Experimental Parasitology 95, 96–105 (2000) doi:10.1006/expr.2000.4523, available online at http://www.idealibrary.com on Leishmania sp: Comparative Study with Toxoplasma gondii and Trypanosoma cruzi in Their Ability to Initialize IL-12 and IFN-g Synthesis Milton A. P. Oliveira,* Helton C. Santiago,* Cristiane R. Lisboa,* Isabela P. Ceravollo,² Giorgio Trinchieri,‡ Ricardo T Gazzinelli,* , ² and Leda Q. Vieira* *Departmento de Bioquı ´mica e Imunologia, Universidade Federal de Minas Gerais, 30161-970, Belo Horizonte, MG, Brazil; ² Centro de Pesquisas Rene ´ Rachou, FIOCRUZ, 30190-002 Belo Horizonte, MG, Brazil; and ‡Wistar Institute of Anatomy and Biology, 3601 Spruce Street, Philadelphia, Pennsylvania 19104, U.S.A. Oliveira, M. A. P., Santiago, H. C., Lisboa, C. R., Ceravollo, I. P., INTRODUCTION Trinchieri, G., Gazzinelli, R. T, and Vieira, L. Q. 2000. Leishmania sp: Comparative study with Toxoplasma gondii and Trypanosoma cruzi in their ability to initialize IL-12 and IFN-g synthesis. Experimental Parasitology 95, 96–105. We compared in vitro and in vivo induction The intracellular parasites Trypanosoma cruzi and Tox- of IL-12 (p40) and IFN-g by mouse cells stimulated with Toxoplasma oplasma gondii, although phylogenetically distinct, display gondii, Trypanosoma cruzi, and different species of Leishmania. Spleen several common features in their biology and life cycle cells cultured in vitro with T. cruzi or T. gondii, but not with Leishmania, in vertebrate hosts. Both parasites infect a wide range of produced IL-12 (p40) and IFN-g. Accordingly, IL-12 (p40) was pro- mammals, including humans. In addition, these intracellular duced by macrophages stimulated in vitro with live T. cruzi or T. gondii or membrane glycoconjugates obtained from trypomastigotes organisms can infect and replicate inside any kind of nucle- or tachyzoites. No IL-12 production was detected when macrophages ated cell from their vertebrate hosts (Frenkel 1988; Burleigh were stimulated with live parasites or glycoconjugates from Leish- and Andrews, 1995). The lack of host cell specificity in the mania, regardless of priming with IFN-g. In vivo, only T. cruzi and T. absence of immunity suggests that these parasites are highly gondii induced the synthesis of IL-12 and IFN-g by mouse spleen cells virulent. Thus, during the early phase of infection and before after intraperitoneal injection of parasites. When injected subcutane- the development of specific immunity, both T. gondii and ously, live Leishmania sp. induced IL-12 (p40) and IFN-g production by draining lymph node cells, albeit the levels were slightly lower than T. cruzi can be found in different tissues and organs and those induced by infection with T. gondii or T. cruzi using the same are normally accompanied by a mononuclear inflammatory route. Together our results indicate that under different conditions, the reaction in small necrotic foci (Frenkel 1988; Silva et al. intracellular protozoa T. gondii and T. cruzi are more potent stimulators 1985). With the rapid onset of parasite-specific CMI, most of IL-12 and IFN-g synthesis by host immune cells than parasites of parasites are cleared from the host tissues, the necrotic loci the genus Leishmania. q 2000 Academic Press regenerate, and the parasite distribution tends to be more Index Descriptors and Abbreviations: Trypanosoma cruzi; Leish- mania amazonensis; Leishmania major; Toxoplasma gondii; Th1; mac- localized (Frenkel 1988; Andrade 1991). In the case of either rophage; membrane glycoconjugates; cell-mediated immunity (CMI); infection, induction of IL-12 and CMI maintained by Th1 interleukin-4 (IL-4); interleukin-12 (IL-12); interferon-g (IFN-g); solu- lymphocytes and IFN-g are thought to be major events pro- ble antigen of T. gondii (STAg); concanavalin A (Con A); lipophospho- tecting the host against rapid parasite multiplication, conse- glycan derived from Leishmania promastigotes (LPG) and glycosyl- quent pathology, and lethality during the acute phase of inositolphospholipids (GIPLs); glycosylphosphatidilinositol-linked infection (Gazzinelli et al. 1993a; Silva et al. 1992). How- mucin-like glycocoproteins derived from T. cruzi trypomastigotes (GPI mucins). ever, the parasite is not eliminated by the immune system and the vertebrate host will act as a reservoir, playing a 0014-4894/00 $35.00 96 Copyright q 2000 by Academic Press All rights of reproduction in any form reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Experimental Parasitology 95, 96–105 (2000)
doi:10.1006/expr.2000.4523, available online at http://www.idealibrary.com on
Leishmania sp: Comparative Study with Toxoplasma gondii and Trypanosomacruzi in Their Ability to Initialize IL-12 and IFN-g Synthesis
Milton A. P. Oliveira,* Helton C. Santiago,* Cristiane R. Lisboa,* Isabela P. Ceravollo,†Giorgio Trinchieri,‡ Ricardo T Gazzinelli,*,† and Leda Q. Vieir
ina2 Bphi
*Departmento de Bioquımica e Imunologia, Universidade Federal de MMG, Brazil; †Centro de Pesquisas Rene Rachou, FIOCRUZ, 30190-00‡Wistar Institute of Anatomy and Biology, 3601 Spruce Street, Philadel
Oliveira, M. A. P., Santiago, H. C., Lisboa, C. R., Ceravollo, I. P.,Trinchieri, G., Gazzinelli, R. T, and Vieira, L. Q. 2000. Leishmaniasp: Comparative study with Toxoplasma gondii and Trypanosoma cruziin their ability to initialize IL-12 and IFN-g synthesis. ExperimentalParasitology 95, 96–105. We compared in vitro and in vivo inductionof IL-12 (p40) and IFN-g by mouse cells stimulated with Toxoplasmagondii, Trypanosoma cruzi, and different species of Leishmania. Spleencells cultured in vitro with T. cruzi or T. gondii, but not with Leishmania,produced IL-12 (p40) and IFN-g. Accordingly, IL-12 (p40) was pro-duced by macrophages stimulated in vitro with live T. cruzi or T.gondii or membrane glycoconjugates obtained from trypomastigotesor tachyzoites. No IL-12 production was detected when macrophageswere stimulated with live parasites or glycoconjugates from Leish-mania, regardless of priming with IFN-g. In vivo, only T. cruzi and T.gondii induced the synthesis of IL-12 and IFN-g by mouse spleen cellsafter intraperitoneal injection of parasites. When injected subcutane-ously, live Leishmania sp. induced IL-12 (p40) and IFN-g productionby draining lymph node cells, albeit the levels were slightly lower thanthose induced by infection with T. gondii or T. cruzi using the sameroute. Together our results indicate that under different conditions, theintracellular protozoa T. gondii and T. cruzi are more potent stimulatorsof IL-12 and IFN-g synthesis by host immune cells than parasites ofthe genus Leishmania. q 2000 Academic Press
Index Descriptors and Abbreviations: Trypanosoma cruzi; Leish-mania amazonensis; Leishmania major; Toxoplasma gondii; Th1; mac-
rophage; membrane glycoconjugates; cell-mediated immunity (CMI);interleukin-4 (IL-4); interleukin-12 (IL-12); interferon-g (IFN-g); solu-ble antigen of T. gondii (STAg); concanavalin A (Con A); lipophospho-glycan derived from Leishmania promastigotes (LPG) and glycosyl-inositolphospholipids (GIPLs); glycosylphosphatidilinositol-linkedmucin-like glycocoproteins derived from T. cruzi trypomastigotes(GPI mucins).96
a*
s Gerais, 30161-970, Belo Horizonte,elo Horizonte, MG, Brazil; anda, Pennsylvania 19104, U.S.A.
INTRODUCTION
The intracellular parasites Trypanosoma cruzi and Tox-oplasma gondii, although phylogenetically distinct, displayseveral common features in their biology and life cyclein vertebrate hosts. Both parasites infect a wide range ofmammals, including humans. In addition, these intracellularorganisms can infect and replicate inside any kind of nucle-ated cell from their vertebrate hosts (Frenkel 1988; Burleighand Andrews, 1995). The lack of host cell specificity in theabsence of immunity suggests that these parasites are highlyvirulent. Thus, during the early phase of infection and beforethe development of specific immunity, both T. gondii andT. cruzi can be found in different tissues and organs andare normally accompanied by a mononuclear inflammatoryreaction in small necrotic foci (Frenkel 1988; Silva et al.1985). With the rapid onset of parasite-specific CMI, mostparasites are cleared from the host tissues, the necrotic lociregenerate, and the parasite distribution tends to be morelocalized (Frenkel 1988; Andrade 1991). In the case of eitherinfection, induction of IL-12 and CMI maintained by Th1
lymphocytes and IFN-g are thought to be major events pro-tecting the host against rapid parasite multiplication, conse-quent pathology, and lethality during the acute phase ofinfection (Gazzinelli et al. 1993a; Silva et al. 1992). How-ever, the parasite is not eliminated by the immune systemand the vertebrate host will act as a reservoir, playing a0014-4894/00 $35.00Copyright q 2000 by Academic Press
All rights of reproduction in any form reserved.

IL-12 AND IFN-g INDUCTION BY INTRACELLULAR PROTOZOA
clearly important role in T. gondii or T. cruzi disseminationand maintenance of their life cycle (Brener and Gazzinelli1997).
The intracellular parasites from the genus Leishmaniaemploy a quite distinct strategy to perpetuate their life cycle.In their vertebrate hosts Leishmania are slow-growing para-sites that reside and proliferate inside macrophage vacuoles,a very hostile environment (Russell and Talamas-Rohana1989). Thus, if parasites are able to compromise the induc-tion or function of IFN-g or suppress macrophage effectorfunctions, they are be more successful at establishing infec-tion, persisting in the vertebrate host, and favoring encounterwith their insect vector and transmission. Thus, it has beenproposed that at early stage of infection parasites from theLeishmania genus are able to evade the induction of CMIin order to persist in host tissues (Carrera et al. 1996).
In order to compare the ability of Leishmania sp., T. cruzi,and T. gondii to induce IL-12 and/or IFN-g, we used differentapproaches employing in vitro or in vivo stimulation ofsplenocytes, peritoneal macrophages, and lymph node cells
from BALB/c or C57BL/6 mice. Our in vitro data show thatT. cruzi and T. gondii but not Leishmania sp. are potentwith 2 mM L-glutamine, 80 mg/ml gentamicin, and 10% heat-inacti-
inducers of IL-12 and IFN-g. In vivo stimulation experimentsindicate that, under the conditions we used, any of the threeprotozoa lead to local production of IL-12 and IFN-g, whileinfection with either T. gondii or T. cruzi, but not withLeishmania sp., triggered an early systemic production ofthese cytokines.
MATERIAL AND METHODS
Animals. Female C3H/HeJ, C57BL/6, and BALB/c mice, 4–8weeks old, were obtained from either FIOCRUZ (Rio de Janeiro, RJ,Brazil) or CEBIO (Instituto de Ciencias Biologicas, UFMG, BeloHorizonte, MG, Brazil). Animals were kept in a conventional animalfacility at controlled temperature, light/dark cycles, and environmen-tal barriers.
Parasites. (1) T. cruzi trypomastigotes: all experiments employedthe Y strain of T. cruzi. Trypomastigotes were obtained after 4–5days of infection of murine fibroblasts L-929 (American Type CultureCollection, Rockville, MD) cultured in D-minimum essential medium
(DMEM, Sigma Chemical Co, St. Louis, MO) supplemented with 2mM L-glutamine (Sigma Chemical Co.), 80 mg/ml gentamicin (SigmaChemical Co.), 10% heat-inactivated fetal calf serum (FCS; NUTRI-CELL, Campinas, SP, Brazil), and 0.3% sodium bicarbonate, at 33Cin a humidified atmosphere containing 5% CO2. Trypomastigotes werecollected from the supernatant of cultures, centrifuged at 40g at 48Cfor 10 min for separation of debris, followed by another centrifugationat 700g at 48C for 10 min. The resulting pellet, containing the livetrypomastigotes, was used to prepare parasite extracts or in experiments97
with live parasites. (2) T. cruzi blood trypomastigotes: blood trypomas-tigotes were obtained by passage through C57BL/6 mice. Mice wereinfected with 5 3 103 blood forms and bled 7 days after infection.Parasites in blood were quantified and injected into C57BL/6 or BALB/c mice, at 20 parasites per mouse, in order to obtain chronically infectedanimals. (3) T. gondii tachyzoites: the RH strain of T. gondii was used.Tachyzoites were obtained from infected human fibroblasts (cell line2C4, American Type Culture Collection—ATCC). The cultures weremaintained in DMEM supplemented with 5% FCS at 338C in 5% CO2.After 4–5 days, tachyzoites were collected and separated from celldebris and concentrated as described for T. cruzi. Cysts were obtainedas previously described from infected C57BL/6 mice (Gazzinelli et al.1994). (4) Leishmania sp. promastigotes: a clone of L. major (WHOMHOM/IL/80/Friedlin) and an isolate of L. amazonensis PH8 (IFLA/Br/67/PH8) were cultured in Grace’s insect medium (GIBCO BRL,Grand Island, NY, USA) supplemented with 20% heat-inactivated FCSand 2 mM L-glutamine, at 258C. Promastigotes were collected at thestationary phase (5th day of culture), centrifuged at 700g at 48C for20 min, and washed in phosphate-buffered saline (PBS). The resultingpellet was used to stimulate cells in vitro or for infections.
Infections. Mice were infected in the peritoneal cavity or subcuta-neously in the footpad with T. gondii, T. cruzi, L. major, or L. amazo-nensis, as described for each experiment under Results.
Parasite antigens. Parasite antigens for in vitro stimulation of cellcultures were prepared from T. cruzi (Aliberti et al. 1996), L. major(Afonso and Scott 1993), and T. gondii (Gazzinelli et al. 1991) aspreviously described. Antigens were added to cultures at 25 mg ofprotein/ml of culture, and protein was assayed using the Lowry method(Lowry et al. 1951).
Purification of T. cruzi glycoconjugates. The glycosylphosphati-dylinositol-anchored mucin-like glycoproteins (GPI mucins) were iso-lated from T. cruzi trypomastigotes as previously described (Almeidaet al. 1994; Serrano et al. 1995; McNeely et al. 1989) using sequentialorganic extraction followed by hydrophobic interaction chromatogra-phy in octyl-Sepharose columns (Pharmacia Biotech, Sweden) andelution with a propan-1-ol gradient (5–60%), and kindly provided byDr. Helia Cannizzaro, Department of Biochemistry and Immunology,Federal University of Minas Gerais, Belo Horizonte, MG, Brazil.
Purification of Leishmania sp. LPG and GIPLs. LPG from L.major promastigotes were kindly provided by Dr. David Sacks fromthe Laboratory of Parasitic Diseases, NIAID (NIH, Bethesda, MD).GIPLs were obtained from amastigote forms of L. donovani followinga protocol previously described (McNeely et al. 1989) and kindlyprovided by Dr. Salvatore Turco, Department of Biochemistry, Univer-sity of Kentucky Medical Center (Lexington, KY).
Macrophage preparations. Mice were inoculated intraperitoneallywith 1 ml of 3% thioglycolate medium and the elicited peritonealexudate cells harvested in cold serum-free RPMI by peritoneal lavage4–5 days later. The medium used in the macrophage cultures (completemedium) consisted of RPMI 1640 (Sigma Chemical Co.) supplemented
vated FCS. Macrophages were ressuspended in complete medium at2 3 106 cells/ml, and 0.1-ml aliquots dispensed into wells of a 96-well plate. The cells were allowed to adhere at 378C in 5% CO2. Aftera 3-h incubation period the cells were washed once with PBS at roomtemperature and 0.1 ml of complete medium was added to each well.In order to assess the effect of live parasites or parasite glycoconjugateson induction of IL-12, macrophages were prepared as described aboveand exposed to different numbers of either T. cruzi tissue culturetrypomastigotes, T. gondii tachyzoites, or Leishmania promastigotes,

98
as indicated in the figure legends. Different T. cruzi (GPI mucins), T.gondii (STAg), L. major (LPG), or L. donovani (GIPLs) glycoconju-gates were added to the macrophage cultures in the presence or absenceof IFN-g (Genzyme Corp., Cambridge, MA). The macrophages werethen incubated with test and control agents in a final volume of 200ml/well. Supernatants were collected after 48 h of culture for IL-12(p40) measurements.
Spleen and lymph node cell preparations. Spleens were removedaseptically and single cell suspensions were prepared in completemedium after lysis of red cells (Johnstone and Thorpe 1982). Spleencells from mice at 5 3 106 were then stimulated with live forms ofT. gondii (tachyzoites), T. cruzi (trypomastigotes), or Leishmania sp.(promastigotes). After incubation for 48 h, supernatants were removedand assayed for IL-12 (p40) and IFN-g. Alternatively, cells were cul-tured with 5 mg/ml Con A or parasite antigen preparations at 25 mg/ml for 48 h. Popliteal lymph node cells were prepared similarly exceptthat red cell lysis was omitted.
Cytokine assays. IL-12 was assayed in cell culture supernatantsby a capture ELISA, as previously described (Vieira et al. 1994), usingthe rat mAb C17.15 (5 mg/ml) and biotinylated C15.6 rat mAb ascapture and detection of bound IL-12, respectively. Recombinant mu-rine IL-12 (a kind gift from Dr. Phillip Scott, School of VeterinaryMedicine, University of Pennsylvania, PA) was used as standard. OPD(Sigma Chemical Co.) was used as substrate for the enzymatic reactionto detect peroxidase-labeled streptavidin bound to biotinylated C15.6.The sensitivity of this method was 0.3 ng/ml. Both anti IL-12 p40antibodies were kind gifts from Dr. Maria Wysocka (Wistar Institute,PA). IFN-g was assayed in a two-site ELISA (Afonso and Scott 1993)using a rat anti-IFN-g mAb R46A2 (ATCC) and a polyclonal rabbitserum specific for the cytokine, a kind gift from Dr. Jay Farrell,University of Pennsylvania, PA). IFN-g levels were calculated by
reference to a standard curve constructed with recombinant cytokine(Genzyme, Cambridge, MA). The sensitivity of this method was 25pg/ml. IL-4 was measured as previously described (Gazzinelli et al. 1991) in a biological assay using the CT4S IL-4-dependent cell linein the presence of neutralizing monoclonal antibody against IL-2 (S4B6,ATCC). IL-4 levels were calculated by reference to a standard curveconstructed with recombinant IL-4 (Genzyme). A neutralizing mono-clonal antibody against murine IL-4 (11B11, ATCC) was used to ensurethat CT4S proliferation was in response to IL-4. The sensitivity of thismethod was 0.1 U/ml.RESULTS
Antigen-specific Th1 response is less intense in experi-mental murine leishmaniasis than in trypanosomiasis and
toxoplasmosis. In order to compare the intensity of theTh1 response in mice infected with intracellular parasites,we infected C57BL/6 and BALB/c mice with L. major, T.gondii, or T. cruzi and compared the production of IL-4 andIFN-g by spleen cells 9 weeks after infection. Doses ofparasites, although different from each other, are those estab-lished as models to study chronic stages of these diseases(e.g., Afonso and Scott 1993; Gazzinelli et al. 1992; ourOLIVEIRA ET AL.
unpublished work). The cytokine profiles found were thoseexpected: both strains of mice responded to T. gondii andT. cruzi with a strong production of IFN-g and no detectableIL-4, as evidenced by culturing spleen cells in vitro in thepresence of parasite antigens (Fig. 1). On the other hand,while infection with L. major triggered the production ofIFN-g by spleen cells from C57BL/6 and no detectable IL-4, BALB/c mice showed the opposite profile: high levels ofIL-4 were produced in response to in vitro antigenic stimulusand low levels of IFN-g were detected (Fig. 1). Interestingly,production of IFN-g in response to Leishmania antigens bymice infected with L. major was systematically lower thanthe response to homologous antigens by spleen cells frommice infected with the other two parasites. When spleencells from infected C57BL/6 mice were stimulated in vitrowith Con A, however, IFN-g and IL-4 levels of productionwere similar in all groups (Fig. 1). In contrast, spleen cellsfrom BALB/c mice infected with L. major produced lowerlevels of IFN-g in response to mitogen than T. gondii- orT. cruzi-infected BALB/c mice. Also, higher levels of IL-4were detected in supernatants from spleen cells from L.major-infected BALB/c mice stimulated in vitro with Con A.
Leishmania promastigotes are incapable of inducingIL-12 and IFN-g production by spleen cells or peritonealmacrophages in vitro. Spleen cells from noninfectedC57BL/6 mice were stimulated in vitro with parasites andIL-12 and IFN-g production levels were assayed 48 h later(Fig. 2). Both T. gondii and T. cruzi induced IL-12 and IFN-g production by naive spleen cells in vitro. T. gondii wascapable of inducing both cytokines at a proportion as lowas 1/103 parasites per spleen cell. The amount of IL-12 foundin supernatants of cultures correlated well with the inductionof IFN-g (compare Figs. 2A and 2B). In contrast, neitherL. major nor L. amazonensis induced IL-12 at any ratio ofparasite to spleen cell except at 1 parasite per 1000 cells.These levels are considered very low and were not presentin all experiments. Since they were absent in higher para-site:cell ratios we consider them insignificant. Spleen cellsfrom IL-12-/- mice were never found to produce IFN-g inresponse to Leishamania stimulation in vitro (data notshown).
Similarly, both T. gondii or T. cruzi induced IL-12 produc-
tion by inflammatory macrophages stimulated in vitro, whileL. major and L. amazonensis did not (Fig. 3A). Macrophagepriming with IFN-g enhanced IL-12 production by macro-phages exposed to T. gondii or T. cruzi (Fig. 3B). The addi-tion of recombinant IFN-g had no effect on the lack ofinduction of IL-12 by L. major or L. amazonensis. Accord-ingly, IL-12 production by thioglycolate-elicited peritonealmacrophages could be achieved by stimulating these cells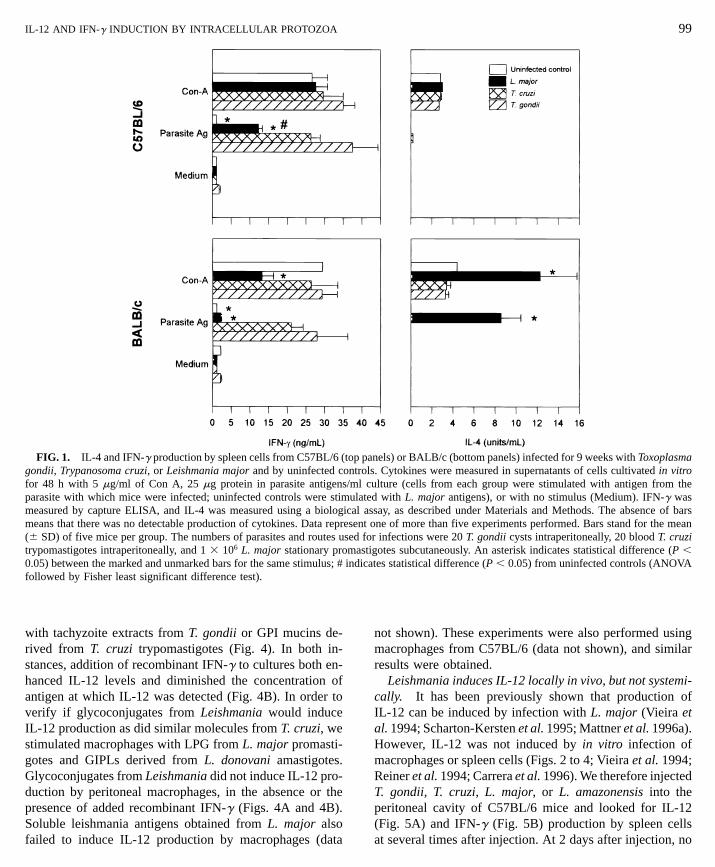
FIG. 1. IL-4 and IFN-g production by spleen cells from C57BL/6 (top panels) or BALB/c (bottom panels) infected for 9 weeks with Toxoplasmagondii, Trypanosoma cruzi, or Leishmania major and by uninfected controls. Cytokines were measured in supernatants of cells cultivated in vitro
ml culture (cells from each group were stimulated with antigen from theulatal a
sented fmasndic
for 48 h with 5 mg/ml of Con A, 25 mg protein in parasite antigens/parasite with which mice were infected; uninfected controls were stimmeasured by capture ELISA, and IL-4 was measured using a biologicmeans that there was no detectable production of cytokines. Data repre(6 SD) of five mice per group. The numbers of parasites and routes ustrypomastigotes intraperitoneally, and 1 3 106 L. major stationary pro0.05) between the marked and unmarked bars for the same stimulus; # ifollowed by Fisher least significant difference test).
with tachyzoite extracts from T. gondii or GPI mucins de-rived from T. cruzi trypomastigotes (Fig. 4). In both in-stances, addition of recombinant IFN-g to cultures both en-hanced IL-12 levels and diminished the concentration ofantigen at which IL-12 was detected (Fig. 4B). In order toverify if glycoconjugates from Leishmania would induce
IL-12 production as did similar molecules from T. cruzi, westimulated macrophages with LPG from L. major promasti-gotes and GIPLs derived from L. donovani amastigotes.Glycoconjugates from Leishmania did not induce IL-12 pro-duction by peritoneal macrophages, in the absence or thepresence of added recombinant IFN-g (Figs. 4A and 4B).Soluble leishmania antigens obtained from L. major alsofailed to induce IL-12 production by macrophages (dataed with L. major antigens), or with no stimulus (Medium). IFN-g wasssay, as described under Materials and Methods. The absence of barsone of more than five experiments performed. Bars stand for the mean
or infections were 20 T. gondii cysts intraperitoneally, 20 blood T. cruzitigotes subcutaneously. An asterisk indicates statistical difference (P ,ates statistical difference (P , 0.05) from uninfected controls (ANOVA
not shown). These experiments were also performed usingmacrophages from C57BL/6 (data not shown), and similarresults were obtained.
Leishmania induces IL-12 locally in vivo, but not systemi-cally. It has been previously shown that production ofIL-12 can be induced by infection with L. major (Vieira et
IL-12 AND IFN-g INDUCTION BY INTRACELLULAR PROTOZOA 99
al. 1994; Scharton-Kersten et al. 1995; Mattner et al. 1996a).However, IL-12 was not induced by in vitro infection ofmacrophages or spleen cells (Figs. 2 to 4; Vieira et al. 1994;Reiner et al. 1994; Carrera et al. 1996). We therefore injectedT. gondii, T. cruzi, L. major, or L. amazonensis into theperitoneal cavity of C57BL/6 mice and looked for IL-12(Fig. 5A) and IFN-g (Fig. 5B) production by spleen cellsat several times after injection. At 2 days after injection, no
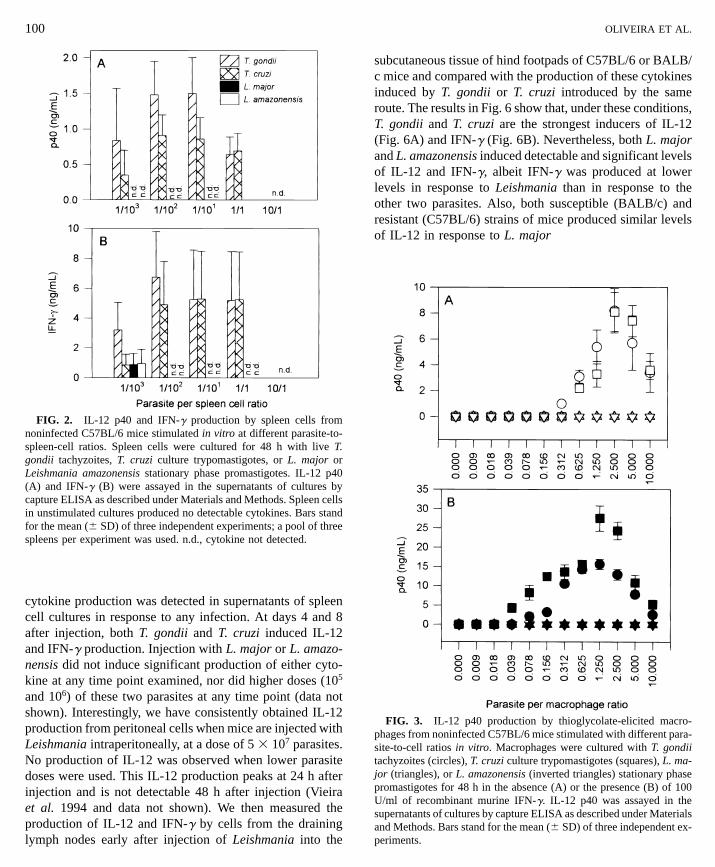
FIG. 2. IL-12 p40 and IFN-g production by spleen cells fromnoninfected C57BL/6 mice stimulated in vitro at different parasite-to-spleen-cell ratios. Spleen cells were cultured for 48 h with live T.
FIG. 3. IL-12 p40 production by thioglycolate-elicited macro-
gondii tachyzoites, T. cruzi culture trypomastigotes, or L. major orLeishmania amazonensis stationary phase promastigotes. IL-12 p40(A) and IFN-g (B) were assayed in the supernatants of cultures bycapture ELISA as described under Materials and Methods. Spleen cellsin unstimulated cultures produced no detectable cytokines. Bars standfor the mean (6 SD) of three independent experiments; a pool of threespleens per experiment was used. n.d., cytokine not detected.
cytokine production was detected in supernatants of spleencell cultures in response to any infection. At days 4 and 8after injection, both T. gondii and T. cruzi induced IL-12and IFN-g production. Injection with L. major or L. amazo-nensis did not induce significant production of either cyto-kine at any time point examined, nor did higher doses (105
and 106) of these two parasites at any time point (data notshown). Interestingly, we have consistently obtained IL-12
production from peritoneal cells when mice are injected withLeishmania intraperitoneally, at a dose of 5 3 107 parasites.No production of IL-12 was observed when lower parasitedoses were used. This IL-12 production peaks at 24 h afterinjection and is not detectable 48 h after injection (Vieiraet al. 1994 and data not shown). We then measured theproduction of IL-12 and IFN-g by cells from the draininglymph nodes early after injection of Leishmania into theOLIVEIRA ET AL.
subcutaneous tissue of hind footpads of C57BL/6 or BALB/c mice and compared with the production of these cytokinesinduced by T. gondii or T. cruzi introduced by the sameroute. The results in Fig. 6 show that, under these conditions,
100
T. gondii and T. cruzi are the strongest inducers of IL-12(Fig. 6A) and IFN-g (Fig. 6B). Nevertheless, both L. majorand L. amazonensis induced detectable and significant levelsof IL-12 and IFN-g, albeit IFN-g was produced at lowerlevels in response to Leishmania than in response to theother two parasites. Also, both susceptible (BALB/c) andresistant (C57BL/6) strains of mice produced similar levelsof IL-12 in response to L. major
phages from noninfected C57BL/6 mice stimulated with different para-site-to-cell ratios in vitro. Macrophages were cultured with T. gondiitachyzoites (circles), T. cruzi culture trypomastigotes (squares), L. ma-jor (triangles), or L. amazonensis (inverted triangles) stationary phasepromastigotes for 48 h in the absence (A) or the presence (B) of 100U/ml of recombinant murine IFN-g. IL-12 p40 was assayed in thesupernatants of cultures by capture ELISA as described under Materialsand Methods. Bars stand for the mean (6 SD) of three independent ex-periments.
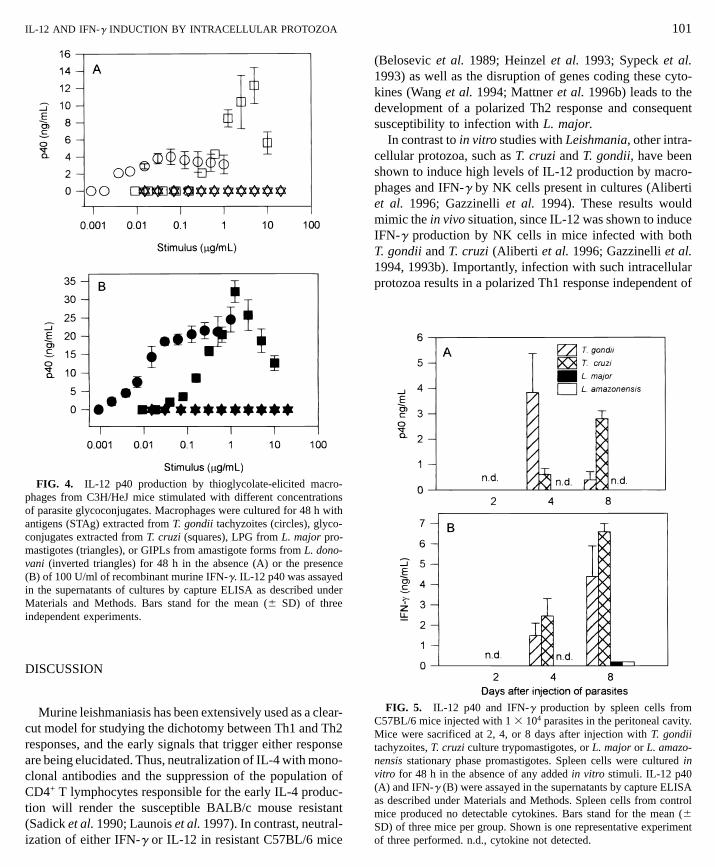
IL-12 AND IFN-g INDUCTION BY INTRACELLULAR PROTOZOA
FIG. 4. IL-12 p40 production by thioglycolate-elicited macro-
phages from C3H/HeJ mice stimulated with different concentrationsof parasite glycoconjugates. Macrophages were cultured for 48 h with antigens (STAg) extracted from T. gondii tachyzoites (circles), glyco-conjugates extracted from T. cruzi (squares), LPG from L. major pro-mastigotes (triangles), or GIPLs from amastigote forms from L. dono-vani (inverted triangles) for 48 h in the absence (A) or the presence(B) of 100 U/ml of recombinant murine IFN-g. IL-12 p40 was assayedin the supernatants of cultures by capture ELISA as described underMaterials and Methods. Bars stand for the mean (6 SD) of threeindependent experiments.DISCUSSION
Murine leishmaniasis has been extensively used as a clear-
cut model for studying the dichotomy between Th1 and Th2responses, and the early signals that trigger either responseare being elucidated. Thus, neutralization of IL-4 with mono-clonal antibodies and the suppression of the population ofCD4+ T lymphocytes responsible for the early IL-4 produc-tion will render the susceptible BALB/c mouse resistant(Sadick et al. 1990; Launois et al. 1997). In contrast, neutral-ization of either IFN-g or IL-12 in resistant C57BL/6 mice101
(Belosevic et al. 1989; Heinzel et al. 1993; Sypeck et al.1993) as well as the disruption of genes coding these cyto-kines (Wang et al. 1994; Mattner et al. 1996b) leads to thedevelopment of a polarized Th2 response and consequentsusceptibility to infection with L. major.
In contrast to in vitro studies with Leishmania, other intra-cellular protozoa, such as T. cruzi and T. gondii, have been
shown to induce high levels of IL-12 production by macro-phages and IFN-g by NK cells present in cultures (Alibertiet al. 1996; Gazzinelli et al. 1994). These results wouldmimic the in vivo situation, since IL-12 was shown to induceIFN-g production by NK cells in mice infected with bothT. gondii and T. cruzi (Aliberti et al. 1996; Gazzinelli et al.1994, 1993b). Importantly, infection with such intracellularprotozoa results in a polarized Th1 response independent ofFIG. 5. IL-12 p40 and IFN-g production by spleen cells fromC57BL/6 mice injected with 1 3 104 parasites in the peritoneal cavity.
Mice were sacrificed at 2, 4, or 8 days after injection with T. gondiitachyzoites, T. cruzi culture trypomastigotes, or L. major or L. amazo-nensis stationary phase promastigotes. Spleen cells were cultured invitro for 48 h in the absence of any added in vitro stimuli. IL-12 p40(A) and IFN-g (B) were assayed in the supernatants by capture ELISAas described under Materials and Methods. Spleen cells from controlmice produced no detectable cytokines. Bars stand for the mean (6SD) of three mice per group. Shown is one representative experimentof three performed. n.d., cytokine not detected.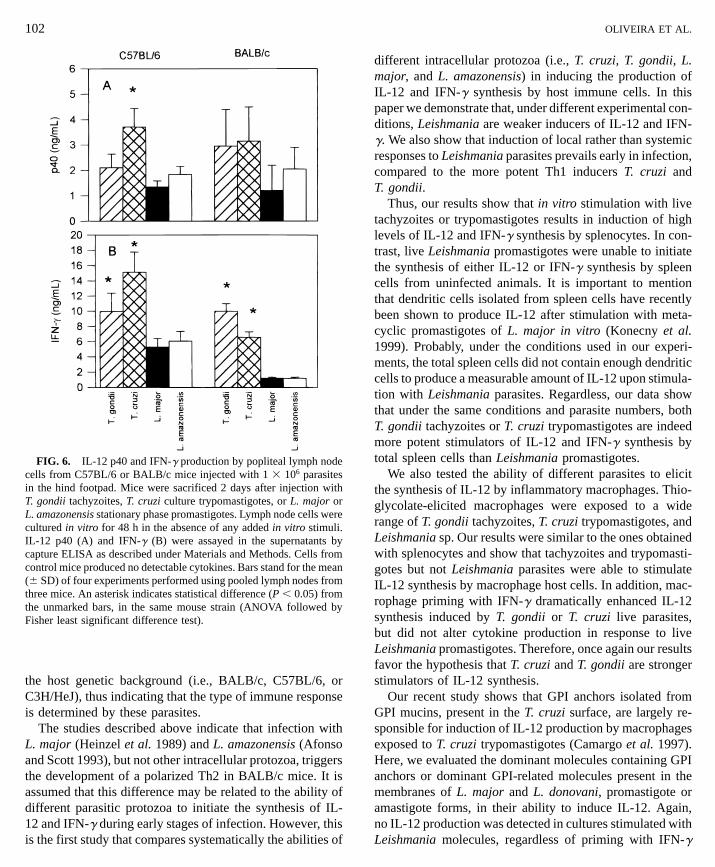
T. gondii tachyzoites, T. cruzi culture trypomastigotes, or L. major orL. amazonensis stationary phase promastigotes. Lymph node cells were
cultured in vitro for 48 h in the absence of any added in vitro stimuli.IL-12 p40 (A) and IFN-g (B) were assayed in the supernatants bycapture ELISA as described under Materials and Methods. Cells fromcontrol mice produced no detectable cytokines. Bars stand for the mean(6 SD) of four experiments performed using pooled lymph nodes fromthree mice. An asterisk indicates statistical difference (P , 0.05) fromthe unmarked bars, in the same mouse strain (ANOVA followed byFisher least significant difference test).the host genetic background (i.e., BALB/c, C57BL/6, orC3H/HeJ), thus indicating that the type of immune responseis determined by these parasites.
The studies described above indicate that infection withL. major (Heinzel et al. 1989) and L. amazonensis (Afonsoand Scott 1993), but not other intracellular protozoa, triggersthe development of a polarized Th2 in BALB/c mice. It isassumed that this difference may be related to the ability ofdifferent parasitic protozoa to initiate the synthesis of IL-12 and IFN-g during early stages of infection. However, thisis the first study that compares systematically the abilities of
OLIVEIRA ET AL.
different intracellular protozoa (i.e., T. cruzi, T. gondii, L.major, and L. amazonensis) in inducing the production ofIL-12 and IFN-g synthesis by host immune cells. In thispaper we demonstrate that, under different experimental con-ditions, Leishmania are weaker inducers of IL-12 and IFN-g. We also show that induction of local rather than systemicresponses to Leishmania parasites prevails early in infection,compared to the more potent Th1 inducers T. cruzi andT. gondii.
Thus, our results show that in vitro stimulation with livetachyzoites or trypomastigotes results in induction of highlevels of IL-12 and IFN-g synthesis by splenocytes. In con-trast, live Leishmania promastigotes were unable to initiatethe synthesis of either IL-12 or IFN-g synthesis by spleencells from uninfected animals. It is important to mentionthat dendritic cells isolated from spleen cells have recentlybeen shown to produce IL-12 after stimulation with meta-cyclic promastigotes of L. major in vitro (Konecny et al.1999). Probably, under the conditions used in our experi-ments, the total spleen cells did not contain enough dendriticcells to produce a measurable amount of IL-12 upon stimula-tion with Leishmania parasites. Regardless, our data showthat under the same conditions and parasite numbers, bothT. gondii tachyzoites or T. cruzi trypomastigotes are indeedmore potent stimulators of IL-12 and IFN-g synthesis bytotal spleen cells than Leishmania promastigotes.
We also tested the ability of different parasites to elicitthe synthesis of IL-12 by inflammatory macrophages. Thio-glycolate-elicited macrophages were exposed to a widerange of T. gondii tachyzoites, T. cruzi trypomastigotes, andLeishmania sp. Our results were similar to the ones obtainedwith splenocytes and show that tachyzoites and trypomasti-gotes but not Leishmania parasites were able to stimulateIL-12 synthesis by macrophage host cells. In addition, mac-rophage priming with IFN-g dramatically enhanced IL-12synthesis induced by T. gondii or T. cruzi live parasites,but did not alter cytokine production in response to liveLeishmania promastigotes. Therefore, once again our resultsfavor the hypothesis that T. cruzi and T. gondii are strongerstimulators of IL-12 synthesis.
Our recent study shows that GPI anchors isolated fromGPI mucins, present in the T. cruzi surface, are largely re-
102
FIG. 6. IL-12 p40 and IFN-g production by popliteal lymph nodecells from C57BL/6 or BALB/c mice injected with 1 3 106 parasitesin the hind footpad. Mice were sacrificed 2 days after injection with
sponsible for induction of IL-12 production by macrophagesexposed to T. cruzi trypomastigotes (Camargo et al. 1997).Here, we evaluated the dominant molecules containing GPIanchors or dominant GPI-related molecules present in themembranes of L. major and L. donovani, promastigote oramastigote forms, in their ability to induce IL-12. Again,no IL-12 production was detected in cultures stimulated withLeishmania molecules, regardless of priming with IFN-g

103
from CAPES, H.C.S. and C.R.L. are Undergraduate Fellows from
IL-12 AND IFN-g INDUCTION BY INTRACELLULAR PROTOZOA
(Fig. 4). In contrast, T. cruzi glycoconjugates induced IL-12 production in a dose-dependent manner. The activityof trypomastigote- and tachyzoite-derived molecules washighly increased by macrophage priming with IFN-g. To-gether, our results are consistent with the hypothesis thatGPI-related molecules derived from Leishmania are poorinducers of IL-12 production by macrophages.
Although we could not detect production of IL-12 stimu-lated by Leishmania in vitro, production of IFN-g by NK(Scharton-Kersten et al. 1995) and later by CD4+ T cells(Scharton-Kersten et al. 1995; Heinzel et al. 1991) is inducedafter infection with this parasite. Here we show that, incontrast to the in vitro studies, all four parasites inducedboth IL-12 and IFN-g production by the draining LN cellsat 48 h postinfection. L. major and L. amazonensis, however,consistently induced lower levels of IL-12 and IFN-g. How-ever, systemic production of IL-12 and IFN-g was foundexclusively upon infection with T. gondii or T. cruzi, asindicated by cytokine synthesis by spleen cells after injectionof parasites in the peritoneal cavity and cytokine measure-ments in the sera from infected animals (data not shown).
The discrepancy between the in vitro and in vivo datarelated to the ability of Leishmania parasites to elicite anIL-12 and IFN-g response is still unknown. It is possiblethat the major cellular source of IL-12 (e.g., neutrophils ordendritic cells) is absent or in low numbers under our invitro conditions. Alternatively, other IL-12-stimulatory fac-tors such as cytokines or cell surface molecules (e.g., CD40ligand), which are required for induction of IL-12 by Leish-mania, may be also absent under our in vitro conditions.Nevertheless, taken together, our data suggest that Leish-mania are poor inducers of IL-12 and IFN-g, compared toT. gondii and T. cruzi. Thus the local stimulus (IL-12 andIFN-g) provided by Leishmania appears not be sufficient to
drive a Th1 response, in a genetic background that biasestoward a Th2 response, and may explain the developmentsoma cruzi trypomastigotes initiate the synthesis of pro-inflammatory
of a susceptible phenotype in BALB/c mice. If, however,the levels of IL-12 are increased either by injection of rIL-12 (Heinzel et al. 1993; Sypeck et al. 1993) or by the effectsof a concomitant infection (Santiago et al. 1999), a Th1response is generated and the switch from susceptibility toresistance is achieved.
ACKNOWLEDGMENTS
This work was in part supported by the PADCT/CNPq (No. 62.0106/95-6-SBIO). R.T.G. is the recipient of a Biotechnology Career Fellow-ship from the Rockefeller Foundation. M.A.P.O. is a Graduate Fellow
FAPEMIG, and L.Q.V. and R.T.G. are Research Fellows from CNPq.The authors are indebted to Drs. David Sacks, Helia Cannizarro, JayFarrell, Maria Wysocka, Phillip Scott, and Salvatore Turco for supply-ing several reagents, to Ms. Soraya Oliveira Silva for keeping theseveral Leishmania strains, and to Dr. Luis Carlos Crocco Afonso forcritical review of the manuscript.
REFERENCES
Afonso, L. C. C., and Scott, P. 1993. Immune responses associated withthe susceptibility of C57BL/10 mice to Leishmania amazonensis.Infection and Immunity 61, 2952–2959.
Aliberti, J. C. S., Cardoso, M. A. G., Martins, G. A., Gazzinelli, R.T., Vieira, L. Q., and Silva, J. S. 1996. Interleukin-12 mediatesresistance to Trypanosoma cruzi in mice and is produced by murinemacrophages in response to live trypomastigotes. Infection and Im-munity 64, 1961–1967.
Almeida, I. C., Ferguson, M. A. I., Schenckman, S., and Travassos,L. R. 1994. Lytic anti-a-galactosyl antibodies from patients withchronic Chagas’ disease recognize novel O-linked oligosaccharideson mucin-like glycosilphosphatidylinositol-anchored glycoproteinsof Trypanosoma cruzi. Biochemical Journal 304, 793–802
Andrade, Z. 1991. Pathogenesis of Chagas’ disease. Research in Immu-nology 142, 130–133.
Belosevic, M., Finbloom, D. S., van der Meid, P. H., Slayter, M. V.,and Nacy, C. A. 1989. Administration of monoclonal anti-IFN-gamma antibodies in vivo abrogates natural resistance of C3H/HeNmice to infection with Leishmania major. Journal of Immunology143, 266–274.
Brener, Z., and Gazzinelli, R. T. 1997. Immunological control of Trypa-nosoma cruzi infection and pathogenesis of Chagas’ disease. Interna-tional Archives of Allergy and Applied Immunology 114, 103–110.
Burleigh, B. A., and Andrews, N. W. 1995. The mechanisms of Trypa-nosoma cruzi invasion of mammalian cells. Annual Review of Micro-biology. 49, 175–200.
Camargo, M. M., Almeida, I. C., Pereira, M. E., Ferguson, M. A.,Travassos, L. R., and Gazzinelli, R. T. 1997 Glycosylphosphatidyli-nositol-anchored mucin-like glycoproteins isolated from Trypano-
cytokines by macrophages. Journal of Immunology 158, 5890–5901.
Carrera, L., Gazzinelli, R. T., Badolato, R., Hieny, S., Muller, W.,Kuhn, R., and Sacks, D. L. 1996. Leishmania promastigotes selec-tively inhibit interleukin 12 induction in bone marrow-derived mac-rophages from susceptible and resistant mice. Journal of Experimen-tal Medicine 183, 515–526.
Frenkel, K. 1988. Pathophysiology of toxoplasmosis. ParasitologyToday 4, 273–278.

104
Gazzinelli, R. T., Hakim, F. T., Hieny, S., Shearer, G. M., and Sher,A. 1991. Synergistic role of CD41 and CD81 T lymphocytesin IFN-gamma production and protective immunity induced by anattenuated Toxoplasma gondii. Journal of Immunology 146, 286–292.
Gazzinelli, R. T., Xu, Y., Hieny, S., Cheever, A., and Sher, A. 1992.Simultaneously depletion of CD41 and CD81 T lymphocytes isrequired to reactivate chronic infection with Toxoplasma gondii.Journal of Immunology 149, 175–180.
Gazzinelli, R. T., Denkers E. Y,. and Sher, A. 1993a. Host resistanceto Toxoplasma gondii: A model for studying the selective inductionof cell mediated immunity by intracellular parasites. InfectiousAgents and Disease 2, 139–149.
Gazzinelli, R. T., Hieny, S., Wynn, T. A., Wolf, S., and Sher, A.1993b. Interleukin-12 is required for the T-lymphocyte-independentinduction of interferon-gamma by an intracellular parasite and in-duces resistance in T-cell-deficient hosts. Proceedings of the NationalAcademy of Sciences USA 90, 6115–6119.
Gazzinelli, R. T., Wysocka, M., Hayashi, S., Denkers, E. Y., Hieny,S., Caspar, P., Trinchieri, G., and Sher, A. 1994. Parasite-inducedIL-12 stimulates early IFN-gamma synthesis and resistance duringacute infection with Toxoplasma gondii. Journal of Immunology153, 2533–2543.
Heinzel, F. P., Sadick, M. D., Holaday B. J., Coffman, R. L.; Locksley,R. M. 1989. Reciprocal expression of interferon gamma or interleu-kin 4 during the resolution or progression of murine leishmaniasis.Evidence for expansion of distinct helper T cell subsets. Journal ofExperimental Medicine 169, 59–72.
Heinzel, F. P., Sadick, M. D., Mutha, S. S., and Locksley R. M. 1991.Production of IFN-g, interleukin-2, interleukin-4 and interleukin-10by CD41 lymphocytes in vivo during healing and progressive murineleishmaniasis. Proceedings of the National Academy of SciencesUSA 88, 7011–1715.
Heinzel, F. P., Schoenhaut, D. S., Rerko, R. M., Rosser, L. E., andGately, M. K. 1993. Recombinant Interleukin-12 cures mice infectedwith Leishmania major. Journal of Experimental Medicine 177,1505–1509.
Johnstone, A., and Thorpe, R. 1982. In “Immunochemistry in Practice”(A. Johnstone and R. Thorpe, Eds.), p. 77. Blackwell Scientific,Oxford, UK.
Konecny, P., Stagg, A. J., Heather, J. English, N., Davidson, R. N.,and Knight, C. C. 1999. Murine dendritic cells internalize Leish-mania major promastigote produce IL-12 p40 and stimulate primaryT cell proliferation in vitro. European Journal of Immunology 29,1803–1811.
Launois, P., Maillard, I., Pingel, S., Swihart, K. G., Xenarios, I., Acha-Orbea, H., Diggelmann, H., Locksley, R. M., MacDonald, H. R.,and Louis, J. A. 1997. IL-4 rapidly produced by Vb4 Va8 CD41T cells instructs Th2 development and susceptibility to Leishmaniamajor in BALB/c mice. Immunity 6, 541–549.
Lowry, O. H., Rosenbrough, N. J., Farr, A. L., and Randall, R. J.1951. Protein measurement with the Folin phenol reagent. Journalof Biological Chemistry 193, 265–275.
OLIVEIRA ET AL.
Mattner, F., Alber, G., Magram, J., and Kopf, M. 1996a. The role ofIL-12 and IL-4 in Leishmania major infection. Chemical Immunol-ogy 68, 86–109.
Mattner, F., Magram, J., Ferrante, J., Launois, P., Di Padova, K., Behin,R., Gately, M. K., Louis, J. A., and Alber, G. 1996b. Geneticallyresistant mice lacking interleukin-12 are susceptible to infectionwith Leishmania major and mount a polarized Th2 cell response.European Journal of Immunology 26, 1553–1559.
McNeely, T. B., Rosen, G., Londner, M. V., and Truco, S. J. 1989.Inhibitory effects on protein kinase C activity by lipophosphoglycanand glycosyl-phosphatidylinositol antigens of the protozoa parasiteLeishmania. Biochemical Journal 259, 601–604.
Reiner, S. L., Zheng, S., Wang, Z. E., Stowinring, L., and Locksley,R. M. 1994. Leishmania promastigotes evade interleukin-12 (IL-12)induction by macrophages and stimulate a broad range of cytokinesfrom CD41 T cells during initiation of infection. Journal of Experi-mental Medicine 179, 447–456.
Russell, D. G., and Talamas-Rohana, P. 1989. Leishmania and themacrophage: a marriage of inconvenience. Immunology Today 10,328–333.
Sadick, M. D., Heinzel, F. P., Holaday, B. J., Pu, R. T., Dawkins, R.S., and Lockslay, R. M. 1990. Cure of murine leishmaniasis with anti-interleukin 4 monoclonal antibody. Evidence for a T cell-dependent,interferon g-independent mechanism. Journal of Experimental Medi-cine 171, 115–127.
Santiago, H. C., Oliveira, M. A. P., Bambirra, E. A., Faria, A. M. C.,Afonso, L. C. C., Vieira, L. Q., and Gazzinelli, R. T. 1999. Coinfec-tion with Toxoplasma gondii inhibits antigen-specific Th2 immuneresponses, tissue inflammation, and parasitism in BALB/c miceinfected with Leishmania major. Infection and Immunity 67, 4939–4944.
Scharton-Kersten, T., Afonso, L. C. C., Wysocka, M., Trinchieri, G.,and Scott, P. 1995. IL-12 is required for natural killer cell activationand subsequent T helper 1 cell development in experimental leishma-niasis. Journal of Immunology 154, 5320–5330
Serrano, A. A., Schenkman, S., Yoshida, N., Mehlert, A., Richardson,J. M., and Ferguson, M. A. 1995. The lipid structure of the glycosyl-phosphatidylisonitol-anchored mucin-like sialic acid acceptors ofTrypanosoma cruzi changes during parasite differentiation from epi-mastigote to infective metacyclic trypomastigote forms. JournalBiological. Chemistry 270, 27244–27253
Silva, J. C., Pirmez, C., Morgado, M. G., and Galvao-Castro, B. 1985.Immunopathological aspects of experimental Trypanosoma cruziinfection: Correlation of immunocomplexes and other serologicalfeatures with muscle lesions during the infection. Parasite Immunol-ogy 7, 457–466.
Silva, J. S., Morrissey, P. J., Grabstein, K. H., Mohler, K. M., Anderson,D., and Reed, S. G. 1992. Interleukin 10 and interferon-g regulationof experimental Trypanosoma cruzi infection. Journal of Experimen-tal Medicine 175, 169–174.
Sypeck, J. P., Chung, C. L., Mayor, S. E. H., Subramanyam, J. M.,Goldman, S. J., Sieburth, D. S., Wolf, S. F., and Shcaub R. G. 1993.Resolution of cutaneous leishmaniasis: Interleukin-12 initiates a pro-tective T helper type1 immune response. Journal of ExperimentalMedicine 177, 1797–1802.

IL-12 AND IFN-g INDUCTION BY INTRACELLULAR PROTOZOA
Vieira, L. Q., Hondowicz, B. D., Afonso, L. C. C., Wysocka, M.,Trinchieri G., and Scott, P. 1994 Infection with Leishmania majorinduces interleukin-12 production in vivo. Immunology Letters 40,157–161.
Wang, Z. E., Reiner, S. L., Zheng, S., Dalton, D. K., and Locksley, R. M.
105
1994. CD41 effector cells default to the Th2 pathway in interferon-gamma-deficient mice infected with Leishmania major. Journal ofExperimental Medicine 179, 1367–1371.
Received 4 August 1999; accepted with revision 28 April 2000
Related Documents











