Vol. 53, No. 4 APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1987, p. 823-827 0099-2240/87/040823-05$02.OO/O Copyright © 1987, American Society for Microbiology Kinetics of Homoacetic Fermentation of Lactate by Clostridium formicoaceticum SHANG-TIAN YANG,'* I-CHING TANG,2 AND MARTIN R. OKOS2 Department of Chemical Engineering, The Ohio State University, Columbus, Ohio 43210,' and Department of Agricultural Engineering, Purdue University, West Lafayette, Indiana 479072 Received 14 October 1986/Accepted 20 December 1986 Clostridiumformicoaceticum homofermentatively converted lactate to acetate at mesophilic temperatures (30 to 42°C) and at pHs between 6.6 and 9.6. The production of acetate was found to be growth associated. Approximately 0.96 g of acetic acid and 0.066 g of cells were formed from each gram of lactic acid consumed at 3rC. The concentration of the substrate (lactate) had little or no effect on the growth rate; however, the fermentation was inhibited by acetic acid. The bacterium grew at an optimal pH of 7.6 and an optimal temperature of 37C. Small amounts of bicarbonate were stimulatory to bacterial growth. Bacterial growth was enhanced, however, by the use of higher concentrations of bicarbonate in the media, only because higher buffer capacities were obtained and proper medium pH could be maintained for growth. Based on its ability to convert lactate to acetate, this homoacetic bacterium may be important in the anaerobic methanogenic process when lactate is a major intermediary metabolite. In the past decade, growing concern about environmental pollution and energy shortages have generated interest in the production of methane through anaerobic digestion of or- ganic wastes. The anaerobic methanogenic process degrades organic matter to the gaseous products CH4 and CO2 (8), which occurs as a result of three distinct but simultaneous metabolic phases (9). First, in the hydrolytic phase, large, complex organic matter is hydrolyzed and fermented to C, to C5 organic acids, alcohols, H2, and CO2. In the second (acetogenic) phase, C3 to C5 organic acids and alcohols are further reduced to acetate, formate, methanol, C02, and H2. These products are then used for methanogenesis in the last phase. The complete process involves at least four metabol- ically distinct groups of bacteria: hydrolytic, H2-producing acetogenic, homoacetogenic, and methanogenic (9, 10). Ace- tic acid is the major immediate precursor for methane formation (8, 9). Acetic acid could be derived from carbo- hydrates by the following methods: (i) homoacetic fermen- tation of glucose to acetate; (ii) heteroacetogenic fermenta- tion of glucose in the presence of H2-utilizing methanogens to shift the metabolism to favor the acetate formation; (iii) homolactic fermentation of glucose or lactose to lactate and then to acetate by homoacetic fermentation or by H2- producing acetogenic fermentation with the coproductions of H2, CO2, and formate; and (iv) ethanol fermentation followed by H2-producing acetogenic fermentation. The ex- act fermentation kinetics is not known yet and could be very complicated and highly dependent on the environmental conditions (S. T. Yang, M. R. Okos, and J. C. Nye, Meet. Am. Soc. Agric. Eng., paper 82-3592, 1982). Recently, results of 14C tracer studies (6) on the interme- diary metabolism during anaerobic degradation of whey lactose have shown that lactate is the major intermediary metabolite of lactose fermentation in a chemostat operated under lactose-limited conditions. Some 82% of the lactose was transformed into lactate during the hydrolytic phase. Lactate and other intermediary products were then trans- formed into acetate and H2-CO2, which were the substrates for methanogens. Although lactate is usually not found in * Corresponding author. large amounts in a normal anaerobic digester under steady- state conditions, large amounts of lactate are accumulated during the anaerobic fermentation when the digester pH is below 6 (11), indicating that lactate may have been produced and converted to acetate or other compounds during the digestion process. Homoacetogenic bacteria can ferment a wide spectrum of multi- or one-carbon compounds to produce acetic acid (1, 3, 4, 7). Because more than 70% of the methane formed in the anaerobic digester is directly derived from acetic acid (8, 9), the homoacetic bacteria may play an important role in the anaerobic digestion process. Although several homoacetic bacteria have been isolated from anaerobic digesters (1, 3, 4, 7), in contrast to the methanogens, very little has been done on characterizing the homoacetic bacteria. To our knowl- edge, no homoacetic bacteria can ferment lactose. However, results of previous research (1, 3) have shown that two homoacetic bacteria, Acetobacterium woodii and Clostrid- ium formicoaceticum, can grow on lactate. In addition, the sulfate-reducing bacteria Desulfovibrio spp. can produce acetate from lactate in the presence of sulfate or H2-utilizing bacteria (5). These bacteria may be responsible for the conversion of lactate to acetate in anaerobic digestion. The growth of homoacetic bacteria on sugars and H2-CO2 has been relatively well studied (1, 3), but very little is known about the growth of these bacteria on lactate. C. formicoaceticum is a gram-negative rod, strictly anaerobic, and mesophilic with a reported optimum temper- ature of about 37°C and a pH of about 7.2 when grown on fructose (1). This bacterium can neither use nor produce hydrogen gas because of a lack of hydrogenase. About 3 mol of acetate was produced from each mole of fructose that was fermented during the exponential phase. However, formate was also produced at the expense of acetate in the stationary phase. This bacterium is also known to grow on lactate, but no work has been reported on the growth kinetics. In this study the fermentation kinetics of C. formicoaceticum grown on lactate was investigated. The effects of tempera- ture, pH, sodium chloride, lactate, acetate, and bicarbonate on bacterial growth were studied and are reported here. The possible functional importance of this bacterium in 823 on July 16, 2018 by guest http://aem.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vol. 53, No. 4APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Apr. 1987, p. 823-8270099-2240/87/040823-05$02.OO/OCopyright © 1987, American Society for Microbiology
Kinetics of Homoacetic Fermentation of Lactate byClostridium formicoaceticum
SHANG-TIAN YANG,'* I-CHING TANG,2 AND MARTIN R. OKOS2Department of Chemical Engineering, The Ohio State University, Columbus, Ohio 43210,' and Department of
Agricultural Engineering, Purdue University, West Lafayette, Indiana 479072
Received 14 October 1986/Accepted 20 December 1986
Clostridiumformicoaceticum homofermentatively converted lactate to acetate at mesophilic temperatures (30to 42°C) and at pHs between 6.6 and 9.6. The production of acetate was found to be growth associated.Approximately 0.96 g of acetic acid and 0.066 g of cells were formed from each gram of lactic acid consumedat 3rC. The concentration of the substrate (lactate) had little or no effect on the growth rate; however, thefermentation was inhibited by acetic acid. The bacterium grew at an optimal pH of 7.6 and an optimaltemperature of 37C. Small amounts of bicarbonate were stimulatory to bacterial growth. Bacterial growth wasenhanced, however, by the use of higher concentrations of bicarbonate in the media, only because higher buffercapacities were obtained and proper medium pH could be maintained for growth. Based on its ability to convertlactate to acetate, this homoacetic bacterium may be important in the anaerobic methanogenic process whenlactate is a major intermediary metabolite.
In the past decade, growing concern about environmentalpollution and energy shortages have generated interest in theproduction of methane through anaerobic digestion of or-ganic wastes. The anaerobic methanogenic process degradesorganic matter to the gaseous products CH4 and CO2 (8),which occurs as a result of three distinct but simultaneousmetabolic phases (9). First, in the hydrolytic phase, large,complex organic matter is hydrolyzed and fermented to C, toC5 organic acids, alcohols, H2, and CO2. In the second(acetogenic) phase, C3 to C5 organic acids and alcohols arefurther reduced to acetate, formate, methanol, C02, and H2.These products are then used for methanogenesis in the lastphase. The complete process involves at least four metabol-ically distinct groups of bacteria: hydrolytic, H2-producingacetogenic, homoacetogenic, and methanogenic (9, 10). Ace-tic acid is the major immediate precursor for methaneformation (8, 9). Acetic acid could be derived from carbo-hydrates by the following methods: (i) homoacetic fermen-tation of glucose to acetate; (ii) heteroacetogenic fermenta-tion of glucose in the presence of H2-utilizing methanogensto shift the metabolism to favor the acetate formation; (iii)homolactic fermentation of glucose or lactose to lactate andthen to acetate by homoacetic fermentation or by H2-producing acetogenic fermentation with the coproductionsof H2, CO2, and formate; and (iv) ethanol fermentationfollowed by H2-producing acetogenic fermentation. The ex-act fermentation kinetics is not known yet and could be verycomplicated and highly dependent on the environmentalconditions (S. T. Yang, M. R. Okos, and J. C. Nye, Meet.Am. Soc. Agric. Eng., paper 82-3592, 1982).
Recently, results of 14C tracer studies (6) on the interme-diary metabolism during anaerobic degradation of wheylactose have shown that lactate is the major intermediarymetabolite of lactose fermentation in a chemostat operatedunder lactose-limited conditions. Some 82% of the lactosewas transformed into lactate during the hydrolytic phase.Lactate and other intermediary products were then trans-formed into acetate and H2-CO2, which were the substratesfor methanogens. Although lactate is usually not found in
* Corresponding author.
large amounts in a normal anaerobic digester under steady-state conditions, large amounts of lactate are accumulatedduring the anaerobic fermentation when the digester pH isbelow 6 (11), indicating that lactate may have been producedand converted to acetate or other compounds during thedigestion process.Homoacetogenic bacteria can ferment a wide spectrum of
multi- or one-carbon compounds to produce acetic acid (1, 3,4, 7). Because more than 70% of the methane formed in theanaerobic digester is directly derived from acetic acid (8, 9),the homoacetic bacteria may play an important role in theanaerobic digestion process. Although several homoaceticbacteria have been isolated from anaerobic digesters (1, 3, 4,7), in contrast to the methanogens, very little has been doneon characterizing the homoacetic bacteria. To our knowl-edge, no homoacetic bacteria can ferment lactose. However,results of previous research (1, 3) have shown that twohomoacetic bacteria, Acetobacterium woodii and Clostrid-ium formicoaceticum, can grow on lactate. In addition, thesulfate-reducing bacteria Desulfovibrio spp. can produceacetate from lactate in the presence of sulfate or H2-utilizingbacteria (5). These bacteria may be responsible for theconversion of lactate to acetate in anaerobic digestion. Thegrowth of homoacetic bacteria on sugars and H2-CO2 hasbeen relatively well studied (1, 3), but very little is knownabout the growth of these bacteria on lactate.
C. formicoaceticum is a gram-negative rod, strictlyanaerobic, and mesophilic with a reported optimum temper-ature of about 37°C and a pH of about 7.2 when grown onfructose (1). This bacterium can neither use nor producehydrogen gas because of a lack of hydrogenase. About 3 molof acetate was produced from each mole of fructose that wasfermented during the exponential phase. However, formatewas also produced at the expense of acetate in the stationaryphase. This bacterium is also known to grow on lactate, butno work has been reported on the growth kinetics. In thisstudy the fermentation kinetics of C. formicoaceticumgrown on lactate was investigated. The effects of tempera-ture, pH, sodium chloride, lactate, acetate, and bicarbonateon bacterial growth were studied and are reported here. Thepossible functional importance of this bacterium in
823
on July 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
APPL. ENVIRON. MICROBIOL.
anaerobic digestion is also discussed, based on its ability toconvert lactate to acetate.
MATERIALS AND METHODS
Culture. C. formicoaceticum ATCC 27076 was isolatedfrom sewage sludge by Andreesen et al. (1) and was usedthroughout this study.Medium. The medium was prepared by the procedures
described by Balch et al. (2). Unless otherwise noted, themedium contained the following, in grams per liter:K2HPO4. 3H20, 0.31; KH2PO4, 0.24; (NH4)2SO4, 0.24;MgSO4 7H20, 0.1; NaCl, 0.48; CaCl2- 2H20, 0.0064;FeSO4 .7H20, 0.002; resazurin, 0.0005; Trypticase (BBLMicrobiology Systems, Cockeysville, Md.), 1.0; yeast ex-tract (Difco Laboratories, Detroit, Mich.), 1.0; L-cysteinehydrochloride H20, 0.5; sodium lactate, 7.4; NaHCO3, 6.0.A vitamin solution and a trace mineral solution were eachadded at 10 ml per liter of medium. The gas phase was 40%C02-60% N2 at 24.7 lb/in2, and the final pH was 7.0.
In the experiments for determining the growth yield,concentrations of sodium lactate in the medium varied from0.038 to 0.188 M. To study the pH range for growth, themedium pH was adjusted to the desired pH range (betweenpH 6 and 10) by adding 4 N HCl or 4 N NaOH. To evaluatethe effect of sodium ion, NaCl solution was added to themedium to a final concentration of 0.05 to 3.5%. To study theeffect of NaHCO3 on bacterial growth, 0.5 M potassiumphosphate buffer (pH 7.6) was used for media containingvarious amounts of NaHCO3 (0 to 16 g/liter). Initial concen-trations of sodium acetate of between 0 and 0.3 M were usedto study the effect of acetate on bacterial growth.
Culture techniques. The anaerobic syringe techniques de-scribed by Balch et al. (2) were used in this study. Cells werecultivated in serum tubes (18 by 150 mm) and 125-ml serumbottles containing 7.5 and 50.0 ml of the medium, respec-tively. The culture was maintained in an active state bytransferring 2% of the culture volume to fresh medium everyweek. All cultures were incubated at 37°C without shaking inthe dark.
Kinetic studies. Batch fermentations were performed inserum tubes, serum bottles, or both under uncontrolled pHconditions to evaluate effects of pH, sodium ion, lactate,acetate, and bicarbonate. A liquid sample (1 ml) was takenfor optical density (OD) reading, pH measurement, andhigh-performance liquid chromatographic (HPLC) assay.
Fermentation kinetics was also studied under controlledconstant pH conditions with 5-liter fermentors (NewBrunswick Scientific Co., Inc., Edison, N.J.). The fermen-tation broth was automatically titrated with membrane-filtered (pore size, 0.2 ,um) 4 N NaOH to maintain a desiredpH (±0.01). A total of 3 liters of medium was used for eachbatch fermentation, and 50 ml of active culture was inocu-lated into the medium. The anaerobiosis was maintained bykeeping a positive N2-CO2 gas pressure (19.7 lb/in2 or 1.34atm) inside the fermentor. At proper time intervals (depend-ing on the fermentation rate), 5 ml of liquid sample waswithdrawn from the sampling port for OD reading and thenfrozen for future HPLC analysis.
Analytical techniques. Lactate, acetate, and other organiccompounds (if present) were identified and quantitated bythe HPLC method. A system (Waters Associates, Inc.,Milford, Mass.) consisting of a pump (6000A; Waters),injector (U6K; Waters), and differential refractometer(R401; Waters) was used. A 10-pd portion of supernatantfrom centrifuged samples was injected onto an organic acid
i pHo -70tCalf.4a -6.8
cf 0. lmn airto ure o tnad fec
Bacterial -6.6wt6.2~
200nm 203 50 60 70 8090ectropotomete 130 -6.250TIME (HOURS)
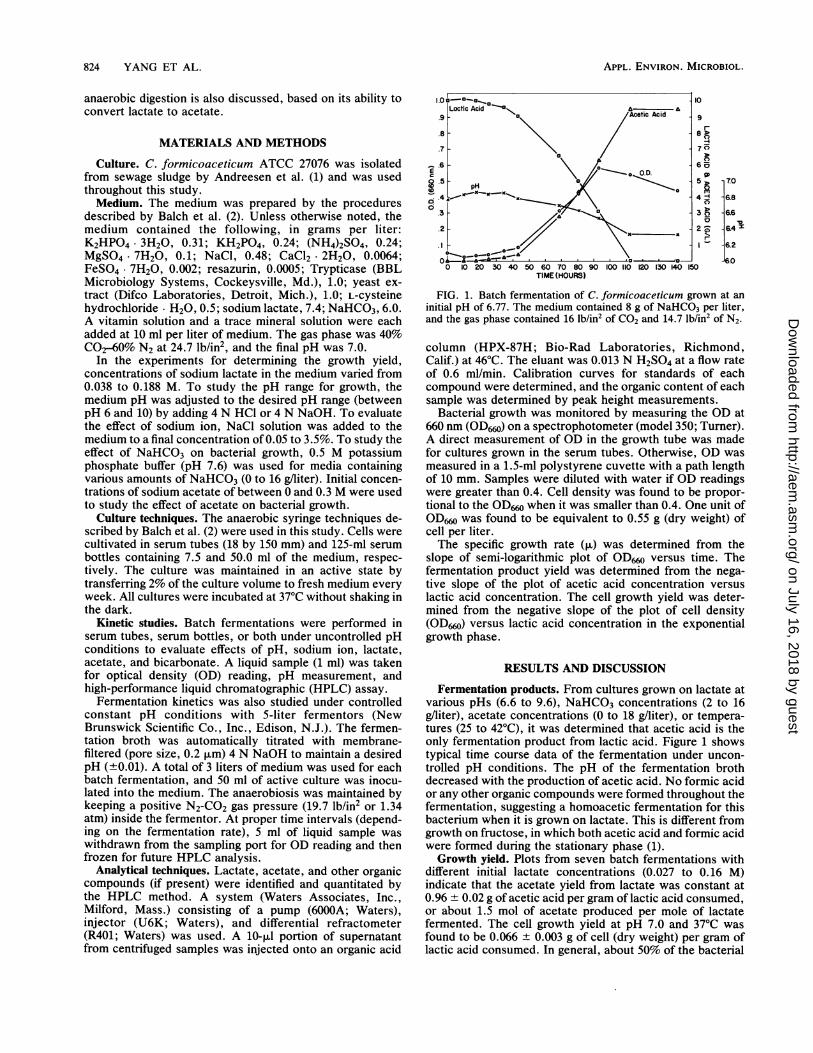
FIG. 1. Batch fermentation of C. formicoaceticum grown at aninitial pH of 6.77. The medium contained 8 g of NaHCO3 per liter,and the gas phase contained 16 lb/in2 Of CO2 and 14.7 lb/in2 of N2.
column (HPX-87H; Bio-Rad Laboratories, Richmond,Calif.) at 46aC. The eluant was 0.013 N H2S04 at a flow rateof 0.6 ml/min. Calibration curves for standards of eachcompound were determined, and the organic content of eachsample was determined by peak height measurements.
Bacterial growth was monitored by measuring the gD at660 nm(OD6l) on a spectrophotometer (model 350; Turner).A direct measurement of GD in the growth tube was madefor cultures grown in the serum tubes. Otherwise, GD wasmeasured in a 1.5-ml polystyrene cuvette with a path lengthof 10 mm. Samples were diluted with water if GD readingswere greater than 0.4. Cell density was found to be propor-tional to the OD60 when it was smaller than 0.4. One unit ofODw0 was found to be equivalent to 0.55 g (dry weight) ofcell per liter.The specific growth rate(ps) was determined from the
slope of semi-logarithmic plot of OD6w versus time. Thefermentation product yield was determined from the nega-tive slope of the plot of acetic acid concentration versuslactic acid concentration. The cell growth yield was deter-mined from the negative slope of the plot of cell density(OD660) versus lactic acid concentration in the exponentialgrowth phase.
RESULTS AND DISCUSSION
Fermentation products. From cultures grown on lactate atvarious pHs (6.6 to 9.6), NaHCO3 concentrations (2 to 16g/liter), acetate concentrations (0 to 18 g/liter), or tempera-tures (25 to 420C), it was determined that acetic acid is theonly fermentation product from lactic acid. Figure 1 showstypical time course data of the fermentation under uncon-trolled pH conditions. The pH of the fermentation brothdecreased with the production of acetic acid. No formic acidor any other organic compounds were formed throughout thefermentation, suggesting a homoacetic fermentation for thisbacterium when it is grown on lactate. This is different fromgrowth on fructose, in which both acetic acid and formic acidwere formed during the stationary phase (1).Growth yield. Plots from seven batch fermentations with
different initial lactate concentrations (0.027 to 0.16 M)indicate that the acetate yield from lactate was constant at0.96 + 0.02 g of acetic acid per gram of lactic acid consumed,or about 1.5 mol of acetate produced per mole of lactatefermented. The cell growth yield at pH 7.0 and 37°C wasfound to be 0.066 + 0.003 g of cell (dry weight) per gram oflactic acid consumed. In general, about 50% of the bacterial
824 YANG ET AL.
on July 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

HOMOACETIC FERMENTATION OF LACTATE 825
cell mass was carbon. Thus, nearly 100% of the carbon andhydrogen in the substrate (lactic acid) was recovered in thefermentation product (acetic acid) and the cells. Both prod-uct formation and cell growth were proportional to thesubstrate consumption, suggesting that the formation ofacetate from lactate is growth associated.
Effect of temperature. It was found that the growth ratedecreased when the temperature was increased from 37 to42°C in a controlled, constant pH experiment. Temperaturedid not affect the acetate yield, but only about 60% of the cellgrowth yield was obtained at 42°C as compared with that at37°C (Fig. 2). The observed lower fermentation rate at 42°Cmight be due largely to the lower cell growth yield. Thisfinding was consistent with the reported optimal temperatureof about 37°C for C. formicoaceticum when grown onfructose (1).
Effect of NaCl. NaCl in the concentration range of 0.05 to3.0% (85 to 510 mM) did not significantly affect the specificgrowth rate. However, a longer lag phase and a slowergrowth rate were found with 3.5% NaCl.
Effect of lactate. The concentration of lactate in the rangeof 0.027 to 0.16 M did not affect the specific growth rate.However, a longer lag phase was found for those culturesgrown in a medium with a higher sodium concentration whencells in the stationary phase were used to initiate thefermentation. This effect was not found for experiments withactive cultures. Because a concentration of NaCl up to 0.51M did not affect bacterial growth, we conclude that substrate(lactate) concentrations between 0.027 and 0.16 M are nei-ther stimulatory nor inhibitory to this fermentation.pH range for growth. Neither bacterial growth nor change
in lactate or acetate concentration was found for cellscultivated at a pH below 6.6 or above 9.6 for a period of 30days, suggesting a growth pH range of between 6.6 and 9.6.
Effects of pH. By using data from the early exponentialgrowth phase, the specific growth rates at various pH valueswere determined, and the effect of the pH on the specificgrowth rate is shown in Fig. 3. Apparently, this bacteriumhas an optimal pH for growth at about 7.6. The fermentationrate was relatively insensitive to pH changes between 7.5and 8.8 but decreased dramatically at a pH below 7.5 orabove 8.8.The medium pH also affected the lag phase. In the pH
1.50
' 1.00
(Dd
0.75
Ct)Z 0.50
C]
-Jw 0.250
0.00-2. 4 6. 8. 10.
LACTIC ACID (G/L)
.20
.1 5
9-
_ .1 0
.05
pH
FIG. 3. Effect of pH on specific growth rate. ,u, Specific growthrate.
range of 6.6 to 7.6, the bacterium had a longer lag phase at alower pH. However, the length of lag phase was also greatlyaffected by the viability of the inoculum and other environ-mental factors such as salt concentration.;Although the growth of C. formicoaceticurn was sensitive
to pH, the acetate yield was not dependent on pH. Thismight be because the production of acetic acid during thefermentation was always growth associated. However,higher ultimate cell densities were observed for culturesgrown at higher pH values. This could be due to either thehigher cell yield or the lower cell decay rate at a higher pH.
Effect of acetate. The specific growth rate was lower whenthe concentration of sodium acetate was higher at the samepH. This effect was mainly due to the acetic acid becauseconcentrations of NaCl up to 0.5 M did not affect thefermentation. The specific growth rate decreased from 0.14to 0.035 h-1 when the concentration of acetic acid increasedfrom 0 to 18 g/liter (0.3 M) at pH 7.0 (Fig. 4). However,concentrations of acetate up to 0.6 M did not completelyinhibit bacterial growth at pH 7.0 or higher. The effect ofacetate was found to be stronger at a lower medium pH,indicating that the undissociated acid may be more inhibitorythan the dissociated acid.
Effect of bicarbonate. The concentration of bicarbonatemay affect bacterial growth either directly or indirectlythrough its buffering effect on the pH. Andreesen et al. (1)
I
C:
n
12. 14. 16. Is.
FIG. 2. Effect of temperature on cell yield. In a batch fermenta-tion under controlled constant pH conditions, the cell growth yielddropped significantly when the temperature was increased from 37to 42°C.
0.16
0.14
0.12
0.10
0.08
0.06
0.04
0.02
0.000. 2. 4. 6. a 10. 12. 14. 16. 18.
ACETIC ACID (G/L)FIG. 4. Effect of acetic acid on specific growth rate at pH 7.0.
I r-I I 1 T ,--I- T
pH 7.0
0 o
.,
VOL. 53, 1987
c
on July 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
826 YANG ET AL.
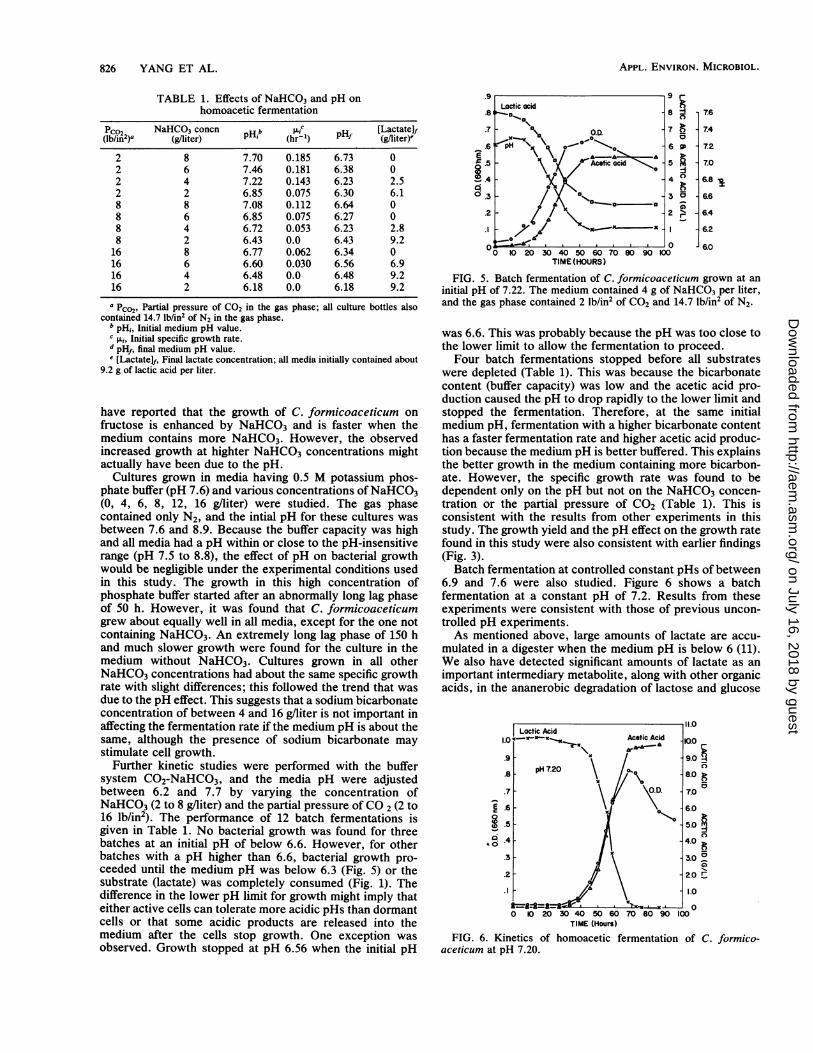
TABLE 1. Effects of NaHCO3 and pH onhomoacetic fermentation
Pco2 NaHCO3 concn b pc[HLactatelf(lb/in2)a (g/liter) (hr'1) (g/fiter)-
2 8 7.70 0.185 6.73 02 6 7.46 0.181 6.38 02 4 7.22 0.143 6.23 2.52 2 6.85 0.075 6.30 6.18 8 7.08 0.112 6.64 08 6 6.85 0.075 6.27 08 4 6.72 0.053 6.23 2.88 2 6.43 0.0 6.43 9.2
16 8 6.77 0.062 6.34 016 6 6.60 0.030 6.56 6.916 4 6.48 0.0 6.48 9.216 2 6.18 0.0 6.18 9.2a pC02, Partial pressure of CO2 in the gas phase; all culture bottles also
contained 14.7 lb/in2 of N2 in the gas phase.b pHi, Initial medium pH value.c .i., Initial specific growth rate.d pHf, final medium pH value.e [Lactatelf, Final lactate concentration; all media initially contained about
9.2 g of lactic acid per liter.
have reported that the growth of C. formicoaceticum onfructose is enhanced by NaHCO3 and is faster when themedium contains more NaHCO3. However, the observedincreased growth at highter NaHCO3 concentrations mightactually have been due to the pH.
Cultures grown in media having 0.5 M potassium phos-phate buffer (pH 7.6) and various concentrations ofNaHCO3(0, 4, 6, 8, 12, 16 g/liter) were studied. The gas phasecontained only N2, and the intial pH for these cultures wasbetween 7.6 and 8.9. Because the buffer capacity was highand all media had a pH within or close to the pH-insensitiverange (pH 7.5 to 8.8), the effect of pH on bacterial growthwould be negligible under the experimental conditions usedin this study. The growth in this high concentration ofphosphate buffer started after an abnormally long lag phaseof 50 h. However, it was found that C. formicoaceticumgrew about equally well in all media, except for the one notcontaining NaHCO3. An extremely long lag phase of 150 hand much slower growth were found for the culture in themedium without NaHCO3. Cultures grown in all otherNaHCO3 concentrations had about the same specific growthrate with slight differences; this followed the trend that wasdue to the pH effect. This suggests that a sodium bicarbonateconcentration of between 4 and 16 g/liter is not important inaffecting the fermentation rate if the medium pH is about thesame, although the presence of sodium bicarbonate maystimulate cell growth.
Further kinetic studies were performed with the buffersystem C02-NaHCO3, and the media pH were adjustedbetween 6.2 and 7.7 by varying the concentration ofNaHCO3 (2 to 8 gfliter) and the partial pressure of CO 2 (2 to16 lb/in2). The performance of 12 batch fermentations isgiven in Table 1. No bacterial growth was found for threebatches at an initial pH of below 6.6. However, for otherbatches with a pH higher than 6.6, bacterial growth pro-ceeded until the medium pH was below 6.3 (Fig. 5) or thesubstrate (lactate) was comnpletely consumed (Fig. 1). Thedifference in the lower pH limit for growth might imply thateither active cells can tolerate more acidic pHs than dormantcells or that some acidic products are released into themedium after the cells stop growth. One exception wasobserved. Growth stopped at pH 6.56 when the initial pH
.7
.2
.1
7.6
7.4
72
7.0
6.8 s
6.6
6.4
6.2
6.0
TIME (HOURS)
FIG. 5. Batch fermentation of C. formicoaceticum grown at aninitial pH of 7.22. The medium contained 4 g of NaHCO3 per liter,and the gas phase contained 2 lb/in2 of CO2 and 14.7 lb/in2 of N2.
was 6.6. This was probably because the pH was too close tothe lower limit to allow the fermentation to proceed.Four batch fermentations stopped before all substrates
were depleted (Table 1). This was because the bicarbonatecontent (buffer capacity) was low and the acetic acid pro-duction caused the pH to drop rapidly to the lower limit andstopped the fermentation. Therefore, at the same initialmedium pH, fermentation with a higher bicarbonate contenthas a faster fermentation rate and higher acetic acid produc-tion because the medium pH is better buffered. This explainsthe better growth in the medium containing more bicarbon-ate. However, the specific growth rate was found to bedependent only on the pH but not on the NaHCO3 concen-tration or the partial pressure of CO2 (Table 1). This isconsistent with the results from other experiments in thisstudy. The growth yield and the pH effect on the growth ratefound in this study were also consistent with earlier findings(Fig. 3).Batch fermentation at controlled constant pHs of between
6.9 and 7.6 were also studied. Figure 6 shows a batchfermentation at a constant pH of 7.2. Results from theseexperiments were consistent with those of previous uncon-trolled pH experiments.As mentioned above, large amounts of lactate are accu-
mulated in a digester when the medium pH is below 6 (11).We also have detected significant amounts of lactate as animportant intermediary metabolite, along with other organicacids, in the ananerobic degradation of lactose and glucose
Lactic Acid .I1.0' _"_se Acetic Acid 10.0
.9 9.0pH1 20 8.0
.7 0T(u7.0
.5
'04 4%0.2 2.0~
.1 1.0
10 20 30 40 50 60 70 80 90 100
TIME (Hours)
homoacetic fermentation of C. formico-FIG. 6. Kinetics ofaceticum at pH 7.20.
0> .5C'A0-
0
C'.I-.
APPL. ENVIRON. MICROBIOL.
on July 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

HOMOACETIC FERMENTATION OF LACTATE 827
(S. T. Yang, Ph.D. thesis, Purdue University, West Lafay-ette, Ind., 1984). While no significant amounts of lactatecould be detected in a normal anaerobic digester at thesteady-state operation, lactate may have been produced andimmediately converted to acetate in the digester byhomoacetic and sulfate-reducing bacteria. Because thegrowth rate of C. formicoaceticum is not dependent on theconcentration of lactate, all the lactate produced during theanaerobic digestion can be readily converted to acetate bythis bacterium at a pH above 6.3. This may partially explainwhy lactate is only present in the sour digester with a pHbelow 6. Results of a study with mixed cultures ofhomolactic, homoacetic, and methanogenic bacteria hasshown that the steady-state lactate concentration in thismixed culture fermentation could be close to 0 if growth wasbalanced (Yang, Ph.D. thesis). About 340 ml of methane wasproduced from each gram of lactose consumed in thisfermentation.
Populations of 105 to 106 homoacetogenic bacteria per mlof sewage sludge have been reported (10). Clostridium andAcetobacterium are two well-recognized genera. The impor-tance of homoacetogenic bacteria in anaerobic digestion hasbeen ascribed to their ability to ferment multicarbon com-pounds (mainly sugars) to acetate. In this study we showedthat C. formicoaceticum can effectively convert lactate toacetate at a pH of between 6.3 and 9.6 and acetate concen-trations ofup to 0.6 M. While A. woodii also ferments lactateto acetate at a pH of between 5 and 7, it only grows attemperatures lower than 32°C and acetate concentrationsbelow 0.075 M (Yang, Ph.D. thesis). Because mostanaerobic digesters are operated at a pH of about 7.0 andacetate concentrations of about 0.1 M, C. formicoaceticumwould be more important than A. woodii for convertinglactate to acetate in the anaerobic digestion process withlactate as a major intermediary metabolite.
Populations of 104 of the sulfate-reducing bacteriaDesulfovibrio spp. have also been reported (10). In theabsence of sulfate, these bacteria convert lactate to acetate,H2, and CO2 when H2-utilizing methanogens (e.g.,Methanobacterium formicicum) are present (5). However,the fermentation rate of a coculture of Desulfovibriodesulfuricans and M. formicicum was slow and was stronglyinhibited by acetate, even at a low acetate concentration, ascompared with that of C. formicoaceticum (Yang, Ph.D.thesis). Nevertheless, one cannot overlook the importanceof sulfate-reducing bacteria in anaerobic digestion becausethey have been inevitably found in the anaerobic digesters.
The potential importance of C. formicoaceticum inanaerobic digestion may be ascribed to its ability to convertlactate to acetate. However, further work is needed to provethis hypothesis because most of the methanogenic ecosys-tems are very complex and the fermentation kinetics ishighly dependent on the environmental conditions.
ACKNOWLEDGMENTS
This study was supported in part by grant 82-CRSR-2-2013 fromthe U.S. Department of Agriculture and a seed grant from the Officeof Research and Graduate Studies, The Ohio State University.
LITERATURE CITED1. Andreesen, J. R., G. Gottschalk, and H. G. Schlegel. 1970.
Clostridium formicoaceticum nov. spec. Isolation, descriptionand distinction from C. aceticum and C. thermoaceticum. Arch.Microbiol. 72:154-174.
2. Balch, W. E., G. E. Fox, L. G. Magrum, C. R. Woese, and R. S.Wolfe. 1979. Methanogens: reevaluation of a unique biologicalgroup. Microbiol. Rev. 43:260-296.
3. Balch, W. E., S. Schoberth, R. S. Tanner, and R. S. Wolfe. 1977.Acetobacterium, a new genus of hydrogen-oxidizing, carbondioxide-reducing, anaerobic bacteria. Int. J. Syst. Bacteriol.27:355-361.
4. Braun, M., F. Mayer, and G. Gottschalk. 1981. Clostridiumaceticum (Wieringa), a microorganism producing acetic acidfrom molecular hydrogen and carbon dioxide. Arch. Microbiol.128:288-293.
5. Bryant, M. P., L. L. Campbell, C. A. Reddy, and M. R. Crabill.1977. Growth of Desulfovibrio in lactate or ethanol media low insulfate in association with H2-utilizing methanogenic bacteria.Appl. Environ. Microbiol. 33:1162-1169.
6. Chartrain, M., and J. G. Zeikus. 1986. Microbial ecophysiologyof whey biomethanation: intermediary metabolism of lactosedegradation in continuous culture. Appl. Environ. Microbiol.51:180-187.
7. Leigh, J. A., F. Mayer, and R. S. Wolfe. 1981. Acetogeniumkivui, a new thermophilic hydrogen-oxidizing, acetogenic bac-terium. Arch. Microbiol. 129:275-280.
8. McCarty, P. L. 1964. Anaerobic waste treatment fundamentals.I. Chemistry and microbiology. Public Works 95:107-122.
9. Winfrey, M. R. 1984. Microbial production of methane, p.153-219. In R. M. Atlas (ed.), Petroleum microbiology. Macmil-lan Publishing Co., New York.
10. Zeikus, J. G. 1980. Microbial populations in digesters, p. 61-87.In D. A. Stafford, B. I. Wheatley, and D. E. Hughes (ed.),Anaerobic digestion. Applied Science Publishers Ltd., London.
11. Zoetemeyer, R. J., A. J. C. M. Matthijsen, J. C. Van den Heuvel,A. Cohen, and C. Boelhouwer. 1982. Anaerobic acidification ofglucose in an upflow reactor. Biomass 2:187-199.
VOL. 53, 1987
on July 16, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
Related Documents











