KINETICS OF SOYBEAN LIPOXYGENASES ARE RELATED TO pH, SUBSTRATE AVAILABILITYAND EXTRACTION PROCEDURES VERONICA S. CHEDEA 1,3 , SIMONA VICAS ¸ 2 and CARMEN SOCACIU 1 1 Department of Chemistry and Biochemistry Faculty of Agriculture University of Agricultural Sciences and Veterinary Medicine 3-5 Ma ˘na ˘s ¸tur Street, 400372, Cluj Napoca, Romania 2 University of Oradea Oradea, Romania Accepted for Publication January 24, 2007 ABSTRACT The kinetic patterns of pure soy lipoxygenase LOX-1 and crude or defat- ted soybean extracts containing LOX isoenzymes (LOX-1, LOX-2 and LOX-3) were studied by UV spectrometry at 234 and 280 nm, depending on their extraction and measurement conditions. Different pHs (from 6.0 to 9.0), cor- responding to specific activation of LOX isoenzymes and the ratios of enzyme protein per substrate were used in order to evaluate the enzyme rates, as indicators of its affinity for substrate in different environments. The crude soy extract contained mainly LOX-1 activity (measured at 234 nm, at pH 9.0) and LOX-3, in an approximate ratio of 3:1. The LOX-2 activity was very low. The defatted extracts buffered at pH 6.8 and 7.1 showed a low LOX-1 and LOX 2 activity, but mostly LOX-3 activity (measured at 280 nm, at pH 7.1), with a mirror-type relation between the enzyme/substrate ratio and their enzymatic specific activity. The results suggest that defatting inhibits specifically the LOX-1 activity and indicate the possibility to modulate LOX activity by modi- fications of enzyme/substrate ratios and modifications of pH in the enzyme environment. PRACTICAL APPLICATIONS Because of the specific kinetic behaviors of the three different LOXs found in crude soy extracts involved in off-flavor generation, one can modulate 3 Corresponding author. TEL: +40 264 595825, 596384 (5,6,7) ext. 213; FAX: +40 264 593792; EMAIL: [email protected], [email protected] Journal of Food Biochemistry 32 (2008) 153–172. All Rights Reserved. © 2008, The Authors Journal compilation © 2008, Blackwell Publishing 153

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

KINETICS OF SOYBEAN LIPOXYGENASES ARE RELATED TOpH, SUBSTRATE AVAILABILITY AND EXTRACTION
PROCEDURES
VERONICA S. CHEDEA1,3, SIMONA VICAS2 and CARMEN SOCACIU1
1Department of Chemistry and BiochemistryFaculty of Agriculture
University of Agricultural Sciences and Veterinary Medicine3-5 Manastur Street, 400372, Cluj Napoca, Romania
2University of OradeaOradea, Romania
Accepted for Publication January 24, 2007
ABSTRACT
The kinetic patterns of pure soy lipoxygenase LOX-1 and crude or defat-ted soybean extracts containing LOX isoenzymes (LOX-1, LOX-2 and LOX-3)were studied by UV spectrometry at 234 and 280 nm, depending on theirextraction and measurement conditions. Different pHs (from 6.0 to 9.0), cor-responding to specific activation of LOX isoenzymes and the ratios of enzymeprotein per substrate were used in order to evaluate the enzyme rates, asindicators of its affinity for substrate in different environments. The crude soyextract contained mainly LOX-1 activity (measured at 234 nm, at pH 9.0) andLOX-3, in an approximate ratio of 3:1. The LOX-2 activity was very low. Thedefatted extracts buffered at pH 6.8 and 7.1 showed a low LOX-1 and LOX 2activity, but mostly LOX-3 activity (measured at 280 nm, at pH 7.1), with amirror-type relation between the enzyme/substrate ratio and their enzymaticspecific activity. The results suggest that defatting inhibits specifically theLOX-1 activity and indicate the possibility to modulate LOX activity by modi-fications of enzyme/substrate ratios and modifications of pH in the enzymeenvironment.
PRACTICAL APPLICATIONS
Because of the specific kinetic behaviors of the three different LOXsfound in crude soy extracts involved in off-flavor generation, one can modulate3 Corresponding author. TEL: +40 264 595825, 596384 (5,6,7) ext. 213; FAX: +40 264 593792;
EMAIL: [email protected], [email protected]
Journal of Food Biochemistry 32 (2008) 153–172. All Rights Reserved.© 2008, The AuthorsJournal compilation © 2008, Blackwell Publishing
153

the inhibition of these isoenzymes during soybean processing. Our experi-ments showed that pH variation could be a simple solution to inhibit the LOXisoenzymes, and therefore, the off-flavor generation.
From the analytical point of view, the techniques described in this articleare designed to be as simple as possible, and easy to use at large-scale level infood industry (food chain control). The idea is to minimize the number ofseparate chemical manipulations and, thereby, minimize errors.
These studies can offer the background of further inhibition experimentsin vitro using natural extracts. The LOX inhibition by natural antioxidants isrelated as well to pH and other factors influencing the enzyme’s activity; thisidea can be also valorized practically in the future.
INTRODUCTION
LOXs (EC 1.13.11.12) are a large family of dioxygenase enzymes con-taining non-heme iron monomeric proteins. LOXs are found in plants, fungiand animals (Funk 1996; Yamamoto et al. 1997; Grechkin 1998), and catalyzethe regio- and stereospecific oxidation of polyunsaturated fatty acids contain-ing at least one 1(Z), 4(Z)-pentadiene unit (e.g., linoleate, linolenate andarachidonate) to yield optically pure (S)-hydroperoxy fatty acids via hydrop-eroxides (Gardner 1991; Siedow 1991). A large variety of LOXs are found inplants, often considered as different groups instead of isoenzymes.
In higher plants, two major pathways involving LOXs have beendescribed in the metabolism of fatty acids, collectively known as the “LOXpathway.” One branch of the LOX pathway produces traumatic acid, a com-pound that may be involved in plant cell wound responses (Zimmerman andCoudron 1979), and volatile C6-aldehydes and C6-alcohols. These volatilecomponds are the major contributors to the characteristic “fresh green” odor ofleaves (Hatanaka 1996) and may play a role in pathogen defense (Croft et al.1993). A second branch produces jasmonic acid, a molecule that may have aregulatory role in plant cells (Staswick 1992; Sembdner and Parthier 1993;Porta and Rocha-Sosa 2002). The biological role of most LOX isoforms is,anyway, still elusive (Ivanov et al. 2001).
Besides their physiological role, plant LOXs are of significant importanceto the food industry, because these enzymes have been implicated in thegeneration of the flavor and aroma in many plant products (Casey and Hughes2004). For instance, they are responsible for the undesirable “beany,” “green”and “grassy” flavors produced during processing and storage of protein prod-ucts derived from legume seeds (Fukushima 1994; Robinson et al. 1995) andthe development of the stale flavor in beer during storage (Kobayashi et al.1993). LOXs also play an important role in the baking industry. They are quite
154 V.S. CHEDEA, S. VICAS and C. SOCACIU

effective as bleaching agents, increase mixing tolerance and improve doughrheology (Nicolas and Potus 1994; Larreta-Garde 1995; Cumbee et al. 1997;Borrelli et al. 1999).
Soybean (Glycine max L.) is a leguminous seed crop widely grown and isa rich source of unsaturated fatty acids and LOXs. Freshly refined soybean oilis practically odorless and bland, but “green, grassy, fishy” off-flavors maydevelop quickly if the oil is heated or stored under conditions that expose it tolight and oxygen or by contamination with pro-oxidant metals such as copperand iron (Berk 1992). “Beany” flavor is the principal inconvenience of tradi-tional soymilk and its products (e.g., tofu) and is caused by some ketonesand aldehydes, particularly hexanals and heptanals, produced through LOX-catalyzed oxidation (Berk 1992).
Plant LOXs are classified as LOX-1, LOX-2 or LOX-3 with respect totheir positional specificity of linoleic acid oxygenation either at C-9 and/or atC-13 of the hydrocarbon backbone of the fatty acid leading to two groups ofcompounds, the (9S)-hydroperoxy- (9-HPOD) and the (13S)-hydroperoxy(13-HPOD), respectively (Feussner and Wasternack 2002). Soybean seedLOX isoenzymes are 94–97 kDa monomeric proteins with distinct isoelectricpoints ranging from about 5.7–6.4, and can be distinguished by their optimumpH, substrate specificity, product formation and stability (Mack et al. 1987;Siedow 1991). LOX-1 is the smallest in size (94 kDa), exhibits maximalactivity at pH 9.0 and converts the free linoleic acid preferentially into the13-hydroperoxide (13-HPOD) derivatives. LOX-2 (97 kDa) is characterizedby a peak of activity at pH 6.8, and forms equal amounts of the 13- and9-hydroperoxides (13-HPOD and 9-HPOD) (Loiseau et al. 2001). In contrastto LOX-1, LOX-2 oxygenates the esterified unsaturated fatty acids in mem-branes (Maccarrone et al. 1994). LOX-3 (96.5 kDa) exhibits its maximalactivity around pH 7.0, and displays a moderate preference for producing9-HPOD (Ramadoss et al. 1978).
The Fe3+/Fe2+ cycling is an essential part of the LOX catalytic mecha-nism, and it is specifically dependent on the pH: at alkaline pH, the active Fe3+
form of the enzyme is reduced to the “inactive” Fe2+ enzyme taking an electronfrom the substrate while the base OH- takes a proton, producing a free radicalform of the 1,4 diene system. The molecular oxygen reacts with the substrateradical and forms peroxyl radical that takes back the electron from the Fe2+
enzyme, releasing the peroxidate anion and regenerating the Fe3+ enzyme. Theperoxidate finally forms the hydroperoxide product by reduction with a proton(de Groot et al. 1975; Kulkarni 2001; Radmark 2002). Studies evaluating thekinetics of soybean LOX-1 catalysis showed different behaviors of Fe2+ andFe3+ LOX (Schilstra et al. 1992; Wang et al. 1993).
Over the years, numerous LOX inhibitors have been discovered withvarying potency and selectivity, and it was shown that the mechanism of
155SOYBEAN LIPOXYGENASES KINETICS

inhibition is strongly correlated to the LOX kinetic behavior (Mogul andHolman 2001; Sadik et al. 2003; Skrzypczak-Jankun et al. 2003).
The specific kinetic behaviors of three different LOXs found in crude soyextracts may explain the off-flavor generation during food processing, and mayrepresent a prerequisite step on their inhibition by natural active compoundswith antioxidant potential. Our experiments investigated the kinetic patterns ofpure soy LOX-1 and different soybean extracts containing LOX isoenzymes,depending on their extraction and testing procedures. The main factors, whichwere considered as modulators of LOX catalysis, were the pH and the ratios ofenzyme protein per substrate, as indicators of enzyme affinity in differentenvironments.
MATERIALS AND METHODS
Chemicals
Pure soybean LOX-1 was purchased from Sigma Chemical Co., St.Louis, MO (L-8383). The standard enzyme contained 46.000 units/mg pro-tein. Linoleic acid and Tween-20 were purchased from Sigma ChemicalCo. (L-1376), while petroleum ether was from S.C. Cristal R Chim SRL,Bucharest, Romania.
Preparation of Substrate (LS) and Standard LOX-1 Solutions(S1 and S2)
The substrate LS solution of 10 mM was prepared from 28 mg of linoleicacid, 28 mg Tween-20 and 1.6 mL O2-free distilled water by homogenizationavoiding air bubbles. Then, a 0.22 mL NaOH 0.5 N solution was added andbrought to 10 mL by the addition of distilled water, to yield a clear LS solution.
Aliquots of 1 mg pure soybean LOX-1 were added to 1 mL 0.2 Mphosphate buffer at pH 6.1 or to 0.2 M borate buffer at pH 9.0, obtainingtwo standard stock solutions (S1 and S2, respectively) corresponding to46.000 units LOX-1/mL buffer. For LOX activity measurements, the S1 and S2
solutions were diluted 1:20, either with 0.2 M phosphate buffer at pH 6.1 orwith 0.2 M borate buffer at pH 9.0 corresponding to 2,300 U enzyme/mL.
Extraction of LOXs from Soybean Samples
The extraction was done using a modified form of the method of Axelrodet al. (1981): an aliquot of 5 g soybean meal was mixed with 30 mL of 0.2 Msodium phosphate buffer, pH 6.1, and stirred for 1 h at room temperature. It
156 V.S. CHEDEA, S. VICAS and C. SOCACIU

was then filtered through cheesecloth and centrifuged 10 min at 16.000 rpm.The supernatant represented the raw extract E1.
Aliquots of 50 g from the same full-fat soybean meal were washed with20 mL petroleum ether (5 ¥ 4 mL, 20 min per washing) until the solventbecame colorless. Aliquots of 10 g defatted sample were extracted with 60 mL0.2 M sodium phosphate buffer at pH 6.1. The mixture was stirred for 1 h atroom temperature, filtered through cheesecloth and centrifuged 10 min at16.000 rpm, resulting in the supernatant extract E2.
Aliquots of 10 g defatted sample were extracted with 75 mL 0.2 Msodium acetate buffer, pH 4.5 for 1 h. After filtration and centrifugation for10 min at 16.000 rpm, the pH was adjusted at 6.8 with NaOH solution, obtain-ing the extract E3.
Aliquots of 7 g defatted sample were extracted for 30 min in 70 mL0.05 M sodium phosphate buffer, pH 7.1. This represented the extractE4.
The total protein content of the four extracts was determined by theGornall method (Gornall et al. 1949). The peroxide identification in all sol-vents and extracts was also checked using the potassium iodide test (http://www.safety.ed.ac.uk), and was negative.
LOX Assay and Activity Calculation
The LOX activity for all standard solutions or soybean extracts wasdetermined by a modified method of Axelrod et al. (1981), and the time courseof the peroxide formation was measured by the increase of absorbance at234 nm, using a JASCO V-500 spectrophotometer (Clvj-Napoca, Romania) at25C. To obtain the kinetic curves and determine the curve parameters, the timecourse measurement mode of the JASCO spectrophotometer was used.
The LOX activity of the S1 or S2 standard solutions after LS substrateaddition was measured either in 0.2 M sodium phosphate buffer (at pH 6.1,7.0, 7.5 and 8.0) or in 0.2 M borate buffer at pH 9.0.
For the measurements of pure LOX activity to 0.16 mL of 1:20 dilutionsof S1 or S2 stock solutions in 0.84 mL 0.2 M phosphate (pH 6.1, 7.0, 7.5, 8.0)or 0.2 M borate buffers (pH 9.0), 8.4 mL of LS was rapidly added and mixed.The increase of absorption DA234 against the blank was recorded. The blankcontained a mixture of 0.84 mL 0.2 M borate buffer (pH 9.0) and 0.16 mLstandard enzyme (1:20).
The time course of the reaction was registered in each case, and thevariation of the product formation (absorption increase at 234 nm) per timeunit and milligrams enzyme was determined (DA234/s/mg protein). The resultswere correlated with the ratios between the enzyme quantity (mg LOX) andsubstrate concentration (nmol LS).
157SOYBEAN LIPOXYGENASES KINETICS

To determine the soy extract LOX activities, we applied three differenttests. The first test (T1) used for the E1–E4 extracts the measurement of LOXactivity at pH 9.0, according to the following procedure: x mL from eachextract was mixed with 3-x mL borate buffer (pH 9.0) and 25 mL LS solution.The kinetic curves were registered at 25C, at 234 nm (indicating the formationof HPODs by LOX-1) up to 300 s.
The second test (T2) followed the formation of reaction products(HPODs), in phosphate buffer at pH 6.1. The protocol was similar to T1 (25C,at 234 nm, for all E1–E4 extracts).
For the third test (T3), the ketodiene formation was measured at 280 nmusing same E1–E4 extracts (x mL of each extract), 2.8-x phosphate bufferpH 7.1 and 0.2 mL LS solution. Each test was performed three times for eachextract.
The time course of the reaction was registered in each case, and theproduct formation was expressed by the ESA defined as the absorption varia-tion (DA234 for T1 and T2 and DA280 for T3) per time unit and milligrams protein(DA/s/mg protein). The ESA was related to the pHm and the E/LS ratio.
There were identified two types of shapes for the kinetic curves: a “con-ventional” shape containing an exponential phase followed by a “plateau” anda “nonconventional” shape containing three regions: an exponential “burst”phase (I), a lag phase (II) represented by an inhibition of the reaction rate andfinalized by a plateau (III).
For curves belonging to the first type, the ESA was calculated as shownearlier, while for the nonconventional curves, the ESA was calculated for eachphase (I, II or III).
RESULTS AND DISCUSSION
Kinetic Curves of Pure Soy LOX-1 (Test T0)
Figure 1 shows comparatively the progression curves (absorptionincrease in time, as indicator of 13-HPOD formation at 234 nm) for standardsolutions S1 and S2, which were prepared at different pH values (pHdil).
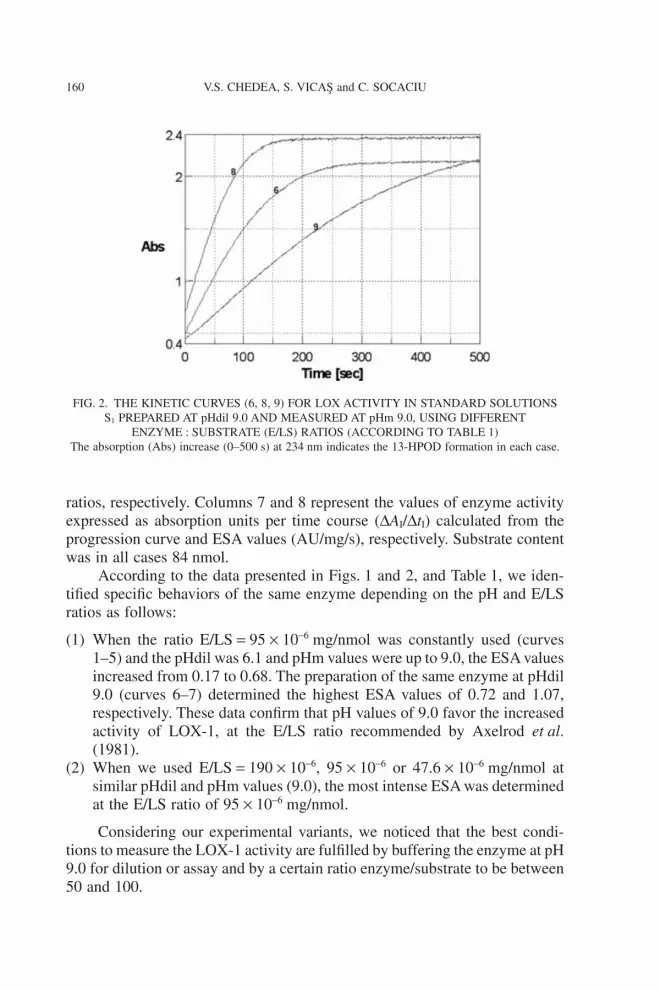
The numbers of the seven curves (column 1) correspond to differentprotocols of preparation (pHdil and pHm) as presented in Table 1 (columns 3and 4). Figure 2 represents the effect of E/LS ratio on the reaction kinetics(curves 6, 8 and 9), and the results are presented in Table 1.
The kinetic curves presented in Figs. 1 and 2 were considered “standardplots” for SLOX-1 activity. Both curves have a “conventional” shape (A)containing an exponential phase followed by a “plateau” phase. The calcula-tion of the specific enzyme activity was done according to the exponential“burst” phase (I).
158 V.S. CHEDEA, S. VICAS and C. SOCACIU
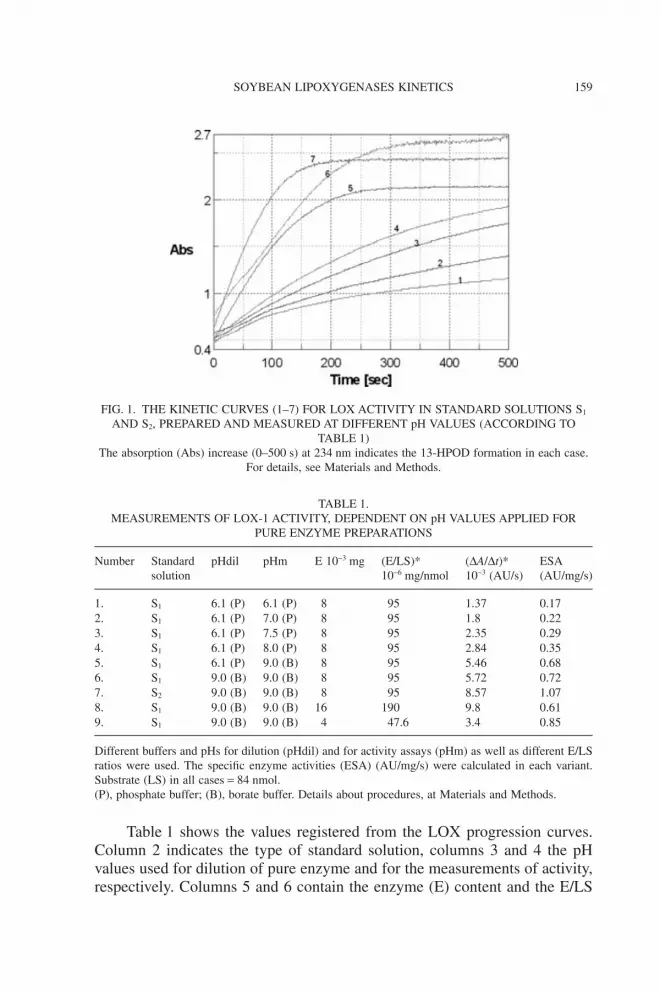
Table 1 shows the values registered from the LOX progression curves.Column 2 indicates the type of standard solution, columns 3 and 4 the pHvalues used for dilution of pure enzyme and for the measurements of activity,respectively. Columns 5 and 6 contain the enzyme (E) content and the E/LS
FIG. 1. THE KINETIC CURVES (1–7) FOR LOX ACTIVITY IN STANDARD SOLUTIONS S1
AND S2, PREPARED AND MEASURED AT DIFFERENT pH VALUES (ACCORDING TOTABLE 1)
The absorption (Abs) increase (0–500 s) at 234 nm indicates the 13-HPOD formation in each case.For details, see Materials and Methods.
TABLE 1.MEASUREMENTS OF LOX-1 ACTIVITY, DEPENDENT ON pH VALUES APPLIED FOR
PURE ENZYME PREPARATIONS
Number Standardsolution
pHdil pHm E 10-3 mg (E/LS)*10-6 mg/nmol
(DA/Dt)*10-3 (AU/s)
ESA(AU/mg/s)
1. S1 6.1 (P) 6.1 (P) 8 95 1.37 0.172. S1 6.1 (P) 7.0 (P) 8 95 1.8 0.223. S1 6.1 (P) 7.5 (P) 8 95 2.35 0.294. S1 6.1 (P) 8.0 (P) 8 95 2.84 0.355. S1 6.1 (P) 9.0 (B) 8 95 5.46 0.686. S1 9.0 (B) 9.0 (B) 8 95 5.72 0.727. S2 9.0 (B) 9.0 (B) 8 95 8.57 1.078. S1 9.0 (B) 9.0 (B) 16 190 9.8 0.619. S1 9.0 (B) 9.0 (B) 4 47.6 3.4 0.85
Different buffers and pHs for dilution (pHdil) and for activity assays (pHm) as well as different E/LSratios were used. The specific enzyme activities (ESA) (AU/mg/s) were calculated in each variant.Substrate (LS) in all cases = 84 nmol.(P), phosphate buffer; (B), borate buffer. Details about procedures, at Materials and Methods.
159SOYBEAN LIPOXYGENASES KINETICS
ratios, respectively. Columns 7 and 8 represent the values of enzyme activityexpressed as absorption units per time course (DAI/DtI) calculated from theprogression curve and ESA values (AU/mg/s), respectively. Substrate contentwas in all cases 84 nmol.
According to the data presented in Figs. 1 and 2, and Table 1, we iden-tified specific behaviors of the same enzyme depending on the pH and E/LSratios as follows:
(1) When the ratio E/LS = 95 ¥ 10-6 mg/nmol was constantly used (curves1–5) and the pHdil was 6.1 and pHm values were up to 9.0, the ESA valuesincreased from 0.17 to 0.68. The preparation of the same enzyme at pHdil9.0 (curves 6–7) determined the highest ESA values of 0.72 and 1.07,respectively. These data confirm that pH values of 9.0 favor the increasedactivity of LOX-1, at the E/LS ratio recommended by Axelrod et al.(1981).
(2) When we used E/LS = 190 ¥ 10-6, 95 ¥ 10-6 or 47.6 ¥ 10-6 mg/nmol atsimilar pHdil and pHm values (9.0), the most intense ESA was determinedat the E/LS ratio of 95 ¥ 10-6 mg/nmol.
Considering our experimental variants, we noticed that the best condi-tions to measure the LOX-1 activity are fulfilled by buffering the enzyme at pH9.0 for dilution or assay and by a certain ratio enzyme/substrate to be between50 and 100.
FIG. 2. THE KINETIC CURVES (6, 8, 9) FOR LOX ACTIVITY IN STANDARD SOLUTIONSS1 PREPARED AT pHdil 9.0 AND MEASURED AT pHm 9.0, USING DIFFERENT
ENZYME : SUBSTRATE (E/LS) RATIOS (ACCORDING TO TABLE 1)The absorption (Abs) increase (0–500 s) at 234 nm indicates the 13-HPOD formation in each case.
160 V.S. CHEDEA, S. VICAS and C. SOCACIU
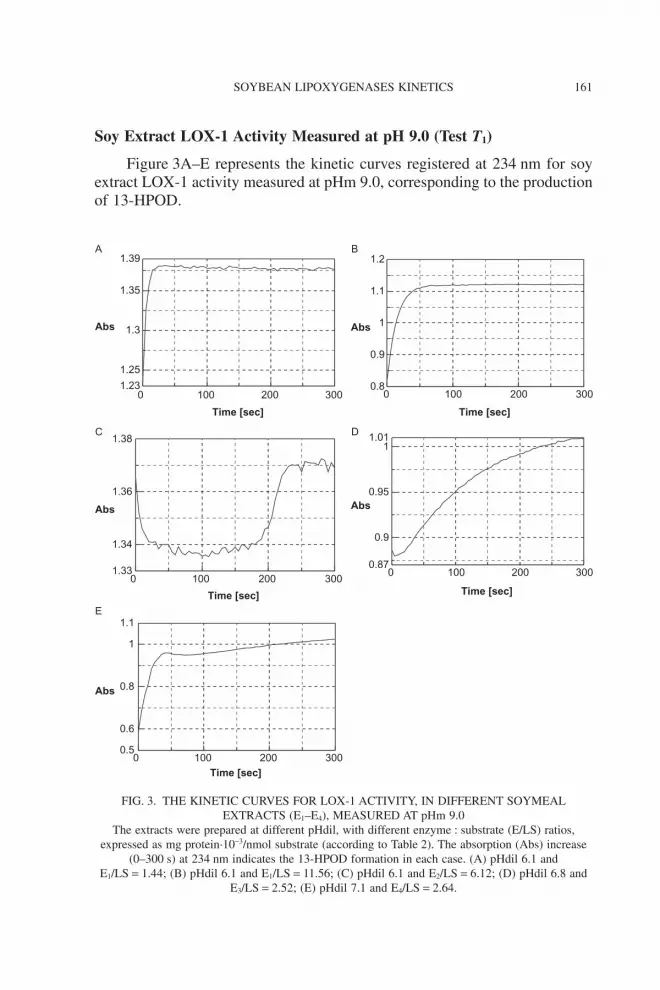
Soy Extract LOX-1 Activity Measured at pH 9.0 (Test T1)
Figure 3A–E represents the kinetic curves registered at 234 nm for soyextract LOX-1 activity measured at pHm 9.0, corresponding to the productionof 13-HPOD.
1.23
1.39A
C D
E
B
1.25
1.3
1.35
3000 100 200
Abs
Time [sec]
0.8
1.2
0.9
1
1.1
3000 100 200
Abs
Time [sec]
1.33
1.38
1.34
1.36
3000 100 200
Abs
Time [sec]
0.87
1.01
0.9
0.95
1
3000 100 200
Abs
Time [sec]
0.5
1.1
0.6
0.8
1
3000 100 200
Abs
Time [sec]
FIG. 3. THE KINETIC CURVES FOR LOX-1 ACTIVITY, IN DIFFERENT SOYMEALEXTRACTS (E1–E4), MEASURED AT pHm 9.0
The extracts were prepared at different pHdil, with different enzyme : substrate (E/LS) ratios,expressed as mg protein·10-3/nmol substrate (according to Table 2). The absorption (Abs) increase
(0–300 s) at 234 nm indicates the 13-HPOD formation in each case. (A) pHdil 6.1 andE1/LS = 1.44; (B) pHdil 6.1 and E1/LS = 11.56; (C) pHdil 6.1 and E2/LS = 6.12; (D) pHdil 6.8 and
E3/LS = 2.52; (E) pHdil 7.1 and E4/LS = 2.64.
161SOYBEAN LIPOXYGENASES KINETICS

Different enzymatic behaviors can be noticed in this test: for extracts E1
and E4 (Fig. 3A,B,E), the kinetic plots have a “conventional” shape, but for E2
and E3 (Fig. 3C,D), the curves show the three-phase type of curve pattern. Inthis later case, a lag phase (II) follows an initial burst (phase I) and wasassociated with a rapid decrease, and then the enzymatic activity increasesuntil it reaches a plateau (phase III). For E2 (Fig. 3C) can be observed a verylong lag phase II (170 s) after the initial burst I. The reaction starts again andreaches the plateau after 40 s. For E3 (Fig. 3D), the kinetic is again different,the lag and the initial burst being much shorter.
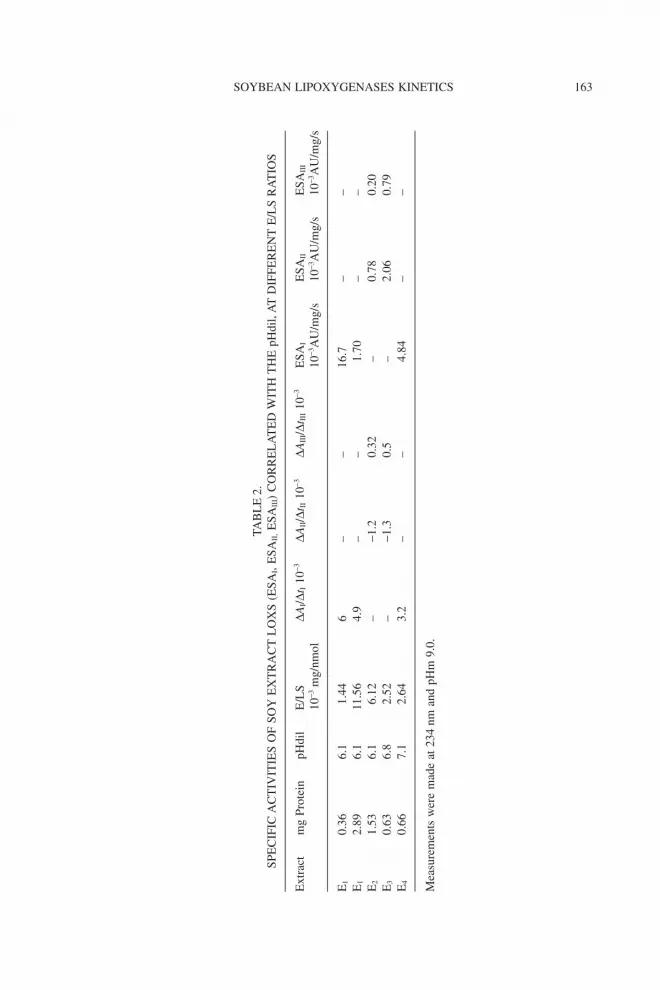
Table 2 presents comparatively the specific activities (ESAI, ESAII,ESAIII), corresponding to each phase of progression curve of soy extractLOX-1 (pHm 9.0), correlated with the pHdil and the different E/LS ratios.
For E1, a conventional shape of kinetic curve was obtained when thepHdil was 6.1 and the ratio E/LS increased from 1.44 to 11.56 ¥ 10-3 mgprotein/nmol the ESA decreased dramatically, around 10 times (from 16.7 to1.7 AU/mg/s). These data demonstrate that the low E/LS ratio favors thecatalytic reaction, while the excess of protein inhibits it. When the soybeanmeal was defatted and solved at pH 6.1 (E2), the enzymatic activity follows thee-phase curve where the lag period affects the total rate, the last phase (III)having only 0.2 AU/mg/s. When the defatted soybean meal was re-extracted inphosphate buffer of pH 4.5 (E3), the enzymatic rate ESAIII reached the value of0.79 AU/mg/s. At pH 7.1, the enzymatic activity of E4 also follows a conven-tional curve with an enzymatic rate of 4.84.
Soy Extract LOX-2 Activity, Registered at pH 6.1 (Test T2)
Figure 4A–F represents the kinetic curves registered at 234 nm for soyextract LOX activity measured at pHm 6.1, corresponding to the production ofa mixture of 13- and 9-HPOD, by favoring the LOX-2 catalysis, confirmed tobe at pH 6.8 (Loiseau et al. 2001).
Axelrod et al. (1981) proposes also this test (T2) for the identification ofLOX-2 isoenzyme. Also, the extraction procedure followed for E4 was the firststep in the isolation of LOX-2 and LOX-3 in the methodology of Axelrod et al.
Table 3 presents comparatively the specific activities (ESA, ESAI, ESAII,ESAIII) of soy extracts’ LOXs (pHm 6.1), correlated with the pHdil, at differentE/LS ratios.
Excepting the curve 4A, all the kinetic plots (4B–4F) follow the curvepattern B with lag phase. The enzymatic rate for E1 at pH 6.1 and E/LS = 2.88was 0.38 AU/mg/s. At slight decreases of E/LS, the curves turned to lag phasetype, and the rate became 0.17 and 0.56, respectively.
We can assume to have a critical threshold (ratio E/LS around 1.5) wherethe enzymatic behavior change.
162 V.S. CHEDEA, S. VICAS and C. SOCACIU
TAB
LE
2.SP
EC
IFIC
AC
TIV
ITIE
SO
FSO
YE
XT
RA
CT
LO
XS
(ESA
I,E
SAII
,E
SAII
I)C
OR
RE
LA
TE
DW
ITH
TH
EpH
dil,
AT
DIF
FER
EN
TE
/LS
RA
TIO
S
Ext
ract
mg
Prot
ein
pHdi
lE
/LS
10-3
mg/
nmol
DAI/D
t I10
-3DA
II/D
t II10
-3DA
III/D
t III
10-3
ESA
I
10-3
AU
/mg/
sE
SAII
10-3
AU
/mg/
sE
SAII
I
10-3
AU
/mg/
s
E1
0.36
6.1
1.44
6–
–16
.7–
–E
12.
896.
111
.56
4.9
––
1.70
––
E2
1.53
6.1
6.12
–-1
.20.
32–
0.78
0.20
E3
0.63
6.8
2.52
–-1
.30.
5–
2.06
0.79
E4
0.66
7.1
2.64
3.2
––
4.84
––
Mea
sure
men
tsw
ere
mad
eat
234
nman
dpH
m9.
0.
163SOYBEAN LIPOXYGENASES KINETICS
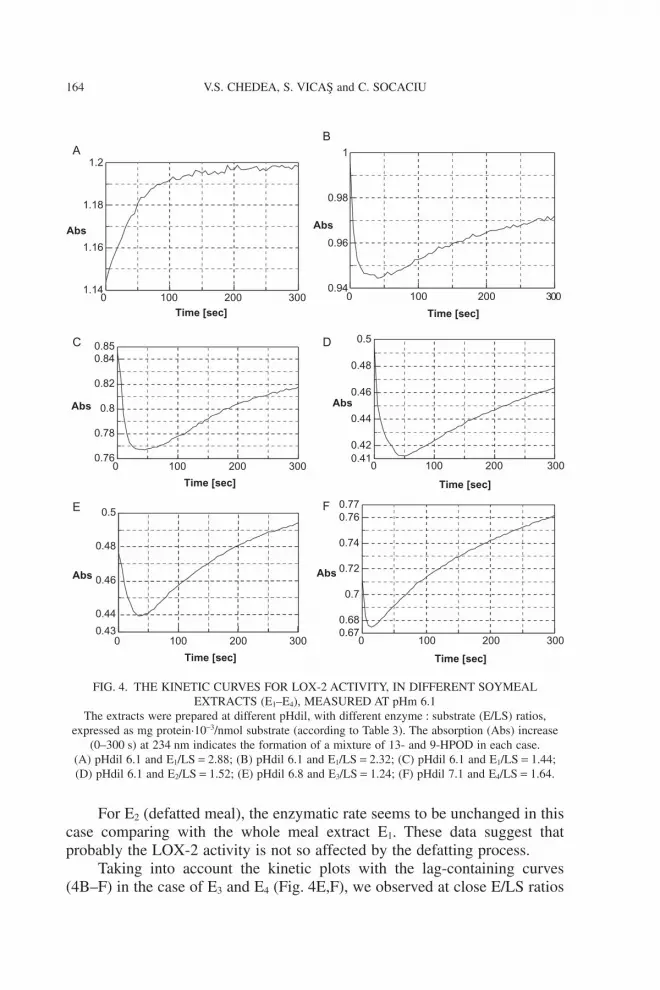
For E2 (defatted meal), the enzymatic rate seems to be unchanged in thiscase comparing with the whole meal extract E1. These data suggest thatprobably the LOX-2 activity is not so affected by the defatting process.
Taking into account the kinetic plots with the lag-containing curves(4B–F) in the case of E3 and E4 (Fig. 4E,F), we observed at close E/LS ratios
1.14
1.2A
C D
FE
B
1.16
1.18
3000 100 200
Abs
Time [sec]
0.94
1
0.96
0.98
0030 100 200
Abs
Time [sec]
0.76
0.85
0.78
0.8
0.82
0.84
3000 100 200
Abs
Time [sec]
0.41
0.5
0.42
0.44
0.46
0.48
3000 100 200
Abs
Time [sec]
0.43
0.5
0.44
0.46
0.48
3000 100 200
Abs
Time [sec]
0.67
0.77
0.68
0.7
0.72
0.74
0.76
3000 100 200
Abs
Time [sec]
FIG. 4. THE KINETIC CURVES FOR LOX-2 ACTIVITY, IN DIFFERENT SOYMEALEXTRACTS (E1–E4), MEASURED AT pHm 6.1
The extracts were prepared at different pHdil, with different enzyme : substrate (E/LS) ratios,expressed as mg protein·10-3/nmol substrate (according to Table 3). The absorption (Abs) increase
(0–300 s) at 234 nm indicates the formation of a mixture of 13- and 9-HPOD in each case.(A) pHdil 6.1 and E1/LS = 2.88; (B) pHdil 6.1 and E1/LS = 2.32; (C) pHdil 6.1 and E1/LS = 1.44;(D) pHdil 6.1 and E2/LS = 1.52; (E) pHdil 6.8 and E3/LS = 1.24; (F) pHdil 7.1 and E4/LS = 1.64.
164 V.S. CHEDEA, S. VICAS and C. SOCACIU

TAB
LE
3.SP
EC
IFIC
AC
TIV
ITIE
S(E
SA)
OF
SOY
EX
TR
AC
TL
OX
SC
OR
RE
LA
TE
DW
ITH
TH
EpH
dil,
AT
DIF
FER
EN
TE
/LS
RA
TIO
S
Ext
ract
pHdi
lm
gPr
otei
n(m
g)E
/LS
mg
10-3
/nm
olDA
I/Dt I
10-3
DAII/D
t II10
-3DA
III/D
t III
10-3
ESA
I
10-3
AU
/mg/
sE
SAII
10-3
AU
/mg/
sE
SAII
I
10-3
AU
/mg/
s
E1
6.1
0.72
2.88
0.27
––
0.38
––
E1
6.1
0.58
2.32
–-1
.10.
1–
-1.8
90.
17E
16.
10.
361.
44–
-2.2
0.2
–-6
.11
0.56
E2
6.1
0.38
1.52
–-2
0.2
–-5
.26
0.53
E3
6.8
0.31
1.24
–-0
.28
0.08
–-0
.90.
26E
47.
10.
411.
64–
-2.5
0.3
–-6
.09
0.73
Mea
sure
men
tsw
ere
mad
eat
234
nman
dpH
m6.
1(H
POD
sfo
rmat
ion)
.
165SOYBEAN LIPOXYGENASES KINETICS
differences between enzymatic rates, the E4 having more enzyme activity,possibly because of the pHdil 7.1 and E/LS = 1.64 closer of the optimumconditions for LOX-2.
In our experiments, the best ESA was identified in E4 extract, with a betteractivity than E1 at almost half value of E/LS ratio. As we cannot certify thepresence of a specific isoenzyme in our extracts, we can suppose that in thiscase more LOX-2 and LOX-3 than LOX-1 were extracted. It also can beremarked that the neutral extraction (E4) of defatted soy meal determined athree times increase of enzyme activity comparing with acidic extraction (E3).
Ketodiene Formation by Soy LOX-3 at pH 7.1 (Test T3)
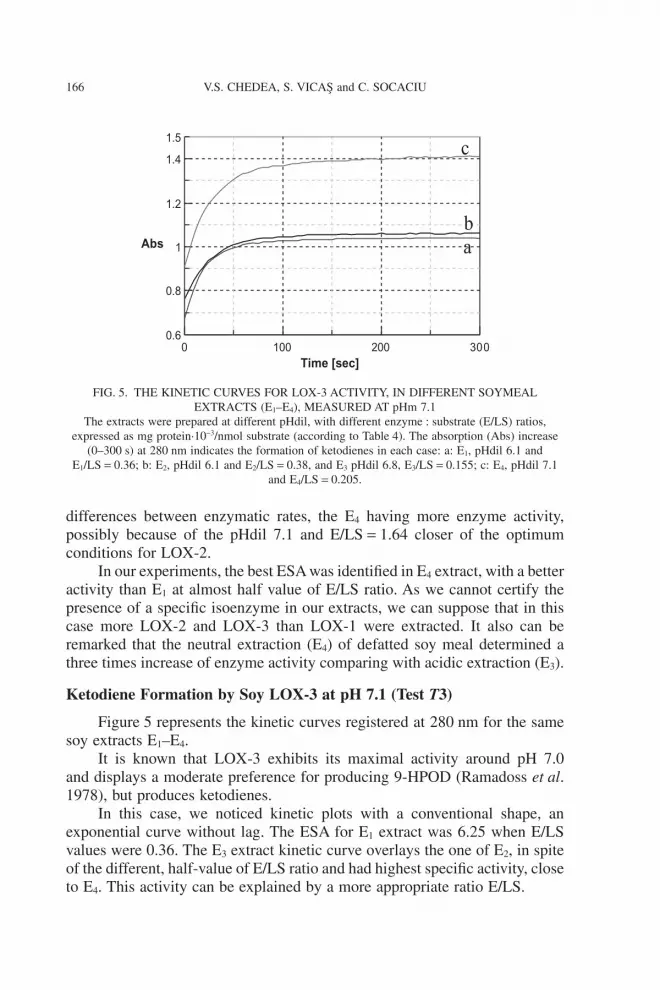
Figure 5 represents the kinetic curves registered at 280 nm for the samesoy extracts E1–E4.
It is known that LOX-3 exhibits its maximal activity around pH 7.0and displays a moderate preference for producing 9-HPOD (Ramadoss et al.1978), but produces ketodienes.
In this case, we noticed kinetic plots with a conventional shape, anexponential curve without lag. The ESA for E1 extract was 6.25 when E/LSvalues were 0.36. The E3 extract kinetic curve overlays the one of E2, in spiteof the different, half-value of E/LS ratio and had highest specific activity, closeto E4. This activity can be explained by a more appropriate ratio E/LS.
0.6
1.5
0.8
1
1.2
1.4
0030 100 200
Abs
Time [sec]
ab
c
FIG. 5. THE KINETIC CURVES FOR LOX-3 ACTIVITY, IN DIFFERENT SOYMEALEXTRACTS (E1–E4), MEASURED AT pHm 7.1
The extracts were prepared at different pHdil, with different enzyme : substrate (E/LS) ratios,expressed as mg protein·10-3/nmol substrate (according to Table 4). The absorption (Abs) increase
(0–300 s) at 280 nm indicates the formation of ketodienes in each case: a: E1, pHdil 6.1 andE1/LS = 0.36; b: E2, pHdil 6.1 and E2/LS = 0.38, and E3 pHdil 6.8, E3/LS = 0.155; c: E4, pHdil 7.1
and E4/LS = 0.205.
166 V.S. CHEDEA, S. VICAS and C. SOCACIU

Table 4 presents comparatively the specific activities (ESA) of soy extractLOXs (pHm 7.1), correlated with the pHdil, at different E/LS ratios. In spiteof low E/LS ratios, the enzymatic activity was higher than for experiments intest T2. In this test, it can be observed that the highest ESA is linked to thelowest value of E/LS, and the more the E/LS increases, the ESA decreases. TheE/LS value of 0.31 reached maximum activities of 10.64 AU/mg/s.
In the model of Axelrod et al. (1981), this test was performed specificallyfor LOX-3 whose presence is identified by the ketodiene formation, as mea-sured at 280 nm. In this case also, the deffating of extract (E2) does not affectthe ketodiene presence as much as the HPODs. We can conclude that we havein E3 an extract rich in LOX-3, and the defatting stimulates the LOX-3 activity.The second ESA is the one of E4 (6.58) at an E/LS = 0.205, and let us remarkthat E4 contains LOX-3 as well.
Relationships Between the pHm, E/LS and LOX Isoenzyme Activity
The kinetic curves at different pHm (6.1, 7.1 and 9.0) aimed the specificactivation of LOX-1, LOX-2 and LOX-3, respectively. Figure 6 shows thegeneral picture of the relationships between the ESA, E/LS and pHm values,for pure LOX or different extracts (E1–E4).
We confirmed some of the previous data regarding the dependence ofLOX isoenzyme activity on the pHm (Siedow 1991; Loiseau et al. 2001;Kosáry et al. 2004).
The influence of pH on the activity of SLOX (soybean LOX-1) as shownin Fig. 6 may be explained by the fact that at different pHm, the catalytic situsof LOX isoenzymes has different conformations and iron oxidation states(Gardner 1989).
Considering our data obtained from T0 (pure enzyme), we could assumethat the best conditions to measure the LOX activity is to use a pHdil and apHm of 9.0 (in 0.2 M borate buffer) at E/LS = 0.095 ¥ 10-3. When E/LS ratios
TABLE 4.SPECIFIC ACTIVITIES OF SOY EXTRACT LOXs (ESA) CORRELATED WITH THE pHdil, AT
DIFFERENT E/LS RATIOS
Extract pHdil mg Protein E/LS mg·10-3/nmol DA280/Dt 10-3 ESAI 10-3AU/mg/s
E1 6.1 0.72 0.36 4.5 6.25E2 6.1 0.76 0.38 3.3 4.34E3 6.8 0.31 0.155 3.3 10.64E4 7.1 0.41 0.205 2.7 6.58
Measurements were made at 280 nm and pH 7.1 (ketodiene formation).
167SOYBEAN LIPOXYGENASES KINETICS
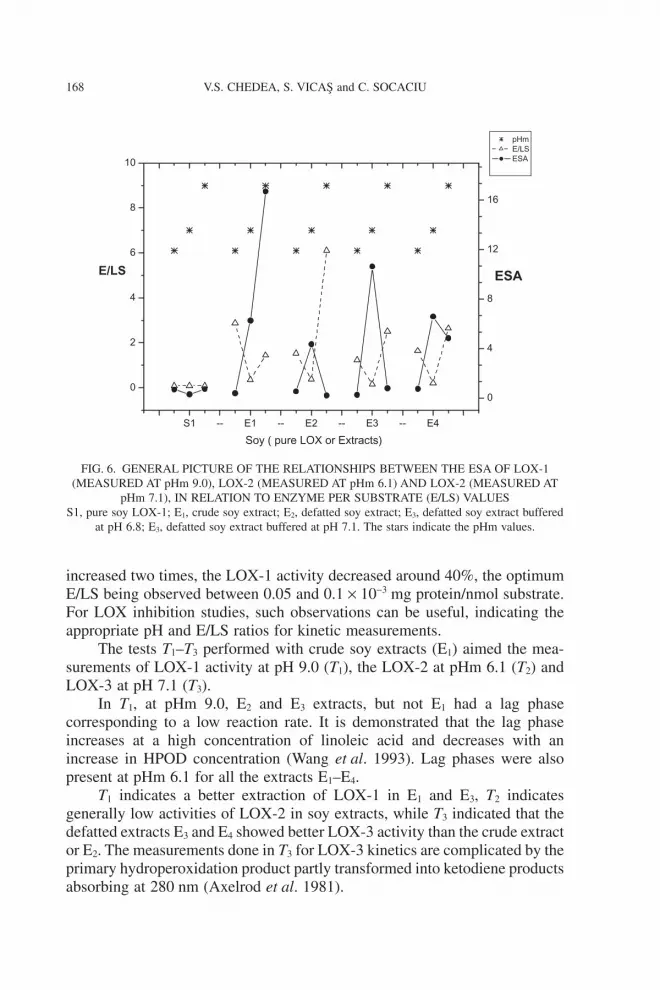
increased two times, the LOX-1 activity decreased around 40%, the optimumE/LS being observed between 0.05 and 0.1 ¥ 10-3 mg protein/nmol substrate.For LOX inhibition studies, such observations can be useful, indicating theappropriate pH and E/LS ratios for kinetic measurements.
The tests T1–T3 performed with crude soy extracts (E1) aimed the mea-surements of LOX-1 activity at pH 9.0 (T1), the LOX-2 at pHm 6.1 (T2) andLOX-3 at pH 7.1 (T3).
In T1, at pHm 9.0, E2 and E3 extracts, but not E1 had a lag phasecorresponding to a low reaction rate. It is demonstrated that the lag phaseincreases at a high concentration of linoleic acid and decreases with anincrease in HPOD concentration (Wang et al. 1993). Lag phases were alsopresent at pHm 6.1 for all the extracts E1–E4.
T1 indicates a better extraction of LOX-1 in E1 and E3, T2 indicatesgenerally low activities of LOX-2 in soy extracts, while T3 indicated that thedefatted extracts E3 and E4 showed better LOX-3 activity than the crude extractor E2. The measurements done in T3 for LOX-3 kinetics are complicated by theprimary hydroperoxidation product partly transformed into ketodiene productsabsorbing at 280 nm (Axelrod et al. 1981).
S1 -- E1 -- E2 -- E3 -- E4
0
2
4
6
8
10
pHm E/LS ESA
ESAE/LS
Soy ( pure LOX or Extracts)
0
4
8
12
16
FIG. 6. GENERAL PICTURE OF THE RELATIONSHIPS BETWEEN THE ESA OF LOX-1(MEASURED AT pHm 9.0), LOX-2 (MEASURED AT pHm 6.1) AND LOX-2 (MEASURED AT
pHm 7.1), IN RELATION TO ENZYME PER SUBSTRATE (E/LS) VALUESS1, pure soy LOX-1; E1, crude soy extract; E2, defatted soy extract; E3, defatted soy extract buffered
at pH 6.8; E3, defatted soy extract buffered at pH 7.1. The stars indicate the pHm values.
168 V.S. CHEDEA, S. VICAS and C. SOCACIU

CONCLUSIONS
Our experiments aimed practical purposes, namely the conditions thatcan keep the stability or inhibit the soybean LOX activity. Considering thepreferences of each soy LOX isoenzyme for specific pHs for their optimumactivity, we tested the effects of pH at different stages of extraction, dilution ormeasurements of enzyme activity. We tested as well the influence of enzyme/substrate ratio on the enzyme activity, using linoleic acid as unique substrate.As for other LOXs, we found out that LOX-1 is most active at pHm 9.0, thatLOX-2 is more active at pHm 6.8 and LOX-3 at pHm 7.1.
The experiences done with pure soybean LOX-1 and soybean extracts(with or without defatting) showed different types of kinetic curves. Thecurves corresponding to pure soy LOX-1 were always of “conventional” type,with an exponential increase followed by a plateau. For crude soy extracts E1,the reaction progression curve had also a conventional shape. For defattedextracts (E2–E4), the curves showed a three-phase type of curve: a very rapidexponential phase (I) followed by an inhibition and lag period (II) and finally,another increase up to a plateau (III). As can be seen in this type of curves, theexponential phase cannot be seen; it happens so rapidly (approximately 15 s)before spectrometry measurements. This time was necessary for the additionand homogenization of the substrate with the enzyme in the spectrometercuvette.
Our experimental observations allow the following conclusions:
(1) The crude soy extract E1 contains mainly LOX-1 isoenzyme, but alsosome LOX-3, in an approximate ratio of 3:1. The LOX-2 in this extract isvery low.
(2) The defatting of E1 inhibited almost completely the LOX-1 activity atacidic pHs. It is rational that defatting eliminates the substrate (linoleate)found in the crude extract, and this is a simple mechanistic effect onLOX-1 inhibition. By reestablishing a neutral or an alkaline pH, a part ofenzyme activity was recovered, a fact which can be explained by theoxidative conversion of Fe2+ to Fe3+, found in active LOX-1.
(3) The three defatted extracts (E2, E3 and E4) buffered at pHs 6.8 and 7.1showed, generally, a similar picture of their LOX activity: low LOX-1 andLOX 2 activity, but mostly LOX-3 activity, for E3 even more intense thanfor E1.
(4) The bilateral dependence of LOX-1 activity on pHm and E/LS suggestspossibilities to modulate this enzyme activity by modifications of theseparameters.
(5) For extracts E2–E4, a mirror-type relation was observed between E/LS andESA. To inhibit LOX activity, high E/LS and low pHm are recommended.
169SOYBEAN LIPOXYGENASES KINETICS

Further studies are necessary to investigate a specific inhibition of soyLOX-1, which is the major isoenzyme in crude soy extracts. Defatting and lowpHs as well as different antioxidants with inhibitory effect on LOX activitywill be investigated.
NOMENCLATURE
ESA enzymatic specific activityHPODs hydroperoxy-octadecadienoic acids9-HPOD 9-hydroperoxy-10E,12Z-octadecadienoic acid13-HPOD 13-hydroperoxy-9Z,11E-octadecadienoic acidLOX lipoxygenaseLS sodium linoleate
REFERENCES
AXELROD, B., CHEESBROUGH, T.M. and LAASKO, S. 1981. Lipoxyge-nase from soybeans. Methods Enzymol. 71, 441–444.
BERK, Z. 1992. Technology of production of edible flours and proteinproducts from soybeans. FAO Agricultural Services Bulletin 97. http://www.fao.org/docrep/t0532e/t0532e01.htm (accessed August 12, 2005).
BORRELLI, G.M., TROCCOLI, A., FONZO, N.D.I. and FARES, C. 1999.Durum wheat lipoxygenase activity and other quality parameters thataffect pasta color. Cereal Chem. 76(3), 335–340.
CASEY, R. and HUGHES, R.K. 2004. Recombinant lipoxygenases and oxy-lipin metabolism in relation to food quality. Food Biotechnol. 18, 135–170.
CROFT, K.P.C., JUTTNER, F. and SLUSARENKO, A.J. 1993. Volatile prod-ucts of the lipoxygenase pathway evolved from Phaseolus vulgaris (L.)leaves inoculated with Pseudomonas syringae pv-Phaseolicola. PlantPhysiol. 101, 13–24.
CUMBEE, B., HILDEBRAND, D.F. and ADDO, K. 1997. Soybean flourlipoxygenase isozymes effects on wheat flour dough rheological andbreadmaking properties. J. Food Sci. 62, 281–283.
DE GROOT, J.J., VELDINK, G.A., VLIEGENTHART, J.F., BOLDINGH, J.,WEVER, R. and VAN GELDER, B.F. 1975. Demonstration by EPRspectroscopy of the functional role of iron in soybean lipoxygenase-1.Biochim. Biophys. Acta 377, 71–79.
FEUSSNER, I. and WASTERNACK, C. 2002. The lipoxygenase pathway.Annu. Rev. Plant Biol. 53, 275–297.
FUKUSHIMA, D. 1994. Recent progress on biotechnology of soybean pro-teins and soybean protein food products. Food Biotechnol. 8, 83–135.
170 V.S. CHEDEA, S. VICAS and C. SOCACIU

FUNK, C.D. 1996. The molecular biology of mammalian lipoxygenases andthe quest for eicosanoid functions using lipoxygenase deficient mice.Biochim. Biophys. Acta 1304, 65–84.
GARDNER, H.W. 1989. Soybean lipoxygenase enzymatically forms both9(S)- and 13(S)-hydroperoxides from linoleic acid by a pH dependentmechanism. Biochim. Biophys. Acta 1001, 274–281.
GARDNER, H.W. 1991. Recent investigations into the lipoxygenase pathwayin plants. Biochim. Biophys. Acta 1084, 221–239.
GORNALL, A., CHARLES, G., BARDAWILL, J. and MAXIMA, M.D.1949. Determination of serum proteins by means of the biuret reaction.J. Biol. Chem 177, 751–766.
GRECHKIN, A. 1998. Recent developments in biochemistry of the plantlipoxygenase pathway. Prog. Lipid Res. 37, 317–352.
HATANAKA, A. 1996. The fresh green odor emitted by plants. Food Rev. Int.12, 303–350.
IVANOV, I., RATHMANN, J., MYAGKOVA, G. and KÜHN, H. 2001.Soybean lipoxygenase-1 oxygenates synthetic polyenoic fatty acids withan altered positional specificity. Evidence for inverse substrate alignment.Biochemistry 40, 10223–10229.
KOBAYASHI, N., KANEDA, H., KANO, Y. and KOSHINO, S. 1993. Theproduction of linoleic and linolenic acid hydroperoxides during mashing.J. Ferment. Bioeng. 76, 371–375.
KOSÁRY, J., KORBÁSZ, M., KISS, I. and BALOGH, T. 2004. Almondlipoxygenases. Proceedings of the 16th International Plant Lipid Sympo-sium, June 1–4, Budapest, Hungary.
KULKARNI, A.P. 2001. Lipoxygenase – a versatile biocatalyst for biotrans-formation of endobiotics and xenobiotics. Cell. Mol. Life Sci. 58, 1805–1825.
LARRETA-GARDE, V. 1995. Lipoxygenase in making breads, cookies andcrackers. Oléagineux Corps Gras Lipides 2, 363–365.
LOISEAU, J., BENOÎT, L.V., MACHEREL, M.-H. and LE DEUNFF, Y. 2001.Seed lipoxygenases: Occurrence and functions. Seed Sci. Res. 11, 199–211.
MACCARRONE, M., VAN AARIE, P.G.M., VELDINK, G.A. andVLIEGENTHART, J.F.G. 1994. In vitro oxygenation of soybean bio-membranes by lipoxygenase-2. Biochim. Biophys. Acta 1190, 164–169.
MACK, A.J., PETERMAN, T.K. and SIEDOW, J.N. 1987. Lipoxygenaseisozymes in higher plants: Biochemical properties and physiological role.Curr. Top. Biol. Med. Res. 13, 127–154.
MOGUL, R. and HOLMAN, T.R. 2001. Inhibition studies of soybean andhuman 15-Lipoxygenases with long-chain alkenyl sulfate substrates.Biochemistry 40, 4391–4397.
171SOYBEAN LIPOXYGENASES KINETICS

NICOLAS, J. and POTUS, J. 1994. Enzymatic oxidation phenomena andcoupled oxidations – effects of lipoxygenase in bread making and ofpolyphenol oxidase in fruit technology. Sci. Aliments 14, 627–642.
PORTA, H. and ROCHA-SOSA, M. 2002. Plant lipoxygenases. Physiologicaland molecular features. Plant Physiol. 130, 15–21.
RADMARK, O. 2002. Arachidonate 5-lipoxygenase. Prostaglandins OtherLipid Mediat. 68–69, 211–234.
RAMADOSS, C.S., PISTORIUS, E.K. and AXELROD, B. 1978. Coupledoxidation of carotene by lipoxygenase requires two isoenzymes. Arch.Biochem. Biophys. 190, 549–552.
ROBINSON, D.S., WU, Z.C., DOMONEY, C. and CASEY, R. 1995. Lipoxy-genases and the quality of foods. Food Chem. 54, 33–43.
SADIK, C.D., SIES, H. and SCHEWE, T. 2003. Inhibition of15-lipoxygenases by flavonoids: Structure–activity relations and mode ofaction. Biochem. Pharmacol. 65, 773–781.
SCHILSTRA, M.J., VELDINK, G.A., VERHAGEN, J. and VLIEGENT-HART, J.F.G. 1992. Effect of lipid hydroperoxide on lipoxygenase kinet-ics. Biochemistry 31, 7692–7699.
SEMBDNER, G. and PARTHIER, B. 1993. The biochemistry and the physi-ological and molecular actions of jasmonates. Annu. Rev. Plant Physiol.Plant Mol. Biol. 44, 569–589.
SIEDOW, J.N. 1991. Plant lipoxygenases. Structure and function. Annu. Rev.Plant Physiol. Plant Mol. Biol. 42, 145–188.
SKRZYPCZAK-JANKUN, E., ZHOU, K., MCCABE, N.P., SELMAN, S.H.and JANKUN, J. 2003. Structure of curcumin in complex with lipoxy-genase and its significance in cancer. Int. J. Mol. Med. 12, 17–24.
STASWICK, P.E. 1992. Jasmonate, genes and fragrant signals. Plant Physiol.99, 804–807.
WANG, Z.-X., DEREK KILLILEA, S. and SRIVASTAVA, D.K. 1993. Kineticevaluation of substrate-dependent origin of the lag phase in soybeanlipoxygenase-1 catalyzed reactions. Biochemistry 32, 1500–1509.
YAMAMOTO, S., SUZUKI, H. and UEDA, N. 1997. Arachidonate 12-lipoxygenases. Prog. Lipid Res. 36, 23–41.
ZIMMERMAN, D.C. and COUDRON, C.A. 1979. Identification of traumatin,a wound hormone, as 12-oxo-trans-10-dodecenoic acid. Plant Physiol.63, 536–541.
172 V.S. CHEDEA, S. VICAS and C. SOCACIU
Related Documents











