85 ± 0.25b
WJ_11 evm.model.scaffold00096.22 1.00 5.90 ± 0.01b 0.55 ± 0.04b
WJ_12 evm.model.scaffold00230.20 1.00 0.17 ± 0.04b 0.81 ± 0.19a
WJ_13 evm.model.scaffold00305.6 1.00 6.77 ± 0.26b 4.06 ± 0.25b
WJ_14 evm.model.scaffold00827.1 1.00 2.64 ± 0.01b 1.75 ± 0.03a
WJ_15 evm.model.scaffold00092.13 1.00 1.51 ± 0.01a 1.65 ± 0.17a
WJ_16 evm.model.scaffold00017.8 1.00 0.45 ± 0.03b 1.40 ± 0.06a
WJ_17 evm.model.scaffold00028.33 1.00 2.68 ± 0.13b 2.11 ± 0.48b
WJ_18 evm.model.scaffold00079.29 1.00 7.36 ± 0.06b 4.00 ± 0.01b
WJ_19 evm.model.scaffold00080.18 1.00 3.12 ± 0.07b 5.13 ± 0.13b
WJ_20 evm.model.scaffold00192.19 1.00 1.34 ± 0.26a 10.93 ± 2.11b
WJ_21 evm.model.scaffold00193.13 1.00 0.05 ± 0.01b 0.15 ± 0.03b
WJ_22 evm.model.scaffold00233.1 1.00 0.07 ± 0.01b 0.89 ± 0.17a
WJ_23 evm.model.scaffold00001.31 1.00 5.82 ± 0.03b 10.34 ± 0.01b
WJ_24 evm.model.scaffold00001.81 1.00 0.20 ± 0.08b 0.15 ± 0.02b
WJ_25 evm.model.scaffold00016.3 1.00 0.36 ± 0.06b 2.33 ± 0.23b
WJ_26 evm.model.scaffold00029.66 1.00 4.38 ± 0.13b 5.06 ± 0.48b
WJ_27 evm.model.scaffold00031.20 1.00 2.48 ± 0.06b 3.78 ± 0.01b
WJ_28 evm.model.scaffold00031.26 1.00 3.66 ± 0.07b 4.35 ± 0.13b
WJ_29 evm.model.scaffold00042.33 1.00 0.01 ± 0.00b 0.01 ± 0.00b
WJ_30 evm.model.scaffold00065.29 1.00 0.06 ± 0.01b 0.16 ± 0.03b
WJ_31 evm.model.scaffold00112.23 1.00 0.64 ± 0.12a 2.53 ± 0.46b
WJ_32 evm.model.scaffold00113.12 1.00 0.12 ± 0.05b 0.56 ± 0.10a
WJ_33 evm.model.scaffold00427.3 1.00 4.03 ± 0.37b 1.77 ± 0.76a
WJ_34 evm.model.scaffold00153.24 1.00 0.14 ± 0.03b 3.82 ± 0.70b
WJ_35 evm.model.scaffold00024.66 1.00 2.04 ± 0.08b 1.15 ± 0.02a
WJ_37 evm.model.scaffold00233.7 1.00 3.03 ± 0.31b 2.85 ± 0.03b
WJ_38 evm.model.scaffold00071.35 1.00 1.11 ± 0.14a 2.57 ± 0.28b
WJ_39 evm.model.scaffold00209.14 1.00 0.17 ± 0.03b 0.53 ± 0.06a
WJ_42 evm.model.scaffold00170.9 1.00 2.13 ± 0.08b 1.85 ± 0.02b
WJ_43 evm.model.scaffold00029.42 1.00 1.87 ± 0.06a 2.51 ± 0.23b
WJ_44 evm.model.scaffold00236.5 1.00 0.62 ± 0.08a 0.30 ± 0.02b
WJ_45 evm.model.scaffold00122.28 1.00 0.28 ± 0.03b 0.08 ± 0.01b
WJ_46 evm.model.scaffold00111.28 1.00 0.37 ± 0.08b 0.14 ± 0.02b
WJ_47 evm.model.scaffold00079.34 1.00 1.57 ± 0.08a 1.03 ± 0.02a
CBS 277.49 114978 1.00 0.14 ± 0.05b 0.26 ± 0.14b
72954 1.00 2.25 ± 0.34b 1.01 ± 0.23a
34010 1.00 2.87 ± 0.56b 1.36 ± 0.21b
143450 1.00 0.78 ± 0.08b 0.29 ± 0.11b
115264 1.00 0.56 ± 0.21b 0.44 ± 0.12b
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022
1478 J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
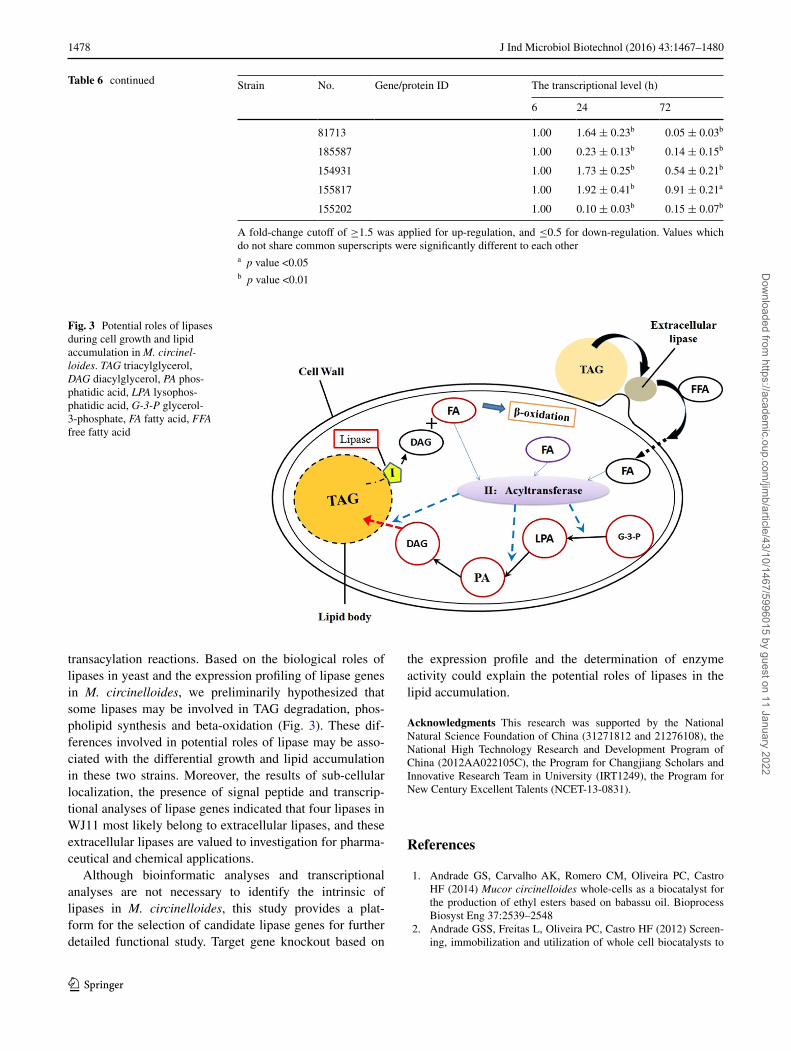
transacylation reactions. Based on the biological roles of lipases in yeast and the expression profiling of lipase genes in M. circinelloides, we preliminarily hypothesized that some lipases may be involved in TAG degradation, phos-pholipid synthesis and beta-oxidation (Fig. 3). These dif-ferences involved in potential roles of lipase may be asso-ciated with the differential growth and lipid accumulation in these two strains. Moreover, the results of sub-cellular localization, the presence of signal peptide and transcrip-tional analyses of lipase genes indicated that four lipases in WJ11 most likely belong to extracellular lipases, and these extracellular lipases are valued to investigation for pharma-ceutical and chemical applications.
Although bioinformatic analyses and transcriptional analyses are not necessary to identify the intrinsic of lipases in M. circinelloides, this study provides a plat-form for the selection of candidate lipase genes for further detailed functional study. Target gene knockout based on
the expression profile and the determination of enzyme activity could explain the potential roles of lipases in the lipid accumulation.
Acknowledgments This research was supported by the National Natural Science Foundation of China (31271812 and 21276108), the National High Technology Research and Development Program of China (2012AA022105C), the Program for Changjiang Scholars and Innovative Research Team in University (IRT1249), the Program for New Century Excellent Talents (NCET-13-0831).
References
1. Andrade GS, Carvalho AK, Romero CM, Oliveira PC, Castro HF (2014) Mucor circinelloides whole-cells as a biocatalyst for the production of ethyl esters based on babassu oil. Bioprocess Biosyst Eng 37:2539–2548
2. Andrade GSS, Freitas L, Oliveira PC, Castro HF (2012) Screen-ing, immobilization and utilization of whole cell biocatalysts to
A fold-change cutoff of ≥1.5 was applied for up-regulation, and ≤0.5 for down-regulation. Values which do not share common superscripts were significantly different to each othera p value <0.05b p value <0.01
Table 6 continued Strain No. Gene/protein ID The transcriptional level (h)
6 24 72
81713 1.00 1.64 ± 0.23b 0.05 ± 0.03b
185587 1.00 0.23 ± 0.13b 0.14 ± 0.15b
154931 1.00 1.73 ± 0.25b 0.54 ± 0.21b
155817 1.00 1.92 ± 0.41b 0.91 ± 0.21a
155202 1.00 0.10 ± 0.03b 0.15 ± 0.07b
Fig. 3 Potential roles of lipases during cell growth and lipid accumulation in M. circinel-loides. TAG triacylglycerol, DAG diacylglycerol, PA phos-phatidic acid, LPA lysophos-phatidic acid, G-3-P glycerol-3-phosphate, FA fatty acid, FFA free fatty acid
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022

1479J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
mediate the ethanolysis of babassu oil. J Mol Catal B Enzym 84:183–188
3. Antczak T (2001) Catalytic properties of fungal lipases from Mucor Circinelloides and Mucor Racemosus. Zeszyty Naukowe. Rozprawy Naukowe/Politechnika Łódzka 298:5–169
4. Bigey F, Bougard TD, Nicaud JM, Moulin G (2003) Identifica-tion of a triacylglycerol lipase gene family in Candida defor-mans: molecular cloning and functional expression. Yeast 20:233–248
5. Carvalho AKF, Faria ELP, Rivaldi JD, Andrade GSS, Oliveira PC (2015) Performance of whole-cells lipase derived from Mucor circinelloides as a catalyst in the ethanolysis of non-edible veg-etable oils under batch and continuous run conditions. Ind Crops Prod 67:287–294
6. Chen H, Hao G, Wang L, Wang H, Gu Z (2014) Identification of a critical determinant that enables efficient fatty acid synthesis in oleaginous fungi. Sci Rep 5:11247. doi:10.1038/srep11247
7. Desfougeres T, Haddouche R, Fudalej F, Neuveglise C, Nicaud JM (2010) SOA genes encode proteins controlling lipase expres-sion in response to triacylglycerol utilization in the yeast Yar-rowia lipolytica. FEMS Yeast Res 10:93–103
8. Destain J, Fickers P, Weekers F, Moreau B, Thonart P (2005) Utilization of methyloleate in production of microbial lipase. Appl Biochem Biotechnol 121–124:269–277
9. Dulermo T, Tréton B, Beopoulos A, Gnankon APK, Haddouche R (2013) Characterization of the two intracellular lipases of Y. lipolytica encoded by TGL3 and TGL4 genes: new insights into the role of intracellular lipases and lipid body organisation. Bio-chim Biophys Acta 1831:1486–1495
10. Fickers P, Marty A, Nicaud JM (2011) The lipases from Yar-rowia lipolytica: genetics, production, regulation, biochemical characterization and biotechnological applications. Biotechnol Adv 29:632–644
11. Fickers P, Nicaud JM, Gaillardin C, Destain J, Thonart P (2004) Carbon and nitrogen sources modulate lipase production in the yeast Yarrowia lipolytica. J Appl Microbiol 96:742–749
12. Garcia-Galan C, Barbosa O, Ortiz C, Torres R, Rodrigues RC (2013) Biotechnological prospects of the lipase from Mucor javanicus. J Mol Catal B Enzym 93:34–43
13. Goncalves C, Lopes M, Ferreira JP, Belo I (2009) Biological treatment of olive mill wastewater by non-conventional yeasts. Bioresour Technol 100:3759–3763
14. Guieysse D, Sandoval G, Faure L, Nicaud JM, Monsan P (2005) New efficient lipase from Yarrowia lipolytica for the resolu-tion of 2-bromo-arylacetic acid esters. Tetrahedron Asymmetry 15(22):3539–3543
15. Ham HJ, Rho HJ, Shin SK, Yoon HJ (2010) The TGL2 gene of Saccharomyces cerevisiae encodes an active acylglycerol lipase located in the mitochondria. J Biol Chem 285:3005–3013
16. Holmes RS, Cox LA, VandeBerg JL (2010) Comparative stud-ies of mammalian acid lipases: evidence for a new gene family in mouse and rat (Lipo). Comp Biochem Physiol Part D Genom Proteom 5:217–226
17. Kamzolova SV, Finogenova TV, Lunina YN, Perevoznikova OA, Minachova LN (2007) Characteristics of the growth on rapeseed oil and synthesis of citric and isocitric acids by Yarrowia lipol-ytica yeasts. Microbiology 76:20–24
18. Kendrick A, Ratledge C (1992) Desaturation of polyunsatu-rated fatty acids in Mucor circinelloides and the involvement of a novel membrane-bound malic enzyme. Eur J Biochem 209:667–673
19. Kurat CF, Natter K, Petschnigg J, Wolinski H, Scheuringer K (2006) Obese yeast: triglyceride lipolysis is functionally con-served from mammals to yeast. J Biol Chem 281:491–500
20. Lazniewski M, Steczkiewicz K, Knizewski L, Wawer I, Ginalski K (2011) Novel transmembrane lipases of alpha/beta hydrolase fold. FEBS Lett 585:870–874
21. Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25:402–408
22. Pleiss J, Scheib H, Schmid RD (2000) The His gap motif in microbial lipases: a determinant of stereoselectivity toward tria-cylglycerols and analogs. Biochimie 82:1043–1052
23. Purwanto MGM, Maretha MV, Wahyudi M, Goeltom MT (2015) Whole cell hydrolysis of sardine (Sardinella Lemuru) oil waste using Mucor Circinelloides NRRL 1405 immobilized in poly-urethane foam. Procedia Chem 14:256–262
24. Rajakumari S, Daum G (2009) Janus-faced enzymes yeast Tgl3p and Tgl5p catalyze lipase and acyltransferase reactions. Mol Biol Cell 21:501–510
25. Rajendran A, Palanisamy A, Thangavelu V (2009) Lipase cata-lyzed ester synthesis for food processing industries. Braz Arch Biol Technol 52:207–219
26. Rosa Amarilis RF, Adrián G, Santiago TM, Victoriano G (2012) Malic enzyme activity is not the only bottleneck for lipid accu-mulation in the oleaginous fungus Mucor circinelloides. Appl Microbiol Biotechnol 97:3063–3072
27. Schmidt C, Athenstaedt K, Koch B, Ploier B, Daum G (2013) Regulation of the yeast triacylglycerol lipase TGl3p by forma-tion of nonpolar lipids. J Biol Chem 288:19939–19948
28. Sona R, Günther D (2010) Multiple functions as lipase, steryl ester hydrolase, phospholipase, and acyltransferase of Tgl4p from the yeast Saccharomyces cerevisiae. J Biol Chem 285:15769–15776
29. Suzzi G, Lanorte MT, Galgano F, Andrighetto C, Lombardi A (2001) Proteolytic, lipolytic and molecular characterisation of Yarrowia lipolytica isolated from cheese. Int J Food Microbiol 69:69–77
30. Szczesna-Antczak M, Antczak T, Piotrowicz-Wasiak M, Rzyska M, Binkowska N, Bielecki S (2006) Relationships between lipases and lipids in mycelia of two Mucor strains. Enzyme Microbial Technol 39:1214–1222
31. Szczesna-Antczak M, Antczak T, Rzyska M, Modrzejewska Z, Patura J, Kalinowska H, Bielecki S (2004) Stabilization of an intracellular Mucor circinelloides lipase for application in non-aqueous media. J Mol Catal B Enzym 29:163–171
32. Tang X, Chen HQ, Chen YQ, Chen W, Victoriano G (2015) Com-parison of biochemical activities between high and low lipid-producing strains of Mucor circinelloides: an explanation for the high oleaginicity of strain WJ11. PLoS One 10(9):e0137543
33. Tang X, Zhao LN, Chen HQ, Chen YQ, Chen W, Song YD, Ratledge C (2015) Complete genome sequence of a high lipid-producing strain of Mucor circinelloides WJ11 and comparative genome analysis with a low lipid-producing strain CBS 277.49. PLoS One 10(9):e0137543. doi:10.1371/journal.pone.0137543
34. Tang X, Zan X, Zhao L, Chen H, Chen YQ, Chen W, Song Y, Ratledge C (2016) Proteomics analysis of high lipid-producing strain Mucor circinelloides WJ11: an explanation for the mecha-nism of lipid accumulation at the proteomic level. Microb Cell Fact 11:15–35
35. Turki S, Mrabet JG, Marouani A, Thonart P (2010) Preliminary safety assessment of Yarrowia lipolytica extracellular lipase: results of acute and 28-day repeated dose oral toxicity studies in rats. Food Chem Toxicol 48:2393–2400
36. Vongsangnak W, Klanchui A, Tawornsamretkit L, Tatiyabor-wornchai W, Laoteng K, Meechai A (2016) Genome-scale meta-bolic modeling of Mucor circinelloides and comparative analysis with other oleaginous species. Gene 583(2):121–129
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022

1480 J Ind Microbiol Biotechnol (2016) 43:1467–1480
1 3
37. Yazawa H, Kumagai H, Uemura H (2012) Characterization of triglyceride lipase genes of fission yeast Schizosaccharomyces pombe. Appl Microbiol Biotechnol 96:981–991
38. Ying Z, Adams IP, Colin R (2007) Malic enzyme: the controlling activity for lipid production? Overexpression of malic enzyme in Mucor circinelloides leads to a 2.5-fold increase in lipid accu-mulation. Microbiology 153:2013–2025
39. Yoon K, Han D, Li Y, Sommerfeld M, Hu Q (2012) Phospho-lipid: diacylglycerol acyltransferase is a multifunctional enzyme involved in membrane lipid turnover and degradation while
synthesizing triacylglycerol in the unicellular green microalga Chlamydomonas reinhardtii. Plant Cell 24:3708–3724
40. Zan XY, Tang X, Zhao LN, Chu LF, Chen HQ, Chen W, Chen YQ, Song YD (2016) Bioinformatical analysis and preliminary study of the role of lipase in lipid metabolism in Mucor circinel-loides. RSC Adv 6:60673–60682
41. Zhao LN, Tang X, Luan X, Chen HQ, Chen YQ, Chen W, Song YD, Ratledge C (2015) Role of pentose phosphate pathway in lipid accumulation of oleaginous fungus Mucor circinelloides. RSC Adv 5:97658–97664
Dow
nloaded from https://academ
ic.oup.com/jim
b/article/43/10/1467/5996015 by guest on 11 January 2022
Related Documents











