Le cytosquelette I - Les principes communs aux trois types de filaments, assemblage, désassemblage II - Régulation des filaments du cytosquelette III - Les moteurs moléculaires IV - Fonctions du cytosquelette dans la cellule

Le cytosquelette
Jan 17, 2016
Le cytosquelette. I - Les principes communs aux trois types de filaments, assemblage, désassemblage II - Régulation des filaments du cytosquelette III - Les moteurs moléculaires IV - Fonctions du cytosquelette dans la cellule. II - Régulation des filaments du cytosquelette. - PowerPoint PPT Presentation
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Le cytosquelette
I - Les principes communs aux trois types de filaments, assemblage, désassemblageII - Régulation des filaments du cytosqueletteIII - Les moteurs moléculairesIV - Fonctions du cytosquelette dans la cellule

II - Régulation des filaments du cytosquelette

33
Dynamisme des éléments du cytosquelette
• Régulation des filaments en longueur, stabilité, nombre et géométrie
• Interagissent les uns avec les autres et avec les autres composants de la cellule
• Parfois liaisons covalentes de protéines accessoires– Soit avec les sous-unités– Soit avec le filament

44
Plan
A. Nucléation
B. Protéines de liaison dynamisme du cytosquelette
C. Organisation d'ordre supérieur
D. Fixation des éléments du cytosquelette à la membrane plasmique

55
A – Nucléation
1. Nucléation des microtubulesa) -tubuline
b) Centrosome
c) Autres cas
2. Nucléation des microfilaments

66
1 - Nucléation des microtubules
• Centre Organisateur des MicroTubules (COMT) = site de nucléation des microtubules dans la cellule
-tubuline = nucléation des microtubules
• De la levure à l'homme
• Présente dans tous les COMT-tubuline ring complex (-TuRC) = COMT
très puissant

77
a) - Nucléation des microtubules par la -tubuline
• Nucléation à l’extrémité moins
• Allongement par l’extrémité plus
• Intervention de deux protéines qui se fixent directement à la -tubuline

88
Fig16-22
• Nucléation d'un microtubule par -TuRC
• Présence des deux protéines plus des protéines accessoires pour aider à la création de l’anneau
-tubuline ring complexes en microscopie électronique
• MT nucléés à partir de -tubuline ring complexes purifiés

99
Moritz,M2001(fig1)
Modèles de nucléation des microtubules
Microtubule nucléé spontanément à partir de
tubuline a/ pure Modèle avec matrice
Modèle protofilament
1010
Moritz,M2001(fig2)
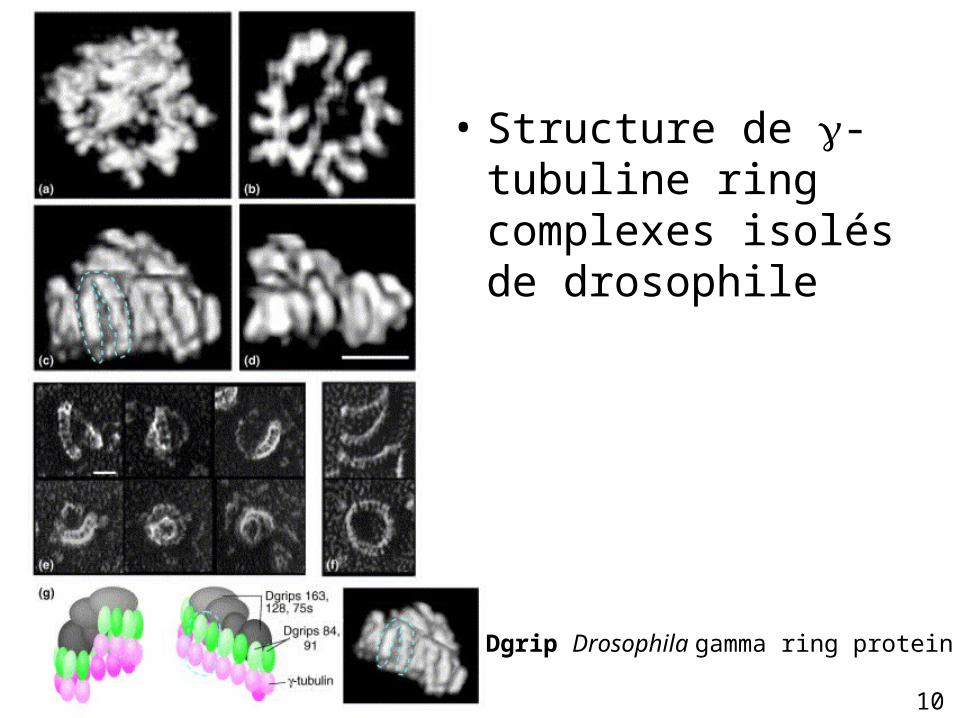
• Structure de -tubuline ring complexes isolés de drosophile
Dgrip Drosophila gamma ring protein

1111
Moritz,M2001(fig4)
New models of microtubule nucleation by the -TuRC or monomeric -tubulin
TuSC : -tubulin small complexDgrip : Drosophila gamma ring protein

1212
b) - Le centrosome : principal centre organisateur des microtubules
• Situé près du noyau
• Émanation des microtubules en étoile à partir du centrosome
• Extrémité - des MT dans le centrosome
• Extrémité + des MT en périphérie
• Contient plus de 50 copies de -TuRC dans sa matrice
• Contient une paire de centrioles

1313
Fig16-23
Le centrosome

1414
Les centrioles
• id corpuscules basaux des cils et des flagelles
• Matériel péricentriolaire = matrice centrosomale
• Duplication des centrioles suivie de la duplication des centrosomes
• cf. mitose

1515
Structure des centrioles
• Cylindre de microtubules + protéines accessoires

1616
Fig16-24
• Un centriole dans un centrosome
• Matrice centrosomale fibreuse

1717
c) - Cas particuliers de COMT
• Champignons et diatomés– pas de centriole– plaques incluses dans l'enveloppe nucléaire :
corps du pôle du fuseau
• Plantes supérieures– pas de centriole– COMT répartis tout autour de l'enveloppe
nucléaire
• Présence de -tubuline dans tous les cas

1818
Orientation des microtubules
• Configuration en étoile des microtubules
• Extrémité plus tournée vers l’extérieur de la cellule
• Extrémité moins tournée vers le noyau
• Dispositif de surveillance – de la périphérie de l’intérieur de la cellule– et de la position centrale du centrosome
• conservé même in vitro…

1919
Fig16-25
Positionnement du centrosome au centre de la cellule
Centrosome isolé + tubuline dans une chambre artificielle en plastique (image toutes les 3 minutes)

2020
•Positioning of MTOCs in microfabricated chambers. (a) Three differential interference contrast images of a centrosome in a square chamber. The chamber is 4 μm deep and the tubulin concentration is 3.2 mg/ml. Images are 3 min apart. The slope of the well dominates the signal close to the edges of the chamber. (b) Three fluorescence images of an AMTOC in a square chamber. The chamber is 6 μm deep and the tubulin concentration is 1.4 mg/ml. Time is indicated in each frame. (c) An aster regrown from an AMTOC in 2.3 mg/ml tubulin, stabilized by diluting with 30% (vol/vol) glycerol/BRB80, spun down through a cushion of 40% glycerol/BRB80 onto a coverslip coated with 3-aminopropyltriethoxysilane, and fixed with glutaraldehyde. Image taken by confocal fluorescence microscopy. (d) Two centrosomes in a square chamber. (All bars are 10 μm.)
Holy,TE1997
Positionnement des COMTs dans des micro-chambres

2121
Fig16-26
Mélanophore de poisson

2222
MT = point de repère dans la cellule
• Permet de retrouver le centre de la cellule
• Permet de disposer les organites dans les cytosol
• Propriété intrinsèque des microtubules

2323
2 - Nucléation des microfilaments (d'actine)
• (Nucléation des MT : près du noyau)• Nucléation des microfilaments : sur la membrane
plasmique accumulation de microfilaments en périphérie définit le cortex cellulaire
• Cortex cellulaire– Couche de filaments d’actine accumulés sous la
membrane plasmique– Détermine la forme et le mouvement de la surface
cellulaire forme • 1-D : microvillosités, spicules, filopodes• 2-D : lamellipodes

2424
Fig16-27
• Bord avant d’un fibroblaste : nucléation des MF• Incubation avec actine-rhodamine visualisant les nouveaux
filaments d’actine
Anciens filaments existant avant la perméabilisation
Nouveaux filaments formés sur le bord avant: site de nucléation des filaments
5 m

2525
Symons,MH1991p503• Control of actin polymerization in live and permeabilized fibroblasts
– MH Symons and TJ Mitchison J. Cell Biol. 1991 114: 503-513
• Figure 2. Localization of rhodamine-actin incorporation in fibroblasts fixed shortly after injection. (a-c) shows a detail of a cell simultaneously permeabilized and fixed 24 s after injection.
– (a) Rhodamine stain, showing newly incorporated actin. The arrows outline the section corresponding to the profiles in (d).
– (b) Fluorescein- phalloidin stain, showing both preexisting and newly incorporated filaments;
– (c) rhodamine/fluorescein ratio image, the arrowheads delineate the lamellipodium, and correspond to the fat arrows marked in the intensity profiles . The appearance of the stress fibers in the ratio image (c) as yellow over a red background, while they are not visible in the rhodamine image (a) itself, is caused by the higher background in the perinuclear part of the rhodamine image, which is divided by the stress fibers in the corresponding area of the fluorescein image (b) .
– (d) Not shown Normalized intensity profiles from the respective channels along the lines delineated by arrows in a. ( - ) Fluorescein profile; ( - - - -) rhodamine profile. The fat arrows in ddelineate the lamellipodium, the thin arrow indicates the front of the wave of incorporated actin. See Materials and Methods for further details. The color scale from green to purple corresponds to low and high rhodamine/fluorescein ratios, respectively.
– Bars, (b) 10 /,m; (d) 2 .5 Am.
• Figure 4. Steady-state incorporationof injected actin, showing a detail of a cell fixed 20 min after injection.
– (a) Rhodamine stain, showing the incorporated actin. The arrows outline the section corresponding to the profiles in d.
– (b) Phalloidin stain .
– (c) Rhodamine/ fluorescein ratio image, arrowheads delineate the lamellipodium boundaries .
– (d) Not shown Normalized intensity profiles along the section marked in a. Full-line fluorescein profile and dashed line, rhodamine profile. Fat arrows delineate the lamellipodium.
– Bars : (b) 10 um; (d) 2.5 gym.
Contrôle de la polymérisation de l’actine dans des fibroblastes vivants et perméabilisés

2626
Symons,MH1991p503
• Control of actin polymerization in live and permeabilized fibroblasts – MH Symons and TJ Mitchison
J. Cell Biol. 1991 114: 503-513
• Figure 5.Rhodamine-actin incorporation in saponin-permeabilized cells.– (a) rhodamine stain, showing exogenous actin incorporation ;– (b) fluorescein-phalloidin stain, showing preexisting and newly
incorporated filaments;– (c) rhodamine/fluorescein ratio, arrowheads outline the lamellipodial
boundary– (d) not shown / intensity profiles along the arrows shown in a. For this
experiment 0.4 t.M RA was added together with 0.2 mg/ml saponin in permeabilization buffer and incubated for 5 min before fixation.
– Bars : (b) 10 I,m; (d) 2.5 um.
Control de la polymérisation de l’actine dans des fibroblastes vivants et perméabilisés

2727
Mécanisme de la nucléation
• Régulée par des signaux externes
• Catalysée par un complexe de protéines qui comprend deux Actine Related Proteins (ARPs)

2828
Actin Related Proteins (ARPs)
• Catalysent la nucléation de l'actine
• 2 protéines proche à 45% de l'actine
• Fonction analogue à -TuRC pour la tubuline
• Comprend le complexe ARP (= Arp 2/3)
• Nuclée le filament à partir de l’extrémité –
• Et allonge rapidement l’extrémité +
• Peut se fixer latéralement embranchement

2929
Fig16-28(AB)
Rôle du complexe ARP dans la nucléation de l'actine

3030
Fig16-28(C)
Rôle du complexe ARP dans la formation du réseau d'actine

3131
Electron micrographs of quick-frozen, deep-etched, and rotary-shadowed samples of Arp2/3 complex (A) and complex mixed with gelsolin-capped actin filaments (B-D). In the presence of Arp2/3, complex actin filaments form branching arbors with numerous end-to-side connections between filaments (B). The branch points appear to be rigid attachments with a fixed 70° angle between actin filaments (C) and frequently contain a globular mass at the point of attachment (C, left arrow in B). (D) Filaments partially decorated with Arp2/3 complex. (E) Globular masses associated with filament pointed ends in the presence of Arp2/3 complex. Conditions: buffer same as Fig. 1Mullins,RD1998
Microscopie électronique de complexes Arp2/3 en congélation sublimation et ombrage rotatoire
(A) et de complexes mélangés avec des filaments d’actine cappés avec de la gelsoline (B-D).

3232
Données évolutives -tubuline et ARP
• Très anciens• Très conservés• Duplication du gène codant pour
tubuline ou actine avec une fonction de nucléation en plus par divergence et spécialisation

3333
B – Protéines de liaison dynamisme du cytosquelette
1. Protéines de liaison aux sous-unités libresa) Actine
i. Thymosine
ii. Profiline
b) Tubuline
i. Stathmine
2. Protéines de liaison latéralea) Tubuline
i. MAPs
b) Actine
i. Tropomyosine
ii. Cofiline
3. Protéines de liaison aux extrémitésa) Actine
b) Tubuline

3434
1 - Protéines de liaison aux sous-unités libres
a) Actine
b) Tubuline

3535
a) Actine
• Dans une cellule (non musculaire) : 50% actine filamenteuse / 50% actine soluble
• [Actine] en monomère dans une cellule : 50-200 M (2-8 mg/ml)
• Cc [Actine] en monomère en tube à essai : <1 M
• Pourquoi l’actine soluble ne polymérise-t-elle pas en filaments dans la cellule ?
• Parce qu’il y a des protéines qui empêchent la polymérisation en se fixant sur les sous-unités

3636
Protéines de liaison aux sous-unités libres d’actine
• Thymosine : la plus abondante
• Profiline
• Ne peuvent être fixées en même temps

3737
Thymosine
• Bloque l'actine libre • Qui ne peut s’associer ni au bout + ni au bout -
• Bloque l'échange et/ou l'hydrolyse de ATP
• Empêche l'allongement
• Comment utiliser cette actine séquestrée ?– Système de régulation de la thymosine ? non– Intervention de la profiline ? oui

3838
Profiline• Se fixe sur l'extrémité + du monomère à l’opposé
de la gorge qui contient l’ATP (extrémité -)• Bloque ainsi le côté du monomère qui
s’associerait normalement à l’extrémité – du filament
• Le complexe actine-profiline peut facilement se fixer sur l’extrémité + libre d’un filament.
• Dès que le complexe actine-profiline est additionné changement de conformation de l’actine diminution de l’affinité actine / profiline profiline part allongement facilité

3939
Résumé thymosine-profiline
• Thymosine– Bloque le monomère partout
• Profiline– Allonge le filament à son bout +
– « Déséquestre » l’actine de la thymosine

4040
Fig16-29
• Profiline liée à un monomère d'actine
• Fixée sur l'extrémité + à l’opposé de la gorge qui contient l’ATP
• Peut allonger l’extrémité + du filament
• Ne peut pas allonger l’extrémité – du filament
+
–

4141
Compétition thymosine – profiline
• Compétition pour se fixer au monomère d’actine
• Activation locale de molécules de profiline libération de l’actine séquestrée par la thymosine

4242
Fig16-30(1)
• Résumé : Effets de la thymosine et de la profiline sur la polymérisation de l'actine

4343
Fig16-30(2)• Effets de la thymosine
et de la profiline sur la polymérisation de l'actine

4444
Régulation de l’activité de la profiline
• Par phosphorylation
• Par liaison aux phospholipides inositol (PI)
• Protéines intra cellulaires avec domaines riches en proline

4545
Sites de régulation de la profiline
• Membrane plasmique
• Se lie aux phospholipides de la membrane plasmique
• En relation étroite avec l’extérieur pour faire croître lamellipodes ou filopodes

4646
b) Tubuline
• C de tubuline libre dans la cellule > Cc [tubuline] en monomère en tube à essais
• Pourquoi la tubuline soluble ne polymérise-t-elle pas en microtubules dans la cellule ?
• Parce qu’il y a des protéines qui séquestrent la tubuline libre

4747
La stathmine
• Se fixe sur deux hétérodimères de tubuline et empêche leur addition aux microtubules
• Diminue ainsi la concentration de tubuline disponible pour la polymérisation (comme la colchicine)

4848
La stathmine
• Taux d’élongation d’un MT = Kon x [tubuline]
• Si [tubuline] taux d’élongation
• Or la stathmine [tubuline] en la séquestrant • Stathmine taux d’élongation du MT

4949
La stathmine
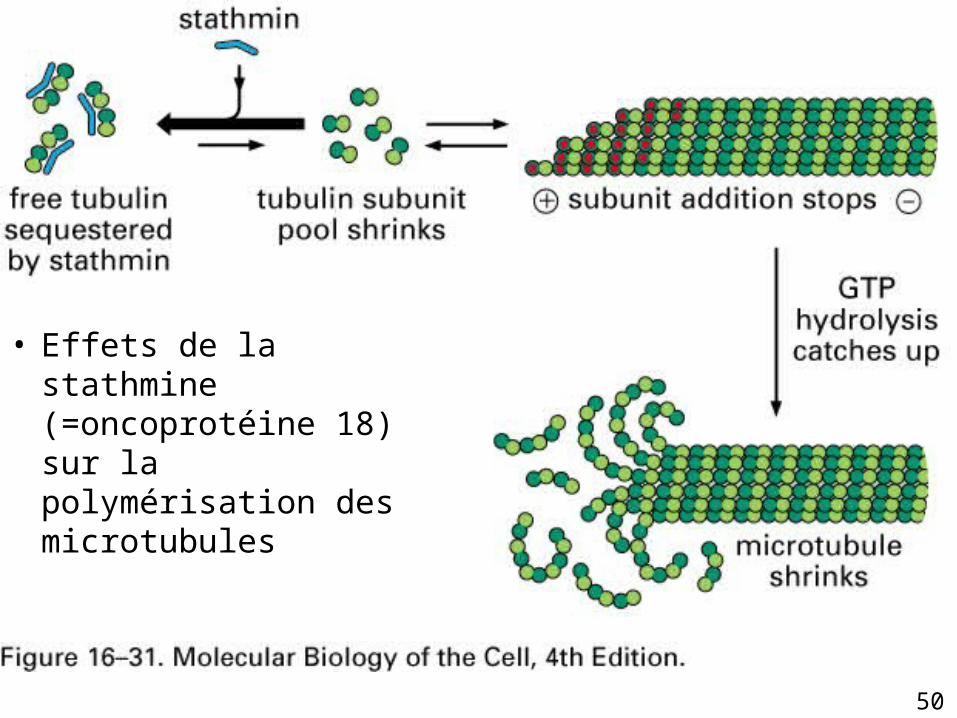
• Le passage de l’état de croissance au désassemblage du microtubule est une course entre l’hydrolyse du GTP et l’allongement du filament
• Comme la stathmine [tubuline] en la séquestrant
• Stathmine le désassemblage du microtubule
5050
Fig16-31• Effets de la stathmine
(=oncoprotéine 18) sur la polymérisation des microtubules

5151
Régulation de l’activité de la stathmine
• Phosphorylation de la stathmine inhibe la fixation à la tubuline
• Phosphorylation de la stathmine taux d’élongation du microtubule le désassemblage du microtubule
(suppression de l’instabilité dynamique)

5252
2 - Protéines de liaison latérale
Protéines qui se fixent le long du polymère pour le stabiliser ou le déstabiliser
a) Tubuline
b) Actine

5353
a) Protéines de liaison latérale : tubuline
• Les Microtubules Associated Proteins (MAPs) = protéines liées le long des microtubules (par définition)
• Peuvent stabiliser les microtubules contre le désassemblage (comme le taxol)
• Interaction des microtubules avec les autres composants de la cellule
• Forment la structure de base des axones et dendrites

5454
Fig16-32
10 m
• Localisation des MAPs dans les axones et les dendrites
• Protéine (vert)axone (ramifié)
• MAP 2corps cellulaire et dendrites

5555
Deux domaines aux MAPs
• Un domaine lié au microtubule
• Un domaine qui se projette en dehorsdont la longueur conditionne le degré d’empaquetage des microtubules– MAP 2 : long domaine externe
faisceaux de microtubules espacés– Protéines : domaine externe court
faisceaux de microtubules compacts

5656
Fig16-33
Organisation des faisceaux de MT par les MAPs
Cellule surexprimant MAP2 Cellule surexprimant tau

5757
Biochimie des MAPS (dont tau et MAP2)
• Nombreuses copies d’un motif de liaison à la tubuline
• Ces MAPs + tubuline pure non polymérisée nucléation par stabilisation des premiers oligomères de tubuline
• Cibles de plusieurs protéine kinases

5858
MAPs
• MAP2 et tau– Localisées dans certains types de cellules
des vertébrés
• Autres MAPs– Rôle central dans la dynamique des
microtubules chez presque tous les eucaryotes
– XMAP215

5959
XMAP215
– Xenopus microtubule associated protein (masse molaire=215)
– Ubiquitaire– De la levure à l’homme– Se lie le long des microtubules (devrait être dans §3)– Capacité particulière à stabiliser les extrémités libres des
microtubules– Inhibe le passage de croissance à raccourcissement des
microtubules– Pendant la mitose il y a phosphorylation de XMAP215
inhibition de cette activité augmentation 10X de instabilité des microtubules

6060
Fig.18-12 A

6161
Fig.18-12 B
• Pendant la mitose, phosphorylation de XMAP215
• Activité inhibée • Instabilité dynamique des
microtubules pendant la mitose multipliée par 10

6262
b) Protéines de liaison latérale : actine
• Tropomyosine
• Cofiline (=actine depolymerising factor)

6363
Tropomyosine
• Dans la plupart des cellules (pas que dans les cellules musculaires)
• Se lie à 7 sous-unités adjacentes d’actine dans le filament
• Empêche l’interaction avec d’autres protéines
• Rôle dans la contraction musculaire

6464
Fig.16-74

6565
Cofiline(=actine depolymerizing factor)
• Déstabilise le filament d’actine
• Peut se lier à la fois sur l’actine et sur le filament (inhabituel)
• Force le filament à s’enrouler plus serré affaiblit les contacts entre les sous-unités d’actine du filament filament plus cassant

6666
Fig16-34
Modification de la morphologie du filament d'actine par la cofiline
Filament plus court et plus épais

6767
Mécanisme d’action 1/2
• Facilite la dissociation d’une sous-unité d’actine-ADP à l’extrémité moins du filament
• Or c’est la lenteur de la dissociation à l’extrémité – qui limite le tapis roulant du filament
• La liaison à la cofiline accélération du tapis roulant
• Le durée de vie des filaments dans la cellule est beaucoup plus courte que de l’actine pure en tube à essais

6868
Mécanisme d’action 2/2
• La cofiline préfère se lier à des filaments contenant de l’actine-ADP plutôt que de l’actine-ATP
• Or l’hydrolyse de l’ATP est plus lente que l’assemblage du filament
• Les nouveaux filaments contiennent surtout de l’ATP-actine
• La cofiline déstabilise plus les anciens filaments que les nouveaux
• Accélère le turn over des filaments

6969
3 - Protéines de liaison aux extrémités du filament
a) Actine
b) Tubuline

7070
Justification de ces protéines• Protéines de liaison latérale (§ 2)
– Modifient la dynamique du filament– Mais il en faut beaucoup (doit recouvrir tout le filament)
• 1 tropomyosine pour 7 actines
• 1 tau pour 4 tubulines
• 1 cofiline pour 1 actine
• Protéines de liaison aux extrémités du filament (§ 3)– Modifient la dynamique du filament– Mais peuvent être en très faible quantité
• 1 molécule par « filament » suffit (eg 1molécule pour 200-500 actines)

7171
a) Protéines de liaison aux extrémités du filament : actine
• Protéines « cappantes »
• « cap » l’extrémité + du filament où ont lieu les changements les plus rapides du filament une fois que l’ATP de l’extrémité s’est transformée en ADP
• Ralentit considérablement les vitesses d’allongement et de raccourcissement du filament en rendant l’extrémité + inactive

7272
Fig16-35
Effet du capping des filaments (actine et pas tubuline) sur leur dynamique

7373
Protéines cappantes
• La plupart des filaments sont « capés » à leur extrémité +
• eg : protéine CapZ (strie Z du muscle)
• Complexe ARP à l’extrémité – d’un filament (celui qui a permis la nucléation)

7474
Régulation du « capping »
• Régulation locale pour adapter le cytosquelette• Signaux intra cellulaires• PIP2 (phosphatidyl Inositol-2 Phosphate) régule les
protéines qui « cappent » l’extrémité + de PIP2 dans le feuillet cytosolique dé « cappe » l’extrémité +
allongement polymérisation en surface
• Dans les cellules musculaires durée de vie très longue des filaments d’actine– CapZ au bout +– Tropomoduline au bout –– (+ Tropomysine tout le long pour stabiliser)

7575
b) Protéines de liaison aux extrémités du filament : microtubule
• Beaucoup plus compliqué– 13 protofilaments– Tube creux
• Rôle dans la dynamique des microtubules
• Rôle dans le contrôle du positionnement des microtubules

7676
Protéines de liaison aux extrémités du microtubule
i. -tubuline ring complex
ii. Complexe protéique spécial qu’on trouve aux extrémités des microtubules des cils (cf. infra)
iii. Catastrophines
iv. MAPs

7777
i - -tubuline ring complex (déjà vu)
• Nuclée le microtubule
• « Cap » l’extrémité –

7878
ii - Complexe protéique spécial qu’on trouve aux extrémités des microtubules
des cils (cf. infra)

7979
iii. Catastrophines
• Jouent sur la fréquence des « catastrophes »
• Tendent à effilocher l’extrémité du filament
• Appartiennent à la superfamille des kinésines
• Synonyme : famille Kin 1 (qui comprend KIF2 cf. moteurs infra)

8080
iv. MAPs
• Action inverse : tendent à favoriser la croissance

8181
Fig16-36(A)
Effets de la liaison de protéines aux extrémités

8282
Régulation des protéines de liaison aux extrémités du microtubule
• Toutes ces protéines sont régulées
• C’est ce qui se passe à la mitose

8383
Fig.18-12 A

8484
Fig.18-12 B
• Pendant la mitose, phosphorylation de XMAP215
• Activité inhibée • Instabilité dynamique des
microtubules pendant la mitose multipliée par 10

8585
Rôle dans le contrôle du positionnement des microtubules
• A l’extrémité – : -tubuline ring complex
• A l’extrémité + : protéines de localisation des microtubules dans le cortex cellulaire– eg :

8686
Fig16-36(B)
• Conséquences de la liaison de protéines aux extrémités des microtubules
• EB1 = protéine cappante de MT (existe chez l’homme)
• Kar9 = protéine localisée vers la membrane
Permet aux MT du fuseau de se diriger dans le bourgeon de croissance pendant la mitose

8787
C - Organisation d'ordre supérieur (cross linking)
• Les protéines vues ci-dessus interviennent dans le dynamisme
• Il faut aussi stabiliser, assurer l’intégrité mécanique, forme– Centrosome (nucléation, orientation des
microtubules, milieu de la cellule…)– MAPs– Tropomyosine …

8888
Différentes organisations faisant appel au cross linking
• Filaments intermédiaires– Association latérale des filaments entre eux– Protéines accessoires
• Actine– Deux types de protéines (tropomyosine pour
stabilisation + autres protéines pour le cross linking)
• Microtubules– MAPs ont deux domaines (MT, extérieur)

8989
Différentes organisations faisant appel au cross linking
1. Filaments intermédiaires
2. Les différents types d’organisation des filaments d’actine
3. Effets de la fragmentation des filaments sur leur dynamique
– Microtubules : catanine– Actine : gelsoline
4. Exemple : activation des plaquettes

9090
1 - Filaments intermédiaires

9191
Organisation du réseau de filaments intermédiaires
• Structure du filament robustesse– Protéine NF-M et NF-H ont un domaine C-
terminal qui forme une projection qui se lie aux FI voisins
• Filagrine : permet le maintien des faisceaux de FI de kératine
• Plectine : nombreuses propriétés

9292
Plectine
• Permet le maintien des faisceaux de vimentine
• Lie les faisceaux de FI (vimentine) aux– Microtubules– Filaments d’actine– Myosine II– Membrane plasmique

9393
Fig16-37
MT
Plectine
FI
• Liaison des filaments intermédiaires aux filaments intermédiaires
• Liaison des filaments intermédiaires aux microtubules• Liaison des filaments intermédiaires aux filaments
épais de myosine• Grâce à la plectine (anticorps marqués à l'or)
Cross linking de la plectine aux différents éléments du cytosquelette

9494
Mutations dans le gène de la plectine
• Maladie très grave– Épidermolyse bulleuse (filaments de kératine)– Dystrophie musculaire (filaments de desmine)– Dégénérescence nerveuse (neurofilaments)
• Chez la souris– Mort en quelques jours– Phlyctènes de la peau– Atteintes musculaire et cardiaque

9595
2 - Différents types d'organisation des filaments d'actine
1. Faisceau de filaments
2. Gel

9696
Fig16-38
Organisation des filaments d'actine
Fibres de stress(tension)
Exploration de l’environnement

9797
2 types de protéines cross linking
1.Protéines formant les faisceaux• Filaments parallèles• Connexion rigide entre les deux domaines de
liaison à l’actine
2.Protéines formant un gel ou un réseau• Filaments à grand angle• Connexion à courbe rigide ou flexible
Les deux ont deux sites identiques de liaison aux filaments d’actine (monomère ou dimère)

9898
Fig16-39
• Quatre exemples de protéines de liaison à l'actine
• Deux sites de liaison à l'actine (avec séquence proche)

9999
Exemples de protéines cross linking
• Faisceau
• Gel ou réseau

100100
Faisceau• Fimbrine
– 14 nm entre deux filaments– Localisation dans les filopodes et bord avant– Microvillosité
a-actinine– 30 nm entre deux filaments (plus lâche)– Fibres de stress– Contacts focaux
• Villine– Deux sites proches de liaison à l’actine ( fimbrine)– Microvillosité– Même famille que la gelsoline (cf. infra)

101101
Gel ou réseau
• Filamine– liaison en V filaments à angle droit
• Spectrine– 200 nm– Forme un réseau
• Les protéines cross linking déterminent le
type de structure d’actine…

102102
Fig16-40
• Formation de deux types de faisceaux d'actine
a-Actinine purifiée

103103
… Les protéines cross linking déterminent le type de structure
d’actine…• …et donc le type des autres molécules qui
vont interagira-actinine myosine II fibres de stress
contractiles
• Fimbrine exclut la myosine les filopodes ne sont pas contractiles
• Villine fimbrine

104104
Microvillosité• Extension en doigt de gant de la membrane
plasmique des cellules épithéliales• 0,08 m x 1 m• Cellules à fonction d’absorption• Augmente la surface d’absorption x20• Entérocyte : plusieurs milliers de microvillosités au
pôle apical• 20 – 30 filaments d’actine en faisceau• Fimbrine + actine• Fibroblaste injecté de villine microvillosité plus
longues et plus nombreuses• Bras latéraux de myosine I entre le faisceau d’actine
et les lipides de la membrane plasmique

105105
Fig16-41(A)
Microvillosité
•Extension en doigt de gant de la membrane plasmique des cellules épithéliales•0,08 m x 1 m•Cellules à fonction d’absorption•Augmente la surface d’absorption x20•Entérocyte : plusieurs milliers de microvillosités au pôle apical•20 – 30 filaments d’actine en faisceau•Fimbrine + villine•Fibroblaste injecté de villine microvillosité plus longues et plus nombreuses•Bras latéraux de myosine I entre le faisceau d’actine et les lipides de la membrane plasmique

106106
Fig16-41(BC)
• Microvillosité (cryofracture)
Contient spectrine et myosine IIAu-dessous : couche de filaments intermédiaires

107107
Karl R. Fath and David R. Burgess Microvillus
Assembly: Not actin alone Current Biology Volume 5, Issue 6 , June 1995, Pages
591-593
• Fig. 1. A summary of the main events that correlate with the localization of the major actin-binding proteins to the microvilli during brush border formation. Three stages in the assembly of microvilli and the terminal web in the apical cytoplasm of a developing chicken enterocyte are illustrated. (a) Early membrane projections expressing both ezrin and villin contain several actin filaments lying near the membrane. (b) Later, as the microvillus elongates, the number of actin filaments increases and myosin-l links the core filaments with the microvillus membrane. Both plasma membrane and myosinmay be supplied to the microvillus by Golgi-derived vesicles. (c) Coincident with the appearance of fimbrin, the number and length of the actin filaments increases and the core filaments become hexagonally packed.

108108
Formation de réseau : spectrine
• Conditionne la formation d’un réseau• Identifiée pour la première fois dans les
globules rouges• Protéine longue et flexible• 4 chaînes polypeptidiques (Deux a et deux )• Les sites d’actine sont séparés de 200 nm• Réseau à 2-D maintenu par de l’actine sous
la membrane plasmique dans le globule rouge
• Protéines membranaires

109109
Fig 10-31

110110
Globule rouge
• Cortex cellulaire rigide
• Reprise de la forme du globule rouge après passage dans un capillaire
• Équivalents de spectrine dans le cortex des autres types cellulaires des vertébrés

111111
Formation d’un gel : filamine
• Servent dans les lamellipodes qui permettent à la cellule de ramper sur un support

112112
Fig16-42
Formation d'un gel (3-D) par la filamine

113113
Fig16-43
• Cellules de mélanome– Peu de filamine (mobilité
anormale)• Pas de lamellipodes• Bulles anormales à la surface• Peu de déplacement• Pas de métastase
– Restauration artificielle de la filamine
• Lamellipodes• Hautement métastasiante

114114
3 - Effets de la fragmentation des filaments sur leur dynamique
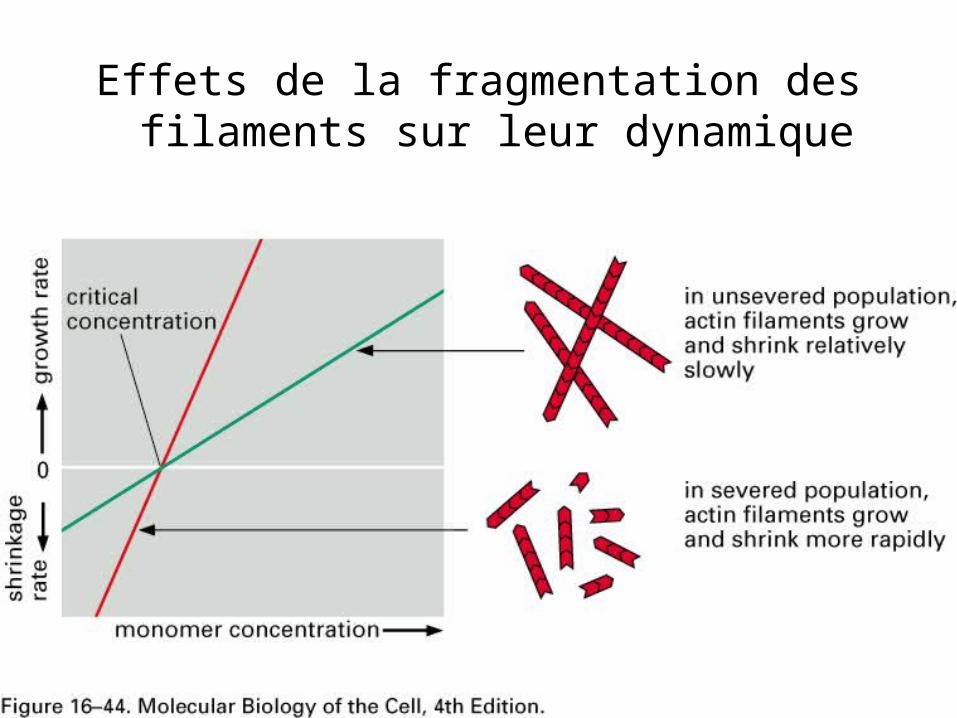
• Cassure d’un filament (en des douzaines de fragments) nombre d’extrémités + et – – Sous certaines conditions : des sites de
nucléation et allongement– Sous d’autres conditions: dépolymérisation des
vieux filaments – Changement des propriétés physiques et
mécaniques du cytoplasme– Les faisceaux et gels deviennent plus fluides
115115
Fig16-44
Effets de la fragmentation des filaments sur leur dynamique

116116
Intervention de protéines cassantes sur la longueur et la dynamique des
MF et microtubulesa) Microtubules
– Katanine
b) Actine– Gelsoline

117117
a - Microtubules : katanine
• « épée » en japonais
• Peut casser les 13 protofilaments d’un microtubule
• Deux sous-unités– Petite : hydrolyse ATP + casse le protofilament– Grosse : dirige la katanine vers le centrosome

118118
Rôle de la katanine
• Libère les microtubules de leur point d’attache à un centre organisateur de microtubules
• Applications– Dépolymérisation rapide des microtubules aux
pôles du fuseau à la mitose– Cellules en prolifération– Neurones

119119
Fig16-45
• Cassure des microtubules par la katanine– 30 s après l'addition de
Katanine
– Même champ 3 minutes après l'addition de la Katanine
MT marqués par la rhodamine et stabilisés par le taxol adsorbés sur une plaque de verre

120120
b - Microfilaments d’actine : gelsoline
• Superfamille des gelsolines
• Peut casser le filament d’actine
• Activité activée par une forte [Ca++]cytosolique
• Ne nécessite pas d’ATP

121121
Gelsoline
• Possède des sous-domaines dont deux sites
• Qui se lient à deux sites différents de la sous-unité d’actine– Un à la surface du filament– Un caché dans la liaison avec la sous-unité
suivante

122122
Mode d’action présumé de la gelsoline
• Liaison sur le côté d’un filament d’actine par un des 2 sites de liaison de la gelsoline à l’actine
• Fluctuation thermique fente entre des sous-unités voisines du protofilament d’actine
• Infiltration de la gelsoline dans la fente • Cassure du filament• « Capping » de la gelsoline à l’extrémité du filament
par son 2ème site de fixation à l’actine• Peut être retirée du filament par une augmentation
de PIP2

123123
Fig16-46
• Modèle de cassure du filament d'actine par la gelsoline

124124
4 - Application : activation des plaquettes
• Très bel exemple de
• Régulation des protéines accessoires à l’actine
• Cassure, « capping », cross linking • Grosses modifications morphologiques

125125
Plaquette au repos
• Circule dans le sang
• Pas de noyau
• Forment le caillot
• Forme discoïde
• Actine sous forme de filaments courts
• « Cappés » par CapZ
• Beaucoup de monomères d’actine liés à la profiline

126126
Activation de la plaquette
• Par contact physique avec un vaisseau endommagé
• Ou agent chimique comme la thrombine

127127
Mécanisme de l’activation
• Cascade de signalisation intra cellulaire – Afflux massif de Ca++ dans le cytosol– Ca++ active gelsoline– Gelsoline casse et « cap » les filaments en
filaments plus petits– Qui sont maintenant « cappés » de gelsoline
• Cascade de signalisation intra cellulaire de PIP2 – Inactivation de gelsoline et CapZ– Qui sont retirées toutes les deux

128128
Mécanisme de l’activation
• Augmentation du nombre d’extrémités libres
• Qui sont rapidement allongés par les nombreuses sous-unités libres d’actine
• Nombreux filaments longs

129129
Intervention des autres protéines de liaison à l’actine
• Ces nombreux filaments longs sont transformés en gel par la gelsoline
• D’autres sont transformés en faisceaux par a-actinine et fimbrine

130130
Fin de l’activation de la plaquette
• Formation de lamellipodes et filopodes pour recouvrir le caillot
• Et fixation des plaquettes au caillot par des intégrines

131131
Retour au repos
• Le signal PIP2 s’affaisse• CapZ retourne aux extrémités des filaments• Les filaments redeviennent stables• Les plaquettes reprennent leur forme étalée• Myosine II hydrolyse de l’ATP pour glisse le
long des filaments d’actine les uns par rapport aux autres
• Contraction de la plaquette qui rapproche les berges de la plaie

132132
Fig16-47(A)
• Activation plaquettaire

133133
Fig16-47(BCD)
• Activation plaquettaire
Avant l’activation Plaquette activée Après contractionde la myosine II

134134
D - Fixation des éléments du cytosquelette à la membrane plasmique
• Rôle du cytosquelette dans la forme et la rigidité de la membrane plasmique de la cellule– Cortex du GR : réseau actine + spectrine– Microvillosité : faisceau actine + villine
• Deux facteurs dans le maintien de ces structures– Maintien des filaments d’actine en faisceau et cross linking
des filaments– Fixation spécifique des filaments d’actine aux protéines ou
lipides de la membrane plasmique• De plus connexion des structures internes de la
cellule à son environnement– Autres cellules– Matrice extra cellulaire

135135
Continuum entre l’intérieur et l’extérieur de la cellule
• Éléments du cytosol >< Actine et filaments intermédiaires >< protéines transmembranaires adhésion cellulaire
• Cf. chapitre matrice extra cellulaire pour connexions membrane matrice extra cellulaire

136136
Famille ERM (Ezérine – Radixine – Moésine)
• Protéines intra cellulaires proches les unes des autres
• Attachent les filaments d’actine à la membrane plasmique– Extrémité – C de la protéine se lie directement au
filament d’actine– Extrémité - N de la protéine se lie à la face
cytoplasmique de glycoprotéines trans-membranaires dont CD44 (récepteur à l’acide hyaluronique)

137137
Neurofibromatose
• Maladie génétique
• Tumeurs nerveuses bénignes multiples
• Mutations dans les protéines ERM
• Mutation perte d’une protéine (appelée merline) de la famille ERM

138138
GENES & DEVELOPMENT 19:2265-2277, 2005 REVIEW Membrane organization and tumorigenesis—the NF2 tumor suppressor, Merlin Andrea I. McClatchey and Marco Giovannini
• Merlin-organized complexes prevent mitogenic signaling and contact-dependent inhibition of proliferation. (Left) At low cell density in cell culture, Merlin is predominantly hyperphosphorylated and inactive and mitogenic signaling proceeds. Similarly, upon cell reattachment to certain ECM substrates, a pool of Merlin is rapidly phosphorylated. (Right) At high cell density in cell culture, Merlin is hypophosphorylated, self-associated, and active in mediating contact-dependent inhibition of proliferation. Merlin is recruited to nascent cell:cell boundaries where it appears to stabilize AJs between cells, perhaps by inhibiting Rac/Pak signaling and/or stabilizing the actin cytoskeleton. Under these conditions, Merlin may inhibit signaling from AJ-associated growth-factor receptors. Hypophosphorylated Merlin can also interact with cell:ECM receptors such as CD44, which binds to hyaluronic acid (HA); increased CD44:HA interaction with increased cell density leads to high levels of active Merlin, which may inhibit associated growth-factor receptors. Notably, complete cell detachment also leads to hypophosphorylation and activation of Merlin. Perhaps inhibited mitogenic signaling from multiple complexes accumulates, reaching a threshold that halts proliferation. The study and manipulation of cell:cell contact in two dimensions in cell culture is nonphysiological; in vivo cells in a tissue are contacting other cells and can override contact-dependent inhibition of proliferation under certain normal conditions or in the context of a tumor. Merlin may coordinate the receipt of physical information from the extracellular milieu with the receipt and processing of mitogenic stimuli.

139139
Régulation des protéines d’attachement des filaments d’actine à la membrane
plasmique• Spectrine ou myosine I régulation par des
signaux intra cellulaires
• Protéines ERM régulation par des signaux intra et extra cellulaires

140140
Les deux conformations des protéines ERM
• Conformation déroulée active : oligomérise et se lie à l’actine et à CD44
• Conformation repliée inactive : les extrémités N – et C – terminales sont liées entre-elles
• Passage d’une conformation à l’autre– Phosphorylation
– Liaison à PIP2
En réponse à des signaux extra cellulaires

141141
Fig16-48
• Rôle des protéines de la famille ERM dans l'attachement des filaments d'actine à la membrane plasmique
(ezrine, radixine, moésine)

142142
Rapport du cytosquelette avec les jonctions cellulaires
1. Cytosquelette matrice extra cellulaire
2. Cytosquelette cellule voisine

143143
1 - Cytosquelette matrice extra cellulaire : contacts focaux
• Terminaison à leur niveau des fibres de stress constituées de faisceaux d’actine et myosine II
• Présence d’intégrines à leur niveau
• Intégrines (cf chapitre MEC)– Protéines d’adhésion trans-membranaire– Se lient indirectement aux faisceaux d’actine

144144
Fonctions des contacts focaux
• Permet à la cellule de tirer sur son support : via fibres de stress
• Relais de signaux de la matrice extra cellulaire vers le cytosol : via FAK

145145
Focal Adhesion Kinase (FAK)
• C’est une tyrosine kinase• Activée par l’agrégation des intégrines des
contacts focaux• Sensible au substrat de la cellule et la force
de tension : comment ??• Cible de la Src tyrosine kinase cytoplasmique• Phosphoryle de nombreuse cibles régule
survie, croissance, prolifération, morphologie, mouvement, différenciation des cellules en réponse à l’environnement

146146
Fig16-49
• Contacts focaux et fibres de stress dans un fibroblaste en culture
Microscopie à contraste interférentiel Anticorps anti-actine

147147
2 - Cytosquelette cellule voisine: cadhérines
• Protéines trans-membranaires
• Queue cytoplasmique se lient aux caténines qui se lient à l’actine
• Jonctions adhérentes = amas de contacts cellules-cellules à cadhérine-actine

148148
Fig 19-24 A-B• Molécule de cadhérine

149149
Fig 19-24 C

150150
Autres structures cytosquelette cellule voisine : desmosomes et
hémidesmosomes• Filaments intermédiaires – membrane
plasmique
• Desmosomes : cellules - cellules
• Hémidesmosomes : cellules - matrice

151151
Retentissement des signaux extra cellulaires sur le cytosquelette :
deux cibles1. Protéines associées aux éléments du
cytosquelette• Longueur, localisation , organisation dynamique• Signal extra cellulaire cytosquelette via
protéines accessoires
2. GTPases monomériques membres de la famille des protéine Rho*
• À l’intérieur de la cellule• Cible de nombreux récepteurs membranaires
* (ne pas confondre avec Rab amarrage des protéines de
transport

152152
Famille des protéines Rho
• Rho GTP-Binding Proteins – Une grande famille de protéines monomériques
de liaison au GTP impliquée• Dans la régulation de l’organisation de l’actine• L’expression des gènes• Et la progression du cycle cellulaire
• This enzyme was formerly listed as EC 3.6.1.47. Year introduced: 2000

153153
• Cells receive extracellular stimuli in the form of soluble molecules (growth factors, cytokines and hormones) that bind to cell surface receptors, adhesive interactions (extracellular matrix and cell-cell adhesion) or mechanical stresses (tension, compression and fluid shear stress). These stimuli act upon guanine-nucleotide-exchange
factors (GEFs) and GTPase-activating proteins (GAPs) to control the activation state of the small GTPases Rho, Rac and Cdc42. Once activated, the GTPases bind to a spectrum of effectors to stimulate downstream signaling pathways. The pathways shown in the poster represent the major effector pathways for these Rho family GTPases. The total number of effectors is too large to be shown here, and I apologize to authors whose work could not be included. [Excellent reviews for these effector pathways are available; for detailed information, please see (Bishop and Hall, 2000; Symons and Settleman, 2000; Bokoch, 2003; Ridley, 2001; Yoshimi et al., 2001; Riento and Ridley, 2003).] Other Rho family proteins are not included but readers are referred to a recent review (Wennerberg and Der, 2004).
• Protein kinase N1 (PKN1, also known as PRK) and PKN2 are Rho effectors involved in endosomal trafficking. Citron is a ROCK-related kinase that is critical for cytokinesis and is also implicated in other aspects of cell cycle progression. Mammalian diaphanous protein 1 (mDia1, also known as dia-related formin or DRF), mDia2 and mDia3 mediate both actin polymerization through a profilin-dependent mechanism and stabilization of microtubule plus ends in cell migration.
• Rho kinase 1 (also known as ROCK1) and ROCK2 are key Rho effectors that have multiple substrates. A partial list includes the myosin-binding subunit of the myosin phosphatase (MBS), which leads to inhibition of phosphatase activity, increased myosin light chain phosphorylation and hence increased tension generation; LIM kinase (LIMK), which phosphorylates cofilin to release actin monomers and promote actin polymerization; and myosin regulatory light chain itself, which again promotes contractility. Rho kinase phosphorylates ezrin-radixin-moesin (ERM) family proteins to activate their function as linkers between actin and the plasma membrane. Rho
kinases also phosphorylate the Na/H+ antiporter NHE-1 to promote actin-membrane interactions and several intermediate filament proteins (desmin, vimentin and glial fibrillary acidic protein) to regulate intermediate filament structure.
• Both Rac and Rho bind to phosphatidylinositol-4-phosphate 5-kinase (PIP5K); activation of PIP5K by Rho also requires Rho kinases. Production of phosphatidylinositol (4,5)-bisphosphate [PtdIns(4,5)P2]
contributes to the activation of ERM proteins and to actin polymerization through WASP, profilin and multiple actin-capping proteins.
Other reported substrates for Rho kinases that are not pictured include MARCKS, EF-1, calponin, CPI-17 and collapsin-response mediator protein 2.
• Rac has numerous effectors that mediate effects on the cytoskeleton and gene expression. Rac binds p67 PHOX to increase activation of the NADPH oxidase system and production of reactive oxygen species (ROS), which mediate activation of NF-B-dependent gene expression, effects of Rac on cell cycle progression and inhibition of Rho activity. Rac binds the WAVE complex (also containing Abi and IRSp53/58), to release active WAVE, which promotes actin polymerization in lamellipodia through activation of the Arp2/3 complex. Both Rac and Cdc42 bind and activate the kinases PAK1, PAK2 and PAK3. PAKs have multiple substrates, including LIM
kinase, which leads to actin polymerization; OP18/stathmin, which stabilizes microtubule plus ends; and Raf-1 and MEK1, whose phosphorylation by PAK enhances transmission of the signal to ERK. PAK activity also regulates myosin phosphorylation and cell contractility through several pathways, including myosin light chain kinase, myosin regulatory light chain, myosin heavy chain and caldesmon. Other pathways not listed on the diagram include filamin A, which cooperates with PAK to promote membrane ruffling; components of the paxillin-GIT/PKL-P1X complex, which regulates cell adhesion and motility; and the apoptotic regulator BAD, which promotes cell survival.
• Rac and Cdc42 are important activators of Jun N-terminal kinase (JNK) and p38, which stimulate AP-1-dependent gene expression. Mixed lineage kinases (MLKs) appear to be the major Rac/Cdc42 effectors leading to JNK and p38 activity. Rac and Cdc42 also bind to the actin-binding protein IQGAP, which is implicated in regulation of cell-cell adhesion. Both Rac and Cdc42 also bind and stimulate PI 3-kinase. The resultant 3'-phosphorylated lipids bind to and stimulate Rac GEFs, creating a positive feedback loop that maintains cell migration.
• WASP (and the more widely expressed N-WASP) are critical downstream effectors of Cdc42 that mediate formation of filopodia. These effectors also require PtdIns(4,5)P2 and interact directly with Arp2/3 complex to promote actin polymerization. Cdc42 activates the serine/threonine kinase MRCK, which is related to Rho kinases and can promote myosin phosphorylation. Cdc42 also activates the tyrosine kinases ACK1 and ACK2; the latter regulates focal adhesion formation and organization of the actin cytoskeleton. Cdc42 promotes oncogenic transformation in part by binding to the and coatamer proteins and regulating membrane trafficking.
Finally, Cdc42 is the critical determinant of polarity in a wide variety of developmental systems; this involves binding of Cdc42 to PAR6. PAR6 is a component of a complex containing PAR3 and atypical protein kinase C (PKC or PKC). This complex regulates positioning of the microtubule-organizing center (MTOC) in migrating cells and early embryos, and tight junction formation in epithelia. Borg proteins are Cdc42 effectors that connect to septins, structural proteins that can polymerize to form filaments involved in cytokinesis in yeast and mammalian cells, and that probably carry out additional structural roles in mammalian cells as well (Joberty et al., 2001).
• Finally, the poster displays some of the major feedback loops among Rho GTPases and their effector pathways. Formation of integrin-dependent cell-ECM adhesion formation, cadherin-dependent cell-cell adhesion formation and PI 3-kinase activation influence the function of GEFs and GAPs for Rho family GTPases. These positive and negative regulatory loops appear to be important for cell motility and perhaps other processes.
Rho signalling at a glance J. Cell Sci 117, 5457-5458 (2004)
Martin Schwartz

154154
Famille des protéines Rho
• Appartiennent à la super famille Ras
• 3 membres de la famille GTPases1. Cdc42 : active WASp
2. Rac : active WASp (dans sa forme GTP)
3. Rho
• Agissent comme des interrupteurs moléculaires
– Actif : liaison au GTP– Inactif : liaison au GDP

155155
Fig. 3-70

156156
Etienne-Manneville,S2002p629
• Figure 1 The Rho GTPase cycle. Twenty mammalian Rho GTPases have been described:• Rho (three isoforms: A, B, C);• Rac (1, 2, 3); Cdc42; TC10; TCL; Chp (1, 2); RhoG; Rnd (1, 2, 3); RhoBTB (1, 2); RhoD; Rif;
and TTF.• They cycle between an active (GTP-bound) and an inactive (GDP-bound) conformation. In the
active state, they interact with one of over 60 target proteins (effectors). The cycle is highly regulated by three classes of protein: in mammalian cells, around 60 guanine nucleotide exchange factors (GEFs) catalyse nucleotide exchange and mediate activation; more than 70 GTPase-activating proteins (GAPs) stimulate GTP hydrolysis, leading to inactivation; and four guanine nucleotide exchange inhibitors (GDIs) extract the inactive GTPase from membranes. All Rho GTPases are prenylated at their C terminus, and this is required for function. Rnd proteins are exceptional in that they do not hydrolyse GTP in vitro, which is an unusual property for a regulatory GTPase. It has been argued that they are controlled by expression, but it is equally possible that a yet to be identified GAP is required for hydrolysis.
Le cycle de la GTPase Rho
GDI : guanine nucleotide exchange inhibitors

157157
1 - Cdc42
• Active WASp

158158
2 - Rac-GTP
• Active WASp• Active PI(4)P-5 kinase • Génère PI(4,5)P2 • Dé - « capping » des filaments dont l’extrémité + est
liée à la gelsoline ou CapZ • Plus de sites d’assemblage à l’actine près de la
membrane plasmique • Lamellipodes et ondulations

159159
3 - Rho-GTP
• Active une protéine kinase qui inhibe une phosphatase qui agit sur les chaînes légères de myosine
• Augmentation des chaînes légères de myosine phosphorylées
• Augmentation de l’activité contractile de la myosine • Augmentation des fibres de stress

160160
Fig 16-67 A
• Illustration

161161
Fig 16-67 B
• Coloration négative en microscopie électronique de filaments de myosine II (plus courts que ceux que l’on trouve dans la cellule musculaire striée) assemblés in vitro par phosphorylation de leurs chaînes légères

162162
Membres de la famille WASp
• Conformation inactive repliée / active dépliée (comme ERM)
• Protéine WASp + cdc 42-GTP stabilisation de la forme ouverte liaison à ARP de l’activité de nucléation du complexe ARP
• Nucléation de l’actine

163163
Dynamique et organisation de l’actine
1. Activation de cdc 42 – Polymérisation de l’actine– Formation de faisceaux – Filopodes et spicules
2. Activation de rac – Polymérisation de l’actine à la périphérie de la cellule – Lamellipodes et ondulations
3. Activation de rho – Formation de faisceaux d’actine avec de la myosine II pour former
des fibres de stress– Formation de contacts focaux par agglomération d’intégrines et
protéines associées

164164
Mode d’action de ces 3 « interrupteurs »
• Nombreuses protéines cibles en aval modifications de l’organisation et de la dynamique
de l’actine

165165
Etienne-Manneville,S2002p629
• Figure 1 The Rho GTPase cycle. Twenty mammalian Rho GTPases have been described: Rho (three isoforms: A, B, C); Rac (1, 2, 3); Cdc42; TC10; TCL; Chp (1, 2); RhoG; Rnd (1, 2, 3); RhoBTB (1, 2); RhoD; Rif; and TTF. They cycle between an active (GTP-bound) and an inactive (GDP-bound) conformation. In the active state, they interact with one of over 60 target proteins (effectors). The cycle is highly regulated by three classes of protein: in mammalian cells, around 60 guanine nucleotide exchange factors (GEFs) catalyse nucleotide exchange and mediate activation; more than 70 GTPase-activating proteins (GAPs) stimulate GTP hydrolysis, leading to inactivation; and four guanine nucleotide exchange inhibitors (GDIs) extract the inactive GTPase from membranes. All Rho GTPases are prenylated at their C terminus, and this is required for function. Rnd proteins are exceptional in that they do not hydrolyse GTP in vitro, which is an unusual property for a regulatory GTPase. It has been argued that they are controlled by expression, but it is equally possible that a yet to be identified GAP is required for hydrolysis.
• Les 3 membres de la famille Rho fonctionnent de façon semblable
• Activation par GEF (Guanine nucleotide Exchange Factor) (on en connaît plus de 20)
Le cycle de la GTPase Rho

166166
Fig16-50
• Effets de Rho, Rac, et Cdc42 sur l'organisation de l'actine dans le fibroblaste (actine colorée à la pholloïdine fluorescente, contacts focaux colorés par AC anti vinculine)
Nombreuses fibres de stress et contacts focaux
Énorme lamellipode qui fait le tour de la cellule Nombreux filopodes en périphérie

167167
Alan Hall
Rho GTPases and the Actin Cytoskeleton Science, Vol 279, Issue 5350, 23 january 1998, p 509
• Fig. 1. Rho, Rac, and Cdc42 control the assembly and organization of the actin cytoskeleton. Quiescent, serum-starved Swiss 3T3 fibroblasts (-) contain very few organized actin filaments (A) or vinculincontaining integrin adhesion complexes (B). The effects of Rho, Rac, or Cdc42 activation in these cells can be observed in several different ways such as with the addition of extracellular growth factors, microinjection of activated GTPases, or microinjection of guanosine diphosphate (GDP)–guanosine triphosphate (GTP) exchange factors. Addition of the growth factor lysophosphatidic acid activates Rho, which leads to stress fiber (C) and focal adhesion formation (D). Microinjection of constitutively active Rac induces lamellipodia (E) and associated adhesion complexes (F). Microinjection of FGD1, an exchange factor for Cdc42, leads to formation of filopodia (G) and the associated adhesion complexes (H). Cdc42 activates Rac; hence, filopodia are intimately associated with lamellipodia, as shown in (G). In (A), (C), (E), and (G), actin filaments were visualized with rhodamine phalloidin; in (B), (D), (F ), and (H), the adhesion complexes were visualized with an antibody to vinculin. Scale: 1 cm 5 25 mm. [Figure courtesy of Kate Nobes]

168168
• Fig. 4. ERM proteins are required for GTPase-mediated cytoskeletal changes. It has been proposed that ERM proteins exist in a closed (inactive) conformation and an open (active) conformation. There is evidence to suggest that this transition can be regulated by Rho, perhaps through the activation of a Ser-Thr kinase or a lipid kinase (through PIP2). In the active conformation, the NH2-terminus of ERM proteins (pink) can interact with transmembrane proteins, such as CD44, whereas the COOH-terminus (blue) interacts with F-actin. This membrane–ERM–F-actin unit is an essential prerequisite for the Rho GTPases to induce cytoskeletal changes. In the case of Rho, this is likely to be mediated by the bundling and reorganization of preexisting actin-myosin filaments to generate stress fibers, whereas in the case of Rac and Cdc42 it is likely that preexisting filaments are uncapped to allow actin polymerization and filament growth leading to the formation of lamellipodia and filopodia.
Alan Hall, Science, Vol 279, Issue 5350, 23 january 1998 p509Rho GTPases and the Actin Cytoskeleton

169169
Alan Hall, Science, Vol 279, Issue 5350, 23 january 1998 p509 Rho GTPases and the Actin Cytoskeleton
• Fig. 5. Cdc42p-associated signaling complexes in S. cerevisiae. Cdc42p is essential for both pheromone- induced mating and for nitrogen starvation–induced filamentous growth. In the pheromone response, Bem1p acts a scaffold protein and interacts with Cdc24p (an exchange factor for Cdc42p), Far1p (a cell cycle inhibitor), actin, Ste20p (a Ser-Thr kinase that can interact with Cdc42p), and Ste5p. Ste5p is a scaffold protein that interacts with components of a MAP kinase cascade. Ste20p activates the MAP kinase cascade, but no interaction with Cdc42p is required. A role of Cdc42p is to localize this signaling complex to the mating projection. Cdc42p and Ste20p are also required to activate a distinct MAP kinase pathway (still not completely defined) in response to nitrogen starvation. In this case, the interaction between Cdc42p and Ste20p is essential.
Related Documents











