1 23 Molecular Biology Reports An International Journal on Molecular and Cellular Biology ISSN 0301-4851 Mol Biol Rep DOI 10.1007/s11033-013-2693-1 Lack of association of Lysyl oxidase (LOX) gene polymorphisms with intracranial aneurysm in a south Indian population Sanish Sathyan, Linda Koshy, K. R. Sarada Lekshmi, H. V. Easwer, S. Premkumar, Jacob P. Alapatt, Suresh Nair, R. N. Bhattacharya, et al.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1 23
Molecular Biology ReportsAn International Journal on Molecularand Cellular Biology ISSN 0301-4851 Mol Biol RepDOI 10.1007/s11033-013-2693-1
Lack of association of Lysyl oxidase (LOX)gene polymorphisms with intracranialaneurysm in a south Indian population
Sanish Sathyan, Linda Koshy,K. R. Sarada Lekshmi, H. V. Easwer,S. Premkumar, Jacob P. Alapatt, SureshNair, R. N. Bhattacharya, et al.

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

Lack of association of Lysyl oxidase (LOX) gene polymorphismswith intracranial aneurysm in a south Indian population
Sanish Sathyan • Linda Koshy • K. R. Sarada Lekshmi •
H. V. Easwer • S. Premkumar • Jacob P. Alapatt •
Suresh Nair • R. N. Bhattacharya • Moinak Banerjee
Received: 13 December 2012 / Accepted: 14 September 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Intracranial aneurysm (IA) accounts for 85 %
of haemorrhagic stroke and is mainly caused due to
weakening of arterial wall. Lysyl oxidase (LOX) is a
cuproenzyme involved in cross linking structural proteins
collagen and elastin, thus providing structural stability to
artery. Using a case–control study design, we tested the
hypothesis whether the variants in LOX gene flanking the
two LD block, can increase risk of aSAH among South
Indian patients, either independently, or by interacting with
other risk factors of the disease. SNPs were genotyped by
fluorescence-based competitive allele-specific PCR (KAS-
Par) chemistry. We selected 200 radiologically confirmed
aneurysmal cases and 235 ethnically and age and gender
matched controls from the Dravidian Malayalam speaking
population of South India. We observed marked interethnic
differences in the genotype distribution of LOX variants
when compared to Japanese and African populations.
However, there was no significant association with any of
the LOX variants with IA. This study also could not
observe any significant role of LOX polymorphisms in
influencing IA either directly or indirectly through its
confounding factors such as hypertension and gender in
South Indian population.
Keywords Intracranial aneurysm � Subarachnoid
haemorrhage � Lysyl oxidase (LOX) � SNP � India
Introduction
Intracranial aneurysm (IA) accounts for 85 % of sub-
arachnoid hemorrhage (SAH), which contributes for
5–15 % of strokes, but occurs at a fairly young age [1].
Reports have shown worldwide incidence of aSAH
(aneurysmal subarachnoid hemorrhage) in 10.5 cases per
100,000 person-years [2]. In general population 2–3 %
individual harbor IA [3]. Prevalence of IA in India based
on autopsies ranges from 0.2 to 10.3 % from various parts
of India, with a mean prevalence of 5.3 % [4–6].
Involvement of genetic component with IA is well
documented. Familial clustering of IA shows four fold
higher risks of aneurysmal rupture in first degree relative
and identical aneurysm location in twins study supports
this cause [7, 8]. Various theories have been proposed for
the development of aneurysm, such as Congenital medial
defects theory, Acquired internal elastic lamina defects
theory [9, 10] and Hemodynamic Stress theory [11], but the
etiology of IA still remains controversial. Most of these
theories reflect on the imbalances in the Extracellular
matrix (ECM) remodeling. ECM remodeling plays an
important role in maintaining the structure and integrity of
the intracranial arteries. Elastin and collagen fibers form
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11033-013-2693-1) contains supplementarymaterial, which is available to authorized users.
S. Sathyan � L. Koshy � K. R. Sarada Lekshmi �M. Banerjee (&)
Human Molecular Genetics Laboratory, Rajiv Gandhi Centre for
Biotechnology, Thiruvananthapuram 695 014, Kerala, India
e-mail: [email protected]
H. V. Easwer
Department of Neurosurgery, Sree Chitra Tirunal Institute for
Medical Science and Technology, Thiruvananthapuram, India
S. Premkumar � J. P. Alapatt � S. Nair � R. N. Bhattacharya
Department of Neurosurgery, Calicut Medical College,
Calicut, India
123
Mol Biol Rep
DOI 10.1007/s11033-013-2693-1
Author's personal copy
the framework for ECM providing strength and elasticity to
the artery. Lysyl oxidase (LOX) is an important class of
copper containing enzyme involved in covalent cross
linking of these fibrous proteins by formation of aldehydes
between lysine residues that insolubilize these extracellular
proteins namely collagen and elastin [12], thus giving
strength to intracranial arteries. Animal models for IA are
mainly based on the LOX inhibitor such as b-aminopro-
pionitrile(BAPN) [13, 14]. These observations strengthens
the role of LOX gene in pathophysiology of IA.
Interestingly, one of the earliest studies on genome wide
linkage analysis identified the loci 5q22-31 comprising of
the LOX gene to be associated with IA [15]. Reduced
extracellular matrix is a prominent feature of cerebral
aneurysms. Therefore, inspite of a strong hypothesis and
genetic linkage to the LOX this gene has not been inves-
tigated for its possible role in IA. The gene has 5 transcripts
and seven exons. The processed product of proteolytic
cleavage of Pro-LOX consist of a functional 32-kDa LOX
enzyme and an 18-kDa LOX propeptide (LOX-PP) [16]. A
non-synonymous SNP rs1800449 (G473A, Arg158Gln)
located in first exon of LOX causing an arginine to gluta-
mine change in the conserved region of LOX propeptide
has been implicated with coronary artery disease (CAD)
[17]. Based on these backgrounds the present study intends
to explore the involvement of the variants in the LOX with
IA in a genetically homogenous Indian population.
Materials and methods
Study population
The study populations consist of radiologically confirmed
200 aneurysmal cases and 235 ethnically age and gender
matched controls from the Dravidian Malayalam speaking
population of South India, as described in our earlier
publication [18]. All patients had angiographically docu-
mented saccular aneurysms with SAH confirmed by cranial
computed tomography (CT), magnetic resonance angiog-
raphy (MRA) and digital subtraction angiography (DSA).
The neurological grade was classified as per World Fed-
eration of Neurological Surgeons (WFNS) scale and all
grades of aSAH were eligible for inclusion. Exclusion
criteria included nonsaccular aneurysm, arteriovenous
malformations or hematological disorders. The control
group consisted of unrelated ethnically matched volunteers
recruited from individuals admitted to the hospitals for any
reason other than neurological diseases. Data were col-
lected by the investigators directly by interviewing the
patients and controls using a structured questionnaire to
obtain information regarding demographic characteristics.
The study was as per the ICMR guidelines and approved by
Institute ethical committee (IEC).
SNP selection and genotyping
SNP’s were selected based on the functional relevance,
tagging status and minor allele frequency in different
populations. Fours SNPs where selected includes a func-
tionally significant variant present in LOX propeptide,
rs1800449, G473A, Arg158Gln and three other SNPs
rs2956540, rs10519694 and rs3792801 which were selec-
ted based on tagging status using genotype data obtained
from Caucasian individuals in the HapMap project (Hap-
Map Data Rel 24/Phase II Nov08, on NCBI B36 assembly,
dbSNP b126) on the basis of pairwise linkage disequilib-
rium with a r2 threshold of 0.8 and minor allele fre-
quency C0.05 to capture all the common SNPs. SNPs
rs2956540, rs10519694 and rs1800449 were present in the
single LD block, while rs379801 and rs379802 formed the
second minor block in caucasian population (Supplemen-
tary Figure 1). Genotyping was performed by fluorescence-
based competitive allele-specific PCR (KASPar) chemistry
(Kbiosciences, UK).
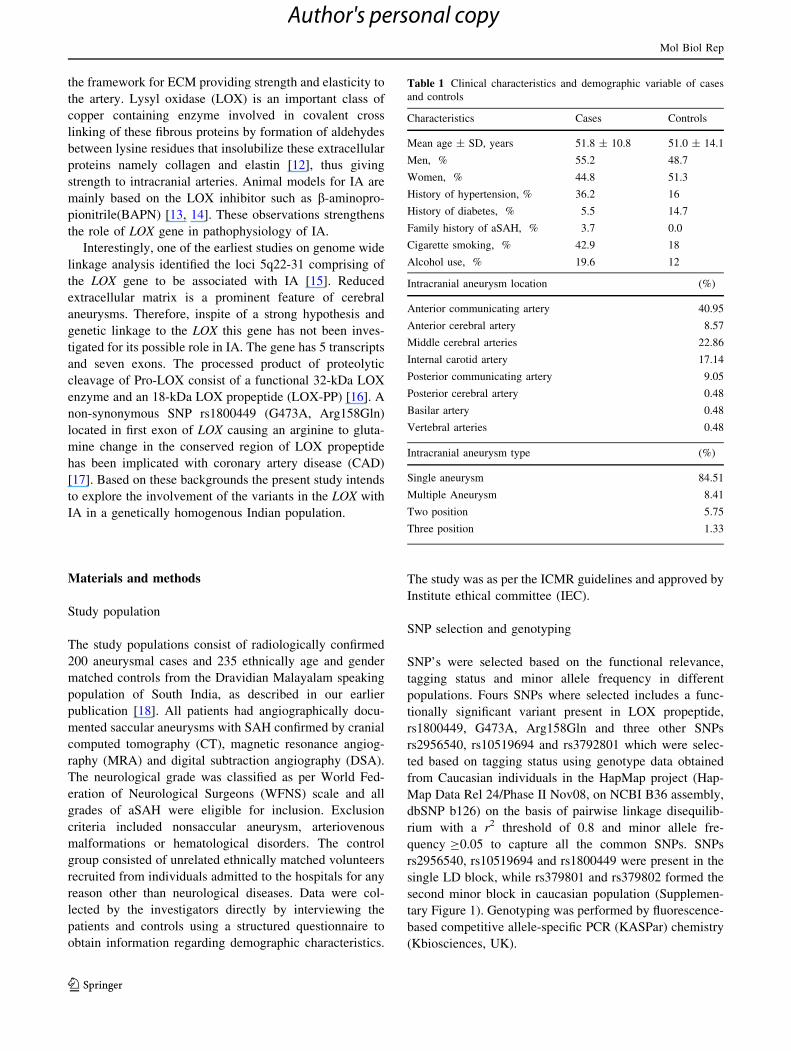
Table 1 Clinical characteristics and demographic variable of cases
and controls
Characteristics Cases Controls
Mean age ± SD, years 51.8 ± 10.8 51.0 ± 14.1
Men, % 55.2 48.7
Women, % 44.8 51.3
History of hypertension, % 36.2 16
History of diabetes, % 5.5 14.7
Family history of aSAH, % 3.7 0.0
Cigarette smoking, % 42.9 18
Alcohol use, % 19.6 12
Intracranial aneurysm location (%)
Anterior communicating artery 40.95
Anterior cerebral artery 8.57
Middle cerebral arteries 22.86
Internal carotid artery 17.14
Posterior communicating artery 9.05
Posterior cerebral artery 0.48
Basilar artery 0.48
Vertebral arteries 0.48
Intracranial aneurysm type (%)
Single aneurysm 84.51
Multiple Aneurysm 8.41
Two position 5.75
Three position 1.33
Mol Biol Rep
123
Author's personal copy
Statistical analysis
Genotype and allelic frequencies were computed and were
checked for deviation from Hardy–Weinberg equilibrium
(http://ihg2.helmholtz-muenchen.de/cgi-bin/hw/hwa1.pl).
Haplotypic association of samples was carried out using
the COCAPHASE program in the UNPHASED v3.1
package (http://www.mrc-bsu.cam.ac.uk/personal/frank/
software/unphased/) [19] with 10,000 permutations. To
estimate LD between pairs of loci in the patient and control
populations, standardized disequilibrium coefficient (D0)and squared correlation coefficient (r2) were calculated
using Haploview 4.1 (www.broad.mit.edu/mpg/haploview/)
[20]. LD blocks were defined in accordance with Gabriel’s
criteria [21]. Further stratification of the patients was done
to understand the role of LOX variant within the genders
and between the hypertensive and non-hypertensive
groups. However due to the exploratory nature of this
study, no adjustment for multiple testing was made, as not
to penalize our data by missing possibility of important
findings.
Results
The demographic and clinical characteristics of the patients
and controls are summarized in Table 1. All the four SNPs
selected for the study was in Hardy–Weinberg equilibrium
(p [ 0.05). Functionally significant non synonymous SNP
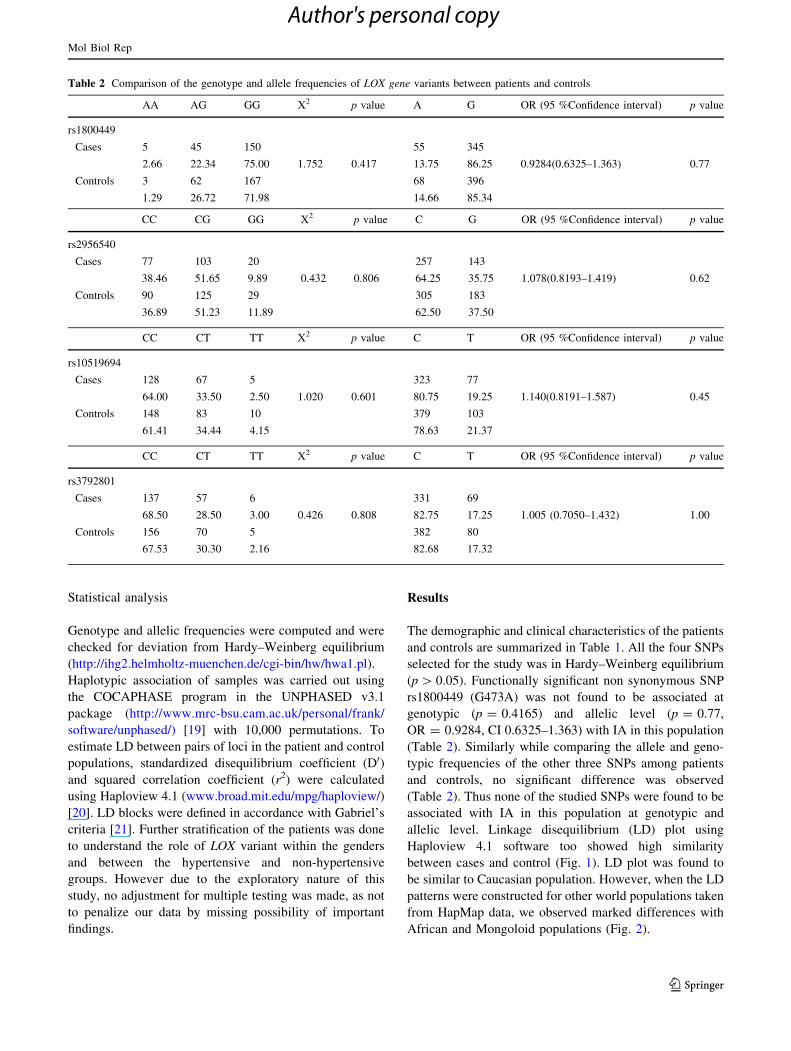
rs1800449 (G473A) was not found to be associated at
genotypic (p = 0.4165) and allelic level (p = 0.77,
OR = 0.9284, CI 0.6325–1.363) with IA in this population
(Table 2). Similarly while comparing the allele and geno-
typic frequencies of the other three SNPs among patients
and controls, no significant difference was observed
(Table 2). Thus none of the studied SNPs were found to be
associated with IA in this population at genotypic and
allelic level. Linkage disequilibrium (LD) plot using
Haploview 4.1 software too showed high similarity
between cases and control (Fig. 1). LD plot was found to
be similar to Caucasian population. However, when the LD
patterns were constructed for other world populations taken
from HapMap data, we observed marked differences with
African and Mongoloid populations (Fig. 2).
Table 2 Comparison of the genotype and allele frequencies of LOX gene variants between patients and controls
AA AG GG X2 p value A G OR (95 %Confidence interval) p value
rs1800449
Cases 5 45 150 55 345
2.66 22.34 75.00 1.752 0.417 13.75 86.25 0.9284(0.6325–1.363) 0.77
Controls 3 62 167 68 396
1.29 26.72 71.98 14.66 85.34
CC CG GG X2 p value C G OR (95 %Confidence interval) p value
rs2956540
Cases 77 103 20 257 143
38.46 51.65 9.89 0.432 0.806 64.25 35.75 1.078(0.8193–1.419) 0.62
Controls 90 125 29 305 183
36.89 51.23 11.89 62.50 37.50
CC CT TT X2 p value C T OR (95 %Confidence interval) p value
rs10519694
Cases 128 67 5 323 77
64.00 33.50 2.50 1.020 0.601 80.75 19.25 1.140(0.8191–1.587) 0.45
Controls 148 83 10 379 103
61.41 34.44 4.15 78.63 21.37
CC CT TT X2 p value C T OR (95 %Confidence interval) p value
rs3792801
Cases 137 57 6 331 69
68.50 28.50 3.00 0.426 0.808 82.75 17.25 1.005 (0.7050–1.432) 1.00
Controls 156 70 5 382 80
67.53 30.30 2.16 82.68 17.32
Mol Biol Rep
123
Author's personal copy

We further investigated the role of these allelic and
genotypic variants in gender stratification and hypertension
to understand whether the genetics of these confounding
factors act as modifiable risk factor for IA. None of the
alleles or genotypes of LOX were found to influence gender
(Supplementary Table-1).and hypertension in causing IA
either directly or through modifiable risk factor (Supple-
mentary Table-2).
Discussion
In the present study the four SNPs rs1800449 (473 G/A),
rs2956540, rs10519694 and rs3792801 within the LOX
were selected based on tagging status comprising of two
LD blocks covering the complete gene based on the
genotype data of Caucasians from phase II HapMap Pro-
ject. LD pattern observed in our control population was
similar to Caucasians but distinctly different from Chinese,
Japanese and Africans. LOX is known to function as the
initiator of crosslinking of elastin and collagen and in the
repair of these ECM molecules during aging and disease.
Thus inactivation of the LOX would have debilitating
consequences. Differences in the LD patterns would sug-
gest that LOX variants might have a strong ethno-genetic
influence in modulating a debilitating consequence of
imbalances in ECM remodeling.
Though functionally and positionally based on linkage
studies, LOX has been implicated in development of IAs in
Japanese [15] but this could not be verified in a validation
study with LOX variants in the same Japanese population
comprising of 172 IA patients and 192 controls [22]. In the
present study too we report lack of association of LOX
variants with IA inspite of distinctly different LD patterns
between our study population and Japanese population.
Lack of association observed in the present study could be
explained with following reasons at positional and func-
tional level. Firstly even though LOX gene loci 5q22-31
was found to be positional candidate gene for IA in linkage
studies [15] this loci was never found to be associated in
Genome wide association studies carried out with higher
sample size [23–26].This conflicting result may be due to
genetic heterogeneity, power of study, difference in sample
size and difference in statistical method employed. Vali-
dating such an observation using a candidate gene
approach in a genetically well stratified population do
confirm that this gene may not play a significant role by
itself.
Secondly, at functional level the gene possesses an
important role in maintaining the structural integrity and
stability of vessel wall. Animal models were also generated
by LOX inhibitor BAPN [13]. The LOX expression was
found to be down regulated in IA arteries [14]. LOX having
a role in endothelial dysfunction [27], a major cause in
pathogenesis of IA, is shown to regulated by number of
factors including homocysteine level [28], low density
lipoprotein (LDLs) [29], TGF-b1 [30] and TNF alpha [31].
The results obtained in the present study suggest that LOX
may not to be a causal variant at genetic level. LOX
expression is known to be regulated by various extraneous
control cases
Fig. 1 Linkage disequilibrium pattern of studied Lysyl oxidase(LOX)
SNPs in cases and controls
Fig. 2 Linkage disequilibrium patterns in different HapMap populations of the world. (S1 South India, CEU European, JPT Japanese, CHB Han
Chinese, YRI Yoruba African)
Mol Biol Rep
123
Author's personal copy

factors. Therefore, one cannot not downplay the impor-
tance of LOX in IA, without understanding the role of
confounding factors involved in regulation of LOX.
Increased LOX activity has been reported in hyperten-
sion which can result in alterations in the degree of cross-
linking of vascular collagens [32]. To address the role of
these confounding factors we further investigated the
association of LOX variants in hypertension and gender as
modifiable factors in influencing IA. Here too we could not
observe any significant role of LOX polymorphisms in
influencing IA either directly or indirectly through its
confounding factors. LOX variant rs1800449 (473AA) has
been reported to be associated with cardiovascular disease
including ischemic stroke [17]. Differences in cerebro-
vascular and cardiovascular stroke could be due to differ-
ence in structural architecture of vessel walls or could be
due to differences in the pathobiology of two strokes.
We conclude that allelic or genotypic variants of LOX
do not play a significant role in South Indian population
either directly or through modifiable factors to result in
IAs. However, differences in allelic and genotypic distri-
bution of LOX were observed in different ethnic popula-
tions of the world.
Acknowledgments SS acknowledges the Council of Scientific and
Industrial Research CSIR), Government of India for providing junior
research fellowship. We also acknowledge all the patients and their
family members who cooperated in this study.
References
1. Bederson JB, Awad IA, Wiebers DO, Piepgras D, Haley EC Jr
et al (2000) Recommendations for the management of patients
with unruptured intracranial aneurysms: a statement for health-
care professionals from the Stroke Council of the American Heart
Association. Circulation 102:2300–2308
2. Ingall T, Asplund K, Mahonen M, Bonita R (2000) A multi-
national comparison of subarachnoid hemorrhage epidemiology
in the WHO MONICA stroke study. Stroke 31:1054–1061
3. Rinkel GJE, Djibuti M, Algra A, Van Gijn J (1998) Prevalence
and risk of rupture of intracranial aneurysms a systematic review.
Stroke 29:251–256
4. Ramamurthi B (1969) Incidence of intracranial aneurysms in
India. J Neurosurg 30:154
5. Banerjee A (2000) Pathology of cerebrovascular disease. Neurol
India 48:305
6. Kapoor K, Kak V (2003) Incidence of intracranial aneurysms in
northwest Indian population. Neurol India 51:22
7. Woo D, Broderick J. Genetics of intracranial aneurysm; 2002.
Elsevier. pp 24–34
8. Krischek B, Inoue I (2006) The genetics of intracranial aneu-
rysms. J Hum Genet 51:587–594
9. Stehbens WE (1989) Etiology of intracranial berry aneurysms.
J Neurosurg 70:823–831
10. Glynn L (1940) Medial defects in the circle of Willis and their
relation to aneurysm formation. J Pathol Bacteriol 51:213–222
11. Cebral JR, Mut F, Weir J, Putman CM (2011) Association of
hemodynamic characteristics and cerebral aneurysm rupture. Am
J Neuroradiol 32:264–270
12. Kagan HM, Li W (2002) Lysyl oxidase: properties, specificity,
and biological roles inside and outside of the cell. J Cell Biochem
88:660–672
13. Hazama F, Hashimoto N (1987) Annotation: an animal model of
cerebral aneurysms. Neuropathol Appl Neurobiol 13:77–90
14. Aoki T, Kataoka H, Ishibashi R, Nozaki K, Morishita R et al
(2009) Reduced collagen biosynthesis is the hallmark of cerebral
aneurysm contribution of interleukin-1b and nuclear factor-jB.
Arterioscler Thromb Vasc Biol 29:1080–1086
15. Onda H, Kasuya H, Yoneyama T, Takakura K, Hori T et al
(2001) Genomewide-linkage and haplotype-association studies
map intracranial aneurysm to chromosome 7q11. Am J Hum
Genet 69:804–819
16. Trackman PC, Bedell-Hogan D, Tang J, Kagan H (1992) Post-
translational glycosylation and proteolytic processing of a lysyl
oxidase precursor. J Biol Chem 267:8666–8671
17. Ma L, Song H, Zhang M, Zhang D (2011) Lysyl oxidase G473A
polymorphism is associated with increased risk of coronary artery
diseases. DNA Cell Biol 30:1033–1037
18. Koshy L, Easwer HV, Premkumar S, Alapatt JP, Marthanda Pillai
A, Nair S, Bhattacharya RN, Banerjee M (2010) Risk factors
attributable to aneurysmal subarachnoid hemorrhage in Indian
population. Cerebrovasc Dis 29:268–274
19. Dudbridge F (2008) Likelihood-based association analysis for
nuclear families and unrelated subjects with missing genotype
data. Hum Hered 66:87–98
20. Barrett JC, Fry B, Maller J, Daly MJ (2005) Haploview: analysis
and visualization of LD and haplotype maps. Bioinformatics
21:263–265
21. Gabriel SB, Schaffner SF, Nguyen H, Moore JM, Roy J, Blu-
menstiel B et al (2002) The structure of haplotype blocks in the
human genome. Science 296:2225–2229
22. Yoneyama T, Kasuya H, Onda H, Akagawa H, Jinnai N, Nak-
ajima T, Hori T, Inoue I (2003) Association of positional and
functional candidate genes FGF1, FBN2, and LOX on 5q31 with
intracranial aneurysm. J Hum Genet 48:309–314
23. Bilguvar K, Yasuno K, Niemela M, Ruigrok YM, von und zu
Fraunberg M et al (2008) Susceptibility loci for intracranial
aneurysm in European and Japanese populations. Nat Genet
40:1472–1477
24. Yasuno K, Bilguvar K, Bijlenga P, Low SK, Krischek B et al
(2010) Genome-wide association study of intracranial aneurysm
identifies three new risk loci. Nat Genet 42:420–425
25. Akiyama K, Narita A, Nakaoka H, Cui T, Takahashi T et al
(2010) Genome-wide association study to identify genetic vari-
ants present in Japanese patients harboring intracranial aneu-
rysms. J Hum Genet 55:656–661
26. Low SK, Takahashi A, Cha PC, Zembutsu H, Kamatani N et al
(2012) Genome-wide association study for intracranial aneurysm
in the Japanese population identifies three candidate susceptible
loci and a functional genetic variant at EDNRA. Hum Mol Genet
21:2102–2110
27. Rodrıguez C, Martınez-Gonzalez J, Raposo B, Alcudia JF,
Guadall A et al (2008) Regulation of lysyl oxidase in vascular
cells: lysyl oxidase as a new player in cardiovascular diseases.
Cardiovasc Res 79:7–13
28. Raposo B, Rodrıguez C, Martınez-Gonzalez J, Badimon L (2004)
High levels of homocysteine inhibit lysyl oxidase (LOX) and
downregulate LOX expression in vascular endothelial cells.
Atherosclerosis 177:1–8
29. Rodrıguez C, Raposo B, Martınez-Gonzalez J, Casanı L, Badi-
mon L (2002) Low density lipoproteins downregulate lysyl
Mol Biol Rep
123
Author's personal copy

oxidase in vascular endothelial cells and the arterial wall. Arte-
rioscler Thromb Vasc Biol 22:1409–1414
30. Boak AM, Roy R, Berk J, Taylor L, Polgar P et al (1994) Reg-
ulation of lysyl oxidase expression in lung fibroblasts by trans-
forming growth factor-beta 1 and prostaglandin E2. Am J Respir
Cell Mol Biol 11:751
31. Rodrıguez C, Alcudia J, Martınez-Gonzalez J, Raposo B, Navarro
M et al (2008) Lysyl oxidase (LOX) down-regulation by TNFa: a
new mechanism underlying TNFa-induced endothelial dysfunc-
tion. Atherosclerosis 196:558–564
32. Ooshima A, Midorikawa O (1977) Increased lysyl oxidase
activity in blood vessels of hypertensive rats and effect of beta-
aminopropionitrile on arteriosclerosis. Jpn Circ J 41:1337
Mol Biol Rep
123
Author's personal copy
Related Documents











