MARINE ECOLOGY PROGRESS SERIES Mar Ecol Prog Ser Vol. 290: 207–221, 2005 Published April 13 INTRODUCTION Body size and maximum life-span are correlated for a broad range of organisms, including mammals, birds and invertebrates, and in general maximum life-span increases with increasing body size (Blueweiss et al. 1978, Stearns 1992). Additionally, Charnov (1993) ob- served that, as an important invariant of life history, growth rate is inversely related to asymptotic size. Therefore, small species are typically expected to exhibit short life-spans and fast growth rates, whereas large species are expected to have greater life-spans and slow growth. Whilst many small species of fish are indeed short-lived, some small temperate and tropical species are actually relatively long-lived (Miller 1996, Munday & Jones 1998), therefore the applicability of general life-history predictions to small reef fish re- quires further investigation. The available data suggest that a positive relation- ship between size and maximum age may not be uni- versally applicable to coral reef fishes (Choat & Robert- son 2002). Firstly, many small species, especially some pomacentrids (e.g. Hill & Radtke 1988, Worthington et al. 1995, Meekan et al. 2001) and small acanthurids (e.g. Choat & Axe 1996, Hart & Russ 1996), are long- lived. Secondly, species with similar maximum ages can differ in maximum size by an order of magnitude (Choat & Robertson 2002). Lastly, within-taxon com- parisons indicate that a positive relationship between size and maximum life-span may not always apply. For example, several small species of acanthurids have greater maximum ages than much larger species in the © Inter-Research 2005 · www.int-res.com *Email: [email protected] Life-history characteristics of coral reef gobies. I. Growth and life-span V. Hernaman 1, 3, * , P. L. Munday 2 1 Department of Marine Science, University of Otago, PO Box 56, Dunedin, New Zealand 2 School of Marine Biology and Aquaculture, James Cook University, Townsville, Queensland, Australia 3 Present address: School of Biological Sciences, Victoria University, PO Box 600, Wellington, New Zealand ABSTRACT: Life-history theory predicts that small species will exhibit short life-spans and fast growth rates; however, previous studies indicate that a positive relationship between size and maxi- mum age may not be universally applicable to coral reef fishes. Here, we investigate the growth and life-span of 5 small species of coral reef goby (family Gobiidae): Istigobius goldmanni, Asterropteryx semipunctatus, Amblygobius bynoensis, Amblygobius phalaena and Valenciennea muralis. All 5 species were relatively short-lived, with the oldest individual sampled ranging from 11 to 16 mo depending on species and sex. Rapid growth occurred over much of the size range of all 5 species and, in contrast to most reef fishes, relatively little or no time was spent at an asymptotic size. Patterns of growth were best described by a Broken Stick model for I. goldmanni, and by either a Broken Stick model or the von Bertalanffy growth function for the other 4 species. Summer-growing individuals had higher growth rates than winter-growing individuals, but this did not affect the overall patterns of growth. Sex-specific differences in growth were evident for I. goldmanni and A. semipunctatus, with males growing faster and attaining a larger maximum size than females. In contrast, there was no significant difference in growth between male and female A. bynoensis, A. phalaena and V. muralis. This pattern may be related to interspecific differences in intensity of sexual selection, mat- ing system, and reproductive behaviour. Overall, the patterns of growth and life-span of these 5 small species conformed to traditional concepts of life-history theory. KEY WORDS: Longevity · Growth · Body size · Life history · Gobiidae · Otolith Resale or republication not permitted without written consent of the publisher

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MARINE ECOLOGY PROGRESS SERIESMar Ecol Prog Ser
Vol. 290: 207–221, 2005 Published April 13
INTRODUCTION
Body size and maximum life-span are correlated fora broad range of organisms, including mammals, birdsand invertebrates, and in general maximum life-spanincreases with increasing body size (Blueweiss et al.1978, Stearns 1992). Additionally, Charnov (1993) ob-served that, as an important invariant of life history,growth rate is inversely related to asymptotic size.Therefore, small species are typically expected toexhibit short life-spans and fast growth rates, whereaslarge species are expected to have greater life-spansand slow growth. Whilst many small species of fish areindeed short-lived, some small temperate and tropicalspecies are actually relatively long-lived (Miller 1996,Munday & Jones 1998), therefore the applicability of
general life-history predictions to small reef fish re-quires further investigation.
The available data suggest that a positive relation-ship between size and maximum age may not be uni-versally applicable to coral reef fishes (Choat & Robert-son 2002). Firstly, many small species, especially somepomacentrids (e.g. Hill & Radtke 1988, Worthington etal. 1995, Meekan et al. 2001) and small acanthurids(e.g. Choat & Axe 1996, Hart & Russ 1996), are long-lived. Secondly, species with similar maximum agescan differ in maximum size by an order of magnitude(Choat & Robertson 2002). Lastly, within-taxon com-parisons indicate that a positive relationship betweensize and maximum life-span may not always apply. Forexample, several small species of acanthurids havegreater maximum ages than much larger species in the
© Inter-Research 2005 · www.int-res.com*Email: [email protected]
Life-history characteristics of coral reef gobies. I. Growth and life-span
V. Hernaman1, 3,*, P. L. Munday2
1Department of Marine Science, University of Otago, PO Box 56, Dunedin, New Zealand2School of Marine Biology and Aquaculture, James Cook University, Townsville, Queensland, Australia
3Present address: School of Biological Sciences, Victoria University, PO Box 600, Wellington, New Zealand
ABSTRACT: Life-history theory predicts that small species will exhibit short life-spans and fastgrowth rates; however, previous studies indicate that a positive relationship between size and maxi-mum age may not be universally applicable to coral reef fishes. Here, we investigate the growth andlife-span of 5 small species of coral reef goby (family Gobiidae): Istigobius goldmanni, Asterropteryxsemipunctatus, Amblygobius bynoensis, Amblygobius phalaena and Valenciennea muralis. All 5species were relatively short-lived, with the oldest individual sampled ranging from 11 to 16 modepending on species and sex. Rapid growth occurred over much of the size range of all 5 speciesand, in contrast to most reef fishes, relatively little or no time was spent at an asymptotic size. Patternsof growth were best described by a Broken Stick model for I. goldmanni, and by either a Broken Stickmodel or the von Bertalanffy growth function for the other 4 species. Summer-growing individualshad higher growth rates than winter-growing individuals, but this did not affect the overall patternsof growth. Sex-specific differences in growth were evident for I. goldmanni and A. semipunctatus,with males growing faster and attaining a larger maximum size than females. In contrast, there wasno significant difference in growth between male and female A. bynoensis, A. phalaena and V.muralis. This pattern may be related to interspecific differences in intensity of sexual selection, mat-ing system, and reproductive behaviour. Overall, the patterns of growth and life-span of these 5 smallspecies conformed to traditional concepts of life-history theory.
KEY WORDS: Longevity · Growth · Body size · Life history · Gobiidae · Otolith
Resale or republication not permitted without written consent of the publisher

Mar Ecol Prog Ser 290: 207–221, 2005
same family (Choat & Robertson 2002), and 3 closelyrelated species of lutjanids that have similar maximumsize differ in maximum age by 12 to 19 yr (Newman etal. 1996, 2000). Therefore, size–age relationships maybe very complex for coral reef fishes, and more infor-mation is required on the growth and life-span of smallspecies to determine to what extent they conform tolife-history predictions and generalisations (Munday &Jones 1998).
An ideal group in which to test life-history predic-tions for small reef fishes is the family Gobiidae (go-bies), which is dominated by species with a maximumtotal length of less than 100 mm. Despite being amongthe most diverse and abundant group of coral reeffishes (Leis & Rennis 1983, Randall et al. 1997, Acker-man & Bellwood 2000), very little is known aboutgrowth rates, patterns of growth and life-span of tropi-cal gobies. Available data suggest that coral reefgobies inhabiting burrows or living in sand/rock habi-tats may be relatively short-lived (<2 yr; Yanagisawa1982, Reavis & Barlow 1998, Kritzer 2002). However,more cryptic species, such as coral-dwelling gobies,appear to have longer life-spans (e.g. >4 yr; Munday2001). Therefore, the type of habitat utilised and itsassociated predation risk might make an importantcontribution to the relationship between life-span andbody size, as has been highlighted for temperate go-bies (Miller 1984) and suggested for tropical gobies(Munday & Jones 1998).
We investigated growth rates and life-span of 5 com-mon coral reef gobies (Asterropteryx semipunctatus,Istigobius goldmanni, Amblygobius bynoensis, Ambly-gobius phalaena and Valenciennea muralis) and com-pared them with predictions from life-history theory.The coral reef gobies in our study are all small species,but the maximum size differs between species, rangingfrom 45 to 120 mm total length. These species also dif-fer in their patterns of habitat use, being foundamongst coral rubble, in sand/rock habitats or in bur-rows. Thus, these 5 species provided an ideal opportu-nity to compare patterns of growth and maximum life-span in relation to both body size and habitat use.
Growth models are often used to describe patterns ofgrowth, and consideration must be given to choosingan appropriate model. Although often subject to criti-cism (e.g. Roff 1980), the von Bertalanffy growth func-tion (VBGF), which describes a growth pattern charac-terised by rapid initial growth that slows towards anasymptotic size, is the most widely used growth curvein the fisheries literature (Chen et al. 1992). The VBGFhas been commonly used to describe the growthpatterns of long-lived coral reef fishes (e.g. Hart & Russ1996, Newman et al. 1996, Gust et al. 2002) that tend tohave a relatively brief period of rapid growth followedby an extended period at asymptotic size. However,
short-lived fishes might tend to have a prolongedperiod of rapid growth with relatively little time spentat asymptotic size, and therefore models other than theVBGF might best describe their pattern of growth. Forthis reason we used a range of growth models to deter-mine the most appropriate descriptor of growth in eachof the 5 goby species. Furthermore, males and femalesoften grow at different rates (e.g. Gladstone & Westoby1988, Newman et al. 1996, McCormick 1998) and theirpatterns of growth may be best described by differentgrowth models. Therefore, growth rates, life-span andgrowth models are examined separately for males andfemales.
Specifically, we addressed 3 questions for each of the5 coral reef gobies considered here: (1) What is themaximum life-span, (2) what is the pattern of growthand which is the most appropriate growth model fordescribing that growth pattern, and (3) are there sex-specific differences in growth rates?
MATERIALS AND METHODS
Collections. Specimens of each species were col-lected at intervals from August 1997 to December 1998from the reef flat at Pioneer Bay, Orpheus Island, Aus-tralia (18°35’ S, 146°29’ E). Fish were caught usinghandnets and a clove oil/alcohol mixture (Munday &Wilson 1997) that caused short-term anaesthesia. Fishwere sacrificed by immersion in an ice slurry, blotteddry, measured (total length [TL] and standard length[SL] to the nearest 0.1 mm) and weighed (wet weight tothe nearest 0.001 g). The head was severed from thebody and stored in 70% ethanol. The body, includingthe reproductive organs, was placed in a solution of4% formaldehyde, 5% glacial acetic acid and 1.3%calcium chloride (FAACC) for at least 1 wk and thentransferred to 70% ethanol.
Length–weight relationship. To enable sex-specificand interspecific comparisons of growth properties, therelationship between fish length and fish weight wasestimated by a power curve, W = aLb, where W isweight (g), L is length (TL, mm) and a and b are con-stants.
Estimation of age using counts of otolith growthincrements. Sagittal otoliths were extracted from thebrain cavity, cleaned, and stored dry in multiwell cul-ture plates until required. To enable enumeration ofgrowth increments, a transverse section of eachotolith was prepared. Using thermoplastic glue (crys-talbond) as the mounting medium, the otolith waspositioned on the end of a glass slide, with the otolithcore on the inner edge of the slide. Holding the slideperpendicular to the sand paper, the otolith was thenhand-ground using 800 grit sand paper. The crystal-
208

Hernaman & Munday: Coral reef gobies. I
bond was softened by reheating on a hotplate and theotolith was repositioned to the middle of a slide withthe ground side facing down. The otolith was thenhand-ground using 800 grit sand paper and 12 µmlapping film until a thin section was obtained thatallowed the growth increments to be counted fromthe core to the edge.
Otolith increments have a daily periodicity in each ofthe study species (Hernaman et al. 2000), thereforecounts of increments provided an estimate of age indays. Counts were conducted using a compoundmicroscope fitted with a polarizing filter. Counts weremade on 3 separate occasions, with each count beingconducted without reference to previous counts, andthe average of the 3 counts used as the age estimate.The otolith was not used if the counts differed by>10%. To minimise interpretation error (Campana2001), 2 additional experienced readers countedgrowth increments on a subset of the sample, includ-ing those of Asterropteryx semipunctatus which werethe most difficult to read. To examine the precision ofage estimates, the coefficient of variation (CV) wasdetermined for each individual, from which a mean CVwas calculated for each species (Campana 2001).
Maximum life-span. The maximum life-span wasdefined as the age of the oldest fish collected.
Growth pattern. To examine patterns and rates ofgrowth, size-at-age plots were constructed for eachspecies where the age estimate (d) for each individualwas plotted against its total length (mm). Twoapproaches were taken to examine the pattern ofgrowth for each species. Firstly, 4 different growthmodels were fitted to the size-at-age data, and 3 mea-sures of goodness-of-fit compared for each model: (1)values of the residual sum of squares (RSS), (2) coeffi-cient of determination (r2) calculated from the residualand explained sums of squares, and (3) plots of theresiduals against age. Secondly, Schnute's (1981)model was fitted to the size-at-age data, and the mostappropriate growth model selected according to theestimated values of the model parameters, a and b.For each species, the value of parameter a was a smallpositive or negative value. Whether the value wasinterpreted as being equal to zero affected the selec-tion of the most appropriate model. To test the nullhypothesis that a = 0, Schnute (1981) suggests fitting asecond growth model (which does not incorporate theparameter a) to the data and examining the varianceratio defined by the residual sum of squares for Model2 divided by the sum of squares for Model 1. The var-iance ratio is approximately F-distributed with theappropriate degrees of freedom under the null hypo-thesis that a = 0. Therefore, if the F-statistic is greaterthan the variance ratio, the null hypothesis is re-tained.
We used 2 models that can describe asymptoticgrowth, the VBGF:
Lt = L∞ (1–e–k(t–t0))
and the Richards growth model:
Lt = L∞ (1 + [1/p] e–k(t–t0))–p
where Lt is mean length at age t, L∞ is the mean asymp-totic length, k is the growth coefficient that describesthe rate at which the asymptotic size (L∞) is approached,and p is the shape parameter for the Richards model.For the VBGF, t0 is the theoretical age at which length iszero, but for the S-shaped Richards growth model t0
corresponds to an inflection point on the curve. These 2models were chosen because they describe differentpatterns of growth in the early life stages. The VBGFdescribes an initial period of rapid growth that thenreaches an asymptote, whereas the Richards model de-scribes an S-shaped growth curve with the most rapidgrowth appearing in the middle of the growth pattern.This type of growth could occur when newly settled fishhave a period of slow growth following settlement.
We also used 2 non-asymptotic growth models, thePower curve (described as: Lt = a[tb], where a and bare constants, and Lt is mean length at age t), and theBroken Stick regression (sometimes referred to asBreakpoint regression) which fits 2 linear models to thedata by determining a breakpoint, x0. All fish aged <x0
have the equation y = a + bx fitted to the size-at-agedata, whilst all fish aged >x0 have the equation y = c +dx fitted. The parameters a and c are points on the y-axis where the 2 linear models cross the y-axis, b and ddescribe the slope of each linear regression. Thebreakpoint, x0, is defined as: x0 = (a – c)/(d – b). TheBroken Stick regression was selected in place of a lin-ear model because, even for species that had indeter-minate adult growth, the rate of adult growth wasslower than for the early life-stages, meaning that 1linear model could not adequately describe both juve-nile and adult growth. The Broken Stick regressionwas fitted to the size-at-age data using an SAS pro-gramme written by Brian Niven (Department of Mathsand Statistics, University of Otago, New Zealand). Theremaining models were fitted using least-squares non-linear regression with the Gauss-Newton algorithm inthe statistical package SYSTAT 10.
Initially, the 4 growth models were fitted separatelyto male and female size-at-age data for each species,and their goodness-of-fit examined. We then tested forsex-specific differences in growth and, where nonewere found, fitted the 4 growth models to combined-sex data. For brevity and clarity, we present growthmodels fitted only to combined-sex data for specieswhere analysis indicated there were no sex-specificdifferences in rates or patterns of growth. Fish were
209
Mar Ecol Prog Ser 290: 207–221, 2005
sexed by the shape and size of the genital papilla(short and blunt in females, long and tapered in males)and/or by macroscopic and histological examination ofthe gonads. It was sometimes difficult to sex smalljuveniles, even using histological techniques, andthese individuals were included in both male-only andfemale-only data when fitting growth models to thesize-at-age data. Specifically, this meant individualsaged <120 d for Asterropteryx semipunctatus, <141 dfor Istigobius goldmanni, <100 d for Amblygobiusbynoensis, <72 d for Amblygobius phalaena, and<76 d for Valenciennea muralis.
To test the null hypothesis of no significant differencebetween male and female growth patterns, 1 of 2 meth-ods was employed depending on the pattern of growth.(1) Analysis of covariance (ANCOVA) was used for spe-cies with linear adult growth. Using this method, wecompared the slopes of linear regressions fitted sepa-rately to adult male and female size-at-age data. (2) Forspecies with attenuating adult growth, a comparison ofgrowth curves between sexes was made using themethods of Chen et al. (1992) and Kimura (1980).Firstly, analysis of residual sum of squares (ARSS)(Chen et al. 1992) was used to determine whether theuse of separate growth curves for females and malesprovided a better fit than 1 common growth curve. Sec-ondly, to examine differences in specific growth char-acteristics, 95% confidence ellipses were generatedaround the parameter estimates of k and L∞ (para-meters of the VBGF) for each sex, with t0 constrained toa common value (Kimura 1980). Non-overlapping con-fidence ellipses were considered to indicate a signifi-cant difference between sexes in these 2 parameters(Kimura 1980). To estimate a common value for t0, theVBGF was fitted to the size-at-age data for males andfemales separately, allowing the model to provide esti-mates for all 3 parameters (L∞, k and t0). The t0 of malesand females was then averaged to provide a commonvalue. Using an average value ensured equal bias indeviation from the original t0 for both sexes. The valueof t0 obtained for males and females was often very sim-ilar (i.e. within 0.01 and 8.57 of each other dependingon species) and, therefore, the impact of using anaverage t0 value calculated in this manner was minimal.
Seasonal comparison of growth. Inspection of thesize-at-age data suggested a possible seasonal influ-ence on growth rate for some species. To test this pos-sibility, the techniques of Kimura (1980) were used tocompare growth curves between seasons. The yearwas divided into periods of increasing (Septemberto February) and decreasing (March to August) seawater temperatures based on average monthly seawater temperatures at Pioneer Bay recordedby the Great Barrier Reef Marine Park Authority(www.gbrmpa.gov.au/corp_site/info_services/science/
seatemp). Average monthly seawater temperaturevaried annually by 7.5 to 9.0°C (maximum averagemonthly temperature 30.3°C, minimum 21.3°C) overthe study period. The months over which each individ-ual had grown were determined by subtracting itsestimated age (minus the period from hatching tosettlement) from its date of capture. Individuals werethen classified according to whether their growth hadoccurred predominantly or wholly through eithersummer or winter. Older individuals that had experi-enced growth in both seasons were categorisedaccording to the season in which most of their initialgrowth had occurred, as the faster growth rates expe-rienced by juveniles and small adults were presumedto have a larger effect on final size-at-age than growthexperienced later in life.
RESULTS
Length–weight relationship
The maximum size of male and female Asterropteryxsemipunctatus and Istigobius goldmanni were moredissimilar than the maximum size of male and femaleAmblygobius bynoensis, Amblygobius phalaena andValenciennea muralis (Table 1). The largest male A.semipunctatus and I. goldmanni were 24 and 16% largerrespectively than the largest females. In contrast, thelargest male A. bynoensis and V. muralis were only 4and 3% larger respectively than the largest females. Thelargest female A. phalaena was 2% larger than thelargest male.
210
Species Sex Max. Length–weight relationshipsize n a b r2
(mm) (×10–5)
Asterropteryx M 56 138 0.95 3.10 0.989semipunctatus F 45 139 0.32 3.42 0.974
Istigobius M 64 99 1.07 2.99 0.991goldmanni F 55 86 0.61 3.15 0.969
Amblygobius M 98 64 0.96 3.01 0.988bynoensis F 94 64 0.82 3.06 0.991
Amblygobius M 102 58 1.33 2.96 0.989phalaena F 105 57 1.07 3.01 0.996
Valenciennea M 116 71 1.32 2.84 0.989muralis F 113 98 0.73 3.00 0.932
Table 1. Maximum body size (total length, mm) andlength–weight relationships of male and female coral reefgobies. Provided are the coefficient of determination (r2) andestimates of the parameters a and b from the relationshipW = aLb, where W is weight and L is length. M: male;
F: female; n: number
Hernaman & Munday: Coral reef gobies. I
Length–weight relationships displayed predictablevalues, with b (where W = aLb) ranging from 2.84 to3.42 (Table 1). Females tended to be heavier thanmales (Table 1), although the difference was least forboth Amblygobius species, intermediate for Istigobiusgoldmanni and V. muralis, and most pronounced forAsterropteryx semipunctatus.
Precision of age estimates
The age estimates for all species generally had avery high level of precision. The CV for each individ-ual ranged from 0 to 16%, but was generally ≤4%. Thevalue of the CV was not related to fish age nor fish sizein any of the species. The mean CV per species rangedfrom 1.95 to 2.79%, depending on species, with thehighest CV associated with Asterropteryx semipuncta-tus and Istigobius goldmanni.
Maximum life-span
All 5 species were relatively short-lived, with theoldest individual in the sample ranging from 11 to16 mo depending on species and sex (Table 2).
Growth patterns
All 5 gobies invested a large proportion of their life-span in somatic growth, with growth occurring overmuch of their size range, and relatively little or no time
spent at asymptotic size (Figs. 1 to 5: note that sepa-rate-sex data are presented for Asterropteryx semi-punctatus and Istigobius goldmanni, because a sex-specific difference in growth was evident for these 2species, see below). The 5 species showed similargrowth rates in relation to their maximum life-spanwith generally about 2/3 of their maximum sizeattained by half of their maximum life-span (Table 2).
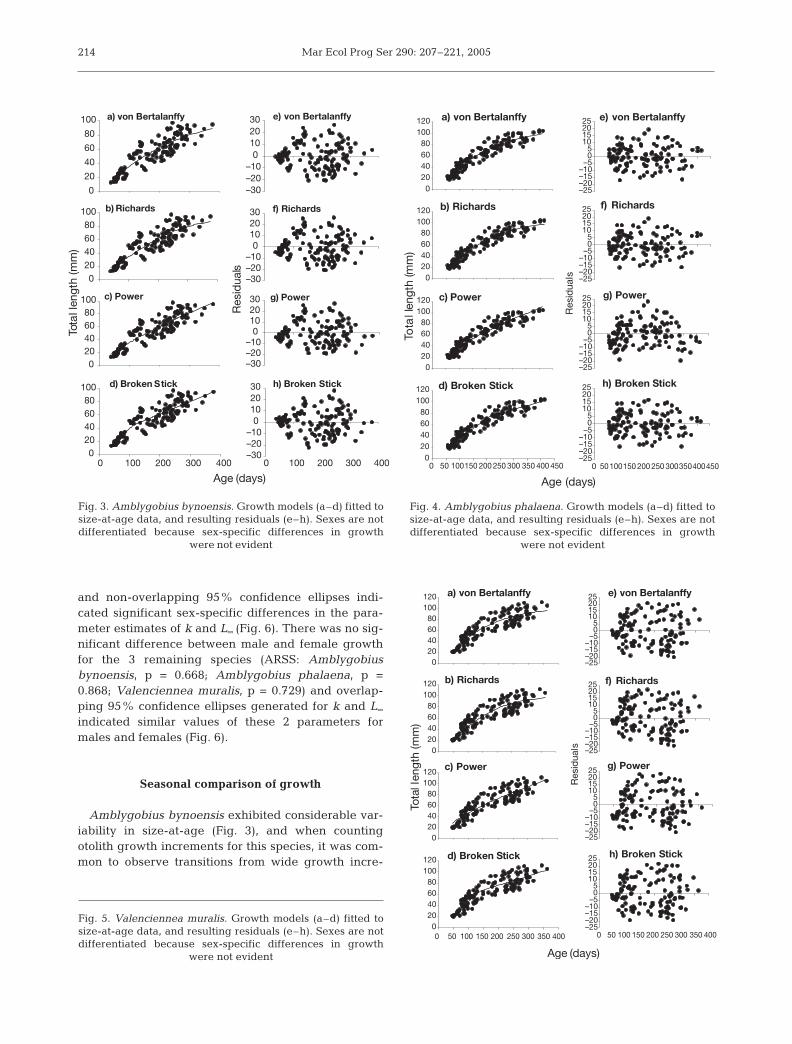
Adult growth was linear for Istigobius goldmanni(Fig. 2), and the Broken Stick regression was clearlythe best descriptor of growth for both sexes (Fig. 2,Tables 3 & 4). Growth of Asterropteryx semipunctatus,Amblygobius bynoensis, Amblygobius phalaena andValenciennea muralis attenuated near the maximumsize of each species (Figs. 1 & 3 to 5). Consequently,growth patterns were neither completely linear norasymptotic, and both the Broken Stick regression andthe VBGF adequately described the growth pattern ofthese 4 species (Figs. 1 & 3 to 5, Tables 3 & 4).
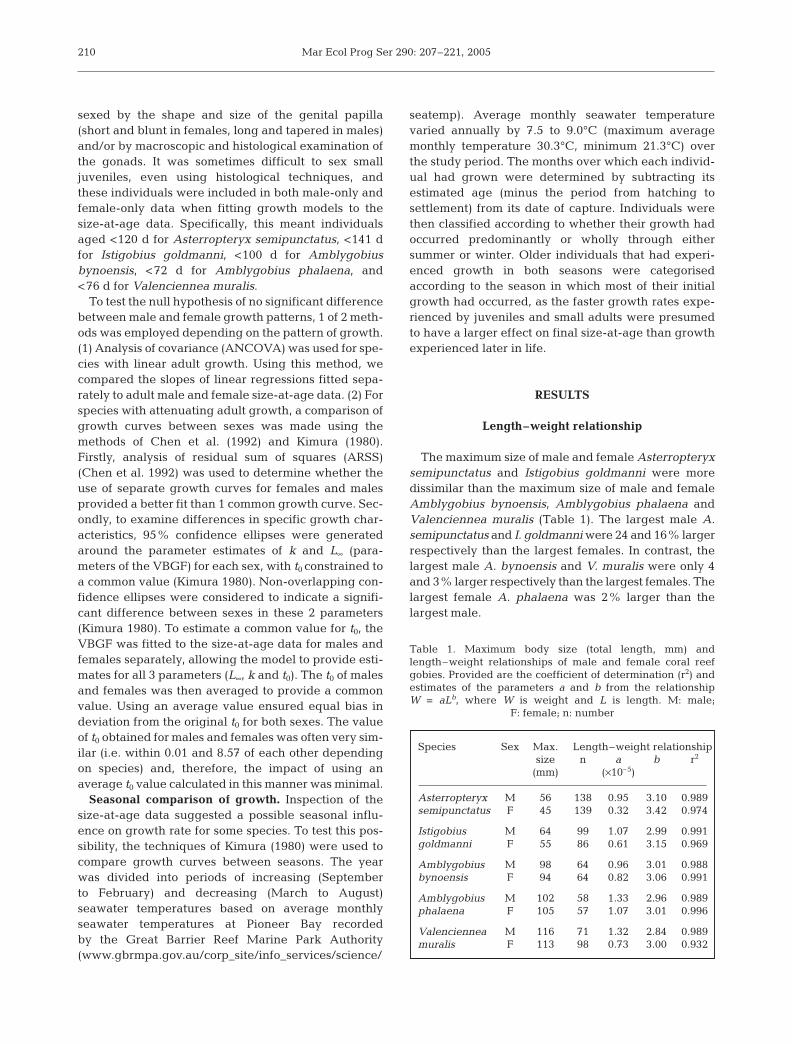
Asterropteryx semipunctatus
The size-at-age relationship showed a period ofrapid juvenile growth followed by a decrease ingrowth rate in fish over 150 d old, and growth attenu-ating at ca. 330 d (females) or 350 d (males) (Fig. 1).The VBGF was the best descriptor of growth forfemales, with an even distribution of residualsthroughout the life-span and the lowest RSS (Table 4,Fig. 1). All models gave a poorer description of malegrowth. In particular, all models tended to overesti-mate the size of males aged 200 to 300 d and >400 d,and tended to underestimate the size of males aged300 to 400 d (Fig. 1). The Broken Stick regression andVBGF had the best combinations of low RSS, high r2
and most even distribution of residuals (Table 4,Fig. 1). It was difficult to distinguish between thesemodels; however, fitting Schnute’s (1981) model to thesize-at-age data suggested that the VBGF was likely tobe the best descriptor of growth for both males andfemales (Table 3).
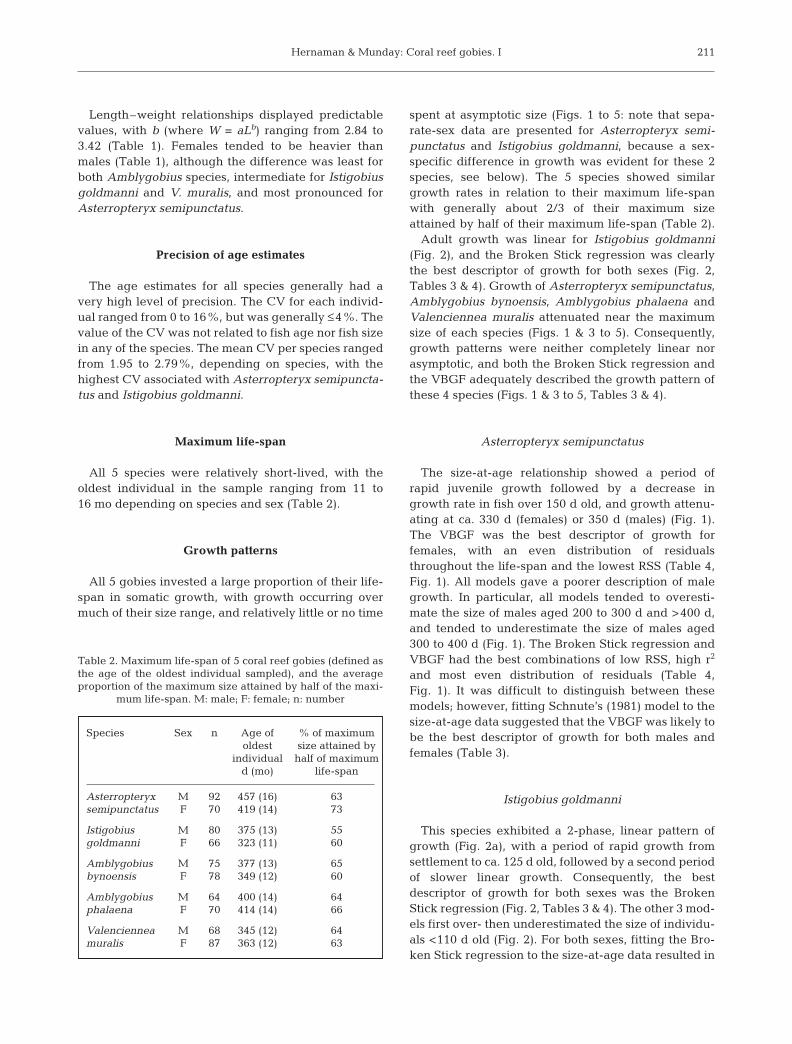
Istigobius goldmanni
This species exhibited a 2-phase, linear pattern ofgrowth (Fig. 2a), with a period of rapid growth fromsettlement to ca. 125 d old, followed by a second periodof slower linear growth. Consequently, the bestdescriptor of growth for both sexes was the BrokenStick regression (Fig. 2, Tables 3 & 4). The other 3 mod-els first over- then underestimated the size of individu-als <110 d old (Fig. 2). For both sexes, fitting the Bro-ken Stick regression to the size-at-age data resulted in
211
Species Sex n Age of % of maximum oldest size attained by
individual half of maximum d (mo) life-span
Asterropteryx M 92 457 (16) 63semipunctatus F 70 419 (14) 73
Istigobius M 80 375 (13) 55goldmanni F 66 323 (11) 60
Amblygobius M 75 377 (13) 65bynoensis F 78 349 (12) 60
Amblygobius M 64 400 (14) 64phalaena F 70 414 (14) 66
Valenciennea M 68 345 (12) 64muralis F 87 363 (12) 63
Table 2. Maximum life-span of 5 coral reef gobies (defined asthe age of the oldest individual sampled), and the averageproportion of the maximum size attained by half of the maxi-
mum life-span. M: male; F: female; n: number
Mar Ecol Prog Ser 290: 207–221, 2005
the lowest RSS (Table 4). The RSS obtained from fittingthe VBGF was not dramatically higher than thatobtained from fitting the Broken Stick regression.However, the VBGF was only able to provide a reason-able description of growth for Istigobius goldmanni byusing unreasonably large predicted values of L∞ (98.1and 64.8 mm TL for males and females respectively),which were much greater than the maximum size ofthis species at Orpheus Island (64 and 56 mm TL formales and females, respectively). Fitting Schnute’smodel to the size-at-age data confirmed that a non-asymptotic growth model was likely to be the bestdescriptor of growth, with an exponential and a linearmodel suggested for males and females respectively(Table 3).
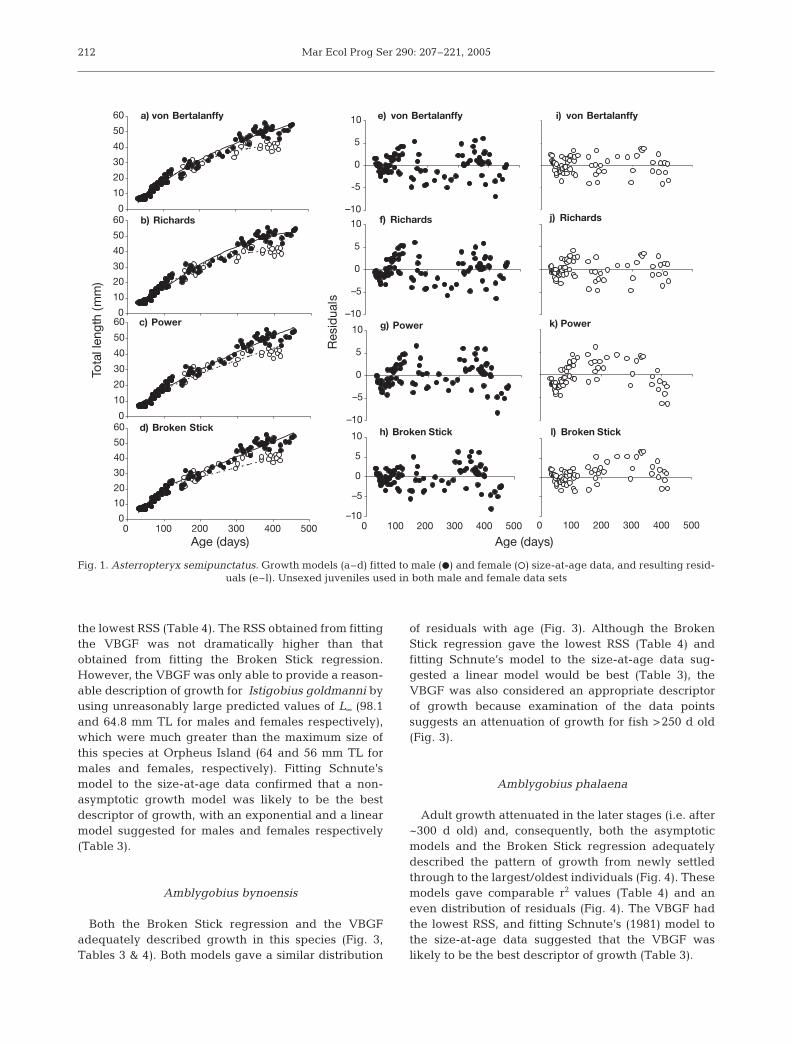
Amblygobius bynoensis
Both the Broken Stick regression and the VBGFadequately described growth in this species (Fig. 3,Tables 3 & 4). Both models gave a similar distribution
of residuals with age (Fig. 3). Although the BrokenStick regression gave the lowest RSS (Table 4) andfitting Schnute’s model to the size-at-age data sug-gested a linear model would be best (Table 3), theVBGF was also considered an appropriate descriptorof growth because examination of the data pointssuggests an attenuation of growth for fish >250 d old(Fig. 3).
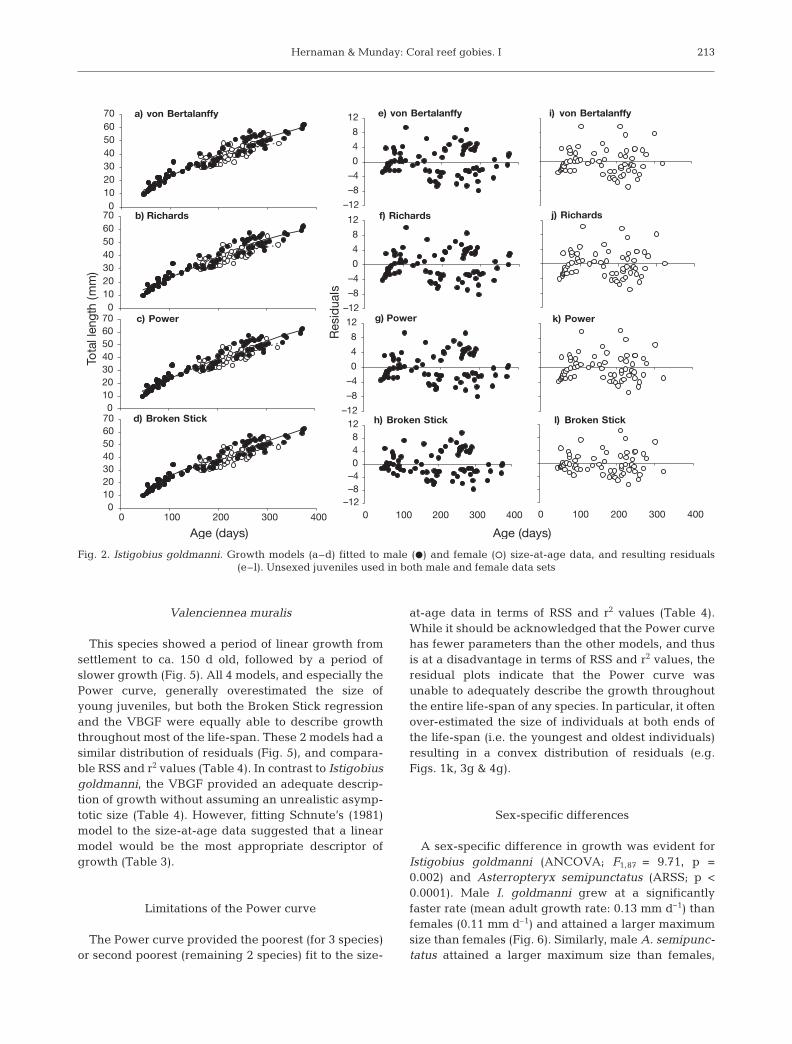
Amblygobius phalaena
Adult growth attenuated in the later stages (i.e. after~300 d old) and, consequently, both the asymptoticmodels and the Broken Stick regression adequatelydescribed the pattern of growth from newly settledthrough to the largest/oldest individuals (Fig. 4). Thesemodels gave comparable r2 values (Table 4) and aneven distribution of residuals (Fig. 4). The VBGF hadthe lowest RSS, and fitting Schnute’s (1981) model tothe size-at-age data suggested that the VBGF waslikely to be the best descriptor of growth (Table 3).
212
a) von Bertalanffy
0
10
20
30
40
50
60
c) Power
0
10
20
30
40
50
60
b) Richards
0
10
20
30
40
50
60
d) Broken Stick
0
10
20
30
40
50
60
0 100 200 300 400 500Age (days)
Tota
llen
gth
(mm
)
i) von Bertalanffy
j) Richards
k) Power
l) Broken Stick
0 100 200 300 400 500
e) von Bertalanffy
–10
-5
0
5
10
f) Richards
–10
–5
0
5
10
g) Power
–10
–5
0
5
10
h) Broken Stick
–10
–5
0
5
10
0 100 200 300 400 500
Age (days)
Res
idua
ls
Fig. 1. Asterropteryx semipunctatus. Growth models (a–d) fitted to male (d) and female (s) size-at-age data, and resulting resid-uals (e–l). Unsexed juveniles used in both male and female data sets
Hernaman & Munday: Coral reef gobies. I
Valenciennea muralis
This species showed a period of linear growth fromsettlement to ca. 150 d old, followed by a period ofslower growth (Fig. 5). All 4 models, and especially thePower curve, generally overestimated the size ofyoung juveniles, but both the Broken Stick regressionand the VBGF were equally able to describe growththroughout most of the life-span. These 2 models had asimilar distribution of residuals (Fig. 5), and compara-ble RSS and r2 values (Table 4). In contrast to Istigobiusgoldmanni, the VBGF provided an adequate descrip-tion of growth without assuming an unrealistic asymp-totic size (Table 4). However, fitting Schnute’s (1981)model to the size-at-age data suggested that a linearmodel would be the most appropriate descriptor ofgrowth (Table 3).
Limitations of the Power curve
The Power curve provided the poorest (for 3 species)or second poorest (remaining 2 species) fit to the size-
at-age data in terms of RSS and r2 values (Table 4).While it should be acknowledged that the Power curvehas fewer parameters than the other models, and thusis at a disadvantage in terms of RSS and r2 values, theresidual plots indicate that the Power curve wasunable to adequately describe the growth throughoutthe entire life-span of any species. In particular, it oftenover-estimated the size of individuals at both ends ofthe life-span (i.e. the youngest and oldest individuals)resulting in a convex distribution of residuals (e.g.Figs. 1k, 3g & 4g).
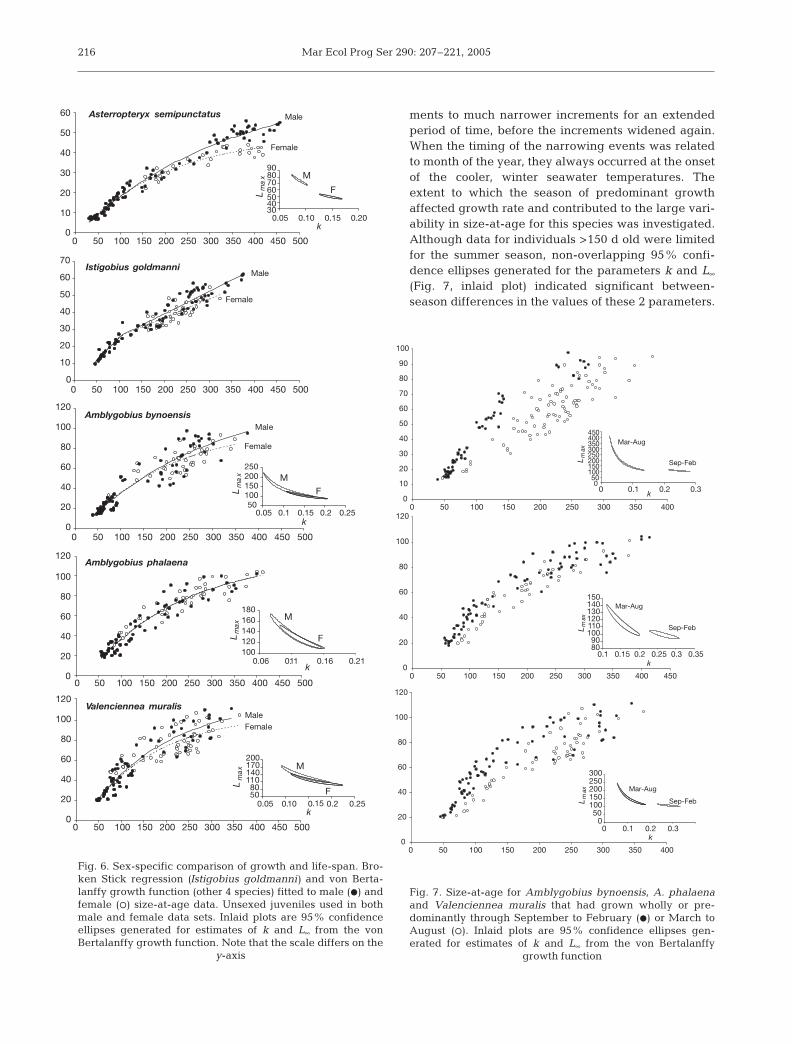
Sex-specific differences
A sex-specific difference in growth was evident forIstigobius goldmanni (ANCOVA; F1,87 = 9.71, p =0.002) and Asterropteryx semipunctatus (ARSS; p <0.0001). Male I. goldmanni grew at a significantlyfaster rate (mean adult growth rate: 0.13 mm d–1) thanfemales (0.11 mm d–1) and attained a larger maximumsize than females (Fig. 6). Similarly, male A. semipunc-tatus attained a larger maximum size than females,
213
a) von Bertalanffy
010203040506070
Age (days)
Tota
llen
gth
(mm
)
b) Richards
010203040506070
c) Power
010203040506070
d) Broken Stick
010203040506070
0 100 200 300 400
i) von Bertalanffye) von Bertalanffy
–12
–8
–4
0
4
8
12
Age (days)
Res
idua
ls
f) Richards
–12
–8
–4
0
4
8
12
g) Power
–12
–8
–4
0
4
8
12
h) Broken Stick
–12–8–4048
12
0 100 200 300 400
j) Richards
k) Power
l) Broken Stick
0 100 200 300 400
Fig. 2. Istigobius goldmanni. Growth models (a–d) fitted to male (d) and female (s) size-at-age data, and resulting residuals (e–l). Unsexed juveniles used in both male and female data sets
Mar Ecol Prog Ser 290: 207–221, 2005
and non-overlapping 95% confidence ellipses indi-cated significant sex-specific differences in the para-meter estimates of k and L∞ (Fig. 6). There was no sig-nificant difference between male and female growthfor the 3 remaining species (ARSS: Amblygobiusbynoensis, p = 0.668; Amblygobius phalaena, p =0.868; Valenciennea muralis, p = 0.729) and overlap-ping 95% confidence ellipses generated for k and L∞
indicated similar values of these 2 parameters formales and females (Fig. 6).
Seasonal comparison of growth
Amblygobius bynoensis exhibited considerable var-iability in size-at-age (Fig. 3), and when countingotolith growth increments for this species, it was com-mon to observe transitions from wide growth incre-
214
a) von Bertalanffy
0
20
40
60
80
100
b) Richards
020406080
100
c) Power
020406080
100
d) Broken Stick
020406080
100
0 100 200 300 400
e) von Bertalanffy
–30–20–10
0102030
f) Richards
–30–20–10
0102030
g) Power
–30–20–10
0102030
h) Broken Stick
–30–20–10
0102030
0 100 200 300 400
Tota
llen
gth
(mm
)
Age (days)
Res
idua
ls
Fig. 3. Amblygobius bynoensis. Growth models (a–d) fitted tosize-at-age data, and resulting residuals (e–h). Sexes are notdifferentiated because sex-specific differences in growth
were not evident
a) von Bertalanffy
020406080
100120
b) Richards
020406080
100120
c) Power
020406080
100120
d) Broken Stick
020406080
100120
0 50 100150 200 250 300 350 400 450
e) von Bertalanffy
–25–20–15–10–505
10152025
f) Richards
–25–20–15–10–505
10152025
g) Power
–25–20–15–10–505
10152025
h) Broken Stick
–25–20–15–10–505
10152025
0 50100150 200 250 300350400450
Age (days)
Tota
llen
gth
(mm
)
Res
idua
ls
Fig. 4. Amblygobius phalaena. Growth models (a–d) fitted tosize-at-age data, and resulting residuals (e–h). Sexes are notdifferentiated because sex-specific differences in growth
were not evident
a) von Bertalanffy
020406080
100120
b) Richards
020406080
100120
c) Power
020406080
100120
d) Broken Stick
020406080
100120
0 50 100 150 200 250 300 350 400
e) von Bertalanffy
–25–20–15–10–505
10152025
f) Richards
–25–20–15–10–505
10152025
g) Power
–25–20–15–10–505
10152025
h) Broken Stick
–25–20–15–10–505
10152025
0 50 100 150 200 250 300 350 400
Age (days)
Tota
llen
gth
(mm
)
Res
idua
ls
Fig. 5. Valenciennea muralis. Growth models (a–d) fitted tosize-at-age data, and resulting residuals (e–h). Sexes are notdifferentiated because sex-specific differences in growth
were not evident
Hernaman & Munday: Coral reef gobies. I 215
Growth RSS r2 Parameter estimates:model VBGF/Richards Power BS
L∞ k t0 a b b d a c x0
Asterropteryx VBGF 590 0.978 77.1 0.003 3.4 – – – – – –semipunctatus Richards 651 0.975 58.6 0.007 132.1 – – – – – –males (n = 92) Power 715 0.973 – – – 0.519 0.767 – – – –
BStick 640 0.976 – – – – – 0.20 0.10 –1.25 10.88 121.3
Asterropteryx VBGF 269 0.976 47.2 0.006 11.2 – – – – – –semipunctatus Richards 278 0.975 42.5 0.011 90.2 – – – – – –females (n = 70) Power 552 0.949 – – – 0.799 0.668 – – – –
BStick 447 0.955 – – – – – 0.20 0.06 –1.25 15.31 118.3
Istigobius VBGF 1125 0.935 98.1 0.003 –9.6 – – – – – –goldmanni Richards 1156 0.934 70.8 0.007 123.5 – – – – – –males (n = 80) Power 1144 0.934 – – – 0.812 0.732 – – – –
BStick 1111 0.935 – – – – – 0.26 0.13 –0.94 12.72 104.4
Istigobius VBGF 771 0.909 64.8 0.004 –6.1 – – – – – –goldmanni Richards 819 0.902 54.9 0.009 87.8 – – – – – –females (n = 66) Power 778 0.906 – – – 1.042 0.674 – – – –
BStick 685 0.919 – – – – – 0.26 0.11 –0.94 15.07 104.2
Amblygobius VBGF 15336 0.806 117.1 0.004 7.3 – – – – – –bynoensis Richards 16015 0.794 95.0 0.009 112.4 – – – – – –(n = 143) Power 15662 0.796 – – – 1.071 0.761 – – – –
BStick 14324 0.819 – – – – – 0.48 0.19 –8.25 23.69 110.1
Amblygobius VBGF 6820 0.908 114.4 0.005 21.1 – – – – – –phalaena Richards 7006 0.905 101.1 0.010 102.8 – – – – – –(n = 122) Power 8307 0.882 – – – 1.452 0.718 – – – –
BStick 7100 0.904 – – – – – 0.33 0.14 3.95 44.73 214.6
Valenciennea VBGF 14856 0.806 106.1 0.007 14.1 – – – – – –muralis Richards 15303 0.798 100.1 0.012 81.3 – – – – – –(n = 130) Power 15765 0.787 – – – 2.525 0.636 – – – –
BStick 14803 0.811 – – – – – 0.47 0.17 0.73 42.41 138.9
Table 4. Comparison of 4 growth models fitted to the size-at-age data for 5 species of coral reef goby. Presented are the residualsum of squares (RSS), coefficient of determination (r2), and parameter estimates (see ‘Materials and methods’ for parameter
definitions). VBGF: von Bertalanffy growth function; BS: Broken Stick. x0: breakpoint
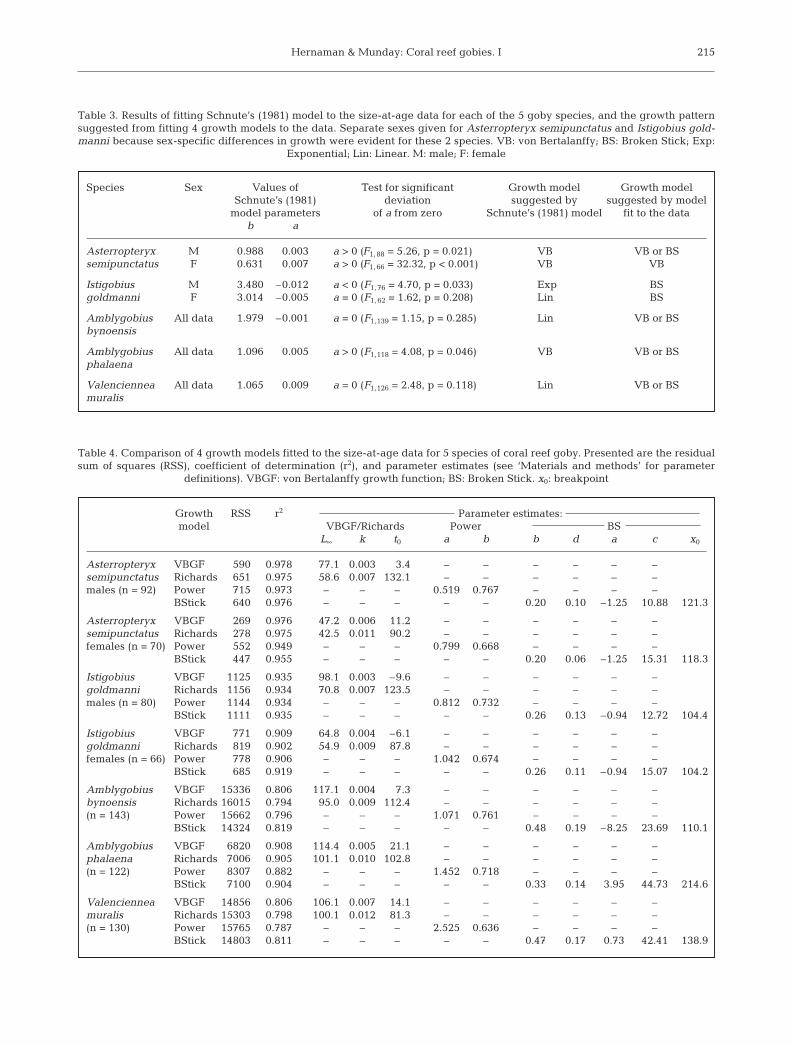
Species Sex Values of Test for significant Growth model Growth model Schnute’s (1981) deviation suggested by suggested by model
model parameters of a from zero Schnute’s (1981) model fit to the datab a
Asterropteryx M 0.988 0.003 a > 0 (F1, 88 = 5.26, p = 0.021) VB VB or BSsemipunctatus F 0.631 0.007 a > 0 (F1, 66 = 32.32, p < 0.001) VB VB
Istigobius M 3.480 –0.012 a < 0 (F1, 76 = 4.70, p = 0.033) Exp BSgoldmanni F 3.014 –0.005 a = 0 (F1, 62 = 1.62, p = 0.208) Lin BS
Amblygobius All data 1.979 –0.001 a = 0 (F1,139 = 1.15, p = 0.285) Lin VB or BSbynoensis
Amblygobius All data 1.096 0.005 a > 0 (F1,118 = 4.08, p = 0.046) VB VB or BSphalaena
Valenciennea All data 1.065 0.009 a = 0 (F1,126 = 2.48, p = 0.118) Lin VB or BSmuralis
Table 3. Results of fitting Schnute’s (1981) model to the size-at-age data for each of the 5 goby species, and the growth patternsuggested from fitting 4 growth models to the data. Separate sexes given for Asterropteryx semipunctatus and Istigobius gold-manni because sex-specific differences in growth were evident for these 2 species. VB: von Bertalanffy; BS: Broken Stick; Exp:
Exponential; Lin: Linear. M: male; F: female
Mar Ecol Prog Ser 290: 207–221, 2005
ments to much narrower increments for an extendedperiod of time, before the increments widened again.When the timing of the narrowing events was relatedto month of the year, they always occurred at the onsetof the cooler, winter seawater temperatures. Theextent to which the season of predominant growthaffected growth rate and contributed to the large vari-ability in size-at-age for this species was investigated.Although data for individuals >150 d old were limitedfor the summer season, non-overlapping 95% confi-dence ellipses generated for the parameters k and L∞
(Fig. 7, inlaid plot) indicated significant between-season differences in the values of these 2 parameters.
216
0
10
20
30
40
50
60
70
80
90
100
0 50 100 150 200 250 300 350 400
050
100150200250300350400450
0 0.1 0.2 0.3k
max
Mar-Aug
Sep-Feb
0
20
40
60
80
100
120
0 50 100 150 200 250 300 350 400 450
8090
100110120130140150
0.1 0.15 0.2 0.25 0.3 0.35k
Mar-Aug
Sep-Feb
0
20
40
60
80
100
120
0 50 100 150 200 250 300 350 400
050
100150200250300
0 0.1 0.2 0.3k
Mar-Aug
Sep-Feb
Lm
axL
max
L
Fig. 7. Size-at-age for Amblygobius bynoensis, A. phalaenaand Valenciennea muralis that had grown wholly or pre-dominantly through September to February (d) or March toAugust (s). Inlaid plots are 95% confidence ellipses gen-erated for estimates of k and L∞ from the von Bertalanffy
growth function
Asterropteryx semipunctatus
0
10
20
30
40
50
60
0 50 100 150 200 250 300 350 400 450 500
Female
Male
30405060708090
0.05 0.10 0.15 0.20k
Lm
ax
F
M
Amblygobius bynoensis
0
20
40
60
80
100
120
0 50 100 150 200 250 300 350 400 450 500
Male
Female
50100150200250
0.05 0.1 0.15 0.2 0.25k
Lm
ax M
F
Amblygobius phalaena
0
20
40
60
80
100
120
0 50 100 150 200 250 300 350 400 450 500
100120140160180
0.06 0.11 0.16 0.21k
Lm
ax
M
F
Valenciennea muralis
0
20
40
60
80
100
120
0 50 100 150 200 250 300 350 400 450 500
MaleFemale
5080
110140170200
0.05 0.10 .150 0.2 0.25k
Lm
ax M
F
Istigobius goldmanni
0
10
20
30
40
50
60
70
0 50 100 150 200 250 300 350 400 450 500
Male
Female
Fig. 6. Sex-specific comparison of growth and life-span. Bro-ken Stick regression (Istigobius goldmanni) and von Berta-lanffy growth function (other 4 species) fitted to male (d) andfemale (s) size-at-age data. Unsexed juveniles used in bothmale and female data sets. Inlaid plots are 95% confidenceellipses generated for estimates of k and L∞ from the vonBertalanffy growth function. Note that the scale differs on the
y-axis

Hernaman & Munday: Coral reef gobies. I
Generally, individuals that had grown wholly or pre-dominantly through the summer months (September toFebruary) had attained a significantly larger size at agiven age than individuals that had grown wholly orpredominantly through the winter months (March toAugust) (Fig. 7). However, there was still substantialwithin-season variation in size-at-age for individualsgrowing predominantly through the winter months(Fig. 7).
The data set for summer-growing adults of Amblygo-bius phalaena and Valenciennea muralis was muchlarger than that for A. bynoensis, and seasonal differ-ences in growth rates of the 2 former species were mostapparent for individuals <130 and <150 d old, respec-tively (Fig. 7). Even so, non-overlapping 95% confi-dence ellipses generated for k and L∞ indicated thatseasonal differences in growth rate resulted in signifi-cant seasonal differences in these 2 growth parameters(Fig. 7, inlaid plots).
Seasonal differences in growth rates were clearlyevident for juveniles of all 3 species. Juveniles growingthrough summer exhibited a significantly highergrowth rate than winter-growing individuals (Ambly-gobius bynoensis, mean growth rate of 0.54 and0.29 mm d–1 for summer and winter growth respec-tively, ANCOVA: F1,61 = 102.14, p < 0.001; A. phalaena,0.63 and 0.45 mm d–1, F1,37 = 95.22, p < 0.001; Valenci-ennea muralis, 0.61 and 0.42 mm d–1, F1,57 = 107.64, p <0.001). These seasonal differences in growth ratestranslate to an average of 25 mm (A. bynoensis), 18 mm(A. phalaena) and 19 mm (V. muralis) difference in sizeafter a period of 100 d.
The data set for Asterropteryx semipunctatus andfemale Istigobius goldmanni was not conducive todetailed examination of seasonal differences in growthrates. Suitable data were available for male I. gold-manni, and ANCOVA indicated that males growingpredominantly through summer had significantlyhigher growth rates than males growing predomi-nantly through winter (F1,49 = 61.12, p < 0.001).
DISCUSSION
The 5 coral reef gobies in this study were all small-bodied and all exhibited a relatively short maximumlife-span and rapid growth rates. Maximum life-spanwas from 11 to 16 mo, and each species attained ca. 2⁄3of its maximum size by half of its expected maximumlife-span. Therefore, it appears that their patterns ofgrowth and life-span conform to traditional concepts oflife-history theory.
The pattern of growth of all 5 coral reef gobiesdiffered considerably from that exhibited by otherrelatively small, but long-lived (up to 32+ yr old) coral
reef fishes, such as certain acanthurids (e.g. Choat &Axe 1996, Hart & Russ 1996) and pomacentrids (e.g.Meekan et al. 2001). These long-lived species tend toattain an asymptotic size very quickly (within 2 to 6 yr)and thus spend much of their life-span (often >75%) atthat size. In contrast, all 5 coral reef gobies invested insomatic growth throughout much or all of their life-span and spent relatively little or no time at asymptoticsize. While there was evidence that growth began toattenuate in the oldest individuals of 4 species, Istigo-bius goldmanni clearly exhibited linear adult growthwith no signs of an asymptotic size. Rapid non-asymptotic growth has also been noted for Istigobiusdecoratus, a short-lived (<1 yr) goby collected from thelagoon at Lizard Island, Great Barrier Reef (Kritzer2002). Similarly, the intertidal Hawaiian goby Bathygo-bius coalitus exhibited a linear pattern of growth for atleast the first 9 mo of life, with no difference in growthrate between immature and mature gobies (Shafer1998). Furthermore, size-at-age data presented for arange of temperate gobies also indicates non-asymp-totic growth (Grossman 1979, Bouchereau & Guelorget1998 and see Kritzer 2002). It seems, therefore, thatnon-asymptotic growth may be relatively commonamong both tropical and temperate water gobies.
Long-lived reef fishes often have a growth patterncharacterised by rapid growth to asymptotic size andsexual maturity, followed by a relatively long period ofreproductive activity, but little additional somaticgrowth (Choat & Robertson 2002). This growth patternmay reflect a physiological trade-off between growthand reproduction. Trade-offs play a central role in life-history theory, and a physiological trade-off betweengrowth and reproduction is expected where these 2processes compete directly for a limited amount ofenergy (Begon & Mortimer 1981, Partridge & Harvey1988, Stearns 1992). However, circumstances mayarise where a trade-off between growth and reproduc-tion is minimised or does not occur. For example,energy allocated to reproduction may not entail agrowth cost if there were limits to the rate at whichenergy can be diverted into the somatic tissues (Roff1992). When fed the same rations, non-reproducingfemale guppies grew more than reproductively activefemales, but most of the growth was in the form of fatreserves and not somatic protein growth (Reznick1983, cited by Wootton 1998). Alternatively, on reach-ing sexual maturity, a reproductive individual might beable to acquire sufficient additional energy to coverthe energetic costs of reproduction, either by an onto-genetic shift in diet to a more profitable or higherquality food source (e.g. MacNeill & Brandt 1990,McCormick 1998), or by an increase in feeding rate.However, increased foraging time may result in in-creased predation risk, potentially resulting in a trade-
217

Mar Ecol Prog Ser 290: 207–221, 2005
off between growth or reproduction and longevity.While the growth rate of the 5 study species decreasedfollowing maturation, investment in somatic growthcontinued for much or all of the adult life-span. There-fore, they appear able to acquire sufficient energy tosatisfy demands for both growth and reproduction.This capacity to continue somatic growth may come ata cost of reduced longevity, as evidenced by theirrelatively short life-spans.
For the 5 study species, continued investment insomatic growth may potentially result in reproductivebenefits for both sexes. A positive relationship be-tween body size and fecundity (Hernaman 2003)means that females can maximise both current andfuture batch fecundity by continued somatic growth.While males do not need a large body size to accom-modate testes capable of fertilising a large number ofeggs, continued investment in somatic growth mayincrease their reproductive success through the com-petitive advantage of large body size in both intra-(Grossman 1980, Yanagisawa 1982, Breitburg 1987,Lindström 1992a,b, Forsgren et al. 1996) and interspe-cific (Breitburg 1987, Robertson 1996) competition forresources (e.g. nest sites, mates, food, territory).Intraspecific competition for nest sites and mates hasbeen recorded for the polygynous Asterropteryx semi-punctatus and Istigobius goldmanni (Hernaman 2003),with a large body size providing a competitive advan-tage in contests over those resources. Intraspecificcompetition for nest sites was not evident for themonogamous pair-spawning Valenciennea muralis,Amblygobius bynoensis and Amblygobius phalaena(Hernaman 2003). However, mate defence by bothsexes was observed occasionally and continued invest-ment in growth may provide a size benefit in contestsover mates. Body size influences mate selection inother monogamous coral reef gobies, with both malesand females preferring larger partners (Reavis 1997b,Takegaki & Nakazono 1999) and pair bonds beingbroken when a mate became available that was largerthan the current partner (Takegaki & Nakazono 1999).Thus, investment of energy in somatic growth is likelyto provide reproductive benefits for both sexes of all5 study species.
Female Asterropteryx semipunctatus and Istigobiusgoldmanni grew more slowly and attained smallermaximum sizes than males. One explanation is thatfemales expend proportionally more energy on theproduction of gametes than males (e.g. Newman et al.1996), and this extra energy requirement is at theexpense of somatic growth. Egg production can consti-tute a considerable drain on energy reserves (Kamler1992). For example, in female Pomatoschistus microps,28 to 44% of the dietary energy content is devoted toegg production (Rogers 1988, cited in Magnhagen
1993). Thus, for the 5 study species that spawn at fre-quent intervals over the summer months (authors’ pers.obs.), sex-specific differences in the cost of gameteproduction could underlie differences in growth ratebetween sexes. However, sex-specific differences ingrowth were not apparent for Valenciennea muralis,Amblygobius bynoensis or Amblygobius phalaena.This disparity among species may be related to inter-specific differences in male egg-guarding behaviour.The eggs of V. muralis, A. bynoensis and A. phalaenaare laid in 1 large cluster, and males appear to spendmuch of the egg-guarding period fanning the eggmass (Hernaman 2003). Males did not leave the nestsite during egg-guarding and did not feed during thistime. In contrast, A. semipunctatus and I. goldmannieggs are deposited in a monolayer, extensive egg fan-ning was not observed, and males fed near the sheltersite during egg-guarding (Hernaman 2003). Thus, theextensive and prolonged egg-fanning activities of V.muralis, A. phalaena and A. bynoensis males, com-bined with the absence of feeding during this time,may result in a relatively higher energetic cost of broodcare, compared to that experienced by male A. semi-punctatus and I. goldmanni, that is equivalent to theextra energetic cost of egg production in females.
The mating system may also contribute to sex-specific differences in growth rate and maximum sizein some species. Sex-specific differences in growthrate were evident for the 2 polygamous species andabsent for the 3 monogamous pair-spawning species.This pattern could be related to differences in matecompetition associated with each of these mating sys-tems. In monogamous gobies, body size of both malesand females appears to be important in mate selectionand the stability of the pair bond (Reavis & Barlow1998). In contrast, for the 2 polygynous species, intra-sexual competition for mates and nest sites is likely tobe much stronger among males than females (Emlen &Oring 1977, Warner 1984). Therefore, there may be aselective premium to rapid male growth and large sizein the 2 polygynous species.
The fast growth rates and short life-span often exhib-ited by small fishes are thought to be the functionalresponse of a high level of unpredictable mortalityacross the entire life-span (Miller 1996). Althoughthere are a variety of causes of mortality in fish (e.g.reproductive demands, senescence and parasitism;Grossman 1979), predation is likely to be the primaryagent of mortality in small species (Pauly 1980, Miller1984, Munday & Jones 1998). Coral reef gobies arepredated upon by a wide range of demersal andmobile piscivorous fishes (Sweatman 1984, Parrish etal. 1986, Norris & Parrish 1988, Kingsford 1992, Clarket al. 2000), and might also be vulnerable to predationby sea snakes, invertebrates (e.g. octopus) and shore
218

Hernaman & Munday: Coral reef gobies. I
birds. Therefore, the short maximum life-span of all 5study species is likely to reflect their vulnerability topredation by a wide range of predators.
Munday & Jones (1998) suggested that patterns ofhabitat use and predation risk might have a substantialinfluence on the maximum life-span of small fishes.Small temperate gobies show 2 divergent life-historystrategies (Grossman 1979), which may be linked to themortality rate associated with habitat type (Miller1996). Nektonic species and those associated with opensandy habitats have a short life-span of <3 yr (Fig. 8),presumably because they experience a high risk of pre-dation (Miller 1984). In contrast, cryptobenthic and bur-row-dwelling species have relatively long lives of 7 to11 yr (Fig. 8), presumably because they experience alower risk of predation (Miller 1984). These differencesin maximum life-span are often unrelated to differ-ences in maximum size (Fig. 8, and see Miller 1979).
Although information for tropical gobies is limited,the available data suggest that gobies with similarpatterns of habitat use to the 5 study species arealso short-lived. For example, Coryphopterus glauco-fraenum and Istigobius decoratus, that attain a similarmaximum size and occur in similar habitats to I. gold-manni, have a maximum life-span of <1 yr (Forrester1995, Kritzer 2002). Gobies that utilise burrows forshelter also have similar maximum life-spans to the3 species in the current study that shelter in burrows.For example, Amblyeletoris japonica and Valencien-nea strigata both have a maximum life-span of <2 yr(Yanagisawa 1982, Reavis 1997a). In contrast, more
cryptic gobies such as obligate coral-dwelling gobieshave longer maximum life-spans. For example, Para-gobiodon echinocephalus can live for >2 yr (Kuwa-mura et al. 1996) and Gobiodon histrio can live at least4 yr (Munday 2001).
A comparison between temperate and tropical go-bies suggests that nektonic and open sand specieshave similarly short lives in both regions (Fig. 8),probably because high rates of predation drive the life-history traits of all these species. In contrast, temperateburrowing and cryptobenthic species tend to belonger-lived than similar sized tropical species with thesame behaviour. This difference may reflect funda-mental differences in life-history traits among lati-tudes. Choat & Robertson (2002) described, for 2species of acanthurid fishes, a similar relationship be-tween maximum age and latitude to that observedhere, and concluded that this pattern may be linked tolatitudinal variation in reproductive rates. The capacityto reproduce successfully throughout the year in thetropics may select for short life-spans, whereas thetruncation of reproduction to just a few months incooler waters may select for longer life-spans. Alterna-tively, there may be an intrinsic association betweengrowth rates and longevity, such that slow growth incool, high-latitude waters leads to a greater maximumage, whereas fast growth in warm, low latitude watersproduces a shorter maximum age (Pauly 1980). How-ever, further studies are required on a range of speciesin each geographic locale to examine interspecificvariation and to separate phylogenetic relationshipsfrom latitudinal differences.
Seasonal variation in growth rate was evident forseveral of the study species. Individuals growingwholly or predominantly through the summer hadattained a significantly larger size for a given age thanindividuals growing wholly or predominantly throughthe winter. This seasonal difference in growth rate wasmost pronounced in juveniles, suggesting that the sea-son of settlement may have a significant impact on thetiming of maturation, the probability of survival tomaturation and reproductive output/success. Repro-ductive activity was highest during the summermonths, but reproductively active females were alsofound in winter. Thus, recruitment to the adult habitatis likely to occur during both summer and winter. If thetiming of maturation is dependent on size, juvenilessettling during early summer will attain that size in ashorter time than juveniles settling during early winter.Thus, the time to first reproduction may be relativelyshorter in summer-recruiting individuals. Fastergrowth, and thus a relatively larger body size, may alsoaffect reproductive output because of the effects ofbody size on fecundity and contests over resources,such as nest sites and reproductive mates.
219
0123456789
1011
0 50 100 150 200 250Maximum body size (mm)
Max
imum
life-
span
(yea
rs)
Temp nektonic/open sand
Temp burrows
Temp cryptobenthic
Trop nektonic/open sand
Trop burrows
Trop cryptobenthic
Trop corals
Fig. 8. Relationship between maximum body size and maxi-mum life-span for a suite of tropical (Trop) and temperate(Temp) gobies with different patterns of habitat use. Dataderived from: Miller (1961, 1979, 1984), Gibson (1970), Gross-man (1979), Yanagisawa (1982), Moreira et al. (1991), Arrudaet al. (1993), Magnhagen (1993), Forrester (1995), Gill et al.(1996), Kuwamura et al. (1996), Iglesias et al. (1997), Reavis(1997b), Sano (1997), Bouchereau & Guelorget (1998), Robert-son & Kaufmann (1998), Kovacic (2001), Munday (2001),Kritzer (2002), and Mazzoldi et al. (2002) and present study

Mar Ecol Prog Ser 290: 207–221, 2005
Acknowledgements. We thank J. H. Choat for his support ofthe project and for providing logistical support at James CookUniversity. We also thank M. Boyle, P. Brewin and D. Craigfor field assistance; W. Robbins for advice on otolith prepara-tion; J. Ackerman, J. Kritzer and W. Robbins for valuable dis-cussions on growth models; 3 anonymous reviewers for con-structive comments. B. Niven, Department of Maths andStatistics, University of Otago, provided statistical advice.This study was funded by a University of Otago ResearchGrant awarded to K. Probert (Department of Marine Science,University of Otago) and by Divisional Funding (Departmentof Marine Science, University of Otago), a PADI AwareResearch Grant, and a Commonwealth Postgraduate Scholar-ship awarded to V.H.
LITERATURE CITED
Ackerman JL, Bellwood DR (2000) Reef fish assemblages: are-evaluation using enclosed rotenone stations. Mar EcolProg Ser 206:227–237
Arruda LM, Azevedo JN, Neto AI (1993) Abundance, age-structure and growth, and reproduction of gobies (Pisces;Gobiidae) in the Ria de Aveiro Lagoon (Portugal). EstuarCoast Shelf Sci 37:509–523
Begon M, Mortimer M (1981) Population ecology: a unifiedstudy of animals and plants. Blackwell Scientific Publica-tions, London
Blueweiss L, Fox H, Kudzma V, Nakashima D, Peters R, SamsS (1978) Relationships between body size and some lifehistory parameters. Oecologia 37:257–272
Bouchereau JL, Guelorget O (1998) Comparison of 3 Gobi-idae (Teleostei) life history strategies over their geograph-ical range. Oceanol Acta 21:503–517
Breitburg DL (1987) Interspecific competition and the abun-dance of nest sites: factors affecting sexual selection.Ecology 68:1844–1855
Campana SE (2001) Accuracy, precision and quality control inage determination, including a review of the use andabuse of age validation methods. J Fish Biol 59:197–242
Charnov EL (1993) Life history invariants: some explorationsof symmetry in evolutionary ecology. Oxford UniversityPress, Oxford
Chen Y, Jackson DA, Harvey HH (1992) A comparison of vonBertalanffy and polynomial functions in modelling fishgrowth data. Can J Fish Aquat Sci 49:1228–1235
Choat JH, Axe LM (1996) Growth and longevity in acanthuridfishes; an analysis of otolith increments. Mar Ecol Prog Ser134:15–26
Choat JH, Robertson DR (2002) Age-based studies. In: Sale PF(ed) Coral reef fishes: dynamics and diversity in a complexecosystem. Academic Press, San Diego, p 57–80
Clark E, Stoll MJ, Alburn TK, Petzold R (2000) Mound-build-ing and feeding behaviour of the twostripe goby, Valenci-ennea helsdingenii, in the south Red Sea. Environ BiolFish 57:131–141
Emlen ST, Oring LW (1977) Ecology, sexual selection and theevolution of mating systems. Science 197:215–223
Forrester GE (1995) Strong density-dependent survival andrecruitment regulate the abundance of a coral reef fish.Oecologia 103:275–282
Forsgren E, Kvarnemo C, Lindström K (1996) Mode of sexualselection determined by resource abundance in two sandgoby populations. Evolution 50:646–654
Gibson RN (1970) Observations on the biology of the giantgoby Gobius cobitis Pallas. J Fish Biol 2:281–288
Gill HS, Wise BS, Potter IC, Chaplin JA (1996) Biannual
spawning periods and resultant divergent patterns ofgrowth in the estuarine goby Pseudogobius olorum: tem-perature-induced? Mar Biol 125:453–466
Gladstone W, Westoby M (1988) Growth and reproduction inCanthigaster valentini (Pisces, Tetraodontidae): a compar-ison of a toxic reef fish with other reef fishes. Environ BiolFish 21:207–221
Grossman GD (1979) Demographic characteristics of an inter-tidal bay goby (Lepidogobius lepidus). Environ Biol Fish 4:207–218
Grossman GD (1980) Food, fights, and burrows: the adaptivesignificance of intraspecific aggression in the bay goby(Pisces: Gobiidae). Oecologia 45:261–266
Gust N, Choat JH, Ackerman JL (2002) Demographic plastic-ity in tropical reef fishes. Mar Biol 140:1039–1051
Hart AM, Russ GR (1996) Response of herbivorous fishes tocrown-of-thorns starfish Acanthaster planci outbreaks. III.Age, growth, mortality and maturity indices of Acanthurusnigrofuscus. Mar Ecol Prog Ser 136:25–35
Hernaman V (2003) A comparative analysis of the life historyand ecology of five species of coral reef goby (Teleostei:Gobiidae). PhD thesis, University of Otago, Dunedin
Hernaman V, Munday PL, Schläppy ML (2000) Validation ofotolith growth-increment periodicity in tropical gobies.Mar Biol 137:715–726
Hill KT, Radtke RL (1988) Gerontological studies of the dam-selfish, Dascyllus albisella. Bull Mar Sci 42:424–434
Iglesias M, Brothers EB, Morales-Nin B (1997) Validation ofdaily increment deposition in otoliths: age and growthdetermination of Aphia minuta (Pisces: Gobiidae) from thenorthwest Mediterranean. Mar Biol 129:279–287
Kamler E (1992) Early life history of fish: an energeticsapproach. Fish and Fisheries Series 4. Chapman & Hall,London
Kimura DK (1980) Likelihood methods for the von Bertalanffygrowth curve. Fish Bull 77:765–776
Kingsford MJ (1992) Spatial and temporal variation in preda-tion on reef fishes by coral trout (Plectropomus leopardus,Serranidae). Coral Reefs 11:193–198
Kovacic M (2001) The biology of Roule's goby in the Kvarnerarea, northern Adriatic Sea. J Fish Biol 59:795–809
Kritzer JP (2002) Stock structure, mortality and growth of thedecorated goby, Istigobius decoratus (Gobiidae), at LizardIsland, Great Barrier Reef. Environ Biol Fish 63:211–216
Kuwamura T, Nakashima Y, Yogo Y (1996) Plasticity in sizeand age at maturity in a monogamous fish: effect of hostcoral size and frequency dependence. Behav Ecol Socio-biol 38:365–370
Leis JM, Rennis DS (1983) The larvae of Indo-Pacific coralreef fishes. New South Wales University Press, Sydney,and University of Hawaii Press, Honolulu, in associationwith The Australian Museum
Lindström K (1992a) The effect of resource holding potential,nest size, and information about resource quality on theoutcome of intruder-owner conflicts in the sandgoby.Behav Ecol Sociobiol 30:53–58
Lindström K (1992b) Female spawning patterns and malemating success in the sand goby Pomatoschistus minutus.Mar Biol 113:475–480
MacNeill DB, Brandt SB (1990) Ontogenetic shift in gill rakermorphology and predicted prey capture efficiency of thealewife Alosa psuedoharengus. Copeia 1990:164–171
Magnhagen C (1993) Conflicting demands in gobies: when toeat, reproduce, and avoid predators. In: Huntingford F,Torricelli P (eds) Behavioural ecology of fishes, Vol 23.Taylor & Francis, London, p 79–90
Mazzoldi C, Poltronieri C, Rasotto MB (2002) Egg size
220

Hernaman & Munday: Coral reef gobies. I
variability and mating system in the marbled gobyPomatoschistus marmoratus (Pisces: Gobiidae). Mar EcolProg Ser 233:231–239
McCormick MI (1998) Ontogeny of diet shifts by a microcar-nivorous fish, Cheilodactylus spectabilis: relationshipbetween feeding mechanics, microhabitat selection andgrowth. Mar Biol 132:9–20
Meekan MG, Ackerman JL, Wellington GM (2001) Demogra-phy and age structures of coral reef damselfishes in thetropical eastern Pacific Ocean. Mar Ecol Prog Ser 212:223–232
Miller PJ (1961) Age, growth, and reproduction of the rockgoby, Gobius paganellus L., in the Isle of Man. J Mar BiolAssoc UK 41:737–769
Miller PJ (1979) Adaptiveness and implications of small sizein teleosts. Symp Zool Soc Lond 44:263–306
Miller PJ (1984) The tokology of gobioid fishes. In: Potts GW,Wootton RJ (eds) Fish reproduction: strategies and tactics.Academic Press, London, p 120–153
Miller PJ (1996) The functional ecology of small fish: someopportunities and consequences. In: Miller PJ (ed) Minia-ture vertebrates: the implications of small body size,Vol 69. The Zoological Society of London, Oxford,p 175–199
Moreira F, Costa JL, Almeida PR, Assis C, Costa MJ (1991)Age determination in Pomatoschistus minutus (Pallas)and Pomatoschistus microps (Kroyer) (Pisces: Gobiidae)from the upper Tagus estuary, Portugal. J Fish Biol 39:433–440
Munday PL (2001) Fitness consequences of habitat use andcompetition among coral-dwelling fishes. Oecologia 128:585–593
Munday PL, Jones GP (1998) The ecological implications ofsmall body size among coral-reef fishes. Oceanogr MarBiol Annu Rev 36:373–411
Munday PL, Wilson SK (1997) Comparative efficacy of cloveoil and other chemicals in anaesthetization of Pomacen-trus amboinensis, a coral reef fish. J Fish Biol 51:931–938
Newman SJ, Williams DM, Russ GR (1996) Age validation,growth and mortality rates of the tropical snappers (Pisces:Lutjanidae) Lutjanus adetii (Castelnau, 1873) and L. quin-quelineatus (Bloch, 1790) from the Central Great BarrierReef, Australia. Mar Freshw Res 47:575–584
Newman SJ, Cappo M, Williams DM (2000) Age, growth andmortality of the stripey, Lutjanus carponatatus (Richard-son) and the brown-stripe snapper, L. vitta (Quoy andGaimard) from the central Great Barrier Reef, Australia.Fish Res 48:263–275
Norris JE, Parrish JD (1988) Predator–prey relationshipsamong fishes in pristine coral reef communities. Proc 6thInt Coral Reef Symp 2:107–113
Parrish JD, Norris JE, Callahan MW, Callahan JK, MagarifujiEJ, Schroeder RE (1986) Piscivory in a coral reef fishcommunity. In: Simenstad CA, Cailliet GM (eds) Contem-porary studies on fish feeding. Dr. W. Junk Publishers,Dordrecht, p 285–297
Partridge L, Harvey PH (1988) The ecological context of lifehistory evolution. Science 241:1449–1455
Pauly D (1980) On the interrelationships between naturalmortality, growth parameters, and mean environmentaltemperature in 175 fish stocks. J Cons Int Explor Mer 39:175–192
Randall JE, Allen GR, Steene RC (1997) Fishes of the GreatBarrier Reef and Coral Sea. Crawford House Publishing,Bathurst
Reavis RH (1997a) The natural history of a monogamouscoral-reef fish, Valenciennea strigata (Gobiidae): 1. Abun-dance, growth, survival and predation. Environ Biol Fish49:239–246
Reavis RH (1997b) The natural history of a monogamouscoral-reef fish, Valenciennea strigata (Gobiidae): 2.Behaviour, mate fidelity, and reproductive success.Environ Biol Fish 49:247–257
Reavis RH, Barlow GW (1998) Why is the coral-reef fishValenciennea strigata (Gobiidae) monogamous? BehavEcol Sociobiol 43:229–237
Robertson DR (1996) Interspecific competition controls abun-dance and habitat use of territorial Caribbean dam-selfishes. Ecology 77:885–899
Robertson DR, Kaufmann KW (1998) Assessing early recruit-ment dynamics and its demographic consequencesamong tropical reef fishes: accommodating variation inrecruitment seasonality and longevity. Aust J Ecol 23:226–233
Roff DA (1980) A motion for the retirement of the Von Berta-lanffy function. Can J Fish Aquat Sci 37:127–129
Roff DA (1992) The evolution of life histories. Chapman &Hall, London
Sano M (1997) Temporal variation in density dependence:recruitment and postrecruitment demography of a tem-perate zone sand goby. J Exp Mar Biol Ecol 214:67–84
Schnute J (1981) A versatile growth model with statisticallystable parameters. Can J Fish Aquat Sci 38:1128–1140
Shafer DJ (1998) Early life history, growth and settlementdynamics of a tropical reef fish (Gobiidae: Bathygobiuscoalitus). PhD thesis, University of Hawaii, Honolulu
Stearns SC (1992) The evolution of life histories. Oxford Uni-versity Press, Oxford
Sweatman HPA (1984) A field study of the predatory behav-iour and feeding rate of a piscivorous coral reef fish, thelizardfish Synodus englemani. Copeia 1984(1):187–194
Takegaki T, Nakazono A (1999) Reproductive behavior andmate fidelity in the monogamous goby, Valenciennealongipinnis. Ichthyol Res 46:115–123
Warner RR (1984) Deferred reproduction as a response to sex-ual selection in a coral reef fish: a test of the life historicalconsequences. Evolution 38:148–162
Wootton RJ (1998) Ecology of teleost fishes. Kluwer AcademicPublishers, Dordrecht
Worthington DG, Doherty PJ, Fowler AJ (1995) Variation inthe relationship between otolith weight and age: implica-tions for the estimation of age of two tropical damselfish(Pomacentrus moluccensis and P. wardi). Can J Fish AquatSci 52:233–242
Yanagisawa Y (1982) Social behaviour and mating system ofthe gobiid fish Amblyeleotris japonica. Jpn J Ichthyol 28:401–422
221
Editorial responsibility: Charles Birkeland (Contributing Editor),Honolulu, Hawaii, USA
Submitted: November 2, 2003; Accepted: September 12, 2004Proofs received from author(s): March 15, 2005
Related Documents











