Lentiviral vector expressing retinoic acid receptor b2 promotes recovery of function after corticospinal tract injury in the adult rat spinal cord Ping K. Yip 1,{ , Liang-Fong Wong 3,4, * ,{ , Damian Pattinson 1 , Anna Battaglia 1 , John Grist 1 , Elizabeth J. Bradbury 1 , Malcolm Maden 2 , Stephen B. McMahon 1 and Nicholas D. Mazarakis 3,{ 1 Neurorestoration Group, Wolfson CARD, 2 MRC Centre for Developmental Biology, King’s College London, Guy’s Campus, London Bridge, London SE1 1UL, UK, 3 Oxford BioMedica (UK) Ltd, Medawar Centre, Robert Robinson Avenue, The Oxford Science Park, Oxford OX4 4GA, UK and 4 Henry Wellcome LINE, Dorothy Hodgkin Building, Whitson Street, Bristol BS1 3NY, UK Received June 9, 2006; Revised and Accepted September 7, 2006 Spinal cord injury often results in permanent and devastating neurological deficits and disability. This is due to the limited regenerative capacity of neurones in the central nervous system (CNS). We recently demon- strated that a transcription factor retinoic acid receptor b2 (RARb2) promoted axonal regeneration in adult sensory neurones located peripherally. However, it is not known if RARb2 can promote axonal regeneration in cortical neurones of the CNS. Here, we demonstrate that delivery of RARb2 via a lentiviral vector to adult dissociated cortical neurones significantly enhances neurite outgrowth on adult cortical cryosections, which normally provide an unfavourable substrate for growth. We also show that lentiviral-mediated transduction of corticospinal neurones resulted in robust transgene expression in layer V corticospinal neurones and their axonal projections in the corticospinal tract (CST) of the spinal cord. Expression of RARb2 in these neurones enhanced regeneration of the descending CST fibres after injury to these axons in the mid-cervical spinal cord. Furthermore, we observed functional recovery in sensory and locomotor behavioural tests in RARb2-treated animals. These results suggest that a direct and selective delivery of RARb2 to the cortico- spinal neurones promotes long-distance functional regeneration of axons in the spinal cord and may thus offer new therapeutic gene strategy for the treatment of human spinal cord injuries. INTRODUCTION Currently, nearly 1000 new cases of spinal cord injury (SCI) occur annually in the UK each year (1). Patients with SCI are often left with permanent and devastating functional dis- abilities due to neurological deficits. Unfortunately, there are no effective therapies that promote regeneration of damaged axons of central nervous system (CNS) neurones and therefore this is still an area of unmet clinical need. The retinoid signalling pathway is important in the survival and differentiation of the vertebrate CNS, and retinoic acid (RA) has been shown to stimulate both neurite number and length in embryonic cultures (2 – 4). The functions of retinoids, however, may extend beyond their well-documented roles in neurodevelopment and play important roles in adult neural function. For example, age-related downregulation of retinoid- mediated transcription events has been postulated to play a role in the cognitive decline observed in aged mice (5–8). A key observation in these studies is the downregulation of mRNA levels for the nuclear receptor RA receptor b (RARb) in the brains of aged rodents. Retinoid ligand # The Author 2006. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] { The authors wish it to be known that, in their opinion, the first two authors should be regarded as joint First Authors. *To whom correspondence should be addressed. Tel: þ44 1173313106; Email: [email protected] ‡ Present Address Department of Gene Therapy, Section of Infectious Diseases, Division of Medicine, Wright – Fleming Institute, Imperial College London, St Mary’s Campus, Norfolk Place, London W2 1PG, UK. Human Molecular Genetics, 2006, Vol. 15, No. 21 3107–3118 doi:10.1093/hmg/ddl251 Advance Access published on September 19, 2006 by guest on October 1, 2014 http://hmg.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Lentiviral vector expressing retinoic acidreceptor b2 promotes recovery of functionafter corticospinal tract injury in the adult ratspinal cord
Ping K. Yip1,{, Liang-Fong Wong3,4,*,{, Damian Pattinson1, Anna Battaglia1, John Grist1,
Elizabeth J. Bradbury1, Malcolm Maden2, Stephen B. McMahon1 and Nicholas D. Mazarakis3,{
1Neurorestoration Group, Wolfson CARD, 2MRC Centre for Developmental Biology, King’s College London, Guy’s
Campus, London Bridge, London SE1 1UL, UK, 3Oxford BioMedica (UK) Ltd, Medawar Centre, Robert Robinson
Avenue, The Oxford Science Park, Oxford OX4 4GA, UK and 4Henry Wellcome LINE, Dorothy Hodgkin Building,
Whitson Street, Bristol BS1 3NY, UK
Received June 9, 2006; Revised and Accepted September 7, 2006
Spinal cord injury often results in permanent and devastating neurological deficits and disability. This is dueto the limited regenerative capacity of neurones in the central nervous system (CNS). We recently demon-strated that a transcription factor retinoic acid receptor b2 (RARb2) promoted axonal regeneration in adultsensory neurones located peripherally. However, it is not known if RARb2 can promote axonal regenerationin cortical neurones of the CNS. Here, we demonstrate that delivery of RARb2 via a lentiviral vector to adultdissociated cortical neurones significantly enhances neurite outgrowth on adult cortical cryosections, whichnormally provide an unfavourable substrate for growth. We also show that lentiviral-mediated transduction ofcorticospinal neurones resulted in robust transgene expression in layer V corticospinal neurones and theiraxonal projections in the corticospinal tract (CST) of the spinal cord. Expression of RARb2 in these neuronesenhanced regeneration of the descending CST fibres after injury to these axons in the mid-cervical spinalcord. Furthermore, we observed functional recovery in sensory and locomotor behavioural tests inRARb2-treated animals. These results suggest that a direct and selective delivery of RARb2 to the cortico-spinal neurones promotes long-distance functional regeneration of axons in the spinal cord and may thusoffer new therapeutic gene strategy for the treatment of human spinal cord injuries.
INTRODUCTION
Currently, nearly 1000 new cases of spinal cord injury (SCI)occur annually in the UK each year (1). Patients with SCIare often left with permanent and devastating functional dis-abilities due to neurological deficits. Unfortunately, there areno effective therapies that promote regeneration of damagedaxons of central nervous system (CNS) neurones and thereforethis is still an area of unmet clinical need.
The retinoid signalling pathway is important in the survivaland differentiation of the vertebrate CNS, and retinoic acid
(RA) has been shown to stimulate both neurite number andlength in embryonic cultures (2–4). The functions of retinoids,however, may extend beyond their well-documented roles inneurodevelopment and play important roles in adult neuralfunction. For example, age-related downregulation of retinoid-mediated transcription events has been postulated to play arole in the cognitive decline observed in aged mice (5–8).A key observation in these studies is the downregulation ofmRNA levels for the nuclear receptor RA receptorb (RARb) in the brains of aged rodents. Retinoid ligand
# The Author 2006. Published by Oxford University Press. All rights reserved.For Permissions, please email: [email protected]
{
The authors wish it to be known that, in their opinion, the first two authors should be regarded as joint First Authors.*To whom correspondence should be addressed. Tel: þ44 1173313106; Email: [email protected]
‡Present Address Department of Gene Therapy, Section of Infectious Diseases, Division of Medicine, Wright–Fleming Institute, Imperial CollegeLondon, St Mary’s Campus, Norfolk Place, London W2 1PG, UK.
Human Molecular Genetics, 2006, Vol. 15, No. 21 3107–3118doi:10.1093/hmg/ddl251Advance Access published on September 19, 2006
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

activation of RARb (or other receptor subtypes) induces genetranscription by interacting with distinct promoter sequencesin target genes (9). The highly conserved sequence homologyof RARb2 between species and not within other subtypessuggests it has a unique and important function regulatingcell growth and differentiation.
We have previously shown that delivery of RARb2 to adultrat sensory neurones in the dorsal root ganglion (DRG) locatedperipherally can stimulate functional axonal regeneration intothe spinal cord (10), suggesting that RARb2 induced growthprogrammes within the injured neurones. In adult spinalcord explants, lentiviral vector-mediated expression ofRARb2 also induced neurite outgrowth (11). However, theeffects of RARb2 on long-distance axonal regeneration inthe adult CNS (known to be severely inhibitory to regrowingaxons) have not yet been examined in vivo.
Here, we test whether RARb2 can prime growth pro-grammes within adult CNS neurones in order for the injuredaxons to overcome the inhibitory CNS environment andthereby promote repair. Using equine infectious anaemiavirus (EIAV)-based lentiviral vectors to deliver RARb2 toadult cortical neurones in vitro, we show that RARb2 caninduce neurite outgrowth from these neurones on differentsubstrata, including substrates that are inhibitory to regener-ation. In the sensorimotor cortex, efficient and long-termexpression in corticospinal neurones was observed followinglentiviral vector delivery. Treatment with EIAV vectorsexpressing RARb2 in the cortex promoted axonal regenerationin the corticospinal tract (CST) carrying descending axonsfrom the sensorimotor cortex to the spinal cord, and improvedfunctional recovery after SCI. These data indicate that RARb2is capable of supporting long-distance axonal regeneration inthe inhibitory adult CNS and may therefore have therapeuticapplications for the treatment of human spinal cord injuries.
RESULTS
EIAV-RARb2 induces neurite outgrowth in adultrat cortical neurones
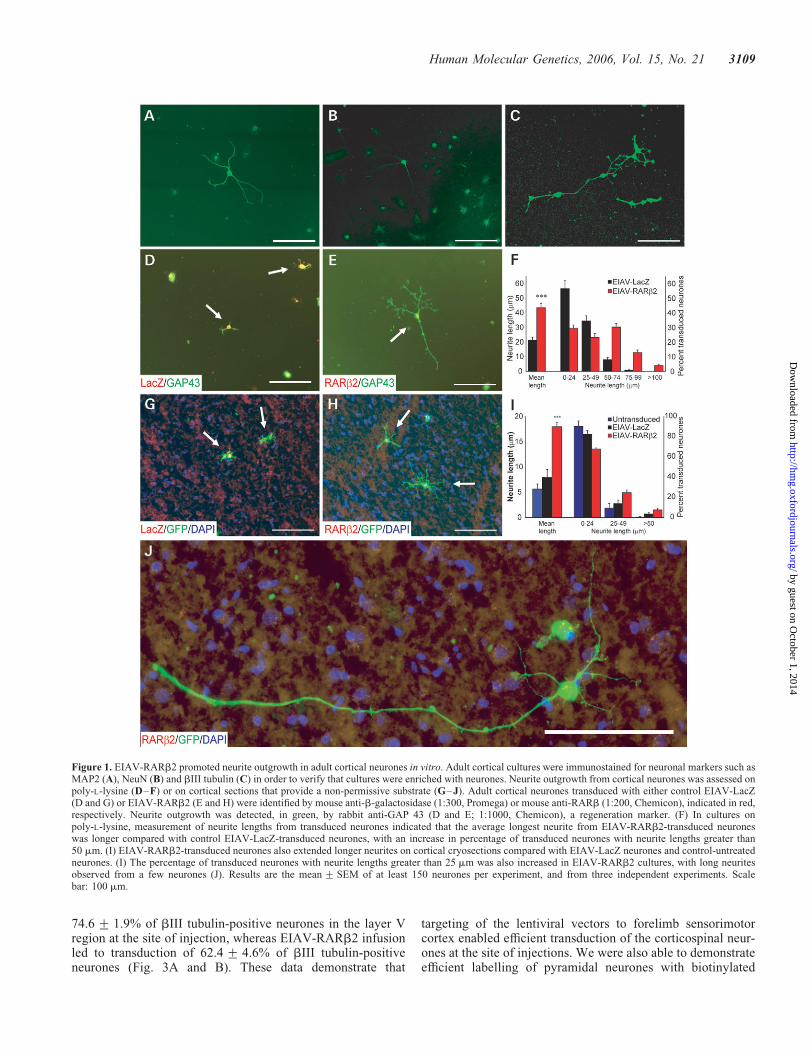
Dissociated neurones were prepared from adult rat cortex andcultured on a limiting substrate (0.01% poly-L-lysine) that wasunfavourable for neurite outgrowth. Plated cells were ident-ified as neurones by immunohistochemical staining withknown neuronal markers such as MAP2 (Fig. 1A), NeuN(Fig. 1B) and bIII tubulin (Fig. 1C). Efficient transductionof cortical neurones was observed with EIAV vectors expres-sing either a reporter gene LacZ (EIAV-LacZ; Fig. 1D) orRARb2 (EIAV-RARb2; Fig. 1E) at a multiplicity of infection(MOI) of 100; 71.0+ 3.4% and 74.4+ 2.1% of the corticalneurons were transduced in EIAV-LacZ and EIAV-RARb2cultures, respectively. EIAV-RARb2 increased neurite out-growth in cortical neurones, compared with controlEIAV-LacZ-transduced neurones (Fig. 1D and E; P , 0.001,Student’s t-test, from three independent experiments, at least150 transduced neurones immunostained with the regenerationmarker GAP 43 were counted per experiment). Neurite out-growth from transduced cortical neurones was assessed bymeasuring the length of the longest neurite from the neurones.In EIAV-RARb2-transduced neurones, the length of the
longest neurite was 43.6+ 3.0 mm (mean+ SEM) comparedwith 21.3+ 2.2 mm in EIAV-LacZ neurones (Fig. 1F).Further analysis of neurite lengths from cortical neuronesrevealed that there was an increase in the percentage of trans-duced neurones with neurite lengths of .50 mm in EIAV-RARb2 cultures, compared with EIAV-LacZ cultures, where91% of transduced neurones retained neurites of ,50 mm.Furthermore, 17% of EIAV-RARb2-transduced neurones pro-duced neurites that were .75 mm, whereas in EIAV-LacZcultures, only 0.8% of neurones produced neurites .75 mm.These data demonstrate that RARb2 has the capacity topromote neurite outgrowth in adult cortical neurones in aserum-free medium without any neurotrophic support.
Cryosections of the cerebrum provide a poor substrate forthe outgrowth of adult rodent cortical neurones (12). Wetested whether expression of EIAV-RARb2 could induce out-growth by plating adult rat green fluorescent protein (GFPþ)rat cortical neurones that were transduced with EIAV-RARb2on adult coronal cerebral cryosections. The GFPþ neuronesenabled us to differentiate between the transduced dissociatedcortical neurones and neurones within the cerebrum cryosec-tions. EIAV-RARb2-transduced neurones were able toextend longer neurites on the grey matter compared withcontrol vector treated or untreated in the presence of10211
M trans-RA (Fig. 1G–I; P , 0.001, Student’s t-test,from three independent experiments, at least 150 transducedneurones were counted per experiment). The mean length ofthe longest neurite was 18.0+ 0.9 mm (mean+ SEM)in EIAV-RARb2 neurones compared with 7.9+ 1.6 mm innon-transduced neurones and 5.6+ 1.0 mm in EIAV-LacZneurones (Fig. 1I). As with cortical neurones grown onpoly-L-lysine substrates, we observed an upward shift in thepercentage of transduced neurones with longer neurites(.25 mm) in EIAV-RARb2 cultures compared with untrans-duced and control EIAV-LacZ cultures. There was an increasein the number of neurones with neurite lengths of greater than50 mm in EIAV-RARb2 cultures; in a few cases, neuroneswere observed to extend neurites of up to 300 mm (Fig. 1J)In contrast, this was not observed in EIAV-LacZ cultures.
Transduction of pyramidal neuronesin the sensorimotor cortex
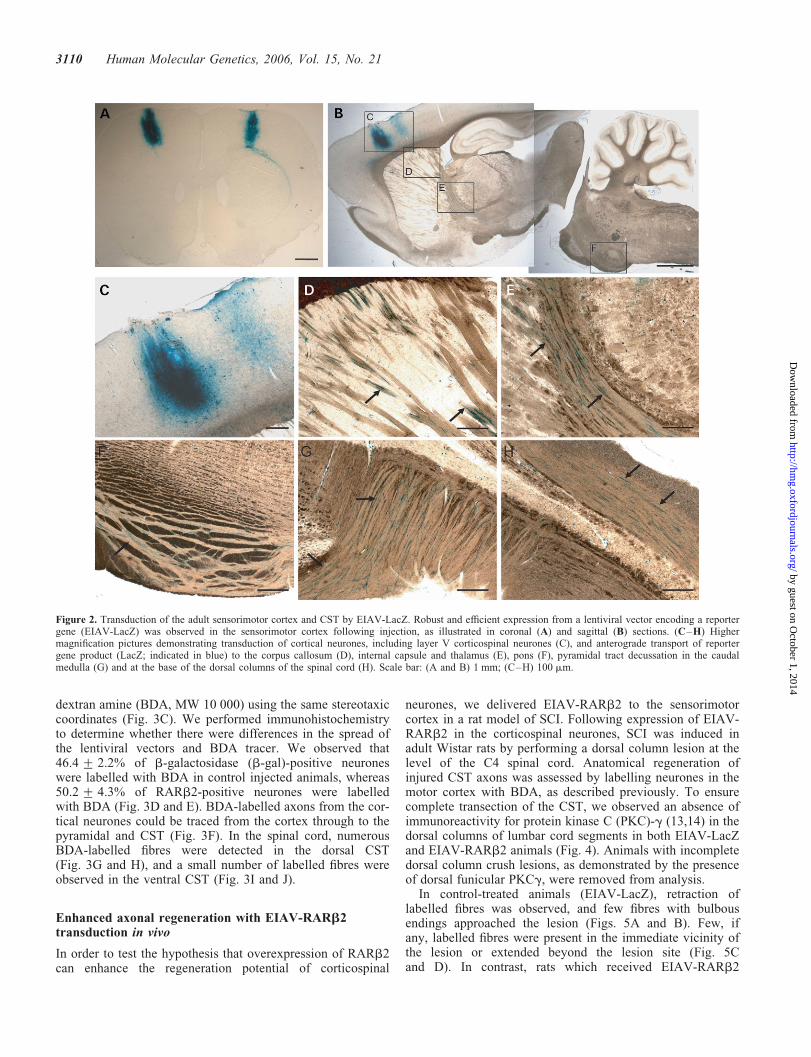
To transduce corticospinal neurones in vivo, EIAV vectorspseudotyped with the VSV-G glycoprotein were injectedinto the sensorimotor cortex. Using an EIAV vector expres-sing the reporter gene LacZ, we observed strong vectorexpression in the layer V pyramidal neurones at the site ofinjection (Fig. 2A–C). Positive X-gal staining was alsodetected in axons as they course through the corpus callosum(Fig. 2D), internal capsule (Fig. 2E), the cerebral peduncle ofthe midbrain and the ventral pons (Fig. 2F). Positively stainedfibres were also observed as the corticospinal axons crossedover at the pyramidal decussation (Fig. 2G) and along theCST in the spinal cord (Fig. 2H). These data provide evidencethat following transduction of corticospinal neurones,b-galactosidase from the EIAV vectors was anterogradelytransported along the fibres in the CST to the spinal cord.
Colocalization studies revealed that injection of EIAV-LacZ into the sensorimotor cortex resulted in transduction of
3108 Human Molecular Genetics, 2006, Vol. 15, No. 21
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
74.6+ 1.9% of bIII tubulin-positive neurones in the layer Vregion at the site of injection, whereas EIAV-RARb2 infusionled to transduction of 62.4+ 4.6% of bIII tubulin-positiveneurones (Fig. 3A and B). These data demonstrate that
targeting of the lentiviral vectors to forelimb sensorimotorcortex enabled efficient transduction of the corticospinal neur-ones at the site of injections. We were also able to demonstrateefficient labelling of pyramidal neurones with biotinylated
Figure 1. EIAV-RARb2 promoted neurite outgrowth in adult cortical neurones in vitro. Adult cortical cultures were immunostained for neuronal markers such asMAP2 (A), NeuN (B) and bIII tubulin (C) in order to verify that cultures were enriched with neurones. Neurite outgrowth from cortical neurones was assessed onpoly-L-lysine (D–F) or on cortical sections that provide a non-permissive substrate (G–J). Adult cortical neurones transduced with either control EIAV-LacZ(D and G) or EIAV-RARb2 (E and H) were identified by mouse anti-b-galactosidase (1:300, Promega) or mouse anti-RARb (1:200, Chemicon), indicated in red,respectively. Neurite outgrowth was detected, in green, by rabbit anti-GAP 43 (D and E; 1:1000, Chemicon), a regeneration marker. (F) In cultures onpoly-L-lysine, measurement of neurite lengths from transduced neurones indicated that the average longest neurite from EIAV-RARb2-transduced neuroneswas longer compared with control EIAV-LacZ-transduced neurones, with an increase in percentage of transduced neurones with neurite lengths greater than50 mm. (I) EIAV-RARb2-transduced neurones also extended longer neurites on cortical cryosections compared with EIAV-LacZ neurones and control-untreatedneurones. (I) The percentage of transduced neurones with neurite lengths greater than 25 mm was also increased in EIAV-RARb2 cultures, with long neuritesobserved from a few neurones (J). Results are the mean+ SEM of at least 150 neurones per experiment, and from three independent experiments. Scalebar: 100 mm.
Human Molecular Genetics, 2006, Vol. 15, No. 21 3109
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
dextran amine (BDA, MW 10 000) using the same stereotaxiccoordinates (Fig. 3C). We performed immunohistochemistryto determine whether there were differences in the spread ofthe lentiviral vectors and BDA tracer. We observed that46.4+ 2.2% of b-galactosidase (b-gal)-positive neuroneswere labelled with BDA in control injected animals, whereas50.2+ 4.3% of RARb2-positive neurones were labelledwith BDA (Fig. 3D and E). BDA-labelled axons from the cor-tical neurones could be traced from the cortex through to thepyramidal and CST (Fig. 3F). In the spinal cord, numerousBDA-labelled fibres were detected in the dorsal CST(Fig. 3G and H), and a small number of labelled fibres wereobserved in the ventral CST (Fig. 3I and J).
Enhanced axonal regeneration with EIAV-RARb2transduction in vivo
In order to test the hypothesis that overexpression of RARb2can enhance the regeneration potential of corticospinal
neurones, we delivered EIAV-RARb2 to the sensorimotorcortex in a rat model of SCI. Following expression of EIAV-RARb2 in the corticospinal neurones, SCI was induced inadult Wistar rats by performing a dorsal column lesion at thelevel of the C4 spinal cord. Anatomical regeneration ofinjured CST axons was assessed by labelling neurones in themotor cortex with BDA, as described previously. To ensurecomplete transection of the CST, we observed an absence ofimmunoreactivity for protein kinase C (PKC)-g (13,14) in thedorsal columns of lumbar cord segments in both EIAV-LacZand EIAV-RARb2 animals (Fig. 4). Animals with incompletedorsal column crush lesions, as demonstrated by the presenceof dorsal funicular PKCg, were removed from analysis.
In control-treated animals (EIAV-LacZ), retraction oflabelled fibres was observed, and few fibres with bulbousendings approached the lesion (Figs. 5A and B). Few, ifany, labelled fibres were present in the immediate vicinity ofthe lesion or extended beyond the lesion site (Fig. 5Cand D). In contrast, rats which received EIAV-RARb2
Figure 2. Transduction of the adult sensorimotor cortex and CST by EIAV-LacZ. Robust and efficient expression from a lentiviral vector encoding a reportergene (EIAV-LacZ) was observed in the sensorimotor cortex following injection, as illustrated in coronal (A) and sagittal (B) sections. (C–H) Highermagnification pictures demonstrating transduction of cortical neurones, including layer V corticospinal neurones (C), and anterograde transport of reportergene product (LacZ; indicated in blue) to the corpus callosum (D), internal capsule and thalamus (E), pons (F), pyramidal tract decussation in the caudalmedulla (G) and at the base of the dorsal columns of the spinal cord (H). Scale bar: (A and B) 1 mm; (C–H) 100 mm.
3110 Human Molecular Genetics, 2006, Vol. 15, No. 21
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

treatment showed thick-labelled fibre bundles that approachedthe proximal edge of the lesion (Fig. 5E–G). At higher mag-nification, we could detect BDA-labelled axons around thelesion site (Fig. 5L). Furthermore, labelled fibres weredetected beyond the lesion (Fig. 5H–K), and some axonswere observed to extend arborizing collaterals from thewhite matter into the grey matter (Fig. 5K, indicated bywhite arrowhead). We also detected some axons with growthcone-like endings in the spinal cord (Figs. 5H and M, indi-cated by white arrowheads). Quantification of the number ofBDA-labelled fibres present in the CST immediately rostralto the lesion showed that there were significantly more
labelled fibres in EIAV-RARb2 compared with controlEIAV-LacZ animals (Fig. 6; P , 0.05, two-way RMANOVA). In the spinal cord caudal to the lesion, thenumber of BDA-labelled fibres in the CST was also signifi-cantly higher in EIAV-RARb2 animals compared with EIAV-LacZ animals (Fig. 6; P , 0.05, two-way RM ANOVA).
Functional recovery associated with histologicalregeneration
The presence of arborizing collaterals in the grey mattersuggested that the regenerated CST axons may establish
Figure 3. Lentiviral expression in the sensorimotor cortex and BDA labelling of corticospinal axons. (A, B) bGal or RARb expression (green) was colocalizedwith bIII tubulin (red) in layer V corticospinal neurones in the cortex in animals after EIAV transduction (inset represents magnified area of boxed region in themain picture). Colocalization of bgal or RARb with bIII tubulin is highlighted with black arrows, whereas bgal- or RARb-positive and bIII tubulin-positiveneurones are indicated by white and black arrowheads, respectively. (C) Efficient anterograde labelling of CST was also achieved by administering BDA(MW 10 000) to the cortex, using identical stereotaxic coordinates as those used for lentiviral transduction. In (A–C), roman numerals represent corticallayers within the cortex. (D, E) bGal- or RARb-overexpressing layer V neurones (red) were also colocalized with BDA-labelled cells (in green) in thecortex. Colocalization of bgal or RARb with BDA is highlighted with black arrows, whereas bgal- or RARb-positive and BDA-positive neurones are indicatedby white and black arrowheads, respectively. Corticospinal fibres were also extensively labelled at the spinal level, and individual fibres with arbours can bevisualized, allowing for quantification of axonal regeneration in the CST (F). (G and H) The majority of BDA-labelled fibres (indicated by white arrows)were present in the dorsal CST (dCST), whereas in (I and J), a few fibres (white arrows) could be detected in the ventral CST (vCST). (H and J) representmagnified areas of the boxed regions in (G and I), respectively. Scale bar: 100 mm.
Human Molecular Genetics, 2006, Vol. 15, No. 21 3111
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

functional connections with post-synaptic neurones. Dorsalcolumn projections are important for discriminative touchand proprioception, and therefore we undertook appropriatebehavioural tests to examine functional recovery. Rats wereassessed for sensorimotor function by testing for their abilityto sense and remove an adhesive tape attached onto theplantar surface of their forepaw. Unlesioned sham controlsperformed these tasks quickly (5.6+ 1.2 s and 24.0+ 5.7 sto sense and remove tape, respectively), whereas EIAV-LacZ-treated animals were significantly impaired throughout testing(Fig. 7A and B; 50.1+ 8.0 s and 75.3+ 8.8 s, respectively, at5 weeks’ post-lesion; P , 0.001, two-way RM ANOVA). Incontrast, EIAV-RARb2-treated animals showed recovery offunction; at 6 weeks’ post-lesion, the time taken to senseand remove tape in these animals was 28.0+ 6.8 s and52.7+ 9.0 s respectively). However, the recovery in EIAV-RARb2 was not complete, as latencies for tape sensing andremoving were significantly different from sham controls(P , 0.05 and P , 0.01, respectively, two-way RMANOVA). EIAV-RARb2 treatment also consistentlyimproved recovery in motor function in a ladder- and grid-crossing task, in which rats were assessed for the number offoot slips as they crossed a horizontal ladder or grid(Fig. 7C and D). In both tasks, unlesioned sham controlsmade few foot slips but EIAV-LacZ-treated rats were severelyand significantly impaired throughout testing. At 5 weeks’post-lesion, the number of foot slips made by EIAV-LacZrats in ladder- and grid-crossing was 9.2+ 1.3 and7.8+ 0.8, respectively, compared with 1.1+ 1.0 and 0+ 0,respectively, in sham controls (P , 0.001, two-way RMANOVA). The EIAV-RARb2 group showed improvement in
task performance from 3 weeks’ post-lesion; and at 5 weeks’post-lesion the number of foot slips made was 5.4+ 1.3 and2.0+ 0.8 in ladder- and grid-crossing tasks, respectively.The improvement demonstrated by EIAV-RARb2 rats waspartial, as the number of foot slips was significantly differentfrom sham controls (P , 0.01, two-way RM ANOVA).Walking patterns were also assessed by analysing footprintspacing. At 5 weeks’ post-lesion, EIAV-LacZ-treated ratstook shorter and wider strides (65.6+ 8.4 mm and37.3+ 1.3 mm, respectively) compared with sham unlesionedcontrols (Fig. 7E and F; 156+ 9.5 mm and 18.1+ 2.8 mm,respectively; P , 0.01 and P , 0.05, respectively, Student’stwo-tailed unpaired t-test). In contrast, the EIAV-RARb2group did not show a significant change in stride length andwidth (125.8+ 7.7 mm and 26.3+ 4.0 mm, respectively)compared with sham controls, suggesting that these ratsmade a significant recovery to their walking patterns.
DISCUSSION
We have investigated the effects of EIAV-RARb2 on axonalregeneration of adult cortical neurones in vitro and in anin vivo rat model of SCI. Overexpression of RARb2 in corticalneurones enabled significant neurite outgrowth in a non-permissive environment and on cortical cryosectionscompared with that of control neurones. In vivo, efficienttransduction of the sensorimotor cortex was demonstratedand resulted in good labelling of the CST. Using this deliverymethod, we showed that overexpression of RARb2 in adult ratcorticospinal neurones increased CST axonal outgrowth up toand beyond a CNS lesion, and improved functional recoveryin sensory and locomotor behavioural tasks.
Previous studies in adult spinal cord regeneration have usedembryonic cells, transformed cells or perinatal cells to evalu-ate neurite outgrowth due to the ease of culture of these cellu-lar types. However, in these studies, the use of neuronesderived from the adult brain is more relevant. Dissociatedadult cortical neurones have been previously employedin other studies in mice (12,15) and rats (16,17), and Tomet al. (12) have demonstrated that adult cortical neuroneswere normally unable to produce long neurites on CNS cryo-sections. In our experiments, RARb2-transduced adult corticalneurones when seeded onto adult cortical cryosections did notdemonstrate significant neurite outgrowth compared with con-trols (data not shown). This was surprising because RARb2-transduced neurones were able to extend neurites in theabsence of exogenous ligand on a different substrate such aspoly-L-lysine (Fig. 1F). The presence of retinol and RA-bindingproteins, such as cellular retinol-binding protein-1 (CRBP-1)and cellular RA-binding protein-1 (CRABP-1), as well as theRA-synthesizing enzyme retinaldehyde dehydrogenase-3(RALDH-3), has been demonstrated in the adult rodentcortex, suggesting that there are internal retinoid reservoirswithin the adult cortical neurones (18–20). In the case of cor-tical neurones grown on the cryocultures, the endogenouslevels of RA may not be sufficient in cortical neurones to over-come the inhibitory environment of the grey matter of the CNS,hence neurite growth was attenuated without additional ligandsupply. Therefore, when we provided exogenous ligand in the
Figure 4. Detection of PKCg in the lumbar spinal cord. Immunostaining forPKCg was carried out in the lumbar spinal cord to ensure that the CSTswere completely lesioned and did not result in spared axon(s) along theCST. (A) In sham (unlesioned) animals, intense labelling of PKCg in thelamina II and at the base of the dorsal columns was observed, whereasanimals with complete lesions demonstrated lack of PKCg labelling at thebase of the dorsal columns (B). Detection of PKCg in lamina II of thespinal cord was unchanged in these animals. (C–F) Higher magnificationmicrographs of dorsal columns of animals with (C) complete, (D and E)incomplete and (F) sham lesions. In animals with incomplete lesions, somePKCg staining at the base of the dorsal columns was observed (indicated bywhite arrows). These animals were subsequently excluded from the study.Scale bar: (A and B) 500 mm; (C–F) 100 mm.
3112 Human Molecular Genetics, 2006, Vol. 15, No. 21
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
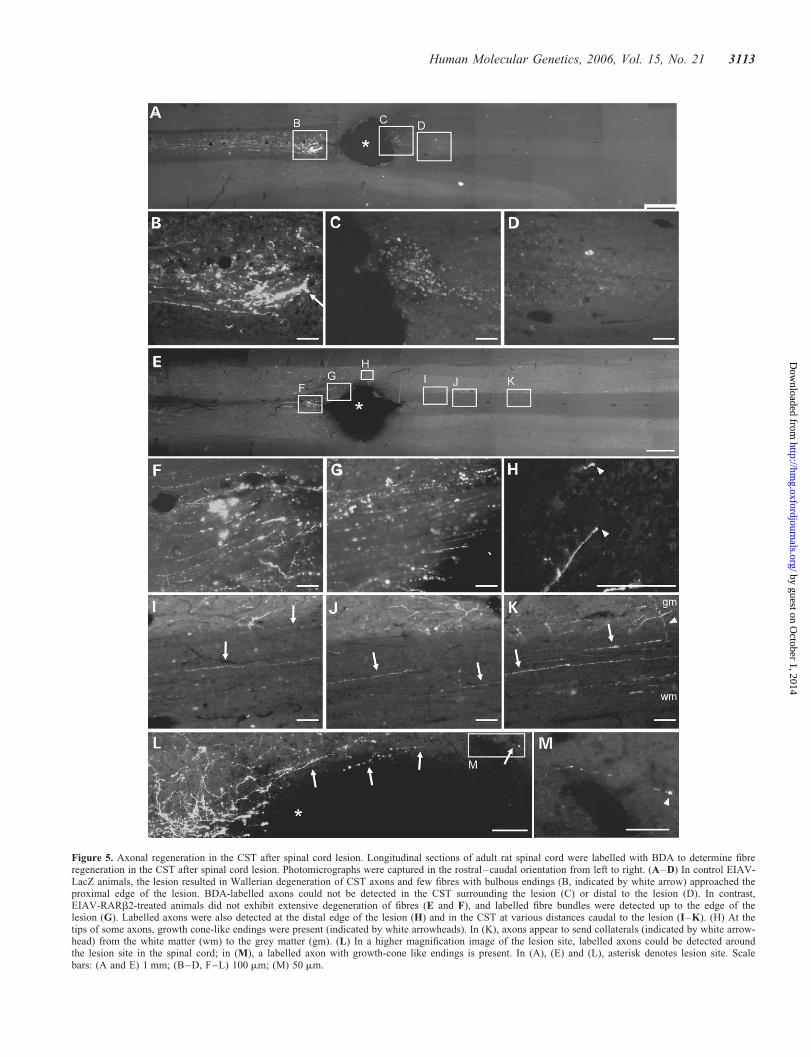
Figure 5. Axonal regeneration in the CST after spinal cord lesion. Longitudinal sections of adult rat spinal cord were labelled with BDA to determine fibreregeneration in the CST after spinal cord lesion. Photomicrographs were captured in the rostral–caudal orientation from left to right. (A–D) In control EIAV-LacZ animals, the lesion resulted in Wallerian degeneration of CST axons and few fibres with bulbous endings (B, indicated by white arrow) approached theproximal edge of the lesion. BDA-labelled axons could not be detected in the CST surrounding the lesion (C) or distal to the lesion (D). In contrast,EIAV-RARb2-treated animals did not exhibit extensive degeneration of fibres (E and F), and labelled fibre bundles were detected up to the edge of thelesion (G). Labelled axons were also detected at the distal edge of the lesion (H) and in the CST at various distances caudal to the lesion (I–K). (H) At thetips of some axons, growth cone-like endings were present (indicated by white arrowheads). In (K), axons appear to send collaterals (indicated by white arrow-head) from the white matter (wm) to the grey matter (gm). (L) In a higher magnification image of the lesion site, labelled axons could be detected aroundthe lesion site in the spinal cord; in (M), a labelled axon with growth-cone like endings is present. In (A), (E) and (L), asterisk denotes lesion site. Scalebars: (A and E) 1 mm; (B–D, F–L) 100 mm; (M) 50 mm.
Human Molecular Genetics, 2006, Vol. 15, No. 21 3113
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

form of trans-RA at a concentration of 10211M, there was a
significant increase in neurite outgrowth from RARb2-transduced adult cortical neurones on cryosections, but notfrom control cultures. The concentration (10211
M) oftrans-RA used in the cryoculture experiments was 100-foldless than previously observed in vivo and it is likely thatin vivo sources of RA, for example, the cerebrospinal fluid orother brain regions such as the striatum, will be sufficient toactivate EIAV-RARb2 in the in vivo experiments.
Following spinal cord lesions, signals derived from myelinsuch as Nogo, myelin-associated glycoprotein (MAG) andoligodendrocyte-myelin glycoprotein (OMgp) (21–27) aswell as extracellular matrix molecules within the glial scarsuch as chondroitin sulphate proteoglycans (CSPGs) (28)prevent regeneration of damaged axons. Many studies haveshown some functional recovery after SCI by neutralizingthese inhibitory signals with treatments including Nogo-blocking peptides (22,29), infusions of chondroitinase ABC(13) or transplantation of olfactory ensheathing cells (30,31).These strategies make the injured spinal cord less hostile to
regeneration, either from lesioned fibres that attempt torepair axonal connections or from surviving intact axons thatpromote functional recovery through plasticity mechanisms.Our approach in this study was to use an EIAV-based lenti-viral vector encoding RARb2 to target the CNS corticospinalneurones in order to increase their regenerative capacitydespite the adverse environment in the spinal cord. Thenuclear transcription factor RARb2 is not normally detectablein corticospinal neurones, as demonstrated by Zetterstromet al. (19), and in our study, by the absence of positiveRARb immunostaining in areas distant from the injectionsite. This indicates that low, if any, amounts of RARb2protein is present in these cells. By targeting the sensorimotorcortex with EIAV-RARb2 and efficiently labelling CST axonsthrough the use of precise stereotaxic coordinates, we wereable to increase RARb2 levels in a large number of corticosp-inal neurones and study its effects on CST regeneration. Wedemonstrated that numerous transduced layer V cortical neur-ones were also labelled with the BDA anterograde tracer, indi-cating that vector transduction and tracer spread were notmutually exclusive. This suggested that many of the anterogra-dely labelled fibres in the spinal cord are likely to arise fromtransduced neurones. By studying this sample population ofcells, we observed that RARb2 treatment increased axonalregeneration in the CST and improved functional recoveryafter SCI.
Assessment of axonal regeneration in the spinal cord usingthe seven-point criteria proposed by Steward et al. (32) indi-cated that several of the criteria were fulfilled. We detectedlabelled axons in the immediate vicinities of the lesion in theRARb2 spinal cord, whereas few, if any, were observed incontrol LacZ cord (Fig. 5L). Astrogliosis in the immediatetissue surrounding the lesion site often generates an environmentinhibitory to regeneration. Therefore, Criteria I and III were ful-filled, where the axon was shown to extend from the site ofinjury into the tissue environment of the scar that develops atthe injury site. Furthermore, we show unusual tracking and mor-phology of regenerated axons, for example, traversing of axonsfrom the white matter into the grey matter and axon branching inthe grey matter (Fig. 5I–K; Criteria IV and VII). We have alsodetermined that the observed axonal regeneration in the RARb2cord did not exceed plausible regeneration rates, as labelledfibres were observed at ,10 mm from the lesion site (on thebasis of a maximal regeneration rate of 1 mm per day inthe CNS, the maximal regeneration distance was 35 mm in theexperimental period) (criterion V). Finally, we detected labelledaxons around the lesion site in the spinal cord, which had growthcone-like endings (Fig. 5H and M; criterion VI). These obser-vations suggest that the labelled fibres in the RARb2 cord arelikely to be regenerated, but not spared axons.
During assessment of functional regeneration using beha-vioural tasks, we observed that the EIAV-RARb2 groupmade partial recoveries in some of the sensory-locomotortasks compared with sham controls and showed a modestincrease in the number of labelled fibres in the CST afterspinal cord lesion. The effects of EIAV-RARb2 we haveseen may be limited by the partial transduction of corti-cospinal neurones we were able to produce. We showedthat 60–75% of the layer V cortical neurones were trans-duced by the lentiviral vectors at the site of injections
Figure 6. Quantification of labelled fibres in the CST after spinal cord lesion.EIAV-RARb2-treated animals showed an increase in the number of BDA-labelled fibres in the CST rostral and caudal to the lesion when comparedwith the EIAV-LacZ-treated animals. In EIAV-LacZ animals, the number oflabelled fibres in the CST decreased as the CST approached the lesion(14.1+ 1.5 per section at 1 mm), whereas in EIAV-RARb2 animals, thenumber of labelled fibres in the CST did not decrease (24.8+ 2.4 persection at 1 mm from lesion). In the CST caudal to the lesion, few BDA-labelled fibres were observed in EIAV-LacZ animals (0.3+ 0.1 per sectionat 1 mm caudal to lesion), whereas, significantly, more fibres were detectedin the EIAV-RARb2 group (2.1+ 0.7 per section at 1 mm caudal to lesion;P, 0.05, two-way RM ANOVA).
3114 Human Molecular Genetics, 2006, Vol. 15, No. 21
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

(Fig. 3A and B); however, fewer neurones were transduced inthe cortical regions distant from the injection sites (data notshown). It has been shown that three unilateral injectionsinto the cortex lead to a tracing efficiency of 1.5% of totalCST fibres (33). Given that we injected at six sites, we are
likely to have transduced only a modest fraction of all CSTneurones. Nevertheless, the partial recovery that was observedin these studies was significant.
The effects of RARb2 overexpression on axonal regener-ation in the inhibitory environment of the injured spinal cord
Figure 7. Assessment of functional regeneration using behavioural tests. Control and treated groups were tested for sensory and locomotor function in order toassess functional regeneration. (A and B) In the tape sensing and removal tests, control EIAV-LacZ animals showed severe deficits compared with the shamcontrols (P , 0.001, two-way RM ANOVA), whereas EIAV-RARb2 showed partial recovery of function (P , 0.05 and P, 0.01 for tape sensing andremoval, respectively, compared with sham controls, two-way RM ANOVA). (C and D) In the ladder and grid crossing tasks, EIAV-LacZ made an increasednumber of foot slips throughout the testing period compared with sham controls (P, 0.001, two-way RM ANOVA); in contrast, the EIAV- RARb2 groupshowed improvement in task performance from 3 weeks’ post-lesion; however, full recovery was not observed, as these animals were significantly differentfrom sham controls (P, 0.01, two-way RM ANOVA). (E and F) In footprint analyses, EIAV-LacZ animals took shorter and wider strides compared withsham controls (P, 0.01 for stride length and P, 0.05 for stride width, Student’s two-tailed unpaired t-test), whereas EIAV-RARb2 took longer and narrowerstrides that were not significantly different from sham controls.
Human Molecular Genetics, 2006, Vol. 15, No. 21 3115
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

may be a result of transcriptional changes that were inducedwithin the corticospinal neurones. We have previouslyshown that EIAV-RARb2 transduction of DRG neuronesincreases cAMP levels in a model of peripheral nerve regener-ation into the CNS (10), and cAMP has been demonstrated topromote regeneration of lesioned dorsal column fibres in rats(34,35). It is therefore possible that EIAV-RARb2 can activatecAMP-signalling systems in corticospinal neurones to increasethe intrinsic regenerative capacity of these neurones. In thisstudy, we observed that EIAV-RARb2 mediated long-distanceregeneration in a modest number of fibres. EIAV-RARb2 wasadministered to the sensorimotor cortex while the lesion wasinduced at the C4 level of the spinal cord, and therefore anyRARb2-mediated effects are likely to occur at the level ofthe cell bodies of the corticospinal neurones and not locallyat the lesion site in the spinal cord. We detected a smallnumber of BDA-labelled fibres in the ventral CST in thespinal cord (Fig. 3I and J) and it is thus possible that the func-tional recovery we observed may be due to spontaneoussprouting from spared fibres in the ventral CST (36). Sparedventral corticospinal fibres have the propensity to extendbranching collaterals into the grey matter areas, as demon-strated by Brosamle and Schwab (33). However, in ourstudies, we have only analysed BDA-labelled fibres in thewhite matter area representing the main CST (in the dorsalcolumns) and not in the grey matter. Furthermore, the popu-lation of BDA fibres in the ventral CST represents a smallfraction of the total labelled fibres (1.8%) and therefore wefeel the majority of the labelled fibres that are present in theRARb2-treated spinal cords, particularly those that arelocated in the dorsal columns, are likely to originate fromregenerated axons and not from the ventral CST.
Although gene therapy approaches are being developedin the clinic for other neurodegenerative diseases such asParkinson’s and motor neurone diseases, gene therapy forspinal cord injuries has not been widely studied and is stillin its infancy. Our study provides a proof-of-concept thatgenetic manipulation of specific neurones can enhance neur-onal function and improve functional regeneration, therebypaving the way for further experimentation with moleculesin the treatment of spinal cord injuries. This approach maybe used in combination with current strategies that aim toprovide a favourable environment in the lesioned spinal cordfor functional regeneration, such as digesting inhibitory pro-teoglycans or providing artificial growth support matrices.
MATERIALS AND METHODS
Minimal lentiviral vector construction and production
EIAV-RARb2 was based on the SMART2 series of EIAVvectors. EIAV-RARb2 comprises minimal long-terminalrepeat sequences, a 50 central polypurine tract element,minimal CMV promoter-driving expression of mouseRARb2 and a 30 woodchuck post-transcriptional regulatoryelement (WPRE) sequence. The RARb is �100% conservedamong mouse, rat and human when its protein homologywas compared between species using the BLAST 2 sequencesin the National Center for Biotechnology Information website(http://www.ncbi.nlm.nih.gov/blast/bl2seq/wblast2.cgi) (37).
Adult cortical neurone cultures
Adult male Wistar rats (220–250 g) (n ¼ 3) were sacrificedand part of their cortices were dissected out for dissociation.The adult cortical neurones were prepared as previously pub-lished (15,38). Briefly, the animal was transcardially perfusedwith heparinized saline, the cortices removed with as littlewhite matter attached as possible. The cortices were cut into0.5 mm longitudinal sections using a McIlwain tissuechopper, white matter was trimmed away before dissociationin 2 mg/ml papain at 308C for 30 min. The cortical neuroneswere mechanically dissociated with a glass Pasteur pipetteand separated from debris by centrifugation in four 1 mlsteps of Optiprep in B27/HibernateA medium (7.5, 10, 12.5and 17.5%) at 600 g for 15 min. Fractions containing neuroneswere collected, washed and resuspended in B27/NeurobasalAmedium with gentamycin, ready for plating at a density ofapproximately 2000 neurones per well on pre-treated coverslips (0.01% poly-L-lysine). The neurones were allowed tosettle onto the poly-L-lysine substrate for 18 h, after whichEIAV-RARb2 pseudotyped with vesicular stomatitis virus(VSV-G) envelope was added to the media at MOI 100 forthe cortical neurones. The neurones were left for a further 3days in vitro before immunohistochemistry.
Adult cortical neurone cryocultures
Studies involving adult cortical neuron cryocultures have beenlimited by the inability to visualize the dissociated neuronesseeded on top of the adult cortical cryosections by immuno-cytochemistry. To overcome this in this study, the dissociatedadult cortical neurones were prepared from enhanced greenflorescent protein (GFP)-expressing rats obtained from Pro-fessor M. Okabe (Sprague Dawley strain, Osaka University,Japan) (39). Adult GFP-expressing rats (250–300 g) (n ¼ 3)were sacrificed, cortices removed and neurones prepared asdescribed earlier. For preparation of cortical cryosections,wild-type Sprague Dawley were perfused as describedearlier, brains were removed, sectioned coronally andquickly frozen on dry ice. Prior to culture, 12 mm coronal sec-tions of cortex were cut on a cryostat and thaw-mounted ontopre-treated poly-L-lysine cover slips. Cryosections were left todry in culture hood for at least 30 min but no more than 3–4 hbefore addition of dissociated adult cortical neurones.
Immunohistochemistry
After 3 days in culture, neurones were fixed with 4% parafor-maldehyde for 20 min and then permeabilized with coldmethanol for 3 min. The neurons were washed with3 � 5 min with PBS before 2 h incubation with rabbitanti-GAP43 (1:1500, Chemicon), mouse anti-RARb (1:200,Chemicon), mouse anti-b-galactosidase (1:500, Sigma) andrabbit anti-GFP (1:1000, Molecular Probes). The cover slipswere washed 3 � 5 min with PBS and then incubated 45 minwith donkey anti-rabbit 488 and -mouse 546, secondary anti-bodies (1:2000, Molecular Probes). After 3 � 5 min PBSwashes, the cover slips were embedded in Flurosave reagent(Calbiochem) with 0.5 ml DAPI (10 mg/ml) to visualize cellnuclei. For quantification of neurite outgrowth, the length of
3116 Human Molecular Genetics, 2006, Vol. 15, No. 21
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

the longest neurite for the first 150 neurones encountered whenscanning in a systematic manner was determined using imageanalysis software (SigmaScan Pro 4.01) and was expressed asmean+ SEM.
Viral vector delivery to the sensorimotor cortex
Adult male Wistar rats (n ¼ 8–9 per group) were anaesthe-tized using a combination of ketamine and medetomidineand fixed in a stereotaxic frame. The skull was exposed andinjections were made at a depth of 2 mm dorsoventrallyinto the sensorimotor cortex region using the following injec-tion coordinates as determined from a microstimulationmapping study (40): (in reference to bregma; AP, anterior–posterior; L, lateral) AP 21.5 mm, L 2.5 mm; AP 20.5 mm,L 3.5 mm; AP þ0.5 mm, L 3.5 mm; AP þ1.0 mm,L 1.5 mm; AP þ1.5 mm, L 2.5 mm; AP þ0.5 mm, L3.5 mm. EIAV-RARb2 pseudotyped with VSV-G was directlyinjected at 1 ml per site via a finely pulled glass micropipette ata rate of 0.2 ml/min, using a microinfusion pump and left for afurther 1 min. EIAV vector pseudotyped with a VSV-G envel-ope produced strong anterograde transgene expression (41,42).Three weeks after viral injection, SCI was induced by per-forming a bilateral dorsal column crush at the level of C4using fine forceps (13). Following injury, regeneration of thedescending motor system was assessed behaviourally (13,43).
Behavioural assessment
The behavioural tests used were grid- and ladder-walking,sticky tape removal and footprint analysis (13,44). Assessmentof all parameters was collected weekly before lesion and for 5weeks post-lesion and compared with the appropriate groupssuch as a control vector (EIAV-LacZ)-treated group as wellas a sham-operated group. At the end of the behaviouralassessment, the animals were sacrificed and perfused transcar-dially with 4% paraformaldehyde and tissue collected forhistology.
Detection of vector transduction and tracer labelling
Thirty-five micrometre coronal sections of the cortex wereobtained on a vibratome and randomly chosen for X-gal(5-bromo-4-chloro-3-indolyl-b-D-galactoside) staining orimmunohistochemistry. Using standard immunohistochemis-try protocols, sections were doubly incubated with eithermouse anti-bIII tubulin (1:1000, Promega) and rabbit anti-bgal/RARb or extrAvidin-FITC (to detect BDA, 1:400,Sigma) and rabbit anti-bgal/RARb, followed by detectionwith secondary antibodies. Dual colour images were obtainedat 10� magnification of the layer V cortical region and ana-lysed using the AxioVision LE 4.2 program. At least six trans-duced sites were analysed and quantified per animal (n¼6 ineach group).
Assessment of axonal regeneration
Regeneration of the descending corticospinal axons wasassessed by injecting 1 ml 10% BDA (MW 10 000, MolecularProbes) bilaterally into the sensorimotor cortex 4 weeks before
termination of experiment using the same coordinates as thoseused to administer the lentiviral vector. Longitudinal sectionsof the spinal cord (30 mm) were collected on a vibratome incold 0.01 M PBS with 0.1% sodium azide. The sections wererinsed in 3 � 5 min PBS-T (0.01 M PBS with 0.3% TritonX-100), then incubated with 0.3% H202 to remove endogenousperoxidases. After a further 3 � 5 min wash with PBS-T, thesections were incubated in avidin–biotin peroxidasecomplex (Vectastain ABC Elite Kit, Vector Laboratories)for 30 min at room temperature. After a 3 � 5 min washingin PBS-T, sections were incubated in tyramide (1:75) for10 min. After 3 � 5 min wash, the sections were incubatedwith extra-avidin FITC (1:400, Sigma) for 2 h before the sec-tions were washed and mounted onto slides and embeddedwith Flurosave reagent (Calbiochem). For quantitative analy-sis, four sections per animal containing the CST were selectedblindly and stained for BDA. Fibres were counted rostral andcaudal to the lesion site at every 0.5 mm under the microscopeat 20� magnification. Only main fibres that were runninglongitudinally in the white matter along the cord werecounted and the fibres tangent to these were not countedgiven that these are collateral fibres, not the main CSTfibres. The number of fibres at the rostral end of the lesion rep-resents the quality and quantity of labelled fibres before theinjury. For detection of BDA-labelled fibres in the ventralCST, 30 mm thick transverse sections of the spinal cordwere obtained and stained for BDA, as described previously.At least six random sections from each animal were selectedand the number of BDA fibres in the contralateral dorsomedialfuniculus (i.e. one side of the CST) and the ipsilateral ventro-medial funiculus were counted (n ¼ 6).
Histological confirmation of CST lesion
To determine whether the CST lesion was complete, trans-verse sections of the lumbar cord were stained for proteinkinase g (PKCg). Animals with staining of PKCg at thebase of the dorsal columns were excluded from the study.
ACKNOWLEDGEMENTS
The authors wish to thank Professor M. Okabe (OsakaUniversity, Japan) for providing the GFP rats and ProfessorW.F. Blakemore (University of Cambridge, U.K.) for supply-ing the GFP rats. The work was supported by ChristopherReeve Paralysis Foundation and Medical Research Council.
Conflict of Interest statement. The authors wish to declarecompeting financial interests.
REFERENCES
1. Grundy, D. and Swain, A. (2002) At the accident. In Grundy, D. andSwain, A. (eds), ABC of Spinal Cord Injury., BMJ Books, London, UK,pp. 1–4.
2. Corcoran, J., Shroot, B., Pizzey, J. and Maden, M. (2000) The role ofretinoic acid receptors in neurite outgrowth from different populations ofembryonic mouse dorsal root ganglia. J. Cell Sci., 113 (Pt 14),2567–2574.
3. Maden, M. (2002) Retinoic acid and limb regeneration—a personal view.Int. J. Dev. Biol., 46, 883–886.
Human Molecular Genetics, 2006, Vol. 15, No. 21 3117
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from

4. Maden, M. and Hind, M. (2003) Retinoic acid, a regeneration-inducingmolecule. Dev. Dyn., 226, 237–244.
5. Enderlin, V., Alfos, S., Pallet, V., Garcin, H., Zais-Braesco, V., Jaffard, R.and Higueret, P. (1997) Aging decreases the abundance of retinoic acid(RAR) and triiodothyronine (TR) nuclear receptor mRNA in rat brain:effect of the administration of retinoids. FEBS Lett., 412, 629–632.
6. Enderlin, V., Pallet, V., Alfos, S., Dargelos, E., Jaffard, R., Garcin, H. andHigueret, P. (1997) Age-related decreases in mRNA for brain nuclearreceptors and target genes are reversed by retinoic acid treatment.Neurosci. Lett., 229, 125–129.
7. Etchamendy, N., Enderlin, V., Marighetto, A., Vouimba, R.M., Pallet, V.,Jaffard, R. and Higueret, P. (2001) Alleviation of a selective age-relatedrelational memory deficit in mice by pharmacologically inducednormalization of brain retinoid signaling. J. Neurosci., 21, 6423–6429.
8. Etchamendy, N., Enderlin, V., Marighetto, A., Pallet, V., Higueret, P. andJaffard, R. (2003) Vitamin A deficiency and relational memory deficit inadult mice: relationships with changes in brain retinoid signalling. Behav.Brain Res., 145, 37–49.
9. Kastner, P., Mark, M., Ghyselinck, N., Krezel, W., Dupe, V.,Grondona, J.M. and Chambon, P. (1997) Genetic evidence that theretinoid signal is transduced by heterodimeric RXR/RAR functional unitsduring mouse development. Development, 124, 313–326.
10. Wong, L.F., Yip, P.K., Battaglia, A., Grist, J., Corcoran, J., Maden, M.,Azzouz, M., Kingsman, S.M., Kingsman, A.J., Mazarakis, N.D. andMcMahon, S.B. (2006) Retinoic acid receptor beta2 promotes functionalregeneration of sensory axons in the spinal cord. Nat. Neurosci., 9,243–250.
11. Corcoran, J., So, P.L., Barber, R.D., Vincent, K.J., Mazarakis, N.D.,Mitrophanous, K.A., Kingsman, S.M. and Maden, M. (2002) Retinoic acidreceptor beta2 and neurite outgrowth in the adult mouse spinal cord invitro. J. Cell Sci., 115, 3779–3786.
12. Tom, V.J., Doller, C.M., Malouf, A.T. and Silver, J. (2004)Astrocyte-associated fibronectin is critical for axonal regeneration in adultwhite matter. J. Neurosci., 24, 9282–9290.
13. Bradbury, E.J., Moon, L.D., Popat, R.J., King, V.R., Bennett, G.S.,Patel, P.N., Fawcett, J.W. and McMahon, S.B. (2002) ChondroitinaseABC promotes functional recovery after spinal cord injury. Nature,416, 636–640.
14. Mori, M., Kose, A., Tsujino, T. and Tanaka, C. (1990)Immunocytochemical localization of protein kinase C subspecies in the ratspinal cord: light and electron microscopic study. J. Comp. Neurol., 299,167–177.
15. Nathan, B.P., Jiang, Y.,Wong,G.K., Shen, F.,Brewer,G.J. andStruble,R.G.(2002) Apolipoprotein E4 inhibits, and apolipoprotein E3 promotes neuriteoutgrowth in cultured adult mouse cortical neurons through the low-densitylipoprotein receptor-related protein. Brain Res., 928, 96–105.
16. Viel, J.J., McManus, D.Q., Cady, C., Evans, M.S. and Brewer, G.J. (2001)Temperature and time interval for culture of postmortem neurons fromadult rat cortex. J. Neurosci. Res., 64, 311–321.
17. Viel, J.J., McManus, D.Q. and Brewer, G.J. (2004) Postmortem effect ofpentobarbital anesthetic on survival of adult cortical neurons in primaryculture. Brain Res., 1009, 219–222.
18. Zetterstrom, R.H., Simon, A., Giacobini, M.M.J., Eriksson, U. andOlson, L. (1994) Localization of cellular retinoid-binding proteinssuggests specific roles for retinoids in the adult central nervous system.Neuroscience, 62, 899–918.
19. Zetterstrom, R.H., Lindqvist, E., Mata de Urquiza, A., Tomac, A.,Eriksson, U., Perlmann, T. and Olson, L. (1999) Role of retinoids in theCNS: differential expression of retinoid binding proteins and receptorsand evidence for presence of retinoic acid. Eur. J. Neurosci., 11,407–416.
20. Wagner, E., Luo, T., Sakai, Y., Parada, L.F. and Drager, U.C. (2006)Retinoic acid delineates the topography of neuronal plasticity in postnatalcerebral cortex. Eur. J. Neurosci., 24, 329–340.
21. Chen, M.S., Huber, A.B., van der Haar, M.E., Frank, M., Schnell, L.,Spillmann, A.A., Christ, F. and Schwab, M.E. (2000) Nogo-A is amyelin-associated neurite outgrowth inhibitor and an antigen formonoclonal antibody IN-1. Nature, 403, 434–439.
22. GrandPre, T., Nakamura, F., Vartanian, T. and Strittmatter, S.M. (2000)Identification of the Nogo inhibitor of axon regeneration as a Reticulonprotein. Nature, 403, 439–444.
23. Kottis, V., Thibault, P., Mikol, D., Xiao, Z.C., Zhang, R., Dergham, P. andBraun, P.E. (2002) Oligodendrocyte-myelin glycoprotein (OMgp) is aninhibitor of neurite outgrowth. J. Neurochem., 82, 1566–1569.
24. McKerracher, L., David, S., Jackson, D.L., Kottis, V., Dunn, R.J. andBraun, P.E. (1994) Identification of myelin-associated glycoprotein as amajor myelin-derived inhibitor of neurite growth. Neuron, 13, 805–811.
25. Mukhopadhyay, G., Doherty, P., Walsh, F.S., Crocker, P.R. andFilbin, M.T. (1994) A novel role for myelin-associated glycoprotein as aninhibitor of axonal regeneration. Neuron, 13, 757–767.
26. Prinjha, R., Moore, S.E., Vinson, M., Blake, S., Morrow, R., Christie, G.,Michalovich, D., Simmons, D.L. and Walsh, F.S. (2000) Inhibitor ofneurite outgrowth in humans. Nature, 403, 383–384.
27. Wang,K.C., Koprivica, V., Kim, J.A., Sivasankaran, R., Guo, Y., Neve, R.L.and He, Z. (2002) Oligodendrocyte-myelin glycoprotein is a Nogo receptorligand that inhibits neurite outgrowth. Nature, 417, 941–944.
28. Davies, S.J., Goucher, D.R., Doller, C. and Silver, J. (1999) Robustregeneration of adult sensory axons in degenerating white matter of theadult rat spinal cord. J. Neurosci., 19, 5810–5822.
29. Schnell, L. and Schwab, M.E. (1990) Axonal regeneration in the rat spinalcord produced by an antibody against myelin-associated neurite growthinhibitors. Nature, 343, 269–272.
30. Li, Y., Field, P.M. and Raisman, G. (1997) Repair of adult ratcorticospinal tract by transplants of olfactory ensheathing cells. Science,277, 2000–2002.
31. Moreno-Flores, M.T., Martin-Aparicio, E., Martin-Bermejo, M.J.,Agudo, M., McMahon, S., Avila, J., Az-Nido, J. and Wandosell, F.(2003) Semaphorin 3C preserves survival and induces neuritogenesisof cerebellar granule neurons in culture. J. Neurochem., 87, 879–890.
32. Steward, O., Zheng, B.H. and Tessier-Lavigne, M. (2003) Falseresurrections: distinguishing regenerated from spared axons in the injuredcentral nervous system. J. Comp. Neurol., 459, 1–8.
33. Brosamle, C. and Schwab, M.E. (1997) Cells of origin, course, andtermination patterns of the ventral, uncrossed component of the mature ratcorticospinal tract. J. Comp. Neurol., 386, 293–303.
34. Neumann, S., Bradke, F., Tessier-Lavigne, M. and Basbaum, A.I. (2002)Regeneration of sensory axons within the injured spinal cord induced byintraganglionic cAMP elevation. Neuron, 34, 885–893.
35. Qiu, J., Cai, D., Dai, H., McAtee, M., Hoffman, P.N., Bregman, B.S. andFilbin, M.T. (2002) Spinal axon regeneration induced by elevation ofcyclic AMP. Neuron, 34, 895–903.
36. Weidner, N., Ner, A., Salimi, N. and Tuszynski, M.H. (2001) Spontaneouscorticospinal axonal plasticity and functional recovery after adult centralnervous system injury. Proc. Natl Acad. Sci. USA, 98, 3513–3518.
37. Tatusova, T.A. and Madden, T.L. (1999) BLAST 2 sequences, a new toolfor comparing protein and nucleotide sequences. FEMS Microbiol. Lett.,174, 247–250.
38. Brewer, G.J. (1997) Isolation and culture of adult rat hippocampalneurons. J. Neurosci. Methods, 71, 143–155.
39. Ito, T., Suzuki, A., Imai, E., Okabe, M. and Hori, M. (2001) Bone marrowis a reservoir of repopulating mesangial cells during glomerularremodeling. J. Am. Soc. Nephrol., 12, 2625–2635.
40. Neafsey, E.J., Bold, E.L., Haas, G., Hurley-Gius, K.M., Quirk, G.,Sievert, C.F. and Terreberry, R.R. (1986) The organization of the ratmotor cortex: a microstimulation mapping study. Brain Res., 396, 77–96.
41. Yip, P.K., Wong, L.F., Grist, J., Kingsman, S.M., Kingsman, A.J.,Mazarakis, N.D. and McMahon, S.B. (2005) Comparison of pattern andefficacy of neuronal transduction in the dorsal root ganglia by lentiviralpseudotypes in vivo. J. Physiol., 565P, C119.
42. Mazarakis, N.D., Azzouz, M., Rohll, J.B., Ellard, F.M., Wilkes, F.J.,Olsen, A.L., Carter, E.E., Barber, R.D., Baban, D.F., Kingsman, S.M.et al. (2001) Rabies virus glycoprotein pseudotyping of lentiviral vectorsenables retrograde axonal transport and access to the nervous system afterperipheral delivery. Hum. Mol. Genet., 10, 2109–2121.
43. Ramer, M.S., Priestley, J.V. and McMahon, S.B. (2000) Functionalregeneration of sensory axons into the adult spinal cord. Nature, 403,312–316.
44. Kunkel-Bagden, E., Dai, H.N. and Bregman, B.S. (1993) Methods toassess the development and recovery of locomotor function after spinalcord injury in rats. Exp. Neurol., 119, 153–164.
3118 Human Molecular Genetics, 2006, Vol. 15, No. 21
by guest on October 1, 2014
http://hmg.oxfordjournals.org/
Dow
nloaded from
Related Documents











