Chapitre 4 Le peuplement végétal cultivé Bruno ANDRIEU, Jérémie LECŒUR, Gilles LEMAIRE, Bertrand NEY Les modes de représentation du peuplement végétal cultivé sont nombreux ct peuvent être classés par le degré d'explicitation des mécanismes impliqués ct par le point de vue théorique de l'observateur. Il peut s'agir d'une représentation ''énergétique», oli le peuplement végét a l est as si mil é ù une surface d'échange thermodynamique avec le milieu qui l'entoure. Cette vision, qualifiée par les Anglo- Saxons de représentation « hig le » et propre ù prédire la croissance en biomasse, est très largement utilisée en raison de sa robustesse dans les modèles de culture actuels. Cette démarche est souvent accompagnée de règles empiri q u es de partage des assimilats po estimer le rendement. Elle est en revanche largement insuffi- sante pour caractériser l'état i nstantané de satisfaction des besoins de la culture pour les différents factes de sa croissance (cau, éléments minéraux ... ). De même, les règles empiriques de ptage des assimilats (entre parties aériennes ct souter- raines ou entre parties végé tatives ct reproductrices) sont très souvent insuffisantes, notamment lors de contraintes environnementales abiotiques. Ces b esoins d'expli- citation nécessitent une approche fonctionnelle du peuplement végétal. Dans ces deux cas, le peuplement végétal est une collection d'individus moyens, feuilles dans le premier, plantes ans le second. Une représentation récente du peup lement végétal en tant que somme d'individus différenciés en compétition ct en relation sociale a été produite grâce à des formalismes nouveaux. Cette représentation « architecturale » relie structure et fonctions par l'intermédiaire elu microphyto- climat. La structure elu peuplement modifie le phytoclimat local qui influence les fonctions de la pl a nte ct la croissance de ses organes. �� L'approche thermodynamique et La production de biomasse À l'origine, l'écophysiologie végétale s'est développée à partir d'une approche des échanges de matière et d'énergie entre le peuplement végétal et son environnement 103

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chapitre 4
Le peuplement végétal cultivé Bruno ANDRIEU, Jérémie LECŒUR,
Gilles LEMAIRE, Bertrand NEY
Les modes de représentation du pe uplement végétal cultivé sont nombreux ct
peuvent être classés par leur degré d'explicitation des mécanismes impliqués ct par le point de vue théorique de l'observateur. Il peut s'agir d'une représentation ''énergétique», oli le peuplement végéta l est assi milé ù une surface d'échange thermodynamique avec le milieu qui l'entoure. Cette vision, qualifiée par les AngloSaxons de représentation « hig leaf » et propre ù prédire la croissance en biomasse, est très largement utilisée en raison de sa robustesse dans les modèles de culture actuels. Cette démarche est souvent accompagnée de règles empiriq u es de partage des assimilats pour estimer le rendement. Elle est en revanche largement insuffisante pour caractériser l'état instantané de satisfaction des besoins de la culture pour les différents facteurs de sa croissance (cau, éléments minéraux ... ). De même, les règles empiriques de partage des assimilats (entre parties aériennes ct souterraines ou entre parties végétatives ct reproductrices) sont très souvent insuffisantes, notamment lors de contraintes environnementales abiotiques. Ces besoins d'explicitation nécessitent une approche fonctionnelle du peuplement végétal. Dans ces
deux cas, le peuplement végétal est une collection d'individus moyens, feuilles dans le premier, plantes clans le second. Une représentation récente du pe up lement végétal en tant que somme d'individus différenciés en compétition ct en relation sociale a été produite grâce à des formalismes nouveaux. Cette représentation « architecturale » relie structure et fonctions par l'intermédiaire elu microphytoclimat. La structure elu peuplement modifie le phytoclimat local qui influence les fonctions de la pla nte ct la croissance de ses organes.
�� L'approche thermodynamique et La production de biomasse À l'origine, l'écophysiologie végétale s'est développée à partir d'une approche des échanges de matière et d'énergie entre le peuplement végétal et son environnement
103
andrieu
Texte tapé à la machine
andrieu
Texte tapé à la machine
Cite as : Andrieu, B., Lecoeur, J., Lemaire, G., Ney, B. (2006). Le peuplement végétal cultivé. In T. Doré, M. Le Bail ,P. Martin, B. Ney, J. Roger-Estrade (Coord.) L'agronomie aujourd'hui (pp. 103-136). Versailles: Editions Quae
andrieu
Texte tapé à la machine
andrieu
Texte tapé à la machine

Analyse et modélisation du peuplement végétal cultivé
physique. L'objectif i nit ial était de pouvoir estimer la productivité pr imaire des peuplements végétaux, c'est-à-elire l 'accumulation de biomasse, et la contribution de ceux-ci aux échanges hydriques avec l 'atmosphère, c'est-à-dire l 'évapotranspiration. Pour ce la , il importait de pouvoi r établ ir un bi lan net des échanges d'énergie ct de matière entre l e peuplement ct son environnement externe en considérant le continuum sol-végétation-atmosphère. La source d'énergie principale étant le rayonnement so laire , il importait avant tout de caractériser ct de quantifier l 'absorption de cette énergie par le peuplement végétal pou r ses deux composantes : - le rayonnement global (RJ déterminant en grande partie la demande évaporatoire par sa fract ion transm['ic au sol ct la demande transpi ra toirc par sa fraction absorbée par la végétat ion :
-le rayonnement visible act if pour la photosynthèse (Par, plzo.\yntlzeticullr uctin' mdiution) dont la fraction absorbée par la végétation déterm ine la photosynthèse brute du couvert végétal, c'est-à-dire sa capacité ù capter ct ù ass imi ler le co_, a tmosphériquc.
L'établ issement d'un tel bi lan rad iat if du peuplement végétal pouvait être réa l i sé expérimenta lement par l 'u t i l isation de capteurs appropriés. disposés au-dessus de la végétation ct au niveau du sol afin de mesur e r les di fférentes composantes du bi lan (fïgure 2.1): le rayonnement incident ( R11) di rect ct d iffus. le rayonnement transmis au sol (R1) ct sa part réfléchie vers la végétation (R1,), enfin le rayonnement réfléchi vers l 'atmosphère (R), afin d'en déduire le rayonnement intercepté par la végétation (Rint' exprimé en MJ/m-') :
Rint = Ro- Rt ( 1)
ou le rayonnement absorbé (R"11J par la végétation :
R"'" = R11 + R1,- R1- Rr (2)
soit pour le rayonnement global R., soit pour sa composante visible, le Par (VarlctGrancher ct Bonhomme, 1 974).
� La modélisation de ces b i l ans a nécessité
R incident (R0)
R refléchi P" lo "' (R,) �"'mi' ao 'oi(RJ ./Rtn Couvert végétal
Sol
Figm·e 2.1. Représentation du bilan radiatif d'un peuplement végétal, soit pour le rayonnement global Rg, soit pour sa composante <<visible>> utile à la photosynthèse.
104
Le peuplement végétal cultivé
d'ass imi ler le peuplement végétal par analogie ù un m il ieu poreux diffusant homogène, ce qui permet d'appl iquer la loi de Beer-Lambert (Monsi et Saek i , 1 953) ct d 'estimer ainsi les composantes du b i lan ù partir d 'une caractérisation des propriétés géométriques du couvert végétal : dens ité des surfaces fol ia i res, orientation angulaire verticale des feu i l les ct propriétés optiques des surfaces fo l ia ires (réflcctancc, absorbancc ct transmittancc) vis-ù-vis des gammes de longueur d 'onde considérées R� ou Par (Bonhomme ct Varlct-Granchcr, 1977).
Flux de carbone entre le peuplement végétal et l'atmosphère
L'estimation ct la mesure des composantes du bi lan rad iat if dans le domaine visib le (Par) ont permis l e déve loppement d 'une approche de la photosynthèse d 'un couvert végéta l , c 'est-à-dire du fl ux de CO, assim i lé par la végétation. Pour ce la, le couvert végétal est décomposé en tranc!Ïcs horizontales de densité de su rface fol ia i re homogène au sein desque l les peuvent être estimées les composantes du bi lan radiati f Par. À ces surfaces foliaires considérées globa lement (hig lm!) est app l iqué un modèle de réponse de l ' unité de surface fol ia ire ù l 'in tensité d 'éclai rement. Cc modè le permet de prendre en compte les différents paramètres de la di ffusion du co_, (résistance stomat iquc. résistance mésophylicnnc. concentration interne. etc. ). ct des processus biochimiques de l 'ass imi lation du carbone : clïïcicncc quantique , activité de la rubisco, etc. (Chartier, 1 %6: Farquhar et ul., llJ�O). La photosynthèse du peuplement végétal est alors calcu lée par intégration des couches successives sous l 'hypothèse d'homogéné ité horizonta le du couvert végétal (!ïgure 2.2). Les pertes resp i ratoi res de CO, du couvert végétal sont estimées par l 'ordonnée ù l 'origine de la courbe de rép()nsc de la feu i l l e ù l 'éclairement ; e l les sont clone i ntégrées également sur l 'ensemble du couvert végétal , cc qui permet d'estimer la photosynthèse nette du couvert végétal, c'est-à-d i re l e bi lan des échanges de CO:! entre le couvert végétal ct l'atmosphère. Cette approche comporte cependant une incertitude importante, qu i est de ne pas prendre en compte des pertes de C02 elues ù la respiration racina i rc . En effet, i l est très d iffic i le de d iscrim iner expérimentalement, clans le flux de CO, en provenance du sol, la part de la respiration des racines de celle des autres conÏposantes b io logiques du sol. L'uti l i sation d 'un témoin << sol nu » n 'est qu 'un pis-al ler, car l a présence des racines clans un sol et leur activité s 'accompagne d 'une pro l i fération de m icro-organismes rhizosphériques qu i contribuent à la respi ration propre du sol. Seules des techniques de traçage isotopique permettent d'appréhender ce problème (Nguycn et al., 1 999). Pour un couvert de fétuque élevée, Bélanger et al. ( 1994) ont montré que la respiration ct l 'exsudation racina ires pouvaient correspondre ensemble à une perte de carbone équivalant à 20 % de l a photosynthèse brute.
Il est possible d 'établi r expérimentalement u ne courbe de réponse de la photosynthèse brute (Pb, en !lillO! C02 · m-2
· s- 1 ) d 'un couvert végétal à l'éclairement (Gastal et Bélanger, 1993) :
Pb= Pmax [1 - exp (- C Par)] (3)
avec Pmax = photosynthèse b rute maximale à Par saturant, et c = coefficient d 'ajustement. La pente in it iale de la courbe égale à (Pmaxc) représente l 'efficience quantique (L Y max), ou rendement l umineux. De p lus, i l est possible de déterminer ces
105

Analyse et modélisation du peuplement végétal cultivé
a)
b)
c)
Photosynthèse nette de la feuille (�tmol co2 m- 2. s- 1)
pmax
Éclairement R0 (pmol photons m- 2 . s- 1)
Photosynthèse nette du couvert (�tmol co2 m- 2. s- 1) p ma<
P du couvert (��;;ol co2 m- 2 . s- 1)
Éclairement incident (,tt mol photons m-2. s- 1)
Indice foliaire Figure 2.2. (a) Courbe de réponse de la photosynthèse de la feuille à l'éclairement. P"'"' : photosyn
thèse nette maximale à éclairement saturant ; E0: éclairement au point de compensation; R0: respirat ion obscure ; a : efficience lumineuse. (b) Courbe de réponse de la photosynthèse nette elu couvert végétal à l'éclairemen t incident. (c)' Réponse elu P"'"' d'un peuplement à l'évolu tion de l'indice foliaire.
courbes de réponse de Pmax au cours du développement du couvert végétal en fonction de son indice foliaire- voir figure 2.2 (c). On observe une augmentation moins que proportionnelle de la photosynthèse maximale avec l'indice foliaire. La même démarche entreprise sur L Y (Gastal et Bélanger, 1993) indique également max une augmentation moins que proportionnelle de l'efficience quantique avec le développement de l'indice foliaire. Cette démarche constitue un changement d'échelle depuis celle de la feuille, et même de l'organite (chloroplaste, stomate), à celui du couvert végétal (population de feuilles considérée comme une surface d'échange), directement, sans passer par l'échelle de l'organisme entier qui est celui
106
Le peuplement végétal cultivé
de la plante ind ividuel le . Cette approche <<saute >> un n iveau d 'organ isation (la plante) ct ne permet pas d'expliciter certa ines régu lations de la photosynthèse par les re l ations sources-puits au sein de la plante imlividucl lc. comme nous le verrons. Montci th ( 1977) c t Varlct-Granchcr cl Bonhomme ( 1974) ont déve loppé une approche permettant de décomposer l 'ef ficience de conversion de l 'énergie solaire en biomasse par un peuplement végétal (f) en : -une efficience cl imatique (�::J correspondant ill! rapport entre l 'énergie incidenlL· d<�ns le spectre visible (Par) ct l 'énergie sol<�irc incidente totale (RJ: - une ellicicncc d 'absorption (r,) correspondant <Ill r<�pporl en tre le P�tr :th�urlx; pc�; le couvert végét<�l ct le P<�r incident : -une cllicicncc biologique (Eh) correspondant i!U rapport ck l'c;ncrgic conte nul: dans la biomasse élaborée (MS ) ct le Par <1bsorbé journa l i e r (Par) : l . .t
soit :
E = (C·MS i ) / R�i (4) (:'i)
(C · MS ) / R = E · E · r J !-!.1 l' (! h (6)
avec C = capacité calor ifique de la biomasse ct Rci = rayonnement solaire global journa l ier.
·
Avec:
d 'où :
E · E · R. =Par c a /:!.J a
(C · MS i) = Eh Par"
(MS i) = (Eh / C) Para
(7)
(S )
(9).
E 1 C a la dimension d'une biomasse produite par unité d 'énergie absorbée par le c�uvcrt végétal (kg MS · nr" · MJ-1) et est appelé communément en anglais « radiation use efficiency >> (RUE).
Cette approche a été largement utilisée sur de nombreuses espèces cultivées (Monteith, 1977; Gosse et al., 1986) . La distinction entre les processus de captation de l'énergie solaire ( E,.) et les processus de conversion de celle-ci en biomasse végétale (Et,) a fourni un cadre conceptuel pour l'analyse des effets du milieu et du génotype sur la croissance en biomasse des peuplements végétaux. La surface foliaire développée par le couvert végétal par unité de surface, c'est-à-dire l'indice foliaire (IF), ou « leaf are index» (LAI) en anglais, est la variable clef pour quantifier les phénomènes d'absorption ou d'interception. Pour un couvert homogène sur le plan horizontal, l'efficience d'absorption (e,.) augmente de façon asymptotique avec l'indice foliaire J·usqu'à une valeur maximale (E. . ) correspondant à l'extinc-d, 111dX tian totale du rayonnement transmis au sol et dépendant des propriétés optiques des feuilles (albédo du couvert végétal dans le Par). On peut alors relier Ea à l'indice foliaire par la relation empirique suivante (figure 2.3) :
E = E ( 1 - e-K. IF) (lü) a a, max avec K (coefficient d'extinction de la lumière), paramètre empirique intégrant la structure du couvert végétal (port dressé vs port horizontal des feuilles), la hauteur du soleil au-dessus de l'horizon, et le rapport diffus/direct dans le rayonnement Par incident. Lorsque Ea est intégré sur une journée, et a fortiori sur une période plus
107
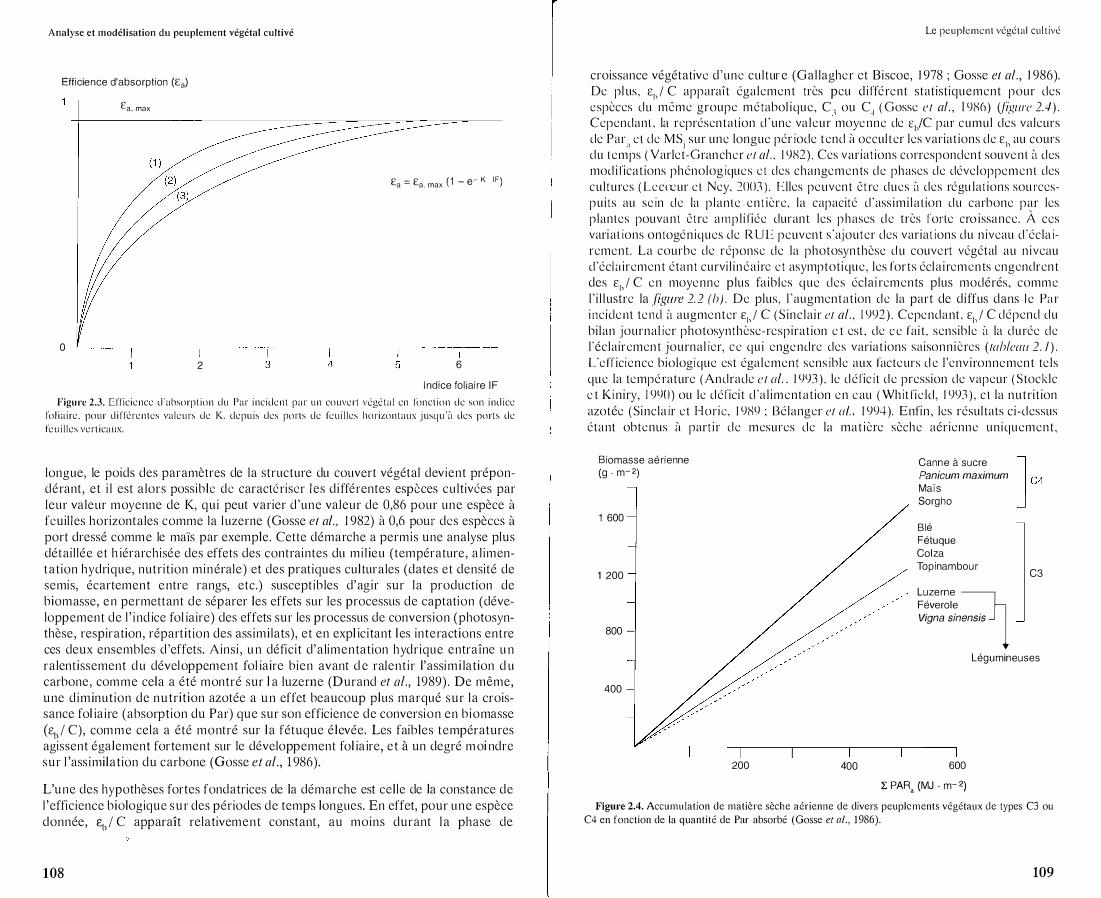
Analyse et modélisation du peuplement végétal cultivé
Efficience d'absorption (êa)
Ea, max
0 1 2 3 4
êa = êa. max ( 1 - e- K IF)
5 6
Indice foliaire IF Figure 2.3. Efficience d'absorption du Par incident par un couvert végétal en fonction de son indice
foliaire. pour différentes valeurs de K. depuis des ports de feuilles horizontaux jusqu'il des ports de
feuilles verticaux.
longue, le poids des paramètres de la structure du couvert végétal devient prépondérant, et i l est alors possible de caractériser les d ifférentes espèces cultivées par leur valeur moyenne de K, qu i peut varier d 'une valeur de 0,86 pour une espèce ù feuilles horizontales comme la l uzerne (Gosse et al., 1982) à 0,6 pour des espèces ù port dressé comme le maïs par exemple. Cette démarche a permis une analyse plus détai l lée et h iérarchisée des effets des contraintes du mi l ieu (température, a l imentation hydrique, nutr it ion minérale) et des pratiques culturales (dates et densité de semis, écartement entre rangs, etc.) susceptibles d'agir sur la production de biomasse, en permettant de séparer les effets sur les processus de captation ( développement de l ' i nd ice fol iaire) des effets sur les processus de convers ion (photosynthèse, respiration, répartit ion des ass imilats), et en expl ic itant l es interactions entre ces deux ensembles d 'effets. Ainsi, u n déficit d'alimentation hydrique entraîne u n ralentissement du développement fol iaire bien avant d e ralent ir l'assim i lation d u carbone, comme cela a été montré sur l a luzerne (Durand et al., 1989) . De même, une d iminut ion de nutrit ion azotée a un effet beaucoup plus marqué sur la croissance fol ia ire (absorption du Par) que sur son efficience de conversion en b iomasse (sb/ C), comme cela a été montré sur la fétuque élevée. Les faibles températures agissent également fortement sur le développement fol ia ire, et à un degré moi ndre sur l 'ass imilation du carbone (Gosse et al., 1 986).
L'une des hypothèses fortes fondatrices de la démarche est celle de la constance de l 'efficience biologique sur des périodes de temps longues. En effet, pour une espèce donnée, ëb/ C apparaît relativement constant, au moins durant la phase de
108
r Le peuplement végétal cul t ivé
croissance végétative d 'une cultur e (Gal laghcr ct Biscoe, 1978 ; Gosse et al., 1986). De pl us, ëh / C apparaît également très peu d ifférent statistiquement pour des espèces du même groupe métabol ique, C1 ou C-l (Gosse et al., l lJ86) (/ïgure 2.4). Cependant, la représentat ion d'une va leur moyenne de ë1/C par cumul des valeurs de Par" ct de MSi sur une longue période tend ù occulter les variations de Eh au cours du temps (Varlet-Granchcr et al . . Jl)82). Ces variations correspondent souvent ù des mod i fications phénologiqucs ct des changements de phases de développement des cultures (Lccœur ct Ney. 2003). Elles peuvent être ducs ù des régu lat ions sourcespui ts au sein de la plante en t ière, la capacité d'ass imi la t ion du carbone par les plantes pouvant être ampl i f iée durant les phases de très forte cro issance. À ces variations ontogén iques de RUE peuvent s'ajouter des variations du n iveau d 'éclairement. La courbe de réponse de la photosynthèse du couvert végétal au n iveau d'éclairement étant curvi l inéa irc ct asymptotique, les forts écla i rements engendrent des Eh / C en moyenne plus fa ibles que des éclai rements plus modérés, comme l ' i l lustre la .fïgure 2.2 (h). De plus , l'augmentation de la part de diffus dans le Par incident tend ù augmenter Eh / C (Sincla ir et al., Jl)l)2). Cependant, Eh / C dépend du bi lan journalier photosynthèse-respiration ct est, de c c fait. sensible ù la durée de l 'éclairement journalier, cc qu i engendre des variations saisonnières (tah!eau 2.1 ) . L'efficience biologique est également sensible aux facteurs d e l 'environnement te ls que la température (Andrade et al .. Jl)l)J). lc déficit de pression de vapeur (Stocklc ct Ki niry, Jl)l)()) ou le déficit d 'a l imentation en cau (Whitlïcld, Jl)l)3 ), c t la nutrition azotée (Sinclair ct Horic, Jl)8l) ; Bélanger et rd., Jl)l)4 ). Enfin , les résultats ci-dessus étant obtenus ù part i r de mesures de la matière sèche aérienne uniquement,
Biomasse aérienne (g . m-2)
1 600
1 200
800-
400-
200 400
Canne à sucre Panicum maximum Maïs Sorgho
Blé Fétuque Colza Topinambour
Féverole Vigna sinensis
Luzerne 4 C3
Légumineuses
600
:E PAR. (MJ . m-2) Figure 2.4. Accumulation de matière sèche aérienne de divers peuplements végétaux de types C3 ou
C4 en fonction de la quantité de Par absorbé (Gosse et al., 1986).
109

Analyse et modélisation du peuplement végétal cultivé
l 'estimation des E11 1 C qui en découle prend en compte également l 'effet direct du m i l ieu sur la répartition de la matière sèche entre les racines ct les parties aériennes. Ai nsi, Bélanger et al. ( 1994) ont pu montrer que le principal effet de la nutrition azotée était bien une modification du rapport biomasse aérienne/biomasse racina irc, ct que l 'efficience biologique pour la biomasse totale n 'était que fa iblement affectée (/ah/eau 2.1). Khaity ct Lemaire ( 1992) ont montré sur la luzerne que E11/ C calculé sur la biomasse tota le (aérien + souterra in ) était beaucoup plus stable entre saisons que celui ca lculé uniquement ù partir de la biomasse aérienne. Cc type de données est cependant assez rare du fa i t de la difficulté de mesure de la biomasse racm<urc .
Tableau 2.1. Effet de la saison de croissance ct de la nutrition azotée sur l 'effi cience de conversion du Par intercepté ( RU E) par un couvert de fétuque élevée (Fetuca arrundinacea) exprimé soit en biomasse aérienne ( R U Ea), so i t en b iomasse totale ( R U Et), soit en photosynthèse nette (RUEpn). soit en photosynthèse brute ( RU Epb ). Source : Bélanger et al .. 1994.
Saison N RUEa (kg· ha-1) (g MS· MJ·1)
]�té {) 0/llJ
1){) l.:lK
240 I.KO
Automne () 0.69
40 LlO
160 l.hK
Printcmp' {) 1.12
(J() 1.73
120 2,07
IKO 2,21
RUEt (g MS· MJ-1)
1.()7
1.97
2,J5
!.OK
1,74
2J2
1.7K
2,2K
2.56
2.65
RUEpn (g MS· MJ-1)
I.Kh
, , , .1,.1.)
:1,47
2.2:'\
2.:'\ 1
2.49
3.17
3,29
3,34
3.K3
RUEpb (g MS· MJ-1)
:u.:�
4.:'\7
:'\.26
:'\,91
5,0.3
:i,K3
4X2
5,()3
:'\,05
5.62
La constance du paramètre C (capacité calorifique de la biomasse) peut également être remise en cause ù certai nes phases du développement sur certaines espèces. Si une valeur commune ct constante de 16 ù 1 g · 103 J · g-1 MS (Varlet-Grancher et al., 1982) peut être admise pour les phases de croissance végétative lorsque l a p lante élabore essentiel lement de la cel lulose et de la l ign i ne ct pour les espèces qui accumulent essentiel lement de l 'amidon clans leurs gra ines, des valeurs plus é levées peuvent être obtenues lors elu rempl issage des grai nes riches en protéines ou en l ipides pour l e s oléoprotéagineux (20 à 22 · 103 J · g-1 MS) . Ces variations de l a valeur de C doivent être prises en compte clans l e s calculs de Eh 1 C ct clans les comparaisons entre espèces.
L'approche énergétique de la conversion de l 'énergie solaire en biomasse par un peup lement végétal a permis d'aborder de m anière plus fonctionnel le l 'effet des facteurs elu m i lieu et des facteurs génétiques sur l a croissance en masse des peuplements végétaux. I l s'agit donc d 'une approche intégrative qui est réalisée à l 'échelle d'un peuplement. Rien a priori n'empêche un changement d'échelle au n iveau de la
110
r Le peuplement végétal cultivé
plante individuel le, isolée ou en peuplement, si cc n'est la difficulté qu' i l y a alors à estimer le rayonnement absorbé par la plante ind ividuelle, cc qui requiert a lors une approche de sa structure en tro is d imensions. Comme nous l'avons vu, la limite de cette approche réside davantage dans le fa it qu'elle ne s'applique en toute rigueur qu'ù l 'accumulation de biomasse totale. ct qu'el le ne permet en aucune manière de considérer la répart ition de la biomasse entre les différents compartiments morphologiques des plantes- feuil les. t iges. racines. gra ines ou fruits ou tubercules. I l s'agit lù d 'un handicap sérieux pour les approches ù caractère agronomique qui visent ù estimer ou ù simuler la biomasse récoltab lc . Néanmoins. même pour une approche stricte d 'écophysiologic visant ù s imuler la productivité prima ire d'un peuplement végétal. le prob lème de la réparti t ion des assimilats au sein d'une plante reste fondamenta l . En effet, clans cette approche, l'al location des assim ilats pour la croissance fol ia ire, ct donc la dynamique du développement de l'ind ice fol ia i re. est primordiale puisqu'elle détermine la quantité de rayonnement absorbé par le couvert végétal .
L'eau et le peuplement végétal:
flux et efficience pour la production de biomasse
L'cau est un facteur essentiel ù la production végétale. El le a p lusieurs fonctions au sein de la plante (Joncs, 1992). Outre l'importance de l 'cau constitutive des tissus. el le est le vecteur de transport de toutes les substances nécessa i res au fonctionnement de la p lante. Enfin, e l le a un rôle essentiel dans le maintien d 'une température de la plante compatible avec son métabolisme, grùcc ù la transpiration. Cette dernière fonction est de loin l a plus consommatrice d 'cau ct participe du bilan d'énergie (cf. ci-dessus) au travers des transports de chaleur latente. Si l 'on ajoute ù ces derniers les transports de chaleur sensible, on obtient la composante advcctivc du b i lan d'énergie. Lù encore, une analogie physique assim ilant le continuum solvégétation-atmosphère ù un mi l ieu conducteur pour l 'eau a perm is d'app l iquer le formalisme de la loi d'Ohm et de dégager deux concepts fondamentaux : celui de potentiel hydrique ct celui de résistance. Ces deux concepts permettent d 'estimer le flux hydrique entre deux points quelconques du continuum sol-végétation-atmosphère (Ritchie, 1981). I l est rapidement apparu que la résistance stomatique, l iée à la régulation de l 'ouverture des stomates des feuil les par les plantes en fonction de l 'environnement - et plus particul ièrement en fonction de leur état hydrique -, est un moyen i mportant pour el les d 'ajuster leur flux transpiratoire ù leur flux d'absorption racinaire ct à la demande c l imatique. L'extension à l 'échelle du peuplement du concept de résistance stomatique - régulation biologique à l 'échelle de l 'organe- a permis d'établir un formalisme générique d'est imation de l 'évapotranspiration réel le (ETR) des peuplements végétaux, en déterminant une résistance stomatique « équivalente » pour un couvert végéta l . L'ETR recouvre à la fois l a transpiration de la p lante et l 'évaporation elu sol . La résistance stomatique elu couvert est une propriété émergente elu couvert végétal, autorégulant son flux d'eau vers l ' atmosphère en fonction des potentiels hydriques appliqués aux bornes elu système sol-végétation-atmosphère. Cette approche a été formalisée par Penmann et Monteith (Jones, 1992). I l s'agit l à d 'un véritabl e changement d 'échel le à partir d'une fonction de régulation qui s'exerce au n iveau de la p lante i ndividuel le et son extension au peuplement végétal sous form e d'analogie physique.
111

Analyse et modélisation du peuplement végétal cultivé
L'estimation simultanée ct paral l è le du flux d 'cau consommé par un couvert végétal (ETR) ct du flux net de carbone assimilé, c'est-à-dire de la biomasse produite ( MS) (si l 'on admet une concentration moyenne constante de 0,44 g C · g-1 MS) permet de calculer une efficience d'uti l isation de l'cau Ew :
Ew = I MS 1 I ETR ( Il ) .
Cette approche peut être mise en o:uvrc ù d i fférentes échel les de temps ct d 'espace. Les incréments de ETR ct cie MS peuvent être estimés de manière quasi instantanée (seconde ou m inute) ou intégrés ù des pas de temps al lant cie la journée au cycle cultur a l complet. Le rapport entre ces deux composantes influence considérablement la valeur de Ew (fïgure 2.5). L'efficience globale d 'uti l isation de l 'cau d 'une culture a donc tendance ù augmenter au cours du clévcloppcmcnt de la culture, au fur et ù mesure que la part de la transpi ration du couvert végétal devient prépondérante du fait de l a d im inution progressive de l 'énergie sola ire transmise au sol. L'approche agronomique globale de l 'efficience de l 'uti l isation de l 'eau ù l 'échel le du système sol-végétation-atmosphère do it donc i ntégrer la var iabi l ité de la composante <<évaporation elu sol >> . Cette dern ière est fonction de la dynamique de développement de l ' ind ice fol i a i re de la culture ct de l 'état hydrique des couches superficielles elu sol. De plus, comme indiqué sur lafïgure 2.5, une cu lture ayant une croissance en biomasse importante aura tendance ù avoir une forte efficience globale d 'uti l i sation de l 'cau, cc qui ne permet pas de prendre cc critère d 'efficience comme base de comparaison entre espèces et génotypes. Pour avoir une analyse plus pertinente de l 'efficience d'util isation de l 'cau par l es espèces végétales ct/ou les génotypes, il convient clone de se l imiter à la composante << transpi ration >> ET puisque, par défi n ition, l 'efficience de l 'évaporation du sol est nulle. I l devient alors nécessaire d 'effectuer un changement d'échel le, elu peuplement (sol-végétationatmosphère) ù la feui l l e , puisque c'est au n iveau de cet organe que le flux d 'cau transpi rée et le flux de carbone ass imi lé sont corégulés. Il est a lors possible soit de mesurer d i rectement ces flux, soit de les estimer en uti l isant l a rel ation fonctionnelle entre l 'effic ience de transpiration de l a feui l le ET ct la d iscr im ination isotop ique elu carbone ass imi lé : 8 13C (Farquhar ct Richards, 1984). Les processus de diffusion et de carboxylation d iscr iminent différemment l ' i sotope lourd 13C. La d iscrim ination isotopique traduit a ins i la part relative de l 'eau uti l isée pour l 'é l imination de la chaleur latente ou pour l 'activité photosynthétique. I l y a u ne relation négative entre 8 13C et Ew· Cette méthode peut être m ise en œuvre à l 'échel le de la plante au sein d'un peuplement, car le 8 13C de sa biomasse totale est révélateur de l 'efficience moyenne de transpiration sur toute l a durée de cro issance correspondante. Cette m éthode a été uti l isée pour comparer la performance des d ifférentes espèces et des différents génotypes clans des mi l ieux à fortes contraintes hydriques. Il est possib le d'ut i liser cette approche sur des i ncréments de temps p lus courts si l 'on est capabl e d'échanti l lonner sur l a p lante la biomasse élaborée durant un i ntervalle L1 t, comme par exemple pour les cernes annuels des arbres ( Guehl et al., 1993), et d'étudier l 'évolution de ET en fonction de facteurs environnementaux. L'efficience de l 'eau peut clone être étudiée à différentes échelles d'organisation -organe, p lante, peuplement- selon l es objectifs poursuivis- agronomique, écologique, physiologique ou génétique -, la connexion entre ces d ifférentes échelles n 'étant pas elu simple domaine de l ' intégration temporelle ou spatiale .
112
r Le peuplement végétal cultivé
Biomasse
(1) (2)
Évaporation Transpiration L: ETR
8 et Ill :variations d'ETR engendrant une variation de Ew pour un ET constant. (1) : culture rapidement couvrante, ou dont le sol reste sec en surface. (2) : culture à couverture lente du sol, ou dont le sol est fréquemment réhumecté
par des pluies en début de cycle. Figure 2.5. Représentation de la dynamique de croissance d'une culture en fonction de sa consomma
tion en cau, ct distinction entre la part de l'évaporation du sol ct de la transpiration du couvert végétal.
Des modèles globaux de s imulation de croissance des peuplements végétaux ont été basés sur l 'hypothèse d'une certa ine constance de Ew lorsque cette variable est intégrée sur un cycle de culture ou une période assez longue. Avec une estimation de l 'ETR, i l est alors possible d 'est imer la p roduction primaire correspondant comme suit :
MS = Ew · ETR ( 12).
Cette approche n'est pertinente que lorsque la composante << évaporation elu sol >> reste faible, comme clans le cas des forêts ou des prairies, et el le est sujette à erreur clans le cas des cultures annuelles.
�� L'approche fonctionnelle
L'approche précédente a été très l argement uti l isée clans les modèles de culture pour prévoir l a production pr imaire des couverts végétaux. Si elle permet une estimation des besoins en eau d'un couvert par exemple - utile pour piloter l ' irrigation -,
113

Analyse et modélisation du peuplement végétal cultivé
el le ne peut en aucun cas donner des i nformations sur l 'état instantané de la plante vis-à-vis de son a l imentation hydrique. De la même façon, ct pour un autre facteur important de l a production agricole- l'azote-, el le ne peut conduire à la précliction des besoins d'un couvert végétal sans comprendre comment cc facteur de la production est absorbé ct uti l isé. Enfin , m is ù part la production fourragère dont le rendement est la b iomasse, les organes récoltés ne représentent souvent qu 'une partie de cette biomasse. La quantifi cation elu partage de cette biomasse entre organes d'échange avec le m i l ieu (feui l les, rac ines) , de soutien des structures (tiges) ct reproducteurs (gra i nes, fruits) est a lors nécessaire.
Pour ces d i fférentes raisons, une approche tenant compte des processus physiologiques ct métabol iques au sein de la plante, isolée ou en peuplement, donc fonctionnel le, devient nécessaire. Nous proposons d ' i l lustrer cette approche complémentaire de la précédente ù travers : -l'étude cie l ' impact cie l a contrainte hydrique sur le fonctionnement de l a plante ct des couverts, et ses impl ications dans l 'é laboration d' ind icateurs de déficit hydrique ; -la compréhension des rapports entre ass im i lation et util isation du carbone ct de l 'azote par le peuplement végétal et leur répartition dans le couvert. Les fondements théoriques de cette répart ition conduisent ù l 'expl ici tation de la <<d i lut ion » cie l 'azote dans l a biomasse d'un couvert, et pratiquement à la détermination d'un ind ice de satisfaction des beso ins en azote pour la croissance ; - l 'analyse de la répartition des assim i lats entre les d ifférents organes constituant la plante à l'aide cie l 'approche morphogénétiquc ; cel le-ci conduit à la modél isation cie l'é laboration du rendement de la plupart des espèces cultivées.
Caractérisation et indicateurs du statut hydrique instantané de la plante
Tous les processus de la plante sont affectés par un déficit hydrique, que cc soit le métabolisme, l 'organogcnèse (production d 'organe par les méristèmes) et la morphogenèse (phénomène de différentiation et de croissance aboutissant �� des organes matures) (Hsiao, 1973). Des exemples non exhaustifs sont donnés, en d istinguant les réponses à court terme affectant généralement le métabolisme des réponses à long terme affectant la structure de la plante. Toutefois, i l est possible d'ordonner ces réponses autour de quatre axes : - l i miter les pertes en eau par une réduction de l a densité de flux, notam ment l a réduction de l 'ouverture des stomates; - l im i ter les pertes en eau par une réduction de la surface d 'échange, au travers de l a tai l le et du nombre des feu i l les ou de leur enroulement ; - augmenter l 'acquisition des ressources en eau grâce à l'exploration racinai re elu sol; - éviter la déshydratation des tissus ou en l imiter ses effets par un ajustement osmot ique ou par la production de protéines de choc thermique qui protègent les organ ites et les membranes.
Ces types de réponses se manifestent dans l 'ordre précisé ci-dessus lors du dessèchement progressif du sol. Suivant l ' intensité du déficit, tout ou partie de ces
114
r Le peuplement végétal cultivé
réponses se produisent. La première peut apparaître en quelques m inutes lors d'un déficit temporaire et réversib le, tandis que les autres nécessitent de p lusieurs heures à plusieurs jours ct sont irréversib les lorsqu'elles affectent l a structure de l a plante . Pour les trois prem iers types de réponse, les tissus de la p lante ne subissent pas d 'altération de leur contenu en cau. Le métabolisme de la plante ù l 'échelle cel l ula ire n 'est pas affecté, et les effets du déficit hydrique résultent d 'une réduction des flux d'cau. La plante sub it alors une contra inte hydrique. Pour le dernier type de réponse, les t issus de la plante commencent à sc déshydrater, provoquant un stress cel lulaire qui affecte le métabolisme cel lula ire. On parle a lors de stress hydrique. Si le dessèchement du sol sc poursuit après l a m ise en ccuvrc de l 'ensemble de ces réponses, la plante peut mour ir par déshydratation de ses tissus. Ces réponses, y compris transitoires, ont des effets induits indésirables comme l a baisse de l 'activité photosynthétique ou la perte d 'organes végétati fs ct reproducteurs qui affectent la production cie biomasse et le rendement (Passioura, 1994 ). Dans le cadre de la production agricole en zone tempérée, la contra inte hydrique est beaucoup plus fréquente que le stress hydrique.
On peut classer les plantes en deux groupes, suivant que la régulat ion de la conductance stomatiquc tend ù mainten ir l 'é ta t hydrique des tissus ou non (Tard ieu ct S imon neau, 199K ). Les plantes d ites « isohydriqucs », comme le maïs, ferment rapidement leurs stomates lors d\111 déficit hydrique. Ccci conduit ù une économie de l ' cau du sol disponib le, mais entraîne une baisse précoce de la photosynthèse. Cc comportement est bien adapté ù des déficits hydriques forts en fi n cie cycle. Il permet ù la p lante de pouvo ir term iner son cycle sans subir un stress hydrique trop prononcé. Les p lantes elites « an i sohydriqucs >>, comme le tourneso l , maintiennent tardivement leurs stomates ouverts. L'état hydrique des tissus de ces plantes tend ù suivre celu i du sol lorsque cc dern ier sc dessèche. Cc comportement <<optimiste >> est b ien adapté ù des situations caractérisées par des successions de déficits hydriques modérés ct temporaires, parce qu' i l permet de valor iser des ressources CI; cau fluctuantes sans l im iter de man ière précoce la photosynthèse. L'impact majeur du déficit hydrique reste une réduction de la production prima ire par le couvert (Joncs, 1992). Cette production est rel iée au bi lan photosynthétiquc elu couvert qu i dépend de la surface fol iaire de ce dernier et de son activité photosynthétique par un ité de surface fol ia ire. Un déficit hydrique affecte ces deux composantes au travers de processus différents dont l ' importance dépend du niveau de ce déficit hydrique, de sa position clans le cycle et de sa durée. Le mode d'action elu déficit hydrique sur ces cieux composantes est donc un déterminant i mportant de l'effet global du déficit hydrique sur la production prima ire.
Chronologiquement, la première cause de l imitation de l 'activité photosynthétique par unité cie surface est la réduction de la fourn iture de C07 aux sites de carboxylation clans les chloroplastes (Jones, 1992) . Cette l imitation se produit très tôt lors du dessèchement du sol . El le est l a conséquence de la fermeture des stomates en réponse à des signaux hormonaux racinaires émis dès l a perception du déficit hydrique (Zhang et Davies, 1990) . Cette l imitation de la fourniture en C02 n 'est pas un mécanisme adaptatif, ma is une conséquence de la réduction des pertes en eau du couvert. Elle est totalement réversible en quelques minutes lorsque la plante est à nouveau bien a l imentée en eau. Lors de cette réduction de transpiration, l a
115

Analyse et modélisation du peuplement végétal cultivé
diffusivité supérieure du CO, par rapport à cel le de la vapeur d'eau permet un maintien temporai re de l ' activité photosynthétique en retardant la baisse de la concentration en CO, de la chambre sous-stomat iquc. Lorsque le dessèchement elu sol sc poursuit, la dégradation de l 'état hydrique interne des cel lules entraîne une baisse de l 'activité des enzymes de la photosynthèse, a insi qu'une augmentation de la résistance au transport du CO, entre la chambre sous-stomatique ct le mésophyllc. L'enzyme prédominante d�ms la réponse au déficit hydrique est la rubisco, dont la concentration ct l 'activité déterminent la photosynthèse maximale (Joncs. 1 992) . Enfin, pour un déficit hydrique sévè re, la << machinerie » photosynthétiquc est détériorée elu fai t de dégradations de la st ructure quaternaire des enzymes ct des photosystèmcs (PSI et PSI!), ainsi que par la modificat ion des propriétés des membranes, notamment de leur perméabilité. Ces événements provoquent une forte chute de production de pouvoir réducteur au n iveau cel lulaire, et des désordres importants dans le métabolisme de la plante.
La ta i l le des organes végétatifs, y compris des feui l les, est réduite par un déficit hydrique quel le que soit la période où i l sc produit au cours du développement de ces organes ct que l le que soit son intensité ( Lccccur et al.. 1995) . Cel a implique que même les organes microscopiques à l'état d'ébauche sont susceptibles d'être affectés par un déficit hydrique. U ne autre conséquence est que les effets d 'un déficit hydrique ponctue l s'exprimeront aussi longtemps que tous les organes présents au moment du déficit n 'auront pas achevé leur expansion . Cela peut prendre plusieurs semaines, étant donnée la durée d'expansion des organes végétatifs. Un déficit hydrique affecte l 'expansion d 'un organe au travers de la rigiclification des parois cel lulaires ct de la réduction de la pression de turgescence i ntracel lulaire (Lockhart, 1965) . Le premier effet est l ié au changement de propriétés mécan iques des parois, en partie sous le contrôle d'une signalétique hormonale. Le second est un effet hydraul ique l i é à l a d im inution elu contenu en cau des cellules. Ces effets entraînent une réduction de la vitesse d 'expansion des tissus elu fai t d'une réduction de la vitesse d'expansion cellulai re et d 'une réduction de la vitesse de d ivision cellulaire. Dans le cas des déficits hydriques modérés, il ne s'agit pas d'un effet d irect sur le cycle cellulaire : la réduction du rythme des d ivisions cellula ires est une conséquence de la réduction de la vitesse d 'expansion. Pour les déficits hydriques sévères, cette réduction résulte du stress troph ique ct de l'altération générale elu métabolisme cellulaire causés par une forte l im i tation de l'activité photosynthétiquc:
La surface photosynthétiquement active peut également être affectée par un déficit au travers de la réduction du nombre des organes et de leur durée de vie. Dans ce cas, c'est la phénologie des organes et non leur morphogenèse qui est déterminante clans la réponse au déficit hydrique. Chez certaines plantes, la réduction de surface foliaire en réponse à un déficit hydrique résulte majoritairement de la réduction elu nombre d'organes de la plante (Belaygue et al. , 1996). Un déficit hydrique accélère également la sénescence de la surface photosynthétique en déclenchant précocement les mécanismes moléculaires de dégradation des protéi nes de la feui l le (par exemple Pic et al., 2002).
Connaître le déficit hydrique subi par une culture est essentiel pour en analyser l ' impact. Malheureusement, il n 'existe pas actuel lement de variable universelle
116
r Le peuplement végétal cultivé
facilement accessible caractérisant l 'état hydrique d 'un couvert (Turner, 1 981 ) . Cette caractérisation peut être abordée sous trois angles auxquels correspondent des variables spécifiques : - mesures de l 'état hydrique de la plante basées sur l 'état énergétique de l'cau dans les tissus (potentiels hydrique, osmotique ct de turgescence); - mesures de l'état hydrique de la plante basées sur le contenu en cau des tissus (teneur en cau des tissus, teneur en cau relative des tissus); -mesures des flux d 'cau au sein de la plante (conductance stomatiquc, flux de sève).
Ces variables appor tent des informations de natures d i fférentes ct sont complémentaires pour avoir une image de l'intensité du déficit hydrique subi et de la réponse de la plante. L'état énergétique de l 'eau indique la difficulté pour la plante à achem iner l 'cau vers ses organes. Le contenu en eau des tissus est bien re l ié à l 'activité métabolique au n iveau cel lulaire (Joncs, 1 992) . Quant aux variables mesurant les flux d 'eau, el les sont des i ndicateurs de la perception précoce par la plante du déficit hydrique. La plupart de ces variables sont rel iées quantitativement à l 'état hydrique du so l . À l 'origine de cette relation, il y a une signalétique hormonale racina i rc (Zhang ct Davics, 19lJO) . Le dessèchement du sol induit la production ct la décompartimcntation d'acide abscissiquc dans les apex racinaircs. Cette hormone migre a lors vers les parties aériennes avec le !lux xylém icn, ct est perçue par les part ies aériennes comme un signal de déficit hydrique. La réponse de la plante, notamment en termes de réduction de la conductance stomatiquc ct de l'expansion fol ia i re, est proportionnelle ù la concentration de la sève xylémiquc en acide abscissiquc. Cette chaîne de réponse est à l 'origine de la relation étroite entre la réponse de la plante ct le déficit hydrique lors des premières phases de dessèchement elu sol . Par l a suite, l ' impact du dessèchement du sol sur l 'état hydrique de la plante explique ces relations pour des n iveaux de déficit hydrique plus importants.
La caractérisation de l 'état hydrique du sol apparaît ainsi comme un indicateur de déficit hydrique subi par la plante ( Ritchie, 1 98 l ) . Pour qu' i l soit pert inent, cet ind icateur doit être représentatif de l'état hydrique moyen du sol perçu par les racines. Deux variables permettent de caractériser cet état. La première est le potentiel hydrique de la plante en fin de nuit, ou « potentiel hydrique de base ». À cc moment, il n 'y a plus de flux d'eau clans l a plante depuis plusieurs heures. Cette dernière est en équil ibre énergétique en tous ses points avec le potentiel hydrique moyen elu sol. Cc potentiel représente clone bien une i ntégration de l 'état énergétique moyen elu sol perçu par la plante. La seconde variable est la quantité d'cau disponible dans la zone de sol explorée par les racines. Cette dernière peut être exprimée en valeur absolue, généralement en mi l l imètres d'eau. On util ise cette expression lorsque l 'on veut indiquer la quantité d 'eau que le sol peut fourn i r pendant tout ou partie elu cycle de la plante. On parle alors de la « réserve hydrique du sol >> ou cl'« état de la réserve uti le >> (voi r partie 3). On considère généralement que la plante n'est pas capable d'extraire l 'eau elu sol s i le potentiel hydrique de cc dernier est i nférieur à -1,5 M Pa1• Toutefois, cette manière d'exprimer l 'eau du sol disponible ne permet pas d'avoi r une i dée elu n iveau de déficit hydrique perçu i nstantanément par la plante, parce que sa valeur dépend de la profondeur
1. Mégapascal.
117

Analyse et modélisation du peuplement végétal cultivé
d'enracinement. Pour pal l ier cet inconvénient, on peut exprimer cette quantité d'cau disponible rel at ivement à la quantité maximale que le sol peut physiquement contenir. On parle alors cl'« état de la réserve hydrique racinairc >> ou de« fraction d'cau transpirablc du sol >> ( Ritch ic , 191\ 1 ; S incla ir ct Lud low, 1 91\6 ).
Ces deux variables ont été largement ut i l isées pour étab l i r des relations quantitat ives robustes entre le niveau de déficit hydrique ct les fonctions essent ie l les de la p lante comme la transpiration. la photosynthèse, l 'expansion des t issus ou la production d'organes (Sincl a ir ct Lud low, 1 91\6; Le cœur ct Sincla ir. 1 9% ) . Ces relat ions ont permis de comparer la sensibi l ité relat ive des processus, mais aussi cel le des espèces (Sadras ct Mi l roy, 1 996) . Ainsi, l 'expansion des t issus est très généralement réduite plus précocement que l 'ouverture des stomates. Pour ces mêmes processus, les monocotylédones sont moins sensibles que les dicotylédones pour l 'expansion des tissus, tout en présentant la même sens ib i l ité pour l 'ouverture des stomates.
La répartition des assimilats dans la plante
Dynamique de croissance et prélèvement d'azote par un peuplement végétal
Comme nous l 'avons vu précédemment. la production de biomasse végétale est d irectement l iée ù la fixation de coê par le peuplement, du fa i t qu 'en première approximation la concentration en carbone de la biomasse végétale peut être considérée comme constante, a insi que les concentrations en oxygène ct en hydrogène. En revanche, la concentration de la biomasse végétale en azote est susceptible de varier clans des gammes de 0,5 ù 6 %. Ces variat ions entraînent b ien évidemment des variations importantes du prélèvement d'azote par les peuplements végétaux. Des variations analogues, bien que de plus faible ampl i tude, sont également observables pour les autres m inéraux constitutifs de la biomasse végétale tels que le phosphore, le potass ium, l e calcium, le manganèse. L'origine principale de ces variations réside dans la disponib i l i té de ces nutriments clans le sol. L'azote, du fai t de son cycle b iologique e t des apports par la ferti l isation, est le nutriment dont la d isponib i l ité dans le sol varie le plus à court terme, cc qu i expl ique en partie la grande variab i l i té de sa concentration clans la b iomasse végéta le . Cependant, Lemaire et Gastal (1997) ont montré que, même en s i tu at ion de forte d isponib i l i té en azote m inéral clans le sol, la concentration en azote de la biomasse d'un peuplement végétal d im inue au cours de sa croissance elu fait d 'une accumulat ion elu carbone plus importante que l ' azote, ce qui condu i t à l a« di lut ion >> de l ' azote clans la plante au cours de sa croissance. Cet effet de d i lut ion a été mis en évidence sur un grand nombre d'espèces, c t une formulation générale a pu être proposée :
N (%) = a (W)-h ( 13)
avec N(% ), teneur en azote de l a b iomasse aérienne elu peuplement; W, biomasse en t · ha-1 ; a, teneu r en azote pour W = 1 t · ha-1 ; et b, coefficient sans d imension.
De cette relation, il est possible de déduire la quantité d 'azote prélevée par le peuplement :
N (kg· ha-1) = 10 a (W) 1 -h exp (14)
118
r 1
Le peuplement végétal cultivé
le coefficient 10 étant nécessaire pour exprimer l 'azote prélevé en kg · ha-1• En situation de d isponibi l i té constante du m i l ieu en azote, le coe fficient a varie avec le niveau de cel le-ci, tand is que le coefficient h reste invariant. I l est alors possible de déterminer trois val eurs remarquables de 1 1 : -une va leur maximale 11m"' qu i est la teneur maximale en azote de la plante pour W = 1, ct qu i correspond ù une satu ration des capaci tés d'absorpt ion d 'azote par la p lante: -une va leur minimale 11min qui correspond au minimum de concent rat ion en azote nécessaire de la p lante pour l'activité de croissance : -une valeur cr i t ique ac qu i correspond au m inimum de concentration en azote de la plante pour une vi tesse de croissance maximale . Ces d i fférc n tes valeurs de a détcrm i ncn t des courbes corrcspondan tes (fïgurc 2. 6). Les courbes correspondant ù umax c t i1 omin sont d i ffic i les i1 déterminer expérimentalement au champ avec précision, car il est d i ffic i le de mainteni r une saturat ion de la nutrition azotée constante dans le temps sur un cycle de culture. ct impossible d 'approcher la s ituation de nutr i t ion minimale du fa i t de la fourniture d'azote par la minéral isat ion du sol. Ces deux valeurs om"' ct omin ne sont donc qu 'approchées par l 'ajustement de courbes-enve loppes encadrant les val eurs maximale ct minimale observées (fïgurc 2.6). En revanche. la valeur de a c peut être déterminée par une approche plus rigoureuse en est imant, par l 'ajustement des courbes de réponse ù l 'azote, l a teneur en azote ù partir de laquel le toute augmentation de d isponibi l ité en azote sc tradui t par une augmentation de la concentration en azote sans augmentation significative de croissance en matière sèche correspondante, selon la méthode présentée ù la .fïgure 2.6 (Justes et al., 1 994; Plénct ct Lema i re , 2000). Cette approche a été réalisée sur d iverses espèces C3 ct C4. Les val eurs de ac ct de h correspondante sont présentées dans l e !ab/cau 2.2 . On peut constater que, pour un groupe métabol ique d 'espèces donné- C3 ou C4 -, la variab i l i té de ac est faible et que la valeur de b varie peu entre l 'ensemble des espèces, avec une valeur moyenne très proche de 1/3, soit de 2/3 pour 1 - b (équation 1 3) . La différence de ac entre les espèces C3 ct C4 est conforme ù cc que l 'on connaît de leur métabolisme, les feu i l les des p lantes C4 étant moins concentrées en azote. Il est possible, ù parti r de l 'équation ( 1 4), de déterminer la demande en azote (c!Ncxp/ dt)c d'un peuplement pour réal iser à chaque instant sa vitesse de croissance maximum (c!W 1 dt) max (Lemaire et al., 2004) :
( 1 5) .
Ainsi, plusieurs conclusions s ' imposent : - la demande instantanée en azote d'un peuplement est proportionnel l e à son maximum de croissance instantanée, les effets du m il ieu sur ce potentiel de croissance entraînent clone des effets équivalents sur la demande en azote ; - le coefficient de proportionnalité entre potentiel de croissance et demande en azote décroît cependant avec l 'augmentation de biomasse du peuplement, ce qui correspond à l a « d i lution >> de l ' azote clans l a biomasse produite ; -lorsque les espèces ou les génotypes d'un même groupe métabolique sont comparés à une même biomasse W, leurs d ifférences de demande instantanée en azote sont essentie l lement expl iquées par leurs différences de vitesse de croissance du fait de la faible variation de ac et de b entre espèces d'un même groupe
119

Analyse et modélisation du peuplement végétal cultivé
N (%)
1 0
9
8
7
5
3
2
courbe de dilution maximale (amaxl
Ar---=:::,.t-___ _j_ courbe de dilution critique ac
o�-------.--------,-------.--------,-------,1-------, 0 2 4 6 8 1 0 1 2
Biomasse ( t · ha- ' ) Figure 2.6. Courbes d e dilution critique. maximale c t m inimale d e l'azote pour un peuplement d e
plantes e n C 3 . Les traits reliant les courbes représentent l a réponse d e l a croissance i t l a nutrition azotée
it différents moments de la croissance ( 1 1 • 1 , . 11 • . . ). Au-dessous de la courbe critique, les plantes s'enri
chissent en awtc ct leur croissance augmc;H� ; au-dessus, les plantes s'enrichissent en azote sans que
leur croissance n'augmente.
Tableau 2.2. Valeurs des paramètres de la courbe crit ique de prélèvement d'azote : Nue = a(W)1 - h (équation 14) pour d ifférentes espèces en C3 ct en C4 (a en kg N · ha-1 pour W en t · ha-1) .
Espèce l Oa b Sources
Graminées C3 48 0,32 Lemaire ct salette, 1 984
Luzerne C3 48 0,33 Lemaire et al. , 1 986
Pois C3 5 1 0,32 Ney et al., 1 997
Blé C3 53 0,44 Justes et al., 1 994
Colza C3 45 0,25 Co lnenne et al., 1 998
Tomate C3 45 0,33 Tei et al., 2002
Maïs C4 34 0,37 Plénet et Lemaire, 2000
Sorgho C4 39 0,39 Plénet et Cruz, 1 997
métabolique ; il ne semble donc pas y avoir de variab i li té génétique de la demande « intrinsèque » en azote, en dehors de celle correspondant au métabolisme C3 ou C4.
Il ressort de cette analyse que la dynamique de croissance en matière sèche d'un peuplement végétal et son prélèvement d'azote dans l e m i l ieu sont l iés . Pour al ler au-delà de cette relation corrélative, i l convient d 'effectuer un changement
120
r 1
Le peuplement végétal cultivé
d'échelle ct de revenir au niveau de la p lante. Comme l 'ont rapporté Lemaire ct Mil lard ( 1999), l 'absorption d'azote m inéral (NO; ct NH;) par les racines ainsi que la fixat ion de N:' sont rétrocontrôlécs par des signaux en provenance des parties aériennes. Interviennent a insi une régu lat ion posi tive l iée à la nutrit ion en carbone ct au flux d 'ass imi lats vers les racines, ct un Jc'ed-hack négat i f lié à la rcc irculat ion vers les racines d'azote organique non u t i l isé par la p lante pour sa croissance ou pour une m ise en réserves. Cc dernier signal est interprété comme un signal de « sat iété » en azote ( Lcjay el al . . 1999 : Forcle . 2002 ) . Ces deux signaux qui interagissent sont l iés à la croissance de la plante. L'act ion de ces signaux sur les deux types de transporteurs de ni trate ou d'ammonium à forte ou à fa ib le affinité (Glass el al., 2002) est ma intenant bien établ ie . Les transporteurs racina i rcs à haute affinité sont seuls act i fs pour les fa ib les concentrations en azote dans la solut ion du sol, tand is que les transporteurs à fa ib le affinité sont actifs ct complètent ceux-ci pour des concentrations é levées. Ainsi, l 'absorpt ion de l 'azote par une plante serait coréguléc à la fois par l 'offre, c'est-à-d i re la disponib i l i té de l 'azote clans le sol au contact des racines (concentrat ion de la so lu t ion) , ct par la demande, c'est-à-d ire la vitesse de croissance potentie l l e de la plante. Devienne-Barret el al. (2000) ont proposé une rel at ion permettant de rendre compte de cette corégu lation :
( 16)
avec (V11' K11) ct (V1, K1 ) , constantes de la formule de Michac l is-Mcntcn pou r les deux systèmes de transport à haute c t faible affinité respectivement : c t C, concentration en NO; de la solution du sol. L'équat ion ( 12) mélange en fai t une approche à l 'échelle du peuplement qu i reprend le formal isme de l 'équation ( I l ) pour définir la demande en azote, ct une approche à l 'échelle de la plante identifiant les constantes de Michael i s-Menten caractérisant le fonctionnement des transporteurs racinai res en fonction de l 'offre. Il y a donc une extension de cette deuxième approche de l 'échelle organe-plante entière à celle du peuplement, qui suppose qu 'une carctctérist iquc moyenne des transporteurs racinaircs des plantes à l 'échel le du peuplement ait un sens, ct que la concentration du sol en nitrates soit spat ia lcment uniforme. Ces deux hypothèses ne sont certainement pas val ides au champ, et seules les vérifications expérimentales de ce modèle (Devienne-Barret et al., 2000) permettent de s'assurer que les approximat ions effectuées par ce changement d 'échelle restent acceptables.
La répartition de l'azote au sein du peup lement végétal et le c han gement d'éc helle plante- peup lement
La d i lution de l 'azote clans l a biomasse é laborée par un peuplement végétal apparaît comme une loi très générale indépendante de d istinctions entre espèces, et a fortiori entre génotypes d'une même espèce. Il importait clone de dépasser l 'approche empirique init iale et de rechercher les déterminants de la d i lution, soit au niveau de l a plante inclivicluelle, soit au niveau d'une propriété émergente elu peuplement végétal . Deux hypothèses pouvaient apporter une explication théorique à la d i lut ion de l 'azote.
Premièrement, les travaux de Hirose et Werger ( 1987) avaient montré que clans un peuplement végétal, la quantité d'azote par unité de surface de feui l le n'était pas
121
Analyse et modélisation du peuplement végétal cultivé
uniformément répartie au sein du couvert, ct q u ' il exi stait un gradient dep u is les feu il le s du sommet les p lus éclairées, p l us riches en azote, vers les feu i l les de la base ombrées, plus pauvre. On pouvait ainsi expl iq uer la baisse de teneu r en azote d 'un pe u p lement en m ê me temps que croissait sa matière sèche par l 'augmentat ion progressive de l 'ombrage entre p lantes vo isines. Cette hypothèse étai t d'au tant p lus sédu isante que cette répart it ion de l'azote dans le couvert végétal cont ribuait ù optim iser la photosynthèse. ct donc sa croissance en m atiè re sèche. L'analyse de cc ph énom ène sur les peuplements de l uzerne ( Lemaire et al . . 199 1 ) a permis de montrer que si cc p hénomène est b i en réel, i l ne suffit pas ù exp liquer l 'ampleur de la di lut ion, car les d i m inu t ions importantes de quantités d'azote par unité de surface fo l iaire entre les feu illes éclairées c t les feu il les ombrées sont accom pagnées de diminution paral lèle de la masse surfaciquc des feu illes, cc q u i atténue notab le ment l'effet l o rsqu ' il est exprimé en teneur en azote.
Deuxièmement, les travaux de Calo in ct Yu ( 1 91\4) p u i s de G rccnwood et al. ( 1990) ont perm is de tester l ' hypot hèse selon laquel le la p lante était const i tuée cie deux compart iments, l 'un métabo l ique correspondant i't la b iomasse de la p lante di rectement im p l i q uée clans les processus cie croissance W111 ct ayant une teneur en azote N111( ';{·) é levée, ct l 'autre structural, correspondant ù la biomasse inerte W, c t ayant de cc fait une concentration en azote NJ'ir, ) faible. La teneur en azote d' une p lante peut alors s'exprimer com me su i t :
avec :
N (%) = ( 1 / W) (N 111W111 + N,W) W = W + W s Ill
( 17)
( U:\). Or par défin it ion, seul le compartiment W111 étant acti f, on peut écrire que la vitesse de croissance de la plante est proport ionne l le ù will :
dW / dt = KW111 ou : Wm / W = dW / (KWdt) ou, selon ( 1 7) c t ( 18) : N (%) = (N - N ) dW 1 KWdt + N Ill s s
( 19)
(20)
(2 1 ) . Ceci implique clone une re
. .
lat�n l inéaire entre la teneur en azote d 'une plante ct sa vitesse de croissance relati� ce qu i a été confirmé par plusieurs auteurs sur de nombreuses espèces (Greènwoocl et al., 199 1 ). Pour une plante isolée, si l 'on admettait une croissance exponentiel le, c'est-à-dire une vitesse de croissance relative constante, on devrait avoir N(%) = Cte, c'est-à-di re ne pas observer de d i lu t ion. Or il n'en est r ien, comme le montre la figure 2. 7 (Lemaire ct Gastal, 1997). Pour cela, i l faut admettre que will augmente de façon moins que proportionnel le à la biomasse totale de l a plante :
avec 0 < b < 1, ou :
soit, d 'ap rès (20) :
et d'après (17) :
122
W = K' (W)I'v Ill '
W 1 W = K' (W)I'v - t Ill
dW 1 Wdt = KK' (W)13v - t
(22)
(23)
(24)
(25)
r !
Le peuplement végétal cultivé
Cette formulation théorique de la d i lut ion se rapproche de la form u lation empirique de l'éq uation ( 13), ù ccci p rêt q u ' il devient nécessaire d' intro d u i re une asympt ote non nu l le ù la loi de d i l u t ion correspondant ù la tene u r en azote du com part im ent structural de la p lante N, q u i. d u fait de sa valeur assez faible ( 0,7 if( ) . n ' a pas d' effet très sensible dans l a gam me des biomasses habi tuel lement explorée ( 1 ù 30 t MS · ha- 1 ). Pou r une p lante isol ée, la vale ur du coefficient �� a été expér i mentalement dé term inée pour d i fférentes espèces ( luzerne. sorgho) avec une gam me de 0,79 ù O,S.:'i ( Lemaire ct GastaL 1997) cc q u i donne une gamme de - 0.2 1 ù - O, l .:'i pour [3, - 1. signilïcat ivcmcnt plus faible que cc q u i est obtenu en peu p lement dense. La .fi:fiure 2. 7 montre q ue la d il u t ion de l 'azote dans une cu l t ur e de sorgho n'est pas monotone. En tout début de croissance, pour des indices fol iaires inférieurs ù 1, la di lut ion moyenne de la p lante suit celle de la plante iso lée, p u is elle s'accé lère pour s'étab l i r avec une pente p roche de - 0,33, caractérist iq ue des peuplements denses. Il existe donc un te rme de passage entre la d i l u t ion de l'azote au sein de la p lante isolée ct la dil u t ion au sein d'un peuplement, ct le changement d'échelle devient alors possible. Ccci confirme que la com pétit ion pour la lu m ière entre les p lantes accé lère la dilu t ion de l'azote. I l ne peul s 'agir d 'une com péti t ion pou r l 'azote, car l'app l ication de doses d'azote plus é levées ne permet pas de d i m i nuer la d i lut ion d u peuple ment. L'hypothèse retenue est que l a cro i ssance des p lantes en peup lement dense accé lère la d i m inut ion du ralJ!JO rt W 1 W. Ill
N (%)
1 0
5
4
3
2
''4
0 Plantes isolées � Plantes en peuplement dense
,'4
�
�'� ._
4.
0�-----------,-----------,-----------,-----------. 0,1 1 0 1 00 1 000
Masse de la plante (g) Figure 2.7. Dilution de l'azote en coordonnées log-log pour des plantes de sorgho isolées ou des
plantes en peuplement dense (Lemaire ct Gastal, 1997, avec la permission de Springer Science and Business Media).
123
Analyse et modélisation du peuplement végétal cultivé
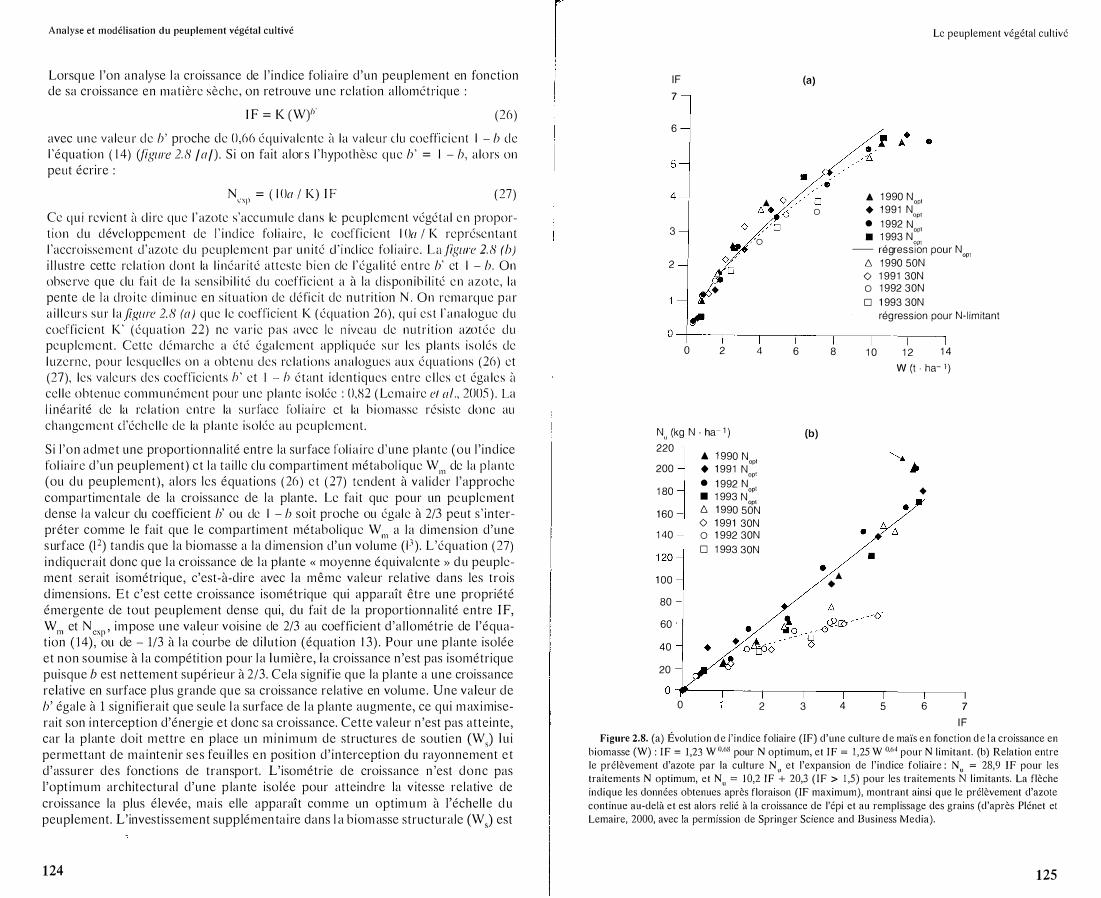
Lorsque l 'on analyse l a croissance de l ' ind ice fol ia i re d'un peuplement en fonction de sa croissance en mat ière sèche, on retrouve une relation allométrique :
I F = K (W)"" (26)
avec une valeur de b' proche de 0,66 équ ivalente à la valeur e lu coeffic ient l - b de l 'équation ( 14) (fïgure 2. 8 /a/) . Si on fait alor s l ' hypothèse que b' = l - b, alors on peut écrire :
Ncxp = ( l Oa 1 K) I F ( 27)
Cc qu i revient à el i re que l 'azote s'accumule dans le peup l ement végétal en proport ion du déve loppemen t de l ' i nd ice fol ia i re, le coeffic ient l Oa 1 K représentant l 'accroissement d'azote elu peuplement par un i té cl ' i ncl icc fol iaire. La .fïgure 2. 8 (b) i l l ustre cette relat ion dont la l i néarité atteste b ien cie l 'éga l i té entre b' ct l - b . On observe que elu fai t de l a sensibi l i té du coeffic ient a à la d isponib i l i té en azote, la pente de l a dro i te d im inue en s i tuat ion de défic i t de nutrition N . On remarque par a i l l eurs sur l a fïgure 2. 8 (a) que le coeffic ient K ( équat ion 26) , qu i est l 'ana logue du
coeffic ient K' (équat ion 22) ne var ie pas avec le n iveau de nutr i t ion azotée du peup lement . Cette démarche a été également appl iquée sur les p lan ts iso lés de
l uzerne, pour lesquel les on a obtenu des re lat ions analogues aux équations (26) ct (27), les valeurs des coefficients h' et 1 - h étant identiques entre e l les c t éga les à cel le obtenue communément pour une plante isolée : 0,1'\2 (Lemaire et al., 2005 ) . La
l i néarité de la re lat ion entre la surface fol ia i re ct la b iomasse résiste clone au
changement cl'échc l lc de la p lante isolée au peuplement .
Si l 'on admet une proportionnal i té entre la surface fol i a i re d'une plante (ou l ' ind ice fol iaire d'un peuplement) c t la tai l le elu compartiment métabo l ique Will de la p lante (ou elu peuplement) , alors les équations (26) c t (27) tendent à val ider l 'approche compartimcntale de la croissance de la plante. Le fai t que pour un peuplemen t dense l a valeur elu coefficient b' ou de l - b soit proche ou égale à 2/3 peut s ' interpréter comme le fai t que le compart iment métabolique Will a la d imension d'une surface (1") tandis que la biomasse a la d imension d'un volume ( !') . L'équation ( 27) indiquerait clone que l a croissance de l a p lante « moyenne équivalente » elu peuplement serait isométrique, c'est-à-dire avec l a même valeur relat ive clans les trois d imensions. Et c'est cette croissance isométrique qui apparaît être une propriété émergente de tout peuplement dense qui, elu fai t de la proportionnal i té entre IF, W et N , impose une valeur voisine de 2/3 au coefficient cl 'a l lométrie de l 'équa-m exp . t ion ( 1 4) , ou de - 1/3 à l a courbe de d i lut ion (équation 13) . Pour une plante isolée et non soumise à l a compétit ion pour l a lumière, la croissance n 'est pas isométrique puisque b est nettement supérieur à 2/3. Cela s ignifie que la p lante a une croissance relative en surface p lus grande que sa croissance relative en volume. Une valeur de b' égale à 1 signifierait que seule l a surface de l a p lante augmente, ce qui max imiserait son in terception d'énergie et donc sa croissance. Cette valeur n 'est pas atteinte, car la p lante doit mettre en place un m in imum de structures de soutien (Ws) l u i permettant de ma intenir ses feu il les en posit ion d' interception elu rayonnement et d'assurer des fonctions de transport. L'isométrie de croissance n 'est donc pas l'opt imum architectural d'une p lante isolée pour atteindre la v itesse relative de croissance la plus élevée, mais elle apparaît comme un optimum à l'échelle du peuplement. L'investissement supplémen ta i re clans l a b iomasse structurale (WJ est
124
r Le peuplement végétal cultivé
IF 7
6
s
 () L> + 0
3 () , ' 0
tP; , '0
2 () ,'
b •
(a)
• - ·
0 0
A 1 990 N001 + 1 991 N opt • 1 992 N001 • 1 993 N opt
-- régression pour Nopt 6 1 990 SON () 1 991 30N o 1 992 30N 0 1 993 30N
régression pour N-limitant 0 -t----,---�--�----�--�----�---
1 0 2 4
N" (kg N . ha- 1) 220
200
1 80
1 60 -
1 40
1 00
80 -
60
40
20
A 1 990 N001 + 1 991 N001 • 1 992 Nopt • 1 993 Nopt 6 1 990 SON 0 1 991 30N o 1 992 30N 0 1 993 30N
6
(b)
•
8
 •
1 0 1 2 1 4 W (t · ha- 1)
• •
•
0 �---,----,----,----,----,----�--� 0 2 3 4 s 6 7
I F Figure 2.8. (a) Évolution d e l'indice foliaire (IF) d'une culture d e maïs e n fonction d e l a croissance en
biomasse (W) : IF = 1,23 W 0·68 pour N optimum, et IF = 1 ,25 W 0·64 pour N limitant. (b) Relation entre le prélèvement d'azote par la culture N et l'expansion de l'indice foliaire · N = 28 9 IF pour les traitements N optimum, et N" = 10,2 IF � 20,3 (IF > 1 ,5) pour les traiteme1;ts N l imit�nts. La flèche indique les données obtenues après floraison (IF maximum), montrant ainsi que le prélèvement d'azote continue au-delà et est alors relié à la croissance de l'épi ct au remplissage des grains (d'après Plénet et Lemaire, 2000, avec la permission de Springer Science and Business Media).
125

Analyse et modélisation du peuplement végétal cultivé
nécessaire à la p lante pour posit ionner ses surfaces fol.ia i res dans les st 1�atcs éclai
rées ct maintenir a insi en activité sa biomasse métabol ique (W"J Le cout dc ccttc
optim isat ion est l 'accé lération de la d i l ut ion de l 'azote ct une réd�JctiOJ� d.c la VItesse
re lative de cro issance clcs p lantes. La fïgurc 2. 9 montre que cette tsomctnc de crots
sance semble être la règle pour toutes les catégories cie plantes (dominées ou domi
nantes) au sein d 'un couvert végétal dense. L'une des conséquences maJeures de
cette d i fférenciat ion au sein du couvert végétal en tre les catégoncs de plantes est
que le partage des ressources azotées semble s 'c !lcctucr au prorata de la masse (ou
de la surface ? ) fol ia ire développée par chacune des p lantes ou. p lus probab lement .
de leur contr ibut ion ù l ï n tcrccption du rayonnement par le peuplement .
(a)
- 4 - 3
(b)
- 4 - 3
- 2 - 1
- - 0
- 2 - 1
-n - - - -
- 1
- 2
- 5
- 6
- 7 Ln N%
0
- 2
- 3
0
- 4 Ln MS feuilles
Ln MS
2
•
+ D : y = 0,687x - 3 , 1 99 R2 = 0,972
0 1 : y = 0,674x - 3,353 R2 = 0,924 S : y = 0 ,747x - 3,645 R2 = 0,862
Ln MS aérienne
2
+ D : y = 0,689x - 0,684 R2 = 0,960
0 1 : y = 0,689x - 0,803 R2 = 0,873 S : y = 0,768x - 1 ,048 R2 = 0,831
Figure 2.9. Relations logarithmiques entre (a) la concentration en azote dans_
la partie aérienne et sa
biomasse sèche, ct (b) entre la biomasse sèche des feuilles et celle de la partie aene��e de plantes ayant
des positions hiérarchiques différentes au sein d'un couvert de luzerne à haute d�ns1te (D : d01111�ante� ; 1 : intermédiaire ; S : dominées). D'après Lemaire et al., 2005, avec la pern1ISSIOn d'Oxford Umvers1ty
Press.
126
r 1
Le peuplement végétal cultivé
Des i n di cateurs du n iveau de sat isfaction des besoi ns en azote
Les fondements théoriques précédents trouvent une appl ication d i recte dans le champ de l 'agronomie . La courbe crit ique, qui conduit à une croissance maximale pour le régime de nutrit ion azotée l e plus bas, permet de caractériser le n iveau de nutrit ion azoté instantané des plantes. Au-dessous de la courbe, le régime de nutrit ion est défici ta ire . tandis qu'au-dessus de la courbe, le régime est excédentaire. Pour quant ifier le déficit . un ind ice de nutri t ion ( INN) est calculé comme suit :
(2� )
où N"h' représente le pourcentage d 'azote observé dans la biomasse de la part ie aér ienne du prélèvement, c t Ncrit le calcul du pourcentage d'azote sur la courbe crit ique pour la biomasse aérienne observée. Cet ind icateur a été largement ut i l isé pour caractériser des états de nutrition de cultures variées ct contribue à l 'évaluation des méthodes de fert i l isat ion azotée des cu l tures (voir partie 1 ) . I l présente le défaut d 'être coCttcux (prélèvements de biomasse, ana lyse d'azote ) ù mettre en œuvre. D'autres ind icateurs des états du n iveau d'a l imentation en N ont été proposés ct sont ut i l isés dans la prat ique . Deux types sont maintenant couramment uti l isés. Le premier fa it appe l ù l 'analyse de la teneur en n i trate du jus de base de tige ù l 'a ide d'un k i t ct est fondé sur la c ircu l at ion de cet é lément dans la plante, clont le n iveau peut être re l ié aux états de nutrit ion (Justes ct al. , 1 <.J<.J7). Le second est basé sur les changements de couleur des feu i l les, cel le-ci étant modifiée par le régime d'al imentation en azote. Cette méthode a donné l ieu à des développements métrologiques différents, de la mesure d i recte sur un échanti l lon de feu i l les par une p ince jusqu'à la mesure embarquée, soit sur tracteur, soit par image satcl l i ta irc . Ces techniques sont prometteuses pour l 'agriculture de précis ion, la modulation de l 'épandage des engrais devenant possib le en fonction des besoins et des zones géographiques. Ces deux ind icateurs doivent être étalonnés par rapport à l ' INN qu i correspond a ins i à une référence de n iveau de nutrit ion azotée.
Répartition des assimilats dans la plante au sein d'un couvert
Les approches précédentes ne rendent pas compte de l a répartit ion des ass imi lats dans l a p lante. Pourtant, l a partie récoltée des plantes ne représente souvent qu 'une fraction de la biomasse totale produite, appelée « i nd ice de récolte » (h(//vest index en anglais), rapport de l a biomasse récoltée et de l a biomasse totale. La compréhension du partage des ass im ilats entre les d ifféren tes parties de l a p lante est donc un enjeu i mportant pour l 'agronomie. C'est pa r l 'augmentation de l ' i nd ice de récolte que de nombreuses espèces cultivées, par exemple les céréales à pail le, ont été améliorées. Une part de la biomasse allouée à la surface fol i ai re et aux tiges, excédentai re dans les variétés anciennes, à été redistribuée au profit des grains sans altérer la capture des ressources du m il ieu . Certa ines pratiques culturales ( la tai l le des arbres fruit iers ou de l a vigne par exemple) visent à assurer un équi l ibre favorable entre parties récoltées, consom matrices d'ass imi lats, et parties productrices de ces assimi lats (racines et feui l les).
127

Analyse et modélisation du peuplement végétal cultivé
La répartit ion des ass im i l ats au sein de l a plante a été abordée dans de nombreux travaux en considérant les relations entre les organes producteurs cl 'ass im i lats ( les « sources >> ) , l eur transport à travers les tubes criblés phloém icns, ct les organes consommateurs ou stockcurs ( les << pu its >> ) . Cc concept << sou rce x pu its >> a été très largement ut i l isé au cours des trente dernières années ct s'est révélé particu l i è rement fécond (Warren-Wilson, 1 972). Le forma l isme permettant de décrire la répartit ion des ass imi lais au sein d'une plante impl ique de connaître l 'activité des sources, à la fo is par l 'absorption ct ! "assimi lation d"éléments présents dans le m i l ieu , ma is aussi par la mobi l isation des réserves accumulées préalablement, le transport des ass imi lats produits ct ! "activité des pui ts , ct leurs influences respect ives. Nous ne considérerons pas ici ! "activité des sources déj it abordée précédemment ct pour lesque l les de nombreux ouvrages de synthèse existent (par exemple Joncs, 1 992) . Le transport entre sources ct puits est souvent négl igé, même s i certains travaux in tègrent des résistances au transport (M inchin el al. , 1 993).
U ne prem ière défin it ion de l a force de puits est donnée par Warren-Wilson en 1 967 : c 'est l e f lux net d 'ass imi lais importés par cc puits . Cette défin i t ion ù été rem ise en question (Patrick, 1 993), car e l l e ne fai t pas la part de lïnfl ucncc du puits l u i-même ct de cel le de la quantité d'ass im i lats d ispon ibles dans la détermination elu fl ux constaté. De nombreux travaux ont été condu its pour tenter dïsolcr cc qu i pourrait proven ir du pui ts, en défin issant une croissance potent ie l le de l 'organe l u imême. Pou r cc fa i re , p lusieurs types de d isposit ifs ont été ut i l isés. Dans certa ins, les ressources d isponibles ont été augmentées (C02, lumière . . . ) , ou bien des organes ont été supprimés (ablation de gousses par exemple) pour saturer les pui ts restant . Dans d'autres, des organes ont été prélevés pour être cult ivés in vitro clans des m i l ieux très riches en é léments nutrit ifs. Ces travaux ont souvent été peu concluants, la valeur de la force de pui ts trouvée restant très conti ngente des condi t ions de m i l ieu et des conditions de culture, très artific iel les in vitro en particu l ier, ou très cléstabi l isantcs pour les ablations d'organes. Pour éviter ces défauts, certains auteurs ont cherché les déterminants elu potentiel de croissance d 'un organe . Thaler ct Pagès ( 1 998), par exemple, montrent que le potentiel de croissance d'une racine peut être rel ié à l a tai l le de son apex lorsque la disponib i l ité en photoassimi lats n 'est pas l im i tantc. Le potent ie l de croissance des organes reproducteurs a été rel ié à leur nombre de cel lu les produ it lors de l 'organogenèsc ( M unier-Jolain et Ney, 1998, pour les grai nes ; Bert in , 2005, pour un frui t de tomate ou Ju l l ien et al. , 200.1 b pour la banane par exemple) . La période de d ivis ion cellu la i re, brève et précoce, fixerait un potentiel u ltérieur : l a v itesse de croissance des graines de pois par exemple apparaît étroitement corrélée à leur nombre de cel l ules coty1éclonaircs lorsque les conditions ne l i mitent pas l a croissance. MunicrJolain et al. ( 1 998) montrent également qu 'un doublement de la concentration elu C07 ambiant clans une chambre de culture après que les d ivis ions cel lula ires des graines d'une plante de lup in (Lupinus albus L.) ont cessé n ' augmente pas leur croissance par rapport au témoin, montrant en cela l 'existence d 'un potentiel précocement déterminé.
Le concept de potent ie l de croissance, même s'il est ut i le pour formaliser en première approche les relations entre sources et puits, notamment clans une modél i sat ion quanti tative de ces rapports, ne résiste en revanche pas à une analyse plus
128
r 1
Le peuplement végétal cul tivé
approfondie, physiologique, des processus. Patrick ( 1 993), parmi d'autres aute urs, préfère défi n i r la force d 'un pui ts comme sa capacité à maintenir un gradient osmot ique entre les sources ct lu i -même. La différence de concentration qui entret ient le flux d 'assim i lais vers l 'organe provient d 'une transformation (dans les zones d 'accumulat ion comme les gra ines par exemple) ou d'une consommation des ass imi lats parvenant au puits (comme dans les zones méristématiqucs, par resp iration) . Les étapes de déchargement du phloèmc puis de transformation sont soum ises ù des processus enzymatiques que ! "on sai t gouvernés par des lois non l i néaires, souvent miclwé l icnncs (M inchin ct Thorpc, l 9lJ3 ) , remettan t en cause ! " idée de proport ionnal i té d'un déchargement des ass imi la i s en fonction des caractérist iques d'un pu its ( ta i l l e . nombre de ce l l u les . . . ) .
Dans les règl es de partage des ass imi lats, ceux produits en excès sont généralement considérés comme mis en réserve dans un compart iment qu i sc rempl it au gré des surp lus ou des besoins en assimi l ais. Dans les forma l ismes courants, cc compartiment sc remplit l orsque les autres pui ts sont saturés, ct sc vide lorsque la production d 'ass imi la is est défic i ta ire . Cc point de vue est en contrad iction avec l ' analyse des processus cie polymérisation ct d 'hydro lyse des composés carbonés stockés dans les tissus. Ces deux processus interviennent au même moment, mais it des in tensités d i ffé rentes selon les besoins, dans les mêmes ce l lu les, ct sont régu lés probablement séparément . Le compart iment << réserves >> pourrait même être considéré comme un puits ù part ent ière , entrant en compéti t ion avec les autres, lors elu partage des ass imi lats, la polymérisat ion sc produ isant a lors même que les puits ne sont pas saturés. On trouvera un exemple d'analyse ct de modél isation de la balance polymérisat ion/hydrolyse ct de la contribution des réserves lors du rempl issage du gra in dans Bancal ct Soltani (2002) . Les formes molécu la i res de m ise en réserves peuvent être très variables ct d iverses. El les peuvent ê tre, pour le carbone, des sucres plus ou moins polyméri sés ou des proté ines (VSP : vegetative .s·torage proteins) chez certaines plantes. On admet souvent par exemple qu'el les contribuent fortement ù l a repousse d 'espèces fou rragères après l a coupe. Pour l 'azote, l 'ensemble des proté ines peut être mobil isé, notamment clans les organes en sénescence . La rubisco, produ i te en quantité excédenta i re , peut être, à certains égards, considérée comme une proté i ne de réserve, au-delà de son rôle fonctionnel clans la photosynthèse. La formation des réserves, ct la compréhension des processus y conduisant, joue un rôle important pu isque ces réserves forment la partie u ti le de certaines cultures comme l a canne à sucre ou l a betterave par exemple. On sait pour la première que l a richesse peut se trouver affa ib l ie par des condit ions favorables à une reprise de croissance q u i mobi l ise ces ressources.
�·�ccumulat ion de réserves carbonées ou azotées rétroagit sur les fonctions d'acqu i Sition correspondant. L'accumulat ion de sucres solubles ct d 'amidon clans l es feu il l es est connue pour d im inuer leur photosynthèse (Crookston et al., 1974) . De même, l ' absorption de NO.J est fortement régulée par l es produi ts de son ass im i lat ion par l a p lante (N02, NHt, et le NO.J iui -même) (Touraine et Gojon, 1997). Ces mécanismes de régulat ion sont encore l 'objet de nombreux travaux, notamment en biologie moléculaire et en physiologie.
Les d ifficultés d'analyse des relations « source x pui ts >> n'ont cependant pas empêché d 'est imer le partage des ass im i lats au sein de la plante ent ière par l a
129

Analyse et modélisation du peuplement végétal cultivé
mesure (emploi d'isotopes par exemple pour le suivi des éléments absorbés) ou la modélisation. Les travaux correspondants sont très variés quant au degré de complexité introduit selon les mécanismes mis en cause (d'une règle de priorité préétablie selon les stades de la plante jusqu'à une gestion précise des flux ), ou les compartiments pris en compte (d'une simple bicompartimcntcltion aéricn-racinaircè ou végétatif-reproducteur, jusqu ' ù un décompte exhaustif des organes en présence,
fourni par la description du développement de la plante ct de ses organes). Pour les plantes herbacées, une approche simple (Fournier ct Andrieu, lt)t)l): Drouet ct Pagès, 2003) est de considérer que tous les organes ont un accès équivalent à un pool un ique . Cette démarche peut être affinée en prenant en compte les distances entre sources ct puits (Balandicr cl al., 2000) . Enfin, l'analogie à un réseau électrique (Minchin el al., llJlJ3) fournit un cadre théorique qui a été récemment implémenté dans des modèles de plantes (Bide! cl al., 2000; D audet el al., 2002; Allen et Richardson, 196/S). Buck-Sorlin el al. (2005) ont développé un modèle d'orge dans lequel l'élongation des cn trcnœuds est p ilotée par un module simulant la biosyn
thèse des gibbércllincs. Toutefois, les exemples de travaux clans cc domaine sont rares. Les approches ''plantes virtuelles» décrites ci-après ont suscité de nombreuses recherches visant ù simuler la prospection des ressources , mais un
important travail demeure nécessaire pour concrétiser les possibilités qu'elles ouvrent dans l 'approche des régulations et des transferts internes à la plante. Ces formalismes, modélisation orientée objet (Drouet ct Pagès, :2003), ou L-.\)'Siems par exemple, offrent clone des perspectives prometteuses.
�� L'approche architecturale Les approches précédentes ont montré leur efficacité pour comprendre et simuler la production des cultures. Mais, dans le même temps, elles conduisent à un certa in nombre de lim ites pour les modèles :
- l ' hypothèse de milieu homogène et continu (figure 2.10 fa/) limite la précision des calculs physiques concernant les échanges avec l'environnement; en outre, les prédictions portent sur des moyennes sur la parcel le alors que la variabilité entre plantes ou au sein de la plante peut avoir des conséquences importantes, notam
ment en termes de qualité des produits ; -la morphologie des organes et l'organisation de l'architecture jouent un rôle dans l'activité fonctionnelle et d��ns les transferts au sein des plantes ; l'assimilation de la structure fonctionnelle à un ensemble de compartiments limite clone la gamme des processus qui peuvent être pris en compte.
Pour contourner ces limites, se développent depuis une quinzaine d'années (de Reffye et al., 1988; Prusinkiewicz et Linden mayer, 1990), des approches dites « pl antes virtuelles» ou «structure-fonction >> qui visent à une modélisation « botaniquement réaliste>> de la plante individuelle (figure 2.10 [b]) (Room et al., 1996; Prusinkiewicz, 2004). Le numéro de juin 2005 de la revue New Phytologist présente un bon aperçu des travaux récents en la matière. Dans ces approches, le peuplement est représenté comme une population de plantes individuelles qui se
2. Par exemple Tabourel-Thyot et Gastal ( 1998) pour le carbone et l'azote.
130
Le peuplement végétal cultivé
(b)
Figur� 2.10. Représentation dc.s L'Oli\'Crts dans les modèll's: <>n;ilo!!ic ;, 111: 111:1icu continu (aL '.'11 rcpréscnlalion nplicilc dcs plantes (h).
dévclopp�nt en parallèle. Les interactions entre plantes sont pri;;es en comprc du ta1t que 1 environnement ct les ressources disponibles pour lllll� nl<'i1ll.' détll'!ldcnt de la façon dont les plantes voisines modifient le milieu physique, .cl ;wum:nctrt les rcssc�urces disponibles. De telles approches nécessitent donc le couplage L'trtre des modeles stmulant les processus biologiques (fonctionnement de la plante) ct des modèles simulant les échanges physiques au sein d'un couvert vénétal, ct calculant les conditions physiques (rayonnement, température, humidité dt� sol) perçues par chacun des organes des plantes. Les apports attendus il terme sont : -une extension du domaine de validité des modèles (plante isolée. plante en peuplement, plante en serre); � un� _plus grande cohérence entre la façon dont le fonctionnement de la plante est ctuciic et celle dont on peut l'exprimer dans les modèles ; -:la possibi lité d'aborder des problèmes nouveaux. Dans cette approche, le fonctionnement d'un peuplement plurispécifique sc représente <<naturellement>>, car chant��e , des pl<�ntcs constituant les peuplements virtuels peut avoir ses propres prop�·Jetcs, rcpr
_csentant une espèce particulière. Les conséquences d 'une coupe de
prame, de la ta�lle d 'un arbre, ne peuvent être simulées sans prendre en compte la structure botamque des plantes.
Un point essenti el clans cette approche est que l ' organisation elu modè le est calquée sur la structure botanique des plantes. Les en tités elu modèle seront clone des éléments botaniques : feuille, pétiole ou gaine, entrenœud, segment raeinaire, méristème, a_ffectés de variables morphologiques (longueur, diamètre, orientation). Les co�pai"ttments et les fonctions sont distribués clans la plante et représentés par des vanables (masse, teneur en carbone, en azote, en eau, etc.) et des lois de comportement associées à ces entités. Ce changement de représentation a des conséquences importantes : d'une part sur les processus et les variables à prendre en compte dans les modèles- puisqu 'il faut notamment prendre en compte l'architecture de la plante -, d'autre part sur la façon de modéliser, du fait du caractère distribué des fonctions et des compartiments et de l'évolution de leur nombre avec le développement de la plante.
131

Analyse et modélisation du peuplement végétal cultivé
Principe de la modélisation
Croissance en dimension des organes
Dans l'approche < <plante virtuelle », l'extension individuelle de chacun des organes doit être simulée. Ccci nécessite de développer des modèles d'extension des différentes entités (limbe, pétiole, cntrcnœud, racine, pétale, etc.), tandis que dans les modèles de cultures, seuls les limbes foliaires sont généralement pris en compte. Ccci conduit donc ù rechercher des modèles génériques pouvant représenter la diversité des cinétiques de croissance ct des dimensions des organes matures au sein de l'architecture (Thaler ct Pagès, 199� ; Dosio et al., 2003 ; Fournier et al., 2005). D'autre part, les coordinations temporelles au sein de l'architecture doivent être simulées. Dans les modèles existant, coordinations ct durées de croissance sont souvent exprimées sous forme d'un<< calendrier>> de développement des organes au sein de la plante, supposé stable en temps thermique'. Cette stabilité n'est qu'approximative, ct les variations de durée de croissance jouent un rôle considérable dans la régulation des dimensions par les contraintes environnementales (Durand et al., 1999). L'importance d'une meilleure compréhension des régulations du développement qui puisse fournir une alternative ù l'approximation d'un <<calendrier» invariant est une des questions importantes soulevées par l'approche <<plantes virtuelle >>. En retour, la prise en compte dans cette approche du fonction
nement des organes dès le début de leur développement ouvre des perspectives pour comprendre les réponses différentielles des génotypes aux stress (Tardieu, 2003) ct inclure les effets différés résultant des répercussions sur les organes matures de conditions subies lors des premiers stades de développement (cf. supra << l'approche architecturale >> ).
Disposition des organes dans l'espace
La disposition des organes dans l'espace dépend de lois morphogénétiques (phylloou rhizotaxie ) , de forces physiques comme la gravité, mais également très largement de régulations actives en réponse à l'environnement perçu comme la quantité et la composition spectrale de la lumière (photomorphogenèse ), la résistance du sol à la pénétration, ou les chocs physiques (thigmomorphogenèse ). En raison de la complexité de ces processus, la plupart des modèles utilisent des descriptions statistiques - c'est-à-dire visant à reproduire au mieux les observations - plutôt que déterministes. Des modélisations déterministes commencent toutefois à se développer (Gautier et al., 2000). L'intégration de réponses dans des modèles de plante virtuelle vise à valider les hypothèses sur les lois de réponse, et à comprendre les conséquences de ces régulations sur le comportement global de la plante.
3. Le temps thermique (ou somme de température) est couramment employé en agronomie pour suivre l'avancée du développement d'une plante. Les stades de développement sont en effet plus stables
lorsque le temps est exprimé par la somme de température. Celle-ci est souvent calculée par la somme
des températures moyennes de chaque jour à laquelle est retranchée la température de base, dépendant de l'espèce et en dessous de laquelle le processus de développement considéré est arrêté.
132
f 1
Le peuplement végétal cultivé
Distribution et intégration du fonctionnement À défaut de mobilité, les plantes s'adaptent ù leur environnement par une forte plasticité, conférée par leur organisation modulaire (Hailé, 1999). Dans une certaine mesure, la plante peut être vue comme un réseau d'organes fonctionnant en parallèle, chacun en fonction de !"environnement dans lequel il sc trouve. Celuici est externe (lumière, température) ct interne (caractéristiques des tissus voisins ct des organes avec lesquels il est lié vasculaircmcnt). Ainsi, la vitesse de croissance d'un organe dépend de la température de sa propre zone de croissance. La teneur en azote foliaire s'ajuste au rayonnement perçu par la feuille, les feuilles les plus éclairées étant aussi les plus aptes ù la photosynthèse. L'orientation ct la croissance des feuilles ct des tiges (Ballaré ct Casa!, 2000) ainsi que des racines (Doussan et al., 2003) sont elles aussi régulées par l'environnement local, permettant des stratégies de recherche de ressources. Pour autant, le fonctionnement des différents organes n'est pas inclépcnclant: les transferts de proximité par diffusion au sein des tissus ou ù plus longue distance par les connexions vasculaires conduisent ù des échanges de matière ct de signaux. Identifier les règles locales cie fonctionnement ct les schémas de communication d'où émerge cette combinaison cie plasticité et cie régularité est certainement un pari et un enjeu majeur de l'approche <<plante virtuelle >>. La représentation des plantes en trois dimensions- conjointement ù des modèles de microclimat -permet de calculer l'environnement perçu par chaque organe. Elle est bien adaptée pour prendre en compte les régulations par l'environnement externe. En revanche, la perception de l'environnement interne par un organe n'est que très partiellement comprise (Pagès, 2000).
Simulation du peuplement
Les approches de simulation du peuplement sc fondent sur l'idée selon laquelle les interactions entre plantes ne sont pas directes entre individus, mais résultent des modifications que chaque plante apporte à l'environnement physico-chimique des autres plantes. L'approche << plante virtuelle >> permet alors d'aborder le fonctionnement du peuplement végétal vu comme un ensemble de plantes dont chacune interagit avec l'environnement qu'elle perçoit (figure 2.11). Cet environnement est caractérisé par des variables globales (durée du jour, gravité) et par des variables locales, distribuées sur l'architecture de chaque plante, (lumière, température, humidité, concentrations en nutriments, etc.) et qui évoluent constamment au cours du temps, notamment en fonction du développement des plantes avoisinantes (par exemple l'ombrage) et de celui de la plante elle-même (auto-ombrage, phototropisme). Un apport majeur de l'approche dans ce domaine est donc de permettre de simuler explicitement les boucles de rétroaction par lesquelles chaque plante perçoit, répond et, à son tour, modifie son environnement physique et celui de ses voisines. Les implémentations élaborées jusqu'à présent n'intègrent qu'une fraction des multiples processus mis en jeu, et sont pour la plupart focalisées sur la simulation des réponses à la lumière (Fournier et Andrieu, 1999; Gautier et al., 2000).
133

Analyse ct modélislllion du peuplement végétal cultivé
Autres plantes 1 Conditions aux bornes
1 l Représentation 30 -+1 Modèle de phytoclimat
1 du peuplement
l Représentation 30 Environnement physique
de la plante distribué sur
T l'architecture
" Modèle de fonctionnement de la plante
Développement croissance assimilation
'
'/ l -'/: r' \,
�· ,· / � l '� ' '
,;! 'ï 1 ' } l
1
/ 1
'Î ,, 41
Il l�;:r ,
1 f ,,
,
c::J variables échangées entre modèles de plante et de microclimat
c::J modèles variables de forçage
Figure 2.11. Principe de modélishtion liant l'environnement local, la réponse de ses organes constitutifs el l'intégration des processus. En bas, résultat de la simulation dans le cas d'un peuplement de maïs (d'après Fournier et Andrieu, t999).
Aspects informatiques
Le développement de l'approche << plante virtuelle >> a été rendu possible par l'évolution des méthodes de modélisation des systèmes dynamiques complexes. Il a lui-même contribué à cette évolution. L'approche« individu-centrée» de modélisation du peuplement, et plus encore celle de la modélisation de la plante en tant que colonie d'organes fonctionnant en réseau, rejoint les approches « multi-agents »
pour la modélisation de sociétés animales (fourmis, coraux). Des méthodes multi-
134
Le peuplement végét<il cultivé
agents ont été mises en œuvre pour la modélisation des interactions entre plantes (La !taud, 2004 ). Les langages objets tels que le C + + sont bien adaptés pour rendre compte du caractère modulaire de l'architecture (Bidcl et al. 2000: Drouet ct Pagès. 20!n). Le caractère dynamique de la structure pose des difficultés informatiques pour certains calcub comme la résolution des équations explicitant les flux par analogie avec un réseau électrique. car il faut maintenir la cohérence des relations entre modules au cours du développement. Toutefois. dans les plantes. l'absence c..le mobilité des cellules contraint fortement le système de relations entre modules: celui-ci est évolutif par ajouts. hranchcmcnts ct différenciations des modules. mai� lcs interconnexions créécs sont prédictiblcs par les « règles » botaniques de développement ct. en première approximation. n'évoluent ensuitc que par mortalité ou abscission. Les travaux de Lindcnmaycr ont fourni un cadre théor ique pour la conservation du réseau de relations au sein de cc type de structure dynamiquc. L'équipe de Prusinkicwicz (université de Calgary, Canada) ct celle de Kunh (université de Cottbus, Allcnwgnc) ont développé lcs fonctionnalités de ces systèmes pour exprimer les processus tic développement, de croissance ct de transferts au sein des plantes, ct pour permettre unc représenta tion en trois dimensions ainsi que l'échange d'information avec des pmgrammcs externes, simulant par exemple le rayonnement ou la température. Les logiciels développés sur celte base (Prusinkiewicz et al .. 2000; Knicmcycr et al., 2004) sont aujourd'hui le support Je plus utilisé pour J'implémentation de modèles dynamiques de plantes. Outre les aspects liés à la simulation, le développement de cette approche a conduit itla mise au point de développements théoriques ct <.l'outils spécifiques permettant l'exploration ct les calculs statistiques au sein de bases de données architec turées (Godin, 2000), qui peuvent être expérimentales ou issues de simulations. Modélisation de l'environnement Comme on l'a vu plus haut. la modélisation de l'environnement a pour mission, compte tenu de la géométrie du peuplement, des propriétés d'échanges (comme les propriétés optiques des feuilles) ainsi que des conditions de forçage (par exemple le rayonnement incident, la température au-dessus du couvert), de déterminer les conditions environnementales perçues en chaque point de la structure. Les variables concernées sont multiples: les transferts de rayonnement, les échanges à J'interface feuilles-atmosphère, ou racines-sol, les transferts par convection dans l'atmosphère, par conduction dans les sols, mais aussi le transport et l'impact de corps particulaires (goutte d'cau, pollen). On pourra se référer à Chelle (2005) pour une revue de ces aspects. Vis-à-vis du couplage avec des modèles de fonctionnement de plante, les modèles les plus opérationnels concernent la distribution du rayonnement el, dans une moindre mesure, de la température des organes végétaux. Les travaux dans cc domaine ont largement bénéficié des approches développées pour d'autres applications impliquant des représentations en trois dimensions (éclairage ou thermique de bâtimenl, vi!\tt�lisation en trois dimensions). En retour, les approches « plantes virtuelles» permettent de faire progresser la modélisation des transferts pour des applications autres que la simulation des plantes. Ainsi, la simulation des signaux mesurés en télédétection et celle des transferts physiques plante-atmosphère progressent via les possibilités de représentation en trois dimensions des plantes. En outre celle-ci permet d'aborder de nouveaux aspects, comme
135

Analyse ct modélisation du peuplement végét<tl cultivé
le transport de l'cau au sein des couverts, qui met en jeu des phénomènes de
ruissellement (Bussière et al., 2002) ct d'éclaboussure (Saint-Jciin ct al., 2004) pour lesquels l'analogie au milieu continu est inadaptée. La meilleure structuration des intcrilctions entre la plante ct son environnement, piir dissociation entre les
modèles ct définition de procédures de communication entre modèles, sc prête aussi au couplage avec des modèles d'environnement biotique : modèle de développement de maladies ou d'insectes (Skirvin, 2004) dont il fournit, en plus de la
représentation architecturée des plantes, des informations clcl's (environnement physique des pathogènes, teneur en azote des feuilles ... ) .
Pour conclure. le développement de l'approche << plante virtuelle >> représente pour la modélisation des peuplements végétaux un changement important vis-ù-vis des modèles de culture usuels. Cc changement élargit grandement les domaines d'application, en particulier vers le fonctionnement de communautés végétales plus variées que les cultures monospécifiqucs, ct vers une meilleure prise en compte des réponses aux contraintes environnementales. Plus fondamentalement, l ' approche
conduit ù ide nti fi e r la plante individuelle comme un système dynamique ù modé
liser. Maillon de départ pour l'étude du fonctionnement des communautés végétales, la plante individuelle est aussi le maillon d'arrivée de la biologie végétale.
136
Related Documents











