Lack of Selective Developmental Neurotoxicity in Rat Pups from Dams Treated by Gavage with Chlorpyrifos Jacques P. J. Maurissen,* ,1 Alan M. Hoberman,² Robert H. Garman,‡ and Thomas R. Hanley, Jr.§ *The Dow Chemical Company, Midland, Michigan 48674; ² Primedica, Argus Research Laboratories, Inc., Horsham, Pennsylvania 19044; ‡CVP, Murrysville, Pennsylvania 15668 – 0068; and §Dow AgroSciences, LLC, Indianapolis, Indiana 46268 Received October 30, 1998; accepted June 28, 2000 Pregnant Sprague-Dawley rats were given chlorpyrifos (O,O- diethyl-O-[3,5,6-trichloro-2-pyridinyl] phosphorothioate; CPF) in corn oil by gavage from gestation day 6 (GD 6) through lactation day 10 (LD 10) at dosages of 0, 0.3, 1, or 5 mg/kg/day in a developmental neurotoxicity study that conformed to U.S. Envi- ronmental Protection Agency 1991 guidelines. GD 0 was the day when evidence of mating was observed and postnatal day 0 (PND 0) was the day of birth. Toxicity was limited to the highest dosage level (5 mg/kg/day) and, in the dams, consisted of muscle fascic- ulation, hyperpnea, and hyperreactivity. A nonsignificant overall trend toward weight gain and feed consumption was also observed in the high-dosage dams, with a statistically significant Group 3 Time interaction for reduced weight gain in the 5-mg/kg/day group near the end of gestation. Although many developmental indices were normal, pups from high-dosage dams had increased mortal- ity soon after birth, gained weight more slowly than controls, and had several indications of slightly delayed maturation. The early deaths and delayed maturation were attributed to maternal tox- icity, though a possible contributing role of direct pup toxicity in delayed development cannot be eliminated. In spite of the appar- ent delay in physical development, high-dosage pups tested just after weaning had normal learning and memory as tested on a T-maze spatial delayed-alternation task. Habituation, a primitive form of learning, was tested in 2 tasks (motor activity and auditory startle) and was not affected. No overt effects were noted in either dams or pups at 1 or 0.3 mg/kg/day. Based on these data, chlor- pyrifos produced maternal and developmental toxicity in the 5-mg/kg/day-dosage group. There was no evidence of selective developmental neurotoxicity following exposure to chlorpyrifos. Key Words: chlorpyrifos; developmental neurotoxicity; spatial delayed alternation; motor activity; auditory startle; habituation; learning; memory; safety evaluation. Chlorpyrifos (O,O-diethyl-O-[3,5,6-trichloro-2-pyridinyl] phosphorothioate; CPF) is a commonly used organophospho- rous insecticide that has been studied extensively. The database on CPF includes, among other studies, a developmental toxic- ity study and 2-generation reproduction study in rats (Breslin et al., 1996) and a developmental toxicity study in mice (Deacon et al., 1980). There was no evidence from these studies that CPF causes a teratogenic response or is a selective develop- mental toxicant. The study described in this paper was conducted to collect supplementary information about the potential selective devel- opmental neurotoxicity of chlorpyrifos to be used in risk as- sessment. This developmental neurotoxicity study expands upon the existing database for chlorpyrifos in that dosing was conducted during both gestation (GDs 6 –21) and the first 10 days of lactation (LDs 1–10), and that extensive neurobehav- ioral and neuropathological evaluations were conducted on the offspring. MATERIALS AND METHODS Overall Study Conduct The potential developmental neurotoxicity of CPF was evaluated in 2 major studies, conducted concurrently in different laboratories. The study reported here evaluated pups for developmental neurotoxicity after exposure of their dams to CPF during gestation and lactation. This study was designed following the requirements of the U.S. Environmental Protection Agency Pesticide Assessment Guidelines (US EPA, 1991). It followed the Guide for the Care and Use of Laboratory Animals (NRC, 1996), and complied with Good Laboratory Practice regulations (US EPA, 1989). In response to the U.S. Animal Welfare Act that was promulgated by the U.S. Department of Agri- culture (CFR, 1985), the Animal Care and Use Activities that were required for the conduct of this study were reviewed and approved by the Institutional Animal Care and Use Committee. A companion study evaluated CPF and CPF-oxon content of blood and milk, 3,5,6-trichloro-2-pyridinol (TCP) in blood, as well as cholinesterase (ChE) inhibition of plasma, erythrocytes, heart, and brain in dams, fetuses, and pups at several time points. These data are reported in a separate publication (Mattsson et al., 2000). Young adult Sprague-Dawley (Crl:CDtBR VAF/plust) rats were obtained from Charles River Laboratories, Inc., Portage, MI, and bred in-house to provide approximately 20 pregnant dams per dosage level for the develop- mental neurotoxicity element of the study. An extra (satellite) group of 5 pregnant dams per dosage level was added for the determination of maternal plasma, erythrocyte, and brain ChE inhibition on GD 20 (to document expo- sure). GD 0 was defined as the day when spermatozoa were observed in a smear of the vaginal contents and/or a copulatory plug observed in situ; and postnatal day 0 (PND 0) was the day of birth. Bred rats were randomly assigned to the various elements of this study. The test material (CPF) was 99.8% pure and was administered daily to dams by gavage in corn oil, from GD 6 through LD 10, at dosage levels of 0, 0.3 (100 1 To whom correspondence should be addressed at The Dow Chemical Company, Health and Environmental Research Laboratories, 1803 Building, Midland, MI 48674. Fax: (517) 638-9863. E-mail: [email protected]. TOXICOLOGICAL SCIENCES 57, 250 –263 (2000) Copyright © 2000 by The Dow Chemical Company 250

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Lack of Selective Developmental Neurotoxicity in Rat Pupsfrom Dams Treated by Gavage with Chlorpyrifos
Jacques P. J. Maurissen,*,1 Alan M. Hoberman,† Robert H. Garman,‡ and Thomas R. Hanley, Jr.§
*The Dow Chemical Company, Midland, Michigan 48674;†Primedica, Argus Research Laboratories, Inc., Horsham, Pennsylvania 19044;‡CVP, Murrysville, Pennsylvania 15668–0068; and§Dow AgroSciences, LLC, Indianapolis, Indiana 46268
Received October 30, 1998; accepted June 28, 2000
Pregnant Sprague-Dawley rats were given chlorpyrifos (O,O-diethyl-O-[3,5,6-trichloro-2-pyridinyl] phosphorothioate; CPF) incorn oil by gavage from gestation day 6 (GD 6) through lactationday 10 (LD 10) at dosages of 0, 0.3, 1, or 5 mg/kg/day in adevelopmental neurotoxicity study that conformed to U.S. Envi-ronmental Protection Agency 1991 guidelines. GD 0 was the daywhen evidence of mating was observed and postnatal day 0 (PND0) was the day of birth. Toxicity was limited to the highest dosagelevel (5 mg/kg/day) and, in the dams, consisted of muscle fascic-ulation, hyperpnea, and hyperreactivity. A nonsignificant overalltrend toward weight gain and feed consumption was also observedin the high-dosage dams, with a statistically significant Group 3Time interaction for reduced weight gain in the 5-mg/kg/day groupnear the end of gestation. Although many developmental indiceswere normal, pups from high-dosage dams had increased mortal-ity soon after birth, gained weight more slowly than controls, andhad several indications of slightly delayed maturation. The earlydeaths and delayed maturation were attributed to maternal tox-icity, though a possible contributing role of direct pup toxicity indelayed development cannot be eliminated. In spite of the appar-ent delay in physical development, high-dosage pups tested justafter weaning had normal learning and memory as tested on aT-maze spatial delayed-alternation task. Habituation, a primitiveform of learning, was tested in 2 tasks (motor activity and auditorystartle) and was not affected. No overt effects were noted in eitherdams or pups at 1 or 0.3 mg/kg/day. Based on these data, chlor-pyrifos produced maternal and developmental toxicity in the5-mg/kg/day-dosage group. There was no evidence of selectivedevelopmental neurotoxicity following exposure to chlorpyrifos.
Key Words: chlorpyrifos; developmental neurotoxicity; spatialdelayed alternation; motor activity; auditory startle; habituation;learning; memory; safety evaluation.
Chlorpyrifos (O,O-diethyl-O-[3,5,6-trichloro-2-pyridinyl]phosphorothioate; CPF) is a commonly used organophospho-rous insecticide that has been studied extensively. The databaseon CPF includes, among other studies, a developmental toxic-ity study and 2-generation reproduction study in rats (Breslinet
al., 1996) and a developmental toxicity study in mice (Deaconet al., 1980). There was no evidence from these studies thatCPF causes a teratogenic response or is a selective develop-mental toxicant.
The study described in this paper was conducted to collectsupplementary information about the potential selective devel-opmental neurotoxicity of chlorpyrifos to be used in risk as-sessment. This developmental neurotoxicity study expandsupon the existing database for chlorpyrifos in that dosing wasconducted during both gestation (GDs 6–21) and the first 10days of lactation (LDs 1–10), and that extensive neurobehav-ioral and neuropathological evaluations were conducted on theoffspring.
MATERIALS AND METHODS
Overall Study Conduct
The potential developmental neurotoxicity of CPF was evaluated in 2 majorstudies, conducted concurrently in different laboratories. The study reportedhere evaluated pups for developmental neurotoxicity after exposure of theirdams to CPF during gestation and lactation. This study was designed followingthe requirements of the U.S. Environmental Protection Agency PesticideAssessment Guidelines (US EPA, 1991). It followed the Guide for the Careand Use of Laboratory Animals (NRC, 1996), and complied with GoodLaboratory Practice regulations (US EPA, 1989). In response to the U.S.Animal Welfare Act that was promulgated by the U.S. Department of Agri-culture (CFR, 1985), the Animal Care and Use Activities that were required forthe conduct of this study were reviewed and approved by the InstitutionalAnimal Care and Use Committee. A companion study evaluated CPF andCPF-oxon content of blood and milk, 3,5,6-trichloro-2-pyridinol (TCP) inblood, as well as cholinesterase (ChE) inhibition of plasma, erythrocytes, heart,and brain in dams, fetuses, and pups at several time points. These data arereported in a separate publication (Mattssonet al., 2000).
Young adult Sprague-Dawley (Crl:CDtBR VAF/plust) rats were obtainedfrom Charles River Laboratories, Inc., Portage, MI, and bred in-house toprovide approximately 20 pregnant dams per dosage level for the develop-mental neurotoxicity element of the study. An extra (satellite) group of 5pregnant dams per dosage level was added for the determination of maternalplasma, erythrocyte, and brain ChE inhibition on GD 20 (to document expo-sure). GD 0 was defined as the day when spermatozoa were observed in asmear of the vaginal contents and/or a copulatory plug observedin situ; andpostnatal day 0 (PND 0) was the day of birth. Bred rats were randomlyassigned to the various elements of this study.
The test material (CPF) was 99.8% pure and was administered daily to damsby gavage in corn oil, from GD 6 through LD 10, at dosage levels of 0, 0.3 (100
1 To whom correspondence should be addressed at The Dow ChemicalCompany, Health and Environmental Research Laboratories, 1803 Building,Midland, MI 48674. Fax: (517) 638-9863. E-mail: [email protected].
TOXICOLOGICAL SCIENCES57, 250–263 (2000)Copyright © 2000 by The Dow Chemical Company
250

times the chronic RfD), 1.0, and 5.0 mg/kg/day. Dosage volume was 1 ml/kg.The dose was adjusted daily for body-weight changes and given at approxi-mately the same time each day (around 11A.M.).
Bred females were initially housed singly in stainless steel, wire-bottomcages beginning on GD 0. Beginning no later than GD 20, bred female ratswere individually housed in nesting boxes with corn cob bedding (Bed-o’-Cobs, The Andersons Industrial Products Group, Maumee, Ohio). Each damand litter were housed in a common nesting box during the postpartum period.After weaning, the pups were individually housed in stainless steel, wire-bottom cages. An automatically controlled fluorescent light cycle was main-tained at 12-h light:12-h dark, with each dark period beginning at 7P.M. Ratswere given Certified Rodent Diet #5002 (PMI Nutrition, International, St.Louis, MO) availablead libitum from individual feeders. Local water that hadbeen processed by passage through a reverse osmosis membrane was availableto the ratsad libitum from an automatic watering system or individual waterbottles. Chlorine was added to the processed water as a bacteriostat.
On PND 0, all pups in a litter were weighed (pup body weights wererecorded after all pups in a litter were delivered and groomed by the dam). OnPND 4, pups to be culled were selected with a table of random units. Theremaining pups were individually marked with a tattoo on the paw, litters werereduced to 10 pups each, and one male and/or one female pup from each litter
was assigned (when possible) to subsets of rats that were grouped for specificevaluations. Whenever possible, the same number of male and female pups perlitter were continued on study. Due to early pup deaths in the high-dosagegroup, on PND 4, 6 high-dosage litters had fewer than 10 pups. In an attemptto evaluate 20 litters/group with similar maternal and lactational demands ineach dosage group, extra pups randomly culled from other high-dosage litterswere added to those litters of less than 10 pups. When no other high-dosagepups were available, culled pups from other litters were used as needed toincrease the litter size to 10 pups (7 pups from 0.3 mg/kg/day litters and 2 pupsfrom control litters). When pups were taken from other litters and placed withhigh-dosage litters, their sole purpose was to keep all of the evaluated littersequal in size. These replacement pups were uniquely identified with a tattoomark on the tail and were not used for any subsequent evaluation.
The time line of the main study activities is presented in Figure 1. Thesubsets of rats formed for specific evaluations were as follows:
Subset 1. Immersion-fixed brain weights, morphometrics, and neuropa-thology were evaluated in pups on PND 11. One male and one female pup/litterwere first selected (i.e., 80 male and 80 female pups). Half of these (one maleor one female/litter) were culled and examined for gross lesions at necropsy.The other half, i.e., 10/sex/dosage (one male or one female per litter) were
FIG. 1. Schematic time line of the main events of the developmental neurotoxicity study.
251CHLORPYRIFOS DEVELOPMENTAL NEUROTOXICITY

retained for brain weight; of these brains, 6/sex/dosage were examined neu-ropathologically and morphometrically. The sample size per litter was there-fore reduced to 4 males and 4 females on PND 11.
Subset 2. Learning and memory were evaluated by a test of Spatial DelayedAlternation, which was conducted post-weaning (PNDs 22 to 24) and in the samepups as adults (PNDs 61 to 90). Sample size was 8/sex/dosage with only one maleor one female pup/litter selected from 16 randomly chosen litters/dosage.
Subset 3. An automated test of motor activity was conducted on PNDs 13,17, 21, and subsequently on PND 60. Body temperature was measured onPNDs 21 and 60. Auditory startle was evaluated on PNDs 22 and 61. One maleand one female pup from each litter were tested (n 5 20/sex/dosage).
Subset 4. Rats (n 5 20/sex/dosage) were evaluated for body weights(PNDs 0, 4, 11, 17, 21, 39, and 65), clinical signs, day of pinna detachment,eye opening, and preputial separation or vaginal opening. On PNDs 65 to 70,10 rats/sex/dosage (one male or one female/litter) were perfused and theirbrains weighed. Of these, the brains of 6/sex/dosage were examined neuro-pathologically and morphometrically.
Evaluation of Dams
Main study. Dams were examined before the daily treatment, inside andoutside the cage, 3 to 4 h after dosing. An individual unaware of treatmentgroup examined the rats for signs of autonomic dysfunction (lacrimation,salivation, palpebral closure, prominence of the eye, piloerection, respiration,urination, and defecation), abnormal postures, abnormal movements, or ab-normal behavior patterns. The dams were evaluated for duration of gestation(GD 0 to the time the first pup was delivered), litter sizes (all pups delivered),and pup viability at birth. The dams’ behavior was observed daily when thepups were examined during the 22-day postpartum observation period.
Body weights were recorded on GD 0, daily during the dosage period, andon the day sacrificed. Feed consumption values were recorded daily from GD0 through LD 14, when it was expected that pups would begin to consume solidfeed, confounding this parameter.
On LD 21, rats that delivered a litter were sacrificed. A gross necropsy of thethoracic, abdominal, and pelvic viscera was performed and the number anddistribution of implantation sites were recorded. Dams with no surviving pupswere sacrificed after the last pup was found dead or missing and presumedcannibalized. Rats that did not deliver a litter were sacrificed on GD 25 andexamined for gross lesions, pregnancy status and uterine contents. Uteri ofapparently non-pregnant rats were stained with 10% ammonium sulfide toconfirm the absence of implantation sites (Salewski, 1964).
Satellite study. On GD 20, 5 dams per dosage level from the ChE-inhibition satellite group were sacrificed via carbon dioxide inhalation 4 to 5 hafter dosage, to document exposure. A more extensive study of ChE inhibitionand quantitative analysis of chlorpyrifos, its oxon, and trichloropyridinol hasbeen reported elsewhere (Mattssonet al., 2000). Blood was collected from theinferior vena cava, and the brain was removed. The maternal blood (5 ml) wascollected into heparinized centrifuge tubes and centrifuged at 2500g for 5 min,and the plasma was removed and stored undiluted at –70°C. The packederythrocytes were washed with normal saline; the total volume was brought to5 ml and re-centrifuged at 2500g for 5 min. The supernatant was discarded and500 ml of the packed cells was diluted to 5 ml with 0.1 M sodium phosphatebuffer (pH 8.0 with 1% Triton X-100). Maternal brains were homogenized onice [1 g tissue per 9 ml 0.1 M sodium phosphate buffer (pH 8.0 with 1% TritonX-100)]. The brain, plasma, and erythrocyte samples were frozen at –70°Cuntil analyzed. Cholinesterase activity was measured using a radiometric assay(Johnson and Russell, 1975) with a final substrate concentration of 1.2 mM.The [3H]acetate produced by the hydrolysis of [3H]acetylcholine was measuredusing a Wallace 1410 scintillation counter with a counting efficiency ofapproximately 50%.
Evaluation of Pups
All pups were examined for vital status at birth (liveborn vs. stillborn). Pupsthat were found dead during the initial examination of the litters were evalu-
ated by removing and immersing the lungs in water. Pups with lungs that sankwere considered stillborn; pups with lungs that floated were considered live-born, and to have died shortly after birth. Each litter was evaluated for viabilityat least twice each day of the 22-day postpartum period. Dead pups wereremoved from the nesting box. When not precluded by autolysis or cannibal-ization by the dam, any pup found dead was necropsied and examined for thecause of death. The pups present in each litter were counted and physical signs(including gross external physical anomalies) were recorded once each dayduring the 22-day postpartum period. Pup body weight and sex were recordedon PNDs 0, 4, 11, 17, and 21 (all pups on PNDs 0 and 4 were used for bodyweight measurement). Pups were observed for viability at least twice dailyduring the postweaning periods. Body weights and clinical observations werealso recorded on PNDs 39 and 65. Feed consumption was measured on PNDs22 to 29, 39 to 46, and 58 to 65.
Learning and memory. Spatial-delayed alternation was tested in rats in amanner similar to the method used by Stantonet al. (1994). The spatial-delayed alternation apparatus (Coulbourn Instruments, Inc., Allentown, PA)consisted of a Plexiglast T-maze that included a start box, a runway leadingfrom the start box to the choice-point, and left and right runways extendingfrom the choice-point to the left and right goals (supplied with liquid dippers).Light cream (with sweet condensed milk) or water was used as a reinforcer forpups or adults, respectively. All runways were fitted with hinged tops made ofclear Plexiglast. Automated guillotine-type doors were located at the start boxand the left and right sides of the choice-point. The test was divided intodiscrete trials. The latency and the performance of the rat for each trial werecomputer-recorded. Pups were tested on PNDs 22–24, and the same pups weretested as adults on PNDs 61–90.
The spatial-delayed alternation testing was conducted blind to treatment onfeed and/or water-deprived rats. Pups were deprived of feed and water, but hadaccess to both in their home cages for 3 h after the end of the last test sessionof the day, in an attempt to achieve a target of 85% free-feeding weight. At theend of each session, the pups were also eventually fed supplementary lightcream to equate the amount of cream received as a reinforcement by all thegroups during the test session. They were also supplemented with light creamwhen they fell below the target weight. Adult rats were deprived of water only,and were given access to it for half an hour, approximately one h after the lasttest session of the day.
The 3 phases of testing included maze acclimation, acquisition training, anddelay testing. Maze acclimation took place on PND 22 and PNDs 61 to 72, andwas divided into 2 parts: goal box training and forced runs (12 trials). Goal boxtraining allowed the rat access to the reward in both the right and left arm ofthe maze. Forced runs allowed the rat to access only one arm of the maze,where the reward was available (either the right or the left).
Acquisition training was comprised of a forced run immediately followed bya choice run (7 blocks of 12 trials on PNDs 23 and 24, and 10 blocks of 6 trialson PNDs 73 to 82). In the forced run, the rat was given access to one arm ofthe T-maze that was baited. In the choice run, the doors to both arms of themaze were raised simultaneously. To have access to the reward, the rat had tochoose the opposite arm from the one previously rewarded in the forced run(spatial alternation). The percent correct was calculated in blocks of 12consecutive trials, and the learning curve was represented by the averagepercent correct (groups of 8 rats each) over successive blocks.
Delay testing was conducted under the same conditions as acquisitiontraining, with time delays of various durations added between the forced runand the choice run (4 blocks of 12 trials on PND 24, and 6 blocks of 6 trialson PNDs 85 to 90). Delays alternated every 6 trials. Each animal was evaluatedevery test day. The percent correct retention could be plotted at each delay(“forgetting” function). The memory component was evaluated by the reten-tion curve and was represented by the slope corresponding to the percentcorrect retention at the tested delays. The intercept of the slope was anextrapolated value (time zero) and gave information about the non-mnemonicaspects of the task (e.g., changes in motivation, sensory-motor functions,attention, and encoding of information). On PND 24, the delays were set at 5,15, and 25 s, while they were set at 5, 65, and 125 s for PNDs 85 to 90.
252 MAURISSEN ET AL.

Motor activity. Motor activity was evaluated on PNDs 13, 17, 21, and 60by counting beam breaks of a passive infrared sensor mounted outside astainless-steel wire-bottom cage with Plexiglast flooring (40.63 25.43 17.8cm). Each test session was one h in duration with the number of beam breakstabulated for each 5-min interval. The apparatus monitored a rack of up to 32cages and sensors during each session, with each rat tested in the same locationon the rack across test sessions. Groups were counterbalanced (sex, dosagegroup) across testing sessions and cages. Data were collected to demonstratethat the test system was capable of detecting increases or decreases in activityproduced by positive control substances. The calibration procedure for themotor activity equipment was performed at least semiannually.
Auditory startle. The auditory startle test was conducted on PNDs 22 and61. Animals were evaluated in sets of 4 within a sound-attenuated chamber,and the amplitude and latency of the startle response were measured (Coul-bourn Instruments, Inc., Allentown, PA). Each rat was placed inside a cagesituated above a platform containing a pressure transducer in its base. The cagehad the overall shape of a hexahedron with an 8-cm base, 8-cm height and16-cm length for younger rats, and with a 9-cm base, 9.5-cm height and18.5-cm length for older rats. A microcomputer sampled the output of thepressure transducer and controlled the test session. The rats were initially givenan adaptation period of five min. During the last minute of this period, ten“blank” trials (no startle auditory stimulus) were given to sample the baselinepressure exerted by the rat. The rats were then presented with 20-ms, 120-dBAbursts of white noise at 10-s intervals for 50 trials. An additional ten blanktrials followed. The peak amplitude of each response was recorded, and theaverage response on baseline trials was subtracted to calculate the peakresponse. The average peak responses and latencies over 10-trial blocks werecompared among the dosage groups. The calibration of auditory signals wasperformed prior to the start of the study. The calibration of the auditory startlepressure transducers was performed daily prior to usage. Groups were coun-terbalanced (sex, dosage group) across testing sessions and cages.
Body temperature. Body temperature was measured on PNDs 21 and 60,immediately following the completion of motor activity testing. The measure-ment was made with a rectal probe (Physitemp Instruments Model RET-2) anddigital thermometer (Physitemp Instruments Model Bat-10 R LOP). The digitalthermometer was calibrated daily prior to usage.
Physical signs of maturation. Pinna unfolding was monitored daily be-ginning on PND 1 (all rats in all litters) and continued until all pups (100%)had the pinna unfolded. In rats of subset 4, eye opening was monitoredbeginning on PND 11 and continued until each pup reached the criterion. Malerats were evaluated for the age at preputial separation beginning on PND 38.Female rats were evaluated for the age at vaginal patency beginning on PND27. A body weight was recorded for the rat on the day the criterion wasattained.
Brain weight, histopathology and morphometrics.At PND 11, 10 maleand 10 female pups (one per litter) were euthanized by carbon dioxideoverexposure. The head of each pup was severed between the first two cervicalvertebrae, the calvaria removed from the top of the skull, and the entire headplaced into neutral buffered 10% formalin. After fixation, brains were removedand weighed. Brains from 6/sex/dosage were subsequently examined his-topathologically. During PNDs 65 to 70, ten rats/sex/dosage group wererandomly selected for brain weight measurements. These grown pups wereadministered a combination of heparin and an anesthetic (39 mg/kg sodiumpentobarbital and 20,000 USP/kg sodium heparin) and perfusedin situ withneutral buffered 10% formalin. Of these 10, 6 were randomly selected forneuropathologic evaluation.
In addition to neuropathologic examinations on PND-11 and PND-65 rats,morphometric measurements (6 rats/sex/dosage) were made of the anterior toposterior length of the cerebrum and cerebellum, height of the cerebellum,thickness of the corpus callosum, frontal cortex, parietal cortex, caudate-putamen, hippocampus, and in pups only, the thickness of the external germi-nal layer of the cerebellum. Linear measurements of the anterior-posteriordimensions of the cerebrum and cerebellum were made with a Vernier caliper.The brains were then weighed and divided into 6 coronal slices. The cuts were
made half way between the ventral base of the olfactory bulbs and the opticchiasm, through the optic chiasm, through the infundibulum, through themidbrain just posterior to the mammillary body, through the cerebellum justanterior to its midpoint, and finally through the anterior portion of the medulla.The anterior faces of these coronal brain slices were sectioned (7mm),paraffin-embedded, and stained with hematoxylin and eosin. After processing,the slides were coded and evaluated, blind to treatment.
Statistical Analyses
Statistical analyses were performed with SAS/STATr (SAS Institute, 1989)programs or by proprietary programs developed for computer use by theTesting Facility. All tests were 2-tailed. Individual motor activity counts weretransformed to square roots. Bartlett’s Tests (Sokal and Rohlf, 1969) forhomogeneity of variance were run ata 5 0.001.
Parametric data were analyzed by factorial analysis of variance (ANOVA,factors of group and sex) or factorial repeated-measure analyses (Rep-ANOVA, factors of group, sex, repeated for time), using the multivariateapproach and the Pillai trace statistic (SAS Institute, 1989). The main effectsand interactions of interest were:Group: A significantp value indicated thatboth male and female pups, taken together, differed in some way among thedifferent groups, independent of time.Group 3 Sex: A significant p valueindicated that male and female pups differed among the different groups(independent of time) in the way they reacted to the test compound.Group3Time: A significantp value indicated that both male and female pups, takentogether, differed in some way among the different groups at some timeinterval. Group 3 Time 3 Block (motor activity and auditory startle):Asignificant p value indicated that group effects were different among thedifferent blocks at some time interval. TheGroup 3 Sex3 Time factor wasalso added by request, and examinedpost hoc. A significantp value indicatedthat group effects were different in males and females at some time interval.
In the event of a statistically significantGroupmain effect (in the presenceor absence of any other statistically significant interaction), one dosage level(beginning with the high-dosage group) was removed, and the same analysiswas re-run. If an overall statistically significant effect was present, but disap-peared when the high-dosage group data were removed, it was inferred that thestatistically significant effect was due to the high-dosage group (Tukeyet al.,1985). This approach minimizes the number of comparisons (and, therefore, ofType-I errors) and also of Type-II errors (by maximizing the sample size). Thefollowing sequential approach was used in an effort to further reduce Type-Iand Type-II errors: IfGroup 3 Sexwas significant, the same analysis wasrerun on separate sexes. If theGroup 3 Time interaction was significant, thedata were examined to identify the time period responsible for the statisticalsignificance. The Type I-error rate was set at 0.02 for all primary analyses.Sequential analyses were conducted ata 5 0.02, following a statisticallysignificant primary planned analysis (consistent with the recommendationsproposed by Tukeyet al.,1985). When one male and one female from the samelitter were used in a test, the litter was used as the unit for analysis. Allp valueswere reported uncorrected.
Brain morphometric data were analyzed by ANOVA for males and femalesseparately, followed by Dunnett’s tests if the ANOVA was statistically sig-nificant. This approach was taken because the number of dosage groupsexamined was not the same, either in males and females, or at the 2 time pointsanalyzed (i.e., male and female pups: morphometric data from control, low-,middle-, and high-dosage groups; male adults: control and high-dosage groups,female adults: control, middle- and high-dosage groups). Such a differenceamong the number of groups selected for morphometric analysis came fromthe fact that, at first, the pups from all dosage groups were prepared formorphometric analyses. However, for adults, the stepwise approach suggestedin the EPA developmental neurotoxicity guideline (US EPA, 1991) was used,which recommends that control and high-dosage groups be examined first, andthen, if evidence of alterations is found, lower dosage groups be examined insequence. Analysis was performed on all tissues prepared for morphometricanalysis. The Type-I error rate was set arbitrarily at 0.05 because this type ofanalysis has less statistical power than the repeated-measure analyses.
253CHLORPYRIFOS DEVELOPMENTAL NEUROTOXICITY

Developmental landmarks were evaluated using the Kruskal-Wallis (Sokaland Rohlf, 1969) (pinna unfolding, vaginal patency, preputial separation, eyeopening;a 5 0.02). Clinical observations were evaluated using the Chi-squaretest for proportions (Snedecor and Cochran, 1967).
More than 200a priori p values were derived in this study, out of whichapproximately 40 were statistically significant. These figures provide a contextfor a better interpretation of the results.
RESULTS
Dams
Gavage administration of 5 mg/kg/day of CPF to pregnant/lactating female rats produced clinical signs of toxicity. Table1 shows the total number of rats exhibiting clinical signs duringGD 6–21 and LD 1–21. Muscle fasciculation was noted duringgestation (p 5 0.001; in all cases these signs occurred post-dosing on the last day of gestation) and during lactation (p 50.001; primarily during the first 4 days of lactation). In addi-tion, signs of hyperreactivity (p 5 0.001) and hyperpnea (p 50.001) were observed in a number of rats in the high-dosage (5mg/kg/day) group during lactation. These clinical signs wereobserved in high-dosage animals immediately prior to delivery,when body weight was the highest and consequently the ab-solute dose in mg administered was the highest, and during thefirst few days of lactation. There were no clinical signs oftreatment-related toxicity in rats given 0.3 or 1.0 mg/kg/day.Though sporadic incidences of hyperreactivity were noted incontrols and the lower dosage groups, these were not dosage-dependent and not considered related to treatment. All otherclinical observations were infrequent, lacked a pattern, andwere considered spontaneous occurrences unrelated to chlor-pyrifos administration (missing/broken incisors, localized alo-pecia, hind limb, head, or tail lesions, red perivaginal substance[data not shown]).
There were no overall statistically significant differences indams’ gestation (p 5 0.94;F(3, 93)5 0.13) or lactation (p 50.48; F(3, 75) 5 0.84) body weights (data not shown). Body
weight gains are depicted in Figure 2. Though there were nostatistically significant differences in body weight gain over theentire gestation period (p 5 0.09;F(3, 93)5 2.20; GDs 0–20)in any dosage group, a statistically significant Group3 Timeinteraction (p 5 0.006;F(9, 279)5 2.64) was present in bodyweight gain, and the 5-mg/kg/day dosage group, toward theend of gestation (GDs 17–20), appeared to be responsible forthis effect (Fig. 2). During lactation, no overall statisticallysignificant differences in body weight gains were noted (p 50.36; F(3, 75) 5 1.08; LDs 0–21), and the Group3 Timeinteraction did not reach statistical significance (p 5 0.04;F(12, 222)5 1.86). However, a transiently lower mean lacta-tion weight gain was observed in the 5-mg/kg/day dosagegroup on LDs 0–3 (6.86 10.7 g in the control group vs. 1.166.5 g in the high-dosage group). This reduced body weight gaintoward the end of gestation and the beginning of lactation(even though not statistically significant) was considered treat-ment related. No differences in body weights and body weightgains were apparent in the 0.3 or 1.0 mg/kg/day dosage groupswhen compared to controls.
There were no overall significant differences in dams’ ab-solute and relative feed consumption (relative to individualbody weight) during gestation (p 5 0.78;F(3, 91)5 0.36, andp 5 0.64;F(3, 91)5 0.57) or during lactation (p 5 0.04;F(3,54) 5 3.07, and 0.12;F(3, 54)5 2.02) in any treatment group(data not shown). During lactation, however, feed consumptionwas 4 to 15% lower in the high-dosage group, with the greatesteffect during the period immediately following parturition.
Table 2 presents ChE activity as a percent of control values,measured in a subgroup of dams sacrificed on GD 20 (satellitestudy). At 5 mg/kg/day of chlorpyrifos, brain and plasma ChEactivity was markedly depressed (approximately 90%) whencompared to controls, and RBC ChE levels were depressedalmost completely. At 1.0 mg/kg/day, dams’ brain ChE activitywas depressed approximately 18% relative to controls, whileRBC ChE levels were inhibited approximately 84% andplasma levels were depressed approximately 69%. At the low
FIG. 2. Maternal body weight gains during gestation and lactation(mean1 SD). Notice the reduced weight gain in the 5-mg/kg/day group duringGDs 17–20 and LDs 0–3. Maternal dosing took place from GD 6 throughLD 10.
TABLE 1Summary of Clinical Observations of Dams
Dosage (mg/kg/day) 0 0.3 1.0 5.0
During gestationNo. animals 25 25 25 25Fasciculations 0 0 0 6a
Ataxia 0 0 0 1Tremors 1 0 0 0Excessive salivation 1 0 0 0
During lactationNo. animals 25 24 24 24Fasciculations 0 0 0 16a
Hyperreactivity 2 7 2 17a
Hyperpnea 0 0 1 8a
Note.Total number of rats showing clinical signs.aStatistically significant (a 5 0.02).
254 MAURISSEN ET AL.

dosage of 0.3 mg/kg/day, there was no indication of effect onbrain cholinesterase activity, while plasma and RBC ChElevels were approximately 45% lower than control values.
Developmental Indices
The developmental parameters measured in dams are pre-sented in Table 3. All pregnant rats (at least 24 per group)delivered litters. The duration of gestation, the number ofimplantations per dam, the number of dams with stillborn pups,and the mean litter size (surviving pups) on PND 0 werecomparable across the groups. A total of 3 dams had total litterloss (accounting for 31 pups) between PNDs 0 and 4. DuringPNDs 1–4, there were 2, 2, 3 and 38 pups cannibalized in the0, 0.3, 1, and 5 mg/kg/day groups respectively. This resulted in
a substantial decrease in litter size on PND 4, and a reducedviability index in the 5 mg/kg/day dosage group. Overall, 30%(88/292) of the fetuses born to dams given 5 mg/kg/day diedduring the PND 0–4 interval, compared to approximately1–2% in all other groups. Following culling, there were nooverall significant differences in mortality in any of the treat-ment groups.
Statistically significant lower male and female pups’ bodyweights were seen at 5 mg/kg/day on PNDs 0–4 (p , 0.001;F(3, 163) 5 11.15); a significant Time3 Group interaction(p , 0.001;F(6, 326)5 5.14) may indicate a larger effect ona particular time period (i.e., PND 4).
When pups’ body weights were evaluated over a longerperiod (i.e., PNDs 0–65), the analysis showed that they werelower in the high-dosage group in both males and femalescompared to controls (p , 0.001;F(3, 75)5 14.07) (Fig. 3).A significant Sex3 Group effect (p 5 0.02;F(3, 71)5 3.52)indicated that males and females were differentially affected bytreatment. However, when the data were analyzed separatelyby sex, the analyses showed that the body weights of bothmales (p , 0.001;F(3, 73)5 11.47) and females (p , 0.001;F(3, 73) 5 8.13) were significantly decreased at the highdosage. A Day3 Group significant effect (p , 0.001;F(18,216) 5 2.80) indicated that the difference between groups forboth sexes taken together changed as a function of day (e.g.,more of a relative effect on PND 65 compared to PND 0). ByPND 65, the weights of high-dosage female offspring were
TABLE 2Cholinesterase Activity in Dams on GD 20
Dosage (mg/kg/day)a 0.3 1.0 5.0
Plasma 56.76 2.7 31.16 4.1 8.56 1.2Erythrocyte 58.76 16.1 15.66 6.8 0.16 0.2Brain 99.76 1.5 82.16 2.8 10.26 1.0
Note.Values are given as percentage of control6 SEM. The control valuesare 861, 653, and 11,296 nmol acetylcholine metabolized/min/ml (or mg) inplasma, erythrocyte and brain, respectively.
aSample size5 5 rats/dosage group.
TABLE 3Summary of Developmental Parameters
Dosage (mg/kg/day) 0 0.3 1.0 5.0
No. animals 25 25 25 25No. pregnant 25 24 24 24No. litters 25 24 24 24Mean gestation length (days6 SD) 23.16 0.5 23.26 0.4 23.06 0.5 23.06 0.2Mean implantations per dam (6 SD) 13.66 3.2 14.56 2.0 14.26 2.4 14.26 1.6No. dams with stillborn pups 1 1 4 1No. dams with total litter loss 0 0 0 3a
Mean litter size (pups6 SD)Liveborn 12.36 3.1 13.36 1.9 13.06 2.4 12.76 2.4Stillborn 0.06 0.2 0.06 0.2 0.26 0.5 0.16 0.6PND 4 (live; pre-cull) 12.26 3.1 13.16 1.7 12.76 2.5 8.96 4.9a
Pup mortalityPND 0 0/308 1/319 2/311 21/292a
PNDs 1–4 4/308 3/318 4/309 67/271a
Mean pup weights (g6 SD)PND 0
Males 6.66 0.6 6.76 0.6 6.46 0.5 6.16 0.7a
Females 6.36 0.6 6.26 0.5 6.16 0.5 5.66 0.6a
PND 4 (precull)Males 9.96 0.9 10.26 1.1 10.36 1.6 8.86 1.6a
Females 9.66 0.9 9.76 1.1 9.76 1.5 8.26 1.6a
Viability index (%)b 98.7 98.7 98.1 69.9a
aStatistically significant (a 5 0.02).bNumber of live pups on PND 4 (preculling)/number of liveborn pups on PND 0.
255CHLORPYRIFOS DEVELOPMENTAL NEUROTOXICITY

comparable to the controls, while the weights in male offspringremained lower.
Overall, the body weight changes followed the pattern ofeffects seen in body weights; however, the effect on males wasstatistically significant (p , 0.001;F(3, 73)5 6.77), but not infemales (p 5 0.08;F(3, 73)5 2.36). The Day3 Sex3 Groupwas also statistically significant (p , 0.008; F(15, 207) 52.17), and may indicate that males and females reacted differ-ently to treatments mainly after day 22. None of the effectsmentioned above were seen when the high-dosage group wasremoved from the analysis.
The feed consumption analyses revealed a main Group ef-fect (p , 0.001; F(3, 75) 5 8.27) in the high-dosage groupwithout a significant Sex3 Group interaction (p 5 0.22;F(3,62) 5 1.53). The biological significance of the Day3 Sex3Group interaction (p , 0.01;F(6, 124)5 2.86) is unclear, butmay reflect a tendency for a small reduction, over time, of thedifference in feed consumption between control and high-dosage groups that may be more accentuated in males than infemales.
The litter losses/reductions and lower body weights of off-spring in the high-dosage group (PNDs 0–4) correspondedtemporally with clinical signs, cannibalization of pups, a trendtoward reduced feed consumption, and body weights in thedams. There were no effects of treatment with chlorpyrifos inthe 0.3- or 1-mg/kg/day dosage groups.
Developmental Landmarks
In addition to the changes in pups’ body weights, delays insome developmental landmarks were present in pups of dams
treated with 5 mg/kg/day (Table 4). The only developmentallandmark which was identified as statistically significantlydifferent from the controls was delayed vaginal opening (p 50.01). However, pinna detachment (p 5 0.03) and preputialseparation (p 5 0.05) also appeared to be somewhat delayed,though there were no statistically significant differences. Therewere no effects at either 0.3 or 1.0 mg/kg/day in any of thesemeasures.
Brain Measurements
The mean absolute and relative brain weights in offspringmeasured on PND 11 and PND 65 (adults) are presented inFigure 4. On PND 11, absolute neonatal brain weight (Table 5)was statistically significantly decreased at 5 mg/kg/day, and thebrain-to-body weight ratio was increased at the same dosage.There were 9 areas of the brain measured in PND-11 pups (8for adult rats), either grossly or microscopically, for length andlayer thickness (Table 5). Five of these linear measures (cere-brum length, cerebellum length and height, parietal cortex,hippocampal gyrus) were statistically significant overall at theANOVA level in males, and 2 (caudate-putamen and cerebel-lum height) in females. The Dunnett’s tests confirmed a sta-tistically significant difference between control and high-dos-age groups in cerebellum length and height in males, and incaudate-putamen in females. Another difference found be-tween control and the medium-dosage group in cerebellum washeight in females. Examination of the data revealed that mostof the absolute measurements (except for the external germinallayer) were smaller in the high-dosage males and femalescompared to controls (Table 5). It is interesting to note that themajority of the smallest layers were found in the smallestbrains, i.e., in the high-dosage pups. The lack of statisticalsignificance in some layer thicknesses is consistent with thesmall sample size (n 5 6/sex/dosage), the relatively small sizeof the effect, and the slightly larger coefficient of variation in
TABLE 4Summary of Developmental Landmarks in Pups
Dosage (mg/kg/day) 0 0.3 1.0 5.0
Pinna detachmenta 2.56 0.7 2.56 0.5 2.66 0.6 3.06 0.6Eye openingb
Males 13.66 0.9 13.66 0.8 13.76 0.7 13.96 1.0Females 13.46 1.0 13.06 0.8 13.46 0.7 13.66 1.1
Vaginal openingc 31.46 1.0 30.56 1.5 31.16 2.3 32.46 2.2d
Preputial separatione 43.26 1.9 42.36 1.9 44.26 3.2 46.06 5.9
Note.Values are given as mean6 SD.aAverage PND on which at least 50% of the pups in a litter exhibited
unfolding of the pinna.bAverage PND on which all pups in every litter were observed with open
eyes.cAverage PND that the vagina was observed to be patent.dStatistically significant (a 5 0.02).eAverage day postpartum that the prepuce was observed to be separated.
FIG. 3. Pup body weights (mean). Overall statistically significant differ-ence in male and female pups of the high-dosage group compared to controls.Maternal dosing took place from GD 6 through PND 10.
256 MAURISSEN ET AL.

layer thickness (in comparison to brain weights’ coefficient ofvariation).
In PND-65 rats, no statistically significant differences wereseen in brain weight or in any linear dimensions of the brain,
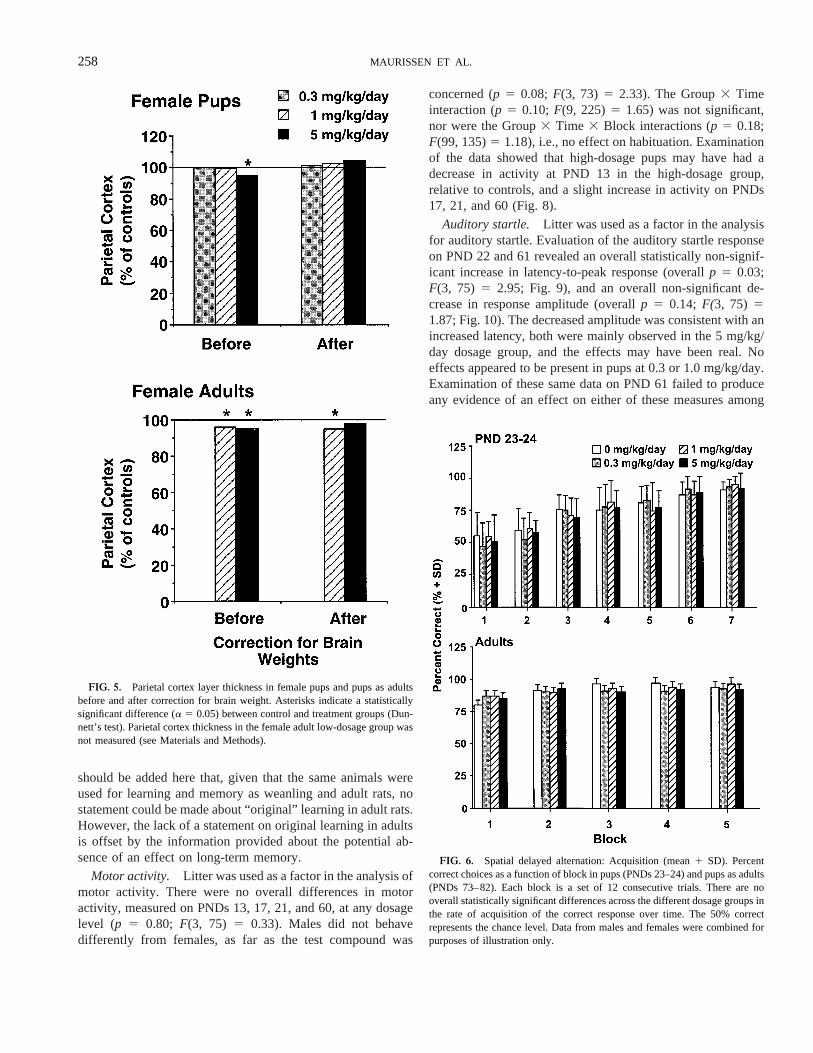
except in the parietal cortex of adult females in medium- andhigh-dosage groups compared to control. These decreases wereabout the same (4.2 and 5.1%, respectively), and were seen inadult females but not in adult males. Given that brain andbrain-layer thickness normally would be correlated, the datafor pup and adult female rats were plotteda posteriori,beforeand after adjustment for brain weight in Figure 5. The upperpanel shows that the 5-mg/kg/day female pup group is signif-icantly different from the corresponding control group. Whenthe same data are plotted after correction for brain weights, thestatistically significant difference disappears and a 4.2% in-crease in relative layer thickness is recorded in the high-dosagegroup. The lower panel shows that, after correction for brainweight, the 5-mg/kg/day female adult group is not statisticallydifferent from the appropriate control group.
Neurobehavioral Assessment
The results of the neurobehavioral assessment of pups arepresented in Figures 6–10.
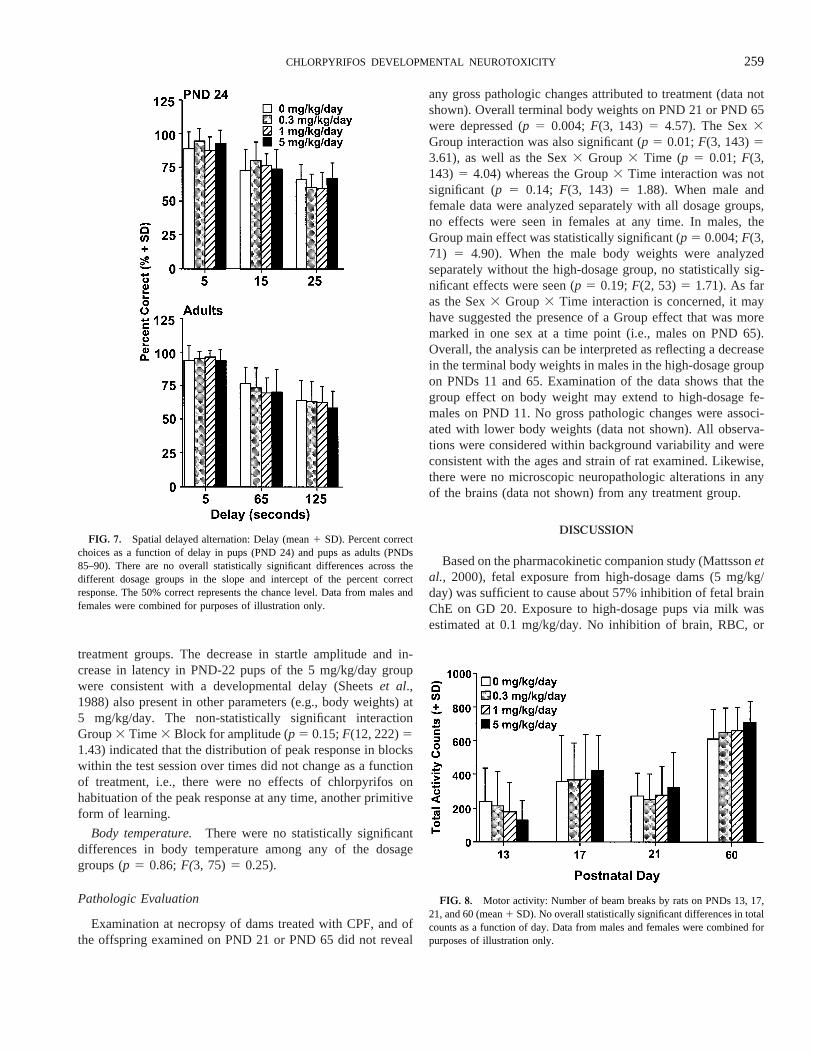
Learning and memory. There were no effects of exposureon cognitive functions in any dosage group on PNDs 22–24 orin the same rats on PNDs 61–91 (adults). Figures 6 and 7demonstrate the lack of any effect of CPF on learning acqui-sition (p 5 0.92;F(3, 55)5 0.17 for pups, andp 5 0.80;F(3,54) 5 0.33 for adults), short-term retention (overallp 5 0.45;F(3, 54) 5 0.89 for the slope of the forgetting curve), ornon-mnemonic factors (overallp 5 0.67;F(3, 54)5 0.52 forintercept), e.g., motivation, attention, sensori-motor perfor-mance, in any of the dosage groups at either time point. Furthera posteriori examination of Figure 6 reveals that the percentcorrect in Block 1 (consisting of the first 12 trials) in adults wasabove random performance and that there did not appear to beany differences in long-term memory among dosage groups. It
FIG. 4. Brain weight in pups (PND 11) and in pups as adults (PND 65)(mean1 SD). Absolute brain weights (upper left panel) were significantlydecreased in the high-dosage group on PND 11. Brain to body weight ratioswere significantly increased in the same group at the same time (upper rightpanel). Data from males and females were combined for purposes of illustra-tion only.
TABLE 5PND 11 Brain Measurements
Dosage(mg/kg/day)
Brain wt(g)
Ant./Post.cerebrum
(mm)
Ant./Post.cerebellum
(mm)
Frontalcortex(mm)
Parietalcortex(mm)
Caudate-putamen
(mm)
Corpuscallosum
(mm)Hippocampus
(mm)Cerebellum ht
(mm)
Ext9l germlayer(mm)
Males0 1.286 0.04 12.56 0.3 3.36 0.3 13486 53 13366 54 22406 84 2936 25 9046 93 35046 129 376 2*0.3 1.416 0.07 13.46 0.5 3.46 0.4 13606 100 14486 58 22406 108 3036 24 10046 114 34566 172 386 41 1.366 0.08 13.16 0.5 3.36 0.2 13526 47 14486 33 23126 93 2906 36* 9726 54 34166 200 406 75 1.176 0.16a,* 11.86 1.0* 2.56 0.6a,* 12726 153* 12566 138* 22246 147* 2936 56 8246 66* 30086 504a,* 386 3
Females0 1.286 0.08 12.46 0.3 3.26 0.2 13766 92 13806 54 23846 131 3076 38 9366 82 35126 200 396 30.3 1.286 0.04 12.76 0.3 3.06 0.3* 13886 79 13766 20 22246 116 2866 27 9126 50 31766 130 366 6*1 1.276 0.11 12.86 0.7 3.36 0.2 13566 54* 13686 80 22886 108 3046 36 9326 96 31206 328a,* 416 65 1.176 0.13a,* 12.26 0.6* 3.06 0.3* 13686 86 13046 72* 21526 134a,* 2746 40* 8286 79* 32086 226 416 6
Notes.Values given are mean6 SD. Overall statistically significant ANOVA for males (Brain wt, Ant./Post. cerebrum, Ant./Post. cerebellum, Parietal cortex,Hippocampus, and Cerebellum ht) and females (Brain wt, Caudate-putamen, and Cerebellum ht) separately (a 5 0.05).
aStatistically significant Dunnett’s test (a 5 0.05).*Smallest values.
257CHLORPYRIFOS DEVELOPMENTAL NEUROTOXICITY
should be added here that, given that the same animals wereused for learning and memory as weanling and adult rats, nostatement could be made about “original” learning in adult rats.However, the lack of a statement on original learning in adultsis offset by the information provided about the potential ab-sence of an effect on long-term memory.
Motor activity. Litter was used as a factor in the analysis ofmotor activity. There were no overall differences in motoractivity, measured on PNDs 13, 17, 21, and 60, at any dosagelevel (p 5 0.80; F(3, 75) 5 0.33). Males did not behavedifferently from females, as far as the test compound was
concerned (p 5 0.08; F(3, 73) 5 2.33). The Group3 Timeinteraction (p 5 0.10; F(9, 225)5 1.65) was not significant,nor were the Group3 Time 3 Block interactions (p 5 0.18;F(99, 135)5 1.18), i.e., no effect on habituation. Examinationof the data showed that high-dosage pups may have had adecrease in activity at PND 13 in the high-dosage group,relative to controls, and a slight increase in activity on PNDs17, 21, and 60 (Fig. 8).
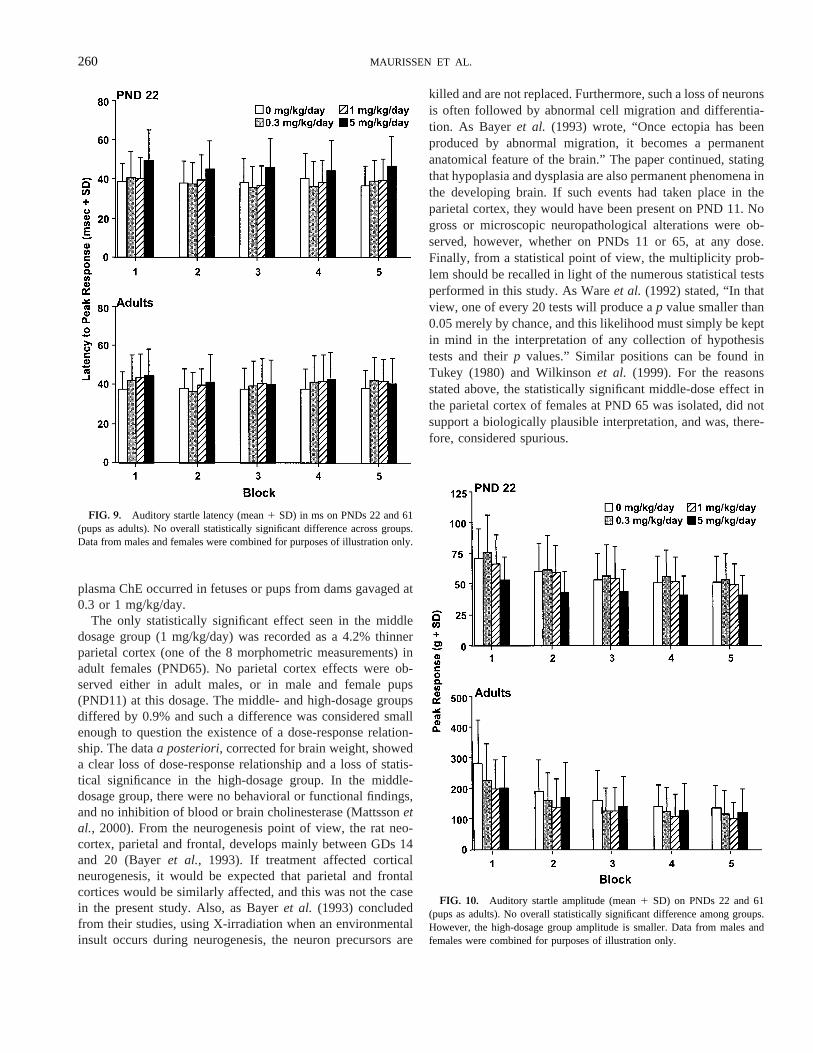
Auditory startle. Litter was used as a factor in the analysisfor auditory startle. Evaluation of the auditory startle responseon PND 22 and 61 revealed an overall statistically non-signif-icant increase in latency-to-peak response (overallp 5 0.03;F(3, 75) 5 2.95; Fig. 9), and an overall non-significant de-crease in response amplitude (overallp 5 0.14; F(3, 75) 51.87; Fig. 10). The decreased amplitude was consistent with anincreased latency, both were mainly observed in the 5 mg/kg/day dosage group, and the effects may have been real. Noeffects appeared to be present in pups at 0.3 or 1.0 mg/kg/day.Examination of these same data on PND 61 failed to produceany evidence of an effect on either of these measures among
FIG. 6. Spatial delayed alternation: Acquisition (mean1 SD). Percentcorrect choices as a function of block in pups (PNDs 23–24) and pups as adults(PNDs 73–82). Each block is a set of 12 consecutive trials. There are nooverall statistically significant differences across the different dosage groups inthe rate of acquisition of the correct response over time. The 50% correctrepresents the chance level. Data from males and females were combined forpurposes of illustration only.
FIG. 5. Parietal cortex layer thickness in female pups and pups as adultsbefore and after correction for brain weight. Asterisks indicate a statisticallysignificant difference (a 5 0.05) between control and treatment groups (Dun-nett’s test). Parietal cortex thickness in the female adult low-dosage group wasnot measured (see Materials and Methods).
258 MAURISSEN ET AL.
treatment groups. The decrease in startle amplitude and in-crease in latency in PND-22 pups of the 5 mg/kg/day groupwere consistent with a developmental delay (Sheetset al.,1988) also present in other parameters (e.g., body weights) at5 mg/kg/day. The non-statistically significant interactionGroup3 Time3 Block for amplitude (p 5 0.15;F(12, 222)51.43) indicated that the distribution of peak response in blockswithin the test session over times did not change as a functionof treatment, i.e., there were no effects of chlorpyrifos onhabituation of the peak response at any time, another primitiveform of learning.
Body temperature. There were no statistically significantdifferences in body temperature among any of the dosagegroups (p 5 0.86;F(3, 75)5 0.25).
Pathologic Evaluation
Examination at necropsy of dams treated with CPF, and ofthe offspring examined on PND 21 or PND 65 did not reveal
any gross pathologic changes attributed to treatment (data notshown). Overall terminal body weights on PND 21 or PND 65were depressed (p 5 0.004; F(3, 143) 5 4.57). The Sex3Group interaction was also significant (p 5 0.01;F(3, 143)53.61), as well as the Sex3 Group 3 Time (p 5 0.01; F(3,143) 5 4.04) whereas the Group3 Time interaction was notsignificant (p 5 0.14; F(3, 143) 5 1.88). When male andfemale data were analyzed separately with all dosage groups,no effects were seen in females at any time. In males, theGroup main effect was statistically significant (p 5 0.004;F(3,71) 5 4.90). When the male body weights were analyzedseparately without the high-dosage group, no statistically sig-nificant effects were seen (p 5 0.19;F(2, 53)5 1.71). As faras the Sex3 Group3 Time interaction is concerned, it mayhave suggested the presence of a Group effect that was moremarked in one sex at a time point (i.e., males on PND 65).Overall, the analysis can be interpreted as reflecting a decreasein the terminal body weights in males in the high-dosage groupon PNDs 11 and 65. Examination of the data shows that thegroup effect on body weight may extend to high-dosage fe-males on PND 11. No gross pathologic changes were associ-ated with lower body weights (data not shown). All observa-tions were considered within background variability and wereconsistent with the ages and strain of rat examined. Likewise,there were no microscopic neuropathologic alterations in anyof the brains (data not shown) from any treatment group.
DISCUSSION
Based on the pharmacokinetic companion study (Mattssonetal., 2000), fetal exposure from high-dosage dams (5 mg/kg/day) was sufficient to cause about 57% inhibition of fetal brainChE on GD 20. Exposure to high-dosage pups via milk wasestimated at 0.1 mg/kg/day. No inhibition of brain, RBC, or
FIG. 7. Spatial delayed alternation: Delay (mean1 SD). Percent correctchoices as a function of delay in pups (PND 24) and pups as adults (PNDs85–90). There are no overall statistically significant differences across thedifferent dosage groups in the slope and intercept of the percent correctresponse. The 50% correct represents the chance level. Data from males andfemales were combined for purposes of illustration only.
FIG. 8. Motor activity: Number of beam breaks by rats on PNDs 13, 17,21, and 60 (mean1 SD). No overall statistically significant differences in totalcounts as a function of day. Data from males and females were combined forpurposes of illustration only.
259CHLORPYRIFOS DEVELOPMENTAL NEUROTOXICITY
plasma ChE occurred in fetuses or pups from dams gavaged at0.3 or 1 mg/kg/day.
The only statistically significant effect seen in the middledosage group (1 mg/kg/day) was recorded as a 4.2% thinnerparietal cortex (one of the 8 morphometric measurements) inadult females (PND65). No parietal cortex effects were ob-served either in adult males, or in male and female pups(PND11) at this dosage. The middle- and high-dosage groupsdiffered by 0.9% and such a difference was considered smallenough to question the existence of a dose-response relation-ship. The dataa posteriori,corrected for brain weight, showeda clear loss of dose-response relationship and a loss of statis-tical significance in the high-dosage group. In the middle-dosage group, there were no behavioral or functional findings,and no inhibition of blood or brain cholinesterase (Mattssonetal., 2000). From the neurogenesis point of view, the rat neo-cortex, parietal and frontal, develops mainly between GDs 14and 20 (Bayeret al., 1993). If treatment affected corticalneurogenesis, it would be expected that parietal and frontalcortices would be similarly affected, and this was not the casein the present study. Also, as Bayeret al. (1993) concludedfrom their studies, using X-irradiation when an environmentalinsult occurs during neurogenesis, the neuron precursors are
killed and are not replaced. Furthermore, such a loss of neuronsis often followed by abnormal cell migration and differentia-tion. As Bayeret al. (1993) wrote, “Once ectopia has beenproduced by abnormal migration, it becomes a permanentanatomical feature of the brain.” The paper continued, statingthat hypoplasia and dysplasia are also permanent phenomena inthe developing brain. If such events had taken place in theparietal cortex, they would have been present on PND 11. Nogross or microscopic neuropathological alterations were ob-served, however, whether on PNDs 11 or 65, at any dose.Finally, from a statistical point of view, the multiplicity prob-lem should be recalled in light of the numerous statistical testsperformed in this study. As Wareet al. (1992) stated, “In thatview, one of every 20 tests will produce ap value smaller than0.05 merely by chance, and this likelihood must simply be keptin mind in the interpretation of any collection of hypothesistests and theirp values.” Similar positions can be found inTukey (1980) and Wilkinsonet al. (1999). For the reasonsstated above, the statistically significant middle-dose effect inthe parietal cortex of females at PND 65 was isolated, did notsupport a biologically plausible interpretation, and was, there-fore, considered spurious.
FIG. 9. Auditory startle latency (mean1 SD) in ms on PNDs 22 and 61(pups as adults). No overall statistically significant difference across groups.Data from males and females were combined for purposes of illustration only.
FIG. 10. Auditory startle amplitude (mean1 SD) on PNDs 22 and 61(pups as adults). No overall statistically significant difference among groups.However, the high-dosage group amplitude is smaller. Data from males andfemales were combined for purposes of illustration only.
260 MAURISSEN ET AL.

Toxicity was limited to the highest dosage level tested, 5 mgCPF/kg/day, and no effects were seen in the pups in theabsence of maternal toxicity. At this dosage level, ChE activityof brain erythrocyte and plasma were depressed by 90% whenmeasured in the dams on GD20. Clinical signs of ChE inhibi-tion, reduced body weight, and feed consumption were seen inhigh-dosage dams primarily at the end of gestation, and duringthe first few days of lactation. No treatment-related effects(other than ChE inhibition) were seen in the dams at dosages of0.3 or 1 mg CPF/kg/day. At 5 mg/kg/day, pup survival wasreduced during the first 4 postnatal days, when maternal effectswere most evident, while pup survival was unaffected frompostnatal day 5 throughout the remainder of the study. Pupsfrom this group also exhibited lower body weight, delayedappearance of some developmental landmarks, and on PND 11,decreased weight and linear dimensions of the brain. Such aneffect on brain does not,ipso facto,qualify the chemical as adevelopmental neurotoxicant when it is related to generalgrowth retardation, as Makriset al. (1998) recognize. Slightchanges in the auditory startle response were present on PND22 (non-statistically significant), but were not detected on PND61. There was some indication that motor activity measured onPND 13 may have been reduced (although no statisticallysignificant differences were found). It would be consistent withthe documented decrease in motor activity caused by chlor-pyrifos in young rats (Moseret al., 1998). Cognitive functionsmeasured in pups from exposed dams were not affected byCPF. The effects that were noted in the pups at 5 mg/kg/daywere considered a result of the lower body weight gain, wereviewed as indicative of a transient maturational delay, andwere considered secondary to the maternal effects, althoughsome contribution from pup toxicity could not be completelyruled out.
Several lines of evidence support that the effects seen in thepups were secondary to effects on the dam. The fetal effectswere observed only at a dosage level that caused clear maternaltoxicity and occurred in the same time interval. Absolute brainweight was decreased by 8–9% when measured on PND 11,although the brain weight relative to body weight was in-creased, and no effect was seen on brain weight when the pupswere 2 months of age. Thus the effects on pup brain weightwere transient, and are consistent with what is expected due todecreased nutrition or maternal care. The relative sparing ofbrain growth has been described by Dobbing and Sands (1971)in undernourished rats produced by creating artificially large(15 or more pups) and small (3 pups) litters. Undernutritionduring early development causes slower body growth, slowerrate of brain growth to a different degree, and contrary topopular belief, depresses the growth rate of various processeswithin the brain to the same extent (Peeling and Smart, 1994).At weaning, offspring from these large litters showed approx-imately a 16% difference in absolute brain weight. This brainweight difference was also transient, but resolved more slowlyin undernourished litters than in the present study.
Some more arguments about maternal toxicity have beenadvanced by Schardein and Scialli (1999) who stated in areview of the present data, “The developmental findings in thehigh-dose pups in this study are consistent with the decreasedpup weight that occurred in parallel with decreased maternalfeed consumption and lactation weight gain. When maternaltoxicity and developmental toxicity occur at the same dose, itis not axiomatic that the maternal toxicity is the cause of thedevelopmental toxicity; however, in this case, the nature of thedevelopmental effects and the lack of persistent neurologicabnormalities strongly suggest that the pup effects were sec-ondary to maternal toxicity. More directly, no evidence ofspecific toxicity of chlorpyrifos for the developing nervoussystem was identified. The maternal and developmentalNOAELs in this study were 1 mg/kg/day.”
Finally, data from Mattssonet al. (2000) showed that brainChE was inhibited by approximately 85% and 78% in dams onLDs 1 and 5. In contrast, brain ChE was inhibited by 35% inpups on PND 1, but was comparable to control levels on PND5. Neonatal mortality was highest between PNDs 1 and 5, andthus, mortality did not correlate with inhibition of neonatalbrain ChE, which was normal by PND 5. However, neonatalmortality did occur concurrently with clinical signs of maternaltoxicity (muscle fasciculation, hyperpnea, hyperreactivity), andthe time when dams’ body weight and body weight gains weredepressed. All these effects pointed at the dams as responsiblefor neonatal mortality and slower growth rather than at thepups.
The results of the present study are consistent with themajority of scientific data published on chlorpyrifos. Nostrandtet al. (1997) concluded that in adult rats given a single oraldose of CPF, a depression in brain ChE in excess of 60–70%was necessary before any association with behavioral changescould be measured. They observed classical cholinergic signs(miosis and salivation) only in adult rats with.90% inhibitionof brain ChE. In the present study, this degree of brain ChEinhibition was seen only in the dams given 5 mg/kg/day. Nocholinergic signs were observed in dams given the 0.3- or1-mg/kg/day dosage, and maternal brain ChE was inhibited byless than 20% at either dose on GD20. Decreased body weightand reduced survival were also observed previously at a 5-mg/kg/day dosage level in pups from the first generation of a2-generation reproduction study (Breslinet al., 1996), but notin the second generation. Reproductive indices were unaf-fected. In the present study, no effects were observed in thepups at the lower dosage levels (i.e., 0.3 and 1 mg/kg/day).
In the present study, cognitive functions, such as learning(i.e., rate of acquisition of a discrimination), short-term mem-ory (i.e., retention), habituation (a primitive form of learning;Cabe and Eckerman, 1982) of motor activity and auditorystartle amplitude, and, potentially, long-term memory (a pos-teriori observation) were not affected by chlorpyrifos. Theabsence of any cognitive effects in the present study was notunexpected in light of data from a previous cognitive study in
261CHLORPYRIFOS DEVELOPMENTAL NEUROTOXICITY

adult rats administered CPF (Maurissenet al., 2000). Theseauthors demonstrated that in adults there were no differences inretention of previously acquired information at dosages of upto 10 mg/kg/day for 28 days, though brain ChE was inhibitedby up to 85% and cholinergic effects, as well as motor slowing,were prominent in the high-dosage group. Stantonet al. (1994)injected PND-21 rats, sc, with CPF, and no effects wereobserved in rats given 90 mg/kg. Rat pups in the 240-mg/kggroup could not be evaluated because of overt toxicity. At 120mg/kg, brain ChE activity was inhibited by approximately80%, and was accompanied by a small (,10%) transient effecton acquisition of information on PND 23, but by PND 26 therewas no discernible effect on acquisition.
The developmental and cognitive data suggest that the tran-sient changes in brain size have no impact on functionalcapabilities. A similar conclusion was expressed by Dobbing(1971), who noted there was no evidence that a transientdecrease in brain size had any functional significance, eventhough it has been argued that a temporary retardation ofdevelopmental processes could lead to long-term conse-quences.
The results of the present study contrast with the conclusionsfrom Campbellet al. (1997) who administered CPF sc to ratpups during the early neonatal period (PNDs 1–4 and 11–14)and predicted that the neurochemical changes they reported“can be expected to have a significant impact on nervoussystem function and may be responsible for neurobehavioraldisturbances seen with developmental chlorpyrifos exposure”.The present study did not report such findings in the nervoussystem of rat pups and adults and did not, therefore, confirmsuch a prediction. The study design and timing of CPF admin-istration made the Campbell study unsuitable for human riskassessment for several reasons: the study was designed tomaximize the rate of absorption rather than to simulate envi-ronmental exposure (chlorpyrifos sc in DMSO), the route ofadministration was subcutaneous, and pups were injected(PND 1–4) when the nervous system development of the ratbrain was at a stage corresponding to the 3rd trimester inhumans.
Pope and Liu (1997) showed that the increased sensitivity(approximately 6 times) of neonatal rats (relative to adults) tochlorpyrifos is associated with the highest dosage compatiblewith no mortality (i.e., maximum tolerated dose). At dosagelevels that cause no cholinergic effects (e.g., 50% inhibition ofbrain ChE), the neonate was no more than twice as sensitive asthe adult. The authors also concluded that adults may be moresensitive than the pups to sub-lethal alterations following re-peated exposures.
Gavage (i.e., bolus dosage leading to high pulses of expo-sure) is often recommended by governmental agencies fordevelopmental neurotoxicology studies, and was the route ofadministration used in the present study. However, the samedosage given as a fractionated dosage will most likely providelower peak concentrations and a less pronounced toxicity pro-
file compared to a bolus administration (Wagner, 1971).Conolly et al. (1999) cautions against use of gavage adminis-tration of chemicals in these terms: “Experimental conveniencehas also been used to justify unrealistic methods of exposuresuch as corn oil gavage and nasal instillation. Both of thesemethods of exposure have the potential to deliver chemicals toa target site at a rate that far exceeds anything that would occurin the real world. [. . .] The relevance of experiments usingdoses that are multiples of conceivable human exposures andunrealistic routes of exposure is, at most, quite dubious. Mech-anisms of action may be elicited under such conditions thatwould not occur with relevant routes and exposure levels.”This admonition appears relevant to our study, because bothdose and mode of administration may exacerbate toxic effects.Such a caution is applicable to all studies using a bolus modeof administration of the test compound, such as those usinggavage or subcutaneous injection, for example.
In summary, chlorpyrifos did not cause any effects in theoffspring when administered to dams during gestation andlactation at dosage levels of 0.3 or 1 mg/kg/day. Administra-tion of chlorpyrifos at a dosage level of 5 mg/kg/day producedclinical evidence of cholinergic toxicity in the dams. Increasedpup mortality, decrease in pup body weight, brain size, brainlayer thickness, potential transient changes in the startle re-sponse, motor activity, pinna detachment, vaginal opening, andpreputial separation were seen only at 5 mg/kg/day in thepresence of maternal effects, and were consistent with delayedmaturation. Cognitive functions (learning, short-term memory,and habituation in 2 different tasks) were not impaired in thepups at any time at any dosage. The no-observed-effect-level(NOEL) for any effects in pups in this study was 1 mg/kg/day.A NOEL was not determined in dams, due to inhibition ofplasma and RBC ChE at 0.3 mg/kg/day. This study, usingthe testing guidelines established by US EPA, indicates thatchlorpyrifos is not a selective developmental neurotoxicant inthe rat.
ACKNOWLEDGMENTS
The authors wish to thank Joel L. Mattsson for the time spent in discussion,consultation and evaluation of the data; Mark E. Stanton (US EPA) for his helpin setting the spatial delayed alternation method; Mildred S. Christian forcommenting on this manuscript; John F. Barnett, Jr. and Eileen M. Tlush forassistance in managing the conduct of the study; Rosalyn Garman and AliceCampos for the histotechnical preparations; Nancy L. Freshour for the analyt-ical chemistry of the test compound; Stephanie Padilla (US EPA) and Rene´e S.Marshall for the cholinesterase assays; Mamtha R. Shankar for assistance inthe preparation of this manuscript; and Ralph R. Albee for preparing thegraphics.
REFERENCES
Bayer, S. A., Altman, J., Russo, R. J., and Zhang, X. (1993). Timetables ofneurogenesis in the human brain based on experimentally determined pat-terns in the rat.Neurotoxicology14, 83–144.
Breslin, W. J., Liberacki, A. B., Dittenber, D. A., and Quast, J. F. (1996).
262 MAURISSEN ET AL.

Evaluation of the developmental and reproductive toxicity of chlorpyrifos inthe rat.Fundam. Appl. Toxicol. 29, 119–130.
Cabe, P. A., and Eckerman, D. A. (1982). Assessment of learning and memorydysfunction in agent-exposed animals. InNervous System Toxicology(C. L.Mitchell, Ed.), pp. 138–198. Raven Press, New York.
Campbell, C. G., Seidler, F. J., and Slotkin, T. A. (1997). Chlorpyrifosinterferes with cell development in rat brain regions.Brain Res. Bull. 43,179–189.
Code of Federal Regulations (CFR) (1985). Title 9: Animals and AnimalProducts, Subchapter A:Animal Welfare. Office of the Federal Register,Washington, D.C.
Conolly, R. B., Beck, B. D., and Goodman, J. I. (1999). Stimulating researchto improve the scientific basis of risk assessment.Toxicol. Sci. 49, 1–4.
Deacon, M. M., Murray, J. S., Pilny, M. K., Rao, K. S., Dittenber, D. A.,Hanley, T. R., Jr., and John, J. A. (1980). Embryotoxicity and fetotoxicity oforally administered chlorpyrifos in mice.Toxicol. Appl. Pharmacol.54,31–40.
Dobbing, J. (1971). Undernutrition and the developing brain: The use ofanimal models to elucidate the human problem. InAdvances in Experimen-tal Medicine and Biology, Chemistry, and Brain Development(R. Paolettiand A. Davison, Eds.), Vol. 13, pp. 399–412. Plenum, New York.
Dobbing, J., and Sands, J. (1971). Vulnerability of developing brain: IX. Theeffect of nutritional growth retardation on the timing of the brain growth-spurt.Biol. Neonate19, 363–378.
Johnson, C. D., and Russell, R. L. (1975). A rapid, simple radiometric assay forcholinesterase, suitable for multiple determinations.Anal. Biochem. 64,229–238.
Makris, S., Raffaele, K., Sette, W., and Seed, J. (1998). A retrospectiveanalysis of twelve developmental neurotoxicity studies submitted to the USEPA Office of Prevention, Pesticides, and Toxic Substances (p. 46)(OPPTS). US EPA Internal Document.
Mattsson, J. L., Maurissen, J. P., Nolan, R. J., and Brzak, K. A. (2000). Lackof differential sensitivity to cholinesterase inhibition in fetuses and neonatescompared to dams treated perinatally with chlorpyrifos.Toxicol. Sci.53,438–446.
Maurissen, J. P., Shankar, M. R., and Mattsson, J. L. (2000). Chlorpyrifos:Lack of cognitive effects in adult Long-Evans rats.Neurotoxicol. Teratol.22, 237–246.
Moser, V. C., and Padilla, S. (1998). Age- and gender-related differences in thetime course of behavioral and biochemical effects produced by oral chlor-pyrifos in rats.Toxicol. Appl. Pharmacol.149,107–119.
Nicholas, K. R., and Hartman, P. E. (1991). Milk secretion in the rat: Progres-sive changes in milk composition during lactation and weaning and theeffect of diet.Comp. Biochem. Physiol. 98A, 535–542.
Nostrandt, A. C., Padilla, S., and Moser, V. C. (1997). The relationship of oralchlorpyrifos effects on behavior, cholinesterase inhibition, and muscarinicreceptor density in the rat.Pharmacol. Biochem. Behav.58, 15–23.
National Research Council (NRC) (1996).Guide for the Care and Use ofLaboratory Animals. National Academy Press, Washington, DC.
Peeling, A. N., and Smart, J. L. (1994). Review of literature showing thatundernutrition affects the growth rate of all processes in the brain to thesame extent. Metabolic Brain Disease9, 33–42.
Pope, C. N., and Liu, J. (1997). Age-related differences in sensitivity toorganophosphorus pesticides.Env. Toxicol. Pharmacol. 4, 309–314.
Salewski, E. (1964). Fa¨rbemethode zum makroskopischen Nachweis von Im-plantationsstellen am Uterus der Ratte.Arch. Pathol. Exp. Pharmakol.247,367.
SAS Institute. (1989).SAS/STAT User’s Guide, Version 6, 4th ed., Volumes 1and 2. SAS Institute, Cary, NC.
Schardein, J. L., and Scialli, A. R. (1999). The legislation of toxicologic safetyfactors: The Food Quality Protection Act with chlorpyrifos as a test case.Reprod. Toxicol. 13, 1–14.
Sheets, L. P., Dean, K. F., and Reiter, L. W. (1988). Ontogeny of the acousticstartle response and sensitization to background noise in the rat.Behav.Neurosci. 102,706–713.
Snedecor, G. W., and Cochran, W. G. (1967).Statistical Methods, 6th Ed, pp.240–241. Iowa State University Press, Ames, IA.
Sokal, R. R., and Rohlf, F. J. (1969).Biometry, pp. 388-389. W. H. Freeman,San Francisco, CA.
Stanton, M. E., Mundy, W. R., Ward, T., Dulchinos, V., and Barry, C. C.(1994). Time-dependent effects of acute chlorpyrifos administration onspatial delayed alternation and cholinergic neurochemistry in weanling rats.Neurotoxicology15, 201–208.
Tukey, J. W. (1980). We need both exploratory and confirmatory.Am. Stat-istician 34, 23–25.
Tukey, J. W., Ciminera, J. L., and Heyse, J. F. (1985). Testing the statisticalcertainty of a response to increasing doses of a drug.Biometrics 41,295–301.
U.S. Environmental Protection Agency (US EPA) (1989). Federal Insecticide,Fungicide, and Rodenticide Act (FIFRA); Good Laboratory Practice Stan-dards; Final Rule. 40 CFR, Part 160.
U.S. Environmental Protection Agency (US EPA) (1991). Pesticide Assess-ment Guidelines. Subdivision F, Hazard Evaluation: Human and DomesticAnimals, Addendum 10, Neurotoxicity. Health Effects Division, Office ofPesticide Program.
Wagner, J. G. (1971).Biopharmaceutics and Relevant Pharmacokinetics, pp.149–150. Drug Intelligence Publications, Hamilton, IL.
Ware, J. H., Mosteller, F., Delgado, F., Donnelly, C., and Ingelfinger, J. A.(1992).P values. InMedical Uses of Statistics(J. C. Bailar and F. Mosteller,Eds.), pp. 181–200. NEJM Books, Boston.
Wilkinson, L., and the Task Force on Statistical Inference. (1999) Statisticalmethods in psychology journals.Am. Psychol.54, 594–604.
263CHLORPYRIFOS DEVELOPMENTAL NEUROTOXICITY
Related Documents