Lactational exposure to inorganic mercury: Evidence of neurotoxic effects Jeferson L. Franco a,b , Hugo de Campos Braga a , Adriana K.C. Nunes a , Camila M. Ribas a , James Stringari a , Aline P. Silva a , Solange C. Garcia Pomblum c , Ângela M. Moro c , Denise Bohrer d , Adair R.S. Santos b , Alcir L. Dafre b , Marcelo Farina a, ⁎ a Departamento de Bioquímica, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, 88040-900, Florianópolis, SC, Brazil b Departamento de Ciências Fisiológicas, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, 88040-900, Florianópolis, SC, Brazil c Departamento de Análises Clínicas e Toxicológicas, Centro de Ciências da Saúde, Universidade Federal de Santa Maria, 97105-900, Santa Maria, RS, Brazil d Departamento de Química, Centro de Ciências Naturais e Exatas, Universidade Federal de Santa Maria, 97105-900, Santa Maria, RS, Brazil Received 26 June 2006; received in revised form 23 November 2006; accepted 24 November 2006 Available online 6 December 2006 Abstract This study examined the effects of inorganic mercury (mercuric chloride – HgCl 2 ) exposure exclusively through maternal milk on biochemical parameters related to oxidative stress (glutathione and thiobarbituric acid reactive substances levels, glutathione peroxidase and glutathione reductase activities) in the cerebellum of weanling mice. These parameters were also evaluated in the cerebellum of mothers, which were subjected to intraperitoneal injections of HgCl 2 (0, 0.5 and 1.5 mg/kg, once a day) during the lactational period. Considering the relationship between cerebellar function and motor activity, the presence of motor impairment was also evaluated in the offspring exposed to HgCl 2 during lactation. After treatments (at weaning), pups lactationally exposed to inorganic mercury showed high levels of mercury in the cerebellar tissue, as well as significant impairment in motor performance in the rotarod task and decreased locomotor activity in the open field. Offspring and dams did not show changes in cerebellar glutathione levels or glutathione peroxidase activity. In pups, lactational exposure to inorganic mercury significantly increased cerebellar lipoperoxidation, as well as the activity of cerebellar glutathione reductase. However, these phenomena were not observed in dams. These results indicate that inorganic mercury exposure through maternal milk is capable of inducing biochemical changes in the cerebellum of weanling mice, as well as motor deficit and these phenomena appear to be related to the pro-oxidative properties of inorganic mercury. © 2006 Published by Elsevier Inc. Keywords: Inorganic mercury; Mercuric chloride; Lactational exposure; Neurotoxicity; Oxidative stress; Motor impairment 1. Introduction Mercury is an important environmental toxicant that causes neurological and developmental impairment in both humans and animals. Within the environment, mercury exists in three different chemical forms (elemental mercury vapor, inorganic mercury salts, and organic mercury) that are all important for human exposure [10]. Gold mining emits elemental mercury vapor that is inhaled and absorbed into the bloodstream [24]. Methylmercury (MeHg), an organic mercury compound, is found in fish and seafood, and fishing communities are highly exposed to MeHg due to the high consumption of MeHg- contaminated fishes [11]. Human exposure to inorganic mer- cury is mainly occupational, which is often related to mining and industrial activities [6]. In addition, inorganic mercury is believed to be the toxic species produced in tissues after inha- lation of mercury vapor [55]. The distribution, metabolism, and toxicity of mercury are largely dependent upon its chemical form. In this regard, it is noteworthy that organomercurials have been pointed to as the most hazardous species when compared to elemental and inorganic mercury compounds [42]. This phenomenon appears to be related, at least in part, to the or- ganomercurials' capability of reaching high levels in the central nervous system (CNS) [1,11]. Inorganic mercury is toxic to the renal, reproductive and nervous systems [21], and human exposure to inorganic mer- cury is often related to specific working conditions e.g. mining, Neurotoxicology and Teratology 29 (2007) 360 – 367 www.elsevier.com/locate/neutera ⁎ Corresponding author. Tel.: +55 48 3331 9795; fax: +55 48 3331 9672. E-mail address: [email protected] (M. Farina). 0892-0362/$ - see front matter © 2006 Published by Elsevier Inc. doi:10.1016/j.ntt.2006.11.009

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

gy 29 (2007) 360–367www.elsevier.com/locate/neutera
Neurotoxicology and Teratolo
Lactational exposure to inorganic mercury: Evidence of neurotoxic effects
Jeferson L. Franco a,b, Hugo de Campos Braga a, Adriana K.C. Nunes a, Camila M. Ribas a,James Stringari a, Aline P. Silva a, Solange C. Garcia Pomblum c, Ângela M. Moro c, Denise Bohrer d,
Adair R.S. Santos b, Alcir L. Dafre b, Marcelo Farina a,⁎
a Departamento de Bioquímica, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, 88040-900, Florianópolis, SC, Brazilb Departamento de Ciências Fisiológicas, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, 88040-900, Florianópolis, SC, Brazil
c Departamento de Análises Clínicas e Toxicológicas, Centro de Ciências da Saúde, Universidade Federal de Santa Maria, 97105-900, Santa Maria, RS, Brazild Departamento de Química, Centro de Ciências Naturais e Exatas, Universidade Federal de Santa Maria, 97105-900, Santa Maria, RS, Brazil
Received 26 June 2006; received in revised form 23 November 2006; accepted 24 November 2006Available online 6 December 2006
Abstract
This study examined the effects of inorganic mercury (mercuric chloride – HgCl2) exposure exclusively through maternal milk on biochemicalparameters related to oxidative stress (glutathione and thiobarbituric acid reactive substances levels, glutathione peroxidase and glutathionereductase activities) in the cerebellum of weanling mice. These parameters were also evaluated in the cerebellum of mothers, which were subjectedto intraperitoneal injections of HgCl2 (0, 0.5 and 1.5 mg/kg, once a day) during the lactational period. Considering the relationship betweencerebellar function and motor activity, the presence of motor impairment was also evaluated in the offspring exposed to HgCl2 during lactation.After treatments (at weaning), pups lactationally exposed to inorganic mercury showed high levels of mercury in the cerebellar tissue, as well assignificant impairment in motor performance in the rotarod task and decreased locomotor activity in the open field. Offspring and dams did notshow changes in cerebellar glutathione levels or glutathione peroxidase activity. In pups, lactational exposure to inorganic mercury significantlyincreased cerebellar lipoperoxidation, as well as the activity of cerebellar glutathione reductase. However, these phenomena were not observed indams. These results indicate that inorganic mercury exposure through maternal milk is capable of inducing biochemical changes in the cerebellumof weanling mice, as well as motor deficit and these phenomena appear to be related to the pro-oxidative properties of inorganic mercury.© 2006 Published by Elsevier Inc.
Keywords: Inorganic mercury; Mercuric chloride; Lactational exposure; Neurotoxicity; Oxidative stress; Motor impairment
1. Introduction
Mercury is an important environmental toxicant that causesneurological and developmental impairment in both humansand animals. Within the environment, mercury exists in threedifferent chemical forms (elemental mercury vapor, inorganicmercury salts, and organic mercury) that are all important forhuman exposure [10]. Gold mining emits elemental mercuryvapor that is inhaled and absorbed into the bloodstream [24].Methylmercury (MeHg), an organic mercury compound, isfound in fish and seafood, and fishing communities are highly
⁎ Corresponding author. Tel.: +55 48 3331 9795; fax: +55 48 3331 9672.E-mail address: [email protected] (M. Farina).
0892-0362/$ - see front matter © 2006 Published by Elsevier Inc.doi:10.1016/j.ntt.2006.11.009
exposed to MeHg due to the high consumption of MeHg-contaminated fishes [11]. Human exposure to inorganic mer-cury is mainly occupational, which is often related to miningand industrial activities [6]. In addition, inorganic mercury isbelieved to be the toxic species produced in tissues after inha-lation of mercury vapor [55]. The distribution, metabolism, andtoxicity of mercury are largely dependent upon its chemicalform. In this regard, it is noteworthy that organomercurials havebeen pointed to as the most hazardous species when comparedto elemental and inorganic mercury compounds [42]. Thisphenomenon appears to be related, at least in part, to the or-ganomercurials' capability of reaching high levels in the centralnervous system (CNS) [1,11].
Inorganic mercury is toxic to the renal, reproductive andnervous systems [21], and human exposure to inorganic mer-cury is often related to specific working conditions e.g. mining,

361J.L. Franco et al. / Neurotoxicology and Teratology 29 (2007) 360–367
spillage of mercury compounds on work clothes or in theworking environment, handling of mercury salts in the chem-ical industry and laboratories [4,7,53]. In addition, accidental[35,43] and intentional [54] events can also contribute to humanexposure to inorganic mercury.
Even though mercury neurotoxicity is not well understood, ithas been shown that alterations in calcium and glutamatehomeostasis [2,46,3,17], oxidative stress [38,20], and oxidationof protein thiols [26] represent important molecular mechan-isms by which both organic and inorganic mercury may causeneurotoxicity. Studies from our [47,20,18] and others groups[2,29,44] have pointed to the antioxidant glutathione systemas a potential target for the deleterious effects of organic andinorganic mercurials.
The developing CNS is particularly susceptible to the neu-rological effects of both inorganic and organic forms of mercury[23,50]. In this regard, some studies have showed that theexposure of pregnant women to methylmercury can lead toindirect embryo/fetal intoxication [27,51] and some studiessuggest that the prenatal exposure to methylmercury causesneurological deficits in the offspring [23,36]. In addition, ex-perimental studies have shown that both inorganic and organicforms of mercury can be excreted into maternal milk [33,20,48].It is noteworthy that inorganic mercury concentrates more indams' breast milk than methylmercury [48]. However, to thebest of our knowledge, only methylmercury (an organomercu-rial) has been shown to display neurotoxic effects after lacta-tional exposure [33,20]. Indeed, there appear to be no studies oflactational exposure to inorganic mercury as a cause of neuro-toxicity in the offspring.
Taking into account that (i) lactational exposure to organicmercury causes neurotoxicity in the offspring; (ii) the earlypostnatal period represents an important phase when the CNS ishighly susceptible to neurotoxicants, including organic andinorganic mercurials; (iii) and there are no studies of the lacta-tional exposure to inorganic mercury as a cause of neurotoxicityin the offspring, this study was aimed at the identification of apotential neurotoxicity induced by the lactational exposure toinorganic mercury in weanling mice. Since organic and in-organic mercurials have been shown to display pro-oxidativeeffects toward the CNS [28,38,19], biochemical parametersrelated to oxidative stress were evaluated in mouse cerebellum.Considering the relationship between cerebellar and motorfunctions, behavioral parameters related to motor performanceof pups were also evaluated.
2. Materials and methods
2.1. Chemicals
Mercuric chloride (HgCl2) was purchased from Merck(Darmstadt, WI). β-nicotinamide adenine dinucleotide phos-phate sodium salt — reduced form, 5-5′-dithio-bis (2-nitroben-zoic) acid, glutathione reductase from baker's yeast, andreduced glutathione were obtained from Sigma (St. Louis, Mo.,USA). All other chemicals were of the highest grade availablecommercially.
2.2. Animals
Adult Swiss Albino mice (male and female), 90 days old,from our own breeding colony were maintained at 22±2 °C, ona 12:12 h light/dark cycle, with free access to food (Nuvital, PR,Brazil) and water. The breeding regimen consisted of groupingthree virgin females with one male for 5 days. Pregnant micewere selected and housed individually in opaque plastic cages.
2.3. Treatment
On the first day after parturition (postnatal day 1), 12 damswere randomly assigned to one of three groups – control (C),low Hg (L) or high Hg (H) – of four animals each. Pups (eightper litter) were maintained with their mothers, which wereimmediately exposed to HgCl2 through intraperitoneal (i.p.)injections. Mothers from groups L and H received i.p. injectionsof HgCl2 (0.5 and 1.5 mg/kg, respectively), once a day, duringthe lactational period. Mercury doses were based on the studyof Sundberg et al., [49]. In addition, based on previous datafrom our group (not published), we have found that the chosendoses of HgCl2 (0.5 and 1.5 mg/kg/day) do not decreasethe body weight of mothers during lactation, suggesting theabsence of a high degree of systemic toxicity. Mothers fromgroup C were submitted to i.p. injections of physiological saline(150 mM NaCl), once a day. Thus, the exclusive means ofoffspring exposure to mercury was through maternal milk.Liquid and solid ingestions of mothers were monitored daily.All experiments were conducted in accordance with the GuidingPrinciples in the Use of Animals in Toxicology, adopted bythe Society of Toxicology in July 1989, and all experimentswere approved by our ethics committee for animal use atthe Universidade Federal de Santa Catarina (313/CEUA;23080.026023/2004-39/UFSC).
2.4. Behavioral tests
On postnatal day 21, two weanling mice (one male and onefemale) from each litter were randomly selected for thebehavioral/functional tests that evaluate the animals' locomotoractivity and coordination (open field and rotarod tasks). Sincetwo weanling mice were selected from each litter and there were4 litters per group, the total number of analyzed pups was 8 pergroup. The remaining 6 pups in each litter were sacrificedby decapitation and were not considered in the analysis. Initial-ly, pups were subjected to the open field test as previouslydescribed [16], with minor modifications. Open field testswere performed in a separated room with no interferencenoise or human activity. Locomotor activity was assessed in asingle session of 6 min using an open field box measuring56 cm×42 cm×40 cm (high) with the floor divided into 12squares. The number of squares crossed with the four paws wasused as measure of locomotor activity. After the open fieldtest, pups were subjected to the rotarod task and this procedurewas based on the study of Duham and Miya [13], with minormodifications. In short, the homemade apparatus consisted ofa bar with a diameter of 2.5 cm, subdivided into 4 compartments

Table 1Body weight (mothers and offspring) at the end of the treatment, and liquid andsolid ingestions of mothers during the treatment
Groups Body weight(g)
Liquid ingestion(ml/day)
Solid ingestion(g/day)
Mothers Offspring Mothers Mothers
Control 35.5±2.4 8.5±0.5 25.6±5.9 28.5±2.3HgCl2 0.5 mg/kg 36.0±2.1 8.9±0.5 25.3±6.1 27.6±1.9HgCl2 1.5 mg/kg 32.5±3.3 5.1±0.3# 26.7±7.8 26.9±3.3
Data are represented as mean±SEM. Mothers (n=4 per group). Offspring(n=4 litters per group; eight⁎ pups per litter).⁎ One female pup plus one male pup were selected from each litter; because nosignificant effects of sex were observed, group mean data are presented.# Statistically different from control.
362 J.L. Franco et al. / Neurotoxicology and Teratology 29 (2007) 360–367
by disks, 25 cm in diameter. The bar rotated at a constant speedof 17 rpm and the length of time pups remained on the apparatuswas recorded. The maximum time allowed on the rotarodapparatus was 60 s. Each pup was subjected to three differenttrials and the mean of their falling latency values was used inthe statistical analysis as actual value.
2.5. Tissue preparation
After the behavioral tests, the two previously selected pups(8 per group) and their respective mothers (4 per group) werekilled by decapitation. Right cerebellar hemispheres werehomogenized (1:5 w/v) in HEPES [N-(2-hydroxyethyl)pipera-zine-N′-(2-ethanesulfonic acid)] 25 mM, pH 7.4 buffer andthe tissue homogenates were rapidly centrifuged at 20,000 ×g,at 4 °C for 30 min. The supernatants obtained were used forthe determinations of enzymatic activities, as well as for thequantifications of glutathione and thiobarbituric acid reactivesubstances (TBARS) levels. Left cerebellar hemispheres wereused for the mercury determinations. It is important to statethat the cerebellum was the chosen encephalic structure dueto the high affinity of mercurials for cerebellar granular cells[31].
2.6. Biochemical determinations
Glutathione reductase (GR) and glutathione peroxidase (GPx)activities were determined based on Carlberg and Mannervik[9] and on Wendel [52], respectively. Glutathione (GSH) wasmeasured as nonprotein thiols based on Ellman [14] with minormodifications [15]. Lipid peroxidation was measured asmalondialdehyde equivalents by the method of Ohkawa andcollaborators [37] with minor modifications [15]. Cerebellarmercury content was measured by cold vapor atomic absorptionspectrometry according to a method previously described [34].This method measures total mercury and does not distinguishbetween organic or inorganic mercury species. Protein concen-tration was determined according to Bradford [8], using a bovineserum albumin as standard.
As already mentioned, two weanling mice (one of each sex)were selected from each litter and there were 4 litters per group:the total number of analyzed pups was 8 per group.
2.7. Statistical analysis
Differences between groups were evaluated by analysis ofvariance (ANOVA), followed by Duncan's multiple range testswhen appropriate. Mothers and pups were independently an-alyzed. Liner regression was also performed in order to detectdose-dependent effects. Since one male pup plus one femalepup were used as representative of the litter, it was possible toanalyze potential gender effects by a mixed model ANOVA(between and within subjects factors). In fact, the litter was theunit of analysis and sex was analyzed as a correlated (repeated)within-litter variable while treatment was a between-litter vari-able. If no effects of sex were found, the data were presented asgroup means.
3. Results
3.1. General health
Liquid and solid ingestions of dams during lactation were notdifferent between groups (Table 1). Signs of overt toxicity werenot observed in lactating dams. Although the body weight ofdams at the end of treatments were not statistically differentbetween groups, pups whose mothers were exposed to thehighest mercury dose (1.5 mg/kg) presented lower body weightwhen compared to the control group when analyzed by a mixedmodel ANOVA [F(2,9)=27.99; pb0.001 — Table 1]. Gendereffects [F(1,9)=0.455; p=0.517] and gender-by-treatmentinteraction [F(2,9)=0.270; p=0.769] were not significant forbody weight.
3.2. Behavioral tests
Fig. 1 depicts the locomotor activity of pups in the open field(A), as well as their motor performance in the rotarod task (B)on postnatal day 21. Animals whose mothers were exposed tothe highest dose of mercuric chloride (1.5 mg/kg) showed asignificant [F(2,9)=5.52; p=0.027] decrease in locomotoractivity (Fig. 1A) and a significant [F(2,9)=4.86; p=0.037]impairment of motor performance (Fig. 1B). Gender effects[F(1,9) =1.629; p=0.234] and gender-by-treatment inter-action [F(2,9)=1.221; p=0.340] were not significant forlocomotor activity (open field). In addition, gender effects[F(1,9)=0.510; p=0.493] and gender-by-treatment interac-tion [F(2,9)=0.504; p=0.620] were not significant for motorperformance (rotarod).
3.3. Biochemical analyses
Fig. 2 depicts data on the glutathione antioxidant systemand lipid peroxidation in mouse pup cerebellum. Glutathionelevels and glutathione peroxidase activity did not change afterlactational exposure to inorganic mercury. However, lactationalexposure to inorganic mercury significantly increased glutathi-one reductase activity [F(2,9)=4.936; p=0.036] and TBARSlevels [F(2,9)=4.766; p=0.039] in the cerebellum of weanlingmice. Gender effects [F(1,9)=0.067; p=0.801] and gender-by-
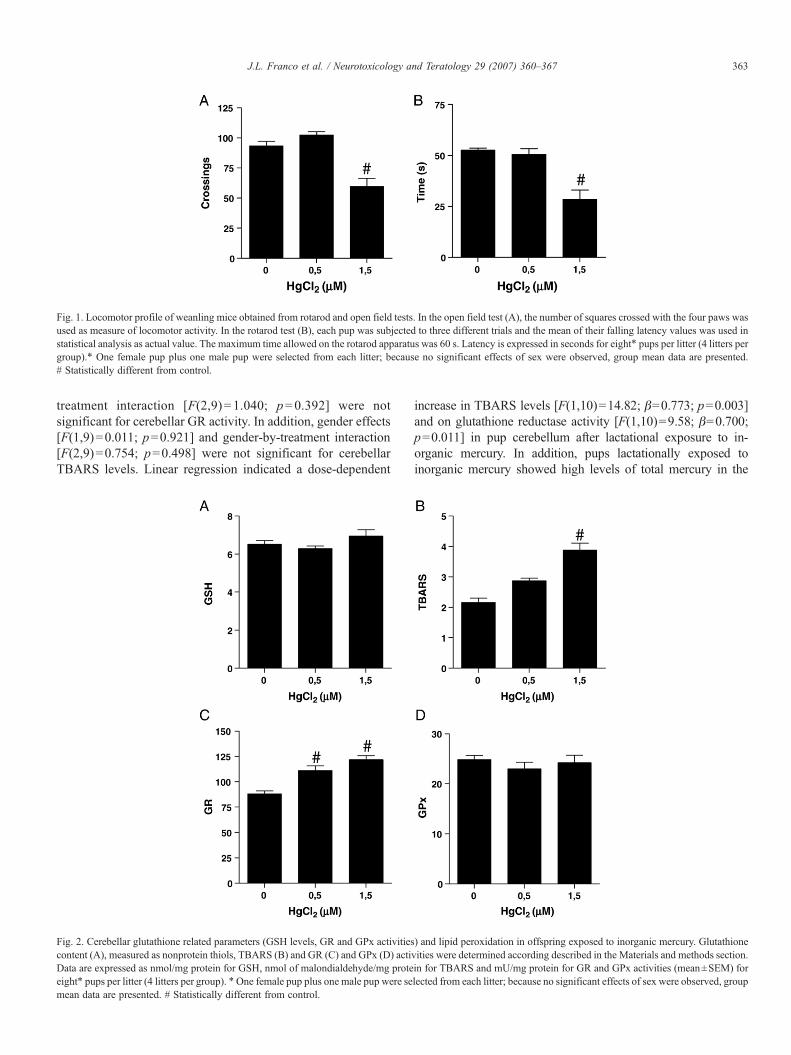
Fig. 1. Locomotor profile of weanling mice obtained from rotarod and open field tests. In the open field test (A), the number of squares crossed with the four paws wasused as measure of locomotor activity. In the rotarod test (B), each pup was subjected to three different trials and the mean of their falling latency values was used instatistical analysis as actual value. The maximum time allowed on the rotarod apparatus was 60 s. Latency is expressed in seconds for eight⁎ pups per litter (4 litters pergroup).⁎ One female pup plus one male pup were selected from each litter; because no significant effects of sex were observed, group mean data are presented.# Statistically different from control.
363J.L. Franco et al. / Neurotoxicology and Teratology 29 (2007) 360–367
treatment interaction [F(2,9)=1.040; p=0.392] were notsignificant for cerebellar GR activity. In addition, gender effects[F(1,9)=0.011; p=0.921] and gender-by-treatment interaction[F(2,9)=0.754; p=0.498] were not significant for cerebellarTBARS levels. Linear regression indicated a dose-dependent
Fig. 2. Cerebellar glutathione related parameters (GSH levels, GR and GPx activitiescontent (A), measured as nonprotein thiols, TBARS (B) and GR (C) and GPx (D) actiData are expressed as nmol/mg protein for GSH, nmol of malondialdehyde/mg proteeight⁎ pups per litter (4 litters per group). ⁎One female pup plus one male pup were semean data are presented. # Statistically different from control.
increase in TBARS levels [F(1,10)=14.82; β=0.773; p=0.003]and on glutathione reductase activity [F(1,10)=9.58; β=0.700;p=0.011] in pup cerebellum after lactational exposure to in-organic mercury. In addition, pups lactationally exposed toinorganic mercury showed high levels of total mercury in the
) and lipid peroxidation in offspring exposed to inorganic mercury. Glutathionevities were determined according described in the Materials and methods section.in for TBARS and mU/mg protein for GR and GPx activities (mean±SEM) forlected from each litter; because no significant effects of sex were observed, group
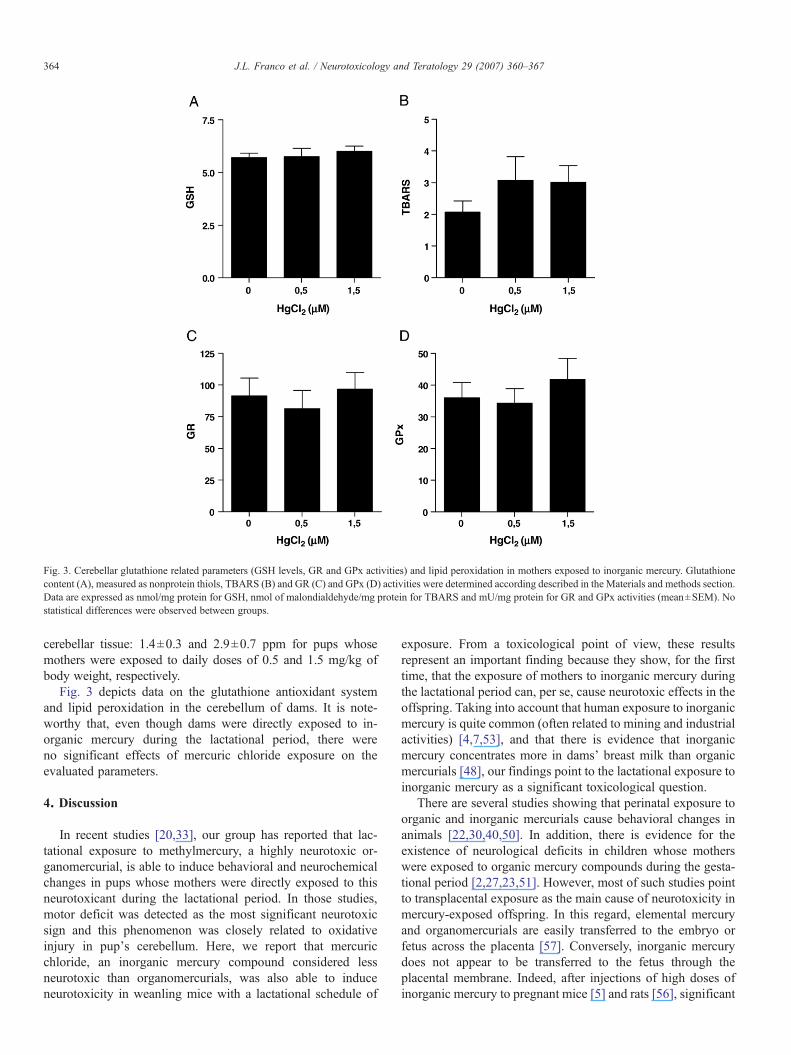
Fig. 3. Cerebellar glutathione related parameters (GSH levels, GR and GPx activities) and lipid peroxidation in mothers exposed to inorganic mercury. Glutathionecontent (A), measured as nonprotein thiols, TBARS (B) and GR (C) and GPx (D) activities were determined according described in the Materials and methods section.Data are expressed as nmol/mg protein for GSH, nmol of malondialdehyde/mg protein for TBARS and mU/mg protein for GR and GPx activities (mean±SEM). Nostatistical differences were observed between groups.
364 J.L. Franco et al. / Neurotoxicology and Teratology 29 (2007) 360–367
cerebellar tissue: 1.4±0.3 and 2.9±0.7 ppm for pups whosemothers were exposed to daily doses of 0.5 and 1.5 mg/kg ofbody weight, respectively.
Fig. 3 depicts data on the glutathione antioxidant systemand lipid peroxidation in the cerebellum of dams. It is note-worthy that, even though dams were directly exposed to in-organic mercury during the lactational period, there wereno significant effects of mercuric chloride exposure on theevaluated parameters.
4. Discussion
In recent studies [20,33], our group has reported that lac-tational exposure to methylmercury, a highly neurotoxic or-ganomercurial, is able to induce behavioral and neurochemicalchanges in pups whose mothers were directly exposed to thisneurotoxicant during the lactational period. In those studies,motor deficit was detected as the most significant neurotoxicsign and this phenomenon was closely related to oxidativeinjury in pup's cerebellum. Here, we report that mercuricchloride, an inorganic mercury compound considered lessneurotoxic than organomercurials, was also able to induceneurotoxicity in weanling mice with a lactational schedule of
exposure. From a toxicological point of view, these resultsrepresent an important finding because they show, for the firsttime, that the exposure of mothers to inorganic mercury duringthe lactational period can, per se, cause neurotoxic effects in theoffspring. Taking into account that human exposure to inorganicmercury is quite common (often related to mining and industrialactivities) [4,7,53], and that there is evidence that inorganicmercury concentrates more in dams' breast milk than organicmercurials [48], our findings point to the lactational exposure toinorganic mercury as a significant toxicological question.
There are several studies showing that perinatal exposure toorganic and inorganic mercurials cause behavioral changes inanimals [22,30,40,50]. In addition, there is evidence for theexistence of neurological deficits in children whose motherswere exposed to organic mercury compounds during the gesta-tional period [2,27,23,51]. However, most of such studies pointto transplacental exposure as the main cause of neurotoxicity inmercury-exposed offspring. In this regard, elemental mercuryand organomercurials are easily transferred to the embryo orfetus across the placenta [57]. Conversely, inorganic mercurydoes not appear to be transferred to the fetus through theplacental membrane. Indeed, after injections of high doses ofinorganic mercury to pregnant mice [5] and rats [56], significant

Table 2Effects of the lactational exposure to methylmercury or inorganic mercury onbehavioral/functional tests and on cerebellar biochemical parameters related tooxidative stress in weanling mice
Behavioral/biochemicalparameters
Lactational exposureto methylmercury
Lactational exposureto inorganic mercury
Cerebellar levels of total mercury ⇑a ⇑Locomotor activity(open field test)
Not changeda ⇓
Motor performance (rotarod test) ⇓a ⇓Cerebellar glutathione levels ⇓a Not changedCerebellar GR activity Not changeda ⇑Cerebellar GPx activity Not changeda Not changedCerebellar lipid peroxidation NE ⇑Cerebellar hydroperoxide levels ⇑b NE
Data from mice lactationally exposed to methylmercury were provided fromprevious studies from our group [20]a and [33]b.⇑: Increased when compared to controls.⇓: Decreased when compared to controls.NE=not evaluated.
365J.L. Franco et al. / Neurotoxicology and Teratology 29 (2007) 360–367
accumulation of mercury was observed in the placenta, and amuch lower accumulation in the fetus, suggesting that theplacental membrane constitutes a barrier against the penetrationof inorganic mercury into the fetus. On the other hand, thelactational transfer of mercury seems to be more efficientfollowing administration of inorganic mercury than after ad-ministration of organic mercury [48]. In fact, Sundberg andcollaborators [48] showed that the lactational transfer of mer-cury in mice was more efficient following administration ofinorganic mercury than after administration of organic mercury(methylmercury), with a five times higher peak concentration inmilk, higher milk:plasma concentration ratios, and 8% of theadministered dose excreted in milk compared with 4% formethylmercury. Our findings, combined with those of Sundbergand collaborators [48], make relevant the lactational exposure toinorganic mercury in terms of health concerns. Nevertheless, itis important to state that the doses used in our study wererelatively high and the equivalent doses in humans have yet tobe established.
It is well known that organic and inorganic mercurials causeneurobehavioral changes in both animals and humans, mainlyduring early phases of development, leading to loss of cognitiveand motor functions in the childhood [50,23,39]. Sanfeliu andcollaborators [42] have shown that cerebellar cells are targetedselectively by mercury compounds in vivo and the relationshipbetween mercury-induced motor deficit and mercury-inducedcerebellar damage is a well-described phenomenon [41]. Here,we observed that the exclusive lactational exposure to inorganicmercury was able to cause a dose-dependent decrease in thelocomotor activity of suckling mice, as well as an impairment inmotor performance. These animals also showed increased lipidperoxidation in the cerebellar tissue, suggesting that the pro-oxidative properties of inorganic mercury could be responsiblefor its neurobehavioral effects. In addition, it is noteworthythat these animals showed high levels of mercury in thecerebellar tissue, confirming the transference of inorganic mer-cury through maternal milk, as well as the entry of thexenobiotic into the CNS of pups. However, it is important tostate that the 40% reduction in offspring body weight, whichwas observed in animals exposed to the highest mercury dose,can represent a significant growth deficit that could by itselfaccount for changes in the locomotor behavior.
Overproduction of reactive oxygen species and furtheroxidative stress is one of the most important consequences ofthe toxicity of metals [26]. Notably, glutathione (GSH) andGSH-related enzymes glutathione peroxidase (GPx) and gluta-thione reductase (GR) figure as major cell defenses to counter-act oxidative stress [45,12]. Particularly important: mercury-induced neurotoxicity is related, at least in part, to its effects onthe GSH antioxidant system. In this regard, studies from ourgroup have reported that organomercurials decrease GSH levelsand increase the levels of peroxides and thiobarbituric acidreactive substances in mouse CNS [33,18]. In addition, organo-mercurial exposure has been reported to change the activities ofthe GSH-related enzymes GPx and GR in mouse CNS [18,47].Here we observed that the lactational exposure of suckling miceto inorganic mercury did not change GSH levels in the cere-
bellum. However, cerebellar TBARS levels were increased inthese animals, pointing to an occurrence of oxidative damage tomembrane lipids. In addition, these animals showed an increasein the cerebellar GR activity in response to inorganic mercuryexposure through maternal milk. GR is an important enzymeinvolved in the reduction of oxidized glutathione (GSSG) toGSH, using reduced nicotinamide adenine dinucleotide phos-phate as a reducing cofactor [25]. Interestingly, the increase inGR activity during organic and inorganic mercury exposure wasalso observed previously [32,18], and such phenomena cancontribute to maintaining GSH levels at physiological levels inthe CNS, as shown in Fig. 2. It is important to note that, in aprevious study by our group [20], the exposure to methylmer-cury caused an increase in cerebellar GR activity in mothersand did not change cerebellar GR activity of pups lacta-tionally exposed to this neurotoxicant. So, the ability of pups inresponding to mercury toxicity varies according to the chemicalform of the metal. Taken together, these results and data fromliterature indicate that GR modulation represents an importantcell response to inorganic mercury toxicity, driving cells to themaintenance of the reducing milieu and homeostasis of thiolstatus, which is essential for cell survival and function. Furtherinvestigations are needed to clarify contrasting responsesrelated to GR and different forms of mercury.
GPx belongs to a class of enzymes that catalyzes the reduc-tion of hydroperoxides by GSH and its main function is toprotect against the damaging effects of endogenously formedhydroperoxides [12]. In previous reports, we have showed thatorganomercurials induce lipid peroxidation and this effectappears to be related, at least in part, to the ability of organo-mercurial compounds to inhibit GPx activity [19,18,33]. Al-though our data do not point to any inhibitory effect of inorganicmercury toward cerebellar GPx, we cannot discard the pos-sibility that this metal could inhibit GPx activity in otherexperimental schedules.
In the present work, we could not observe any change inneurochemical parameters evaluated in the cerebellum ofmothers injected with inorganic mercury. In contrast, offspring

366 J.L. Franco et al. / Neurotoxicology and Teratology 29 (2007) 360–367
lactationally exposed to inorganic mercury showed neurochem-ical and behavioral signs of neurotoxicity. In an elegant studyperformed by Sundberg and collaborators [48], it was shownthat inorganic mercury concentrates more in dams' breast milkthan methylmercury. So, a higher load of mercury could betaken up by offspring leading to the observed behavioral andbiochemical changes. Since the developing CNS is more sus-ceptible to metals and oxidative challenge, exposure to in-organic mercury appears to be a health problem during breast-feeding period. Table 2 depicts a comparison between theeffects of the lactational exposure to methylmercury [20,33]and inorganic mercury on behavioral/functional tests and oncerebellar biochemical parameters related to oxidative stress inweanling mice.
In conclusion, the presented results indicate that inorganicmercury exposure through maternal milk is capable of inducingbiochemical changes in the cerebellum of weanling mice, aswell as motor deficits. In addition, these phenomena appear tobe related to the pro-oxidative properties of inorganic mercuryand make relevant the lactational exposure to inorganic mercuryin terms of health concerns.
Acknowledgements
This study was supported by grants from CNPq to M. Farina(475329/2004-0 and 474796/2006-0). J.L. Franco was arecipient of a CAPES fellowship. J. Stringari was a recipientof a CNPq/PIBIC fellowship. H.C. Braga was a recipient of aFAPESC fellowship.
References
[1] M. Aschner, T.W. Clarkson, Distribution of mercury 203 in pregnant ratsand their fetuses following systemic infusions with thiol-containing aminoacids and glutathione during late gestation, Teratology 38 (1988) 145–155.
[2] M. Aschner, K.J. Mullaney, D. Wagoner, L.H. Lash, H.K. Kimelberg,Intracellular glutathione (GSH) levels modulate mercuric chloride (MC)-and methylmercuric chloride (MeHgCl)-induced amino acid release fromneonatal rat primary astrocytes cultures, Brain Res. 664 (1994) 133–140.
[3] M. Aschner, C.P. Yao, J.W. Allen, K.H. Tan, Methylmercury altersglutamate transport in astrocytes, Neurochem. Int. 37 (2000) 199–206.
[4] M. Berlin, Mercury, in: Friberg, G.F. Nordberg, V. Vouk (Eds.), Handbookon the toxicology of metals, II ed., vol. 16, Elsevier Science Publications,1986, pp. 387–445.
[5] M. Berlin, S. Ullberg, Accumulation and retention of mercury in themouse. I. An autoradiographic study after a single intravenous injection ofmercuric chloride, Arch. Environ. Health 6 (1963) 589–601.
[6] J.J. Berzas Nevado, L.F. García Bermejo, R.C. Rodríguez Martín-Doimeadios, Distribution of mercury in the aquatic environment at Almaden,Spain, Environ. Pollut. 122 (2003) 261–271.
[7] R.E. Bluhm, R.G. Bobbitt, L.W. Welch, A.J.J. Wood, J.F. Bonfiglio, C.Sarzen, A.J. Heath, R.A. Branch, Elemental mercury vapor toxicity,treatment, and prognosis after acute, intensive exposure in chloralkali plantworkers. Part I: history, neuropsychological findings and chelator effects,Hum. Exp. Toxicol. 11 (1992) 201–210.
[8] M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein–dyebinding, Anal. Biochem. 72 (1976) 248–254.
[9] I. Carlberg, B. Mannervik, Glutathione reductase, Methods Enzymol. 113(1985) 484–490.
[10] T.W. Clarkson, The toxicology of mercury, Crit. Rev. Clin. Lab. Sci. 34(1997) 369–403.
[11] T.W. Clarkson, L. Magos, G.J. Myers, The toxicology of mercury—current exposures and clinical manifestations, N. Engl. J. Med. 349 (2003)1731–1737.
[12] R. Dringen, P.G. Pawlowski, J. Hirrlinger, Peroxide detoxification by braincells, J. Neurosci. Res. 79 (2005) 157–165.
[13] N.W. Duhan, T.S. Miya, A note on a simple apparatus for detectingneurological deficit in rats and mice, J. Am. Pharm. Assoc. 46 (1957)208–209.
[14] G.L. Ellman, Tissue sulfhydryl groups, Arch. Biochem. Biophys. 82 (1959)70–77.
[15] M. Farina, R. Brandao, F.S. de Lara, L.B. Pagliosa, F.A. Soares, D.O.Souza, J.B. Rocha, Profile of nonprotein thiols, lipid peroxidation anddelta-aminolevulinate dehydratase activity in mouse kidney and liver inresponse to acute exposure to mercuric chloride and sodium selenite,Toxicology 184 (2003) 179–187.
[16] M. Farina, V. Cereser, L.V. Portela, A. Mendez, L.O. Porciuncula,J. Fornaguera, C.A. Goncalves, S.T. Wofchuk, J.B.T. Rocha, D.O. Souza,Methylmercury increases S100B content in rat cerebrospinal fluid, Environ.Toxicol. Pharmacol. 19 (2005) 249–253.
[17] M. Farina, K.C.Dahm, F.D. Schwalm, A.M.Brusque,M.E. Frizzo, G. Zeni,D.O. Souza, J.B. Rocha, Methylmercury increases glutamate release frombrain synaptosomes and glutamate uptake by cortical slices from sucklingrat pups: modulatory effect of ebselen, Toxicol. Sci. 73 (2003) 135–140.
[18] M. Farina, J.L. Franco, C.M. Ribas, F.C. Meotti, F.C. Missau, M.G.Pizzolatti, A.L. Dafre, A.R. Santos, Protective effects of Polygalapaniculata extract against methylmercury-induced neurotoxicity in mice,J. Pharm. Pharmacol. 57 (2005) 1503–1508.
[19] M. Farina, F.A. Soares, G. Zeni, D.O. Souza, J.B. Rocha, Additive pro-oxidative effects of methylmercury and ebselen in liver from suckling ratpups, Toxicol. Lett. 146 (2004) 227–235.
[20] J.L. Franco, A. Teixeira, F.C. Meotti, C.M. Ribas, J. Stringari, S.C. GarciaPomblum, A.M.Moro, D. Bohrer, A.V. Bairros, A.L. Dafre, et al., Cerebellarthiol status and motor deficit after lactational exposure to methylmercury,Environ. Res. 102 (2006) 22–28.
[21] H. Frumkin, R. Letz, P.L. Williams, F. Gerr, M. Pierce, A. Sanders, L.Elon, C.C. Manning, J.S. Woods, V.S. Hertzberg, P. Mueller, B.B. Taylor,Health effects of long-term mercury exposure among chloralkali plantworkers, Am. J. Ind. Med. 39 (2001) 1–18.
[22] S. Goulet, F.Y. Dore, M.E. Mirault, Neurobehavioral changes in micechronically exposed to methylmercury during fetal and early postnataldevelopment, Neurotoxicol. Teratol. 25 (2003) 335–347.
[23] P. Grandjean, P. Weihe, R.F. White, F. Debes, S. Araki, K. Yokoyama, K.Murata, N. Sorensen, R. Dahl, P.J. Jorgensen, Cognitive deficit in 7-year-old children with prenatal exposure to methylmercury, Neurotoxicol.Teratol. 19 (1997) 417–428.
[24] P. Grandjean, R.F. White, A. Nielsen, D. Cleary, E.C. de Oliveira Santos,Methylmercury neurotoxicity in Amazonian children downstream fromgold mining, Environ. Health Perspect. 107 (1999) 587–591.
[25] M. Gul, F.Z. Kutay, S. Temocin, O. Hanninen, Cellular and clinicalimplications of glutathione, Indian J. Exp. Biol. 38 (2000) 625–634.
[26] J.M. Hansen, H. Zhang, D.P. Jones, Differential oxidation of thioredoxin-1, thioredoxin-2, and glutathione by metal ions, Free. Radic. Biol. Med. 40(2006) 138–145.
[27] M. Harada, Minamata disease: methylmercury poisoning in Japan causedby environmental pollution, Crit. Rev. Toxicol. 25 (1995) 1–24.
[28] S. Hussain, D.A. Rodgers, H.M. Duhart, S.F. Ali, Mercuric chloride-induced reactive oxygen species and its effect on antioxidant enzymes indifferent regions of rat brain, J. Environ. Sci. Health, Part B, Pestic. FoodContam. Agric. Wastes 32 (1997) 395–409.
[29] P. Kaur, M. Aschner, T. Syversen, Glutathione modulation influencesmethyl mercury induced neurotoxicity in primary cell cultures of neuronsand astrocytes, Neurotoxicology 27 (2006) 492–500.
[30] C.Y. Kim, K. Nakai, Y. Kasanuma, H. Satoh, Comparison of neurobeha-vioral changes in three inbred strains of mice prenatally exposed tomethylmercury, Neurotoxicol. Teratol. 22 (2000) 397–403.
[31] R. Klein, S.P. Herman, P.E. Brubaker, G.W. Lucier, M.R. Krigman, Amodel of acute methyl mercury intoxication in rats, Arch. Pathol. 93(1972) 408–418.

367J.L. Franco et al. / Neurotoxicology and Teratology 29 (2007) 360–367
[32] L.H. Lash, R.K. Zalups, Alterations in renal cellular glutathionemetabolism after in vivo administration of a subtoxic dose of mercuricchloride, J. Biochem. Toxicol. 11 (1996) 1–9.
[33] C.B. Manfroi, F.D. Schwalm, V. Cereser, F. Abreu, A. Oliveira, L. Bizarro,J.B. Rocha, M.E. Frizzo, D.O. Souza, M. Farina, Maternal milk asmethylmercury source for suckling mice: neurotoxic effects involved withthe cerebellar glutamatergic system, Toxicol. Sci. 81 (2004) 172–178.
[34] M.B. Moretto, J. Franco, T. Posser, C.W. Nogueira, G. Zeni, J.B. Rocha,Ebselen protects Ca2+ influx blockage but does not protect glutamateuptake inhibition caused by Hg2+, Neurochem. Res. 29 (2004) 1801–1806.
[35] D.Y. Moromisato, N.G. Anas, G. Goodman, Mercury inhalation poisoningand acute lung injury in a child, Chest 105 (1945) 613–615.
[36] K. Murata, M. Sakamoto, K. Nakai, P. Weihe, M. Dakeishi, T. Iwata, X.J.Liu, T. Ohno, T. Kurosawa, K. Kamiya, et al., Effects of methylmercury onneurodevelopment in Japanese children in relation to the Madeiran study,Int. Arch. Occup. Environ. Health 77 (2004) 571–579.
[37] H. Ohkawa, N. Ohishi, K. Yagi, Assay for lipid peroxides in animal tissuesby thiobarbituric acid reaction, Anal. Biochem. 95 (1979) 351–358.
[38] Y.C. Ou, C.C. White, C.M. Krejsa, R.A. Ponce, T.J. Kavanagh, E.M.Faustman, The role of intracellular glutathione in methylmercury-inducedtoxicity in embryonic neuronal cells, Neurotoxicology 20 (1999) 793–804.
[39] C.S. Roegge, V.C. Wang, B.E. Powers, A.Y. Klintsova, S. Villareal, W.T.Greenough, S.L. Schantz, Motor impairment in rats exposed to PCBs andmethylmercury during early development, Toxicol. Sci. 77 (2004)315–324.
[40] M. Sakamoto, A. Kakita, K. Wakabayashi, H. Takahashi, A. Nakano, H.Akagi, Evaluation of changes in methylmercury accumulation in thedeveloping rat brain and its effects: a study with consecutive and moderatedose exposure throughout gestation and lactation periods, Brain Res. 949(2002) 51–59.
[41] M. Sakamoto, A. Nakano, Y. Kajiwara, I. Naruse, T. Fujisaki, Effects ofmethyl mercury in postnatal developing rats, Environ. Res. 61 (1993)43–50.
[42] C. Sanfeliu, J. Sebastia, R. Cristofol, E. Rodriguez-Farre, Neurotoxicity oforganomercurial compounds, Neurotox. Res. 5 (2003) 283–305.
[43] D.J. Shamley, J.S. Sack, Mercury poisoning. A case report and commenton 6 other cases, SAMT 76 (1989) 114–116.
[44] G. Shanker, T. Syversen, J.L. Aschner, M. Aschner, Modulatory effect ofglutathione status and antioxidants on methylmercury-induced free radical
formation in primary cultures of cerebral astrocytes, Brain Res. Mol. BrainRes. 137 (2005) 11–22.
[45] H. Sies, Glutathione and its role in cellular functions, Free. Radic. Biol.Med. 27 (1999) 916–921.
[46] J.E. Sirois, W.D. Atchison, Methylmercury affects multiple subtypes ofcalcium channels in rat cerebellar granule cells, Toxicol. Appl. Pharmacol.167 (2000) 1–11.
[47] J. Stringari, F.C. Meotti, D.O. Souza, A.R. Santos, M. Farina, Postnatalmethylmercury exposure induces hyperlocomotor activity and cerebellaroxidative stress in mice: dependence on the neurodevelopmental period,Neurochem. Res. 31 (2006) 563–569.
[48] J. Sundberg, S. Jonsson, M.O. Karlsson, I.P. Hallen, A. Oskarsson,Kinetics of methylmercury and inorganic mercury in lactating andnonlactating mice, Toxicol. Appl. Pharmacol. 151 (1998) 319–329.
[49] J. Sundberg, S. Jonsson, M.O. Karlsson, A. Oskarsson, Lactationalexposure and neonatal kinetics of methylmercury and inorganic mercury inmice, Toxicol. Appl. Pharmacol. 154 (1999) 160–169.
[50] A. Szasz, B. Barna, Z. Gajda, G. Galbacs, M. Kirsch-Volders, M. Szente,Effects of continuous low-dose exposure to organic and inorganic mercuryduring development on epileptogenicity in rats, Neurotoxicology 23(2002) 197–206.
[51] P. Weihe, J.C. Hansen, K. Murata, F. Debes, P. Jorgensen, U. Steuerwald,R.F. White, P. Grandjean, Neurobehavioral performance of Inuit childrenwith increased prenatal exposure to methylmercury, Int. J. CircumpolarHealth 61 (2002) 41–49.
[52] A. Wendel, Glutathione peroxidase, Methods Enzymol. 77 (1981)325–333.
[53] C. Wide, Mercury hazards arising from the repair of sphygmomanometers,Br. Med. J. 293 (1986) 1409–1410.
[54] K.A.Winship, Toxicity of mercury and its inorganic salts, Adv. Drug ReactAc Pois Rev., vol. 3, 1985, pp. 129–160.
[55] World Health Organization, Inorganic mercury — Environmental healthcriteria, Geneva, vol. 118, 1991.
[56] J.M. Yang, X.Z. Jiang, Q.Y. Chen, P.J. Li, Y.F. Zhou, Y.L. Wang, Thedistribution of HgCl2 in rat body and its effects on fetus, Biomed. Environ.Sci. 9 (1996) 437–442.
[57] M. Yoshida, Placental to fetal transfer of mercury and fetotoxicity, TohokuJ. Exp. Med. 196 (2002) 79–88.
Related Documents











