Let the Force Be With Us: Dyads Exploit Haptic Coupling for Coordination Robrecht P. R. D. van der Wel, Guenther Knoblich, and Natalie Sebanz Radboud University Nijmegen People often perform actions that involve a direct physical coupling with another person, such as when moving furniture together. Here, we examined how people successfully coordinate such actions with others. We tested the hypothesis that dyads amplify their forces to create haptic information to coordinate. Participants moved a pole (resembling a pendulum) back and forth between two targets at different amplitudes and frequencies. They did so by pulling on cords attached to the base of the pole, one on each side. In the individual condition, one participant performed this task bimanually, and in the joint condition two participants each controlled one cord. We measured the moment-to-moment pulling forces on each cord and the pole kinematics to determine how well individuals and dyads performed. Results indicated that dyads produced much more overlapping forces than individuals, especially for tasks with higher coordination requirements. Thus, the results suggest that dyads amplify their forces to generate a haptic information channel. This likely reflects a general coordination principle in haptic joint action, where force amplification allows dyads to perform at the same level as individuals. Keywords: joint action, interpersonal coordination, haptics Much of what people do concerns coordinating their own ac- tions with those of others, including actions such as carrying furniture, building bridges, playing sports, and dancing together (Sebanz, Bekkering, & Knoblich, 2006). These examples indicate that people can do many things together that they could not do alone. Owing to this observation, research on the mechanisms underlying joint action is a rapidly developing research domain. Here, we focus on one aspect of joint action, namely joint action coordination. We use this term to refer to situations in which two or more actors coordinate their actions under real time constraints with or without the explicit intention to do so. Joint action coordination takes on many different forms, includ- ing rowing a boat, walking hand in hand, having a conversation with another person, or performing a wave in a stadium full of people. The diversity of these examples indicates that such coor- dination tasks may differ from one another in several ways. First, information about other actors may be available and exchanged to varying degrees through different modalities (Clark, 1996). The usefulness of these modalities for achieving successful coordina- tion may differ between tasks. Second, asymmetries may exist between coactors (see Brennan, Chen, Dickinson, Neider, & Ze- linsky, 2008; Brennan & Hanna, 2009), both at the level of the information available to them (e.g., when two people row a boat while one is sitting behind the other) and in relation to the possible roles coactors fulfill for coordination. For example, in some cases one actor may be the leader and another actor a follower (e.g., in dance), while in other cases different actors may have the same roles. Third, joint action coordination tasks may require continu- ous coordination with others (e.g., while dancing a waltz) or may require coordination at particular points in time (e.g., when taking turns in a conversation). A complete theory of joint action coordination will ultimately require an understanding of which role different modalities as well as social factors and different time constraints play for such coordination (Galantucci & Sebanz, 2009; Schmidt, Fitzpatrick, Caron, & Mergeche, in press; Vesper, Butterfill, Knoblich, & Sebanz, in press). One issue in this regard concerns how providing different opportunities for coupling between coactors within a dyad may shape the enfolding dynamics of joint coordination. The notion of coupling refers to the extent to which the movements of two or more parts of a system (e.g., two actors in a dyad) show regularities in their relative behavior over time. Such coupling plays a key role in the Haken-Kelso-Bunz (HKB) model of coor- dination dynamics (Haken, Kelso, & Bunz, 1985). This coupled- oscillator model was originally developed to account for rhythmic interlimb coordination, but has since been extended to rhythmic interpersonal coordination as well. Within the model, a coupling function quantifies the extent to which the component parts (e.g., two limbs) of the moving system are linked. Coupling may emerge from the presence of a mechanical link- age (as in interlimb coupling or in physically connected dyads) or from the presence of an informational linkage between component parts of the system. Informational coupling could rely on visual, auditory, and haptic information. Theoretically, providing more This article was published Online First March 21, 2011. Robrecht P. R. D. van der Wel, Guenther Knoblich, and Natalie Sebanz, Donders Institute for Brain, Cognition, and Behaviour, Radboud University Nijmegen, Nijmegen, the Netherlands. The research was performed with support from a EURYI grant awarded to Natalie Sebanz. We thank Chris Frith, Uta Frith, Ruud Meulenbroek, Claire Michaels, Michael Richardson, Howie Zelaznik, and an anonymous reviewer for helpful comments. We also thank the Instrumentmakerij at Radboud University for building the experimental apparatus. Correspondence concerning this article should be addressed to Robrecht P. R. D. van der Wel, Donders Institute for Brain, Cognition, and Behav- iour, Radboud University, Nijmegen, Montessorilaan 3, 6525 HR Nijme- gen. E-mail: [email protected] Journal of Experimental Psychology: © 2011 American Psychological Association Human Perception and Performance 2011, Vol. 37, No. 5, 1420 –1431 0096-1523/11/$12.00 DOI: 10.1037/a0022337 1420

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Let the Force Be With Us:Dyads Exploit Haptic Coupling for Coordination
Robrecht P. R. D. van der Wel, Guenther Knoblich, and Natalie SebanzRadboud University Nijmegen
People often perform actions that involve a direct physical coupling with another person, such as whenmoving furniture together. Here, we examined how people successfully coordinate such actions withothers. We tested the hypothesis that dyads amplify their forces to create haptic information tocoordinate. Participants moved a pole (resembling a pendulum) back and forth between two targets atdifferent amplitudes and frequencies. They did so by pulling on cords attached to the base of the pole,one on each side. In the individual condition, one participant performed this task bimanually, and in thejoint condition two participants each controlled one cord. We measured the moment-to-moment pullingforces on each cord and the pole kinematics to determine how well individuals and dyads performed.Results indicated that dyads produced much more overlapping forces than individuals, especially fortasks with higher coordination requirements. Thus, the results suggest that dyads amplify their forces togenerate a haptic information channel. This likely reflects a general coordination principle in haptic jointaction, where force amplification allows dyads to perform at the same level as individuals.
Keywords: joint action, interpersonal coordination, haptics
Much of what people do concerns coordinating their own ac-tions with those of others, including actions such as carryingfurniture, building bridges, playing sports, and dancing together(Sebanz, Bekkering, & Knoblich, 2006). These examples indicatethat people can do many things together that they could not doalone. Owing to this observation, research on the mechanismsunderlying joint action is a rapidly developing research domain.Here, we focus on one aspect of joint action, namely joint actioncoordination. We use this term to refer to situations in which twoor more actors coordinate their actions under real time constraintswith or without the explicit intention to do so.
Joint action coordination takes on many different forms, includ-ing rowing a boat, walking hand in hand, having a conversationwith another person, or performing a wave in a stadium full ofpeople. The diversity of these examples indicates that such coor-dination tasks may differ from one another in several ways. First,information about other actors may be available and exchanged tovarying degrees through different modalities (Clark, 1996). Theusefulness of these modalities for achieving successful coordina-tion may differ between tasks. Second, asymmetries may exist
between coactors (see Brennan, Chen, Dickinson, Neider, & Ze-linsky, 2008; Brennan & Hanna, 2009), both at the level of theinformation available to them (e.g., when two people row a boatwhile one is sitting behind the other) and in relation to the possibleroles coactors fulfill for coordination. For example, in some casesone actor may be the leader and another actor a follower (e.g., indance), while in other cases different actors may have the sameroles. Third, joint action coordination tasks may require continu-ous coordination with others (e.g., while dancing a waltz) or mayrequire coordination at particular points in time (e.g., when takingturns in a conversation).
A complete theory of joint action coordination will ultimatelyrequire an understanding of which role different modalities as wellas social factors and different time constraints play for suchcoordination (Galantucci & Sebanz, 2009; Schmidt, Fitzpatrick,Caron, & Mergeche, in press; Vesper, Butterfill, Knoblich, &Sebanz, in press). One issue in this regard concerns how providingdifferent opportunities for coupling between coactors within adyad may shape the enfolding dynamics of joint coordination. Thenotion of coupling refers to the extent to which the movements oftwo or more parts of a system (e.g., two actors in a dyad) showregularities in their relative behavior over time. Such couplingplays a key role in the Haken-Kelso-Bunz (HKB) model of coor-dination dynamics (Haken, Kelso, & Bunz, 1985). This coupled-oscillator model was originally developed to account for rhythmicinterlimb coordination, but has since been extended to rhythmicinterpersonal coordination as well. Within the model, a couplingfunction quantifies the extent to which the component parts (e.g.,two limbs) of the moving system are linked.
Coupling may emerge from the presence of a mechanical link-age (as in interlimb coupling or in physically connected dyads) orfrom the presence of an informational linkage between componentparts of the system. Informational coupling could rely on visual,auditory, and haptic information. Theoretically, providing more
This article was published Online First March 21, 2011.Robrecht P. R. D. van der Wel, Guenther Knoblich, and Natalie Sebanz,
Donders Institute for Brain, Cognition, and Behaviour, Radboud UniversityNijmegen, Nijmegen, the Netherlands.
The research was performed with support from a EURYI grant awardedto Natalie Sebanz. We thank Chris Frith, Uta Frith, Ruud Meulenbroek,Claire Michaels, Michael Richardson, Howie Zelaznik, and an anonymousreviewer for helpful comments. We also thank the Instrumentmakerij atRadboud University for building the experimental apparatus.
Correspondence concerning this article should be addressed to RobrechtP. R. D. van der Wel, Donders Institute for Brain, Cognition, and Behav-iour, Radboud University, Nijmegen, Montessorilaan 3, 6525 HR Nijme-gen. E-mail: [email protected]
Journal of Experimental Psychology: © 2011 American Psychological AssociationHuman Perception and Performance2011, Vol. 37, No. 5, 1420–1431
0096-1523/11/$12.00 DOI: 10.1037/a0022337
1420

ways for dyads to informationally couple their individual actioncontributions to each other should stabilize the dynamics of jointcoordination and create greater similarity between interpersonaland interlimb coordination. In this article, we focus on the rolehaptic (touch) information may play for achieving successful co-ordination in a continuous interpersonal coordination task. Beforewe discuss the possible role of haptic information for joint coor-dination, we will shortly review the insights obtained in previousresearch on interpersonal coordination based on visual informationexchange.
Many previous studies on continuous interpersonal coordinationhave focused on coordination based on visual information ex-change. In line with the HKB model, these studies indicated thatthe macroscopic dynamical principles for visually guided interper-sonal coordination are very similar to those underlying intraper-sonal interlimb coordination. Thus, when two individuals producerhythmic movements while they can see each other, their move-ments tend to become coupled in much the same way as thecoupling that emerges between two moving limbs of one individ-ual. Such visually mediated coupling is reflected in the tendencyfor actors to unintentionally synchronize their actions and thetendency for in-phase movements to be more stable than antiphasemovements. These tendencies appear to be quite general, becausethey emerge when people walk (van Ulzen, Lamoth, Daffertshofer,Semin, & Beek, 2008), swing pendulums (Amazeen, Schmidt, &Turvey, 1995; Richardson, Marsh, & Schmidt, 2005; Schmidt,Bienvenu, Fitzpatrick, & Amazeen, 1998) or legs (Schmidt,Carello, & Turvey, 1990), or rock in rocking chairs (Richardson,Marsh, Isenhower, Goodman, & Schmidt, 2007) alongside eachother. The amount of visual information available has been shownto modulate the coupling strength between the movements of twopeople (Richardson et al., 2007). Thus, when participants rockchairs alongside each other while they only have peripheral infor-mation about each other’s movements, their coordination is lessstable than when they have full visual information. For verbalinformation exchange, Shockley and colleagues (Shockley, Baker,Richardson, & Fowler, 2007) similarly found that increasingspeaking rate (which could be conceived of as providing a strongercoupling) unintentionally resulted in an increase in shared posturalactivity between two speaking partners (but not between speakersof different pairs). Thus, increasing the available informationthrough different modalities could lead to stronger coupling be-tween components of a system.
The research on visually mediated coupling between individualssuggests strong similarities between individual and joint continu-ous coordination. However, these two forms of coordination ex-hibit important differences as well. Interpersonal coordinationbased on visual information alone typically shows a weaker cou-pling than interlimb coordination (e.g., Richardson et al., 2007;Richardson et al., 2005; Schmidt & O’Brien, 1997), which in partcould be because of the absence of a mechanical linkage betweenthe component parts. Perhaps resulting from this weaker coupling,increased coordination difficulties have been observed for inten-tional joint continuous coordination based exclusively on the shar-ing of visual information (e.g., Bosga & Meulenbroek, 2007;Knoblich & Jordan, 2003; Newman-Norlund, Bosga, Meulen-broek, & Bekkering, 2008). Although these initial difficulties canbe overcome, they suggest that being able to share informationthrough additional channels (i.e., not only visual) could potentially
help dyads to coordinate more effectively. In terms of the HKBmodel, adding information exchange through additional modalitiescould increase the coupling strength between components of thesystem (i.e., the actors within the dyad). Increasing the couplingstrength between actors could make interpersonal coordinationmore similar to interlimb coordination, which then could supportsuccessful performance.
How may information exchange through modalities other thanvision support intentional coordination in dyads? For some tasks,verbal information exchange forms a natural medium for coordi-nating with others. For example, people coordinate at discretepoints in time based on verbal information when they take turnsduring a phone conversation. On a longer time scale, people useverbal exchange to make plans to meet each other at some partic-ular time and place (Clark, 1996). Thus, verbal exchange is usefulfor establishing plans to coordinate in the absence of continuousreal-time constraints.
Verbal information exchange may be less useful when twoactors intentionally perform continuous coordination tasks, how-ever. Especially when people perform a joint action coordinationtask with a direct physical coupling between actors, adjustmentsneed to be made quickly and continuously. For example, when twopeople move a couch together the movements of each actor di-rectly influence the forces that operate on the couch as well as onthe other actor. For such tasks, sharing a haptic channel mayprovide a quick informational coupling that may enable dyads tocoordinate successfully. The role haptic information may play forcontinuous interpersonal coordination in the presence of visualinformation (such as when carrying a couch) has rarely beenexamined. Most studies on joint action to date have used para-digms that did not involve a direct physical coupling betweenactors. Here, we focus on coordination of physically coupled jointactions, in particular the use of haptic information for this kind ofcoordination.
Only a handful of studies to date have used a task in which twoactors were physically coupled to one another directly. In onestudy, Harrison and Richardson (2009) examined whether similargait dynamics emerge for the legs of a quadruped (e.g., horse)compared with the legs of two people whose upper bodies wereeither unconnected, visible for each other, or connected with afoam appendage, thus resembling the middle part of the body of ahorse. The physical connection between actors provided an oblig-atory coupling in this experiment, which should make interper-sonal coordination more similar to interlimb coordination. Theauthors asked pairs of participants to walk at different speeds andexamined the extent to which participants adopted one of fourpossible patterns that quadrupeds tend to adopt at different speeds.The results indicated that participants adopted quadruped-like gaitpatterns, and did so most strongly while they were physicallycoupled. Therefore, the authors argued that interpersonal and intra-personal coordination dynamics emerge from the same lawfulprinciples.
In a second study, Braun and colleagues (Braun, Ortega, &Wolpert, 2009) asked whether dyads behaved according to gametheoretical solutions (e.g., Camerer, 2003; Fudenberg & Tirole,1991; Sanfey, 2007) when they together influenced the dynamicsof a force field. In particular, in one condition two participantseach moved a robot arm in a two-dimensional force field to arectangular target area. In an individual control condition, one
1421LET THE FORCE BE WITH US

participant performed this same task bimanually. The task was tocontrol one dimension (e.g., x) with one robot arm, and to controlthe other dimension (e.g., y) with the other robot arm. However,both robot arms could also be used to control movement in therespective other dimension. Because of the range of target loca-tions created through the use of a rectangular target a range ofdifferent endpoints could be used to complete the task success-fully. It is important to note that the amount of movement in theirrelevant dimension for one robot arm directly affected the springstiffness of the other robot arm. Therefore, movements in theirrelevant dimension for each robot arm could ease the movementsof the other robot arm. Participants were told to achieve the task aseasily as possible. The question of interest was whether individualsand dyads would adjust their endpoints in the target to easemovements of the other robot arm.
The results indicated that whereas individuals tended to adoptsolutions that reduced the amount of force required for movingboth robot arms by moving in both dimensions, dyads predomi-nantly moved only in the dimension that was assigned to them.This solution implied that the movements for each robot armrequired more force, but also that the amount of force required forone participant of the dyad did not depend on the endpointsadopted by the other participant.
The results by Braun and colleagues raise the possibility thatwhen people need to coordinate forces with one another, theystrive for solutions that reduce the amount of interdependence.This possibility is difficult to square with the results of a thirdstudy on physically coupled joint actions, however. Reed et al.(2006) examined how pairs of participants coordinated their forceswhen they performed a joint target aiming task compared withwhen they performed the aiming task by themselves. In this task,participants manipulated a two-handled rigid crank such that theymoved it from a start orientation to a target orientation. A givenparticipant always controlled only one handle on the crank. Theother handle was either left uncontrolled (in the individual condi-tion), or moved by another participant (in the dyad condition). Inthe dyad condition, the movements to the targets always requiredthe same direction and extent of rotation of the crank for the twoparticipants. To ensure that a participant could in principle com-plete the task with the same amount of force in the individual anddyad conditions, the inertia of the crank was doubled in the dyadcondition.
Although participants were instructed to complete the targetaiming movements as quickly as possible in both conditions, theresults indicated that dyads performed the aiming task faster thanthe individual participants did. In fact, dyads performed the taskfaster than the faster of the two individuals in the pair performingthe task alone. Of interest, this finding emerged despite the factthat participants in the dyad condition generated large opposingforces. Based on the large overlapping forces and the observationthat dyads specialized such that one member mostly acceleratedand the other member mostly decelerated, the authors argued thatdyads used a haptic channel to cooperate.
The possibility that people overlap their force profiles to obtaincontinuous haptic information is highly interesting, because it mayindicate that people construct a haptic channel in the service ofcoordination. Within the HKB model, creating such a channelcould increase the coupling strength between actors and in turnsupport successful coordination performance, For individuals, it is
well known that haptic information can be very effective in guid-ing performance. For example, the lawful increase in choice reac-tion time with an increase in the number of stimulus-responsealternatives, known as Hick-Hyman law (Hick, 1952; Hyman,1953), does not hold when stimuli are presented haptically (Leon-hard, 1959). For bimanual coordination, the strong interferencethat is observed when people plan to simultaneously producedifferent movement patterns with their two hands, for example acircle with one hand and a straight line with the other hand (Franz,Zelaznik, & McCabe, 1991), is abolished when people merelyhaptically track moving disks creating different patterns for thetwo hands (Rosenbaum, Dawson, & Challis, 2006). Thus, hapticinformation can be an extremely powerful source to overcomeintra-individual coordination difficulties.
The study by Reed and colleagues raises the possibility thatdyads may exploit the haptic channel for effective coordination ofphysically coupled joint actions. However, the study has an im-portant caveat. Because the control condition involved movementsof one actor on only one side of the manipulandum, force overlapcould not exist in the individual case. As a result, it was notpossible to know whether the large force overlap in the dyadcondition arose because of the interaction with another actor, orbecause the device needed to be controlled on both sides.
In the current study, we tested the hypothesis that dyads showmore force overlap than individuals when they coordinate physi-cally coupled actions. Creating such overlap would generate ahaptic information exchange that could, in turn, support successfulcoordination. To ensure that we could evaluate the amount of forceoverlap of dyads against a meaningful individual baseline, wedeveloped a task that people either performed bimanually (con-trolling both sides, one with each hand), or together with anotherperson (controlling one side per person). In particular, we askedparticipants to rotate a pole that moved around a fixed axis (i.e., amodified pendulum) back and forth between two targets. They didso by pulling on cords attached to the base of the pole, one on eachside. This formed a challenging task as the timing and force ofpulls on either cord influenced the displacement of the pole. Thus,the task required continuous coordination between the two sides ofthe apparatus. By measuring the forces on each cord, we couldevaluate the proportion and strength of overlapping forces. Thetask also allowed us to determine how quickly and successfullyindividuals and dyads adopted a coordination pattern for a noveltask.
Given that individuals have and dyads do not have internalinformation about what the other hand is doing, one may expectthat individuals will learn a novel task more quickly than dyadsand that dyads gradually approach bimanual performance. Such afinding would be consistent with previous literature on joint actioncoordination tasks that rely exclusively on visual information (e.g.,Bosga & Meulenbroek, 2007; Knoblich & Jordan, 2003; Newman-Norlund et al., 2008). However, it may be that providing dyadswith a haptic linkage (in addition to visual information) will allowdyads to reach the same coordination performance as would oneperson performing the same task bimanually from the start.
We tested the generality of the tendency to increase forceoverlap in dyads. In particular, we asked participants to performthe task for different target amplitudes and at different speeds. Thismanipulation allowed us to evaluate whether the adopted coordi-nation pattern and the amount of force overlap changed as a
1422 VAN DER WEL, KNOBLICH, AND SEBANZ

function of the spatiotemporal task requirements, in particular, thefrequency and amplitude of the required movements. With in-creases in the required frequency and decreases in the requiredamplitudes, the two sides of the task may require different coor-dination patterns because of the spatial and/or temporal con-straints. Thus, different coordination patterns may emerge acrossthe spatiotemporal continuum, as has been observed for bimanualcoordination (e.g., Kelso, 1984). If so, these different coordinationpatterns may differentially rely on force overlap. Alternatively, itmay be that generating more force overlap forms a general char-acteristic of dyadic coordination.
Finally, besides examining whether dyads would show moreforce overlap than individuals we also examined how dyads usethe haptic channel to coordinate. In particular, we examinedwhether dyads use the haptic channel in an intermittent or in acontinuous way to coordinate. At one extreme, dyads may use anintermittent coordination strategy such that they produce forceoverlap at specific parts of the task. At the other extreme, dyadsmay produce force overlap for the full duration of the task toobtain haptic information continuously.
Method
Participants
Fifty-four participants took part in this experiment. Eighteenparticipants performed the task individually (6 males and 12 fe-males between the ages of 17 and 32) and 36 participants per-formed the task in dyads (1 male dyad, 11 female dyads, and 6mixed gender dyads, all between the ages of 17 and 38). Allparticipants were right-handed and none reported any neurologicaldeficits. Participants were compensated for their time, either mon-etarily or though course credit. Data from three individuals wereremoved from the data set because of recording error.
Apparatus and Procedure
Figure 1 shows the experimental apparatus. In the individualgroup, participants controlled the cord on the left with their lefthand and the cord on the right with their right hand. In dyads,participants sat next to each other at a distance of approximately 30
cm. The participant on the left controlled the cord on the left withtheir left hand, and the participant on the right controlled the cordon the right with their right hand.
At the beginning of the experiment, participants were told thaton each trial they would be asked to move the pole back and forthbetween two targets by pulling on each of the two cords. Theywere also told to do this at a frequency that approximated asequences of tones played before the onset of the trial (detailsfollow). No indication was given about how participants shouldaccomplish this task. Participants were then given a chance to tryout the task for approximately one minute. Dyads were also askednot to communicate with each other verbally.
During the experimental trials, participants started with the poleresting on the left side, and they could start moving the pole afterhearing an isochronous sequence of tones that indicated at whichspeed the movement should be performed. We used a customizedMatlab program to play a sound file that indicated the pace atwhich the pole should approximately be moved back and forthbetween the targets. The sounds participants heard consisted ofeight alternating 700 and 850 Hertz tones (corresponding to 4movement cycles) at the relevant target period. Participants lis-tened to these tones, and were told to start moving the pole atapproximately the same rate after the tones had played. Thus, theparticipants did not hear any tones during pole movement.
Participants were told to continue moving the pole back andforth until the experimenter told them to stop. The experimenterdid so after participants completed 15 back and forth movementsbetween the targets. None of the participants appeared to havetrouble understanding the instructions.
Participants completed a total of 9 conditions that differed withrespect to their spatiotemporal coordination requirements, suchthat participants moved the pole over different amplitudes and atdifferent periods. Our rationale for introducing these manipula-tions was to have a way of testing the generality of the use ofhaptic information. For bimanual coordination, the spatiotemporalrequirements influence the stability of the emerging coordinationpatterns (e.g., Kelso, 1984). As interpersonal coordination is alsosensitive to the spatiotemporal demands (e.g., Richardson et al.,2007), it provided a useful vehicle for testing generalities in forceoverlap. We used all combinations of 3 amplitudes (4, 10, or 16 cm
Figure 1. Overview of the experimental apparatus, and the task for individuals and dyads.
1423LET THE FORCE BE WITH US

between the targets) and 3 target periods (462, 546, or 667 msperiods corresponding to fast, medium, and slow, respectively).We chose the particular amplitudes and periods because theyseemed to cover the range for which the task was possible yetdifferentially challenging (based on informal pilot testing). Wecounterbalanced the order of conditions for individuals and dyads.Each condition was completed as a block of 5 repetitions, thuscreating 45 trials in total. The experiment lasted about 45 to 50minutes.
The pole was made from a solid PVC pipe (length � 46.6 cm,diameter � 1.0 cm, mass � 50.7 g), and was affixed to a rotatingaxis positioned at its base, thus creating a pivot point. A cord wasattached at 0.5 cm below the pivot point on each side of the pole.The cords were 65 cm long. Participants always held each cord inthe same location, at 30 cm from the pole, between their thumb andindex finger. To ensure a fixed relation between the pulling angleand the pole, we ran each cord through a small hole at the sameheight and 17 cm away from the pivot point of the pole. From rest,the pole required approximately 1.27 N of pulling force to startmoving.
Each target region was 3 cm in width and indicated by a coloredarea. The target regions were drawn on a white piece of cardboardthat was placed on a flat platform positioned 4.0 cm above thepivot point of the pole. Different pieces of cardboard were used forthe different amplitude conditions. The target regions on both sideswere always equidistant from the balance point of the pole.
A goniometer (Encoder Rotary 360 PPR, Avago Technologies)recorded the pole kinematics. We recorded the forces exerted oneach cord with force sensors (Low profile universal load cell,model LC703-25) that were positioned between the axis and thecord on each side. Both the goniometer and the force sensorsrecorded data at 200 Hz. They were connected to a HewlettPackard Compaq dc7900 computer through a USB connection.The force data were amplified with a strain gage amplifier(INA125P, Texas Instruments).
Data Analysis
Before calculating our dependent variables, we first filtered thepole kinematics and force data with a 20-Hz low-pass Butterworthfilter to remove noise. From the pole kinematics, we then calcu-lated the mean amplitude of the pole movements by calculating thedistance between direction reversal points of the pole. We used athreshold criterion of 20 degrees of movement for the amplitudevalues to avoid including small corrective movements in measur-ing amplitudes. We calculated the times between successive di-rection reversals (the same ones we used for the amplitude calcu-lations) to calculate the movement periods.
From the force data, we calculated two dependent variables totest the hypothesis that dyads display more force overlap thanindividual participants. First we calculated the mean proportion ofinstances of force overlap. We classified all data samples for whichthe pulling force exerted on each separate cord exceeded a thresh-old of 0.5 N as instances of force overlap. We chose this thresholdbased on the rationale that at this force level coordination based onhaptic information is possible (e.g., Rosenbaum et al., 2006). Forthe instances of force overlap, we also calculated the mean amountof force overlap to evaluate possible differences in the amount offorce exerted on both cords. To evaluate whether individuals and
dyads used haptic information intermittently or continuously, weconsidered how the instances of force overlap were distributedacross different parts of the movements by dividing each move-ment trajectory into quadrants.
Results
The results are reported in two sections. In the first section, wereport on the pole kinematics to evaluate how closely individualsand dyads matched the required amplitudes and target periods. Inthe second section, we provide the analyses on the force data toevaluate force overlap for individuals and dyads in each of theconditions. For both types of analyses (pole kinematics and forcedata), we examined the effects of our experimental manipulationson performance. For the pole kinematics, we also tested for learn-ing effects by examining the effect of the number of completedtrials on performance. To do so, we considered trials 1 to 15 asPhase 1, trials 16 to 30 as Phase 2, and trials 31 to 45 as Phase 3of the Experiment. For the force data, we performed a similaranalysis to determine whether force overlap changed as partici-pants learned the task. We applied a Greenhouse-Geisser correc-tion to the degrees of freedom when the assumption of sphericitywas violated.
Pole Kinematics
We calculated the mean absolute difference of the pole location(in degrees) relative to the alternating target locations to evaluatehow closely individuals and dyads matched the required ampli-tudes. We also used this mean absolute endpoint error to evaluatethe rate of learning in individuals and dyads. We used the durationsbetween pole reversal points to estimate the mean movementperiod in each trial.
Movement Amplitudes
To evaluate the effect of target amplitude and target period onperformance in individuals and dyads, we calculated the meanabsolute endpoint error for the five trial repetitions within a con-dition. Thus, we reduced the data for each individual and dyad toone value per condition. We then performed a 2 (individual vs.dyad) � 3 (target amplitude; small, medium, or large) � 3 (targetperiod; slow, medium, or fast) repeated-measures analysis of vari-ance (ANOVA) on the resulting values.
Figure 2 shows the results. The results indicated a main effect oftarget amplitude, F(2, 62) � 3.181, p � .05, partial �2 � .093,such that the mean absolute endpoint error significantly increasedfor the large amplitude (M � 4.22, SE � .41) compared with themedium (M � 3.28, SE � .30) and small (M � 3.33, SE � .38)amplitude conditions. The results showed no other significant maineffects or interactions, p � .10. Importantly, mean absolute end-point error did not differ for individuals (M � 3.61, SE � .40) anddyads (M � 3.61, SE �.37).
Movement Periods
We analyzed the movement periods in terms of their absolutevalues rather than in terms of their deviation from the prescribedtarget periods. The rationale for this approach was that participantsdid not hear the metronome while they performed the task. As a
1424 VAN DER WEL, KNOBLICH, AND SEBANZ
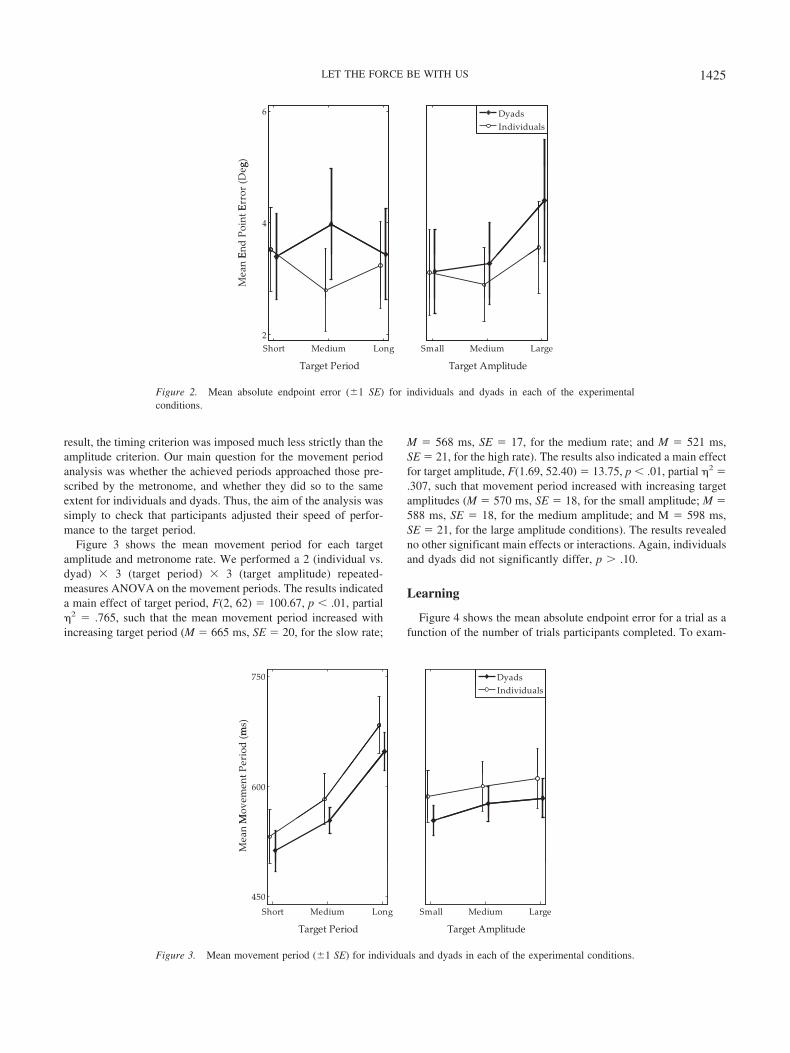
result, the timing criterion was imposed much less strictly than theamplitude criterion. Our main question for the movement periodanalysis was whether the achieved periods approached those pre-scribed by the metronome, and whether they did so to the sameextent for individuals and dyads. Thus, the aim of the analysis wassimply to check that participants adjusted their speed of perfor-mance to the target period.
Figure 3 shows the mean movement period for each targetamplitude and metronome rate. We performed a 2 (individual vs.dyad) � 3 (target period) � 3 (target amplitude) repeated-measures ANOVA on the movement periods. The results indicateda main effect of target period, F(2, 62) � 100.67, p � .01, partial�2 � .765, such that the mean movement period increased withincreasing target period (M � 665 ms, SE � 20, for the slow rate;
M � 568 ms, SE � 17, for the medium rate; and M � 521 ms,SE � 21, for the high rate). The results also indicated a main effectfor target amplitude, F(1.69, 52.40) � 13.75, p � .01, partial �2 �.307, such that movement period increased with increasing targetamplitudes (M � 570 ms, SE � 18, for the small amplitude; M �588 ms, SE � 18, for the medium amplitude; and M � 598 ms,SE � 21, for the large amplitude conditions). The results revealedno other significant main effects or interactions. Again, individualsand dyads did not significantly differ, p � .10.
Learning
Figure 4 shows the mean absolute endpoint error for a trial as afunction of the number of trials participants completed. To exam-
6
Dyads
g)
Individuals
Erro
r (D
eg
4
End
Poin
t EM
ean
E
2Short Medium Long
Target Period
Small Medium Large
Target Amplitude
Figure 2. Mean absolute endpoint error (�1 SE) for individuals and dyads in each of the experimentalconditions.
750
Dyads
ms)
Individuals
Peri
od (m
600
Mov
emen
t M
ean
M
450Short Medium Long
Target Period
Small Medium Large
Target Amplitude
Figure 3. Mean movement period (�1 SE) for individuals and dyads in each of the experimental conditions.
1425LET THE FORCE BE WITH US
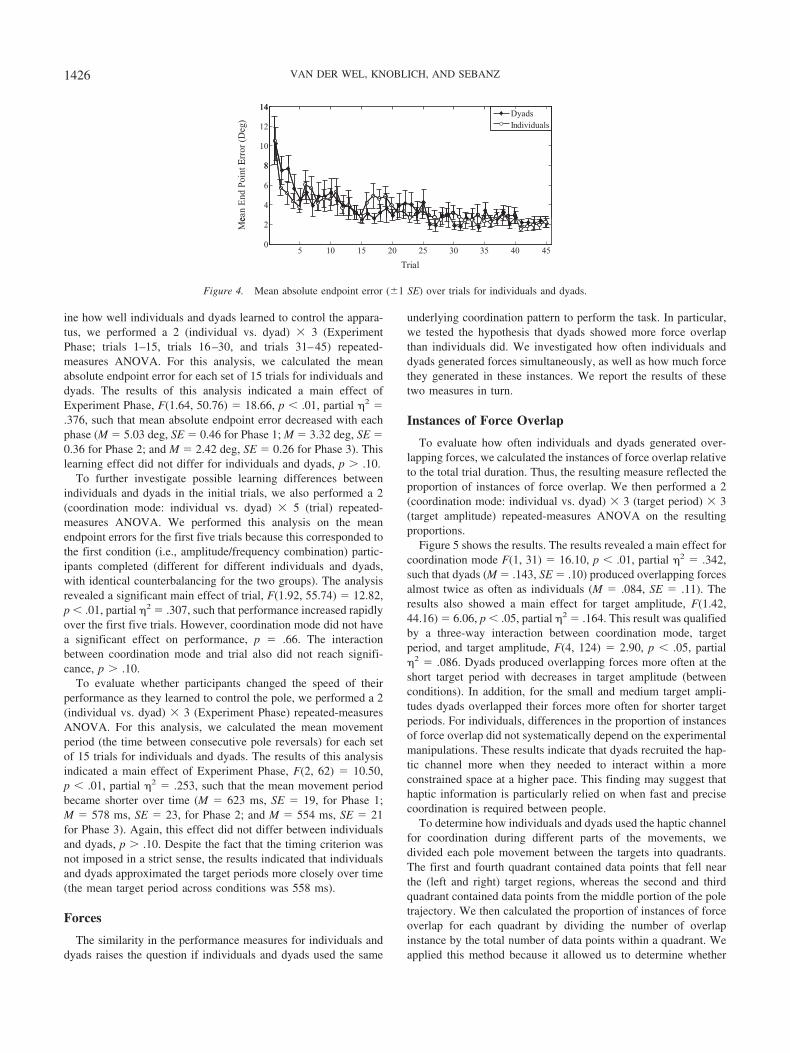
ine how well individuals and dyads learned to control the appara-tus, we performed a 2 (individual vs. dyad) � 3 (ExperimentPhase; trials 1–15, trials 16–30, and trials 31–45) repeated-measures ANOVA. For this analysis, we calculated the meanabsolute endpoint error for each set of 15 trials for individuals anddyads. The results of this analysis indicated a main effect ofExperiment Phase, F(1.64, 50.76) � 18.66, p � .01, partial �2 �.376, such that mean absolute endpoint error decreased with eachphase (M � 5.03 deg, SE � 0.46 for Phase 1; M � 3.32 deg, SE �0.36 for Phase 2; and M � 2.42 deg, SE � 0.26 for Phase 3). Thislearning effect did not differ for individuals and dyads, p � .10.
To further investigate possible learning differences betweenindividuals and dyads in the initial trials, we also performed a 2(coordination mode: individual vs. dyad) � 5 (trial) repeated-measures ANOVA. We performed this analysis on the meanendpoint errors for the first five trials because this corresponded tothe first condition (i.e., amplitude/frequency combination) partic-ipants completed (different for different individuals and dyads,with identical counterbalancing for the two groups). The analysisrevealed a significant main effect of trial, F(1.92, 55.74) � 12.82,p � .01, partial �2 � .307, such that performance increased rapidlyover the first five trials. However, coordination mode did not havea significant effect on performance, p � .66. The interactionbetween coordination mode and trial also did not reach signifi-cance, p � .10.
To evaluate whether participants changed the speed of theirperformance as they learned to control the pole, we performed a 2(individual vs. dyad) � 3 (Experiment Phase) repeated-measuresANOVA. For this analysis, we calculated the mean movementperiod (the time between consecutive pole reversals) for each setof 15 trials for individuals and dyads. The results of this analysisindicated a main effect of Experiment Phase, F(2, 62) � 10.50,p � .01, partial �2 � .253, such that the mean movement periodbecame shorter over time (M � 623 ms, SE � 19, for Phase 1;M � 578 ms, SE � 23, for Phase 2; and M � 554 ms, SE � 21for Phase 3). Again, this effect did not differ between individualsand dyads, p � .10. Despite the fact that the timing criterion wasnot imposed in a strict sense, the results indicated that individualsand dyads approximated the target periods more closely over time(the mean target period across conditions was 558 ms).
Forces
The similarity in the performance measures for individuals anddyads raises the question if individuals and dyads used the same
underlying coordination pattern to perform the task. In particular,we tested the hypothesis that dyads showed more force overlapthan individuals did. We investigated how often individuals anddyads generated forces simultaneously, as well as how much forcethey generated in these instances. We report the results of thesetwo measures in turn.
Instances of Force Overlap
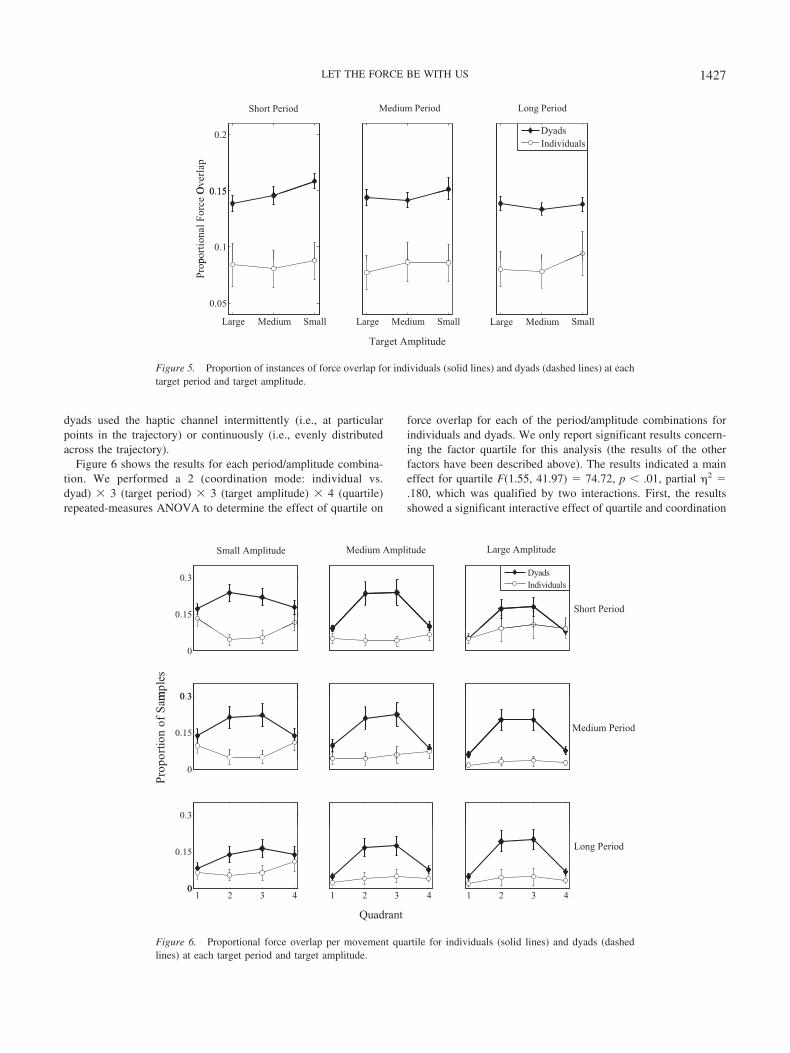
To evaluate how often individuals and dyads generated over-lapping forces, we calculated the instances of force overlap relativeto the total trial duration. Thus, the resulting measure reflected theproportion of instances of force overlap. We then performed a 2(coordination mode: individual vs. dyad) � 3 (target period) � 3(target amplitude) repeated-measures ANOVA on the resultingproportions.
Figure 5 shows the results. The results revealed a main effect forcoordination mode F(1, 31) � 16.10, p � .01, partial �2 � .342,such that dyads (M � .143, SE � .10) produced overlapping forcesalmost twice as often as individuals (M � .084, SE � .11). Theresults also showed a main effect for target amplitude, F(1.42,44.16) � 6.06, p � .05, partial �2 � .164. This result was qualifiedby a three-way interaction between coordination mode, targetperiod, and target amplitude, F(4, 124) � 2.90, p � .05, partial�2 � .086. Dyads produced overlapping forces more often at theshort target period with decreases in target amplitude (betweenconditions). In addition, for the small and medium target ampli-tudes dyads overlapped their forces more often for shorter targetperiods. For individuals, differences in the proportion of instancesof force overlap did not systematically depend on the experimentalmanipulations. These results indicate that dyads recruited the hap-tic channel more when they needed to interact within a moreconstrained space at a higher pace. This finding may suggest thathaptic information is particularly relied on when fast and precisecoordination is required between people.
To determine how individuals and dyads used the haptic channelfor coordination during different parts of the movements, wedivided each pole movement between the targets into quadrants.The first and fourth quadrant contained data points that fell nearthe (left and right) target regions, whereas the second and thirdquadrant contained data points from the middle portion of the poletrajectory. We then calculated the proportion of instances of forceoverlap for each quadrant by dividing the number of overlapinstance by the total number of data points within a quadrant. Weapplied this method because it allowed us to determine whether
14
8
10
12
14
Erro
r (D
eg)
DyadsIndividuals
4
6
8
ean
End
Poin
t E5 10 15 20 25 30 35 40 45
0
2
Trial
Me
Figure 4. Mean absolute endpoint error (�1 SE) over trials for individuals and dyads.
1426 VAN DER WEL, KNOBLICH, AND SEBANZ
dyads used the haptic channel intermittently (i.e., at particularpoints in the trajectory) or continuously (i.e., evenly distributedacross the trajectory).
Figure 6 shows the results for each period/amplitude combina-tion. We performed a 2 (coordination mode: individual vs.dyad) � 3 (target period) � 3 (target amplitude) � 4 (quartile)repeated-measures ANOVA to determine the effect of quartile on
force overlap for each of the period/amplitude combinations forindividuals and dyads. We only report significant results concern-ing the factor quartile for this analysis (the results of the otherfactors have been described above). The results indicated a maineffect for quartile F(1.55, 41.97) � 74.72, p � .01, partial �2 �.180, which was qualified by two interactions. First, the resultsshowed a significant interactive effect of quartile and coordination
Short Period Medium Period
0 15
0.2
Ove
rlap
DyadsIndividuals
0.1
0.15po
rtion
al F
orce
O
Large Medium Small
0.05
Prop
Large Medium Small Large Medium Small
Target Amplitude
Long Period
Figure 5. Proportion of instances of force overlap for individuals (solid lines) and dyads (dashed lines) at eachtarget period and target amplitude.
Small Amplitude Medium Amplitude Large Amplitude
0.15
0.3 DyadsIndividuals
Short Period
0
0 3mpl
es
0.15
0.3
ortio
n of
Sam
Medium Period
0
Prop
0.3
0
0.15 Long Period
1 2 3 40
1 2 3 4
Quadrant
1 2 3 4
Figure 6. Proportional force overlap per movement quartile for individuals (solid lines) and dyads (dashedlines) at each target period and target amplitude.
1427LET THE FORCE BE WITH US
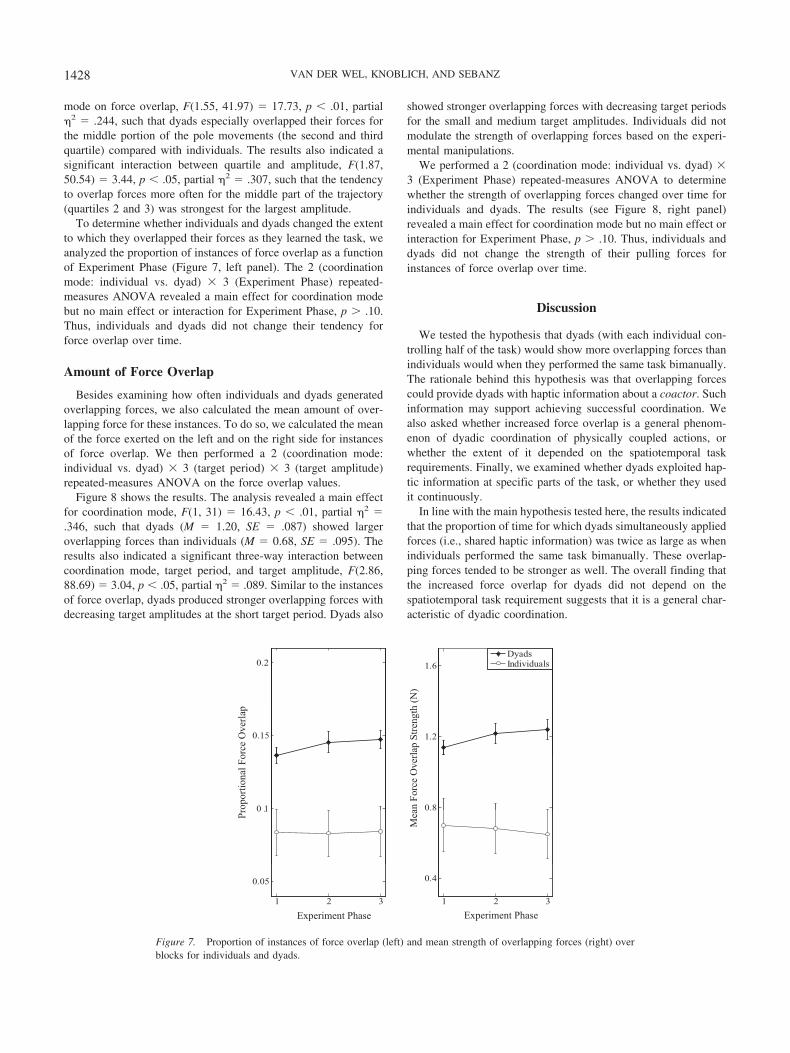
mode on force overlap, F(1.55, 41.97) � 17.73, p � .01, partial�2 � .244, such that dyads especially overlapped their forces forthe middle portion of the pole movements (the second and thirdquartile) compared with individuals. The results also indicated asignificant interaction between quartile and amplitude, F(1.87,50.54) � 3.44, p � .05, partial �2 � .307, such that the tendencyto overlap forces more often for the middle part of the trajectory(quartiles 2 and 3) was strongest for the largest amplitude.
To determine whether individuals and dyads changed the extentto which they overlapped their forces as they learned the task, weanalyzed the proportion of instances of force overlap as a functionof Experiment Phase (Figure 7, left panel). The 2 (coordinationmode: individual vs. dyad) � 3 (Experiment Phase) repeated-measures ANOVA revealed a main effect for coordination modebut no main effect or interaction for Experiment Phase, p � .10.Thus, individuals and dyads did not change their tendency forforce overlap over time.
Amount of Force Overlap
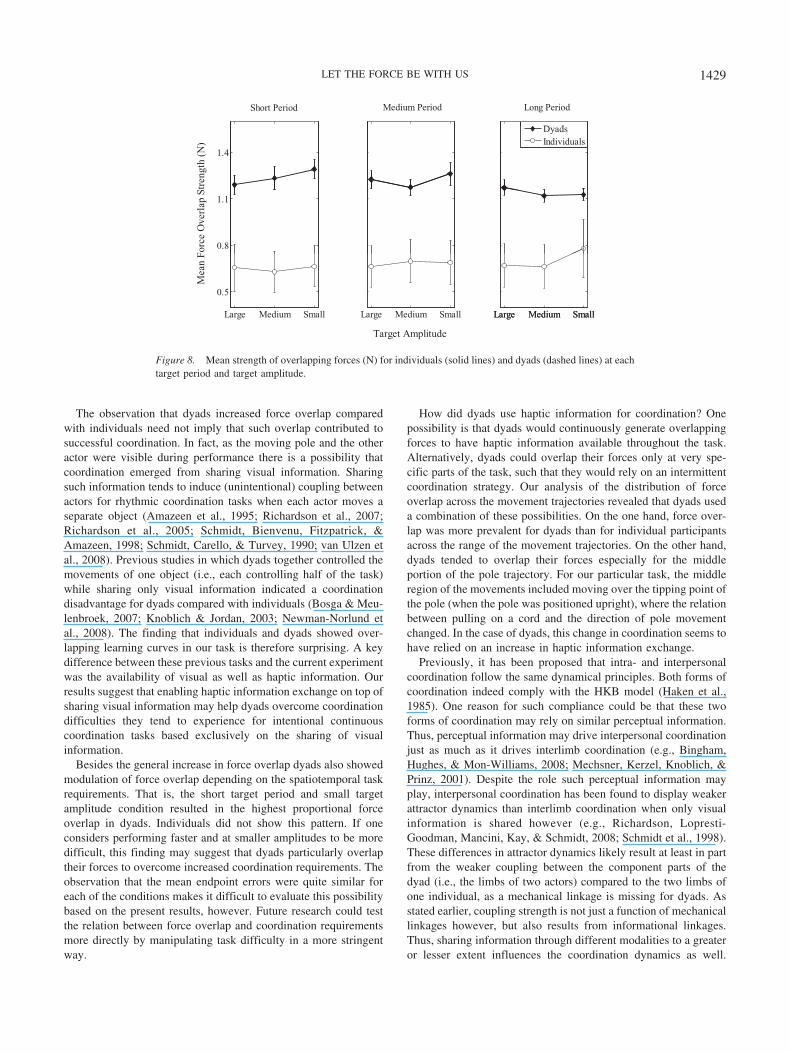
Besides examining how often individuals and dyads generatedoverlapping forces, we also calculated the mean amount of over-lapping force for these instances. To do so, we calculated the meanof the force exerted on the left and on the right side for instancesof force overlap. We then performed a 2 (coordination mode:individual vs. dyad) � 3 (target period) � 3 (target amplitude)repeated-measures ANOVA on the force overlap values.
Figure 8 shows the results. The analysis revealed a main effectfor coordination mode, F(1, 31) � 16.43, p � .01, partial �2 �.346, such that dyads (M � 1.20, SE � .087) showed largeroverlapping forces than individuals (M � 0.68, SE � .095). Theresults also indicated a significant three-way interaction betweencoordination mode, target period, and target amplitude, F(2.86,88.69) � 3.04, p � .05, partial �2 � .089. Similar to the instancesof force overlap, dyads produced stronger overlapping forces withdecreasing target amplitudes at the short target period. Dyads also
showed stronger overlapping forces with decreasing target periodsfor the small and medium target amplitudes. Individuals did notmodulate the strength of overlapping forces based on the experi-mental manipulations.
We performed a 2 (coordination mode: individual vs. dyad) �3 (Experiment Phase) repeated-measures ANOVA to determinewhether the strength of overlapping forces changed over time forindividuals and dyads. The results (see Figure 8, right panel)revealed a main effect for coordination mode but no main effect orinteraction for Experiment Phase, p � .10. Thus, individuals anddyads did not change the strength of their pulling forces forinstances of force overlap over time.
Discussion
We tested the hypothesis that dyads (with each individual con-trolling half of the task) would show more overlapping forces thanindividuals would when they performed the same task bimanually.The rationale behind this hypothesis was that overlapping forcescould provide dyads with haptic information about a coactor. Suchinformation may support achieving successful coordination. Wealso asked whether increased force overlap is a general phenom-enon of dyadic coordination of physically coupled actions, orwhether the extent of it depended on the spatiotemporal taskrequirements. Finally, we examined whether dyads exploited hap-tic information at specific parts of the task, or whether they usedit continuously.
In line with the main hypothesis tested here, the results indicatedthat the proportion of time for which dyads simultaneously appliedforces (i.e., shared haptic information) was twice as large as whenindividuals performed the same task bimanually. These overlap-ping forces tended to be stronger as well. The overall finding thatthe increased force overlap for dyads did not depend on thespatiotemporal task requirement suggests that it is a general char-acteristic of dyadic coordination.
0.2 1.6DyadsIndividuals
0.15 1.2
0 1 0.8.
Prop
ortio
nal F
orce
Ove
rlap
Mea
n Fo
rce
Ove
rlap
Stre
ngth
(N)
1 2 3
0.05
Experiment Phase1 2 3
0.4
Experiment Phase
Figure 7. Proportion of instances of force overlap (left) and mean strength of overlapping forces (right) overblocks for individuals and dyads.
1428 VAN DER WEL, KNOBLICH, AND SEBANZ
The observation that dyads increased force overlap comparedwith individuals need not imply that such overlap contributed tosuccessful coordination. In fact, as the moving pole and the otheractor were visible during performance there is a possibility thatcoordination emerged from sharing visual information. Sharingsuch information tends to induce (unintentional) coupling betweenactors for rhythmic coordination tasks when each actor moves aseparate object (Amazeen et al., 1995; Richardson et al., 2007;Richardson et al., 2005; Schmidt, Bienvenu, Fitzpatrick, &Amazeen, 1998; Schmidt, Carello, & Turvey, 1990; van Ulzen etal., 2008). Previous studies in which dyads together controlled themovements of one object (i.e., each controlling half of the task)while sharing only visual information indicated a coordinationdisadvantage for dyads compared with individuals (Bosga & Meu-lenbroek, 2007; Knoblich & Jordan, 2003; Newman-Norlund etal., 2008). The finding that individuals and dyads showed over-lapping learning curves in our task is therefore surprising. A keydifference between these previous tasks and the current experimentwas the availability of visual as well as haptic information. Ourresults suggest that enabling haptic information exchange on top ofsharing visual information may help dyads overcome coordinationdifficulties they tend to experience for intentional continuouscoordination tasks based exclusively on the sharing of visualinformation.
Besides the general increase in force overlap dyads also showedmodulation of force overlap depending on the spatiotemporal taskrequirements. That is, the short target period and small targetamplitude condition resulted in the highest proportional forceoverlap in dyads. Individuals did not show this pattern. If oneconsiders performing faster and at smaller amplitudes to be moredifficult, this finding may suggest that dyads particularly overlaptheir forces to overcome increased coordination requirements. Theobservation that the mean endpoint errors were quite similar foreach of the conditions makes it difficult to evaluate this possibilitybased on the present results, however. Future research could testthe relation between force overlap and coordination requirementsmore directly by manipulating task difficulty in a more stringentway.
How did dyads use haptic information for coordination? Onepossibility is that dyads would continuously generate overlappingforces to have haptic information available throughout the task.Alternatively, dyads could overlap their forces only at very spe-cific parts of the task, such that they would rely on an intermittentcoordination strategy. Our analysis of the distribution of forceoverlap across the movement trajectories revealed that dyads useda combination of these possibilities. On the one hand, force over-lap was more prevalent for dyads than for individual participantsacross the range of the movement trajectories. On the other hand,dyads tended to overlap their forces especially for the middleportion of the pole trajectory. For our particular task, the middleregion of the movements included moving over the tipping point ofthe pole (when the pole was positioned upright), where the relationbetween pulling on a cord and the direction of pole movementchanged. In the case of dyads, this change in coordination seems tohave relied on an increase in haptic information exchange.
Previously, it has been proposed that intra- and interpersonalcoordination follow the same dynamical principles. Both forms ofcoordination indeed comply with the HKB model (Haken et al.,1985). One reason for such compliance could be that these twoforms of coordination may rely on similar perceptual information.Thus, perceptual information may drive interpersonal coordinationjust as much as it drives interlimb coordination (e.g., Bingham,Hughes, & Mon-Williams, 2008; Mechsner, Kerzel, Knoblich, &Prinz, 2001). Despite the role such perceptual information mayplay, interpersonal coordination has been found to display weakerattractor dynamics than interlimb coordination when only visualinformation is shared however (e.g., Richardson, Lopresti-Goodman, Mancini, Kay, & Schmidt, 2008; Schmidt et al., 1998).These differences in attractor dynamics likely result at least in partfrom the weaker coupling between the component parts of thedyad (i.e., the limbs of two actors) compared to the two limbs ofone individual, as a mechanical linkage is missing for dyads. Asstated earlier, coupling strength is not just a function of mechanicallinkages however, but also results from informational linkages.Thus, sharing information through different modalities to a greateror lesser extent influences the coordination dynamics as well.
D d
Medium Period Long PeriodShort Period
1.4
Stre
ngth
(N)
DyadsIndividuals
0.8
1.1Fo
rce
Ove
rlap
Large Medium Small
0.5
Mea
n
Large Medium Small Large Medium Small
Target Amplitude
Large Medium Small
Figure 8. Mean strength of overlapping forces (N) for individuals (solid lines) and dyads (dashed lines) at eachtarget period and target amplitude.
1429LET THE FORCE BE WITH US

Additional access to haptic information may shift the attractordynamics for interpersonal coordination towards greater similaritywith interlimb coordination. Our finding that individuals and dyadsperformed our task equally well and improved their performance atapproximately the same rate is in line with this proposal.
A complete understanding of how people perform joint actionswill require explication of how and when different modalities areused to support performance. The present experiment indicates thathaptic information is very powerful for guiding joint coordinationtasks such as lifting and moving a couch together. In general, itwould be useful to know when dyads profit from having sharedaccess to different information sources, and how such sourcesinteract to guide performance. Thus, parametric manipulationsconcerning the access to different modalities available for planningand real-time coordination across dyads could reveal how differentmodalities are used and how their information is integrated forjoint actions. For instance, using the present experimental para-digm, one could manipulate the presence and quality of informa-tion available through different modalities to determine how in-formation exchange through each modality influencesperformance, and how different modalities interact in this regard.This could help to answer the question of whether optimal inter-personal coupling is achieved in particular modalities, whetherredundancies between information in different modalities boostscoupling, and how the usefulness of information from differentmodalities interacts with task demands. In terms of the HKBmodel, taking this route could result in a quantification of howinformational coupling through different modalities influences thecoupling function in the model.
Another issue that requires further investigation is whether theincreased force overlap in dyadic interaction provides a functionalmechanism to enhance haptic information, or whether it emergesas a by-product of lacking the internal information that is availableto individuals in bimanual coordination. It is tempting to speculatethat force amplification serves a communicative purpose (Reed etal., 2006), whereby instrumental actions are modulated in order toserve as coordination signals.
From an implementation perspective, understanding how peoplecoordinate physically coupled actions is of central importance forthe development of interactive robots (e.g., Feth, Groten, Peer,Hirsche, & Buss, 2009). The development of robots that smoothlyinteract with people requires an understanding of the use of dif-ferent information channels for interpersonal action planning andcoordination. Our findings provide a modest but important steptowards understanding how people may exploit the haptic channelto achieve such smooth interpersonal coordination.
References
Amazeen, P. G., Schmidt, R. C., & Turvey, M. T. (1995). Frequencydetuning of the phase entrainment dynamics of visually coupled rhyth-mic movements. Biological Cybernetics, 72, 511–518.
Bingham, G. P., Hughes, K., & Mon-Williams, M. (2008). The coordina-tion patterns observed when two hands reach-to-grasp separate objects.Experimental Brain Research, 184(3), 283–293.
Bosga, J., & Meulenbroek, R. G. J. (2007). Joint-action coordination ofredundant force contributions in a virtual lifting task. Motor Control, 11,235–258.
Braun, D. A., Ortega, P. A., & Wolpert, D. M. (2009). Nash equilibria in
multi-agent motor interactions. PLoS Computational Biology, 5,e1000468.
Brennan, S. E., Chen, X., Dickinson, C., Neider, M., & Zelinsky, G.(2008). Coordinating cognition: The costs and benefits of shared gazeduring collaborative search. Cognition, 106, 1465–1477.
Brennan, S. E., & Hanna, J. E. (2009). Partner-specific adaptation indialogue. Topics in Cognitive Science, 1, 274–291.
Camerer, C. (2003). Behavioral game theory: Experiments in strategicinteraction. Princeton, NJ: University Press.
Clark, H. H. (1996). Using language. Cambridge, England: CambridgeUniversity Press.
Feth, D., Groten, R., Peer, A., Hirsche, S., & Buss, M. (2009, March).Performance related energy exchange in haptic human-human interac-tion in a shared virtual object manipulation task. Third Joint EurohapticsConference and Symposium on Haptic Interfaces for Virtual Environ-ment and Teleoperator Systems, Salt Lake City, UT.
Franz, E. A., Zelaznik, H. N., & McCabe, G. (1991). Spatial topologicalconstraints in a bimanual task. Acta Psychologica, 77, 137–151.
Fudenberg, D., & Tirole, J. (1991). Game theory. Cambridge, MA: MITPress.
Galantucci, B., & Sebanz, N. (2009). Joint action: Current perspectives.Topics in Cognitive Science, 1, 255–259.
Haken, H., Kelso, J. A. S., & Bunz, H. (1985). A theoretical model of phasetransitions in human hand movements. Biological Cybernetics, 51, 347–356.
Harrison, S. J., & Richardson, M. (2009). Horsing around: Spontaneousfor-legged coordination. Journal of Motor Behavior, 41, 519–524.
Hick, W. E. (1952). On the rate of gain of information. Quarterly Journalof Experimental Psychology, 4, 11–26.
Hyman, R. (1953). Stimulus information as a determinant of reaction time.Journal of Experimental Psychology, 45, 188–196.
Kelso, J. A. S. (1984). Phase transitions and critical behavior in humanbimanual coordination. American Journal of Physiology: Regulatory,Integrative and Comparative, 246, R1000–R1004.
Knoblich, G., & Jordan, S. (2003). Action coordination in groups andindividuals: Learning anticipatory control. Journal of Experimental Psy-chology: Learning, Memory, & Cognition, 29, 1006–1016.
Leonhard, J. (1959). Tactile choice reactions: I. Quarterly Journal ofExperimental Psychology, 11, 76–83.
Mechsner, F., Kerzel, D., Knoblich, G., & Prinz, W. (2001). Perceptualbasis of bimanual coordination. Nature, 414, 69–73.
Newman-Norlund, R. D., Bosga, J., Meulenbroek, R. G. J., & Bekkering,H. (2008). Anatomical substrates of cooperative joint-action in a con-tinuous motor task: Virtual lifting and balancing. NeuroImage, 41,169–177.
Reed, K., Peshkin, M., Hartmann, M. J., Grabowecky, M., Patton, J., &Vishton, P. M. (2006). Haptically linked dyads: Are two motor-controlsystems better than one? Psychological Science, 17, 365–366.
Richardson, M. J., Lopresti-Goodman, S., Mancini, M., Kay, B., &Schmidt, R. C. (2008). Comparing the attractor strength of intra- andinterpersonal interlimb coordination using cross-recurrence analysis.Neuroscience Letters, 438, 340–345.
Richardson, M. J., Marsh, K. L., Isenhower, R., Goodman, J., & Schmidt,R. C. (2007). Rocking together: Dynamics of intentional and uninten-tional interpersonal coordination. Human Movement Science, 26, 867–891.
Richardson, M. J., Marsh, K. L., & Schmidt, R. C. (2005). Effects of visualand verbal interaction on unintentional interpersonal coordination. Jour-nal of Experimental Psychology: Human Perception and Performance,31, 62–79.
Rosenbaum, D. A., Dawson, A. M., & Challis, J. H. (2006). Haptictracking permits bimanual independence. Journal of Experimental Psy-chology: Human Perception and Performance, 32, 1266–1275.
1430 VAN DER WEL, KNOBLICH, AND SEBANZ

Sanfey, A. G. (2007). Social decision-making: Insights from game theoryand neuroscience. Science, 318, 598–602.
Schmidt, R. C., Bienvenu, M., Fitzpatrick, P. A., & Amazeen, P. G. (1998).A comparison of intra- and interpersonal interlimb coordination: Coor-dination breakdowns and coupling strength. Journal of ExperimentalPsychology: Human Perception and Performance, 24, 884–900.
Schmidt, R. C., Carello, C., & Turvey, M. T. (1990). Phase transitions andcritical fluctuations in the visual coordination of rhythmic movementsbetween people. Journal of Experimental Psychology: Human Percep-tion and Performance, 16, 227–247.
Schmidt, R. C., Fitzpatrick, P., Caron, R., & Mergeche, J. (in press).Understanding social motor coordination. Human Movement Science.
Schmidt, R. C., & O’Brien, B. (1997. Evaluating the dynamics of unin-tended interpersonal coordination. Ecological Psychology, 9, 189–206.
Sebanz, N., Bekkering, H., & Knoblich, H. (2006). Joint action: Bodies andminds moving together. Trends in Cognitive Sciences, 10, 70–76.
Shockley, K., Baker, A. A., Richardson, M. J., & Fowler, C. A. (2007).Articulatory constraints on interpersonal postural coordination. Journalof Experimental Psychology: Human Perception and Performance, 33,201–208.
van Ulzen, N. R., Lamoth, C. J. C., Daffertshofer, A., Semin, G. R., &Beek, P. J. (2008). Characteristics of instructed and uninstructed inter-personal coordination while walking side-by-side. Neuroscience Letters,432, 88–93.
Vesper, C., Butterfill, S., Knoblich, G., & Sebanz, N. (in press). A minimalarchitecture for joint action. Neural Networks.
Received April 8, 2010Revision received October 26, 2010
Accepted November 2, 2010 �
1431LET THE FORCE BE WITH US
Related Documents











