UNIVERSIDAD DE OVIEDO Programa de Doctorado en Ciencias de la Salud La Matriz Extracelular en Mecanorreceptores de Vertebrados Jorge García Piqueras Tesis doctoral, 2019

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

UNIVERSIDAD DE OVIEDO
Programa de Doctorado en Ciencias de la Salud
La Matriz Extracelular en
Mecanorreceptores de Vertebrados
Jorge García Piqueras
Tesis doctoral, 2019


UNIVERSIDAD DE OVIEDO
Programa de Doctorado en Ciencias de la Salud
La Matriz Extracelular en
Mecanorreceptores de Vertebrados
Jorge García Piqueras
Tesis doctoral, 2019


FO
R-M
AT
-VO
A-0
10 (
Reg
.2018)
RESUMEN DEL CONTENIDO DE TESIS DOCTORAL
1.- Título de la Tesis
Español: LA MATRIZ EXTRACELULAR EN MECANORRECEPTORES DE VERTEBRADOS
Inglés: THE EXTRACELLULAR MATRIX IN VERTEBRATE MECHANORECEPTORS
2.- Autor
Nombre: JORGE GARCÍA PIQUERAS
DNI/Pasaporte/NIE:
Programa de Doctorado: CIENCIAS DE LA SALUD
Órgano responsable: CENTRO INTERNACIONAL DE POSTGRADO
RESUMEN (en español)
Los mecanorreceptores son estructuras nerviosas especializadas que se encargan de percibir los estímulos táctiles. Se encuentran distribuidos por todos los tejidos de los vertebrados, en el inicio de las vías aferentes sensitivas. En la piel, los corpúsculos de Meissner y Pacini son mecanorreceptores de adaptación rápida y bajo umbral encargados del tacto ligero y tacto profundo respectivamente. Ambos están formados por un axón rodeado por células de Schwann modificadas, y en el caso de los corpúsculos de Pacini, también por tejido conectivo organizado formando una cápsula. Los complejos célula de Merkel-axón son mecanorreceptores de adaptación lenta y bajo umbral responsables de diferentes sensaciones del tacto fino; están formados por una asociación sinapsis-like entre un axón y una célula de Merkel epitelial que se localiza en la base de la epidermis. Dos de los aspectos menos conocidos sobre los corpúsculos sensitivos cutáneos son, por un lado, la matriz extracelular y, por otro, los cambios en estas estructuras durante el proceso de envejecimiento. La primera parte de la tesis se compone de cuatro publicaciones distintas relacionadas con la matriz extracelular. En la primera se ha investigado el origen endoneural o perineural de los distintos compartimentos de los corpúsculos de Pacini humanos; el principal resultado ha sido la identificación de una lámina intermedia CD34-positiva alrededor del núcleo interno, de origen endoneural. En la segunda se ha analizado la expresión del glicosaminoglicano condroitín sulfato en los corpúsculos sensitivos: mientras que los corpúsculos de Meissner no expresan proteoglicanos de condroitin sulfato, en los corpúsculos de Pacini se expresan asociados a la lámina intermedia endoneural CD34-positiva. En el tercer trabajo se ha estudiado el patrón de expresión de otro glicosaminoglicano, el heparán sulfato: tanto en los corpúsculos de Meissner como en los de Pacini, los proteoglicanos de heparan sulfato están asociados a la membrana basal, la cual es especialmente prominente en las primeras lamelas del núcleo externo de los corpúsculos de Pacini. En el cuarto estudio se ha determinado la expresión de los Pequeños Proteoglicanos Ricos en Leucina (SLRPs) de clase I y clase II, ambas de localización extracelular, en los corpúsculos de Pacini: los proteoglicanos biglicano, decorina, lumicano, osteomodulina y fibromodulina se expresan en el núcleo interno y en el núcleo externo; todos ellos excepto la osteomodulina también están presentes en la cápsula externa; contrariamente, la osteomodulina sí se expresa en la lámina intermedia, donde los demás están ausentes. La segunda parte de la tesis tiene un capítulo metodológico, para determinar datos cuantitativos y cualitativos sobre los mecanorreceptores cutáneos, que se ha utilizado en el estudio del envejecimiento y establece las bases para su utilización para la validación de la biopsia cutánea en el diagnóstico de neuropatías periféricas. En el quinto y último trabajo se ha realizado un análisis de los cambios cualitativos y cuantitativos que aparecen en los corpúsculos de Pacini y de Meissner y en los complejos célula de Merkel-axón como consecuencia del envejecimiento. Además, se ha evaluado la expresión de la mecanoproteína Piezo2 (principal canal iónico implicado en la función mecanorreptora) y la expresión del sistema neurotrófico TrkB-BDNF (responsable del desarrollo y mantenimiento de los mecanorreceptores cutáneos) en sujetos de edad avanzada.
Carmen
Rectángulo

RESUMEN (en Inglés)
Mechanoreceptors are specialized nervous structures involved in sensing tactile stimuli. They are localized throughout all the vertebrate tissues at the beginning of the sensory afferent nerve fibres. In the skin, Meissner’s and Pacinian corpuscles are rapidly adapting low-threshold mechanoreceptors, responsible for fine and deep touch respectively. Both of them consist of an axon surrounded by modified Schwann cells, and in the case of Pacinian corpuscles, also by organized connective tissue forming a capsule. Merkel’s cells-axon complexes are slowly adapting low-threshold mechanoreceptors responsible for several light-touch sensations; they consist of a synapsis-like association between an axon and an epithelial Merkel’s cell, which is localized in the basal epidermis. Two of the least known topics about cutaneous sensory corpuscles are, on the one hand, the extracellular matrix, and on the other hand, the changes on these structures during the aging process. The first part of the thesis is compound by four different publications related to the extracellular matrix. In the first one, it has been investigated the endoneural or perineural origin of the different human Pacinian corpuscle compartments; the main result has been the identification of an endoneurial intermediate CD34-positive layer around the inner core. In the second one, it has been analysed the expression of chondroitin sulfate glycosaminoglycan: whereas Meissner’s corpuscles do not express chondroitin sulfate proteoglycans, in Pacinian corpuscles they are expressed in association to the endoneural CD34-positive intermediate layer. In the third work it has been studied the expression pattern of another glycosaminoglycan, the heparan sulfate: in both Meissner and Pacinian corpuscles, the heparan sulfate proteoglycans are associated to the basement membrane, which is specially prominent in the first lamellae of the outer core of the Pacinian corpuscles. In the fourth study, it has been established the expression of extracellular classes I and II Small Leucine-Rich Proteoglycans (SLRPs) in Pacinian corpuscles: biglycan, decorin, lumican, osteomodulina and fibromodulin are expressed in both the inner and outer core; all of them except osteomodulina are present in the external capsule; contrary, osteomodulina is expressed in the intermediate layer, where the others are absent. The second part of the thesis includes a methodological chapter, in order to define quantitative and qualitative data about cutaneous mechanoreceptors, which has been used in the study of aging and it sets the basis for its utilization for the validation of cutaneous biopsy in the diagnosis of peripheral neuropathies. In the fifth and last work, it has been analysed the qualitative and quantitative changes occurring in Pacinian and Meissner’s corpuscles and Merkel’s cells-axon complexes with the aging process. Furthermore, it has been evaluated the expression of Piezo2 mechanoprotein (main ionic canal involved in the mechanoreceptive function) and the expression of TrkB-BDNF neurotrophic system (responsible for the development and maintenance of cutaneous mechanoreceptors) in elderly subjects.
SR. PRESIDENTE DE LA COMISIÓN ACADÉMICA DEL PROGRAMA DE DOCTORADO EN CIENCIAS DE LA SALUD

ÍNDICE
1.- Introducción 2.- Estado actual del problema 2.1. Sistema somatosensorial del tacto 2.2. Formaciones nerviosas sensitivas: corpúsculos sensitivos
2.2.1. Corpúsculos de Meissner 2.2.2. Corpúsculos de Pacini 2.2.3. Complejos célula de Merkel-axón
2.3. Matriz extracelular: glicosaminoglicanos y proteoglicanos 2.3.1. Conceptos generales de la matriz extracelular 2.3.2. Proteoglicanos y glicosaminoglicanos de la matriz extracelular 2.3.3. Localización de los proteoglicanos en el sistema nervioso periférico 2.3.4 Función de los proteoglicanos en el sistema nervioso periférico
2.4. Edad y tacto 2.4.1. Envejecimiento del sistema somatosensorial del tacto 2.4.2. Cambios edad-dependientes en los mecanorreceptores cutáneos de mamíferos
3.- Hipótesis de trabajo y objetivos 4.- Material y técnicas 4.1. Material 4.2. Técnicas 4.2.1. Tinción hematoxilina-eosina 4.2.2. Inmunohistoquímica simple indirecta 4.2.3. Doble inmunohistoquímica indirecta
4.2.4. Doble inmunofluorescencia indirecta con microscopía confocal 4.2.5. Análisis cuantitativo 4.2.6. Análisis estadístico
5.- Resultados 5.1. CD34 5.2. Condroitín sulfato 5.3. Heparán sulfato 5.4. SLRPs 5.5 Capítulo libro 5.6. Envejecimiento
6.- Discusión 7.- Conclusiones 8.- Bibliografía

1. INTRODUCCIÓN

El grupo de investigación en el que se ha realizado el presente trabajo de tesis
doctoral, denominado SINPOS (Sistema Nervioso Periférico y Órganos de los Sentidos)
perteneciente al Cluster de Biomedicina del Campus de Excelencia Internacional de la
Universidad de Oviedo, tiene entre sus líneas de investigación prioritarias el estudio de
los corpúsculos sensitivos cutáneos. Los análisis incluyen la estructura, perfil
inmunohistoquímico, desarrollo y envejecimiento, así como diferentes patologías del
sistema nervioso en las que se ven alterados. El resultado de 30 años de investigación
ha dado lugar a numerosas publicaciones científicas en prestigiosas revistas
internacionales (ver revisiones Vega y Cols., 1996; 2009; 2012; Montaño y Cols., 2010;
del Valle y Cols., 2012).
Los conocimientos acerca de los corpúsculos sensitivos están en continua expansión. A
lo largo de los años, se ha tratado de descubrir la naturaleza de las células que forman
los distintos compartimentos de los corpúsculos sensitivos cutáneos, así como de
determinar marcadores específicos para cada uno de ellos. Un área de especial interés
ha sido el estudio del patrón de expresión proteico en los constituyentes celulares de
los corpúsculos sensitivos. El grupo SINPOS ha contribuido notablemente en este
campo de investigación, demostrando la expresión de numerosas proteínas en los
distintos tipos celulares de los corpúsculos: proteínas ligantes del calcio, filamentos
intermedios, receptores de factores de crecimiento, neurotrofinas o canales iónicos.
Teniendo en cuenta que los corpúsculos sensitivos son funcionalmente
mecanorreceptores, la presencia de canales iónicos en ellos da apoyo molecular a la
mecanorrecepción y mecanotransducción (García-Añoveros y Cols., 2001; Cabo y Cols.,
2012; 2015; Alonso-González y Cols., 2017; García-Mesa y Cols., 2017).
Sin embargo, tras varias décadas estudiando los corpúsculos sensitivos en nuestro
grupo de trabajo todavía existen vacíos de conocimiento sobre estas estructuras
sensitivas, y con esta tesis doctoral se pretende llenar o completar alguno de ellos.
En primer lugar, se tratará de establecer la naturaleza del núcleo externo de los
corpúsculos de Pacini humanos, desconocida hasta el momento, mediante
inmunohistoquímica para marcadores de células endoneural o perineural.

En segundo término, estudiar la matriz extracelular de los corpúsculos de Meissner y
de Pacini humanos, la cual ha sido ignorada casi por completo en los estudios sobre
estas estructuras, en lo que concierne a uno de sus componentes principales: los
proteoglicanos.
Y por último, las variaciones inmunohistoquímicas en algunas proteínas presentes en
los corpúsculos sensitivos asociadas con el envejecimiento, ya que se ha demostrado
que con la edad se produce un deterioro de la sensibilidad táctil que podría deberse, al
menos en parte, a alteraciones en los corpúsculos sensitivos. Actualmente, tan solo se
han publicado datos aislados y poco congruentes sobre los cambios edad-
dependientes del aparato sensitivo de la piel. En el presente trabajo de tesis doctoral
se estudian en detalle los cambios cuantitativos, cualitativos y morfológicos que
aparecen en los corpúsculos sensitivos cutáneos humanos con la edad. La consecución
de los objetivos planteados, los cuales se detallarán más adelante, contribuirá a
ampliar el conocimiento sobre los corpúsculos sensitivos. El trabajo permitirá
determinar el origen celular del núcleo externo de los corpúsculos de Pacini, con
especial interés de la lámina intermedia (referida como zona de crecimiento) y de
forma indirecta, identificar marcadores específicos para ella.
Por otra parte, tomando como referencia las funciones de los proteoglicanos en el
sistema nervioso periférico (regeneración/degeneración axonal, mantenimiento
estructural, mecanotransducción, etc.; ver para una revisión Chen y Birck, 2013), los
resultados que se aporten sobre la expresión y distribución de estas moléculas dentro
de los corpúsculos sensitivos permitirán hipotetizar sobre el papel que desempeñan en
estas formaciones sensitivas. Finalmente, el análisis de los corpúsculos sensitivos
humanos a diferentes edades, proporcionará datos más fiables que los existentes
hasta la fecha para establecer el papel de la porción más periférica del sistema
nervioso periférico en la disminución de la sensibilidad táctil que ocurre con el
envejecimiento.
En la última década, los corpúsculos sensitivos se han convertido en un elemento clave
para el diagnóstico clínico mediante biopsia cutánea de algunas neuropatías periféricas
y enfermedades neurodegenerativas (Herrmann y Cols., 2007; Nolano y Cols., 2008;
Almodovar y cols., 2012; Nolano y Cols., 2017). Este estudio constituirá una base

fundamental para establecer qué variaciones en estas formaciones sensitivas son
propias de la edad y no están asociadas a patologías, lo cual permitirá incluir de forma
sistemática a los corpúsculos sensitivos en la biopsia cutánea con fines diagnósticos

2. ESTADO ACTUAL DEL PROBLEMA

El eje central de esta tesis doctoral está constituido por los corpúsculos sensitivos
cutáneos, considerados funcionalmente mecanorreceptores. Con el fin de poder
abordar los objetivos del trabajo, realizar una adecuada interpretación y discusión de
los resultados es necesario realizar una revisión detallada sobre la estructura y
características funcionales de estas formaciones sensitivas.
2.1. Sistema somatosensorial del tacto
De los cinco sistemas sensoriales clásicos (auditivo, olfatorio, somatosensorial,
gustativo, y visual) solo el sistema somatosensorial es multimodal, es decir, es el único
capaz de detectar diferentes tipos de estímulos específicos tales como la posición de
las articulaciones (propiocepción), dolor, temperatura y tacto. Este último, a su vez,
incluye una gran variedad de componentes entre los que se encuentran la detección
de la curvatura, dureza, forma textura y presión.
La piel humana posee una rica inervación sensitiva a expensas de las prolongaciones
periféricas de las neuronas sensitivas localizadas a nivel de los ganglios sensitivos de
los pares craneales y de los ganglios raquídeos. Los extremos terminales de dichas
prolongaciones finalizan mayoritariamente a nivel de la dermis, formando los
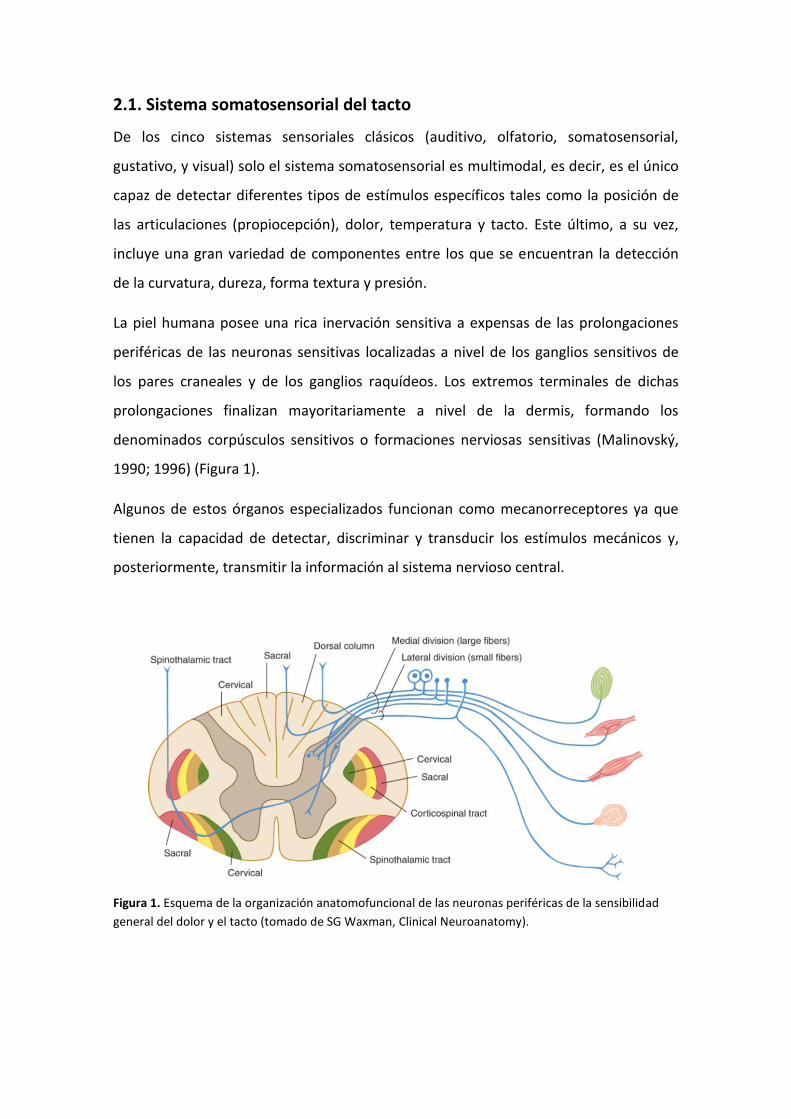
denominados corpúsculos sensitivos o formaciones nerviosas sensitivas (Malinovský,
1990; 1996) (Figura 1).
Algunos de estos órganos especializados funcionan como mecanorreceptores ya que
tienen la capacidad de detectar, discriminar y transducir los estímulos mecánicos y,
posteriormente, transmitir la información al sistema nervioso central.
Figura 1. Esquema de la organización anatomofuncional de las neuronas periféricas de la sensibilidad
general del dolor y el tacto (tomado de SG Waxman, Clinical Neuroanatomy).

Los corpúsculos sensitivos están formados por las mismas estructuras básicas que
constituyen las fibras nerviosas:
a) Axón central (zona dendrítica): formado a partir del extremo terminal de la
prolongación periférica del axón de las neuronas sensitivas periféricas. Puede
pertenecer a cualquier subtipo de fibra nerviosa (Lawson, 1992), aunque en el caso
de los corpúsculos sensitivos cutáneos de los mamíferos, suelen ser fibras Aα, Aβ y
Aδ (Dalsgaard, 1988; Perl, 1992).
b) Células de Schwann modificadas (células gliales periaxónicas): constituyen el
núcleo interno de los corpúsculos de Pacini y las células lamelares de los
corpúsculos de Meissner, y la glia asociada a los demás tipos de corpúsculos
sensitivos (Vega y Cols., 1992, 1993). Se originan a partir de células de la cresta
neural (Renehan y Munger, 1990; Saxod, 1996; Feito y Cols., 2018).
c) Estructuras relacionadas con el endoneuro: corresponden al estrato intermedio de
los corpúsculos de Pacini del mesenterio del gato descrito por Ide y Hayashi (1987)
y se originan a partir de fibroblastos endoneurales (Munger e Ide, 1988).
d) Estructuras relacionadas con el perineuro: corresponden a la cápsula de los
corpúsculos (en el caso de estar presente) y las lamelas del núcleo externo de los
corpúsculos de Pacini (Munger e Ide, 1988). Se originan por condensación a partir
del mesénquima local (Halata y Cols., 1990).
Debido a la continuidad estructural de los elementos que los forman, los corpúsculos
sensitivos y las fibras nerviosas comparten la mayoría de sus características
inmunohistoquímicas (Vega y Cols., 1994; 1996) (Figura 2).
En los mamíferos, los corpúsculos de Meissner, los corpúsculos de Pacini y los
complejos célula de Merkel-axón representan tres tipos de formaciones nerviosas
sensitivas cutáneas diferentes en base a su estructura y organización tisular. Los
corpúsculos de Pacini son el prototipo de corpúsculos capsulados mientras que los
corpúsculos de Meissner representan a los no capsulados o parcialmente capsulados.
Funcionalmente, ambos son receptores de bajo umbral y adaptación rápida y
dependen de fibras nerviosas sensitivas Aα y Aβ originadas de neuronas sensitivas de
tamaño grande o intermedio (Perl, 1992; Zelená, 1994; Johnson, 2001). Los complejos
célula de Merkel-axón son una asociación, a modo de sinapsis, entre una célula
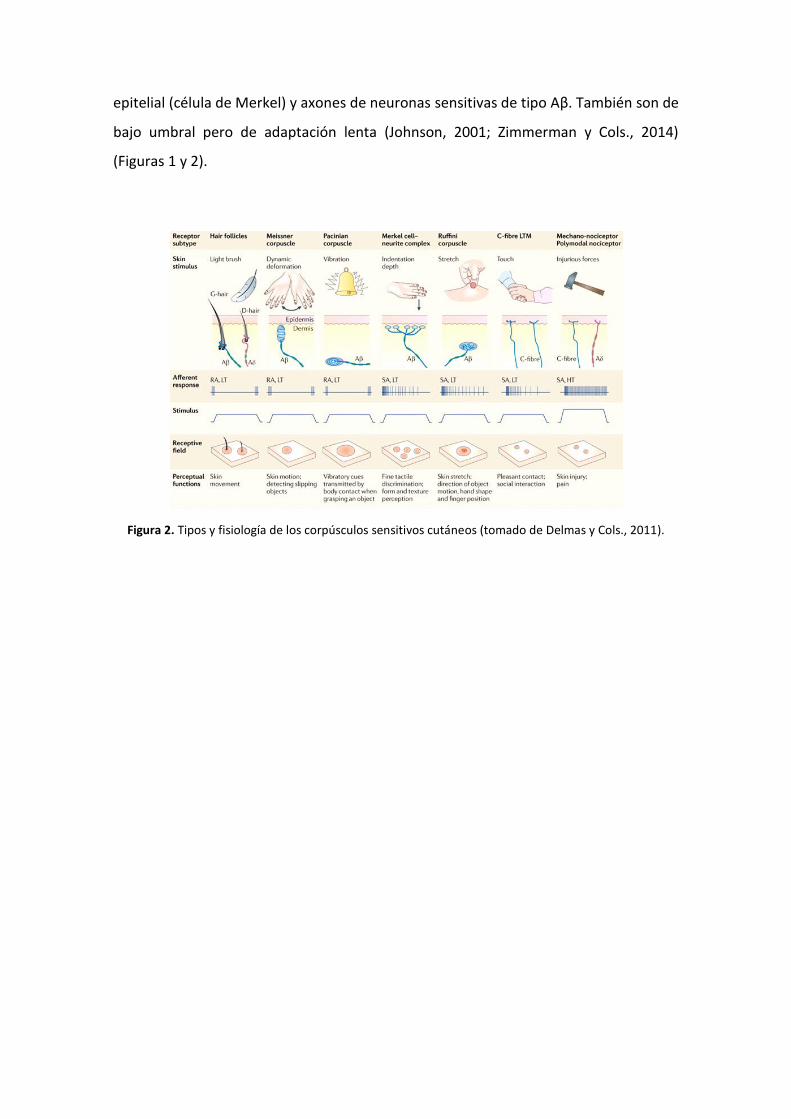
epitelial (célula de Merkel) y axones de neuronas sensitivas de tipo Aβ. También son de
bajo umbral pero de adaptación lenta (Johnson, 2001; Zimmerman y Cols., 2014)
(Figuras 1 y 2).
Figura 2. Tipos y fisiología de los corpúsculos sensitivos cutáneos (tomado de Delmas y Cols., 2011).

2.2. Formaciones nerviosas sensitivas: corpúsculos sensitivos
2.2.1. Corpúsculos de Meissner
Los corpúsculos de Meissner son estructuras sensitivas complejas localizadas en el
interior de las papilas dérmicas de la piel glabra con su eje mayor dispuesto
perpendicularmente a la superficie epidérmica. Suelen tener forma ovoide y su
tamaño varía entre 30-200 micras de longitud y 25-70 micras de diámetro (Guinard y
Cols., 2000; Vega y Cols., 2012) (Figura 3).
Están formados por terminales axónicos y células no nerviosas de naturaleza glial,
denominadas células lamelares o laminares, dispuestas a modo de pila de monedas
(lamelas) con su eje mayor paralelo a la superficie cutánea y núcleos de disposición
periférica. Pueden presentar una cápsula incompleta, sobre todo en la parte basal del
corpúsculo, formada por fibroblastos.
Una de las características morfológicas de los corpúsculos de Meissner es el curso
serpenteante del axón entre las lamelas. Este axón contiene numerosos filamentos
intermedios (neurofilamentos) en su parte central y abundantes mitocondrias
dispuestas preferentemente bajo el axolema. Cerca de la membrana hay vesículas de
transcitosis que recuerdan a las sinápticas. Además del axón principal, los corpúsculos
de Meissner pueden contener axones accesorios, en número variable, considerados de
naturaleza nociceptiva en base a los marcadores que expresan (Paré y Cols., 2001).
Normalmente, las fibras nerviosas que forman los corpúsculos de Meissner pierden la
vaina de mielina cuando entran en el corpúsculo, en el que pueden penetrar por la
base, por un lado o por la parte distal (Figuras 3 y 4). No obstante, se ha demostrado
que el axón del 25% de los corpúsculos de Meissner de la piel digital humana presenta
segmentos de mielina en su interior (García-Suárez y Cols., 2009).
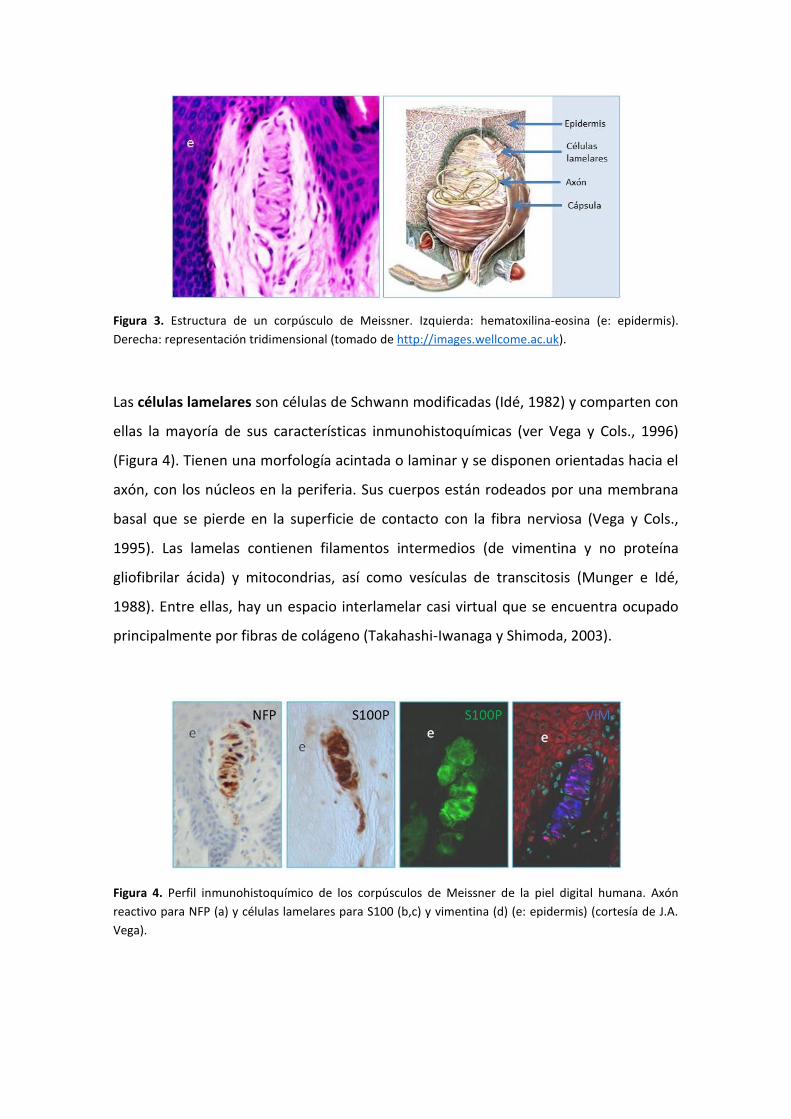
Figura 3. Estructura de un corpúsculo de Meissner. Izquierda: hematoxilina-eosina (e: epidermis).
Derecha: representación tridimensional (tomado de http://images.wellcome.ac.uk).
Las células lamelares son células de Schwann modificadas (Idé, 1982) y comparten con
ellas la mayoría de sus características inmunohistoquímicas (ver Vega y Cols., 1996)
(Figura 4). Tienen una morfología acintada o laminar y se disponen orientadas hacia el
axón, con los núcleos en la periferia. Sus cuerpos están rodeados por una membrana
basal que se pierde en la superficie de contacto con la fibra nerviosa (Vega y Cols.,
1995). Las lamelas contienen filamentos intermedios (de vimentina y no proteína
gliofibrilar ácida) y mitocondrias, así como vesículas de transcitosis (Munger e Idé,
1988). Entre ellas, hay un espacio interlamelar casi virtual que se encuentra ocupado
principalmente por fibras de colágeno (Takahashi-Iwanaga y Shimoda, 2003).
Figura 4. Perfil inmunohistoquímico de los corpúsculos de Meissner de la piel digital humana. Axón
reactivo para NFP (a) y células lamelares para S100 (b,c) y vimentina (d) (e: epidermis) (cortesía de J.A.
Vega).

Ocasionalmente, los corpúsculos de Meissner poseen una cápsula incompleta formada
por fibroblastos y fibras de colágeno. La cápsula está ausente a nivel superior del
corpúsculo, mientras que en la base inferior está compuesta por 2-4 estratos de
células capsulares aplanadas con fibrillas elásticas de colágeno dispuestas en los
espacios intercelulares. Estas células capsulares de la base pueden estar cubiertas por
láminas basales que estarían ausentes del resto del corpúsculo (Zelená, 1994).
Funcionalmente, los corpúsculos de Meissner son los responsables principales del
tacto fino. Se trata de mecanorreceptores de adaptación rápida, que responden a
estímulos comprendidos entre los 10 y 400 Hz, con una sensibilidad máxima para 100-
200 Hz (Johnson, 2001).
Por lo que se refiere a su composición proteica, la figura 5 resume las principales
proteínas descritas en los corpúsculos de Meissner según su localización.
El axón presenta inmunorreacción para marcadores neuronales generales como la
enolasa neuronal específica y la proteína PGP9.5 (Vega y Cols., 1996; Pawson y Cols.,
2000), además de para la subunidad de 200 kDa del neurofilamento, lo cual subraya el
carácter mecánico del mismo (Vega y Cols., 1996). También contiene proteínas ligantes
de calcio (calbindina D-28K, parvalbúmina, calretinina, y neurocalcina), las cuales
regulan presumiblemente la homeostasis del calcio axónico en la transducción
mecanoeléctrica (Vega y Cols., 1996; Albuerne y cols., 1998; Galeano y cols., 2000). La
β-arrestin-1 axónica juega un papel importante en la desensibilización e internalización
de los receptores asociados a proteínas G. Esta molécula emigra desde las neuronas de
los ganglios espinales a los corpúsculos que las suplen (Komori y cols., 2003). Otras
moléculas relacionadas con el crecimiento axónico, como la proteína GAP-43, también
están presentes en los corpúsculos cutáneos de Meissner (Verzé y cols., 2003).

Figura 5. Representación de las proteínas que se expresan en los corpúsculos de Meissner según su
localización (azul: células lamelares; rojo: axones accesorios; amarillo: axón principal).
Desde principios de este siglo, diferentes familias de canales iónicos, especialmente los
pertenecientes a las superfamilias de degenerina/epitelial- Na+ (DEG/ENaC) y Transient
Receptor Potential (TRP) han sido foco de interés en el estudio de los corpúsculos
sensitivos debido a su implicación en el proceso de mecanotransducción. En este
sentido, nuestro grupo de investigación ha centrado sus esfuerzos en la localización de
diferentes potenciales mecanoproteínas en los corpúsculos sensitivos, ampliando
considerablemente los conocimientos sobre este tema. A nivel proteico, se han
localizado BNaC1α, ASIC2, TRPV4 y TRPC6 en el axón principal de los corpúsculos de
Meissner de la piel digital humana (García-Añoveros y cols., 2001; Cabo y cols., 2012;
2015; Alonso-González y cols., 2017). Recientemente, hemos detectado en esta
situación el canal PIEZO2, perteneciente a una familia de canales iónicos reciente

identificada y caracterizada, relacionado de forma directa con la mecanotransducción
del tacto fino (García-Mesa y cols., 2017).
Respecto a los axones accesorios que ocasionalmente suplen los corpúsculos, se han
detectado en ellos diferentes neuropéptidos como la sustancia P, CGRP, hormona
estimulante de los melanocitos, galanina o somatostatina, todos ellos relacionados con
la nocicepción. Por supuesto estos axones también presentan inmunorreactividad para
anticuerpos neuronales específicos, especialmente la PGP9.5 (Johansson y Cols., 1999;
Paré y Cols., 2001).
En cuanto a las células lamelares, la vimentina es el principal filamento intermedio de
su citoesqueleto, y no la proteína gliofibrilar ácida (GFAP) como cabría esperar dada la
naturaleza glial de estas células (Vega y Cols., 1996). Además, Calavia y Cols. (2012)
demostraron la presencia de otro filamento intermedio, la nestina, en una pequeña
población de las células lamelares vimentina positivas, relacionándola con un posible
nicho de células neurales pluripotenciales presentes en los corpúsculos sensitivos. La
proteína ligante de calcio S100 es específica de las células lamelares y se co-localiza
con la parvalbúmina o calbindina D-28k (Vega y Cols., 1996). Por otro lado, los
receptores para las neurotrofinas p75, TrkA y TrkB también se detectan en este tipo
celular (Vega y Cols., 1993; Vega y Cols., 1994a; Bronzetti y Cols., 1995; Calavia y Cols.,
2010), así como el receptor para el factor de crecimiento epidérmico (Vega y Cols.,
1994b). Además, la membrana basal de las células lamelares contiene laminina y
colágeno tipo IV (Pawson y cols., 2000).
Existen algunas proteínas que no son específicas de ninguno de los componentes de
los corpúsculos y aparecen tanto en el axón como en las células lamelares. Es el caso
del canal iónico ASIC2 y el de algunas proteínas ligantes de calcio, que como se ha
mencionado previamente, aparecen en el axón pero también en una pequeña
población de las células lamelares (Vega y Cols., 2012; Cabo y Cols., 2015).
2.2.2. Corpúsculos de Pacini
Los corpúsculos de Pacini son los corpúsculos sensitivos de mayor tamaño (hasta 4 mm
su eje mayor y 2 mm su eje menor), tienen forma ovoide y detectan la presión y la

vibración. Se distribuyen por la mayoría de los órganos y tejidos, como páncreas,
mesenterio, mesocolon, adventicia de vasos y anastomosis arteriovenosas, piel,
tendones y ligamentos. En la piel, los corpúsculos de Pacini se sitúan en la dermis
profunda (Zelená, 1994).
Al microscopio óptico muestran un aspecto típico en “bulbo de cebolla”, debido a una
serie de formaciones laminares, más o menos concéntricas, dispuestas en torno al
axón. Se diferencian en ellos dos compartimentos denominados núcleo interno y
núcleo externo, ambos rodeados por una cápsula densa constituida por un número
variable de láminas (Figura 6 y 7).
Figura 6. Estructura de un corpúsculo de Pacini. Izquierda: hematoxilina-eosina (c: cápsula; ne: núcleo externo; ni: núcleo interno). Derecha: representación tridimensional (modificado de http://images.wellcome.ac.uk).
En la parte central del núcleo interno se dispone el axón, generalmente único, aunque
ocasionalmente puede contener hasta cinco o más (García-Suárez y Cols., 2010) (Figura
7). De forma característica, un axón mielínico de tipo Aα o Aβ entra por uno de los
polos del corpúsculo y termina en un engrosamiento con espículas denominado zona
dendrítica. El axón contiene neurofilamentos en su parte central, microtúbulos
dispersos y mitocondrias en grupos bajo el axolema. La fibra nerviosa que suple el
corpúsculo, en su trayecto intracorpuscular, mantiene uno o dos segmentos de mielina
y luego se rodea por las prolongaciones de las células laminares que forman el núcleo
interno (Bell y Cols., 1994). Además del axón principal, mecánico, los corpúsculos de
Pacini pueden tener fibras nerviosas accesorias, finas y amielínicas (fibras C o fibras
postganglionares simpáticas) que siguen un curso serpenteante en contraposición al
trayecto longitudinal de la fibra principal (Malinovský y Pác, 1982).

Las láminas o lamelas que forman el núcleo interno son células de Schwann
modificadas (Figura 7). Tienen una disposición muy compacta ya que están muy
próximas entre ellas. Permanecen separadas únicamente por espacios casi virtuales
que contienen fibrillas de colágeno muy finas (Idé y Cols., 1988; Malinovský y Cols.,
1990) y diferentes componentes de la matriz extracelular (Dubový y Bednárová, 1999;
Pawson y Cols., 2000; Sames y Cols., 2001). Las lamelas del núcleo interno carecen de
lámina basal, salvo las más externas (Munger e Idé, 1988).
Figura 7. Perfil inmunohistoquímico de los corpúsculos de Pacini de la piel digital humana. Axones
reactivos para el neurofilamento (NFP) y células del núcleo interno para S100P (cortesía de J.A. Vega).
El núcleo externo está formado por células aplanadas que rodean por completo al
núcleo interno, formando capas separadas entre sí por un compartimento con
diferentes componentes de la matriz extracelular (Dubový y Bednárová, 1999; Pawson
y Cols., 2000; Sames y Cols., 2001). Las células del núcleo interno muestran uniones
tipo “gap” o comunicantes, mientras que las lamelas del núcleo externo tienen
uniones intercelulares de tipo “tigh junction” o estrechas (Munger e Ide, 1988).
Entre los núcleos interno y externo se dispone un estrato celular intermedio, que ha
sido demostrado de manera fehaciente en los corpúsculos de Pacini del mesenterio del
gato (Idé y Cols., 1988; Munger e Idé, 1988; Pawson y Cols., 2000) y, recientemente, en
los corpúsculos de Pacini humanos por nuestro grupo de investigación (García-
Piqueras y Cols., 2017).
La cápsula es continuación del perineuro del tronco nervioso que inerva el corpúsculo
(Zelená, 1994). Su función podría ser la de aportar cierta tensión al núcleo externo
(Munger e Idé, 1988). En ella se localizan vasos sanguíneos y macrófagos. Se ha
sugerido que la cápsula y sus lamelas no solo tienen importancia en el procesamiento

de los estímulos mecánicos, sino que constituyen una parte del sistema metabólico del
corpúsculo (Malinovský y Cols., 1990).
La estructura de un corpúsculo de Pacini no es uniforme a lo largo del recorrido del
axón, sino que se distinguen tres zonas de acuerdo a las relaciones entre él y las células
periaxónicas (Malinovský y Cols., 1986) (Figura 8):
a) segmento preterminal: situado dentro de la cápsula y en la que el axón aún
permanece envuelto por la vaina de mielina y las células de Schwann;
b) segmento terminal: las láminas se disponen formando dos mitades simétricas,
separadas entre sí por las denominadas “clefts” o hendiduras. En esta zona, el axón
se caracteriza por emitir pequeñas prolongaciones, espículas, que se introducen en
dichas hendiduras. Las espículas siempre encaran las dos hendiduras del núcleo
interno en la porción terminal del corpúsculo (Munger e Idé, 1988); y
c) segmento ultraterminal: se corresponde con la parte final del axón, que está
engrosado y presenta numerosas espículas en su superficie. A este nivel, las células
del núcleo interno pierden la simetría bilateral y se disponen de forma irregular.
Figura 8. Componentes estructurales de un corpúsculo de Pacini, representado en cortes transversales a
nivel de la zona preterminal (A), terminal (B) y ultraterminal (C) (modificado de Malinovský y Cols.,
1986).
Funcionalmente, los corpúsculos de Pacini son mecanorreceptores de adaptación
rápida que responden a estímulos vibratorios comprendidos entre los 20 y los 1500 Hz,
con sensibilidad máxima en los 200-400 Hz, y por lo tanto, detectan presión y
vibración.
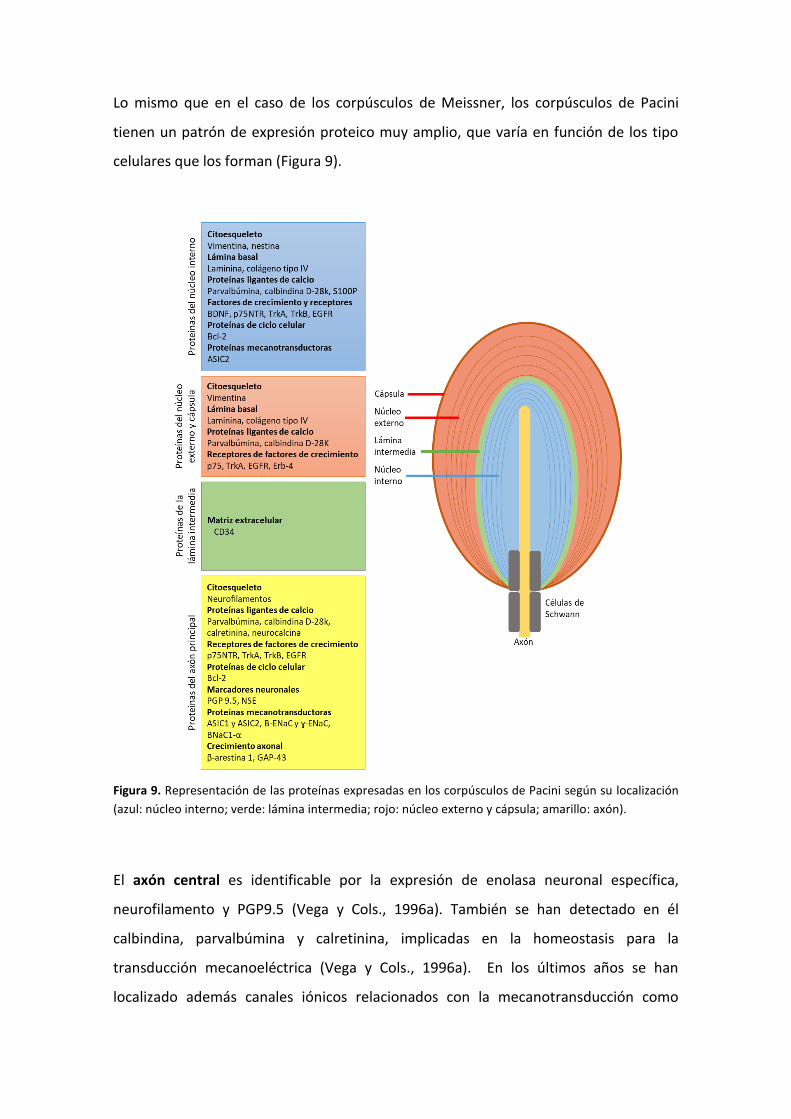
Lo mismo que en el caso de los corpúsculos de Meissner, los corpúsculos de Pacini
tienen un patrón de expresión proteico muy amplio, que varía en función de los tipo
celulares que los forman (Figura 9).
Figura 9. Representación de las proteínas expresadas en los corpúsculos de Pacini según su localización
(azul: núcleo interno; verde: lámina intermedia; rojo: núcleo externo y cápsula; amarillo: axón).
El axón central es identificable por la expresión de enolasa neuronal específica,
neurofilamento y PGP9.5 (Vega y Cols., 1996a). También se han detectado en él
calbindina, parvalbúmina y calretinina, implicadas en la homeostasis para la
transducción mecanoeléctrica (Vega y Cols., 1996a). En los últimos años se han
localizado además canales iónicos relacionados con la mecanotransducción como

ASIC2 y ASC1 en el axón de los corpúsculos de Pacini de Macaca fascicularis (Cabo y
Cols., 2012) y humanos respectivamente (Calavia y Cols., 2010). Por otro lado, se ha
descrito la proteína Bcl-2 en el axón principal (González-Martínez y Cols., 2006).
El resto de elementos que constituyen los corpúsculos de Pacini son el núcleo interno,
el núcleo externo y la cápsula. Una contribución importante de nuestro grupo de
investigación ha sido la identificación de antígenos que son específicos de
determinados compartimentos del corpúsculo de Pacini, como la proteína S100 para el
núcleo interno (Vega y cols., 1996b) y el CD34 para la lámina intermedia (García-
Piqueras y Cols., 2017). Hasta el momento actual, sólo se ha encontrado expresión de
la mecanoproteína ASIC2 en el núcleo interno de los corpúsculos de Pacini humanos
(Calavia y Cols., 2010; Cabo y Cols., 2015), al igual que la proteína básica de la mielina,
cuya expresión se restringe al segmento preterminal del núcleo interno (García-Suárez
y Cols., 2009).
Sin embargo, la mayoría de las proteínas suelen expresarse en varios elementos
corpusculares: 1) los filamentos intermedios vimentina y nestina aparecen en el
núcleo interno y núcleo externo (la vimentina también en la cápsula) (Calavia y Cols.,
2012); 2) las proteínas de membrana basal colágeno IV y laminina se localizan en
todos los elementos no neurales del corpúsculo (Vega y Cols., 1995); 3) los receptores
de neurotrofinas TrkA, p75 y EGFR se encuentran en todos los compartimentos (Vega
y Cols., 1994a; 1994b), TrkB en el axón y ocasionalmente en las células lamelares del
núcleo interno de los corpúsculos de Pacini de Macaca fascicularis (Cabo y Cols., 2015),
y su ligando BDNF en el núcleo interno y núcleo externo (Cabo y Cols., 2015).
No obstante, también se dan situaciones contradictorias. Mientras que para Britsch
(2007) el axón central y las células de Schwann presentan inmunorreactividad para los
receptores de neorregulinas erb2, erb3, erb4, nuestro grupo de investigación ha
demostrado que erb4 exclusivamente aparece en la cápsula y en el núcleo externo de
los corpúsculos de Pacini (González-Martínez y Cols., 2007).
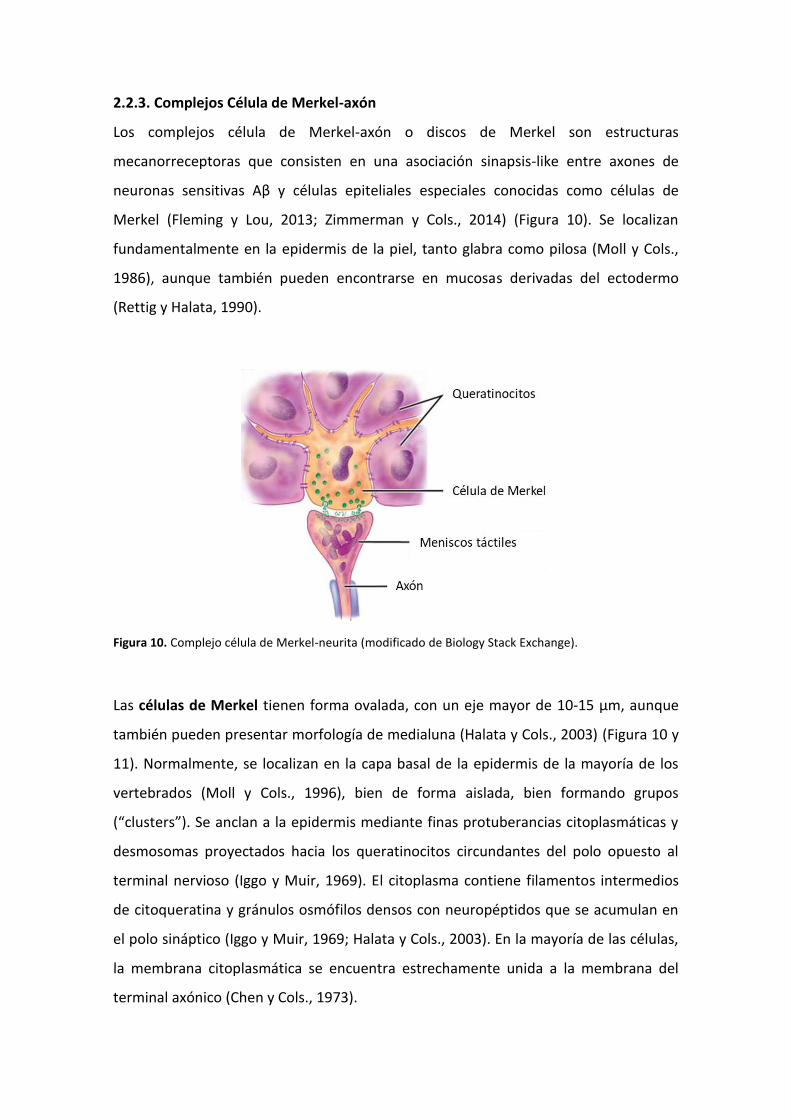
2.2.3. Complejos Célula de Merkel-axón
Los complejos célula de Merkel-axón o discos de Merkel son estructuras
mecanorreceptoras que consisten en una asociación sinapsis-like entre axones de
neuronas sensitivas Aβ y células epiteliales especiales conocidas como células de
Merkel (Fleming y Lou, 2013; Zimmerman y Cols., 2014) (Figura 10). Se localizan
fundamentalmente en la epidermis de la piel, tanto glabra como pilosa (Moll y Cols.,
1986), aunque también pueden encontrarse en mucosas derivadas del ectodermo
(Rettig y Halata, 1990).
Figura 10. Complejo célula de Merkel-neurita (modificado de Biology Stack Exchange).
Las células de Merkel tienen forma ovalada, con un eje mayor de 10-15 µm, aunque
también pueden presentar morfología de medialuna (Halata y Cols., 2003) (Figura 10 y
11). Normalmente, se localizan en la capa basal de la epidermis de la mayoría de los
vertebrados (Moll y Cols., 1996), bien de forma aislada, bien formando grupos
(“clusters”). Se anclan a la epidermis mediante finas protuberancias citoplasmáticas y
desmosomas proyectados hacia los queratinocitos circundantes del polo opuesto al
terminal nervioso (Iggo y Muir, 1969). El citoplasma contiene filamentos intermedios
de citoqueratina y gránulos osmófilos densos con neuropéptidos que se acumulan en
el polo sináptico (Iggo y Muir, 1969; Halata y Cols., 2003). En la mayoría de las células,
la membrana citoplasmática se encuentra estrechamente unida a la membrana del
terminal axónico (Chen y Cols., 1973).

La fibra nerviosa de tipo Aβ que contacta con las células de Merkel es el axón de una
neurona mecanosensitiva de bajo umbral y adaptación lenta de tipo I, cuyas
terminaciones forman una ampliación discoide en su parte distal para hacer conexión
sinapsis-like con el polo basal de las células de Merkel. Esta asociación es la que
permite que el complejo sea capaz de iniciar la respuesta para la discriminación táctil
(Maksimovic y Cols., 2013; Zimmerman y Cols., 2014).
Figura 11. Perfil inmunohistoquímico del complejo célula de Merkel-Neurita. Izquierda: células de
Merkel inmunorreactivas para CK20. Derecha: arriba, célula de Merkel marcada con fluorescencia para
CK20 (rojo); abajo, célula de Merkel marcada con fluorescencia para PIEZO2 (verde) y enolasa neuro-
específica (rojo; las flechas blancas denotan la fibra nerviosa) (tomado de García-Mesa y Cols., 2017).
Respecto al origen embrionario de las células de Merkel se han barajado dos hipótesis
diferentes. La primera apostaba por un origen de la cresta neural, dado que son células
excitables con capacidad de sintetizar neuropéptidos y expresar moléculas
presinápticas (Grim y Halata, 2000; Szeder y Cols., 2003). La segunda hipótesis afirma
que las células de Merkel son de origen ectodérmico debido a que expresan
citoqueratinas de bajo peso molecular, no tienen capacidad mitótica y establecen
contactos mediante desmosomas con los queratinocitos (Munger, 1965; Compton y
Cols., 1990; Moll y Cols., 1990; Kim y Holbrook, 1995). Actualmente se acepta esta
segunda teoría, ya que se ha demostrado que las células de Merkel provienen de
células madre epidérmicas (Morrison y Cols., 2009; Van Keymeulen y Cols., 2009).

Además, en la epidermis se ha identificado una población celular con fenotipo distinto
al de los queratinocitos que residen junto a las células de Merkel y con capacidad de
regenerar este tipo celular (Woo y Cols., 2010).
La función principal del complejo célula de Merkel-axón es la mecanosensación, pero
existe una gran controversia sobre el papel que desempeñan las células de Merkel en
este proceso. Algunos autores detallan que la fibra nerviosa de tipo Aβ es la
responsable de la mecanotransducción en el complejo, mientras que la célula de
Merkel ocupa un papel modulador (Kinkelin y Cols., 1999). Otros autores, por el
contrario, establecen que la célula de Merkel es el componente indispensable del
complejo para la transducción del estímulo mecánico en la señal química que
posteriormente activará la fibra nerviosa a la que se encuentra unida (Maricich y Cols.,
2009). Una tercera alternativa defiende que ambos son imprescindibles y necesarios,
es decir, que tanto la célula de Merkel como la fibra nerviosa están implicadas en el
proceso de mecanotransducción (Yamashita y Ogawa, 1991; Fleming y Luo, 2013;
Maksimovic y Cols., 2014).
En estudios con ratones que carecen de células de Merkel (knock-out para Atoh1),
pero que conservan las terminaciones nerviosas de tipo Aβ, no se produce reacción
ante estímulos de presión sostenida, resultando la sensación del tacto ligero
completamente alterada (Maricich y Cols., 2009). También existen evidencias de que
las células de Merkel producen neurotransmisores capaces de actuar sobre la
terminación nerviosa a modo de sinapsis-like (Hartschuh y Weihe, 1980; Tachibana y
Nawa, 2002; Haeberle y Cols., 2004; Hitchcock y Cols., 2004). Recientemente, se ha
demostrado la presencia del canal iónico mecanosensitivo PIEZO2 (Ranade y Cols.,
2014; Xiao y Cols., 2014) en las células de Merkel de piel glabra digital humana, pero
no en el terminal axónico (García-Mesa y Cols., 2017). Todos estos resultados avalan la
segunda de las hipótesis mencionadas anteriormente, según la cual, las células de
Merkel transducen los estímulos mecánicos en señales eléctricas mediante PIEZO2 y,
en consecuencia, se inducen los potenciales de acción en las fibras aferentes mediante
la activación de canales de calcio dependientes de voltaje. Aunque a día de hoy,
tampoco puede descartarse la tercera alternativa, en la que ambos componentes del
complejo serían mecanosensibles y, por tanto, ambos participarían en la
mecanotransducción (Maksimovic y Cols., 2013).
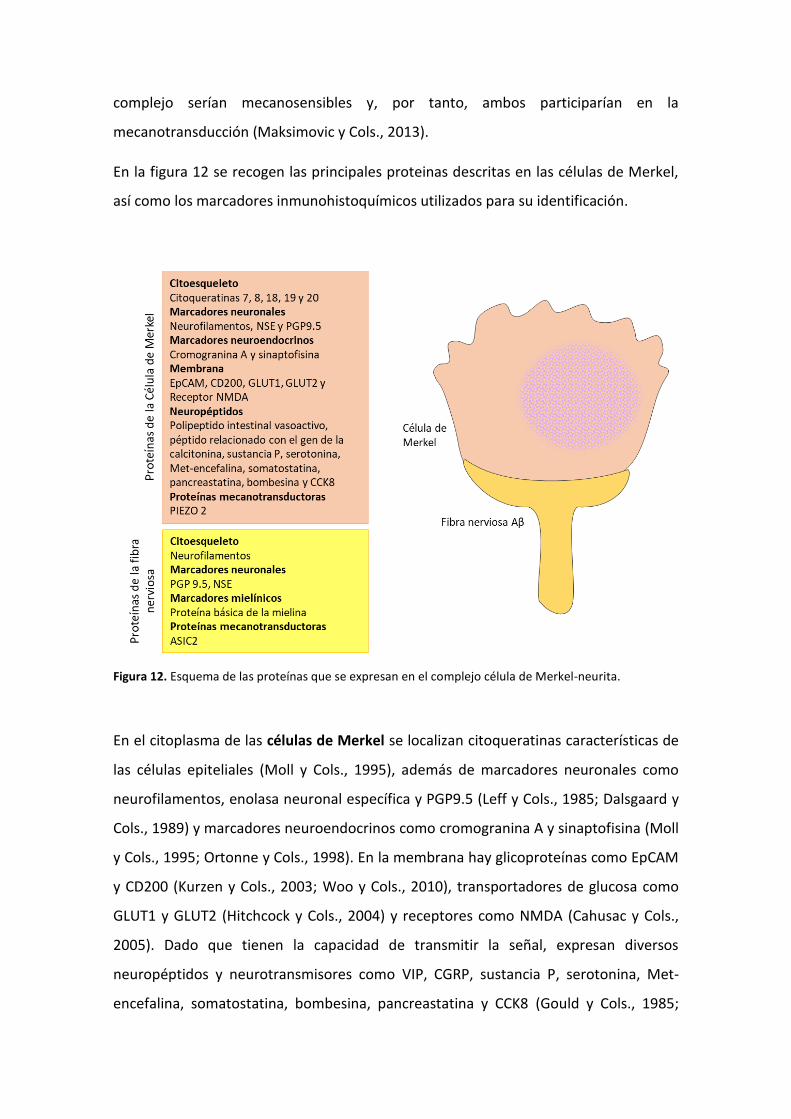
En la figura 12 se recogen las principales proteinas descritas en las células de Merkel,
así como los marcadores inmunohistoquímicos utilizados para su identificación.
Figura 12. Esquema de las proteínas que se expresan en el complejo célula de Merkel-neurita.
En el citoplasma de las células de Merkel se localizan citoqueratinas características de
las células epiteliales (Moll y Cols., 1995), además de marcadores neuronales como
neurofilamentos, enolasa neuronal específica y PGP9.5 (Leff y Cols., 1985; Dalsgaard y
Cols., 1989) y marcadores neuroendocrinos como cromogranina A y sinaptofisina (Moll
y Cols., 1995; Ortonne y Cols., 1998). En la membrana hay glicoproteínas como EpCAM
y CD200 (Kurzen y Cols., 2003; Woo y Cols., 2010), transportadores de glucosa como
GLUT1 y GLUT2 (Hitchcock y Cols., 2004) y receptores como NMDA (Cahusac y Cols.,
2005). Dado que tienen la capacidad de transmitir la señal, expresan diversos
neuropéptidos y neurotransmisores como VIP, CGRP, sustancia P, serotonina, Met-
encefalina, somatostatina, bombesina, pancreastatina y CCK8 (Gould y Cols., 1985;

Hartschuh y Weihe; 1989). Maksimovic y Cols., 2003). La prueba fehaciente de su
carácter mecanotransductor es la expresión de PIEZO2 (García-Mesa y Cols., 2017).
La fibra nerviosa de tipo Aβ presenta inmunorreacción positiva para marcadores
neuronales generales como cabría esperar (Dalsgaard y Cols., 1989; Gould y Cols.,
1985; Narisawa y Cols., 1994). Se ha demostrado también la expresión de la proteína
mecanotransductora ASIC2 (Cabo y Cols., 2015).

2.3. Matriz extracelular: glicosaminoglicanos y proteoglicanos
2.3.1. Conceptos generales sobre la matriz extracelular
Los tejidos de los organismos pluricelulares están formados, no solo por un conjunto
organizado de células, sino también por componentes no celulares que se localizan en
el espacio que rodea a las mismas, donde forman una estructura ordenada
denominada matriz extracelular (Alberts y Cols., 2002; Theocharis y Cols., 2016).
La matriz extracelular es una red compleja formada por moléculas de diferente
naturaleza que pueden ser clasificadas en tres grupos (Alberts y Cols., 2002; Bosman y
Stamenkovic, 2003; Theocharis y Cols., 2016) (Figura 13):
a) Proteínas fibrosas: diferentes tipos de colágenos y elastinas;
b) Proteínas de adhesión: lamininas, fibronectinas, tenascinas, fibulinas y nidógeno; y
c) Proteoglicanos constituidos por diferentes tipos de glicosaminoglicanos.
No obstante, esta composición es diferente en cada tejido, en cada situación
fisiológica/patológica y en cada especie (Frantz y Cols., 2010; Theocharis y Cols., 2016).
Dentro de la matriz extracelular de los tejidos, aparecen frecuentemente dos motivos
estructurales diferentes (Figura 13):
a) Membrana basal: estructura bien definida que se encuentra en el exterior celular,
pero inmediatamente a continuación de la membrana citoplasmática (LeBleu y
Cols., 2007). Sus componentes más conocidos son colágeno IV y laminina, pero
también los proteoglicanos agrina, perlecano y endostatina, y otras glicoproteínas
como nidógeno y fibulinas (Sasaki y Cols., 2004).
b) Matriz intersticial: correspondiente al resto de moléculas de matriz que rellenan el
espacio extracelular sin relación con la membrana basal (Alberts y Cols., 2002;
Bosman y Stamenkovic, 2003). Su composición tiene mayor variabilidad según el
tejido en el que se encuentra (Couchman y Pataki, 2012).
Una de las peculiaridades de la matriz extracelular es su alto grado de dinamismo, es
decir, las enzimas son capaces de producir, modificar y degradar la matriz extracelular
permanentemente (Page-McCaw y Cols., 2007; Pengfei y Cols., 2011; Bonnans y Cols.,
2014).
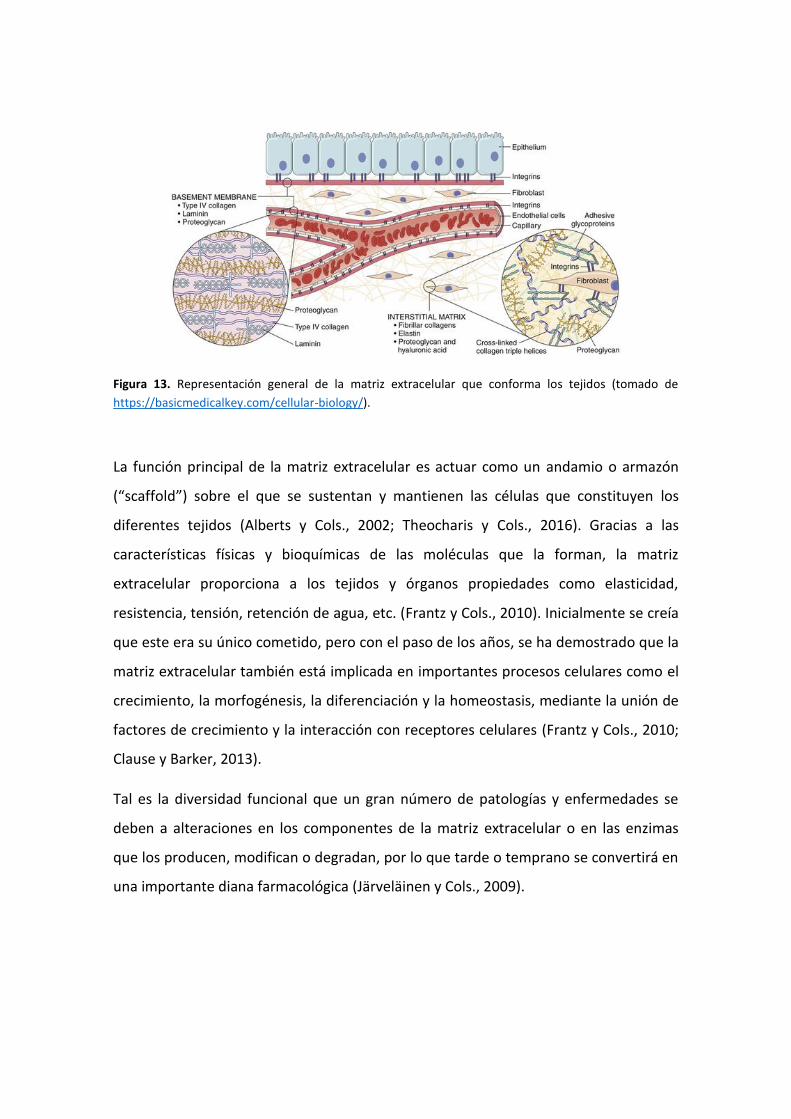
Figura 13. Representación general de la matriz extracelular que conforma los tejidos (tomado de
https://basicmedicalkey.com/cellular-biology/).
La función principal de la matriz extracelular es actuar como un andamio o armazón
(“scaffold”) sobre el que se sustentan y mantienen las células que constituyen los
diferentes tejidos (Alberts y Cols., 2002; Theocharis y Cols., 2016). Gracias a las
características físicas y bioquímicas de las moléculas que la forman, la matriz
extracelular proporciona a los tejidos y órganos propiedades como elasticidad,
resistencia, tensión, retención de agua, etc. (Frantz y Cols., 2010). Inicialmente se creía
que este era su único cometido, pero con el paso de los años, se ha demostrado que la
matriz extracelular también está implicada en importantes procesos celulares como el
crecimiento, la morfogénesis, la diferenciación y la homeostasis, mediante la unión de
factores de crecimiento y la interacción con receptores celulares (Frantz y Cols., 2010;
Clause y Barker, 2013).
Tal es la diversidad funcional que un gran número de patologías y enfermedades se
deben a alteraciones en los componentes de la matriz extracelular o en las enzimas
que los producen, modifican o degradan, por lo que tarde o temprano se convertirá en
una importante diana farmacológica (Järveläinen y Cols., 2009).

2.3.2. Proteoglicanos y glicosaminoglicanos de la matriz extracelular
Los glicosaminoglicanos son polisacáridos lineares constituidos por una unidad
disacárida que se repite a lo largo de su estructura molecular. Según el azúcar del
disacárido, existen varios tipos de glicosaminoglicanos: hialuronano, heparán sulfato,
heparina, condroitín sulfato, dermatán sulfato y keratan sulfato (Schaefer y Schaefer,
2010) (Figura 14). Los glicosaminoglicanos se encuentran unidos a una proteína
principal, formando moléculas denominadas proteoglicanos; todos, excepto el
hialuronano, que es sintetizado en la superficie celular y liberado directamente en el
medio extracelular (Schaefer y Schaefer, 2010).
Los proteoglicanos forman parte de la matriz extracelular de todos los tejidos de los
mamíferos, ya que todas las células son capaces de sintetizarlos y secretarlos al
exterior, insertarlos en la membrana plasmática o empaquetarlos intracelularmente
(Gandhi y Mancera, 2008). Pero no todas las matrices extracelulares contienen los
mismos proteoglicanos (Couchman y Pataki, 2012).
En la matriz extracelular de los mamíferos, se han identificado más de 40
proteoglicanos con una gran variabilidad a nivel genético y proteico, lo cual ha
dificultado su clasificación. Según se ha propuesto recientemente, los proteoglicanos
pueden agruparse en base a la localización celular y subcelular, la homología de
secuencia genética/proteica y la presencia de motivos proteicos particulares (Iozzo y
Schaefer, 2015) (Figura 15).

Figura 14. Esquema molecular de los glicosaminoglicanos (tomado de Papakonstantinou y Karakiulakis,
2009).
En general, los proteoglicanos de heparán sulfato, como por ejemplo las familias de
los sindecanos y de los glipicanos, localizados en la superficie celular, actúan como
receptores moleculares y participan en diferentes vías de transducción de la señal
relacionadas con el desarrollo y la proliferación celular (Carey, 1997; Filmus y Cols.,
2008; Lambaerts y Cols., 2009; Filmus y Capurro, 2014). Otro grupo de proteoglicanos
de heparán sulfato son los que forman parte de las membranas basales: agrina,
perlecano, colágeno XVIII (endostatina) y colágeno XV (Yurchenco y Patton, 2009).
Como ya se ha expuesto anteriormente, la membrana basal es una diferenciación de la
matriz extracelular que se encuentra en el exterior de la mayoría de células
(epiteliales, musculares, adipocitos, células de Schwann, etc.) (Bosman y Stamenkovic,
2003) y que cumple funciones específicas en cada tejido (LeBleu y Cols., 2007).
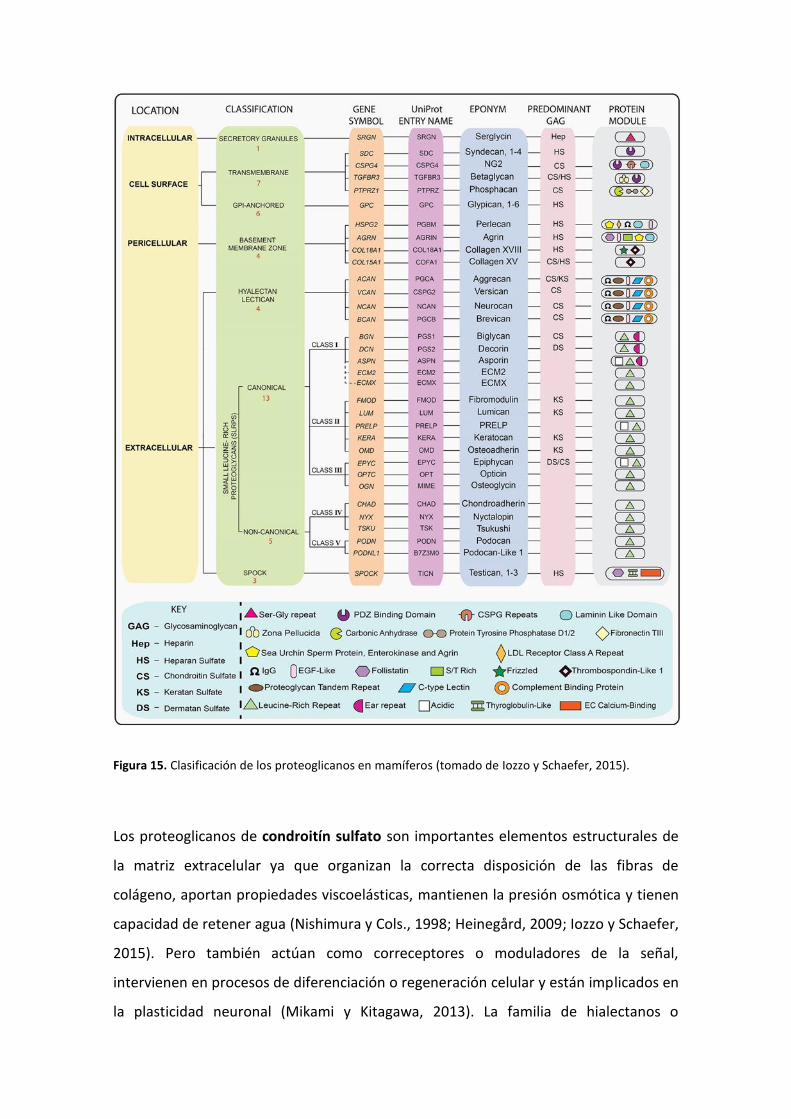
Figura 15. Clasificación de los proteoglicanos en mamíferos (tomado de Iozzo y Schaefer, 2015).
Los proteoglicanos de condroitín sulfato son importantes elementos estructurales de
la matriz extracelular ya que organizan la correcta disposición de las fibras de
colágeno, aportan propiedades viscoelásticas, mantienen la presión osmótica y tienen
capacidad de retener agua (Nishimura y Cols., 1998; Heinegård, 2009; Iozzo y Schaefer,
2015). Pero también actúan como correceptores o moduladores de la señal,
intervienen en procesos de diferenciación o regeneración celular y están implicados en
la plasticidad neuronal (Mikami y Kitagawa, 2013). La familia de hialectanos o

lecticanos es un ejemplo de proteoglicanos de condroitín sulfato de localización
extracelular, especialmente relevantes en el sistema nervioso central (Crespo-
Santiago, 2004; Howell y Gottschall, 2012). Sin embargo, fosfacano, NG2 y β-glicano
son proteoglicanos de condroitín sulfato insertados en la membrana citoplasmática y,
por tanto, asociados a la superficie celular (López-Casillas y Cols., 1991; Nishiyama y
Cols., 1991; Maurel y Cols., 1994).
La familia más grande de proteoglicanos la constituyen los SLRP (“small leucine-rich
proteoglycans”), también de localización extracelular, dentro de la cual se distinguen
cinco clases. Su similar estructura proteica hace que tengan características y funciones
comunes, como por ejemplo, regular la fibrilogénesis mediante la unión a colágenos
(Kalamajski y Oldberg, 2010; Chen y Birk, 2013) o influenciar la actividad celular
mediante la unión a receptores de superficie celular, factores de crecimiento,
citoquinas u otros componentes de la matriz extracelular (Schaefer y Iozzo, 2008).
Participan en importantes vías de señalización celular como la del factor de
crecimiento transformante β (TGF-β) y la proteína morfogénica ósea (BMP)
(Hildebrand y Cols., 1994). Sin embargo, cada uno de ellos también puede desempeñar
funciones únicas en tejidos específicos gracias a la variabilidad del extremo N-terminal
y a la unión de los diferentes glicosaminoglicanos (Iozzo y Schaefer, 2015).
2.3.3. Localización de los proteoglicanos en el sistema nervioso periférico
En el sistema nervioso periférico, alrededor de las células de Schwann se dispone una
membrana basal sobre la que se cimientan el resto de los constituyentes de la matriz
extracelular (Feltri y Wrabetz, 2005), entre ellos, los proteoglicanos.
Como ya se ha mencionado, algunos de los proteoglicanos de heparan sulfato son
constituyentes importantes de la estructura de las membranas basales. Mediante
cultivos celulares se ha demostrado que las células de Schwann y las neuronas
sensitivas expresan proteoglicanos de heparán sulfato en su membrana basal (Mehta y
Cols., 1985); en este sentido, se han detectado tres de ellos (perlecano, agrina y
endostatina) en las membranas basales de las células de Schwann de nervios y ganglios
periféricos (Halfter y Cols., 1998; Ma y Cols., 1994; Halfter y Cols., 1997; Chernousov y

Cols., 1998). Además, se ha localizado perlecano en los nódulos de Ranvier de
neuronas periféricas (Bangratz y Cols., 2012). Los otros dos grandes grupos de
proteoglicanos de heparán sulfato (sindecanos y glipicanos) también se han descrito en
el sistema nervioso periférico. Los glipicanos 1 y 3 se expresan en las neuronas de los
ganglios raquídeos (Litwack y Cols. 1994; Bloechlinger y Cols., 2004; Iglesias y Cols.,
2008), mientras que los sindecanos 3 y 4 en la periferia de las células de Schwann
(Carey y Cols., 1992; Erdman y Cols., 2002; Goutebroze y Cols., 2003) y en los nódulos
de Ranvier (Melendez-Vasquez y Cols., 2005). El receptor del factor de crecimiento
transformante β3, también conocido como betaglicano, el cual está formado por
cadenas de heparan sulfato y de condroitín sulfato, también se expresa en las células
de Schwann (Thomas y De Vries, 2007).
Los proteoglicanos de condroitín sulfato son mayoritarios en el sistema nervioso
central (Bandtlow y Zimmermann, 2000), mientras que en el periférico su distribución
es menor o no ha sido tan estudiada hasta la fecha. Mediante microscopia electrónica
e inmunohistoquímica, se ha demostrado en nervios periféricos su expresión en la
lámina basal de las células de Schwann (Aquino y Cols., 1984) y en el endoneuro
alrededor del axón (Tona y Cols., 1993; Morrison y Cols., 1994); concretamente son
versicano, agrecano y NG2 (Neural/glial antigen 2; Bode-Lesniewska y Cols., 1996;
Rezajooi y Cols., 2004; Ali y Cols., 2011). Además, también se ha encontrado versicano
en los nódulos de Ranvier (Melendez-Vasquez y Cols., 2005).
Todavía es más desconocido el patrón de expresión de los SLRP en el sistema nervioso
periférico y, en la literatura científica, son escasas las menciones a estos
proteoglicanos. Estudios de hibridación in situ han demostrado que las células de
Schwann expresan decorina (Hanemann y Cols., 1993). También se han detectado
decorina y lumicano en ganglios raquídeos al final de la segunda semana de desarrollo
del ratón (Wilda y Cols., 2000) y biglicano en el nervio óptico adulto (Ali y Cols., 2011).
En relación a los corpúsculos sensitivos, los proteoglicanos se encuentran embebidos
en la matriz extracelular correspondiente a cada compartimento corpuscular, aunque a
día de hoy, la información existente sobre la misma es escasa. Actualmente, se conoce
que tanto los corpúsculos de Pacini como los corpúsculos de Meissner expresan
laminina y colágeno tipo IV (componentes de lámina basal) en las células periaxónicas,

así como en la cápsula (Halata 1975; Zelená 1994; Vega y Cols., 1995). Los estudios de
Dubový y Bednárová (1999a) y Chouchkov y Cols. (2003) confirmaron la presencia de
distintas lamininas en las zonas de los corpúsculos de Pacini en las que ya se había
descrito la lámina basal (próxima a las células perineurales y lamelas), pero también en
las células de Schwann modificadas de los corpúsculos de Pacini (Dubový y Bednárová,
1999a; 1999b). Por otro lado, en los espacios interlamelares del núcleo externo y de la
cápsula de los corpúsculos de Pacini del mesenterio del gato predomina el colágeno
tipo II, mientras que la lámina intermedia contiene colágeno tipo IV (Pawson y Cols.,
2000). Los espacios equivalentes de los corpúsculos de Meissner de Macaca fuscata
fueron estudiados por Takahashi-Iwanaga y Shimoda (2003), demostrando de igual
forma la existencia de colágeno. Todavía resultan más desconocidos los proteoglicanos
que componen dicha matriz extracelular en los corpúsculos sensitivos. Los primeros
estudios en los que se documenta la presencia de proteoglicanos son los realizados por
Dubový y Svízenská (1993) y Dubový y Bednárová (1999). Tan sólo existe otro estudio
en corpúsculos de Pacini del mesenterio del gato en el que fueron localizados decorina
y biglicano en las lamelas perineurales (Sames y Cols., 2001). No obstante, el contenido
de proteoglicanos de la matriz extracelular es complejo y varía en cada zona del
corpúsculo (Dubový y Bednárová, 1999).
2.3.4. Funciones de los proteoglicanos en el sistema nervioso periférico
En el sistema nervioso periférico, la matriz extracelular proporciona el entorno físico-
químico adecuado para el desarrollo y la supervivencia de las neuronas y de las células
de la glía ya que está implicada en múltiples actividades y procesos celulares (Barros y
Cols., 2001; Gardiner, 2011):
a) Desarrollo embrionario: el sistema nervioso periférico se forma tras una serie de
procesos migratorios coordinados que experimentan las células de cresta neural en
los que los proteoglicanos juegan un papel fundamental. La expresión de los
proteoglicanos de condroitín sulfato en determinadas zonas, se relaciona
directamente con la ausencia de células de cresta neural durante el desarrollo
(Perris y Cols., 1991). Por otra parte, los SLRP están prácticamente ausentes

durante el movimiento de las células de cresta neural, pero muestran un aumento
de expresión en la unión dermoepidérmica, lo que les relaciona presumiblemente
con la guía de las terminaciones nerviosas sensitivas que inervan la piel (Perris,
1997). Además, los sindecanos podrían estar implicados en la proliferación de las
células de Schwann antes de la formación del nódulo de Ranvier (Melendez-
Vasquez y Cols., 2005), así como en las interacciones axogliales en el desarrollo
(Goutebroze y Cols., 2003).
b) Crecimiento axonal y mielinización: La lámina basal de las células de Schwann, de
la que forman parte diferentes proteoglicanos de heparán sulfato, interviene en la
migración y diferenciación de las células de Schwann, en su disposición alrededor
del axón y, al mismo tiempo junto a integrinas, en la mielinización (McGarvey y
Cols., 1984; Anton y Cols., 1994; Shorer y Cols., 1995; Feltri y Cols., 2002). Se ha
sugerido que la agrina participa en el proceso de crecimiento axonal mediante la
unión a factores de crecimiento y a proteínas de adhesión (Halfter y Cols., 1997).
c) Regeneración axonal: En un nervio adulto intacto predominan los componentes de
matriz inhibitorios para el crecimiento axonal, de forma que se controla la
proliferación neurítica. Sin embargo, en un nervio lesionado, la matriz extracelular
cambia su composición para crear un ambiente favorable a la regeneración. Tras
una lesión en un nervio periférico, se produce un aumento de proteoglicanos de
condroitín sulfato, que impiden la regeneración axonal al inhibir el efecto positivo
de las lamininas de las células de Schwann sobre el crecimiento neurítico (Muir y
Cols., 1989; Zuo y Cols., 1998). En este mismo sentido, se ha demostrado que la
degradación de proteoglicanos de condroitín sulfato en neuronas sensitivas,
favorece la regeneración axónica tras una lesión nerviosa (Udina y Cols., 2010). Por
el contrario, la expresión de proteoglicanos de heparan sulfato está relacionada
con la permisividad para el crecimiento y prolongación axonal, como se ha sugerido
con los glipicanos en el proceso de regeneración (Litwack y Cols. 1994; Bloechlinger
y Cols., 2004).
Actualmente, poco se conoce sobre la matriz extracelular de los corpúsculos sensitivos
de la piel glabra digital humana. La existencia de lámina basal (estructura derivada de
la matriz extracelular) en los componentes perineurales de los corpúsculos sensitivos

(Vega y Cols., 1995; Dubový y Bednárová, 1999; Chouchkov y Cols., 2003), podría estar
relacionada con la organización celular (Beck y Cols., 1990) o con la interacción celular
mediante receptores de superficie (Edgar, 1989). Tampoco puede descartarse su
implicación en el crecimiento axonal (Baron van Evercooren y Cols., 1982; Wang y
Cols., 1992; Feltri y Cols., 2002). Los distintos tipos de colágenos caracterizados en los
espacios interlamelares de los corpúsculos de Pacini del mesenterio del gato (Pawson y
Cols., 2000) y en los espacios equivalentes de los corpúsculos de Meissner de Macaca
fuscata (Takahashi-Iwanaga y Shimoda, 2003) pueden desempeñar una función
estructural o de regulación como se ha demostrado en las células de Schwann de
nervios periféricos (Koopmans y Cols., 2009). Sin embargo, todavía se conoce menos
sobre los proteoglicanos que conforman la matriz extracelular de los corpúsculos
sensitivos (Dubový y Svízenská, 1993; Dubový y Bednárová, 1999; Sames y Cols., 2001).
Estas moléculas son especialmente importantes para las capacidades físico-químicas
de los corpúsculos sensitivos, como son la retención de agua, viscoelasticidad,
transmisión de la presión, etc. (Sames y Cols., 2001), aunque tampoco pueden
descartarse otras funciones.

2.4. Edad y tacto
2.4.1. Envejecimiento en el sistema somatosensorial del tacto
En la piel, la pérdida de sensibilidad es una consecuencia inevitable asociada a la edad,
debida fundamentalmente a un proceso de denervación y deterioro de las estructuras
nerviosas periféricas (Kelly y Cols., 2005; Chang y Cols., 2004; Panoutsopoulou y Cols.,
2009). Los estudios realizados hasta la fecha se han centrado preferentemente en las
terminaciones nerviosas libres (Wickremaratchi y Llewelyn, 2006; Taguchi y Cols.,
2010; Decorps y Cols., 2014). Sin embargo, poco se conoce acerca de los cambios que
tienen lugar en los corpúsculos sensitivos, principales estructuras encargadas del tacto,
como consecuencia de la edad.
En el sistema nervioso periférico, con el envejecimiento, se producen cambios
estructurales en las fibras nerviosas, menor densidad de las mismas, afectación de los
umbrales de detección y disminución de la velocidad de conducción nerviosa (Cerimele
y Cols., 1990; Stevens y Patterson, 1995; Verrillo y Cols., 2002; Perry, 2006). Los
cambios estructurales guardan relación con la reducción del diámetro del axón,
degeneración axónica, aumento de formas irregulares y aumento del número de
fibrillas de colágeno en el endoneuro y el perineuro (Chase y Cols., 1992). La
disminución en la velocidad de conducción nerviosa se debe a una mayor
vulnerabilidad de la vaina de mielina (Bouche y Cols., 1993; Verdú y Cols., 2000; Di
Iorio y Cols., 2006).
Los estudios funcionales han demostrado que con la edad se produce una alteración
en las habilidades para detectar vibraciones, lo que implica que se requiera mayor
amplitud de vibración para percibir la misma magnitud de sensación (Verrillo, 1979;
Gescheider y Cols., 1994; Goble y Cols., 1996; Verrillo y Cols., 2002; Perry, 2006).
Además, se ha comprobado que con el envejecimiento se deterioran la agudeza y la
discriminación espacial táctil (Stevens y Patterson, 1995; Leveque y Cols., 2000),
mientras que aumenta el umbral para el tacto fino (Bruce, 1980; Thornbury y
Mistretta, 1981).
2.4.2. Cambios edad-dependientes en los morfotipos de mecanorreceptores
cutáneos
La variabilidad en la metodología empleada así como la utilización de modelos
animales muy dispares hacen que los escasos resultados existentes sobre los cambios
cuantitativos y cualitativos de los corpúsculos sensitivos como consecuencia de la edad
sean imprecisos y dispares.
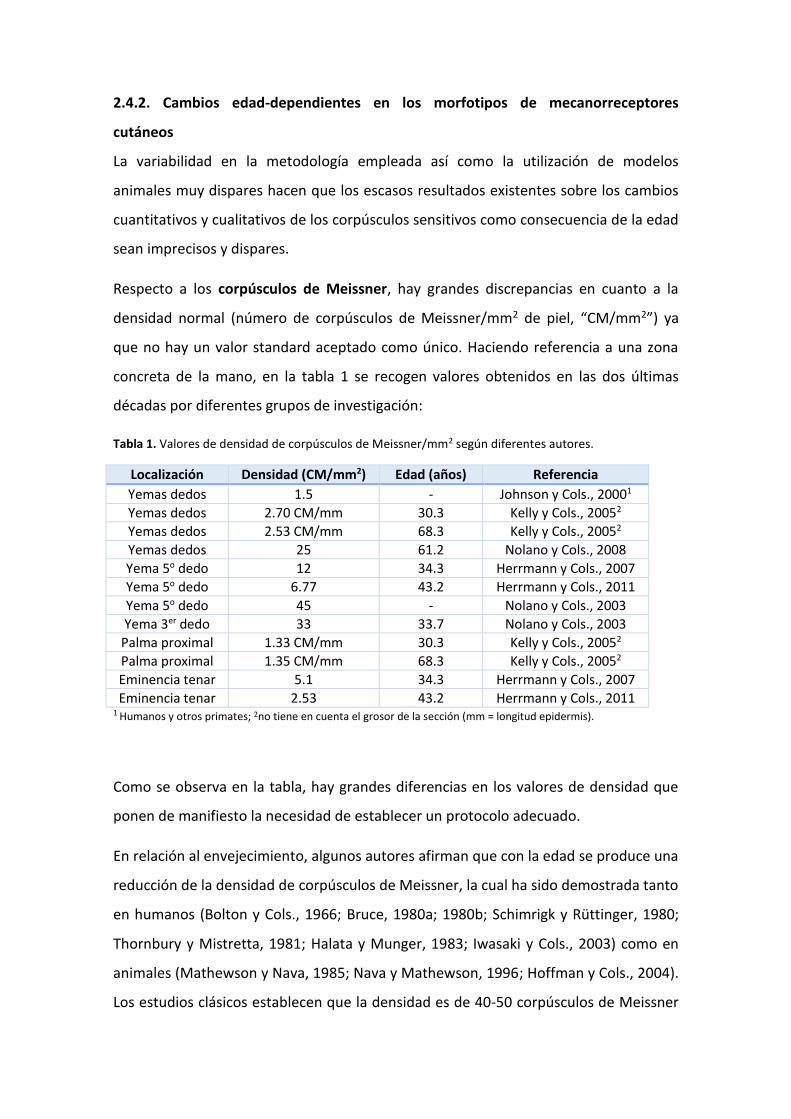
Respecto a los corpúsculos de Meissner, hay grandes discrepancias en cuanto a la
densidad normal (número de corpúsculos de Meissner/mm2 de piel, “CM/mm2”) ya
que no hay un valor standard aceptado como único. Haciendo referencia a una zona
concreta de la mano, en la tabla 1 se recogen valores obtenidos en las dos últimas
décadas por diferentes grupos de investigación:
Tabla 1. Valores de densidad de corpúsculos de Meissner/mm2 según diferentes autores.
Localización Densidad (CM/mm2) Edad (años) Referencia
Yemas dedos 1.5 - Johnson y Cols., 20001
Yemas dedos 2.70 CM/mm 30.3 Kelly y Cols., 20052
Yemas dedos 2.53 CM/mm 68.3 Kelly y Cols., 20052
Yemas dedos 25 61.2 Nolano y Cols., 2008
Yema 5o dedo 12 34.3 Herrmann y Cols., 2007
Yema 5o dedo 6.77 43.2 Herrmann y Cols., 2011
Yema 5o dedo 45 - Nolano y Cols., 2003
Yema 3er dedo 33 33.7 Nolano y Cols., 2003
Palma proximal 1.33 CM/mm 30.3 Kelly y Cols., 20052
Palma proximal 1.35 CM/mm 68.3 Kelly y Cols., 20052
Eminencia tenar 5.1 34.3 Herrmann y Cols., 2007
Eminencia tenar 2.53 43.2 Herrmann y Cols., 2011 1 Humanos y otros primates; 2no tiene en cuenta el grosor de la sección (mm = longitud epidermis).
Como se observa en la tabla, hay grandes diferencias en los valores de densidad que
ponen de manifiesto la necesidad de establecer un protocolo adecuado.
En relación al envejecimiento, algunos autores afirman que con la edad se produce una
reducción de la densidad de corpúsculos de Meissner, la cual ha sido demostrada tanto
en humanos (Bolton y Cols., 1966; Bruce, 1980a; 1980b; Schimrigk y Rüttinger, 1980;
Thornbury y Mistretta, 1981; Halata y Munger, 1983; Iwasaki y Cols., 2003) como en
animales (Mathewson y Nava, 1985; Nava y Mathewson, 1996; Hoffman y Cols., 2004).
Los estudios clásicos establecen que la densidad es de 40-50 corpúsculos de Meissner

por mm2 al final de la infancia (Thornbury y Mistretta, 1981), la cual disminuye
progresivamente hasta los 50 años llegando a 10-25 corpúsculos de Meissner por mm2
(Bolton y Cols., 1966; Bruce 1980a; Thornbury y Mistretta, 1981; Halata y Munger,
1983) y alcanza valores mínimos de 5-8 corpúsculos de Meissner por mm2 a partir de
los 60 años (Bolton y Cols., 1966; Bruce, 1980b).
Contrariamente, otros grupos de investigación no constatan un cambio edad-
dependiente en la densidad de corpúsculos sensitivos (Nolano y Cols., 2003; Kelly y
Cols., 2005).
Además de los cambios cuantitativos, se han descrito en detalle, principalmente en
modelos murinos, ciertos cambios morfológicos que afectan a la capacidad funcional
de los corpúsculos sensitivos: los corpúsculos de Meissner de menor tamaño
corresponden a los ratones más jóvenes, los ratones de mediana edad presentan los
más grandes y los ratones de edad más avanzada tienen corpúsculos desorganizados y
lobulados que decrecen en tamaño hasta atrofiarse por completo. En relación a los
componentes corpusculares, las lamelas de los corpúsculos se atenúan y disminuyen
en número mientras que el material de la lámina basal se duplica al aumentar las
fibrillas de colágeno; el axón se vuelve más complejo con ramificaciones hasta la
mediana edad, mientras que en la vejez se pierde la forma característica serpenteante.
Además se ha contabilizado un mayor número de papilas dérmicas desocupadas en la
piel de los ratones de mayor edad (Nava y Mathewson, 1996; Mathewson y Nava,
1985).
Muchas de estas variaciones cualitativas también se han encontrado en los
corpúsculos de Meissner humanos (Bolton y Cols., 1966; Bruce, 1980; Matsuoka y
Cols., 1983; Iwasaki y Cols., 2003): distribución más irregular, con formas lobuladas u
alargadas y alejados de la epidermis u orientados oblicuamente en la papila, mayor
tamaño con la edad pero reducción a partir de los 70 años debido a cambios en el
tamaño de las células lamelares y formas retorcidas a partir de los 60 años.
Los corpúsculos de Pacini de la piel digital humana también sufren un proceso de
degeneración y atrofia con la edad, así como una disminución en número (Cauna y
Mannan, 1958; Gescheider y Cols., 1994). El tamaño del corpúsculo de Pacini es de

500-700 µm al nacer y se incrementa hasta alcanzar los 3-4 mm; pero a partir de los 70
años, el corpúsculo disminuye en dimensión y se vuelve más irregular (Cauna y
Mannan, 1958). Además, se ha demostrado que la sensibilidad vibrotáctil que implica a
los corpúsculos de Pacini se reduce con la edad (Verrillo, 1979).
En el caso de las células de Merkel, también se produce un descenso en número con el
envejecimiento tanto en humanos como en modelos murinos (Moll y Cols., 1984;
Fundin y Cols., 1997). Nolano y Cols. (2003) determinan que hay una densidad de 4
células de Merkel por mm2 de piel en individuos con edad media de 33.7 años, y
además, especifica que el número de corpúsculos de Meissner es 5 veces mayor que el
de células de Merkel en la piel digital humana. Otros resultados muy diferentes
establecen que hay una densidad media de 1700 células de Merkel por mm2 en crestas
glandulares de piel plantar de fetos humanos de 18-24 semanas, la cual se reduce
progresivamente en recién nacido y en adulto (Moll y Cols., 1984). De igual forma, Kim
y Holbrook (1995) establecen que en la epidermis palmar de fetos humanos, la
densidad llega hasta 1400 células de Merkel por mm2 de piel a las 8 semanas de edad
gestacional, momento en el cual el número empieza a caer.

3. HIPÓTESIS DE TRABAJO Y OBJETIVOS

En la presente tesis doctoral se han planteado los siguientes objetivos:
El objetivo general es contribuir al conocimiento de los corpúsculos sensitivos de la
piel glabra humana aportando nuevos datos sobre el origen de las células que los
componen, estudiando los proteoglicanos y glicosaminoglicanos de su matriz
extracelular y analizando las variaciones morfológicas, cuantitativas y cualitativas que
tienen lugar como consecuencia de proceso de envejecimiento.
Los objetivos específicos son los siguientes:
1. Investigar la presencia y distribución de antígenos específicos de células
endoneurales (CD34) o perineurales (Glut1 y EMA) en los corpúsculos de Pacini de
la piel digital humana con el fin de determinar el origen de las células del núcleo
externo y de la cápsula de los mismos.
2. Analizar la presencia de condroitín sulfato en la matriz extracelular de los
corpúsculos sensitivos humanos determinando su localización en los corpúsculos
sensitivos mediante doble inmunofluorescencia.
3. Analizar la presencia de heparán sulfato en la matriz extracelular de los
corpúsculos sensitivos humanos determinando su localización en los corpúsculos
sensitivos mediante doble inmunofluorescencia.
4. Identificar qué miembros de la familia SLRP se encuentran presentes en los
corpúsculos de Pacini humanos y analizar mediante inmunofluorescencia asociada
a microscopía confocal su distribución en los mismos.
5. Determinar los cambios cuantitativos, cualitativos y morfológicos que aparecen en
los mecanorreceptores cutáneos (corpúsculos de Meissner y células de Merkel) de
la piel digital humana como consecuencia del envejecimiento.

4. MATERIAL Y TÉCNICAS
4.1. Material
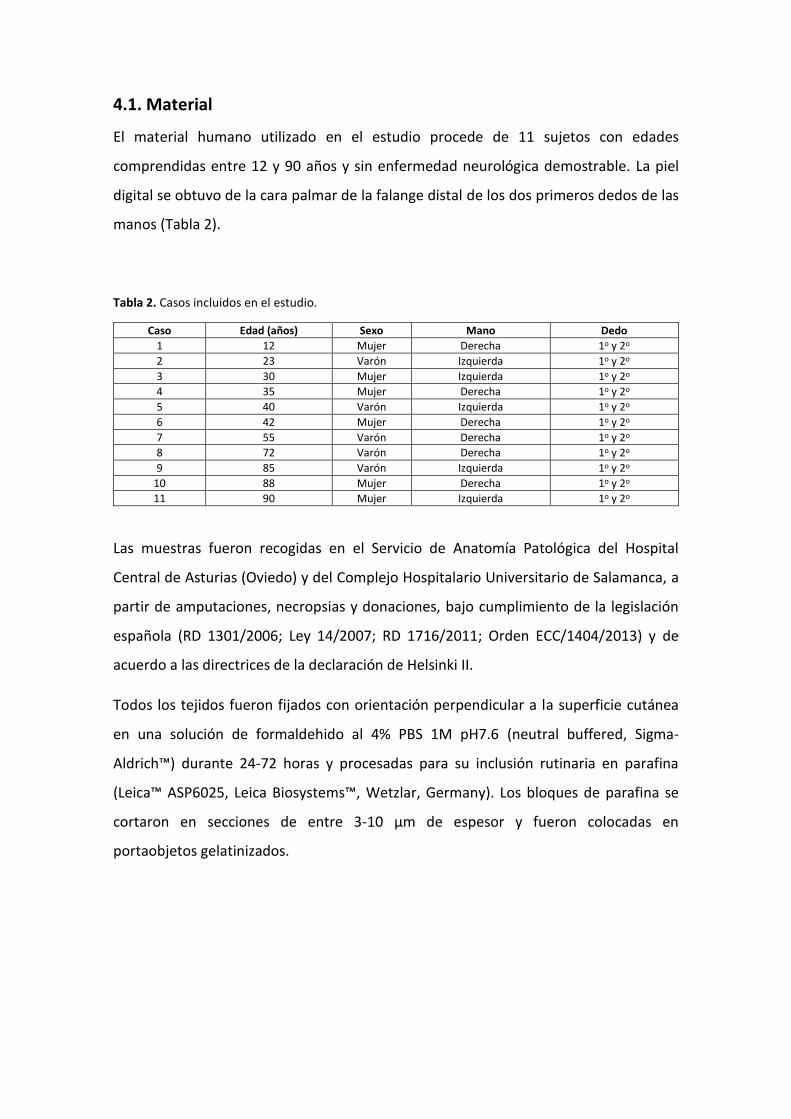
El material humano utilizado en el estudio procede de 11 sujetos con edades
comprendidas entre 12 y 90 años y sin enfermedad neurológica demostrable. La piel
digital se obtuvo de la cara palmar de la falange distal de los dos primeros dedos de las
manos (Tabla 2).
Tabla 2. Casos incluidos en el estudio.
Caso Edad (años) Sexo Mano Dedo
1 12 Mujer Derecha 1o y 2o
2 23 Varón Izquierda 1o y 2o
3 30 Mujer Izquierda 1o y 2o
4 35 Mujer Derecha 1o y 2o
5 40 Varón Izquierda 1o y 2o
6 42 Mujer Derecha 1o y 2o
7 55 Varón Derecha 1o y 2o
8 72 Varón Derecha 1o y 2o
9 85 Varón Izquierda 1o y 2o
10 88 Mujer Derecha 1o y 2o
11 90 Mujer Izquierda 1o y 2o
Las muestras fueron recogidas en el Servicio de Anatomía Patológica del Hospital
Central de Asturias (Oviedo) y del Complejo Hospitalario Universitario de Salamanca, a
partir de amputaciones, necropsias y donaciones, bajo cumplimiento de la legislación
española (RD 1301/2006; Ley 14/2007; RD 1716/2011; Orden ECC/1404/2013) y de
acuerdo a las directrices de la declaración de Helsinki II.
Todos los tejidos fueron fijados con orientación perpendicular a la superficie cutánea
en una solución de formaldehido al 4% PBS 1M pH7.6 (neutral buffered, Sigma-
Aldrich™) durante 24-72 horas y procesadas para su inclusión rutinaria en parafina
(Leica™ ASP6025, Leica Biosystems™, Wetzlar, Germany). Los bloques de parafina se
cortaron en secciones de entre 3-10 µm de espesor y fueron colocadas en
portaobjetos gelatinizados.

4.2. Técnicas
4.2.1. Tinción hematoxilina-eosina
Se realizaron manualmente tinciones con hematoxilina-eosina para identificar las
secciones de tejido que contenían los corpúsculos sensitivos de interés. El protocolo
empleado fue el siguiente: desparafinado en xilol y rehidratación con alcoholes de
concentración decreciente hasta llegar a agua corriente; baño en hematoxilina durante
10 segundos y lavado en agua corriente; baño en eosina durante 60 segundos y lavado
con agua destilada; deshidratación con alcoholes de concentración creciente,
diafanización en xilol y montaje del cubreobjetos con Entellan®.
4.2.2. Inmunohistoquímica simple indirecta
Las secciones de tejido se desparafinaron con xilol y se rehidrataron mediante una
batería de alcoholes de concentración decreciente hasta llegar al agua. A continuación,
para bloquear la actividad peroxidasa endógena, se trataron las muestras con H2O2 al
3% durante 10 minutos, seguido de un lavado en tampón PBS 1M a pH 7.6 con Tween-
20 al 0.5% (PBS-T) para permeabilizar las membranas celulares. Después de ello, se
bloquearon las uniones inespecíficas con albumina bovina (BSA) al 3% durante 30
minutos. Posteriormente, las muestras se incubaron toda la noche a 4oC y en cámara
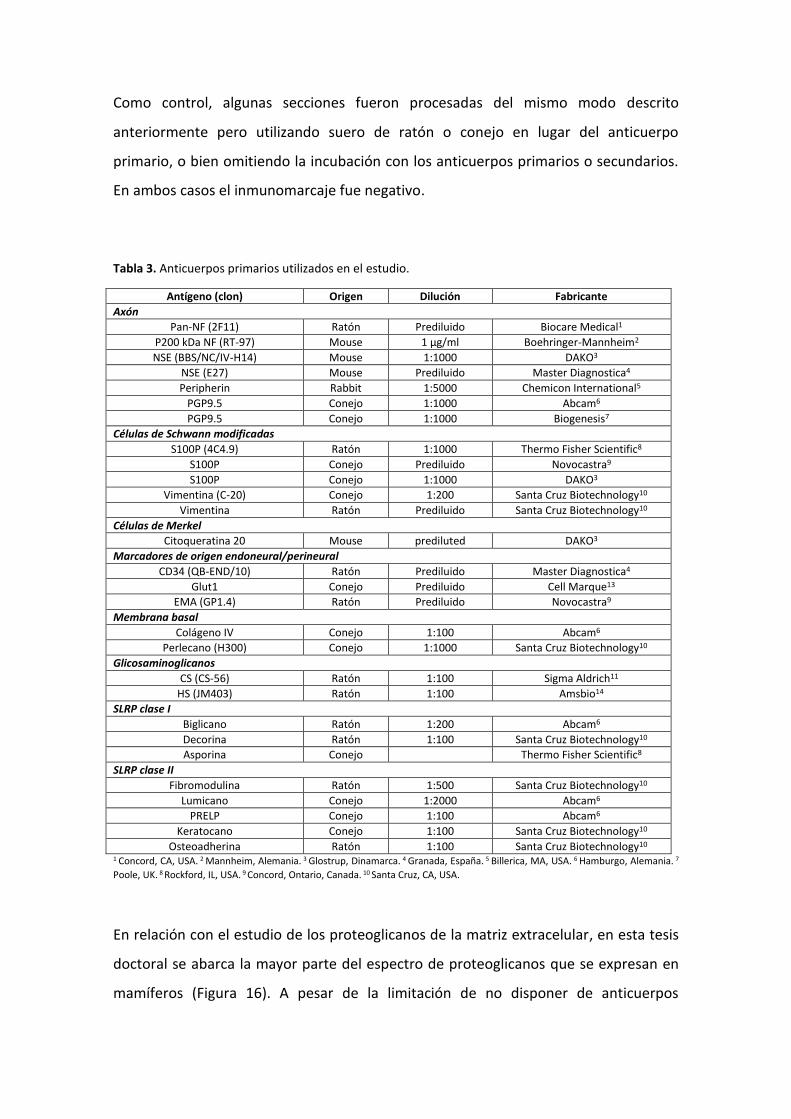
húmeda con los anticuerpos primarios según cada estudio (Tabla 3).
Tras la incubación, las secciones se lavaron en PBS-T durante 15 minutos y se
incubaron a temperatura ambiente con el anticuerpo secundario correspondiente
conjugado con peroxidasa (Dako EnVision labelled polymer-HRP anti-conejo IgG o de
IgG anti-ratón) durante 90 minutos. Tras un lavado en PBS-T, se reveló la
inmunorreacción con una solución de 3-3’ diaminobencidina (Leica Bond™ Polymer
Refine Detection Kit, Leica Biosystems™). Las secciones se contrastaron con
hematoxilina, se lavaron en agua, se deshidrataron con alcoholes de concentración
creciente, se diafanizaron en xilol y se montaron con Entellan®. Por último, se
fotografiaron en un microscopio óptico Nikon Eclipse® 80i acoplado a una cámara
Nokia® DS-5M.
Como control, algunas secciones fueron procesadas del mismo modo descrito
anteriormente pero utilizando suero de ratón o conejo en lugar del anticuerpo
primario, o bien omitiendo la incubación con los anticuerpos primarios o secundarios.
En ambos casos el inmunomarcaje fue negativo.
Tabla 3. Anticuerpos primarios utilizados en el estudio.
Antígeno (clon) Origen Dilución Fabricante
Axón
Pan-NF (2F11) Ratón Prediluido Biocare Medical1
P200 kDa NF (RT-97) Mouse 1 μg/ml Boehringer-Mannheim2
NSE (BBS/NC/IV-H14) Mouse 1:1000 DAKO3
NSE (E27) Mouse Prediluido Master Diagnostica4
Peripherin Rabbit 1:5000 Chemicon International5
PGP9.5 Conejo 1:1000 Abcam6
PGP9.5 Conejo 1:1000 Biogenesis7
Células de Schwann modificadas
S100P (4C4.9) Ratón 1:1000 Thermo Fisher Scientific8
S100P Conejo Prediluido Novocastra9
S100P Conejo 1:1000 DAKO3
Vimentina (C-20) Conejo 1:200 Santa Cruz Biotechnology10
Vimentina Ratón Prediluido Santa Cruz Biotechnology10
Células de Merkel
Citoqueratina 20 Mouse prediluted DAKO3
Marcadores de origen endoneural/perineural
CD34 (QB-END/10) Ratón Prediluido Master Diagnostica4
Glut1 Conejo Prediluido Cell Marque13
EMA (GP1.4) Ratón Prediluido Novocastra9
Membrana basal
Colágeno IV Conejo 1:100 Abcam6
Perlecano (H300) Conejo 1:1000 Santa Cruz Biotechnology10
Glicosaminoglicanos
CS (CS-56) Ratón 1:100 Sigma Aldrich11
HS (JM403) Ratón 1:100 Amsbio14
SLRP clase I
Biglicano Ratón 1:200 Abcam6
Decorina Ratón 1:100 Santa Cruz Biotechnology10
Asporina Conejo Thermo Fisher Scientific8
SLRP clase II
Fibromodulina Ratón 1:500 Santa Cruz Biotechnology10
Lumicano Conejo 1:2000 Abcam6
PRELP Conejo 1:100 Abcam6
Keratocano Conejo 1:100 Santa Cruz Biotechnology10
Osteoadherina Ratón 1:100 Santa Cruz Biotechnology10
1 Concord, CA, USA. 2 Mannheim, Alemania. 3 Glostrup, Dinamarca. 4 Granada, España. 5 Billerica, MA, USA. 6 Hamburgo, Alemania. 7
Poole, UK. 8 Rockford, IL, USA. 9 Concord, Ontario, Canada. 10 Santa Cruz, CA, USA.
En relación con el estudio de los proteoglicanos de la matriz extracelular, en esta tesis
doctoral se abarca la mayor parte del espectro de proteoglicanos que se expresan en
mamíferos (Figura 16). A pesar de la limitación de no disponer de anticuerpos

específicos para todos los proteoglicanos, se ha podido realizar un estudio lo
suficientemente exhaustivo como para llegar a analizar un gran número de las
moléculas totales: proteoglicanos de heparan sulfato, proteoglicanos de condroitín
sulfato y proteoglicanos de la familia SLRP de las clases I y II.
Figura 16. Proteoglicanos analizados de forma general o específica y proteoglicanos no analizados.
4.2.3. Doble inmunohistoquímica indirecta
Esta técnica fue empleada exclusivamente para la inmunodetección de las proteínas
Glut1/EMA y CD34 correspondiente al primer resultado de la tesis (ver apartado 5.1).
Las secciones fueron procesadas con el sistema de tinción automático Leica Bond™
Max (Leica Biosystems™) ubicado en el Servicio de Anatomía Patológica del Complejo
Hospitalario Universitario de Salamanca. Una vez visualizada la inmunorreacción
correspondiente al primer antígeno con diaminobencidina como cromógeno (color
marrón) (Leica Bond™ Polymer Refine Detection Kit, Leica Biosystems™), se volvió a
realizar un segundo proceso de inmunohistoquímica para el segundo antígeno. En esta
ocasión, se utilizó un método de revelado diferente: el anticuerpo secundario está
conjugado con fosfatasa alcalina y el cromógeno utilizado es Fast Red (Leica Bond™
Polymer Red Refine Detection Kit, Leica Biosystems™), de forma que la segunda
inmunorreacción fue visualizada en color rojo.

4.2.4. Doble inmunofluorescencia indirecta con microscopía confocal
Las muestras de 10 µm se desparafinaron y rehidrataron como se describe en el
apartado de la inmunohistoquímica simple. Seguidamente, se lavaron en PBS-T
durante 20 minutos y se incubaron durante toda la noche a 4oC en cámara húmeda
con una mezcla 1:1 de dos anticuerpos primarios para la detección simultanea de dos
antígenos. Posteriormente las preparaciones se lavaron con PBS-T durante 30 minutos
y se incubaron con los anticuerpos secundarios durante 90 minutos: en primer lugar
IgG de cabra anti-conejo conjugado con Alexa Fluor 488 (1:1000; Serotec™, Oxford,
Reino Unido) y a continuación, con IgG de asno anti-ratón conjugado con Cy3 (1:50;
Jackson-ImmunoResearch™, Baltimore, MD, USA). Ambas incubaciones se realizaron
en cámara húmeda, en oscuridad y a temperatura ambiente. Se realizó un lavado con
PBS-T entre ambas incubaciones y luego se montó la preparación con DAPI (4',6-
diamino-2-fenilindol; 10 ng/ml) diluido en medio de montaje Fluoromount-G
(Southern-Biotech, Alabama, USA) para contrastar los núcleos (color azul).
En el caso de la doble inmunofluorescencia en las que ambos anticuerpos primarios
eran monoclonales de ratón, primero las muestras se incubaron con uno de los dos
anticuerpos contra el primer epítopo. Al día siguiente, se incubaron con IgG de conejo
anti-ratón (Novolink™ Polymer Detection Systems, Leica Biosystems™) durante 90
minutos y, tras un lavado en PBS-T, con anticuerpo secundario IgG de cabra anti-
conejo conjugado con Alexa Fluor 488 durante otros 90 minutos. Posteriormente, las
secciones fueron incubadas con el segundo anticuerpo primario para el otro epítopo
durante toda la noche. El tercer día, las preparaciones se incubaron con anticuerpo
secundario IgG de asno anti-ratón conjugado con Cy3 durante 90 minutos y se
montaron con DAPI diluido en medio de montaje Fluoromount-G.
La inmunofluorescencia fue detectada mediante un microscopio automático de
fluorescencia Leica DMR-XA acoplado a un software de captación de fluorescencia
Leica Confocal Software v2.5 (Leica Microsystems, Heidelberg GmbH, Germany),
ubicado en el Servicio de Procesado de Imágenes de la Universidad de Oviedo. Los
controles específicos de la reacción fueron ejecutados de la misma forma que para la
inmunohistoquímica simple. Además, se llevaron a cabo controles adicionales con el

fin de confirmar la ausencia de procesos de autofluorescencia del tejido o producidos
por el proceso de fijación omitiendo ambos anticuerpos.
4.2.5. Análisis cuantitativo
Se realizaron estudios cuantitativos para los corpúsculos de Meissner y las células de
Merkel en secciones independientes, pero no para los corpúsculos de Pacini debido a
su distribución irregular en la dermis y a su gran tamaño.
Para detectar los corpúsculos de Meissner y las células de Merkel, se realizó
inmunohistoquímica simple para la proteína S100 y CK20 respectivamente, en varios
cortes de 5 µm de espesor, separados entre sí una distancia mínima de 100 µm. Una
vez visualizada la inmunorreacción y montado el cubreobjetos, las secciones fueron
escaneadas mediante el escáner SCN400F (Leica Biosystems™) ubicado en el Instituto
Universitario de Oncología del Principado de Asturias (IUOPA) y procesadas
informáticamente con el programa SlidePath Gateway LAN (Leica Biosystems™).
Para determinar la densidad, el método cuantitativo aplicado fue el utilizado
previamente por Verendeev y Cols. (2015) en un estudio comparativo sobre los
corpúsculos de Meissner de las yemas de los dedos de diferentes primates.
En cada sección, se contabilizó el número de corpúsculos de Meissner o de células de
Merkel y se midió la longitud de la epidermis de la piel tranzando una línea sobre la
misma con una de las herramientas del programa informático anterior (Figura 16).
Para estimar la densidad de forma adecuada, hay que tener en cuenta que en una
sección de tejido podemos estar observando células de Merkel completas pero
también incompletas (el diámetro de una célula de Merkel es de 5-15 µm y el grosor
de la muestra es de 5 µm). Esto se conoce como el efecto del grosor de la sección en
relación al diámetro de la célula (Figura 17). En el caso de los corpúsculos de Meissner,
debido a su mayor tamaño, observamos exclusivamente corpúsculos incompletos
(diámetro de 20-60 µm y grosor de la sección de 5 µm).

Figura 16. Esquema representativo del método empleado para la cuantificación de corpúsculos
sensitivos.
Figura 17. Efecto del grosor de la muestra en relación al diámetro de la célula. Corrección de
Abercrombie (modificado de Técnicas de análisis de imagen: Aplicaciones en Biología).
Por lo que el número obtenido directamente de la cuantificación deber ser corregido
mediante un factor matemático para evitar una sobreestimación de la densidad. Dicho
factor de corrección fue formulado por Abercrombie en 1946 (Verendeev y Cols.,

2015) para estimar poblaciones nucleares a partir de secciones cortadas con
micrótomo y su fórmula es la siguiente:
𝑁 =𝑛 ∗ 𝑇
𝑇 + 𝐻
N = valor medio corregido; n = valor medio observado; T = espesor medio de las
secciones cuantificadas; H = diámetro medio de las células de Merkel o de los
corpúsculos de Meissner cuantificados en las secciones.
El diámetro de las células de Merkel o de los corpúsculos de Meissner fue medido con
la misma herramienta del software SlidePath Gateway LAN, trazando una línea recta
sobre el eje transversal de la célula de Merkel o el corpúsculo de Meissner
respectivamente.
La densidad media (nº de estructuras sensitivas/mm2 de piel) se calculó dividiendo el
valor medio corregido (N) entre el área superficial de piel (mm2), la cual se obtuvo a
partir del grosor medio de las secciones (5 µm = 0,005 mm) y la longitud media de la
piel de las muestras (X mm):
𝐷𝑒𝑛𝑠𝑖𝑑𝑎𝑑 =𝑁ú𝑚𝑒𝑟𝑜 𝑒𝑠𝑡𝑟𝑢𝑐𝑡𝑢𝑟𝑎𝑠 𝑠𝑒𝑛𝑠𝑖𝑡𝑖𝑣𝑎𝑠
Á𝑟𝑒𝑎 𝑑𝑒 𝑝𝑖𝑒𝑙
Para el estudio correspondiente al envejecimiento, los individuos fueron clasificados
en tres grupos de edad: a) [20-39 años], b) [40-59 años] y c) [60-90 años]. Se
determinó la densidad media para cada grupo y se extrapolaron los datos
construyendo las gráficas correspondientes mediante el programa Excel.
El desarrollo del proceso de cuantificación, aplicado al presente trabajo, es susceptible
de ser utilizado en la evaluación de los cambios en la inervación cutánea debidos a
neuropatías periféricas y centrales, lo cual dio lugar al capítulo de un libro que se
incluye en esta Tesis doctoral.
4.2.6. Análisis estadístico
Las diferencias en la densidad de corpúsculos de Meissner y células de Merkel entre
los tres grupos de edad pre-establecidos se valoró mediantes el test H de Kruskal-

Wallis H test, y valores de p ≤0.05 se consideraron como estadísticamente
significativos (marcados en las figuras como *p<0.05, **p<0.01).

5. RESULTADOS
Esta parte del capítulo se corresponde con el artículo:
García Piqueras, Jorge et al. Endoneurial-CD34 positive cells define an intermediate layer in
human digital Pacinian corpuscles. En Annals of Anatomy, 211, p. 55-60 (2017);
doi:10.1016/j.aanat.2017.01.006
Debido a la política de autoarchivo de la publicación la versión de la editorial está disponible, únicamente para usuarios con suscripción de pago a la revista, en el siguiente enlace: http://dx.doi.org/10.1016/j.aanat.2017.01.006
Información facilitada por equipo RUO
Esta parte del capítulo se corresponde con el artículo:
García Piqueras, Jorge et al. Chondroitin Sulfate in Human Cutaneous Meissner and Pacinian
Sensory Corpuscles. En Anatomical Record, 302, p. 325-331 (2018); doi:10.1002/ar.23951
Debido a la política de autoarchivo de la publicación la versión de la editorial está disponible, únicamente para usuarios con suscripción de pago a la revista, en el siguiente enlace: http://dx.doi.org/10.1002/ar.23951
Información facilitada por equipo RUO

For Peer ReviewHEPARAN SULFATE IS ASSOCIATED TO THE BASEMENT MEMBRANE IN HUMAN CUTANEOUS MEISSNER’S AND
PACINIAN CORPUSCLES
Journal: Anatomical Record
Manuscript ID Draft
Wiley - Manuscript type: Full Length Article
Date Submitted by the Author: n/a
Complete List of Authors: García-Piqueras, Jorge; Universidad de Oviedo , Morfología y Biología Celular García-Suarez, Olivia; Universidad de Oviedo, Departamento de Morfología y Biología CelularGarcia-Mesa , Yolanda ; Universidad de Oviedo , Morfología y Biología Celular García-Fernandez, Beatriz; Universidad de OviedoQuiros , Luis; Universidad de Oviedo , Biología Funcional Cobo, Juan ; Universidad de Oviedo , Morfología y Biología Celular Martín-Biedma, Benjamin ; Universidade de Santiago de Compostela, Departamento de Cirugía y Especialidades Médico-QuirúrgicasFeito , Jorge; Complejo Hostipalatio Salamanca , Anatomia Patologica Vega, Jose; Universidad de Oviedo , Morfología y Biología Celular
Keywords: cutaneous sensory corpuscles, basement membrane, extracellular matrix, heparan sulfate proteoglycans, human
John Wiley & Sons, Inc.
The Anatomical Record

For Peer ReviewHEPARAN SULFATE IS ASSOCIATED TO THE BASEMENT
MEMBRANE IN HUMAN CUTANEOUS MEISSNER’S AND
PACINIAN CORPUSCLES
J. García-Piqueras1*, O. García-Suárez1*, Y. García-Mesa1, B. García2, L.
Quirós2,
J.L. Cobo1,3, B. Martín-Biedma4, J. Feito5, J.A. Vega1,6
1Departamento de Morfología y Biología Celular, Grupo SINPOS Universidad
de Oviedo, Spain2Departamento de Biología Funcional, Área de Microbiología, Universidad de
Oviedo, Spain 3Servicio de Cirugía Maxilofacial, Hospital Universitario Central de Asturias,
Oviedo, Spain 4Departamento de Cirugía y Especialidades Médico-Quirúrgicas, Universidad
de Santiago de Compostela, Santiago de Compostela, Spain5Servicio de Anatomía Patológica, Hospital Universitario de Salamanca,
Salamanca, Spain
6Facultad de Ciencias de la Salud, Universidad Autónoma de Chile, Chile
*Both authors contributed equally to this paper
Page 1 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Short title: Extracellular matrix in Pacinian corpuscles
Address for correspondence
José A. Vega, M.D., Ph.D. Departamento de Morfología y Biología Celular Facultad de Medicina y Ciencias de la Salud – Universidad de Oviedo Avd. JualianClaveria, 6, Planta 9ª 33006 OVIEDO – Spain Telf. (0034) 985104097 E-mail: [email protected]
Abstract.- Heparan sulfate proteoglycans are pericellular/cellular-surface
molecules involved in somatosensory axon guidance in the peripheral nervous
system. However, the distribution of heparan sulfate proteoglycans in the
extracellular matrix of human cutaneous sensory corpuscles is unknown.
Immunohistochemistry and immunofluorescence were performed in order to
define the localization of heparan sulfate proteoglycans in Meissner’s and
Pacinian corpuscles using an anti-heparan sulfate antibody together with anti-
type IV collagen, anti-S100 protein, anti-PGP9.5 and anti-CD34 antibodies (to
immunolabel basement membranes, Schwann cells, axon and the endoneurial
intermediate layer of the Pacinian corpuscles, respectively). In Meissner’s
Page 2 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
corpuscles heparan sulfate proteoglycans were found co-localized with type IV
collagen. In Pacinian corpuscles heparan sulfate proteoglycans co-localized
with type IV collagen of the outer core lamellae and capsule, but not in the inner
core or the intermediate layer. Present results strongly suggest that heparan
sulfate proteoglycans are associated to basement membranes of the lamellar
cells in Meissner’s corpuscles, and the complex outer core-capsule in Pacinian
corpuscles. The functional significance of these results, if any, remains to be
elucidated.
Key words.- cutaneous sensory corpuscles, basement membrane, extracellular
matrix, heparan sulfate proteoglycans, human
Introduction
Page 3 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Heparan Sulfate (HS) proteoglycans (HSPGs) are basic constituents of the
extracellular matrix (ECM). Structurally they are glycoproteins formed by a
variable number of HS chains attached covalently to a core protein localized at
the cell surface (i.e., syndecan or glypican families) or in the pericellular basal
membrane (BM; i.e., agrin, perlecan, type XV collagen and type XVIII) (Sarrazin
et al., 2011; Iozzo and Schaefer, 2015) that act principally as structural scaffold,
but also are involved in other cellular processes such as differentiation,
proliferation or survival (Schwarzbauer, 1999; LeBleu et al., 2007). The BM
architecture is the result of polymeric assemblies constituted by two principal
components that are laminins and type IV collagen, and other molecules as
nidogen glycoproteins, HSPGs, integrins or dystroglycan (Sasaki et al., 2004;
Yurchenco and Patton, 2009).
HSPGs are essential molecules for nervous system development and axonal
regeneration (Bovolenta and Fernaud-Espinosa, 2000; Yamaguchi, 2001;
Poulain and Yost, 2015), and are required for somatosensitive axons guidance
to the skin (Wang et al., 2012). Consistently, HSPGs have been detected in the
peripheral nerves and sensory ganglia (Ma et al., 1994; Halfter et al., 1997,
Page 4 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
1998; Chernousov et al., 1998). Nevertheless, the occurrence and distribution
of HSPGs in human cutaneous mechanoreceptors, the most peripheral
segment of the somatosensory system, have been never investigated.
Meissner’s and Pacinian sensory corpuscles are low-threshold
mechanoreceptors that consist of one axon surrounded by Schwann-related
cells and in the case of the Pacinian corpuscles, by endoneurial- and
perineurial-related cells (Munger and Idé, 1988; Zelená, 1994). Each corpuscle
cell-type is continuous with its counterpart of the nerve fibre supplying the
corpuscle, thus sharing their immunohistochemical features (Vega et al., 1996,
2009; García-Piqueras et al., 2017). Despite the scarce knowledge of the
protein composition of ECM in sensory corpuscles (Vega et al., 1995; Dubový
and Bednárová, 1999; Sames et al., 2001), a BM has been described and
partially characterized in cutaneous sensory corpuscles (Malinovský and Pac,
1982; Malinovský, 1986; Ide et al., 1987; Ide and Hayashi, 1987; Munger and
Ide, 1988; Chouchkov et al., 2003). In particular a BM was found underlying the
surface of the lamellar cells of Meissner’s corpuscles and in the inner core,
intermediate layer, outer core and capsule cells of Pacinian corpuscles
Page 5 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
(Malinovský, 1986; Vega et al., 1995). Nevertheless, other studies have shown
that BM is absent or discontinuous in the inner core and the intermediate layer
(Ide et al., 1987; Ide and Hayashi, 1987; Munger and Ide, 1988). Of particular
interest is the fact that some BM components, and other ECM molecules could
take part in the mechanotransduction process in sensory neurons (Wetzel et al.,
2007; Hu and Chiang, 2010; Chiang et al., 2011).
Since the occurrence and distribution of HSPGs in sensory corpuscles are
known only partially, we have designed the present research on human
cutaneous Meissner’s and Pacinian corpuscles. The study was aimed to
analyze the composition of ECM in the sensory corpuscles, in particular its
presence in BM.
Material and methods
Skin samples were obtained from the palmar aspect of the distal phalanx of the
first and second fingers of both hands (n = 11). Human samples were collected
from autopsies of non-demonstrable neurological disease subjects (age range
20 to 55 years) at the Departments of Pathology of the Hospital Universitario
Central de Asturias (Oviedo, Spain) and Complejo Hospitalario Universitario de
Page 6 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Salamanca (Salamanca, Spain). These materials were obtained according to
the guidelines of the Helsinki Declaration (World Medical Association, 2013)
and in compliance with Spanish Laws. Tissue samples were fixed in 10%
formaldehyde in 1 M phosphate buffer saline (pH 7.4) for 24 h, dehydrated and
routinely embedded in paraffin. The pieces were cut into 7 μm thick sections
perpendicular to the skin surface and mounted on gelatine-coated microscope
slides. To confirm the presence of Pacinian and Meissner’s corpuscles in the
skin samples, several random sections were staining with hematoxylin and
eosin.
Immunohistochemistry.- Deparaffinized and rehydrated 7-μm thick sections
were processed for immunohistochemistry using EnVision antibody complex
detection kit (Dako, Copenhagen, Denmark) following manufacturer’s
instructions. The endogenous peroxidase activity and the non-specific binding
were blocked with 3% H2O2 and 10% foetal calf serum respectively for 30 min
each one. After that, the sections were incubated overnight in a humid chamber
at 4° C with the following primary antibodies: mouse monoclonal anti-HS
antibody (1:100; clone JM-403, AMSBIO; Abingdon, UK); rabbit polyclonal anti-
Page 7 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
type IV collagen (1:100; Abcam; Hamburg, Germany); mouse monoclonal anti-
CD34 (prediluted; clone QB-END/10, Master Diagnóstica™; Granada, Spain).
Moreover, rabbit polyclonal antibodies against S100 protein (1:1000; DAKO;
Glostrup, Denmark) and PGP9.5 (1:500; abcam; Cambridge, UK) were used to
selectively label the lamellar cells and inner-core cells, and the axon,
respectively. After incubation, the sections were rinsed in TBS-T and incubated
with anti-rabbit and anti-mouse EnVision system-labelled polymer
(DakoCytomation) for 60 minutes. After rising again, the immunoreaction was
visualized using 3-3’-diaminobenzidine as a chromogen. Finally, sections were
counterstained with hematoxylin-eosin to ascertain structural details,
dehydrated and mounted with Entellan (Merck, Dramstadt, Germany). Controls
were made by omitting the primary antibody or replacing it with a non-immune
rabbit or mouse serum. No immunostaining was observed (data not shown).
Double immunofluorescence.- Sections were processed as described before
but the primary antibodies were incubated simultaneously overnight at 4oC in a
humid chamber with a 1:1 v/v mixture of anti-HS antibody with anti-S100P
antibody, anti-HS antibody with anti-PGP9.5 antibody (1:300; abcam;
Page 8 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Cambridge, UK) and anti-HS antibody with anti-type IV collagen antibody. After
rising with TBS, sections were incubated with Alexa fluor 488-conjugated goat
anti-rabbit IgG (1:1000; Serotec, Oxford, UK) for 90 minutes, then rinsed again
and incubated other 90 minutes with Cy3-conjugated donkey anti-mouse
antibody (1:50; Jackson-ImmunoResearch, Baltimore, MD, USA).
In the case of double immunodetection of HS and CD34, sections were
incubated overnight in a humid chamber at 4o C with primary anti-CD34
antibody (to label the intermediate layer; García-Piqueras et al., 2017, 2018).
After a TBS-rinsing, sections were incubated with Post-Primary (Novolink
Polymer Detection System®, Leica-Novocastra Biosystems, Newclastle, UK) in
a humid chamber at room temperature for 90 minutes. Post-Primary is a
prediluted rabbit IgG which recognizes mouse antibodies. After that, sections
were incubated for 90 minutes at room temperature with Alexa fluor 488-
conjugated goat anti-rabbit IgG as described above. Then, sections were
incubated again overnight at 4o C with mouse anti-HS antibody, followed by
incubation with Cy3-conjugated donkey anti-mouse antibody for 90 minutes.
Finally, to label nuclei, sections were counterstained with DAPI (10 ng/ml) and
Page 9 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
mounted with Fluoromount Gold, as previously. Immunofluorescence was
detected using a Leica TCS AOBS SP8 X confocal microscopy (Leica
Microsystem GmbH, Heidelberg, Germany; Servicios Científico-Técnicos de la
Universidad de Oviedo, Oviedo, Spain) and images processed by Leica
Application Suite X software v1.8.1.13759 (Leica Microsystem Gmbh,
Heidelberg, Germany). Representative samples for control purposes were
carried out as described before (data not shown).
Results
Human Meissner’s and Pacinian corpuscles display immunoreactivity for
HSPGs. Both types of sensory corpuscles analysed display immunoreactivity
for HSPGs. In Meissner’s corpuscles, positivity was observed in the surface of
the lamellar cells forming parallel lines which presumably corresponds to a BM
pattern (Figs. 1a-c). In Pacinian corpuscles, HSPGs immunoreactivity occurred
in the outer core and the capsule being especially intense at the innermost
lamellae of the outer core (Figure 2a). No HSPGs immunostaining was
observed in the inner core.
Page 10 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
In order to establish the precise location of HSPGs in Meissner’s and Pacinian
corpuscles, double immunofluorescence using different combinations of anti-HS
with anti-S100P or anti-PGP9.5 antibodies (to label lamellar/inner core cells and
axon, respectively) was performed. No co-localization of HSPGs and S100P
was observed in Meissner’s corpuscles thus supporting the idea that HSPGs
are localized in the pericellular BM of the lamellar cells (Figs. 1d-f).
Nevertheless, in some areas there was partial co-localization HSPGs and
S100P that presumably identified non-BM HSPGs. Moreover, the no co-
localization of HSPGs and PGP9.5 confirmed that HSPGs do not encircle the
axon (Figure 1g-i). Double immunofluorescence also confirmed that HSPGs are
not present in the inner core of Pacinian corpuscles (Figs. 2b-g).
HSPGs in Meissner’s and Pacinian corpuscles are localized in BM. The
immunohistochemical patterns of distribution of HSPGs and type IV collagen
were similar in both Meissner’s and Pacinian corpuscles (Figs. 3a-d; 4a,b
respectively), and also in the BM of skin epidermis (Figs. 3a-d).
To verify this almost identical localization, double immunofluorescence for
HSPGs and type IV collagen was performed. The results demonstrate that they
Page 11 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
were mainly co-localized at BM of Meissner’s (Figs. 3e-g) and Pacinian
corpuscles (Figs. 4c-h). In Pacinian corpuscles, HSPGs immunoreactivity was
detected in the outer part of the capsule, not co-localized with type IV collagen,
which indicates the occurrence of HSPGs independent of BM (Figure 4c-e).
HSPGs are localized in a specific perineural area in the innermost lamellae of
the outer core of Pacinian corpuscles. Since human Pacinian corpuscles show
an intermediate layer between the inner and the outer cores, and the intensity of
immunostaining for HSPGs (as well for type IV collagen) was strong in this
limiting zone, we decided to elucidate where HSPGs are localized. Light
microscopy images shown that HSPGs immunoreactivity coincides with the
endoneural CD34-positive intermediate layer (Figure 5a,b). Nevertheless,
double immunofluorescence showed no co-localization of HSPGs and CD34
(Figure 5c-h). This together with the no co-localization of HSPGs and S100
protein that exclude the presence of HSPGs in the outer lamellae of the inner
core (Figs. 2b-g), strongly suggesting that HSPGs in this corpuscular region are
localized in the innermost lamellae of the outer core.
Discussion
Page 12 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
In the present study we have demonstrate the presence of HSPGs
immunoreactivity in human cutaneous Meissner’s and Pacinian corpuscles,
primarily associated to BM. It must be clarified that the antibody used here
recognizes an epitope that includes N-unsubstituted glucosamine residues of
HS chains. Thus the results apply only for that molecule and not for the different
HSPGs BM or cell surface (Sarrazin et al., 2011; Iozzo and Schaefer, 2015).
Nevertheless, it can be assumed that HS present in basal membrane represent
the HSPGs perlecan, agrin, endostatin and type XV collagen (Yurchenco and
Patton, 2009; Iozzo and Schaefer, 2015).
BMs have been described of Meissner’s and Pacinian corpuscles using
transmission electron microscopy (Malinovsky and Pac, 1982; Malinovsky,
1986; Ide et al., 1987; Ide and Hayashi, 1987; Munger and Ide, 1988) as well as
immunohistochemistry (Vega et al., 1995). Two main components of BM in
human sensory corpuscles are laminin and type IV collagen (Vega et al., 1995).
Here we used as a criterion to determine that HSPGs are present in BM their
co-localization with type IV collagen. Conversely, whereas the absence of co-
localization was interpreted as an evidence of the presence of HSPGs outside
Page 13 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
BM presumably associated to HSPGs like syndecan or glypican families (Iozzo
and Schaefer, 2015).
In Meissner’s corpuscles, HSPGs were mostly found as a part of BM, although
occasionally HSPGs were detected in the cytoplasm of the lamellar cells. This
may be due to cytoplasmic production of HSPGs (Carey et al., 1993; Mehta et
al., 1985; Dziadek et al., 1986). The distribution of type IV collagen matched
and confirms previous studies form our group (Vega et al., 1995). Regarding
Pacinian corpuscles, HSPGs were exclusively co-localized with type IV
collagen, indicating localization in BM. Nevertheless, whether or not BM is
present in all the compartments of Pacinian corpuscle is still matter of
discussion. Based on the present results alone, it seems that the cells of the
inner core and those of the intermediate layer as well lack BM, although it has
been described using transmission electron microscopy (Ide and Hayashi,
1987; Munger and Ide, 1988; Malinovsky, 1986), and the presence of type IV
collagen and laminin was reported in those localizations (Vega et al., 1995).
The present study partially agrees with that by Vega and co-workers since we
have not found immunohistochemical evidences of BM in the inner core or the
Page 14 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
intermediate layer. So, the matter remains still open. In fact, while Ide et al.
(1987), Ide and Hayashi (1987) and Munger and Ide (1988) failed to
demonstrate BM in the inner core cells, Malinovsky and Pac (1982) and
Malinovsky (1986) found it regularly in these places. Further studies are
necessaries to elucidate these discrepancies. On the other hand, and
supporting present finding, there is consensus that the outer core and the
capsule have BM (Munger and Ide, 1988; Malinovsky, 1986; Ide et al., 1987).
The most interesting finding of our study refers to a strong localization of
HSPGs in a corpuscular area that comprises the innermost lamellae of the outer
core. This coincides with the so-called transitional area by Vega et al. (1995)
regarded of perineurial origin (Halata et al., 1990), although recently it was
demonstrated that this zone (also denominated intermediate layer) shares
endoneurial characteristics (García-Piqueras et al., 2017). Here we
demonstrate that the intermediate layer lack BM.
The cells forming Pacinian corpuscles are coupled by different types of
membrane junctions. Whilst the inner core cells communicate among
themselves by gap junctions, the outer core cells are coupled with thigh
Page 15 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
junctions, which are “especially prominent in the first and second (or sometimes
third) innermost lamellae of the outer core” (Ide and Hayashi, 1987). So, the
lamellae with the stronger expression of type IV collagen and HSPGs
correspond to the lamellae with prominent thigh junctions. Special organization
of thigh junction in these first outer core lamellae avoid the leak of fluid and ions
between the inner and outer core, and prevent a perturbation of the electrical
composition in the inner core from changings in the outer core composition (Ide
and Hayashi, 1987; Munger and Ide, 1988). Therefore, BM of the outer core
itself (Bush and Allt, 1990) or HSPGs components (Piña-Oviedo and Ortiz-
Hidalgo, 2008) might act as a barrier that isolates the axon and the inner core,
and presumably also the intermediate layer, from the outer core core-capsule.
ECM molecules of perineural cells are able to regulate the endoneural pressure
(Piña-Oviedo and Ortiz-Hidalgo, 2008).
It is also accepted now that some BM components, as well as ECM molecules,
are implicated in mechanotransduction (Wetzel et al., 2007; Hu and Chiang,
2010; Chiang et al., 2011). Whether or not HSPGs are involved in those
functions remains to be established.
Page 16 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
As a summary, in the present study we have demonstrated that HSPGs are
mainly associated to BM in human cutaneous Meissner’s and Pacinian
corpuscles, and that the inner core and the intermediate layer lack HSPGs thus
suggesting the lack BM.
Acknowledgements.- This study was supported by a grant from Gerencia
Regional de Salud de Castilla y León to JF (GRS 1615/A/17). The authors
thanks Dr. Marta Guervos (Servicios Comunes de Investigación, Microscopia
Confocal, Universidad de Oviedo) and Marta Sánchez-Pitiot (Grupo de
Histopatología molecular, Instituto Universitario de Oncología del Principado de
Asturias) for technical assistance.
Page 17 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
References
Bovolenta, P., Fernaud-Espinosa, I., 2000. Nervous system proteoglycans as
modulators of neurite outgrowth. Prog. Neurobiol. 61, 113-132.
Bush, M.S., Allt, G., 1990. Blood-nerve barrier: distribution of anionic sites on
the endothelial plasma membrane and basal lamina. Brain. Res. 535, 181-
188.
Carey, D.J., Stahl, R.C., Asundi, V.K., Tucker, B., 1993. Processing and
subcellular distribution of the Schwann cell lipid-anchored heparan sulfate
proteoglycan and identification as glypican. Exp. Cell. Res. 208, 10-18.
Chernousov, M.A., Stahl, R.C., Carey, D.J., 1998. Schwann cells use a novel
collagen-dependent mechanism for fibronectin fibril assembly. J. Cell. Sci.
111, 2763-2777.
Chiang, L.Y., Poole, K., Oliveira, B.E., Duarte, N., Sierra, Y.A., Bruckner-
Tuderman, L., Koch, M., Hu, J., Lewin, G.R., 2011. Laminin-332 coordinates
mechanotransduction and growth cone bifurcation in sensory neurons. Nat.
Neurosci. 14, 993-1000.
Page 18 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Chouchkov, C., Andreev, D., Dandov, A., 2003. Localization and distribution of
laminin in the basal lamina of certain mechanoreceptors--an immunogold
study. Somatosens. Mot. Res. 20, 265-270.
Dubový, P., Bednárová, J., 1999. The extracellular matrix of rat pacinian
corpuscles: an analysis of its fine structure. Anat. Embryol. (Berl.) 200, 615-
623.
Dziadek, M., Edgar, D., Paulsson, M., Timpl, R., Fleischmajer, R., 1986.
Basement membrane proteins produced by Schwann cells and in
neurofibromatosis. Ann. N. Y. Acad. Sci. 486, 248-259.
García-Piqueras, J., García-Suárez, O., Rodríguez-González, M.C., Cobo, J.L.,
Cabo, R., Vega, J.A., Feito, J., 2017. Endoneurial-CD34 positive cells define
an intermediate layer in human digital Pacinian corpuscles. Ann. Anat. 211,
55-60.
García-Piqueras, J., Carcaba, L., Feito, J., García, B., Viña, E., Suárez-
Quintanilla, J., Cobo, J., Vega, J.A., García-Suárez, O., 2018. Chondroitin
Sulfate in Human Cutaneous Meissner and Pacinian Sensory Corpuscles.
Anat. Rec. (Hoboken). doi: 10.1002/ar.23951. [Epub ahead of print].
Page 19 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Halata, Z., Grim, M., Christ, B., 1990. Origin of spinal cord meninges, sheaths of
peripheral nerves, and cutaneous receptors including Merkel cells. An
experimental and ultrastructural study with avian chimeras. Anat. Embryol.
(Berl.) 182, 529-537.
Halfter, W., Schurer, B., Yip, J., Yip, L., Tsen, G., Lee, J.A., Cole, G.J., 1997.
Distribution and substrate properties of agrin, a heparan sulfate proteoglycan
of developing axonal pathways. J. Comp. Neurol. 383, 1-17.
Halfter, W., Dong, S., Schurer, B., Cole, G.J., 1998. Collagen XVIII is a
basement membrane heparan sulfate proteoglycan. J. Biol. Chem. 273,
25404-25412.
Hu, J., Chiang, L.Y., Koch, M., Lewin, G.R., 2010. Evidence for a protein tether
involved in somatic touch. EMBO. J. 29, 855-867.
Ide, C., Hayashi, S., 1987. Specializations of plasma membranes in Pacinian
corpuscles: implications for mechano-electric transduction. J. Neurocytol. 16,
759-773.
Page 20 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Ide, C., Nitatori, T., Munger, B.L., 1987. The cytology of human Pacinian
corpuscles: evidence for sprouting of the central axon. Arch. Histol. Jpn. 50,
363-383.
Iozzo, R.V., Schaefer, L., 2015. Proteoglycan form and function: A
comprehensive nomenclature of proteoglycans. Matrix. Biol. 42, 11-55.
LeBleu, V.S., Macdonald, B., Kalluri, R., 2007. Structure and function of
basement membranes. Exp. Biol. Med. (Maywood) 232, 1121-1129.
Ma, E., Morgan, R., Godfrey, E.W., 1994. Distribution of agrin mRNAs in the
chick embryo nervous system. J. Neurosci. 14, 2943-2952.
Malinovský, L., Pác, L., 1982. Morphology of sensory corpuscles in mammals.
Acta Fac. Med. Univ. Brunensis. 79, 5-179.
Malinovský, L., 1986. Mechanoreceptors and free nerve endings. In: Biology of
the integument, 2. Vertebrates. Springer, Berlin-Heidelberg, 535-560.
Mehta, H., Orphe, C., Todd, M.S., Cornbrooks, C.J., Carey, D.J., 1985.
Synthesis by Schwann cells of basal lamina and membrane-associated
heparan sulfate proteoglycans. J Cell Biol. 101, 660-666.
Page 21 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Munger, B.L., Ide, C., 1988. The structure and function of cutaneous sensory
receptors. Arch. Histol. Cytol. 51, 1-34.
Piña-Oviedo, S., Ortiz-Hidalgo, C., 2008. The normal and neoplastic
perineurium: a review. Adv. Anat. Pathol. 15, 147-164.
Poulain, F.E., Yost, H.J., 2015. Heparan sulfate proteoglycans: a sugar code for
vertebrate development? Development. 142, 3456-3467.
Sames, K., Halata, Z., Jojovic, M., van Damme, E.J., Peumans, W.J., Delpech,
B., Asmus, B., Schumacher, U., 2001. Lectin and proteoglycan
histochemistry of feline pacinian corpuscles. J. Histochem. Cytochem. 49,
19-28.
Sarrazin, S., Lamanna, W.C., Esko, J.D., 2011. Heparan sulfate proteoglycans.
Cold. Spring. Harb. Perspect. Biol. 3:a004952.
Sasaki, T., Fässler, R., Hohenester, E., 2004. Laminin: the crux of basement
membrane assembly. J. Cell. Biol. 164, 959-963.
Schwarzbauer, J., 1999. Basement membranes: Putting up the barriers. Curr.
Biol. 9, R242-R244.
Page 22 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Vega, J.A., Esteban, I., Naves, F.J., del Valle, M.E., Malinovsky, L., 1995.
Immunohistochemical localization of laminin and type IV collagen in human
cutaneous sensory nerve formations. Anat. Embryol. (Berl.) 191, 33-39.
Vega, J.A., Haro, J.J., del Valle, M.E., 1996. Immunohistochemistry of human
cutaneous Meissner and Pacinian corpuscles. Micros. Res. Tech. 34, 351-
361.
Vega, J.A., García-Suárez, O., Montaño, J.A., Pardo, B., Cobo, J.M., 2009. The
Meissner and Pacinian sensory corpuscles revisited new data from the last
decade. Microsc. Res. Tech. 72, 299-309.
Wang, F., Wolfson, S.N., Gharib, A., Sagasti, A., 2012. LAR receptor tyrosine
phosphatases and HSPGs guide peripheral sensory axons to the skin. Curr.
Biol. 22, 373-382.
Wetzel, C., Hu, J., Riethmacher, D., Benckendorff, A., Harder, L., Eilers, A.,
Moshourab, R., Kozlenkov, A., Labuz, D., Caspani, O., Erdmann, B.,
Machelska, H., Heppenstall, P.A., Lewin, G.R., 2007. A stomatin-domain
protein essential for touch sensation in the mouse. Nature. 445, 206-209.
Page 23 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
Yamaguchi, Y., 2001. Heparan sulfate proteoglycans in the nervous system:
their diverse roles in neurogenesis, axon guidance, and synaptogenesis.
Semin. Cell. Dev. Biol. 12, 99-106.
Yurchenco, P.D., Patton, B.L., 2009. Developmental and pathogenic
mechanisms of basement membrane assembly. Curr. Pharm. Des. 15,
1277-1294.
Zelená, J., 1994. Nerves and mechanoreceptors. Chapman & Hall, London
LEGENDS FOR FIGURES
Figure 1. Human Meissner’s corpuscles showing immunoreactivity for heparan
sulfate. Objective 40x (a-c). Double immunofluorescence using anti-heparan
sulfate antibody conjugated with CyTM3 (d, g) and anti-S100P (e) or PGP9.5
(h) antibodies conjugated with Alexa Fluor 488. Predominately no colocalization
or partially colocalization between heparan sulfate and S100P was indicative of
a Schwann cell pericellular expression pattern (f). No colocalization with
Page 24 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
PGP9.5 excluded the presence of heparan sulfate in the axon (i). Objective
63x/1.40 oil; pinhole 1.37; XY resolution 139.4 nm and Z resolution 235.8 nm
(d-i).
Figure 2.- Single immunohistochemistry for heparan sulfate in human
cutaneous Pacinian corpuscles. Heparan sulfate is present in all outer core-
lamellae including the capsule and more intensely in a thin layer around the
inner core. Objective 20x (a). Double immunofluorescence for heparan sulfate
(b, e) and S100P (c, f) confirmed absence of heparan sulfate in the inner core of
Pacinian corpuscles (d, g). Objective 20x/1.25 oil; pinhole 1.30; XY resolution
156 nm and Z resolution 266 nm (b-d); Objective 63x/1.40 oil; pinhole 1.37; XY
resolution 139.4 nm and Z resolution 235.8 (e-g).
Figure 3. Single immunohistochemistry for heparan sulfate (a, b) and type IV
collagen (c, d). Heparan sulfate expression was observed in basement
membranes of Meissner’s corpuscles similar to type IV collagen pattern. Note
equal immunoreactivity in basement membrane of epidermis. Objective 10x (a-
d); Colocalization of heparan sulfate (e) and type IV collagen (f) using double
immunofluorescence (secondary antibodies conjugated with CyTM3 and Alexa
Page 25 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
fluor 488 respectively) confirmed the presence of heparan sulfate in basement
membranes of Meissner’s corpuscles (g). Objective 63x/1.40 oil; pinhole 1.37;
XY resolution 139.4 nm and Z resolution 235.8 nm (e-g).
Figure 4. Single immunohistochemistry for type IV collagen (a) and heparan
sulfate (b) antibodies in Pacinian corpuscles. A similar pattern was observed:
immunoreactivity in all outer core lamellae including the capsule and more
intensive expression in a layer at the periphery of the inner core. Objective 20x
(a, b); Double immunofluorescence confirmed the co-localization of heparan
sulfate (c, f) and type IV collagen (d, g) in Pacinian corpuscles (e, h). Note that
some outer core lamellae were immunopositive just for heparan sulfate and vice
versa. Objective 20x/1.25 oil; pinhole 1.30; XY resolution 156 nm and Z
resolution 266 nm (c-e). Objective 63x/1.40 oil; pinhole 1.37; XY resolution
139.4 nm and Z resolution 235.8 nm (f-h).
Figure 5. Single immunohistochemistry for heparan sulfate (a) and CD34 (b) in
Pacinian corpuscles. HS-immunoreactivity was observed more intensively in a
thin layer around the inner core, similar to the intermediate layer described by
CD34-positive cells. Objective 20x (a, b); Double immunofluorescence using
Page 26 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
anti-HS (c, f) and anti-CD34 (d, g) antibodies (conjugated with CyTM3 and
Alexa fluor 488 respectively) showed no colocalization between them (e, h). HS
is expressed in the first lamellae of the outer core but not in the positive-CD34
intermediate layer of Pacinian corpuscles. Objective 20x/1.25 oil; pinhole 1.30;
XY resolution 156 nm and Z resolution 266 nm µm (c-e). Objective 63x/1.40 oil;
pinhole 1.37; XY resolution 139.4 nm and Z resolution 235.8 nm (f-h).
Page 27 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
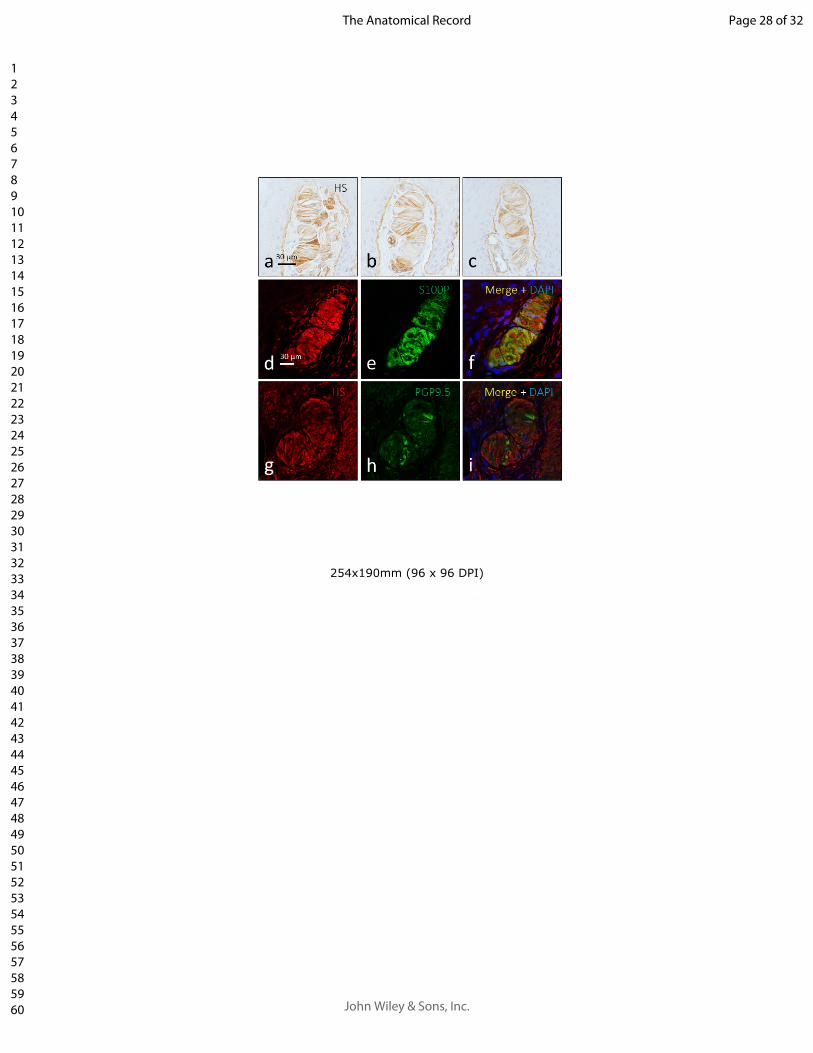
For Peer Review
254x190mm (96 x 96 DPI)
Page 28 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960
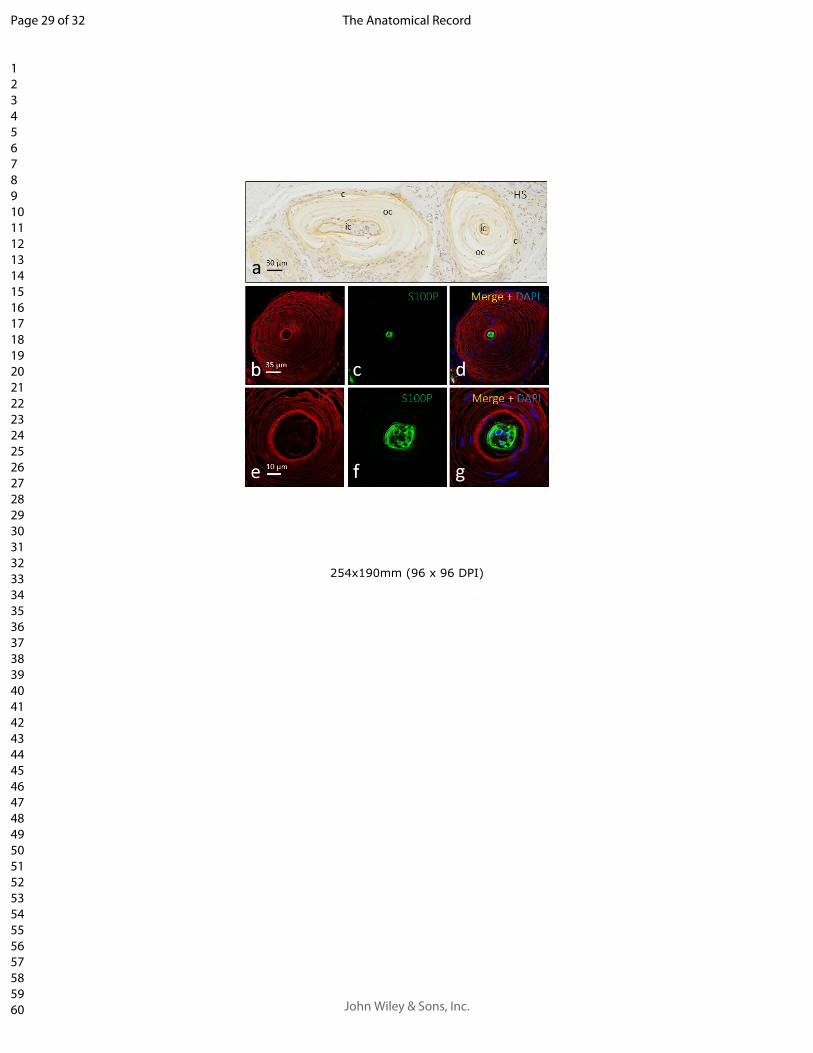
For Peer Review
254x190mm (96 x 96 DPI)
Page 29 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
254x190mm (96 x 96 DPI)
Page 30 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
254x190mm (96 x 96 DPI)
Page 31 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

For Peer Review
254x190mm (96 x 96 DPI)
Page 32 of 32
John Wiley & Sons, Inc.
The Anatomical Record
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354555657585960

Elsevier Editorial System(tm) for Annals of
Anatomy
Manuscript Draft
Manuscript Number:
Title: CLASS I AND CLASS II SMALL LEUCINE-RICH PROTEOGLYCANS IN HUMAN
CUTANEOUS PACINIAN CORPUSCLES
Article Type: Research Article
Keywords: Pacinian corpuscles; extracellular matrix; small leucine-rich
proteoglycans; human
Corresponding Author: Professor Jose A Vega, Ph.D, M.D.
Corresponding Author's Institution: Universidad de Oviedo
First Author: Jorge Garcia-Piqueras
Order of Authors: Jorge Garcia-Piqueras; Yolanda García-Mesa; Jorge
Feito; Beatriz Garcia; Luis M Quiros; Benjamin Martin-Biedma; Juan Cobo;
Jose A Vega, Ph.D, M.D.; Olivia Garcia
Abstract: Pacinian corpuscles are onion bulb-like multilayered
mechanoreceptors that consist of a complicated structure of axon
terminals, Schwann related cells (inner core), endoneural related cells
(intermediate layer) and perineurial related cells (outer core-capsule).
The cells forming those compartments are continuous and share the
properties of that covering the nerve fibers. Small leucine-rich
proteoglycans are major proteoglycans of the extracellular matrix and
regulate collagen fibrillogenesis, cell signalling pathways and
extracellular matrix assembly. Here we used immunohistochemistry to
investigate the distribution of class I (biglycan, decorin, asporin, ECM2
and ECMX) and class II (fibromodulin, lumican, prolargin, keratocan and
osteoadherin) small leucine-rich proteoglycans in human cutaneous
Pacinian corpuscles. The distribution of these compounds was: the inner
core express decorin, biglycan, lumican, fibromodulin, osteoadherin; the
intermediate layer display immunoreactivity for osteoadherin; the outer
core biglycan, decorin, lumican, fibromodulin and osteoadherine; and the
capsule contains biglycane, decorin, fibromoduline, and lumican.
Aspronin, prolargin and keratocan were undetectable. These results
complement our knowledge about the distribution of small leucine-rich
proteoglycans in human Pacinian corpuscles, and help to understand the
composition of the extracellular matrix in these sensory formations.

December 21, 2018
Dear Editor,
We are pleased to submit for publication in Annals of Anatomy a manuscript entitled
“CLASS I AND CLASS II SMALL LEUCINE-RICH PROTEOGLYCANS IN HUMAN CUTANEOUS
PACINIAN CORPUSCLES”, by J. García-Piqueas et al. The manuscript consists of 22
typewritten pages (including 1 table, references and legends for figures), and 8 figures.
The manuscript was prepared following the Authors Guide of Annals of Anatomy. I hope
de manuscript can be of interest for the Journal.
I am looking forward to hearing from you, and I would like to thank you in advance for
your attention and your time. I wish you Merry Christmas and a Happy New Year.
Sincerely yours.
Best regards,
Prof. J.A. Vega
Cover Letter

December 21, 2018
Dear Editor,
Manuscript: “CLASS I AND CLASS II SMALL LEUCINE-RICH PROTEOGLYCANS IN HUMAN
CUTANEOUS PACINIAN CORPUSCLES”, by J. García-Piqueas et al.
Conflict of Interest: The authors declare no conflict of interest
Conflict of Interest

CLASS I AND CLASS II SMALL LEUCINE-RICH PROTEOGLYCANS IN HUMAN
CUTANEOUS PACINIAN CORPUSCLES
J. García-Piqueras1, Y. García-Mesa1, J. Feito1,2, B. García3, L.M. Quiros3,4,
B. Martín-Biedma5, J. Cobo6,7, J.A. Vega1,8, O. García-Suárez1
1Departamento de Morfología y Biología Celular, Grupo SINPOS Universidad de Oviedo,
Spain 2Servicio de Anatomía Patológica, Hospital Universitario de Salamanca, Salamanca, 3 Instituto Universitario Fernández-Vega, Universidad de Oviedo & Fundación de
Investigación Oftalmológica, Oviedo, Spain 4Departmento de Biología Funcional, Universidad de Oviedo, Spain 5Departamento de Cirugía y Especialidades Médico-Quirúrgicas, Universidad de
Santiago de Compostela, Spain 6Departamento de Cirugía y Especialidades Médico-Quirúrgicas, Universidad de Oviedo,
Spain 7Instituto Asturiano de Odontología, Oviedo, Spain 8Facultad de Ciencias de la Salud, Universidad Autónoma de Chile, Chile
Short title: Extracellular matrix in Pacinian corpuscles
Address for correspondence José A. Vega, M.D., Ph.D. Departamento de Morfología y Biología Celular Facultad de Medicina y Ciencias de la Salud – Universidad de Oviedo Avd. JualianClaveria, 6, Planta 9ª 33006 OVIEDO – Spain Telf. (0034) 985104097 E-mail: [email protected]
*Manuscript

ABSTRACT
Pacinian corpuscles are onion bulb-like multilayered mechanoreceptors that consist of a
complicated structure of axon terminals, Schwann related cells (inner core), endoneural
related cells (intermediate layer) and perineurial related cells (outer core-capsule). The
cells forming those compartments are continuous and share the properties of that
covering the nerve fibers. Small leucine-rich proteoglycans are major proteoglycans of
the extracellular matrix and regulate collagen fibrillogenesis, cell signalling pathways
and extracellular matrix assembly. Here we used immunohistochemistry to investigate
the distribution of class I (biglycan, decorin, asporin, ECM2 and ECMX) and class II
(fibromodulin, lumican, prolargin, keratocan and osteoadherin) small leucine-rich
proteoglycans in human cutaneous Pacinian corpuscles. The distribution of these
compounds was: the inner core express decorin, biglycan, lumican, fibromodulin,
osteoadherin; the intermediate layer display immunoreactivity for osteoadherin; the
outer core biglycan, decorin, lumican, fibromodulin and osteoadherin; and the capsule
contains biglycan, decorin, fibromodulin, and lumican. Asporin, prolargin and keratocan
were undetectable. These results complement our knowledge about the distribution of
small leucine-rich proteoglycans in human Pacinian corpuscles, and help to understand
the composition of the extracellular matrix in these sensory formations.
Keywords: Pacinian corpuscles; extracellular matrix; small leucine-rich proteoglycans;
human.

INTRODUCTION
The Pacini’s corpuscles are rapidly-adapting low-threshold mechanoreceptors
functionally connected with myelinated A or A nerve fibres that detect gross
pressure changes and vibrations (Johnson, 2001; Roudaut et al., 2012; Fleming and Luo,
2013; Jones and Smith, 2014). They are widely distributed throughout the body (Bell et
al., 1994; Zelena, 1994), and their structure is independent of the anatomical
localization (Pawson et al., 2008). Typically they are oval-shaped multilayered
capsulated formations that consist of one central axon surrounded by non-neuronal
cells arranged in concentric lamellae, that form the inner core, intermediate layer, outer
core, and capsule (see Bell et al., 1994; Zelena, 1994). The inner core is composed of
terminal non-myelinating Schwann-like cells, the intermediate layer is formed by
endoneurial-related cells, and the outer core-capsule cells are perineurial-related cells
(Munger and Ide, 1988; Zelena, 1994, Vega et al., 2009; Feito et al., 2016; García-
Piqueras et al., 2017). Consistently, the different parts of the Pacinian corpuscles share
the immunohistochemical profile of their counterparts in nerves (Vega et al., 1996,
2009). Interposed between the different cell layers forming the Pacinian corpuscles
there is an extracellular matrix (ECM) occasionally organized as a basement membrane
(BM). Thus, it has been described a BM in relation to the inner core, outer core and
capsule of Pacinian corpuscles of different mammalian species (Munger and Ide, 1988;
Bell et al., 1994; Zelena, 1994; Vega et al., 1995; Dubový and Bednarova, 1999).
Recently, the presence of chondroitin sulfate proteoglycan chains was found associated
to the intermediate layer in human cutaneous Pacinian corpuscles (García-Piqueras et
al., 2018).

ECM is an organized complex network primarily formed of fibrillary proteins, such as
collagens and laminins, and proteoglycans (PGs; Iozzo and Schaefer, 2015; Theocharis et
al., 2016). Collagens are localized in the interlamellar spaces of Pacinian corpuscles:
type II collagen predominates in the outer core and capsule, type V collagen in the
intermediate layer, and collagen IV is present in all three extra-neuronal compartments
(Vega et al., 1995; Pawson et al., 2000). Conversely, little information is available about
the occurrence and distribution of other ECM components, especially PGs, in Pacinian
corpuscles.
PGs fall into two main groups: small and large. The group of small PGs (also known as
small leucine-rich proteoglycans; SLRPs), contains 18 members that are subclassified
into classes I to V. In particular class I is formed by biglycan (BGN), decorin (DCN),
asporin (ASPN), extracellular matrix protein 2 (ECM2) and ECMX, whereas class II is
formed by fibromodulin (FMOD), lumican (LUM), prolargin (PRELP), keratocan (KERA)
and osteoadherin (OMD) (McEwan et al., 2006; Schaefer and Schaefer, 2010; Chen and
Birk, 2013). Sames et al. (2001) are the only authors reporting the presence of DCN and
BGN in the cat mesentery Pacinian corpuscles.
SLRPs are expressed mainly during neural development but also in adult neural tissues
(Le Goff and Bishop, 2007; Dellett et al., 2012). In the peripheral nervous system,
including cat Pacinian corpuscles (Sames et al., 2001), some SLRPs have been detected
(Hanemann et al., 1993; Wilda et al., 2000). Nevertheless, as far as we know, SLRPs have
not been systematically explored in the human Pacinian corpuscles. Thus, the present
study was designed to investigate the occurrence and distribution of classes I and II of
SLRPs in human digital Pacinian corpuscles, and thus to contribute to the knowledge of
ECM composition in sensory corpuscles.

MATERIAL AND METHODS
Skin samples were obtained from the palmar side of the distal phalanx of amputated
hand fingers (n = 15) collected within 6-8 h after the accident from otherwise healthy
subjects (age range 24–42 years). Also, samples of finger tip skin (n = 7; age range 8–
56 years) were obtained from necropsies of neurological-disease-free subjects at the
Department of Pathology of the Hospital Universitario Central de Asturias (Oviedo,
Spain) and Complejo Hospitalario Universitario de Salamanca (Salamanca, Spain). The
specimens were fixed in 4% formaldehyde in 0.1 M phosphate buffer saline (pH 7.4) for
24 h, dehydrated, and embedded in paraffin.
All the tissue samples used in this study were obtained in compliance with Spanish Law
and the guidelines of the Helsinki Declaration II, and the study was approved by the
Ethical Committee for Biomedical Research of the Principality of Asturias, Spain (Ref.
266/18). The paraffin-embedded digital skin was cut into 10 μm-thick sections
perpendicular to the skin surface and mounted on gelatine-coated microscope slides.
The presence of Pacinian corpuscles was ensured staining randomly selected sections
with hematoxylin & eosin.
Immunohistochemistry.- Sections were deparaffinized, rehydrated and rinsed in 0.05 M
HCl Tris buffer (pH 7.5) containing 0.1% bovine serum albumin and 0.1% Triton X-100.
Thereafter, the endogenous peroxidase activity (3% H2O2) and non-specific binding
(10% fetal calf serum) were blocked, and the sections were incubated overnight in a
humid chamber at 4° C with the primary antibodies listed in Table 1. The antibodies
against protein gene product 9.5 (PGP9.5), neurofilaments proteins (NFP), peripherin,
S100 protein, CD34 and vimentin, were used to immunolabel the axon, inner core,
intermediate layer and outer core-capsule, respectively, within Pacinian corpuscles

(Vega et al., 2009; García-Piqueras et al., 2017). For immunohistochemical detection of
BGN the sections were heated in Envision FLEX target retrieval solution of pH6 (Dako) at
65°C for 20 min and then incubated for another 20 min at room temperature in the
same solution, previously to incubation of the primary antibody. After incubation, the
sections were rinsed in the same buffer as above and incubated with Dako EnVision
System labeled polymer-HR anti-rabbit IgG or anti-mouse IgG (DakoCytomation,
Denmark) for 30 minutes at room temperature. Finally, sections were washed, and the
immunoreaction visualized using 3-3’-diaminobenzidine as a chromogen. To ascertain
structural details, sections were slightly counterstained with hematoxylin & eosin.
For control purposes, representative sections were processed in the same way as
described above using non-immune rabbit or mouse sera instead of the primary
antibodies or omitting the primary antibodies in the incubation. Under these conditions,
no positive immunostaining was observed (data not shown).
Double immunofluorescence.- Based on the results of positivity for SLRPs in simple
immunohistochemistry, double immunofluorescence was carried out for DCN or BGN or
LUM or FMOD or OMD with axonal markers, S100 protein, vimentin and CD34.
Deparaffinized and rehydrated 10 µm thick sections were processed for reduction of
non-specific bindings (30 minutes with a solution of 5% bovine serum albumin in tris-
buffer saline –TBS- pH 7,4). Them they were incubated overnight, at 4° C in a humid
chamber with a 1:1 v/v mixture of the selected antibodies. After rinsing with TBS, the
sections were incubated for 1 hour with Alexa fluor 488-conjugated goat anti-rabbit IgG
(Invitrogen, Eugene, OR, USA), diluted 1:1000 in TBS containing 5% mouse serum
(Serotec, Oxford, UK), then rinsed again and incubated for another hour with Cy 3-
conjugated donkey anti-mouse antibody (Jackson-ImmunoResearch, Baltimore, MD,

USA) diluted 1:50 in TBS. Both steps were performed at room temperature in a dark
humid chamber. Sections were then washed and mounted with Fluoromount Gold.
Finally, to ascertain structural details sections were counterstained with DAPI (10
ng/ml). In the case of double immunodetection of CD34 with DCN, FMOD or OMD,
sections were incubated overnight in a humid chamber at 4o C with primary mouse anti-
DCN, anti-FMOD or anti-OMD, followed by incubation with Cy3-conjugated donkey anti-
mouse antibody for 90 minutes. Thereafter samples were incubated overnight with
mouse anti-CD34 antibody. After rinsing with TBS, sections were incubated with
prediluted rabbit Post-Primary IgG, which recognizes mouse antibodies (Novolink
Polymer Detection System®, Leica-Novocastra Biosystems, Newclastle, UK) at room
temperature for 90 minutes. After that, sections were incubated for 90 minutes at room
temperature with Alexa fluor 488-conjugated goat anti-rabbit IgG. Finally, to label
nuclei, sections were counterstained with DAPI (10 ng/ml) and mounted with
Fluoromount Gold as described above. For control purposes representative sections
were processed in the same way as described above using non-immune rabbit or mouse
sera instead of the primary antibodies or omitting the primary antibodies in the
incubation. Triple staining was detected using a Leica DMR-XA automatic fluorescence
microscope coupled with a Leica Confocal Software, version 2.5 (Leica Microsystems,
Heidelberg GmbH, Germany) and the images captured were processed using the
software Image J version 1.43 g Master Biophotonics Facility, Mac Master University
Ontario (www.macbiophotonics.ca).
RESULTS
Immunohistochemistry was performed to investigate the occurrence and localization of
some members of the class I and class II SLRPs in human digital Pacinian corpuscles. So,

we investigated the distribution within them of ASPN, BGN, DCN, FMOD, OMD, LUM,
PRELP and KERA. As a rule ASPN, PRELP and KERA were undetectable (data not shown)
whereas specific immunostaining was detected for the other SLRPs investigated with
different patterns of distribution within the corpuscles.
Class I SLRPs.- Serial sections of Pacinian corpuscles showed different patterns of
distribution for DCN and BGN. In light microscopy DCN and BGN immunoreactivity was
observed in all the corpuscular compartments including the intermediate layer, inner
and the outer core (Figs. 1a-e). Double immunofluorescence for DCN in combination
with specific markers of Pacinian corpuscular constituents (S100 protein, vimentin and
CD34) demonstrate that DCN surround the S100 protein positive cells, thus it is
localized outside the inner core (Fig. 2a-c). In addition, the relation of DCN
immunoreactive structures with the intermediate layer suggests that DCN is also
outside this layer (Fig. 2d-f). Therefore, DCN should be expressed in the interlamellar
spaces of the outer core and the capsule (Fig. 2g-i).
A strong and specific immunoreactivity for BGN was found in the inner core, outer core
and capsule (Fig. 1a-d). However, because the lack of immunoreactivity observed
encircling the inner core, the presence of BGN in the intermediate layer cannot be
ensured (Fig. 1d). To ascertain whether BGN is localized in the inner segment of the
Pacinian corpuscles simultaneous detection of BGN and S100 protein, or BGN and
peripherin, was performed. The results demonstrated that while BGN highly co-localizes
with S100 protein, thus supporting its presence in the inner core (Figs. 3a-c), it was
absent from the central axon (Fig. 3d-f). In addition, co-localization of BGN and
vimentin, but not of BGN and CD34, demonstrates that BGN is present in the outer core
lamellae and capsule (Fig. 3j-l) but not in the intermediate layer (Fig 3g-i).

Class II SLRPs.- Comparing serial sections processed for detection of S100P (Fig. 4a),
CD34 (Fig. 4c) and vimentin (Fig. 4e) with those used for detection of class II SLRPs, it is
apparent that the distribution of LUM immunoreactivity overlaps with the inner core,
intermediate layer and the outermost lamellae of the capsule (Fig. 4b); OMD has the
same distribution observed with LUM without the capsule´s outermost enhancement
(Fig. 4f); FMOD was only detected in the inner core and outer core-capsule (Fig. 4d).
In order to determine the precise localization of these molecules we performed double
immunofluorescence with specific antigens of the different corpuscular compartments
(Fig. 5). The results of these experiments demonstrated that LUM is absent from the
axon (Fig. 5a-c) and the intermediate layer (Fig. 5g-i) while it is localized in the inner
core (Fig. 5d-f), and widely in the outer core and capsule (Fig. 5l-k). Regarding FMOD, it
was confirmed that FMOD is absent from the axon (Fig. 6a-c) and the intermediate layer
(Fig. 6g-i). Conversely, a faint but specific reaction was detected in the inner core,
although in many cases do not match completely with S100 protein (Fig. 6d-f); the outer
core-capsule display solid co-localization (Fig. 6j-l). The inner core, intermediate layer
and outer core displayed OMD immunoreactivity (Fig. 7). Although in most cases co-
localization of OMD and S100P was observed, we have occasionally found that a
subpopulation of Schwan cells OMD positive do not express S100 and vice versa (Fig.
7c).
The results, based on the results of double immunofluorescence, are summarized in the
figure 8.
DICUSSION
ECM is composed by network of secreted proteins surrounding cells, either as an
interstitial matrix or organized into basement membranes. In Pacinian corpuscles ECM

fills the spaces between lamellae that form the inner core, intermediate layer and outer
core-capsule (Ide, 1986). In some of these compartments ECM is organized as a BM
(Malinovský et al., 1986; Vega et al., 1995; Dubový and Aldskogius, 1996; García-
Piqueras et al., 2018). The current information about the composition of ECM in
vertebrate Pacinian corpuscles regards to different types of collagen, laminin and two
PGs (Malinovský et al., 1986; Vega et al., 1995, 1996; Sames et al., 2001). Thus, in the
present study we have investigated the occurrence of other constituents of ECM, in
particular class I (DCN, BGN and ASPN) and Class II (LUM, FMOD, OMD, KERA and
PRELP) SLRPs.
A previous study carried out on Pacinian corpuscles from the cat mesentery detected
DCN and BGN (Sames et al., 2001), being found DCN in the outer core and BGN in the
inner core. Present results in human digital Pacinian corpuscles matches those for DCN
in the cat, but differs with respect to BGN distribution since they found it restricted to
the inner core while we observed immunoreactivity in the inner core and also in the
outer core. The role of these two molecules in Pacinian corpuscles is unknown.
Evidence of the influence of DCN PGs in the cellular morphology via the stabilization of
the cytoskeletal vimentin intermediate filaments has been recently reported (Jungmann
et al., 2012) but the importance of this relationship in the Pacinian corpuscles outer
core must be elucidated in additional studies.
As far as we know the occurrence and distribution of class II SLRPs in vertebrate
Pacinian corpuscles was never investigated. Therefore, data provided in the present
study cannot be contrasted with other studies.
Biochemical and cell culture assays have shown that SLRPs modulate ECM organization
by binding to various collagens (I, II, III, IV, V, VI, XII and XIV; Chen and Birk 2013;

Gubbiotti et al., 2016). In particular, LUM binds collagen II, BGN and FMOD interact with
collagens II and VI, and DCN binds collagens II, IV, V and VI (Chen and Birk, 2013). The
interlamellar spaces of Pacinian corpuscles contain different types of collagen (II, IV, V,
VI; Tachibana et al., 1995; Vega et al., 1996; Pawson et al., 2000) and members of the
class I and class II SLRPs proteins participate in the regulation of collagen fibril diameter
and interfibrillar spacings (Chen and Birk, 2013; Tashima et al., 2015). SLRPs modulate
ECM fibrillogenesis, organization and assembly (Kalamajski and Oldberg, 2010; Chen
and Birk, 2013; Gubbiotti et al., 2016).
On the other hand, SLRPs regulate cell activity interacting with growth factor receptors
(Hocking et al., 1998; Svensson et al., 1999; Ezura et al., 2000; Merline et al., 2009;
Dellett et al., 2012; Iozzo and Schaefer 2015; Neill et al., 2015). The relative proportion
of collagen and SLRPs determine the mechanical properties of the tissues (Hansen et al.,
2015). DCN and BGN bind to epidermal growth factor receptor (EGFR) and Erb4
(Schaefer and Iozzo, 2008; Schaefer and Schaefer, 2010; Iacob and Cs-Szabo, 2012) and
the localization of those SLRPs in human Pacinian corpuscles observed here is similar to
that reported for EGFR in human Pacinian corpuscles (Vega et al., 1994) or Erb4 in the
murine Pacinian corpuscles (González-Martínez et al., 2007). Interestingly DCN interacts
with EGFR to regulate collagen fibrillogenesis (Mohan et al., 2011).
SLRPs are expressed during neural development and in adult neural tissues (Le Goff and
Bishop, 2007; Dellett et al., 2012), including Schwann cells (Hanemann et al., 1993),
dorsal root ganglia (Wilda et al., 2000), and cat mesentery Pacinian corpuscles (Sames
et al., 2001). PGs, including the molecules studied here, are involved in regenerative
process of peripheral nervous system, both as neuritogenic molecules and as inhibitors
of axonal regeneration (Braunewell et al 1995; Oohira et al., 2000; Bradbury et al.,

2002; Maeda et al., 2011; Soleman et al., 2013; Silver and Silver, 2014). DCN inhibits
neurite outgrowth (Stichel et al., 1995; Davies et al., 2004), whereas BGN is a
neurotrophic molecule (Junghans et al., 1995; Koops et al., 1996).
In conclusion, this study provides a comprehensive analysis of SLRPs localization in
human cutaneous Pacinian corpuscle. Their complex expression patterns suggest a
compartment-specific regulation of collagen fibrillogenesis and stromal matrix assembly
and hydration. Considering the numerous SLRPs functions, they could be essential for
determining Pacinian corpuscle elasticity, mechanical properties and structural integrity
for the correct transmission of pressure resulting from indentation or stretch. The roles
of all the SLRPs in the human Pacinian corpuscles await elucidation in the future.
Acknowledgements.- This study was partially supported by a grant from Gerencia
Regional de Salud de Castilla y León to JF (GRS 1615/A/17). The authors thank Dr. Marta
Guervos (Servicios Comunes de Investigación, Microscopia Confocal, Universidad de
Oviedo) and Marta Sánchez-Pitiot (Grupo de Histopatología molecular, Instituto
Universitario de Oncología del Principado de Asturias) for technical assistance.

REFERENCES
Bell, J., Bolanowski, S., Holmes, M.H., 1994. The structure and function of Pacinian
corpuscles: a review. Prog. Neurobiol. 42, 79–128.
Bradbury, E.J., Moon, L.D., Popat, R.J., King, V.R., Bennett, G.S., Patel, P.N., Fawcett,
J.W., McMahon, S.B., 2002. Chondroitinase ABC promotes functional recovery after
spinal cord injury. Nature. 416, 636-640.
Braunewell, K.H., Pesheva, P., McCarthy, J.B., Furcht, L.T., Schmitz, B., Schachner, M.,
1995. Functional involvement of sciatic nerve-derived versican- and decorin-like
molecules and other chondroitin sulphate proteoglycans in ECM-mediated cell
adhesion and neurite outgrowth. Eur. J. Neurosci. 7, 805-814.
Chen, S., Birk, D.E., 2013. The regulatory roles of small leucine-rich proteoglycans in
extracellular matrix assembly. FEBS J 280, 2120-2137.
Davies, J.E., Tang, X., Denning, J.W., Archibald, S.J., Davies, S.J., 2004. Decorin
suppresses neurocan, brevican, phosphacan and NG2 expression and promotes
axon growth across adult rat spinal cord injuries. Eur. J. Neurosci. 19, 1226-1242.
Dellett, M., Hu, W., Papadaki, V., Ohnuma, S., 2012. Small leucine rich proteoglycan
family regulates multiple signalling pathways in neural development and
maintenance. Dev. Growth Differ. 54, 327-340.
Dubový, P., Aldskogius, H., 1996. Degeneration and regeneration of cutaneous sensory
nerve formations. Microsc. Res. Tech. 34, 362-375.
Dubový, P., Bednárová, J., 1999. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat. Embryol. (Berl.). 200, 615-623.

Ezura, Y., Chakravarti, S., Oldberg, A., Chervoneva, I., Birk, D.E., 2000. Differential
expression of lumican and fibromodulin regulate collagen fibrillogenesis in
developing mouse tendons. J. Cell Biol. 151, 779-788.
Feito, J., Ramos-García, J.L., Gago, Á., Cobo, J.L., García-Suárez, O., Junquera, L.M., Vega,
J.A., 2016. Pacinian Corpuscles in a Cervical Chondrocutaneous Remnant: A Case
Report and Update of Pacinian Corpuscles. Am. J. Dermatopathol. 38, 231-235.
Fleming, M.S., Luo, W., 2013. The anatomy, function and development of mammalian
Aβ low-threshold mechanoreceptors. Front. Biol. (Beijing) 8.
García-Piqueras, J., García-Suárez, O., Rodríguez-González, M.C., Cobo, J.L., Cabo, R.,
Vega, J.A., Feito, J., 2017. Endoneurial-CD34 positive cells define an intermediate
layer in human digital Pacinian corpuscles. Ann. Anat. 211, 55-60.
García-Piqueras, J., Carcaba, L., García-Mesa, Y., Feito, J., García, B., Viña, E., Suárez-
Quintanilla, J., Cobo, J., Vega, J.A., García-Suárez, O., 2018. Chondroitin Sulfate in
Human Cutaneous Meissner and Pacinian Sensory Corpuscles. Anat. Rec.
(Hoboken). [Epub ahead of print]. doi: 10.1002/ar.23951.
González-Martínez, T., Germana, A., Catania, S., Cobo, T., Ochoa-Erena, F.J., de Carlos,
F., Díaz-Esnal, B., Vega, J.A., 2007. Postnatal developmental changes in the
expression of ErbB receptors in murine Pacinian corpuscles. Neurosci. Lett. 420, 90-
95.
Gubbiotti, M.A., Vallet, S.D., Ricard-Blum, S., Iozzo, R.V., 2016. Decorin interacting
network: A comprehensive analysis of decorin-binding partners and their versatile
functions. Matrix Biol. 55, 7-21.

Hanemann, C.O., Kuhn, G., Lie, A., Gillen, C., Bosse, F., Spreyer, P., Müller, H.W., 1993.
Expression of decorin mRNA in the nervous system of rat. J. Histochem. Cytochem.
41, 1383-1391.
Hansen, M.L., Beck, H.C., Irmukhamedov, A., Jensen, P.S., Olsen, M.H., Rasmussen, L.M.,
2015. Proteome analysis of human arterial tissue discloses associations between
the vascular content of small leucine-rich repeat proteoglycans and pulse wave
velocity. Arterioscler. Thromb. Vasc. Biol. 35, 1896-1903.
Hocking, A.M., Shinomura, T., McQuillan, D.J., 1998. Leucine-rich repeat glycoproteins
of the extracellular matrix. Matrix Biol. 17, 1-19.
Iacob, S., Cs-Szabo, G., 2010. Biglycan regulates the expression of EGF receptors
through EGF signaling pathways in human articular chondrocytes. Connect. Tissue
Res. 51, 347-358.
Ide, C., 1986. Basal laminae and Meissner corpuscle regeneration. Brain Res. 384, 311-
322.
Iozzo, R.V., Schaefer, L., 2015. Proteoglycan form and function: A comprehensive
nomenclature of proteoglycans. Matrix. Biol. 42, 11-55.
Johnson, K.O., 2001. The roles and functions of cutaneous mechanoreceptors. Curr.
Opin. Neurobiol. 11, 455–461.
Jones, L.A., Smith, A.M, 2014. Tactile sensory system: encoding from the periphery to
the cortex. Wiley Interdiscip. Rev. Syst. Biol. Med. 6, 279-287.
Junghans, U., Koops, A., Westmeyer, A., Kappler, J., Meyer, H.E., Müller, H.W., 1995.
Purification of a meningeal cell-derived chondroitin sulphate proteoglycan with
neurotrophic activity for brain neurons and its identification as biglycan. Eur. J.
Neurosci. 7, 2341-2350.

Jungmann, O., Nikolovska, K., Stock, C., Schulz, J.N., Eckes, B., Riethmüller, C., Owens,
R.T., Iozzo, R.V., Seidler, D.G., 2012. The dermatan sulfate proteoglycan decorin
modulates α2β1 integrin and the vimentin intermediate filament system during
collagen synthesis. PLoS One. 7:e50809.
Kalamajski, S., Oldberg, A., 2010. The role of small leucine-rich proteoglycans in collagen
fibrillogenesis. Matrix Biol. 29, 248-253.
Koops, A., Kappler, J., Junghans, U., Kuhn, G., Kresse, H., Müller, H.W., 1996. Cultured
astrocytes express biglycan, a chondroitin/dermatan sulfate proteoglycan
supporting the survival of neocortical neurons. Brain Res. Mol. Brain Res. 41, 65-73.
Le Goff, M.M., Bishop, P.N., 2007. Focus on molecules: opticin. Exp. Eye Res. 85, 303-
304.
Maeda, N., Ishii, M., Nishimura, K., Kamimura, K., 2011. Functions of chondroitin sulfate
and heparan sulfate in the developing brain. Neurochem. Res. 36, 1228-1240.
Review.
Malinovský, L., 1986. Mechanoreceptors and free nerve endings. In: Bereiter-Hahn, J.,
Matoltsy, A.G., Richards, K.S. (Eds.), Biology of the Integument. 2. Vertebrates.
Springer-Verlag, Berlin, pp. 535–560.
McEwan, P.A., Scott, P.G., Bishop, P.N., Bella, J., 2006. Structural correlations in the
family of small leucine-rich repeat proteins and proteoglycans. J. Struct. Biol. 155:,
94-305.
Merline, R., Schaefer, R.M., Schaefer, L., 2009. The matricellular functions of small
leucine-rich proteoglycans (SLRPs). J. Cell Commun. Signal. 3, 323-335.
Mohan, R.R., Tovey, J.C., Gupta, R., Sharma, A., Tandon, A., 2011. Decorin biology,
expression, function and therapy in the cornea. Curr. Mol. Med. 11, 110-128.

Munger, B.L., Yoshida, Y., Hayashi, S., Osawa, T., Ide, C., 1988. A re-evaluation of the
cytology of cat Pacinian corpuscles. I. The inner core and clefts. Cell Tissue Res. 253,
83–93.
Neill, T., Schaefer, L., Iozzo, R.V., 2015. Decoding the Matrix: Instructive Roles of
Proteoglycan Receptors. Biochemistry. 54, 4583-4598.
Oohira, A., Matsui, F., Tokita, Y., Yamauchi, S., Aono, S., 2000. Molecular interactions of
neural chondroitin sulfate proteoglycans in the brain development. Arch. Biochem.
Biophys. 374, 24-34. Review.
Pawson, L., Slepecky, N.B., Bolanowski, S.J., 2000. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens. Mot. Res. 17, 159-170.
Pawson, L., Checkosky, C.M., Pack, A.K., Bolanowski, S.J., 2008. Mesenteric and tactile
Pacinian corpuscles are anatomically and physiologically comparable. Somatosens.
Mot. Res. 25, 194-206.
Roudaut, Y., Lonigro, A., Coste, B., Hao, J., Delmas, P., Crest, M., 2012. Touchsense:
functional organization and molecular determinants of mechanosensitive
receptors. Channels (Austin) 6, 234–245.
Sames, K., Halata, Z., Jojovic, M., van Damme, E.J., Peumans, W.J., Delpech, B., Asmus,
B., Schumacher, U., 2001. Lectin and proteoglycan histochemistry of feline pacinian
corpuscles. J. Histochem. Cytochem. 49, 19-28.
Schaefer, L., Iozzo, R.V., 2008. Biological functions of the small leucine-rich
proteoglycans: from genetics to signal transduction. J. Biol. Chem. 283, 21305-
21309.
Schaefer, L., Schaefer, R.M., 2010. Proteoglycans: from structural compounds to
signaling molecules. Cell Tissue Res. 339, 237-246.

Silver, D.J., Silver, J., 2014. Contributions of chondroitin sulfate proteoglycans to
neurodevelopment, injury, and cancer. Curr. Opin. Neurobiol. 27, 171-178.
Soleman, S., Filippov, M.A., Dityatev, A., Fawcett, J.W., 2013. Targeting the neural
extracellular matrix in neurological disorders. Neuroscience. 253, 194-213.
Stichel, C.C., Kappler, J., Junghans, U., Koops, A., Kresse, H., Müller, H.W., 1995.
Differential expression of the small chondroitin/dermatan sulfate proteoglycans
decorin and biglycan after injury of the adult rat brain. Brain Res. 704, 263-274.
Svensson, L., Aszódi, A., Reinholt, F.P., Fässler, R., Heinegård, D., Oldberg, A., 1999.
Fibromodulin-null mice have abnormal collagen fibrils, tissue organization, and
altered lumican deposition in tendon. J. Biol. Chem. 274, 9636-9647.
Tachibana, T., Iida, S., Nawa, T., 1995. Type-VI collagen as a major component in the
interstitial matrix of cutaneous mechanoreceptors. Arch. Histol. Cytol. 58, 507-515.
Tashima, T., Nagatoishi, S., Sagara, H., Ohnuma, S., Tsumoto, K., 2015. Osteomodulin
regulates diameter and alters shape of collagen fibrils. Biochem. Biophys. Res.
Commun. 463, 292-296.
Theocharis, A.D., Skandalis, S.S., Gialeli, C., Karamanos, N.K., 2016. Extracellular matrix
structure. Adv. Drug. Deliv. Rev. 97, 4-27
Vega, J.A., Vazquez, E., Naves, F.J., Calzada, B., del Valle, M.E., Represa, J.J., 1994.
Expression of epidermal growth factor receptor (EGFr) immunoreactivity in human
cutaneous nerves and sensory corpuscles. Anat. Rec. 240, 125-130.
Vega, J.A., Esteban, I., Naves, F.J., del Valle, M.E., Malinovsky, L., 1995.
Immunohistochemical localization of laminin and type IV collagen in human
cutaneous sensory nerve formations. Anat. Embryol. (Berl.). 191, 33-39.

Vega, J.A., Haro, J.J., Del Valle, M.E., 1996. Immunohistochemistry of human cutaneous
Meissner and Pacinian corpuscles. Microsc. Res. Tech. 34, 351–361.
Vega, J.A., García-Suárez, O., Montaño, J.A., Pardo, B., Cobo, J.M., 2009. The Meissner
and Pacinian sensory corpuscles revisited: new data from the last decade. Microsc.
Res. Tech. 72, 299–309.
Wilda, M., Bächner, D., Just, W., Geerkens, C., Kraus, P., Vogel, W., Hameister, H., 2000.
A comparison of the expression pattern of five genes of the family of small leucine-
rich proteoglycans during mouse development. J. Bone Miner. Res. 15, 2187-2196.
Zelená, J., 1994. Nerves and Mechanoreceptors. Chapman & Hall, London.

LEGENDS FOR FIGURES
Figure 1.- Serial sections (a-e) of a human digital Pacinian corpuscle showing the
immunohistochemical distribution of BGN (d) and DCN (e). Figures a-c correspond to
immunodetection of VIM, CD34 and S100P respectively. DCN was mainly found in
outer-core cells and capsule, in a thin layer between both inner and outer cores and in
the S100P-positive inner core (e). BGN was expressed in all the compartments of
Pacinian corpuscles except the axon and the intermediate layer (d). Negative control (f).
ic: inner core; oc-c: outer core-capsula; il: intermediate layer. Scale bar: 100 µm, a to f.
Objective: 20X, a to f.
Figure 2.- Double immunofluorescence in Pacinian corpuscles showing the simultaneous
localization of DCN in red (a, d and g) with S100P (b), CD34 (e) and VIM (h) in green.
Although DCN did not colocalize with S100P (a-c), CD34 (d-f) or VIM (g-i), it was found in
both inner and outer cores. In the inner core, DCN was localized around the Schwann-
related cells (a-c), and sometimes its immunostaining was weaker (d). Moreover, a close
association between DCN and CD34 or VIM was observed, especially at the periphery of
the inner core (d-f) and in the outer core (g-i), respectively. Scale bar: 20 µm, a to f; 40
µm, g to i. Objective: 63X/1.40 oil, pinhole 1.37, XY resolution 139.4 nm and Z resolution
235.8 nm, a to f; 40X/1.25 oil, pinhole 1.00, XY resolution 156 nm and Z resolution 334
nm, g to i.
Figure 3.- Double immunofluorescence using anti-BGN antibodies (a, d, g and j) and anti-
S100P (b), Peripherin (e), VIM (h) or CD34 (k) antibodies in Pacinian corpuscles. The
expression of BGN was observed in both the inner core (a-c) and the outer core-capsule
(g-i). Conversely, BGN was not present in the intermediate layer (j-l), but its presence in
the axon can be ruled out (d-f). Scale bar: 20 µm, a to c, g to i, j to l; 10 µm, d to f.

Objective: 63X/1.40 oil, pinhole 1.37, XY resolution 139.4 nm and Z resolution 235.8 nm,
a to f, j to l; 40X, g to i.
Figure 4.- Immunodetection of S100P (a), LUM (b), CD34 (c), FMOD (d), VIM (e) and
OMD (f) in consecutive sections of a Pacinian corpuscle using single
immunohistochemistry. LUM was found in the inner core, the intermediate layer and
the outermost lamellae of the outer core-capsule (b). The expression patterns of FMOD
(d) and OMD (f) were similar, and both proteoglycans were present in the inner core
and outer core. However OMD was detected in the intermediate layer but not in the
capsule, contrary to FMOD. ic: inner core; oc: outer core; c: capsule; il: intermediate
layer. Scale bar: 100 µm, a to f. Objective: 20X, a to f.
Figure 5.- Double immunofluorescence in Pacinian corpuscles for LUM in green (a, d, g
and j) and NFP (b), S100P (e), CD34 (h) or VIM (k) in red. LUM colocalized with S100P in
the inner core (d-f) but not in the axon with NFP (a-c). It was also observed in the outer
core and capsule associated to VIM (j-l), but not in the CD34-positive intermediate layer
(g-i). Scale bar: 20 µm, a to i; 40 µm, j to l. Objective: 63X/1.40 oil, pinhole 1.37, XY
resolution 139.4 nm and Z resolution 235.8 nm, a to i; 40X/1.25 oil, pinhole 1.00, XY
resolution 156 nm and Z resolution 334 nm, j to l.
Figure 6.- Double detection of FMOD in red (a, d, g and j) and PGP9.5 (b), S100P (e),
CD34 (h) or VIM (k) in green by using confocal immunofluorescence in Pacinian
corpuscles. Immunolabeling of FMOD was observed in the inner core but colocalizing
irregularly with S100P (d-f). It was also found in the outer core and capsule, adjacent to
VIM (j-l). On the other hand, FMOD was neither present in the axon (a-c), nor in the
intermediate layer (g-i). Scale bar: 10 µm, a to l. Objective: 63X/1.40 oil, pinhole 1.37, XY
resolution 139.4 nm and Z resolution 235.8 nm, a to l.

Figure 7.- Double immunofluorescence for simultaneous detection of OMD in red (a, d,
g and j) with S100P (b), PGP9.5 (e), CD34 (h) and VIM (k) in green, in Pacinian
corpuscles. OMD was expressed in the axon weakly (d-f), the inner core with an
undefined pattern (a-c), the intermediate layer (g-i) and the outer core (j-l). VIM and
OMD do not colocalized in the outer core, but both molecules were adjacent in every
single lamella (j-l). Scale bar: 10 µm, a to f and g to i; 40 µm, j to l. Objective: 63X/1.40
oil, pinhole 1.37, XY resolution 139.4 nm and Z resolution 235.8 nm, a to I; 40X/1.25 oil,
pinhole 1.00, XY resolution 156 nm and Z resolution 334 nm, j to l.
Figure 8.- Schematic representation of the transverse section of a Pacinian corpuscle
showing the distribution of the analyzed SLRPs in the different corpuscular
compartments, based on the results of double immunofluorescence.
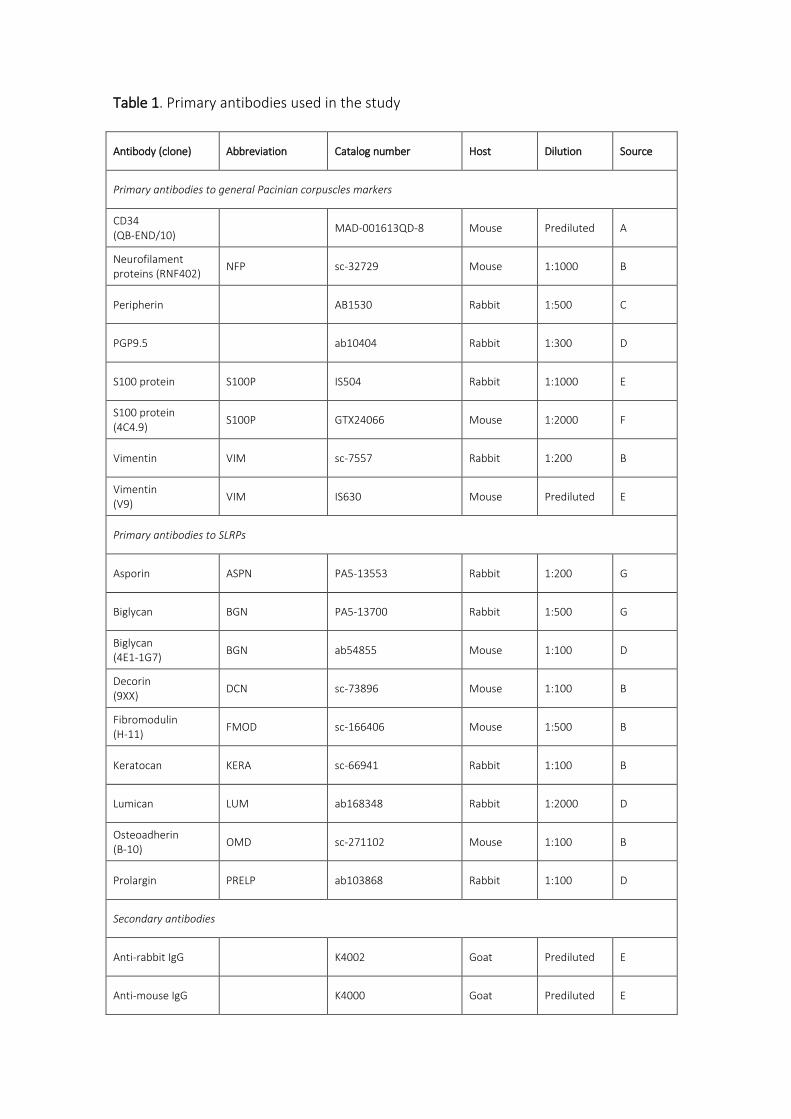
Table 1. Primary antibodies used in the study
Antibody (clone) Abbreviation Catalog number Host Dilution Source
Primary antibodies to general Pacinian corpuscles markers
CD34 (QB-END/10)
MAD-001613QD-8 Mouse Prediluted A
Neurofilament proteins (RNF402)
NFP sc-32729 Mouse 1:1000 B
Peripherin AB1530 Rabbit 1:500 C
PGP9.5 ab10404 Rabbit 1:300 D
S100 protein S100P IS504 Rabbit 1:1000 E
S100 protein (4C4.9)
S100P GTX24066 Mouse 1:2000 F
Vimentin VIM sc-7557 Rabbit 1:200 B
Vimentin (V9)
VIM IS630 Mouse Prediluted E
Primary antibodies to SLRPs
Asporin ASPN PA5-13553 Rabbit 1:200 G
Biglycan BGN PA5-13700 Rabbit 1:500 G
Biglycan (4E1-1G7)
BGN ab54855 Mouse 1:100 D
Decorin (9XX)
DCN sc-73896 Mouse 1:100 B
Fibromodulin (H-11)
FMOD sc-166406 Mouse 1:500 B
Keratocan KERA sc-66941 Rabbit 1:100 B
Lumican LUM ab168348 Rabbit 1:2000 D
Osteoadherin (B-10)
OMD sc-271102 Mouse 1:100 B
Prolargin PRELP ab103868 Rabbit 1:100 D
Secondary antibodies
Anti-rabbit IgG K4002 Goat Prediluted E
Anti-mouse IgG K4000 Goat Prediluted E
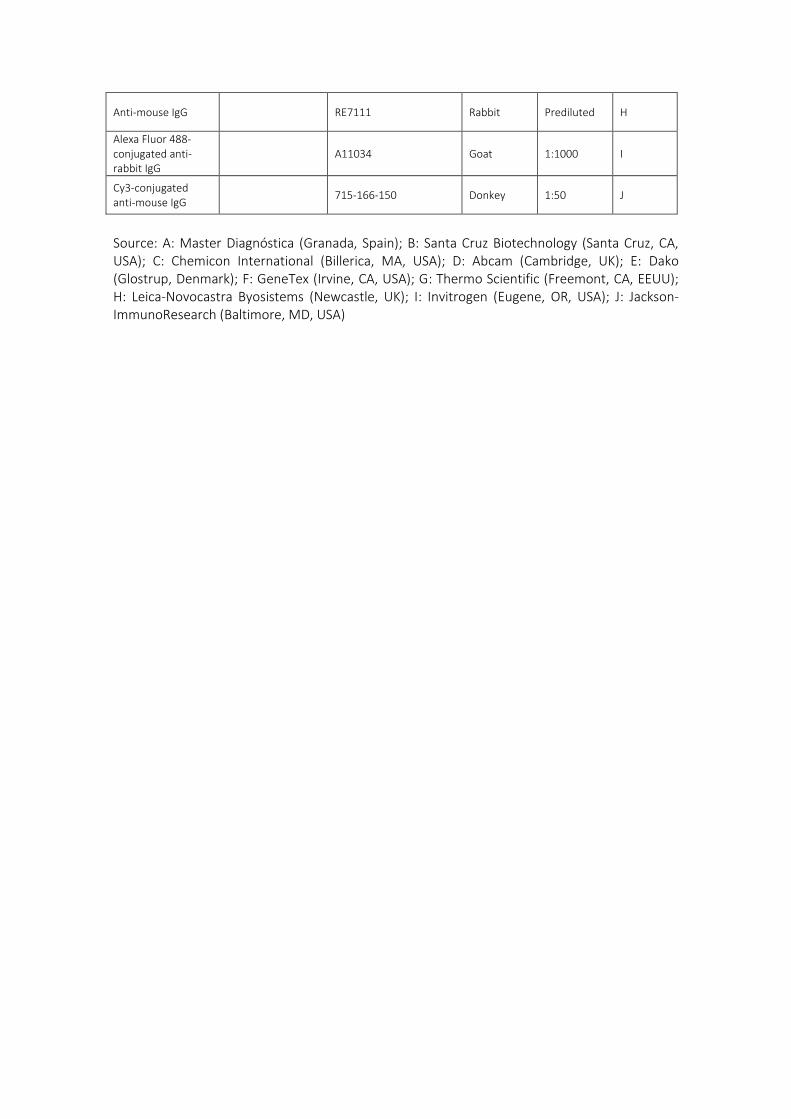
Anti-mouse IgG RE7111 Rabbit Prediluted H
Alexa Fluor 488-conjugated anti-rabbit IgG
A11034 Goat 1:1000 I
Cy3-conjugated anti-mouse IgG
715-166-150 Donkey 1:50 J
Source: A: Master Diagnóstica (Granada, Spain); B: Santa Cruz Biotechnology (Santa Cruz, CA, USA); C: Chemicon International (Billerica, MA, USA); D: Abcam (Cambridge, UK); E: Dako (Glostrup, Denmark); F: GeneTex (Irvine, CA, USA); G: Thermo Scientific (Freemont, CA, EEUU); H: Leica-Novocastra Byosistems (Newcastle, UK); I: Invitrogen (Eugene, OR, USA); J: Jackson-ImmunoResearch (Baltimore, MD, USA)
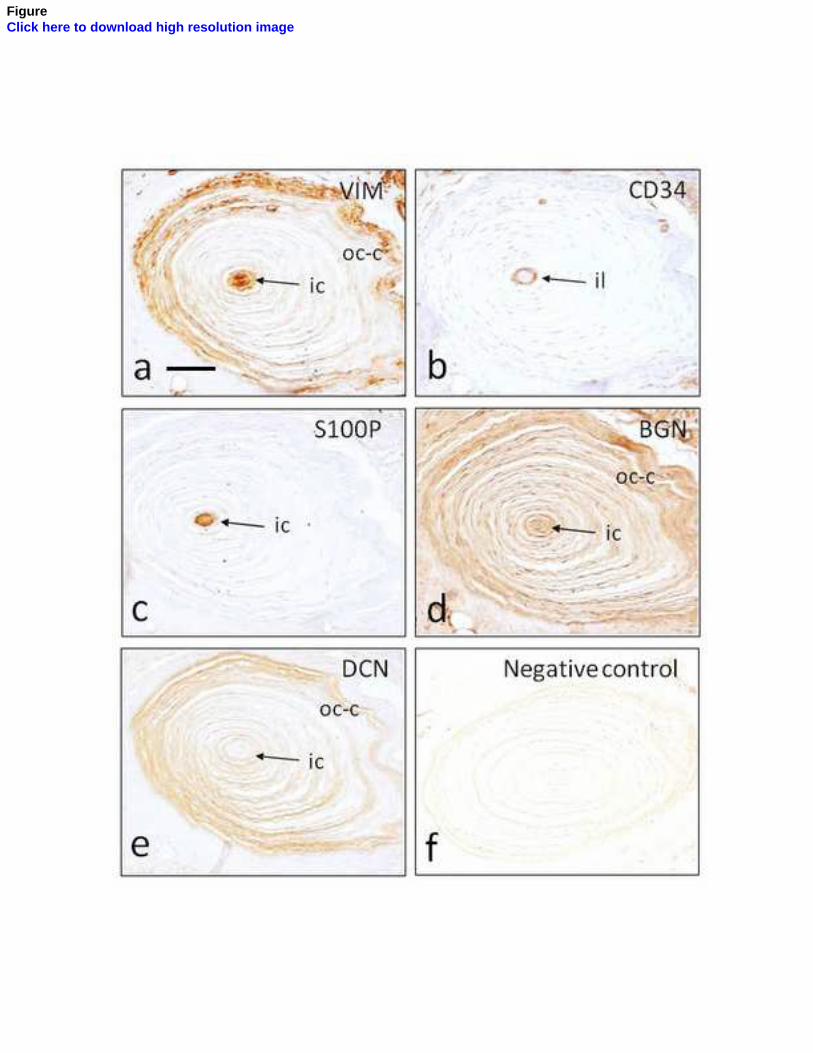
FigureClick here to download high resolution image
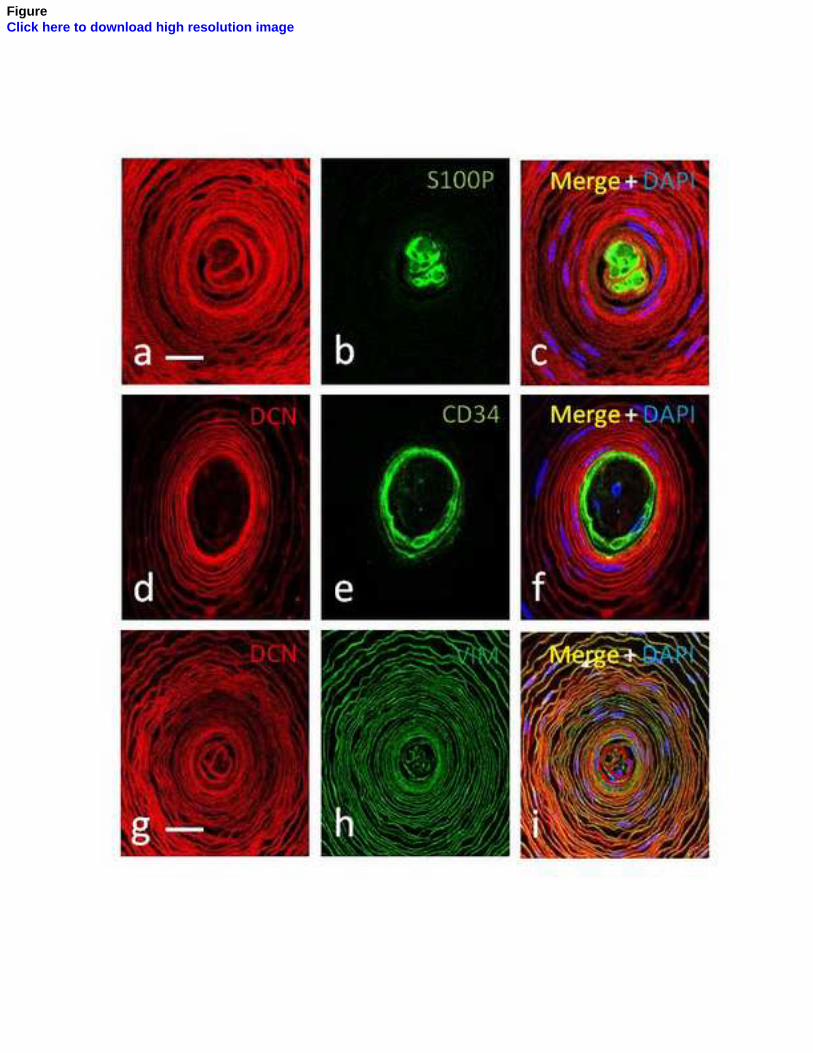
FigureClick here to download high resolution image
FigureClick here to download high resolution image
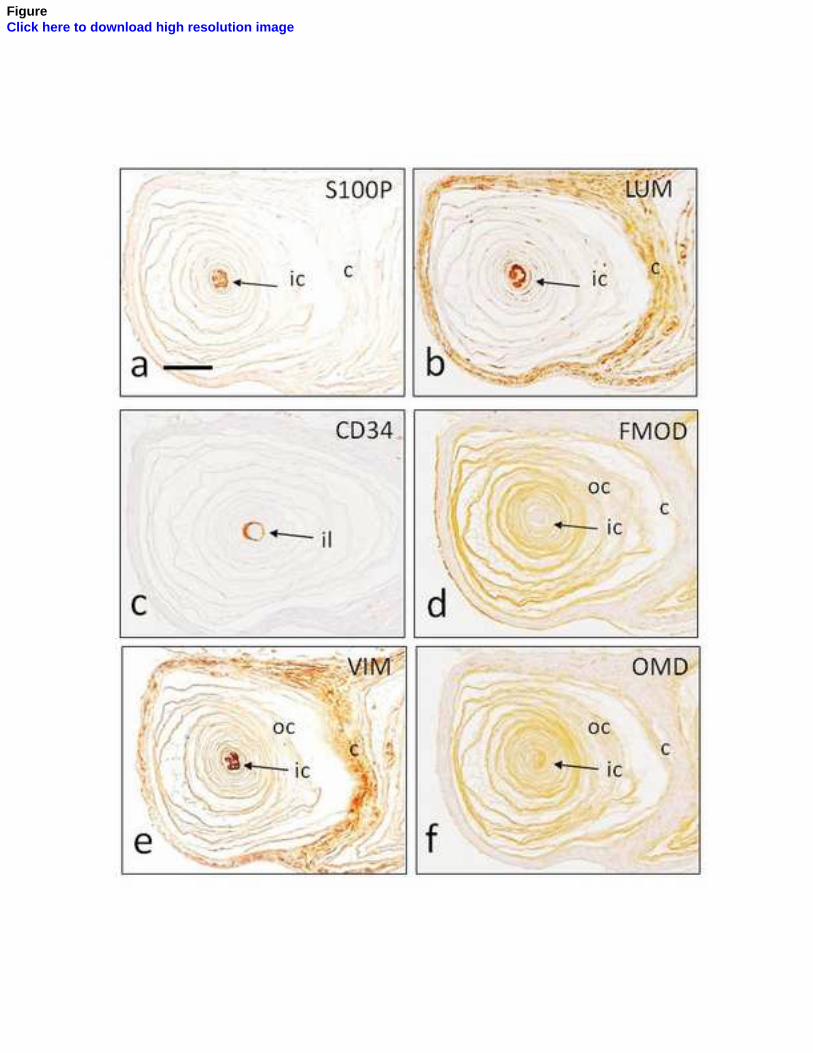
FigureClick here to download high resolution image
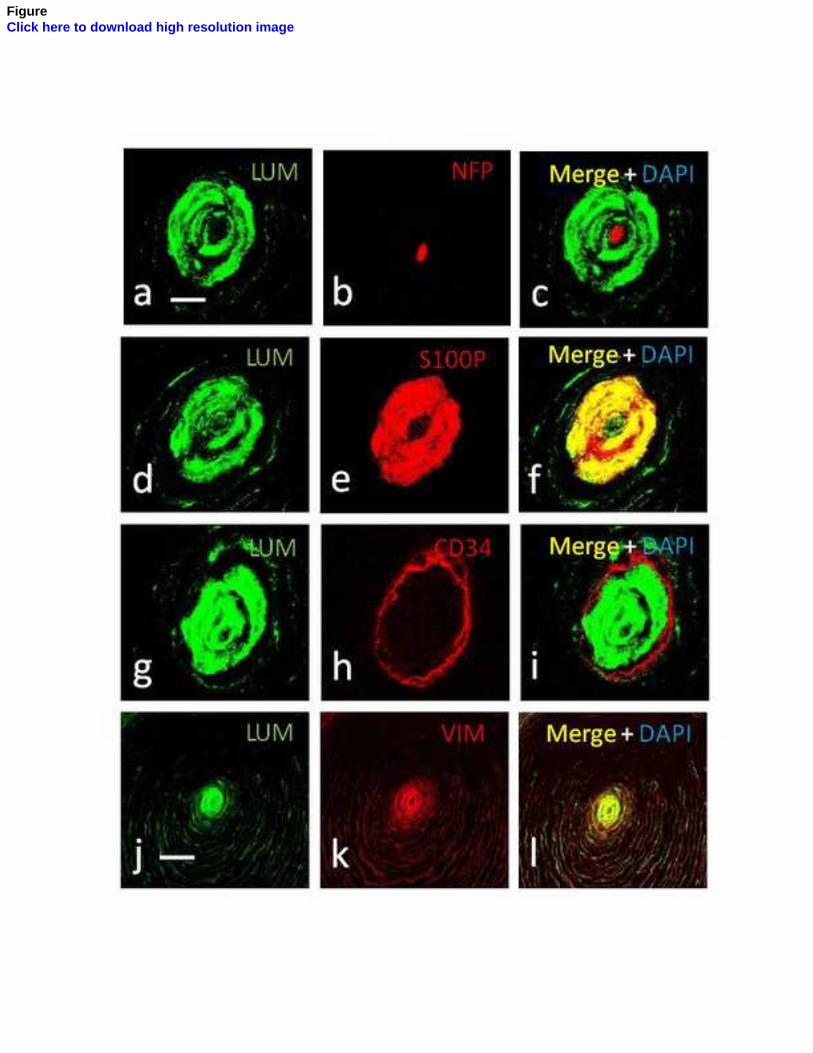
FigureClick here to download high resolution image

FigureClick here to download high resolution image
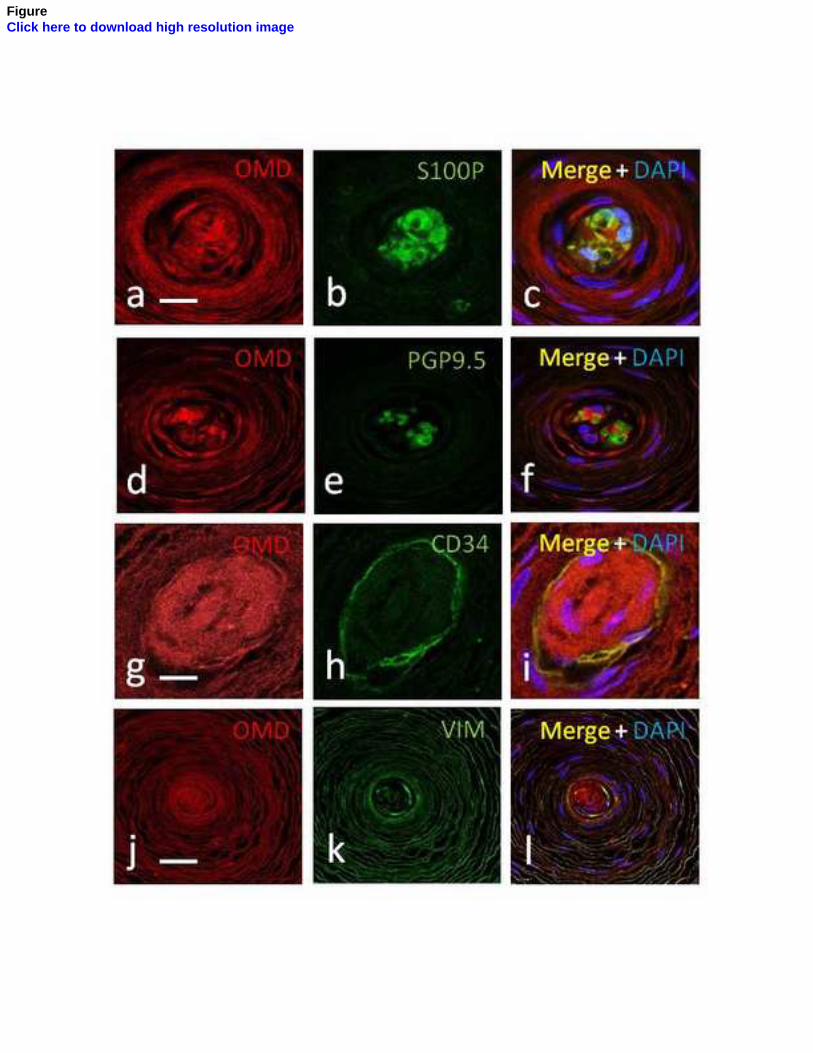
FigureClick here to download high resolution image
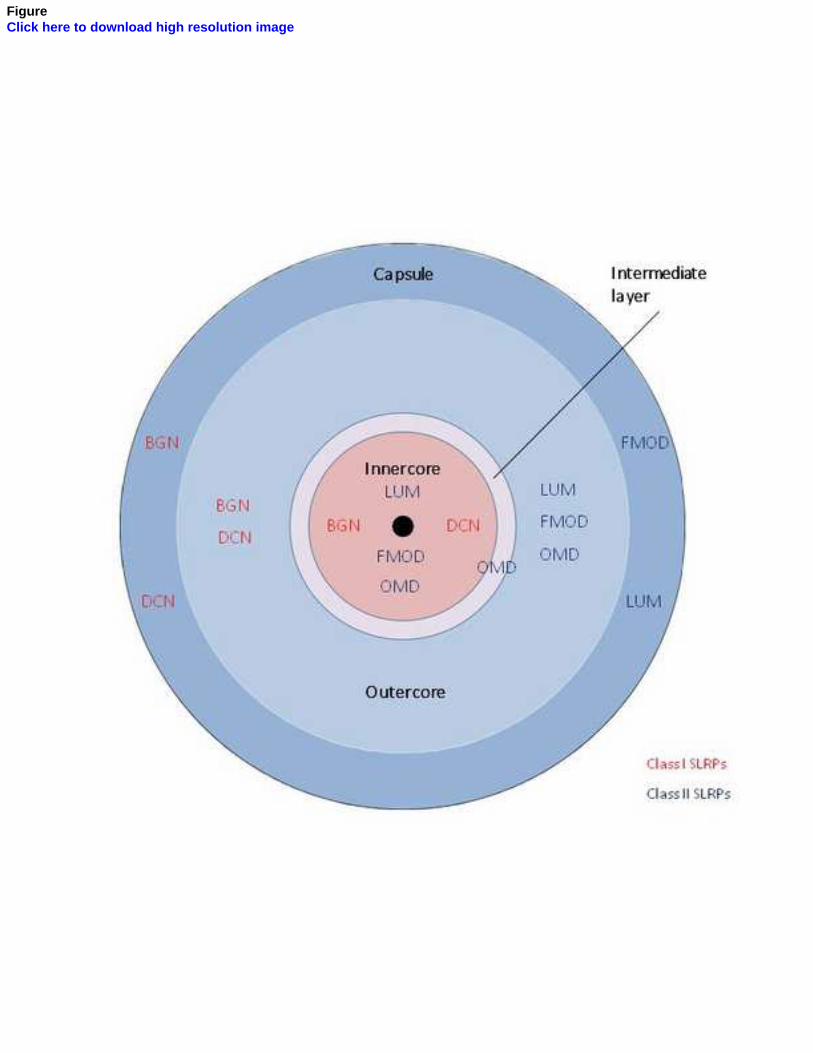
FigureClick here to download high resolution image

December 21, 2018
Dear Editor,
Manuscript: “CLASS I AND CLASS II SMALL LEUCINE-RICH PROTEOGLYCANS IN HUMAN
CUTANEOUS PACINIAN CORPUSCLES”, by J. García-Piqueas et al.
Ethical statement: this study was performed in compliance with Spanish Law and the
guidelines of the Helsinki Declaration II, and the study was approved by the Ethical
Committee for Biomedical Research of the Principality of Asturias, Spain (Ref. 266/18).
*ethical statement

Chapter
The Cutaneous Biopsy for theDiagnosis of PeripheralNeuropathies: Meissner’sCorpuscles and Merkel’s CellsOlivia García-Suárez, Yolanda García-Mesa,Jorge García-Piqueras, Giuseppina Salvo, Juan L. Cobo,Elda Alba, Ramón Cobo, Jorge Feito and José A. Vega
Abstract
Cutaneous biopsy is a complementary method, alternative to peripheral nervebiopsy, for the analysis of nerve involvement in peripheral neuropathies, systemicdiseases, and several pathologies of the central nervous system. Most of theseneuropathological studies were focused on the intraepithelial nerve fibers (thin-myelinated Aδ fibers and unmyelinated C fibers), and few studies investigated thevariations in dermal innervation, that is, large myelinated fibers, Merkel’s cell-neurite complexes, and Meissner’s corpuscles. Here, we updated and summarizedthe current data about the quantitative and qualitative changes that undergo MCsand MkCs in peripheral neuropathies. Moreover, we provide a comprehensiverationale to include MCs in the study of cutaneous biopsies when analyzing theperipheral neuropathies and aim to provide a protocol to study them.
Keywords: skin biopsy, peripheral neuropathy, Meissner’s corpuscles,Merkel’s cells
1. Introduction
Since the last half of the past century, the analysis in the cutaneous biopsy ofnerves, Merkel’s cells (MkCs), and sensory corpuscles, especially Meissner’s cor-puscles (MCs), become a complementary method to diagnose peripheral neuropa-thies [1] and a reliable alternative to peripheral nerve biopsy. Nevertheless, it hasbeen during the last decade that numerous studies have provided consistent evi-dence to support this technique as a valuable tool to understand the etiologies ofsome neurological diseases and to follow up clinical trials [2–4] (Figures 1 and 2).
Most of the neuropathological studies on cutaneous biopsies were focused onintraepithelial nerve fibers, which are thin-myelinated Aδ fibers or unmyelinated Cfibers [2, 3, 5–9]. Conversely, few studies have investigated the large myelinated fibers(although it can offer notable advantages over the unmyelinated ones [10]). Also, thequantitative and qualitatively changes in MCs and MkCs associated to peripheralneuropathies are poorly known although the study of MCs has gained interest [11–13].
1
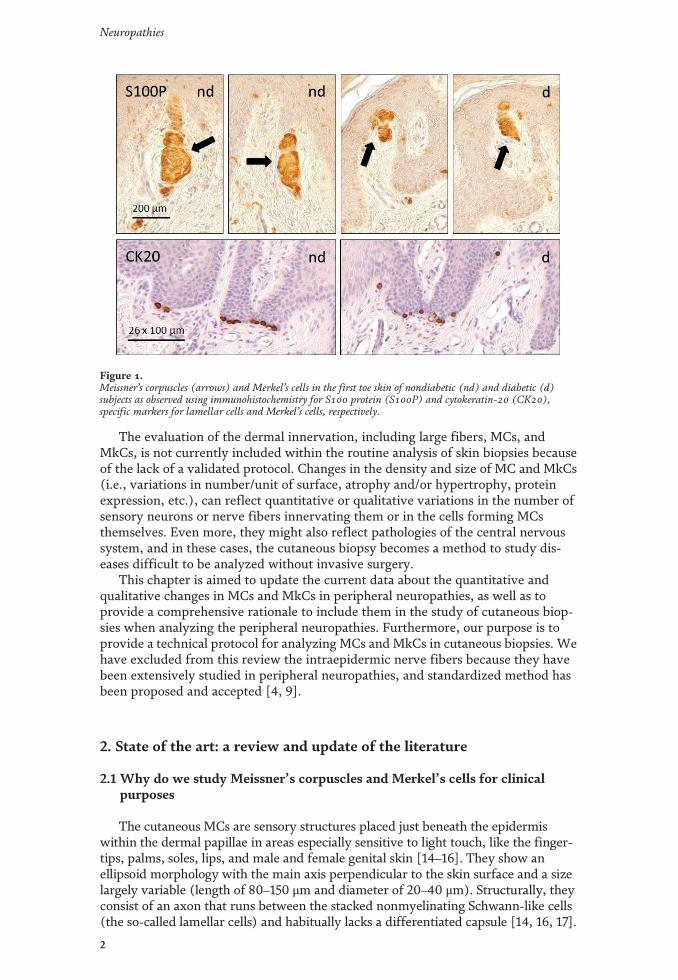
The evaluation of the dermal innervation, including large fibers, MCs, andMkCs, is not currently included within the routine analysis of skin biopsies becauseof the lack of a validated protocol. Changes in the density and size of MC and MkCs(i.e., variations in number/unit of surface, atrophy and/or hypertrophy, proteinexpression, etc.), can reflect quantitative or qualitative variations in the number ofsensory neurons or nerve fibers innervating them or in the cells forming MCsthemselves. Even more, they might also reflect pathologies of the central nervoussystem, and in these cases, the cutaneous biopsy becomes a method to study dis-eases difficult to be analyzed without invasive surgery.
This chapter is aimed to update the current data about the quantitative andqualitative changes in MCs and MkCs in peripheral neuropathies, as well as toprovide a comprehensive rationale to include them in the study of cutaneous biop-sies when analyzing the peripheral neuropathies. Furthermore, our purpose is toprovide a technical protocol for analyzing MCs and MkCs in cutaneous biopsies. Wehave excluded from this review the intraepidermic nerve fibers because they havebeen extensively studied in peripheral neuropathies, and standardized method hasbeen proposed and accepted [4, 9].
2. State of the art: a review and update of the literature
2.1 Why do we study Meissner’s corpuscles and Merkel’s cells for clinicalpurposes
The cutaneous MCs are sensory structures placed just beneath the epidermiswithin the dermal papillae in areas especially sensitive to light touch, like the finger-tips, palms, soles, lips, and male and female genital skin [14–16]. They show anellipsoid morphology with the main axis perpendicular to the skin surface and a sizelargely variable (length of 80–150 μm and diameter of 20–40 μm). Structurally, theyconsist of an axon that runs between the stacked nonmyelinating Schwann-like cells(the so-called lamellar cells) and habitually lacks a differentiated capsule [14, 16, 17].
Figure 1.Meissner’s corpuscles (arrows) and Merkel’s cells in the first toe skin of nondiabetic (nd) and diabetic (d)subjects as observed using immunohistochemistry for S100 protein (S100P) and cytokeratin-20 (CK20),specific markers for lamellar cells and Merkel’s cells, respectively.
2
Neuropathies
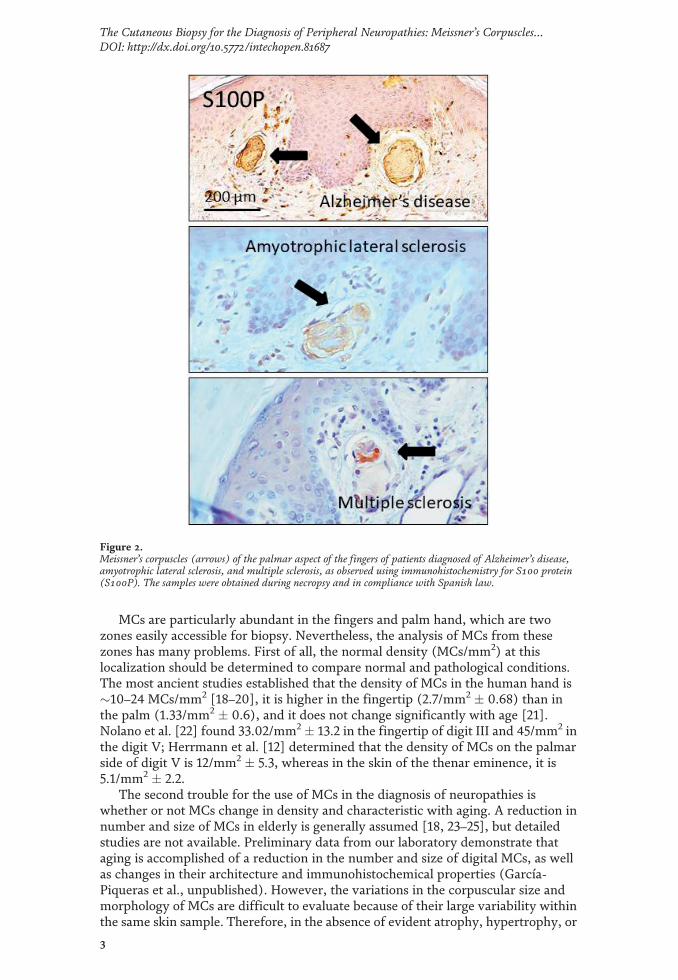
MCs are particularly abundant in the fingers and palm hand, which are twozones easily accessible for biopsy. Nevertheless, the analysis of MCs from thesezones has many problems. First of all, the normal density (MCs/mm2) at thislocalization should be determined to compare normal and pathological conditions.The most ancient studies established that the density of MCs in the human hand is�10–24 MCs/mm2 [18–20], it is higher in the fingertip (2.7/mm2 � 0.68) than inthe palm (1.33/mm2 � 0.6), and it does not change significantly with age [21].Nolano et al. [22] found 33.02/mm2 � 13.2 in the fingertip of digit III and 45/mm2 inthe digit V; Herrmann et al. [12] determined that the density of MCs on the palmarside of digit V is 12/mm2 � 5.3, whereas in the skin of the thenar eminence, it is5.1/mm2 � 2.2.
The second trouble for the use of MCs in the diagnosis of neuropathies iswhether or not MCs change in density and characteristic with aging. A reduction innumber and size of MCs in elderly is generally assumed [18, 23–25], but detailedstudies are not available. Preliminary data from our laboratory demonstrate thataging is accomplished of a reduction in the number and size of digital MCs, as wellas changes in their architecture and immunohistochemical properties (García-Piqueras et al., unpublished). However, the variations in the corpuscular size andmorphology of MCs are difficult to evaluate because of their large variability withinthe same skin sample. Therefore, in the absence of evident atrophy, hypertrophy, or
Figure 2.Meissner’s corpuscles (arrows) of the palmar aspect of the fingers of patients diagnosed of Alzheimer’s disease,amyotrophic lateral sclerosis, and multiple sclerosis, as observed using immunohistochemistry for S100 protein(S100P). The samples were obtained during necropsy and in compliance with Spanish law.
3
The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles…DOI: http://dx.doi.org/10.5772/intechopen.81687

corpuscular disruption, the evaluation of these parameters must be cautiously con-sidered when evaluating cutaneous biopsies.
The main constituents of MCs, that is, the axon and lamellar cells, contain specificproteins as widely demonstrated using immunohistochemistry [17, 26, 27]. Thesestudies reported a large volume of information, but they are purely descriptive and donot consent to quantify those proteins and their possible variations in neuropathies.The central axon displays immunoreactivity for general neuronal markers (neuron-specific enolase, protein gene product 9.5, neurofilament subunit proteins). They alsoexpress Ca2+-binding proteins such as calbindin D28k, parvalbumin, calretinin, andneurocalcin, which presumably regulate the axonic Ca2+ homeostasis and thereforeparticipate in the mechanoelectric transduction. Recently, our research’s groupdetected axonic TRPC6, TRPV4, ASIC2, and Piezo2 ion channels that work as puta-tive mechanoproteins [28–30]. Regarding lamellar cells, the vimentin is the interme-diate filament filling their cytoplasm, while the glial fibrillary acidic protein is alwaysabsent. They strongly express S100 protein colocated with parvalbumin or calbindinD-28 kDa. The lamellar cells also display immunoreactivity for TrkB, the signalingreceptor for the neurotrophins BDNF/NT-4 [31]. Apart from axon- or lamellar cell-specific proteins, there are some others shared by both corpuscular constituents. Theyinclude p75NTR and TrkA (low-affinity pan-neurotrophin receptor and the high-affinity receptor for nerve growth factor, respectively; [32, 33]), the epidermalgrowth factor receptor [34], or cell death protein Bcl-2 [35]. The presence of some ionchannels in the lamellar cells has been also reported [28–30]. It is possible that someof these proteins undergo changes during peripheral neuropathies, but limited infor-mation is so far available in this topic (see [17]). The proteins present in human MCsare summarized in Table 1.
The cutaneous MkCs are special epidermal cells placed in the basal layer of theepidermis, isolated or forming clusters, in both the glabrous and hairy skin. Theyare innervated by Aβ sensory axons connected through synapse-like contactsforming the so-called MkCs-neurite complexes. MkCs are involved in fine touchworking as a part of slowly adapting type I low-threshold mechanoreceptors andexpress specific mechanoproteins [16, 30, 36–39]. MkCs have an epithelial originand do not originate from the neural crest, as classically accepted [40–42].
Using immunohistochemistry, diverse proteins have been detected in the MkC-neurite complexes. They include low-molecular-weight cytokeratins and a reper-tory of synaptic vesicles-related proteins (chromogranin A, synaptophysin), differ-ent neuropeptides as well as neurotransmitter receptors, neurotrophin receptors,ion channels (ASIC2 and Piezo2), and neuron-specific enolase [28, 43–46]. Theaxon of the MkC-neurite complexes displays immunoreactivity for general neuronalmarkers (Table 1).
The density of MCs varies from an anatomical region to another, and it is directlyrelated to the sensibility of those zones [47]. In terms of density as far as we know, noage-dependent changes have been communicated. Recently, we have found signifi-cant reduction in of digital MkCs with aging (García-Piqueras et al., unpublished).On the other hand, whether or not MkCs, or the nerve fibers innervating them, areinvolved in peripheral neuropathies has been poorly studied, but this possibilityshould be explored because the easily accessibility to MkCs-neurite complexes.
2.2 Variations in MCs and MKCs in peripheral neuropathies
Data reporting changes in MCs in peripheral neuropathies are scarce and arerestricted to diabetes and other rare inheretary neuropathies, HIV infection,mechanical or traumatic nerve entrapment, and a miscellaneous group of systemicdiseases with neurological symptoms.
4
Neuropathies
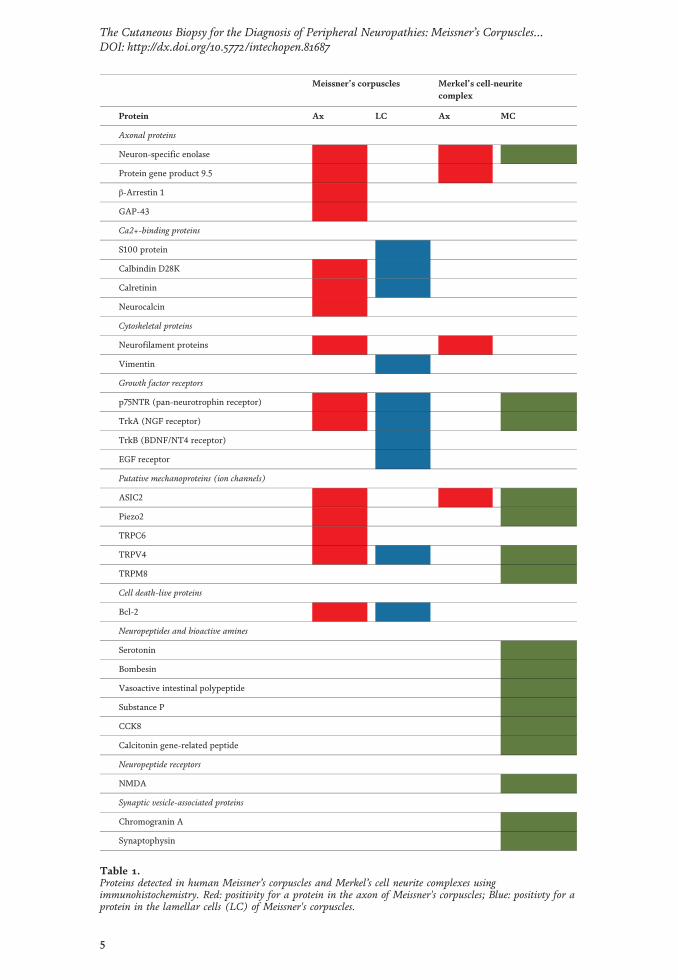
Meissner’s corpuscles Merkel’s cell-neuritecomplex
Protein Ax LC Ax MC
Axonal proteins
Neuron-specific enolase
Protein gene product 9.5
β-Arrestin 1
GAP-43
Ca2+-binding proteins
S100 protein
Calbindin D28K
Calretinin
Neurocalcin
Cytoskeletal proteins
Neurofilament proteins
Vimentin
Growth factor receptors
p75NTR (pan-neurotrophin receptor)
TrkA (NGF receptor)
TrkB (BDNF/NT4 receptor)
EGF receptor
Putative mechanoproteins (ion channels)
ASIC2
Piezo2
TRPC6
TRPV4
TRPM8
Cell death-live proteins
Bcl-2
Neuropeptides and bioactive amines
Serotonin
Bombesin
Vasoactive intestinal polypeptide
Substance P
CCK8
Calcitonin gene-related peptide
Neuropeptide receptors
NMDA
Synaptic vesicle-associated proteins
Chromogranin A
Synaptophysin
Table 1.Proteins detected in human Meissner’s corpuscles and Merkel’s cell neurite complexes usingimmunohistochemistry. Red: positivity for a protein in the axon of Meissner's corpuscles; Blue: positivty for aprotein in the lamellar cells (LC) of Meissner's corpuscles.
5
The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles…DOI: http://dx.doi.org/10.5772/intechopen.81687

2.2.1 Diabetic neuropathy
Distal symmetric peripheral neuropathy is one of the most common complica-tions of diabetes [48] and involves motor, autonomic, and sensory nerve fibers. Thehistopathological studies have provided evidence that both the thin unmyelinated Cfibers and the large myelinated ones are affected in on diabetic neuropathy. Con-sistently, the two most prominent complaints are peripheral pain and changes intouch [13, 49–52]. The intraepidermic nerve fibers as well as the nerve apparatus ofthe dermis are reduced in the diabetic neuropathy, and the reduction of the dermicnerves involves MCs. Importantly, although some authors have argued their inter-est in studying MCs and MkCs to better understand the diabetic neuropathy [53],only few studies have approached this topic.
In cutaneous biopsies, it was shown that the density of MCs is significantlyreduced in diabetic patients with respect to the controls (10.2 � 8.4 vs.16.2� 9.4/mm2, more evidently in type I than in type II diabetes), and this correlatedwith a reduction inmedian and ulnar nerves sensory amplitude; moreover, someMCswere hypertrophic or showed anomalies in their architecture (disorganization of thelamellar cells and increase in the irregularity of the axons) [54]. Similar findings asthose obtained from cutaneous biopsy were observed using in vivo reflectance confo-cal microscopy at the thenar eminence and digit V [55]. We have recently communi-cated that long-term diabetic neuropathy courses with a reduction in the numberand size of MCs and changes in their immunohistochemical profile [56] (Figure 1).
Nevertheless, the number and size of MCs are probably related with the time ofevolution of the neuropathy. In fact, in an animal model of diabetes that developneuropathy, MCs were found more abundant and hypertrophic during the first fewyears of hyperglycemia, whereas after a long time, the hypertrophy declines but thenumber of corpuscles remained higher than in age-matched nondiabetic subjects;furthermore, the MCs from the diabetic animals found had abnormal structure andimmunochemistry properties [57].
On the other hand, as far as we know, the only study reporting a reduction in thenumber of immunohistochemically demonstrable MkCs in diabetic neuropathy wasfrom our laboratory [56].
2.2.2 Charcot-Marie-Tooth disease
Charcot-Marie-Tooth (CMT) disease is a common inherited neuromuscular dis-order characterized by neuropathies without known metabolic alterations. In theskin of patients with common and rare forms of CMT caused by different muta-tions, the density of MCs is reduced compared with normal controls [58–60].Similar findings were reported by Almodovar et al. [61] using in vivo reflectanceconfocal microscopy.
2.2.3 Human immunodeficiency virus (HIV) neuropathy
HIV-sensory neuropathy is a common complication of HIV infection and may beassociated with significant morbidity due to neuropathic pain [62]. Severalapproaches exist for quantitative assessment of human HIV-associated distal sen-sory polyneuropathy, and some of them have analyzed both unmyelinated andmyelinated nerve fibers, as well as MCs. Using in vivo reflectance confocal micros-copy, it was found a marked reduction in MCs [12, 63] in HIV+ subjects with andwithout distal sensory neuropathy [64].
6
Neuropathies

2.2.4 Entrapment neuropathies
Surprisingly, little is known about the impact of entrapment neuropathy ontarget innervation. More than 20 years ago, we reported that human digital MCssurvive to entrapment or section of peripheral nerves for more than 10 years, andalthough its number remains relatively stable, denervated MCs lack some antigensor change the pattern of expression of some others [65–67]. These data were con-firmed recently in subjects undergoing carpal tunnel syndrome [68].
2.2.5 Miscellaneous
A reduction in density or loss of MCs has also been reported in the skin ofpatients suffering from Ross syndrome (a rare disorder of sweating associated withareflexia and tonic pupil) [69], POEMS syndrome (polyneuropathy, organomegaly,endocrinopathy, monoclonal gammopathy, and skin changes) [70], systemic scle-rosis [71], pachyonychia congenita (in contrast, MkC densities are higher) [72],chronic inflammatory demyelinating polyradiculoneuropathy [73], and systemiclupus erythematosus [12].
2.3 MCs are also altered in central nervous system disorders
In addition to the abovementioned peripheral neuropathies, changes in MCshave been reported in Parkinson’s disease associated or not with dementia [74–76],spinobulbar muscular atrophy [77], Friedreich’s ataxia [78], amyotrophic lateralsclerosis [79], or Guillain-Barré syndrome [73]. Furthermore, altered cutaneousinnervation also has been observed in some psychiatric disorders [80] and mentaldeficiencies [81] (Figure 2).
3. Proposal of a method to systematically study MCs and MkCs incutaneous biopsies
MCs are only present in glabrous skin, and therefore fingers or toes are appro-priate regions to take cutaneous biopsies focused to evaluate them; in spite of thediscrepancies regarding their density in these places, they are abundant enough.
In our opinion, the palmar aspect of fingertip IV would be an ideal region to bebiopsied, because it is not involved in handling; the lateral borders should beexcluded to avoid damaging the digital nerves and the formation of neuromas. Onthe other hand, toe pad biopsies can be also useful, but they contain a lower densityof MCs than fingers [82].
The Joint Task Force EFNS/PNS [9] recommends to perform a 3 mm punch skinbiopsy (including epidermis and the subpapillary and reticular dermis), using asterile technique and under local anesthesia. A sample of these dimensions does notneed sutures, heal completely within 1 week, and this normally guaranties no sideeffects or complaints. Informed consent is required, and information on the possiblerisks must be always provided. The fixation of the skin samples is recommended in2% PLP (2% paraformaldehyde, 0.075 M lysine, 0.037 M sodium phosphate, 0.01 Mperiodate) or Zamboni’s solution. We have also obtained excellent structural resultsand good antigen preservation using Bouin’s fixative and buffered 10% formalde-hyde. Conversely, 4% paraformaldehyde masked most of the antigens present inMCs. The thickness of the sections is also important. The Joint Task Force EFNS/
7
The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles…DOI: http://dx.doi.org/10.5772/intechopen.81687
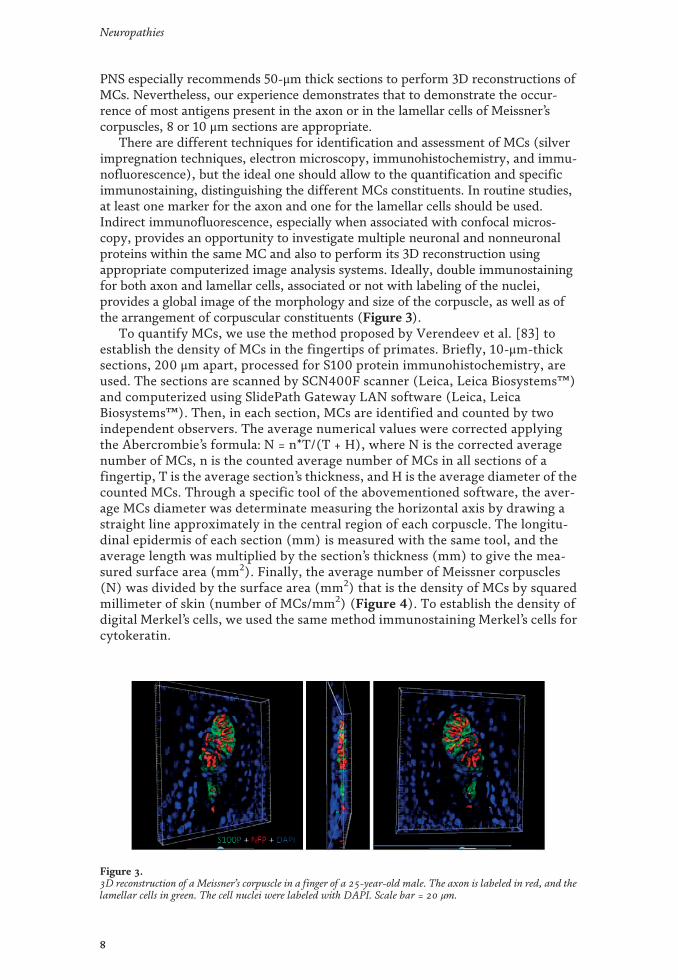
PNS especially recommends 50-μm thick sections to perform 3D reconstructions ofMCs. Nevertheless, our experience demonstrates that to demonstrate the occur-rence of most antigens present in the axon or in the lamellar cells of Meissner’scorpuscles, 8 or 10 μm sections are appropriate.
There are different techniques for identification and assessment of MCs (silverimpregnation techniques, electron microscopy, immunohistochemistry, and immu-nofluorescence), but the ideal one should allow to the quantification and specificimmunostaining, distinguishing the different MCs constituents. In routine studies,at least one marker for the axon and one for the lamellar cells should be used.Indirect immunofluorescence, especially when associated with confocal micros-copy, provides an opportunity to investigate multiple neuronal and nonneuronalproteins within the same MC and also to perform its 3D reconstruction usingappropriate computerized image analysis systems. Ideally, double immunostainingfor both axon and lamellar cells, associated or not with labeling of the nuclei,provides a global image of the morphology and size of the corpuscle, as well as ofthe arrangement of corpuscular constituents (Figure 3).
To quantify MCs, we use the method proposed by Verendeev et al. [83] toestablish the density of MCs in the fingertips of primates. Briefly, 10-μm-thicksections, 200 μm apart, processed for S100 protein immunohistochemistry, areused. The sections are scanned by SCN400F scanner (Leica, Leica Biosystems™)and computerized using SlidePath Gateway LAN software (Leica, LeicaBiosystems™). Then, in each section, MCs are identified and counted by twoindependent observers. The average numerical values were corrected applyingthe Abercrombie’s formula: N = n*T/(T + H), where N is the corrected averagenumber of MCs, n is the counted average number of MCs in all sections of afingertip, T is the average section’s thickness, and H is the average diameter of thecounted MCs. Through a specific tool of the abovementioned software, the aver-age MCs diameter was determinate measuring the horizontal axis by drawing astraight line approximately in the central region of each corpuscle. The longitu-dinal epidermis of each section (mm) is measured with the same tool, and theaverage length was multiplied by the section’s thickness (mm) to give the mea-sured surface area (mm2). Finally, the average number of Meissner corpuscles(N) was divided by the surface area (mm2) that is the density of MCs by squaredmillimeter of skin (number of MCs/mm2) (Figure 4). To establish the density ofdigital Merkel’s cells, we used the same method immunostaining Merkel’s cells forcytokeratin.
Figure 3.3D reconstruction of a Meissner’s corpuscle in a finger of a 25-year-old male. The axon is labeled in red, and thelamellar cells in green. The cell nuclei were labeled with DAPI. Scale bar = 20 μm.
8
Neuropathies
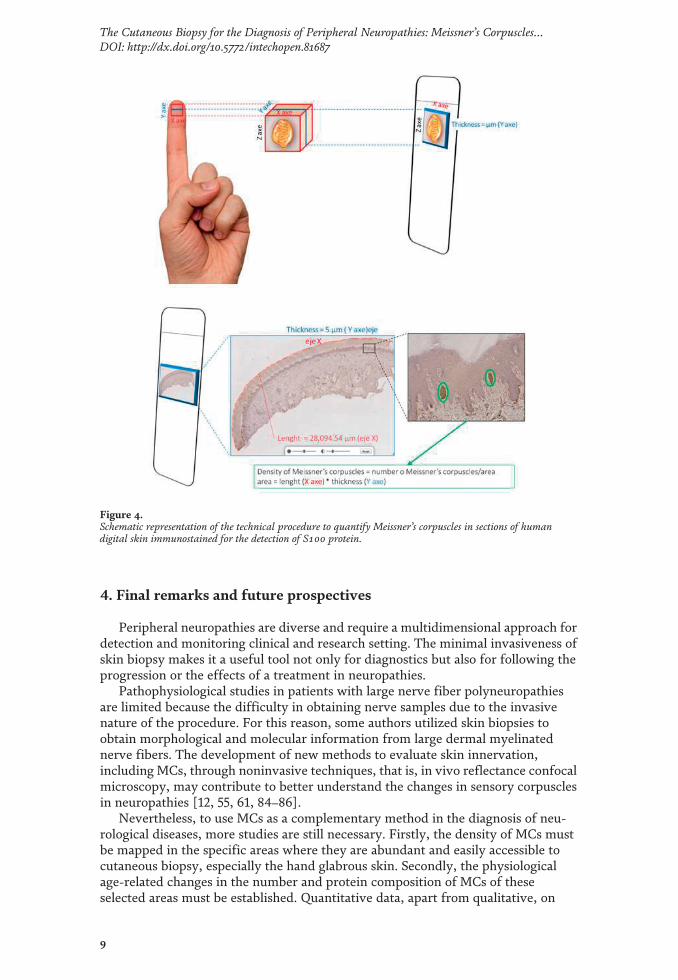
4. Final remarks and future prospectives
Peripheral neuropathies are diverse and require a multidimensional approach fordetection and monitoring clinical and research setting. The minimal invasiveness ofskin biopsy makes it a useful tool not only for diagnostics but also for following theprogression or the effects of a treatment in neuropathies.
Pathophysiological studies in patients with large nerve fiber polyneuropathiesare limited because the difficulty in obtaining nerve samples due to the invasivenature of the procedure. For this reason, some authors utilized skin biopsies toobtain morphological and molecular information from large dermal myelinatednerve fibers. The development of new methods to evaluate skin innervation,including MCs, through noninvasive techniques, that is, in vivo reflectance confocalmicroscopy, may contribute to better understand the changes in sensory corpusclesin neuropathies [12, 55, 61, 84–86].
Nevertheless, to use MCs as a complementary method in the diagnosis of neu-rological diseases, more studies are still necessary. Firstly, the density of MCs mustbe mapped in the specific areas where they are abundant and easily accessible tocutaneous biopsy, especially the hand glabrous skin. Secondly, the physiologicalage-related changes in the number and protein composition of MCs of theseselected areas must be established. Quantitative data, apart from qualitative, on
Figure 4.Schematic representation of the technical procedure to quantify Meissner’s corpuscles in sections of humandigital skin immunostained for the detection of S100 protein.
9
The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles…DOI: http://dx.doi.org/10.5772/intechopen.81687

changes in protein composition of MCs with aging are necessary as a baseline forpossible pathological changes. In addition to immunohistochemical studies, skinbiopsy is amenable to the extraction of mRNA, RT-PCR, or microarrays for genesinvolved in neuropathies, and these methods should be used and standardized tostudy MCs. Finally, future studies should include not only neuropathies such asneurofibromatosis [85], or other rare metabolic neuropathies such as Gauchertype 1 disease [86], but also central nervous system diseases such as Alzheimer’sdisease.
Author details
Olivia García-Suárez1, Yolanda García-Mesa1, Jorge García-Piqueras1,Giuseppina Salvo1, Juan L. Cobo1,2, Elda Alba3, Ramón Cobo1, Jorge Feito1,4 andJosé A. Vega1,5*
1 Departamento de Morfología y Biología Celular, Grupo SINPOS, Universidad deOviedo, Spain
2 Servicio de Cirugía Máxilofacial, Hospital Universitario Cental de Astrias, Oviedo,Spain
3 Servicio de Neurología, Hopital Universitario “La Paz”, Madrid, Spain
4 Servicio de Anatomía Patológica, Complejo Hospitalario de Salamanca,Salamanca, Spain
5 Facultad de Ciencias de la Salud, Universidad Autónoma de Chile, Santiago deChile, Chile
*Address all correspondence to: [email protected]
© 2018 The Author(s). Licensee IntechOpen. This chapter is distributed under the termsof the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution, and reproduction in any medium,provided the original work is properly cited.
10
Neuropathies

References
[1] Dyck PJ, Winkelmann RK, BoltonCF. Quantitation of Meissner'scorpuscles in hereditary neurologicdisorders. Charcot-Marie-Tooth disease,Roussy-Levy syndrome, Dejerine-Sottasdisease, hereditary sensory neuropathy,spinocerebellar degenerations, andhereditary spastic paraplegia.Neurology. 1966;16:10-17
[2] Sommer C. Skin biopsy as adiagnostic tool. Current Opinion inNeurology. 2008;21:562-568
[3] Lauria G, Lombardi R. Small fiberneuropathy: Is skin biopsy the holygrail? Current Diabetes Reports. 2012;12:384-392
[4] Mellgren SI, Nolano M, Sommer C.The cutaneous nerve biopsy: Technicalaspects, indications, and contribution.Handbook of Clinical Neurology. 2013;115:171-188
[5] Lauria G, Devigili G. Skin biopsy as adiagnostic tool in peripheralneruropathy. Nature Clinical Practice.Neurology. 2007;3:546-557
[6] Sommer C, Lauria G. Skin biopsy inthe management of peripheralneuropathy. Lancet Neurology. 2007;6:632-642
[7] Lauria G, Lombardi R, Camozzi F,Devigili G. Skin biopsy for the diagnosisof peripheral neuropathy.Histopathology. 2009;54:273-385
[8] Lauria G, Bakkers M, Schmitz C,Lombardi R, Penza P, Devigili G, et al.Intraepidermal nerve fiber density atthe distal leg: A worldwide normativereference study. Journal of thePeripheral Nervous System. 2010;15:202-207
[9] Lauria G, Hsieh ST, Johansson O,Kennedy WR, Leger JM, Mellgren SI,et al. European Federation of
Neurological Societies/Peripheral NerveSociety Guideline on the use of skinbiopsy in the diagnosis of small fiberneuropathy. Report of a joint task forceof the European Federation ofNeurological Societies and the PeripheralNerve Society. European Journal ofNeurology. 2010;17:903-912, e44-9
[10] Saporta MA, de Moraes Maciel R.Analysis of myelinating Schwann cellsin human skin biopsies. Methods inMolecular Biology. 2018;1739:359-369
[11] Dyck PJ. Enumerating Meissnercorpuscles: Futute gold standard of largefiber sensoriomotor polyneuropathy?Neurology. 2007;69:2116-2118
[12] Herrmann DN, Boger JN, Jansen C,Alessi-Fox C. In vivo confocalmicroscopy of Meissner corpuscles as ameasure of sensory neuropathy.Neurology. 2007;69:2121-2127
[13] Myers MI, Peltier AC, Li J.Evaluating dermal myelinated nervefibers in skin biopsy. Muscle & Nerve.2013;47:1-11
[14] Munger BL, Ide C. The structureand function of cutaneous sensoryreceptors. Archives of Histology andCytology. 1988;51:1-34
[15] Zelena J. Nerves andMechanoreceptors. London: Chapman& Hall; 1994
[16] Zimmerman A, Bai L, Ginty DD.The gentle touch receptors ofmammalian skin. Science. 2014;346:950-954
[17] Vega JA, López-Muñiz A, CalaviaMG, García-Suárez O, Cobo J, Otero J,et al. Clinical implication of Meissner’scorpuscles. CNS & NeurologicalDisorders Drug Targets. 2012;11:856-868
11
The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles…DOI: http://dx.doi.org/10.5772/intechopen.81687

[18] Bolton CF, Winkelmann RK, DyckPJ. A quantitative study of Meissner'scorpuscles in man. Neurology. 1966;16:1-9
[19] Halata Z, Munger BL. The terminalmyelin segments of afferent axons tocutaneous mechanoreceptors. BrainResearch. 1985;347:177-182
[20] Johnson KO, Yoshioka T, Vega-Bermudez F. Tactile functions ofmechanoreceptive afferents innervatingthe hand. Journal of ClinicalNeurophysiology. 2000;17:539-558
[21] Kelly EJ, Terenghi G, Hazari A,Wiberg M. Nerve fiber and sensory endorgan density in the epidermis andpapillary dermis of human hand. BritishJournal of Plastic Surgery. 2005;58:774-779
[22] Nolano M, Provitera V, Crisci C.Quantification of myelinated endingsand mechanoreceptors in human digitalskin. Annals of Neurology. 2003;54:197-205
[23] Ridley A. Silver staining of nerveendings in human digital glabrous skin.Journal of Anatomy. 1969;104:41-48
[24] Bruce MF. The relation of tactilethresholds to histology in the fingers ofelderly people. Journal of Neurology,Neurosurgery, and Psychiatry. 1980;43:730-734
[25] Iwasaki T, Goto N, Goto J, Ezure H,Moriyama H. The aging of humanMeissner's corpuscles as evidenced byparallel sectioning. Okajimas FoliaAnatomica Japonica. 2003;79:185-189
[26] Vega JA, Haro JJ, Del Valle ME.Immunohistochemistry of humancutaneous Meissner and Paciniancorpuscles. Microscopy Research andTechnique. 1996;34:351-361
[27] Vega JA, García-Suárez O, MontañoJA, Pardo B, Cobo JM. The Meissner and
Pacinian sensory corpuscles revisitednew data from the last decade.Microscopy Research and Technique.2009;72:299-309
[28] Cabo R, Alonso P, Viña E, VázquezG, Gago A, Feito J, et al. ASIC2 ispresent in human mechanosensoryneurons of the dorsal root ganglia and inmechanoreceptors of the glabrous skin.Histochemistry and Cell Biology. 2015;143:267-276
[29] Alonso-González P, Cabo R, SanJosé I, Gago A, Suazo IC, García-SuárezO, et al. Human digital Meissnercorpuscles display immunoreactivity forthe multifunctional ion channels Trpc6and Trpv4. Anatomical Record(Hoboken). 2017;300:1022-1031
[30] García-Mesa Y, García-Piqueras J,García B, Feito J, Cabo R, Cobo J, et al.Merkel cells and Meissner's corpusclesin human digital skin display Piezo2immunoreactivity. Journal of Anatomy.2017;231:978-989
[31] Calavia MG, Feito J, López-IglesiasL, de Carlos F, García-Suarez O, Pérez-Piñera P, et al. The lamellar cells inhuman Meissner corpuscles expressTrkB. Neuroscience Letters. 2010;468:106-109
[32] Vega JA, Del Valle ME, Haro JJ,Calzada B, Suarez-Garnacho S,Malinovsky L. Nerve growth factorreceptor immunoreactivity in Meissnerand Pacinian corpuscles of the humandigital skin. The Anatomical Record.1993;236:730-736
[33] Vega JA, Vazquez E, Naves FJ, DelValle ME, Calzada B, Represa JJ.Immunohistochemical localization ofthe high-affinity NGF receptor (gp140-trkA) in the adult human dorsal rootand sympathetic ganglia and in thenerves and sensory corpuscles supplyingdigital skin. The Anatomical Record.1994;240:579-588
12
Neuropathies

[34] Vega JA, Vazquez E, Naves FJ,Calzada B, del Valle ME, Represa JJ.Expression of epidermal growth factorreceptor (EGFr) immunoreactivity inhuman cutaneous nerves and sensorycorpuscles. The Anatomical Record.1994;240:125-130
[35] González-Martínez T, Monjil DF,Aguado-Barrios A, Cobo J, Germanà G,Vega JA. Bcl-2 immunoreactivity inhuman cutaneous Meissner andPacinian corpuscles. NeuroscienceLetters. 2006;394:13-16
[36] Woo SH, Lumpkin EA, PatapoutianA. Merkel cells and neurons keep intouch. Trends in Cell Biology. 2015;25:74-81
[37] Feng J, Luo J, Yang P, Du J, Kim BS,Hu H. Piezo2 channel-Merkel cellsignaling modulates the conversionof touch to itch. Science. 2018;360:530-533
[38] Maricich SM, Wellnitz SA, NelsonAM, Lesniak DR, Gerling GJ, LumpkinEA, et al. Merkel cells are essential forlight-touch responses. Science. 2009;324:1580-1582
[39] Maksimovic S, Nakatani M, Baba Y,Nelson AM, Marshall KL, Wellnitz SA,et al. Epidermal Merkel cells aremechanosensory cells that tunemammalian touch receptors. Nature.2014;509:617-621
[40] Morrison KM, Miesegaes GR,Lumpkin EA, Maricich SM. MammalianMerkel cells are descended from theepidermal lineage. DevelopmentalBiology. 2009;336:76-83
[41] Van Keymeulen A, Mascre G,Youseff KK, Harel I, Michaux C, DeGeest N, et al. Epidermal progenitorsgive rise to Merkel cells duringembryonic development and adulthomeostasis. The Journal of CellBiology. 2009;187:91-100
[42]Woo SH, Stumpfova M, Jensen UB,Lumpkin EA, Owens DM. Identificationof epidermal progenitors for the Merkelcell lineage. Development. 2010;137:3965-3971
[43] Sieber-Blum M, Szeder V, Grim M.The role of NT-3 signaling in Merkel celldevelopment. Progress in BrainResearch. 2004;146:63-72
[44] Boulais N, Pereira U, Lebonvallet N,Gobin E, Dorange G, Rougier N, et al.Merkel cells as putative regulatory cellsin skin disorders: An in vitro study.PLoS One. 2009;4(8):e6528
[45] Maksimovic S, Baba Y, LumpkinEA. Neurotransmitters and synapticcomponents in the Merkel cell-neuritecomplex, a gentle-touch receptor.Annals of the New York Academy ofSciences. 2013;1279:13-21
[46] Isgrò MA, Bottoni P, Scatena R.Neuron-specific enolase as a biomarker:Biochemical and clinical aspects.Advances in Experimental Medicine andBiology. 2015;867:125-143
[47] Kim DK, Holbrook KA. Theappearance, density, and distribution ofMerkel cells in human embryonic andfetal skin: Their relation to sweat glandand hair follicle development. TheJournal of Investigative Dermatology.1995;104:411-416
[48] Vinik AI, Nevoret ML, Casellini C,Parson H. Diabetic neuropathy.Endocrinology and Metabolism Clinicsof North America. 2013;42:747-787
[49] Bierhaus A, Haslbeck KM, HumpertPM, Liliensiek B, Dehmer T, Morcos M,et al. Loss of pain perception in diabetesis dependent on a receptor of theimmunoglobulin superfamily. TheJournal of Clinical Investigation. 2004;114:1741-1751
[50] Bril V, Buchanan RA. Long-termeffects of ranirestat (AS-3201) on
13
The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles…DOI: http://dx.doi.org/10.5772/intechopen.81687

peripheral nerve function in patientswith diabetic sensorimotorpolyneuropathy. Diabetes Care. 2006;29:68-72
[51] Christianson JA, Ryals JM, JohnsonMS, Dobrowsky RT, Wright DE.Neurotrophic modulation of myelinatedcutaneous innervation and mechanicalsensory loss in diabetic mice.Neuroscience. 2007;145:303-313
[52] Obrosova IG, Stavniichuk R, DrelVR, Shevalye H, Vareniuk I, Nadler JL,et al. Different roles of 12/15-lipoxygenase in diabetic large and smallfiber peripheral and autonomicneuropathies. The American Journal ofPathology. 2010;177:1436-1447
[53] Alsunousi S, Marrif HI. Diabeticneuropathy and the sensory apparatus“Meissner corpuscle and Merkel cells”.Frontiers in Neuroanatomy. 2014;8:79
[54] Peltier AC, Myers MI, Artibee KJ,Hamilton AD, Yan Q, Guo J, et al.Evaluation of dermal myelinated nervefibers in diabetes mellitus. Journal of thePeripheral Nervous System. 2013;18:162-167
[55] Creigh PD, McDermott MP, SowdenJE, Ferguson M, Herrmann DN. In-vivoreflectance confocal microscopy ofMeissner's corpuscles in diabetic distalsymmetric polyneuropathy. Journal ofthe Neurological Sciences. 2017;378:213-219
[56] García-Mesa Y, García-Piqueras J,Feito J, Cabo R, Vega JA, García-SuárezO. Evaluation of Meissner corpusclesand Merkel cells in diabetic neuropathy.European Journal of ClinicalInvestigation. 2018;48(Sup. 1):195-196
[57] Paré M, Albrecht PJ, Noto CJ,Bodkin NL, Pittenger GL, Schreyer DJ,et al. Differential hypertrophy andatrophy among all types of cutaneousinnervation in the glabrous skin of themonkey hand during aging and
naturally occurring typ. 2 diabetes. TheJournal of Comparative Neurology.2007;501:543-567
[58] Saporta MA, Katona I, Lewis RA,Masse S, Shy ME, Li J. Shortenedinternodal length of dermal myelinatednerve fibres in Charcot-Marie-Toothdisease typ. 1A. Brain. 2009;132(Pt 12):3263-3273
[59] Manganelli F, Nolano M, PisciottaC, Provitera V, Fabrizi GM1, CavallaroT, et al. Charcot-Marie-Tooth disease:New insights from skin biopsy.Neurology. 2015;85:1202-1208
[60] Nolano M, Manganelli F, ProviteraV, Pisciotta C, Stancanelli A, CaporasoG, et al. Small nerve fiber involvementin CMT1A. Neurology. 2015;84:407-414
[61] Almodovar JL, Ferguson M,McDermott MP, Lewis RA, Shy ME,Herrmann DN. In vivo confocalmicroscopy of Meissner corpuscles as anovel sensory measure in CMT1A.Journal of the Peripheral NervousSystem. 2011;16:169-174
[62] Aziz-Donnelly A, Harrison TB.Update of HIV-associated sensoryneuropathies. Current TreatmentOptions in Neurology. 2017;19:36
[63] Herrmann DN. Noninvasive andminimally invasive detection andmonitoring of peripheral neuropathies.Expert Review of Neurotherapeutics.2008;8:1807-1816
[64] Almodovar JL, Schifitto G,McDermott MP, Ferguson M,Herrmann DN. HIV neuropathy: Anin vivo confocal microscopic study.Journal of Neurovirology. 2012;18:503-510
[65] Albuerne M, López S, Naves FJ,Martínez-Almagro A, Represa J, VegaJA. S100α and S100β proteins in humancutaneous sensory corpuscles: Effects of
14
Neuropathies

nerve and spinal cord injury. TheAnatomical Record. 1998;251:351-359
[66] Márquez J, Pérez-Pérez M, NavesFJ, Vega JA. Effect of spinal cord andperipheral nerve injury on humancutaneous sensory corpuscles. Animmunohistochemical study. Journal ofthe Peripheral Nervous System. 1997;2:49-59
[67] López SM, Pérez-Pérez M, MárquezJM, Naves FJ, Represa J, Vega JA. p75and TrkA neurotrophin receptors inhuman skin after spinal cord andperipheral nerve injury, with specialreference to sensory corpuscles. TheAnatomical Record. 1998;251:371-383
[68] Schmid AB, Bland JD, Bhat MA,Bennett DL. The relationship of nervefibre pathology to sensory function inentrapment neuropathy. Brain. 2014;137(Pt 12):3186-3199
[69] Nolano M, Provitera V, Perretti A,Stancanelli A, Saltalamacchia AM,Donadio V, et al. Ross syndrome:A rare or a misknown disorder ofthermoregulation? A skin innervationstudy on 12 subjects. Brain. 2006;129:2119-2131
[70] Santoro L, Manganelli F, Bruno R,Nolano M, Provitera V, Barbieri F. Suralnerve and epidermal vascularabnormalities in a case of POEMSsyndrome. European Journal ofNeurology. 2006;13:99-102
[71] Provitera V, Nolano M, Pappone N,di Girolamo C, Stancanelli A, Lullo F,et al. Distal degeneration of sensory andautonomic cutaneous nerve fibres insystemic sclerosis. Annals of theRheumatic Diseases. 2005;64:1524-1526
[72] Pan B, Byrnes K, Schwartz M,Hansen CD, Campbell CM, KrupiczojcM, et al. Peripheral neuropathic changesin pachyonychia congenita. Pain. 2016;157:2843-2853
[73] Hutton EJ, Carty L, Laura M,Houlden H, Lunn MP, Brandner S,et al. c-Jun expression in humanneuropathies: A pilot study. Journal ofthe Peripheral Nervous System. 2011;16:295-303
[74] Nolano M, Provitera V, Estraneo A,Selim MM, Caporaso G, Stancanelli A,et al. Sensory deficit in Parkinson'sdisease: Evidence of a cutaneousdenervation. Brain. 2008;131:1903-1911
[75] Nolano M, Provitera V, ManganelliF, Iodice R, Stancanelli A, Caporaso G,et al. Loss of cutaneous large and smallfibers in naive and l-dopa-treated PDpatients. Neurology. 2017;89(8):776-784
[76] Ikemura M, Saito Y, Sengoku R,Sakiyama Y, Hatsuta H, Kanemaru K,et al. Lewy body pathology involvescutaneous nerves. Journal ofNeuropathology and ExperimentalNeurology. 2008;67:945-953
[77] Manganelli F, Iodice V, Provitera V,Pisciotta C, Nolano M, Perretti A, et al.Small-fiber involvement in spinobulbarmuscular atrophy (Kennedy's disease).Muscle & Nerve. 2007;36:816-820
[78] Nolano M, Provitera V, Crisci C,Saltalamacchia AM, Wendelschafer-Crabb G, Kennedy WR, et al. Smallfibers involvement in Friedreich'sataxia. Annals of Neurology. 2001;50:17-25
[79] Ren Y, Liu W, Li Y, Sun B, Li Y,Yang F, et al. Cutaneous somatic andautonomic nerve TDP-43 deposition inamyotrophic lateral sclerosis. Journal ofNeurology. 2018;265:1753-1763
[80] Symons FJ, Wendelschafer-CrabbG, Kennedy W, Hardrict R, Dahl N,Bodfish JW. Evidence of aleteredepidermal nerve fiber morphology inadults with self-injurious behaviour andneurodevelopmental disorders. Pain.2008;134:232-237
15
The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles…DOI: http://dx.doi.org/10.5772/intechopen.81687

[81] Taylor SW, Laughlin RS, Kumar N,Goodman B, Klein CJ, Dyck PJ, et al.Clinical, physiological and pathologicalcharacterisation of the sensorypredominant peripheral neuropathy incopper deficiency. Journal ofNeurology, Neurosurgery, andPsychiatry. 2017;88:839-845
[82] Kennedy WR, Selim MM, Brink TS,Hodges JS, Wendelschafer-Crabb G,Foster SX, et al. A new device toquantify tactile sensation in neuropathy.Neurology. 2011;76:1642-1649
[83] Verendeev A, Thomas C, McFarlinSC, Hopkins WD, Phillips KA,Sherwood CC. Comparative analysis ofMeissner's corpuscles in the fingertips ofprimates. Journal of Anatomy. 2015;227:72-80
[84] Li J, Bai Y, Ghandour K, Qin P,Grandis M, Trostinskaia A, et al. Skinbiopsies in myelin-related neuropathies:Bringing molecular pathology to thebedside. Brain. 2005;128:1168-1177
[85] Terracciano C, Pachatz C, Rastelli E,Pastore FS, Melone MAB, Massa R.Neurofibromatous neuropathy: Anultrastructural study. UltrastructuralPathology. 2018;42:312-316
[86] Devigili G, De Filippo M, Ciana G,Dardis A, Lettieri C, Rinaldo S, et al.Chronic pain in Gaucher disease:Skeletal or neuropathic origin?Orphanet Journal of Rare Diseases. 2017;12:148
16
Neuropathies

The ageing of the somatosensory system at the periphery: Age-related changes in
cutaneous mechanoreceptors
Short running page heading: Aging of sensory corpuscles
J. García-Piqueras1*, Y. García-Mesa1*, L. Cárcaba1, J. Feito1,2, I. Torres-Parejo1, B.
Martín-Biedma3, J. Cobo4,5, O. García-Suárez1, J. A. Vega1,6
1Departamento de Morfología y Biología Celular, Grupo SINPOS Universidad de
Oviedo, Spain
2Servicio de Anatomía Patológica, Hospital Universitario de Salamanca, Spain
3Departamento de Cirugía y Especialidades Médico-Quirúrgicas, Universidad de
Santiago de Compostela, Spain
4Departamento de Cirugía y Especialidades Médico-Quirúrgicas, Área de
Estomatología, Universidad de Oviedo, Spain
5Instituto Asturiano de Odontología, Oviedo, Spain
6Facultad de Ciencias de la Salud, Universidad Autónoma de Chile, Chile
*These authors contributed equally to this paper

ABSTRACT
Decline of tactile sensation associated to aging depends on modifications in skin and
both central and peripheral nervous systems. Nowadays, age-related changes at the
periphery of the somatosensory system, especifically in human mechanoreceptors is
poorly known. Here we used immunohistochemistry to analyze the age-dependent
changes in Meissner’s and Pacinian corpuscles as well as in Merkel’s cells-neurite
complexes. Moreover, the variations in the neurotrophic TrkB-BDNF system and the
mechanoprotein Piezo2 (involved in maintenance of cutaneous mechanoreceptors and
light touch respectively) were evaluated. The number of Meissner’s corpuscles and
Merkel´s cells decreased progressively with aging. Meissner’s corpuscles were smaller,
rounded in morphology and located deeper in the dermis and evidences of corpuscular
denervation were found in the oldest subjects. Pacinian corpuscles showed generally no
relevant age-related alterations. Ageing was accomplished of reduced expression of
Piezo2 expression in the axon of Meissner’s corpuscles and Merkel’s cells, as well as of
a decline in the BDNF-TrkB neurotrophic system. This study demonstrates that
cutaneous Meissner’s corpuscles and Merkel’s cell-neurite complexes (and less
evidently Pacinian corpuscles) undergo morphological and size changes during the
aging process, as well as a reduction in terms of density. Furthermore, the
mechanoprotein Piezo2 and the neurotrophic TrkB-BDNF system are reduced in aged
corpuscles. All together, these alterations might explain part of the impairment of the
somatosensory system associated to aging.
KEYWORDS
ageing, glabrous skin innervation, sensory corpucles, Piezo2, BDNF-TrkB system,
human

INTRODUCTION
The sensory decline in the main sensory modalities is a consequence of the
ageing process. Nevertheless, sensory ageing is not homogeneous across sensory
systems, and large differences exist among these systems (Andersen, 2012; Jayakody et
al., 2018). The experimental and clinical findings regarding the ageing of the
somatosensory system (involving mechanoreception, thermoreception and nociception)
are not conclusive, probably because of the involvement of the central nervous system,
the peripheral nervous system and skin (Shaffer & Harrison, 2007; Goble et al., 2009;
Tseng et al., 2013; Decorps et al., 2014; Heft & Robinson, 2017). In any case, elderly
people have reduced fine touch discrimination ability (Skedung et al., 2018).
Age-dependent changes in the structure and physiology of the somatosensory
cortex are now rather well known (Blatow et al., 2007; Brodoehl et al., 2013; Cheng &
Lin, 2013; Gröschel et al., 2013; Hagiwara et al., 2014). Additionally, the occurrence of
morphological and histological changes in aged skin due to genetic and environmental
factors is widely acknowledged (Khavkin & Ellis, 2011; Rittie & Fisher, 2015; Kanaki
et al., 2016; Lephart, 2016; Krutmann et al., 2017). Conversely, the effects of ageing on
the somatosensory peripheral nervous system (dorsal root ganglia, large nerve fibres,
and sensory corpuscles; Vega et al., 1993; Ulfhake et al., 2002; see Rittie & Fisher,
2015), especially at the periphery, are largely unknown. Skin ageing is associated with
an overall reduction in nerve fibre endings in the epidermis and dermis (Besné et al.,
2002; Panoutsopoulou et al., 2009; Fromy et al., 2010; Namer, 2010), although
photoaging is characterized by increased sensory nerves in skin (Toyoda et al., 2005).
Tactile sensation is one of the most important components of mechanosensation
and is sensed by specific sensory formations localized in the skin and known
collectively as cutaneous sensory corpuscles or receptors (Zimmerman et al., 2014).

Functionally, these receptors fall into two categories: rapidly adapting
mechanoreceptors and slowly adapting mechanoreceptors, which each have two
variants, type I and type II, which differ in terms of their electrophysiological and
morphological characteristics (Jones & Smith, 2014). Slowly adapting type I
mechanoreceptors (SAI) are associated with epidermal Merkel cell-neurite complexes,
and slowly adapting type II mechanoreceptors (SAII) are thought to be located in
dermal Ruffini’s corpuscles, although the function of SAII can be carried out by other
sensory corpuscles in addition to or different from Ruffini’s corpuscles (Wu, 1998; Paré
et al., 2003; Olson et al., 2016). Merkel cells are specialized epidermal cells (Van
Keymeulen et al., 2009) functionally connected to Aβ nerve fibres that act as SAI and
express mechanoproteins (Ikeda et al., 2014; Maksimovic et al., 2014; García-Mesa et
al., 2017). In mammals, rapidly adapting mechanoreceptors are found in Meissner’s and
Pacinian sensory corpuscles (Zimmerman et al., 2014). Meissner’s corpuscles (MCs)
consist of an axon that spirals between stacked Schwann-related cells, are located in the
superficial papillary dermis and represent rapidly adapting type I mechanoreceptors
(RAI; Vega et al., 2012). Cutaneous Pacinian corpuscles are located in the superficial
hypodermis or deep reticular dermis and represent rapidly adapting type II
mechanoreceptors (RAII); their morphology resembles an onion bulb, with a central
axon surrounded by thickly packed lamellar Schwann-related cells that form the inner
core and are surrounded by many loosely packed endoneurial-perineurium-related cells
that constitute the intermediate layer, the outer core and the capsule outer core (Vega et
al., 2009; García-Piqueras et al., 2017).
Because age-dependent impairments in mechanosensory function have been
reported and because limited information is available about age-dependent changes in
mechanosensors, we conducted research on human digital skin to evaluate the

quantitative and qualitative changes that occur in Merkel’s cells, Meissner’s corpuscles
and Pacinian corpuscles with ageing. We have recently studied the development of
these structures until maturity (Feito et al., 2018), demonstrating that human cutaneous
sensory corpuscles are not stable but are dynamic structures that undergo permanent
age-related changes. On the other hand, because the ion channel Piezo2 provides at the
basis of mechanotransduction (see for a review Anderson et al., 2017) and is located in
both Meissner’s corpuscles and Merkel’s cells of human digital skin (García-Mesa et
al., 2017), we investigated whether the pattern of expression of this protein changes
with ageing. Furthermore, it is well known that cutaneous mechanoreceptors depend on
the neurotrophin system of the brain-derived neurotrophic factor (BDNF) and its high-
affinity receptor TrkB for development, growth and maintenance (Botchkarev et al.,
1999; LeMaster et al., 1999; González-Martinez et al., 2004; Perez-Pinera et al., 2008;
Reed-Geaghan et al., 2016). Additionally, both BDNF and TrkB have been detected in
developing and adult sensory corpuscles in different species, including humans (Stark et
al., 2001; Sedy et al., 2004; Calavia et al., 2010; Cabo et al., 2015a). Therefore, we
analysed whether the localization of BDNF and TrkB in mechanoreceptors varies with
age. Reduced expression of BDNF and TrkB has been observed in the central nervous
system of old subjects compared with young and adult subjects (Erickson et al., 2010;
Forlenza et al., 2015), suggesting that these molecules have a role in normal ageing.
The present study aimed to add information about the mechanisms that underly
sensory deficits in elderly subjects and is focused on the peripheral touch components of
somatosensory perception, with analysis of the effects of ageing on the structural and
neurotrophic features of cutaneous sensory corpuscles.
METHODS

Skin samples were obtained from the palmar aspect of the distal phalanx of the
first and second hand fingers during autopsy at the Service of Pathology of the Hospital
Universitario Central de Asturias, Oviedo, Spain (n = 6) and the Service of Pathology of
the Complejo Hospitalario Universitaro de Salamanca (n = 9) and from incidental
amputation at the Service of Plastic Surgery of the Hospital Universitario Central de
Asturias (n = 6). The samples were collected within 12 h after demise from subjects free
of neurological diseases and without a clinical history of fibromatosis or labour-induced
repeated trauma. The age range was 23 to 90 years, and at least one case per decade of
life was analysed.
The specimens were fixed in 4% formaldehyde in 0.1 M phosphate-buffered
saline (pH 7.4) for 24 h, dehydrated and routinely embedded in paraffin. The pieces
were cut to 7 µm thickness in sections perpendicular to the skin surface and mounted on
gelatin-coated microscope slides. The presence of Pacinian and Meissner’s corpuscles
in the skin samples was evaluated by staining with haematoxylin and eosin. The
occurrence of Merkel’s cells was assessed with immunohistochemistry. These materials
were all obtained in compliance with Spanish Law and the guidelines of the Declaration
of Helsinki.
The automated diagnostic platform Leica Bond III was used for
immunohistochemistry with the Leica Bond™ Polymer Refine Detection Kit (Leica
Biosystems™, New Clastle upon Tyne, UK ) according to the manufacturer’s
instructions. The primary antibodies were directed against the main corpuscular
constituents: axons (neurofilament, PGP 9.5) and Schwann cell-derived lamellar and
inner core cells (S100 protein). Moreover, to identify Merkel cells, an antibody against
cytokeratin 20 (CK20) was used. The BDNF-TrkB neurotrophin system was detected
using specific antibodies against specific amino acid sequences within these molecules.

Indirect immunohistochemistry included several negative and positive controls, as well
as the internal positive and negative controls. Additionally, representative sections were
processed in the same way as described above using non-immune rabbit or mouse sera
instead of the primary antibodies or by omitting the primary antibodies during the
incubation.
Data on case material and the characteristics of the primary antibodies used are
summarized in Tables 1 and 2, respectively.
Double immunofluorescence was performed on deparaffinized and rehydrated
sections. Endogenous peroxidase activity and non-specific binding were reduced by
incubating sections with 3% H2O2 and a solution of 10% bovine serum albumin in Tris
buffer solution (TBS), respectively. The sections were then incubated overnight at 4°C
in a humid chamber with a 1:1 mixture of a polyclonal antibody against S100-protein
(diluted 1:1000), PGP 9.5 (diluted 1:1000), BDNF (diluted 1:200) or TrkB (diluted
1:200) and a monoclonal antibody against NFP (diluted 1:1000) or CK20 (prediluted
and 1:200 in the blocking solution). After rinsing with TBS, the sections were incubated
for 1 h with Alexa Fluor 488-conjugated goat anti-rabbit IgG (Serotec, Oxford, UK),
diluted 1:1000 in TBS containing 5% mouse serum (Serotec), rinsed again and
incubated for another hour with Cy3-conjugated donkey anti-mouse antibody (Jackson-
ImmunoResearch, Baltimore, MD, USA) diluted 1:50 in TBS. Both steps were
performed at room temperature in a dark humid chamber. Finally, to ascertain structural
details, sections were counterstained and mounted with DAPI diluted in glycerol
medium (10 ng/ml). Triple fluorescence was detected using a Leica DMR-XA
automatic fluorescence microscope (Photonic Microscopy Service, University of
Oviedo) with Leica Confocal Software, version 2.5 (Leica Microsystems, Heidelberg
GmbH, Germany), and the captured images were processed using the software ImageJ,

version 1.43 g Master Biophotonics Facility, Mac Master University Ontario
(www.macbiophotonics.ca). For control purposes, representative sections were
processed in the same way as described above using non-immune rabbit or mouse sera
instead of the primary antibodies or by omitting the primary antibodies during the
incubation. Furthermore, when available, additional controls were carried out using
specifically preabsorbed antisera. Under these conditions, no positive immunostaining
was observed (data not shown).
Quantitative analyses
Quantitative analyses were performed to determine the density of cutaneous
digital Meissner’s corpuscles and Merkel cells at different ages but not the density of
Pacinian corpuscles because of their irregular distribution in the skin dermis. To
quantify Meissner’s corpuscles, we used the method proposed by Verendeev et al.
(2015) to determine the density of Meissner’s corpuscles in the fingertips of primates.
The samples included in the study were divided into three age groups: 20 to 39 years, 40
to 59 years, and 60 years or older (see Table 1). Briefly, six 7 µm thick sections of each
skin sample that were 200 µm apart and processed for S100 protein
immunohistochemistry were evaluated. The sections were scanned by an SCN400F
scanner (Leica, Leica Biosystems™), and the scans were computerized using SlidePath
Gateway LAN software (Leica, Leica Biosystems™). Then, in each section, Meissner’s
corpuscles were identified and counted by two independent observers (JG-P and JAV).
The average numerical values were corrected by applying Abercrombie’s formula: N =
n*T/(T+H), where N is the corrected average number of Meissner’s corpuscles, n is the
counted average number of Meissner´s corpuscles in all sections of a fingertip, T is the
average section thickness, and H is the average diameter of the counted Meissner

corpuscles. Through a specific tool of the previously mentioned software, the average
Meissner corpuscle’s diameter was determined by measuring its horizontal axis by
drawing a straight line approximately in the central region of each Meissner’s corpuscle.
The epidermal length of each section (mm) was measured with the same tool, and the
average length was multiplied by the section’s thickness (mm) to give the measured
surface area (mm2). Finally, the average number of Meissner corpuscles (N) was
divided by the surface area (mm2) to calculate the density of Meissner corpuscles per
squared millimetre of skin (number of Meissner’s corpuscles/mm2). Afterward, the
average density for each pre-established age group was calculated from the individual
densities. To determine the density of digital Merkel cells, we used the same Merkel
cell immunostaining method for CK20. Significant differences among the three pre-
established age groups were assessed with the Kruskal-Wallis H test, and p-values
<0.05 were considered statistically significant (marked in the figures as *p<0.05,
**p<0.01).
RESULTS
Ageing of digital Meissner’s corpuscles
Meissner’s corpuscles were identified at all ages investigated (Figs. 1-3), but
differences in the morphology, placement within the dermis, intensity of
immunostaining for the evaluated markers, size and number were noted between the
three pre-established age groups. In the age group of 20-39 years, Meissner’s corpuscles
were elongated and always localized in the apex of the dermal papillae. The lamellar
cells were packed, arranged in parallel and displayed strong S100 protein
immunoreactivity (Figs. 1a, 2b and d); within the corpuscle, the axon showed a tortuous

trajectory (Fig. 2a and c). By 40-59 years, the predominant localization of Meissner’s
corpuscles was also within the dermal papillae, but some were also highly displaced
(Fig. 1b); the corpuscles showed no reduction in size, and the architecture and
immunohistochemical profile of the axon and lamellar cells did not vary (Fig. 2e-h).
The scenario changed dramatically in the older subjects (Fig. 1c). In this group, the size
of the Meissner’s corpuscles was reduced, the morphology was rounded (only a few
showed a typical elongated morphology), and most were localized behind rete pegs
instead of inside the dermal papillae. In those corpuscles, the lamellar cells showed
reduced immunostaining for S100 protein, and the axon was sometimes
undistinguishable (Figs. 2i-l and 3). This picture applied to approximately 70% of the
corpuscles, whereas the 30% remaining had a size, morphology and localization similar
to that of the younger subjects.
All these findings were confirmed by confocal microscopy of the Meissner’s
corpuscles of the three age groups, which showed that ageing induced changes in the
size, morphology and immunohistochemical profile of these structures (Fig. 4). Of
particular interest was the absence of axon profiles in a large percentage of Meissner’s
corpuscles from older subjects (Fig. 4i).
In addition to the above-described qualitative changes, ageing is accompanied
by a progressive reduction in the number of Meissner’s corpuscles. Significant
differences were observed between age groups, especially between young subjects vs.
adult and old subjects (Fig. 1).
Ageing of digital Pacinian corpuscles
No evident noticeable variations in the Pacinian corpuscles were found between
the three established age groups (Fig. 5). The arrangement of all corpuscular

components was almost identical, and no age-related changes were found in the number
of concentric lamellae forming the outer core and the capsule. The axon displayed NFP
immunoreactivity (Figs. 5a, c, e, g), whereas the lamellae of the inner core were positive
for S100 protein (Figs. 5b, d, f, h, j). Nevertheless, in some Pacinian corpuscles of older
subjects (approximately 15-25%), no immunoreactivity for NFP was detected, and S100
protein immunoreactivity was restricted to the outer lamellae of the inner core (Figs. 5i-
l). Additionally, in some other corpuscles, the neural compartment of the corpuscles
(i.e., the axon and the inner core) was disarranged compared with the typical
arrangement of these structures (Fig. 6).
Ageing of cutaneous Merkel’s cells
Digital Merkel cells are epidermal cells primarily localized in the basal strata
that selectively express the intermediate filament CK20. Using the expression of this
protein as a marker, Merkel’s cells were found in the epidermal rete pegs of all subjects,
and the number of cells progressively decreased with age (Fig. 7). The quantitative
analysis showed significant differences between the young subjects and the adult and
old subjects (graph in Fig. 7).
A graphic summary of the age-related changes in Meissner’s corpuscles and
Merkel’s cells described above is presented in Fig. 8.
Ageing of the BDNF-TrkB system in cutaneous mechanoreceptors
The development and maintenance of Meissner’s corpuscles and, to a lesser
extent, Merkel’s cells and Merkel’s cell-axon complexes are under the control of the
BDNF-TrkB neurotrophin system. Thus, we investigated whether the components of
that system are present in those sensory structures and whether these components

undergo age-dependent variations. In human digital Meissner’s corpuscles from adult
subjects, immunoreactivities for BDNF and TrkB were detected in the lamellar cells
(Supplementary Fig. 1a-c) and the axon (Supplementary Fig. 1d-f), respectively. In the
Pacinian corpuscles, BDNF was found in the inner core cells (Supplementary Fig. 2),
and TrkB was found in the axons (data not shown). Regarding Merkel’s cells, BDNF
was undetectable in these cells (Supplementary Fig. 3a-c), while TrkB
immunoreactivity regularly occurred in their cytoplasms (Supplementary Fig. 3d-f).
The pattern of localization of both BDNF and TrkB with age was previously
described. However, in younger subjects, BDNF immunoreactivity also showed
localization in axons (Fig. 9a). With ageing, the intensity of immunostaining for BDNF
(Figs. 9b-d) and TrkB (Figs. 9e-h) progressively decreased, and TrkB immunoreactivity
was undetectable in older subjects (Fig. 9h). These results applied to approximately
85% of Meissner’s corpuscles, and no apparent age-dependent changes were observed
in the remaining corpuscles (15%). The percentage of TrkB-positive Merkel cells also
declined with age, with a reduction of approximately 60% in the old age group
compared with the young and adult age groups (data not shown).
Piezo2 levels in Meissner’s corpuscles and Merkel’s cells decrease with age
The occurrence of Piezo2 immunoreactivity in the axons of human cutaneous
Meissner’s corpuscles and the cytoplasm of Merkel’s cells has been demonstrated
recently by our research group (García-Mesa et al., 2017). Here, we observed that the
axon of most of the Meissner’s corpuscles of the younger subjects displayed Piezo2
positivity (Figs. 10a and b), which remained unchanged in the middle-age group and
was markedly reduced in the older subject age group (Fig. 10d and e). No Piezo2
immunostaining was noted in the lamellar cells. In agreement with the above data

regarding the age-dependent decrease in the density of Merkel’s cells, there was an age-
dependent decrease in the density of Piezo2-positive Merkel’s cells (Fig. 10c and f). On
the other hand, although no direct contacts between Merkel’s cells and axons were
observed, the density of axons in the vicinity or apparently contacting Merkel’s cell also
decreased with age (Fig. 11).
DISCUSSION
In human skin, a series of sensory formations, collectively known as sensory
corpuscles (Zimmerman et al., 2014), are able to detect different qualities of
somatosensation. In particular, Merkel’s cell-neurite complexes, Meissner’s corpuscles
and Pacinian corpuscles discriminate unique aspects of touch with SAI and SAII
mechanoreceptors (Jones & Smith, 2014; Zimmerman et al., 2014). Ageing is
accompanied by a progressive decrease in the somatosensory system that affects the
quality of life of elderly subjects (Skedung et al., 2018). This deterioration presumably
involves all levels of the somatosensory pathways from the skin to the cerebral cortex,
including the cutaneous receptors in which the mechanical input is transduced into
electrical energy. The present study was designed to investigate the age-dependent
changes in some types of cutaneous mechanoreceptors and in one mechanoprotein
directly involved in mechanotransduction (Piezo2), as well as the variations in a
neurotrophic system (BDNF-TrkB) that is essential for their development and
maintenance.
The effects of ageing on touch have been reviewed in detail by Wickremaratchi
and Llewelyn (2006), Decorps et al. (2014), and Heft and Robinson (2017). Ageing is
also associated with a progressive decline in cutaneous thermic (Guergova & Dufour,
2011) and mechanical (Wickremaratchi & Llewelyn, 2006; Wu et al., 2011) perception,

as well as a marked degradation of tactile spatial acuity (Kalisch et al., 2009). As a
whole, these authors affirm that tactile perceptual ability as well as spatial acucity
decrease with age, and this impairment could be due at least in part to a reduction in the
density and distribution of Meissner’s and Pacinian corpuscles (Wickremaratchi &
Llewelyn, 2006), as well as Merkel’s disc (Lumpkin et al., 2003).
Age-dependent changes in cutaneous Meissner’s and Pacinian corpuscles
Classic studies have demonstrated that Pacinian and Meissner’s corpuscles
decrease in number and undergo structural deterioration upon ageing in humans (Bolton
et al., 1966; Kennedy et al., 2011), monkey (Paré et al., 2007) and mice (Mathewson &
Nava, 1985). Furthermore, deterioration of the discs of Merkel was also observed with
ageing in rats (Fundin et al., 1997), but deterioration was not observed in Merkel’s cell-
neurite complexes (Bolton et al., 1966; Mathewson & Nava, 1985; Paré et al., 2007).
Studies in humans are limited and do not use specific immunohistochemistry
assays for each corpuscular component. Here, we observed that the size, morphology
and structure of Meissner’s corpuscles remained essentially unaltered until 60 years or
age, and these corpuscles then progressively shrank, changed morphology and
topographical localization, and lost their immunohistochemical profile. Interestingly,
the absence of immunoreactivity for axonal markers in older subjects suggests
denervation of those corpuscles, which is supported by the decrease in the expression of
S100 protein by the lamellar cells. In fact, it is well known that lamellar cells of rat pad
Meissner-like corpuscles lack S100 protein (del Valle et al., 1993), with a similar result
in humans (Marquez et al., 1997; Albuerne et al., 1998). Globally, our results are in
good agreement with the data reported by Nava and Mathewson (1996) in Meissner-like
corpuscles of murine forepaw digital pads. These authors observed that in mice aged to

their maximum life expectancy, Meissner-like corpuscles decrease and become
disorganized and lobulated compared with those in middle-age animals, and these
changes are attributed to distal axonopathy and atrophy of the sensory neurons. On the
other hand, these changes in Meissner’s corpuscles affect most but not all the
corpuscles, suggesting that tactile texture discrimination remains partially intact with
age, irrespective of cutaneous condition. The findings observed here are difficult to
apply to the glabrous skin of the entire body. Presumably, differences in ageing exist
between different anatomical zones depending on the environment. In addition,
decreased epidermal innervation of facial skin was found, epidermal innervation of
abdominal skin did not change with age, and an age-associated increase in innervation
was observed in mammary skin (Besné et al., 2002). A recent study by Skedung et al.
(2018) evaluated the density of Meissner’s corpuscles in two groups of aged subjects
with high and low performance, showing that the number was reduced approximately
50% in the subjects with lower performance. Thus, the lower performance of this group
can be confidently linked to a neural decline, although we stress that we do not link the
reduced acuity directly to the lower Meissner’s corpuscle density because there are no
data on the other tactile receptors. Thus, the age-related decline in neural properties is
the primary explanation for the reduction in active touch acuity. This reduction results
in an age-related change in of hand movement perception (Landelle et al., 2018)
Regarding Pacinian corpuscles, the effects of ageing were not evident either in
the number or in the structure. However, a small population of Pacinian corpuscles from
the older subjects seemed to be denervated (no labelling of the axon and absence of
S100 protein in the whole inner core) or the neural compartment was disarranged.
Pacinian corpuscles have been related to the detection of vibration (Bell et al., 1994;
Zelena, 1994; Zimmerman et al., 2014), and these variations might be correlated with

the decreased vibration sensitivity reported in elderly people (see Shaffer & Harrison,
2007; Landelle et al., 2018).
The exhaustive review on the Pacinian corpuscles carried out by Zelena (1994)
in the book “Nerves and Mechanoreceptors” does not mention the age-dependent
changes in Pacinian corpuscles in humans or other mammalian species.
Age-related changes in the expression of basic corpuscular antigens
We observed that in some Meissner’s corpuscles from the aged subjects, there
was a reduction in the immunoreactivity for S100 protein along with an absence of
immunohistochemically detectable axons. These two findings suggest denervation of
these corpuscles because the expression of S100 protein by lamellar cells depends on
the functional integrity of the axon (Marquez et al., 1997; Albuerne et al., 1998).
Age-dependent changes in Merkel’s cells and Merkel’s cell-neurite complexes
In aged mice, we found a reduction in the number of palatal Merkel cells
number at key time-points that correlate with impaired oral abilities, such as swallowing
and mastication. Collectively, this work identified the mechanosensory architecture of
oral tissues involved in feeding (Moayedi et al., 2018). A study carried out in the
Meissner-like corpuscles of the murine palatine mucosa showed that only rudiments of
corpuscles were encountered due to an atrophy of the axon and lamellae (Iida &
Tachibana, 1996). Recently, Moayedi et al. (2018) reported that Merkel cells undergo a
dramatic reduction in density with ageing in murine palatine mucosa, whereas the
authors did not examine variations in Meissner’s corpuscles.
Piezo2 expression is reduced in aged cutaneous mechanoreceptors

Activation of mechanically gated ion channels is at the origin of the detection of
low- or high-threshold mechanical stimuli. Different candidates have been proposed to
be the mechanotransducers in human sensory corpuscles and Merkel’s cells (Calavia et
al., 2010b; Cabo et al., 2015b; Alonso-González et al., 2017). However, only Piezo2 has
been shown to be an essential component of distinct stretch-activated ion channels
involved in the mechanotransduction (see Roudaut et al., 2012; Delmas & Coste, 2013;
Wu et al., 2017. Piezo2 is present in mechanosensory neurons (Coste et al., 2010;
Ranade et al., 2014) and different low-threshold cutaneous mechanoreceptors, including
human Meissner’s corpuscles and Merkel’s cells (García-Mesa et al., 2017). Although
the ageing effects on the structure and/or the function of these mechanosensitive ion
channels have not been described, one can speculate that these channels could
contribute to age-related tactile defects.
The BDNF-TrkB neurotrophin complex is altered in the skin of aged subjects
The development, survival and maintenance of cutaneous sensory corpuscles are
under the control of different neurotrophins and their Trk signalling receptors (see
Montano et al., 2010). The BDNF-TrkB system is involved in different modalities of
touch because it controls Meissner’s corpuscles and, to a lesser extent, Pacinian
corpuscles and Merkel’s cell-neurite complexes (González-Martınez et al., 2004; Sedy
et al., 2004; de Carlos et al., 2006; Perez-Pinera et al., 2008). The BDNF-TrkB system
over sensory corpuscles is controlled by retrograde axonal transport in the sensory
neurons supplying these components or directly by non-neuronal cells surrounding the
peripheral tip of the sensory axon.
BDNF-TrkB continues to be expressed in some neurons of adult sensory ganglia
and in some types of sensory corpuscles throughout the whole lifespan (see Montano et

al., 2010). The present and previous studies (Calavia et al., 2010a) demonstrate that
human cutaneous sensory corpuscles contain immunohistochemically detectable BDNF
and TrkB. Our results strongly suggest that in Meissner’s and Pacinian corpuscles,
retrograde trophic signalling occurs to provide trophic support to the sensory neurons
supplying these components: the lamellar cells and the inner core cells seem to be the
source of BDNF, and the axons that express TrkB are the recipients.
As far as we know, the effects of ageing in this neurotrophic system have never
been investigated. Our results demonstrate that the ageing of sensory corpuscles is
accompanied by a decrease in the immunohistochemically detectable BDNF and TrkB,
which were absent in some Meissner’s corpuscles. Whether the absence of BDNF-TrkB
is responsible, at least in part, for the size and morphological changes observed in
Meissner’s corpuscles and in the density of Merkel cells remains to be investigated. In
any case, our results could explain the reduction in sensory neurons of the dorsal root
ganglia that occurs with ageing (Vega et al., 1993). In the human central nervous
system, low levels of BDNF and reduced expression of TrkB are related to ageing and
the related diseases (Erickson et al., 2010; Forlenza et al., 2015; Nunes et al., 2018).
Overall, the present study demonstrates that the cutaneous Meissner’s corpuscles
and Merkel cell-neurite complexes and, less evidently, Pacinian corpuscles, which
represent the most peripheral part of the sensory nervous system, undergo age-
dependent morphological and size changes as well as a decrease in their density. The
mechanoprotein Piezo2 involved in fine touch is also reduced in aged corpuscles, as is
the trophic system responsible for their maintenance. These changes partially explain
the impairment of the somatosensory with ageing.
ACKNOWLEDGEMENTS

This study was supported in part by a grant from Gerencia Regional de Salud de Castilla
y León (GRS 1615/A/17). The authors thank Dr. Marta Guervos (Servicios Comunes de
Investigación, Microscopia Confocal, Universidad de Oviedo) and Marta Sánchez-Pitiot
(Grupo de Histopatología Molecular, Instituto Universitario de Oncología del
Principado de Asturias) for technical assistance.
AUTHOR CONTRIBUTIONS
J.G-P, Y.G-M and I.TP performed the experiments
O.G-S and JAV designed the stdy abd analysed the data
JF, JC and BM-B collected the material under legal conditions and performed part of
the experiments
J.G-P and JAV wrote the manuscript

REFERENCES
Albuerne M, López S, Naves FJ, Martínez-Almagro A, Represa J, Vega JA. (1998)
S100α and S100β proteins in human cutaneous sensory corpuscles: effects of
nerve and spinal cord injury. Anat Rec 251, 351–359.
Alonso-González P, Cabo R, José I, et al. (2017) Human digital Meissner corpuscles
display immunoreactivity for the multifunctional ion channels Trpc6 and Trpv4.
Anat Rec 300, 1022–1031.
Andersen GJ. (2012) Aging and vision: changes in function and performance from
optics to perception. Wiley Interdiscip Rev Dev Biol 3, 403–410.
Anderson EO, Schneider ER, Bagriantsev SN. (2017) Piezo2 in cutaneous and
proprioceptive mechanotransduction in vertebrates. Curr Top Membr 79, 197–
217.
Bell J, Bolanowski S, Holmes MH. (1994)The structure and function of Pacinian
corpuscles: a review. Prog Neurobiol. 42, 79-128.
Besné I, Descombes C, Breton L. (2002) Effect of age and anatomical site on density
of sensory innervation in human epidermis. Arch Dermatol 138, 1445–1450.
Blatow M, Nennig E, Durst A, Sartor K, Stippich C. (2007) fMRI reflects functional
connectivity of human somatosensory cortex. NeuroImage 37, 927–936.
Bolton CF, Winkelmann RK, Dyck PJ. (1966) A quantitative study of Meissner's
corpuscles in man. Neurology 16, 1–9.
Botchkarev VA, Kief S, Paus R, Moll I. (1999) Overexpression of brain-derived
neurotrophic factor increases Merkel cell number in murine skin. J Invest
Dermatol 113, 691–692.

Brodoehl S, Klingner C, Stieglitz K, Witte OW. (2013) Age-related changes in the
somatosensory processing of tactile stimulation—An fMRI study. Behav Brain
Res 238, 259–264.
Cabo R, Alonso P, José IS, et al. (2015a) Brain-derived neurotrofic factor and its
receptor TrkB are present, but segregated, within mature cutaneous pacinian
corpuscles of Macaca fascicularis. Anat Rec 298, 624–629.
Cabo R, Alonso P, Viña E, et al. (2015b) ASIC2 is present in human mechanosensory
neurons of the dorsal root ganglia and in mechanoreceptors of the glabrous skin.
Histochem Cell Biol 143, 267–276.
Calavia MG, Feito J, López-Iglesias L, et al. (2010a) The lamellar cells in human
Meissner corpuscles express TrkB. Neurosci Lett 468, 106–109.
Calavia MG, Montaño JA, García-Suárez O, et al. (2010b) Differential localization
of acid-sensing ion channels 1 and 2 in human cutaneus pacinian corpuscles.
Cell Mol Neurobiol 30, 841–848.
Cheng CH, Lin YY. (2013) Aging-related decline in somatosensory inhibition of the
human cerebral cortex. Exp Brain Res 226, 145–152.
Coste B, Mathur J, Schmidt M, et al. (2010) Piezo1 and Piezo2 are essential
components of distinct mechanically activated cation channels. Science 330, 55–
60.
De Carlos F, Cobo J, Germanà G, et al. (2006) Abnormal development of pacinian
corpuscles in double trkB-trkC knockout mice. Neurosci Lett 410, 157–161.
Decorps J, Saumet JL, Sommer P, Sigaudo-Roussel D, Fromy B. (2014) Effect of
ageing on tactile transduction processes. Ageing Res Rev 13, 90–99.
Delmas P, Coste B. (2013) Mechano-gated ion channels in sensory systems. Cell 155,
278–284.

Del Valle ME, Cabal A, Alvarez-Mendez JC, et al. (1993) Effect of denervation on lamellar
cells of Meissner-like sensory corpuscles of the rat. An immunohistochemical study.
Cell Mol Biol (Noisy-le-grand) 39, 801-807.
Erickson KI, Prakash RS, Voss MW, et al. (2010) Brain-derived neurotrophic factor
is associated with age-related decline in hippocampal volume. J Neurosci 30,
5368–5375.
Feito J, Garcia-Suarez O, Garcia-Piqueras J, et al. (2018) The development of
human digital Meissner's and Pacinian corpuscles. Ann Anat 219, 8–24.
Forlenza OV, Miranda AS, Guimar I, et al. (2015) Decreased neurotrophic support is
associated with cognitive decline in non-demented subjects. J Alzheimers Dis
46, 423–429.
Fromy B, Sigaudo-Roussel D, Gaubert-Dahan ML, et al. (2010) Aging-associated
sensory neuropathy alters pressure-induced vasodilation in humans. J Investig
Dermatol 130, 849–855.
Fundin BT, Bergman E, Ulfhake B. (1997) Alterations in mystacial pad innervation in
the aged rat. Exp Brain Res 117, 324–340.
García-Mesa Y, García-Piqueras J, García B, et al. (2017) Merkel cells and
Meissner's corpuscles in human digital skin display Piezo2 immunoreactivity. J
Anat 231, 978–989.
García-Piqueras J, García-Suárez O, Rodríguez-González MC, et al. (2017)
Endoneurial-CD34 positive cells define an intermediate layer in human digital
Pacinian corpuscles. Ann Anat 211, 55–60.
Goble DJ, Coxon JP, Wenderoth N, Van Impe A, Swinnen SP. (2009)
Proprioceptive sensibility in the elderly: degeneration, functional consequences
and plastic-adaptive processes. Neurosci Biobehav Rev 33, 271–278.

González-Martínez T, et al. (2004) Absence of Meissner corpuscles in the digital pads
of mice lacking functional TrkB. Brain Res 1002, 120–128.
Gröschel S, Sohns JM, Schmidt-Samoa C, et al. (2013) Effects of age on negative
BOLD signal changes in the primary somatosensory cortex. NeuroImage 71,
10–18.
Guergova S, Dufour A. (2011) Thermal sensitivity in the elderly: a review. Ageing Res
Rev 10, 80–92.
Hagiwara K, Ogata K, Okamoto T, et al. (2014) Age-related changes across the
primary and secondary somatosensory areas: an analysis of neuromagnetic
oscillatory activities. Clin Neurophysiol 125, 1021–1029.
Heft MW, Robinson ME. (2017) Somatosensory function in old age. J Oral Rehabil
44, 327–332.
Iida S, Tachibana T. (1996) Age-related changes in Meissner corpuscles in the mouse
palate: a histochemical and ultrastructural study. Arch Histol Cytol 59, 281–290.
Ikeda R, Cha M, Ling J, Jia Z, Coyle D, Gu J. (2014) Merkel cells transduce and
encode tactile stimuli to drive Aβ-afferent impulses. Cell 157, 664–675.
Jayakody DMP, Friedland PL, Martins RN, Sohrabi HR. (2018) Impact of aging on
the auditory system and related cognitive functions: a narrative review. Front
Neurosci 12, 125.
Jones LA, Smith AM. (2014) Tactile sensory system: encoding from the periphery to
the cortex. Wiley Interdiscip Rev Dev Biol 6, 279–287.
Kalisch T, Ragert P, Schwenkreis P, Dinse HR, Tegenthoff M. (2009) Impaired
tactile acuity in old age is accompanied by enlarged hand representations in
somatosensory cortex. Cereb Cortex 19, 1530–1538.

Kanaki T, Makrantonaki E, Zouboulis CC. (2016) Biomarkers of skin aging. Rev
Endocr Metab Disord 17, 433–442.
Kennedy WR, Selim MM, Brink TS, et al. (2011) A new device to quantify tactile
sensation in neuropathy. Neurology 76, 1642–1649.
Khavkin J, Ellis DAF. (2011) Aging skin: histology, physiology, and pathology.
Facial Plast Surg Clin North Am 19, 229–234.
Krutmann J, Bouloc A, Sore G, Bernard BA, Passeron T. (2017) The skin aging
exposome. J Dermatol Sci 85, 152–161.
Landelle C, El Ahmadi A, Kavounoudias A. (2018) Age-related impairment of hand
movement perception based on muscle proprioception and touch. Neuroscience
381, 91–104.
LeMaster AM, Krimm RF, Davis BM, et al. (1999) Overexpression of brain-derived
neurotrophic factor enhances sensory innervation and selectively increases
neuron number. J Neurosci 19, 5919–5931.
Lephart ED. (2016) Skin aging and oxidative stress: equol’s anti-aging effects via
biochemical and molecular mechanisms. Ageing Res Rev 31, 36–54.
Lumpkin EA, Collisson T, Parab P, et al. (2003) Math1-driven GFP expression in the
developing nervous system of transgenic mice. Gene Expr Patterns 3, 389–395.
Maksimovic S, Nakatani M, Baba Y, et al. (2014) Epidermal Merkel cells are
mechanosensory cells that tune mammalian touch receptors. Nature 509, 617–
621.
Marquez J, Perez-Perez M, Naves FJ, Vega JA. (1997) Effect of spinal cord and
peripheral nerve injury on human cutaneous sensory corpuscles. An
immunohistochemical study. J Peripher Nerv Syst 2, 49–59.

Mathewson RC, Nava PB. (1985) Effects of age on meissner corpuscles: a study of
silver-impregnated neurites in mouse digital pads. J Comp Neurol 231, 250–259.
Moayedi Y, Duenas-Bianchi LF, Lumpkin EA. (2018) Somatosensory innervation of
the oral mucosa of adult and aging mice. Sci Rep 8, 9975.
Montano JA, Perez-Pinera P, Garcia-Suarez O, Cobo J, Vega JA. (2010)
Development and neuronal dependence of cutaneous sensory nerve formations:
lessons from neurotrophins. Microsc Res Tech 73, 513–529.
Namer B. (2010) Age related changes in human C-fiber function. Neurosci Lett 470,
185–187.
Nava PB, Mathewson RC. (1996) Effect of age on the structure of Meissner corpuscles
in murine digital pads. Microsc Res Tech. 34, 376-389.
Nunes PV, Nascimento CF, Kim HK, et al. (2018) Low brain-derived neurotrophic
factor levels in post-mortem brains of older adults with depression and dementia
in a large clinicopathological sample. J Affect Disord 241, 176–181.
Olson W, Dong P, Fleming M, Luo W. (2016) The specification and wiring of
mammalian cutaneous low-threshold mechanoreceptors. Wiley Interdiscip Rev
Dev Biol 5, 389–404.
Panoutsopoulou IG, Wendelschafer-Crabb G, Hodges JS, Kennedy WR. (2009)
Skin blister and skin biopsy to quantify epidermal nerves: a comparative study.
Neurology 72, 1205–1210.
Paré M, Albrecht PJ, Noto CJ, et al. (2007) Differential hypertrophy and atrophy
among all types of cutaneous innervation in the glabrous skin of the monkey
hand during aging and naturally occurring type 2 diabetes. J Comp Neurol 501,
543–567.

Paré M, Behets C, Cornu O. (2003) Paucity of presumptive ruffini corpuscles in the
index finger pad of humans. J Comp Neurol 456, 260–266.
Perez-Pinera P, García-Suarez O, Germanà A, et al. (2008) Characterization of
sensory deficits in TrkB knockout mice. Neurosci Lett 433, 43–47.
Ranade SS, Woo SH, Dubin AE, et al. (2014) Piezo2 is the major transducer of
mechanical forces for touch sensation in mice. Nature 516, 121-125.
Reed-Geaghan EG, Wright MC, See LA, et al. (2016) Merkel cell-driven BDNF
signaling specifies SAI neuron molecular and electrophysiological phenotypes. J
Neurosci 36, 4362–4376.
Rittie L, Fisher GJ. (2015) Natural and sun-induced aging of human skin. Cold Spring
Harb Perspect Med 5, a015370.
Roudaut Y, Lonigro A, Coste B, Hao J, Delmas P, Crest M. (2012) Touch sense:
functional organization and molecular determinants of mechanosensitive
receptors. Channels 6, 234–245.
Sedy J, Szeder V, Walro JM, et al. (2004) Pacinian corpuscle development involves
multiple Trk signaling pathways. Dev Dyn 231, 551–563.
Shaffer SW, Harrison AL. (2007) Aging of the somatosensory system: a translational
perspective. Phys Ther 87, 193–207.
Skedung L, El Rawadi C, Arvidsson M, et al. (2018) Mechanisms of tactile sensory
deterioration amongst the elderly. Sci Rep 8, 5303.
Stark B, Risling M, Carlstedt T. (2001) Distribution of the neurotrophin receptors p75
and trkB in peripheral mechanoreceptors; observations on changes after injury.
Exp Brain Res 136, 101–107.

Toyoda M, Nakamura M, Nakada K, Nakagawa H, Morohashi M. (2005)
Characteristic alterations of cutaneous neurogenic factors in photoaged skin. Br
J Dermatol 153, 13–22.
Tseng MT, Chiang MC, Yazhuo K, Chao CC, Tseng WYI, Hsieh ST. (2013) Effect
of aging on the cerebral processing of thermal pain in the human brain. Pain
154, 2120–2129.
Ulfhake B, Bergman E, Fundin BT. (2002) Impairment of peripheral sensory
innervation in senescence. Auton Neurosci 96, 43–49.
Van Keymeulen A, Mascre G, Youseff KK, et al. (2009) Epidermal progenitors give
rise to Merkel cells during embryonic development and adult homeostasis. J
Cell Biol 187, 91–100.
Vega JA, Calzada B, Del Valle ME. (1993) Age-induced changes in the mammalian
autonomic and sensory ganglia. In Aging of the autonomic nervous system (ed.
Amenta F), pp. 37–67. Boca Raton, FL: CRC Press.
Vega JA, García-Suárez O, Montaño JA, Pardo B, Cobo JM. (2009) The Meissner
and Pacinian sensory corpuscles revisited new data from the last decade.
Microsc Res Tech 72, 299–309.
Vega JA, Lopez-Muniz A, Calavia MG, et al. (2012) Clinical implication of
Meissner’s corpuscles. CNS Neurol Disord Drug Targets 11, 856–868.
Verendeev A, Thomas C, McFarlin SC, Hopkins WD, Phillips KA, Sherwood CC.
(2015) Comparative analysis of Meissner's corpuscles in the fingertips of
primates. J Anat 227, 72–80.
Wickremaratchi MM, Llewelyn JG. (2006) Effects of ageing on touch. Postgrad Med
J 82, 301–304.

Wu G. (1998) Clustering of slowly adapting type II mechanoreceptors in human
peripheral nerve and skin. Brain 121, 265–279.
Wu J, Lewis AH, Grandl J. (2017) Touch, tension, and transduction – the function and
regulation of Piezo ion channels. Trends Biochem Sci 42, 57–71.
Wu M, Fannin J, Rice KM, Wang B, Blough ER. (2011) Effect of aging on cellular
mechanotransduction. Ageing Res Rev 10, 1–15.
Zelená J. (1994) Nerves and Mechanoreceptors. Chapman & Hall, London.
Zimmerman A, Bai L, Ginty DD. (2014) The gentle touch receptors of mammalian
skin. Science 346, 950–954.

SUPPLEMENTARY MATERIAL
Supplementary Fig. 1. Double immunofluorescence for BDNF (a) and TrkB (d) (green
fluorescence) and S100 protein (red fluorescence) in human digital Meissner’s
corpuscles of one 30-year-old subject. BDNF and S100 protein were colocalized,
suggesting that BDNF was present in the lamellar cells. Conversely, no colocalization
of TrkB and S100 protein was detected, indicating that TrkB is mainly located in axons.
Objective 63×/1.40 oil; pinhole 1.37; XY resolution 139.4 nm and Z resolution 235.8
nm. Scale bar: 30 µm.
Supplementary Fig. 2. Double immunofluorescence for BDNF (green fluorescence)
and NFP (b) and S100 protein (e) (red fluorescence) in human digital Meissner’s
corpuscles of one 30-year-old subject. BDNF and S100 protein were colocalized,
suggesting that BDNF is present in inner core cells. Conversely, no colocalization of
BDNF and NFP was detected, indicating that BDNF is not present in the axon.
Objective 63×/1.40 oil; pinhole 1.37; XY resolution 139.4 nm and Z resolution 235.8
nm. ic: inner core; arrow indicates the axon. Scale bar: 50 µm.
Supplementary Fig. 3. Double immunofluorescence for BDNF (a) and TrkB (d) (green
fluorescence) and CK20 (red fluorescence) in human digital Merkel cells (arrows).
BDNF and CK20 never colocalized, whereas TrkB and CK20 colocalized in the
cytoplasm of Merkel cells. Objective 63×/1.40 oil; pinhole 1.37; XY resolution 139.4
nm and Z resolution 235.8 nm. ic: inner core; arrow indicates the axon. Scale bar: 30
µm.
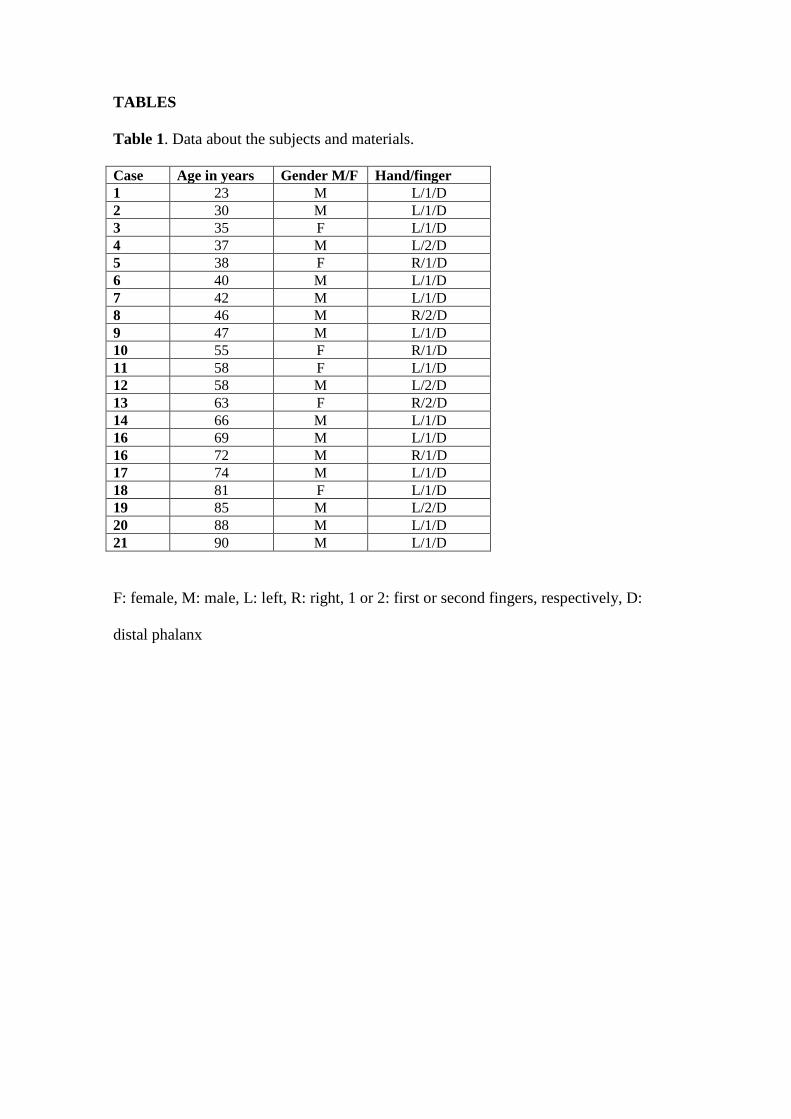
TABLES
Table 1. Data about the subjects and materials.
Case Age in years Gender M/F Hand/finger
1 23 M L/1/D
2 30 M L/1/D
3 35 F L/1/D
4 37 M L/2/D
5 38 F R/1/D
6 40 M L/1/D
7 42 M L/1/D
8 46 M R/2/D
9 47 M L/1/D
10 55 F R/1/D
11 58 F L/1/D
12 58 M L/2/D
13 63 F R/2/D
14 66 M L/1/D
16 69 M L/1/D
16 72 M R/1/D
17 74 M L/1/D
18 81 F L/1/D
19 85 M L/2/D
20 88 M L/1/D
21 90 M L/1/D
F: female, M: male, L: left, R: right, 1 or 2: first or second fingers, respectively, D:
distal phalanx

Table 2. Primary antibodies used in the study.
Antigen (clone) Origin Dilution Supplier Catalogue number
S100P Rabbit 1:1000 DAKO 1 Z 0311
P200 kDa NF (RT-97) Mouse 1:1000 Boehringer-Mannheim2 1178709
PGP 9.5 Rabbit 1:300 Abcam3 ab10404
Cytokeratin 20 Mouse prediluted Leica 4 PA0022
Piezo2 Rabbit 1:500 Sigma Aldrich5 HPA040616
BDNF Rabbit 1:200 Chemicon Int6 AB1534SP
TrkB Rabbit 1:200 Santa Cruz Biotechnology7 SC12
1Glostrup, Denmark; 2Mannheim, Germany; 3Hamburg, Germany; 4Newcastke, UK;
5Saint Louis, MO, USA; 6Temecula, CA, USA; 7Santa Cruz, CA, USA.
BDNF: brain-derived neurotrophic factor; NF: neurofilament; NSE: neuron-specific
enolase.
The antibody against BDNF is directed against the sequence H2N-HSDPARRGEL-
COOH (manufacturer’s notice); the antibody anti-TrkB was directed against the
residues 794-808 of the intracytoplasmic domain of human TrkB (manufacturer’s
notice); the amino acid sequence recognized by the antibody against Piezo2 is
FEDENKAAVRIMAGDNVEICMNLDAASFSQHNP (manufacturer’s information).

FIGURE LEGENDS
Fig. 1. Sections of glabrous digital skin from 3 subjects with ages of 23 (a), 55 (b) and
88 (c) years; immunostained was performed to evaluate S100 protein (S100P). Nerves
and Meissner’s corpuscles were immunolabelled (arrows). e: epidermis. Scale bar: 100
µm. The density of Meissner corpuscles in the three pre-established age ranges is shown
in the graph. A significant reduction in the density of Meissner’s corpuscles was
observed with ageing. *p<0.05, **p<0.01.
Fig. 2. Immunohistochemical localization of neurofilament protein (NFP) and S100
protein (S100P) in the axon and lamellar cells, respectively, of human digital
Meissner’s corpuscles of subjects of different ages: 23 (a, b), 30 (c, d), 40 (e, f), 55 (g,
h), 85 (I, j) and 90 (k, l) years. The localization, size, morphology and arrangement of
corpuscular constituents varied with ageing. Moreover, the Meissner’s corpuscles from
old subjects showed a marked decrease in the intensity of the immunoreaction for
S100P, and in some of these corpuscles, it was impossible to identify the axon. e:
epidermis. Scale bar: 60 µm.
Fig. 3. Digital Meissner’s corpuscles from three subjects with ages of 85 (a-d), 88 (e-h)
and 90 (i-l) years. Meissner’s corpuscles were localized in the dermal papillae, but most
were found in the reticular dermis. Those found at this location were smaller in the old
subjects than in the adults, showed a rounded morphology and lacked the typical
immunohistochemical profile. e: epidermis. Scale bar: 60 µm.
Fig. 4. Double immunofluorescence for S100 protein (green fluorescence) and
neurofilament protein (red fluorescence) in human digital Meisner’s corpuscles of
subjects with ages of 23 (a-c), 58 (d-f) and 90 (g-i) years. Sections were counterstained
with DAPI to ascertain structural details. Objective 63×/1.40 oil; pinhole 1.37; XY
resolution 139.4 nm and Z resolution 235.8 nm. Scale bar: 20 µm.

Fig. 5. Immunohistochemical localization of neurofilament protein (NFP) and S100
protein (S100P) in the axon and the inner core cells, respectively, of human digital
Pacinian corpuscles of subjects with ages of 23 (a-d), 55 (e-h), and 88 (i-l) years. The
axon supplying the corpuscles was identifiable in all corpuscles of 23- and 55-year-old
subjects and was not identified in the 88-year-old subject. The inner core cells displayed
S100P immunoreactivity in all lamellae in the 23- and 55-year-old subjects, whereas in
the older subjects, S100P immunoreactivity was restricted to the outer core lamellae.
Scale bar: 50 µm.
Fig. 6. Double immunofluorescence for S100 protein (green fluorescence) and
neurofilament protein (red fluorescence, arrows in b and e) in human digital Pacinian
corpuscles of subjects with ages of 23 (a-c) and 88 (d-f) years. Sections were
counterstained with DAPI to ascertain structural details. The axon and the inner core of
the 88-year-old subject are disarranged, although the corpuscle retained the general
organization. Objective 63×/1.40 oil; pinhole 1.37; XY resolution 139.4 nm and Z
resolution 235.8 nm. Scale bar: 50 µm.
Fig. 7. Immunohistochemical characterization of CK20 in human digital cells, identified
based on their localization as Merkel’s cells, in subjects of different ages: 23 (a), 30 (b),
55 (c), 72 (d), 88 (e) and 90 (f) years. Scale bar: 50 µm. The density of Merkel’s cells in
the three pre-established age ranges is shown in the graph. A significant reduction in the
density of Meissner’s corpuscles was observed with ageing. *p<0.05, **p<0.01.
Fig. 8. Schematic representation of Meissner’s corpuscles (MC) and Merkel’s cell-
neurite complexes (McNc) in the human digital glabrous skin of young-adult (top) and
old (bottom) subjects. Ageing results in a reduction in the density of both types of
mechanoreceptors as well as changes in the placement, size, morphology and
immunohistochemical profile of Meissner’s corpuscles. e: epidermis.

Fig. 9. Immunohistochemical detection of brain-derived neurotrophic factor (BDNF)
and its signalling receptor TrkB in Meissner’s corpuscles of subjects with ages of 23 (a,
e), 55 (b, f), 88 (c, g) and 90 (d, h) years. Scale bar: 60 µm.
Fig. 10. Sections of glabrous digital skin from 2 subjects with ages of 30 (a) and 88 (d)
years immunostained to detect Piezo2. The axon of most Meissner’s corpuscles
displayed Piezo2 immunoreactivity in young individuals (arrows in a, b), whereas this
immunoreactivity was restricted to segments of the corpuscle, or even absent in the
older subjects (e). The density of Piezo2-positive Merkel cells in young subjects (c) was
higher than that in the old subjects (f, arrow). e: epidermis. Scale bar: 100 µm a and d;
20 µm b-c and e-f.
Fig. 11. Double immunofluorescence for neuron-specific enolase (green fluorescence a
and b), neurofilament proteins (green fluorescence in c-f; red fluorescence in g),
cytokeratin 20 (red fluorescence in a-f), and Piezo2 (green fluorescence in g) in human
Merkel’s cell-neurite complexes of subjects with ages of 30 (a-c), and 88 (d-f) years.
Axonal profiles are indicated by arrows. e: epidermis. Objective 63×/1.40 oil; pinhole
1.37; XY resolution 139.4 nm and Z resolution 235.8 nm. Scale bar: 30 µm a,c,d,e and
f; 20 µm b and g.
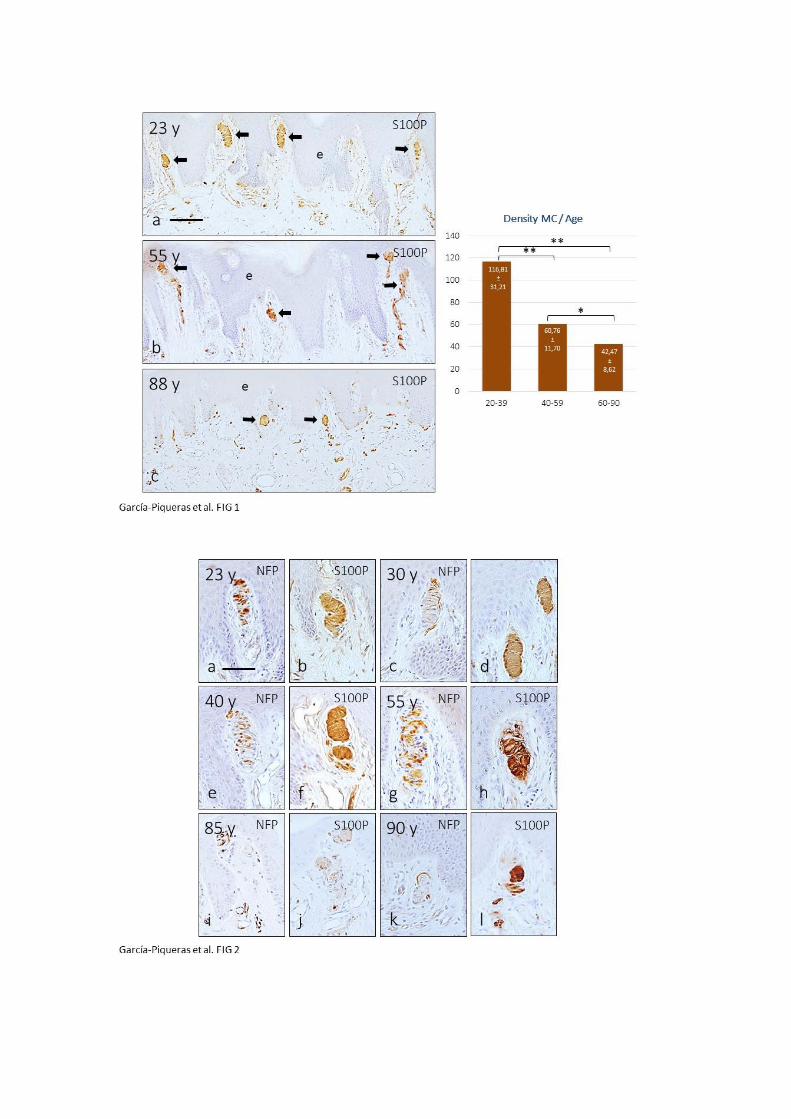

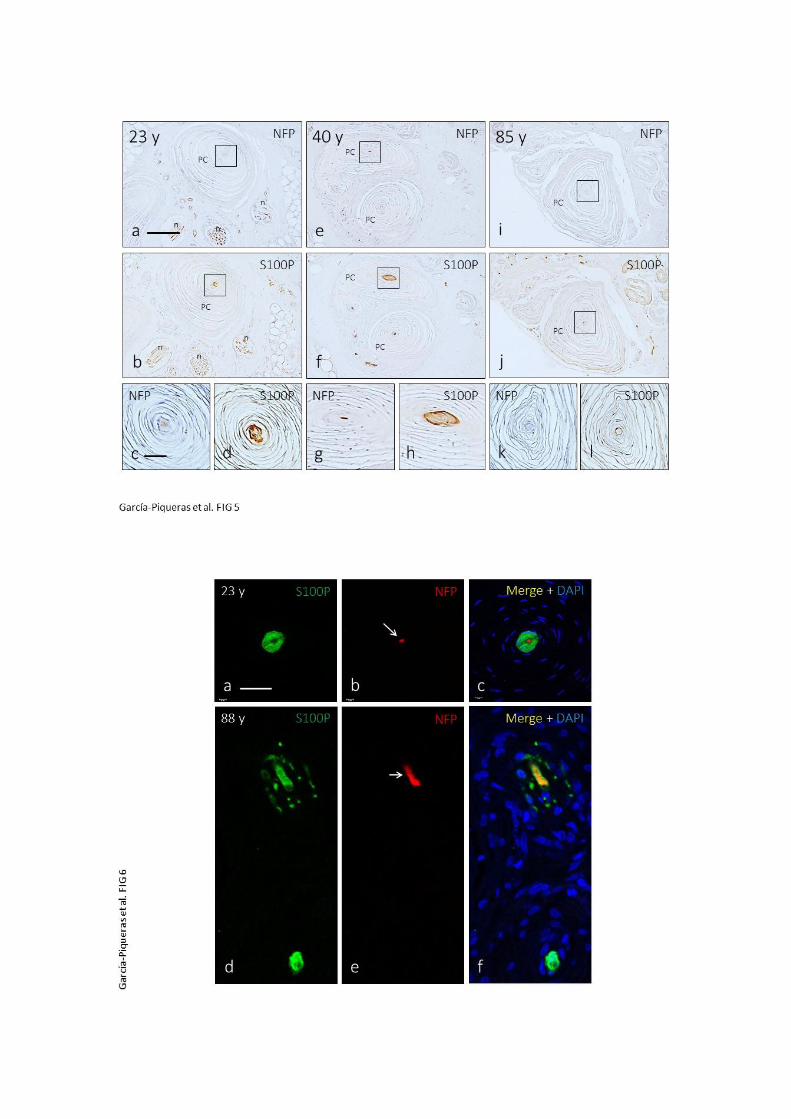
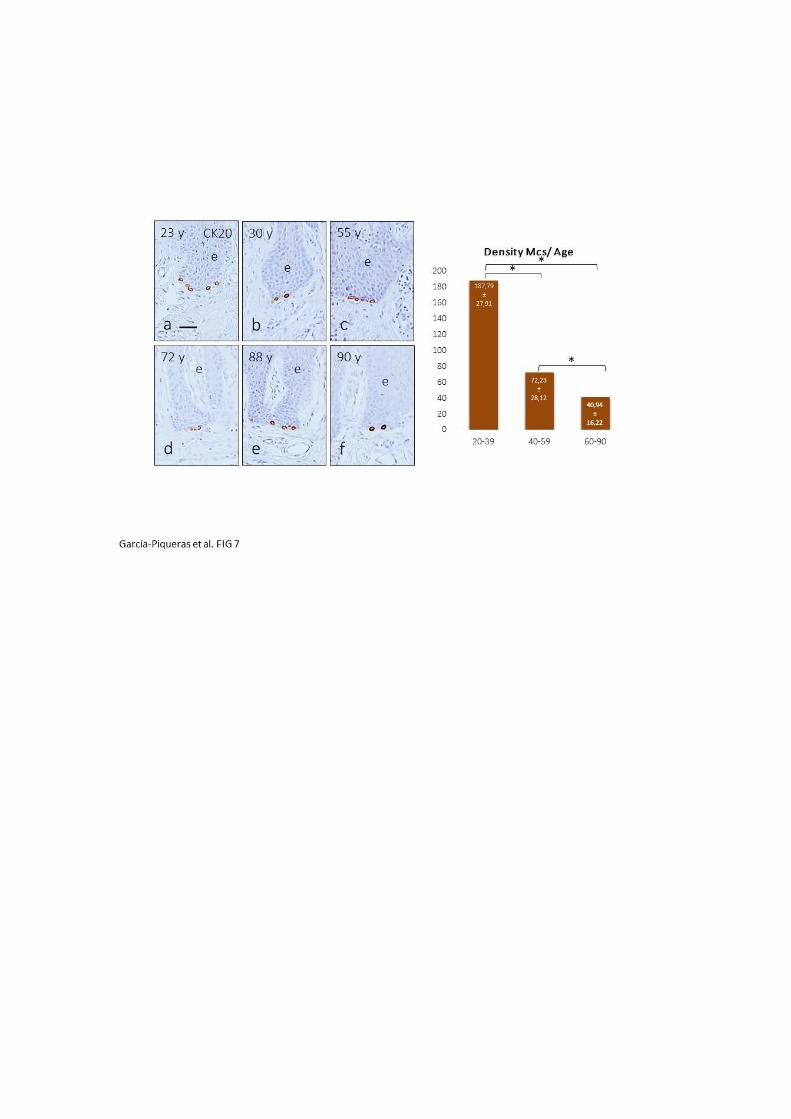
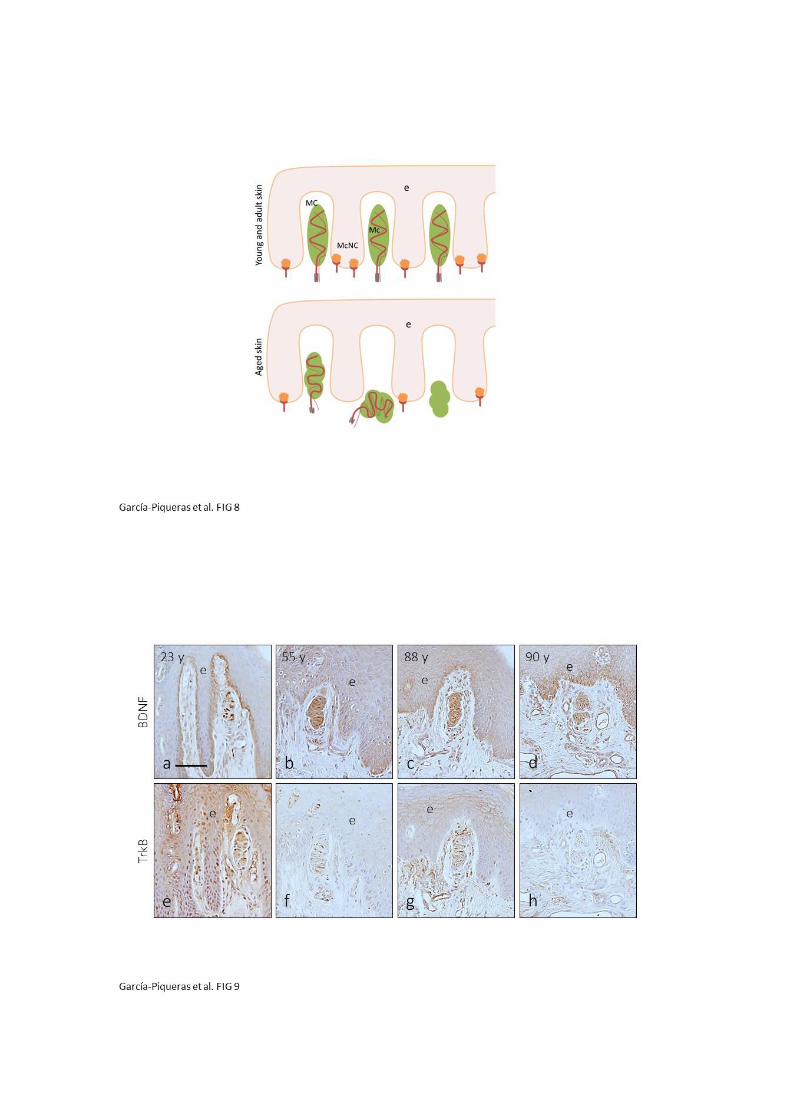
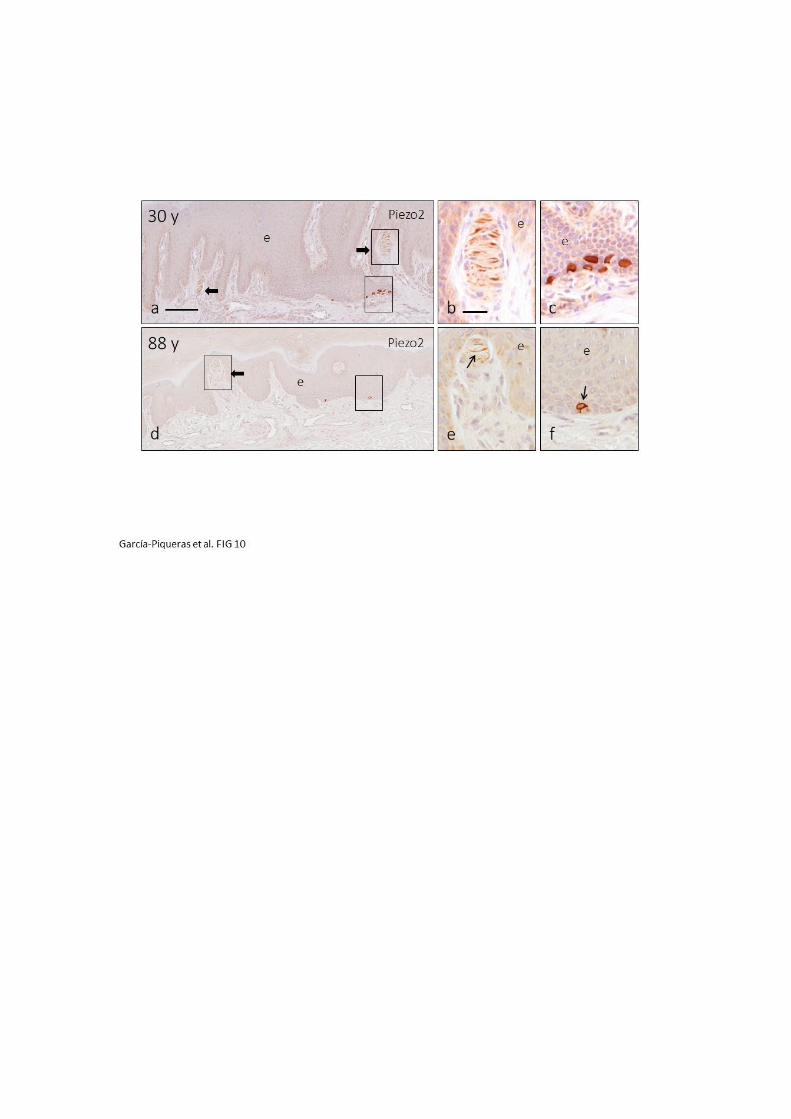
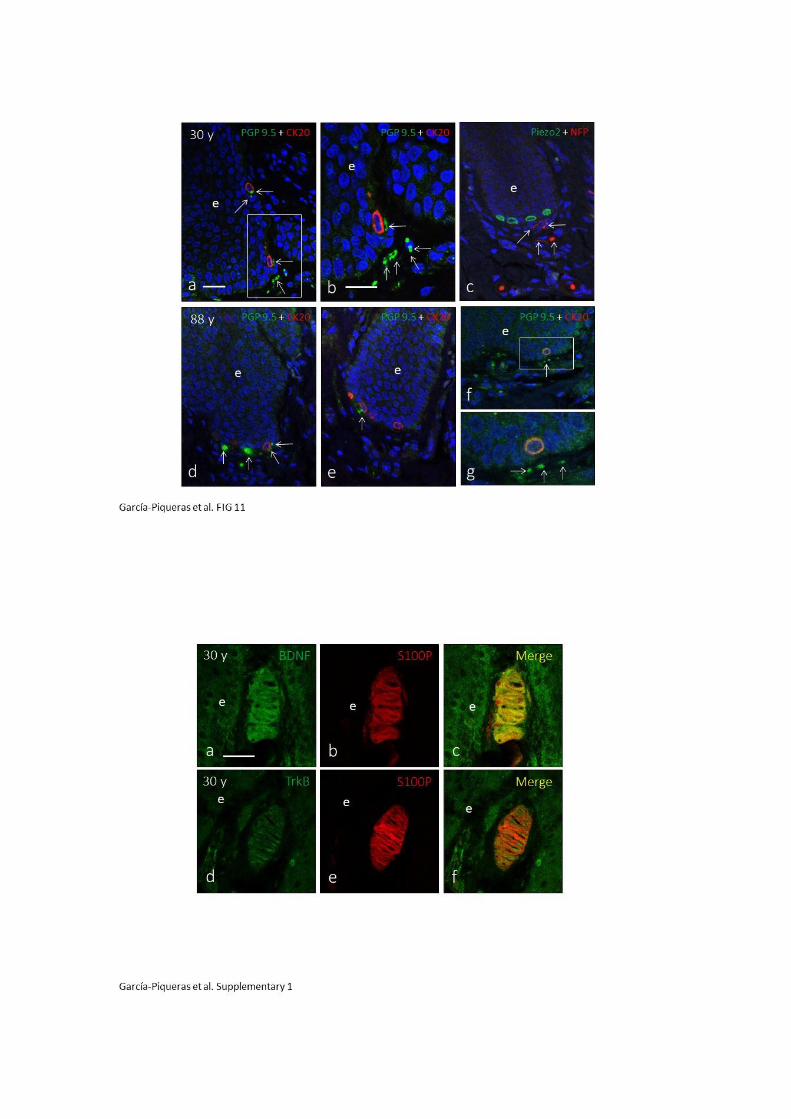
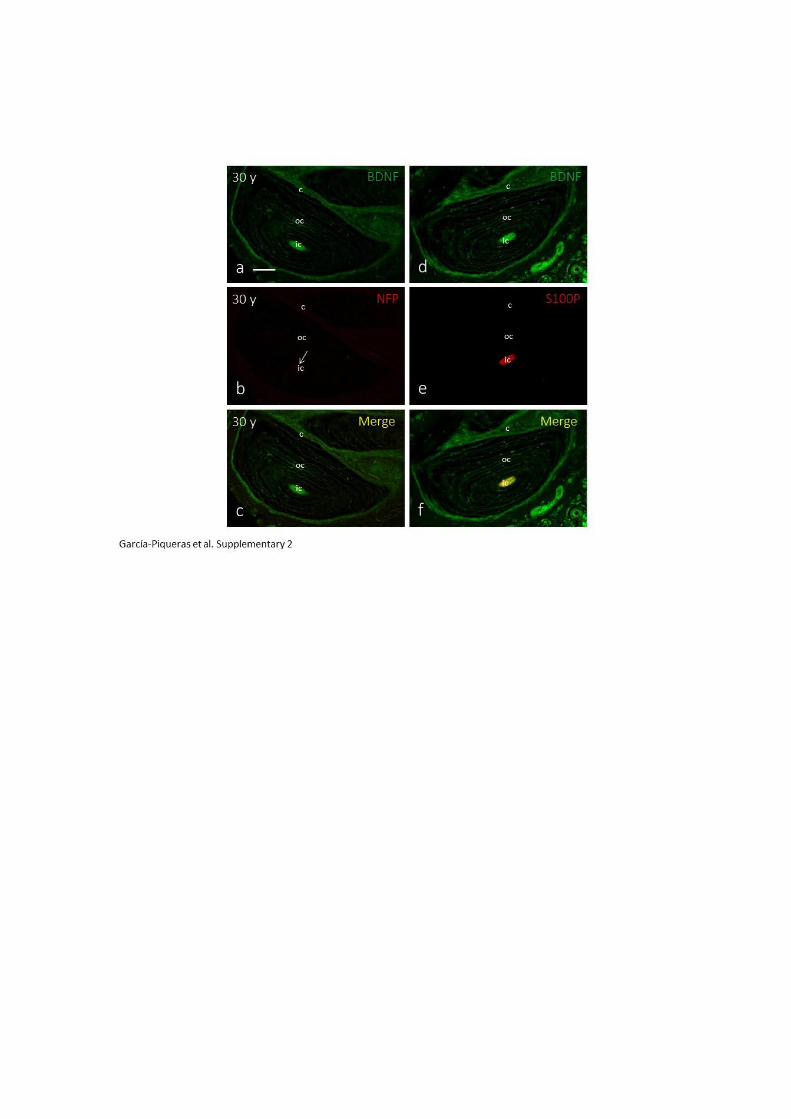


6. DISCUSIÓN

6.1. Consideraciones generales
El presente trabajo de tesis doctoral se enmarca en una de las líneas de investigación
que, durante los últimos 35 años, desarrolla el grupo SINPOS (Cluster de Biomedicina,
Campus de Excelencia internacional de la Universidad de Oviedo) en el Área de
Anatomía y Embriología Humana del Departamento de Morfología y Biología Celular
de la Universidad de Oviedo, y que se dedica al estudio de los corpúsculos sensitivos
los cutáneos. La mayor parte de las contribuciones de esta línea de investigación se
recogen en tres revisiones publicadas por J.A. Vega y colaboradores en 1996, 2009 y
2012.
La supervivencia de un organismo depende en gran medida de su capacidad de
respuesta y adaptación al medio ambiente que lo rodea y una de las formas de
interactuar con él es a través del tacto. Los mamíferos han desarrollado estructuras
sensitivas especializadas que les permiten detectar tacto fino, vibraciones y presión
(Zelena, 1994; Munger e Idé, 1998; Johnson, 2001). En conjunto, estas formaciones
nerviosas se denominan corpúsculos sensitivos y funcionalmente son
mecanorreceptores.
Los corpúsculos de Meissner y de Pacini son dos tipos de formaciones nerviosas
sensitivas que, aunque son diferentes en estructura y organización celular, se
comportan fisiológicamente de forma casi idéntica, ya que ambos son receptores de
bajo umbral y adaptación rápida y dependen de fibras nerviosas sensitivas Aα o Aβ
(Johnson, 2001). En cambio, los complejos célula de Merkel-neurita son estructuras
mecanorreceptoras formadas por una asociación sinapsis-like entre axones de
neuronas sensitivas Aβ y células epiteliales especializadas (Fleming y Lou, 2013). A
pesar del conocimiento sobre las propiedades y características neurofisiológicas, no ha
sido hasta estos últimos años cuando se han empezado a identificar los mecanismos
moleculares y celulares mediante los cuales un estímulo mecánico se convierte en una
señal eléctrica. Detrás de esta transformación mecanoeléctrica se encuentran canales
iónicos que se expresan en las células de los mecanorreceptores y que son capaces de
activarse mediante factores mecánicos (Del Valle y Cols., 2012; Kuipers y Cols., 2012;
Zimmerman y Cols., 2014).

La estructura celular y la composición proteica de los mecanorreceptores sensoriales
están bien definidas y caracterizadas (Vega y Cols., 1996; 2009; Pawson y Cols., 2000).
No obstante, el conocimiento sobre algunos aspectos como el origen celular, la matriz
extracelular y los cambios edad-dependientes de los corpúsculos sensitivos resultan
incompletos o son totalmente desconocidos a día de hoy.
Con el presente trabajo de investigación, se aportan nuevos datos sobre la naturaleza
de las células que componen el núcleo externo de los corpúsculos de Pacini, sobre los
proteoglicanos y glicosaminoglicanos de la matriz extracelular de los corpúsculos
sensitivos y sobre las variaciones cuantitativas, cualitativas y morfológicas que cursan
los mecanorreceptores sensoriales en función de la edad.

6.2. Las células CD34-positivas definen una lámina intermedia en los
corpúsculos de Pacini digitales humanos
En el primer trabajo de esta tesis, se ha analizado el origen celular del núcleo externo
de los corpúsculos de Pacini de la piel digital glabra humana. Para ello se han tenido en
cuenta las siguientes premisas: 1) las vainas endoneurales o perineurales de los nervios
periféricos son continuas con las capas periaxónicas de los corpúsculos de Pacini
(Malinovsky, 1986); 2) el endoneuro de los troncos de los nervios periféricos expresa el
antígeno CD34 (Weiss et al., 1993; Khalifa et al., 2000; Hirose et al., 2003; Richard et
al., 2014); 3) los antígenos Glut1 y EMA se expresan en el núcleo externo y en la
cápsula de los corpúsculos de Pacini (Vega y Cols., 2009; Feito y Cols., 2016).
Diversos estudios realizados en corpúsculos de Pacini de diferentes especies animales
han constatado mediante inmunohistoquímica y, sobre todo, mediante microscopía
electrónica de transmisión, la existencia de una lámina intermedia en los corpúsculos
de Pacini. González-Martínez y Cols. (2007) describieron una lámina Erb4-positiva en el
exterior del núcleo interno de corpúsculos de Pacini de ratón; una lámina intermedia o
de crecimiento ha sido caracterizada al detalle con microscopía electrónica en varias
ocasiones en corpúsculos de Pacini del mesenterio del gato (Pease y Quilliam, 1957;
Munger y Cols., 1988) y de la pata delantera de perro (Rico y Cols., 1996); Chouchkov
(1978) describió un espacio subcapsular estrecho con fibroblastos y macrófagos
dispersos y fibrillas de colágeno alrededor del núcleo interno de corpúsculos de Pacini
del mesenterio de gato; un patrón especial compuesto por lectinas, proteoglicanos y
glicosaminoglicanos fue también descrito en la zona equivalente a la lámina
intermedia en corpúsculos de Pacini del peritoneo de gato (Pawson y Cols., 2000;
Sames y Cols., 2001); una zona inmunorreactiva para proteína quinasa C fue
identificada con la lámina intermedia en corpúsculos de Pacini de las patas traseras de
la rata (Kawakita y Cols., 1992).
Sin embargo, en corpúsculos de Pacini humanos, esta lámina intermedia nunca había
sido identificada como tal. Tan sólo fue sugerida en un primer momento en base a la
presencia de algunos antígenos específicos (como la vimentina) entre el núcleo interno
y el núcleo externo (García-Suárez y Cols., 2010).

La lámina intermedia CD34-positiva identificada y descrita en este trabajo se asemeja a
la lámina intermedia o de crecimiento del corpúsculo de Pacini del mesenterio de gato
y con las células del espacio subcapsular estrecho (Chouchkov, 1978).
Estructuralmente divide el corpúsculo de Pacini en dos compartimentos: el
compartimento interno neural y avascular (axón y células de Schwann-modificadas) y
el compartimento externo no neural y vascular (células perineurales y epineurales,
vasos sanguíneos y macrófagos). Aunque su función queda por concretar, podría estar
relacionada con el potencial crecimiento del corpúsculo de Pacini (Zelena, 1994).
Además, las células que expresan CD34 tienen capacidades potenciales de
diferenciación y proliferación (Sidney y Cols., 2014). CD34 es una glicoproteína de
membrana que actúa principalmente como molécula de adhesión (Silva y Cols., 2010),
conocida principalmente por expresarse en las células madre hematopoyéticas, pero
también en células mesenquimales potencialmente diferenciables (Sidney et al., 2014).
Por otro lado, se puede hipotetizar que CD34 interacciona con la membrana basal de
lamela más externa del núcleo interno o con otros componentes de la matriz
extracelular (Dubový y Brednarova, 1999), aunque se necesitan más estudios sobre la
composición de la matriz extracelular de los corpúsculos de Pacini, ya que se
desconoce casi por completo.
Por otro lado, en base al diámetro de las fibras de colágeno del espacio extracelular,
algunos autores han sugerido que la lámina intermedia de los corpúsculos de Pacini de
felinos es de origen endoneural (Osawa e Ide, 1986), así como en los de rata, según la
expresión de la proteína quinasa C beta (Kawakita et al., 1992). Los resultados
obtenidos en este trabajo demuestran el origen endoneural de la lámina intermedia ya
que se corresponde con la expresión de CD34 en las células endoneurales de los
troncos nerviosos. Además, siguiendo la continuidad anatómica entre los nervios
periféricos y los corpúsculos sensitivos (Malinovsky, 1986) y sus perfiles
inmunohistoquímicos comunes (Vega y Cols., 1996, 2009), se establece una relación
entre el perineuro y el núcleo externo de los corpúsculos de Pacini en base a la
expresión de los antígenos EMA y Glut1 (Vega y Cols., 2009; Feito y Cols., 2016).

6.3. Condroitín sulfato en los corpúsculos sensitivos de Meissner y de
Pacini humanos
En el segundo trabajo que conforma el cuerpo de la tesis, se ha estudiado la expresión
de condroitín sulfato en los corpúsculos sensitivos de piel glabra digital humana.
Las moléculas que conforman la matriz extracelular se clasifican en dos categorías:
proteinas fibrosas (colágenos, elastina, laminina, fibronectina, etc.) y proteoglicanos
(entre ellos, los formados por condroitín sulfato). Dubovy y Bednárová (1999)
demostraron que el núcleo interno y la capsula de los corpúsculos de Pacini de rata
tienen diferentes patrones estructurales de proteoglicanos y que, probablemente,
estén implicados en diferentes funciones.
En esta investigación se ha determinado que los corpúsculos de Pacini humanos
expresan una lámina de condroitín sulfato dispuesta entre el núcleo interno y el núcleo
externo, la cual es similar a la descrita en los corpúsculos de Pacini del mesenterio de
gato (Sames y Cols., 2001).
Además, se ha demostrado que esta lámina de condroitín sulfato está asociada a la
lámina CD34 positiva descrita en el apartado anterior (García-Piqueras y Cols., 2017).
Ocurre lo mismo en el caso de los nervios periféricos de los vertebrados, en los que los
proteoglicanos de condroitín sulfato se asocian a las células endoneurales (CD34-
positivas; García-Piqueras y Cols., 2017) pero no a las epineurales (Graham y Muir,
2016). Este resultado es una evidencia más que avala la continuidad de los corpúsculos
de Pacini con los nervios periféricos y, al mismo tiempo, que están formados por los
mismos elementos (axón, células de Schwann, endoneuro, perineuro y
epineuro/cápsula).
Otro resultado relevante es la no expresión de condroitín sulfato y CD34 en los
corpúsculos de Meissner. La ausencia de elementos perineurales (cápsula) en los
corpúsculos de Meissner puede estar relacionada con la ausencia de elementos
endoneurales (objeto de investigación). El endoneuro actúa probablemente como zona
de crecimiento a partir de la cual se genera el perineuro, lo cual puede ser importante
para los corpúsculos de Pacini pero no para los de Meissner.

A pesar de que la función de los proteoglicanos de condroitín sulfato en los nervios
periféricos ha sido determinada y es bastante bien conocida (Graham y Muir, 2016), su
papel en los corpúsculos sensitivos es todavía dudoso. Los proteoglicanos de
condroitín sulfato pueden estar relacionados con la separación del segmento neural
(axón y núcleo interno) del no neural (núcleo externo y cápsula) del corpúsculo de
Pacini, pero también con la mecanotransducción. Aunque los canales iónicos son los
actores principales de la mecanotransducción, se ha demostrado que en el proceso
también participan algunas moléculas de la matriz extracelular (Silver y Siperko, 2003;
Lumpkin y Caterina, 2007; Arnadottir y Chalfie, 2010; DuFort y Cols., 2011). En este
sentido, Matthews y Cols. (2006) afirman que el potencial de acción depende de
específicas deformaciones mecánicas de la membrana axonal y de interconexiones con
la matriz extracelular en terminales nerviosos. Se desconoce todavía qué moléculas de
dicha matriz pueden estar implicadas directamente en la mecanotransducción.
Respecto a la función de control del crecimiento axónico, los proteoglicanos de
condroitín sulfato son inhibidores del mismo, ya que su degradación enzimática
elimina la actividad inhibitoria (Muir, 2010; Graham y Muir, 2016; Man y Cols., 2014).
Se ha demostrado que aplicando condroitinasa in vivo, se puede mejorar el resultado
de la reparación nerviosa (Graham y Muir, 2016). La asociación de los proteoglicanos
de condroitín sulfato a las células endoneurales CD34-positivas en los corpúsculos de
Pacini probablemente regule el correcto crecimiento axónico dentro del
compartimento neural del corpúsculo, aunque no hay datos sobre ello. Sin embargo,
esto es respaldado parcialmente por el hecho de que los corpúsculos de Pacini
denervados cursan con reinervación poliaxonal (Zelená, 1984; Zelená y Zacharova,
1997; Zelená y Jirmanova, 1988, 1995). La reinervación poliaxonal se debe
probablemente a la lámina intermedia, en concreto a los proteoglicanos de condroitin
sulfato, que resultan alterados gravemente e incapaces de inhibir correctamente el
crecimiento axonal (sprouting) durante la reinervación. También, algunos de estos
axones reinervan el núcleo interno orginal (Zelená, 1994).

6.4. El heparán sulfato está asociado a la membrana basal de los
corpúsculos de Meissner y Pacini cutáneos humanos
El tercer artículo continúa con el estudio de otro de los glicosaminoglicanos, el heparán
sulfato, en los corpúsculos sensitivos cutáneos humanos.
En este estudio, se ha demostrado la expresión de heparán sulfato en los corpúsculos
de Meissner y de Pacini, asociada casi exclusivamente a proteoglicanos que forman
parte de la membrana basal con localización pericelular y que corresponden a
perlecano, agrina, colágeno XV y colágeno XVIII (Yurchenco y Patton, 2009; Iozzo y
Schaefer, 2015).
La membrana basal de los corpúsculos de Meissner y de Pacini ya ha sido estudiada
previamente mediante microscopía electrónica (Malinovsky y Pac, 1982; Malinovsky,
1986; Ide y Cols., 1987; Ide y Hayashi, 1987; Munger y Ide, 1988) y por métodos
inmunohistoquímicos (Vega y Cols., 1995). En el presente trabajo, se ha utilizado la
microscopía confocal, utilizando como base la presencia de colágeno IV (uno de los
componentes principales de las membranas basales junto a las lamininas; Vega y Cols.,
1995) para asociar la expresión de heparán sulfato a la membrana basal. En
consecuencia, la colocalización de heparán sulfato con colágeno IV se interpreta como
proteoglicanos de heparán sulfato pertenecientes a la membrana basal, mientras que
la no colocalización indica la existencia de otros proteoglicanos de heparan sulfato de
superficie celular (no pertenecientes a la membrana basal), como son las familias de
sindecanos o glipicanos (Iozzo y Schaefer, 2015).
En los corpúsculos de Meissner, los proteoglicanos de heparán sulfato se encuentran
asociados de la membrana basal, lo cual confirma estudios previos de nuestro
laboratorio realizados mediante la inmunodetección de colágeno IV y laminina (Vega y
Cols., 1995). Sin embargo, también se encontraron otros proteoglicanos de heparán
sulfato no asociados a la membrana basal.
En los corpúsculos de Pacini, los proteoglicanos de heparan sulfato se encuentran
asociados exclusivamente a la membrana basal. No obstante, la presencia de
membrana basal en todos los compartimentos del corpúsculo sigue siendo objeto de
debate. Mientras que algunos autores han descrito la membrana basal tanto en las

células del núcleo interno como en la lámina intermedia, bien con microscopía
electrónica (Malinovsky y Pac, 1982; Malinovsky, 1986), bien con inmunohistoquímica
para colágeno IV y laminina (Vega y Cols., 1995), otros afirman que dichos
compartimentos celulares carecen de membrana basal (Ide y Hayashi, 1987; Munger y
Ide, 1988). Los resultados de este trabajo apoyan esta segunda teoría, ya que no se ha
detectado ni heparan sulfato ni colágeno IV en estas dos localizaciones. A pesar de
ello, sí hay acuerdo sobre la existencia de membrana basal alrededor de las células del
núcleo externo y de la cápsula (Malinovsky, 1986; Ide y Cols., 1987; Munger y Ide,
1988). Esta investigación también confirma este hecho, a partir de la colocalización del
heparán sulfato con el colágeno IV en todas las lamelas del núcleo externo y la cápsula.
Pero el resultado más relevante es la detección una intensa inmunorreacción de
heparán sulfato en las lamelas más internas del núcleo externo. Este área coincide con
la denominada “zona de transición” (Vega y Cols., 1995), la cual se pensaba
previamente que era de origen perineural (Halata y Cols., 1990), pero que
recientemente se ha caracterizado como endoneural (García-Piqueras y Cols., 2017).
Por lo que a partir de la no colocalización de CD34 y HS, se puede concluir que dicha
lámina intermedia carece de membrana basal.
Por otra parte, es conocido que las células que forman los compartimentos del
corpúsculo de Pacini se comunican entre ellas por diferentes tipos de unión celular en
sus membranas: entre las células del núcleo interno existen uniones tipo “gap”,
mientras que en las células del núcleo externo hay uniones tipo “thigh”. Hide y Hayashi
(1987) describieron que estas uniones tipo “thigh” eran “especialmente prominentes
en la primera y segunda lamelas más internas del núcleo externo (a veces también en
la tercera)”. Estas primeras lamelas del núcleo externo con mayor número de uniones
tipo “thigh” coinciden con la intensa expresión de heparan sulfato y colágeno IV
observada en este trabajo. La especial organización de uniones tipo “thigh” presentes
en las primeras lamelas del núcleo externo guarda relación con el aislamiento del
núcleo interno respecto al externo, en modo tal de evitar pérdidas de iones y fluidos
entre ambos, y prevenir que una posible perturbación en el núcleo externo pueda
cambiar la composición eléctrica del núcleo interno (Ide y Hayashi, 1987; Munger e
Ide, 1988). Es por ello que, la membrana basal por sí misma y/o los proteoglicanos de

heparan sulfato inmunodetectados más intensamente en esta zona podrían participar
en dicho aislamiento del núcleo interno-axón (e incluso también de la lámina
intermedia) respecto al núcleo externo-cápsula. Además, se sabe que las moléculas de
la matriz extracelular de las células perineurales son capaces de regular la presión
endoneural (Piña-Oviedo y Ortiz-Hidalgo, 2008).
Diferentes investigaciones han determinado que algunos componentes de la
membrana basal, así como de la matriz extracelular en general, están implicados en el
proceso de la mecanotransducción (Wetzel y Cols., 2007; Hu y Chiang, 2010; Chiang y
Cols., 2011). Queda por establecer ahora si los proteoglicanos de heparan sulfato
también están implicados en esta función.
En resumen, se ha demostrado que los proteoglicanos de heparán sulfato están
vinculados fundamentalmente a la membrana basal en ambos tipos de corpúsculos y
que la ausencia de los mismos en el núcleo interno y la lámina intermedia de los
corpúsculos de Pacini sugiere que estos compartimentos carecen de membrana basal.

6.5. SLRPs
Otro los de los aspectos abordados en el presente trabajo de tesis doctoral en relación
con la matriz extracelular de los corpúsculos sensitivos ha sido la distribución en ellos
de los componentes de las familias I y II de los proteoglicanos denominados pequeños
y ricos en leucina (small leucine-rich proteoglycans, SLRPs). Al igual que el resto de los
componentes de la matriz extracelular su localización en los corpúsculos es,
necesariamente, en los espacios dispuestos entre las láminas que los forman.
La composición molecular de la matriz extracelular en los corpúsculos sensitivos es
relativamente poco conocida, y no existe casi ninguna información en humanos salvo
el trabajo de Vega y Cols. (1995). En la actualidad se conoce bastante bien la
distribución de los distintos tipos de colágeno en los espacios intercelulares de los
corpúsculos de Pacini: el colágeno de tipo II predomina en el núcleo externo y la
cápsula, el de tipo V en la capa intermedia, y el de tipo IV está presente en todos los
compartimentos (Vega y Cols., 1995; Pawson y Cols., 2000). También se conoce que en
algunas zonas de los corpúsculos la matriz extracelular se organiza para formar una
membrana basal (Malinosky y Cols., 1986; Vega y Cols., 1995; Dubovy y Aldskogius,
1996; García-Piqueras y Cols., 2018). Sin embargo, la distribución en los corpúsculos de
los otros componentes principales de la matriz extracelular, es decir, los
proteoglicanos (PGs; Iozzo y Schaefer, 2015; Theocharis y Cols., 2016), es poco
conocida. Sólo el trabajo de Sames y Cols. (2001) aporta datos muy parciales en los
corpúsculos de Pacini del mesenterio del gato.
El grupo de los SLRPs está formado por, al menos, 18 miembros subclasificados en las
clases de la I a la V. En concreto, las clases I y II que son las analizadas en este trabajo
incluyen al biglicano (BGN), decorina (DCN), asporina (ASPN), proteína 2 de la matriz
extracelular (ECM2), la proteína X de la matriz extracelular (ECMX), la fibromodulina
(FMOD), el lumicano (LUM), la prolargina (PRELP), el keratocano (KERA) y la
osteoadherina (OMD) (McEwan et al., 2006; Schaefer and Schaefer, 2010; Chen and
Birk, 2013).
Se ha detectado DCN en el núcleo externo y BGN en el núcleo interno; lo primero está
en consonancia con lo observado por Sames y Cols. (2001) en los corpúsculos de Pacini

del mesenterio del gato. La distribución del BGN difiere de la de estos autores ya que
ellos lo encuentran restringido al núcleo interno y en nuestro trabajo la hemos
detectado tanto en el interno como en el externo. La función de estas dos moléculas
en los corpúsculos de Pacini es desconocida en la actualidad. Es posible que DCN
participe en la estabilización de la vimentina del citoesqueleto (Jungmann y Cols.,
2012), que es el filamento intermedio de las células de ambos núcleos corpusculares
(ver Vega y Cols., 2009), pero ello tiene que ser confirmado en estudios posteriores.
Los datos que se aportan sobre la distribución del resto de los SLRPs estudiados son
completamente nuevos y, por tanto, no permite la comparación ni discusión con
estudios previos.
En cuanto al papel de los SLRPs en los corpúsculos de Pacini, todo lo que se puede
hacer en la actual situación, es hipotetizar. Algunos ensayos bioquímicos han
demostrado que los SLRPs modulan la organización de la matriz extracelular uniéndose
a los diferentes tipos de colágeno (Kalamajski and Oldberg, 2010; Chen and Birk 2013;
Gubbiotti et al., 2016). En concreto LUM se une al colágeno II, BGN y FMOD a los
colágenos II y VI, y DCN a los colágenos II, IV, V y VI (Chen and Birk, 2013). La
distribución de SLRPs observada en este trabajo y la previamente aportada por otros
estudios acerca de la localización corpuscular de los diferentes colágenos (Tachibana y
Cols., 1995; Vega y Cols., 1995; Pawson y Cols., 2000) apoya los datos bioquímicos.
Por otro lado, SLRPs regulan la actividad celular al interactuar con receptores para
diferentes factores de crecimiento (Merline y Cols., 2009; Dellet y Cols., 2012; Iozzo y
Schaefer 2015; Neill y Cols., 2015). DCN y BGN se unen al receptor del factor de
crecimiento epidérmico (EGFR) y a Erb4 (Schaefer y Iozzo, 2008; Schaefer y Schaefer,
2010; Jacob y Cs-Szabo, 2012) y la localización de estos SLRPs observada en los
corpúsculos de Pacini es similar a la del ERGR tanto en humanos (Vega et al., 1994)
como en ratón (Gonzalez-Martinez et al., 2007). Y se ha demostrado que la interacción
de DCN con EGFR regula la formación de colágeno (Mohan et al., 2011). Además, la
proporción relativa de colágenos y SLRPs determina las propiedades mecánicas de los
tejidos (Hansen et al., 2015).

En la conclusión, esta parte de nuestro estudio proporciona por primera vez un mapa
de la localización de los SLRPs en los corpúsculos de Pacini humanos. Sus complejos
patrones de expresión sugieren funciones muy diversas desde determinar la
elasticidad de los corpúsculos de Pacini, hasta condicionar sus características
mecánicas para la transmisión correcta de la presión.

6.6. El envejecimiento en la periferia del sistema somatosensorial:
cambios edad-dependientes en los mecanorreceptores cutáneos
En el quinto trabajo de esta tesis se ha realizado un estudio de los cambios edad-
dependientes que aparecen en los mecanorreceptores cutáneos (en concreto, células
de Merkel, corpúsculos de Pacini y corpúsculos de Meissner) como consecuencia del
envejecimiento. Además, se han analizado a diferentes edades, por un lado, el sistema
neurotrófico TrkB-BDNF, del cual depende el desarrollo y mantenimiento de los
corpúsculos sensitivos y, por otro lado, la expresión de la proteína Piezo2, principal
mecanotransductor de los corpúsculos sensitivos para el tacto fino.
Los corpúsculos sensitivos de la piel son capaces de detectar diferentes cualidades de
la somatosensibilidad. Los complejos célula de Merkel-neurita, los corpúsculos de
Meissner y los corpúsculos de Pacini actúan como mecanorreceptores SAI y SAII para
discriminar diferentes aspectos del tacto (Zimmerman et al., 2014). Con la edad, se
experimenta un progresivo deterioro del sistema somatosensorial a todos los niveles,
desde la piel hasta la corteza cerebral, incluyendo los recetores cutáneos en los que
tiene lugar la transducción de los estímulos mecánicos en impulsos eléctricos.
Evidentemente, dicha perdida de sensibilidad táctil afecta a la calidad de vida de las
personas (Skedung et al., 2018).
Diferentes autores afirman que la capacidad de percepción del tacto y por
consiguiente, la agudeza espacial táctil, se deterioran con la edad (Decorps y Cols.,
2014; Heft y Robinson, 2017). Esto puede deberse al menos en parte a una reducción
de la densidad de los corpúsculos de Meissner y de Pacini (Wickremaratchi y Llewelyn,
2006), así como de las células de Merkel (Lumpkin y Cols., 2003).
Mediante estudios inmunohistoquímicos se ha observado que los corpúsculos de
Meissner permanecen invariables en morfología y estructura hasta aproximadamente
los 60 años. A partir de entonces, tienen menor tamaño, adquieren forma redondeada,
aparecen topográficamente más profundos en la dermis (fuera de la papila dérmica) y
pierden parte de su perfil inmunohistoquímico característico. Así sucede en muchos
casos correspondientes a sujetos de edad más avanzada, en los que no se ha
detectado inmunorreacción para los marcadores axonales, lo que sugiere que se

produce una denervación del corpúsculo que, además se refleja en la reducción de
expresión de proteína S100 en las células lamelares. Es conocido que la expresión de la
proteína S100 por las células lamelares depende de la integridad funcional del axón
(Márques y Cols., 1997; Albuerne y Cols., 1998). Estos resultados concuerdan con los
cambios morfológicos descritos por Nava y Mathewson (1996) en los corpúsculos de
Meissner de las almohadillas digitales de ratones de avanzada edad, como
consecuencia de axonopatía distal y atrofia de las neuronas sensitivas. Sin embargo,
hay que mencionar que estos cambios afectan a un gran número de los corpúsculos de
Meissner analizados, pero no a todos, y que seguramente, la denervación observada
no se produzca por igual a lo largo de toda la piel en sus diferentes áreas anatómicas
del cuerpo (Besné y Cols., 2002). Recientemente, un estudio en el que se ensaya un
método para testar la agudeza táctil ha demostrado que los individuos con menor
rendimiento (los de mayor edad) poseen una densidad de corpúsculos de Meissner un
50% menor que los individuos con mayor rendimiento (los más jóvenes) (Skedung y
Cols., 2018). La primera explicación para esta pérdida de agudeza táctil podría ser el
deterioro asociado a la edad de las propiedades neurales, aunque habría que
considerar también el análisis de otros corpúsculos sensitivos.
Respecto a los corpúsculos de Pacini, no se han observado diferencias relevantes ni en
número ni en estructura en función de la edad, aunque en algunos casos
correspondientes a los sujetos de edad más avanzada, no ha sido posible detectar el
axón o presentaban un núcleo interno desestructurado (con pérdida parcial de
inmunorreacción para la proteína S100 en el núcleo interno). Esto podría explicar la
reducida capacidad para detectar las vibraciones que aparece en personas ancianas
(Shaffer y Harrison, 2007; Landelle y Cols., 2018).
Las células de Merkel sufren una reducción significativa asociada a la edad en los
individuos estudiados. Este hecho coincide con estudios realizados en ratones viejos,
en los que se observó una reducción de células de Merkel en la mucosa del paladar y
que precisamente, se correlaciona con pérdidas de las capacidades orales (Moayedi y
Cols., 2018).
La activación de los canales iónicos controlados mecánicamente supone el origen de la
detección de los estímulos mecánicos. Entre todos los candidatos propuestos como

posibles mecanotransductores principales que actúan en los mecanorreceptores
humanos (ASIC2, TRPV4, TRPC6; Calavia y Cols., 2010; Cabo y Cols., 2015; Alonso-
González y Cols., 2017), la mecanoproteína Piezo2 parece ser la única esencial para
que el proceso de mecanotransducción tenga lugar (Roudeaut y Cols., 2012; Delmas y
Coste, 2013; Wu y Cols., 2017). Piezo2 se expresa tanto en las neuronas
mecanosensitivas (Coste y Cols., 2010; Ranade y Cols., 2014) como en los
mecanorreceptores de bajo umbral (células de Merkel y corpúsculos de Meissner;
García-Mesa y Cols., 2017). Los resultados obtenidos demuestran que la expresión de
Piezo2 disminuye con el envejecimiento en ambos tipos de mecanorreceptores
cutáneos y que presumiblemente, dicho déficit esté detrás de la pérdida de
sensibilidad táctil asociada a la edad.
El desarrollo y mantenimiento de los corpúsculos sensitivos cutáneos está controlado
por el sistema de neurotrofinas TrkB-BDNF (González-Martínez y Cols., 2004; Sedý y
Cols., 2004; De Carlos y Cols., 2006; Pérez-Piñera y Cols., 2008; Montaño y Cols., 2010).
En adultos, TrkB y BDNF se expresan en algunas neuronas de ganglios sensitivos
(Montaño y Cols., 2010), así como en los corpúsculos sensitivos cutáneos (Calavia y
Cols., 2010). Los resultados de este trabajo sugieren que en los corpúsculos de
Meissner y de Pacini, tiene lugar la señalización retrograda de TrkB-BDNF para
proporcionar soporte trófico a las neuronas sensitivas de las cuales que derivan. De
esta forma, las células lamelares y el núcleo interno serían la fuente de BDNF, mientras
que el axón sería el receptor al expresar TrkB. El efecto de la edad en este sistema de
señalización TrkB-BDNF nunca ha sido estudiado. A partir de esta investigación se
puede notar un evidente paralelismo entre los cambios que aparecen en los
corpúsculos sensitivos asociados al envejecimiento y la decadencia del sistema TrkB-
BDNF; en algunos corpúsculos de Meissner, su inmunodetección fue completamente
negativa. Queda por determinar si el deterioro del sistema TrkB-BDNF es el
responsable principal de los cambios morfológicos observados en los corpúsculos de
Meissner y la perdida de densidad de los mismos y de las células de Merkel a edades
avanzadas. En el sistema nervioso central, la reducida expresión de TrkB y BDNF está
asociada al envejecimiento y enfermedades relacionadas (Erickson y Cols., 2010;
Forlenza y Cols., 2015; Nunes y Cols., 2018).

En resumen, todos estos resultados demuestran que los corpúsculos de Meissner y las
células de Merkel, y en menor medida los corpúsculos de Pacini, experimentan
cambios morfológicos edad-dependientes, así como una reducción en su densidad.

7. CONCLUSIONES

En base a los resultados obtenidos en este trabajo de tesis doctoral y tras la discusión
de los mismos, se han realizado las siguientes conclusiones:
1. Una lámina intermedia de origen endoneural, definida por células CD34-positivas,
se dispone entre el núcleo interno y el núcleo externo de los corpúsculos de Pacini
cutáneos humanos.
2. Los proteoglicanos de condroitín sulfato se localizan en la lámina intermedia de
origen endoneural CD34-positiva de los corpúsculos de Pacini cutáneos humanos,
pero no se expresan en los de Meissner.
3. El heparán sulfato está asociado a proteoglicanos de membrana basal en los
corpúsculos de Meissner y de Pacini humanos.
4. Los proteoglicanos de heparán sulfato y el colágeno IV de la membrana basal
denotan una fuerte expresión en las primeras lamelas del núcleo externo de los
corpúsculos de Pacini, hecho que vincula la membrana basal al aislamiento del
núcleo interno.
5. Los corpúsculos de Pacini de la piel digital humana expresan inmunorreacción para
los proteoglicanos pequeños ricos en leucina de las familias I y II, con un patrón de
distribución complejo: decorina, biglicano, lumicano, fibromodulina y
osteoadherina se detectan en el núcleo interno; la capa intermedia contiene
osteoadherina; el núcleo externo posee biglicano, decorina, lumicano,
fibromodulina y osteoadherina; y la cápsula contiene biglicano, decorina,
fibromodulina y lumicano. No se ha detectado la presencia de aspronina,
prolargina y keratocano.
6. La densidad de corpúsculos de Meissner y de células de Merkel en piel glabra
digital humana desciende drásticamente con el envejecimiento.
7. Los corpúsculos de Meissner cutáneos presentan una morfología alterada en
individuos de edad avanzada: se reducen en tamaño, se redondean en forma y se
localizan más profundos en la dermis. Los corpúsculos de Pacini no presentan
generalmente cambios morfológicos relevantes.
8. Los corpúsculos sensitivos de Meissner y de Pacini humanos experimentan
procesos de denervación y desorganización lamelar asociado a la edad. Además,

los complejos célula de Merkel-axón presentan menor número de axones en las
proximidades de las células de Merkel.
9. La expresión de la mecanoproteína Piezo2 disminuye en los corpúsculos de
Meissner y células de Merkel de la piel glabra digital de sujetos de edad avanzada.
10. El sistema de neurotrofinas TrkB-BDNF, del que depende el desarrollo y
mantenimiento de los corpúsculos sensitivos, decae con la edad mostrando
patrones de expresión reducidos.

8. BIBLIOGRAFÍA

- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Vega JA, García-Suárez O, Montaño JA, Pardo B, Cobo JM. The Meissner and
Pacinian sensory corpuscles revisited new data from the last decade. Microsc Res
Tech. 2009; 72:299-309.
- Vega JA, López-Muñiz A, Calavia MG, García-Suárez O, Cobo J, Otero J, Arias-
Carrión O, Prez-Piñera P, Menéndez-González M. Clinical implication of Meissner’s
corpuscles. CNS Neurol Disord Drug Targets. 2012; 11:856-868.
- Montaño JA, Pérez-Piñera P, García-Suárez O, Cobo J, Vega JA. Development and
neuronal dependence of cutaneous sensory nerve formations: Lessons from
neurotrophins. Microsc Res Tech. 2010; 73:513-29.
- Del Valle ME, Cobo T, Cobo JL, Vega JA. Mechanosensory neurons, cutaneous
mechanoreceptors, and putative mechanoproteins. Microsc Res Tech. 2012; 75:
1033-43.
- García-Añoveros J, Samad TA, Zuvela-Jelaska L, Woolf CJ, Corey DP. Transport and
localization of the DEG/ENaC ion channel BNaC1alpha to peripheral
mechanosensory terminals of dorsal root ganglia neurons. J Neurosci. 2001;
21:2678-2686.
- Cabo R, Gálvez MA, San José I, Laurà R, López-Muñiz A, García-Suárez O, Cobo T,
Insausti R, Vega JA. Immunohistochemical localization of acid-sensing ion channel 2
(ASIC2) in cutaneous Meissner and Pacinian corpuscles of Macaca fascicularis.
Neurosci Lett. 2012; 516:197-201.
- Cabo R, Alonso P, Viña E, Vázquez G, Gago A, Feito J, Pérez-Moltó FJ, García-Suárez
O, Vega JA. ASIC2 is present in human mechanosensory neurons of the dorsal root
ganglia and in mechanoreceptors of the glabrous skin. Histochem Cell Biol. 2015;
143:267-276.
- Alonso-González P, Cabo R, San José I, Gago A, Suazo IC, García-Suárez O, Cobo J,
Vega JA. Human Digital Meissner Corpuscles Display Immunoreactivity for the
Multifunctional Ion Channels Trpc6 and Trpv4. Anat Rec (Hoboken). 2017;
300:1022-1031.
- García-Mesa Y, García-Piqueras J, García B, Feito J, Cabo R, Cobo J, Vega JA, García-
Suárez O. Merkel cells and Meissner's corpuscles in human digital skin display
Piezo2 immunoreactivity. J Anat. 2017; 231:978-989.
- Chen S, Birk DE. The regulatory roles of small leucine-rich proteoglycans in
extracellular matrix assembly. FEBS J. 2013; 280:2120-3217.
- Herrmann DN, Boger JN, Jansen C, Alessi-Fox C. In vivo confocal microscopy of
Meissner corpuscles as a measure of sensory neuropathy. Neurology. 2007;
69:2121-2127.
- Nolano M, Provitera V, Estraneo A, Selim MM, Caporaso G, Stancanelli A,
Saltalamacchia AM, Lanzillo B, Santoro L. Sensory deficit in Parkinson's disease:
evidence of a cutaneous denervation. Brain. 2008; 131:1903-1111.
- Almodovar JL, Schifitto G, McDermott MP, Ferguson M, Herrmann DN. HIV
neuropathy: an in vivo confocal microscopic study. J Neurovirol. 2012; 18:503-510.

- Nolano M, Provitera V, Manganelli F, Iodice R, Stancanelli A, Caporaso G,
Saltalamacchia A, Califano F, Lanzillo B, Picillo M, Barone P, Santoro L. Loss of
cutaneous large and small fibers in naive and l-dopa-treated PD patients.
Neurology. 2017; 89:776-784.
- Malinovský L, Pác L, Vega-Alvarez JA, Bozilow W. The capsule structure of Pacinian
corpuscles from the cat mesentery. Z Mikrosk Anat Forsch. 1990; 104:193-201.
- Malinovský L. Sensory nerve formations in the skin and their classification. Microsc
Res Tech. 1996; 34:283-301.
- Lawson SN. 1992. Morphological and biochemical cell types of sensory neurons. In:
Scott SA, editor. Sensory Neurons: Diversity, development, and plasticity. New York
(USA): Oxford University Press. p. 27-52.
- Dalsgaard C-J. 1988. The sensory system. In: Bjorklund A, Hokfelt T, Owmann C
(eds.). Handbook of Chemical Neuroanatomy, vol 6. The Peripheral Nervous
System. Elsevier, Amsterdam, pp 599-636.
- Perl ER. 1992. Function dorsal root ganglion neurons: An overview. In: Scott SA,
editor. Sensory neurons: Diversity, development and plasticity. New York: Oxford
University Press. pp. 3–23.
- Vega JA, Del Valle ME, Calzada B, Bengoechea ME, Perez-Casas A. Expression of
nerve growth factor receptor immunoreactivity in the rat choroid plexus. Cell Mol
Biol. 1992 Apr;38(2):145-9.
- Vega JA, Del Valle ME, Haro JJ, Calzada B, Suarez-Garnacho S, Malinovsky L. Nerve
growth factor receptor immunoreactivity in Meissner and Pacinian of the human
digital skin. Anat Rec. 1993; 236:730-6.
- Renehan WE, Munger BL. The development of Meissner corpuscles in primate
digital skin. Brain Res Dev Brain Res. 1990; 51: 35-44.
- Saxod R. Ontogeny of the cutaneous sensory organs. Microsc Res Tech. 1996;
34:313-333.
- Ide C, Hayashi S. Specializations of plasma membranes in Pacinian corpuscles:
implications for mechano-electric transduction. J Neurocytol. 1987; 16:759-773.
- Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch
Histol Cytol. 1988; 51:1–34.
- Halata Z, Grim M, Christ B. Origin of spinal cord meninges, sheaths of peripheral
nerves, and cutaneous receptors including Merkel cells. An experimental and
ultrastructural study with avian chimeras. Anat Embryol (Berl). 1990; 182:529-537.
- Vega JA, Del Valle ME, Haro JJ, Naves FJ, Calzada B, Uribelarea R. The inner-core,
outer-core and capsule cells of the human Pacinian corpuscles: an
immunohistochemical study. Eur J Morphol. 1994; 32:11-18.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Perl ER. 1992. Function dorsal root ganglion neurons: An overview. In: Scott SA,
editor. Sensory neurons: Diversity, development and plasticity. New York: Oxford
University Press. pp. 3–23.
- Zelená J. Nerves and mechanoreceptors. Chapman & Hall, London. 1994.

- Johnson KO. The roles and functions of cutaneous mechanoreceptors. Curr Opin
Neurobiol. 2001; 11:455-461.
- Johnson KO. The roles and functions of cutaneous mechanoreceptors. Curr Opin
Neurobiol. 2001; 11:455-461.
- Zimmerman A, Bai L, Ginty DD. The gentle touch receptors of mammalian skin.
Science. 2014; 346:950-954.
- Guinard D, Usson Y, Guillermet C, Saxod R. PS100 and NF 70-200 double
immunolabeling for human digital skin meissner corpuscle 3D imaging. J Histochem
Cytochem. 2000; 48:295-302.
- Vega JA, López-Muñiz A, Calavia MG, García-Suárez O, Cobo J, Otero J, Arias-
Carrión O, Pérez-Piñera P, Menéndez-González M. Cinical implication of Meissner’s
corpuscles. CNS Disord Drug Targets. 2012; 11:856-68.
- Paré M, Elde R, Mazurkiewicz JE, Smith AM, Rice FL. The Meissner corpuscle
revised: a multiafferented mechanoreceptor with nociceptor immunochemical
properties. J Neurosci. 2001; 21:7236-46.
- García-Suárez O, Montaño JA, Esteban I, González-Martínez T, Alvarez-Abad C,
López-Arranz E, Cobo J, Vega JA. Myelin basic protein-positive nerve fibres in
human Meissner corpuscles. J Anat. 2009; 214:888-893.
- Ide C. Histochemical study of lamellar cell development of Meissner
corpuscles.Arch Histol Jpn. 1982; 45: 83-97.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Vega JA, Esteban I, Naves FJ, del Valle ME, Malinovsky L. Immunohistochemical
localization of laminin and type IV collagen in human cutaneous sensory nerve
formations. Anat Embryol (Berl). 1995; 191: 33-39.
- Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch
Histol Cytol. 1988; 51:1–34.
- Takahashi-Iwanaga H, Shimoda H. The three-dimensional microanatomy of
Meissner corpuscles in monkey palmar skin. J Neurocytol. 2003; 32:363-371.
- Zelená J. Nerves and mechanoreceptors. Chapman & Hall, London. 1994.
- Johnson KO. The roles and functions of cutaneous mechanoreceptors. Curr Opin
Neurobiol. 2001; 11:455-461.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Pawson L, Slepecky NB, Bolanowski SJ. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens Mot Res. 2000; 17:159-170.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Albuerne M, López S, Naves FJ, Martínez-Almagro A, Represa J, Vega JA. S100alpha
and S100beta proteins in human cutaneous sensory corpuscles: effects of nerve
and spinal cord injury. Anat Rec. 1998; 25:351-359.

- Galeano RM, Germanà A, Vázquez MT, Hidaka H, Germanà G, Vega JA.
Immunohistochemical localization of neurocalcin in human sensory neurons and
mechanoreceptors. Neurosci Lett. 2000; 279:89-92.
- Komori N, Neal J, Cain SD, Logan J, Wirsig C, Miller KE. Presence of beta-arrestin-1
immunoreactivity in the cutaneous nerve fibers of rat glabrous skin. Brain Res.
2003; 988:121-9.
- Verzé L, Viglietti-Panzica C, Maurizo S, Sica M, Panzica G. Distribution of GAP-43
nerve fibers in the skin of the adult human hand. Anat Rec A Discov Mol Cell Evol
Biol. 2003; 272:467-73
- García-Añoveros J, Samad TA, Zuvela-Jelaska L, Woolf CJ, Corey DP. Transport and
localization of the DEG/ENaC ion channel BNaC1alpha to peripheral
mechanosensory terminals of dorsal root ganglia neurons. J Neurosci. 2001;
21:2678-2686.
- Cabo R, Gálvez MA, San José I, Laurà R, López-Muñiz A, García-Suárez O, Cobo T,
Insausti R, Vega JA. Immunohistochemical localization of acid-sensing ion channel 2
(ASIC2) in cutaneous Meissner and Pacinian corpuscles of Macaca fascicularis.
Neurosci Lett. 2012; 516:197-201.
- Cabo R, Alonso P, Viña E, Vázquez G, Gago A, Feito J, Pérez-Moltó FJ, García-Suárez
O, Vega JA. ASIC2 is present in human mechanosensory neurons of the dorsal root
ganglia and in mechanoreceptors of the glabrous skin. Histochem Cell Biol. 2015;
143:267-276.
- Alonso-González P, Cabo R, San José I, Gago A, Suazo IC, García-Suárez O, Cobo J,
Vega JA. Human Digital Meissner Corpuscles Display Immunoreactivity for the
Multifunctional Ion Channels Trpc6 and Trpv4. Anat Rec (Hoboken). 2017;
300:1022-1031.
- García-Mesa Y, García-Piqueras J, García B, Feito J, Cabo R, Cobo J, Vega JA, García-
Suárez O. Merkel cells and Meissner's corpuscles in human digital skin display
Piezo2 immunoreactivity. J Anat. 2017; 231:978-989.
- Johansson O, Fantini F, Hu H. Neuronal structural proteins, transmitters,
transmitter enzymes and neuropeptides in human Meissner's corpuscles: a
reappraisal using immunohistochemistry. Arch Dermatol Res. 1999; 291:419-424.
- Paré M, Elde R, Mazurkiewicz JE, Smith AM, Rice FL. The Meissner corpuscle
revised: a multiafferented mechanoreceptor with nociceptor immunochemical
properties. J Neurosci. 2001; 21:7236-72346.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Calavia M, Viña E, Menéndez-González M, López-Muñiz A, Alonso-Guervós M, Cobo
J, Otero J, Arias-Carrión O, Vega JA, García-Suárez O. Evidence of nestin-positive
cells in the human cutaneus Meissner and Pacinian corpuscles. CNS Neurol Disord
Drug Targets. 2012; 11: 869-77.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.

- Vega JA, Del Valle ME, Haro JJ, Calzada B, Suarez-Garnacho S, Malinovsky L. Nerve
growth factor receptor immunoreactivity in Meissner and Pacinian of the human
digital skin. Anat Rec. 1993; 236:730-736.
- Vega JA, Del Valle ME, Haro JJ, Naves FJ, Calzada B, Uribelarea R. The inner-core,
outer-core and capsule cells of the human Pacinian corpuscles: an
immunohistochemical study. Eur J Morphol. 1994; 32:11-18.
- Bronzetti E, Ciriaco E, Germanà G, Vega JA. Immunohistochemical localization of
neurotrophin receptor proteins in human skin. Ital J Anat Embryol. 1995;100 Suppl
1:565-571.
- Calavia MG, Feito J, López-Iglesias L, de Carlos F, García-Suarez O, Pérez-Piñera P,
Cobo J, Vega JA. The lamellar cells in human Meissner corpuscles express TrkB.
Neurosci Lett. 2010; 468:106-109.
- Vega JA, Del Valle ME, Haro JJ, Naves FJ, Calzada B, Uribelarea R. The inner-core,
outer-core and capsule cells of the human Pacinian corpuscles: an
immunohistochemical study. Eur J Morphol. 1994; 32:11-18.
- Pawson L, Slepecky NB, Bolanowski SJ. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens Mot Res. 2000; 17:159-170.
- Vega JA, López-Muñiz A, Calavia MG, García-Suárez O, Cobo J, Otero J, Arias-
Carrión O, Pérez-Piñera P, Menéndez-González M. Cinical implication of Meissner’s
corpuscles. CNS Disord Drug Targets. 2012; 11:856-68.
- Cabo R, Alonso P, Viña E, Vázquez G, Gago A, Feito J, Pérez-Moltó FJ, García-Suárez
O, Vega JA. ASIC2 is present in human mechanosensory neurons of the dorsal root
ganglia and in mechanoreceptors of the glabrous skin. Histochem Cell Biol. 2015;
143:267-76.
- Cabo R, Gálvez MA, San José I, Laurà R, López-Muñiz A, García-Suárez O, Cobo T,
Insausti R, Vega JA. Immunohistochemical localization of acid-sensing ion channel 2
(ASIC2) in cutaneous Meissner and Pacinian corpuscles of Macaca fascicularis.
Neurosci Lett. 2012; 516:197-201.
- Vega JA, Del Valle ME, Haro JJ, Naves FJ, Calzada B, Uribelarea R. The inner-core,
outer-core and capsule cells of the human Pacinian corpuscles: an
immunohistochemical study. Eur J Morphol. 1994; 32:11-18.
- Vega JA, Vazquez E, Naves FJ, Calzada B, del Valle ME, Represa JJ.
- Expression of epidermal growth factor receptor (EGFr) immunoreactivity in human
cutaneous nerves and sensory corpuscles. Anat Rec. 1994; 240:125-130.
- Cabo R, Alonso P, Viña E, Vázquez G, Gago A, Feito J, Pérez-Moltó FJ, García-Suárez
O, Vega JA. ASIC2 is present in human mechanosensory neurons of the dorsal root
ganglia and in mechanoreceptors of the glabrous skin. Histochem Cell Biol. 2015;
143:267-76.
- Cabo R, Alonso P, Viña E, Vázquez G, Gago A, Feito J, Pérez-Moltó FJ, García-Suárez
O, Vega JA. ASIC2 is present in human mechanosensory neurons of the dorsal root
ganglia and in mechanoreceptors of the glabrous skin. Histochem Cell Biol. 2015;
143:267-76.
- Zelená J. Nerves and mechanoreceptors. Chapman & Hall, London. 1994.

- García-Suárez O, Calavia MG, Pérez-Moltó FJ, Alvarez-Abad C, Pérez-Piñera P, Cobo
JM, Vega JA. Immunohistochemical profile of human pancreatic pacinian
corpuscles. Pancreas. 2010; 39:403-410.
- Bell J, Bolanowski S, Holmes MH. The structure and function of Pacinian corpuscles:
a review. Prog Neurobiol. 1994; 42:79-128.
- Malinovský L, Pác L. Morphology of sensory corpuscles in mammals. Acta facultatis
Medicae Universitatis Brunensis, Brno. 1982.
- Ide C, Yoshida Y, Hayashi S, Takashio M, Munger BL. A re-evaluation of the cytology
of cat Pacinian corpuscles II. The extreme tip of the axon. Cell Tissue Res. 1988;
253:95-103.
- Malinovský L, Pác L, Vega-Alvarez JA, Bozilow W. The capsule structure of Pacinian
corpuscles from the cat mesentery. Z Mikrosk Anat Forsch. 1990; 104:193-201.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.
- Pawson L, Slepecky NB, Bolanowski SJ. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens Mot Res. 2000; 17:159-170.
- Sames K, Halata Z, Jojovic M, van Damme EJ, Peumans WJ, Delpech B, Asmus B,
Schumacher U. Lectin and proteoglycan histochemistry of feline pacinian
corpuscles. J Histochem Cytochem. 2001; 49:19-28.
- Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch
Histol Cytol. 1988; 51:1-34.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.
- Pawson L, Slepecky NB, Bolanowski SJ. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens Mot Res. 2000; 17:159-170.
- Sames K, Halata Z, Jojovic M, van Damme EJ, Peumans WJ, Delpech B, Asmus B,
Schumacher U. Lectin and proteoglycan histochemistry of feline pacinian
corpuscles. J Histochem Cytochem. 2001; 49:19-28.
- Ide C, Yoshida Y, Hayashi S, Takashio M, Munger BL. A re-evaluation of the cytology
of cat Pacinian corpuscles II. The extreme tip of the axon. Cell Tissue Res. 1988;
253:95-103.
- Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch
Histol Cytol. 1988; 51:1-34.
- Pawson L, Slepecky NB, Bolanowski SJ. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens Mot Res. 2000; 17:159-170.
- García-Piqueras J, García-Suárez O, Rodríguez-González MC, Cobo JL, Cabo R, Vega
JA, Feito J. Endoneurial-CD34 positive cells define an intermediate layer in human
digital Pacinian corpuscles. Ann Anat. 2017; 211:55-60.
- Zelená J. Nerves and mechanoreceptors. Chapman & Hall, London. 1994.
- Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch
Histol Cytol. 1988; 51:1-34.
- Malinovský L, Pác L, Vega-Alvarez JA, Bozilow W. The capsule structure of Pacinian
corpuscles from the cat mesentery. Z Mikrosk Anat Forsch. 1990; 104:193-201.

- Malinovský L. Classification of sensory nerve endings in vertebrates brought up to
date. Folia Morphol (Praha). 1986, 34:261-4.
- Munger BL, Ide C. The structure and function of cutaneous sensory receptors. Arch
Histol Cytol. 1988; 51:1-34.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- Vega JA, Llamosas MM, Huerta JJ, García-Fernández JM. Study of human cutaneous
sensory corpuscles using double immunolabelling and confocal laser scanning
microscopy. Anat Rec. 1996; 246:557-760.
- Calavia MG, Feito J, López-Iglesias L, de Carlos F, García-Suarez O, Pérez-Piñera P,
Cobo J, Vega JA. The lamellar cells in human Meissner corpuscles express TrkB.
Neurosci Lett. 2010; 468:106-109.
- González-Martínez T, Monjil DF, Aguado-Barrios A, Cobo J, Germanà G, Vega JA.
Bcl-2 immunoreactivity in human cutaneous Meissner and Pacinian corpuscles.
Neurosci Lett. 2006; 394:13-6.
- Vega JA, Haro JJ, Del Valle ME. Immunohistochemistry of human cutaneous
Meissner and pacinian corpuscles. Microsc Res Tech. 1996; 34:351-361.
- García-Piqueras J, García-Suárez O, Rodríguez-González MC, Cobo JL, Cabo R, Vega
JA, Feito J. Endoneurial-CD34 positive cells define an intermediate layer in human
digital Pacinian corpuscles. Ann Anat. 2017; 211:55-60.
- Calavia MG, Feito J, López-Iglesias L, de Carlos F, García-Suarez O, Pérez-Piñera P,
Cobo J, Vega JA. The lamellar cells in human Meissner corpuscles express TrkB.
Neurosci Lett. 2010; 468:106-109.
- Cabo R, Alonso P, Viña E, Vázquez G, Gago A, Feito J, Pérez-Moltó FJ, García-Suárez
O, Vega JA. ASIC2 is present in human mechanosensory neurons of the dorsal root
ganglia and in mechanoreceptors of the glabrous skin. Histochem Cell Biol. 2015;
143:267-276.
- García-Suárez O, Montaño JA, Esteban I, González-Martínez T, Alvarez-Abad C,
López-Arranz E, Cobo J, Vega JA. Myelin basic protein-positive nerve fibres in
human Meissner corpuscles. J Anat. 2009; 214:888-893.
- Calavia M, Viña E, Menéndez-González M, López-Muñiz A, Alonso-Guervós M, Cobo
J, Otero J, Arias-Carrión O, Vega JA, García-Suárez O. Evidence of nestin-positive
cells in the human cutaneus Meissner and Pacinian corpuscles. CNS Neurol Disord
Drug Targets. 2012; 11: 869-77.
- Vega JA, Esteban I, Naves FJ, del Valle ME, Malinovsky L. Immunohistochemical
localization of laminin and type IV collagen in human cutaneous sensory nerve
formations. Anat Embryol (Berl). 1995; 191: 33-39.
- González-Martínez T, Germana A, Catania S, Cobo T, Ochoa-Erena FJ, de Carlos F,
Díaz-Esnal B, Vega JA. Postnatal developmental changes in the expression of ErbB
receptors in murine Pacinian corpuscles. Neurosci Lett. 2007; 420:90-5.
- Fleming MS, Luo W. The anatomy, function, and development of mammalian Aβ
low-threshold mechanoreceptors. Front Biol (Beijing). 2013; 8.
- Zimmerman A, Bai L, Ginty DD. The gentle touch receptors of mammalian skin.
Science. 2014; 346:950-954.

- Moll I, Moll R, Franke WW. Formation of epidermal and dermal Merkel cells during
human fetal skin development. J Invest Dermatol. 1986; 87:779-787.
- Rettig T, Halata Z. Structure of the sensory innervation of the anal canal in the pig.
A light- and electron-microscopical study. Acta Anat (Basel).1990; 137:189-201.
- Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel and his "Merkel cell",
morphology, development, and physiology: review and new results. Anat Rec A
Discov Mol Cell Evol Biol. 2003; 271:225-239.
- Moll I, Paus R, Moll R. Merkel cells in mouse skin: intermediate filament pattern,
localization, and hair cycle-dependent density. J Invest Dermatol. 1996; 106:281-
286.
- Iggo A, Muir AR. The structure and function of a slowly adapting touch corpuscle in
hairy skin. J Physiol. 1969; 200:763-796.
- Iggo A, Muir AR. The structure and function of a slowly adapting touch corpuscle in
hairy skin. J Physiol. 1969; 200:763-796.
- Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel and his "Merkel cell",
morphology, development, and physiology: review and new results. Anat Rec A
Discov Mol Cell Evol Biol. 2003; 271:225-239.
- Chen SY, Gerson S, Meyer J. The fusion of Merkel cell granules with a synapse-like
structure. J Invest Dermatol. 1973; 61:290-292.
- Maksimovic S, Baba Y, Lumpkin EA. Neurotransmitters and synaptic components in
the Merkel cell-neurite complex, a gentle-touch receptor. Ann N Y Acad Sci. 2013;
1279:13-21.
- Zimmerman A, Bai L, Ginty DD. The gentle touch receptors of mammalian skin.
Science. 2014; 346:950-954.
- Grim M, Halata Z. Developmental origin of avian Merkel cells. Anat Embryol (Berl).
2000; 202:401-410.
- Szeder V, Grim M, Halata Z, Sieber-Blum M. Neural crest origin of mammalian
Merkel cells. Dev Biol. 2003; 253:258-263.
- Munger BL. The intraepidermal innervation of the snout skin of the opossum. A
light and electron microscope study, with observations on the nature of Merkel's
Tastzellen. J Cell Biol. 1965; 26:79-97.
- Compton CC, Regauer S, Seiler GR, Landry DB. Human Merkel cell regeneration in
skin derived from cultured keratinocyte grafts. Lab Invest. 1990; 63:233-241.
- Moll I, Lane AT, Franke WW, Moll R. Intraepidermal formation of Merkel cells in
xenografts of human fetal skin. J Invest Dermatol. 1990; 94:359-364.
- Kim DK, Holbrook KA. The appearance, density, and distribution of Merkel cells in
human embryonic and fetal skin: their relation to sweat gland and hair follicle
development. J Invest Dermatol. 1995; 104:411-416.
- Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Mammalian Merkel cells
are descended from the epidermal lineage. Dev Biol. 2009; 336:76-83.
- Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N, Szpalski C,
Achouri Y, Bloch W, Hassan BA, Blanpain C. Epidermal progenitors give rise to
Merkel cells during embryonic development and adult homeostasis. J Cell Biol.
2009; 187:91-100.

- Woo SH, Stumpfova M, Jensen UB, Lumpkin EA, Owens DM. Identification of
epidermal progenitors for the Merkel cell lineage. Development. 2010; 137:3965-
3971.
- Kinkelin I, Stucky CL, Koltzenburg M. Postnatal loss of Merkel cells, but not of
slowly adapting mechanoreceptors in mice lacking the neurotrophin receptor p75.
Eur J Neurosci. 1999; 11:3963-3969.
- Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA, Zoghbi
HY. Merkel cells are essential for light-touch responses. Science. 2009. 19;
324:1580-1582.
- Yamashita Y, Ogawa H. Slowly adapting cutaneous mechanoreceptor afferent units
associated with Merkel cells in frogs and effects of direct currents. Somatosens
Mot Res. 1991; 8:87-95.
- Fleming MS, Luo W. The anatomy, function, and development of mammalian Aβ
low-threshold mechanoreceptors. Front Biol (Beijing). 2013; 8.
- Maksimovic S, Nakatani M, Baba Y, Nelson AM, Marshall KL, Wellnitz SA, Firozi P,
Woo SH, Ranade S, Patapoutian A, Lumpkin EA. Epidermal Merkel cells are
mechanosensory cells that tune mammalian touch receptors. Nature. 2014;
509:617-621.
- Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA, Zoghbi
HY. Merkel cells are essential for light-touch responses. Science. 2009. 19;
324:1580-1582.
- Hartschuh W, Weihe E. Fine structural analysis of the synaptic junction of Merkel
cell-axon-complexes. J Invest Dermatol. 1980; 75:159-165.
- Tachibana T, Nawa T. Recent progress in studies on Merkel cell biology. Anat Sci
Int. 2002; 77:26-33.
- Haeberle H, Fujiwara M, Chuang J, Medina MM, Panditrao MV, Bechstedt S,
Howard J, Lumpkin EA. Molecular profiling reveals synaptic release machinery in
Merkel cells. Proc Natl Acad Sci U S A. 2004; 101:14503-14508.
- Hitchcock IS, Genever PG, Cahusac PM. Essential components for a glutamatergic
synapse between Merkel cell and nerve terminal in rats. Neurosci Lett. 2004;
362:196-199.
- Ranade SS, Woo SH, Dubin AE, Moshourab RA, Wetzel C, Petrus M, Mathur J, Bégay
V, Coste B, Mainquist J, Wilson AJ, Francisco AG, Reddy K, Qiu Z, Wood JN, Lewin
GR, Patapoutian A. Piezo2 is the major transducer of mechanical forces for touch
sensation in mice. Nature. 2014; 516:121-125.
- Xiao Y, Williams JS, Brownell I. Merkel cells and touch domes: more than
mechanosensory functions? Exp Dermatol. 2014; 23:692-695.
- García-Mesa Y, García-Piqueras J, García B, Feito J, Cabo R, Cobo J, Vega JA, García-
Suárez O. Merkel cells and Meissner's corpuscles in human digital skin display
Piezo2 immunoreactivity. J Anat. 2017; 231:978-989.
- Maksimovic S, Baba Y, Lumpkin EA. Neurotransmitters and synaptic components in
the Merkel cell-neurite complex, a gentle-touch receptor. Ann N Y Acad Sci. 2013;
1279:13-21.

- Moll I, Kuhn C, Moll R. Cytokeratin 20 is a general marker of cutaneous Merkel cells
while certain neuronal proteins are absent. J Invest Dermatol. 1995; 104:910-915.
- Leff EL, Brooks JS, Trojanowski JQ. Expression of neurofilament and neuron-specific
enolase in small cell tumors of skin using immunohistochemistry. Cancer. 1985;
56:625-631.
- Dalsgaard CJ, Rydh M, Haegerstrand A. Cutaneous innervation in man visualized
with protein gene product 9.5 (PGP 9.5) antibodies. Histochemistry. 1989; 92:385-
390.
- Moll I, Kuhn C, Moll R. Cytokeratin 20 is a general marker of cutaneous Merkel cells
while certain neuronal proteins are absent. J Invest Dermatol. 1995; 104:910-915.
- Ortonne JP, Petchot-Bacque JP, Verrando P, Pisani A, Pautrat G, Bernerd F. Normal
Merkel cells express a synaptophysin-like immunoreactivity. Dermatologica. 1988;
177:1-10.
- Kurzen H, Kaul S, Egner U, Deichmann M, Hartschuh W (2003). Expression of MUC-
1 and EpCAM in Merkel cell carcinomas: implications for immunotherapy. Arch
Dermatol Res 295:146-154.
- Woo SH, Stumpfova M, Jensen UB, Lumpkin EA, Owens DM. Identification of
epidermal progenitors for the Merkel cell lineage. Development. 2010; 137:3965-
3971.
- Hitchcock IS, Genever PG, Cahusac PM. Essential components for a glutamatergic
synapse between Merkel cell and nerve terminal in rats. Neurosci Lett. 2004;
362:196-199.
- Cahusac PM, Senok SS, Hitchcock IS, Genever PG, Baumann KI. Are unconventional
NMDA receptors involved in slowly adapting type I mechanoreceptor responses?
Neuroscience. 2005; 133:763-773.
- Gould VE, Moll R, Moll I, Lee I, Franke WW. Neuroendocrine (Merkel) cells of the
skin: hyperplasias, dysplasias and neoplasms. Lab Invest. 1985; 52:334-353.
- Hartschuh W, Weihe E. Pancreastatin-like immunoreactivity in epidermal Merkel
cells of pig and man. Neurosci Lett. 1989; 98:258-263.
- Maksimovic S, Baba Y, Lumpkin EA. Neurotransmitters and synaptic components in
the Merkel cell-neurite complex, a gentle-touch receptor. Ann N Y Acad Sci. 2013;
1279:13-21.
- García-Mesa Y, García-Piqueras J, García B, Feito J, Cabo R, Cobo J, Vega JA, García-
Suárez O. Merkel cells and Meissner's corpuscles in human digital skin display
Piezo2 immunoreactivity. J Anat. 2017; 231:978-989.
- Dalsgaard CJ, Rydh M, Haegerstrand A. Cutaneous innervation in man visualized
with protein gene product 9.5 (PGP 9.5) antibodies. Histochemistry. 1989; 92:385-
390.
- Gould VE, Moll R, Moll I, Lee I, Franke WW (1985). Neuroendocrine (Merkel) cells
of the skin: hyperplasias, dysplasias and neoplasms. Lab Invest 52:334-353.
- Narisawa Y, Hashimoto K, Kohda H (1994c). Immunohistochemical demonstration
of the expression of neurofilament proteins in Merkel cells. Acta Dermatol
Venereol (Stockh.) 74:441-443.

- Cabo R, Alonso P, Viña E, Vázquez G, Gago A, Feito J, Pérez-Moltó FJ, García-Suárez
O, Vega JA. ASIC2 is present in human mechanosensory neurons of the dorsal root
ganglia and in mechanoreceptors of the glabrous skin. Histochem Cell Biol. 2015;
143:267-276.
- Muir D, Engvall E, Varon S, Manthorpe M. Schwannoma cell-derived inhibitor of the
neurite-promoting activity of laminin. Cell Biol. 1989; 109:2353-2362.
- Zuo J, Hernandez YJ, Muir D. 1998. Chondroitin sulfate proteoglycan with neurite-
inhibiting activity is up-regulated following peripheral nerve injury. J Neurobiol.
1998; 34:41-54.
- Udina E, Ladak A, Furey M, Brushart T, Tyreman N, Gordon T. Rolipram-induced
elevation of cAMP or chondroitinase ABC breakdown of inhibitory proteoglycans in
the extracellular matrix promotes peripheral nerve regeneration. Exp Neurol. 2010;
223:143-152.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the
Cell. edn 4.. Garland; London: 2002.
- Theocharis AD, Skandalis SS, Gialeli C, Karamanos NK. Extracellular matrix
structure. Adv Drug Deliv Rev. 2016; 97:4-27.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the
Cell. edn 4.. Garland; London: 2002.
- Bosman FT, Stamenkovic I. Functional structure and composition of the
extracellular matrix. J Pathol. 2003; 200:423-8.
- Theocharis AD, Skandalis SS, Gialeli C, Karamanos NK. Extracellular matrix
structure. Adv Drug Deliv Rev. 2016; 97:4-27.
- Frantz C, Stewart KM, Weaver VM. The extracellular matrix at a glance. J Cell Sci.
2010; 123:4195-4200.
- Theocharis AD, Skandalis SS, Gialeli C, Karamanos NK. Extracellular matrix
structure. Adv Drug Deliv Rev. 2016; 97:4-27.
- LeBleu VS, Macdonald B, Kalluri R. Structure and function of basement membranes.
Exp Biol Med (Maywood). 2007; 232:1121-1129.
- Sasaki T, Fässler R, Hohenester E. Laminin: the crux of basement membrane
assembly. J Cell Biol. 2004; 164:959-963.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the
Cell. edn 4.. Garland; London: 2002.
- Bosman FT, Stamenkovic I. Functional structure and composition of the
extracellular matrix. J Pathol. 2003; 200:423-8.
- Couchman JR, Pataki CA. An introduction to proteoglycans and their localization. J
Histochem Cytochem. 2012; 60:885-97.
- Page-McCaw A, Ewald AJ, Werb Z. Matrix metalloproteinases and the regulation of
tissue remodelling. Nat Rev Mol Cell Biol. 2007; 8:221-233.
- Pengfei Lu, Ken Takai, Valerie M. Weaver, Zena Werb. Extracellular Matrix
Degradation and Remodeling in Development and Disease. Cold Spring Harb
Perspect Biol. 2011; 3: a005058.
- Bonnans C, Chou J, Werb Z. Remodelling the extracellular matrix in development
and disease. Nat Rev Mol Cell Biol. 2014; 15:786-801.

- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular Biology of the
Cell. edn 4.. Garland; London: 2002.
- Theocharis AD, Skandalis SS, Gialeli C, Karamanos NK. Extracellular matrix
structure. Adv Drug Deliv Rev. 2016; 97:4-27.
- Frantz C, Stewart KM, Weaver VM. The extracellular matrix at a glance. J Cell Sci.
2010; 123:4195-4200.
- Frantz C, Stewart KM, Weaver VM. The extracellular matrix at a glance. J Cell Sci.
2010; 123:4195-4200.
- Clause KC, Barker TH. Extracellular matrix signaling in morphogenesis and repair.
Curr Opin Biotechnol. 2013; 24:830-833.
- Järveläinen H, Sainio A, Koulu M, Wight TN, Penttinen R. Extracellular matrix
molecules: potential targets in pharmacotherapy. Pharmacol Rev. 2009; 61:198-
223.
- Schaefer L, Schaefer RM. Proteoglycans: from structural compounds to signaling
molecules. Cell Tissue Res. 2010; 339:237-246.
- Schaefer L, Schaefer RM. Proteoglycans: from structural compounds to signaling
molecules. Cell Tissue Res. 2010; 339:237-246.
- Gandhi NS, Mancera RL. The structure of glycosaminoglycans and their interactions
with proteins. Chem Biol Drug Des. 2008; 72:455-482.
- Couchman JR, Pataki CA. An introduction to proteoglycans and their localization. J
Histochem Cytochem. 2012; 60:885-97.
- Iozzo RV, Schaefer L. Proteoglycan form and function: A comprehensive
nomenclature of proteoglycans. Matrix Biol. 2015; 42:11-55.
- Carey DJ. Syndecans: multifunctional cell-surface co-receptors. Biochem J. 1997;
327:1-16.
- Filmus J, Capurro M, Rast J. Glypicans. Genome Biol. 2008; 9:224.
- Lambaerts K, Wilcox-Adelman SA, Zimmermann P. The signaling mechanisms of
syndecan heparan sulfate proteoglycans. Curr Opin Cell Biol. 2009; 21:662-669.
- Filmus J, Capurro M. The role of glypicans in Hedgehog signaling. Matrix Biol. 2014;
35:248-252.
- Yurchenco PD, Patton BL. Developmental and pathogenic mechanisms of basement
membrane assembly. Curr Pharm Des. 2009; 15:1277-94.
- Bosman FT, Stamenkovic I. Functional structure and composition of the
extracellular matrix. J Pathol. 2003; 200:423-428.
- LeBleu VS, Macdonald B, Kalluri R. Structure and function of basement membranes.
Exp Biol Med (Maywood). 2007; 232:1121-1129.
- Nishimura M, Yan W, Mukudai Y, Nakamura S, Nakamasu K, Kawata M, Kawamoto
T, Noshiro M, Hamada T, Kato Y. Role of chondroitin sulfate-hyaluronan
interactions in the viscoelastic properties of extracellular matrices and fluids.
Biochim Biophys Acta. 1998; 1380:1-9.
- Heinegård D. Fell-Muir Lecture: Proteoglycans and more--from molecules to
biology. Int J Exp Pathol. 2009; 90:575-586.
- Iozzo RV, Schaefer L. Proteoglycan form and function: A comprehensive
nomenclature of proteoglycans. Matrix Biol. 2015; 42:11-55.

- Mikami T, Kitagawa H. Biosynthesis and function of chondroitin sulfate. Biochim
Biophys Acta. 2013; 1830:4719-4133.
- Crespo-Santiago D. [The extracellular matrix of the central nervous system:
chondroitin sulphate type proteoglycans and neural repair]. Rev Neurol. 2004;
38:843-51.
- Howell MD, Gottschall PE. Lectican proteoglycans, their cleaving
metalloproteinases, and plasticity in the central nervous system extracellular
microenvironment. Neuroscience. 2012; 217:6-18.
- López-Casillas F, Cheifetz S, Doody J, Andres JL, Lane WS, Massagué J. Structure and
expression of the membrane proteoglycan betaglycan, a component of the TGF-
beta receptor system. Cell. 1991; 67:785-795.
- Nishiyama A, Dahlin KJ, Prince JT, Johnstone SR, Stallcup WB. The primary structure
of NG2, a novel membrane-spanning proteoglycan. J Cell Biol. 1991; 114:359-371.
- Maurel P, Rauch U, Flad M, Margolis RK, Margolis RU.
- Phosphacan, a chondroitin sulfate proteoglycan of brain that interacts with
neurons and neural cell-adhesion molecules, is an extracellular variant of a
receptor-type protein tyrosine phosphatase. Proc Natl Acad Sci U S A. 1994;
91:2512-2516.
- Kalamajski S, Oldberg A. The role of small leucine-rich proteoglycans in collagen
fibrillogenesis. Matrix Biol. 2010; 29:248-53.
- Chen S, Birk DE. The regulatory roles of small leucine-rich proteoglycans in
extracellular matrix assembly. FEBS J. 2013; 280:2120-2137.
- Schaefer L, Iozzo RV. Biological functions of the small leucine-rich proteoglycans:
from genetics to signal transduction. J Biol Chem. 2008; 283:21305-21309.
- Hildebrand A, Romarís M, Rasmussen LM, Heinegård D, Twardzik DR, Border WA,
Ruoslahti E. Interaction of the small interstitial proteoglycans biglycan, decorin and
fibromodulin with transforming growth factor beta. Biochem J. 1994; 302:527-534.
- Iozzo RV, Schaefer L. Proteoglycan form and function: A comprehensive
nomenclature of proteoglycans. Matrix Biol. 2015; 42:11-55.
- Feltri ML, Wrabetz L. Laminins and their receptors in Schwann cells and hereditary
neuropathies. J Peripher Nerv Syst. 2005; 10:128-143.
- Mehta H, Orphe C, Todd MS, Cornbrooks CJ, Carey DJ. Synthesis by Schwann cells
of basal lamina and membrane-associated heparan sulfate proteoglycans. J Cell
Biol. 1985; 101:660-6.
- Halfter W, Dong S, Schurer B, Cole GJ. Collagen XVIII is a basement membrane
heparan sulfate proteoglycan. J Biol Chem. 1998; 273:25404-12.
- Ma E, Morgan R, Godfrey EW. Distribution of agrin mRNAs in the chick embryo
nervous system. J Neurosci. 1994 May;14(5 Pt 2):2943-52.
- Halfter W, Schurer B, Yip J, Yip L, Tsen G, Lee JA, Cole GJ. Distribution and substrate
properties of agrin, a heparan sulfate proteoglycan of developing axonal pathways.
J Comp Neurol. 1997; 383:1-17.
- Chernousov MA, Stahl RC, Carey DJ. Schwann cells use a novel collagen-dependent
mechanism for fibronectin fibril assembly. Journal of cell science. 1998; 111:2763–
2777.

- Bangratz M, Sarrazin N, Devaux J, Zambroni D, Echaniz-Laguna A, René F, Boërio D,
Davoine CS, Fontaine B, Feltri ML, Benoit E, Nicole S. A mouse model of Schwartz-
Jampel syndrome reveals myelinating Schwann cell dysfunction with persistent
axonal depolarization in vitro and distal peripheral nerve hyperexcitability when
perlecan is lacking. Am J Pathol. 2012; 180:2040-55.
- Litwack ED, Stipp CS, Kumbasar A, Lander AD. Neuronal expression of glypican, a
cell-surface glycosylphosphatidylinositol-anchored heparan sulfate proteoglycan, in
the adult rat nervous system. J Neurosci. 1994; 14:3713-24.
- Bloechlinger S, Karchewski LA, Woolf CJ. Dynamic changes in glypican-1 expression
in dorsal root ganglion neurons after peripheral and central axonal injury. Eur J
Neurosci. 2004; 19:1119-32.
- Iglesias BV, Centeno G, Pascuccelli H, Ward F, Peters MG, Filmus J, Puricelli L, de
Kier Joffé EB. Expression pattern of glypican-3 (GPC3) during human embryonic and
fetal development. Histol Histopathol. 2008; 23:1333-1340.
- Carey DJ, Evans DM, Stahl RC, Asundi VK, Conner KJ, Garbes P, Cizmeci-Smith G.
Molecular cloning and characterization of N-syndecan, a novel transmembrane
heparan sulfate proteoglycan. J Cell Biol. 1992; 117:191-201.
- Erdman R, Stahl RC, Rothblum K, Chernousov MA, Carey DJ. Schwann cell adhesion
to a novel heparan sulfate binding site in the N-terminal domain of alpha 4 type V
collagen is mediated by syndecan-3. J Biol Chem. 2002; 277:7619-7625.
- Goutebroze L, Carnaud M, Denisenko N, Boutterin MC, Girault JA. Syndecan-3 and
syndecan-4 are enriched in Schwann cell perinodal processes. BMC Neurosci. 2003;
4:29.
- Melendez-Vasquez C, Carey DJ, Zanazzi G, Reizes O, Maurel P, Salzer JL. Differential
expression of proteoglycans at central and peripheral nodes of Ranvier. Glia. 2005;
52:301-308.
-
- Carey DJ, Evans DM, Stahl RC, Asundi VK, Conner KJ, Garbes P, Cizmeci-Smith G.
Molecular cloning and characterization of N-syndecan, a novel transmembrane
heparan sulfate proteoglycan. J Cell Biol. 1992; 117:191-201.
- Melendez-Vasquez C, Carey DJ, Zanazzi G, Reizes O, Maurel P, Salzer JL. Differential
expression of proteoglycans at central and peripheral nodes of Ranvier. Glia. 2005;
52:301-308.
- Thomas SL, De Vries GH. Angiogenic expression profile of normal and
neurofibromin-deficient human Schwann cells. Neurochem Res. 2007; 32:1129-
1141.
- Bandtlow CE, Zimmermann DR. Proteoglycans in the developing brain: new
conceptual insights for old proteins. Physiol Rev. 2000; 80:1267-1290.
- Aquino DA, Margolis RU, Margolis RK. Immunocytochemical localization of a
chondroitin sulfate proteoglycan in nervous tissue. I. Adult brain, retina, and
peripheral nerve. J Cell Biol. 1984; 99:1117-1129.
- Tona A, Perides G, Rahemtulla F, Dahl D. Extracellular matrix in regenerating rat
sciatic nerve: a comparative study on the localization of laminin, hyaluronic acid,

and chondroitin sulfate proteoglycans, including versican. J Histochem Cytochem.
1993 Apr;41(4):593-9.
- Morrison JC, Rask P, Johnson EC, Deppmeier L. Chondroitin sulfate proteoglycan
distribution in the primate optic nerve head. Invest Ophthalmol Vis Sci. 1994;
35:838-845.
- Bode-Lesniewska B, Dours-Zimmermann MT, Odermatt BF, Briner J, Heitz PU,
Zimmermann DR. Distribution of the large aggregating proteoglycan versican in
adult human tissues. J Histochem Cytochem. 1996 Apr;44(4):303-12.
- Rezajooi K, Pavlides M, Winterbottom J, Stallcup WB, Hamlyn PJ, Lieberman AR,
Anderson PN. NG2 proteoglycan expression in the peripheral nervous system:
upregulation following injury and comparison with CNS lesions. Mol Cell Neurosci.
2004; 25:572-584.
- Ali SA, Hosaka YZ, Uehara M. Spatiotemporal distribution of chondroitin sulfate
proteoglycans in the developing mouse retina and optic nerve. J Vet Med Sci. 2011;
73:13-8.
- Melendez-Vasquez C, Carey DJ, Zanazzi G, Reizes O, Maurel P, Salzer JL. Differential
expression of proteoglycans at central and peripheral nodes of Ranvier. Glia. 2005;
52:301-308.
- Hanemann CO, Kuhn G, Lie A, Gillen C, Bosse F, Spreyer P, Müller HW. Expression
of decorin mRNA in the nervous system of rat. J Histochem Cytochem. 1993;
41:1383-1391.
- Wilda M, Bächner D, Just W, Geerkens C, Kraus P, Vogel W, Hameister H. A
comparison of the expression pattern of five genes of the family of small leucine-
rich proteoglycans during mouse development. J Bone Miner Res. 2000; 15:2187-
2196.
- Ali SA, Hosaka YZ, Uehara M. Spatiotemporal distribution of chondroitin sulfate
proteoglycans in the developing mouse retina and optic nerve. J Vet Med Sci. 2011;
73:13-8.
- Halata Z. The mechanoreceptors of the mammalian skin ultrastructure and
morphological classification. Adv Anat Embryol Cell Biol. 1975; 50:3-77.
- Zelená J (1994) Nerves and mechanoreceptors. Chapman and Hall, London Glasgow
Weinheim, pp 1–355
- Vega JA, Esteban I, Naves FJ, del Valle ME, Malinovsky L. Immunohistochemical
localization of laminin and type IV collagen in human cutaneous sensory nerve
formations. Anat Embryol (Berl). 1995; 191: 33-39.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.
- Chouchkov C, Andreev D, Dandov A. Localization and distribution of laminin in the
basal lamina of certain mechanoreceptors--an immunogold study. Somatosens Mot
Res. 2003; 20:265-70.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.

- Dubový P, Svízenská I, Jancálek R, Klusáková I. Immunohistochemical localization of
some extracellular molecules and their integrin receptors in the rat Pacinian
corpuscles. Gen Physiol Biophys. 1999; 18 Suppl 1:66-8.
- Pawson L, Slepecky NB, Bolanowski SJ. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens Mot Res. 2000; 17:159-170.
- Takahashi-Iwanaga H, Shimoda H. The three-dimensional microanatomy of
Meissner corpuscles in monkey palmar skin. J Neurocytol. 2003; 32:363-371.
- Dubový P, Svízenská I, Jancálek R, Klusáková I. Immunohistochemical localization of
some extracellular molecules and their integrin receptors in the rat Pacinian
corpuscles. Gen Physiol Biophys. 1999; 18 Suppl 1:66-8.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.
- Sames K, Halata Z, Jojovic M, van Damme EJ, Peumans WJ, Delpech B, Asmus B,
Schumacher U. Lectin and proteoglycan histochemistry of feline pacinian
corpuscles. J Histochem Cytochem. 2001; 49:19-28.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.
- Barros CS, Franco SJ, Müller U. Extracellular matrix: functions in the nervous
system. Cold Spring Harb Perspect Biol. 2011; 3(1):a005108.
- Gardiner NJ. Integrins and the extracellular matrix: key mediators of development
and regeneration of the sensory nervous system. Dev Neurobiol. 2011; 71:1054-
1072.
- Perris R, Krotoski D, Lallier T, Domingo C, Sorrell JM, Bronner-Fraser M. Spatial and
temporal changes in the distribution of proteoglycans during avian neural crest
development. Development. 1991; 111:583-599.
- Perris R. The extracellular matrix in neural crest-cell migration. Trends Neurosci.
1997; 20:23-31.
- Melendez-Vasquez C, Carey DJ, Zanazzi G, Reizes O, Maurel P, Salzer JL. Differential
expression of proteoglycans at central and peripheral nodes of Ranvier. Glia. 2005;
52:301-308.
- Syndecan-3 and syndecan-4 are enriched in Schwann cell perinodal processes.
Goutebroze L, Carnaud M, Denisenko N, Boutterin MC, Girault JA. BMC Neurosci.
2003; 4:29.
- McGarvey ML, Baron-Van Evercooren A, Kleinman HK, Dubois-Dalcq M. Synthesis
and effects of basement membrane components in cultured rat Schwann cells. Dev
Biol. 1984; 105:18-28.
- Anton ES, Sandrock AW Jr, Matthew WD. Merosin promotes neurite growth and
Schwann cell migration in vitro and nerve regeneration in vivo: evidence using an
antibody to merosin, ARM-1. Dev Biol. 1994; 164:133-146.
- Shorer Z, Philpot J, Muntoni F, Sewry C, Dubowitz V. Demyelinating peripheral
neuropathy in merosin-deficient congenital muscular dystrophy. J Child Neurol.
1995; 10:472-475.
- Feltri ML, Graus Porta D, Previtali SC, Nodari A, Migliavacca B, Cassetti A,
Littlewood-Evans A, Reichardt LF, Messing A, Quattrini A, Mueller U, Wrabetz L.

Conditional disruption of beta 1 integrin in Schwann cells impedes interactions
with axons. J Cell Biol. 2002; 156:199-209.
- Halfter W, Schurer B, Yip J, Yip L, Tsen G, Lee JA, Cole GJ. Distribution and substrate
properties of agrin, a heparan sulfate proteoglycan of developing axonal pathways.
J Comp Neurol. 1997; 383:1-17.
- Litwack ED, Stipp CS, Kumbasar A, Lander AD. Neuronal expression of glypican, a
cell-surface glycosylphosphatidylinositol-anchored heparan sulfate proteoglycan, in
the adult rat nervous system. J Neurosci. 1994; 14:3713-24.
- Bloechlinger S, Karchewski LA, Woolf CJ. Dynamic changes in glypican-1 expression
in dorsal root ganglion neurons after peripheral and central axonal injury. Eur J
Neurosci. 2004; 19:1119-32.
- Vega JA, Esteban I, Naves FJ, del Valle ME, Malinovsky L. Immunohistochemical
localization of laminin and type IV collagen in human cutaneous sensory nerve
formations. Anat Embryol (Berl). 1995; 191: 33-39.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.
- Chouchkov C, Andreev D, Dandov A. Localization and distribution of laminin in the
basal lamina of certain mechanoreceptors--an immunogold study. Somatosens Mot
Res. 2003; 20:265-70.
- Beck K, Hunter I, Engel J. Structure and function of laminin: anatomy of a
multidomain glycoprotein. FASEB J. 1990; 4:148-60.
- Edgar D. Neuronal laminin receptors. Trends Neurosci. 1989; 12:248-51.
- Baron-Van Evercooren A, Kleinman HK, Ohno S, Marangos P, Schwartz JP, Dubois-
Dalcq ME. Nerve growth factor, laminin, and fibronectin promote neurite growth in
human fetal sensory ganglia cultures. J Neurosci Res. 1982; 8:179-193.
- Wang GY, Hirai K, Shimada H. The role of laminin, a component of Schwann cell
basal lamina, in rat sciatic nerve regeneration within antiserum-treated nerve
grafts. Brain Res. 1992 Jan 20;570(1-2):116-25.
- Feltri ML, Graus Porta D, Previtali SC, Nodari A, Migliavacca B, Cassetti A,
Littlewood-Evans A, Reichardt LF, Messing A, Quattrini A, Mueller U, Wrabetz L.
Conditional disruption of beta 1 integrin in Schwann cells impedes interactions
with axons. J Cell Biol. 2002; 156:199-209.
- Pawson L, Slepecky NB, Bolanowski SJ. Immunocytochemical identification of
proteins within the Pacinian corpuscle. Somatosens Mot Res. 2000; 17:159-170.
- Takahashi-Iwanaga H, Shimoda H. The three-dimensional microanatomy of
Meissner corpuscles in monkey palmar skin. J Neurocytol. 2003; 32:363-371.
- Koopmans G, Hasse B, Sinis N. Chapter 19: The role of collagen in peripheral nerve
repair. Int Rev Neurobiol. 2009;87:363-79.
- Dubový P, Svízenská I, Jancálek R, Klusáková I. Immunohistochemical localization of
some extracellular molecules and their integrin receptors in the rat Pacinian
corpuscles. Gen Physiol Biophys. 1999; 18 Suppl 1:66-8.
- Dubový P, Bednárová J. The extracellular matrix of rat pacinian corpuscles: an
analysis of its fine structure. Anat Embryol (Berl). 1999; 200:615-23.

- Sames K, Halata Z, Jojovic M, van Damme EJ, Peumans WJ, Delpech B, Asmus B,
Schumacher U. Lectin and proteoglycan histochemistry of feline pacinian
corpuscles. J Histochem Cytochem. 2001; 49:19-28.
- Sames K, Halata Z, Jojovic M, van Damme EJ, Peumans WJ, Delpech B, Asmus B,
Schumacher U. Lectin and proteoglycan histochemistry of feline pacinian
corpuscles. J Histochem Cytochem. 2001; 49:19-28.
- Kelly EJ, Terenghi G, Hazari A, Wiberg M. Nerve fibre and sensory end organ density
in the epidermis and papillary dermis of the human hand. Br J Plast Surg. 2005;
58:774-779.
- Chang YC, Lin WM, Hsieh ST. Effects of aging on human skin innervation.
Neuroreport. 2004; 15:149-153.
- Panoutsopoulou IG, Wendelschafer-Crabb G, Hodges JS, Kennedy WR. Skin blister
and skin biopsy to quantify epidermal nerves: a comparative study. Neurology.
2009; 72:1205-1210.
- Wickremaratchi MM, Llewelyn JG. Effects of ageing on touch. Postgrad Med J.
2006; 82:301-304.
- Taguchi T, Ota H, Matsuda T, Murase S, Mizumura K. Cutaneous C-fibernociceptor
responses and nociceptive behaviors in aged Sprague-Dawley rats.Pain. 2010;
151:771-782.
- Decorps J, Saumet JL, Sommer P, Sigaudo-Roussel D, Fromy B. Effect of ageing on
tactile transduction processes. Ageing Res Rev. 2014; 13:90-99.
- Cerimele D, Celleno L, Serri F. Physiological changes in ageing skin. Br J Dermatol.
1990; 122 Suppl 35:13-20.
- Stevens JC, Patterson MQ. Dimensions of spatial acuity in the touch sense: changes
over the life span. Somatosens Mot Res. 1995; 12:29-47.
- Verrillo RT, Bolanowski SJ, Gescheider GA. Effects of ageing on the subjective
magnitude of vibration. Somatosens Mot Res 2002; 19:238-244.
- Perry SD. Evaluation of age-related plantar-surface insensitivity and onset age of
advanced insensitivity in older adults using vibratory and touch sensation tests.
Neurosci Lett. 2006; 392:62-67.
- Chase MH, Engelhardt JK, Adinolfi AM, Chirwa SS. Age-dependent changes in cat
masseter nerve: an electrophysiological and morphological study. Brain Res. 1992;
586:279-288.
- Bouche P, Cattelin F, Saint-Jean O, Léger JM, Queslati S, Guez D, Moulonguet A,
Brault Y, Aquino JP, Simunek P. Clinical and electrophysiological study of the
peripheral nervous system in the elderly. J Neurol. 1993; 240:263-268.
- Verdú E, Ceballos D, Vilches JJ, Navarro X. Influence of aging on peripheral nerve
function and regeneration. J Peripher Nerv Syst. 2000; 5:191-208.
- Di Iorio A, Cherubini A, Volpato S, Sparvieri E, Lauretani F, Franceschi C, Senin U,
Abate G, Paganelli R, Martin A, Andres-Lacueva C, Ferrucci L. Markers of
inflammation, vitamin E and peripheral nervous system function: the InCHIANTI
study. Neurobiol Aging. 2006; 27:1280-1288.
- Verrillo RT. Change in vibrotactile thresholds as a function of age. Sens Processes.
1979; 3:49-59.

- Gescheider GA, Bolanowski SJ, Hall KL, Hoffman KE, Verrillo RT. The effects of aging
on information-processing channels in the sense of touch: I. Absolute sensitivity.
Somatosens Mot Res. 1994; 11:345-357.
- Goble AK, Collins AA, Cholewiak RW. Vibrotactile threshold in young and old
observers: the effect of spatial summation and the presence of a rigid surround. J
Acoust Soc Am 1996; 99:2256-2269.
- Verrillo RT, Bolanowski SJ, Gescheider GA. Effects of ageing on the subjective
magnitude of vibration. Somatosens Mot Res 2002; 19:238-244.
- Perry SD. Evaluation of age-related plantar-surface insensitivity and onset age of
advanced insensitivity in older adults using vibratory and touch sensation tests.
Neurosci Lett. 2006; 392:62-67.
- Stevens JC, Patterson MQ. Dimensions of spatial acuity in the touch sense: changes
over the life span. Somatosens Mot Res. 1995; 12:29-47.
- Leveque JL, Dresler J, Ribot-Ciscar E, Roll JP, Poelman C. Changes in tactile spatial
discrimination and cutaneous coding properties by skin hydration in theelderly. J.
Invest. Dermatol. 2000; 115:454-458.
- Bruce MF. The relation of tactile thresholds to histology in the fingers of elderly
people. J Neurol Neurosurg Psychiatry 1980; 43:730-734.
- Thornbury JM, Mistretta CM. Tactile sensitivity as a function of age. J Gerontol.
1981; 36:34-39.
- Johnson KO, Yoshioka T, Vega-Bermudez F. Tactile functions of mechanoreceptive
afferents innervating the hand. J Clin Neurophysiol. 2000; 17:539-558.
- Kelly EJ, Terenghi G, Hazari A, Wiberg M. Nerve fibre and sensory end organ density
in the epidermis and papillary dermis of the human hand. Br J Plast Surg. 2005;
58:774-779.
- Nolano M, Provitera V, Estraneo A, Selim MM, Caporaso G, Stancanelli A,
Saltalamacchia AM, Lanzillo B, Santoro L. Sensory deficit in Parkinson's disease:
evidence of a cutaneous denervation. Brain. 2008; 131:1903-1111.
- Herrmann DN, Boger JN, Jansen C, Alessi-Fox C. In vivo confocal microscopy of
Meissner corpuscles as a measure of sensory neuropathy. Neurology. 2007;
69:2121-2127.
- Nolano M, Provitera V, Crisci C, Stancanelli A, Wendelschafer-Crabb G, Kennedy
WR, Santoro L. Quantification of myelinated endings and mechanoreceptors in
human digital skin. Ann Neurol. 2003; 54:197-205.
- Kelly EJ, Terenghi G, Hazari A, Wiberg M. Nerve fibre and sensory end organ density
in the epidermis and papillary dermis of the human hand. Br J Plast Surg. 2005;
58:774-779.
- Herrmann DN, Boger JN, Jansen C, Alessi-Fox C. In vivo confocal microscopy of
Meissner corpuscles as a measure of sensory neuropathy. Neurology. 2007;
69:2121-2127.
- Bolton CF, Winkelmann RK, Dyck PJ. A quantitative study of Meissner’s corpuscles
in man. Neurology 1966; 16:1-9.
- Bruce MF. The relation of tactile thresholds to histology in the fingers of elderly
people. J Neurol Neurosurg Psychiatry 1980; 43:730-734.

- Schimrigk K, Ruttinger H. The touch corpuscles of the plantar surface of the big toe.
Histological and histometrical investigations with respect to age. Eur Neurol 1980;
19:49-60.
- Thornbury JM, Mistretta CM. Tactile sensitivity as a function of age. J Gerontol.
1981; 36:34-39.
- Halata Z, Munger BL. The sensory innervation of primate facial skin. II. Vermilion
border and mucosa of lip. Brain Res. 1983; 286:81-107.
- Iwasaki T, Goto N, Goto J, Ezure H, Moriyama H. The aging of human Meissner's
corpuscles as evidenced by parallel sectioning. Okajimas Folia Anat Jpn. 2003;
79:185-189.
- Mathewson RC, Nava PB. Effects of age on Meissner corpuscles: a study of silver-
impregnated neurites in mouse digital pads. J Comp Neurol. 1985; 231:250-259.
- Nava PB, Mathewson RC. Effect of age on the structure of Meissner corpuscles in
murine digital pads. Microsc Res Tech. 1996; 34:376-389.
- Thornbury JM, Mistretta CM. Tactile sensitivity as a function of age. J Gerontol.
1981; 36:34-39.
- Bolton CF, Winkelmann RK, Dyck PJ. A quantitative study of Meissner’s corpuscles
in man. Neurology 1966; 16:1-9.
- Bruce MF. The relation of tactile thresholds to histology in the fingers of elderly
people. J Neurol Neurosurg Psychiatry 1980; 43:730-734.
- Thornbury JM, Mistretta CM. Tactile sensitivity as a function of age. J Gerontol.
1981; 36:34-39.
- Halata Z, Munger BL. The sensory innervation of primate facial skin. II. Vermilion
border and mucosa of lip. Brain Res. 1983; 286:81-107.
- Bolton CF, Winkelmann RK, Dyck PJ. A quantitative study of Meissner’s corpuscles
in man. Neurology 1966; 16:1-9.
- Bruce MF. The relation of tactile thresholds to histology in the fingers of elderly
people. J Neurol Neurosurg Psychiatry 1980; 43:730-734.
- Nolano M, Provitera V, Crisci C, Stancanelli A, Wendelschafer-Crabb G, Kennedy
WR, Santoro L. Quantification of myelinated endings and mechanoreceptors in
human digital skin. Ann Neurol. 2003; 54:197-205.
- Kelly EJ, Terenghi G, Hazari A, Wiberg M. Nerve fibre and sensory end organ density
in the epidermis and papillary dermis of the human hand. Br J Plast Surg. 2005;
58:774-779.
- Nava PB, Mathewson RC. Effect of age on the structure of Meissner corpuscles in
murine digital pads. Microsc Res Tech. 1996; 34:376-389.
- Mathewson RC, Nava PB. Effects of age on Meissner corpuscles: a study of silver-
impregnated neurites in mouse digital pads. J Comp Neurol. 1985; 231:250-259.
- Bolton CF, Winkelmann RK, Dyck PJ. A quantitative study of Meissner’s corpuscles
in man. Neurology 1966; 16:1-9.
- Bruce MF. The relation of tactile thresholds to histology in the fingers of elderly
people. J Neurol Neurosurg Psychiatry 1980; 43:730-734.

- Matsuoka S, Suzuki H, Morioka S, Ogawa Y, Kojima T. Quantitative and qualitative
studies of Meissner's corpuscles in human skin, with special reference to
alterations caused by aging. J Dermatol. 1983; 10:205-216.
- Iwasaki T, Goto N, Goto J, Ezure H, Moriyama H. The aging of human Meissner's
corpuscles as evidenced by parallel sectioning. Okajimas Folia Anat Jpn. 2003;
79:185-189.
- Cauna N, Mannan G. The structure of human digital pacinian corpuscles (corpus
cula lamellosa) and its functional significance. J Anat. 1958; 92:1-20.
- Gescheider GA, Beiles EJ, Checkosky CM, Bolanowsky SJ, Verrillo RT. The effects of
ageing on information processing channels in the sense of touch: II. Temporal
summation in the P channel. Somatosens Mot Res 1994; 11:359-365.
- Cauna N, Mannan G. The structure of human digital pacinian corpuscles (corpus
cula lamellosa) and its functional significance. J Anat. 1958; 92:1-20.
- Verrillo RT. Change in vibrotactile thresholds as a function of age. Sens Processes.
1979; 3:49-59.
- Moll R, Moll I, Franke WW. Identification of Merkel cells in human skin by specific
cytokeratin antibodies: changes of cell density and distribution in fetal and adult
plantar epidermis. Differentiation. 1984; 28:136-154.
- Fundin BT, Bergman E, Ulfhake B. Alterations in mystacial pad innervationin the
aged rat. Exp. Brain Res. 1997; 117:324-340.
- Nolano M, Provitera V, Crisci C, Stancanelli A, Wendelschafer-Crabb G, Kennedy
WR, Santoro L. Quantification of myelinated endings and mechanoreceptors in
human digital skin. Ann Neurol. 2003; 54:197-205.
- Moll R, Moll I, Franke WW. Identification of Merkel cells in human skin by specific
cytokeratin antibodies: changes of cell density and distribution in fetal and adult
plantar epidermis. Differentiation. 1984; 28:136-154.
- Kim DK, Holbrook KA. The appearance, density, and distribution of Merkel cells in
human embryonic and fetal skin: their relation to sweat gland and hair follicle
development. J Invest Dermatol. 1995; 104:411-416.

Curriculum Vitae Jorge García-Piqueras
Personal information
First and last names: Jorge García Piqueras
Nationality: Spanish
Date and place of birth: 15/07/1992, Calatayud (Spain)
Telephone number: 637048404
Current residence: Catedrático Doctor Flemng 6 1B
Calatayud (Spain), 50300
Work experience
October 2017 – February 2018:
Research Laboratory Technician. Department of Surgery and Medical Surgical Specialities of the University of Oviedo (Spain) and Fernández Vega Ophthalmological Institute (Oviedo, Spain). National Research Project: MINECO (Innovative solutions for the treatment of dry eye combined with ocular pain). Immunohistochemical tasks consisted in comparative studies on the innervation of elderly/young mouse eyes.
University laboratory experience
September 2016 – Currently:
PhD thesis. Department of Cellular Biology and Morphology of the University of Oviedo
(Spain). “The extracellular matrix in vertebrate mechanoreceptors”.
Immunohistochemical study of vertebrate sensory corpuscles: quantitative/qualitative
changes in relation to aging and the proteoglycan/glycosaminoglycan composition of
the extracellular matrix.
February 2016 – July 2016:
Master’s thesis. Department of Experimental, Diagnostic and Specialty Medicine of the
University of Bologna (Italy) and S.Orsola-Malpighi Hospital (Bologna, Italy).
“Identification of microRNA expression profile in the adenocarcinoma of the
esophagus and cardia”. Characterization of deregulated microRNA in association to
different histotypes of esophageal-cardia adenocarcinoma, which were determined
previously by immunohistochemical and clinical parameters.
February 2015 – July 2015:
Practicum. Department of Cellular Biology and Morphology of the University of Oviedo
(Spain). Several immunohistochemical/immunocytochemical studies on human tissues
and Zebrafish model: peripheral nervous system, cancer research, virus infection, etc.
March 2014 – June 2014:
Bachelor's thesis. Department of Biosciences of the University of Milano (Italy).
“Cloning of a protozoan protein in E.coli and genotyping of transgenic plants”.

Curriculum Vitae Jorge García-Piqueras
Education and degrees
September 2017 – Currently:
PhD in Health Sciences: University of Oviedo (Spain).
September 2014 – July 2016:
International Double Master’s Degree in (120 ECTS)*:
- Biotechnology of Environment and Health: University of Oviedo (Spain).
- Medical Biotechnology: University of Bologna (Italy).
* 1st academic year at Oviedo University. 2nd academic year at Bologna University.
September 2010 – July 2014:
Bachelor's Degree in Biotechnology (240 ECTS): Universidad of Zaragoza (Spain).
*4th academic year at Milano University (Erasmus Programme).
Supplementary courses
December 2016:
Formation in Laboratory Animal Science (by Animalaria SL according to EU legislation):
- Function B: Euthanasia.
- Function C: Experimental procedures.
- Function D: Procedure and project design.
October 2015:
Diploma of University Expert in Assisted Reproduction (15 ECTS): UNED (Spain).
Diploma of University Expert in Biomedical Research and Law (15 ECTS): UNED (Spain).
Personal skills
Mother tongue: Spanish
Other languages:
- English: B2 level. First Certificate in English: Cambridge University (England).
- Italian: B2 level. CALCIF Certificate: Milano University (Italy).
Grants/Scholarships
1. Erasmus grant (September 2015 - July 2016): Bologna University (Italy).
For the second academic year of the Master’s Degree.
2. Erasmus grant (September 2013 - July 2014): Milano University (Italy).
For the fourth academic year of the Bachelor’s Degree.

Curriculum Vitae Jorge García-Piqueras
Scientific publications
1. García-Piqueras J, Carcaba L, García-Mesa Y, Feito J, García B, Viña E, Suárez-Quintanilla J, Cobo J, Vega JA, García-Suárez O. Chondroitin Sulfate in Human Cutaneous Meissner and Pacinian Sensory Corpuscles. Anat Rec (Hoboken). 2018 Oct 9. doi: 10.1002/ar.23951. [Epub ahead of print].
2. Olivia García-Suárez, Yolanda García-Mesa, Jorge García-Piqueras, Giuseppina Salvo, Juan L. Cobo, Elda Alba, Ramón Cobo, Jorge Feito and José A. Vega. The Cutaneous Biopsy for the Diagnosis of Peripheral Neuropathies: Meissner’s Corpuscles and Merkel’s Cells. DOI: 10.5772/intechopen.81687
3. Feito J, García-Suárez O, García-Piqueras J, García-Mesa Y, Pérez-Sánchez A, Suazo I, Cabo R, Suárez-Quintanilla J, Cobo J, Vega JA. The development of human digital Meissner's and Pacinian corpuscles. Ann Anat. 2018 May 26;219:8-24.
4. García-Mesa Y, García-Piqueras J, García B, Feito J, Cabo R, Cobo J, Vega JA, García-Suárez O. Merkel cells and Meissner's corpuscles in human digital skin display Piezo2 immunoreactivity. J Anat. 2017 Dec;231(6):978-989.
5. García-Piqueras J, García-Suárez O, Rodríguez-González MC, Cobo JL, Cabo R, Vega
JA, Feito J. Endoneurial-CD34 positive cells define an intermediate layer in human digital Pacinian corpuscles. Ann Anat. 2017 May;211:55-60.
6. Mohamedi Y, Fontanil T, Solares L, Garcia-Suárez O, García-Piqueras J, Vega JA, Cal
S, Obaya AJ. Fibulin-5 downregulates Ki-67 and inhibits proliferation and invasion
of breast cancer cells. Int J Oncol. 2016 Apr;48(4):1447-56.
Congresses
1. García-Piqueras J, García B, García-Mesa Y, Cabo R, Feito J, Cárcaba-García L, Vega JA, García-Suárez O. Keratan sulfate proteoglycans in the extracellular matrix of human Meissner and Pacinian corpuscles. 28th Congress of the Spanish Anatomical Society. 1-3 February, 2018, Badajoz (Spain).
2. García-Piqueras J, García-Mesa Y, Muriel-Rodríguez JD, Torres-Parejo I, Feito J,
Cabo R, Vega JA, García-Suárez O. Quantitative and qualitative age-dependent changes in Meissner’s corpuscles and Merkel cells in human digital glabrous skin. 28th Congress of the Spanish Anatomical Society. 1-3 February, 2018, Badajoz (Spain).
3. García-Mesa Y, García-Piqueras J, Feito J, Cabo P, García-Suárez O, Vega JA. The
Meissner’s corpuscles and Merkel’s cells in multiple sclerosis and amyotrophic lateral sclerosis. 28th Congress of the Spanish Anatomical Society. 1-3 February, 2018, Badajoz (Spain).
4. García-Mesa Y, García-Piqueras J, Feito J, Torres-Parejo Isidro, Cobo R, Cabo P,
García-Suárez O, Vega JA. The validity of Meissner’s corpuscles and Merkel’s cells

Curriculum Vitae Jorge García-Piqueras
analysis in skin biopsies for the diagnosis of peripheral neuropathies. 28th Congress of the Spanish Anatomical Society. 1-3 February, 2018, Badajoz (Spain).
5. García-Suárez O, García-Piqueras J, Cárcaba-García L, Viña E, Cobo J, Pérez-Moltó
FJ, Vega JA. The extracellular matrix composition of the Pacinian and Meissner’s corpuscles in digital skin. 27th Congress of the Spanish Anatomical Society. 2-4 September 2016, Oviedo (Spain).
Related Documents











