Review Knowledge discovery in metabolomics: An overview of MS data handling While metabolomics attempts to comprehensively analyse the small molecules char- acterising a biological system, MS has been promoted as the gold standard to study the wide chemical diversity and range of concentrations of the metabolome. On the other hand, extracting the relevant information from the overwhelming amount of data generated by modern analytical platforms has become an important issue for knowledge discovery in this research field. The appropriate treatment of such data is therefore of crucial importance in order, for the data, to provide valuable information. The aim of this review is to provide a broad overview of the methodologies developed to handle and process MS metabolomic data, compare the samples and highlight the relevant metabo- lites, starting from the raw data to the biomarker discovery. As data handling can be further separated into data processing, data pre-treatment and data analysis, recent advances in each of these steps are detailed separately. Keywords: Data mining / Data processing / Metabolomics / MS DOI 10.1002/jssc.200900609 1 Metabolomics 1.1 Systems biology Metabolomics (also known as metabonomics) is a recent discipline that attempts to globally study metabolites and their concentrations, interactions and dynamics within complex samples [1]. It constitutes one of the tools of the post-genomic era [2], which is concerned with the study of the different functional levels of a biological system, i.e. the transcriptome, the proteome and the metabolome [3, 4]. As metabolites can be considered the downstream products of cellular regulatory processes [5], metabolomics data can precisely characterise cells, tissues, biofluids or whole organisms by defining specific biochemical phenotypes that are representative of physiological or developmental states. The metabolome is the holistic quantitative set of low- molecular-weight compounds ( o1000 Da), including many hundreds or thousands of molecules such as carbohydrates, vitamins, lipids and amino or fatty acids. These metabolites participate in the metabolic reactions necessary for normal functions, the maintenance or growth of a cell [2]. Their origin can be either endogenous such as the products of biosynthesis and catabolism or exogenous such as nutrients or pharmaceutical compounds degradation products [6, 7]. The chemical properties of the organic or inorganic constituents of this set are greatly variable and this diversity is a critical aspect to consider [8]. The variability in the molecular weight, polarity or solubility and the wide dynamic range of concentrations (from pmol to mmol) constitute additional difficulties when analysing metabolites while no amplification process is available. 1.2 Applications of metabolomics Measuring metabolites changes could offer deeper insights into biological mechanisms by describing the responses of systems to environmental or genetic modifications. Meta- bolomic analyses constitute a potent tool for the discovery of biomarkers related to a physiological response and the diagnosis of complex phenotypes. Numerous applications have already been developed in a variety of research fields of the post-genomic era, from medical science to agriculture. Although early metabolic studies focused on pre-selec- ted specific compounds, modern untargeted approaches represent efficient tools for the early detection of diseases [9–11]. The screening of drug candidates constitutes another prominent application through the assessment of the effects of metabolic modifications or toxicity [12, 13]. In addition, metabolomics has recently received an increasing interest from the nutrition research field [14]. Several applications Julien Boccard Jean-Luc Veuthey Serge Rudaz School of Pharmaceutical Sciences, University of Geneva, University of Lausanne, Geneva, Switzerland Received September 29, 2009 Revised November 3, 2009 Accepted November 3, 2009 Abbreviations: ANN, artificial neural networks; DIMS, direct infusion mass spectrometry; HCA, hierarchical cluster analysis; PCA, principal component analysis; PLS, projection to latent structures by means of partial least squares; SVM, support vector machine; UV, unit variance Correspondence: Dr. Serge Rudaz, School of Pharmaceutical Sciences – EPGL, University of Geneva, 20 Bd d’Yvoy, 1211 Geneva 4, Switzerland E-mail: [email protected] Fax: 141-22-379-68-08 & 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com J. Sep. Sci. 2010, 33, 1–15 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Review
Knowledge discovery in metabolomics: Anoverview of MS data handling
While metabolomics attempts to comprehensively analyse the small molecules char-
acterising a biological system, MS has been promoted as the gold standard to study the
wide chemical diversity and range of concentrations of the metabolome. On the other
hand, extracting the relevant information from the overwhelming amount of data
generated by modern analytical platforms has become an important issue for knowledge
discovery in this research field. The appropriate treatment of such data is therefore of
crucial importance in order, for the data, to provide valuable information. The aim of this
review is to provide a broad overview of the methodologies developed to handle and
process MS metabolomic data, compare the samples and highlight the relevant metabo-
lites, starting from the raw data to the biomarker discovery. As data handling can be
further separated into data processing, data pre-treatment and data analysis, recent
advances in each of these steps are detailed separately.
Keywords: Data mining / Data processing / Metabolomics / MSDOI 10.1002/jssc.200900609
1 Metabolomics
1.1 Systems biology
Metabolomics (also known as metabonomics) is a recent
discipline that attempts to globally study metabolites and
their concentrations, interactions and dynamics within
complex samples [1]. It constitutes one of the tools of the
post-genomic era [2], which is concerned with the study of
the different functional levels of a biological system, i.e. the
transcriptome, the proteome and the metabolome [3, 4]. As
metabolites can be considered the downstream products of
cellular regulatory processes [5], metabolomics data can
precisely characterise cells, tissues, biofluids or whole
organisms by defining specific biochemical phenotypes that
are representative of physiological or developmental states.
The metabolome is the holistic quantitative set of low-
molecular-weight compounds (o1000 Da), including many
hundreds or thousands of molecules such as carbohydrates,
vitamins, lipids and amino or fatty acids. These metabolites
participate in the metabolic reactions necessary for normal
functions, the maintenance or growth of a cell [2]. Their
origin can be either endogenous such as the products of
biosynthesis and catabolism or exogenous such as nutrients
or pharmaceutical compounds degradation products [6, 7].
The chemical properties of the organic or inorganic
constituents of this set are greatly variable and this diversity
is a critical aspect to consider [8]. The variability in the
molecular weight, polarity or solubility and the wide
dynamic range of concentrations (from pmol to mmol)
constitute additional difficulties when analysing metabolites
while no amplification process is available.
1.2 Applications of metabolomics
Measuring metabolites changes could offer deeper insights
into biological mechanisms by describing the responses of
systems to environmental or genetic modifications. Meta-
bolomic analyses constitute a potent tool for the discovery of
biomarkers related to a physiological response and the
diagnosis of complex phenotypes. Numerous applications
have already been developed in a variety of research fields of
the post-genomic era, from medical science to agriculture.
Although early metabolic studies focused on pre-selec-
ted specific compounds, modern untargeted approaches
represent efficient tools for the early detection of diseases
[9–11]. The screening of drug candidates constitutes another
prominent application through the assessment of the effects
of metabolic modifications or toxicity [12, 13]. In addition,
metabolomics has recently received an increasing interest
from the nutrition research field [14]. Several applications
Julien BoccardJean-Luc VeutheySerge Rudaz
School of PharmaceuticalSciences, University of Geneva,University of Lausanne, Geneva,Switzerland
Received September 29, 2009Revised November 3, 2009Accepted November 3, 2009
Abbreviations: ANN, artificial neural networks; DIMS, directinfusion mass spectrometry; HCA, hierarchical clusteranalysis; PCA, principal component analysis; PLS,
projection to latent structures by means of partial leastsquares; SVM, support vector machine; UV, unit variance
Correspondence: Dr. Serge Rudaz, School of PharmaceuticalSciences – EPGL, University of Geneva, 20 Bd d’Yvoy, 1211Geneva 4, SwitzerlandE-mail: [email protected]: 141-22-379-68-08
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com
J. Sep. Sci. 2010, 33, 1–15 1
are foreseen, such as food composition analyses, quality and
authenticity assessments and the monitoring of the
physiological consequences of specific diets. Additionally,
metabolomics is expected to greatly benefit agriculture,
plant biochemistry, phytomedicine and natural product
approaches [4, 15]. Metabolome analysis represents a key
tool to uncover the roles of genes in functional genomics
[16]. Genotyping and phenotyping will certainly provide
precious information to link the sequence of a gene to its
function.
2 Analytical methods
2.1 General considerations
Due to the high chemical diversity of the metabolome,
complementary analytical techniques are required to
monitor it completely [17]. Moreover, sensitivity constitutes
a major concern, because there is no available method of
amplification as in the case of transcriptome analysis.
Multiple analytical platforms have already been applied to
metabolomic studies [18], such as direct infusion MS
(DIMS), hyphenated separation technique MS platforms,
i.e. GC [19–23], LC [24–26] or CE [27–30], as well as NMR [1,
31, 32], Fourier transform-infrared spectroscopy [33] or
accelerator MS [34]. The selection of the most appropriate
methodology can be considered as a compromise between
the chemical selectivity, sensitivity and speed of the different
techniques [35].
2.2 MS
MS is a well-established detection method employed in
many fields and is one of the essential instruments in
analytical laboratories. By measuring the m/z of elemental
or molecular species, it allows the simultaneous detection of
multiple analytes with high sensitivities. MS has
been demonstrated as a potent platform for metabolomics,
thanks to its ability to detect metabolites present at
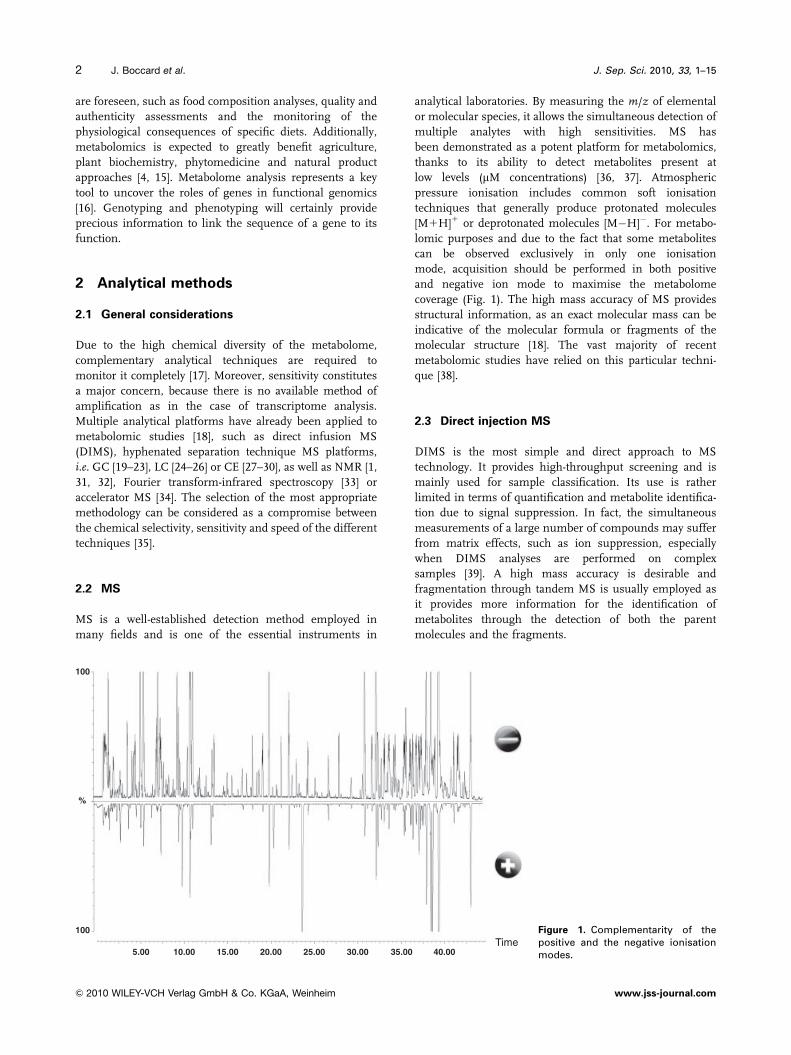
low levels (mM concentrations) [36, 37]. Atmospheric
pressure ionisation includes common soft ionisation
techniques that generally produce protonated molecules
[M1H]1 or deprotonated molecules [M�H]�. For metabo-
lomic purposes and due to the fact that some metabolites
can be observed exclusively in only one ionisation
mode, acquisition should be performed in both positive
and negative ion mode to maximise the metabolome
coverage (Fig. 1). The high mass accuracy of MS provides
structural information, as an exact molecular mass can be
indicative of the molecular formula or fragments of the
molecular structure [18]. The vast majority of recent
metabolomic studies have relied on this particular techni-
que [38].
2.3 Direct injection MS
DIMS is the most simple and direct approach to MS
technology. It provides high-throughput screening and is
mainly used for sample classification. Its use is rather
limited in terms of quantification and metabolite identifica-
tion due to signal suppression. In fact, the simultaneous
measurements of a large number of compounds may suffer
from matrix effects, such as ion suppression, especially
when DIMS analyses are performed on complex
samples [39]. A high mass accuracy is desirable and
fragmentation through tandem MS is usually employed as
it provides more information for the identification of
metabolites through the detection of both the parent
molecules and the fragments.
100
100
%
Time5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00
Figure 1. Complementarity of thepositive and the negative ionisationmodes.
J. Sep. Sci. 2010, 33, 1–152 J. Boccard et al.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com
2.4 Chromatography-MS
The hyphenation of MS with separative techniques such as
chromatography greatly increases the quality of the raw data
generated [18, 40]. The sequential introduction of
compounds into the mass spectrometer enables a higher
sensitivity and allows more metabolites to be detected but it
also involves an increase in the analysis time. Two main
separative methods are currently extensively used in
metabolomics, namely GC and LC.
2.4.1 GC-MS
GC-MS is well established and already employed [41–43].
The typical ionisation techniques are chemical ionisation,
which minimises fragmentation, and fragmentation
through electron impact [23]. GC-MS provides reproducible
and accurate measurements of volatile compounds and the
fragmentation pattern of these molecules [19, 21]. The
chemical derivatisation of semi-volatile compounds is
required to increase the volatility and produces ions that
can be separated in the GC column [44–46]. GC-MS was
historically the method-of-choice for the early development
of metabolomic studies [47, 48], and libraries have been
built to facilitate the identification of compounds [49].
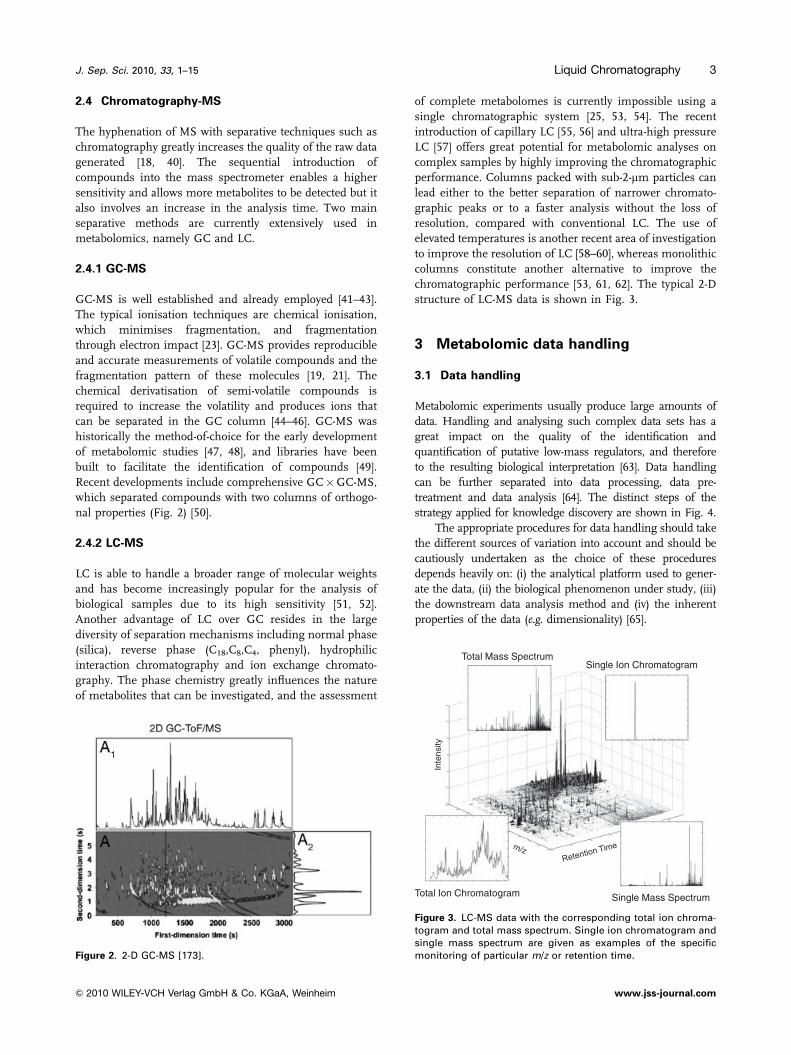
Recent developments include comprehensive GC�GC-MS,
which separated compounds with two columns of orthogo-
nal properties (Fig. 2) [50].
2.4.2 LC-MS
LC is able to handle a broader range of molecular weights
and has become increasingly popular for the analysis of
biological samples due to its high sensitivity [51, 52].
Another advantage of LC over GC resides in the large
diversity of separation mechanisms including normal phase
(silica), reverse phase (C18,C8,C4, phenyl), hydrophilic
interaction chromatography and ion exchange chromato-
graphy. The phase chemistry greatly influences the nature
of metabolites that can be investigated, and the assessment
of complete metabolomes is currently impossible using a
single chromatographic system [25, 53, 54]. The recent
introduction of capillary LC [55, 56] and ultra-high pressure
LC [57] offers great potential for metabolomic analyses on
complex samples by highly improving the chromatographic
performance. Columns packed with sub-2-mm particles can
lead either to the better separation of narrower chromato-
graphic peaks or to a faster analysis without the loss of
resolution, compared with conventional LC. The use of
elevated temperatures is another recent area of investigation
to improve the resolution of LC [58–60], whereas monolithic
columns constitute another alternative to improve the
chromatographic performance [53, 61, 62]. The typical 2-D
structure of LC-MS data is shown in Fig. 3.
3 Metabolomic data handling
3.1 Data handling
Metabolomic experiments usually produce large amounts of
data. Handling and analysing such complex data sets has a
great impact on the quality of the identification and
quantification of putative low-mass regulators, and therefore
to the resulting biological interpretation [63]. Data handling
can be further separated into data processing, data pre-
treatment and data analysis [64]. The distinct steps of the
strategy applied for knowledge discovery are shown in Fig. 4.
The appropriate procedures for data handling should take
the different sources of variation into account and should be
cautiously undertaken as the choice of these procedures
depends heavily on: (i) the analytical platform used to gener-
ate the data, (ii) the biological phenomenon under study, (iii)
the downstream data analysis method and (iv) the inherent
properties of the data (e.g. dimensionality) [65].
Figure 2. 2-D GC-MS [173].
Total Ion Chromatogram Single Mass Spectrum
Single Ion ChromatogramTotal Mass Spectrum
Inte
nsity
Figure 3. LC-MS data with the corresponding total ion chroma-togram and total mass spectrum. Single ion chromatogram andsingle mass spectrum are given as examples of the specificmonitoring of particular m/z or retention time.
J. Sep. Sci. 2010, 33, 1–15 Liquid Chromatography 3
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

3.2 Data pre-processing
3.2.1 Data files
Raw data are recorded as a single file corresponding to a
specific sample and hyphenated data files consist of a set of
mass spectra recorded in sequence. The processing of such
raw data therefore constitutes the initial step of data
handling, and its main goal is to extract the relevant
information and summarise the multiple files in a single
table [66]. Such data summary requires the characterisation
of each sample by the same number of variables, and
every variable must correspond to the same metabolite
measured in all of the samples in the data set.
This methodology includes several data-processing steps,
such as noise filtering, data binning, automatic peak
detection and chromatographic alignment, to allow the
appropriate comparison of multiple samples by statistical
methods.
3.2.2 Centroid data
A common method of data acquisition is to centroid the data
by combining the multiple m/z values corresponding to a
given peak into a single data point characterised by one
m/z value, the weighted centre of mass and its associated
intensity. The intensity is provided by the normalised
area of the peak calculated with the intensities
of the associated data points. The main peak of an isotopic
cluster can be identified and selected as a monoisotopic
peak instead of as profile data. Centroiding allows a
better mass assignment and usually increases the data
quality [67].
3.2.3 Resolution and data binning
Raw hyphenated data are usually binned in intervals of fixed
values to generate regularly sampled data. This step is
crucial for resolution tuning and allows the proper
representation and processing of the data. Ion intensities
are summed for a given m/z and retention time interval
[68]. The data are therefore converted into a 2-D array
with one axis describing the retention times values and the
other the m/z values. The values within the array are
naturally the measured intensities for a given m/z at a given
retention time. Despite a loss in m/z resolution, this data
representation allows for easier data handling and
treatment.
Ctrl
Experiment
Filtering
Feature Detection
Alignment
NormalisationDat
a P
repr
oces
sing
& P
retre
atm
ent
Dat
a A
naly
sis
Training
Algorithm Selection
Evaluation
Model Examination
Biological Interpretation
Scaling
A
B
A B A B
A B A B
Figure 4. Data mining and knowledgediscovery pipeline.
J. Sep. Sci. 2010, 33, 1–154 J. Boccard et al.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

3.2.4 Filtering data
The filtering process is used to suppress or at least reduce
random analytical noise or baseline drift in the data. Signal-
processing techniques are usually used, such as moving-
window filtering [69, 70], Savitzky–Golay filters [71] or
wavelet transforms [72]. During the baseline correction, the
shape of the baseline is identified and then subtracted from
the data.
3.2.5 Peak detection
Several strategies are possible to distinguish real signals
from noise, and the simplest one relies on an intensity
threshold, with the selection of all peaks detected above a
defined noise level. Either the peak area or the peak height
can be used as a quantitative measure of the real
concentration and can be further associated with the
corresponding m/z and retention time information. This
threshold can be applied independently in both the m/z and
the retention time directions using all intensity values along
one direction [71, 73]. Constraints on the peak shapes in the
chromatographic direction can refine the detection; data
points meeting this shape criterion are defined as peaks [74].
The second possibility is based on the examination of single
ion chromatograms, defined by a given m/z resolution. The
chromatograms are processed independently, either with a
threshold level or with a Gaussian-filter to detect the apex
and inflection points for the area integration [75, 76]. The
third approach depends on the application of a model-fitting
procedure to the data, which involves isotopes detection [77,
78]. The comparison of the peaks among different samples
constitutes another approach for peak detection but it
involves the proper alignment of the chromatograms. The
true peaks are consequently defined by an intensity above a
given threshold, and they must be present at least in a given
proportion of the samples. The signal-processing procedure
and the two main approaches for generating variables are
shown in Fig. 5.
3.2.6 Deconvolution
When soft ionisation methods are employed, a single peak
is expected for each compound. Fragments, adducts and
molecule isotopes can complicate the peak-detection
process, and an accurate detection procedure is needed.
Curve resolution or deconvolution algorithms use both the
spectral and the chromatographic profile resolution. Based
on the correlation of the profiles among the samples and the
retention time coincidence, deconvolution algorithms such
as the component detection algorithm [79, 80] attempt to
regroup ions coming from the same metabolite. In addition,
these ions are supposed to have related biological variations.
The use of these methods is however problematic when
applied to complex biological analyses such as metabolomic
data because numerous highly correlated signals can be
observed at similar retention times, leading to overlapping
peaks with close biological variations.
3.2.7 Alignment
There must be a one-to-one correspondence between the
variables being compared in order to accurately ascertain the
differences in the chemical composition between samples
from their chromatograms. Thus, one mandatory require-
ment is the proper alignment in both the m/z and the
Raw 2D data
Raw ion map
Filtered ion map
Peak Detection
Intensity
m/z
Time
Total Mass Spectra
Summing
Time
Peak Detection
Intensity
m/z
List of peaks
Figure 5. The first steps of data handlinginvolving signal processing and featuredetection with the two principal strategiesfor generating variables.
J. Sep. Sci. 2010, 33, 1–15 Liquid Chromatography 5
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

retention time dimensions in order to combine results.
Shifts along the m/z axis are usually easily corrected using
calibration, but changes in retention times are more
problematic. Progress in chromatographic techniques has
reduced these shifts but variability in the retention time of a
given metabolite is frequently encountered [81]. These drifts
can be caused by multiple factors, including temperature
changes, pH, pump pressure fluctuations or column
clogging. An alignment algorithm usually requires a
reference chromatogram to correct these retention time
differences, and its choice has a great influence on the
results. Additionally, numerous issues need to be tackled
when performing such corrections. For instance, the
method must preserve the chemical selectivity between
samples of distinct compositions while minimising run-to-
run shifts [82]. Moreover, corresponding peaks are expected
to be shifted by less than the representative distance
between adjacent peaks. On the other hand, an alignment
algorithm must be fast enough to deal quickly with a large
amount of data to supply a high sample throughput.
Three approaches are commonly utilised to align chro-
matograms [83]. The simplest solution resides in summing
or binning data along the chromatographic dimension. No
loss of information is required and the errors are pushed
back towards the bin boundaries. Indeed peaks can be
occasionally allocated to neighbouring m/z bins alternatively
[75].
Secondly, an alignment without peak detection can be
performed by compressing or stretching the retention time
axis of all samples to a common reference. As it needs only a
minimal manual contribution, this approach is interesting
from an automation point of view [84]. The mapping of total
ion chromatograms is one of the most investigated align-
ment methods, and warping methods are well-known
alternatives for that purpose, allowing the stretching and
shrinking of segments of the chromatograms [85]. Numer-
ous warping methods are available [86–88], but two algo-
rithms were initially developed, namely correlation
optimised warping [89] and dynamic time warping [90].
Unfortunately, such iterative alignment procedures are
considerably time consuming when dealing with the large
data sets of long chromatograms.
Another way to match the corresponding peaks among
samples relies on the detection of independent signals using
curve resolution. Several algorithms are available such as
progressive clustering [91], GENTLE [92] or MEND [93]. The
correspondence between the detected peaks of different
samples can then be assigned by the use of a time and m/ztolerance to regroup similar peaks. Another solution is to
match components with high-spectral similarity in a given
time window.
3.2.8 Normalisation
Prior to analysis, data are often pre-treated to remove
systematic variations between samples [64]. Normalisation
is a critical step, because it aims to remove bias in the data
while preserving biological information. It is a difficult task
mainly due to the great chemical diversity of complex data,
leading to discrepancies in the extraction or ionisation
efficiencies. Statistical models represent simple methods to
estimate the scaling factors among samples, such as the
unit norm [94] or median intensities [71]. The addition of a
set of internal or external standards that reasonably cover
the retention time range constitutes another way to perform
normalisation in a practical manner [95–97].
3.2.9 Software packages
The need of powerful data-processing methods gave rise to
numerous commercial or free tools, implementing either
one specific part or combining several steps of the data-
processing pipeline. Most of the tools include standard
procedures of noise filtering, peak detection, alignment and
normalisation, while they output a data matrix containing
the peak intensities or areas for each detected ion among all
the samples.
Integrated platforms for hyphenated MS data pre-
processing and analysis include free packages such as
MZmine [74], MAthDAMP [98], metAlign [99], MSFacts
[100], XCMS [75] and MeltDB [101] or commercial solutions
as, for example, MS Resolver [102], MarkerLynx [103] or
Sieve based on the ChromAlign algorithm [104]. Moreover,
comparisons of performance have been published recently
[105, 106].
3.3 Metabolomic data pre-treatment
The data pre-treatment phase constitutes another crucial
step that can drastically change the pertinence and the
outcome of the data analysis. Centring aims to estimate the
model parameters with an observed bias. Mean centring
across the first mode is a classical pre-processing practice
prior to model fitting. It provides simpler models by
reducing the rank, increasing the fit to the data, removing
offsets and avoiding algorithmic problems [107]. The
fluctuations of the variables are centred around zero instead
of their mean value to focus on the variation between
samples. Scaling is used to adjust the importance allocated
to the elements of the data in fitting the model. Most scaling
methods adjust the weight of each variable with a scaling
factor that can be estimated by either a dispersion criterion
or a size measure. It may be obtained by calculating the
inverse of its standard deviation so that all variables have the
same chance to contribute to the model as they have an
equal unit variance (UV). This technique is called UV
scaling, or auto-scaling when prior mean centring is
performed [108]. Further analyses are then based on
correlations instead of covariances. UV scaling is often
suitable when no prior information is available but equal
weights are allocated to the baseline noise and proper
signals. Pareto scaling involves a similar procedure based on
the dispersion except that the square root of the standard
J. Sep. Sci. 2010, 33, 1–156 J. Boccard et al.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

deviation is used as scaling factor [109]. It is an intermediate
situation as it gives a variance equal to the standard
deviation of each variable instead of UV. Large fold changes
have less influence than when raw data are used, but the
reasonable weight allocated to small values can avoid the
detrimental effects caused by a similar error increase. Other
methods such as variable stability scaling [110] or range
scaling [111] are available, but UV and Pareto scaling are the
prevailing methods applied in metabolomic MS-based
studies.
Furthermore, as many statistical procedures are based
on the assumption of normally distributed values, a trans-
formation can be desirable to address skewed data. Log or
power transformation are well-known functions generally
applied to correct heteroscedasticity [112]. The log function
is useful when the interactions between variables are not
only additive but also multiplicative and its use allows the fit
of linear models [113].
4 Data analysis
4.1 Modelling metabolomic data
Metabolomic experiments exploit computerised instru-
ments leading to the simultaneous detection of a great
number of variables, and the generation of large multi-
variate data sets [114]. From a statistical point of view,
metabolomic data sets constitute a great challenge, as
reliable and robust approaches are necessary to handle and
extract the relevant information from the vast amount of
data generated [115, 116]. The outputs of a metabolomic
data analysis may differ greatly depending on the purposes
of the investigation. Although very few highly reliable
metabolites can be sufficient for diagnostic purposes (as in
the case of disease evaluation), an extended set of
compounds may be desired when a biochemical network
is under examination. Alternatively, an overview of the
distribution of samples can be desirable to assess a data set
globally. The choice of the data analysis strategy depends
heavily on the questions that are asked. Changes in
metabolite levels may be drastic or subtle, and statistical
processing is required to determine the relevance of an
observed change. The choice of the most appropriate model
for a given data set constitutes an important issue. The
structural descriptions and the explicit knowledge that are
obtained by examining the model are more important than
the predictive ability of new samples in many applications.
The description of the underlying concept has therefore to
be both operational and intelligible. The methods are
expected to constitute high-throughput and potent auto-
mated tools to highlight trends and patterns among samples
or relevant biomarkers within large metabolomic data sets.
Metabolomic commercial solutions include built-in
statistical packages for data analysis. Common chemometric
tools [117, 118] such as principal component analysis (PCA)
[119] are generally proposed for display and exploratory
analysis purposes, whereas univariate statistical tests such
as the Student’s t-test are used to identify the relevant
variables. These methods are useful by providing an over-
view of the pre-processed data but are rather limited [120].
There is, therefore, a considerable opportunity to increase
the understanding of biological phenomena through the use
of data mining methods with respect to their strengths and
applicability.
4.2 Univariate hypothesis testing
Univariate significance tests such as the Student’s t-test, the
one-way analysis of variance or the non-parametric equiva-
lents can be used to identify statistical differences between
samples of distinct classes [121]. The predictive power of
each variable is assessed by finding statistically significant
differences between the mean intensity values of a given
signal, and the calculated p-value is a straightforward
indicator. Such procedures are easily understandable but
their use is rather limited when dealing with thousands of
highly correlated variables. False positives (type I error) are
likely to occur when performing multiple comparisons and
procedures such as the Bonferroni correction have been
introduced to address this issue [122]. The vast majority of
metabolomics-dedicated software provides statistical
hypothesis testing [99, 123, 124].
4.3 Exploratory analysis by unsupervised learning
Unsupervised methods attempt to analyse a set of observa-
tions without measuring or observing any related outcome.
As there is no specified class label or response, the data set
is considered as a collection of analogous objects. Unsu-
pervised learning uses procedures that attempt to find the
natural partitions of patterns to facilitate the understanding
of the relationship between the samples and to highlight the
variables that are responsible for these relationships. By
providing means for visualisation, unsupervised learning
aids in the discovery of unknown but meaningful categories
of samples or variables that naturally fall together. The
success of such approaches is frequently evaluated subjec-
tively by the interpretability and usefulness of the results
with respect to a given problem.
4.3.1 PCA
PCA can be considered as the starting point of multivariate
data analyses. PCA is an orthogonal transformation of
multivariate data first formulated by Pearson [125] mostly
used for exploratory analyses by extracting and displaying
systematic variations. PCA attempts to uncover hidden
internal structures by building principal components
describing the maximal variance of the data [119]. This
method represents a very useful tool for display purposes as
it provides a low-dimension projection of the data, i.e. a
J. Sep. Sci. 2010, 33, 1–15 Liquid Chromatography 7
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

window into the original K-dimensional space, by a
transformation into a new coordinate system. The basic
concept relies on areas of signal variance in the data where
underlying phenomena can be observed. This principle
leads to a focus on a small number of uncorrelated
independent signals that explain a large part of the total
variance in a compact and insightful manner. In practice,
PCA builds hyperplanes in the original feature space that
are linear combinations of the original variables and
describes the data in a least squares view. The inspection
of PCA scores and loading plots highlights the relationships
among the distribution of samples that may reveal group-
ings, trends or outliers and the corresponding variables.
Moreover, more effective data analyses can be performed on
the reduced dimensional space, such as clustering, pattern
recognition or classification. The vast majority of metabo-
lomic studies involve PCA as a first exploratory step [102,
126–131].
4.3.2 Hierarchical cluster analysis
When there is no class or response to be predicted, cluster
analysis can help to partition a data set into natural groups.
It intends to group together observations that are similar to
one another by producing a hierarchical structure dividing
the instance space. These subsets may reveal underlying
patterns in the whole data structure, highlighting inner
mechanisms of a phenomenon. There is a wide variety of
clustering methods, but hierarchical cluster analysis (HCA)
constitutes a straightforward and general approach when
the number of clusters is unknown a priori. It can either be
agglomerative (bottom-up, i.e. iteratively grouping objects)
or divisive (top-down, i.e. dividing a data set), but in practice,
the agglomerative methods are of wider use. A key part of
HCA is the calculation of similarity measures between
observations, and between clusters once observations begin
to be grouped into clusters. This similarity is estimated by
both a set of distances and a linkage function. The distances
are computed using a specific metric, such as Euclidean,
correlation or Manhattan. The linkage function defines the
criteria chosen to compute the distance between objects. In
all cases, the pair of clusters with highest cluster-to-cluster
similarity is iteratively fused into a single cluster. The
outcome is represented graphically as a dendrogram, i.e. a
tree where each leaf corresponds to an object and the
branching depicts the hierarchy of clusters [132]. Several
applications of HCA have been used for the clustering of
metabolomic data [133, 134]. The learning principles of PCA
and HCA are shown in Fig. 6.
4.4 Classification by supervised learning
Supervised learning considers each object with respect to an
observed response and includes regression and classification
problems depending on the output type under consideration,
i.e. a numerical value in the first case and a class label in the
second. Classification thus aims to produce general hypoth-
eses based on a training set of examples that are described by
several variables and identified by known labels corresponding
to the class information. The task is to learn the mapping
from the former to the latter and numerous techniques, based
either on statistics or on artificial intelligence, have been
developed for that purpose.
4.4.1 Partial least squares projection to latent struc-
tures
Projection to latent structures by means of partial least
squares (PLS) [135, 136] is a well-established regression-
based method [136]. This technique is particularly adapted
to situations where fewer observations (i.e. number of
different observed objects, N) than measured variables (e.g.m/z, detected features, K) are available. Its use has become
very popular, thanks to its ability to deal with many
correlated and noisy variables forming megavariate data
structures (K�N) [137, 138]. PLS builds a low dimensional
sub-space based on linear combinations of the original X-
variables and makes use of the additional Y information by
adjusting the model to capture the Y-related variation in X.
A PLS-based classification therefore has the property that it
builds data structures with an intrinsic prediction power, by
maximising the covariance between the data and the class
assignment. The decomposition relies on latent variables
that are computed sequentially to provide a good correlation
with the remaining unexplained fraction of Y. In the context
of classification, PLS-DA is performed to sharpen the
partition between groups of observations, such that a
maximum separation among classes is obtained. The model
Principal Components Analysis
Hierarchical Cluster Analysis
Obj
ects
X1 X2 X3
X1 X2 X3
X2
X1
X3
t1
t2
t2
t1
Scores (objects)
Loadings (variables)
p2
p1
X1
X3X2
Obj
ects
X2
X1
X3
Figure 6. PCA and HCA learning principles.
J. Sep. Sci. 2010, 33, 1–158 J. Boccard et al.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

can then be analysed to understand which variables carry
the class-separating information [139]. PLS-DA was demon-
strated as a potent tool for the classification of metabolomic
data [65]. Orthogonal PLS analysis [140] and O2-PLS [141]
are recent extensions of the PLS method and several recent
studies applied these techniques to metabolomic data [126].
4.4.2 Decision trees
Decision trees are logic-based classification algorithms that
are able to sort the training examples by class, using feature
values to build a tree based on a divide-and-conquer strategy.
The root node constitutes the starting point, and the
instances are classified sequentially down the tree according
to the values of attributes tested at each node. Decision trees
build models that are based on a hierarchy of test and are
consequently usually straightforward to interpret. The
assumption made in such a method is that at least one
feature shows dissimilar values among different classes.
Features can be ranked by their capacity to divide the
remaining data subset at a given node. The best (i.e. the
most informative) feature can then be selected, and the
procedure is repeated recursively with each branch delimit-
ing a partition of the data, producing sub-trees from the
splits. Decision trees are intrinsically univariate methods,
because each node corresponds to a single feature splitting
the data set. The most popular univariate decision tree
algorithm is C4.5 [142]. Multivariate alternatives have,
however, been proposed, such as CART [143] or alternatives
relying either on logical operators [144] or on constructive
induction [145]. The random forest constitutes another
approach to build such classifiers, as it is a collection of
individual decision trees obtained by a bagging procedure
(bootstrap aggregating). All of the trees are applied in
parallel to build a consensus predicted class as the most
frequent output produced by the independent trees [146].
4.4.3 Artificial neural networks
Artificial neural networks (ANN) are well-established
methods that are able to build models by the use of levels
of units, called neurons, linked together by a network of
connections of adjustable weights in a hierarchical structure
[147]. The neural networks are thus named due to the
similarity they share with biological neurons, which receive
an input that is a weighted sum of the outputs of other
elements. The signal starts from the input units, propagates
through the connections and determines activation values
when the output units are reached. Connection weights are
first set to random values and then adjusted to progress
slowly in the direction of the correct output. The interpret-
ability of ANNs remains a critical issue because their
outputs are usually not easily comprehensible. The most
popular ANN algorithms include the multilayer perceptron
[148] and radial basis functions [149]. The learning
principles of the PLS regression, the decision trees and
the multilayer perceptron are shown in Fig. 7.
4.4.4 Probabilistic learning algorithms
Probabilistic algorithms rely on an explicit probability model
by allocating a probability to each class during the training
phase. The predicted class label then corresponds to the
class with the greatest probability. Bayesian networks
constitute well-known methods of statistical learning, and
their most simple version is the Naıve Bayes classifier.
Despite being very simple and assuming the almost always
wrong principles of independence between variables, Naıve
Bayes often performs well in the presence of complex
problems, making it useful in practice [150].
4.4.5 Instance-based learning algorithms
Instance-based learning algorithms can be considered
the simplest form of learning [151]. The nearest neighbour
classification method is one of the most popular and
straightforward approaches. Its k-Nearest Neighbour
implementation relies on the regrouping of instances with
similar properties in the high-dimensional space of the
data and a majority vote. Rather than inferring classification
rules, the learned concept is encapsulated in the training
set without explicitly illustrating the data patterns. Despite
multiple possibilities, most instance-based classifiers use
Euclidean distances as a good compromise to balance
large and small differences, while weighting schemes
have been developed to balance the influence of either the
set of attributes or the neighbouring instances [152]. Only a
few applications of the k-Nearest Neighbour approach
have been presented for the classification of metabonomic
data [134].
Multilayer PerceptronDecision tree
PLS Regression
X1>θ1
X3>θ3X2≤θ2
X4≤θ4Input units
Output units
Hidden units
wij wjk
X space Y spacePoint i
Projection
1st
LatentVariable
i
i
A B C D E
Figure 7. PLS regression, decision trees and multilayer percep-tron learning principles.
J. Sep. Sci. 2010, 33, 1–15 Liquid Chromatography 9
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

4.4.6 Support vector machines
Support vector machines (SVMs) are recent and increas-
ingly popular classification methods with a great ability for
generalisation [153, 154]. These algorithms possess several
interesting qualities as they are generally robust to noisy
data or to the presence of outliers and they can be applied to
non-linear classification problems by making use of a kernel
function. They rely on the selection of a small number of
critical boundary instances called support vectors. SVMs
were originally developed by Vapnik [155] and they rely on a
projection of the data in a higher-dimensional space, i.e. the
feature space. This transformation is properly selected to
provide a separation of the training set classes, and a linear
separation in the feature space can represent a non-linear
decision boundary in the original data space. Kernel
functions are used to map new points in the feature space
for classification, and hence their choice is critical.
Commonly used kernel functions include the linear,
polynomial and radial basis function kernels. A hyperplane
separating two classes with a tolerance value expressed as a
margin on both sides of this limit is then built in the feature
space. The algorithm attempts to maximise the value of this
geometrical margin separating the hyperplane and the
instances on both sides. Data points positioned on this
margin are called the support vectors, and the solution is
computed as a linear combination of these points. If no
solution can be found, as when the classes are not linearly
separable, a soft margin can be used to introduce a tolerance
and accept errors of classification. Although SVMs
were originally conceived to deal with binary classification
problems, several algorithms exist to resolve multi-
class cases [154]. Thanks to promising results [156], SVMs
will probably be increasingly applied in the field of
metabolomics [157]. The learning principles of the
SVM and the nearest neighbours algorithms are shown in
Fig. 8.
4.5 Variable selection
4.5.1 Dimensionality reduction
The number of features is a key aspect that determines
the size of the hypothesis space of the data [158].
Augmenting the former increases the latter exponentially
and correct hypotheses then become harder to find. The
number of variables necessary to build a model depends
heavily on the algorithm under consideration, ranging from
the inclusion of all features (e.g. nearest neighbours
classifiers) to very few variables (e.g. decision trees) [159].
In practice, the performance of most classifiers can be
improved by a prior variable selection, as their accuracy is
often negatively influenced by a high number of correlated
or irrelevant variables. This dimensionality reduction is
therefore often very useful when mining large metabolomic
data sets, and the selection of a subset of representative
features that retain the salient characteristics of the data is
therefore a fundamental issue. Moreover, circumventing the
inclusion of irrelevant attributes having negative effects
provides a more compact and interpretable image of a
phenomenon, together with reduced computing time
requirements.
4.5.2 Algorithmic aspects
Although both supervised and unsupervised approaches are
available, attribute selection procedures usually combine a
search algorithm that generates the subsets of variables and
an estimation method that detects the most appropriate
combination of variables using some quality index. This
indicator measures either the predictive or the clustering
ability of the variables, depending on whether the selection
is supervised or not. Moreover, the selection method can
either take each variable individually or a group of features
+
Nearest NeighboursX1 X2 X3
+ ?
? K=3K=5K=10
Support Vector Machines
Φ
Margin
Hyper-plane Figure 8. SVM and nearest neighbours algo-rithms learning principles.
J. Sep. Sci. 2010, 33, 1–1510 J. Boccard et al.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

into consideration. Due to its nature, individual feature
evaluation is unable to detect feature interactions. Moreover,
a totally uninformative variable can significantly enhance
the predictive ability in the context of other variables.
Two main approaches are usually proposed, the filter
and the wrapper methodologies [160]. The filter selection
relies generally on a pre-processing evaluation system that is
independent from the learning phase. A quality index can
be measured for individual or groups of attributes, and
although it can be derived from a classifier, it does not
directly rely on the prediction accuracy. Among several
methods, feature ranking represents a simple supervised
approach to filter the most valuable variables. Several
ranking criteria are available to find variables that discri-
minate between the observed classes, including the analysis
of variance [161] and Recursive Elimination of Features [162,
163]. On the other hand, algorithms that build linear models
provide rankings by comparing the coefficients of features,
such as orthogonal signal correction [164]. Other methods
use different indicators, such as the information gain [165]
based on entropy, to quantify the information content of a
variable. Such rankings imply, however, the determination
of a suitable number of features to select. Methods that
eliminate redundant variables by allocating a goodness
measure to subsets of the attributes are able to take the
inter-correlations into account [166]. The correlation issue
can be addressed either by selecting only one of the corre-
lated variables within a group, such as correlation-based
feature selection, or by building new features starting from
the original correlated variables (feature construction/
transformation).
On the other hand, wrapper selection relies on the
predictive accuracy of a learning algorithm as a quality index
of a features subset, e.g. SVM-based feature selection. As the
assortment giving raise to the best performance is kept,
the selection is an integrated part of the learning phase and
the number of features can be automatically determined.
The wrapper model is computationally very expensive when
dealing with a large number of features and the selection is
evidently oriented by the choice of the classifier used as a
black box [167]. Finally, by combining complementary filter
selection and learning algorithms relying on distinct learn-
ing principles, performance can be improved by the
strengths of both methods.
4.6 Algorithms comparison
Selecting a particular classification algorithm from the wide
variety of available methods constitutes a critical operation.
Despite numerous measures of performance, such as
sensitivity, specificity and the Kappa coefficient [168, 169],
the performance of a classifier is most often based on its
prediction accuracy, i.e. the percentage of correctly predicted
examples over the total number of predictions. However,
this single value is not sufficiently informative when dealing
with a limited number of observations, as this estimate will
be completely optimistic. Several schemes are available for
the robust evaluation of this accuracy to avoid overfitting,
but the most popular include the repeated cross-validation
and the prediction of an independent test set. By evaluating
the accuracies of multiple algorithms using cross-validation,
each experiment provides an independent error estimate
[170]. Moreover, distinct costs of misclassification errors can
be defined to avoid a given type of error, e.g. false positives
or negatives. These results are then used to determine
whether the average accuracy value of a particular algorithm
is significantly greater or lower than another by performing
a Student’s t-test for statistical comparison [171]. The sub-
sampling of the whole data set to generate subsets, however,
is in contradiction with the independency statement
required by hypothesis testing, and a test should not
depend on the partitioning of the data set. Several heuristic
solutions have been proposed to counteract this difficulty,
such as the corrected resampled t-test [172]. Repeated
random partitioning of ten folds is commonly used to
evaluate the prediction accuracies by examining their
replicability.
5 Concluding remarks
Deciphering complex biological systems and processes
requires detailed descriptions of the systems under study
as well as data mining techniques suitable to handle large
amounts of information. As continued data accumulation is
inevitable, finding automatically the valuable but hidden
information in metabolomic data of high dimensionality is
already a crucial issue. Data mining, therefore, has a central
role to play by making sense of data and inferring the
structures governing biological phenomena. Indeed, these
methods will have more and more opportunities to prove
their worth when dealing with massive data sets and may
lead to fruitful results by helping to decode complex
phenomena. However, the relevant answers will not come
from computer programs but from the people working with
the data and understanding its related characteristics and
issues.
On the other hand, an increasing number of metabo-
lomic studies rely on multiple analytical platforms
combining GC-, LC-, CE-MS or NMR data. Merging the
information from data of different natures and structures
and extracting the common traits will undoubtedly consti-
tute a key issue in the perspective of a more comprehensive
vision of metabolomics. In addition, the merging with data
from other Omic technologies, such as transcriptomics and
proteomics will probably offer a deeper insight into the
regulatory networks in systems biology and will definitely
represent another great challenge. Modelling all types
of regulatory events occurring within a cell in a holistic
manner is expected to provide a global view of the
system.
The authors have declared no conflict of interest.
J. Sep. Sci. 2010, 33, 1–15 Liquid Chromatography 11
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

6 References
[1] Nicholson, J. K., Lindon, J. C., Holmes, E., Xenobiotica1999, 29, 1181–1189.
[2] Sumner, L. W., Mendes, P., Dixon, R. A., Phytochem-istry 2003, 62, 817–836.
[3] Hollywood, K., Brison, D. R., Goodacre, R., Proteomics2006, 6, 4716–4723.
[4] Oksman-Caldentey, K. M., Saito, K., Curr. Opin.Biotechnol. 2005, 16, 174–179.
[5] Oresic, M., Clish, C. B., Davidov, E. J., Verheij, E.,Vogels, J., Havekes, L. M., Neumann, E., Adourian, A.,Naylor, S., van der Greef, J., Plasterer, T., Appl.Bioinformatics 2004, 3, 205–217.
[6] Keun, H. C., Pharmacol. Ther. 2006, 109, 92–106.
[7] Plumb, R. S., Dear, G. J., Mallett, D. N., Higton, D. M.,Pleasance, S., Biddlecombe, R. A., Xenobiotica 2001,31, 599–617.
[8] Forster, J., Famili, I., Fu, P., Pjzalsson, B. O., Nielsen, J.,Genome Res. 2003, 13, 244–253.
[9] Ackermann, B. L., Hale, J. E., Duffin, K. L., Curr. DrugMetab. 2006, 7, 525–539.
[10] van der Greef, J., Martin, S., Juhasz, P., Adourian, A.,Plasterer, T., Verheij, E. R., McBurney, R. N.,J. Proteome Res. 2007, 6, 1540–1559.
[11] Sreekumar, A., Poisson, L. M., Rajendiran, T. M., Khan,A. P., Cao, Q., Yu, J. D., Laxman, B., Mehra, R., Lonigro,R. J., Li, Y., Nyati, M. K., Ahsan, A., Kalyana-Sundaram,S., Han, B., Cao, X. H., Byun, J., Omenn, G. S., Ghosh,D., Pennathur, S., Alexander, D. C., Berger, A., Shuster,J. R., Wei, J. T., Varambally, S., Beecher, C., Chinnai-yan, A. M., Nature 2009, 457, 910–914.
[12] Lindon, J. C., Nicholson, J. K., Holmes, E., Antti, H.,Bollard, M. E., Keun, H., Beckonert, O., Ebbels, T. M.,Reilly, M. D., Robertson, D., Stevens, G. J., Luke, P.,Breau, A. P., Cantor, G. H., Bible, R. H., Niederhauser,U., Senn, H., Schlotterbeck, G., Sidelmann, U. G.,Laursen, S. M., Tymiak, A., Car, B. D., Lehman-McKeeman, L., Colet, J. M., Loukaci, A., Thomas, C.,Toxicol. Appl. Pharmacol. 2003, 187, 137–146.
[13] Dieterle, F., Schlotterbeck, G. T., Ross, A., Niederhauser,U., Senn, H., Chem. Res. Toxicol. 2006, 19, 1175–1181.
[14] Wishart, D. S., Trends Food Sci. Technol. 2008, 19,482–493.
[15] Weckwerth, W., Annu. Rev. Plant. Biol. 2003, 54,669–689.
[16] Fiehn, O., Plant Mol. Biol. 2002, 48, 155–171.
[17] Atherton, H. J., Bailey, N. J., Zhang, W., Taylor, J.,Major, H., Shockcor, J., Clarke, K., Griffin, J. L., Physiol.Genomics 2006, 27, 178–186.
[18] Dunn, W. B., Ellis, D., Trends Anal. Chem. 2005, 24,285–294.
[19] Jonsson, P., Gullberg, J., Nordstrom, A., Kusano, M.,Kowalczyk, M., Sjostrom, M., Moritz, T., Anal. Chem.2004, 76, 1738–1745.
[20] Sinha, A. E., Hope, J. L., Prazen, B. J., Nilsson, E. J.,Jack, R. M., Synovec, R. E., J. Chromatogr. A 2004,1058, 209–215.
[21] Veriotti, T., Sacks, R., Anal. Chem. 2001, 73, 4395–4402.
[22] Wagner, C., Sefkow, M., Kopka, J., Phytochemistry2003, 62, 887–900.
[23] Kopka, J., J. Biotechnol. 2006, 124, 312–322.
[24] Huhman, D. V., Sumner, L. W., Phytochemistry 2002,59, 347–360.
[25] Idborg, H., Zamani, L., Edlund, P. O., Schuppe-Koisti-nen, I., Jacobsson, S. P., J. Chromatogr. B 2005, 828,9–13.
[26] Wolfender, J. L., Terreaux, C., Hostettmann, K., Pharm.Biol. 2000, 38, 41–54.
[27] Ramautar, R., Demirci, A., de Jong, G. J., Trends Anal.Chem. 2006, 25, 455–466.
[28] Garcia-Perez, I., Vallejo, M., Garcia, A., Legido-Quigley,C., Barbas, C., J. Chromatogr. A 2008, 1204, 130–139.
[29] Soga, T., Ohashi, Y., Ueno, Y., Naraoka, H., Tomita, M.,Nishioka, T., J. Proteome Res. 2003, 2, 488–494.
[30] von Roepenack-Lahaye, E., Degenkolb, T., Zerjeski, M.,Franz, M., Roth, U., Wessjohann, L., Schmidt, J., Scheel,D., Clemens, S., Plant Physiol. 2004, 134, 548–559.
[31] Krishnan, P., Kruger, N. J., Ratcliffe, R. G., J. Exp. Bot.2005, 56, 255–265.
[32] Wolfender, J. L., Ndjoko, K., Hostettmann, K., Phyto-chem. Anal. 2001, 12, 2–22.
[33] Brown, S. C., Kruppa, G., Dasseux, J. L., Mass Spec-trom. Rev. 2005, 24, 223–231.
[34] Brown, K., Tompkins, E. M., White, I. N. H., MassSpectrom. Rev. 2006, 25, 127–145.
[35] Brown, M., Dunn, W. B., Ellis, D. I., Goodacre, R.,Handl, J., Knowles, J. D., O’Hagan, S., Spasic, I., Kell,D. B., Metabolomics 2005, 1, 39–51.
[36] van der Greef, J., van der Heijden, R., Verheij, E. R.,Adv. Mass Spectrom. 2004, 16, 145–165.
[37] Werner, E., Heilier, J. F., Ducruix, C., Ezan, E., Junot, C.,Tabet, J. C., J. Chromatogr. B 2008, 871, 143–163.
[38] Dunn, W. B., Phys. Biol. 2008, 5, 11001.
[39] Taylor, A. J., Linforth, R. S. T., Int. J. Spectrom. 2003,223, 179–191.
[40] van der Greef, J., Niessen, W. M. A., Int. J. MassSpectrom. Ion Process. 1992, 118, 857–873.
[41] Villas-Boas, S. G., Mas, S., Akesson, M., Smedsgaard,J., Nielsen, J., Mass Spectrom. Rev. 2005, 24,613–646.
[42] Robards, K., Haddad, P. R., Jackson, P. E., Principlesand Practice of Modern Chromatographic Methods,Academic Press, London 1996.
[43] Roessner, U., Wagner, C., Kopka, J., Trethewey, R. N.,Willmitzer, L., Plant J. 2000, 23, 131–142.
[44] Halket, J. M., Zaikin, V. G., Eur. J. Mass Spectrom.2003, 9, 1–21.
[45] Halket, J. M., Waterman, D., Przyborowska, A. M.,Patel, R. K. P., Fraser, P. D., Bramley, P. M., J. Exp. Bot.2005, 56, 219–243.
[46] Zaikin, V. G., Halket, J. M., Eur. J. Mass Spectrom.2003, 9, 421–434.
[47] Horning, E. C., Clin. Chem. 1968, 14, 777.
[48] Pauling, L., Robinson, A. B., Teranish, R., Cary, P., Proc.Natl. Acad. Sci. USA 1971, 68, 2374–2376.
J. Sep. Sci. 2010, 33, 1–1512 J. Boccard et al.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

[49] Schauer, N., Steinhauser, D., Strelkov, S., Schomburg,D., Allison, G., Moritz, T., Lundgren, K., Roessner-Tunali, U., Forbes, M. G., Willmitzer, L., Fernie, A. R.,Kopka, J., FEBS Lett. 2005, 579, 1332–1337.
[50] Beens, J., Brinkman, U. A. T., Analyst 2005, 130, 123–127.
[51] Idborg, H., Edlund, P. O., Jacobsson, S. P., RapidCommun. Mass. Spectrom. 2004, 18, 944–954.
[52] Niessen, W. M. A., J. Chromatogr. A 1999, 856,179–197.
[53] Tolstikov, V. V., Lommen, A., Nakanishi, K., Tanaka, N.,Fiehn, O., Anal. Chem. 2003, 75, 6737–6740.
[54] Wang, C., Kong, H. W., Guan, Y. F., Yang, J., Gu, J. R.,Yang, S. L., Xu, G. W., Anal. Chem. 2005, 77,4108–4116.
[55] Ding, J., Sorensen, C. M., Zhang, Q. B., Jiang, H. L.,Jaitly, N., Livesay, E. A., Shen, Y. F., Smith, R. D., Metz,T. O., Anal. Chem. 2007, 79, 6081–6093.
[56] Dear, G. J., Ayrton, J., Plumb, R., Fraser, I. J., RapidCommun. Mass. Spectrom. 1999, 13, 456–463.
[57] Swartz, M. E., J. Liquid Chromatogr. Rel. Technol.2005, 28, 1253–1263.
[58] Plumb, R. S., Rainville, P., Smith, B. W., Johnson, K. A.,Castro-Perez, J., Wilson, I. D., Nicholson, J. K., Anal.Chem. 2006, 78, 7278–7283.
[59] Stoll, D. R., Cohen, J. D., Carr, P. W., J. Chromatogr.A 2006, 1122, 123–137.
[60] Grata, E., Boccard, J., Guillarme, D., Glauser, G.,Carrupt, P. A., Farmer, E. E., Wolfender, J. L., Rudaz, S.,J. Chromatogr. B Analyt. Technol. Biomed. Life Sci.2008, 871, 261–270.
[61] Maruska, A., Kornysova, O., J. Chromatogr. A 2006,1112, 319–330.
[62] Pham-Tuan, H., Kaskavelis, L., Daykin, C. A., Janssen,H. G., J. Chromatogr. B 2003, 789, 283–301.
[63] Hendriks, M. M. W. B., Cruz-Juarez, L., De Bont, D.,Hall, R., Anal. Chim. Acta 2005, 545, 53–64.
[64] van den Berg, R. A., Hoefsloot, H. C. J., Westerhuis,J. A., Smilde, A. K., van der Werf, M. J., Biomed.Chromatogr. Genomics 2006, 7.
[65] Jonsson, P., Bruce, S. J., Moritz, T., Trygg, J., Sjoes-troem, M., Plumb, R., Granger, J., Maibaum, E.,Nicholson, J. K., Holmes, E., Antti, H., Analyst 2005,130, 701–707.
[66] Hilario, M., Kalousis, A., Pellegrini, C., Muller, M., MassSpectrom. Rev. 2006, 25, 409–449.
[67] Katajamaa, M., Oresic, M., J. Chromatogr. A 2007,1158, 318–328.
[68] Davis, R. A., Charlton, A. J., Godward, J., Jones, S. A.,Harrison, M., Wilson, J. C., Chemometric Intell. Lab.Syst. 2007, 85, 144–154.
[69] Fleming, C. M., Kowalski, B. R., Apffel, A., Hancock,W. S., J. Chromatogr. A 1999, 849, 71–85.
[70] Radulovic, D., Jelveh, S., Ryu, S., Hamilton, T. G., Foss,E., Mao, Y., Emili, A., Mol. Cell. Proteom. 2004, 3,984–997.
[71] Wang, W., Zhou, H., Lin, H., Roy, S., Shaler, T. A., Hill,L. R., Norton, S., Kumar, P., Anderle, M., Becker, C. H.,Anal. Chem. 2003, 75, 4818–4826.
[72] Li, X. J., Yi, E. C., Kemp, C. J., Zhang, H., Aebersold, R.,Mol. Cell. Proteom. 2005, 4, 1328–1340.
[73] Hastings, C. A., Norton, S. M., Roy, S., Rapid Commun.Mass. Spectrom. 2002, 16, 462–467.
[74] Katajamaa, Mi., Oresic, M., Biomed. Chromatogr.Bioinformatics 2005, 6, 179.
[75] Smith, C. A., Want, E. J., O’Maille, G., Abagyan, R.,Siuzdak, G., Anal. Chem. 2006, 78, 779–787.
[76] Eanes, R. C., Marcus, R. K., Spectrochim. Acta B. At.Spectrosc. 2000, 55B, 403–428.
[77] Hermansson, M., Uphoff, A., Kakela, R., Somerharju,P., Anal. Chem. 2005, 77, 2166–2175.
[78] Leptos, K. C., Sarracino, D. A., Jaffe, J. D., Krastins, B.,Church, G. M., Proteomics 2006, 6, 1770–1782.
[79] Windig, W., Phalp, J. M., Payne, A. W., Anal. Chem.1996, 68, 3602–3606.
[80] Windig, W., Smith, W. F., J. Chromatogr. A 2007, 1158,251–257.
[81] Gika, H. G., Macpherson, E., Theodoridis, G. A.,Wilson, I. D., J. Chromatogr. B 2008, 871, 299–305.
[82] Lange, E., Tautenhahn, R., Neumann, S., Gropl, C.,Biomed Chromatogr. Bioinformatics 2008, 9, 375.
[83] Nordstrom, A., O’Maille, G., Qin, C., Siuzdak, G., Anal.Chem. 2006, 78, 3289–3295.
[84] Johnson, K. J., Wright, B. W., Jarman, K. H., Synovec,R. E., J. Chromatogr. A 2003, 996, 141–155.
[85] Wang, C. P., Isenhour, T. L., Anal. Chem. 1987, 59,649–654.
[86] Bylund, D., Danielsson, R., Malmquist, G., Markides,K. E., J. Chromatogr. A 2002, 961, 237–244.
[87] Tomasi, G., van den Berg, F., Andersson, C.,J. Chemometrics 2004, 18, 231–241.
[88] Prince, J. T., Marcotte, E. M., Anal. Chem. 2006, 78,6140–6152.
[89] Nielsen, N. P. V., Carstensen, J. M., Smedsgaard, J.,J. Chromatogr. A 1998, 805, 17–35.
[90] Pravdova, V., Walczak, B., Massart, D. L., Anal. Chim.Acta 2002, 456, 77–92.
[91] De Souza, D. P., Saunders, E. C., McConville, M. J.,Likic, V. A., Bioinformatics 2006, 22, 1391–1396.
[92] Shen, H. L., Grung, B., Kvalheim, O. M., Eide, I., Anal.Chim. Acta 2001, 446, 313–328.
[93] Andreev, V. P., Rejtar, T., Chen, H. S., Moskovets, E. V.,Ivanov, A. R., Karger, B. L., Anal. Chem. 2003, 75,6314–6326.
[94] Scholz, M., Gatzek, S., Sterling, A., Fiehn, O., Selbig, J.,Bioinformatics 2004, 20, 2447–2454.
[95] Bijlsma, S., Bobeldijk, L., Verheij, E. R., Ramaker, R.,Kochhar, S., Macdonald, I. A., van Ommen, B., Smilde,A. K., Anal. Chem. 2006, 78, 567–574.
[96] Wu, L., Mashego, M. R., van Dam, J. C., Proell, A. M.,Vinke, J. L., Ras, C., van Winden, W. A., van Gulik,W. M., Heijnen, J. J., Anal. Biochem. 2005, 336,164–171.
[97] Birkemeyer, C., Luedemann, A., Wagner, C.,Erban, A., Kopka, J., Trends Biotechnol. 2005, 23,28–33.
J. Sep. Sci. 2010, 33, 1–15 Liquid Chromatography 13
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

[98] Baran, R., Kochi, H., Saito, N., Suematsu, M., Soga, T.,Nishioka, T., Robert, M., Tomita, M., Biomed Chro-matogr. Bioinformatics 2006, 7, 530.
[99] Vorst, O., De Vos, C. H. R., Lommen, A., Staps, R. V.,Visser, R. G. F., Bino, R. J., Hall, R. D., Metabolomics2005, 1, 169–180.
[100] Duran, A. L., Yang, J., Wang, L., Sumner, L. W.,Bioinformatics 2003, 19, 2283–2293.
[101] Neuweger, H., Albaum, S. P., Dondrup, M., Persicke,M., Watt, T., Niehaus, K., Stoye, J., Goesmann, A.,Bioinformatics 2008, 24, 2726–2732.
[102] Idborg-Bjorkman, H., Edlund, P. O., Kvalheim, O. M.,Schuppe-Koistinen, I., Jacobsson, S. P., Anal. Chem.2003, 75, 4784–4792.
[103] Lenz, E. M., Bright, J., Knight, R., Westwood, F. R.,Davies, D., Major, H., Wilson, I. D., Biomarkers 2005,10, 173–187.
[104] Sadygov, R. G., Maroto, F. M., Huhmer, A. F. R., Anal.Chem. 2006, 78, 8207–8217.
[105] Idborg, H., Zamani, L., Edlund, P. O., Schuppe-Koistinen,I., Jacobsson, S. P., J. Chromatogr. B 2005, 828, 14–20.
[106] Kind, T., Tolstikov, V., Fiehn, O., Weiss, R. H., Anal.Biochem. 2007, 363, 185–195.
[107] Bro, R., Smilde, A. K., J. Chemometrics 2003, 17, 16–33.
[108] Jackson, J. E., A User’s Guide to Principal Compo-nents, Wiley Interscience, New York 2004.
[109] Eriksson, L., Johansson, E., Kettaneh-Wold, N., Wold,S., Multi- and Megavariate Data Analysis: Principlesand Applications, Umetrics Academy, Umea, Sweden2002.
[110] Keun, H. C., Ebbels, T. M. D., Antti, H., Bollard, M. E.,Beckonert, O., Holmes, E., Lindon, J. C., Nicholson,J. K., Anal. Chim. Acta 2003, 490, 265–276.
[111] Smilde, A. K., van der Werf, M. J., Bijlsma, S., van derWerff-van der Vat, B., Jellema, R. H., Anal. Chem. 2005,77, 6729–6736.
[112] Kvalheim, O. M., Brakstad, F., Liang, Y. Z., Anal. Chem.1994, 66, 43–51.
[113] Sokal, R. R., Rohlf, F. J., Biometry: The Principles andPractice of Statistics in Biological Research, W. H.Freeman and Company, New York 1995.
[114] Weckwerth, W., Morgenthal, K., Drug Discov. Today2005, 10, 1551–1558.
[115] Broadhurst, D. I., Kell, D. B., Metabolomics 2006, 2,171–196.
[116] Holmes, E., Antti, H., Analyst 2002, 127, 1549–1557.
[117] Massart, D. L., Vandeginste, B. G. M., Buydens,L. M. C., De Jong, S., Lewi, P. J., Smeyers-Verbeke, J.(Eds.), Handbook of Chemometrics and Qualimetrics,Part A, Elsevier, Oxford, UK 1997.
[118] Eriksson, L., Antti, H., Gottfries, J., Holmes, E., Johans-son, E., Lindgren, F., Long, I., Lundstedt, T., Trygg, J.,Wold, S., Anal. Bioanal. Chem. 2004, 380, 419–429.
[119] Hotelling, H., J. Educat. Psychol. 1933, 24, 417–441.
[120] Trygg, J., Gullberg, J., Johansson, A. I., Jonsson, P.,Moritz, T., Biotechnol. Agri. Forestry 2006, 57, 117–128.
[121] Wiener, M. C., Sachs, J. R., Deyanova, E., Yates, N. A.,Anal. Chem. 2004, 76, 6085–6096.
[122] Shaffer, J. P., Ann. Rev. Psychol. 1995, 46, 561–584.
[123] Schauer, N., Zamir, D., Fernie, A. R., J. Exp. Bot. 2005,56, 297–307.
[124] Serkova, N. J., Jackman, M., Brown, J. L., Liu, T.,Hirose, R., Roberts, J. P., Maher, J. J., Niemann, C. U.,J. Hepatol. 2006, 44, 956–962.
[125] Pearson, K., Philos. Mag. 1901, 2, 559–572.
[126] Major, H. J., Williams, R., Wilson, A. J., Wilson, I. D.,Rapid Commun. Mass Spectrom. 2006, 20, 3295–3302.
[127] Pierce, K. M., Hope, J. L., Hoggard, J. C., Synovec,R. E., Talanta 2006, 70, 797–804.
[128] Bylund, D., Samskog, J., Markides, K. E., Jacobsson,S. P., J. Am. Soc. Mass Spectrom. 2003, 14, 236–240.
[129] Belton, P. S., Colquhoun, I. J., Kemsley, E. K., Delga-dillo, I., Roma, P., Dennis, M. J., Sharman, M., Holmes,E., Nicholson, J. K., Spraul, M., Food Chem. 1998, 61,207–213.
[130] Jansen, J. J., Hoefsloot, H. C. J., Boelens, H. F. M., vander Greef, J., Smilde, A. K., Bioinformatics 2004, 20,2438–2446.
[131] Chan, E. C. Y., Yap, S. L., Lau, A. J., Leow, P. C., Toh,D. F., Koh, H. L., Rapid Commun. Mass Spectrom.2007, 21, 519–528.
[132] Eisen, M. B., Spellman, P. T., Brown, P. O., Botstein, D.,Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868.
[133] Tikunov,Y., Lommen, A., De Vos, C. H. R., Verhoeven,H. A., Bino, R. J., Hall, R. D., Bovy, A. G., Plant Physiol.2005, 139, 1125–1137.
[134] Beckonert, O., Bollard, M. E., Ebbels, T. M. D., Keun,H. C., Antti, H., Holmes, E., Lindon, J. C., Nicholson, J.K., Anal. Chim. Acta 2003, 490, 3–15.
[135] Wold, S., Johansson, E., Cocchi, M., 3D QSAR in DrugDesign: Theory, Methods and Applications, EscomScience Publisher, Leiden 1993, pp. 523–550.
[136] Wold, S., Sjostrom, M., Eriksson, L., ChemometricIntell. Lab. Syst. 2001, 58, 109–130.
[137] Kettaneh-Wold, N., Chemometric Intell. Lab. Syst.1992, 14, 57–69.
[138] Wold, S., Ruhe, A., Wold, H., Dunn, W. J., Siam J., Sci.Stat. Comput. 1984, 5, 735–743.
[139] Daszykowski, M., Walczak, B., Massart, D. L., Chemo-metric Intell. Lab. Syst. 2003, 65, 97–112.
[140] Trygg, J., Wold, S., J. Chemometrics 2002, 16, 119–128.
[141] Trygg, J., Wold, S., J. Chemometrics 2003, 17, 53–64.
[142] Quinlan, J. R., J. Artif. Intell. Res. 1996, 4, 77–90.
[143] Breiman, L., Friedman, J. H., Olshen, R. A., Stone, C. J.,Classification and Regression Trees, Wadsworth &Brooks, Monterey, CA, USA 1984.
[144] Zheng, Z. J., Knowl-Based Syst. 1998, 10, 421–430.
[145] Gama, J., Adv. Intell. Data Anal. 1997, 1280, 187–198.
[146] Breiman, L., Mach. Learn. 2001, 45, 5–32.
[147] Gallant, S. I., IEEE Trans. Neural Netw. 1990, 1,179–191.
[148] Taylor, J., King, R. D., Altmann, T., Fiehn, O., Bioin-formatics 2002, 18, S241–S248.
[149] Poggio, T., Girosi, F., Proc. IEEE 1990, 78, 1481–1497.
[150] Hand, D. J., Yu, K. M., Int. Stat. Rev. 2001, 69, 385–398.
J. Sep. Sci. 2010, 33, 1–1514 J. Boccard et al.
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com

[151] Aha, D. W., Kibler, D., Albert, M. K., Mach. Learn. 1991,6, 37–66.
[152] Wettschereck, D., Aha, D. W., Mohri, T., Artif. Intell.Rev. 1997, 11, 273–314.
[153] Keerthi, S. S., Gilbert, E. G., Mach. Learn. 2002, 46,351–360.
[154] Platt, J., IEEE Intell. Syst. 1998, 13, 26–28.
[155] Vapnik, V. N., IEEE Trans. Neural Netw. 1999, 10, 988–999.
[156] Guyon, I., Weston, J., Barnhill, S., Vapnik, V., Mach.Learn. 2002, 46, 389–422.
[157] Mahadevan, S., Shah, S. L., Marrie, T. J., Slupsky,C. M., Anal. Chem. 2008, 80, 7562–7570.
[158] Mitchell, T. M., Machine Learning, McGraw Hill HigherEducation, New York 1997.
[159] Hall, M. A., Holmes, G., IEEE Trans. Knowl. Data En.2003, 15, 1437–1447.
[160] Guyon, I., Elisseeff, A., J. Mach. Learn. Res. 2003, 3,1157–1182.
[161] Fisher, R. A., Statistical Methods for Research Workers,Oliver and Boyd, Edinburgh, UK 1925.
[162] Robnik-Sikonja, M., Kononenko, I., Mach. Learn. 2003,53, 23–69.
[163] Kononenko, I., Simec, E., Robnik-Sikonja, M., Appl.Intell. 1997, 7, 39–55.
[164] Wold, S., Antti, H., Lindgren, F., Ohman, J., Chemo.Intell. Lab. Syst. 1998, 44, 175–185.
[165] Kullback, S., Ann. Math. Stat. 1952, 23, 88–102.
[166] Yu, L., Liu, H., J. Mach. Learn. Res. 2004, 5, 1205–1224.
[167] Kohavi, R., John, G. H., Artif. Intell. 1997, 97, 273–324.
[168] Cohen, J., Edu. Psychol. Meas. 1960, 20, 37–46.
[169] Fleiss, J. L., Psychol. Bull. 1971, 76, 378–382.
[170] Bousquet, O., Elisseeff, A., J. Mach. Learn. Res. 2002,2, 499–526.
[171] Dietterich, T. G., Neural Comput. 1998, 10,1895–1923.
[172] Nadeau, C., Bengio, Y., Mach. Learn. 2003, 52,239–281.
[173] Pierce, K. M., Hoggard, J. C., Mohler, R. E., Synovec, R.E., J. Chromatogr. A 2008, 1184, 341–352.
J. Sep. Sci. 2010, 33, 1–15 Liquid Chromatography 15
& 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jss-journal.com
Related Documents











