LATERAL FLORET 1 induced the three-florets spikelet in rice Ting Zhang a,1 , Yunfeng Li a,1,2 , Ling Ma a , Xianchun Sang a , Yinghua Ling a , Yantong Wang a , Peng Yu a , Hui Zhuang a , Junyang Huang a , Nan Wang a , Fangming Zhao a , Changwei Zhang a , Zhenglin Yang a , Likui Fang a , and Guanghua He a,2 a Rice Research Institute, Key Laboratory of Application and Safety Control of Genetically Modified Crops, Academy of Agricultural Sciences, Southwest University, Chongqing 400715, China Edited by Jiayang Li, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China, and approved August 4, 2017 (received for review January 11, 2017) The spikelet is a unique inflorescence structure in grass. The molecular mechanisms behind the development and evolution of the spikelet are far from clear. In this study, a dominant rice mutant, lateral florets 1 (lf1), was characterized. In the lf1 spikelet, lateral floral meristems were promoted unexpectedly and could generally blossom into relatively normal florets. LF1 encoded a class III homeodomain-leucine zipper (HD-ZIP III) protein, and the site of mutation in lf1 was located in a putative miRNA165/166 target sequence. Ectopic expression of both LF1 and the meristem maintenance gene OSH1 was detected in the axil of the sterile lemma primordia of the lf1 spikelet. Furthermore, the promoter of OSH1 could be bound directly by LF1 protein. Collectively, these results indicate that the mutation of LF1 induces ectopic expres- sion of OSH1, which results in the initiation of lateral meristems to generate lateral florets in the axil of the sterile lemma. This study thus offers strong evidence in support of the “three-florets spike- let” hypothesis in rice. lateral floret | three-florets spikelet | evolution | yield | rice F lower development is a key process in the reproduction of angiosperms. Under suitable conditions, flowering signals are transmitted to shoot apical meristems (SAMs), which are transformed first into inflorescence meristems (IMs). Floral meristems (FMs) are then initiated on the top and/or lateral domains of the IMs and subsequently transformed into the four whorls of floral organs. The spikelet is a unique unit of in- florescence architecture in grasses and consists of a pair of glumes and a fixed or variable number of florets. Some grass- specific genes are involved in regulating spikelet development. For example, FRIZZY PANICLE (FZP) functions in regulating spikelet meristem (SM) identity in rice. In the fzp mutant, axil- lary meristems (AMs) are formed instead of FMs, and these then develop into higher-order branches (1). Three genes that encode members of the AP2/ERF superfamily, SUPERNUMERARY BRACT (SNB), INDETERMINATE SPIKELET 1 (OsIDS1), and MULTI-FLORETS SPIKELET 1 (MFS1), are involved in regu- lating spikelet determinacy in rice (2, 3). In these mutants, the transition from SM to FM is delayed, and extra organs or florets are produced. However, our knowledge about the details of spikelet development in rice remains limited. In most members of Oryzeae, the spikelet is composed of one pair of rudimentary glumes, one pair of sterile lemmas, and one terminal fertile floret, which consists of one pair of hulls (lemma and palea) and inner floral organs (4). Regarding the origin of the sterile lemmas, the “three-florets spikelet” hypothesis pro- poses that the putative ancestor of the rice spikelet contained two lateral florets in addition to a terminal fertile floret. Sub- sequently, the lemmas of the two lateral florets degenerated into sterile lemmas, and the inner floral organs and palea degen- erated markedly and disappeared during evolution (5). In recent years, several reports have supported this hypothesis. The genes LONG STERILE LEMMA (G1), EXTRA GLUME 1 (EG1), PANICLE PHYTOMER 2 (PAP2)/ OsMADS34, NONSTOP GLUMES (NSG), and ABERRANT SPIKELET AND PANICLE 1 (ASP1) are involved in the regulation of sterile lemma identity. Loss of function of these genes results in lemma-like sterile lemmas (6–10). In addition, ectopic expression of OsMADS1 causes lemma-like sterile lemmas (11). All these results suggest that the sterile lemma is homologous to the lemma. However, there is no direct evidence that these lemma-like organs are derived from the lemmas of two lateral florets, because no lateral florets with inner floral organs have been observed in these mutants. Given the lack of further evidence, the three-florets spikelet hypothesis remains widely debated. In this study, we characterized a gain-of-function mutant, lat- eral florets 1 (lf1), in which the lateral florets were surprisingly formed in the axil of the sterile lemmas. By map-based cloning, we isolated the LF1 gene, which encodes a homeodomain- leucine zipper class III (HD-ZIP III) transcriptional activator. In lf1, LF1 was expressed ectopically because miRNA165/166 could not act on the mutated LF1 mRNA. LF1 then directly activated the expression of the meristem maintenance gene OSH1 to induce the initiation of lateral FMs. Interestingly, the results of the study provide strong evidence in support of the three-florets spikelet hypothesis in rice. Results The lf1 Spikelet Developed Lateral Florets. The WT rice spikelet consisted of one pair of rudimentary glumes, one pair of sterile lemmas, and one terminal fertile floret (Fig. 1 A, 1). The Significance In cereal crops, the number of florets in a spikelet is an im- portant factor affecting the grain number per panicle and then the grain yield. In wild-type rice, one spikelet produces one fertile floret. This study characterized a gain-of-function mu- tant lateral florets 1 (lf1) in rice. In lf1, the spikelet developed lateral florets with proper floral organ identities in the axil of the sterile lemma, showing that the rice spikelet has the po- tential to restore the “three-florets spikelet” which may have existed in ancestors. Therefore, it provides strong evidence supporting the three-florets spikelet hypothesis and presents a prospect for increasing grain number per panicle by breeding rice with three-floret spikelets. Author contributions: T.Z., Y. Li, and G.H. designed research; T.Z., Y. Li, L.M., X.S., Y. Ling, Y.W., P.Y., H.Z., and J.H. performed research; T.Z., Y. Li, F.Z., C.Z., Z.Y., and L.F. contributed new reagents/analytic tools; T.Z., Y. Li, X.S., Y. Ling, N.W., C.Z., and Z.Y. analyzed data; and T.Z., Y. Li, and G.H. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. Freely available online through the PNAS open access option. 1 T.Z. and Y. Li contributed equally to this work. 2 To whom correspondence may be addressed. Email: [email protected] or liyf1980@swu. edu.cn. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1700504114/-/DCSupplemental. 9984–9989 | PNAS | September 12, 2017 | vol. 114 | no. 37 www.pnas.org/cgi/doi/10.1073/pnas.1700504114

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

LATERAL FLORET 1 induced the three-florets spikeletin riceTing Zhanga,1, Yunfeng Lia,1,2, Ling Maa, Xianchun Sanga, Yinghua Linga, Yantong Wanga, Peng Yua, Hui Zhuanga,Junyang Huanga, Nan Wanga, Fangming Zhaoa, Changwei Zhanga, Zhenglin Yanga, Likui Fanga, and Guanghua Hea,2
aRice Research Institute, Key Laboratory of Application and Safety Control of Genetically Modified Crops, Academy of Agricultural Sciences, SouthwestUniversity, Chongqing 400715, China
Edited by Jiayang Li, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China, and approved August 4, 2017 (receivedfor review January 11, 2017)
The spikelet is a unique inflorescence structure in grass. Themolecular mechanisms behind the development and evolution ofthe spikelet are far from clear. In this study, a dominant ricemutant, lateral florets 1 (lf1), was characterized. In the lf1 spikelet,lateral floral meristems were promoted unexpectedly and couldgenerally blossom into relatively normal florets. LF1 encoded aclass III homeodomain-leucine zipper (HD-ZIP III) protein, and thesite of mutation in lf1 was located in a putative miRNA165/166target sequence. Ectopic expression of both LF1 and the meristemmaintenance gene OSH1 was detected in the axil of the sterilelemma primordia of the lf1 spikelet. Furthermore, the promoterof OSH1 could be bound directly by LF1 protein. Collectively, theseresults indicate that the mutation of LF1 induces ectopic expres-sion of OSH1, which results in the initiation of lateral meristems togenerate lateral florets in the axil of the sterile lemma. This studythus offers strong evidence in support of the “three-florets spike-let” hypothesis in rice.
lateral floret | three-florets spikelet | evolution | yield | rice
Flower development is a key process in the reproduction ofangiosperms. Under suitable conditions, flowering signals
are transmitted to shoot apical meristems (SAMs), which aretransformed first into inflorescence meristems (IMs). Floralmeristems (FMs) are then initiated on the top and/or lateraldomains of the IMs and subsequently transformed into the fourwhorls of floral organs. The spikelet is a unique unit of in-florescence architecture in grasses and consists of a pair ofglumes and a fixed or variable number of florets. Some grass-specific genes are involved in regulating spikelet development.For example, FRIZZY PANICLE (FZP) functions in regulatingspikelet meristem (SM) identity in rice. In the fzp mutant, axil-lary meristems (AMs) are formed instead of FMs, and these thendevelop into higher-order branches (1). Three genes that encodemembers of the AP2/ERF superfamily, SUPERNUMERARYBRACT (SNB), INDETERMINATE SPIKELET 1 (OsIDS1), andMULTI-FLORETS SPIKELET 1 (MFS1), are involved in regu-lating spikelet determinacy in rice (2, 3). In these mutants, thetransition from SM to FM is delayed, and extra organs or floretsare produced. However, our knowledge about the details ofspikelet development in rice remains limited.In most members of Oryzeae, the spikelet is composed of one
pair of rudimentary glumes, one pair of sterile lemmas, and oneterminal fertile floret, which consists of one pair of hulls (lemmaand palea) and inner floral organs (4). Regarding the origin ofthe sterile lemmas, the “three-florets spikelet” hypothesis pro-poses that the putative ancestor of the rice spikelet containedtwo lateral florets in addition to a terminal fertile floret. Sub-sequently, the lemmas of the two lateral florets degenerated intosterile lemmas, and the inner floral organs and palea degen-erated markedly and disappeared during evolution (5). In recentyears, several reports have supported this hypothesis. The genesLONG STERILE LEMMA (G1), EXTRA GLUME 1 (EG1),PANICLE PHYTOMER 2 (PAP2)/OsMADS34, NONSTOP
GLUMES (NSG), and ABERRANT SPIKELET AND PANICLE1 (ASP1) are involved in the regulation of sterile lemma identity.Loss of function of these genes results in lemma-like sterilelemmas (6–10). In addition, ectopic expression of OsMADS1causes lemma-like sterile lemmas (11). All these results suggestthat the sterile lemma is homologous to the lemma. However,there is no direct evidence that these lemma-like organs arederived from the lemmas of two lateral florets, because no lateralflorets with inner floral organs have been observed in thesemutants. Given the lack of further evidence, the three-floretsspikelet hypothesis remains widely debated.In this study, we characterized a gain-of-function mutant, lat-
eral florets 1 (lf1), in which the lateral florets were surprisinglyformed in the axil of the sterile lemmas. By map-based cloning,we isolated the LF1 gene, which encodes a homeodomain-leucine zipper class III (HD-ZIP III) transcriptional activator.In lf1, LF1 was expressed ectopically because miRNA165/166could not act on the mutated LF1 mRNA. LF1 then directlyactivated the expression of the meristem maintenance geneOSH1 to induce the initiation of lateral FMs. Interestingly, theresults of the study provide strong evidence in support of thethree-florets spikelet hypothesis in rice.
ResultsThe lf1 Spikelet Developed Lateral Florets. The WT rice spikeletconsisted of one pair of rudimentary glumes, one pair of sterilelemmas, and one terminal fertile floret (Fig. 1 A, 1). The
Significance
In cereal crops, the number of florets in a spikelet is an im-portant factor affecting the grain number per panicle and thenthe grain yield. In wild-type rice, one spikelet produces onefertile floret. This study characterized a gain-of-function mu-tant lateral florets 1 (lf1) in rice. In lf1, the spikelet developedlateral florets with proper floral organ identities in the axil ofthe sterile lemma, showing that the rice spikelet has the po-tential to restore the “three-florets spikelet” which may haveexisted in ancestors. Therefore, it provides strong evidencesupporting the three-florets spikelet hypothesis and presents aprospect for increasing grain number per panicle by breedingrice with three-floret spikelets.
Author contributions: T.Z., Y. Li, and G.H. designed research; T.Z., Y. Li, L.M., X.S., Y. Ling,Y.W., P.Y., H.Z., and J.H. performed research; T.Z., Y. Li, F.Z., C.Z., Z.Y., and L.F. contributednew reagents/analytic tools; T.Z., Y. Li, X.S., Y. Ling, N.W., C.Z., and Z.Y. analyzed data;and T.Z., Y. Li, and G.H. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.1T.Z. and Y. Li contributed equally to this work.2To whom correspondence may be addressed. Email: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1700504114/-/DCSupplemental.
9984–9989 | PNAS | September 12, 2017 | vol. 114 | no. 37 www.pnas.org/cgi/doi/10.1073/pnas.1700504114

rudimentary glumes and sterile lemmas were inserted on therachilla in an alternate phyllotaxy, whereas the terminal floretwas produced on the top of the rachilla (Fig. 1 A, 1). The ter-minal floret consisted of four whorls of floral organs: one pistil inwhorl 4; six stamens in whorl 3; two lodicules at the lemma sidein whorl 2, and a lemma and palea that surround the inner floralorgans in whorl 1 (Fig. 1A and Fig. S1C). The sterile lemmaswere two small white glume-like organs with a smooth surface onthe abaxial side (Fig. 1 A, 2–4). The two rudimentary glumessubtended below the sterile lemmas and were degenerate withabundant trichomes on the abaxial side (Fig. 1 A, 2 and 3).In the lf1 spikelet, no significant abnormalities were detected
in the terminal floret and rudimentary glumes (Fig. 1 B, 1 and 2and C, 1 and 2 and Fig. S1 A, 1–3). However, the lateral floretscontained inner floral organs that grew obliquely in the axil ofthe upper and/or lower sterile lemma in an alternate phyllotaxyin the lf1 spikelet (Fig. 1 B and C). The statistical analysis showedthat 78% of lf1 spikelets developed one lateral floret only in theaxil of the upper sterile lemma, and 4% developed two lateralflorets in the axils of both the upper and lower sterile lemmas(Fig. S1B). In these lateral florets, all types of inner floral organswere discovered. The pistil with an ovary and bifid stigmas wassame as that in the WT. The transparent and oval-shaped lodi-cules and the yellow stamens with anthers growing in the slenderfilaments were very similar to those in the WT, although theirnumbers were decreased in some lateral florets. The palea,which had three vascular bundles that consisted of the body ofthe palea (bop) and two marginal regions (mrps), was similar tothose in the WT, even though some of them showed abnormalshapes (Fig. S1 C and D). Furthermore, known genes for floralorgan identity were expressed at high levels in the WT terminalfloret and lf1 lateral floret, whereas almost no expression wasdetected in the WT sterile lemma by qRT-PCR (Fig. S1E).
These genes included the lemma and palea identity geneOsMADS1 (12), lodicule and stamen identity genes OsMADS4and OsMADS16 (13, 14), stamen and carpel identity genesOsMADS3 and OsMADS58 (15), pistil identity gene DROOPINGLEAF (DL) (16), and the ovule identity gene OsMADS13 (17).Therefore, these results indicated that the identities of the floralorgans were specified correctly in lf1 lateral florets, althoughdefects in the shape and number of floral organs were observedin some lateral florets. They also suggested that the reversion tolateral florets occurred in the lf1 spikelet. Unfortunately, nosterile lemmas were transformed into lemmas, and in fact someeven degenerated further in some spikelets (Fig. 1 B and C).
The lf1 Mutation Induced Lateral FMs During Early SpikeletDevelopment. In the WT, the primordia of the rudimentaryglumes, sterile lemmas, lemma, and palea had formed or initi-ated in an alternate phyllotaxis during spikelet stage 4 (Sp4) (Fig.2A). During stages Sp5–Sp8 the inner floral organs developed inthe following order: two lodicules developed first, followed by sixstamens and a pistil (Fig. 2 B–D). In lf1, no significant differencewas observed during stage Sp4 compared with the WT (Fig. 2E).However, during stage Sp5, when lodicule and stamen primordiawere forming in the terminal floret, the lateral FM became vis-ible as a protrusion at the axil of the sterile lemma primordia inlf1 (Fig. 2F). During approximately stages Sp6/Sp7, when thepistil primordium was forming in the terminal florets, the for-mation of the palea primordium was completed in the lateralflorets (Fig. 2G). The initiation of the inner floral organ pri-mordia, e.g., the stamen and pistil, was not finally finished in thelateral florets until stage Sp8, after the lemma and palea hadclosed in the terminal floret (Fig. 2H). Therefore, the de-velopmental stage of the lateral florets was delayed by approxi-mately two spikelet stages compared with the terminal floret(Fig. 2). This delay might be caused by the florets in the three-florets spikelet developing in acropetal succession (18). In ad-dition, degradation of the lf1 sterile lemma was observed duringstages Sp4–Sp8 (Fig. S2).To confirm the identity of the organs in lf1 lateral florets
during the early developmental stages of the spikelet, the ex-pression of OsMADS16 and DL was investigated by in situ hy-bridization. Expression of OsMADS16 was detected in thelodicule and stamen of terminal florets in both the WT and lf1during stages Sp4–Sp8 (Fig. S3 A and B). High levels of ex-pression were also detected in the lodicule and stamen of lf1lateral florets during stages Sp5–Sp8 (Fig. S3 B, 2–4). In the
Fig. 1. Phenotypes of spikelets in the WT and lf1. (A) WT spikelets. (B and C)lf1 spikelets. (A1, B1, and C1) Whole spikelets. (A2, B2, and C2) Scanningelectron micrographs of the lower part of spikelets. (A3, B3, and C3) Lon-gitudinal sections of spikelets. (A4, B4, and C4) Transverse sections ofspikelets. Red dotted lines represent lateral florets; le, lemma; lf, lateralfloret; lo, lodicule; pa, palea; pi, pistil; rg, rudimentary glume; sl, sterilelemma; st, stamen. (Scale bars, 500 μm.)
Fig. 2. Scanning electron micrographs of spikelets at early developmentalstages in the WT and lf1. (A–D) WT spikelets. (E–H) lf1 spikelets. (A and E)Stage Sp4. (B and F) Stage Sp5. (C and G) Stage Sp6–Sp7. (D and H) Stage Sp8.Stars indicate stamens. fm, floral meristem; other abbreviations are as in Fig.1. (Scale bars, 100 μm.)
Zhang et al. PNAS | September 12, 2017 | vol. 114 | no. 37 | 9985
PLANTBIOLO
GY
terminal florets of the WT and lf1, DL was expressed first inlemma primordia during stages Sp4–Sp6 (Fig. S3 C, 1 and D, 1)and then was expressed in pistil primordia during stages Sp7–Sp8(Fig. S3 C, 2–4 and D, 2–4). Ectopic expression of DL was alsodetected in the lateral florets of lf1 during stages Sp6–Sp8 (Fig. S3D, 2–4). In conclusion, the initiation of FMs and subsequent for-mation of the primordia of the floral organs in lateral florets oc-curred during the early developmental stages of spikelets in lf1.
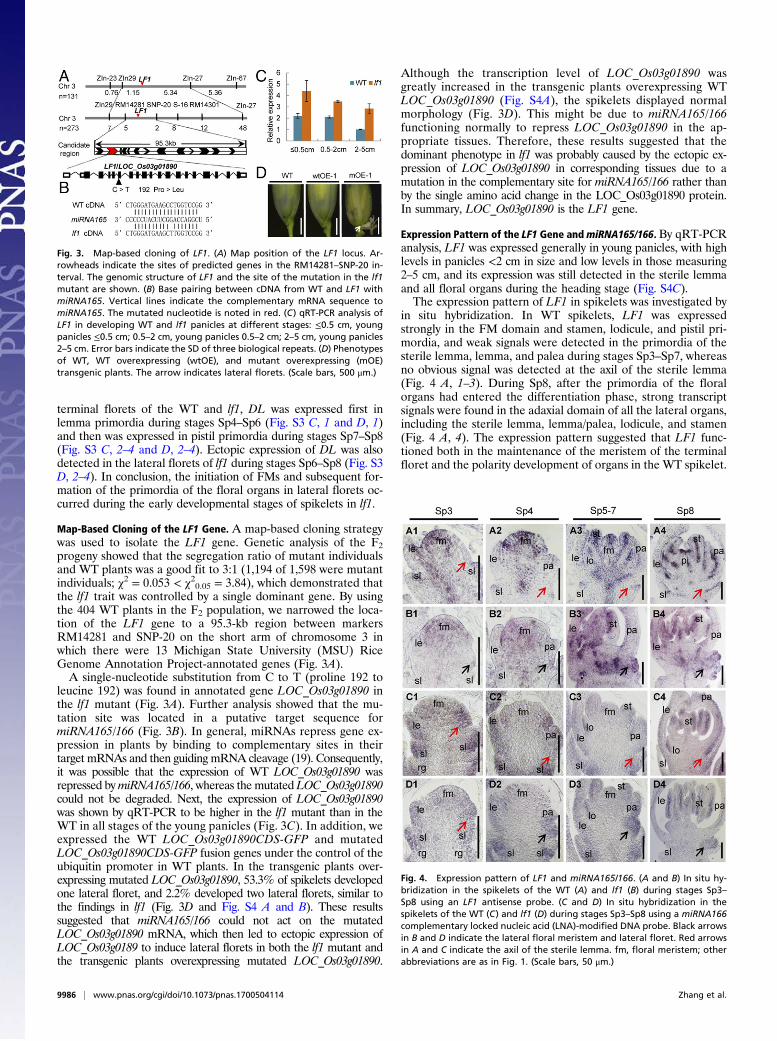
Map-Based Cloning of the LF1 Gene. A map-based cloning strategywas used to isolate the LF1 gene. Genetic analysis of the F2progeny showed that the segregation ratio of mutant individualsand WT plants was a good fit to 3:1 (1,194 of 1,598 were mutantindividuals; χ2 = 0.053 < χ20.05 = 3.84), which demonstrated thatthe lf1 trait was controlled by a single dominant gene. By usingthe 404 WT plants in the F2 population, we narrowed the loca-tion of the LF1 gene to a 95.3-kb region between markersRM14281 and SNP-20 on the short arm of chromosome 3 inwhich there were 13 Michigan State University (MSU) RiceGenome Annotation Project-annotated genes (Fig. 3A).A single-nucleotide substitution from C to T (proline 192 to
leucine 192) was found in annotated gene LOC_Os03g01890 inthe lf1 mutant (Fig. 3A). Further analysis showed that the mu-tation site was located in a putative target sequence formiRNA165/166 (Fig. 3B). In general, miRNAs repress gene ex-pression in plants by binding to complementary sites in theirtarget mRNAs and then guiding mRNA cleavage (19). Consequently,it was possible that the expression of WT LOC_Os03g01890 wasrepressed bymiRNA165/166, whereas the mutated LOC_Os03g01890could not be degraded. Next, the expression of LOC_Os03g01890was shown by qRT-PCR to be higher in the lf1 mutant than in theWT in all stages of the young panicles (Fig. 3C). In addition, weexpressed the WT LOC_Os03g01890CDS-GFP and mutatedLOC_Os03g01890CDS-GFP fusion genes under the control of theubiquitin promoter in WT plants. In the transgenic plants over-expressing mutated LOC_Os03g01890, 53.3% of spikelets developedone lateral floret, and 2.2% developed two lateral florets, similar tothe findings in lf1 (Fig. 3D and Fig. S4 A and B). These resultssuggested that miRNA165/166 could not act on the mutatedLOC_Os03g01890 mRNA, which then led to ectopic expression ofLOC_Os03g0189 to induce lateral florets in both the lf1 mutant andthe transgenic plants overexpressing mutated LOC_Os03g01890.
Although the transcription level of LOC_Os03g01890 wasgreatly increased in the transgenic plants overexpressing WTLOC_Os03g01890 (Fig. S4A), the spikelets displayed normalmorphology (Fig. 3D). This might be due to miRNA165/166functioning normally to repress LOC_Os03g01890 in the ap-propriate tissues. Therefore, these results suggested that thedominant phenotype in lf1 was probably caused by the ectopic ex-pression of LOC_Os03g01890 in corresponding tissues due to amutation in the complementary site for miRNA165/166 rather thanby the single amino acid change in the LOC_Os03g01890 protein.In summary, LOC_Os03g01890 is the LF1 gene.
Expression Pattern of the LF1 Gene andmiRNA165/166. By qRT-PCRanalysis, LF1 was expressed generally in young panicles, with highlevels in panicles <2 cm in size and low levels in those measuring2–5 cm, and its expression was still detected in the sterile lemmaand all floral organs during the heading stage (Fig. S4C).The expression pattern of LF1 in spikelets was investigated by
in situ hybridization. In WT spikelets, LF1 was expressedstrongly in the FM domain and stamen, lodicule, and pistil pri-mordia, and weak signals were detected in the primordia of thesterile lemma, lemma, and palea during stages Sp3–Sp7, whereasno obvious signal was detected at the axil of the sterile lemma(Fig. 4 A, 1–3). During Sp8, after the primordia of the floralorgans had entered the differentiation phase, strong transcriptsignals were found in the adaxial domain of all the lateral organs,including the sterile lemma, lemma/palea, lodicule, and stamen(Fig. 4 A, 4). The expression pattern suggested that LF1 func-tioned both in the maintenance of the meristem of the terminalfloret and the polarity development of organs in the WT spikelet.
Fig. 4. Expression pattern of LF1 and miRNA165/166. (A and B) In situ hy-bridization in the spikelets of the WT (A) and lf1 (B) during stages Sp3–Sp8 using an LF1 antisense probe. (C and D) In situ hybridization in thespikelets of the WT (C) and lf1 (D) during stages Sp3–Sp8 using a miRNA166complementary locked nucleic acid (LNA)-modified DNA probe. Black arrowsin B and D indicate the lateral floral meristem and lateral floret. Red arrowsin A and C indicate the axil of the sterile lemma. fm, floral meristem; otherabbreviations are as in Fig. 1. (Scale bars, 50 μm.)
Fig. 3. Map-based cloning of LF1. (A) Map position of the LF1 locus. Ar-rowheads indicate the sites of predicted genes in the RM14281–SNP-20 in-terval. The genomic structure of LF1 and the site of the mutation in the lf1mutant are shown. (B) Base pairing between cDNA from WT and LF1 withmiRNA165. Vertical lines indicate the complementary mRNA sequence tomiRNA165. The mutated nucleotide is noted in red. (C) qRT-PCR analysis ofLF1 in developing WT and lf1 panicles at different stages: ≤0.5 cm, youngpanicles ≤0.5 cm; 0.5–2 cm, young panicles 0.5–2 cm; 2–5 cm, young panicles2–5 cm. Error bars indicate the SD of three biological repeats. (D) Phenotypesof WT, WT overexpressing (wtOE), and mutant overexpressing (mOE)transgenic plants. The arrow indicates lateral florets. (Scale bars, 500 μm.)
9986 | www.pnas.org/cgi/doi/10.1073/pnas.1700504114 Zhang et al.

Likewise, although the expression of the LF1 signal was en-hanced in the spikelets of transgenic plants overexpressing WTLF1, it was not expressed at the axil of the sterile lemma duringstages Sp4–Sp8 (Fig. S5 A, 1–3). In the lf1 spikelet, the expres-sion domain of LF1 was obviously expanded (Fig. 4B). First, theexpression of LF1 was detected in cells located between thesterile lemma and palea, which might be precursors of the lateralFM during stages Sp3–Sp4 (Fig. 4 B, 1 and 2). Next, LF1 wasexpressed in the same domain where the FM or floral organsof the lateral floret were formed during stages Sp5–Sp8 (Fig.4 B, 3 and 4).The expression pattern of miRNA165/166 was also investi-
gated. In the WT spikelet, miRNA165/166 was expressed mainlyin the precursor cells or primordia of organs, such as the sterilelemma, lemma/palea, lodicule, and stamen, during stages Sp3–Sp7, whereas no obvious signal was detected in the central do-main of the FM (Fig. 4 C, 1–3). During stage Sp8, the expressionof miRNA165/166 was observed mainly in the abaxial sides of allorgans in the spikelet except the pistil (Fig. 4 C, 4). In thespikelets of transgenic plants overexpressing WT LF1, the ex-pression pattern of miRNA165/166 was similar to that of WTduring stages Sp4–Sp8 (Fig. S5 A, 4–6). In the lf1 spikelet, theexpression pattern of miRNA165/166 was also similar to that ofthe WT (Fig. 4D), except that obvious expression of miRNA165/166 was detected in the lateral FM domain at the axil of the sterilelemma (Fig. 4 D, 2–4). Thus, the data indicated that expression ofmiRNA165/166 and LF1 is mutually exclusive in the WT spikeletbut not in the lf1 mutant, which suggests that miRNA165/166 isresponsible for the restriction of WT LF1 expression.In particular, during stage Sp3, before the palea primordium
was initiated, strong expression of miRNA165/166 was observedin the margin domain of the FM located next to the primordiumof the upper sterile lemma in the spikelet of WT and lf1 (Fig. 4C, 1 and D, 1). Therefore, it is possible that the precursor cells ofthe lateral FM in the WT were repressed due to degradation ofthe LF1 mRNA by miRNA165/166 in this domain at stage Sp3.The fact that no signal for miRNA165/166 was detected at theaxil of the sterile lemma in the WT might be due to the absenceof target cells here after stage Sp3 (Fig. 4 C, 2–4), and the serialsections of the picture in Fig. 4 C, 2 further demonstrate thatthere was no expression of the miRNA165/166 signal (Fig. S5B).However, it was expressed in the lf1 mutant due to the presenceof target cells after stage Sp3 (Fig. 4 D, 2–4). Taken togetherwith all the above data, these results suggest that the accumu-lation of miRNA165/166 in the presumptive lateral FM regionsuppresses the expression of LF1 in the WT, whereas once amutation had occurred in the miRNA165/166 target sequence ofLF1, the latter was expressed ectopically to induce initiation ofthe lateral FM at the axil of the sterile lemma in lf1.
LF1 Encodes a HD-ZIP III Transcriptional Activator. LF1 was found toencode the rice HD-ZIP III transcription factor OsHB1 andshared high sequence similarity with other HD-ZIP III genes inmonocots and dicots (Fig. S6). The results of phylogeneticanalysis revealed that the angiosperm HD-ZIP III genes formedthree strongly supported clades, referred to as the “REV,” “PHB/PHV,” and “CNA/HB8” clades, and that LF1/OsHB1 belonged tothe REV clade (Fig. S7A). Previous studies showed that thisfamily of genes regulates embryo patterning, meristem function,vascular development, lateral organ polarity, and interfascicularfiber differentiation in angiosperms (20–23). In this study, weidentified a function of LF1/OsHB1 that is associated with lateralFM fate in rice.Next, constructs that encoded GFP and the GFP-LF1ORF
fusion protein were expressed transiently in rice protoplasts. ForGFP, green fluorescence was detected consistently throughoutthe cell, apart from in the vacuole, whereas the GFP-LF1ORFfusion protein was localized in the nucleus (Fig. S7B). Further, a
GAL4 DNA-binding domain and LF1 fusion protein (pGBKT7-LF1) was expressed in Y2HGold yeast cells. The yeast cells thatcontained pGBKT7-LF1 and the positive control yeast cells wereable to grow in the absence of adenine, histidine, and tryptophanand showed X-α-Gal activity, whereas yeast cells that containedthe negative controls did not (Fig. S7C). These results suggestedthat LF1 is a transcriptional activator.
LF1 Regulates (Floral) Meristem-Related Genes to Initiate the LateralFM. Previous studies have shown that REV (the ortholog of LF1in rice) up-regulates the expression of STM (the ortholog ofOSH1 in rice) in leaf axil meristem cells directly to ensure theinitiation of axillary meristems in Arabidopsis (24). In the presentstudy, the expression level of OSH1 was increased significantly inthe panicles of lf1 compared with the WT by qRT-PCR analysis(Fig. 5A). Next, in situ hybridization was used to compare theexpression pattern of OSH1 in WT and lf1. In the WT, OSH1 wasexpressed specifically in the FM and receptacle at a high levelduring stages Sp4–Sp6 (Fig. 5 B, 1 and 2). After stage Sp7, whenthe carpel primordium formed, only a slight signal was detectedin the rachilla (Fig. 5 B, 3). In the terminal floret of lf1, theexpression pattern of OSH1 was generally similar to that in theWT during stages Sp4–Sp8 (Fig. 5 B, 4–6). As expected, a strongexpression signal for OSH1 was observed in the lf1 lateral FMduring stages Sp5–Sp8 (Fig. 5 B, 5 and 6). Subsequently, ChIPassays were used to examine whether LF1 protein can bind di-rectly to the OSH1 promoter. Chromatin isolated from youngpanicles of the mutated LF1-GFP (mLF1-GFP) transgenic and
Fig. 5. LF1 regulates the meristem maintenance gene OSH1. (A) qRT-PCRanalysis of OSH1 in WT and lf1 panicles. (B) In situ hybridization of OsH1 inWT (B1–B3) and lf1 (B4–B6) panicles. Black arrows indicate gene expressionin the lateral FM of lf1 panicles. fm, floral meristem. (C) Distribution of theconserved binding site ATGAT in the promoter regions of OsH1. Underliningindicates the DNA fragments amplified in the ChIP assays. (D and E) ChIP en-richment compared with the input sample was tested by qPCR. (D) Chromatinisolated from fresh young WT panicles and immunoprecipitated with anti-LF1 antibody. (E) Chromatin isolated from fresh young panicles of mLF1CDS-GFP transgenic plants and immunoprecipitated with anti-GFP antibody. Errorbars indicate the SD of three biological repeats. (Scale bars, 50 μm.)
Zhang et al. PNAS | September 12, 2017 | vol. 114 | no. 37 | 9987
PLANTBIOLO
GY

WT plants was immunoprecipitated with an anti-GFP antibodyand an anti-LF1 antibody, respectively, and then was subjected toqRT-PCR analysis. The results showed that both mLF1-GFP andthe WT LF1 protein cloud bound to four regions of the OSH1promoter, which contained one to six conserved binding motifs(ATGAT) for the HD-ZIP III protein (Fig. 5 C–E). Thesefindings suggested that LF1 might regulate the OSH1 gene di-rectly in both the lf1 and WT spikelet.Analysis by qRT-PCR also showed that expression of the FM
identity genes OsMADS6 and OsMADS15 was increased signifi-cantly in the panicles of lf1 compared with that in the WT (Fig.S8A). Next, the expression patterns of OsMADS6 and OsMADS15were determined in detail by in situ hybridization. During stageSp4, strong expression ofOsMADS6 and OsMADS15 was detectedin the terminal FM in both the WT and lf1 (Fig. S8 B, 1; C, 1; D, 1;and E, 1). From stages Sp5–Sp8, in the terminal florets of both theWT and lf1, OsMADS6 was expressed mainly in the mrp, lodicule,and carpel (Fig. S8 B, 2 and 3 and C, 2 and 3), whereasOsMADS15 was expressed in all the floral organs (Fig. S8D, 2 and3 and E, 2 and 3). Therefore, no differences in expression patternwere found between the WT and lf1 terminal florets. However,clear expression signals for OsMADS6 and OsMADS15 were de-tected in the lf1 lateral FM from stages Sp5–Sp8 (Fig. S8 C, 2 and3 and E, 2 and 3). Thus, it is suspected that the FM identity genesOsMADS6 and OsMADS15 might also be involved in initiation ofthe lateral FM in lf1.
DiscussionOur data suggested that themiRNA165/166–LF1–OSH1 pathwayplays a key role in regulating the induction of lateral florets.First, a single-nucleotide substitution in the complementary sitefor miRNA165/166 in the LF1 gene and the ectopic expression ofthe latter in the lateral FM was observed in lf1. The expressiondomains of miRNA165/166 and LF1 were mutually exclusive inthe WT spikelet. These results fit well with the assumption that
the LF1 mRNA was degraded by miRNA165/166 in the precursorcells of the lateral FM at the axil of the sterile lemma in the WT,whereas miRNA165/166 could not act on the mutated LF1 mRNA,resulting in its ectopic expression at the axil of the sterile lemma toinduce the formation of lateral florets in lf1. Second, in transgenicplants overexpressing the mutated LF1 gene, the spikelet developedlateral florets, similarly to lf1, whereas the spikelet showed a normalmorphology in the transgenic plants overexpressing WT LF1. Thisindicated that the gain-of-function phenotypes that were observedin the lf1 mutant were probably due to ectopic expression ratherthan to overexpression of LF1. In addition, the LF1 gene belongs tothe REV clade of theHD-ZIP III gene family. Previous studies haveshown that most HD-ZIP III genes share conserved complemen-tary sites for miRNA165/166 and similar regulatory mecha-nisms (25). In several gain-of-function mutants of HD-ZIP IIIgenes, such as phb-d, phv-d, and rev in Arabidopsis and Rld1 inmaize, the mutation sites were all located in the complementarysite for miRNA165/166, and the gain of function was shown to becaused by the disruption of miRNA165/166 binding rather thanby the amino acid substitution in the ligand-binding domain (20,21, 26). Furthermore, the function of miRNA165/166 in main-taining the correct expression domain of these HD-ZIP III genesis conserved between dicots and monocots. In the present study,OSH1 was expressed ectopically in the axil of the lf1 sterilelemma where LF1 was expressed ectopically, and its promotercould be bound directly by the LF1 protein, which suggested thatOSH1 might be activated by LF1 to promote initiation of thelateral FM in the lf1 spikelet. Previous studies have demon-strated that REV is required for initiation of the lateral meristemby promoting the expression of the meristem maintenance geneSTM in the axil in Arabidopsis (24). Therefore, the miRNA165/166–LF1–OSH1 pathway in rice is similar to the miRNA165/166–REV–STM pathway in Arabidopsis. In WT rice, miRNA165/166represses the expression of LF1 in the axil of the sterile lemma,which prevents the activation of OSH1 so that a single-floret WTspikelet is produced. In the lf1 mutant, the point mutation in thecomplementary site for miRNA165/166 resulted in the ectopicexpression of LF1, which in turn induced the ectopic expressionof OSH1 and finally resulted in initiation of the lateral FM.From the findings of morphological studies, it has been hy-
pothesized that a putative ancestor of the Oryzeae spikelet hadthree florets (two lateral and one terminal); the two lateral floretsdegenerated during the course of evolution, leaving only onefertile terminal floret (5). This is referred to as the “early three-florets spikelet” hypothesis. Previous studies have demonstratedthat the sterile lemmas in extant Oryza species probably evolvedfrom the lemmas in the two lateral florets. Mutation of G1, EG1,OsMADS34, NSG, or ASP1 or ectopic expression of OsMADS1causes homeotic transformation of the sterile lemma into a lemmato various degrees (6–11), which suggests that sterile lemmas arehomologous to lemmas and partly supports the above hypothesis.This paper reports the observation of lateral florets with normalfloral organs in rice and other Oryzeae (Fig. 6), which constitutesmuch stronger evidence in support of the three-florets spikelethypothesis than the transformation of the sterile lemma into alemma. It is notable that the sterile lemma was not restored to alemma-like organ in the lf1 mutant, in contrast to mutants such asg1, eg1, and pap2. In contrast, the lf1 mutant developed innerfloral organs in lateral florets, whereas those other mutants didnot. This interesting finding has two implications. First, in theprocess of evolution from the three-florets to one-floret spikelet,the degradation of lemmas was controlled by the G1, EG1, andOsMADS34 genes, among others, whereas the degradation ofother floral organs as a whole was controlled by the miRNA165/166–LF1–OSH1 pathway (Fig. 6), although it is not clear whichdegradation occurred earlier in the evolutionary process. Theother is that the lemma (or sterile lemma) is not a real floral organbut is a bract-like organ. In fact, the identities of the lemma and
Fig. 6. Model of hypothesized changes in spikelet architecture duringOryza evolution and putative function of LF1. During the evolutionaryprocess the G1, EG1, OsMADS34, NSG, ASP1, and OsMADS1 genes determinedthe degeneration from the lemma to sterile lemma in lateral florets, whereasthe miRNA165/166–LF1–OSH1 pathway controlled the degeneration of floralorgans in lateral florets. Red arrows indicate positive regulation, the red “T”shape patterning indicates negative regulation, and green arrows show thetwo evolutionary lines of the WT spikelet. The blue arrows and “X” shapepatterning suggest a possible method to breed a rice hybrid with a three-florets spikelet by combining the lf1 mutant and other materials with alemma-like sterile lemma.
9988 | www.pnas.org/cgi/doi/10.1073/pnas.1700504114 Zhang et al.

palea have long been debated (4). In one opinion, their positionson the axis suggest distinct identities of the lemma and palea as abract (formed on a spikelet axis) and a prophyll (formed on afloret axis), respectively (5, 27). In the other opinion, the lemmaand palea are perceived as modified outer tepals (sepals) (28, 29).Our results obviously support the first point of view. Therefore,both the lemma and sterile lemma might be bract-like organs, withthe real floret consisting of the palea and other inner floral organsthat form in the axil of the lemma or sterile lemma.Finally, the three-florets spikelet has potential value in rice
breeding. Increasing the grain number per panicle is one of thekey factors to increase yield. In rice, the grain number per panicleis correlated mainly with the first and secondary branches. Somepreviously reported genes, such as OsCKX2/Gn1a, Ghd7, DENSEAND ERECT PANICLE 1 (DEP1), DEP2, and OsSPL14/IPA1,which have been used to make major contributions to increasingyield, are all involved in regulating panicle branching in rice (30–34). According to the present study, it should be possible to breeda three-florets spikelet in rice, for example by combining the lf1mutant and other lemma-like sterile lemma mutants or transgenicplants (Fig. 6), and this presents the prospect for increasing grainnumber per panicle.
Materials and MethodsThe rice mutant lf1 was identified from an ethylmethane-sulfonate–treatedpopulation of Oryza sativa subsp. indica cultivar Jinhui 10. All plant materialswere grown in experimental fields of the Rice Research Institute of South-west University in Chongqing and Hainan, China. A map-based cloningstrategy was used to isolate the LF1 gene. Expression of LF1 was investigatedin different organs at different growth stages by real-time RT-PCR and in situhybridization. See SI Materials and Methods and Table S1 for full details. Thefold enrichment of the mLF1-GFP and WT LF1 target promoter regionspulled down by the anti-GFP antibody and anti-LF1antibody, respectively,was compared with the input sample, and an IgG antibody was included as anegative control. Three biological repeats with three technical repeats eachwere used to produce data for the statistical analysis. Experimental proce-dures for ChIP-qPCR were performed as described previously (35). DNAfragments of the OSH1 promoter regions, which were positively enriched inChIP-qPCR, were amplified using a SYBR Premix Ex Taq II Kit (TaKaRa).
ACKNOWLEDGMENTS. This work was supported by National Natural ScienceFoundation of China Grants 31730063 and 31271304, National Key Researchand Development Project Grant 2016YFD0101107, Fundamental ResearchFunds for the Central Universities Grant XDJK2016A013, Project of Chongq-ing Science & Technology Commission Grants CSTC2016shms-ztzx0017 andCSTC2017jcyjBX0062, and National Natural Science Foundation of ChinaGrant 31501377.
1. Komatsu M, Chujo A, Nagato Y, Shimamoto K, Kyozuka J (2003) FRIZZY PANICLE isrequired to prevent the formation of axillary meristems and to establish floral mer-istem identity in rice spikelets. Development 130:3841–3850.
2. Lee DY, An G (2012) Two AP2 family genes, supernumerary bract (SNB) and Osinde-terminate spikelet 1 (OsIDS1), synergistically control inflorescence architecture andfloral meristem establishment in rice. Plant J 69:445–461.
3. Ren D, et al. (2013) MULTI-FLORET SPIKELET1, which encodes an AP2/ERF protein,determines spikelet meristem fate and sterile lemma identity in rice. Plant Physiol 162:872–884.
4. Yoshida H, Nagato Y (2011) Flower development in rice. J Exp Bot 62:4719–4730.5. Arber AR (1934) The Gramineae: A Study of Cereal, Bamboo, and Grass (Cambridge
Univ Press Archive, Cambridge, UK).6. Yoshida A, Suzaki T, Tanaka W, Hirano HY (2009) The homeotic gene long sterile
lemma (G1) specifies sterile lemma identity in the rice spikelet. Proc Natl Acad Sci USA106:20103–20108.
7. Li H, et al. (2009) A putative lipase gene EXTRA GLUME1 regulates both empty-glumefate and spikelet development in rice. Plant J 57:593–605.
8. Lin X, et al. (2014) The pleiotropic SEPALLATA-like gene OsMADS34 reveals that the‘empty glumes’ of rice (Oryza sativa) spikelets are in fact rudimentary lemmas. NewPhytol 202:689–702.
9. Wang N, et al. (2013) nonstop glumes (nsg), a novel mutant affects spikelet devel-opment in rice. Genes Genomics 35:149–157.
10. Yoshida A, Ohmori Y, Kitano H, Taguchi-Shiobara F, Hirano HY (2012) Aberrantspikelet and panicle1, encoding a TOPLESS-related transcriptional co-repressor, isinvolved in the regulation of meristem fate in rice. Plant J 70:327–339.
11. Wang L, et al. (2016) Ectopic expression of OsMADS1 caused dwarfism andspikelet alteration in rice. Plant Growth Regul 81:433–442.
12. Wang K, et al. (2010) DEP and AFO regulate reproductive habit in rice. PLoS Genet 6:e1000818.
13. Yao SG, Ohmori S, Kimizu M, Yoshida H (2008) Unequal genetic redundancy of ricePISTILLATA orthologs, OsMADS2 and OsMADS4, in lodicule and stamen development.Plant Cell Physiol 49:853–857.
14. Nagasawa N, et al. (2003) SUPERWOMAN1 and DROOPING LEAF genes control floralorgan identity in rice. Development 130:705–718.
15. Yamaguchi T, et al. (2006) Functional diversification of the two C-class MADS boxgenes OSMADS3 and OSMADS58 in Oryza sativa. Plant Cell 18:15–28.
16. Yamaguchi T, et al. (2004) The YABBY gene DROOPING LEAF regulates carpel spec-ification and midrib development in Oryza sativa. Plant Cell 16:500–509.
17. Dreni L, et al. (2007) The D-lineage MADS-box gene OsMADS13 controls ovuleidentity in rice. Plant J 52:690–699.
18. Sparks DL (2012) Advances in Agronomy (Academic, Cambridge, MA).
19. Rhoades MW, et al. (2002) Prediction of plant microRNA targets. Cell 110:513–520.20. Mallory AC, et al. (2004) MicroRNA control of PHABULOSA in leaf development:
Importance of pairing to the microRNA 5′ region. EMBO J 23:3356–3364.21. Zhong R, Ye ZH (2004) Amphivasal vascular bundle 1, a gain-of-function mutation of
the IFL1/REV gene, is associated with alterations in the polarity of leaves, stems andcarpels. Plant Cell Physiol 45:369–385.
22. Zhong R, Ye ZH (1999) IFL1, a gene regulating interfascicular fiber differentiationin Arabidopsis, encodes a homeodomain-leucine zipper protein. Plant Cell 11:2139–2152.
23. Otsuga D, DeGuzman B, Prigge MJ, Drews GN, Clark SE (2001) REVOLUTA regulatesmeristem initiation at lateral positions. Plant J 25:223–236.
24. Shi B, et al. (2016) Two-step regulation of a meristematic cell population acting inshoot branching in Arabidopsis. PLoS Genet 12:e1006168.
25. Floyd SK, Zalewski CS, Bowman JL (2006) Evolution of class III homeodomain-leucinezipper genes in streptophytes. Genetics 173:373–388.
26. Juarez MT, Kui JS, Thomas J, Heller BA, Timmermans MC (2004) microRNA-mediatedrepression of rolled leaf1 specifies maize leaf polarity. Nature 428:84–88.
27. Kellogg EA (2001) Evolutionary history of the grasses. Plant Physiol 125:1198–1205.28. Stebbins GL (1951) Natural selection and the differentiation of angiosperm families.
Evolution 5:299–324.29. Bowman JL (1997) Evolutionary conservation of angiosperm flower development at
the molecular and genetic levels. J Biosci 22:515–527.30. Ashikari M, et al. (2005) Cytokinin oxidase regulates rice grain production. Science
309:741–745.31. Xue W, et al. (2008) Natural variation in Ghd7 is an important regulator of heading
date and yield potential in rice. Nat Genet 40:761–767.32. Yan CJ, et al. (2007) Identification and characterization of a major QTL responsible for
erect panicle trait in japonica rice (Oryza sativa L.). Theor Appl Genet 115:1093–1100.33. Li F, et al. (2010) Rice DENSE AND ERECT PANICLE 2 is essential for determining
panicle outgrowth and elongation. Cell Res 20:838–849.34. Miura K, et al. (2010) OsSPL14 promotes panicle branching and higher grain pro-
ductivity in rice. Nat Genet 42:545–549.35. Xu J, et al. (2010) The ABORTED MICROSPORES regulatory network is required for
postmeiotic male reproductive development in Arabidopsis thaliana. Plant Cell 22:91–107.
36. Sang X, et al. (2012) CHIMERIC FLORAL ORGANS1, encoding a monocot-specific MADSbox protein, regulates floral organ identity in rice. Plant Physiol 160:788–807.
37. Tamura K, et al. (2011) MEGA5: Molecular evolutionary genetics analysis usingmaximum likelihood, evolutionary distance, and maximum parsimony methods. MolBiol Evol 28:2731–2739.
Zhang et al. PNAS | September 12, 2017 | vol. 114 | no. 37 | 9989
PLANTBIOLO
GY
Related Documents











