KARAKTERISASI DAN POTENSI JAMUR ENDOFIT DAN SAPROFIT ANTAGONISTIK SEBAGAI AGENS PENGENDALI HAYATI JAMUR Fusarium oxysporum f. sp. vanillae PADA TANAMAN VANILI DI PULAU LOMBOK NTB RINGKASAN DISERTASI Oleh I MADE SUDANTHA NIM 0430100012 PROGRAM DOKTOR ILMU PERTANIAN KEKHUSUSAN HAMA DAN PENYAKIT TANAMAN PROGRAM PASCASARJANA FAKULTAS PERTANIAN UNIVERSITAS BRAWIJAYA MALANG 2007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

KARAKTERISASI DAN POTENSI JAMUR ENDOFIT DAN SAPROFIT ANTAGONISTIK SEBAGAI AGENS PENGENDALI HAYATI JAMUR
Fusarium oxysporum f. sp. vanillae PADA TANAMAN VANILI DI PULAU LOMBOK NTB
RINGKASAN DISERTASI
Oleh
I MADE SUDANTHA
NIM 0430100012
PROGRAM DOKTOR ILMU PERTANIAN
KEKHUSUSAN HAMA DAN PENYAKIT TANAMAN
PROGRAM PASCASARJANA FAKULTAS PERTANIAN
UNIVERSITAS BRAWIJAYA
MALANG
2007

2
Tim Penguji
JUDUL DISERTASI:
Karakterisasi dan Potensi Jamur Endofit dan Saprofit Antagonistik Sebagai Agens Pengendali Hayati Jamur Fusarium oxysporum f. sp. vanillae Pada Tanaman Vanili di
Pulau Lombok NTB
Nama Mahasiswa : I Made Sudantha NIM : 0430100012 Program Doktor : Ilmu Pertanian Kekhususan : Hama dan Penyakit Tanaman KOMISI PROMOTOR:
Promotor : Prof. Dr. Ir. Tutung Hadiastono, MS. Ko-Promotor : Prof. Dr. Ir. Abdul Latief Abadi, MS. Ko-Promotor : Dr. Ir. Syamsuddin Djauhari, MS. TIM DOSEN PENGUJI:
Dosen Penguji 1 : Prof. Dr. Ir. Siti Rasminah Ch. Sy. Dosen Penguji 2 : Prof. Dr. Ir. Soemarno, MS. Dosen Penguji 3 : Prof. (Riset) Dr. Ir. Nasir Saleh, APU. Dosen Penguji Luar : Prof. (Riset) Dr. Ir. Gatot Kartono, APU. Tanggal Seminar : 1 Februari 2007 Tanggal Ujian Tertutup : 16 Maret 2007 Tanggal Ujian Terbuka : 2 April 2007

3
Karakterisasi dan Potensi Jamur Endofit dan Saprofit Antagonistik Sebagai Agens Pengendali Hayati Jamur Fusarium oxysporum f. sp.
vanillae Pada Tanaman Vanili di Pulau Lombok NTB
I Made Sudantha
ABSTRAK
Penelitian bertujuan untuk mengetahui karakteristik jamur endofit dan saprofit antagonis pada tanaman vanili; pengaruh pH medium, suhu inkubasi dan seresah daun terhadap pertumbuhannya; mekanisme antagonismenya terhadap jamur F. oxysporum f. sp. vanillae; efektivitasnya dalam mengendalikan penyakit busuk batang; sinergismenya dalam mengendalikan penyakit busuk batang dan potensinya dalam meningkatkan ketahanan induksi tanaman vanili terhadap penyakit busuk batang.
Penelitian menggunakan metode deskriptif dan eksperimental. Metode deskriptif dilaksanakan melalui survei dan eksplorasi di lapang yang dilakukan di enam lokasi kebun vanili di Pulau Lombok yaitu Timbenuh, Banok, Selebung, Jurang Malang, Lingsar dan Celelos. Metode eksperimental dilaksanakan di Laboratorium Proteksi Tumbuhan, Rumah Plastik dan Kebun Percobaan Fakultas Pertanian Universitas Mataram. Percobaan di laboratorium meliputi: karakterisasi jamur endofit dan saprofit; uji pertumbuhan jamur endofit dan saprofit pada berbagai pH medium, suhu inkubasi dan serersah daun; uji antagonisme dan uap biakan jamur endofit dan saprofit terhadap F. oxysporum f. sp. vanillae. Percobaan di rumah plastik meliputi: uji efektivitas jamur endofit dan cara aplikasi terhadap penyakit busuk batang vanili, uji efektivitas jamur saprofit dan cara aplikasi terhadap penyakit busuk batang vanili, uji sinergisme jamur endofit dan saprofit terhadap penyakit busuk batang vanili, dan uji sinergisme jamur endofit dan saprofit vanili terhadap penyakit busuk batang pada beberapa klon vanili. Percobaan di kebun percobaan meliputi: uji sinergisme jamur endofit dan saprofit terhadap penyakit busuk batang pada beberapa klon vanili di pembibitan dan di kebun.
Hasil penelitian menunjukan bahwa: (1) Pada tanaman sehat ditemukan 19
isolat jamur endofit dan pada rhizosfer ditemukan 19 isolat jamur saprofit dengan karakteristik yang berbeda. (2) Jamur endofit dan saprofit yang ditemukan tumbuh
dengan baik pada suhu optimum 25o C dan pH optimum 6,0, kedua kelompok jamur ini dapat tumbuh dengan baik pada seresah daun kopi, kemiri, lamtoro dan gamal. (3)
Terdapat delapan isolat jamur endofit dan 12 isolat jamur saprofit efektif menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae secara in-vitro. Penghambatan pertumbuhan oleh jamur endofit dan saprofit dengan cara fisik (kompetisi ruang dan mikoparasit) dan antibiosis (mengeluarkan antibiotik yang mudah menguap). (4) Penggunaan lima isolat jamur endofit yaitu Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum), Trichoderma sp. ENDO-05 batang Selebung (T. pseudokoningii), Trichoderma sp. ENDO-06 batang Celelos (T. viride), dan Rhizoctonia sp. ENDO-07 batang Timbenuh dengan cara perendaman stek vanili dengan suspensi spora dan

4
infestasi substrat yang mengandung spora ke medium tanah menyebabkan bibit vanili tidak terinfeksi penyakit busuk batang. Demikian pula penggunaan 10 isolat jamur saprofit menyebabkan bibit vanili tidak terinfeksi penyakit busuk batang, empat diantaranya dapat memacu pembungaan lebih awal yaitu Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum) dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum). (5) Penggunaan tiga isolat jamur endofit yaitu Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang (T. polysporum ) dan Trichoderma sp. ENDO-06 batang Celelos (T. viride) dapat bersinergisme dengan lima isolat jamur saprofit yaitu Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-05 vanili Celelos (T. piluliferum), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum), dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum),
menyebabkan bibit dan tanaman vanili baik klon Timbenuh NTB, klon Celelos NTB maupun klon Malang Jawa Timur tidak terinfeksi penyakit busuk batang, sehingga dapat meningkatkan ketahanan induksi terhadap penyakit busuk batang.
ABSTRACT
This research was aimed to characterize antagonistic endophytic and saprophytic fungi found on vanilla plants; to know the effects of medium pH, incubation temperature and leaf manures on their growth; to characterize their antagonistic mechanisms against F. oxysporum f. sp. vanillae fungus; to measure their effectiveness in controlling vine rot
disease; to mesuare levels of their synergism in controlling vine rot disease and their potentials in improving induced resistance on vanilla plants to vine rot disease.
This research applied both descriptive and experimental methods. Descriptive methods were applied through surveys and field exploration. Surveys were conducted in six locations of vanilla plantation on Lombok island, including Timbenuh, Banok, Selebung, Jurang Malang, Lingsar dan Celelos. Experimental methods were applied to the experiments conducted at the Plant Protection Lab., plastic house and experiment station of the Faculty of Agriculture, University of Mataram. Lab experiments consisted of characterization of endophytic and saprophytic fungi; growth test for the endophytic and saprophytic fungi on medium varying in pH levels, incubation temperatures and on various types of leaf manures; test of antagonism and medium favor of the endophytic and saprophytic fungi against F. oxysporum f. sp. Vanillae. The plastic house experiments consisted of test of endophytic fungi effectiveness and their application techniques on vine rot disease; test of saprophytic fungi effectiveness and their application techniques on vine rot disease synergism test of endophytic and saprophytic fungal on vine rot disease; and synergism test of endophytic and saprophytic fungal to the vanilla vine rot disease on vanilla clones. Experiments on the experiment station consisted of synergism test of endophytic and saprophytic fungal to vanilla vine rot disease on vanilla clones in a nursery and in plantation farm.

5
Results indicated that: (1) In the healthy vanilla are found 19 isolates of
endophytic fungi and in the rhizospere are found 19 isolates of saprophytic fungi with different characteristics. (2) Endophytic and saprophytic fungi need optimum
temperature 25o C and optimum pH 6,0 to ideal growth, and booth of them can good growth on leaf manures of coffee, Aleurites, Leucaena and Glyricida. (3) There are eight isolate endophytic fungi and 12 isolate saprophytic fungi effective in suppressing F. oxysporum f. sp. vanillae in-vitro. Growth obstruction by endophytic and saprophytic fungi used were physical competition (in the forms of spatial competition and as myco-parasites) and antibiosis (through exudation of favorable antibiotics). (4) Treatments with five isolates of endophytic fungi as Trichoderma sp. ENDO-02 vine Timbenuh (T. koningii), Trichoderma sp. ENDO-04 vine Jurang Malang (T. polysporum), Trichoderma sp. ENDO-05 vine Selebung (T. pseudokoningii), Trichoderma sp. ENDO-06 vine Celelos (T. viride), dan Rhizoctonia sp. ENDO-07 vine Timbenuh either through soaking of vanilla cuttings or through soil infestation caused the vanilla cuttings uninfected by vine rot disease. Thus, treatments with 10 isolates of saprophytic fungi caused the vanilla cuttings uninfected by vine rot disease, among four isolates that surprisingly induced early flowering , i.e. Trichoderma sp. SAPRO-03 vanilla Timbenuh (T. harzianum), Trichoderma sp. SAPRO-07 vanilla Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanilla Lingsar (T. hamatum) dan Trichoderma sp. SAPRO-11 vanilla Selebung (T. hamatum). (5) Treatments three isolates endophytic fungi as Trichoderma sp. ENDO-02 vine Timbenuh (T. koningii), Trichoderma sp. ENDO-04 vine Jurang (T. polysporum ) dan Trichoderma sp. ENDO-06 vine Celelos (T. viride) can sinergism with five isolates saprophytic fungi as Trichoderma sp. SAPRO-03 vanilla Timbenuh (T. harzianum), Trichoderma sp. SAPRO-05 vanilla Celelos (T. piluliferum), Trichoderma sp. SAPRO-07 vanilla Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanilla Lingsar (T. hamatum), dan Trichoderma sp. SAPRO-11 vanilla Selebung (T. hamatum), causing vanilla
cuttings and plants Timbenuh NTB clones, Celelos NTB clones and Malang East Java clones uninfected by vine rot disease, so can improving induced resistance to vine rot disease.
I. PENDAHULUAN
1.1. Latar Belakang
Daerah pengembangan dan produksi vanili (Vanilla planifolia Andrews) di
Indonesia saat ini adalah Sumatera Utara, Lampung, Jawa Barat, Jawa Timur, Bali, Nusa Tenggara Barat, Nusa Tenggara Timur, Sulawesi Utara, Sulawesi Tengah, dan Sulawesi Selatan. Data dari Ditjen Perkebunan Deptan tahun 2001 luas areal penanaman vanili di Indonesia sekitar 15.937 ha. Diperkirakan saat ini luas areal penanaman vanili tinggal 50 % dan sebagaian besar dalam keadaan rusak dan kurang produktif, salah satu penyebabnya adalah adanya penyakit busuk batang (Ruhnayat, 2004).

6
Penyakit busuk batang yang disebabkan oleh jamur Fusarium oxysporum f. sp. vanillae merupakan salah satu penyakit penting pada tanaman vanili. Jamur ini
menyerang semua bagian tanaman mulai dari akar, batang, daun dan buah. Pada tanaman dewasa tingkat kematian akibat serangan jamur ini mencapai 50 – 100 %, memperpendek umur produksi dari 10 kali panen menjadi dua kali bahkan tidak dapat berproduksi, sedang pada bibit ditemukan 7 – 32 % telah terkontaminasi oleh jamur ini, walaupun tanaman induknya tidak menunjukkan gejala serangan (Hadisutrisno, 2005).
Khusus di Pulau Lombok NTB, areal penanaman vanili pada tahun 2002 seluas 534,60 ha dengan produksi 320,76 ton polong kering dengan nilai Rp 481.140.000.000 yang tersebar di Kabupaten Lombok Barat (Kecamatan Gangga dan Narmada), Kabupaten Lombok Tengah (Kecamatan Batukliang) dan Kabupaten Lombok Timur (Kecamatan Pringgasela) (Balai Laboratorium Perlindungan Tanaman Perkebunan NTB, 2003). Akibat serangan penyebab penyakit busuk batang pada tahun 2003 sekitar 93,00 ha tanaman vanili dalam keadaan rusak dan tidak berproduksi, sedang tanaman vanili lainnya yang masih ada terjadi penurunan hasil sampai 50 %, sehingga diperkirakan terjadi kehilangan produksi mencapai 188,42 ton polong kering dengan nilai Rp 282.420.000.000.
Sampai saat ini penyakit busuk batang vanili sulit untuk dikendalikan, karena jamur F. oxysporum f. sp. vanillae memiliki struktur bertahan berupa klamidospora yang dapat bertahan dalam tanah sebagai saprofit dalam waktu relatif lama sekitar tiga sampai empat tahun (Sukamto dan Tombe, 1995; Nurawan, Tombe dan Matsumoto, 1995). Menurut Hadisutrisno (2005), sulitnya pengendalian penyakit ini disebabkan karena penularannya melalui stek yang sudah terinfeksi, sehingga penyebarannya menjadi cepat dan meluas. Ruhnayat (2004) mengatakan bahwa sampai saat ini belum ditemukan klon vanili yang tahan atau toleran terhadap penyakit ini.
Dengan demikian perlu dicari alternatif pengendalian yang dapat menekan perkembangan penyakit busuk batang. Berdasarkan hasil survei pendahuluan pada daerah endemik penyakit busuk batang vanili di Dusun Timbenuh Desa Pengadangan Kecamatan Pringgasela Kabupaten Lombok Timur dan Dusun Celilos Desa Bentek Kecamatan Gangga Kabupaten Lombok Barat ditemukan beberapa tanaman vanili sehat di antara beberapa tanaman vanili yang terinfeksi penyakit busuk batang. Hasil isolasi dari jaringan tanaman sehat dan contoh tanah ditemukan beberapa jamur endofit dan saprofit, sehingga diduga fenomena tanaman vanili sehat tersebut disebabkan karena ketahanan induksi (terimbas) karena adanya jamur saprofit dan jamur endofit antagonistik (Sudantha, 2005). Ketahanan induksi merupakan ketahanan tanaman terhadap infeksi patogen karena tanaman telah terinfeksi oleh mikroorganisme lain sebelumnya, baik dari jenis yang sama maupun dari jenis lain (Abadi, 2003).
Mekanisme antagonisme jamur endofit dalam menekan perkembangan patogen sehingga tanaman menjadi tahan karena antibiosis. Petrini (1993) melaporkan bahwa jamur endofit menghasilkan alkaloid dan mikotoksin sehingga memungkinkan digunakan untuk meningkatkan ketahanan tanaman terhadap penyakit. Menurut Dahlam, Eichenseer dan Siegel (1991), dan Brunner dan Petrini (1992), jamur endofit menghasilkan senyawa aktif biologis secara in-vitro antara lain alkaloid, paxillin, lolitrems dan tetranone steroid. Selain itu menurut Photita (2003 dalam Lumyong et al., 2004), jamur endofit antagonis mempunyai aktivitas tinggi dalam menghasilkan enzim yang dapat digunakan untuk mengendalikan patogen. Jamur endofit Neotyphodium sp.

7
menghasilkan enzim β-1,6-glucanase yang menyerupai enzim yang sama yang dihasilkan oleh jamur Trichoderma harzianum dan T. virens (Moy et al., 2002).
Penelitian tentang jamur saprofit antagonis untuk pengendalian patogen tular tanah yang menyerang berbagai tanaman di Indonesia telah banyak dilakukan, namun penggunaannya di lapangan masih terbatas dalam skala percobaan. Abadi (1987) melaporkan bahwa Trichoderma harzianum, T. viride dan Penicillium citrinum merupakan jamur yang bersifat antagonistik terhadap Ganoderma boninense pada kelapa sawit. Arifin, Dahlan dan Dahlan (1989) juga melaporkan bahwa jamur Trichoderma spp. merupakan jamur antagonis yang berpotensi mengendalikan jamur G. pseudoferrum pada tanaman teh.
Jamur saprofit antagonis dapat menekan jamur patogen tular tanah melalui tiga mekanisme, seperti jamur T. viride mampu hidup sebagai mikoparasit yang dapat melakukan penetrasi ke miselium dan klamidospora jamur patogen sehingga terjadi lisis dan pengkristalan, menghasilkan antibiotik (gliotoksin dan viridin) yang dapat menghambat pertumbuhan jamur patogen, dan mempunyai kemampuan tumbuh yang lebih cepat sehingga terjadi persaingan dalam ruang dan nutrisi dengan jamur lainnya (Baker dan Cook, 1982).
Ketahanan induksi pada berbagai tanaman karena keberadaan jamur endofit dan saprofit telah banyak diteliti, namun belum ada penelitian pada tanaman vanili. Oleh karena itu dilakukan penelitian tentang karakterisasi dan potensi jamur endofit dan saprofit sebagai agens pengendali hayati jamur F. oxysporum f. sp. vanillae pada
tanaman vanili di Pulau Lombok NTB.
1.2. Tujuan Penelitian
Penelitian bertujuan untuk mengetahui karakteristik jamur endofit dan saprofit
antagonis pada tanaman vanili; pengaruh pH medium, suhu inkubasi dan seresah daun terhadap pertumbuhannya; mekanisme antagonismenya terhadap jamur F. oxysporum f. sp. vanillae; efektivitasnya dalam mengendalikan penyakit busuk batang;
sinergismenya dalam mengendalikan penyakit busuk batang dan potensinya dalam meningkatkan ketahanan induksi tanaman vanili terhadap penyakit busuk batang.
1.3. Manfaat Penelitian
Hasil penelitian dapat menjelaskan karakteristik jamur endofit dan saprofit antagonis pada tanaman vanili, dan mekanisme antagonismenya terhadap jamur F. oxysporum f. sp. vanillae dan mekanisme ketahanan induksi terhadap penyakit busuk batang. Selain itu sinergisme jamur endofit dan saprofit antagonis dapat digunakan sebagai alternatif untuk pengendalian penyakit busuk batang vanili dengan cara meningkatkan ketahanan induksi.
8
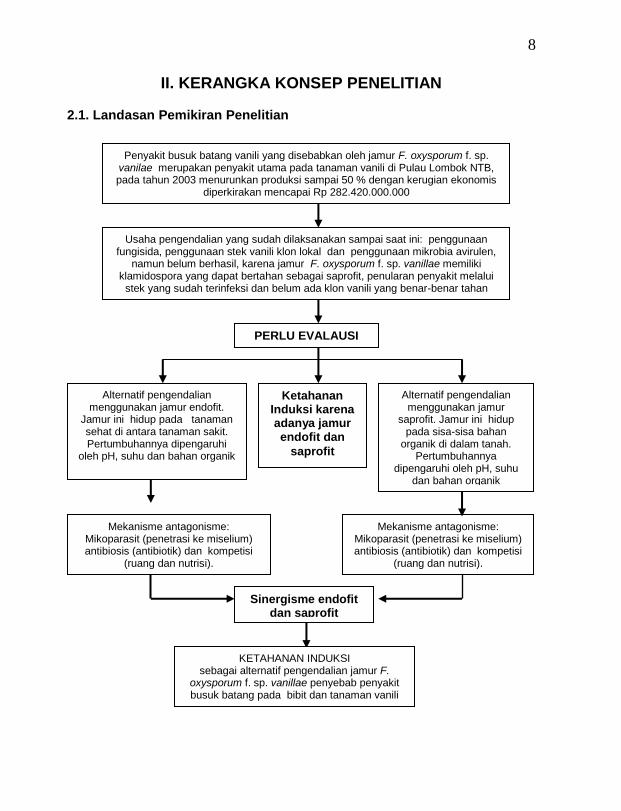
II. KERANGKA KONSEP PENELITIAN
2.1. Landasan Pemikiran Penelitian
Penyakit busuk batang vanili yang disebabkan oleh jamur F. oxysporum f. sp. vanilae merupakan penyakit utama pada tanaman vanili di Pulau Lombok NTB, pada tahun 2003 menurunkan produksi sampai 50 % dengan kerugian ekonomis
diperkirakan mencapai Rp 282.420.000.000
Alternatif pengendalian menggunakan jamur
saprofit. Jamur ini hidup pada sisa-sisa bahan
organik di dalam tanah. Pertumbuhannya
dipengaruhi oleh pH, suhu dan bahan organik
Alternatif pengendalian menggunakan jamur endofit.
Jamur ini hidup pada tanaman sehat di antara tanaman sakit. Pertumbuhannya dipengaruhi
oleh pH, suhu dan bahan organik
Mekanisme antagonisme: Mikoparasit (penetrasi ke miselium) antibiosis (antibiotik) dan kompetisi
(ruang dan nutrisi).
Mekanisme antagonisme: Mikoparasit (penetrasi ke miselium) antibiosis (antibiotik) dan kompetisi
(ruang dan nutrisi).
Ketahanan Induksi karena adanya jamur endofit dan
saprofit
Sinergisme endofit dan saprofit
Usaha pengendalian yang sudah dilaksanakan sampai saat ini: penggunaan fungisida, penggunaan stek vanili klon lokal dan penggunaan mikrobia avirulen,
namun belum berhasil, karena jamur F. oxysporum f. sp. vanillae memiliki klamidospora yang dapat bertahan sebagai saprofit, penularan penyakit melalui
stek yang sudah terinfeksi dan belum ada klon vanili yang benar-benar tahan
PERLU EVALAUSI
KETAHANAN INDUKSI sebagai alternatif pengendalian jamur F.
oxysporum f. sp. vanillae penyebab penyakit busuk batang pada bibit dan tanaman vanili

9
2.2. Hipotesis Penelitian
Hipotesis yang akan dibuktikan pada penelitian ini adalah sebagai berikut:
1. Pada tanaman vanili sehat terdapat jamur endofit antagonis dan pada rhizosfer terdapat jamur saprofit antagonis dengan karakteristik yang berbeda.
2. Jamur endofit dan saprofit dapat tumbuh pada kisaran pH medium 4,0 sampai dengan 8,0, dan kisaran suhu inkubasi 15o sampai dengan 40 o C, serta dapat tumbuh pada berbagai seresah daun pohon pelindung.
3. Mekanisme antagonistik jamur endofit dan saprofit terhadap jamur F. oxysporum f. sp. vanillae secara fisik (kompetisi ruang atau nutrisi dan mikoparasit) dan antibiosis (mengeluarkan antibiotik atau alkaloid yang mudah menguap).
4. Terdapat lebih dari satu jenis jamur endofit atau saprofit antagonis potensial efektif mengendalikan penyakit busuk batang vanili dan dapat memacu pembentukan tunas bunga vanili.
5. Terdapat sinergisme antara jamur endofit dan saprofit antagonis dalam mengendalikan penyakit busuk batang vanili dan dapat meningkatkan ketahanan induksi terhadap penyakit busuk batang.
10
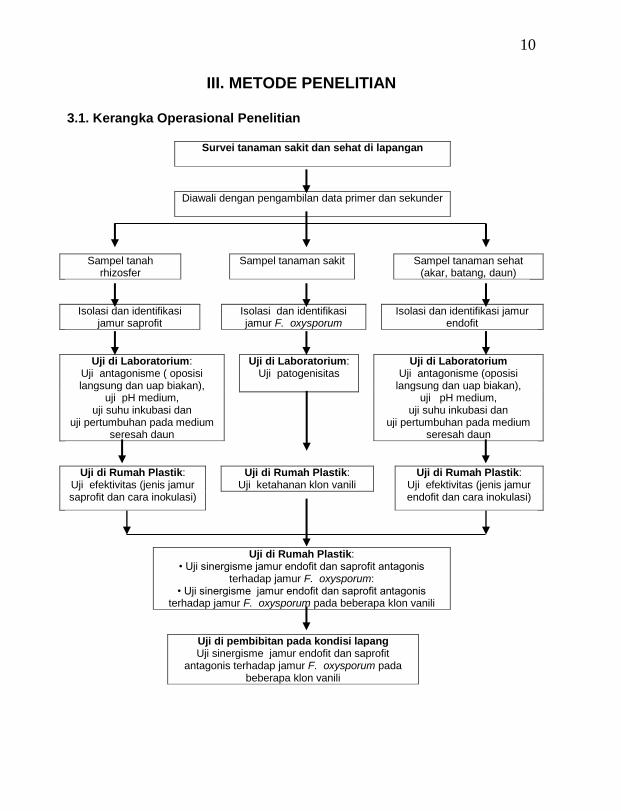
III. METODE PENELITIAN 3.1. Kerangka Operasional Penelitian
Survei tanaman sakit dan sehat di lapangan
Diawali dengan pengambilan data primer dan sekunder
Sampel tanah rhizosfer
Sampel tanaman sakit Sampel tanaman sehat (akar, batang, daun)
Isolasi dan identifikasi jamur saprofit
Isolasi dan identifikasi jamur F. oxysporum
Isolasi dan identifikasi jamur endofit
Uji di Laboratorium: Uji antagonisme ( oposisi langsung dan uap biakan),
uji pH medium, uji suhu inkubasi dan
uji pertumbuhan pada medium seresah daun
Uji di Laboratorium: Uji patogenisitas
Uji di Laboratorium Uji antagonisme (oposisi
langsung dan uap biakan), uji pH medium,
uji suhu inkubasi dan uji pertumbuhan pada medium
seresah daun
Uji di Rumah Plastik: Uji efektivitas (jenis jamur saprofit dan cara inokulasi)
Uji di Rumah Plastik: Uji ketahanan klon vanili
Uji di Rumah Plastik: Uji efektivitas (jenis jamur endofit dan cara inokulasi)
Uji di Rumah Plastik: • Uji sinergisme jamur endofit dan saprofit antagonis
terhadap jamur F. oxysporum: • Uji sinergisme jamur endofit dan saprofit antagonis
terhadap jamur F. oxysporum pada beberapa klon vanili
Uji di pembibitan pada kondisi lapang Uji sinergisme jamur endofit dan saprofit
antagonis terhadap jamur F. oxysporum pada beberapa klon vanili

11
3.2. Survei Tanaman Sakit dan Sehat di Lapang
3.2.1. Pengambilan data primer dan sekunder Pengambilan data primer di lapang meliputi intensitas penyakit busuk batang,
gejala penyakit busuk batang, jenis pohon pelindung dan panjatan tanaman vanili, kandungan bahan organik tanah, pH tanah, kelembaban dan suhu, dan cara budidaya vanili. Data sekunder diambil dari Dinas Perkebunan Kabupaten Lombok Barat, Kabupaten Lombok Tengah, Kabupaten Lombok Timur, Balai Laboratorium Perlindungan Tanaman Perkebunan NTB dan Balai Perbenihan Tanaman Perkebunan NTB meliputi data luas kebun vanili, luas kebun yang terinfeksi penyakit busuk batang, klon-klon vanili yang ditanam di kebun, jenis tanah, dan ketinggian tempat dari permukaan air laut.
3.2.2. Pengambilan sampel tanah, tanaman sakit dan sehat
Pengambilan sampel tanah, tanaman vanili yang terinfeksi penyakit busuk batang dan sehat dilakukan pada enam lokasi kebun di Pulau Lombok. Tanaman vanili yang diduga mengandung jamur endofit diambil dari tanaman vanili yang sehat di antara tanaman yang sakit, sedang jamur F. oxysporum f.sp. vanillae diisolasi dari tanaman
vanili yang menunjukkan gejala busuk batang vanili. Pada masing-masing klon dan lokasi dipilih lima tanaman, kemudian bagian akar, pangkal batang dan daun dari masing-masing tanaman tersebut dimasukkan ke dalam kantong plastik, selanjutnya dibawa ke laboratorium.
3.3. Isolasi dan Identifikasi
Jamur endofit diisolasi dari contoh akar, batang dan daun vanili yang sehat atau tidak terinfeksi penyakit busuk batang. Metode isolasi yang digunakan serupa dengan yang digunakan oleh Petrini dan Carroll (1991 dalam Arnold, 2000). Akar, batang dan daun vanili dipotong-potong sepanjang 1,0 - 1,5 cm, dilakukan sterilisasi permukaannya dengan menggunakan Klorox (sodium hypochlorite) 10 % selama dua menit dan alkohol absolut 96 % (selama 30 detik) sebanyak tiga kali serta aquades steril sebanyak tiga kali.
Isolasi jamur saprofit dilakukan dengan mengisolasi dari rhizosfer atau tanah sekitar perakaran tanaman vanili. Metode isolasi yang akan digunakan adalah metode cawan pengenceran dengan tingkat pengenceran sampai 10-4.
Jamur endofit dan saprofit yang tumbuh, masing-masing dipindahkan ke dalam cawan Petri yang berisi medium PDA dengan teknik transfer konidium tunggal atau teknik transfer ujung hifa (Burgess, Liddell dan Summerell, 1988), kemudian diberi tanda. Pengamatan dilakukan secara makroskopis meliputi warna koloni, arah pertumbuhan koloni, ketebalan koloni, diameter koloni dan kecepatan pertumbuhan koloni; dan secara mikroskopis meliputi warna hifa, bentuk konidia, warna konidia, ada atau tidaknya phialide dan kerapatan phialide.

12
3.4. Percobaan Laboratorium Jamur endofit dan saprofit yang diperoleh selanjutnya diuji pertumbuhannya
pada berbagai pH medium PDA (4 – 8), berbagai suhu inkubasi (15o C – 40 o C), berbagai seresah daun pohon pelindung, uji antagonisme terhadap jamur F. oxysporum f. sp. vanillae menggunakan metode oposisi langsung dan uap biakan jamur endofit dan saprofit. Percobaan dilakukan di Laboratorium Proteksi Tumbuhan dan Laboratorium Mikrobiologi Fakultas Pertanian Universitas Mataram. Percobaan menggunakan Rancangan Acak Lengkap dengan tiga ulangan. Data hasil pengamatan dianalisis menggunakan analisis keragaman pada taraf nyata 0,05 dan uji BNJ pada taraf nyata yang sama.
3.5. Percobaan Rumah Plastik
Jamur endofit dan saprofit yang secara in-vitro efektif menekan pertumbuhan jamur F. oxysporum f. sp. vanillae diaplikasi pada stek vanili melalui perendaman stek dan infestasi ke medium tanah. Selanjutnya jamur endofit dan saprofit yang diketahui efektif mengendalikan penyakit busuk batang vanili diaplikasi secara bersama. Semua percobaan ini dilakukan di Rumah Plastik Fakultas Pertanian Universitas Mataram menggunakan Rancangan Acak Lengkap dengan percobaan faktorial yang diulang tiga kali. Data hasil pengamatan dianalisis menggunakan analisis keragaman pada taraf nyata 0,05 dan uji BNJ pada taraf nyata yang sama.
3.6. Percobaan Lapangan
Jamur endofit dan saprofit yang dapat bersinergis selanjutnya diaplikasi pada stek vanili klon Timbenuh NTB, klon Celelos NTB dan klon Malang Jawa Timur. Percobaan dilaksanakan di Kebun Percobaan Fakultas Pertanian Universitas Mataram menggunakan Rancangan Acak Lengkap dengan percobaan faktorial dengan tiga ulangan. Pengamatan pengamatan penyakit busuk batang dilakukan sampai tanaman berumur 16 minggu. Data hasil pengamatan dianalisis menggunakan analisis keragaman pada taraf nyata 0,05 dan uji BNJ pada taraf nyata yang sama.

13
IV. HASIL DAN PEMBAHASAN
4.1. Karakterisasi jamur endofit, uji pertumbuhan dan uji antagonisme
Berdasarkan hasil isolasi dari jaringan akar tanah, akar udara, batang, daun dan
buah tanaman vanili sehat ditemukan 19 jamur endofit dengan karakteristik yang berbeda (Tabel 1). Dari kebun Timbenuh ditemukan sembilan isolat, kebun Selebung ada lima isolat, kebun Jurang Malang ada dua isolat, kebun Lingsar ada satu isolat dan kebun Celelos ada dua isolat, sedang dari kebun Banok tidak ditemukan jamur endofit. Irawati (2005), melaporkan bahwa jamur Rhizoctonia sp. ditemukan pada akar tanaman vanili sehat, namun belum dimanfaatkan untuk pengendalian penyakit, sedang pada pada tanaman lain dilaporkan oleh Sulistyowati et al. (2005), yaitu jamur endofit Trichoderma asperellum ditemukan pada jaringan batang jeruk sehat. Budi, Mariana dan Rachmadi (2005) menemukan jamur endofit Trichoderma spp. dan Penicillium sp. pada jaringan batang dan akar padi rawa pasang surut. Mekkamol (1998 dalam Lumyong et al., 2004) menemukan jamur Cladosporium sp. pada jaringan batang pohon jati, sedang Busarakum (2002 dalam Manoch, 2004) menemukan jamur Gliocladium penicilloides pada anggrek.
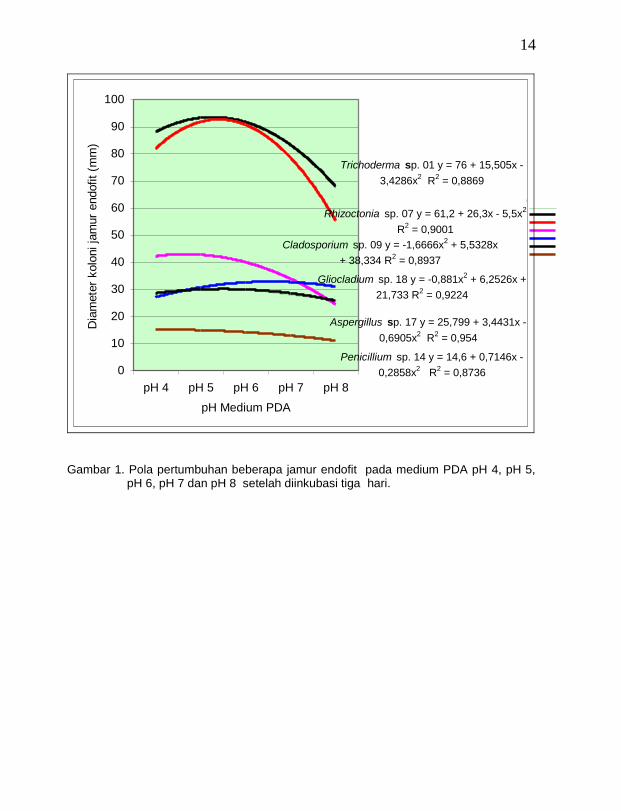
Semua jamur endofit yang ditemukan pada tanaman vanili dapat tumbuh pada pH medium 4 – 8 dan pertumbuhan optimum terjadi pada pH 6 (Gambar 1). Secara umum jamur dapat tumbuh pada kisaran pH yang luas, namun jamur tumbuh optimum apabila substrat mempunyai keasaman antara 5,0 – 6,0. (Barnet dan Hunter (1998). Jamur Trichoderma spp. dapat tumbuh pada kisaran pH medium yang luas dari pH 3,0
sampai dengan pH 8,0, namun pH 5,6 sampai dengan pH 6,0 merupakan pH ideal untuk pertumbuhan (Domsch, Gams dan Anderson, 1980). Jamur T. viride aktif sebagai antagonis di dalam tanah pada pH 5,6 sampai dengan pH 6,0 (Cook dan Baker, 1983). Jamur Rhizoctonia sp. dapat tumbuh pada kisaran pH 2,5 sampai dengan pH
8,5, (Barnett dan Hunter, 1998), namum pertumbuhan yang baik pada medium yang mempunyai pH asam (Carling dan Sumner, 1992 dalam Singleton et al., 1992). pH optimum untuk pertumbuhan jamur Cladosporium sp. mendekati netral (Domsch et al., 1980). Jamur Penicillium spp. dapat tumbuh pada kisaran pH 1,9 sampai dengan pH 9,3, sedang jamur Aspergillus spp. dapat tumbuh pada kisaran pH 4,0 sampai dengan pH 8,0 dan pH optimum sekitar 6,5. Untuk jamur Gliocladium spp. dapat tumbuh pada kisaran pH 3,0 sampai dengan pH 8,2 dan pH optimum 5,6 (Barnett dan Hunter, 1998).
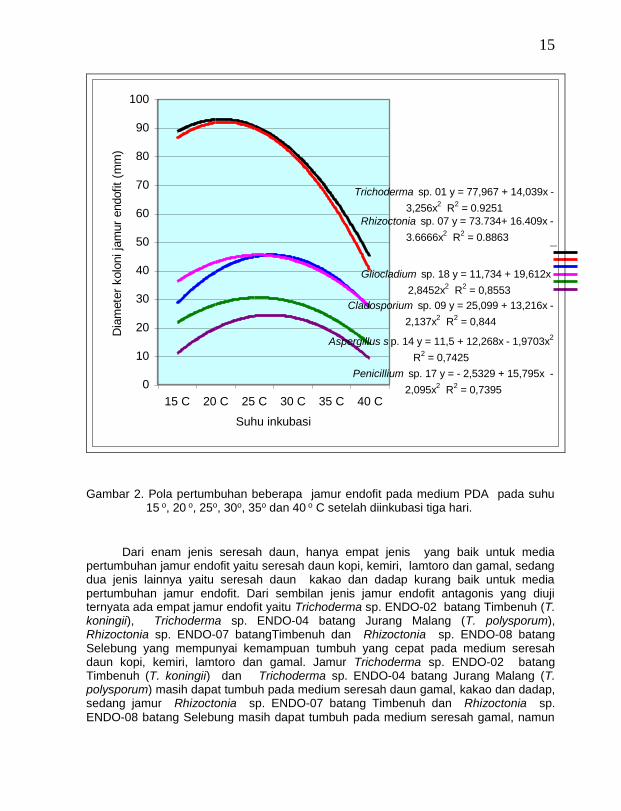
Semua jamur endofit yang ditemukan pada tanaman vanili dapat tumbuh pada kisaran suhu 15o C – 40 o C dan pertumbuhan yang terbaik pada suhu optimum 25 o C (Gambar 2). Suhu merupakan salah satu faktor yang berpengaruh terhadap ketahanan hidup, pertumbuhan dan reproduksi jamur. Untuk pertumbuhan yang ideal jamur memerlukan suhu antara 20 o sampai 30 o C (Barnett dan Hunter, 1998). Suhu optimum untuk pertumbuhan jamur Trichoderma spp. yaitu sekitar 25 o C (Rifai, 1969). Jamur Rhizoctonia spp. dapat tumbuh dengan baik pada suhu kamar (Carling dan Sumner, 1992 dalam Singleton, et al., 1992). Jamur Cladosporium spp. memerlukan suhu optimal sekitar 25 o C untuk pertumbuhannya (Domsch et al., 1980). Suhu optimum untuk pertumbuhan jamur G. viride yaitu 25o C, sedang pertumbuhan jamur P. citrinum memerlukan suhu optimum 24o C dan untuk jamur A. flavus dan A. niger juga memerlukan suhu optimum 25o C untuk pertumbuhannya (Domsch et al., 1980).
14
Trichoderma sp. 01 y = 76 + 15,505x -
3,4286x2 R
2 = 0,8869
Rhizoctonia sp. 07 y = 61,2 + 26,3x - 5,5x2
R2 = 0,9001
Cladosporium sp. 09 y = -1,6666x2 + 5,5328x
+ 38,334 R2 = 0,8937
Gliocladium sp. 18 y = -0,881x2 + 6,2526x +
21,733 R2 = 0,9224
Aspergillus sp. 17 y = 25,799 + 3,4431x -
0,6905x2 R
2 = 0,954
Penicillium sp. 14 y = 14,6 + 0,7146x -
0,2858x2 R
2 = 0,87360
10
20
30
40
50
60
70
80
90
100
pH 4 pH 5 pH 6 pH 7 pH 8
pH Medium PDA
Dia
mete
r kolo
ni ja
mur
endofit
(mm
)
Gambar 1. Pola pertumbuhan beberapa jamur endofit pada medium PDA pH 4, pH 5,
pH 6, pH 7 dan pH 8 setelah diinkubasi tiga hari.
15
Trichoderma sp. 01 y = 77,967 + 14,039x -
3,256x2 R
2 = 0.9251
Rhizoctonia sp. 07 y = 73.734+ 16.409x -
3.6666x2 R
2 = 0.8863
Gliocladium sp. 18 y = 11,734 + 19,612x -
2,8452x2 R
2 = 0,8553
Cladosporium sp. 09 y = 25,099 + 13,216x -
2,137x2 R
2 = 0,844
Aspergillus s p. 14 y = 11,5 + 12,268x - 1,9703x2
R2 = 0,7425
Penicillium sp. 17 y = - 2,5329 + 15,795x -
2,095x2 R
2 = 0,7395
0
10
20
30
40
50
60
70
80
90
100
15 C 20 C 25 C 30 C 35 C 40 C
Suhu inkubasi
Dia
mete
r kolo
ni ja
mur
endofit
(mm
)
Gambar 2. Pola pertumbuhan beberapa jamur endofit pada medium PDA pada suhu 15 o, 20 o, 25o, 30o, 35o dan 40 o C setelah diinkubasi tiga hari.
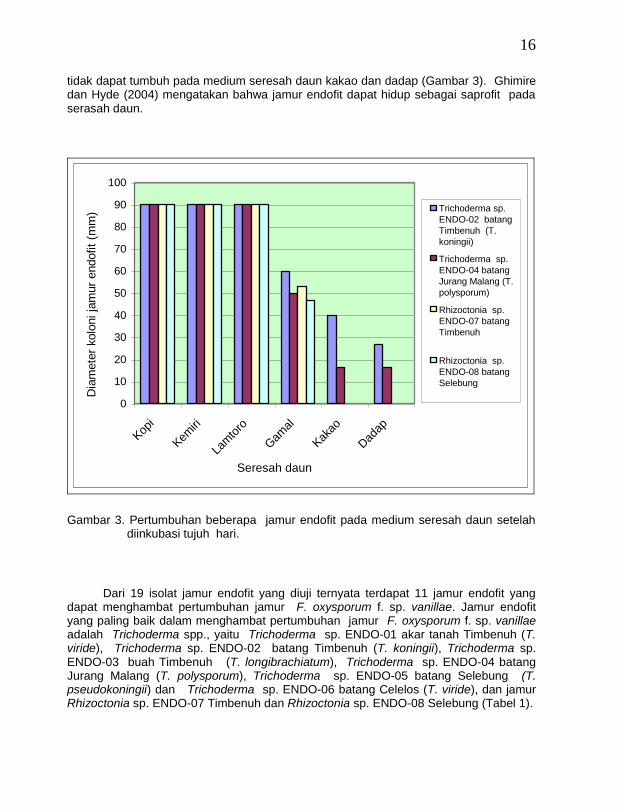
Dari enam jenis seresah daun, hanya empat jenis yang baik untuk media pertumbuhan jamur endofit yaitu seresah daun kopi, kemiri, lamtoro dan gamal, sedang dua jenis lainnya yaitu seresah daun kakao dan dadap kurang baik untuk media pertumbuhan jamur endofit. Dari sembilan jenis jamur endofit antagonis yang diuji ternyata ada empat jamur endofit yaitu Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum), Rhizoctonia sp. ENDO-07 batangTimbenuh dan Rhizoctonia sp. ENDO-08 batang Selebung yang mempunyai kemampuan tumbuh yang cepat pada medium seresah daun kopi, kemiri, lamtoro dan gamal. Jamur Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii) dan Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum) masih dapat tumbuh pada medium seresah daun gamal, kakao dan dadap, sedang jamur Rhizoctonia sp. ENDO-07 batang Timbenuh dan Rhizoctonia sp.
ENDO-08 batang Selebung masih dapat tumbuh pada medium seresah gamal, namun
16
tidak dapat tumbuh pada medium seresah daun kakao dan dadap (Gambar 3). Ghimire dan Hyde (2004) mengatakan bahwa jamur endofit dapat hidup sebagai saprofit pada serasah daun.
0
10
20
30
40
50
60
70
80
90
100
Kop
i
Kem
iri
Lam
toro
Gam
al
Kak
ao
Dad
ap
Seresah daun
Dia
mete
r kolo
ni ja
mur
endofit
(mm
) Trichoderma sp.
ENDO-02 batang
Timbenuh (T.
koningii)
Trichoderma sp.
ENDO-04 batang
Jurang Malang (T.
polysporum)
Rhizoctonia sp.
ENDO-07 batang
Timbenuh
Rhizoctonia sp.
ENDO-08 batang
Selebung
Gambar 3. Pertumbuhan beberapa jamur endofit pada medium seresah daun setelah
diinkubasi tujuh hari.
Dari 19 isolat jamur endofit yang diuji ternyata terdapat 11 jamur endofit yang dapat menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae. Jamur endofit yang paling baik dalam menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae adalah Trichoderma spp., yaitu Trichoderma sp. ENDO-01 akar tanah Timbenuh (T. viride), Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-03 buah Timbenuh (T. longibrachiatum), Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum), Trichoderma sp. ENDO-05 batang Selebung (T. pseudokoningii) dan Trichoderma sp. ENDO-06 batang Celelos (T. viride), dan jamur Rhizoctonia sp. ENDO-07 Timbenuh dan Rhizoctonia sp. ENDO-08 Selebung (Tabel 1).
17
Hasil pengamatan terhadap hifa semua isolat jamur endofit Trichoderma spp. yang mulai kontak dengan jamur F. oxysporum f. sp. vanillae di bawah mikroskop menunjukkan bahwa ± 90 % hifa jamur F. oxysporum f. sp. vanillae mengalami lisis dan pengkristalan (Gambar 4), sedang untuk isolat jamur endofit Rhizoctonia spp. menyebabkan terjadi lisis ± 90 % pada hifa jamur F. oxysporum f. sp. vanillae, namun tidak terjadi pengkristalan. Hasil yang sama pernah dilaporkan oleh Basuki (1985), bahwa jamur T. koningii menyebabkan hifa jamur Rigidiporus microporus mengalami lisis apabila terjadi kontak hifa antar kedua jamur tersebut. Demikian pula Sukamto dan Tombe (1995) melaporkan bahwa isolat Trichoderma sp. (diduga T. viride) asal Manado mampu menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae. Selain itu Abd-El Moity dan Shatla (1981) melaporkan bahwa Trichoderma merupakan mikoparasit yang dapat melakukan penetrasi ke miselium dan sclerotia jamur S. rolfsii sehingga terjadi lisis dan pengkristalan. Lebih lanjut Papavizas (1985) menyatakan bahwa mekanisme mikoparasitisme dimulai dengan pelunakan sel inang oleh enzim yang dihasilkan oleh mikoparasit sebelum kerusakan dan kematian sel inang. Menurut Hadar, Chet dan Henis (1979), jamur T. harzianum memproduksi enzim ekstra selluler ß-(1,3) glucanase
dan chitinase yang mampu merusak dinding sel R. solani.
Tabel 1. Rata-rata persentase hambatan pertumbuhan jamur F. oxysporum f.
sp. vanillae yang beroposisi dengan beberapa jamur endofit
No. Perlakuan jamur F. oxysporum f. sp. vanillae yang beroposisi
dengan jamur endofit Rata-rata
hambatan (%) 1 Trichoderma sp. ENDO-01 akar tanah Timbenuh (T. viride) 45,22 c *)
2 Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum) 45,22 c
3 Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii) 44,44 c
4 Trichoderma sp. ENDO-03 buah Timbenuh (T. longibrachiatum) 43,83 c
5 Trichoderma sp. ENDO-05 batang Selebung (T. pseudokoningii) 43,28 c
6 Trichoderma sp. ENDO-06 batang Celelos (T. viride) 43,28 c
7 Rhizoctonia sp. ENDO-07 batang Timbenuh 41,59 c
8 Rhizoctonia sp. ENDO-08 batang Selebung 41,43 c
9 Gliocladium sp. ENDO-19 batang Lingsar (G. Viride) 31,06 b
10 Gliocladium sp. ENDO-18 batang Jurang Malang (G. catenulatum) 24,33 b
11 Penicllium sp. ENDO-14 akar tanah Timbenuh (P. citrinum) 13,56 b
12 Aspergillus sp. ENDO-17 akar tanah Selebung (A. niger) 6,72 a
13 Aspergillus sp. ENDO-15 akar udara Timbenuh (A. flavus) 5,39 a
14 Aspergillus sp. ENDO-16 daun Timbenuh (A. flavus) 5,33 a
15 Cladosporium sp. ENDO-09 batang Timbenuh 2,67 a
16 Cladosporium sp. ENDO-10 daun Timbenuh 2,67 a
17 Cladosporium sp. ENDO-12 akar udara Selebung 2,67 a
18 Cladosporium sp. ENDO-13 batang Celelos 2,67 a
19 Cladosporium sp. ENDO-11 buah Selebung 1,33 a
20 Kontrol (tanpa jamur endofit) 0,00 a
*) Angka yang diikuti huruf yang sama tidak berbeda nyata pada p p ≤ 0,05
18
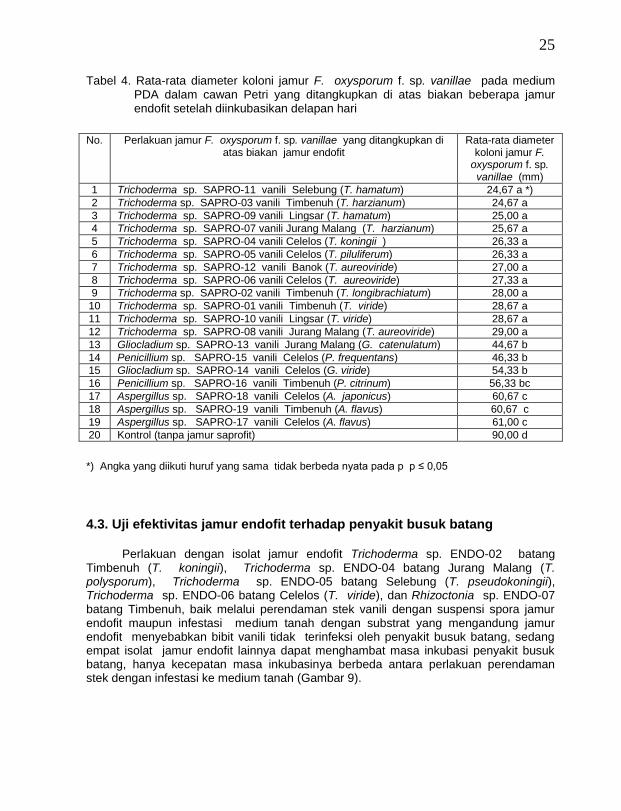
Pada Tabel 2 terlihat bahwa pertumbuhan jamur F. oxysporum f. sp. vanillae pada medium PDA terhambat secara nyata bila biakan tersebut ditangkupkan di atas biakan jamur endofit antagonis dibandingkan dengan bila biakan yang sama ditangkupkan di atas medium PDA tanpa jamur endofit antagonis. Jamur endofit yang terbaik dalam menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae adalah jamur endofit Trichoderma spp. dan Rhizoctonia spp., hal ini diperlihatkan dengan kecilnya diameter koloni jamur F. oxysporum f. sp. vanillae (Gambar 4). Jamur T. viride diketahui menghasilkan gliotoksin pertama kali oleh Weindling dan Emerson (1936 dalam Cook dan Baker, 1983), kemudian Brian dan McGowan (1945 dalam Cook dan Baker, 1983) mengatakan bahwa selain gliotoksin, jamur T. viride juga menghasilkan viridin seperti yang dihasilkan oleh jamur G. virens. Rifai (1969) melaporkan bahwa jamur T. viride mengeluarkan bau minyak kelapa terutama pada biakan yang sudah tua.
Tabel 2. Rata-rata diameter koloni jamur F. oxysporum f. sp. vanillae pada medium PDA dalam cawan Petri yang ditangkupkan di atas biakan beberapa jamur endofit setelah diinkubasikan delapan hari
No. Perlakuan jamur F. oxysporum f. sp. vanillae yang ditangkupkan
di atas biakan jamur endofit Rata-rata diameter
koloni jamur F. oxysporum f. sp. vanillae (mm)
1 Trichoderma sp. ENDO-06 batang Celelos (T. viride) 30,00 a *)
2 Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum) 31,00 ab
3 Rhizoctonia sp. ENDO-08 batang Selebung 33,33 bc
4 Trichoderma sp. ENDO-05 batang Selebung (T. pseudokoningii) 34,67 bc
5 Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii) 35,33 bc
6 Trichoderma sp. ENDO-01 akar tanah Timbenuh (T. viride) 37,00 bc
7 Trichoderma sp. ENDO-03 buah Timbenuh (T. longibrachiatum) 38,00 c
8 Rhizoctonia sp. ENDO-07 batang Timbenuh 40,33 cd
9 Gliocladium sp. ENDO-19 batang Lingsar (G. viride) 45,67 de
10 Gliocladium sp. ENDO-18 batang Jurang Malang (G. catenulatum) 47,67 e
11 Penicllium sp. ENDO-14 akar tanah Timbenuh (P. citrinum) 50,00 e
12 Cladosporium sp. ENDO-13 batang Celelos 55,67 e
13 Cladosporium sp. ENDO-12 akar udara Selebung 56,33 ef
14 Cladosporium sp. ENDO-10 daun Timbenuh 60,67 f
15 Cladosporium sp. ENDO-11 buah Selebung 64,67 f
16 Cladosporium sp. ENDO-09 batang Timbenuh 65,00 f
17 Aspergillus sp. ENDO-15 akar udara Timbenuh (A. flavus) 74,00 g
18 Aspergillus sp. ENDO-16 daun Timbenuh (A. flavus) 75,33 g
19 Aspergillus sp. ENDO-17 akar tanah Selebung (A. niger) 80,33 g
20 Kontrol (tanpa jamur endofit) 90,00 h
*) Angka yang diikuti huruf yang sama tidak berbeda nyata pada p p ≤ 0,05

19
Jamur Trichoderma sp. menghambat
pertumbuhan F. oxysporum f. sp. vanillae (tanda panah) (A) dan lisis pada hifa jamur F.
oxysporum f. sp. vanillae (tanda panah) (B)
Jamur Trichoderma sp. menghambat pertumbuhan F. oxysporum f. sp. vanillae
(tanda panah) (C) dan kontrol (D) pada uji uap biakan
Gambar 4. Uji antagonisme dan uji uap biakan jamur endofit terhadap jamur F.
oxysporum f. sp. vanillae
4.2. Karakterisasi jamur saprofit, uji pertumbuhan dan uji antagonisme
Berdasarkan hasil isolasi dari rhizosfer atau tanah sekitar perakaran tanaman vanili sehat ditemukan 19 jamur saprofit dengan karakteristik yang berbeda (Tabel 3). Dari kebun Timbenuh ditemukan lima isolat, kebun Banok ada satu isolat, kebun Selebung ada satu isolat, kebun Jurang Malang ada tiga isolat, Kebun Lingsar ada dua isolat dan kebun Celelos ada tujuh isolat. Beberapa peneliti terdahulu juga pernah melaporkan, seperti Sastrahidayat (1991) menemukan Trichoderma viride dan Aspergillus sp. di rhizosfer tanaman vanili di Malang. Sukamto dan Tombe (1995) melaporkan bahwa isolat jamur Trichoderma sp. (diduga T. viride) asal Manado dan Bali juga ditemukan pada rhizsosfer tanaman vanili yang digunakan untuk percobaan pengendalian penyakit busuk batang vanili.
Sama halnya dengan jamur endofit, ternyata semua jamur saprofit yang ditemukan dapat tumbuh pada kisaran pH medium 4 – 8 dan pertumbuhan yang terbaik terjadi pada pH 6, demikian pula semua jamur saprofit dapat tumbuh pada kisaran suhu 15o C – 40 o C dan pertumbuhan terbaik terjadi pada suhu 25 o (Gambar 5 dan 6).
A B C D
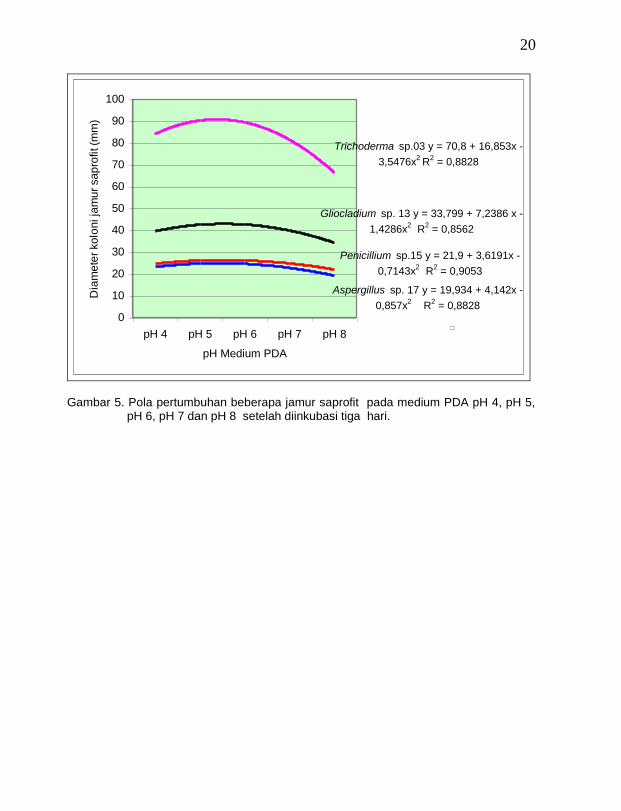
20
Trichoderma sp.03 y = 70,8 + 16,853x -
3,5476x2
R2 = 0,8828
Gliocladium sp. 13 y = 33,799 + 7,2386 x -
1,4286x2 R
2 = 0,8562
Penicillium sp.15 y = 21,9 + 3,6191x -
0,7143x2
R2 = 0,9053
Aspergillus sp. 17 y = 19,934 + 4,142x -
0,857x2 R
2 = 0,8828
0
10
20
30
40
50
60
70
80
90
100
pH 4 pH 5 pH 6 pH 7 pH 8
pH Medium PDA
Dia
me
ter
ko
lon
i ja
mu
r sap
rofit (m
m)
Gambar 5. Pola pertumbuhan beberapa jamur saprofit pada medium PDA pH 4, pH 5,
pH 6, pH 7 dan pH 8 setelah diinkubasi tiga hari.
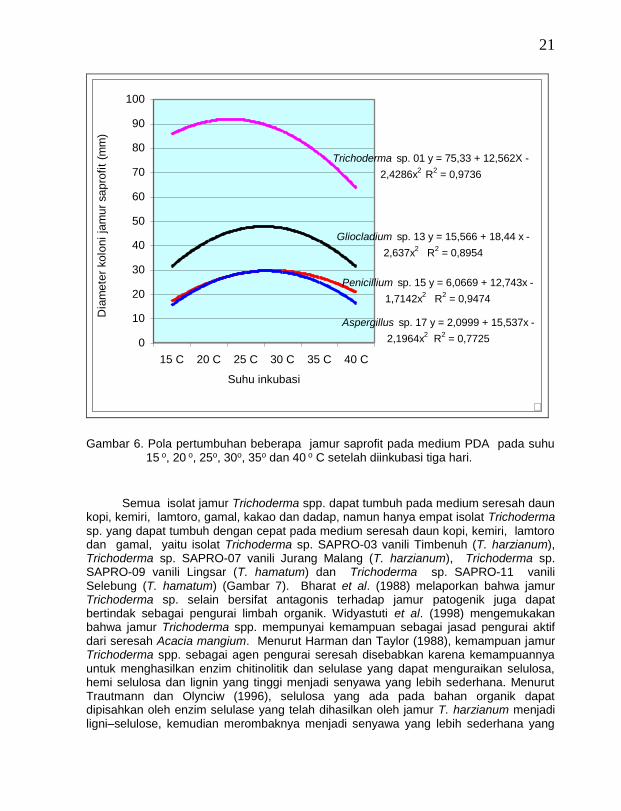
21
Trichoderma sp. 01 y = 75,33 + 12,562X -
2,4286x2
R2 = 0,9736
Gliocladium sp. 13 y = 15,566 + 18,44 x -
2,637x2
R2 = 0,8954
Penicillium sp. 15 y = 6,0669 + 12,743x -
1,7142x2
R2 = 0,9474
Aspergillus sp. 17 y = 2,0999 + 15,537x -
2,1964x2 R
2 = 0,77250
10
20
30
40
50
60
70
80
90
100
15 C 20 C 25 C 30 C 35 C 40 C
Suhu inkubasi
Dia
mete
r kolo
ni
jam
ur
sap
rofi
t (m
m)
Gambar 6. Pola pertumbuhan beberapa jamur saprofit pada medium PDA pada suhu
15 o, 20 o, 25o, 30o, 35o dan 40 o C setelah diinkubasi tiga hari.
Semua isolat jamur Trichoderma spp. dapat tumbuh pada medium seresah daun kopi, kemiri, lamtoro, gamal, kakao dan dadap, namun hanya empat isolat Trichoderma
sp. yang dapat tumbuh dengan cepat pada medium seresah daun kopi, kemiri, lamtoro dan gamal, yaitu isolat Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum) dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum) (Gambar 7). Bharat et al. (1988) melaporkan bahwa jamur Trichoderma sp. selain bersifat antagonis terhadap jamur patogenik juga dapat bertindak sebagai pengurai limbah organik. Widyastuti et al. (1998) mengemukakan bahwa jamur Trichoderma spp. mempunyai kemampuan sebagai jasad pengurai aktif dari seresah Acacia mangium. Menurut Harman dan Taylor (1988), kemampuan jamur Trichoderma spp. sebagai agen pengurai seresah disebabkan karena kemampuannya untuk menghasilkan enzim chitinolitik dan selulase yang dapat menguraikan selulosa, hemi selulosa dan lignin yang tinggi menjadi senyawa yang lebih sederhana. Menurut Trautmann dan Olynciw (1996), selulosa yang ada pada bahan organik dapat dipisahkan oleh enzim selulase yang telah dihasilkan oleh jamur T. harzianum menjadi ligni–selulose, kemudian merombaknya menjadi senyawa yang lebih sederhana yang
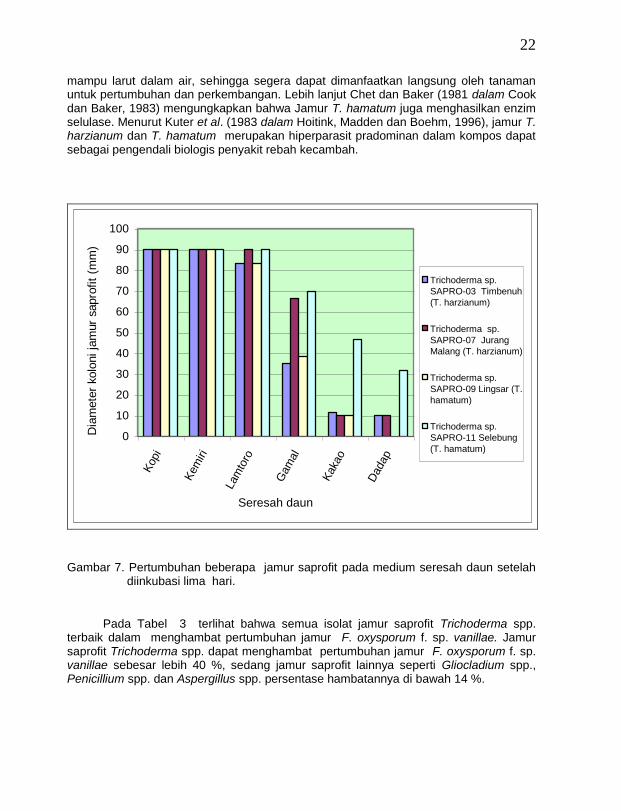
22
mampu larut dalam air, sehingga segera dapat dimanfaatkan langsung oleh tanaman untuk pertumbuhan dan perkembangan. Lebih lanjut Chet dan Baker (1981 dalam Cook dan Baker, 1983) mengungkapkan bahwa Jamur T. hamatum juga menghasilkan enzim selulase. Menurut Kuter et al. (1983 dalam Hoitink, Madden dan Boehm, 1996), jamur T. harzianum dan T. hamatum merupakan hiperparasit pradominan dalam kompos dapat sebagai pengendali biologis penyakit rebah kecambah.
0
10
20
30
40
50
60
70
80
90
100
Kopi
Kem
iri
Lam
toro
Gam
al
Kaka
o
Dadap
Seresah daun
Dia
mete
r kolo
ni ja
mur
sapro
fit
(mm
)
Trichoderma sp.
SAPRO-03 Timbenuh
(T. harzianum)
Trichoderma sp.
SAPRO-07 Jurang
Malang (T. harzianum)
Trichoderma sp.
SAPRO-09 Lingsar (T.
hamatum)
Trichoderma sp.
SAPRO-11 Selebung
(T. hamatum)
Gambar 7. Pertumbuhan beberapa jamur saprofit pada medium seresah daun setelah diinkubasi lima hari.
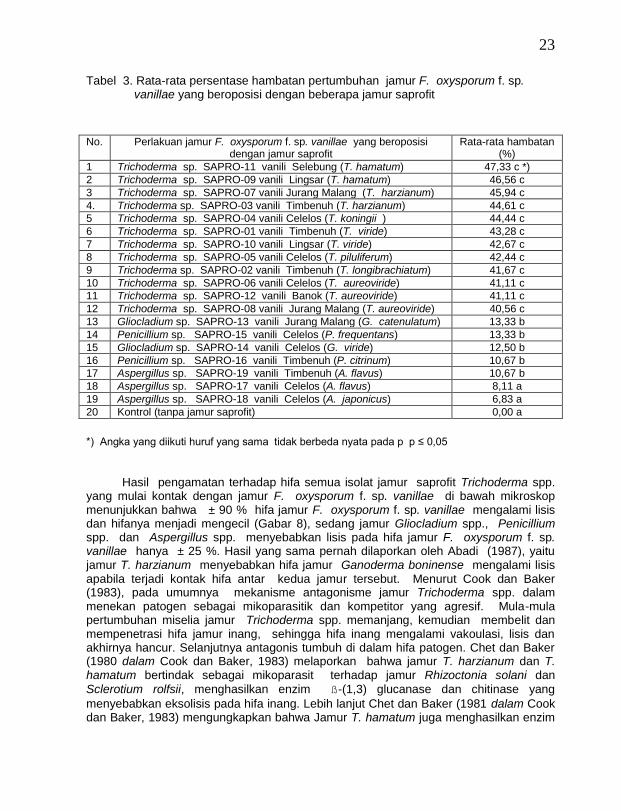
Pada Tabel 3 terlihat bahwa semua isolat jamur saprofit Trichoderma spp.
terbaik dalam menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae. Jamur saprofit Trichoderma spp. dapat menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae sebesar lebih 40 %, sedang jamur saprofit lainnya seperti Gliocladium spp., Penicillium spp. dan Aspergillus spp. persentase hambatannya di bawah 14 %.
23
Tabel 3. Rata-rata persentase hambatan pertumbuhan jamur F. oxysporum f. sp. vanillae yang beroposisi dengan beberapa jamur saprofit
No. Perlakuan jamur F. oxysporum f. sp. vanillae yang beroposisi
dengan jamur saprofit Rata-rata hambatan
(%)
1 Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum) 47,33 c *)
2 Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum) 46,56 c
3 Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum) 45,94 c
4. Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum) 44,61 c
5 Trichoderma sp. SAPRO-04 vanili Celelos (T. koningii ) 44,44 c
6 Trichoderma sp. SAPRO-01 vanili Timbenuh (T. viride) 43,28 c
7 Trichoderma sp. SAPRO-10 vanili Lingsar (T. viride) 42,67 c
8 Trichoderma sp. SAPRO-05 vanili Celelos (T. piluliferum) 42,44 c
9 Trichoderma sp. SAPRO-02 vanili Timbenuh (T. longibrachiatum) 41,67 c
10 Trichoderma sp. SAPRO-06 vanili Celelos (T. aureoviride) 41,11 c
11 Trichoderma sp. SAPRO-12 vanili Banok (T. aureoviride) 41,11 c
12 Trichoderma sp. SAPRO-08 vanili Jurang Malang (T. aureoviride) 40,56 c
13 Gliocladium sp. SAPRO-13 vanili Jurang Malang (G. catenulatum) 13,33 b
14 Penicillium sp. SAPRO-15 vanili Celelos (P. frequentans) 13,33 b
15 Gliocladium sp. SAPRO-14 vanili Celelos (G. viride) 12,50 b
16 Penicillium sp. SAPRO-16 vanili Timbenuh (P. citrinum) 10,67 b
17 Aspergillus sp. SAPRO-19 vanili Timbenuh (A. flavus) 10,67 b
18 Aspergillus sp. SAPRO-17 vanili Celelos (A. flavus) 8,11 a
19 Aspergillus sp. SAPRO-18 vanili Celelos (A. japonicus) 6,83 a
20 Kontrol (tanpa jamur saprofit) 0,00 a
*) Angka yang diikuti huruf yang sama tidak berbeda nyata pada p p ≤ 0,05
Hasil pengamatan terhadap hifa semua isolat jamur saprofit Trichoderma spp. yang mulai kontak dengan jamur F. oxysporum f. sp. vanillae di bawah mikroskop menunjukkan bahwa ± 90 % hifa jamur F. oxysporum f. sp. vanillae mengalami lisis dan hifanya menjadi mengecil (Gabar 8), sedang jamur Gliocladium spp., Penicillium spp. dan Aspergillus spp. menyebabkan lisis pada hifa jamur F. oxysporum f. sp. vanillae hanya ± 25 %. Hasil yang sama pernah dilaporkan oleh Abadi (1987), yaitu jamur T. harzianum menyebabkan hifa jamur Ganoderma boninense mengalami lisis apabila terjadi kontak hifa antar kedua jamur tersebut. Menurut Cook dan Baker (1983), pada umumnya mekanisme antagonisme jamur Trichoderma spp. dalam menekan patogen sebagai mikoparasitik dan kompetitor yang agresif. Mula-mula pertumbuhan miselia jamur Trichoderma spp. memanjang, kemudian membelit dan mempenetrasi hifa jamur inang, sehingga hifa inang mengalami vakoulasi, lisis dan akhirnya hancur. Selanjutnya antagonis tumbuh di dalam hifa patogen. Chet dan Baker (1980 dalam Cook dan Baker, 1983) melaporkan bahwa jamur T. harzianum dan T. hamatum bertindak sebagai mikoparasit terhadap jamur Rhizoctonia solani dan
Sclerotium rolfsii, menghasilkan enzim ß-(1,3) glucanase dan chitinase yang
menyebabkan eksolisis pada hifa inang. Lebih lanjut Chet dan Baker (1981 dalam Cook dan Baker, 1983) mengungkapkan bahwa Jamur T. hamatum juga menghasilkan enzim
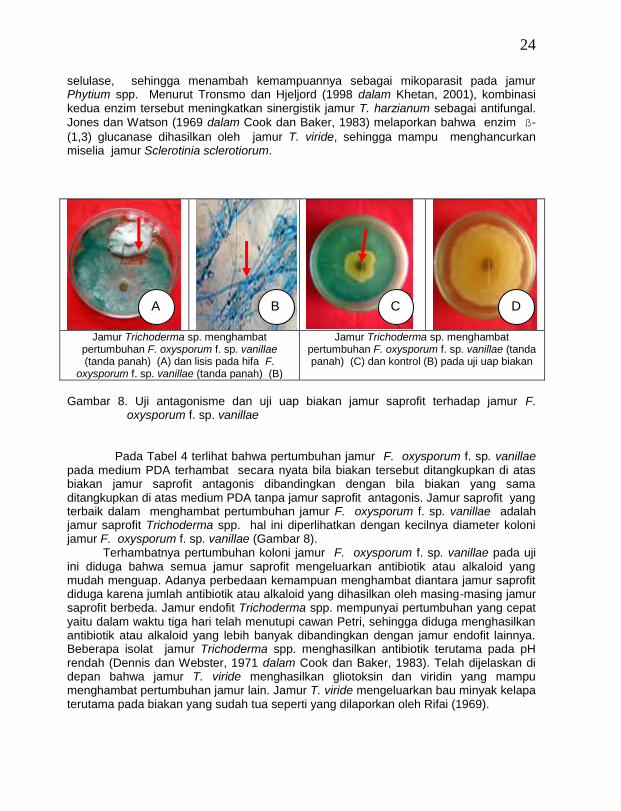
24
selulase, sehingga menambah kemampuannya sebagai mikoparasit pada jamur Phytium spp. Menurut Tronsmo dan Hjeljord (1998 dalam Khetan, 2001), kombinasi kedua enzim tersebut meningkatkan sinergistik jamur T. harzianum sebagai antifungal.
Jones dan Watson (1969 dalam Cook dan Baker, 1983) melaporkan bahwa enzim ß-
(1,3) glucanase dihasilkan oleh jamur T. viride, sehingga mampu menghancurkan miselia jamur Sclerotinia sclerotiorum.
Jamur Trichoderma sp. menghambat
pertumbuhan F. oxysporum f. sp. vanillae (tanda panah) (A) dan lisis pada hifa F.
oxysporum f. sp. vanillae (tanda panah) (B)
Jamur Trichoderma sp. menghambat pertumbuhan F. oxysporum f. sp. vanillae (tanda panah) (C) dan kontrol (B) pada uji uap biakan
Gambar 8. Uji antagonisme dan uji uap biakan jamur saprofit terhadap jamur F.
oxysporum f. sp. vanillae
Pada Tabel 4 terlihat bahwa pertumbuhan jamur F. oxysporum f. sp. vanillae pada medium PDA terhambat secara nyata bila biakan tersebut ditangkupkan di atas biakan jamur saprofit antagonis dibandingkan dengan bila biakan yang sama ditangkupkan di atas medium PDA tanpa jamur saprofit antagonis. Jamur saprofit yang terbaik dalam menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae adalah jamur saprofit Trichoderma spp. hal ini diperlihatkan dengan kecilnya diameter koloni jamur F. oxysporum f. sp. vanillae (Gambar 8).
Terhambatnya pertumbuhan koloni jamur F. oxysporum f. sp. vanillae pada uji ini diduga bahwa semua jamur saprofit mengeluarkan antibiotik atau alkaloid yang mudah menguap. Adanya perbedaan kemampuan menghambat diantara jamur saprofit diduga karena jumlah antibiotik atau alkaloid yang dihasilkan oleh masing-masing jamur saprofit berbeda. Jamur endofit Trichoderma spp. mempunyai pertumbuhan yang cepat yaitu dalam waktu tiga hari telah menutupi cawan Petri, sehingga diduga menghasilkan antibiotik atau alkaloid yang lebih banyak dibandingkan dengan jamur endofit lainnya. Beberapa isolat jamur Trichoderma spp. menghasilkan antibiotik terutama pada pH rendah (Dennis dan Webster, 1971 dalam Cook dan Baker, 1983). Telah dijelaskan di depan bahwa jamur T. viride menghasilkan gliotoksin dan viridin yang mampu menghambat pertumbuhan jamur lain. Jamur T. viride mengeluarkan bau minyak kelapa terutama pada biakan yang sudah tua seperti yang dilaporkan oleh Rifai (1969).
A
B
C
D
25
Tabel 4. Rata-rata diameter koloni jamur F. oxysporum f. sp. vanillae pada medium PDA dalam cawan Petri yang ditangkupkan di atas biakan beberapa jamur endofit setelah diinkubasikan delapan hari
No. Perlakuan jamur F. oxysporum f. sp. vanillae yang ditangkupkan di
atas biakan jamur endofit Rata-rata diameter
koloni jamur F. oxysporum f. sp. vanillae (mm)
1 Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum) 24,67 a *)
2 Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum) 24,67 a
3 Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum) 25,00 a
4 Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum) 25,67 a
5 Trichoderma sp. SAPRO-04 vanili Celelos (T. koningii ) 26,33 a
6 Trichoderma sp. SAPRO-05 vanili Celelos (T. piluliferum) 26,33 a
7 Trichoderma sp. SAPRO-12 vanili Banok (T. aureoviride) 27,00 a
8 Trichoderma sp. SAPRO-06 vanili Celelos (T. aureoviride) 27,33 a
9 Trichoderma sp. SAPRO-02 vanili Timbenuh (T. longibrachiatum) 28,00 a
10 Trichoderma sp. SAPRO-01 vanili Timbenuh (T. viride) 28,67 a
11 Trichoderma sp. SAPRO-10 vanili Lingsar (T. viride) 28,67 a
12 Trichoderma sp. SAPRO-08 vanili Jurang Malang (T. aureoviride) 29,00 a
13 Gliocladium sp. SAPRO-13 vanili Jurang Malang (G. catenulatum) 44,67 b
14 Penicillium sp. SAPRO-15 vanili Celelos (P. frequentans) 46,33 b
15 Gliocladium sp. SAPRO-14 vanili Celelos (G. viride) 54,33 b
16 Penicillium sp. SAPRO-16 vanili Timbenuh (P. citrinum) 56,33 bc
17 Aspergillus sp. SAPRO-18 vanili Celelos (A. japonicus) 60,67 c
18 Aspergillus sp. SAPRO-19 vanili Timbenuh (A. flavus) 60,67 c
19 Aspergillus sp. SAPRO-17 vanili Celelos (A. flavus) 61,00 c
20 Kontrol (tanpa jamur saprofit) 90,00 d
*) Angka yang diikuti huruf yang sama tidak berbeda nyata pada p p ≤ 0,05
4.3. Uji efektivitas jamur endofit terhadap penyakit busuk batang
Perlakuan dengan isolat jamur endofit Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum), Trichoderma sp. ENDO-05 batang Selebung (T. pseudokoningii), Trichoderma sp. ENDO-06 batang Celelos (T. viride), dan Rhizoctonia sp. ENDO-07 batang Timbenuh, baik melalui perendaman stek vanili dengan suspensi spora jamur endofit maupun infestasi medium tanah dengan substrat yang mengandung jamur endofit menyebabkan bibit vanili tidak terinfeksi oleh penyakit busuk batang, sedang empat isolat jamur endofit lainnya dapat menghambat masa inkubasi penyakit busuk batang, hanya kecepatan masa inkubasinya berbeda antara perlakuan perendaman stek dengan infestasi ke medium tanah (Gambar 9).

26
0
10
20
30
40
50
60
Tricho
ENDO-0
1 ak
ar T
imbe
nuh
Tricho
ENDO-0
2 ba
tang
Tim
benu
h
Tricho
ENDO-0
3 bu
ah T
imbe
nuh
Tricho
ENDO-0
4 ba
tang
Jur
ang
Malan
g
Tricho
ENDO-0
5 ba
tang
Sel
ebun
g
Tricho
ENDO-0
6 ba
tang
Cel
elos
Rhi
zoct E
NDO
-07
bata
ng T
imbe
nuh
Rhi
zoct E
NDO
-08
bata
ng S
eleb
ung
Cla
dosp
ENDO-1
3 ba
tang
Celel
os
Glio
clad
ENDO-1
9 ba
tang
Lings
ar
Kon
trol
Jamur endofit
Masa inkubasi penyakit b
usuk b
ata
ng v
anili
(hari
)
Perendaman stek
Investasi media tanah
Gambar 9. Histogram masa inkubasi penyakit busuk batang vanili sebagai akibat
perlakuan jamur endofit dan cara aplikasinya
Adanya perbedaan masa inkubasi penyakit busuk batang karena perlakuan jamur endofit diduga erat kaitannya kemampuan menghambat yang berbeda dari masing-masing jamur endofit antagonis terhadap jamur F. oxysporun f. sp. vanillae. Berdasarkan hasil percobaan uji antagonis jamur endofit terhadap jamur F. oxysporum f. sp. vanillae secara in-vitro di laboratorium melalui metode oposisi langsung dan uji uap biakan, ternyata semua isolat jamur endofit dapat menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae dengan persentase hambatan yang tertinggi oleh Trichoderma spp. disusul oleh Rhizoctonia spp., Gliocladium spp. dan Cladosporium

27
spp. Diduga mekanisme penghambatan yang terjadi secara in-vitro juga terjadi secara in-vivo pada jaringan batang vanili yang terinfeksi oleh penyakit busuk batang, yaitu
secara fisik (kompetisi ruang dan mikoparasit) dan mengeluarkan antibiotik yang didifusikan ke ruang antar sel dalam jaringan.
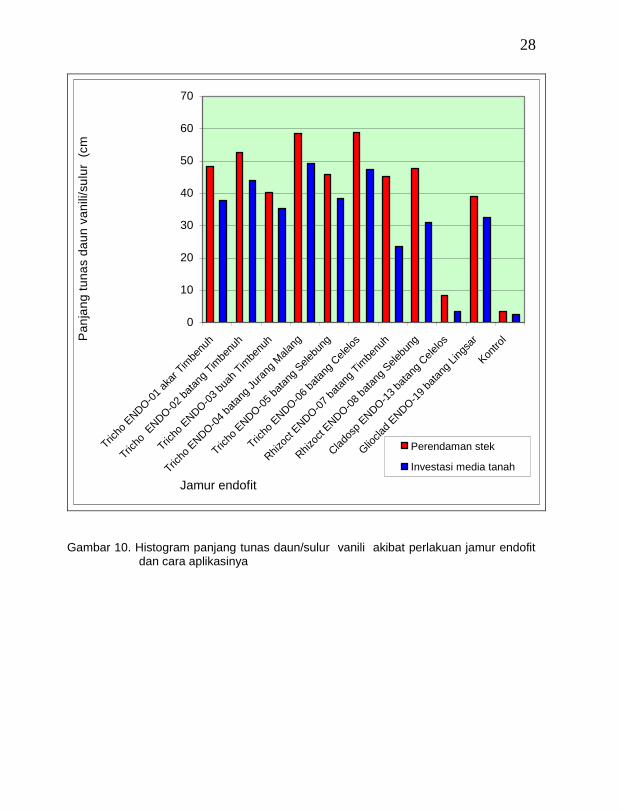
Panjang tunas/sulur pada bibit vanili umur delapan minggu karena perlakuan isolat jamur endofit melalui perendaman stek vanili berbeda nyata dengan infestasi ke medium tanah. Tunas/sulur pada bibit yang diperlakukan dengan jamur endofit melalui perendaman stek lebih panjang dibandingkan dengan perlakuan infestasi ke medium tanah (Gambar 10). Adanya perbedaan panjang tunas/sulur daun pada bibit vanili ini diduga erat kaitannya dengan kemampuan kolonisasi dari jamur endofit setelah diperlakukan dengan kedua cara tersebut. Jamur endofit yang diperlakukan dengan cara perendaman stek lebih cepat mengkolonisasi pada jaringan tanaman dibandingkan dengan infestasi ke medium tanah. Dengan makin cepatnya melakukan kolonisasi pada jaringan tanaman maka besar kemampuannya dalam mengendalikan penyakit busuk batang. Jamur endofit Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum) dan Trichoderma sp. ENDO-06 batang Celelos (T. viride) menyebabkan tunas daun/sulur pada bibit vanili lebih panjang dibandingkan dengan jamur endofiit lainnya baik yang diperlakukan melalui perendaman stek maupun infestasi ke medium tanah.
Dari pengujian penggunaan jamur endofit untuk pengendalian penyakit busuk batang ini dapat dikatakan bahwa penggunaan jamur endofit baik melalui perendaman stek vanili maupun infestasi ke medium tanah dapat meningkatkan ketahanan induksi bibit vanili terhadap penyakit busuk batang (Gambar 11). Hal ini sesuai dengan pendapat Abadi (2003) bahwa ketahanan induksi dapat terjadi karena tanaman telah terinfeksi oleh mikroorganisme lain sebelumnya, baik dari jenis yang sama maupun dari jenis lain. Lebih lanjut Guest (2005) mengatakan bahwa ketahanan induksi terjadi karena kombinasi dari rintangan pasif dengan respon lokal karena adanya peristiwa matinya sel dan akumulasi antibiotik yang dapat berupa fitoaleksin. Telah dijelaskan di depan bahwa jamur T. viride menghasilkan gliotoksin dan viridin (Weindling dan Emerson, 1936, dan Brian dan McGowan, 1945 dalam Cook dan Baker, 1983) Rifai (1969) melaporkan bahwa jamur T. viride mengeluarkan bau minyak kelapa terutama pada biakan yang sudah tua. Selain itu menurut Jones dan Watson (1969 dalam Cook
dan Baker, 1983)), jamur T. viride menghasilkan enzim enzim ß-(1,3) glucanase,
sehingga mampu menghancurkan miselia jamur patogenik.
28
0
10
20
30
40
50
60
70
Tricho
ENDO-0
1 ak
ar T
imbe
nuh
Tricho
ENDO-0
2 ba
tang
Tim
benu
h
Tricho
ENDO-0
3 bu
ah T
imbe
nuh
Tricho
ENDO-0
4 ba
tang
Jur
ang
Malan
g
Tricho
ENDO-0
5 ba
tang
Seleb
ung
Tricho
ENDO-0
6 ba
tang
Cel
elos
Rhi
zoct E
NDO-0
7 ba
tang
Tim
benu
h
Rhi
zoct E
NDO-0
8 ba
tang
Seleb
ung
Cla
dosp
ENDO-1
3 ba
tang
Cel
elos
Glio
clad
ENDO-1
9 ba
tang
Lin
gsar
Kon
trol
Jamur endofit
Pa
nja
ng
tu
na
s d
au
n v
an
ili/s
ulu
r (
cm
)
Perendaman stek
Investasi media tanah
Gambar 10. Histogram panjang tunas daun/sulur vanili akibat perlakuan jamur endofit dan cara aplikasinya

29
Perlakuan jamur Trichoderma sp. ENDO-06
batang Celelos (T. viride) Bibit vanili terinfeksi pada kontrol (tanpa
perlakuan jamur endofit)
Gambar 11. Bibit vanili sehat (A) akibat penggunaan isolat jamur endofit dan bibit
vanili yang terinfeksi penyakit busuk batang pada kontrol (B)
4.4. Uji efektivitas jamur saprofit terhadap penyakit busuk batang
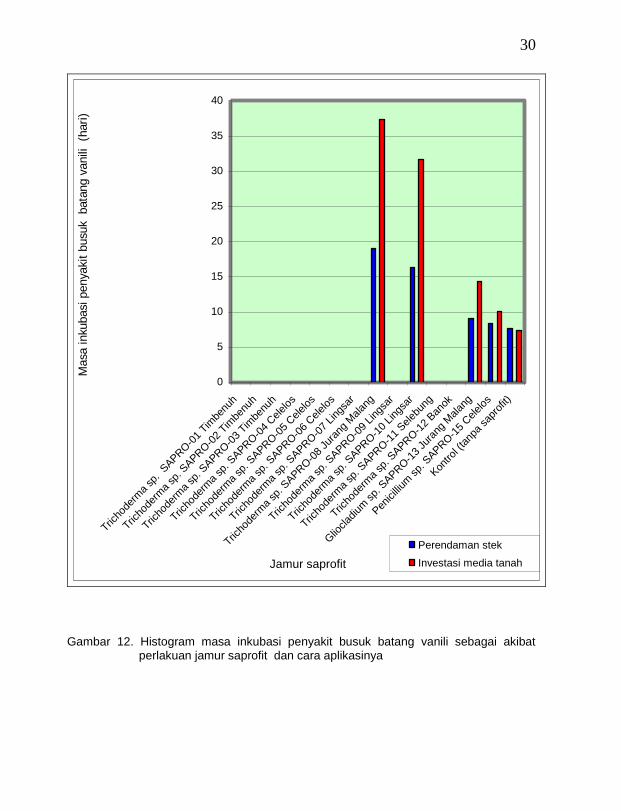
Dari 14 isolat jamur saprofit antagonistik yang diperlakukan dengan cara
perendaman stek vanili dan infestasi ke medium tanah ternyata ada delapan isolat yang menyebabkan bibit vanili tidak terinfeksi oleh penyakit busuk batang, yaitu Trichoderma sp. SAPRO-01 vanili Timbenuh (T. viride), Trichoderma sp. SAPRO-02 vanili Timbenuh (T. longibrachiatum), Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-04 vanili Celelos (T. koningii), Trichoderma sp. SAPRO-05 vanili Celelos (T. piluliferum), Trichoderma sp. SAPRO-06 vanili Celelos (T. aureoviride), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum), Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum), dan Trichoderma sp. SAPRO-12 vanili Banok (T. aureoviride) (Gambar 12).
A B
30
0
5
10
15
20
25
30
35
40
Tricho
derm
a sp
. SAP
RO-0
1 Tim
benu
h
Tricho
derm
a sp
. SAPR
O-0
2 Tim
benu
h
Tricho
derm
a sp
. SAPR
O-0
3 Tim
benu
h
Tricho
derm
a sp
. SAPR
O-0
4 Celelos
Tricho
derm
a sp
. SAPR
O-0
5 Celelos
Tricho
derm
a sp
. SAPR
O-0
6 Celelos
Tricho
derm
a sp
. SAPR
O-0
7 Li
ngsa
r
Tricho
derm
a sp
. SAPR
O-0
8 Ju
rang
Malan
g
Tricho
derm
a sp
. SAPR
O-0
9 Li
ngsa
r
Tricho
derm
a sp
. SAPR
O-1
0 Li
ngsa
r
Tricho
derm
a sp
. SAPR
O-1
1 Seleb
ung
Tricho
derm
a sp
. SAPR
O-1
2 Ban
ok
Glio
clad
ium
sp.
SAP
RO-1
3 Ju
rang
Malan
g
Pen
icillium
sp.
SAP
RO-1
5 Celelos
Kon
trol (
tanp
a sa
prof
it)
Jamur saprofit
Masa
inku
basi penya
kit b
usu
k b
ata
ng v
anili
(h
ari
)
Perendaman stek
Investasi media tanah
Gambar 12. Histogram masa inkubasi penyakit busuk batang vanili sebagai akibat
perlakuan jamur saprofit dan cara aplikasinya

31
Adanya perbedaan masa inkubasi penyakit busuk batang karena perlakuan jamur saprofit diduga erat kaitannya kemampuan menghambat yang berbeda dari masing-masing jamur saprofit antagonis terhadap jamur F. oxysporun f. sp. vanillae. Berdasarkan hasil percobaan uji antagonis jamur saprofit secara in-vitro di laboratorium melalui metode oposisi langsung dan uji uap biakan, ternyata persentase hambatan pertumbuhan jamur F. oxysporun f. sp. vanillae yang tertinggi dilakukan oleh jamur Trichoderma spp. disusul oleh Gliocladium spp. dan Penicillium spp. Mekanisme penghambatan terhadap penyakit busuk batang pada pengujian ini diduga terjadi secara fisik (kompetisi ruang dan mikoparasit) dan mengeluarkan antibiotik yang didifusikan ke ruang antar sel dalam jaringan, seperti yang terjadi secara in-vitro pada percobaan di laboratorium. Abd-El Moity dan Shatla (1981) menyatakan bahwa Trichoderma spp. merupakan mikoparasit yang dapat melakukan penetrasi ke miselium dan sclerotia jamur S. rolfsii sehingga terjadi lisis dan pengkristalan. Lebih lanjut Papavizas (1985) menyatakan bahwa mekanisme mikoparasitisme dimulai dengan pelunakan sel inang oleh enzim yang dihasilkan oleh mikoparasit sebelum kerusakan dan kematian sel inang. Menurut Hadar et al. (1979), jamur T. harzianum memproduksi enzim ekstra
selluler ß-(1,3) glucanase dan chitinase yang mampu merusak dinding sel R. solani.
Cook dan Baker (1983) mengatakan bahwa strain tertentu dari Trichoderma menghasilkan antibiotik viridin yang dapat menghambat pertumbuhan jamur lain. Elfina et al. (2001) juga melaporkan bahwa jamur T. harzianum mengeluarkan senyawa anti mikroba yang mampu menghambat pertumbuhan jamur S. rolfsii.
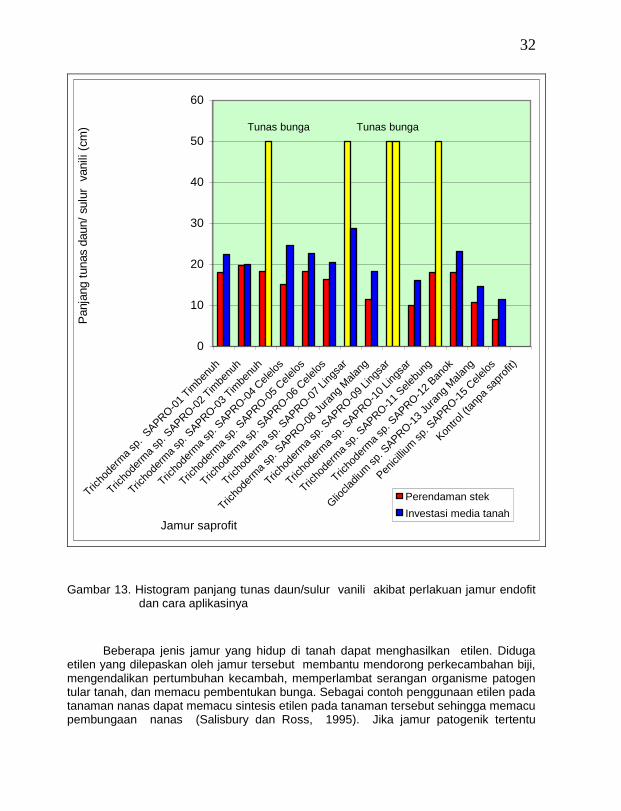
Pada pengujian penggunaan jamur saprofit antagonis dan cara aplikasinya untuk pengendalian penyakit busuk batang vanili, terjadi fenomena menarik sebagai dampak positif penggunaan jamur saprofit antagonis, yaitu ada empat isolat jamur saprofit antagonis yang dapat merangsang pembungaan lebih awal pada fase pembibitan di polibag sedang 10 isolat lainnya hanya merangsang pembentukan tunas daun/sulur (Gambar 12).
Keempat isolat jamur tersebut yaitu Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum) yang diinfestasi ke medium tanah membentuk tunas bunga pada umur 45 hari setelah pemberian antagonis, sedang tiga isolat lainnya membentuk tunas bunga pada umur 30 hari setelah pemberian antonis, yaitu Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum) yang diberikan dengan cara perendaman stek vanili, Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum) dengan cara perendaman stek vanili dan infestasi ke medium tanah, dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum) dengan cara infestasi ke medium tanah (Gambar 13). Penggunaan jamur Trichoderma spp. lainnya dalam hal ini cenderung merangsang
pembentukan tunas daun/ sulur, sedang pada kontrol (tanpa jamur saprofit) bibit vanili terinfeksi penyakit busuk batang. Fenomena ini terjadi diduga karena keempat isolat jamur saprofit Trichoderma spp. ini mengeluarkan substansi kimia atau hormon yang didifusikan ke dalam jaringan tanaman vanili yang dapat memacu pembungaan. Windham et al. (1986) pernah melaporkan bahwa jamur T. harzianum dapat meningkatkan perkecambahan benih dan pertumbuhan tanaman. Tronsmo dan Dennis (1977 dalam Cook dan Baker, 1983) melaporkan bahwa penyemprotan konidia jamur T. viride dan T. polysporum untuk melindungi tanaman strawberi dari penyakit busuk ternyata dapat memacu pembungaan lebih awal.
32
0
10
20
30
40
50
60
Tricho
derm
a sp
. SA
PRO-0
1 Tim
benu
h
Tricho
derm
a sp
. SAP
RO-0
2 Tim
benu
h
Tricho
derm
a sp
. SAP
RO-0
3 Tim
benu
h
Tricho
derm
a sp
. SAP
RO-0
4 Cel
elos
Tricho
derm
a sp
. SAP
RO-0
5 Cel
elos
Tricho
derm
a sp
. SAP
RO-0
6 Cel
elos
Tricho
derm
a sp
. SAP
RO-0
7 Ling
sar
Tricho
derm
a sp
. SAP
RO-0
8 Ju
rang
Malan
g
Tricho
derm
a sp
. SAP
RO-0
9 Ling
sar
Tricho
derm
a sp
. SAP
RO-1
0 Ling
sar
Tricho
derm
a sp
. SAP
RO-1
1 Seleb
ung
Tricho
derm
a sp
. SAP
RO-1
2 Ban
ok
Glio
clad
ium
sp.
SAP
RO-1
3 Ju
rang
Mal
ang
Pen
icillium
sp.
SAP
RO-1
5 Cel
elos
Kon
trol (
tanp
a sa
prof
it)
Jamur saprofit
Panja
ng t
unas
daun/
sulu
r v
anili
(cm
)
Perendaman stek
Investasi media tanah
Tunas bunga Tunas bunga
Gambar 13. Histogram panjang tunas daun/sulur vanili akibat perlakuan jamur endofit
dan cara aplikasinya
Beberapa jenis jamur yang hidup di tanah dapat menghasilkan etilen. Diduga etilen yang dilepaskan oleh jamur tersebut membantu mendorong perkecambahan biji, mengendalikan pertumbuhan kecambah, memperlambat serangan organisme patogen tular tanah, dan memacu pembentukan bunga. Sebagai contoh penggunaan etilen pada tanaman nanas dapat memacu sintesis etilen pada tanaman tersebut sehingga memacu pembungaan nanas (Salisbury dan Ross, 1995). Jika jamur patogenik tertentu

33
menyerang sel, etilen menginduksi tanaman untuk membentuk dua macam enzim yang menguraikan dinding sel jamur tersebut, yaitu ß-(1,3) glucanase dan chitinase (Boller,
1988 dalam Salisbury dan Ross, 1995). Dengan demikian dikatakan bahwa etilen
dapat mengaktifkan mekanisme ketahanan induksi tanaman terhadap jamur patogenik. Selain itu menurut Chet dan Baker (1980 dalam Cook dan Baker, 1983), jamur T.
harzianum dan T. hamatum menghasilkan enzim ß-(1,3) glucanase dan chitinase yang
menyebabkan eksolisis pada hifa inang jamur patogenik jamur Rhizoctonia solani dan Sclerotium rolfsii. Lebih lanjut Chet dan Baker (1980 dalam Cook dan Baker, 1983) mengungkapkan bahwa Jamur T. hamatum juga menghasilkan enzim cellulase, sehingga menambah kemampuannya sebagai mikoparasit pada jamur Phytium spp.
Perlakuan dengan jamur Trichoderma
sp. SAPRO-07 Jurang Malang (T. harzianum)
Perlakuan dengan jamur Trichoderma sp. SAPRO-09 Jurang Malang (T. hamatum)
Gambar 14. Pertumbuhan tunas bunga dan pembuahan pada bibit vanili dengan
perlakuan jamur saprofit Trichoderma spp.

34
4.5. Uji Sinergisme jamur endofit dan saprofit terhadap penyakit busuk batang
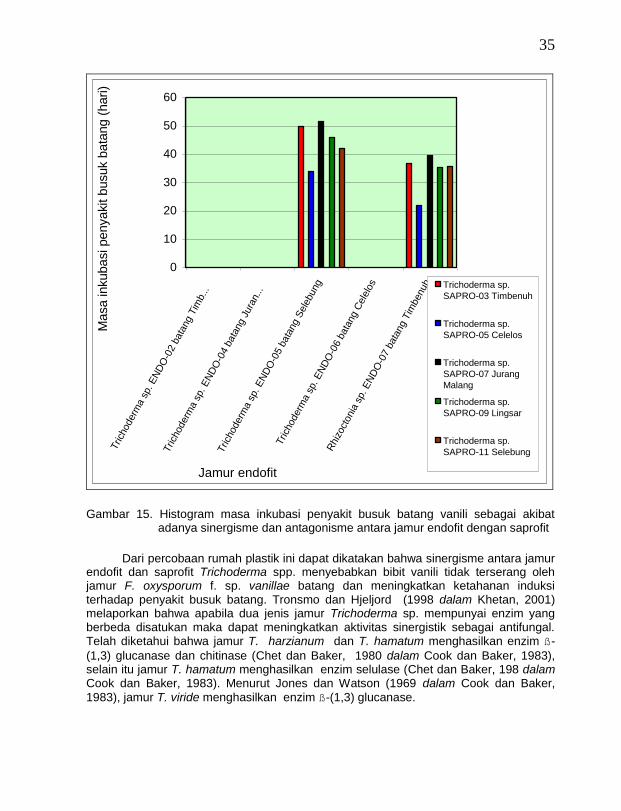
Ada tiga isolat jamur endofit antagonis yaitu Trichoderma sp. ENDO-02 batang
Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang (T. polysporum ) dan Trichoderma sp. ENDO-06 batang Celelos (T. viride) apabila penggunaannya dikombinasikan dengan lima isolat jamur saprofit antagonis yaitu Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-05 vanili Celelos (T. piluliferum), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum), dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum) menyebabkan bibit vanili tidak terinfeksi oleh penyakit busuk batang. Sementara itu ada dua isolat jamur endofit antagonis yaitu Trichoderma sp. ENDO-05 batang Selebung (T. pseudokoningii) dan Rhizoctonia sp. ENDO-07 batang Timbenuh, apabila dikombinasikan dengan kelima isolat jamur saprofit antagonis tersebut menjadi tidak efektif menghambat infeksi penyakit busuk batang, padahal pada percobaan sebelumnya perlakuan dengan isolat jamur endofit antagonis atau saprofit antagonis tersebut secara sendiri-sendiri menyebabkan bibit vanili tidak terinfeksi penyakit busuk batang. Kejadian ini diduga terjadi kompetisi atau saling menghambat antara kedua isolat jamur endofit antagonis tersebut dengan kelima isolat jamur saprofit antagonis tersebut (Gambar 15).
35
0
10
20
30
40
50
60Trich
oderm
a s
p. E
ND
O-0
2 b
ata
ng T
imb...
Trich
oderm
a s
p. E
ND
O-0
4 b
ata
ng J
ura
n...
Trich
oderm
a s
p. E
ND
O-0
5 b
ata
ng S
ele
bung
Trich
oderm
a s
p. E
ND
O-0
6 b
ata
ng C
ele
los
Rhiz
oct
onia
sp. E
ND
O-0
7 b
ata
ng T
imbenuh
Jamur endofit
Masa inkubasi penyakit b
usuk b
ata
ng (
hari)
Trichoderma sp.
SAPRO-03 Timbenuh
Trichoderma sp.
SAPRO-05 Celelos
Trichoderma sp.
SAPRO-07 Jurang
Malang
Trichoderma sp.
SAPRO-09 Lingsar
Trichoderma sp.
SAPRO-11 Selebung
Gambar 15. Histogram masa inkubasi penyakit busuk batang vanili sebagai akibat adanya sinergisme dan antagonisme antara jamur endofit dengan saprofit
Dari percobaan rumah plastik ini dapat dikatakan bahwa sinergisme antara jamur
endofit dan saprofit Trichoderma spp. menyebabkan bibit vanili tidak terserang oleh jamur F. oxysporum f. sp. vanillae batang dan meningkatkan ketahanan induksi terhadap penyakit busuk batang. Tronsmo dan Hjeljord (1998 dalam Khetan, 2001) melaporkan bahwa apabila dua jenis jamur Trichoderma sp. mempunyai enzim yang berbeda disatukan maka dapat meningkatkan aktivitas sinergistik sebagai antifungal. Telah diketahui bahwa jamur T. harzianum dan T. hamatum menghasilkan enzim ß-
(1,3) glucanase dan chitinase (Chet dan Baker, 1980 dalam Cook dan Baker, 1983), selain itu jamur T. hamatum menghasilkan enzim selulase (Chet dan Baker, 198 dalam Cook dan Baker, 1983). Menurut Jones dan Watson (1969 dalam Cook dan Baker,
1983), jamur T. viride menghasilkan enzim ß-(1,3) glucanase.
36
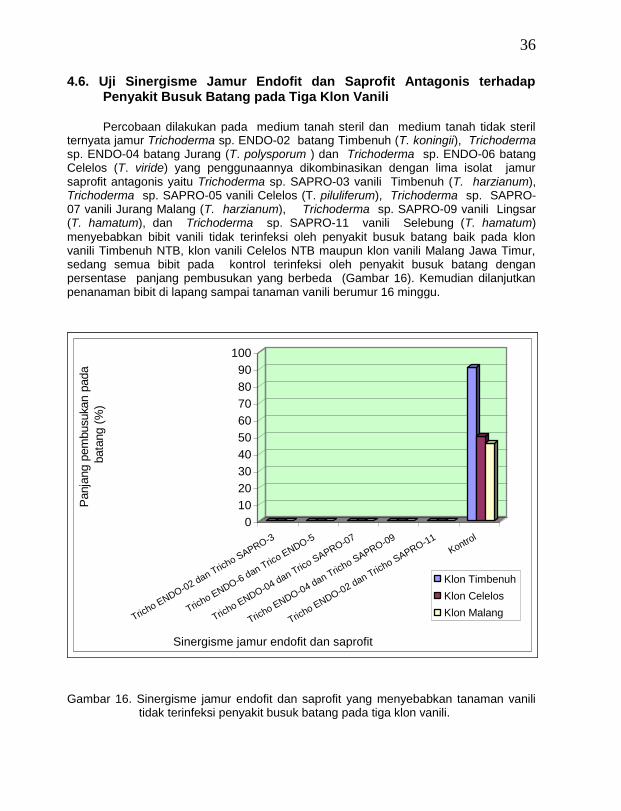
4.6. Uji Sinergisme Jamur Endofit dan Saprofit Antagonis terhadap Penyakit Busuk Batang pada Tiga Klon Vanili
Percobaan dilakukan pada medium tanah steril dan medium tanah tidak steril
ternyata jamur Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang (T. polysporum ) dan Trichoderma sp. ENDO-06 batang Celelos (T. viride) yang penggunaannya dikombinasikan dengan lima isolat jamur saprofit antagonis yaitu Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-05 vanili Celelos (T. piluliferum), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum), dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum)
menyebabkan bibit vanili tidak terinfeksi oleh penyakit busuk batang baik pada klon vanili Timbenuh NTB, klon vanili Celelos NTB maupun klon vanili Malang Jawa Timur, sedang semua bibit pada kontrol terinfeksi oleh penyakit busuk batang dengan persentase panjang pembusukan yang berbeda (Gambar 16). Kemudian dilanjutkan penanaman bibit di lapang sampai tanaman vanili berumur 16 minggu.
0
10
20
30
40
50
60
70
80
90
100
Panja
ng p
em
busuka
n p
ada
bata
ng (
%)
Tricho ENDO-02 dan Tric
ho SAPRO-3
Tricho ENDO-6 dan Tric
o ENDO-5
Tricho ENDO-04 dan Tric
o SAPRO-07
Tricho ENDO-04 dan Tric
ho SAPRO-09
Tricho ENDO-02 dan Tric
ho SAPRO-11
Kontrol
Sinergisme jamur endofit dan saprofit
Klon Timbenuh
Klon Celelos
Klon Malang
Gambar 16. Sinergisme jamur endofit dan saprofit yang menyebabkan tanaman vanili tidak terinfeksi penyakit busuk batang pada tiga klon vanili.

37
Keberhasilan penggunaan jamur Trichoderma spp. sebagai agens pengendalian hayati patogen tanaman di rumah kaca juga dilaporkan oleh beberapa peneliti terdahulu. Marshal (1982 dalam Cook dan Baker, 1983) melaporkan bahwa inokulasi jamur T. harzianum pada benih kacang buncis untuk pengendalian jamur Rhizoctonia solani dapat mengurangi penyakit rebah kecambah sampai 65 %. Harman et al. (1981 dalam Cook dan Baker, 1983) mengatakan bahwa perlakuan benih lobak dengan spora jamur T. hamatum dapat mencegah terjadinya infeksi oleh jamur Phytium spp. Rachmawati, Ambarwati dan Martoredjo (1995) melaporkan bahwa inokulasi jamur T. viride ke dalam medium tanam dapat mencegah bibit vanili terinfeksi penyakit busuk batang. Keberhasilan penggunaan jamur Trichoderma spp. untuk pengendalian penyakit tanaman pada kondisi lapang dilaporkan oleh beberapa penelti terdahulu. Elad et al. (1982 dalam Cook dan Baker, 1983) melaporkan bahwa perlakuan benih kapas dengan jamur T. hamatum atau T. harzianum dapat menurunkan kejadian penyakit rebah kecambah di lapang. Chet dan Baker (1980 dalam Cook dan Baker, 1983) melaporkan bahwa penambahan spora jamur T. harzianum dan T. hamatum ke dalam tanah dapat meniadakan jamur R. solani. Keberhasilan penggunaan jamur Trichoderma spp. dalam pengendalian berbagai penyakit tanaman mendorong dilakukan formulasi komersial seperti jamur T. harzianum dan T. polysporum diproduksi dalam bentuk pelet dengan nama dagang Binab T yang digunakan untuk pengendalian jamur Endothia parasitica dan Verticillium malthousei pada tanaman apel atau T. harzianum dan T. viride diformulasi dengan nama dagang Promote untuk pengendalian jamur Phytium sp., Rhizoctonia sp. dan Fusarium sp. pada tanaman hias dan pembibitan (Kethan, 2001).
Ketahanan induksi tanaman terhadap penyakit setelah diperlakukan dengan jamur endofit non-patogenik dilaporkan oleh beberapa peneliti terdahulu. Biles dan Martyn (1989 dalam Baker dan Paulitz, 1996) melaporkan bahwa inokulasi jamur non-patogenik F. oxysporum dapat menekan keganasan penyakit layu pada tanaman melon. Freeman dan Rodriguez (1993 dalam Baker dan Paulitz, 1996) juga melaporkan bahwa jamur endofit non-patogenik Colletotrichum magna dapat meningkatkan ketahanan induksi bibit melon terhadap jamur F. oxysporum f. sp. niveum. Menurut Wymore dan Baker (1982 dalam Baker dan Paulitz, 1996), mikroorganisme endofit non-patogenik yang menyebabkan ketahanan induksi secara cepat dapat melindungi tanaman dari serangan jamur patogenik di dalam jaringan dengan cara berpindah dari satu tempat ke tempat lainnya, namun tidak selalu mengadakan kontak dengan jamur patogenik. Lebih lanjut Sequeira (1990 dalam Baker dan Paulitz, 1996) mengatakan bahwa segera setelah jamur endofit non-patogenik berada dalam tanaman, maka akan
mengadakan respon dengan merangsang sintesis fitoaleksin oleh enzim ß-(1,3)
glucanase, chitinase dan peroksidase yang dihasilkannya. Fitoaleksin ini menyebabkan terjadinya akumulasi suberin, lignin dan selulosa dalam dinding sel tanaman. Selain itu ke tiga enzim tersebut dapat menghambat secara langsung pertumbuhan jamur patogenik.

38
V. KESIMPULAN DAN SARAN
5.1. Kesimpulan
Berdasarkan hasil dan pembahasan maka dapat diambil suatu kesimpulan sebagai berikut: 1. Pada tanaman sehat ditemukan 19 isolat jamur endofit dan pada rhizosfer ditemukan
19 isolat jamur saprofit dengan karakteristik yang berbeda. 2. Jamur endofit dan saprofit yang ditemukan tumbuh dengan baik pada suhu optimum
25o C dan pH optimum 6,0, kedua kelompok jamur ini dapat tumbuh dengan baik pada seresah daun kopi, kemiri, lamtoro dan gamal.
3. Terdapat delapan isolat jamur endofit dan 12 isolat jamur saprofit efektif menghambat pertumbuhan jamur F. oxysporum f. sp. vanillae secara in-vitro. Penghambatan pertumbuhan oleh jamur endofit dan saprofit dengan cara fisik (kompetisi ruang dan mikoparasit) dan antibiosis (mengeluarkan antibiotik yang mudah menguap).
4. Perendaman bibit vanili sebelum tanam menggunakan suspensi spora jamur antagonis dan infestasi substrat yang mengandung jamur antagonis ke medium tanah (lima isolat jamur endofit dan 10 isolat jamur saprofit hasil isolasi dari kebun vanili Timbenuh, Selebung, Jurang Malang, Lingsar dan Celelos) menyebabkan bibit vanili tidak terinfeksi penyakit busuk batang sampai umur delapan minggu setelah tanam stek.
5. Kombinasi isolat-isolat jamur endofit dan saprofit (tiga isolat jamur endofit dan lima isolat jamur saprofit) menyebabkan bibit vanili tidak terinfeksi penyakit busuk batang baik pada tanah steril maupun tidak steril, dan pada tanaman vanili di lapang baik klon Timbenuh dan Celelos NTB, maupun klon Malang Jawa Timur, juga tidak terinfeksi penyakit busuk batang, sehingga dapat meningkatkan ketahanan induksi terhadap penyakit busuk batang sampai tanaman berumur 16 minggu setelah tanam bibit.
6. Penggunaan empat isolat jamur saprofit dengan cara infestasi ke medium tanah ternyata menyebabkan pembungaan awal pada bibit vanili setelah 30 hari tanam stek vanili di polibag.
5.2. Saran
1. Untuk meningkatkan ketahanan induksi bibit dan tanaman vanili terhadap penyakit busuk batang terutama pada daerah-daerah endemis penyakit busuk batang, maka perlu penggunaan isolat jamur endofit antagonis Trichoderma sp. ENDO-02 batang Timbenuh (T. koningii), Trichoderma sp. ENDO-04 batang Jurang Malang (T. polysporum), dan Trichoderma sp. ENDO-06 batang Celelos (T. viride) dengan cara perendaman stek batang dengan suspensi spora jamur endofit, sedang untuk perlakuan medium tanah dapat digunakan jamur saprofit antagonis Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum) dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum).

39
2. Perlu dilakukan penelitian mekanisme ketahanan induksi bibit dan tanaman vanili terhadap penyakit busuk batang akibat penggunaan jamur endofit dan saprofit antagonis terutama kaitannya dengan kemampuan jamur endofit dan saprofit sebagai mikoparasit dan peran alkaloid yang dihasilkan, sehingga dapat menekan pertumbuhan jamur F. oxysporum f. sp. vanillae.
3. Perlu dilakukan penelitian mekanisme pembungaan lebih awal pada bibit vanili karena rangsangan isolat jamur saprofit antagonis seperti Trichoderma sp. SAPRO-03 vanili Timbenuh (T. harzianum), Trichoderma sp. SAPRO-07 vanili Jurang Malang (T. harzianum), Trichoderma sp. SAPRO-09 vanili Lingsar (T. hamatum) dan Trichoderma sp. SAPRO-11 vanili Selebung (T. hamatum).
DAFTAR PUSTAKA
Abadi, A. L. 1987. Biologi Ganoderma boninense Pat. Pada Kelapa Sawit (Elaes
guineensis Jacq) dan Pengaruh Beberapa Mikroba Tanah Antagonistik Terhadap
Pertumbuhannya. Fakultas Pasca Sarjana IPB. Disertasi (tidak dipublikasikan). 147 hal.
Abadi, A. L. 2003. Ilmu Penyakit Tumbuhan I Edisi Pertama. Bayumedia Publishing dan
Fakultas Pertanian Universitas Brawijaya, Malang Jawa Timur – Indonesia. 137 hal.
Abd-El Moity, H. and M. N. Shatla.1981. Biological Control of White Rot Disease of
Onion (Sclerotium cepivorum) by Trichoderma harzianum. Phytopathologiche Zeitschrift 100: 29 - 35.
Arifin, I. S., B. Dahlan dan U. Dahlan. 1989. Potensi Antagonisme Jamur Tanah pada
Areal tanaman Teh terhadap Jamur Ganoderma pseudoferrum in-vitro. Kongres
Nasional X PFI, Denpasar Bali. Arnold, A. E., Z. Maynard, G. S. Gilbert, P. D. Coley and T. A. Kursar. 2000. A Tropical
Fungal Endophytes Hyperdiverse ?. (Abstract). http://www.ingentaconnect.com/;isesionid=m0ca7cutigbp.victoria, (19 Maret 2005).
Balai Laboratorium Perlindungan Tanaman Perkebunan NTB, 2003. Laporan Tahunan
Balai Laboratorium Perlindungan Tanaman Perkebunan NTB. Tahun 2003. Mataram.
Baker, K. F. and R. J. Cook. 1982. Biological Control of Plant Pathogens. The American
Phytopathology Society. Minnessota. 433 p.

40
Baker, R. and T. C. Paulitz. 1996. Theoretical Basis for Microbial Interactions Leading to Biological Control of Soilborne Plant Pathogens. In. Hal. R (Ed.) Principles and
Practice of Managing Soilborne Plant Pathogens. APS Press, The American Phytopathological Society. St. Paul, Minnesota. 330 p.
Barnett, H. L. And B. B. Hunter. 1998. Illustrated Genera of Imperfect Fungi. Fourth
Edition. APS Press, The American Phytopathological Society, St. Paul, Minnesota.218 p.
Basuki. 1985. Peranan Belerang Sebagai Pemacu Pengendalian Biologi Penyakit Akar
Putih Pada Karet. Disertasi Doktor, UGM Yogyakarta. Bharat, R., R. S. Upadhayay and A. K. Srivastava. 1988. Utilization of Cellulose and
Gallic Acid by Litter Inhabiting Fungi and Its Possible Implication in Litter Decomposition of A Tropical Deciduous Forest, Pedobiologia. Dept. Bot. Banaes Hindu University, Varanasi, India.
Brunner, F. and O. Petrini. 1992. Taxonomic Studies of Xylaria jenis and Xylariaceous
Endophytes by Izozyme Electrophoresis. Mycological Research 96: 723 – 733.
Budi, I. S. Mariana and Rachmadi. 2005. Exploration of Tidal Swamp Rice Endophytic
Fungi from South Kalimantan and Biological Control of Rhizoctonia solani. In
Program and Abstract The 1st International Conference of Crop Security, Brawijaya University, Malang, September 20th – 22nd, 2005. 264 p.
Burgess, L. W., C. M. Liddell and B. A. Summerell. 1988. Laboratory Mannual For
Fusarium Research. Second Edition. Fusarium Research Laboratory Departement of Plant Pathology and Agricultural Entomology The University of Sydney. 156 p.
Carling, D. E. and D. R. Sumner, 1992. Rhizoctonia. In Singleton, L. L., J. D. Mihail and
C. M. Rush (Ed). Methods for Research on Soilborne Phytopathogenic Fungi. APS Press, The American Phytopathological Society, St. Paul, Minnesota. 157 – 165.
Cook, R. J. and K. F. Baker. 1983. The Nature and Practice of Biological Control of
Plant Pathogens. The American Phytopathol. Society, St. Paul MN. 539 p.
Dahlam, D. L., H. Eichenseer and M. R. Siegel. 1991. Chemical Perspectives on Endophyte-Grass Interaction and Their Implications to Insect Herbivory. In Barbosa, P., V. A. Krichil and C. G. Jones (Ed). Microbial Mediation of Plant-Herbivore Interaction, Jhon Wiley & Sons Inc., New York: 227 – 252.
Domsch, K. H.; W. Gams and T. Anderson. 1980. Compendium of Soil Fungi. Academic
Press. New York. 859 p.

41
Elfina, Y., Mardinus, T. Habazar dan A. Bachtiar. 2001. Studi Kemampuan Isolat-isolat Jamur Trichoderma spp. yang Beredar di Sumatera Barat untuk Pengendalian Jamur Patogen Sclerotium rolfsii pada Bibit Cabai. Dalam Purwantara, A. et al. (Penyunting), Prosiding Kongres Nasional XVI dan Seminar Ilmiah PFI, di Bogor. 167 - 173.
Ghimire, S. R. and K. D. Hyde, 2004. Fungal Endophyt. In Varma, A., L. Abboot, D.
Werner and R. Hampp (Ed.), Plant Surface Microbiology Springer-Verlag, Berlin: 281 – 292.
Guest, D. 2005. Induced Disease Resistance in Plants. In Program and Abstract The 1st
International Conference of Crop Security 2005, Brawijaya University, Malang, September 20th – 22nd, 2005. 264 p.
Hadar, Y.; I Chet and Y. Henis. 1979. Biological Control of Rhizoctonia solani Damping-
Off with Wheat Bran Culture of Trichoderma harzianum. Phytopathology 69 ; 64 -
69. Hadisutrisno, B. 2005. Budidaya Vanili Tahan Penyakit Busuk Batang. Penerbit Penebar
Swadaya, Depok. 87 p. Harman, G. E. and A. Taylor, 1988. Improved seedling performance by integration of
biological control agents at favourable pH levels with solid matrix priming. Phytopatholgy 78: 520 – 525.
Hoitink, H. A. J., L. V. Madden and M. J. Boehm. 1996. Relationships Among Organic
Matter Decomposition Level, Microbial Species Diversity, and Soilborne Disease Severity. In. Hal. R (Ed.) Principles and Practice of Managing Soilborne Plant Pathogens. APS Press, The American Phytopathological Society. St. Paul, Minnesota. 330 p.
Irawati, A. F. C. 2005. Characterization and Hypovirulent Test of Rhizoctonia sp. from
Heahlty Vanilla Roots. Paper Presented on The 1st International Conference of Crop Security 2005, Brawijaya University, Malang, September 20th – 22nd, 2005. 17 p.
Kethan, S. K. 2001. Microbial Pest Control. Marcel Dekker, Inc. New York – Basel. 300 p.
Lumyong, S., P. Lumyong and K. D. Hyde, 2004. Endophytes. In Jones, E. B. G., M.
Tantichareon and K. D. Hyde (Ed.), Thai Fungal Diversity. Published by BIOTEC Thailand and Biodiversity Research and Training Program (BRTI/TRF. Biotec). 197 – 212.
Manoch, L. 2004. Soil Fungi. In Jones, E. B. G., M. Tantichareon and K. D. Hyde (Ed.),
Thai Fungal Diversity. Published by BIOTEC Thailand and Biodiversity Research and Training Program (BRTI/TRF. Biotec). 141 – 154.

42
Moy, M., H. M. Li, R. Sullivan, J. F. White Jr, and F. C. Belanger. 2002. Endophytic Fungal β-1,6-Glucanase Expression in the Infected Host Grass. Plant Physiol. Vol. 130: 1298 – 1308. http://www.plantphysiol.org/cgi/content/full/130/3/1298,
(18 Maret 2005). Nurawan, A., M. Tombe dan K. Matsumoto. 1995. Penelitian Daya Antagonisme Isolat
Bakteri yang Diisolasi Dari Rhizosfera Berbagai Jenis Tanaman Terhadap Patogen Busuk Batang Vanili. Dalam Parman et al. (Penyunting), Peran Fitopatologi dalam Pembangunan Pertanian Berkelanjutan di Kawasan Timur Indonesia. Risalah Kongres Nasional XIII dan Seminar Ilmiah Perhimpunan Fitopatologi Indonesia di Mataram. 356 – 359.
Papavizas, G. C. 1985. Trichoderma and Gliocladium: Biology, Ecology and Potential for
Biocontrol. Ann. Rev. Phytopathology 23: 23 - 54. Petrini, O. 1993. Endophyt of Pteridium spp.: Some Considerations for Biological
Control. Sydowia 45: 330 –338.
Rahmawati, A., H. T. Ambarwati dan T. Martoredjo. 1995. Kajian Pengendalian Penyakit
Busuk Batang Vanili dengan Trichoderma viride. Dalam Parman et al. (Penyunting), Risalah Kongres Nasional XIII dan Seminar Ilmiah Perhimpunan Fitopatologi Indonesia di Mataram. 207 – 213.
Rifai, M. A. 1969. A revision of the marga Trichoderma. Commonwealth Mycological
Institute, Mycol. Papers 116: 1 - 56.
Ruhnayat, A. 2004. Kiat Mengatasi Permasalahan Praktis Bertanam Vanili Si Emas Hijau nan Wangi. Agromedia Pustaka, Jakarta. 51 hal.
Salisbury, F. B. Dan C. W. Ross, 1995. Fisiology Tumbuhan Jilid 3. Perkembangan tumbuhan dan fisiologi Tumbuhan (Terjemahan D. R. Lukman dan Sumaryono). Penerbit ITB Bandung.
Sastrahidayat, I. R. 1991. Penggunaan Energi Sinar Matahari dan Mikroorganisme Untuk Menanggulangi Serangan Fusarium batatis var. vanillae Penyebab Penyakit Busuk Batang pada Tanaman Vanili di Pesemaian. Dalam Sarbini, G. et al. (Penyunting), Prosiding Kongres Nasional XI dan Seminar Ilmiah PFI di Ujung Pandang. 201 – 206.
Sudantha, I. M. 2005. Laporan Hasil Survei Pendahuluan Penyakit Busuk Batang Vanili di Dusun Timbenuh Desa Pengadangan Kecamatan Pringgasela Lombok Timur dan Dusun Celelos Desa Bentek Kecamatan Gangga Lombok Barat dalam Rangka Penyusunan Proposal Disertasi Program Pascasarjana Universitas Brawijaya Malang. Fakultas Pertanian Universitas Mataram, Mataram.

43
Sukamto dan M. Tombe. 1995. Antagonisme Trichoderma viride terhadap Fusarium oxysporum f. sp. vanillae secara In-Vitro. Dalam Parman et al. (Penyunting),
Peran Fitopatologi dalam Pembangunan Pertanian Berkelanjutan di Kawasan Timur Indonesia. Risalah Kongres Nasional XIII dan Seminar Ilmiah Perhimpunan Fitopatologi Indonesia di Mataram. 600 – 604.
Sulistyowati, L., N. F. Deci and A. R. Gendall. 2005. Isolation and Sequencing of
Chitinase and Glucanase Genes of Endophytic Trichoderma asperellum from Citrus Stem. In Program and Abstract The 1st International Conference of Crop Security 2005, Brawijaya University, Malang, September 20th – 22nd, 2005. 264 p.
Trautman, N. and E. Olynciw, 1996. Compost microorganism. Cornell Composting.
Science and Engineering. Cornell University. 16 hal. Widyastuti, S. M., Sumardi dan N. Hidayat. 1998. Kemampuan Trichoderma spp. untuk
Pengendalian Hayati Jamur Akar Putih pada Acacia mangium secara In-vitro. Buletin Kehutanan No. 36. 24 – 38.
Windham, M., Y. Elad and R. Baker. 1986. A Mechanism of Increased Plant Growth
Induced by Trichoderma spp. Phytopathology 76: 518 - 521.

44
Lampiran 5. Karakterisasi isolat jamur endofit yang ditemukan pada jaringan akar tanah, akar udara, batang, daun dan buah tanaman vanili di Pulau Lombok NTB
No. Nama Isolat
Karakerisasi makroskopis Karakterisasi mikroskopis
1 Trichoderma sp. isolat ENDO-01 akar tanah vanili Timbenuh (Trichoderma viride Pers. Ex S. F. Gray aggr.)
Koloni jamur tumbuh cepat, tiga hari setelah inokulasi telah menutupi cawan Petri (90,00 mm). Mula-mula koloni jamur berwarna putih, setelah terbentuk konidia berubah menjadi hijau tua sampai hijau kebiruan. Koloni tumbuh tebal dan padat.
Miselia hyaline, bersekat, bercabang, Phialospore berbentuk bulat berwarna hijau, berdiameter 3 – 5 µ. Konidiofor bercabang banyak. Phialide terbentuk lebih 2 – 3 pada ujung percabangan konidiofor, tiap ujung phialide terbentuk phialospore
Morfologi T. viride (Rifai, 1969) Koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
2 Trichoderma sp. isolat ENDO-02 batang vanili Timbenuh (Trichoderma koningii Oud. aggr.)
Koloni jamur tumbuh cepat tiga hari setelah inokulasi menutupi cawan Petri (90 mm). Mula-mula berwarna putih dan setelah terbentuk phialospore berubah menjadi putih kehijauan sampai hijau tua. Koloni tumbuh tebal dan padat.
Miselia hyaline, bercabang banyak dan bersekat., Phialospore bentuk bulat
sampai elips berukuran 3 – 5 µ x 2 – 3 µ, berwarna hijau muda. Pada cabang utama konidiofor terdapat 2 – 3 kelompok cabang. Sistem percabangan seperti piramid. Phialide bentuk kerucut, pada ujungnya terdapat phialospore.
10 µ
1
2
3
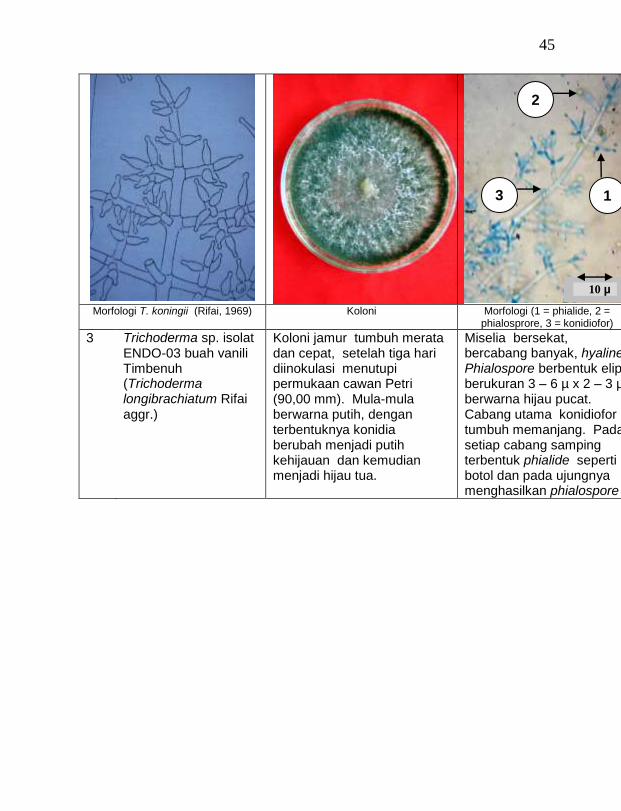
45
Morfologi T. koningii (Rifai, 1969) Koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
3 Trichoderma sp. isolat ENDO-03 buah vanili Timbenuh (Trichoderma longibrachiatum Rifai aggr.)
Koloni jamur tumbuh merata dan cepat, setelah tiga hari diinokulasi menutupi permukaan cawan Petri (90,00 mm). Mula-mula berwarna putih, dengan terbentuknya konidia berubah menjadi putih kehijauan dan kemudian menjadi hijau tua.
Miselia bersekat, bercabang banyak, hyaline Phialospore berbentuk elips berukuran 3 – 6 µ x 2 – 3 µ berwarna hijau pucat. Cabang utama konidiofor tumbuh memanjang. Pada setiap cabang samping terbentuk phialide seperti botol dan pada ujungnya menghasilkan phialospore
10 µ
2
1 3

46
Morfologi T. longibrachiatum
(Rifai, 1969) Koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
4 Trichoderma sp. isolat ENDO-04 batang vanili Jurang Malang (Trichoderma polysporum (Link ex Pers.) Rifai aggr.)
Koloni jamur ini pada medium PDA tumbuh cepat, menutupi seluruh permukaan cawan (90,00 mm) setelah tiga hari inokulasi dengan miselia berwarna putih dan tetap berwarna putih pada koloni yang sudah tua.
Hifa bercabang, hyalin, bersepta. Pada konidiofor fertil terdapat banyak cabang dan pada ujungnya terdapat phialide, konidiofor steril tumbuh memanjang dengan cabang antara 2 – 3. Bentuk phialide seperti buah per, berukuran 4 – 6,5 x 3 – 3,5 µ. Phialospore tidak berwarna berbentuk elips berukuran 2,7 – 3,7 x 1,5 – 2,2 µ
10 µ
3
1
2
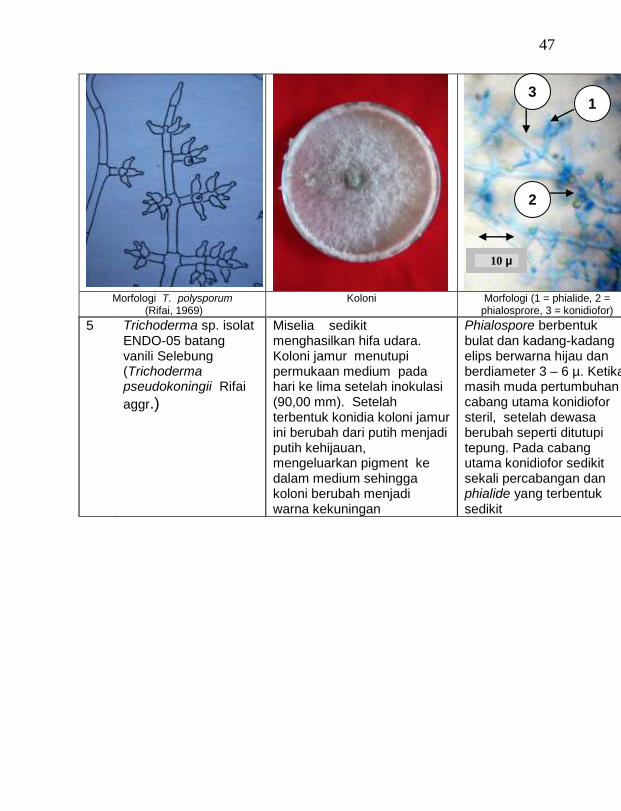
47
Morfologi T. polysporum
(Rifai, 1969) Koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
5 Trichoderma sp. isolat ENDO-05 batang vanili Selebung (Trichoderma pseudokoningii Rifai
aggr.)
Miselia sedikit menghasilkan hifa udara. Koloni jamur menutupi permukaan medium pada hari ke lima setelah inokulasi (90,00 mm). Setelah terbentuk konidia koloni jamur ini berubah dari putih menjadi putih kehijauan, mengeluarkan pigment ke dalam medium sehingga koloni berubah menjadi warna kekuningan
Phialospore berbentuk bulat dan kadang-kadang elips berwarna hijau dan berdiameter 3 – 6 µ. Ketika masih muda pertumbuhan cabang utama konidiofor steril, setelah dewasa berubah seperti ditutupi tepung. Pada cabang utama konidiofor sedikit sekali percabangan dan phialide yang terbentuk sedikit
10 µ
1
2
3
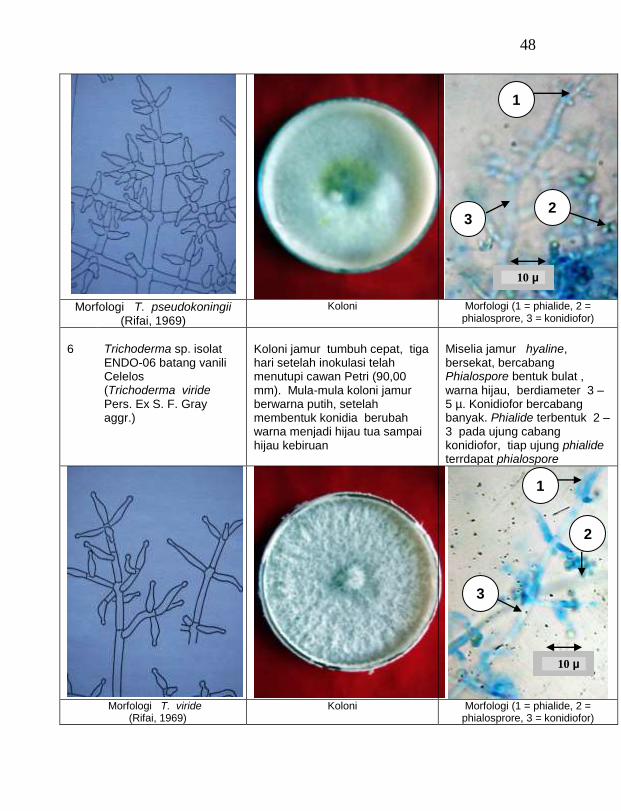
48
Morfologi T. pseudokoningii
(Rifai, 1969)
Koloni Morfologi (1 = phialide, 2 = phialosprore, 3 = konidiofor)
6
Trichoderma sp. isolat ENDO-06 batang vanili Celelos (Trichoderma viride Pers. Ex S. F. Gray aggr.)
Koloni jamur tumbuh cepat, tiga hari setelah inokulasi telah menutupi cawan Petri (90,00 mm). Mula-mula koloni jamur berwarna putih, setelah membentuk konidia berubah warna menjadi hijau tua sampai hijau kebiruan
Miselia jamur hyaline, bersekat, bercabang Phialospore bentuk bulat ,
warna hijau, berdiameter 3 – 5 µ. Konidiofor bercabang banyak. Phialide terbentuk 2 – 3 pada ujung cabang konidiofor, tiap ujung phialide terrdapat phialospore
Morfologi T. viride
(Rifai, 1969) Koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
10 µ
1
2 3
10 µ
3
2
1

49
7 Rhizoctonia sp. isolat ENDO-07 batang vanili Timbenuh (Rhizoctonia sp.)
Koloni jamur tumbuh cepat, tiga hari setelah inokulasi telah menutupi cawan Petri (90,00 mm). Mula-mula koloni jamur berwarna putih, kemudian berubah menjadi coklat kehitaman.
Jamur mempunyai hifa yang kasar dan panjang, berwarna hyaline sampai hitam, tidak mempunyai konidia (miselium steril). Miselia bercabang siku-siku, membentuk sclerotia berwarna hitam di atas permukaan koloni
.Morfologi Rhizoctonia sp. (Barnett
dan Hunter, 1998). Koloni Morfologi (1 = hifa siku)
8
Rhizoctonia sp. isolat ENDO-08 batang Selebung (Rhizoctonia sp.)
Koloni jamur tumbuh cepat, tiga hari setelah inokulasi telah menutupi cawan Petri (90,00 mm). Mula-mula koloni jamur berwarna putih, kemudian berubah menjadi coklat kehitaman.
Jamur mempunyai hifa yang kasar dan panjang, berwarna hyaline sampai hitam, tidak mempunyai konidia (miselium steril). Miselia bercabang siku-siku, membentuk sclerotia berwarna hitam di atas permukaan koloni
1
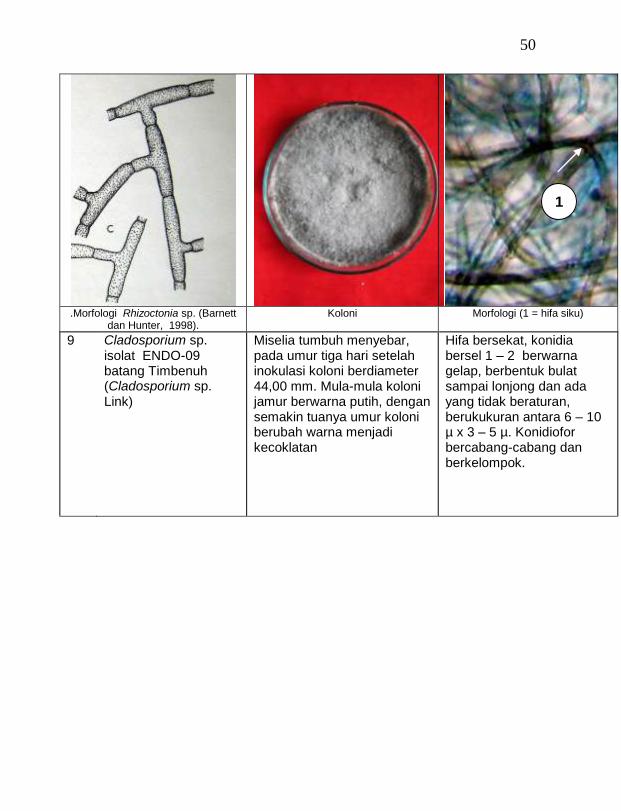
50
.Morfologi Rhizoctonia sp. (Barnett
dan Hunter, 1998). Koloni Morfologi (1 = hifa siku)
9 Cladosporium sp. isolat ENDO-09 batang Timbenuh (Cladosporium sp. Link)
Miselia tumbuh menyebar, pada umur tiga hari setelah inokulasi koloni berdiameter 44,00 mm. Mula-mula koloni jamur berwarna putih, dengan semakin tuanya umur koloni berubah warna menjadi kecoklatan
Hifa bersekat, konidia bersel 1 – 2 berwarna gelap, berbentuk bulat sampai lonjong dan ada yang tidak beraturan, berukukuran antara 6 – 10 µ x 3 – 5 µ. Konidiofor bercabang-cabang dan berkelompok.
1
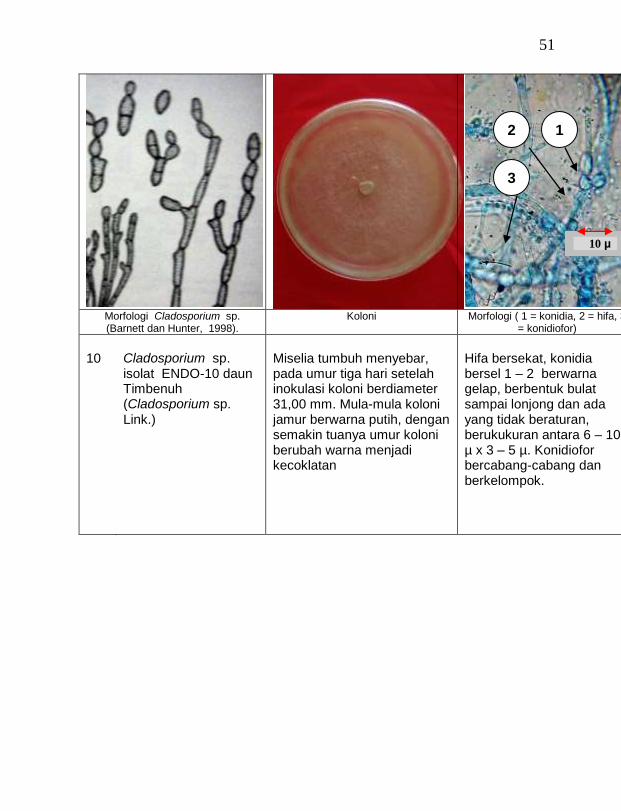
51
Morfologi Cladosporium sp. (Barnett dan Hunter, 1998).
Koloni Morfologi ( 1 = konidia, 2 = hifa, 3 = konidiofor)
10
Cladosporium sp. isolat ENDO-10 daun Timbenuh (Cladosporium sp. Link.)
Miselia tumbuh menyebar, pada umur tiga hari setelah inokulasi koloni berdiameter 31,00 mm. Mula-mula koloni jamur berwarna putih, dengan semakin tuanya umur koloni berubah warna menjadi kecoklatan
Hifa bersekat, konidia bersel 1 – 2 berwarna gelap, berbentuk bulat sampai lonjong dan ada yang tidak beraturan, berukukuran antara 6 – 10 µ x 3 – 5 µ. Konidiofor bercabang-cabang dan berkelompok.
10 µ
2 1
3
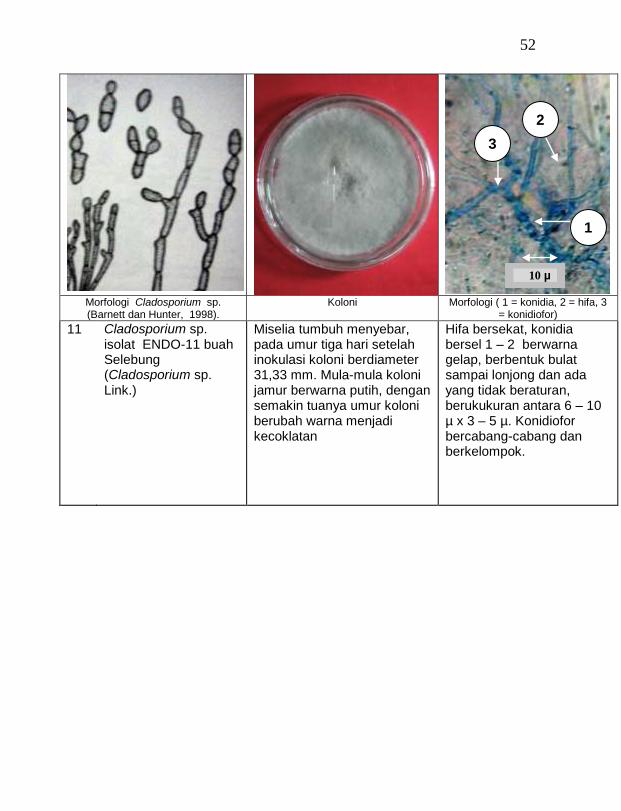
52
Morfologi Cladosporium sp. (Barnett dan Hunter, 1998).
Koloni Morfologi ( 1 = konidia, 2 = hifa, 3 = konidiofor)
11 Cladosporium sp. isolat ENDO-11 buah Selebung (Cladosporium sp. Link.)
Miselia tumbuh menyebar, pada umur tiga hari setelah inokulasi koloni berdiameter 31,33 mm. Mula-mula koloni jamur berwarna putih, dengan semakin tuanya umur koloni berubah warna menjadi kecoklatan
Hifa bersekat, konidia bersel 1 – 2 berwarna gelap, berbentuk bulat sampai lonjong dan ada yang tidak beraturan, berukukuran antara 6 – 10 µ x 3 – 5 µ. Konidiofor bercabang-cabang dan berkelompok.
10 µ
3
1
2
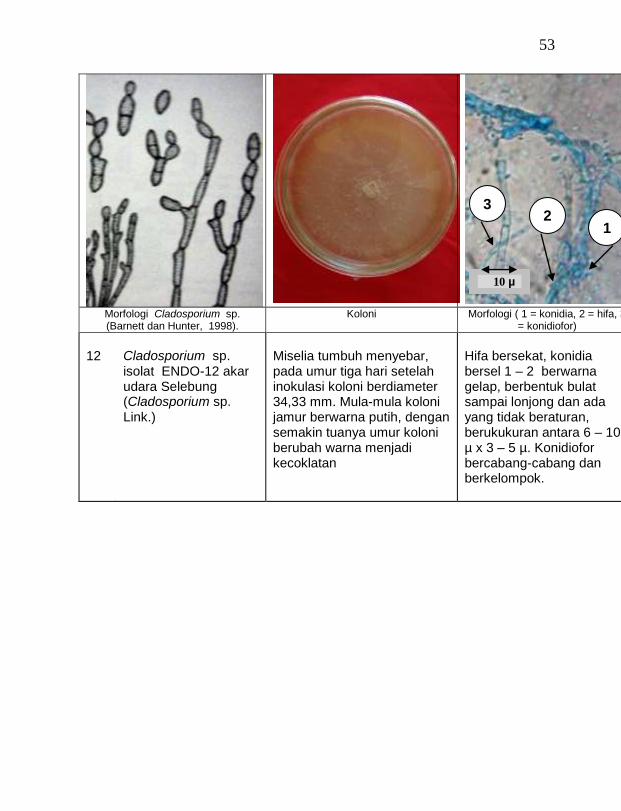
53
Morfologi Cladosporium sp. (Barnett dan Hunter, 1998).
Koloni Morfologi ( 1 = konidia, 2 = hifa, 3 = konidiofor)
12
Cladosporium sp. isolat ENDO-12 akar udara Selebung (Cladosporium sp. Link.)
Miselia tumbuh menyebar, pada umur tiga hari setelah inokulasi koloni berdiameter 34,33 mm. Mula-mula koloni jamur berwarna putih, dengan semakin tuanya umur koloni berubah warna menjadi kecoklatan
Hifa bersekat, konidia bersel 1 – 2 berwarna gelap, berbentuk bulat sampai lonjong dan ada yang tidak beraturan, berukukuran antara 6 – 10 µ x 3 – 5 µ. Konidiofor bercabang-cabang dan berkelompok.
10 µ
1 2
3
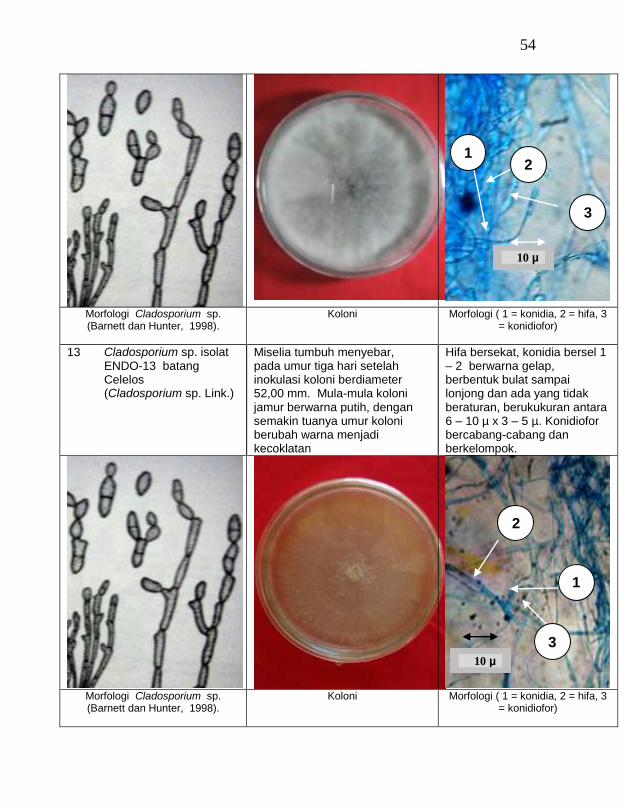
54
Morfologi Cladosporium sp. (Barnett dan Hunter, 1998).
Koloni Morfologi ( 1 = konidia, 2 = hifa, 3 = konidiofor)
13 Cladosporium sp. isolat ENDO-13 batang Celelos (Cladosporium sp. Link.)
Miselia tumbuh menyebar, pada umur tiga hari setelah inokulasi koloni berdiameter 52,00 mm. Mula-mula koloni jamur berwarna putih, dengan semakin tuanya umur koloni berubah warna menjadi kecoklatan
Hifa bersekat, konidia bersel 1 – 2 berwarna gelap, berbentuk bulat sampai lonjong dan ada yang tidak beraturan, berukukuran antara 6 – 10 µ x 3 – 5 µ. Konidiofor bercabang-cabang dan berkelompok.
Morfologi Cladosporium sp. (Barnett dan Hunter, 1998).
Koloni Morfologi ( 1 = konidia, 2 = hifa, 3 = konidiofor)
10 µ
1 2
3
10 µ
1
3
2
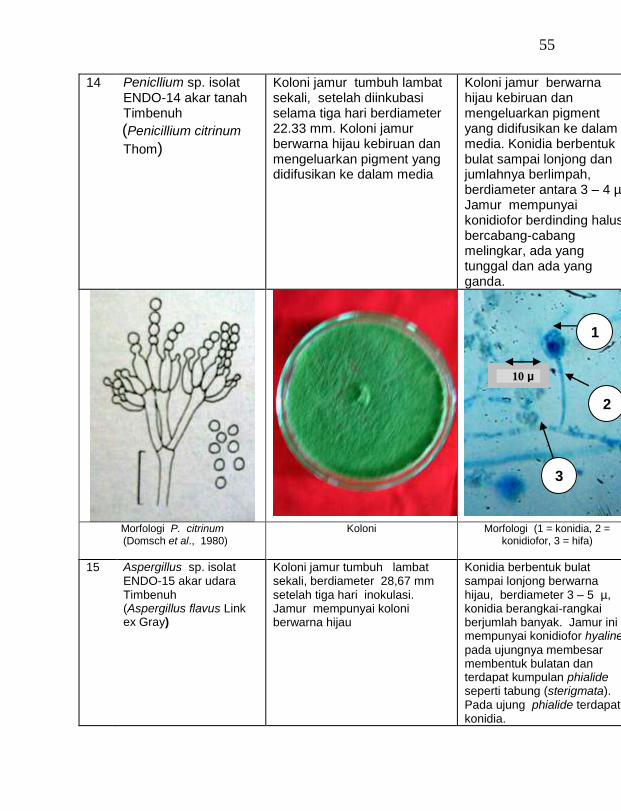
55
14 Penicllium sp. isolat ENDO-14 akar tanah Timbenuh
(Penicillium citrinum
Thom)
Koloni jamur tumbuh lambat sekali, setelah diinkubasi selama tiga hari berdiameter 22.33 mm. Koloni jamur berwarna hijau kebiruan dan mengeluarkan pigment yang didifusikan ke dalam media
Koloni jamur berwarna hijau kebiruan dan mengeluarkan pigment yang didifusikan ke dalam media. Konidia berbentuk bulat sampai lonjong dan jumlahnya berlimpah, berdiameter antara 3 – 4 µ. Jamur mempunyai konidiofor berdinding halus, bercabang-cabang melingkar, ada yang tunggal dan ada yang ganda.
Morfologi P. citrinum (Domsch et al., 1980)
Koloni Morfologi (1 = konidia, 2 = konidiofor, 3 = hifa)
15 Aspergillus sp. isolat ENDO-15 akar udara Timbenuh (Aspergillus flavus Link ex Gray)
Koloni jamur tumbuh lambat sekali, berdiameter 28,67 mm setelah tiga hari inokulasi. Jamur mempunyai koloni berwarna hijau
Konidia berbentuk bulat sampai lonjong berwarna hijau, berdiameter 3 – 5 µ, konidia berangkai-rangkai berjumlah banyak. Jamur ini mempunyai konidiofor hyaline,
pada ujungnya membesar membentuk bulatan dan terdapat kumpulan phialide seperti tabung (sterigmata). Pada ujung phialide terdapat konidia.
10 µ
1
2
3

56
Morfologi A. flavus (Domsch et al., 1980)
Koloni Morfologi (1 = konidia, 2 = kumpulan phialide, 3 = konidiofor)
16 Aspergillus sp. isolat ENDO-16 daun Timbenuh (Aspergillus flavus
Link ex Gray)
Koloni jamur tumbuh lambat sekali, berdiameter 28,33 mm setelah tiga hari inokulasi. Jamur mempunyai koloni berwarna hijau
Konidia berbentuk bulat sampai lonjong berwarna hijau berdiameter 3 – 5 µ, konidia berangkai-rangkai berjumlah banyak. Jamur ini mempunyai konidiofor hyaline, pada ujungnya membesar membentuk bulatan dan terdapat kumpulan phialide seperti tabung (sterigmata). Pada ujung phialide terdapat konidia.
10 µ
2
3
1
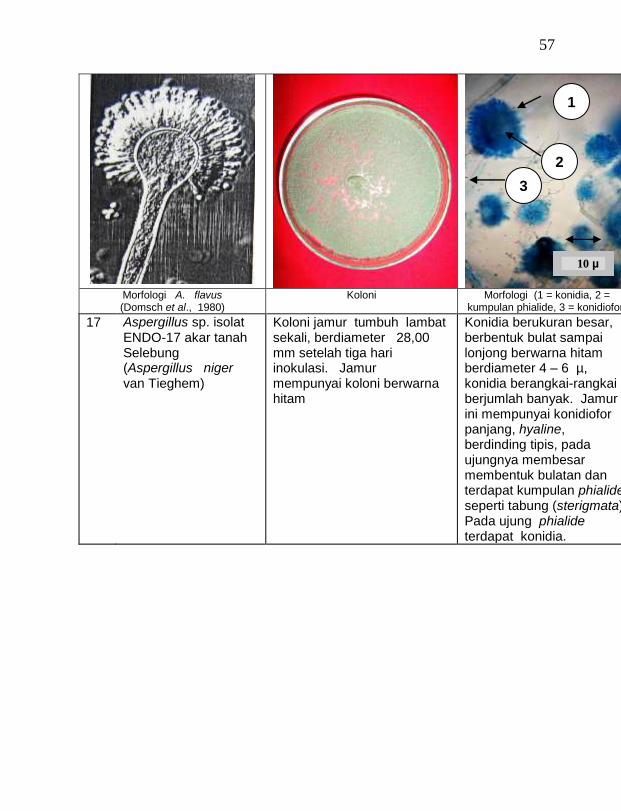
57
Morfologi A. flavus (Domsch et al., 1980)
Koloni Morfologi (1 = konidia, 2 = kumpulan phialide, 3 = konidiofor)
17 Aspergillus sp. isolat ENDO-17 akar tanah Selebung (Aspergillus niger van Tieghem)
Koloni jamur tumbuh lambat sekali, berdiameter 28,00 mm setelah tiga hari inokulasi. Jamur mempunyai koloni berwarna hitam
Konidia berukuran besar, berbentuk bulat sampai lonjong berwarna hitam berdiameter 4 – 6 µ, konidia berangkai-rangkai berjumlah banyak. Jamur ini mempunyai konidiofor panjang, hyaline, berdinding tipis, pada ujungnya membesar membentuk bulatan dan terdapat kumpulan phialide seperti tabung (sterigmata). Pada ujung phialide terdapat konidia.
10 µ
1
3
2
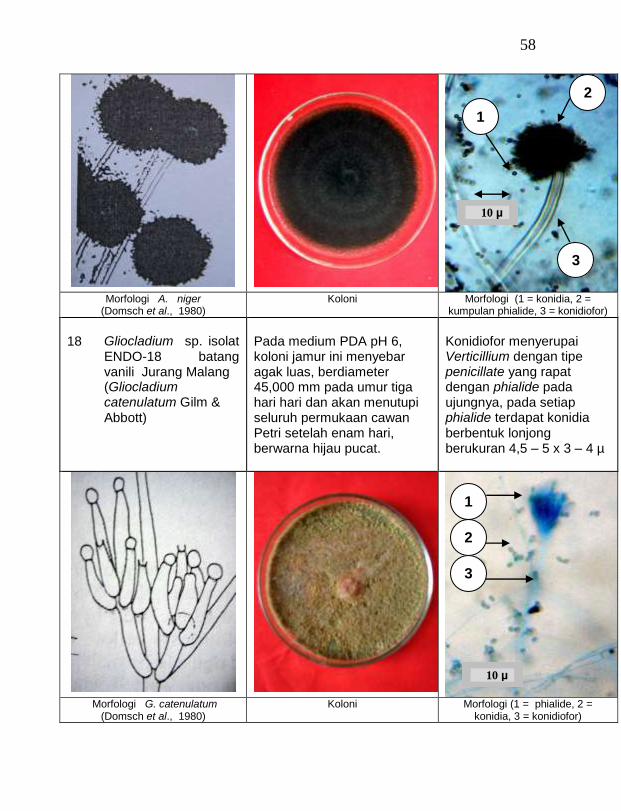
58
Morfologi A. niger
(Domsch et al., 1980) Koloni Morfologi (1 = konidia, 2 =
kumpulan phialide, 3 = konidiofor)
18
Gliocladium sp. isolat ENDO-18 batang vanili Jurang Malang (Gliocladium catenulatum Gilm & Abbott)
Pada medium PDA pH 6, koloni jamur ini menyebar agak luas, berdiameter 45,000 mm pada umur tiga hari hari dan akan menutupi seluruh permukaan cawan Petri setelah enam hari, berwarna hijau pucat.
Konidiofor menyerupai Verticillium dengan tipe penicillate yang rapat dengan phialide pada ujungnya, pada setiap phialide terdapat konidia berbentuk lonjong berukuran 4,5 – 5 x 3 – 4 µ
Morfologi G. catenulatum
(Domsch et al., 1980) Koloni Morfologi (1 = phialide, 2 =
konidia, 3 = konidiofor)
10 µ
1
3
2
10 µ
2
3
1
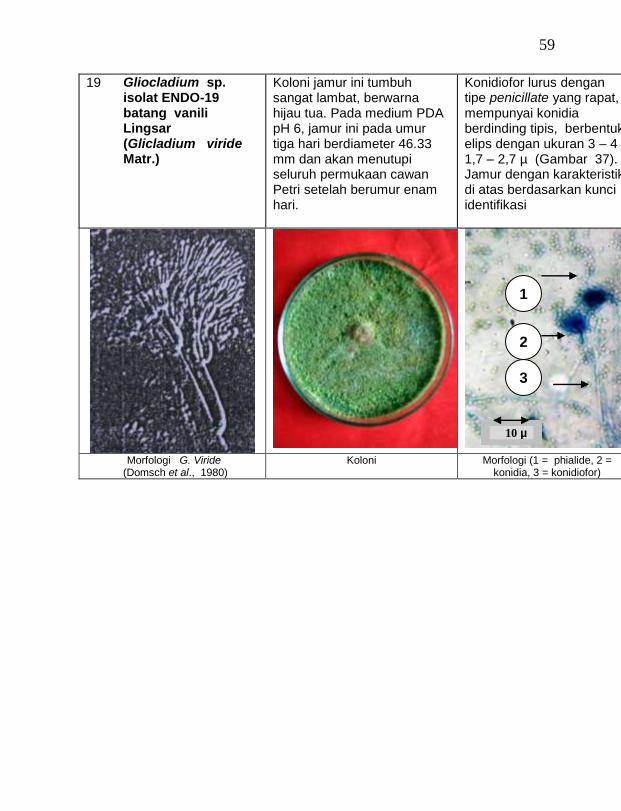
59
19 Gliocladium sp. isolat ENDO-19 batang vanili Lingsar (Glicladium viride Matr.)
Koloni jamur ini tumbuh sangat lambat, berwarna hijau tua. Pada medium PDA pH 6, jamur ini pada umur tiga hari berdiameter 46.33 mm dan akan menutupi seluruh permukaan cawan Petri setelah berumur enam hari.
Konidiofor lurus dengan tipe penicillate yang rapat, mempunyai konidia berdinding tipis, berbentuk elips dengan ukuran 3 – 4 x 1,7 – 2,7 µ (Gambar 37). Jamur dengan karakteristik di atas berdasarkan kunci identifikasi
Morfologi G. Viride (Domsch et al., 1980)
Koloni Morfologi (1 = phialide, 2 = konidia, 3 = konidiofor)
10 µ
3
2
1
60
Lampiran 6. Karakterisasi isolat jamur saprofit yang ditemukan pada rhizosfer atau daerah sekitar perakaran tanaman vanili di Pulau Lombok NTB
No. Jenis jamur saprofit
Karakerisasi makroskopis Karakterisasi mikroskopis
1 Trichoderma sp.
SAPRO-01 vanili Timbenuh Trichoderma viride Pers. Ex S. F. Gray aggr.
Koloni jamur tumbuh cepat, tiga hari setelah inokulasi menutupi cawan Petri (90,00 mm). Koloni jamur berwarna putih, setelah terbentuk konidia berubah menjadi hijau tua sampai hijau kebiruan. Koloni tumbuh tebal dan padat.
Miselia hyaline, berdinding
halus, bersekat, bercabang, Phialospore bentuk bulat, warna hijau dan diameter 3 – 5 µ. Konidiofor bercabang banyak. Phialide terbentuk 2 – 3 pada ujung percabangan konidiofor, dan pada tiap ujung phialide terbentuk phialospore
Morfologi T. viride
(Rifai, 1969) koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
2 Trichoderma sp. SAPRO-02 vanili Timbenuh Trichoderma longibrachiatum Rifai aggr.
Koloni jamur tumbuh cepat, setelah tiga hari diinokulasi menutupi permukaan cawan Petri (90,00 mm). Mula-mula berwarna putih, dengan terbentuknya konidia berubah menjadi putih kehijauan dan kemudian menjadi hijau tua.
Miselia bersekat, bercabang banyak, hyaline, dan berdinding halus. Phialospore bentuk elips berukuran 3 – 6 µ x 2 – 3 µ berwarna hijau pucat. Cabang utama konidiofor tumbuh memanjang dengan sedikit cabang samping. Pada setiap cabang samping terbentuk phialide seperti botol dan pada ujungnya menghasilkan phialospore
22
1 3
10 µ
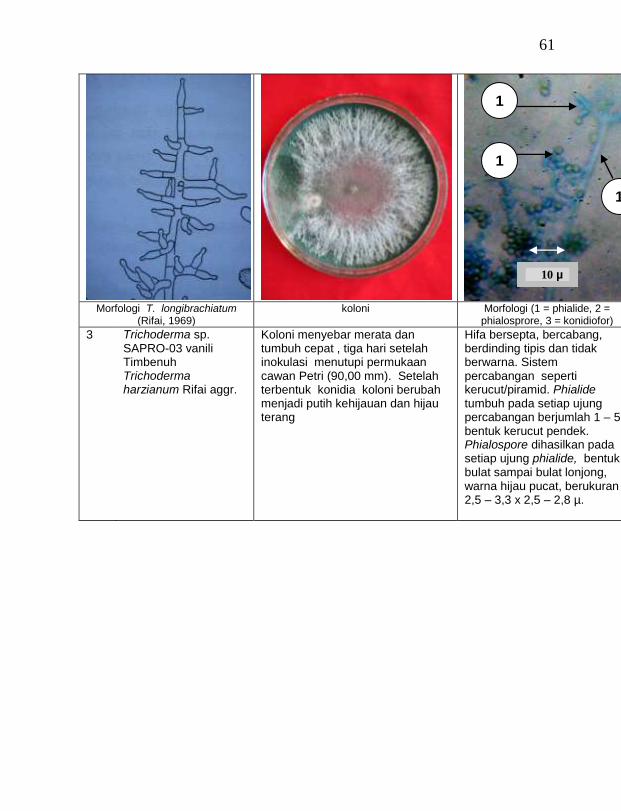
61
Morfologi T. longibrachiatum
(Rifai, 1969) koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
3 Trichoderma sp. SAPRO-03 vanili Timbenuh Trichoderma harzianum Rifai aggr.
Koloni menyebar merata dan tumbuh cepat , tiga hari setelah inokulasi menutupi permukaan cawan Petri (90,00 mm). Setelah terbentuk konidia koloni berubah menjadi putih kehijauan dan hijau terang
Hifa bersepta, bercabang, berdinding tipis dan tidak berwarna. Sistem percabangan seperti kerucut/piramid. Phialide tumbuh pada setiap ujung percabangan berjumlah 1 – 5, bentuk kerucut pendek. Phialospore dihasilkan pada setiap ujung phialide, bentuk bulat sampai bulat lonjong, warna hijau pucat, berukuran 2,5 – 3,3 x 2,5 – 2,8 µ.
1
1
1
10 µ
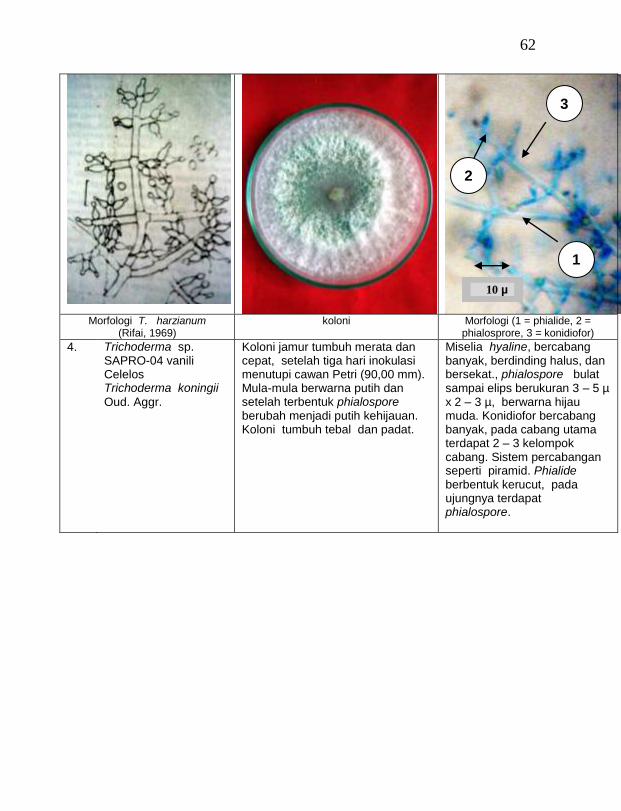
62
Morfologi T. harzianum (Rifai, 1969)
koloni Morfologi (1 = phialide, 2 = phialosprore, 3 = konidiofor)
4. Trichoderma sp. SAPRO-04 vanili Celelos Trichoderma koningii
Oud. Aggr.
Koloni jamur tumbuh merata dan cepat, setelah tiga hari inokulasi menutupi cawan Petri (90,00 mm). Mula-mula berwarna putih dan setelah terbentuk phialospore berubah menjadi putih kehijauan. Koloni tumbuh tebal dan padat.
Miselia hyaline, bercabang banyak, berdinding halus, dan bersekat., phialospore bulat sampai elips berukuran 3 – 5 µ x 2 – 3 µ, berwarna hijau muda. Konidiofor bercabang banyak, pada cabang utama terdapat 2 – 3 kelompok cabang. Sistem percabangan seperti piramid. Phialide berbentuk kerucut, pada ujungnya terdapat phialospore.
3
1
2
10 µ
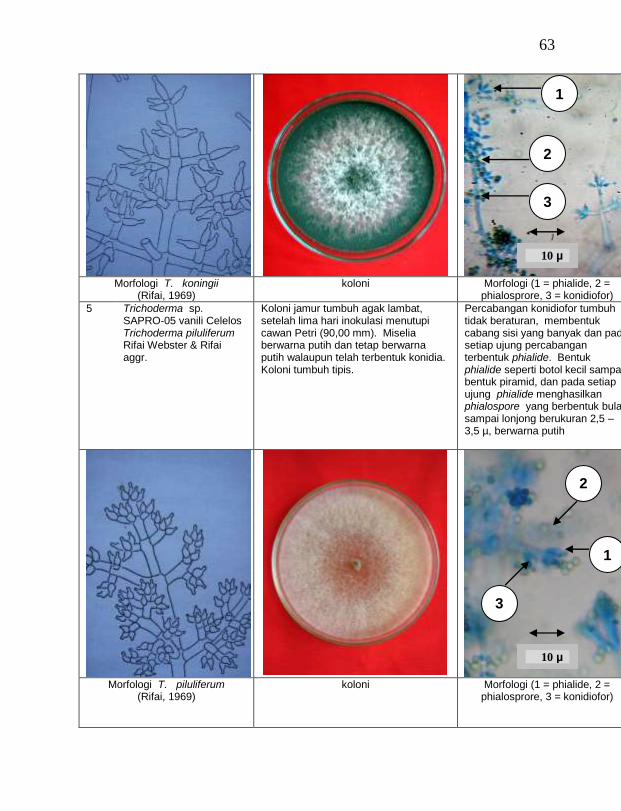
63
Morfologi T. koningii
(Rifai, 1969) koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
5 Trichoderma sp. SAPRO-05 vanili Celelos Trichoderma piluliferum Rifai Webster & Rifai aggr.
Koloni jamur tumbuh agak lambat, setelah lima hari inokulasi menutupi cawan Petri (90,00 mm). Miselia berwarna putih dan tetap berwarna putih walaupun telah terbentuk konidia. Koloni tumbuh tipis.
Percabangan konidiofor tumbuh tidak beraturan, membentuk cabang sisi yang banyak dan pada setiap ujung percabangan terbentuk phialide. Bentuk phialide seperti botol kecil sampai bentuk piramid, dan pada setiap ujung phialide menghasilkan phialospore yang berbentuk bulat sampai lonjong berukuran 2,5 – 3,5 µ, berwarna putih
Morfologi T. piluliferum
(Rifai, 1969)
koloni Morfologi (1 = phialide, 2 = phialosprore, 3 = konidiofor)
3
2
1
10 µ
3
1
2
10 µ
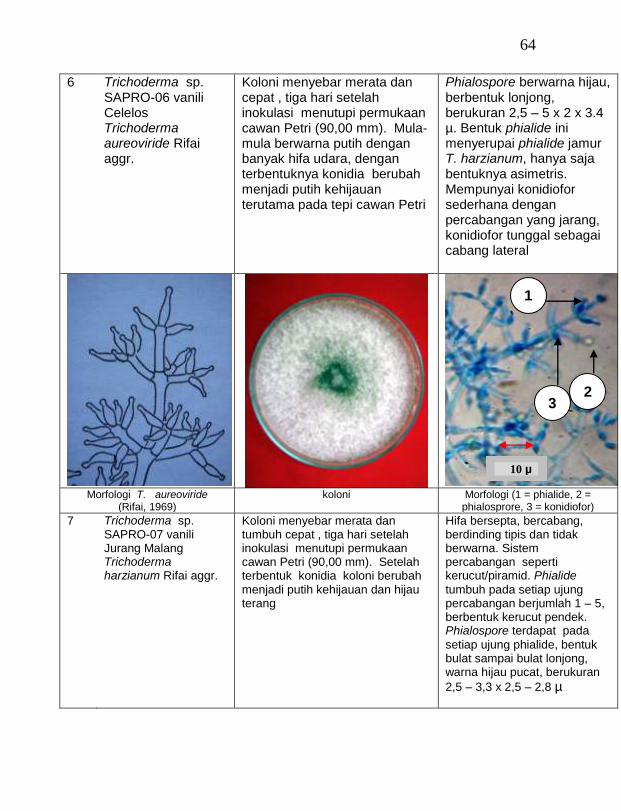
64
6 Trichoderma sp. SAPRO-06 vanili Celelos Trichoderma aureoviride Rifai aggr.
Koloni menyebar merata dan cepat , tiga hari setelah inokulasi menutupi permukaan cawan Petri (90,00 mm). Mula-mula berwarna putih dengan banyak hifa udara, dengan terbentuknya konidia berubah menjadi putih kehijauan terutama pada tepi cawan Petri
Phialospore berwarna hijau, berbentuk lonjong, berukuran 2,5 – 5 x 2 x 3.4 µ. Bentuk phialide ini menyerupai phialide jamur T. harzianum, hanya saja bentuknya asimetris. Mempunyai konidiofor sederhana dengan percabangan yang jarang, konidiofor tunggal sebagai cabang lateral
Morfologi T. aureoviride
(Rifai, 1969) koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
7 Trichoderma sp. SAPRO-07 vanili Jurang Malang Trichoderma harzianum Rifai aggr.
Koloni menyebar merata dan tumbuh cepat , tiga hari setelah inokulasi menutupi permukaan cawan Petri (90,00 mm). Setelah terbentuk konidia koloni berubah menjadi putih kehijauan dan hijau terang
Hifa bersepta, bercabang, berdinding tipis dan tidak berwarna. Sistem percabangan seperti kerucut/piramid. Phialide
tumbuh pada setiap ujung percabangan berjumlah 1 – 5, berbentuk kerucut pendek. Phialospore terdapat pada
setiap ujung phialide, bentuk bulat sampai bulat lonjong, warna hijau pucat, berukuran
2,5 – 3,3 x 2,5 – 2,8 µ
3 2
1
10 µ

65
Morfologi T. harzianum (Rifai, 1969)
koloni Morfologi (1 = phialide, 2 = phialosprore, 3 = konidiofor)
8 Trichoderma sp. SAPRO-08 vanili Jurang Malang
Trichoderma aureoviride Rifai aggr.
Koloni menyebar merata dan cepat , tiga hari setelah inokulasi menutupi permukaan cawan Petri (90,00 mm). Mula-mula berwarna putih dengan banyak hifa udara, dengan terbentuknya konidia berubah menjadi putih kehijauan terutama pada tepi cawan Petri
Phialospore berwarna hijau, berbentuk lonjong, berukuran 2,5 – 5 x 2 x 3.4 µ. Bentuk phialide ini menyerupai phialide jamur T. harzianum, hanya bentuknya asimetris. Mempunyai konidiofor sederhana dengan percabangan yang jarang, konidiofor tunggal sebagai cabang lateral
2
3
1
10 µ
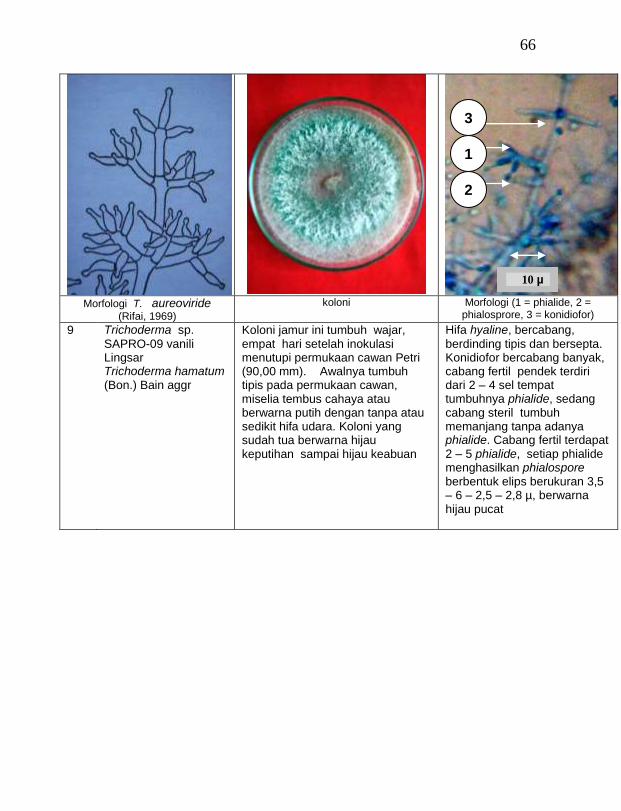
66
Morfologi T. aureoviride
(Rifai, 1969)
koloni Morfologi (1 = phialide, 2 = phialosprore, 3 = konidiofor)
9 Trichoderma sp.
SAPRO-09 vanili Lingsar Trichoderma hamatum (Bon.) Bain aggr
Koloni jamur ini tumbuh wajar, empat hari setelah inokulasi menutupi permukaan cawan Petri (90,00 mm). Awalnya tumbuh tipis pada permukaan cawan, miselia tembus cahaya atau berwarna putih dengan tanpa atau sedikit hifa udara. Koloni yang sudah tua berwarna hijau keputihan sampai hijau keabuan
Hifa hyaline, bercabang,
berdinding tipis dan bersepta. Konidiofor bercabang banyak, cabang fertil pendek terdiri dari 2 – 4 sel tempat tumbuhnya phialide, sedang cabang steril tumbuh memanjang tanpa adanya phialide. Cabang fertil terdapat 2 – 5 phialide, setiap phialide menghasilkan phialospore berbentuk elips berukuran 3,5 – 6 – 2,5 – 2,8 µ, berwarna hijau pucat
2
1
3
10 µ
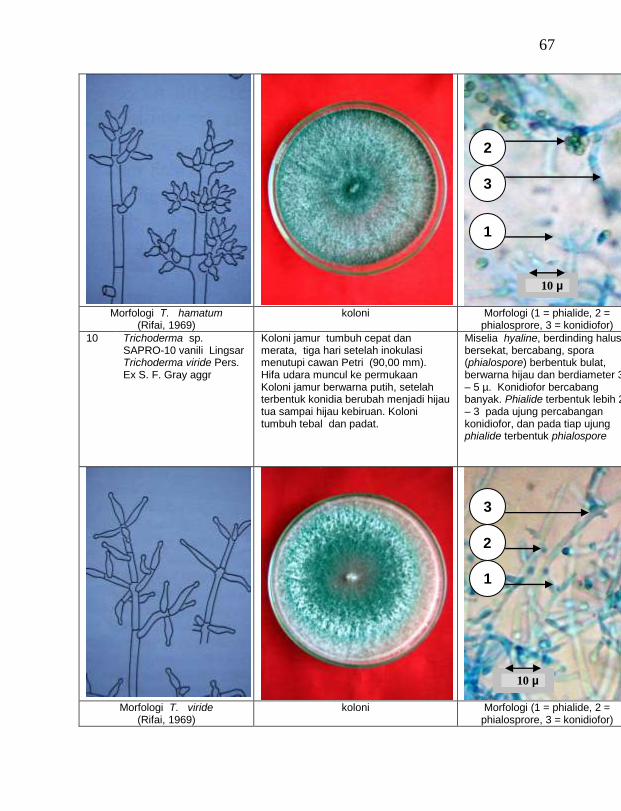
67
Morfologi T. hamatum
(Rifai, 1969) koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
10 Trichoderma sp. SAPRO-10 vanili Lingsar Trichoderma viride Pers. Ex S. F. Gray aggr
Koloni jamur tumbuh cepat dan merata, tiga hari setelah inokulasi menutupi cawan Petri (90,00 mm). Hifa udara muncul ke permukaan Koloni jamur berwarna putih, setelah terbentuk konidia berubah menjadi hijau tua sampai hijau kebiruan. Koloni tumbuh tebal dan padat.
Miselia hyaline, berdinding halus, bersekat, bercabang, spora (phialospore) berbentuk bulat, berwarna hijau dan berdiameter 3 – 5 µ. Konidiofor bercabang banyak. Phialide terbentuk lebih 2 – 3 pada ujung percabangan konidiofor, dan pada tiap ujung phialide terbentuk phialospore
Morfologi T. viride
(Rifai, 1969) koloni Morfologi (1 = phialide, 2 =
phialosprore, 3 = konidiofor)
1
3
2
10 µ
2
1
3
10 µ
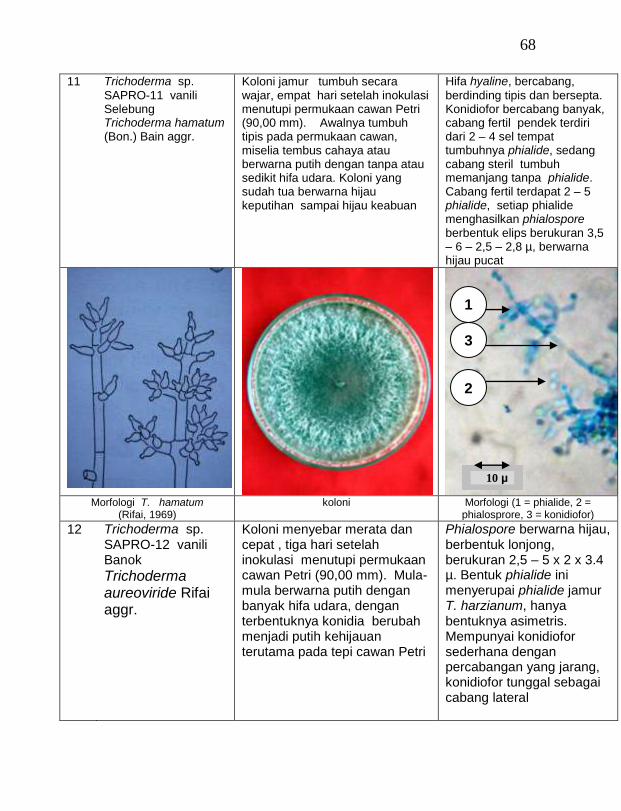
68
11 Trichoderma sp.
SAPRO-11 vanili Selebung Trichoderma hamatum (Bon.) Bain aggr.
Koloni jamur tumbuh secara wajar, empat hari setelah inokulasi menutupi permukaan cawan Petri (90,00 mm). Awalnya tumbuh tipis pada permukaan cawan, miselia tembus cahaya atau berwarna putih dengan tanpa atau sedikit hifa udara. Koloni yang sudah tua berwarna hijau keputihan sampai hijau keabuan
Hifa hyaline, bercabang,
berdinding tipis dan bersepta. Konidiofor bercabang banyak, cabang fertil pendek terdiri dari 2 – 4 sel tempat tumbuhnya phialide, sedang cabang steril tumbuh memanjang tanpa phialide. Cabang fertil terdapat 2 – 5 phialide, setiap phialide menghasilkan phialospore berbentuk elips berukuran 3,5 – 6 – 2,5 – 2,8 µ, berwarna hijau pucat
Morfologi T. hamatum (Rifai, 1969)
koloni Morfologi (1 = phialide, 2 = phialosprore, 3 = konidiofor)
12 Trichoderma sp. SAPRO-12 vanili Banok
Trichoderma aureoviride Rifai aggr.
Koloni menyebar merata dan cepat , tiga hari setelah inokulasi menutupi permukaan cawan Petri (90,00 mm). Mula-mula berwarna putih dengan banyak hifa udara, dengan terbentuknya konidia berubah menjadi putih kehijauan terutama pada tepi cawan Petri
Phialospore berwarna hijau, berbentuk lonjong, berukuran 2,5 – 5 x 2 x 3.4 µ. Bentuk phialide ini menyerupai phialide jamur T. harzianum, hanya bentuknya asimetris. Mempunyai konidiofor sederhana dengan percabangan yang jarang, konidiofor tunggal sebagai cabang lateral
2
3
1
10 µ
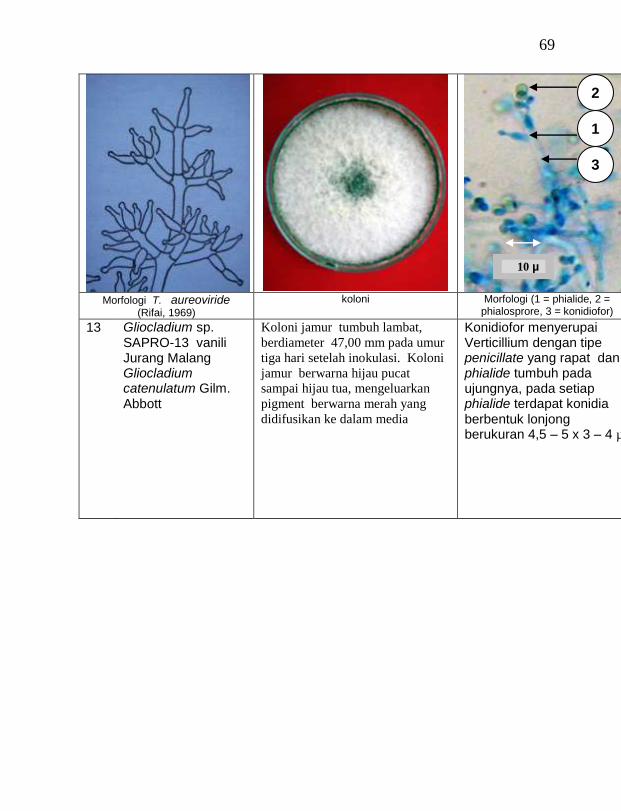
69
Morfologi T. aureoviride
(Rifai, 1969)
koloni Morfologi (1 = phialide, 2 = phialosprore, 3 = konidiofor)
13 Gliocladium sp. SAPRO-13 vanili Jurang Malang Gliocladium catenulatum Gilm. Abbott
Koloni jamur tumbuh lambat,
berdiameter 47,00 mm pada umur
tiga hari setelah inokulasi. Koloni
jamur berwarna hijau pucat
sampai hijau tua, mengeluarkan
pigment berwarna merah yang
didifusikan ke dalam media
Konidiofor menyerupai Verticillium dengan tipe penicillate yang rapat dan phialide tumbuh pada ujungnya, pada setiap phialide terdapat konidia berbentuk lonjong berukuran 4,5 – 5 x 3 – 4 µ
3
1
2
10 µ
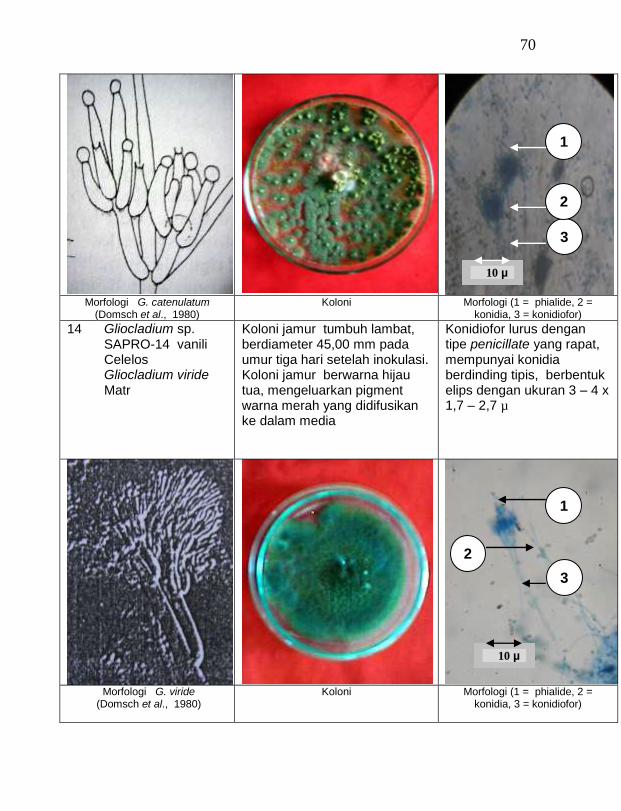
70
Morfologi G. catenulatum
(Domsch et al., 1980) Koloni Morfologi (1 = phialide, 2 =
konidia, 3 = konidiofor)
14 Gliocladium sp. SAPRO-14 vanili Celelos Gliocladium viride Matr
Koloni jamur tumbuh lambat, berdiameter 45,00 mm pada umur tiga hari setelah inokulasi. Koloni jamur berwarna hijau tua, mengeluarkan pigment warna merah yang didifusikan ke dalam media
Konidiofor lurus dengan tipe penicillate yang rapat, mempunyai konidia berdinding tipis, berbentuk elips dengan ukuran 3 – 4 x 1,7 – 2,7 µ
Morfologi G. viride
(Domsch et al., 1980)
Koloni Morfologi (1 = phialide, 2 = konidia, 3 = konidiofor)
3
2
1
10 µ
3
2
1
10 µ
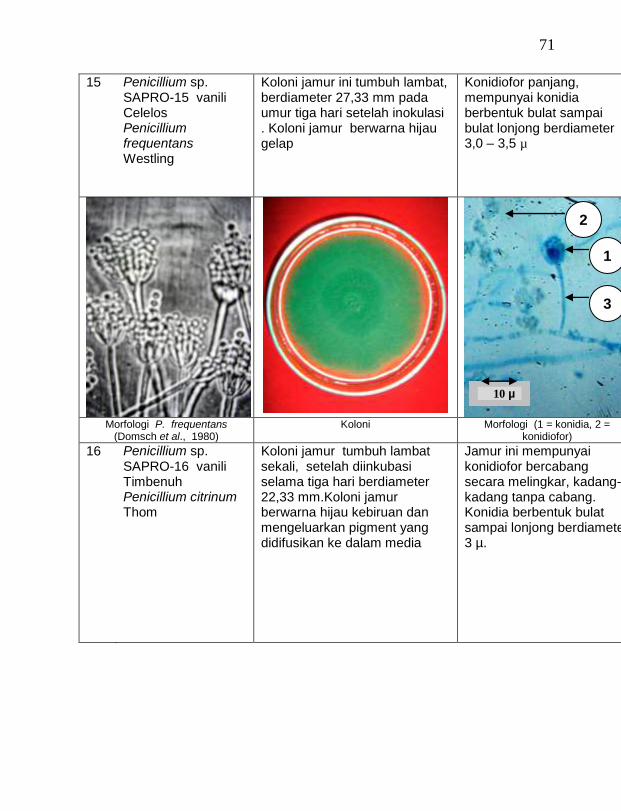
71
15 Penicillium sp. SAPRO-15 vanili Celelos Penicillium frequentans Westling
Koloni jamur ini tumbuh lambat, berdiameter 27,33 mm pada umur tiga hari setelah inokulasi . Koloni jamur berwarna hijau gelap
Konidiofor panjang, mempunyai konidia berbentuk bulat sampai bulat lonjong berdiameter 3,0 – 3,5 µ
Morfologi P. frequentans
(Domsch et al., 1980) Koloni Morfologi (1 = konidia, 2 =
konidiofor)
16 Penicillium sp. SAPRO-16 vanili Timbenuh Penicillium citrinum Thom
Koloni jamur tumbuh lambat sekali, setelah diinkubasi selama tiga hari berdiameter 22,33 mm.Koloni jamur berwarna hijau kebiruan dan mengeluarkan pigment yang didifusikan ke dalam media
Jamur ini mempunyai konidiofor bercabang secara melingkar, kadang-kadang tanpa cabang. Konidia berbentuk bulat sampai lonjong berdiameter 3 µ.
2
3
1
10 µ

72
Morfologi P. citrinum (Domsch et al., 1980)
Koloni Morfologi (1 = konidia, 2 = konidiofor)
17 Aspergillus sp. SAPRO-17 vanili Celelos Aspergillus flavus Link ex Gray
Koloni jamur tumbuh lambat sekali, berdiameter 28,67 mm setelah tiga hari inokulasi. Jamur mempunyai koloni berwarna hijau
Jamur ini mempunyai konidiofor hyalin, pada ujungnya membesar membentuk bulatan dan terdapat kumpulan phialide seperti tabung (sterigmata). Pada ujung phialide terdapat konidia berbentuk bulat sampai lonjong berwarna hijau berdiameter 3 – 5 µ, konidia berangkai-rangkai berjumlah banyak dan secara keseluruhan tampak membulat
3
1
2
10 µ
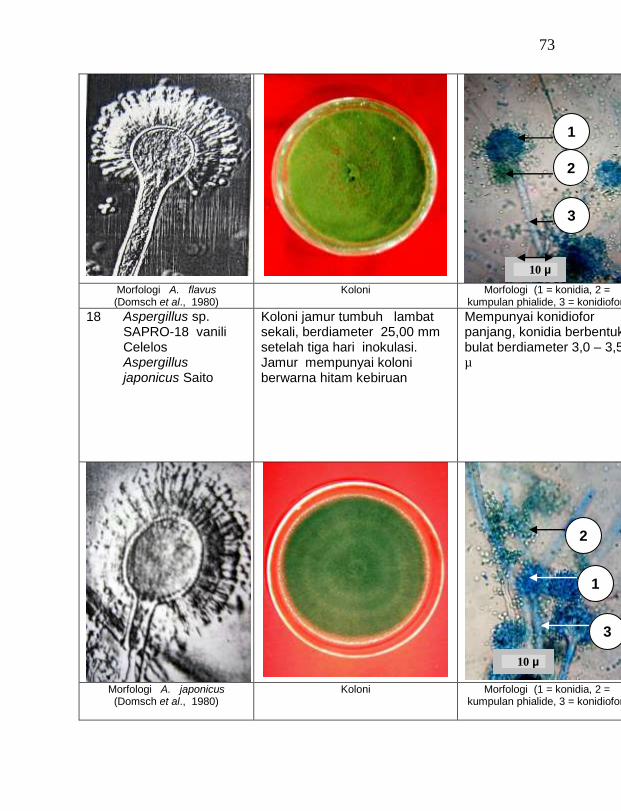
73
Morfologi A. flavus (Domsch et al., 1980)
Koloni Morfologi (1 = konidia, 2 = kumpulan phialide, 3 = konidiofor)
18 Aspergillus sp. SAPRO-18 vanili Celelos Aspergillus japonicus Saito
Koloni jamur tumbuh lambat sekali, berdiameter 25,00 mm setelah tiga hari inokulasi. Jamur mempunyai koloni berwarna hitam kebiruan
Mempunyai konidiofor panjang, konidia berbentuk bulat berdiameter 3,0 – 3,5 µ
Morfologi A. japonicus (Domsch et al., 1980)
Koloni Morfologi (1 = konidia, 2 = kumpulan phialide, 3 = konidiofor)
2
3
1
10 µ
2
1
3
10 µ
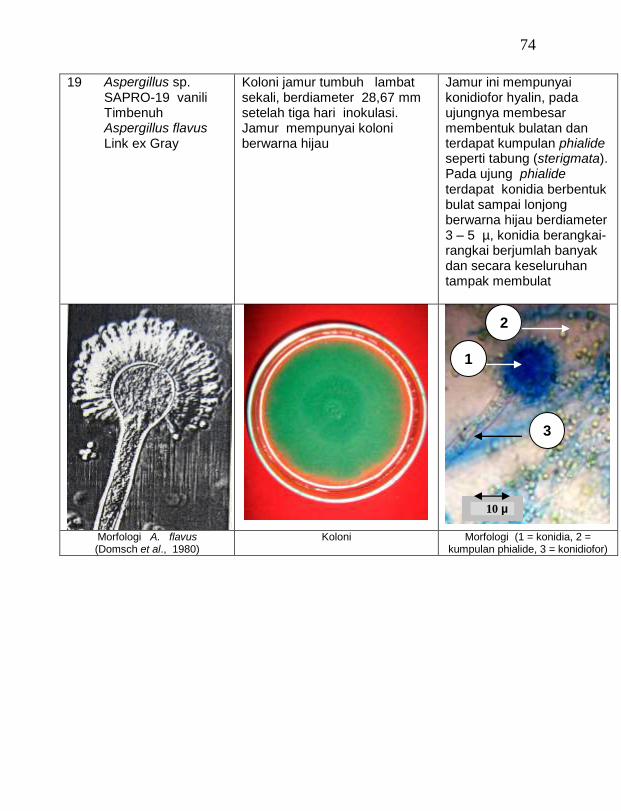
74
19 Aspergillus sp. SAPRO-19 vanili Timbenuh Aspergillus flavus Link ex Gray
Koloni jamur tumbuh lambat sekali, berdiameter 28,67 mm setelah tiga hari inokulasi. Jamur mempunyai koloni berwarna hijau
Jamur ini mempunyai konidiofor hyalin, pada ujungnya membesar membentuk bulatan dan terdapat kumpulan phialide seperti tabung (sterigmata). Pada ujung phialide terdapat konidia berbentuk bulat sampai lonjong berwarna hijau berdiameter 3 – 5 µ, konidia berangkai-rangkai berjumlah banyak dan secara keseluruhan tampak membulat
Morfologi A. flavus (Domsch et al., 1980)
Koloni Morfologi (1 = konidia, 2 = kumpulan phialide, 3 = konidiofor)
3
1
2
10 µ
Related Documents











