LC8 dynein light chain (DYNLL1) binds to the C-terminal domain of ATM-interacting protein (ATMIN/ASCIZ) and regulates its subcellular localization Péter Rapali b , María Flor García-Mayoral e , Mónica Martínez-Moreno a , Krisztián Tárnok c , Katalin Schlett c , Juan Pablo Albar d , Marta Bruix e , László Nyitray b,⇑ , Ignacio Rodriguez-Crespo a,⇑ a Dept. Biochemistry and Molecular Biology I, Universidad Complutense, Madrid, Spain b Dept. Biochemistry, Eötvös Loránd University, Budapest, Hungary c Dept. Physiology and Neurobiology, Eötvös Loránd University, Budapest, Hungary d Proteomics Facility, CNB, CSIC, Madrid, Spain e Dept. Biological Physical Chemistry, IQFR, CSIC, Madrid, Spain article info Article history: Received 16 September 2011 Available online 24 September 2011 Keywords: Microtubules Dynein Intracellular transport ATMIN abstract LC8 dynein light chain (now termed DYNLL1 and DYNLL2 in mammals), a dimeric 89 amino acid protein, is a component of the dynein multi-protein complex. However a substantial amount of DYNLL1 is not associated to microtubules and it can thus interact with dozens of cellular and viral proteins that display well-defined, short linear motifs. Using DYNLL1 as bait in a yeast two-hybrid screen of a human heart library we identified ATMIN, an ATM kinase-interacting protein, as a DYNLL1-binding partner. Interest- ingly, ATMIN displays at least 18 SQ/TQ motifs in its sequence and DYNLL1 is known to bind to proteins with KXTQT motifs. Using pepscan and yeast two-hybrid techniques we show that DYNLL1 binds to mul- tiple SQ/TQ motifs present in the carboxy-terminal domain of ATMIN. Recombinant expression and puri- fication of the DYNLL1-binding region of ATMIN allowed us to obtain a polypeptide with an apparent molecular mass in gel filtration close to 400 kDa that could bind to DYNLL1 in vitro. The NMR data-driven modelled complexes of DYNLL1 with two selected ATMIN peptides revealed a similar mode of binding to that observed between DYNLL1 and other peptide targets. Remarkably, co-expression of mCherry- DYNLL1 and GFP-ATMIN mutually affected intracellular protein localization. In GFP-ATMIN expressing- cells DNA damage induced efficiently nuclear foci formation, which was partly impeded by the presence of mCherry-DYNLL1. Thus, our results imply a potential cellular interference between DYNLL1 and ATMIN functions. Ó 2011 Elsevier Inc. All rights reserved. 1. Introduction LC8 dynein light chain (DYNLL) is a small, highly conserved eukaryotic dimeric protein of 89 amino acids. Initially, it was iden- tified as a subunit of dynein and myosin Va motor proteins and, due to its two binding sites, it was assumed to function as a cargo adaptor [1–3]. However, a significant amount of DYNLL is not asso- ciated with any molecular motor [4]. In fact, recent results indicate that DYNLL acts as a hub protein in several cellular events having functions unrelated to cargo transport, for instance in nuclear transport, apoptosis and cancer development [5,6]. DYNLL has more than sixty known protein binding partners with a wide vari- ety of functions thought to be regulated by DYNLL binding. Several X-ray and NMR structures of apo- and DYNLL complexes with partner peptides are available [7,8]. Structural data, mutagenesis, pepscan and in vitro binding assays have revealed that DYNLL has a well-defined homodimer structure in which two parallel binding grooves form at the dimerization interface. These binding grooves recognize a short linear motif with a loose consensus se- quence [D/S]KX[T/V/I]Q[T/V][D/E] that adopts a b-strand confor- mation. The central, highly conserved glutamine is, in many cases, flanked by either threonines (TQT) or hydrophobic residues (e.g. IQV) [7,9–15]. The DYNLL recognizing motif is generally lo- cated in disordered regions of partner proteins, frequently in the vicinity of coiled coil regions or potential dimerization domains [5,16]. As a matter of fact, structural, kinetic and thermodynamic data suggest that DYNLL might be a dimerization engine [5,7,8]. Using yeast two-hybrid techniques and DYNLL1 as bait, we re- trieved ATMIN as a new interacting partner protein. ASCIZ/ATMIN (hereafter called ATMIN) share an ATM-interaction motif present near the carboxy-terminus and homologous to that present in NBS1 [17]. Besides, ATMIN has four N-terminal zinc finger domains followed by a central core domain which is necessary for Rad51- containing focus formation after ssDNA damage, and a stretch with 11 SQ/TQ motifs at the carboxy terminal end (17 SQ/TQ in the full- length polypeptide) which implicate ATMIN as a potential ATM 0006-291X/$ - see front matter Ó 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.bbrc.2011.09.093 ⇑ Corresponding authors. E-mail addresses: [email protected] (L. Nyitray), [email protected] (I. Rodri- guez-Crespo). Biochemical and Biophysical Research Communications 414 (2011) 493–498 Contents lists available at SciVerse ScienceDirect Biochemical and Biophysical Research Communications journal homepage: www.elsevier.com/locate/ybbrc

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biochemical and Biophysical Research Communications 414 (2011) 493–498
Contents lists available at SciVerse ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
LC8 dynein light chain (DYNLL1) binds to the C-terminal domain ofATM-interacting protein (ATMIN/ASCIZ) and regulates its subcellular localization
Péter Rapali b, María Flor García-Mayoral e, Mónica Martínez-Moreno a, Krisztián Tárnok c,Katalin Schlett c, Juan Pablo Albar d, Marta Bruix e, László Nyitray b,⇑, Ignacio Rodriguez-Crespo a,⇑a Dept. Biochemistry and Molecular Biology I, Universidad Complutense, Madrid, Spainb Dept. Biochemistry, Eötvös Loránd University, Budapest, Hungaryc Dept. Physiology and Neurobiology, Eötvös Loránd University, Budapest, Hungaryd Proteomics Facility, CNB, CSIC, Madrid, Spaine Dept. Biological Physical Chemistry, IQFR, CSIC, Madrid, Spain
a r t i c l e i n f o
Article history:Received 16 September 2011Available online 24 September 2011
Keywords:MicrotubulesDyneinIntracellular transportATMIN
0006-291X/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.bbrc.2011.09.093
⇑ Corresponding authors.E-mail addresses: [email protected] (L. Nyitray), na
guez-Crespo).
a b s t r a c t
LC8 dynein light chain (now termed DYNLL1 and DYNLL2 in mammals), a dimeric 89 amino acid protein,is a component of the dynein multi-protein complex. However a substantial amount of DYNLL1 is notassociated to microtubules and it can thus interact with dozens of cellular and viral proteins that displaywell-defined, short linear motifs. Using DYNLL1 as bait in a yeast two-hybrid screen of a human heartlibrary we identified ATMIN, an ATM kinase-interacting protein, as a DYNLL1-binding partner. Interest-ingly, ATMIN displays at least 18 SQ/TQ motifs in its sequence and DYNLL1 is known to bind to proteinswith KXTQT motifs. Using pepscan and yeast two-hybrid techniques we show that DYNLL1 binds to mul-tiple SQ/TQ motifs present in the carboxy-terminal domain of ATMIN. Recombinant expression and puri-fication of the DYNLL1-binding region of ATMIN allowed us to obtain a polypeptide with an apparentmolecular mass in gel filtration close to 400 kDa that could bind to DYNLL1 in vitro. The NMR data-drivenmodelled complexes of DYNLL1 with two selected ATMIN peptides revealed a similar mode of binding tothat observed between DYNLL1 and other peptide targets. Remarkably, co-expression of mCherry-DYNLL1 and GFP-ATMIN mutually affected intracellular protein localization. In GFP-ATMIN expressing-cells DNA damage induced efficiently nuclear foci formation, which was partly impeded by the presenceof mCherry-DYNLL1. Thus, our results imply a potential cellular interference between DYNLL1 andATMIN functions.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction has a well-defined homodimer structure in which two parallel
LC8 dynein light chain (DYNLL) is a small, highly conservedeukaryotic dimeric protein of 89 amino acids. Initially, it was iden-tified as a subunit of dynein and myosin Va motor proteins and,due to its two binding sites, it was assumed to function as a cargoadaptor [1–3]. However, a significant amount of DYNLL is not asso-ciated with any molecular motor [4]. In fact, recent results indicatethat DYNLL acts as a hub protein in several cellular events havingfunctions unrelated to cargo transport, for instance in nucleartransport, apoptosis and cancer development [5,6]. DYNLL hasmore than sixty known protein binding partners with a wide vari-ety of functions thought to be regulated by DYNLL binding. SeveralX-ray and NMR structures of apo- and DYNLL complexes withpartner peptides are available [7,8]. Structural data, mutagenesis,pepscan and in vitro binding assays have revealed that DYNLL
ll rights reserved.
[email protected] (I. Rodri-
binding grooves form at the dimerization interface. These bindinggrooves recognize a short linear motif with a loose consensus se-quence [D/S]KX[T/V/I]Q[T/V][D/E] that adopts a b-strand confor-mation. The central, highly conserved glutamine is, in manycases, flanked by either threonines (TQT) or hydrophobic residues(e.g. IQV) [7,9–15]. The DYNLL recognizing motif is generally lo-cated in disordered regions of partner proteins, frequently in thevicinity of coiled coil regions or potential dimerization domains[5,16]. As a matter of fact, structural, kinetic and thermodynamicdata suggest that DYNLL might be a dimerization engine [5,7,8].
Using yeast two-hybrid techniques and DYNLL1 as bait, we re-trieved ATMIN as a new interacting partner protein. ASCIZ/ATMIN(hereafter called ATMIN) share an ATM-interaction motif presentnear the carboxy-terminus and homologous to that present inNBS1 [17]. Besides, ATMIN has four N-terminal zinc finger domainsfollowed by a central core domain which is necessary for Rad51-containing focus formation after ssDNA damage, and a stretch with11 SQ/TQ motifs at the carboxy terminal end (17 SQ/TQ in the full-length polypeptide) which implicate ATMIN as a potential ATM

494 P. Rapali et al. / Biochemical and Biophysical Research Communications 414 (2011) 493–498
and ATR kinase substrate. Fine mapping studies revealed that sev-eral DYNLL1 dimers might associate with the full-length ATMIN.We describe herein this interaction and discuss its possible biolog-ical significance.
2. Materials and methods
2.1. Yeast two-hybrid screen, b-galactosidase assay, pepscan andexpression of DYNLL1 and ATMIN in Escherichia coli
We have followed our own published protocols [7,10,12,15,18].Residues 378–823 and 475–823 of ATMIN were cloned in a pET-15b vector using NdeI and EcoRI and expressed in frame with ahexa-His tag in bacteria. Expression was performed at 25 �C andpurification involved binding to an affinity Ni–NTA resin followedby gel filtration using a Serva D-6900 Polyol Si300 column.
2.2. Chemical crosslinking
ATMIN fragment (378–823) and DYNLL1 were dialyzed againstPBS buffer (pH 7.3). 20-fold molar excess of dimethyl pimelimi-date�2 HCl (DMP) was added to the proteins. After the reaction2� SDS running buffer supplied by 3-mercaptoethanol was added
Fig. 1. Identification of ATMIN as a DYNLL1-interacting protein and fine mapping of the bfingers in green, core domain in yellow, DYNLL1 binding motifs (SQ/TQ) in blue and thecarboxy-terminus are shown. Using a yeast two-hybrid screen of a human library, resiATMIN constructs were confronted against DYNLL1 and all of them rendered a positivconfronted with empty plasmid (ep). (B) Pepscan analysis of the putative DYNLL1 bindinshown to bind to DYNLL1. These DYNLL1 binding motifs within ATMIN were highly conselength GFP-tagged ATMIN and DYNLL1 were co-immunoprecipitated in COS7 cells usingreferences to color in this figure legend, the reader is referred to the web version of thi
to the samples and analyzed by tricine or glycine SDS gelelectrophoresis.
2.3. NMR experiments and sample preparation
Two N15-labelled DYNLL1 samples of 109 and 85 lM concentra-tion were prepared in aqueous solutions of 90% H2O/10% D2O in100 mM potassium phosphate buffer, pH 7.0, 1 mM DTT. Concen-trated solutions of ATMIN peptides 488-GVSRETQTSGIE-499 and666-ESLDIETQTDFL-677 were prepared in the same buffer withestimated concentrations of 8.8 and 4.0 mM, respectively. Seriesof N15-HSQC spectra were recorded at 25 �C at the different stepsof the titration experiments in a Bruker Avance 800 NMR spec-trometer (Bruker, Rheinstetten, Germany) equipped with a z-gradi-ent cryoprobe as previously reported [14,19].
2.4. Gel-filtration chromatography
ATMIN (378–823) fragment was analyzed by gel-filtration usingServa Polyol Si-300 column. The measurements were performed in20 mM phosphate buffer (pH 7.3) containing 200 mM NaCl. Theflow rate was 0.5 ml per minute. The hydrodynamic size was deter-mined using Bio-Rad Gel-Filtration Standard.
inding site. (A) Schematic representation of the various modules found in ATMIN (ZnATM kinase binding site in red). Only the 11 DYNLL1 binding sites located at the
dues 362–823 of ATMIN (carboxy-terminus) were shown to bind to DYNLL1. Foure interaction. Conversely, no interactions were obtained when either protein wasg site within the ATMIN sequence. At least 11 ATMIN isolated dodecapeptides wererved among mammals as revealed by a Clustal alignment of the sequences. (C) Full-an anti-DYNLL1 antibody but not a pre-immune serum. (For interpretation of the
s article.)
P. Rapali et al. / Biochemical and Biophysical Research Communications 414 (2011) 493–498 495
2.5. Isothermal titration calorimetry
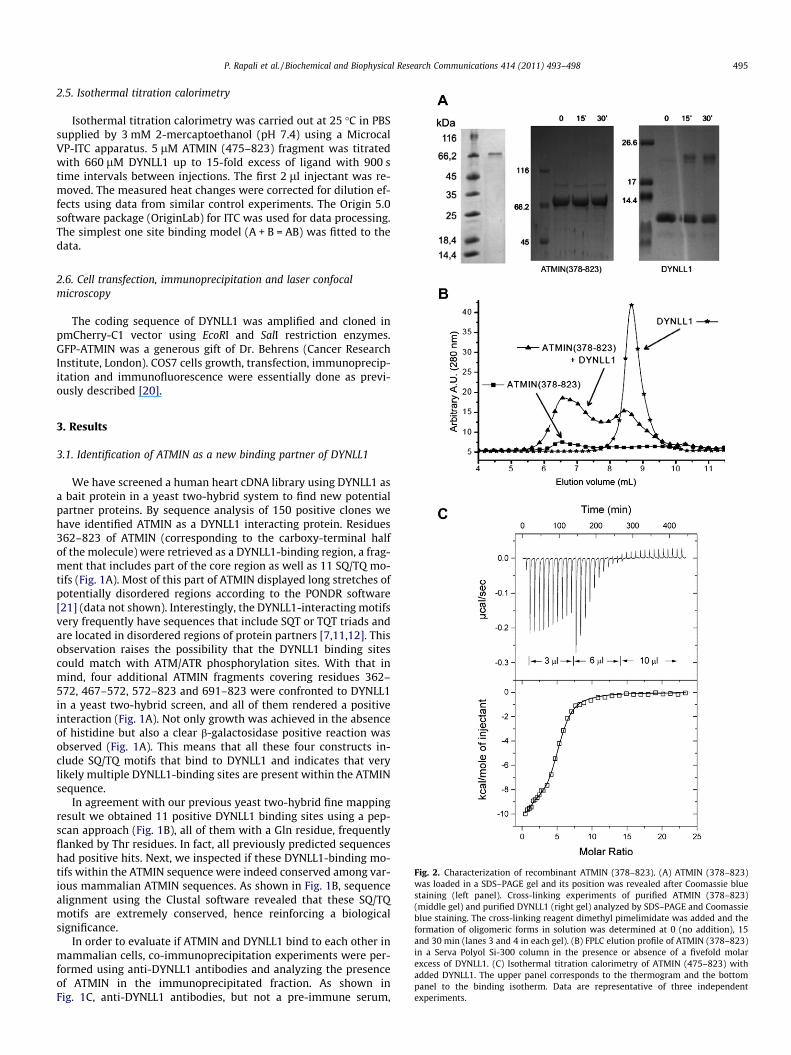
Isothermal titration calorimetry was carried out at 25 �C in PBSsupplied by 3 mM 2-mercaptoethanol (pH 7.4) using a MicrocalVP-ITC apparatus. 5 lM ATMIN (475–823) fragment was titratedwith 660 lM DYNLL1 up to 15-fold excess of ligand with 900 stime intervals between injections. The first 2 ll injectant was re-moved. The measured heat changes were corrected for dilution ef-fects using data from similar control experiments. The Origin 5.0software package (OriginLab) for ITC was used for data processing.The simplest one site binding model (A + B = AB) was fitted to thedata.
2.6. Cell transfection, immunoprecipitation and laser confocalmicroscopy
The coding sequence of DYNLL1 was amplified and cloned inpmCherry-C1 vector using EcoRI and SalI restriction enzymes.GFP-ATMIN was a generous gift of Dr. Behrens (Cancer ResearchInstitute, London). COS7 cells growth, transfection, immunoprecip-itation and immunofluorescence were essentially done as previ-ously described [20].
ig. 2. Characterization of recombinant ATMIN (378–823). (A) ATMIN (378–823)as loaded in a SDS–PAGE gel and its position was revealed after Coomassie blue
taining (left panel). Cross-linking experiments of purified ATMIN (378–823)iddle gel) and purified DYNLL1 (right gel) analyzed by SDS–PAGE and Coomassie
lue staining. The cross-linking reagent dimethyl pimelimidate was added and thermation of oligomeric forms in solution was determined at 0 (no addition), 15
nd 30 min (lanes 3 and 4 in each gel). (B) FPLC elution profile of ATMIN (378–823)a Serva Polyol Si-300 column in the presence or absence of a fivefold molar
xcess of DYNLL1. (C) Isothermal titration calorimetry of ATMIN (475–823) withdded DYNLL1. The upper panel corresponds to the thermogram and the bottomanel to the binding isotherm. Data are representative of three independentxperiments.
3. Results
3.1. Identification of ATMIN as a new binding partner of DYNLL1
We have screened a human heart cDNA library using DYNLL1 asa bait protein in a yeast two-hybrid system to find new potentialpartner proteins. By sequence analysis of 150 positive clones wehave identified ATMIN as a DYNLL1 interacting protein. Residues362–823 of ATMIN (corresponding to the carboxy-terminal halfof the molecule) were retrieved as a DYNLL1-binding region, a frag-ment that includes part of the core region as well as 11 SQ/TQ mo-tifs (Fig. 1A). Most of this part of ATMIN displayed long stretches ofpotentially disordered regions according to the PONDR software[21] (data not shown). Interestingly, the DYNLL1-interacting motifsvery frequently have sequences that include SQT or TQT triads andare located in disordered regions of protein partners [7,11,12]. Thisobservation raises the possibility that the DYNLL1 binding sitescould match with ATM/ATR phosphorylation sites. With that inmind, four additional ATMIN fragments covering residues 362–572, 467–572, 572–823 and 691–823 were confronted to DYNLL1in a yeast two-hybrid screen, and all of them rendered a positiveinteraction (Fig. 1A). Not only growth was achieved in the absenceof histidine but also a clear b-galactosidase positive reaction wasobserved (Fig. 1A). This means that all these four constructs in-clude SQ/TQ motifs that bind to DYNLL1 and indicates that verylikely multiple DYNLL1-binding sites are present within the ATMINsequence.
In agreement with our previous yeast two-hybrid fine mappingresult we obtained 11 positive DYNLL1 binding sites using a pep-scan approach (Fig. 1B), all of them with a Gln residue, frequentlyflanked by Thr residues. In fact, all previously predicted sequenceshad positive hits. Next, we inspected if these DYNLL1-binding mo-tifs within the ATMIN sequence were indeed conserved among var-ious mammalian ATMIN sequences. As shown in Fig. 1B, sequencealignment using the Clustal software revealed that these SQ/TQmotifs are extremely conserved, hence reinforcing a biologicalsignificance.
In order to evaluate if ATMIN and DYNLL1 bind to each other inmammalian cells, co-immunoprecipitation experiments were per-formed using anti-DYNLL1 antibodies and analyzing the presenceof ATMIN in the immunoprecipitated fraction. As shown inFig. 1C, anti-DYNLL1 antibodies, but not a pre-immune serum,
Fws(mbfoaineape
496 P. Rapali et al. / Biochemical and Biophysical Research Communications 414 (2011) 493–498
can immunoprecipitate ATMIN, hence indicating that the two pro-teins interact in a physiological environment.
3.2. Recombinant expression and characterization of ATMIN (378–823)
Next, we produced a recombinant ATMIN fragment consistingof residues 378–823 of ATMIN (corresponding to the domain en-riched in SQ/TQ motifs) in E. coli which migrated in a SDS–PAGEgel at approximately 66 kDa, whereas its theoretical molecularweight is �50.9 kDa (Fig. 2A). This is usually indicative of an intrin-sically disordered protein in solution [21]. We next decided to ana-lyze if ATMIN (378–823) was an oligomeric protein. Treatment ofATMIN (378–823) with crosslinking agents for 0, 15 or 30 min ren-dered no oligomers according to SDS–PAGE analysis (Fig. 2A, mid-dle panel). On the other hand, under identical treatmentconditions, the crosslinked protomers of DYNLL1, which are alwaysin dimeric form, could be clearly observed (Fig. 2B, right panel).
When analyzed using gel filtration (Fig. 2B) ATMIN (378–823)appeared at approximately 6.5 ml, corresponding to 363 kDa whenits elution volume was interpolated in a calibration curve (data notshown). Consequently, at this point we considered that ATMIN isable to adopt a non-globular or disordered conformation responsi-ble for its apparent large molecular weight. Addition of a five molarexcess of recombinant DYNLL1 to recombinant purified ATMIN(378–823) resulted in no displacement to larger apparent molecu-lar masses in gel filtration experiments (Fig. 2B), although a clearconcomitant decrease in the amount of free DYNLL1 could be ob-served. In addition, SDS–PAGE revealed that DYNLL1 bound tothe ATMIN (378–823) fractions that eluted at �6.5 ml (data notshown). This further proves DYNLL1 binding to ATMIN (378–823)and is also indicative that this binding does not induce a significantchange in the hydrodynamic behavior of the later.
A
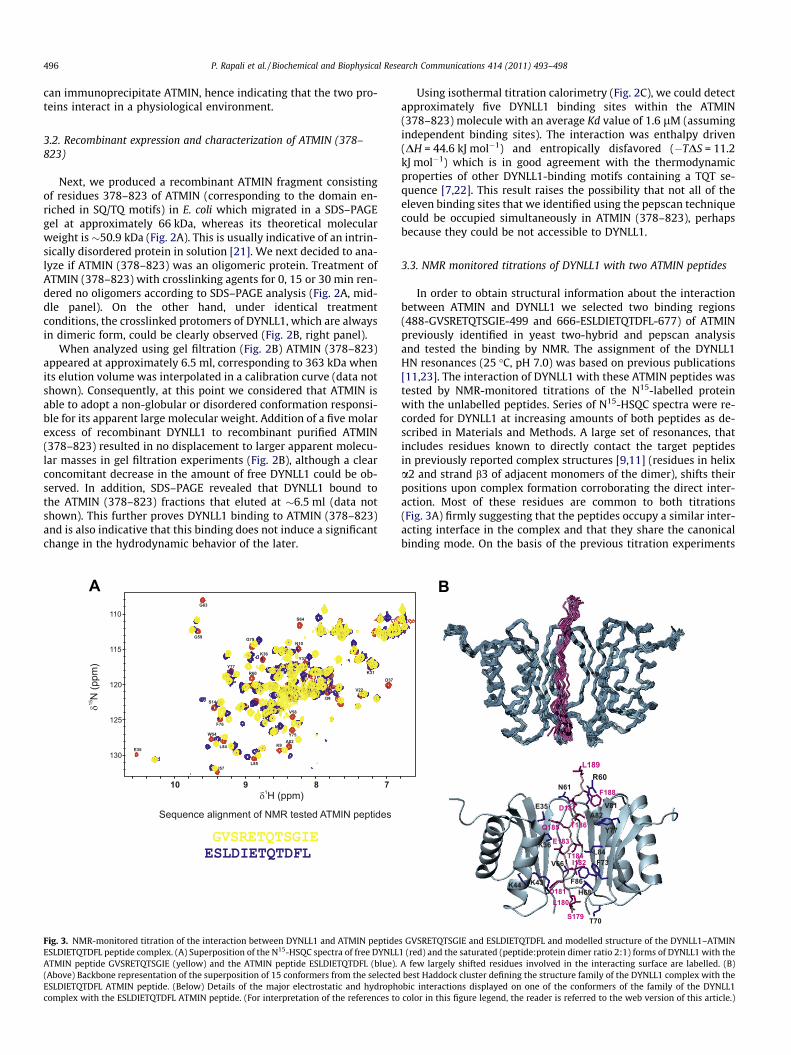
Fig. 3. NMR-monitored titration of the interaction between DYNLL1 and ATMIN peptideESLDIETQTDFL peptide complex. (A) Superposition of the N15-HSQC spectra of free DYNLLATMIN peptide GVSRETQTSGIE (yellow) and the ATMIN peptide ESLDIETQTDFL (blue).(Above) Backbone representation of the superposition of 15 conformers from the selectedESLDIETQTDFL ATMIN peptide. (Below) Details of the major electrostatic and hydrophcomplex with the ESLDIETQTDFL ATMIN peptide. (For interpretation of the references to
Using isothermal titration calorimetry (Fig. 2C), we could detectapproximately five DYNLL1 binding sites within the ATMIN(378–823) molecule with an average Kd value of 1.6 lM (assumingindependent binding sites). The interaction was enthalpy driven(DH = 44.6 kJ mol�1) and entropically disfavored (�TDS = 11.2kJ mol�1) which is in good agreement with the thermodynamicproperties of other DYNLL1-binding motifs containing a TQT se-quence [7,22]. This result raises the possibility that not all of theeleven binding sites that we identified using the pepscan techniquecould be occupied simultaneously in ATMIN (378–823), perhapsbecause they could be not accessible to DYNLL1.
3.3. NMR monitored titrations of DYNLL1 with two ATMIN peptides
In order to obtain structural information about the interactionbetween ATMIN and DYNLL1 we selected two binding regions(488-GVSRETQTSGIE-499 and 666-ESLDIETQTDFL-677) of ATMINpreviously identified in yeast two-hybrid and pepscan analysisand tested the binding by NMR. The assignment of the DYNLL1HN resonances (25 �C, pH 7.0) was based on previous publications[11,23]. The interaction of DYNLL1 with these ATMIN peptides wastested by NMR-monitored titrations of the N15-labelled proteinwith the unlabelled peptides. Series of N15-HSQC spectra were re-corded for DYNLL1 at increasing amounts of both peptides as de-scribed in Materials and Methods. A large set of resonances, thatincludes residues known to directly contact the target peptidesin previously reported complex structures [9,11] (residues in helixa2 and strand b3 of adjacent monomers of the dimer), shifts theirpositions upon complex formation corroborating the direct inter-action. Most of these residues are common to both titrations(Fig. 3A) firmly suggesting that the peptides occupy a similar inter-acting interface in the complex and that they share the canonicalbinding mode. On the basis of the previous titration experiments
B
s GVSRETQTSGIE and ESLDIETQTDFL and modelled structure of the DYNLL1–ATMIN1 (red) and the saturated (peptide:protein dimer ratio 2:1) forms of DYNLL1 with theA few largely shifted residues involved in the interacting surface are labelled. (B)best Haddock cluster defining the structure family of the DYNLL1 complex with the
obic interactions displayed on one of the conformers of the family of the DYNLL1color in this figure legend, the reader is referred to the web version of this article.)
P. Rapali et al. / Biochemical and Biophysical Research Communications 414 (2011) 493–498 497
we selected one of the ATMIN peptides (ESLDIETQTDFL) to modelthe structure of the complex with DYNLL1 since, as mentionedabove, a large overall similitude is expected for the complexes withboth tested peptides. Superposition of 15 conformers from the bestcluster that defines the family of structures of the DYNLL1–ATMINESLDIETQTDFL peptide complex is shown in Fig. 3B. The Haddockscore for this cluster is �129.2, the electrostatic contribution tothe energy �378 kcal/mol, and the buried surface area 1573 Å2.The orientation of the peptide is well-defined with an RMSD valueof 0.65 Å for the backbone, and 1.11 Å when the heavy atoms areincluded. Similarly to other complexes [7,8,15], the peptide runsparallel to the DYNLL1 binding groove extending the central b-sheet with and additional sixth strand establishing characteristicH-bond networks.
3.4. Subcellular localization of DYNLL1 and ATMIN: DYNLL1interference of ATMIN-dependent foci formation
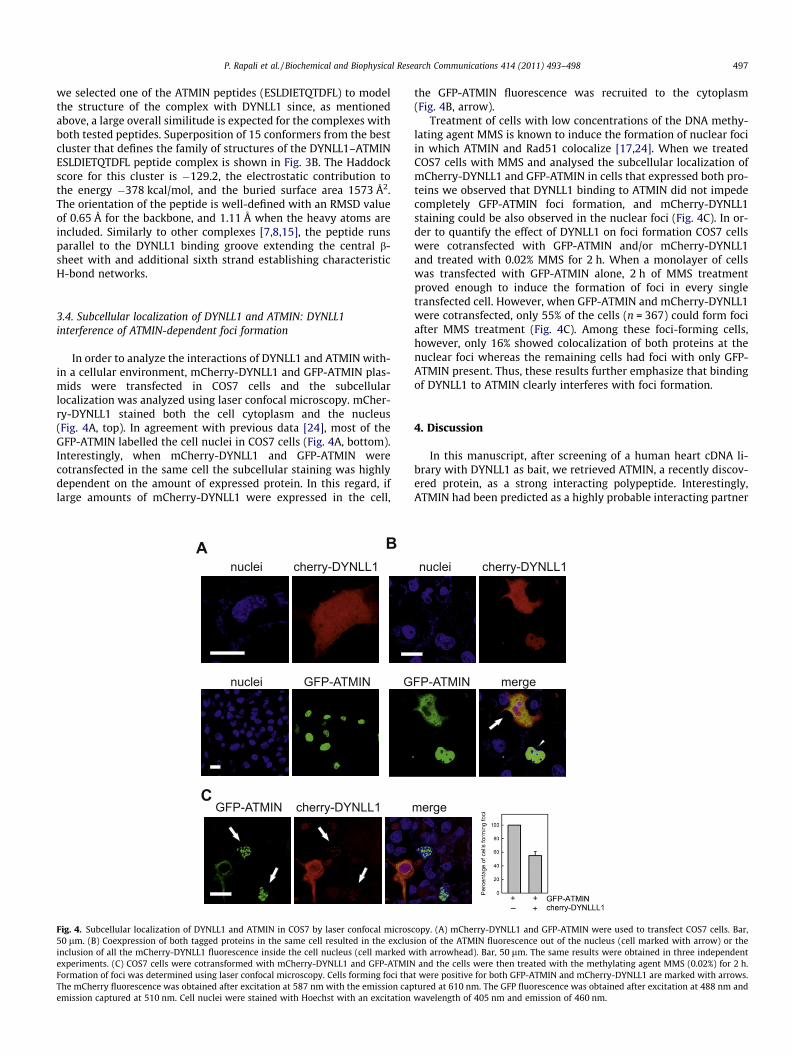
In order to analyze the interactions of DYNLL1 and ATMIN with-in a cellular environment, mCherry-DYNLL1 and GFP-ATMIN plas-mids were transfected in COS7 cells and the subcellularlocalization was analyzed using laser confocal microscopy. mCher-ry-DYNLL1 stained both the cell cytoplasm and the nucleus(Fig. 4A, top). In agreement with previous data [24], most of theGFP-ATMIN labelled the cell nuclei in COS7 cells (Fig. 4A, bottom).Interestingly, when mCherry-DYNLL1 and GFP-ATMIN werecotransfected in the same cell the subcellular staining was highlydependent on the amount of expressed protein. In this regard, iflarge amounts of mCherry-DYNLL1 were expressed in the cell,
nuclei cherry-DYNLL1
nuclei GFP-ATMIN
BA
CGFP-ATMIN cherry-DYNLL1
G
Fig. 4. Subcellular localization of DYNLL1 and ATMIN in COS7 by laser confocal micros50 lm. (B) Coexpression of both tagged proteins in the same cell resulted in the exclusinclusion of all the mCherry-DYNLL1 fluorescence inside the cell nucleus (cell marked wexperiments. (C) COS7 cells were cotransformed with mCherry-DYNLL1 and GFP-ATMINFormation of foci was determined using laser confocal microscopy. Cells forming foci thaThe mCherry fluorescence was obtained after excitation at 587 nm with the emission capemission captured at 510 nm. Cell nuclei were stained with Hoechst with an excitation
the GFP-ATMIN fluorescence was recruited to the cytoplasm(Fig. 4B, arrow).
Treatment of cells with low concentrations of the DNA methy-lating agent MMS is known to induce the formation of nuclear fociin which ATMIN and Rad51 colocalize [17,24]. When we treatedCOS7 cells with MMS and analysed the subcellular localization ofmCherry-DYNLL1 and GFP-ATMIN in cells that expressed both pro-teins we observed that DYNLL1 binding to ATMIN did not impedecompletely GFP-ATMIN foci formation, and mCherry-DYNLL1staining could be also observed in the nuclear foci (Fig. 4C). In or-der to quantify the effect of DYNLL1 on foci formation COS7 cellswere cotransfected with GFP-ATMIN and/or mCherry-DYNLL1and treated with 0.02% MMS for 2 h. When a monolayer of cellswas transfected with GFP-ATMIN alone, 2 h of MMS treatmentproved enough to induce the formation of foci in every singletransfected cell. However, when GFP-ATMIN and mCherry-DYNLL1were cotransfected, only 55% of the cells (n = 367) could form fociafter MMS treatment (Fig. 4C). Among these foci-forming cells,however, only 16% showed colocalization of both proteins at thenuclear foci whereas the remaining cells had foci with only GFP-ATMIN present. Thus, these results further emphasize that bindingof DYNLL1 to ATMIN clearly interferes with foci formation.
4. Discussion
In this manuscript, after screening of a human heart cDNA li-brary with DYNLL1 as bait, we retrieved ATMIN, a recently discov-ered protein, as a strong interacting polypeptide. Interestingly,ATMIN had been predicted as a highly probable interacting partner
merge
nuclei cherry-DYNLL1
FP-ATMIN merge
copy. (A) mCherry-DYNLL1 and GFP-ATMIN were used to transfect COS7 cells. Bar,ion of the ATMIN fluorescence out of the nucleus (cell marked with arrow) or theith arrowhead). Bar, 50 lm. The same results were obtained in three independentand the cells were then treated with the methylating agent MMS (0.02%) for 2 h.
t were positive for both GFP-ATMIN and mCherry-DYNLL1 are marked with arrows.tured at 610 nm. The GFP fluorescence was obtained after excitation at 488 nm andwavelength of 405 nm and emission of 460 nm.

498 P. Rapali et al. / Biochemical and Biophysical Research Communications 414 (2011) 493–498
of DYNLL [7]. ATMIN is a DNA damage response protein located inthe cell nucleus that colocalizes with the recombinase Rad51,though it does not interact directly with it [17,24]. ATM kinase isbelieved to respond primarily to DNA double strand breaks suchas those caused by ionizing radiation whereas ATR kinase is be-lieved to be mostly activated by single-stranded DNA gaps [25].Once activated, both kinases phosphorylate several hundred effec-tor proteins, in many cases in SQ or TQ motifs [26]. It is assumedthat these linear SQ/TQ motifs are generally located in intrinsicallydisordered regions of these effector molecules [17,27]. Since theSQ/TQ repeats are hallmarks of DNA damage response proteinsand potential substrates for the checkpoint kinases ATM and ATR[28] the behavior of ATMIN/ASCIZ was analyzed under conditionsof methylating DNA damage.
The simultaneous binding of multiple DYNLL1 or multipleDYNLL2 units to the same polypeptide chain is not without prece-dents. For instance, the DID domain of Nup159 displays six consec-utive binding sites for Dyn2 (the yeast dynein light chain), five ofwhich also bind in a pepscan analysis and in solution [29]. In addi-tion, two units of DYNLL1 or DYNLL2 can bind to two tandem mo-tifs in GKAP1 [12,30]. Using pepscan approaches, we have alsoshown previously that some other proteins, such as human adeno-virus-associated BS69 protein or human herpesvirus U19 displaytandem binding sites for DYNLL1 [10]. Finally, p53BP1 displaystwo sequential DYNLL1 binding motifs [31].
Whether all of these 11 sites are simultaneously occupiedalongside the ATMIN sequence remains to be established. In trans-fected cells DYNLL1 binding to ATMIN also interferes with foci for-mation after MMS treatment. It is then conceivable that ATMINphosphorylation by ATM/ATR kinases under conditions of DNAdamage might occur at SQ/TQ motifs that serve simultaneouslyas kinase phosphorylation spots and DYNLL1 binding sites. There-fore, our results indicate that DYNLL1 binding to certain cellulartargets interferes with ATM/ATR phosphorylation and vice versa.Moreover, this reciprocal regulation could be present in multiplecellular proteins.
Acknowledgments
This work was supported by Grants MICINN BFU2009-10442(I.R.C.), Consolider-centrosoma 3D (CSD2006-00023) (M.B.),CTQ2008-0080 (M.B.), OTKA NK81950 (L.N.), K81934 (K.S.), TÁMOP4.2.1./B-09/KMR-2010-0003. We would like to thank FernandoRoncal for synthesis of the pepscan cellulose membranes. We areindebted to Dr. Axel Behrens (Cancer Research Institute, London)for his generous gift of the ATMIN plasmid and Dr. József kardosfor his continuous advice. We also thank Dr. János Kovács andAndrás Patthy for preliminary EM experiments and in-housepeptide synthesis, respectively.
References
[1] S.E. Benashski, A. Harrison, R.S. Patel-King, S.M. King, Dimerization of thehighly conserved light chain shared by dynein and myosin V, J. Biol. Chem. 272(1997) 20929–20935.
[2] F.S. Espindola, D.M. Suter, L.B. Partata, T. Cao, J.S. Wolenski, R.E. Cheney, S.M.King, M.S. Mooseker, The light chain composition of chicken brain myosin-Va:calmodulin, myosin-II essential light chains, and 8-kDa dynein light chain/PIN,Cell. Motil. Cytoskelet. 47 (2000) 269–281.
[3] J. Merino-Gracia, M.F. Garcia-Mayoral, I. Rodriguez-Crespo, The association ofviral proteins with host cell dynein components during virus infection, FEBS J.278 (2011) 2997–3011.
[4] S.M. King, E. Barbarese, J.F. Dillman 3rd, R.S. Patel-King, J.H. Carson, K.K. Pfister,Brain cytoplasmic and flagellar outer arm dyneins share a highly conserved Mr8000 light chain, J. Biol. Chem. 271 (1996) 19358–19366.
[5] E. Barbar, Dynein light chain LC8 is a dimerization hub essential in diverseprotein networks, Biochemistry 47 (2008) 503–508.
[6] Z.R. Hodi, P. Radnai, L. Molnar, T. Szenes, A. Kardos, J. Buday, L. Stafford, W.F., L.Nyitray, The LC8 family of dynein light chains: multifunctional chaperon-likeproteins, FEBS J. 274 (2007) 106 (Suppl. 1 Meeting Abstract: B1-372007).
[7] P. Rapali, L. Radnai, D. Suveges, V. Harmat, F. Tolgyesi, W.Y. Wahlgren, G.Katona, L. Nyitray, G. Pal, Directed evolution reveals the binding motifpreference of the LC8/DYNLL hub protein and predicts large numbers ofnovel binders in the human proteome, PLoS One 6 (2011) e18818.
[8] P. Rapali, A. Szenes, L. Radnai, A. Bakos, G. Pal, L. Nyitray, DYNLL/LC8: a lightchain subunit of the dynein motor complex and beyond, FEBS J. 278 (2011)2980–2996.
[9] G. Benison, P.A. Karplus, E. Barbar, Structure and dynamics of LC8 complexeswith KXTQT-motif peptides: swallow and dynein intermediate chain competefor a common site, J. Mol. Biol. 371 (2007) 457–468.
[10] M. Martinez-Moreno, I. Navarro-Lerida, F. Roncal, J.P. Albar, C. Alonso, F.Gavilanes, I. Rodriguez-Crespo, Recognition of novel viral sequences thatassociate with the dynein light chain LC8 identified through a pepscantechnique, FEBS Lett. 544 (2003) 262–267.
[11] J. Fan, Q. Zhang, H. Tochio, M. Li, M. Zhang, Structural basis of diversesequence-dependent target recognition by the 8 kDa dynein light chain, J. Mol.Biol. 306 (2001) 97–108.
[12] I. Rodriguez-Crespo, B. Yelamos, F. Roncal, J.P. Albar, P.R. Ortiz deMontellano, F. Gavilanes, Identification of novel cellular proteins that bindto the LC8 dynein light chain using a pepscan technique, FEBS Lett. 503(2001) 135–141.
[13] K.W. Lo, S. Naisbitt, J.S. Fan, M. Sheng, M. Zhang, The 8-kDa dynein light chainbinds to its targets via a conserved (K/R)XTQT motif, J. Biol. Chem. 276 (2001)14059–14066.
[14] M.F. Garcia-Mayoral, I. Rodriguez-Crespo, M. Bruix, Structural models ofDYNLL1 with interacting partners: African swine fever virus protein p54 andpostsynaptic scaffolding protein gephyrin, FEBS Lett. 585 (2011) 53–57.
[15] L. Radnai, P. Rapali, Z. Hodi, D. Suveges, T. Molnar, B. Kiss, B. Becsi, F. Erdodi, L.Buday, J. Kardos, M. Kovacs, L. Nyitray, Affinity, avidity and kinetics of targetsequence binding to LC8 dynein light chain isoforms, J. Biol. Chem. 285 (2010)38649–38657.
[16] J.C. Williams, P.L. Roulhac, A.G. Roy, R.B. Vallee, M.C. Fitzgerald, W.A.Hendrickson, Structural and thermodynamic characterization of acytoplasmic dynein light chain-intermediate chain complex, Proc. Natl. Acad.Sci. USA 104 (2007) 10028–10033.
[17] N. Kanu, A. Behrens, ATMIN defines an NBS1-independent pathway of ATMsignalling, EMBO J. 26 (2007) 2933–2941.
[18] I. Rodriguez-Crespo, W. Straub, F. Gavilanes, P.R. Ortiz de Montellano, Bindingof dynein light chain (PIN) to neuronal nitric oxide synthase in the absence ofinhibition, Arch. Biochem. Biophys. 359 (1998) 297–304.
[19] M.F. Garcia-Mayoral, M. Martinez-Moreno, J.P. Albar, I. Rodriguez-Crespo, M.Bruix, Structural basis for the interaction between dynein light chain 1 and theglutamate channel homolog GRINL1A, FEBS J. 277 (2010) 2340–2350.
[20] I. Navarro-Lerida, M. Martinez-Moreno, I. Ventoso, A. Alvarez-Barrientos, I.Rodriguez-Crespo, Binding of CAP70 to inducible nitric oxide synthase andimplications for the vectorial release of nitric oxide in polarized cells, Mol. Biol.Cell 18 (2007) 2768–2777.
[21] P. Romero, Z. Obradovic, X. Li, E.C. Garner, C.J. Brown, A.K. Dunker, Sequencecomplexity of disordered protein, Proteins 42 (2001) 38–48.
[22] J. Hall, A. Hall, N. Pursifull, E. Barbar, Differences in dynamic structure of LC8monomer, dimer, and dimer–peptide complexes, Biochemistry 47 (2008)11940–11952.
[23] W. Wang, K.W. Lo, H.M. Kan, J.S. Fan, M. Zhang, Structure of the monomeric 8-kDa dynein light chain and mechanism of the domain-swapped dimerassembly, J. Biol. Chem. 278 (2003) 41491–41499.
[24] C.J. McNees, L.A. Conlan, N. Tenis, J. Heierhorst, ASCIZ regulates lesion-specificRad51 focus formation and apoptosis after methylating DNA damage, EMBO J.24 (2005) 2447–2457.
[25] K. Hiom, DNA repair: how to PIKK a partner, Curr. Biol. 15 (2005) R473–R475.[26] S. Matsuoka, B.A. Ballif, A. Smogorzewska, E.R. McDonald 3rd, K.E. Hurov, J.
Luo, C.E. Bakalarski, Z. Zhao, N. Solimini, Y. Lerenthal, Y. Shiloh, S.P. Gygi, S.J.Elledge, ATM and ATR substrate analysis reveals extensive protein networksresponsive to DNA damage, Science 316 (2007) 1160–1166.
[27] J. Heierhorst, Mdt1/ASCIZ: a new DNA damage response protein family, CellCycle 7 (2008) 2654–2660.
[28] A. Traven, J. Heierhorst, SQ/TQ cluster domains: concentrated ATM/ATR kinasephosphorylation site regions in DNA-damage-response proteins, Bioessays 27(2005) 397–407.
[29] P. Stelter, R. Kunze, D. Flemming, D. Hopfner, M. Diepholz, P. Philippsen, B.Bottcher, E. Hurt, Molecular basis for the functional interaction of dynein lightchain with the nuclear-pore complex, Nat. Cell Biol. 9 (2007) 788–796.
[30] S. Naisbitt, J. Valtschanoff, D.W. Allison, C. Sala, E. Kim, A.M. Craig, R.J.Weinberg, M. Sheng, Interaction of the postsynaptic density-95/guanylatekinase domain-associated protein complex with a light chain of myosin-V anddynein, J. Neurosci. 20 (2000) 4524–4534.
[31] K.W. Lo, H.M. Kan, L.N. Chan, W.G. Xu, K.P. Wang, Z. Wu, M. Sheng, M. Zhang,The 8-kDa dynein light chain binds to p53-binding protein 1 and mediatesDNA damage-induced p53 nuclear accumulation, J. Biol. Chem. 280 (2005)8172–8179.
Related Documents











