Universidade Federal do Rio de Janeiro Programa de Pós-graduação em Produtos Bioativos e Biociências Juliana Silva Nascimento De Novais Caracterização da matéria prima vegetal e atividade biológica das folhas de Senna alata (L.) Roxb. Em acessos na caatinga (BA) e restinga (RJ), Brasil MACAÉ-RJ, 2017

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Universidade Federal do Rio de Janeiro Programa de Pós-graduação em Produtos Bioativos e Biociências
Juliana Silva Nascimento De Novais
Caracterização da matéria prima vegetal e atividade
biológica das folhas de Senna alata (L.) Roxb. Em acessos
na caatinga (BA) e restinga (RJ), Brasil
MACAÉ-RJ, 2017

Universidade Federal do Rio de Janeiro Programa de Pós-graduação em Produtos Bioativos e Biociências
Juliana Silva Nascimento De Novais
Caracterização da matéria prima vegetal e atividade
biológica das folhas de Senna alata (L.) Roxb. Em acessos
na caatinga (BA) e restinga (RJ), Brasil
Dissertação a ser apresentada pela aluna Juliana Silva
Nascimento de Novais ao Programa de Pós Graduação
em Produtos Bioativos e Biociencias, da Universidade
Federal do Rio de Janeiro, Campus UFRJ Macaé, como
requisito parcial para a obtenção do título de Mestre em
Ciências.
Orientador: Prof Dr Gilberto Dolejal Zanetti
Co-orientação: Prof. Drª Elaine dos Anjos da Cruz da
Rocha
Macaé, RJ 2017


CARACTERIZAÇÃO DA MATÉRIA PRIMA VEGETAL E ATIVIDADE
BIOLÓGICA DAS FOLHAS DE SENNA ALATA (L.) ROXB. EM ACESSOS
NA CAATINGA (BA) E RESTINGA (RJ), BRASIL
Juliana Silva Nascimento de Novais
Dissertação apresentada ao Programa de Pós Graduação em Produtos Bioativos e Biociencias da Universidade Federal do Rio de Janeiro, como parte das exigências para a obtenção do título de Mestre em Ciências.
Aprovada em 30 de Outubro de 2017.
_______________________________________________ Prof Dr Arídio Mattos Junior
_______________________________________________
Prof Dr Edison Luis Santana de Carvalho
_______________________________________________ Prof Dra Michelle Frazão Muzitano
________________________________________________
Orientador: Prof Dr Gilberto Dolejal Zanetti
_______________________________________________ Coorientadora: Profa Dra Elaine dos Anjos da Cruz da Rocha
Macaé, RJ 2017

AGRADECIMENTOS
Agradeço a Deus por ter me presenteado com o dom da vida. Gostaria de agradecer a diversas pessoas pela ajuda, apoio, contribuição,
atenção e paciência, sem os quais tornaria a realização desse trabalho imensamente árduo. Estas são:
Meu esposo, por apoiar, participar e está sempre disposto a me ajudar em uma participação direta na concretização dos meus sonhos. Pela paciência durante incontáveis horas em que tive que está no laboratório ou escrevendo durante madrugadas. Pelo amor que ameniza a dor e torna tudo possível.
À minha mãe e minha irmã pelo amor incondicional, pelas palavras doces e duras de amor e de encorajamento, por sempre se fazerem tão presentes mesmo a distância sendo um abismo.
Sou grata ao meu orientador Gilberto Zanetti por nunca escolher o caminho mais fácil ou conveniente, mas por sempre escolher o caminho certo. Por ter acreditado em mim quando eu própria não podia. Pelo carinho e amizade dispensados a mim. Por cumprir de maneira graciosa e exemplar o sentido da palavra “mentor” na sua definição mais completa. Pela sua brilhante capacidade científica. Pela sua incansável dedicação no desenvolvimento da carreira de cada um dos seus alunos. Pelas suas aulas que iam além do assunto proposto, de forma didática e interessante. Pelo exemplo de inspiração.
Agradeço também a minha coorientadora Elaine da Rocha, por generosamente abrir as portas do seu laboratório e compartilhar seu conhecimento e recurso no desenvolvimento desse trabalho. Por ter me abraçado e por não ter medido esforços em me ajudar e apoiar. Por não olhar simplesmente naquilo que se investiga, mas se preocupar de forma generosa com quem está investigando. Pelo exemplo de paciência, sabedoria e liderança.
Aos técnicos e amigos do Instituto de ciência e tecnologia de Macaé, em especial, Jobert e Jéssyca, que foram sempre presentes durante esses anos e me ajudaram no desenvolvimento desse trabalho.
Aos docentes, Denise, Edison e Michelle, por me ensinarem que além de cérebros brilhantes haviam corações generosos e dispostos a ajudar. Por toda auxílio, conselhos e apoio no desenvolvimento desse estudo. Ao professor Arídio que aceitou contribuir com esse trabalho.
Agradeço universidade de Taubaté em especial a colaboração da professora Josseara Beralto, que mesmo sem me conhecer me apoiou no que foi preciso para concluir os experimentos.
A equipe da secretaria de coordenação do PRODGRADBIO, principalmente, a Andreia e o Leonardo pela atenção, paciência e carinho.
Enfim a todas as pessoas que contribuíram direta e indiretamente na realização desse trabalho e que porventura tenham sido omitidas, meus sinceros agradecimentos.

RESUMO
NOVAIS, Juliana Silva Nascimento. Caracterização da Matéria prima vegetal e atividade biológica das folhas de Senna alata (L.) Roxb. em acessos na caatinga (BA) e restinga (RJ), Brasil. Macaé, RJ, 2017. Dissertação (Mestrado em ciências) – Universidade Federal do Rio de Janeiro, Macaé, 2017.
Senna alata (L.) Roxb. é uma leguminosa arbustiva da família Fabaceae introduzida na caatinga e na restinga do Brasil. Essa espécie possui utilização tradicional para fins medicinais o que motivou este estudo que visa caracterizar as folhas e identificar, comparativamente, as atividades biológicas de Senna alata (L.) Roxb. em acessos na caatinga (BA) e restinga (RJ). Após a coleta, o material foi desidratado e levado a pó. Obtiveram-se extratos etanólicos a 40 % por maceração e, este extrato bruto seco, foi resuspenso em diferentes solventes de acordo com o ensaio a ser realizado. Optou-se pelo extrato etanólico a 40 % uma vez que os acessos apresentaram maior teor extrativo nesta concentração de solventes. As folhas nos dois acessos apresentaram morfo-anatomia semelhante pelas técnicas farmacopeias, destacando-se como marcador os tricomas tectores e a presença de estômatos nas duas faces da epiderme. Também se obteve resultados de pureza satisfatórios aos padrões farmacopeicos. Detectou-se flavonoides, saponinas hemolíticas, antraquinonas e mucilagem para os dois acessos e taninos apenas no acesso da restinga. O teor total de polifenois e de flavonoides foi semelhante nos dois acessos. O perfil cromatográfico foi obtido em cromatoplacas de sílica gel com sistema eluente constituído por hexano: acetato de etila (1:1) e vanilina sulfúrica mantida a 100º C por 5 minutos como agente cromogênico, revelando 2 bandas semelhantes para os acessos. A determinação da atividade antibacteriana por bioautografia demonstrou que os dois acessos inibiram o crescimento de Staphylococcus aureus e Escherichia coli. A atividade alelopática frente a diásporos de Lactuca sativa demonstrou potencial alelopático em extratos nas concentrações a partir de 0,5 %, sujerindo atividade inseticida e herbicida.
O extrato dos acessos não demonstrou citotoxidade nas concentrações 50 e 100 μg/mL
pelo método de {brometo de [3-(4,5-dimetiltiazol-2-il)-2,5-difenil tetrazólio]}, nas concentrações testadas. A atividade anti-inflamatória avaliada pela dosagem de óxido nítrico e fator de necrose tumoral alfa por células intraperitoneais de camundongo Swiss, induzido por lipopolissacarídeo e o polissacarídeo Zymosan. Observou-se um potencial efeito anti-inflamatório sobre macrófagos peritoneais quando estimulados com lipopolissacarídeo e tratados com os 2 acessos na concentração 100 µg/mL, e em macrófagos estimulados por Zymosan, após tratamento a partir de 50 µg/mL do acesso caatinga e 100 µg/mL do acesso restinga. Os extratos apresentam baixa toxidez aguda via intraperitoneal em camundongos Swiss, com concentração letal de 40 e 60 g/kg nos acessos da caatinga e restinga, respectivamente e os níveis de lactato desidrogenase mantiveram-se normais. Os resultados obtidos foram semelhantes para os acessos de Senna alata na caatinga e restinga, sendo que os extratos estudados revelaram baixa toxidez e resultados promissores nas atividades biológicas testadas, conferindo para as folhas de à Senna alata potencialidade real no desenvolvimento de um fitoterápico. Palavras chaves: Fabaceae; Senna alata; Farmacobotânica; Plantas medicinais; Atividades biológicas; Anti-inflamatório; Toxidez; Citotoxidade.

ABSTRACT
NOVAIS, Juliana Silva Nascimento. Characterization of vegetable raw materials and biological activity of the leaves of Senna alata (L.) Roxb. in access in the caatinga (BA) and restinga (RJ), Brazil. Macaé, RJ, 2017. Dissertation (master of science)-Federal University of Rio de Janeiro, Macaé, 2017.
Senna alata (L.) Roxb. is a shrubby legume of the Fabaceae family introduced in the caatinga and the restinga areas of Brazil. This species is traditionally used for medicinal purposes. Therefore this study aims to compare the characterization and identification of leaves, the biological activities of Senna alata (L.) Roxb. in access from the caatinga (Bahia) and restinga (Rio de Janeiro). After collection, the material was dehydrated and taken to dust. Ethanolic extracts were obtained at concentration of 40% by maceration. This crude extract, was ressuspended in various solvents according to the test being performed. We opted for the 40% ethanolic extract once the hits presentedthe most extractive content in this concentration of solvents. The leaves showed morpho-anatomy by using Pharmacopoeias techniques, including tectores and estomatic trichomes types. It was also observed satisfactory results of purity according to Pharmacopeia standards. It were detected flavonoids, haemolytic saponins, anthraquinones and mucilage for both access and tannins only in resting access. The total content of polyphenols and flavonoids was similar both accesses. The chromatographic profile was retrieved from cromatoplates of silica gel with eluent system of hexane: ethyl acetate (1:1) and sulfuric vanillin at 100 ºC for 5 minutes as chromogenic agent, revealing two similar bands to both accesses. The determination of antibacterial activity obtained by bioautography showed that the two approaches accesses inhibited the growth of Staphylococcus aureus and Escherichia coli. Alelopatic activity, were evaluated in Lactuca sativa diaspores and showed allelopathic potential in extracts at concentrations of 0.5 %, which suggested insecticide and herbicide activity. The cytotoxic potential was evaluated by {bromide [3-(4.5-dimetiltiazol-2yl)-2.5-diphenyl tetrazolium]} and both accesses were not cytotoxic at tested concentrations. The in vitro anti-inflammatory activity was evaluated by determination of nitric oxide and tumor necrosis factor alpha produced by intraperitoneais mouse cells induced by lipopolysaccharide and Zymosan polysaccharide. The results indicated that both accesses of Senna alata have an effective anti-inflammatory activity on peritoneal macrophages at 100 µg/mL after lipopolysaccharide stimulation. Using the Zymosan stimuli, it was observed an anti-inflammatory activity with 50 µg/mL of in the caatinga access and at 100 µg/mL of restinga access. The extracts feature low acute toxicity after intraperitoneal administration in Swiss mice, with lethal dose at 40 mg/kg and 60 mg/kg for access of the caatinga and restinga, respectively. The lactate dehydrogenase levels remained normal. The results were similar to those of Senna alata in caatinga and restinga, being extracts studied revealed low toxicity and promising results on biological activities tested, giving to the leaves of the Senna alata real capability in developing a phytotherapeutic. Key words: Fabaceae; Senna alata; Farmacobotânica; Medicinal plants; Biological activities; Anti-inflammatory; Toxicity; Cytotoxicity.

LISTA DE FIGURAS
Figura 1 Aspecto geral de Senna alata (L.) Roxb..................................................................... 23
Figura 2 Estruturas químicas encontradas em Senna alata (L.) Roxb...................................... 25
Figura 3 Estrutura química básica da antraquinona.................................................................. 25
Figura 4 Transformação de {brometo de [3-(4,5-dimetiltiazol-2-il)-2,5-difenil tetrazólio]} (MTT) em { E, Z- 1- (4,5-dimetiltiazol-2-il) -1,3-difenilformasan } (Formasan)............ 31
Figura 5 Aspecto geral das folhas de Senna alata (L.) Roxb.................................................... 53
Figura 6 Venação da folha de Senna alata (L.) Roxb. acesso caatinga................................... 54
Figura 7 Venação da folha de Senna alata (L.) Roxb. acesso restinga.................................... 55
Figura 8 Aspecto geral das folhas desidratadas de Senna alata (L.) Roxb.............................. 55
Figura 9 Secção paradérmica da lamina foliar de Senna alata (L.) Roxb. acesso caatinga..... 56
Figura 10 Secção paradérmica da lamina foliar de Senna alata (L.) Roxb. acesso restinga...... 57
Figura 11 Glanulometria por tamisação das folhas em pó de Senna alata (L.) Roxb................. 61
Figura 12 Reta obtida a partir da curva padrão de ácido gálico no ensaio de quantificação de fenolicos totais de Senna alata (L.) Roxb. nos acesso caatinga e restinga................ 68
Figura 13 Gráfico obtido a partir da curva de calibração realizada com rutina na obtenção do teor total de flavonoides nos extratos das folhas de Senna alata (L.) Roxb. nos acessos caatinga e restinga........................................................................................
69
Figura 14 Representação do perfil cromatográfico obtido para os extratos etanólicos a 40 % a partir das folhas de Senna alata (L.) Roxb., sobre cromatofolhas de sílica gel G 60 F254.......................................................................................................................
71
Figura 15 Germinabilidade (%) de Lactuca sativa L. frente extratos de Senna alata (L.) Roxb. obtidas na caatinga (A) e na restinga (B) nas concentrações de 0,5, 1 e 2 %.................................................................................................................................
74
Figura 16 Teste de viabilidade do extrato de Senna alata (L.) Roxb. das soluções extrativas dos acessos catinga e restinga nas concentrações de 50 µg/mL e 100 µg/mL em macrófagos pelo método de 3-(4,5dimetiltiazol-2-il)-2,5-difenil tetrazolio................... 76
Figura 17 Inibição da citocina TNF-α após tratamento com extrato hidroetanólico de Senna alata (L.) Roxb, caatinga e restinga, das células RAW-264-7, estimuladas com LPS 78
Figura 18 Produção de óxido nítrico os grupos tratados com Senna alata(L.) Roxb.................. 79
Figura 19 Inibição da citocina TNF-α após tratamento com extrato hidroetanólico de Senna alata (L.) Roxb. caatinga e restinga, das células RAW-264-7, estimuladas com Zymosan...................................................................................................................... 81
Figura 20 Produção de NO nos grupos tratados com Senna alata (L.) Roxb. nos acessos de restinga e caatinga...................................................................................................... 82
Figura 21 Potencial tóxico de Senna alata (L.) Roxb. acesso caatinga e restinga..................... 83
Figura 22 Liberação da LDH em soro sanguíneo de camundongos Swiss tratados com extratos de Senna alata (L.) Roxb............................................................................... 84

LISTA DE TABELAS
Tabela 1 Microorganismos utilizados nos ensaios de bioautografia........................................ 39
Tabela 2 Características macroscópicas das folhas de Senna alata (L.) Roxb....................... 54
Tabela 3 Características microscópicas em das folhas de Senna alata (L.) Roxb. nos acessos restinga e caatinga......................................................................................
56 Tabela 4 Descrição macroscópica e organoléptica das folhas em pó de Senna alata (L.)
Roxb............................................................................................................................ 60
Tabela 5 Determinação físico-química referente à pureza das folhas de Senna alata (L.) Roxb............................................................................................................................ 63
Tabela 6 Determinação do teor extrativo dos extratos hidroetanólicos obtidos a partir das folhas de Senna alata (L.) Roxb..................................................................................
64 Tabela 7 Triagem fitoquimica dos extratos etanólicos a 40% obtidos a partir das folhas de
Senna alata (L.) Roxb. dos acessos restinga e caatinga............................................ 65
Tabela 8 Conteúdo de fenólicos totais de Senna alata caatinga e restinga em mg de ácido gálico/g por amostra....................................................................................................
68 Tabela 9 Conteúdo de flavonoides totais de Senna alata caatinga e restinga em mg de
rutina/g por amostra.................................................................................................... 70
Tabela 10 Tabela 10 Halos de inibição obtidos em bioautografia dos extratos de Senna alata (L.) Roxb., acesso caatinga e restinga, frente a Staphilococcus aureus e Escherichia coli........................................................................................................... 73

LISTA DE ABREVIATURAS E SIGLAS
A3P Programa da agenda ambiental na administração pública
AAs Ácido acetilsalicílico
AcOEt Acetato de Etíla
AINEs Anti-inflamatórios não esteroidais
AIEs Anti-inflamatórios esteroidais
ANOVA Análise de variância
ARMDS Antirreumáticos modificadores da doença
AVC Acidente vascular cerebral
Ca Caatinga
CCD Cromatografia de camada delgada
CEUA Comissão de ética no uso de animais
CONAMA Conselho nacional do meio ambiente
CP Controle positivo
DC Células dendríticas
DL50 Dose letal de 50 %
DMSO Dimetilsufoxido
DMSO-c Dimetilsulfoxido completo
EtOH Etanol
FMS Fenazina metosulfato
IE Índice estomático
Ig Imunoglobulina
IkB Inibidor de Kappa B
IL-2 Imugnoglobina 2
I.p Intra peritoneal
LDH Lactato desidrogenase
LES Lúpus eritematoso sistêmico
LPS Lipopolissacarideo
MTS Tetrazólio
MTT {Brometo de [3-(4,5-dimetiltiazol-2il)-2,5-difenil tetrazólio]}
NAD Difosfopiridino nucleotídeo
NF-KB Fator nuclear kappa B
NO Óxido nítrico
OMS Organização mundial da saúde
PA Padrão analítico
PIB Produto interno bruto
PBS Tampão fosfato salino
PNPMF Programa nacional de plantas medicinal e fitoterápico
PNPIC Política nacional de práticas integrativas e complementares

Re Restinga
Rf Fator de retenção
PRRs Receptores de reconhecimento padrões
SUS Sistema único de saúde
TNFα Fator de necrose tumoral-alfa
UFC Unidade formadora de colônia
UV Ultra violeta
XTT 2,3-bis (2-metoxi-4nitro-5-sulfofenil)-5-[(fenilamino)carbonil]-2H-hidróxido de
tetrazólio
Zy Zymosan

SUMÁRIO
1. INTRODUÇÃO ............................................................................................ 14
1.1 Caatinga x Restinga ............................................................................... 18
1.2 Família Fabaceae .................................................................................. 19
1.3 Senna alata (L.) Roxb. ........................................................................... 22
1.4 Emprego medicinal de Senna alata (L.) Roxb. ....................................... 23
1.5 Substâncias químicas no gênero Senna e Senna alata (L) Roxb. .......... 23
1.6 Atividades biológicas .............................................................................. 26
1.6.1 Atividade antimicrobiana em bioautografia ...................................... 27
1.6.2 Alelopatia: interferências sobre comunidades vegetais.................... 28
1.6.3 Atividade citotóxica pelo método MTT ............................................. 30
1.6.4 O processo inflamatório e o estimulo por lipopolissacarídeo (LPS) e
zymosan (Zy) ....................................................................................................... 31
1.6.5 Toxidez aguda ................................................................................. 35
1.7 A importância de estudos sobre Senna alata como medicinal ................ 36
2. OBJETIVOS ................................................................................................ 38
2.1 Objetivo Geral ........................................................................................ 38
2.2 Objetivos Específicos ............................................................................. 38
3. MATERIAL E MÉTODOS ............................................................................ 39
3.1 Material vegetal ...................................................................................... 39
3.2 Solventes, reagentes e substâncias referência utilizadas ...................... 39
3.3 Micro-organismos .................................................................................. 39
3.4 Cultura de Macrofagos ........................................................................... 39
3.5 Animais .................................................................................................. 40
3.6 Análise morfo-anatômica das folhas de Senna alata .............................. 40
3.6.1 Análise Morfológica ......................................................................... 40
3.6.2 Análise Anatômica ........................................................................... 41
3.7 Determinação do teor de extrativos ........................................................ 42
3.8 Análise físico-química das folhas pulverizadas de Senna alata .............. 43

3.9 Análise fitoquímica dos extratos das folhas de Senna alata ................... 44
3.9.1 Determinação do pH dos extratos ................................................... 45
3.9.2 Determinação de antraquinona ........................................................ 45
3.9.3 Determinação de saponinas ............................................................ 45
3.9.4 Determinação de saponinas hemolíticas ......................................... 45
3.9.5 Determinação de flavonoides .......................................................... 46
3.9.6 Determinação de mucilagem ........................................................... 46
3.9.7 Determinação de taninos ................................................................. 47
3.9.8 Determinação do teor de polifenois totais ........................................ 47
3.9.9 Determinação do teor de flavonoides totais ..................................... 47
3.9.10 Perfil cromatográfico dos extratos em cromatografia de camada
delgada (CCD) ..................................................................................................... 48
3.10Determinação de atividades biológicas dos extratos das folhas de
Senna alata......... ............................................................................................... 48
3.10.1 Determinação da atividade antibacteriana ..................................... 48
3.10.2 Determinação da atividade alelopática .......................................... 49
3.10.3 Viabilidade celular pelo MTT .......................................................... 49
3.10.4 Avaliação da atividade anti-inflamatória in vitro ............................. 50
3.10.5 Determinação da toxidez intraperitoneal aguda ............................. 51
3.11 Análise estatística dos resultados ........................................................ 52
4. RESULTADOS E DISCUSSÃO ................................................................... 53
4.1 Morfo-anatomia e caraterização organoléptica das folhas de Senna alata .
............................................................................................................... 53
4.1.1 Análise macroscópica ...................................................................... 53
4.1.2 Análise microscópica ....................................................................... 56
4.1.3 Análise macro-microscópica e organoléptica das folhas em pó ....... 59
4.2 Análise físico-química ............................................................................ 62
4.3 Determinação do teor extrativo .............................................................. 64
4.4 Triagem fitoquímica dos extratos ........................................................... 65
4.4.1 Determinação de polifenois totais .................................................... 67

4.4.2 Determinação de flavonoides totais ................................................. 69
4.4.3 Perfil cromatográfico em cromatografia de camada delgada ........... 71
4.5 Atividades biológicas .............................................................................. 73
4.5.1 Atividade antibacteriana .................................................................. 73
4.5.2 Atividade alelopática ........................................................................ 74
4.5.3 Viabilidade celular pelo método MTT ............................................... 76
4.5.4 Atividade anti-inflamatória in vitro com LPS ..................................... 77
4.5.5 Atividade anti-inflamatória in vitro com Zymosan ............................. 80
4.5.6 Toxidez aguda ................................................................................. 83
5. CONCLUSÕES ........................................................................................... 86
REFERENCIAS ............................................................................................... 88

14
1. INTRODUÇÃO
A valorização da medicina tradicional nos mais diferentes países
começou a ser mais estimulada quando a Organização Mundial de Saúde
(OMS), no ano de 1978, reconheceu e recomendou as plantas como
medicinais, como medida para minimizar a falta de medicamentos,
principalmente no atendimento primário à saúde (Akerelle, 1988; Gottlieb,
1993). A utilização de plantas com objetivos medicinais é bastante difundida
em todo o mundo. Na Alemanha, por exemplo, já no final do século passado,
70 % dos médicos em clínica geral prescreviam fitoterápicos (Blumentahl,
1998). No Brasil, conforme Veiga Junior (2008) as plantas medicinais são muito
utilizadas em diversas formas de tratamentos de saúde, apesar da
disponibilidade de medicamentos alopáticos. Sempre é ressaltado que há a
necessidade de aliar esse conhecimento popular a estudos científicos para
minimizar situações preocupantes que a falta de informações pode gerar, tanto
para a população quanto para a ciência, a exemplo dos possíveis efeitos da
automedicação (Ferreira et al., 2005; Aquino, 2008).
Em 2006, foram publicadas políticas governamentais voltadas para o
setor de plantas medicinais e, desde então, o governo federal do Brasil, em
parceria com um número elevado de prefeituras, têm estruturado programas
para o uso de fitoterápicos por meio do decreto 5813/2006, a exemplo da
Política Nacional de Plantas Medicinais e Fitoterápicas (PNPMF), portaria
interministerial 2960/2008 e a Política Nacional de Práticas Integrativas e
complementares (PNPIC), portaria 971/2006, integrado ao Sistema Único de
Saúde (SUS). Essas iniciativas foram tomadas com o objetivo claro de
contribuir consideravelmente para o uso racional de fitoterápico, e na promoção
de ações efetivas que constitui na distribuição de medicamentos fitoterápicos.
Tais políticas trazem como diretrizes, dentre outras ações, o incentivo à
pesquisa sobre as plantas medicinais, priorizando a biodiversidade do país,
motivando as instituições de pesquisas e as indústrias farmacêuticas buscarem
estratégias para avançar etapas na obtenção de novos fitomedicamentos,
obedecendo ao controle de qualidade (Brasil, 2014; Brasil, 2016).
Existem requisitos de qualidade que a produção de medicamentos
obrigatoriamente obedece, os quais passam, por exemplo, pela avaliação dos
componentes empregados na sua produção. Para cumprir essas exigências, a

15
Farmacopéia Brasileira, inclui monografias de diversas plantas utilizadas como
medicinais destacando experimentos e as padronizações necessárias que
garantem minimamente a qualidade dessas plantas como drogas vegetais
numa linha de produção de medicamentos (Farmacopeia Brasileira, Ed. V,
2010).
Mais de 70 espécies vegetais que ocorrem no Brasil possuem
monografias publicadas na quinta edição da Farmacopeia Brasileira (2010),
onde são apresentados os critérios para definir a identidade, bem como pureza
e teor de constituintes químicos. Ressalta-se que a preparação de uma
monografia oficial sobre determinada planta medicinal exige estabelecer as
características botânicas, químicas, farmacológicas e toxicológicas, além de
estudos científicos mais detalhados, incluindo a analise morfo-anatômica, a
padronização química e testes de pureza nos procedimentos de fabricação de
um fitoterápico a partir de drogas vegetais (Simões e Schenkel, 2001; Maciel et
al., 2002; Veiga e Pinto, 2005; Brasil, 2014). As análises físico-químicas, por
exemplo, são procedimentos que aperfeiçoam a qualidade na produção de
medicamentos e, consequentemente, imprimem uma maior segurança na
qualidade na obtenção de um fitoterápico (Abifito, 2003; Brasil, 2014; Kroes,
2014).
O estudo farmacobotânico em conjunto com avaliações físico químicas,
contribui no controle de qualidade, padronização das composições química das
drogas, tornando o seu uso eficaz e seguro, pois é comum que espécies
distintas, pertencentes a diferentes famílias, sejam confundidas e empregadas
para um mesmo fim terapêutico pelo fato de apresentarem características
morfológicas semelhantes ou nome popular comum. Assim é que as análises
morfo-anatômicas fornecem subsídios que contribui na padronização dos
insumos e permitem a diferenciação entre as amostras, inclusive entre
espécies botanicamente próximas (Amaral et al., 2005; Kremer et al., 2005).
Por outro lado, tem-se de ter claro que o perfil químico e
consequentemente a atividade biológica de plantas medicinais pode ser
afetado pelas condições de cultivo, manufatura, comercialização e distribuição.
A biossíntese dos metabólitos secundários sofre variações em função de
diferenças fisiológicas, genéticas no qual se encontra o vegetal. Também,
horário de colheita, condições de armazenagem, secagem, métodos de

16
extração, a forma como é feito o processamento e o tipo embalagem interfere
no resultado final utilizada (Sahoo et al., 2010; Folashade et al., 2012).
Além de possíveis falsificações e equívocos com a droga vegetal que se
fará uso em uma pesquisa ou na linha de produção de um medicamento,
destaca-se que a época, local e condição em que uma droga é coletada são
fatores de suma importância no que se refere a quantificação de seus
metabólitos secundários. Estudos relatam que a quantidade e, às vezes, até
mesmo a natureza dos constituintes químicos ativos não é constante durante
todo o ano, nem em todos os locais em que a planta ocorre naturalmente ou é
cultivada. É comum o relato de variações sazonais no conteúdo de
praticamente todas as classes de metabólitos secundários. A temperatura, luz,
disponibilidade de água também estão diretamente envolvidos na composição
química das plantas (Grace, 1998; Hogedal, 2000; Agerbirk, 2001; Tattini,
2004; Voltolino e Santos, 2011).
Para garantir que os medicamentos disponíveis no mercado
farmacêutico sejam seguros e eficazes é necessário que estes cumpram as
normas técnicas para a produção e o controle de qualidade. Na indústria
farmacêutica, o controle de qualidade das drogas de origem vegetal é de
fundamental importância para garantir a qualidade do produto final, e,
consequentemente, do medicamento comercializado ao paciente. Além dos
ensaios para aprovação, todos os medicamentos devem passar também por
estudos de estabilidade, conforme determinado pela resolução RE nº 01 de 29
de julho de 2005 (Brasil, 2005; Oliveira, 2009). Nos procedimentos rotineiros de
análise
da qualidade, geralmente é preconizado o emprego de metodologias em
controle físico-químico e farmacobotânica, sendo necessária à correlação entre
os parâmetros analisados e a finalidade a que se destina o medicamento. O
controle de qualidade procura avaliar as drogas, quanto a identificação correta
(parte usada e espécie botânica), pesquisas qualitativa e quantitativa de
marcadores químicos (muitos deles, princípios ativos) e de impurezas e
falsificações, segundo as monografias das farmacopeias e publicações
científicas. Na ausência de parâmetros estabelecidos, padrões de drogas são
utilizados para comparação (Simões et al., 2001). Para isso são feitos muitos
tipos de teste tais como: índice de intumescência, densidade relativa, testes de

17
pureza e impurezas, teste de metais pesados, chumbo, umidade, perda por
secagem, cinzas totais, saponinas. Todos os procedimentos devem ser
realizados de acordo com os protocolos das farmacopeias e darão resultados
que devem ser padronizados para cada droga vegetal diante de suas
singularidades (Brasil, 2010).
O uso popular e mesmo tradicional não são suficientes para validar as
plantas medicinais como medicamentos eficazes e seguros (Simões et al.,
2001). Neste sentido, as plantas medicinais não se diferenciam de qualquer
outro produto sintético. Se a intenção é utilizar uma planta medicinal como
medicamento, ela deve ter sua ação comprovada e sua toxicidade potencial
avaliada cientificamente como qualquer outro medicamento (Brandão, et al.,
2006; Melo et al., 2007; Brandão et al., 2009a). Contudo, a partir do
estabelecimento dos parâmetros de qualidade para a matéria-prima, e
considerando-se um planejamento adequado e um controle do processo de
produção do produto final, a qualidade do medicamento estará, em grande
parte, assegurada. Portanto, a qualidade da matéria-prima vegetal é a
determinante inicial da qualidade do fitoterápico, mas não garante a eficácia, a
segurança e a qualidade do produto final (Oliveira, 2009). Ressalta-se ainda
que o controle de qualidade de drogas vegetais deve ser realizado em todas as
etapas que envolvem a produção das mesmas, como parte do cultivo racional
(solo, luminosidade, temperatura, irrigação), colheita e pós colheita,
embalagem e armazenagem adequadas, para que o produto final atenda aos
requisitos de qualidade para um medicamento (Brasil, 2005; Oliveira, 2009).
Questões como as descritas acima demarcam o quanto é importante
considerar tanto o manejo, como o local onde a planta é cultivada, por
constituírem determinantes importantes para garantir a qualidade, a eficiência e
a eficácia de um medicamento, podendo interferir diretamente na quantidade e
características dos metabolitos secundários presentes na planta (Souza et al.,
2010). Nesse sentido, este trabalho traz um estudo comparativo
farmacobotânico, físico-químico, fotoquímico e biológico de Senna alata (L.)
Roxb. de acessos da caatinga (BA) e da restinga do Brasil.

18
1.1 Caatinga x Restinga
O Brasil abriga vários biomas, como por exemplo, a restinga e a
caatinga. A Caatinga ocorre exclusivamente em território brasileiro, com
abrangência de 70 % da região nordeste e ainda parte de Minas Gerais,
totalizando 11 % do território nacional (Giulietti et al., 2003; Leal et al., 2003;
Araújo e Sobrinho, 2009). Na linguagem tupi, caatinga significa Mata Branca
(caa = Mata + tinga = branca). A razão para esta denominação reside no fato
dessa vegetação apresentar-se verde somente em pequenas épocas de chuva,
o que é raro. No restante do ano, a caatinga é vista sem folhas, normalmente
em vegetação baixa, o que permite que a vista penetre sem dificuldade até
grande distância, sobressaindo a cor clara dos caules que ficam
esbranquiçados na ausência da folhagem (Maciel et al., 2010).
Outras características peculiares das plantas da caatinga são
encontradas na presença de caules e raízes suculentas que armazenam água
e nutrientes, além do ciclo de vida dessas plantas que em geral é curto e
comumente há dormência nas sementes. Essas plantas, em grande número
são utilizadas pelos sertanejos como alimentos, remédios, plantas forrageiras e
fontes de madeira e de energia (Maciel et al., 2010; EMBRAPA, 2012).
Giulietti, 2003, afirmou que apesar desse bioma encontrar-se bastante
alterado, especialmente nas terras mais baixas, contém uma variedade de tipos
vegetacionais, com elevado número de espécies vegetais e remanescentes de
vegetação ainda bem preservada, que incluem um número expressivo de
táxons endêmicos.
Diferente da caatinga, em um espaço geográfico formado por depósitos
à linha da costa, onde se encontram diferentes comunidades que recebem
influência marinha, encontra-se a restinga. Essa vegetação está localizada ao
longo da faixa litorânea, na Zona Costeira (mais de 8,5 mil Km de extensão em
terras brasileiras), elas são marcadas por uma grande variedade de paisagens
que se repetem ao longo do litoral (EMBRAPA, 2012).
O CONAMA (Conselho Nacional de Meio Ambiente) em 2002, na
resolução 3303/2002, art.2°, definiu restinga como:

19
Depósito arenoso paralelo a linha da costa, de forma geralmente alongada, produzido por processos de sedimentação, onde se encontram diferentes comunidades que recebem influência marinha, também consideradas comunidades edáficas por dependerem mais da natureza do substrato do que do clima. A cobertura vegetal nas restingas ocorre mosaico, e encontra-se em praias, cordões arenosos, dunas e depressões, apresentando, de acordo com o estágio sucessional, estrato herbáceo, arbustivos e abóreo, este último mais
interiorizado. (CONAMA, 2002, resolução 3303/2002, art. 2°)
Assim é que a restinga pode ser encontrada como resultado de
diferentes fenômenos naturais, onde as principais formações deste bioma que
ocorrem no extenso litoral brasileiro encontram-se de São Paulo a Alagoas
(Amaral, 2008). Frequentemente faz-se referência ao fato das restingas no
Brasil apresentarem poucos endemismos, o que comumente é justificado pelo
fato das áreas da planície costeira ser relativamente recentes do ponto de vista
geológico (Rizzini, 1979; Araújo e Henriques, 1984, Silva, 1990). No Rio de
Janeiro, a restinga está formada a partir de sedimentos depositados pelo mar,
trazidos ou da plataforma continental ou por retrabalhado nos sedimentos
quem vem pelos rios depositados na praia (Silva, 1990).
Conforme autores como Irwin e Barneby (1982), citado por Rodrigues e
colaboradores (2005), apesar das variabilidades geológicas e climáticas,
existem espécies vegetais que são comumente encontradas tanto na caatinga
quanto na restinga, e outras espécies vegetais são introduzidas nestes biomas,
como é o caso de Senna alata. Essa coexistência em biomas diferentes
acarreta algumas peculiaridades à pesquisa, pelo fato de os vegetais
apresentarem características específicas de acordo o ambiente onde estão
inseridos (Souza et al., 2010).
1.2 Família Fabaceae
A família botânica Fabaceae é popularmente conhecida como
leguminosa, e constitui-se na terceira maior família de angiospermas, com
19325 espécies abrigadas em 727 gêneros (Lewis et al., 2005). É comum
encontrá-las em uma grande extensão do globo terrestre, principalmente em
regiões com condições tropicais de alta temperatura e umidade, estando
também presente em menor número nas regiões temperadas (Lima et al.,
1994). As leguminosas são reconhecidas por ser uma família de “múltiplo uso”,
o que provavelmente está relacionado com sua elevada diversidade em

20
espécies. O potencial econômico dessa família é amplamente conhecido, pois
muitas espécies fornecem madeiras, óleos e resinas de boa qualidade.
Produzem matéria prima para perfumes, tinturas e fármacos, sendo muitas
espécies utilizadas como ornamental. É comum também o consumo dos seus
frutos. Além disso, devido associação que fazem com bactérias capazes de
fixar nitrogênio, diversas espécies desta família são empregadas como adubo
natural (Lewis e Owen, 1989; Costa et al., 2008).
Uma particularidade das espécies que constituem a família Fabaceae
está na plasticidade de hábitos de crescimento, o que incluem ervas, lianas e
plantas arbustivas de diferentes tamanhos. O caule apresenta folha composta
ou recomposta e as suas flores são tipicamente chamativas para atrair
polinizadores, além disso, essa família é normalmente reconhecida devido a
presença de frutos com formato de vagem (Judd, 2002).
Para Fabaceae são reconhecidas as subfamílias: Caesalpinioideae,
Mimosoideae e Papilionoideae (ou Faboideae), conforme Lorenzi (2002) e Judd
(2002). Nesse trabalho destaca-se a subfamília Caesalpinoidea que possui
aproximadamente 154 gêneros e 2800 espécies, com ocorrências nas regiões
tropicais e subtropicais, principalmente na América do Sul, África tropical e
sudeste da Ásia, sendo pouco representada na América do norte e em outras
regiões temperadas (Cowan, 1981; Lewis, 1987; Lewis e Polhill, 1998, Judd,
2002). No Brasil foram catalogados aproximadamente 178 gêneros e
aproximadamente 1550 espécies (Barroso et al., 1978; Lorenzi, 2002).
De acordo com Lewis e Polhill (2005), existem 4 tribos atualmente
aceitas para Caepinoideae: Caesalpinieae, Cassieae, Cercideae e Detarieae. A
espécie vegetal que trata esse estudo, Senna alata, pertence a tribo Cassieae
que compreende seis gêneros (Apuleia, Cassia, Chamaecrista, Dialium,
Martiodendron e Senna) muito similares, principalmente Cassia e Senna,
(Lorenzi, 2002; Costa et al., 2008). Irwin e Barneby (1982) fizeram
diferenciações indispensáveis a serem observadas e incluídas nas chaves de
identificação para estes dois gêneros. Senna distingue-se de Cassia
principalmente pelos filetes retos, mais curtos ou até duas vezes o
comprimento das anteras, pelas anteras basifixas e pela presença de nectários
extraflorais na maioria das espécies. Por outro lado, Senna difere de
Chamaecrista principalmente pela ausência de bractéolas (excepcionalmente

21
presentes), pelo androceu zigomorfo e pelos legumes que podem ser
indeiscentes (Lorenzi, 2002).
O gênero Senna é monofilético e é o segundo maior gênero da tribo
Cassieae com aproximadamente 300 espécies de distribuição pantropical, das
quais cerca de 200 espécies são americanas (Irwin e Barneby, 1982; Lewis,
2005). No Brasil tem-se cerca 80 espécies, das quais, 26 ocorrem
exclusivamente neste país (Souza e Bortoluzzi, 2012). O trabalho realizado por
Irwin e Barneby (1982), o mais abrangente para o gênero, caracteriza o mesmo
pelas flores amarelas, normalmente assimétricas, sem bractéolas no pedicelo,
androceu heteromórfico e anteras basifixas, bem como folhas comumente com
nectários entre os pares de folíolos e frutos predominantemente indeiscentes.
Segundo Lewis e colaboradores (2005), o gênero possui muitas
espécies com propriedades medicinais. Outras têm sua madeira empregada na
construção civil ou como ornamental, no paisagismo e arborização de ruas,
parques e jardins, devido a grande beleza no período do seu florescimento
(Lorenzi, 2002). Varias espécies de Senna são utilizadas na medicina popular.
A maioria delas foi descritas e classificadas cientificamente pelos botânicos
Irwin e Barneby, Philip Miller (Mill), pelo médico e botânico escocês William
Roxburgh (Roxb.) e pelo alemão Carl Ludwig Willdenow (Will) também
botânico, médico e farmacêutico. Entre as espécies catalogadas por esses
autores, destaca-se S. alexandrina, S. corymbosa, S. didymobotrya, S. italica,
S. reticulata, S. spectabilis e S. sophera, todas com nome popular como sene,
fedegoso ou cássia. As folhas da maioria destas plantas citadas são utilizadas
como laxante por apresentar antraquinonas em sua constituição química
(Ibrahim e Osman, 1995; Khan et al., 2001; Somchit et al., 2003; Ordoñez et al.,
2004; Barrese et al., 2005; Plantamed, 2016).
Dentre estas espécies, há destaque para Senna alexandrina, também
cognominada de Cassia angustifolia, descrita pelo botânico inglês de origem
escocesa Philip Miller (Mill). Senna alexandrina é popularmente conhecida
como Sene, e é considerada como planta medicinal, com vasta utilização em
fitoterapia sendo prescrito para as atividades laxativa, purgativa e em
tratamento de constipação intestinal, além de possuir componentes que
auxiliam no combate de verme e alivio de flatulência. Nativa no Egito, onde é
cultivada comercialmente, S. alexandrina também cresce em vários outros

22
locais do oriente, no entanto o potencial dos seus constituintes químicos não se
revela quando cultivado fora das condições climáticas adaptativas (Blumenthal
et al., 1998; Heber, 2004; ANVISA, 2016).
1.3 Senna alata (L.) Roxb.
Senna alata (L.) Roxb. (Figura 1), pertence à família Fabaceae. Nativa
do norte da América do Sul, essa espécie vegetal se tornou cosmopolita
tropical, com vasta distribuição geográfica na África, Ásia, além de toda a
América tropical, onde é cultivada desde os Estados Unidos da América até a
Argentina. Esta planta é também conhecida pela sinonímia de Cassia alata, e
possui denominações populares como alcapulco, “fedegoso gigante”,
“fedegosão (ao odor dessa espécie que não se compara a nenhum outro,
considerando Suigeneres)”, “mangerioba-do-pará”, “mangerioba-grande
(devido quando torradas são utilizadas no preparo de uma bebida semelhante
ao café)”, “mata-pasto-grande (sugestivo ao potêncial contaminante em áreas
de pastoreiro)”, “café-beirão”, “candelabro amarelo e dartrial” (Irwin e Barneby,
1982; Nascimento e Queiroz, 2012; Souza e Bortoluzzi, 2012). Constitui-se
uma espécie perene e arbustiva muito frequente dentro e no entorno de
cidades, ocorrendo naturalmente em áreas de pastagens, beira de estradas e
terrenos baldios, preferencialmente em lugares úmidos, havendo tendência a
formação de estandes puros (Souza F., 1995; Lorenzi, 2000). A planta
apresenta caule fino com circunferência no nível do solo de 34,2 cm, com 2 m
de altura e copa esgalhada e espalhada em todas as direções, algumas vezes
muito ramificada a partir da base. Trata-se de uma planta pioneira e perene de
crescimento rápido declinando o seu desenvolvimento após 2-3 anos de
estabelecimento no local (Irwin e Barneby, 1982; Rodrigues et al., 2005; Souza
e Bortoluzzi, 2012).
Em geral é tratada como uma planta daninha, constituindo-se em um
problema de ordem bioeconômica a limitar o desempenho produtivo e a
rentabilidade da atividade agrícola (Rodrigues et al., 2009). Contudo, por outro
lado, tem uso etnofarmacológico como já descrito neste trabalho e ainda,
conforme Viana e colaboradores (2008) e Medeiros e colaboradores (2011)
esta planta pode ser uma importante ferramenta no controle de pragas
agrícolas, eficaz no controle de micro-organismos, através do uso dos seus

23
extratos, uma vez que a quimiotaxonomia aponta constituinte como flavonoides
e outros polifenois no gênero.
1.4 Emprego medicinal de Senna alata (L.) Roxb.
As folhas de Senna alata são utilizadas popularmente devido as suas
propriedades medicinais. Na Índia, todas as partes da planta são utilizadas nos
sistemas de medicina ayurvedica, unani e alopática, principalmente como
finalidade digestiva (Damodaran e Venkataraman, 1994). Pesquisas apontaram
que também são utilizadas popularmente no tratamento de anemia,
blenorragia, congestão do fígado, hemorróidas, herpes, malária, pano branco,
tratamento e prevenção da erisipela, sarnas, tumores, leucemia, tuberculose,
câncer e inflamações (Ibrahim e Osman, 1995; Khan et al., 2001, Somchit et
al., 2003, Awal et al., 2004; Barrese et al., 2005 e Pieme et al., 2006). As
folhas, inflorescências e raízes de S. alata têm atividade biológica comprovada
como sendo antimicrobiana eficaz no tratamento de dermatites, blenorragia e
herpes, além de ser diurética e laxativa. No entanto, além dessas propriedades
terapêuticas, essa planta deve ser ministrada com cuidado, pois é suspeita de
ser tóxica aos rins e, ainda, considerada abortiva (Lorenzi, 2000; Plantamed,
2016).
1.5 Substâncias químicas no gênero Senna e Senna alata (L) Roxb.
Em vegetais, tem-se muitos metabólitos secundários que são
substâncias químicas capazes de atuarem diretamente no crescimento,
desenvolvimento, reprodução e defesa dos organismos principalmente contra a
A B
Figura 1 Aspecto geral de Senna alata (L.) Roxb. Legenda : (A) Acesso caatinga; (B) Acesso Restinga.
Fonte: Nascimento, 2016.

24
herbivoria e ao ataque do oxigênio produzido dentro da planta pela máquina
fotossintetizante (Foye e Williams, 1995; Fraga, 2001; Stamp, 2003).
Segundo Gupta e Singh (1991), espécies do gênero Senna são ricas em
flavonoides, antraquinonas, taninos e saponinas. Além disso, foram relatados
para esse gênero esteroides, alcaloides piperidínicos e isoquinolínicos,
cromonas, lactonas, estilbenos e triterpenos (Alemayehu e Abegaz, 1996;
Valencia et al., 2000). Apesar da correlação de compostos entres os gêneros
da subfamília Caesalpinoide, as espécies se diferem entre si devido a
existência ou ausência dessas substâncias químicas naturais (Alemayehu et
al., 1998).
As folhas de S. alata são ricas em cristais de oxalato de cálcio, ao longo
de suas nervuras - característica da subfamília Caesalpinioideae, além de
flavonoides e antraquinonas que foram encontrados em células epidérmicas e
em células dispersas no parênquima paliçádico, especialmente nas
proximidades da nervura mediana (Barrese et al., 2005). Segundo Ordoñez
(2004), as folhas possuem flavononas, flavonas e xantonas além de esteroides
livres e taninos catéquicos.
No sistema biológico, os flavonoides apresentam capacidade
antioxidativa (esta constitui a atividade mais elucidada pelos estudos até agora
desenvolvidos), atividades anti-inflamatórias e de efeito vasodilatador, ação
antialérgica, atividade contra o desenvolvimento de tumores, anti-hepatotó-
xica, antiulcerogênica, atuação antiplaquetária, bem como ações
antimicrobianas e antivirais. Os flavonoides podem inibir vários estágios dos
processos que estão diretamente relacionados com o início da aterosclerose,
como ativação de leucócitos, adesão, agregação e secreção de plaquetas
(Lemmens, 2015), além de atividades hipolipidêmicas e aumento de atividades
de receptores de LDL (Koes et al., 1994; Peterson, 1998; Zuanazzi e
Montanha, 2010).
Moriyama e colaboradores (2001; 2003) reportaram a presença
kampferol-3-O-gentiobioside e Luteolina (Figura 2) em folhas de S. alata,
demonstrando que durante o período de outubro a dezembro há decréscimo no
conteúdo destes flavonoides.
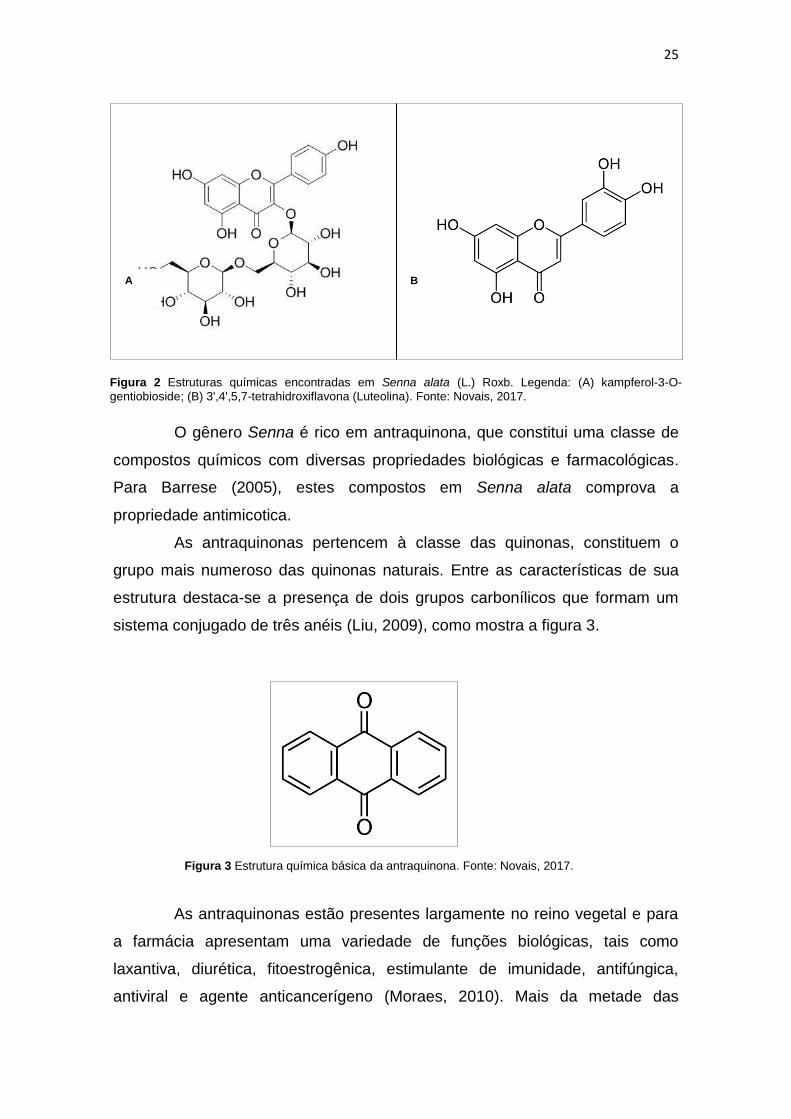
25
O gênero Senna é rico em antraquinona, que constitui uma classe de
compostos químicos com diversas propriedades biológicas e farmacológicas.
Para Barrese (2005), estes compostos em Senna alata comprova a
propriedade antimicotica.
As antraquinonas pertencem à classe das quinonas, constituem o
grupo mais numeroso das quinonas naturais. Entre as características de sua
estrutura destaca-se a presença de dois grupos carbonílicos que formam um
sistema conjugado de três anéis (Liu, 2009), como mostra a figura 3.
As antraquinonas estão presentes largamente no reino vegetal e para
a farmácia apresentam uma variedade de funções biológicas, tais como
laxantiva, diurética, fitoestrogênica, estimulante de imunidade, antifúngica,
antiviral e agente anticancerígeno (Moraes, 2010). Mais da metade das
A
A B
Figura 2 Estruturas químicas encontradas em Senna alata (L.) Roxb. Legenda: (A) kampferol-3-O-
gentiobioside; (B) 3',4',5,7-tetrahidroxiflavona (Luteolina). Fonte: Novais, 2017.
Figura 3 Estrutura química básica da antraquinona. Fonte: Novais, 2017.

26
antraquinonas naturais são encontradas em fungos, a exemplo das espécies
Penicillium e Aspergillus e em liquens. Existem alguns casos isolados
encontrados em insetos, como o ácido carmínico, extraído das fêmeas da
colchinilha (Dactylopius coccus), um inseto hemíptero da família dos coccídeos,
muito perniciosos às plantas cultivadas (Liu, 2009; Moraes, 2010).
Oliveira (2012) identificou a presença de 53 antraquinonas descritas
para o gênero Senna. Entre esses, 24,5 % dos compostos presentes nos
registros encontrados são dímeros, 18,9 % são glicosídeos e 56,6 % são
antraquinonas livres. Alemayehu e colaboradores (1996) e Valência e
colaboradores (2000), constataram em S. alata, a presença de 1,5,7-trihidroxi-3
metilantraquinona (emodina) e senosidio A e B, o que justifica o potencial
laxativo de suas folhas.
Devido à proximidade das espécies, S. alata apresenta composição
química similar à S. alexandrina, a exemplo da presença de emodina,
antraquinona, flavonoide do tipo campferol, mucilagens, cristais de oxalato de
cálcio e senosídio A e B (Blumenthal et al., 1998; Heber, 2004; Ordoñez, 2004;
Barrese et al., 2005; Liu, 2009). Essas substâncias quimicas podem estar
diretamente relacionadas à função biológica desta planta.
1.6 Atividades biológicas
Estudar as atividades biológicas ou farmacológicas que uma
determinada droga apresenta é essencial para descrever os efeitos benéficos
ou adversos dela sobre os seres vivos. Esses efeitos podem ser revelados a
partir de modelos in vitro ou in vivo (Fraga, 2001).
A utilização de modelos in vitro como estratégia de redução do número
de animais é proposto em clássicos como o livro “The Principle of Humane
Experimental Technique” de Russel e Bruch em 1959. Assim é que, em
princípio, deve-se utilizar a atividade in vivo para confirmação dos resultados
encontrados nos testes in vitro quanto satisfatória ou, quando o teste in vitro
não é suficiente para avaliação da atividade em questão, a exemplo do estudo
dos agentes esquistossomicida (Araújo, et al., 2016).

27
1.6.1 Atividade antimicrobiana em bioautografia
Os métodos mais utilizados para detecção da atividade antimicrobiana
são ensaios in vitro, que normalmente não trazem informações sobre o modo
de ação do extrato vegetal. As avaliações in vitro podem ser realizadas por
difusão, diluição ou bioautografia. Essas metodologias objetivam identificar se
um extrato, fração ou molécula possui ou não atividade antimicrobiana, além de
avaliar a intensidade dessa atividade e identificar os componentes do extrato
da planta com atividade frente a bactérias ou fungos (Zachino, 2001; Ostrosky
et al., 2008).
A bioautografia é um protocolo comumente utilizado na pesquisa de
produtos naturais desde 1946, quando foi utilizado pela primeira vez por
Goodall e Levi. Constitui-se um método prático que basea-se no
desenvolvimento dos micro-organismos, junto a uma cromatografia em camada
delgada formando zonas de inibição em bandas específicas em seus Rfs na
cromatoplaca. Esta prática permite o biomonitoramento para o isolamento e
purificação de substâncias químicas que tenham atividade antimicrobiana
desejada a patir de um extrato vegetal. Para tanto, deve-se atentar à qualidade
da cromatografia, da suspensão bacteriana e as condições de incubação das
cromatoplacas (Botz et al., 2005; Colorado et al., 2007; Hostettmann et al.,
2008). Por outro lado, apesar de ser considerado simples e de baixo custo,
esse ensaio, quando utilizado em meio ágar, não é apropriado para análise de
compostos que não se difundem facilmente nesse hidrocoloide (Khurram et al.,
2009). Para tanto, existem três métodos bioautográficos que são utilizados:
bioautografia de contato, método de sobreposição do ágar e a biautografia
direta (Scorzoni et al., 2007). Nos dois primeiros métodos utiliza-se ágar
enquanto que no terceiro método esse hidrocoloide não é utilizado.
Na bioautografia de contato (método realizado neste estudo), as
cromatoplacas são colocadas nas placas de petri e cobre-se a mesma com o
meio de cultura onde os micro-organismos são inoculados. Já no ensaio de
sobreposição do ágar, a placa de petri é coberta primeiramente com o meio de
cultura onde os micro-organismos são inoculados e as cromatoplacas são
sobrepostas a ele. Estes experimentos são vantajosos principalmente pelo fato
de apresentar resultados satisfatórios a compostos polares ou moderadamente
polares (Botz et al., 2005; Choma, 2005; Khurram et al., 2009). A bioautografia

28
direta utiliza apenas a cultura líquida do micro organismo, que cresce
diretamente sobre a cromatoplaca, de forma que este terceiro subtipo de
bioautografia é apropriado para análise de compostos que não se difundem
facilmente no ágar (Horváth et al., 2010).
Nas três variações da bioautografia, após o período de incubação dos
micro-organismos, em que a cromatoplaca dentro da placa de petri com o meio
onde o micro organismo está se desenvolvendo, é mantida em estufa, borrifa-
se sal tetrazolico que é convertido quase que imediatamente em formasan,
corando de vermelho os locais com atividade antimicrobiana. Assim é que se
formam manchas claras pela inibição de crescimento do micro-organismo
utilizado e, essas manchas, apresentam-se bem localizadas, demarcando no
cromatograma as zonas de inibição nos respectivos Fatores de retenção (Rfs)
e dessa forma, apontando as bandas com atividade antimicrobiana para cada
micro-organismo utilizado (Choma, 2005; Valgas et al., 2007).
1.6.2 Alelopatia: interferências sobre comunidades vegetais
Nas comunidades vegetais, as plantas interagem entre elas de maneira
positiva, negativa ou neutra. Comumente, as plantas vizinhas interagem de
maneira negativa, por exemplo, enquanto algumas espécies absorvem maior
quantidade de nutrientes outras não conseguem absorver o suficiente para sua
sobrevivência, ocorrendo, consequentemente o crescimento de uma planta ou
a inibição de ambas. Os metabólitos secundários sintetizados por plantas,
frequentemente geram efeitos em outros organismos e, em muitos casos,
possuem a função de compostos químicos de defesa ou aleloquímicos (Mota,
1975; Chom e Kim, 2004; Macias, 2007). No ano de 1969, Muller determinou o
termo interferência como as interações entre os organismos vizinhos. Por este
termo ser muito abrangente, em 1977, Szczepanski o subdividiu em alelopatia,
alelospolia e alelomediação. Contudo, há de se destacar que o termo
alelopatia, utilizado para denominar essas interferências, foi criado por Molisch
em 1937 tendo origem no grego allelon = de um para outro, pathós = sofrer,
conforme Souza (2006).
A alelopatia é uma interferência caracterizada pela introdução de
substâncias químicas por um organismo vivo de forma que estes componentes
venham a prejudicar o desenvolvimento normal de outros componentes da

29
comunidade. A alelospolia, também denominada de competição, é
caracterizada toda vez que um organismo vivo, consegue absorver o máximo
de nutrientes, luz e água, prejudicando o desenvolvimento das plantas vizinhas.
A alelomediação ou interferência indireta, se caracterizada por ser uma
modificação física no ambiente que é provocada por um organismo com
reflexos aos seres vivos vizinhos (Szczepanski, 1977; Muller, 1969; Formagio,
2010).
É importante ressaltar que o conceito de alelopatia descreve a
capacidade de um vegetal interferir de forma natural no desenvolvimento da
vegetação adjacente, por meio de substâncias químicas denominadas
“aleloquímicos” liberados no ambiente, quem venham promover qualquer efeito
direto ou indireto, danoso ou benéfico, sobre outra planta (Rice, 1984; Rizvi et
al., 1992; Rodrigues et al. 1999; Macías, 2007).
Os aleloquímicos são liberados na atmosfera ou no solo por quatro vias:
exsudação radicular, lixiviação, volatilização e decomposição dos resíduos da
planta. A exsudação radicular é a liberação de compostos na forma líquida
(aminoácidos, nucleotídeos), através das raízes, diretamente no solo. A
lixiviação ocorre com a liberação de compostos exsudados na forma líquida
(compostos fenolicos e alcaloides como é o caso da cafeína e a nicotina),
normalmente pelas folhas que são lavados da planta por ação da água da
chuva ou irrigação e carregados até o solo. Na volatilização há a liberação de
compostos como o limoneno, mirceno e o mentol que são micromoléculas que
se localizam normalmente em estrutras epidérmicas e se volatizam com
facilidade. Por fim, a última via de liberação da-se pela decomposição de restos
vegetais, havendo a liberação de compostos químicos, como isoflavonas e
antocianinas, na medida em que os restos vegetais vão sendo decompostos
por micro-organismos ao longo do tempo (Carvalho, 2013).
Independente da forma que ocorre a inibição, a interferência pode ser
aproveitada de forma positiva na produção de inseticidas, repelentes de
insetos, venenos usados na pesca, para o desenvolvimento de produtos
biológicos com ação sobre plantas (bio-herbicidas) na agricultura e +no
controle de plantas daninhas a uma determinada cultura através da
rotação/sucessão de culturas ou no uso de cobertura viva na entressafra e no
uso de cobertura morta (Chom e Kim, 2004; Souza, 2006). Conforme Malheiros

30
e Peres (2001) e Macías (2007), entre os aleloquímicos comumente citados
como responsáveis por causarem efeitos diretos e indiretos estão os terpenos,
alcaloides, flavonoides, ácidos graxos de cadeia longa e lactonas insaturadas.
Os terpenos, por exemplo, podem atuar como inseticidas, como por exemplo, o
pineno, o limoneno, o mirceno e os peretroides além de algumas saponinas.
Podem, ainda, atuarem como repelentes ou atrativos de insetos como muitos
óleos essenciais e lactonas sesquiterpênicas, ou mesmo como compostos
tóxicos e exemplo do phorbol, algumas saponinas e resinas. Dentre os
flavonoides destacam-se os isoflavonoides que funcionam como inseticidas e
as antocianinas que funcionam como atrativos de insetos.
1.6.3 Atividade citotóxica pelo método MTT
O termo citotoxicidade, segundo NIH (2001), significa causa de efeitos
tóxicos como morte celular, devido, por exemplo, alterações na permeabilidade
da membrana celular ou inibição enzimática. Com isso testes de viabilidade
celular in vitro são utilizados para verificar citotoxicidade aguda, dentre os quais
está o ensaio de proliferação celular {brometo de [3-(4,5-dimetiltiazol-2-il)-2,5-
difenil tetrazólio]} (MTT), conforme NIH (2001) e Lazzarini (2015).
Em geral, os ensaios de atividade citotóxica medem a prorção de células
viáveis após um procedimento traumático. Conforme definido na ISO 10993-5
(2009), pode-se avaliar a citotoxicidade por análise morfológica das celulas ou
por analises de aspectos referentes ao metabolismo celular. Os testes
validados para esse fim são o 3T3 NRU, teste de citotoxicidade para formação
de colônias, MTT, MTS e XTT (2,3-bis (2-metoxi-4nitro-5-sulfofenil)-5-
[(fenilamino) carbonil]-2H-hidroxido de tetrazólio) de citotoxicidade (NIH, 2001;
ICCVAM, 2006).
O ensaio do MTT é um teste colorimétrico usado para avaliar a
viabilidade celular em que as desidrogenases mitocondriais, presentes apenas
em células metabolicamente viáveis, clivam o anel de tetrazólio, transformando
sua coloração amarela em violeta, pela formação do composto formasan {E, Z-
1-(4,5-dimetiltiazol-2-il)-1,3-difenilformasan}, cristais insolúveis em soluções
aquosas (figura 4). Dessa forma, a produção de formasan reflete o estado
funcional da cadeia respiratória, sendo sua formação diretamente proporcional

31
à atividade mitocondrial e a viabilidade celular (Rutala e Weber, 1999;
Lazzarini, 2015).
1.6.4 O processo inflamatório e o estimulo por lipopolissacarídeo
(LPS) e zymosan (Zy)
A inflamação ou flogose (do latim inflamare, do grego fhlogos, que
significa pegar fogo) é uma reação complexa dos tecidos conjuntivos
vascularizados em resposta a estímulos endógenos ou exógenos que podem
causar dano celular. Os estímulos endógenos são derivados de degenerações
ou necroses tissulares e de alterações na resposta imunológica, por
imunocomplexos ou reações autoimunes. Os estímulos exógenos referem-se à
indução por agentes físicos químicos e biológicos (Lima et al., 2009; Brasileiro
Filho, 2011).
Um fenômeno irritativo como um trauma, promove uma resposta rápida
do organismo, resposta esta que é mediada principalmente pelos leucócitos.
Assim é que num quadro inflamatório existem sinais marcantes, os chamados
danos teciduais, que provocam no local da inflamação calor, rubor, edema e
dor seguido da perda de função dos tecidos em casos mais avançados. Cada
um destes sinais corresponde ao avanço do quadro inflamatório (Iwalewa et al.,
2007; Lima et al., 2009).
A inflamação pode ser aguda ou crônica, tanto uma quanto a outra
possui os mesmos mecanismos iniciais, o que as diferem são o tempo de
exposição ao agente agressor, o tipo de agente e a resposta imune. A
Redução
Mitocondrial
MTT Formazan
Figura 4 Transformação de {brometo de [3-(4,5-dimetiltiazol-2-il)-2,5-difenil tetrazólio]} (MTT) em { E, Z- 1- (4,5-dimetiltiazol-2-il) -1,3-difenilformazan } (Formazan). Fonte: Novais, 2017.

32
inflamação aguda é de curta duração, totalizando alguns minutos a no máximo
dois dias, dependendo do estímulo causal. Suas principais características são
a exsudação de fluidos e proteínas do plasma e emigração de leucócitos,
predominantemente neutrófilos. A inflamação crônica é menos uniforme do que
a inflamação aguda, apresentando duração mais longa e caracterizando-se
histologicamente pela presença de linfócitos e macrófagos, proliferação de
vasos sanguíneos (neoangiogênese) e tecido conjuntivo denominado
fibroplasia (Janeway, 2002; Bechara, 2006; Lopes e Amaral, 2013).
Quando o estímulo desencadeante do processo inflamatório é
controlado por tratamento, a inflamação é inibida. Contudo, toda vez que o
estímulo não for devidamente tratado, o indivíduo que sofre a inflamação pode
chegar a um estado crônico desse processo inflamatório, a exemplo das
neoplasias, esclerose múltipla, lúpus eritematoso sistêmico (LES), acidente
vascular cerebral (AVC), Alzheimer e artrite reumatoide (Ballou et al., 1993;
Baumann et al., 1994; Ticani, 2003).
Na defesa contra agentes infecciosos a resposta imune é fundamental
em todo o processo, contituindo o principal impedimento para a ocorrência de
infecções disseminadas. Assim é que o quadro de infecção envolve uma ação
coordenada entre o sistema imunológico e o tecido no qual ocorreu a lesão,
sendo comum o surgimento de outras doenças caso a inflamação não seja
controlada (Tilley et al., 2001; Machado, 2004; Zhou et al., 2007; Rang e Dale,
2008; Lima et al., 2009).
Experimentalmente, uma inflamação pode ser induzida por diferentes
métodos in vivo ou in vitro (Mogilski et al., 2016). Um método frequentemente
utilizado para o estímulo da inflamação in vitro tem sido realizado com o
lipopolissacarídeo (LPS). O LPS é uma endotoxina presente em bactérias
gram-negativas que consiste de uma estrutura composta por duas camadas de
açúcar e uma camada lipídica, uma parte hidrofílica e outra hidrofóbica,
respectivamente, sendo que a porção lipídica é considerada a responsável pela
maior ação antigênica. A presença de LPS, mesmo que em pequena
quantidade, é identificada pelos monócitos, que através de uma resposta
imunológica inata passa a expressar diferentes moléculas A sinalização
intracelular induzida pela interação do LPS com seus receptores desencadeia a
ação de genes relacionados à resposta inflamatória aguda por intermédio de

33
citocinas pró-inflamatórias a exemplo do TNF-α (Rietschel et al., 1994; Chen e
Greene, 2004; Tuin et al., 2006; Gosh e Hayden, 2008).
Assim como o estímulo à resposta imunológica frente a um processo
inflamatório realizado com o LPS, tem-se o polissacarídeo zymosan (Zy) como
sendo outro estímulo à essa resposta imunológica. Zy é um polissacarídeo
proveniente da parede de Saccharomyces cerevisiae, capaz de ativar
macrófagos e induzir a liberação de mediadores inflamatórios. O Zy é utilizado
em ensaios por administração via intraperitoneal em animais de laboratório,
pois estudos demonstram que a inflamação induzida por estímulos com o Zy é
dependente da presença de células tipo macrófagos e mastócitos residentes na
cavidade peritoneal.
Estas células são altamente secretórias e produzem mediadores
inflamatórios e anti-inflamatórios. O polissacarídeo Zy é reconhecido pelos
macrófagos através do receptor dectina-1, que é expresso predominantemente
em monócitos, macrófagos e neutrófilos e em uma população de células T do
baço. Ressalta-se também que o Zy é capaz de estimular a produção de
citocinas inflamatórias e pode ativar o sistema complemento na ausência de
imunoglobulinas. Além disso, esse polissacarídeo é capaz de induzir a
maturação de células dendríticas in vitro e estimular a produção de IL-2 por
essas células, indicando a ligação entre as respostas imunológica inata e
adaptativa (Underhill et al., 1999; Ribeiro et al., 2000; Granucci et al., 2003;
Mazur-Bialy et al., 2012; Mogilski et al., 2016).
Neste processo onde se tem LPS ou Zy como estímulos à resposta
imunologica frente à inflamação, as células de defesa do organismo são
ativadas em meio inflamatório, promovendo a liberação de citocinas, como
TNF-α, que acabam por induzir a morte de células tumorais combatendo a
inflamação (Meffert e Baltimore, 2005; Mogilski et al., 2016).
O TNF-α é uma citocina altamente pró-inflamatória que apresenta
propriedades benéficas ou injuriantes no processo inflamatório, constituindo-se
como principal mediador da resposta inflamatória aguda a bactérias gram-
negativas e outros patógenos. Essa citocina é produzida em macrófagos,
linfócitos, neutrófilos e células epiteliais e atua estimulando o recrutamento de
neutrófilos e monócitos para locais da infecção, tornando-os ativos para na

34
eliminação dos micro-organismos invasores (Sordillo et al., 1991; Abbas e
Lichtman, 2005; Bannerman, 2009).
Em ensaios biológicos o TNF-α é utilizado como sinalizador da inflanção.
A produção dessa citocina pode ser estimulada tanto in vivo quanto in vitro.
Quando testado in vivo, o TNF-α exerce atividade antitumoral e induz a síntese
de mediadores como prostaglandinas e glicocorticoides. Quando testado in
vitro, essa citocina estimula mitose de fibroblastos, células musculares lisas e
células do tipo B e T, indicando citotoxicidade, através da atividade
imunomodulatória anti-inflamatória, além de induzir a ação de genes envolvidos
com a inflamação, reparo tecidual, resposta imune e efeitos antitumorais
(Beyaert e Fiers, 1998; Esteves, 2017).
Com a ativação do TNF-α, provocada pelo quadro infeccioso, os
macrófagos e neutrófilos são estimulados a produzir óxido nítrico (NO), que
pode ser utilizado como outro marcador da resposta inflmatória. Ocorre que,
como efeito da inflamação, a molécula de NO combina-se com moléculas de
oxigênio e provoca a morte dos micro-organismos presentes nos macrófagos e
neutrófilos. Sendo assim, essa molécula citotóxica, atua como alerta do
organismo, promovendo uma resposta imune durante a infecção. O surgimento
de moléculas de NO é uma característica marcante nos processos infecciosos,
podendo está relacionado à patogenia de diversas doenças, tais como a
esclerose múltipla, o mal de parkinson, o alzheimer e o diabetes (Arturo et al.,
2010; Hart e Tapping, 2012; Salemme et al., 2016).
Apesar da descoberta de medicamentos eficazes do tratamento da
inflamação, existem os efeitos colaterais e a suspeita de efeitos
cardiovasculares potencialmente tóxicos, como foi o caso do medicamento
rofecoxib, retirado do mercado farmacêutico em setembro de 2004. Com isso,
novas opções terapêuticas no tratamento de inflamações são comumente
exploradas (Juni e Dieppe, 2004; Wannmacher, 2005).
Para contornar o quadro clínico de uma inflamação, tem-se fármacos
extensamente empregados, sendo estes os não-esteroidais (AINEs), os anti-
reumáticos (ARMDs), os glicocorticoides e os anti-inflamatórios estereoidais
(AIEs), que também estão sendo incluídos na condição de fármacos
imunossupressores devido à inibição da transcrição de citocinas iniciadoras da
inflamação, algumas quimiocinas e, a expressão das enzimas cicloxigenases.

35
Dessa forma, por exemplo, os AIEs atuam na diminuição do acúmulo e função
de células que participam das reações inflamatórias e imunes, como linfócitos,
monócitos, macrófagos, eosinófilos, neutrófilos, mastócitos e basófilos (Rang e
Dale, 2008; Balbino, 2011).
1.6.5 Toxidez aguda
A toxidez de uma planta medicinal ou de suas partes é um dos critérios
relevantes a serem avaliados em estudos de viabilidade na produção racional
de um medicamento. Essa avaliação tem como objetivo verificar os efeitos
tóxicos das substâncias químicas da planta num outro organismo vivo,
determinar a dosagem segura, bem como o tempo e a via de administração
(Klaassen et al., 2012; Ruppenthal, 2013). Ocorre que qualquer planta
medicinal apresenta algum grau de toxidez, contudo, esse fato não invalida o
seu uso em tratamento de enfermidades. O que distingue esses dois universos
(medicinal e tóxico), é a dose segura (Luand, 1976). Por isso, o estudo da
atividade toxicológica de uma planta ou substâncias químicas é relevante, a fim
de encontrar a dose segura da droga vegetal, o seu tempo e a via de
administração, a tornando candidata a um medicamento.
Em contato com o agente biológico, o agente químico com potêncial
tóxico, imediatamente inicia os sintomas de intoxicação que rapidamente evolui
para sinais visíveis que vão de simples irritação nos olhos e perda de pêlos a
sintomas mais severos como diarreia, vômitos, dificultade de locomoção e
óbito. A avaliação da dose com efetivo potêncial tóxico se da pelo valor da
dose letal (DL50), onde é verificada a quantidade de mortes de animais de uma
mesma espécie quando em exposição com a substância química (Brito, 1994;
Hayes, 1994; Ruppenthal, 2013).
A intoxicação pode ser do tipo aguda, quando decorre de um único
contato com a/as substâncias danosas em um período máximo de 24 horas
com efeitos que vão de imediato a no máximo duas semanas. Esse tipo de
estudo busca verificar órgãos alvo de intoxicação da substâcia química para
delinear os níveis de dose em estudos mais prolongados. Intoxicação do tipo
sobreaguda ou subcrônica, se da em exposições repetidas a substâncias
químicas em um período inferior ou igual a 1 mês para o tipo sobreaguda e 1 a
3 meses para o tipo subcrônica, com estudos efetivos nos órgãos alvo,

36
demarcando aqueles mais suceptiveis, além do que, esse periodo de 3 meses
de estudo permite delimitar a dose máxima na qual não observa-se efeito. Para
exposição prolongada, superior a 3 meses, classificada como intoxicação
crônica, pode-se prever a dose máxima livre de toxicidez, além dos efeitos
tóxicos a partir de exposição prolongada, e o mecanismo de ação da
substância química estudada (Fukushima, 2008; Ruppenthal, 2013).
Para estudos de toxidade com plantas medicinais deve-se considerar
não apenas a dose, mas também outros fatores que interferem na sua toxidez
como as propriedades físico-quimicas do extrato, local e período de coleta e
mesmo as condições de cultivo da planta que foi administrada, uma vez que a
química da planta é uma resposta ao ambiente externo e sua fenologia,
geralmente, trás diferenças marcantes em sua composição química. Soma-se a
estas questões a via de introdução dos extratos da planta, bem como a dose e
a concetração extrativa por dose, além da frequência de administração
(Fukushima, 2008; Silva, 2014).
A administração de uma droga pode se dar por via enteral ou parenteral,
a primeira refere-se às vias oral, sublingual, cutâneo e retal e a segunda refere-
se às vias diretas à corrente sanguínea. Nos organismos, estudos de
toxicologia são realizados principalmente por via gastrintestinal (ingestão),
pulmonar (inalação) e cutânea (contato), havendo também estudos realizados
por via endovenosa, intraperitoneal, subcultânea, intramuscular, intradérmica e
oral. A via a ser utilizada deve ser escolhida de acordo com a forma
farmaceutica (pomada, comprimido, injeção, supositório, xarope, etc) que irá se
estabelecer pela ação da droga no organismo, bem como de acordo com área
de toxicologia em estudo, por exemplo, estudos de toxicologia ambiental ou
ocupacional sugerem-se a via pulmonar ou cultânea (Lima, 2008; ANVISA,
2013; Montanha et al., 2013; Brito, s.d.).
1.7 A importância de estudos sobre Senna alata como medicinal
Senna alata age como planta pioneira em biomas muito adversos como
a caatinga e a restinga. Nesses ambientes, assim como outras plantas, é
utilizada tradicionalmente como medicinal pelas populações locais e apresenta

37
estudos iniciais referentes principalmente a sua química, com algumas
descrições de flavonoides e antraquinonas.
Apesar da utilização de Senna alata como medicinal, para que a mesma
venha a ser inserir no mercado de drogas vegetais e mesmo de produtos
farmacêuticos, os extratos devem ser avaliados nas diversas áreas como a
físico-química, fitoquímica, farmacologia e tecnologia farmacêutica, além de
produzir padrões em que sua utilização como fitoterápico seja otimizada. Por
outro lado, a quinta edição programa da agenda ambiental na administração
pública (A3P), lançada em 2009, afirma que 50 % do PIB (Produto Interno
Bruto) brasileiro dependem da biodiversidade encontrada no país. Este
indicativo aponta para a necessidade de se buscar formas eficazes de pensar
em desenvolvimento, preservando os recursos naturais, seja para produção de
medicamentos, melhoramento genético ou em outras questões (Scheffer, 1999;
Correia, 2015).
Sendo assim, este trabalho objetiva determinar padrões referentes aos
acessos de Senna alata nas regiões de restinga e caatinga, fundamentalmente
em controle botânico de qualidade e parâmetros físico-químico das folhas
pulverizadas ou em extratos obtidos a partir das suas folhas, bem como
explorar algumas atividades biológicas na tentativa de avaliar semelhanças e
diferenças entre os dois acessos citados. Com isso contribui-se com
informações substanciais sobre o potencial dessa planta em ser utilizada
futuramente como um possível anti-inflamatório de baixa toxidez ou mesmo
como antimicrobiano ou bio-herbicida.

38
2. OBJETIVOS
2.1 Objetivo Geral
Este trabalho tem como objetivo geral determinar as características
farmacobotânicas, físico-química, e investigar as atividades biológicas de
Senna alata (L.) Roxb. obtidas em acessos da caatinga (BA) e da restinga (RJ),
comparando possíveis semelhanças ou diferenças entre as mesmas.
2.2 Objetivos Específicos
Determinar características organolépticas, macro e microscópicas das
folhas de Senna alata como insumos farmacêuticos;
Determinar o teor extrativo a partir das folhas pulverizadas de Senna
alata;
Determinar parâmetros físico-quimicos das folhas pulverizadas ou em
extrato de Senna alata;
Determinar a atividade antibacteriana do extrato a 40 % de Senna alata
por bioautografia;
Determinar a atividade alelopática de Senna alata frente à Lactuca
sativa;
Determinar a atividade citotóxica dos extratos de Senna alata pela
técnica MTT;
Determinar a atividade anti-inflamatória de Senna alata in vitro induzida
por LPS e Zymosan;
Determinar a toxidez aguda de Senna alata na administração por via
intraperitoneal em camundongos.

39
3. MATERIAL E MÉTODOS
3.1 Material vegetal
O material vegetal deste trabalho constituiu-se de folhas de Senna alata
(L.) Roxb., coletado em patrimônio privado na zona rural da cidade de Mucugê,
BA, e nas proximidades do bairro Barreto na Cidade de Macaé, RJ. As coletas
foram realizadas no mês de agosto de 2015. O material foi identificado por Dra
Ana Paula Lima do Couto Santos, conforme exsicatas 186333 e 225664,
depositadas junto ao Herbário da Universidade Estadual de Feira de Santana –
HUEFS.
Parte do material vegetal foi desidratada, pulverizado em moinho de
facas modelo Modelo 340, Adamo® e armazenado em frasco âmbar para
posterior obtenção dos extratos. O material restante foi utilizado na
investigação macroscópica da folha íntegra.
3.2 Solventes, reagentes e substâncias referência utilizadas
Os solventes utilizados foram do grau de padrão analítico (P.A.) da
Tedia® ou Sigma-Aldrich®. Todas as soluções foram preparadas com água
destilada e purificada no sistema Milli Q. Plus®. O meio de cultura utilizada para
os microorganismos foi ágar Muller Hinton (HIMEDIA®). Todas as substâncias
referência utilizadas foram do tipo pró analise de procedência Merck®.
3.3 Micro-organismos
Para determinação da atividade antimicrobiana, utilizou-se bactérias
Gram + e Gram -, conforme tabela 1.
Tabela1 Micro organismos utilizados no ensaio de bioautografia.
3.4 Cultura de Macrofagos
Para esse estudo, foi utilizada a linhagem de macrófagos RAW 264.7 de
camundongos, obtida do laboratório no laboratório de investigação biológica da
Universidade de Taubaté (UNITAU). As células foram mantidas em nitrogênio
liquido (criopreservação) em solução de congelamento composta por meio
Micro organismos Coleção de Cultura
Staphylococcus aureus BAA-1026
Escherichia coli 33876

40
DMEM suplementado com 10 % de Dimetilsufóxido (DMSO) e 90 % de soro
bovino fetal (SBF). Para os ensaios de estimulação in vitro as células foram
descongelas a temperatura ambiente e expandida em dois frascos de cultura
celular em meio DMEM completo (DMEM-c), mantidos a temperatura de 37° C
em estufa incubadora umitificada contendo 95 % de ar e 5 % de CO2.
Após a formação da monocamada de células, o meio DMEM-c foi
descartado e adicionado novo meio (15 mL de DMEM-c). O meio contendo as
células foi retirado e pipetado em dois tubos esterilizados de 50 mL.
Centrifugou-se os frascos a 1500 rpm, durante 10 min, para separação das
células do meio. Após a centrifugação, o sobrenadante foi descartado e 10 mL
de DMEM-c foi adicionado em cada tubo contendo as células.
A contagem das células foi efetuada na câmara Neubauer, partindo-se
da suspensão de 1 mL diluída em 10 vezes em solução azul Trypan para
avaliação da viabilidade celular e para contagem total do número de
macrófagos. Para os ensaios, as células forma colocadas em meio de cultura
de 96 poços de acordo com a concetração exigida em cada experimento e
incubadas overnight em DMEM-c a 37° C, com 95 % de O2 e 5 % de CO2.
3.5 Animais
Camundongos Swiss machos adultos foram utilizados com 6 semanas
de vida, provenientes do biotério de criação do Campus UFRJ-Macaé. Os
camundongos foram mantidos no biotério de experimentação do LPBio/UFRJ
– Macaé em temperatura aproximada de 25° C e promoveu-se o ciclo de 12
horas-luz no biotério. Água filtrada e ração peletizada em livre acesso. O
experimento foi aprovado pela comissão de ética em uso de animais, sob
protocolo n° MAC042.
3.6 Análise morfo-anatômica das folhas de Senna alata
3.6.1 Análise Morfológica
A análise morfológica das folhas fresca e da droga foi efetuada a olho
nú, quando necessário fez-se uso de estereomicroscópio binocular (aumento
de 10 x), tomando por base para descrição macroscópica Hickey (1974),
Oliveira e Akisue (1991), Gattuso e Gattuso (1999).

41
3.6.2 Análise Anatômica
A analise anatômica foi realizada em secções paradermica. O estudo da
epiderme na face adaxial e abaxial dos folíolos foi obtido por lâminas
temporárias com secções a mão livre de material fresco, corados com azul de
toluidina a 0,05 % (p/v) segundo Oliveira e Akisue (1989), Gattuso e Gattuso
(1999) e Farmacopéia Brasileira V ed. (2010). O material foi observado em
microscópio binocular de campo claro e registrado em fotomicrografia obtidas
em câmara digital e analisados segundo Esau (1985), Cutter (1987), Gattuso e
Gattuso (1999), Dickison (2000), Appezzato-da-Glória e Carmello-Guerreiro
(2003).
3.6.2.1 Determinação do índice Estomático
A determinação do índice estomático foi realizada em secções
paradermica segundo descrito por Gattuso e Gattuso (1999). As observações
para a contagem dos estômatos foram feitas em microscópio com aumento de
10 X. Para cada amostra realizou-se 10 determinações e o índice estomático
se deu conforme a média obtida. O resultado foi calculado conforme a equação
abaixo:
Equação 1:
3.6.2.2 Análise das folhas pulverizadas
As características de cor, textura, odor e sabor foram analisados tomando
por base Oliveira e Akisue (2005) e Farmacopéia Brasileira V ed. (2010). As
características macroscópicas (forma, ângulos, bordos, coloração, superfície e
consistência das fraturas) da droga em pó foram analisadas em
esteriomicroscópio binocular tomando por base a técnica utilizada por Gattuso
e Gattuso (1999) e Oliveira e Akisue (2005). As características microscópicas
da droga em pó foram analisadas em lâminas semi-temporárias, com glicerina
I = S x 100
E + S
Em que:
I = Índice de estômatos S = Número de estômatos na superfície determinada da folha; E = Número total de células epidérmicas na mesma área (incluindo os
Tricomas que apareceram).

42
a 50 % (v/v) como meio de inclusão, utilizando como corante o azul de toluidina
a 0,05 % (v/v), tomando por base Gattuso e Gattuso (1999).
3.6.2.3 Análise de Granulometria
Para a análise granulométrica, seguiu-se a metodologia apresentada por
OMS (1992). Para a tamisação usou-se 100 gramas da droga submetida à
passagem forçada, realizada a 60 vibrações por segundo durante 30 minutos,
em uma sequencia de tamises (abertura de malhas correspondente a 70, 92,
165, 232 e 331 µm), de maior a menor abertura de malha. Em seguida, as
frações da droga em pó foram retiradas dos tamises, bem como do coletor do
tamisador, e calculadas as porcentagens de retenção em cada tamis. Para o
cálculo do resultado foi considerada a média das triplicatas expressas em
porcentagem (%, p/p). Os resultados foram expressos graficamente, sendo que
o diâmetro médio da malha do tamis foi obtido por regressão linear (Alberton e
Ribeiro, 2001) e a classificação da droga em pó seguiu Who (1992) e
Farmacopéia Brasileira V ed. (2010).
3.7 Determinação do teor de extrativos
De acordo com a metodologia descrita por Simões e Spetizer (2004), as
folhas desidratadas e pulverizadas, foram submetidas ao preparo do extrato
etanólico nas concentrações 20, 40, 60, 80 e 95 % (m/v), por maceração. Estes
extratos foram preparados a 10 %, ou seja, utilizou-se 10 g da droga para 100
mL de solvente nas diferentes concentrações de etanol descritas acima. Foi
adotada a extração pelo período de 4 horas com agitação constante a cada 30
minutos com bastão de vidro. Para evitar incidência de luz, as vidrarias foram
envolvidas em papel alumínio. O material foi filtrado por 3 vezes e levado para
secagem em estufa de ar circundante a 40° C. Ao término de 24 horas, obteve-
se o extrato bruto seco que foi quantificado e pesado em balança de alta
precisão, depositado em recipiente de vidro âmbar hermeticamente fechado e
armazenado a 4° C. O teor de extrativos foi calculado em massa percentual,
pela a média de três determinações segundo a equação 2:

43
Equação 2:
3.8 Análise físico-química das folhas pulverizadas de Senna alata
Os ensaios físico-quimicos referentes à pureza e integrudade da droga
em pó abrangeram a pesquisa de material estranho, umidade e cinzas totais e
insolúveis em ácido, de acordo com os critérios farmacopeicos e
recomendações da Organização Mundial de Saúde. Todos os ensaios foram
realizados em triplicata com os resultados obtidos pela média dos valores
obtidos nos ensaios (Who, 1998; Farmacopeia Brasileira, V ed.).
3.8.1 Determinação de material estranho
Com o auxilio de uma lupa (aumento de 10 X, as amostras foram
analisadas pelo método de quarteamento). Todo o material estranho foi
separado e pesado em balança eletrônica de precisão. O resultado foi
expresso em porcentagem de elementos estranhos, com base no peso da
tomada de ensaio.
3.8.2 Determinação da umidade
A determinação da umidade da droga em pó foi realizada pelo método
gravimétrico em que 3 g das folhas em pó foram dessecadas em estufa a 105º
C, por 5 horas, e após este tempo, foram colocadas em dessecador para
posterior pesagem. Continuou-se pesando o material, em intervalos de 1 hora,
até massa constante, calculando-se a porcentagem de água em relação a
amostra seca ao ambiente, utilizando a equação 3 abaixo:
Equação 3:
% umidade = Pu – Ps / Pa x 100
Em que: Pu = peso úmido (béquer + amostra) Ps = peso seco (béquer após 5 horas de estufa) Pa = peso da amostra
% Te = Ps x 100 / Pf
Em que: Te = Teor de extrativos (%; m/m) Ps = Peso inicial da amostra Pf = Peso final da amostra

44
3.8.3 Cinzas totais
Nesse ensaio, 3 g da folha desidratada e pulverizada foram colocadas
em cadinho de cálcio devidamente pesado. O material foi incinerado em mufla,
aumentando gradativamente a temperatura até 450º C, em seguida, foi
resfriado em dessecador e pesado. Para determinação das cinzas totais,
calculou-se a porcentagem das cinzas resultantes após o processo em relação
a amostra úmida inicial, utilizando a equação 4 abaixo.
Equação 4:
3.8.4 Cinzas insolúveis em ácido
O resíduo obtido no ensaio de cinzas totais (item 3.7.1.3) foi fervido
durante 5 minutos com 25 mL de solução de ácido clorídrico a 7 % (p/v), em
um cadinho coberto com vidro de relógio. O vidro de relógio foi lavado com 5
mL de água quente e as amostras (resíduo e lavagem do vidro de relógio)
foram filtradas em papel de filtro isento de cinzas. Em seguida, lavou-se o
papel de filtro com água quente até pH neutro e transferiu para o cadinho
original, cujo conteúdo foi seco sob chapa quente e incinerado em mufla a 500
°C, durante 4 h. O teor de cinzas insolúveis em ácido foi calculado como
percentual pela média de três determinações em relação.
3.9 Análise fitoquímica dos extratos das folhas de Senna alata
Para a determinação do teor de constituintes químicos utilizou-se as
metodologias empregadas por Namba (1988), OMS (1992), por Sharapin
(2000) e pela Farmacopéia Brasileira V ed. (2010). Todos os experimentos
foram analisados em triplicata.
% cinzas totais = Pu – Ps / Pa x 100
Em que: Pu= peso úmido (cadinho + amostra) Ps= peso seco (cadinho após a incineração) Pa= peso da amostra

45
3.9.1 Determinação do pH dos extratos
Para determinação do pH do extrato das folhas de S. alata foi realizado
a leitura em pHmetro calibrado em pH de 4 a 9. O experimento foi realizado em
triplicata e os resultados equivalem à média dessas medições.
3.9.2 Determinação de antraquinona
Para determinação de antraquinona, agitou-se 1 g da droga em pó com
10 mL de éter etílico. Na solução etérea adicionou-se 1 mL de amônia diluída e
agitou-se novamente. Esse constitui um teste qualitativo onde como efeito da
mistura ocorre a mudança para coloração rósea no caso positivo. Sendo
caracterizado pela presença ou ausência dessa substância.
3.9.3 Determinação de saponinas
Para a determinação de saponina, realizou-se teste quantitativo, onde
aqueceu-se 2 g do material vegetal pulverizado com 10 mL de água destilada
sob refluxo, durante 10 minutos. Esfriou-se e filtrou-se para um tubo de ensaio,
completando-se para 10 mL o volume final. Agitou-se o tubo no sentido do seu
comprimento, vedado com uma rolha, durante 15 segundos. Deixou-se em
repouso por 30 minutos. Verificou-se em seguida o aparecimento de um anel
de espuma persistente por 2 horas ao menos, de aproximadamente 1 cm de
altura no caso positivo, conforme equação 5.
Equação 5:
3.9.4 Determinação de saponinas hemolíticas
Para avaliação citotóxica de S. alata sobre eritrócito, utilizou-se o teste
de hemólise seguindo o protocolo descrito por Rangel e colaboradores, 1997.
Para isso, utilizou-se sangue de camundongo, cedido pela equipe de pesquisa
e experimentação Universidade Federal do Rio de Janeiro (UFRJ-IMCT,
Macaé). Assim, uma amostra do sangue (2 mL) foi misturado a NaCl 0,96 % na
proporção 1:30, e centrifugado a 2000 rpm por 3 vezes de 5 minutos para
IE= 1000 (1) A
Em que: IE= Indice de espuma A= Volume do decocto usado para preparação da diluição

46
obtenção dos eritrócitos. O sedimento da ultima centrifugação foi ressuspenso
em uma concentração final de 0,5 %. Às amostras extrativas foram adicionados
Tampão fosfato salino (PBS) pH 7,0 e procedeu-se a diluição seriada onde
obteve-se soluções com concentrações finais igual a 62,5, 125, 250, 500 e
1000 μg/mL, a elas foram adicionadas 2 mL da suspensão de eritrócitos. Como
controle negatigo utilizou-se suspensão de eritrocitos com NaCl a 0,96 %
(0 % de hemólise) e como controle positivo utilizou-se a suspensão de
eritrócitos mais 100 μg/mL de Triton X-100 a 1 % (100 % de hemólise). As
misturas foram incubadas sob agitação lenta e constante (100rpm) por 1 hora a
22 ± 2° C. Decorrido esse tempo, as amostras foram centrifugadas por 5
minutos a 2000 rpm e a hemólise foi quantificada por espectrofotometria a
540 nm. Considerou-se como resultado a média das triplicadas.
3.9.5 Determinação de flavonoides
Para determinação de flavonoides, utilizou-se método qualitativo
colorimétrico, onde foram dissolvidos 25 mg de extrato bruto seco em 5 mL de
metanol em um tubo de ensaio. Adicionou-se fragmentos de magnésio seguro
por uma pinça e 0,5 mL de ácido clorídrico (HCl) concentrado. Ao final foi
determinada a presença de flavonoides pela efervescência e o surgimento de
uma coloração avermelhada na mistura.
3.9.6 Determinação de mucilagem
Para a pesquisa de mucilagens mediu-se o volume ocupado por 1 g da
droga pulverizada, em proveta de 25 mL. Adicionou 25 mL de água (agente
intumescente) e agitou-se por 1 minuto a cada 10 minutos, por 1 hora. Como
efeito do experimento, após 3 horas, em temperatura ambiente, a diferença do
volume (mL) inicial e final ocupado pelo material vegetal, constitui o índice de
intumescência da amostra calculado a partir da equação 6:
Equação 6
IT= Vi –Vf Em que: Vi= Volume inicial ocupado pelo material vegetal Vf= Volume final ocupado pelo material vegetal

47
3.9.7 Determinação de taninos
Para a determinação de taninos, utilizou-se de método qualitativo. Para
isso, dissolveu-se 5 mg do extrato etanólico a 40 % em 5 mL de água destilada,
e posteriormente este material foi filtrado. Adicionou-se 2 gotas de solução
etanólica de FeCl3 a 1 % (v/v). Qualquer mudança na coloração ou formação
de precipitado é indicativo de reação positiva para taninos.
3.9.8 Determinação do teor de polifenois totais
O teor de Polifenois totais dos extratos a 40 % (v/v) foi determinado com o
reagente Folin-Ciocalteau (uma mistura de fosfomolibdato de amônio e ácido
fosfotungstato), descrito por Chandra e Meija (2004).
Neste ensaio, as amostras de extrato bruto seco a 40 % (v/v) obtidos a
partir das folhas de S. alata em acessos na restinga e caatinga, foram diluídas
em água, adicionando-se uma mesma alíquota de reativo de folin-Ciocalteau,
levando-se a incubação no escuro por 5 minutos, quando acrescentou-se igual
volume de carbonato de sódio a 20 % (v/v). Após incubação de 10 minutos,
mediu-se a absorvância das amostras a 760 nm em um espectrofotômetro
UV/visível.
Como branco substituiu-se a amostra por água. A curva padrão foi
obtida utilizando-se soluções referência de ácido gálico nas concentrações de
12,5, 25, 50, 100 e 200 µg/mL.
A partir dos resultados da absorvância e das concentrações de ácido
gálico dos extratos, foi construído um gráfico da equação da reta por regressão
linear para calcular o teor de polifenois totais do analito.
3.9.9 Determinação do teor de flavonoides totais
A dosagem total de flavonoides foi realizada de acordo com Rio (1996) e
Markhan (1982), a partir da construção de uma curva de calibração, utilizando
rutina como substância referência.
As amostras de extrato bruto seco a 40 % (v/v) obtidos a partir das
folhas de Senna alata obtidos em acessos na restinga e caatinga foram
diluídas em metanol a 70 % (p/v), adicionou-se 7,5 % (m) de cloreto de
alumínio a 5 % em metanol (v) e, após repouso de 30 minutos no escuro,
realizou-se a leitura a 425 nm em espectrofotômetro UV/visível.

48
Como branco substituiu-se a amostra por água. A curva padrão foi
obtida utilizando-se a solução referência rutina nas concentrações de 12,5, 25,
50, 100 e 200 µg/mL.
A partir dos resultados da absorvância das concentrações de rutina dos
extratos, foi construído um gráfico da equação da reta por regressão linear para
calcular o teor de flavonoides totais.
3.9.10 Perfil cromatográfico dos extratos em cromatografia de
camada delgada (CCD)
Para obtenção do perfil cromatográfico os extratos obtidos a partir das
folhas de S. alata foram cromatografados em CCD, sobre cromatofolhas de
sílica gel G 60 F254, empregando-se sistemas eluentes e reveladores segundo
Wagner e Bladt (1996). Com auxilio de tubos capilares de vidro, os extratos
foram aplicados nas cromatoplacas, formando manchas semelhantes e
regulares. A fase móvel escolhida neste estudo foi a de uma mistura de hexano
e acetato de etila (1:1). A migração cromatográfica foi de 5 cm. Após eluição
utilizou-se vanilina sulfúrica em estufa a 100º C por 5 minutos como agente
homogênico. As placas foram observadas sob luz ultravioleta, sendo
registrados os resultados em Fator de retenção (Rfs). Utilizou-se rutina e
quercetina como substâncias referência.
3.10 Determinação de atividades biológicas dos extratos das folhas de
Senna alata
3.10.1 Determinação da atividade antibacteriana
A atividade antimicrobiana do extrato hidroetanólico de S. alata foi
determinada pelo método de bioautografia conforme Holetz (2002). Utilizaram-
se bactérias do tipo Gram-positivas (Staphylococcus aureus) e Gram-negativas
(Escherichia coli), conforme tabela 1. Os extratos foram cromatografados (item
3.9.10) e após a migração dos extratos aplicou-se nas placas cromatográfica o
padrão antibiótico. A placa cromatográfica foi depositada em placa de Petri
estéril em câmara de fluxo laminar. A suspensão padrão dos micro-organismos
a 1,5 x 108 UFC/mL foi inoculada sobre a mesma com o auxílio do swab em
seguida foram mantidas em estufa a 35° C por 24 h. Passado o período de
incubação o bioautograma foi revelado por nebulização de solução aquosa de

49
trifenil-2,3,5 cloreto de tetrazóleo a 2 % e novamente levado a estufa a 35º C
por 1 h. A presença de substâncias químicas ativas foi caracterizada pela
visualização de halo de inibição em torno das bandas cromatográficas. O
experimento foi realizado em triplicata e registrado por fotografia digital. Como
padrão de antibiótico, utilizou-se amoxicilina tri-hidratada (875 µg/mL).
3.10.2 Determinação da atividade alelopática
A atividade alelopática das amostras foi determinada por bioensaios de
inibição da germinação de diásporos de Lactuca sativa L. var. Grand rapids, em
solução aquosa com extrato de S. alata a 0,5, a 1,0 e a 2,0 % (v/v), à
temperatura ambiente de 25° C ± 2º C. Em cada experimento foram semeadas
20 diásporos por placa de Petri (9 mm de diâmetro) sobre camada tripla de
papel filtro umedecido com 5 mL da solução análise. A contagem dos diásporos
que germinaram deu-se em intervalos de 3, 6, 9, 12, 15, 18, 24 e 48 horas após
a distribuição das sementes, baseando-se na emissão de radícula com cerca
de 2 mm além do estado da plântula (cor, tamanho, consistência e espessura)
observadas sob lupa com 10 x de aumento (Hartmann et al., 2001; Souza et
al., 2005; Ferreira et al., 2007; Gusman et al., 2008). Como controle positivo,
utilizou-se água destilada. Os resultados foram representados graficamente
com a média das triplicatas.
Determinou-se a viabilidade dos diásporos não germinados pelo
desenvolvimento de coloração rósea na parte interna dos mesmos expostos
por secção transversal. Para tanto, as sementes passaram 24 h no escuro e à
temperatura de 30° C, em solução aquosa de cloro-2,3,5-trifenil cloreto de
tetrazólio a 1 % (v/v).
3.10.3 Viabilidade celular pelo MTT
O teste de citotoxidez in vitro foi efetuado utilizando o ensaio de
viabilidade celular através do MTT, conforme NIH (2001). Os sobrenadantes
das culturas celulares de macrófagos RAW 264.7 foram obtidos em cultura
destas células na concentração de 5 x 104 células/mL, tratadas com os extratos
de S. alata nas concentrações 50 e 100 µg/mL. Após 24 horas, adicionou-se o
MTT (5 mg/mL) em PBS durante 4 horas a 37° C. Após essa incubação, a
placa foi centrifugada a 1800 rpm por 5 minutos. Retirou-se o sobrenadante e
adicionou-se à placa 200 µL de DMSO. A absorvância foi medida em

50
espectrofotômetro de placas de cultura (SLT Spectra®) a um comprimento de
onda de 490 nm. Como controle branco, utilizou-se 200 µL de DMSO. Os
valores da viabilidade celular foram expressos em porcentagem média
conforme a absorbância determinada nos grupos estudados.
A liberação específica foi calculada de acordo com os controles:
macrófagos não-tratados e estimulados (controle negativo), e macrófagos
tratados com 1 % (v/v) de Triton X-100 (controle positivo).
3.10.4 Avaliação da atividade anti-inflamatória in vitro
Os ensaios foram realizados no laboratório de produtos bioativos (LPBio)
no Campus Macaé UFRJ seguindo a metodologia descrita por Mori e
colaboradores (2000) e Zhao e colaboradores (2008).
3.10.4.1 Estímulo com LPS
Em uma placa de 96 poços foi depositado 100 µL de macrofagos da
linhagem RAW 264.7 na quantidade de 2 x 105 células/mL. Após duas horas na
estufa a (37° C, 5 % de CO2), a placa foi centrifugada a 2000 rpm por 10
minutos a 10° C retirou-se o sobrenadante e aplicou-se os estímulos de LPS na
concentração de 1 µg/mL e os extratos etanolicos a 40 % (v/v) de S. alata
acesso caatinga e restinga retomado em água nas concentrações 25, 50 e 100
µg/mL (1:1) v/v. Após 24 horas em estufa, o sobrenadante foi retirado e
estocado à -80° C para dosagem de TNF-α e NO.
3.10.4.1.1 Avaliação da capacidade inibitória de Senna alata sobre a
produção de TNF-α
Para quantificação de TNF-α, os sobrenadantes dos macrofagos RAW
264.7 tratados com LPS conforme item 3.9.4.1, foram desgelados em
temperatura ambiente para a dosagem do TNF-α por kit de ELISA, de acordo
com as especificações do fabricante (ReD Systems Quantikine). Como controle
negativo utilizou-se sobrenadante da cultura de macrófagos não tratados com
extratos de Senna alata e não estimulados com LPS e como controle positivo
utilizou-se macrófagos estimulados com LPS (1 µg/mL) e não tratados com os
extratos de Senna alata. A leitura foi realizada em espectofotômetro UV a 450
nm.

51
3.10.4.1.2 Avaliação da capacidade inibitória de Senna alata sobre a
produção de óxido nítrico
Para quantificação de nitrito, os sobrenadantes dos macrofagos RAW
264.7 tratados com LPS conforme item 3.10.4.1, foram desgelados em
temperatura ambiente. Como controle negativo utilizou-se sobrenadante da
cultura de macrófagos não tratados com extratos de Senna alata e não
estimulados com LPS e como controle positivo utilizou-se macrófagos
estimulados com LPS (1 µg/mL) e não tratados com os extratos de Senna
alata. A dosagem de NO foi realizada pelo método de Griess. A leitura foi
realizada em espectrofotômero a 540 nm. Os resultados foram expressos
graficamente.
3.10.4.2 Estímulo com Zymosan
A indução de inflamação a partir do polissacarídeo Zy, foi realizada
conforme Rocha (2003). Em uma placa de 96 poços foi depositado 100 µL de
suspensão de células peritoneais de camundongos Swiss por poço, na
quantidade de 2 x 105 células. Após duas horas na estufa a 37° C e 5 % de CO2
para adesão dos macrófagos na placa, a mesma foi centrifugada a 2000 rpm
por 10 min a 10° C. Foi retirado o sobrenadante e adicionado os estímulos de
Zy na concentração de 1 µg/mL e os extratos etanolicos a 40 % (v/v) de S.
alata acesso caatinga e restinga retomado em água nas concentrações de 25,
50 e 100 µg/mL, (1:1) v/v. Após esse procedimento, levou-se o material
novamente à estufa para incubação por 24 horas, quando foi retirado o
sobrenadante para as dosagens de TNF e NO, conforme os itens 3.10.4.1.1 e
3.10.4.1.2.
3.10.5 Determinação da toxidez intraperitoneal aguda
O ensaio de toxidez foi realizado pela metodologia de Brito (1994).
Utilizou-se 45 camundongos Swiss adultos (45 ± 2 dias de vida). Foram
utilizados 1 grupo controle negativo (recebendo apenas solução salina), e 4
grupos teste tratados com o extrato de Senna alata obtidas do acesso caatinga,
4 grupos teste tratados com o extrato de Senna alata obtidas do acesso
restinga, em diferentes concentrações. O extrato etanólico de S. alata a 40 %
(v/v) seco foi dissolvido em PBS e os grupos testados receberam os
respectivos extratos nas doses 16, 32, 40 e 60 g/kg, por via intraperitoneal.

52
Realizou-se a observação ininterrupta nas primeiras 2 horas e em seguida em
4, 6, 8, 12, 24, 32 e 48 horas. Os parâmetros de toxidez avaliados e registrados
foram: sinais de depressão, excitação, convulsão, salivação, lacrimejamento,
efeitos sobre a defecação, micção, respiração, locomoção e óbito dos animais,
além da concentração do LDH.
3.10.5.1 Quantificação da liberação de enzima citoplasmática lactato
desidrogenase (LDH) em animais submetidos à toxidez aguda
O sangue dos animais que foram submetidos ao teste de toxidez aguda
nas várias concentrações de extrato de S. alata dos acessos caatinga e
restinga foi coletado por punção cardíaca logo após a eutanásia dos animais
utilizados no ensaio 3.10.5. O soro do sangue foi separado por centrifugação a
2000 rpm em 10 minutos e utilizou-se o kit ezimático Doles® para a
quantificação do LDH. Adicionou-se ao soro 250 µL de alúmen férrico em 4 mL
de substrato do kit enzimático (solução de 0,1 M de lactato e 0,05 M de o-
fenantrolina em solução tampão tris 0,2 M, pH 8,8), mantendo-se em estufa a
37° C por 2 minutos. Em seguida foi adicionado fenazina metosulfato (FMS) e
difosfopiridino nucleotído (NAD) na proporção de 1:20, mantendo-se em estufa
por mais 5 minutos, para então determinar a concentração de LDH, que
representa uma indicação indireta de citotoxicidade na absorvância de 510 nm.
Os reagentes foram manipulados na ausência de luz. Como controle positivo
utilizou-se amostra de soro sanguíneo em DMSO a 1 % na proporção 1:1 (v/v).
Como controle negativo utilizou-se o soro sanguíneo dos animais.
3.11 Análise estatística dos resultados
Neste trabalho todos os resultados são apresentados como médias ±
erro padrão da média (SEM) do número de experiências (n) indicado nas
figuras ou nas suas legendas. A significância das diferenças entre dois grupos
de valores foi calculada pelo teste t de Student não emparelhado. Quando um
grupo de valores foi comparado com três ou mais grupos, a significância das
diferenças foi analisada pelo teste ANOVA seguido do teste de Bonferroni. Os
valores de p inferiores a 0,05 (p < 0,05) foram considerados como
representando diferenças significativas. Para construção dos gráficos, utilizou-

53
se o software GraphPad Prism 7.0. Todos os experimentos foram realizados
em triplicatas (n = 3), sendo as médias consideradas como resultados.
4. RESULTADOS E DISCUSSÃO
4.1 Morfo-anatomia e caraterização organoléptica das folhas de Senna
alata
4.1.1 Análise macroscópica
Uma das atribuições da Farmacobotânica é realizar a analise morfo-
anatômica de drogas vegetais, visando a sua autenticidade e padronização de
um material vegetal. Neste trabalho realizou-se um estudo para enfatizar as
características morfoanatômicas marcantes nas folhas de S. alata que sejam
úteis na autenticidade das mesmas como insumo farmacêutico.
S. alata é um subarbusto contituido por pares de folíolos. Ressalta-se
que as características macroscópicas dessa espécie nos acessos restinga e
caatinga são em geral muito semelhantes para estes dois acessos,
diferenciando-se, principalmente, pela quantidade e tamanho dos folíolos, que
apresentam-se maiores e em maior quantidade no acesso caatinga, além
disso, diferenciam-se pela presença de um maior número de tricomas no
acesso caatinga, onde ocorre aglomeração de tricomas na face adaxial. Essas
características demarcam as diferenças mais consistentes entre os acessos,
conforme pode ser verificado na figura 5 e tabela 2.
.
Figura 5 Aspecto geral das folhas de Senna alata (L.) Roxb. Legenda: A. face adaxial e abaxial do acesso Caatinga; B. face adaxial e abaxial do acesso restinga. Regua: 1 cm.

54
Tabela 2 Determinação de características macroscópicas das folhas de Senna alata (L.) Roxb.
O padrão de venação em uma folha também é uma característica
consistente na descrição das espécies e drogas de origem vegetal. Tomando
por base Hickey (1974), nos dois acessos (restinga e caatinga) de S. alata,
pode-se observar que a nervação apresenta o padrão geral do tipo
camptódroma, com a nervura central proeminente apenas na face adaxial. A
venação terciária é reticulada. A última marginal é incompleta. A areolação é
desenvolvida, geralmente quadrangular (por vezes pentagonal) e as vênulas
são ramificadas, figura 6 e 7.
ESTRUTURAS
PARTES CONSTINTUINTES
CARACTERÍSTICA
Limbo
Tamanho
CAATINGA RESTINGA
Possui 10-12 cm de comprimento e 3-5 cm de
largura
Possui 5-7,5 cm de comprimento e 2,3-4,4 cm
de largura
Coloração Discolores, na face adaxial
Verde-oliva opaco =
Nervação Camptódroma =
Consistência Membranácea =
Superfície
Lisa, com tricomas, esparços na face adaxial e aglomerados na face abaxial próximo a nervura
central
Lisa, com tricomas esparsos na face adaxial e
face abaxial
Forma Oblongas =
Bordo Inteiro =
Ápice Retuso =
Base Assimétrica =
Divisão do Limbo Folha penaticomposta bifoliada do tipo paripenada, constituíd a
por 7 a 13 pares de folíolos.
Folha penaticomposta bifoliada do tipo
paripenada, constituída por 5 a 10 pares de folíolos.
Estípulas Triangulares, persistentes,
assimétricas com 1,4-1,6 x 0,7-1,1 cm
Triangulares, persistentes, assimétricas com 1,2-1,6 x
0,4-0,9 cm
Pecíolo Tamanho Reto e curto com 0,4 a 1,2 cm
de comprimento =
Figura 6 Venação da folha de Senna alata (L.) Roxb. acesso caatinga. Legenda: P. Detalhes da venação primária; S. Detalhe da venação secundária; T. Detalhe da venação Terciária. Régua: 1 mm.

55
Figura 7 Venação da folha de Senna alata (L.) Roxb. acesso restinga. Legenda: P. Detalhes da venação primária; S. Detalhe da venação secundária; T. Detalhe da venação Terciária . Régua: 1 mm.
Macroscopicamente, a folha de S. alata segue o padrão geral da
subfamília Caesalpinoideae apresentando-se composta por folíolos de bordo
inteiro e com a venação principal camptódroma, conforme Metcalfe e Chalk
(1950).
As características macroscópicas de S. alata diferem de S. alexandrina,
conforme a Farmacopéia Brasileira V Ed. (2010), basicamente na forma dos
folíolos e ápice, que para esta espécie é elíptica e aguda, respectivamente. O
padrão geral de venação, assim como a assimetria da base do limbo nos
folíolos também são o mesmo para estas duas espécies. S. alexandrina
também apresenta tricomas esparsos, mas neste caso estes tricomas possuem
textura áspera, o que não foi observada em S. alata.
Os folíolos dos acessos, quando desidratados, apresentam cor verde-
mate, podendo se verificar além das características descritas anteriormente,
ondulações marcantes no limbo que as tornam suavemente côncavas, como
podem ser observadas na figura 8.
Figura 8 Aspecto geral das folhas desidratadas de Senna alata (L.) Roxb. Legenda: A. face abaxial e
adaxial dos folíolos do acesso caatinga; B.face abaxial e adaxial do folíolo do acesso restinga. Régua: 1 cm.

56
O padrão que os folíolos de S. alata assumem quando desidratados
difere do padrão da droga de S. alexandrina, uma vez que nesta planta os
folíolos permanecem planos, praticamente sem dobraduras, e a coloração
também difere uma vez que em S. alexandrina assume um tom de verde-
amarelado, conforme Oliveira e Akisue (2005).
4.1.2 Análise microscópica
Em secção paradérmica os acessos restinga e caatinga de S. alata são
semelhantes (Tabela 3 e figuras 9 e 10). Pôde-se verificar as células
epidérmicas ondeadas com as paredes espessas tanto na face adaxial quanto
na face abaxial. Os estômatos se fazem presentes em ambas as faces
epidérmicas, demarcando o padrão anfiestomático. Estes estômatos são do
tipo paracítico e muito raramente anisocítico, com índice estomático maior na
face abaxial. Observou-se tricomas tectores unicelulares pouco alongado e
eretos, com o ápice afilado.
Tabela 3 Determinação das características microscópicas das folhas de Senna alata (L.) Roxb.
em secção paradérmica nos acessos restinga e caatinga.
ACESSO
ESTRUTURA
SUPERFICIE DO LIMBO
Adaxial Abaxial
CAATINGA
Estômatos Paracítico =
Tricomas tectores Unicelular de ápice afilado e cutícula espessa e lisa
=
Células epidérmicas Ondeadas =
Índice estomático 43,0 ± 3 73,49 ± 2
RESTINGA
Estômatos Paracítico =
Tricomas tectores Mais esparsos. Unicelular de ápice afilado e cutícula espessa e lisa
=
Células epidérmicas Ondeadas =
Índice estomático 44,13 ± 2 71,02 ± 2
Figura 9 Secção paradérmica da lamina foliar de Senna alata (L.) Roxb. acesso caatinga. Legenda: A.
Epiderme na face abaxial; B. Epiderme na face adaxial; C. Detalhe evidenciando tricoma tector unicelular na face adaxial; 1. Células epidérmica; 2. Estômato; 3. Tricoma tector; Régua: 20 μm. Aumento de 40 x.
1
2 3

57
Notou-se a presença de tricoma na face adaxial e abaxial de Senna
alata nos acessos estudados, no entanto, na superfície adaxial esses tricomas
apresentam-se débeis, podendo-se perceber a inserção do mesmo na
epiderme em sua extremidade inferior, como um ponto preto. A presença de
tricomas tectores é uma característica microscópica semelhante a S.
alexandrina, a presença de tricomas tectores, conforme a Farmacopéia
Brasileira V ed. (2010). No entanto, essses tricomas se diferenciam nas duas
espécies em relação a sua forma, pois, em S. alexandrina, os tricomas se
apresentam curvos e ásperos, o que não foi observado em S. alata Luna
(2013) relata que em Senna occidentallis, os tricomas se limitam a face abaxial.
As células epidérmicas são diferentes entre estas duas espécies
vegetais, uma vez que em S. alexandrina estas células apresentam a forma
poligonal e em S. alata são ondeadas. Esses dados mostram que apesar da
similaridade das espécies, que são facilmente confundidas no uso
populacional, existem diferenças microscópicas que auxiliam a diferenciação
da espécie.
Solereder (1908), Metcalfe e Chalk (1950) e Watson (1981) comentam
que o tipo mais comum de estômato na subfamília Caesalpinoideae é o
paracítico, como foi verificado neste trabalho para S. alata, dados que
corroboram com os estudos realizados com S. alexandrina. Contudo, Metcalfe
e Chalk (1950) mencionam que muitos gêneros desta subfamília, em especial
Senna, apresentam estômatos confinados à superfície inferior, obedecendo ao
padrão denominado hipoestomático. S. alata não segue este padrão usual para
Caesalpinoideae e sim o padrão anfiestomático, fato observado nos dois
Figure 10 Secção paradérmica da lamina foliar de Senna alata (L.) Roxb. acesso restinga. Legenda: A.
Epiderme na face abaxial; B. Epiderme na face adaxial; C. Detalhe evidenciando tricoma tector unicelular na face adaxial; 1. Células epidérmica; 2.Estômatos; 3. Tricoma tector. Régua: 20 μm. Aumento de 40 x.
1
2 3

58
acessos estudados de S. alata, o que demarca uma singularidade relevante
para esta espécie vegetal.
Em relação ao índice estomático sabe-se que o mesmo é um caracter
anatômico constante para as espécies vegetais. Esta questão é relevante uma
vez que pode ser utilizado na autenticação de uma droga vegetal, uma vez que
não é o índice e sim o desempenho dos estômatos que vai variar com a
influência da luz, da umidade do solo e do ar. Segundo Larcher (2006) e
Tatagiba e colaboradores (2007) essas características ambientais provocam
um deficit de pressão de vapor próximo a lâmina foliar, influenciando na
abertura do estomato e consequentemente na função dessas estruturas.
A partir dos estudos realizados por Larcher (2006) e Tatagiba e
colaboradores (2007), pode-se entender que a característica anfietomática de
S. alata, é uma das justificativas do potencial adaptativo e pioneirismo dessa
espécie vegetal que consegue apresentar alto desenvolvimento mesmo em
condições relevantes de salinidade e intenso calor. Por outro lado, levando em
conta Marabesi (2007), pode-se indicar que o rápido desenvolvimento de S.
alata tem relação com a alta taxa de fotossíntese alinhada a condutância
estomática em nível foliar, fato que abastece a planta de amido e assegura seu
desenvolvimento. Outras carcaterísticas foliares descritas para essa S. alata
sugere mecanismo de adaptação a ambientes com excesso de calor,
favorecendo a menor reflexão de luz e proteção dos estomatos, a exemplo da
epiderme apresentar parede celular expessa, característica adaptativa
ressaltada por Monteiro e colaboradores (1985) e Fahn (1990).
Em relação aos tricomas salienta-se que são apêndices da epiderme
estão presentes em diversos órgãos das plantas, segundo Fahn (1990). Os
tricomas tectores que são os de cobertura, como os que foram observados nas
folhas de S. alata são úteis para afastar ou fazer chamariz de alguns
animáculos e ainda por promover um microclima na camada epidérmica,
evitando a transpiração excessiva da planta. Assim é que essas estruturas,
além de proteção contra herbívoros, aumentam a superfície de absorção de
água da planta, o que é uma característica marcante de espécies da caatinga
(Henrique, 2008). Essa informação referente a presença de tricomas nas
espécies da caatinga corrobora ao fato de ter-se encontrado um maior número

59
de tricomas tectores nas folhas do acesso caatinga em relação ao acesso
restinga, como pode ser verificado na tabela 2.
Ressalta-se que os espécimes estudados foram retirados de biomas de
clima tropical, e assim é que caatinga e restinga, por apresentarem diferenças
diferenças climática marcantes ditarão caracteristicas morfo-anatômicas
adaptativas as mesmas variantes nas espécies locais. Contudo, salienta-se
que S. alata é uma espécie introduzida nestes biomas, logo suas
características morfo-anatômicas gerais serão preservadas, mas por adaptação
local, seus tecidos e órgãos estarão adaptados em algumas questões como
número de tricomas e espessamento cuticular. Com média de temperatura
anual em nos ultimos 3 anos entre 29,8 ± 4,2° C, a caatinga apresentou nos
meses mais secos (entre junho e agosto) a temperatura do solo com média de
60º C ± 10 º C durante todo o ano. Além dessas condições climáticas rigorosas,
a umidade desse bioma apresentou valores entre 45 % e 60 % com ventos
fortes e secos. Essa característica, associada aos demais elementos
climáticos, determinam a aridez dessa paisagem. Na restinga a temperatura
anual encontra-se entre 24 ± 4° C, com meses mais secos também entre Junho
e Agosto. A temperatura do solo desse bioma, variou entre 45 e 50º C, com
umidade relativa do ar entre 60 e 80 %. Outro fator que também difere nesses
biomas é a salinidade presente na restiga. Essas caracteristicas ambientais
salientadas acabam por promover a retenção de água pela planta
resguardando as funções fotossinteticas e conservando a homeostase iônica
(Esteves, 2008; EMBRAPA, 2012; Clima tempo, 2017; Climate, 2017).
4.1.3 Análise macro-microscópica e organoléptica das folhas em pó
O pó das folhas de S. alata Ca e Re apresentaram características
macroscópicas e organolépticas similares entre si, salientando-se que o pó é
homogêneo, as fraturas são predominantemente ovaladas com os bordos
sinuosos e reentrâncias suaves. A característica que varia é o sabor, pois o
acesso da caatinga apresentou-se marcadamente amargo enquanto que o
acesso da restinga é levemente amargo (tabela 4).
Microscopicamente os dois acessos (restinga e caatinga) também
apresentaram-se semelhantes, com estruturas que seguem o padrão já

60
descrito neste trabalho, ressaltando-se a presença dos tricomas tectores
unicelulares de ápice afilado, fragmentos de epiderme com as células
epidérmicas ondeadas e paredes celulares expessas, bem como os estômatos
do tipo paracíticos.
Tabela 4 Descrição macroscópica e organoléptica das folhas pulverizadas de Senna alata (L.)
Roxb.
CAATINGA
Tamanho Fratura
Coloração Forma Borda
Pó fino A maioria com forma ovalada
Suaves reentrâncias
Homogênea em tom verde-mate
RESTINGA Pó fino A maioria com forma ovalada
Suaves reentrâncias
Homogênea em tom de verde-mate
CAATINGA
Sabor Odor Superfície Consistência
Marcadamente amargo
Suave, Suigeneres
Lisa Membranácea
RESTINGA Levemente
amargo Suave,
Suigeneres Lisa Membranácea
Tomando-se por base Oliveira e Akisue (2005) e a Farmacopeia
Brasileira V Ed. (2010) e Oliveira e colaboradores (2012), os folíolos
pulverizados de S. alata em relação a S. alexandrina diferem basicamente no
que se refere a cor do pó, pois S. alexandrina possui o pó dos folíolos na cor
verde-amarelado, enquanto que S. alata possui o pó dos folíolos na cor verde-
mate. O pó dos folíolos destas duas espécies em questão é similar no sabor
amargo, contudo a diferença encontrada no sabor (mais ou menos amargo)
para os folíolos em pó de S. alata, entre os dois acessos, deve estar
relacionada à constituição química desses acessos em estudo pela variação de
classes químicas ou mesmo devido a maior ou menor proporção de
metabólitos das mesmas classes químicas.
. A forma das céluas epidérmicas e a cutícula do tricoma tector
apresentam-se com diferenças microscópicas marcantes no pó de S.
alexandrina e S. alata. Em S. alexandrina a forma das células epidérmicas é
poligonal e a cutícula do tricoma tector é rugosa, enquanto que na análise
microscópica do pó de S. alata neste trabalho pode-se verificar que as células
epidérmicas possuem forma ondeada e os tricomas tectores apresentam
cutícula expessa, mas sem rugosidades. Por fim há de se diferenciar também
pelo fato de o tricoma em S. alata ser reto e em S. alexandrina ser inclinado.
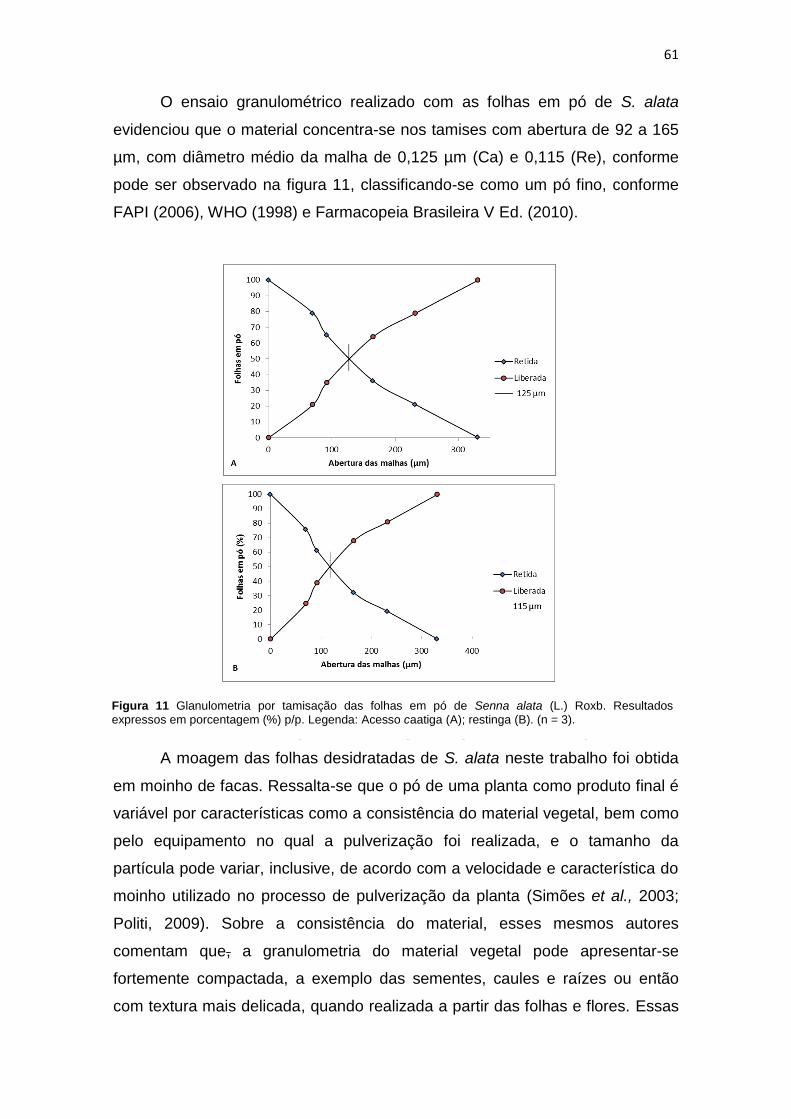
61
O ensaio granulométrico realizado com as folhas em pó de S. alata
evidenciou que o material concentra-se nos tamises com abertura de 92 a 165
µm, com diâmetro médio da malha de 0,125 µm (Ca) e 0,115 (Re), conforme
pode ser observado na figura 11, classificando-se como um pó fino, conforme
FAPI (2006), WHO (1998) e Farmacopeia Brasileira V Ed. (2010).
As estruturas histológicas de dos órgãos vegetais são heterogêneos.
A moagem das folhas desidratadas de S. alata neste trabalho foi obtida
em moinho de facas. Ressalta-se que o pó de uma planta como produto final é
variável por características como a consistência do material vegetal, bem como
pelo equipamento no qual a pulverização foi realizada, e o tamanho da
partícula pode variar, inclusive, de acordo com a velocidade e característica do
moinho utilizado no processo de pulverização da planta (Simões et al., 2003;
Politi, 2009). Sobre a consistência do material, esses mesmos autores
comentam que, a granulometria do material vegetal pode apresentar-se
fortemente compactada, a exemplo das sementes, caules e raízes ou então
com textura mais delicada, quando realizada a partir das folhas e flores. Essas
Figura 11 Glanulometria por tamisação das folhas em pó de Senna alata (L.) Roxb. Resultados expressos em porcentagem (%) p/p. Legenda: Acesso caatiga (A); restinga (B). (n = 3).

62
características influenciam diretamente na eficiência da extração. O resultado
da granulometria obtido neste trabalho para os acessos de S. alata,
apresentou-se como sendo um pó fino, contudo para S. alexandrina, Almeida
(2005) e Severo (2013) obtiveram pó das folhas classificado como grosso,
enquanto que Luna (2013), para S. occidentalis obteve pó das folhas de
classificado como médio. Ressalta-se que as duas espécies vegetais citadas
apresentam consistência membranácea, e também, no processo, foi utilizado
moinho de facas para a moagem, em todos os trabalhos citados. Trata-se de
três espécies de plantas do mesmo gênero e, possivelmente, a velocidade
utilizada ou mesmo o tamis de recolhimento do pó e, mesmo o tempo de
secagem da planta antes da pulverização pode ter sido fatores determinantes
para esses resultados.
Também deve-se levar em consideração de que o material de S.
alexandrina, em geral, apresenta uma quantidade considerável de frutos junto
as folhas, conforme Farmacopéia Brasileira V ed. (2010), o que deve ser
determinante para os resultados destes trabalhos serem classificados com
granulometria diferente.
4.2 Análise físico-química
Os ensaios de pureza são essenciais para comprovar a integridade da
droga em pó e os interferentes que podem ocorrer devido o cultivo e manuseio
no processo de colheita e pós-colheita da mesma. A caracterização da pureza
da droga, quanto à presença de material estranho, teor de umidade e cinzas
das amostras analisadas, teve seus valores correspondentes a quantidades
permitidas pela Farmacopeia Brasileira V Ed. (2010), que corresponde a no
máximo 2,0 % de material estranho, 12,0 % de umidade, 13,0 % de cinzas
totais e 10,0 % de cinzas insolúveis em ácido, como pode ser verificado na
tabela 5.

63
Tabela 5 Determinação físico-química referente à pureza das folhas de Senna alata (L.) Roxb. (n = 3).
Ensaios de pureza Senna alata Ca (%)
Senna alata Re (%)
Material estranho 0,28 ± 0,02 0,23 ± 0,03
Água 11,8 ± 1,10 9,10 ± 1,20
Cinzas totais 8,12 ± 0,40 12,87 ± 1,00
Cinzas insolúveis em ácido 4,72 ± 0,50 6,22 ± 0,70
pH 7,0 6,0
A análise dos valores referentes à pureza dos folíolos como drogas
vegetais constitui resultado inédito para S. alata.
A avaliação do teor de umidade presente na droga vegetal é importante
na medida em que serve de parâmetro para a produção industrial, pois entre
outras questões revela dados sobre as condições da conservação da droga
vegetal para que se venha ter os cuidados necessários em sua preservação. O
excesso de umidade acarreta em ação de enzimática e consequente
degradação de constituintes químicos, além disso, facilita a proliferação de
micro-organismos (Galina, 2003).
Dependendo das condições de processamento e da composição
química do vegetal, minerais podem aparecem nas cinzas sob a forma de
óxidos, sulfatos, fosfatos, silicatos, cloretos. A determinação de cinzas em
amostras vegetais é importante por indicar equilíbrio microbiano e estabilidade
físico-química da droga. Figueiredo (2005) afirma que o teor de cinzas
insolúveis em ácido está relacionado principalmente com a presença de areia
no extrato. Essa informação pode justificar o fato do acesso restinga ter
apresentado maior quantidade em comparação à caatinga.
Com relação ao teor de cinzas entre os extratos do experimento neste
trabalho, os valores foram elevados, no entanto dentro do permitido pela
Farmacopéia Brasileira V Ed. (2010). Estas cinzas são resíduos inorgânicos
remanescentes da queima da matéria orgânica, que é transformada em CO2,
H2O e NO2. A composição das cinzas corresponde à quantidade de
substâncias minerais presentes, devido às perdas por volatilização ou mesmo
pela reação entre os componentes (Chaves et al., 2004).
Salienta-se que muitos dos minerais são vitais para a saúde, visto que
atuam no crescimento do corpo e seu metabolismo, nas contrações musculares

64
e também no balanceamento da água, entre muitos outros processos
(Wardilaw et al., 1994). Os resultados da tabela 4 de S. alata, mostram-se
similares quando comparados com os dados de S. alexandrina descrina na
Farmacopéia brasileira V Ed. (2010), uma vez que se tem para esta espécie
comercial o valor de 12 % para as cinzas totais e 3% para as cinzas insolúveis
em ádico.
4.3 Determinação do teor extrativo
A determinação do teor extrativo de S. alata permitiu verificar que o
rendimento do extrato a 10 % da droga vegetal (p/v), usando como solvente
etanol nas concentrações a 20, 40, 60, 80 e 95 %, obtiveram maior rendimento
em etanol a 40 % (v/v). Este resultado foi muito próximo às extrações com
etanol a 20 e 60 %, conforme apresentado na tabela 6.
Tabela 6 Determinação do teor extrativo dos extratos hidroetanólicos obtidos a partir das folhas de Senna alata (L.) Roxb. Ensaio realizado em triplicatas nas concentrações 20, 40, 60, 80 e
95 % de etanol. (n = 3).
O ensaio referente à determinação do teor extrativo é de suma
importância, pois antes de executar uma extração, deve-se levar em
consideração uma série de fatores que interferem nesta operação tais como
granulometria do material em pó, polaridade do solvente, acidez do meio,
agitação, temperatura, tempo de extração e tocixidade do solvente (Soares et
al., 1998), pois a estrutura histológica de um órgão vegetal é bastante
heterogênea o que influencia diretamente na eficiência da extração.
A composição química do gênero Senna, rico principalmente em
flavonoides e antracênicos, fez com que neste trabalho se optasse pela
extração com etanol. Também se utilizou etanol devido a polaridade e menor
índice de toxicidade dessa substância em relação a outros solventes
Solução extrativa em etanol (%) v/v
Teor extrativo de Senna alata (L.) Roxb. Ca
(%) p/p
Teor extrativo de Senna alata (L.) Roxb. Re
(%) p/p
20 21,54 ± 1,02 21,18 ± 1,40
40 23,89 ± 1,07 22,57 ± 1,00
60 20,98 ± 2,00 19,10 ± 1,12
80 16,10 ± 2,00 16,12 ± 2,00
95 09,90 ± 1,00 10,12 ± 1,00

65
comumente utilizado em extrações. Waksmundzka e colaboradores (2008)
ressalta que, como os extratos vegetais possuem moléculas polares e
apolares, a utilização de solvente com polaridade intermediária contribui para
separar os compostos químicos de maneira mais eficiente, a não ser que se
objetive uma determinada classe química de metabólitos.
Na escolha de um método extrativo, deve-se avaliar a eficiência, a
seletividade, a estabilidade das substâncias extraídas e o custo do processo
escolhido, considerando principalmente a finalidade do extrato que se quer
preparar (Costa, 1994; Simões e Spetizer, 2003). Assim é que, apesar de não
ter havido grandes diferenças entre o teor extrativo nos extratos entre etanol a
20 %, 40 % e 60 %, optou-se por utilizar a extração a 40 % de etanol por
representar uma economia média em etanol e em tempo e energia ao se
evaporar o solvente.
4.4 Triagem fitoquímica dos extratos
Em relação ao perfil fitoquímico qualitativo de S. alata, com a
demarcação da presença de classes químicas nos extratos obtidos a partir das
folhas, pode-se afirmar que os acessos da caatinga e restinga, apresentaram
perfis semelhantes, conforme tabela 7.
Tabela 7 Triagem fitoquimica dos extratos etanólico a 40 % obtidos a partir das folhas de Senna alata (L.) Roxb. acesso caatinga e restinga.
Ensaios S. alata caatinga S. alata restinga
Mucilagem - índice de intumescência 1,3 ± 0,1 mL 1,0 ± 0,1 mL
Saponinas - índice de Espuma 1,0 ± 0,1 cm 1,0 ± 0,1 cm
Saponinas hemolíticas - grau de Hemólise < 10 % < 10 %
Antraquinonas Presente Presente
Flavonoides Presente Presente
Taninos Taninos Ausente
A prospecção fitoquímica de uma planta geralmente tem como objetivo
principal o conhecimento das classes dos compostos químicos presentes no
extrato vegetal obtido a partir de um órgão específico da mesma. No que se
refere ao gênero Senna sabe-se que o mesmo possui um perfil químico em que
se sobressai os flavonoides e as antraquinonas, relatado por autores como
Viegas e colaboradores (2005). Neste trabalho, as classes químicas
encontradas nos extratos analisados foram fiéis aos padrões para o gênero e

66
subfamília de S. alata, encontrando-se as mesmas classes químicas que em S.
obtusifolia (L.) H.S. Irwin e Barneby e S. angustifolia Vahl. e S. occidentalis
(Lorenzzi, 2002; Lombardo et al., 2009; Schulz, 2012). Ressalta-se que os
resultados obtidos também corroboram com os ensaios histoquímicos de
Rodrigues e colaboradores (2011) para as folhas de S. alata, em que
identificaram antraquinonas, flavonoides, saponinas e taninos em material
obtido na Amazonia Oriental.
Ocorreu presença marcante de mucilagem nos dois acessos de S. alata,
com maior índice no acesso caatinga. A mucilagem é um polímero de alta
massa molecular que apresenta a capacidade de se complexar com moléculas
de água e por isso a presença marcante em cactos e outras plantas suculentas
ou não que ocorrem naturalmente em locais sob sol pleno e baixo regime de
águas. Estes metabólitos, por vezes, formam uma solução viscosa, produzidas
como exsudatos para a proteção de danos mecânicos ou a ataques
microbianos, que a planta tenha sofrido. Um exemplo de ocorrência de
mucilagens é o gênero Aloe, em que estes metabólitos ocorrem em grande
quantidade no tecido parenquimático das folhas (Thaiz e zeiger, 2002; Lulinski
et al., 2003). Farmacologicamente é uma classe química que apresenta
propriedade cicatrizante, anti-inflamatória, laxativa, expectorante e
antiespasmódica.
A determinação da presença de saponinas neste trabalho foi obtida pelo
índice de espuma e/ou determinação do grau da atividade hemolítica (Simões e
Spetizer 2003; Farmacopeia Brasileira Ed. V, 2010). Destaca-se que a
determinação da presença destes metabólitos em S. alata foi obtida pela
intensa formação de espuma e também pela atividade hemolítica em grau
baixo por hemólise de eritrócitos mesmo que em baixo grau (tabela 6) para os
dois acessos de S. alata. Ressalta-se que a palavra hemólise é definida na
literatura como um rompimento da membrana da hemácia, causando a
liberação da hemoglobina e outros componentes internos no líquido
circundante indicando que não está havendo danos à membrana celular dos
eritrócitos (Brandão et al., 2006), representando grau de toxidez dos extratos o
que necessariamente deve ser analisado em outros ensaios - o que foi
averiguado neste trabalho em testes seguintes. A presença de saponinas está
relatada para S. occidentalis e S. reticulata por Okwu (2001), Ogunkunle e

67
colaboradores (2006), Rodrigues e colaboradores (2009) e Alonso e
colaboradores (2015). Esta classe química de metabólitos apresenta um amplo
espectro de atividades biológicas e farmacológicas, estando associadas a
toxidez quando apresentam a capacidade de produzir hemólise e, muitas
vezes, estão relacionadas a atividade antimicrobiana (Lacaille et al., 1996). A
atividade hemolítica das saponinas faz parte do sistema de proteção do vegetal
contra ataques de predadores como insetos e outros animáculos (Bruneton,
1999), e pode estar ligada a atividade espermicida, anti-inflamatória e
antimicrobiana apresentada por uma variedade de plantas (Lacaille et al.,
1996).
Em outro ensaio, os extratos de S. alata em contato com o NH4OH as
amostras apresentaram coloração rósea o que constitui reação positiva para
antraquinona. Este grupo de compostos químicos é marcante para esta família
botânica e reflete a possibilidade de um maior potencial alelopático, bem como
atividade anti-inflamatória, vasodilatadora, ou ainda atividade anticarcinogênica
(Alemayehu et al., 1998; Machado, 2011; Rasul et al., 2011; Lu et al., 2013;
Vitorino et al., 2013).
Quanto à presença de taninos, foi detectada pelo aparecimento de
precipitado exposto ao cloreto férrico. Segundo Costa (1975), soluções
aquosas de cloreto férrico em contato com taninos podem causar alteração
para a cor verde-enegrecida, o que foi observado apenas para o acesso
caatinga de S. alata. Os taninos, algumas vezes, são relacionados ao sistema
de defesa das plantas em representantes da subfamília Caesalpinoideae
(Polhill et al., 1981; Ordoñes, 2004; Barrese et al., 2005).
4.4.1 Determinação de polifenois totais
Na determinação dos compostos fenólicos totais das amostras de S.
alata, fez-se a curva de calibração com ácido gálico e obteve-se a equação da
reta para o cálculo dos compostos fenólicos totais como equivalentes de ácido
gálico nas amostras. O coeficiente de determinação (r2) entre x e y, no valor de
0,9987, comprovou a correlação positiva entre a massa de ácido gálico (mg) e
a absorvância (760 nm) dos acessos estudados (figura 12).
O conteúdo de polifenois encontrados nas amostras em análise
demonstrou resultados semelhantes para os dois acessos de S. alata com
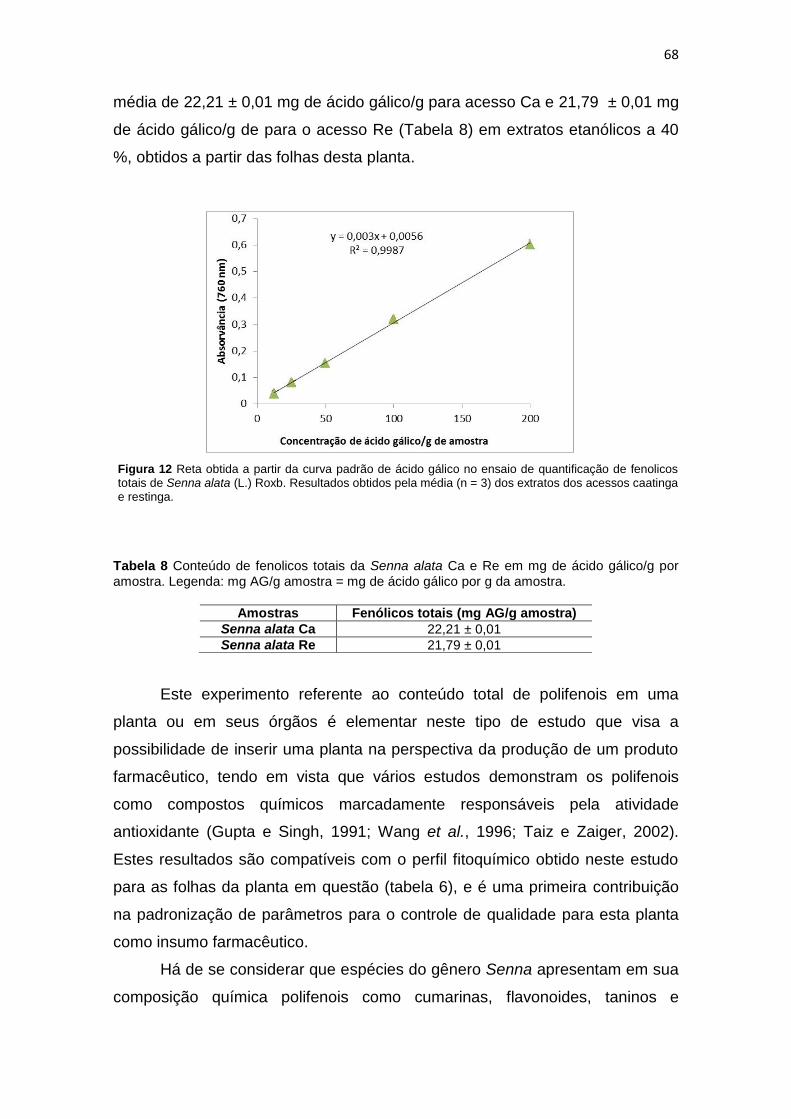
68
média de 22,21 ± 0,01 mg de ácido gálico/g para acesso Ca e 21,79 ± 0,01 mg
de ácido gálico/g de para o acesso Re (Tabela 8) em extratos etanólicos a 40
%, obtidos a partir das folhas desta planta.
Tabela 8 Conteúdo de fenolicos totais da Senna alata Ca e Re em mg de ácido gálico/g por
amostra. Legenda: mg AG/g amostra = mg de ácido gálico por g da amostra.
Este experimento referente ao conteúdo total de polifenois em uma
planta ou em seus órgãos é elementar neste tipo de estudo que visa a
possibilidade de inserir uma planta na perspectiva da produção de um produto
farmacêutico, tendo em vista que vários estudos demonstram os polifenois
como compostos químicos marcadamente responsáveis pela atividade
antioxidante (Gupta e Singh, 1991; Wang et al., 1996; Taiz e Zaiger, 2002).
Estes resultados são compatíveis com o perfil fitoquímico obtido neste estudo
para as folhas da planta em questão (tabela 6), e é uma primeira contribuição
na padronização de parâmetros para o controle de qualidade para esta planta
como insumo farmacêutico.
Há de se considerar que espécies do gênero Senna apresentam em sua
composição química polifenois como cumarinas, flavonoides, taninos e
Amostras Fenólicos totais (mg AG/g amostra)
Senna alata Ca 22,21 ± 0,01
Senna alata Re 21,79 ± 0,01
Figura 12 Reta obtida a partir da curva padrão de ácido gálico no ensaio de quantificação de fenolicos totais de Senna alata (L.) Roxb. Resultados obtidos pela média (n = 3) dos extratos dos acessos caatinga e restinga.

69
antraquinonas (Lombardo et al., 2009). Por outro lado, ressalta-se, que os
polifenois estão diretamente envolvidos com a disponibilidade hídrica e
radiação solar, fatores relevantes tanto na restinga como na caatinga,
interferindo diretamente na composição química e mesmo na fenologia das
plantas (Mota, 1975; Taiz e Zaiger, 2002).
A presença de polifenois que este ensaio demarca para as folhas de S.
alata corroboram com os resultados em estudos fitoquímicos para o gênero
Senna. Viegas e colaboradores (2006) constataram que as folhas e frutos de
Senna occidentalis são fontes majoritárias de polifenois, destacando-
sesobretudo pela presença de compostos antraquinônicos e flavonoídicos A
presença de compostos fenolicos também foi confirmada em Senna reticulata
por Santos, (2004) e Zuanassi (2004), onde o estudo fitoquímico de suas raízes
revelou a presença de cumarinas, taninos e flavanonas. A presença de
polifenois pode estar associada ao potêncial anti-inflamatório e relaxante
muscular, além da inibição de peroxidação lipídica, o que já foi constatado para
S. occidentallis.
4.4.2 Determinação de flavonoides totais
Com a curva padrão, obtida a partir das absorvâncias referentes a
concentrações crescentes de rutina através da equação da reta, com R2 de
0,997 foi possível obter as concentrações de (mg/mL) de flavonoides presentes
nas amostras de S. alata expressos em rutina (Figura 13). Os resultados do
doseamento de flavonoides foram de 10,12 ± 0,08 mg de rutina/g para acesso
Ca e 9,78 ± 0,1 mg de rutina/g para o acesso Re, como consta na tabela 9.
Figura 13 Gráfico obtido a partir da curva de calibração realizada com rutina na obtenção do teor total de flavonoides nos extratos das folhas de Senna alata (L.) Roxb. nos acessos caatinga e restinga. (n = 3).

70
Tabela 9 Conteúdo de flavonoides totais de Senna alata Caatinga e Restinga em mg de
rutina/g por amostra. Legenda: mg rutina/g amostra = mg de ácido gálico por g da amostra.
A determinação do teor de uma classe de substâncias químicas pode
servir como parâmetro, mesmo que inicial, tanto para um controle de qualidade
como também para se otimizar as quantidades desta classe química através de
variáveis nos processos de cultivo, colheita e pós colheita da planta. Ocorre
que a marcação das quantidades de uma classe química presente na
constituição química de uma planta, registra como a mesma irá reagir frente às
condições do habitat, e, consequentemente, demarcará as melhores
características climáticas-edáficas a serem observadas em seu cultivo e outros
processos com vistas a uma maior produção destes constituintes químicos
(Vigo et al., 2004; Freire et al., 2006). Apesar de a espectrofotometria no
UV/Vis apresentar limitações de seletividade e especificidade, principalmente
tratando-se de matrizes complexas como as drogas vegetais, esta técnica
apresenta confiabilidade suficiente para a sua adoção no controle de qualidade
da espécie em questão, além da simplicidade operacional e rapidez na
obtenção dos resultados (Moffat et al., 2004).
A presença de flavonoides é relatada em todas as partes das plantas do
gênero Senna (Gobbo-Neto e Lopes, 2007), sendo que esta classe de
substâncias químicas (além das antraquinonas) são os marcadores químicos
responsáveis por diversas atividades farmacológicas desempenhadas no
gênero Senna (Viega, J., 2005; Rodrigues et al. 2009).
Os resultados obtidos neste estudo identificam que entre o conteúdo
fenólico existente nas amostras de S. alata, cerca de 50 % constitui-se de
flavonoides. O teor de flavonoides em S. alata também foi obtido por Moriyama
e colaboradores (2013) com a planta coletada na Indonésia. Estes autores
demonstraram que o teor de flavonoides é maior entre janeiro a setembro, com
pico (4 %) em Janeiro. Assim é que o teor de flavonoides nos acessos testados
(caatinga e restinga) são praticamente o dobro do que se obtem na indonésia
para esta planta (Moriyama et al., 2003; Rodrigues et al., 2009). Por outro lado,
Amostras Fenólicos totais (mg rutina/g amostra)
Senna alata Ca 10,12 ± 0,08
Senna alata Re 9,78 ± 0,10

71
os resultados obtidos neste trabalho para o teor de flavonoides são próximos
ao que Nogueira (2009) obteve para o extrato hidroetanólico de Senna
macranthera em que o teor de flavonoides chegou a aproximadamente 50 %
dos fenois totais. Por vez, este mesmo autor chegou ao resultado de 70 % de
flavonoides do conteúdo total de polifenais ao utilizar metanol como solvente
extrator.
4.4.3 Perfil cromatográfico em cromatografia de camada delgada
Neste estudo objetivou-se verificar o comportamento químico dos
extratos de S. alata em diferentes fases móveis para se obter a presença do
maior número de bandas, com seus Rfs, além de marcar as que são comuns e
diferentes aos dois acessos testados (Re e Ca). A fase móvel escolhida foi
hexano: acetato de etila (1:1) e como agente cromogênico utilizou-se vanilina
sulfúrica a 100º C por 5 minutos (Figura 14). Neste sistema cromatográfico
obteve-se 6 bandas para o acesso caatinga (Rf: 0,21, 0,23, 0,38, 0,52, 0,77 e
0,97) e 4 bandas para o acesso restinga (Rf: 0,17, 0,42, 0,77, 0,97), tendo-se
somente duas bandas com Rf semelhantes (Rf: 0,77, 0,97).
A cromatografia em camada delgada visa, principalmente, desenvolver
um perfil cromatográfico para os extratos em estudo bem como, demonstrar as
diferenças e similaridades que se pode encontrar nos dois acessos em questão
Figura 14 Representação do perfil cromatográfico obtido para os extratos etanólicos a 40 % a partir das folhas de Senna alata (L.) Roxb., sobre cromatofolhas de sílica gel G 60 F254. Fase móvel constituída por
uma mistura de hexano: acetato de etila (1:1) e vanilina sulfúrica com tratamento de temperatura a 100º C por 5 min como agente cromogênico. Legenda: 1. Banda cromatográfica do extrato de S. alata Ca; 2. Bandas cromatográficas do extrato de S. alata Re; 3. Bandas cromatográficas do padrão rutina; 4. Bandas cromatrográficas do padrão quercetina.

72
através das bandas detectadas. Para tanto, se fez um estudo com diversas
fases móveis e agentes cromogênicos para um estudo do comportamento dos
extratos obtidos a partir das folhas de S. alata sobre cromatofolhas de sílica gel
G 60 F254, tomando por base o estudo de Wagner e Bladt (1996).
Em que pese as diferença de fatores ambientais existente entre os
acessos estudados, ocorrem duas bandas que são comum nesses acessos, o
que sugere que apesar da diferença climática entre os biomas, existem
substâncias em comum entre os dois acessos.
O estabelecimento de um perfil cromatográfico em cromatografia de
camada delgada (CCD) é corriqueiramente utilizado como uma técnica
farmacopeica que, apesar de ser simples, caracteriza inúmeras drogas
vegetais, de acordo com a Farmacopeia Brasileira Ed. V (2010).
Engel e colaboradores (2008) encontraram diferenças nos perfis
químicos em plantas do gênero Bauhinia, comercializadas como sendo pata-
de-vaca, através da observação de perfil cromatográfico em CCD. Estes
resultados mostram uma falta de padronização do material vegetal
comercializado. As amostras neste caso devem corresponder a plantas de
espécies diferentes ou a cultivares, épocas e manejos diferentes. Esses fatores
citados interferem nos resultados de análise em uma CCD e mostram a
necessidade de uma padronização em processos como na coleta e tratamento
do material vegetal. Deve-se comentar ainda que, por outro lado, a época de
coleta e a fase de desenvolvimento da planta podem desenvolver perfis
cromatográficos diferentes entre o extrato de uma mesma espécie devido a
variação climpatica existência em cada estação do ano.
Tendo acesso a esses resultados, que confirma a presença de
compostos químicos capazes de atuar no sistema biológico de forma positiva,
nos estudos seguintes, são realizados teste de atividade biológica, que buscará
verificar se esses compostos são eficazes no controle antibacteriano e no
tratamento da inflamação, sendo também testado o potencial citotóxico e tóxico
dos extratos em diferentes concentrações.

73
4.5 Atividades biológicas
4.5.1 Atividade antibacteriana
A bioautografia é considerada um ensaio qualitativo, que permite
detectar na cromatografia as bandas que apresentam atividade antimicrobiana
a partir da formação de halos de inibição (Holetz, 2002). O ensaio
bioautográfico demonstrou que o extrato etanolico a 40 % obtido a partir das
folhas de S. alata dos acessos caatinga e restinga, apresentam bandas
cromatográficas com atividade antibacteriana para as duas cepas utilizadas
(Tabela 10).
Tabela 10 Halos de inibição obtidos em cromatografia dos extratos de Senna alata, acesso
caatinga e restinga, frente a Staphilococcus aureus e Escherichia coli.
Linhagens bacterianas
DIÂMETRO DOS HALOS DE INIBIÇÃO (cm)
Senna alata Ca (Rf) Senna alata Re (Rf) Amoxicilina
0,21 0,23 0,38 0,52 0,77 0,97 0,17 0,42 0,77 0,97
Staphylococcus aureus
- 1,26 - - 0,62 -
0,89 - 0,74 - 1,2
Escherichia coli 0,12 - 0,91 - 0,52 0,61 0,20 - 0,54 0,94 1,4
Para a bactéria gram negativa Escerichia coli, observou-se inibição do
crescimento em 4 e 3 bandas no acesso Ca e Re, respectivamente. Para a
bactéria gram positiva Staphylococcus aureus, obteve-se inibição em 2 bandas
para os acessos em estudo. Destaca-se que duas bandas, Rf 0,17 e 0,77,
inibiram o crescimento das duas sepas em análise e, para S. aureus, o Rf 0,23
do acesso Ca demonstrou halo de inibição superior ao encontrado no padrão
utilizado de amoxicilina.
Os resultados obtidos somam-se ao estudo de Silva e colaboradores
(2017), em que extratos de S. alata mostraram-se eficiente como agente
antibacteriano frente à Xanthomonas axonopodis, com redução de 37,82 % da
doença do maracujazeiro, correspondendo a um resultado muito significativo
quando comparado ao padrão utilizado no controle fitopatogenico (Óxicloreto
de cobre). Por outro lado, o potencial antibacteriano de extratos obtidos a partir
de caule e folhas de S. obtusifolia foi negativo para A. aureus e E. coli nos
estudos de Cordeiro (2006) e Rodrigues e colaboradores (2011) , diferente dos
resultados para S. alata, neste trabalho.
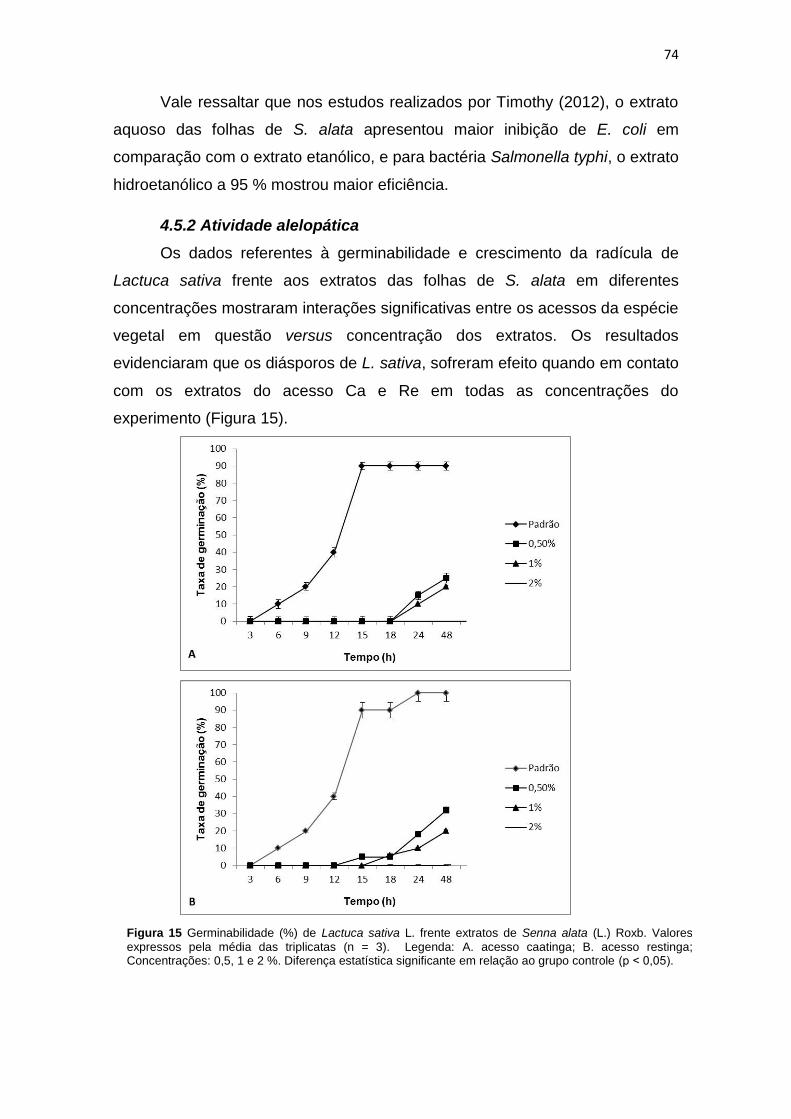
74
Vale ressaltar que nos estudos realizados por Timothy (2012), o extrato
aquoso das folhas de S. alata apresentou maior inibição de E. coli em
comparação com o extrato etanólico, e para bactéria Salmonella typhi, o extrato
hidroetanólico a 95 % mostrou maior eficiência.
4.5.2 Atividade alelopática
Os dados referentes à germinabilidade e crescimento da radícula de
Lactuca sativa frente aos extratos das folhas de S. alata em diferentes
concentrações mostraram interações significativas entre os acessos da espécie
vegetal em questão versus concentração dos extratos. Os resultados
evidenciaram que os diásporos de L. sativa, sofreram efeito quando em contato
com os extratos do acesso Ca e Re em todas as concentrações do
experimento (Figura 15).
Figura 15 Germinabilidade (%) de Lactuca sativa L. frente extratos de Senna alata (L.) Roxb. Valores
expressos pela média das triplicatas (n = 3). Legenda: A. acesso caatinga; B. acesso restinga; Concentrações: 0,5, 1 e 2 %. Diferença estatística significante em relação ao grupo controle (p ˂ 0,05).

75
A viabilidade das sementes não germinadas foi verificada pela secção
das sementes transversalmente, após tratamento com solução aquosa de
cloro-2,3,5-trifenil cloreto de tetrazólio a 1 % (v/v). Neste ensaio detectou-se
que 10 % das sementes estavam inviáveis e o restante (90 % das sementes
não germinadas) desenvolveram cor rosa, indicando a viabilidade das mesmas.
Os diásporos de L. Sativa frente à água pura completou o processo
germinativo em 12 horas com 98 % dos diásporos germinados. No entanto, as
características analisadas indicaram que os extratos de S. alata tiveram uma
influência negativa na germinação dos diásporos de L. sativa, visto que nas
concentrações 0,5 e 1 %, houve retardo germinativo e protrusão radicular após
12 horas para o acesso caatinga e 18 horas para o acesso restinga e, após 48
horas, para os extratos nas concentrações de 0,5 e 1 % teve-se
germinabilidade de inferior a 10 %, nos dois testados. Já, na concentração do
extrato a 2 %, mesmo após 48 horas, não se verificou nenhum
desenvolvimento germinativo dos diásporos de L. sativa, constatando completa
inibição da germinação.
Com o teste do tetrazólio nos diásporos não germinados constatou-se
que um percentual muito baixo (10 %) estava viáveis o que indica de fato a
atividade alelopática dos extratos utilizados.
Em relação ao comprimento radicular, os resultados indicaram que
houve efeito negativo no desenvolvimento das sementes germinadas frente aos
extratos de S. alata, pois quando comparadas com o controle (água pura) o
desenvolvimento radicular foi comprometido severamente. Estes dois fatos
apontam para um efeito alelopático do extrato em análise.
Com os resultados obtidos neste ensaio pode-se inferir que os extratos
testados desenvolvem atividade alelopática e que os mesmos apresentam
substâncias aleloquimicas, provocando forte interferência no desenvolvimento
dos diásporos de L. sativa. Este resultado corrobora com os estudos realizados
por Agbagwa e colaboradores (2003), em testes de alelopatia para extratos de
S. alata frente à celosia argentea e, como no presente estudo, também
obtiveram efeitos alelopáticos. Com isso tem-se o indicativo de uma possível
atividade inseticida e herbicida para o extrato testado, conforme Bansal e Bhan
(1993), Souza (1995), Lorenzi, (2000) e Rodrigues (2011). Neste sentido,
soma-se a informação de Barrese e colaboradores (2005) que registra o cultivo

76
na África de S. alata ao redor das casas para espantar formigas. Por outro
lado, Polhill e colaboradores (1981) e Taiz e Zeiger (2002) explicam que uma
planta pode reduzir o crescimento das plantas vizinhas pela liberação de
aleloquímicos no solo, isso pode ter como consequência a chance maior de
acesso à luz, à água e aos nutrientes e, portanto, propiciar sua maior
adaptação evolutiva.
Com base no estudo fitoquimico realizado nesse trabalho, pode-se
afirmar que S. alata possui classes de compostos orgânicos que apresentam
atividade fitoquímica o que pode justificar o potencial alelopático dessa planta,
a exemplo das antraquinonas, flavonoides e taninos (encontrado no acesso
Ca).
4.5.3 Viabilidade celular pelo método MTT
Não foi observada redução da viabilidade celular após o tratamento de
macrófagos com os extratos de S. alata. Os extratos Ca e Re não causaram
citotocidez nas concentrações testadas, em comparação com macrófagos
lisados com Triton-X (controle de lise celular máxima) e, não tratado, conforme
figura 16.
A citotoxicidade, em geral, é avaliada com a finalidade de verificar o
quão seguro as diferentes substâncias podem ser e em quais concentrações
essa segurança e citotoxidez são observadas para que permaneça na triagem
como candidatos a nova droga.
Os resultados dos testes efetuados nas células tratadas com os extratos
de S. alata não apresentaram diferenças consideráveis entre os acessos Ca e
Figura 16 Viabilidade do extrato de Senna alata (L.) Roxb. nos acessos catinga e restinga. Nas
concentrações de 50 e 100 µg/mL em macrófagos pelo método de 3-(4,5dimetiltiazolio-2-il)-2,5-difenil tetrazólio. Valores expressos pela média das triplicatas. Legenda: C +. controle positivo: macrófagos estimulados e não tratados; Controle negativo: Triton X. Leitura a 570 nm; Diferença estatística significante em relação ao grupo controle (p˂0,05).

77
Re, nas concentrações testadas, apresentando-se próximos ao controle
positivo. Esse resultado sugere citotoxicidade baixa ou ausente para esses
extratos, podendo assim ser considerado seguro em formulações
farmacêuticas.
Esta foi a primeira vez que estudos de citoxidade por MTT no gênero
Senna foram abordados em pesquisa científica, no entanto, outras técnicas de
verificação de citotoxicidade já foram testadas com plantas do gênero Senna
onde também apresentaram baixa mortalidade celular. Lombardo (2008)
avaliou o potêncial citotóxico de Senna occidentallis a partir do método MTS,
que se difere do MTT principalmente pela formação de cristais solúveis em
água. O autor verificou que, a concentração hidroetanólica testada superior a
0,38 % (p/v), é considerada tóxica, causando morte da população celular, logo
nas primeiras 24 horas de contato com a cultura apresentando viabilidade
celular inferior a 50 %.
Estudos realizados com outros gêneros da subfamília Caesalpinoide
podem ser também compados a esses resultados. Kayano e colaboradores
(2011) e Zanin e colaboradores (2012) verificaram, a partir de espécies do
gênero Caesalpinia resultados satisfatórios em relação a citotoxidade das
espécies testadas (C. pyramidalis, Caesalpinia pluviosa e C. férrea), onde
demostraram toxicidade a protozoários causadores da malária severa em
humanos e ao mesmo tempo não apresentaram efeito citotoxico significativo
em células de camundongos Swiss.
Rosário (2010) também verificou o potencial citotóxico de espécie da
sunfamília Caesalpinoideae. Segundo o autor, a Copaifera langsdorffii, não
exerceu efeito citotóxico significativo nas concentrações de 10 a 100 μg/mL
para macrófagos estimulados com LPS.
Esses resultados foram importantes para o conhecimento da dosagem
segura de S. alata nos estudos conseguintes e, além disso, motivam a
continuidade da investigação a fim de verificar o potêncial anti-inflamatorio dos
acessos estudados.
4.5.4 Atividade anti-inflamatória in vitro com LPS
Verificou-se a produção de NO e TNF-α induzidos por LPS,
demonstrando os efeitos anti-inflamatórios dos extratos das folhas de S.alata
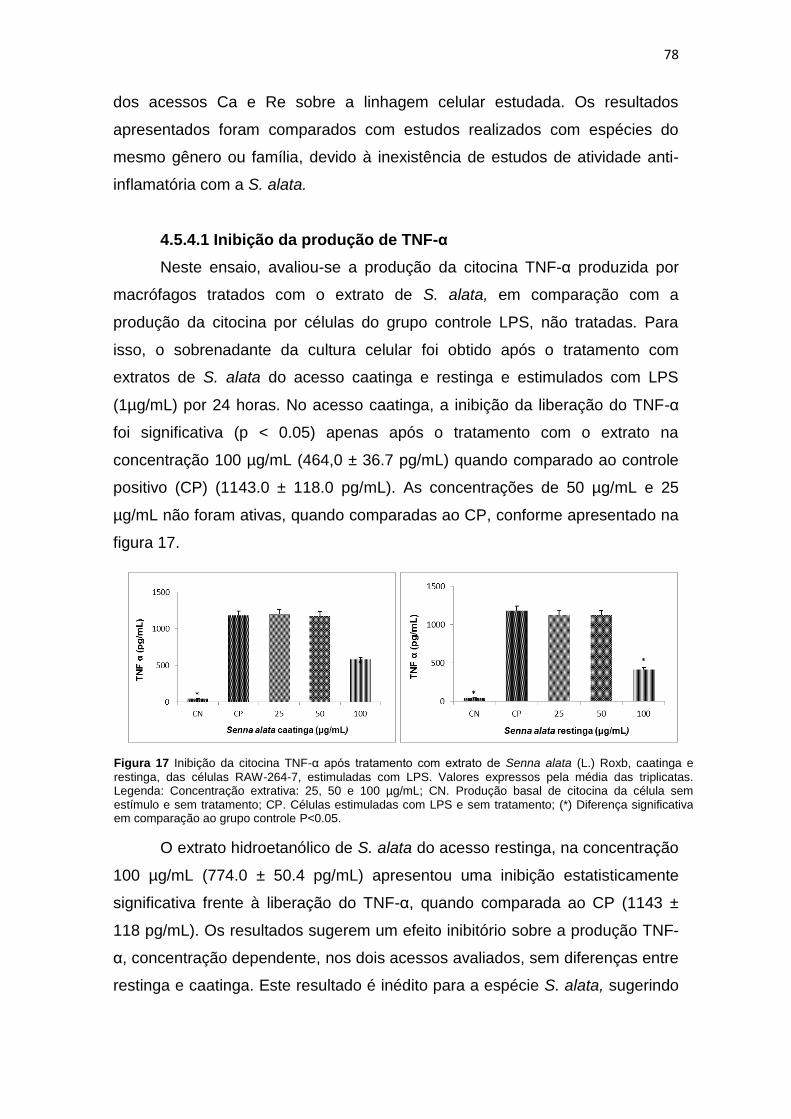
78
dos acessos Ca e Re sobre a linhagem celular estudada. Os resultados
apresentados foram comparados com estudos realizados com espécies do
mesmo gênero ou família, devido à inexistência de estudos de atividade anti-
inflamatória com a S. alata.
4.5.4.1 Inibição da produção de TNF-α
Neste ensaio, avaliou-se a produção da citocina TNF-α produzida por
macrófagos tratados com o extrato de S. alata, em comparação com a
produção da citocina por células do grupo controle LPS, não tratadas. Para
isso, o sobrenadante da cultura celular foi obtido após o tratamento com
extratos de S. alata do acesso caatinga e restinga e estimulados com LPS
(1µg/mL) por 24 horas. No acesso caatinga, a inibição da liberação do TNF-α
foi significativa (p < 0.05) apenas após o tratamento com o extrato na
concentração 100 µg/mL (464,0 ± 36.7 pg/mL) quando comparado ao controle
positivo (CP) (1143.0 ± 118.0 pg/mL). As concentrações de 50 µg/mL e 25
µg/mL não foram ativas, quando comparadas ao CP, conforme apresentado na
figura 17.
O extrato hidroetanólico de S. alata do acesso restinga, na concentração
100 µg/mL (774.0 ± 50.4 pg/mL) apresentou uma inibição estatisticamente
significativa frente à liberação do TNF-α, quando comparada ao CP (1143 ±
118 pg/mL). Os resultados sugerem um efeito inibitório sobre a produção TNF-
α, concentração dependente, nos dois acessos avaliados, sem diferenças entre
restinga e caatinga. Este resultado é inédito para a espécie S. alata, sugerindo
Figura 17 Inibição da citocina TNF-α após tratamento com extrato de Senna alata (L.) Roxb, caatinga e
restinga, das células RAW-264-7, estimuladas com LPS. Valores expressos pela média das triplicatas. Legenda: Concentração extrativa: 25, 50 e 100 µg/mL; CN. Produção basal de citocina da célula sem estímulo e sem tratamento; CP. Células estimuladas com LPS e sem tratamento; (*) Diferença significativa em comparação ao grupo controle P<0.05.

79
um potencial efeito anti-inflamatório associado à inibição da produção desta
citocina.
A produção da citocina TNF-α foi citada por Galdino (2012) em estudo
realizado com S. angustifólia, onde o extrato de Sene estimulou a produção
das citocinas IL-1, IL-6 e TNF-α, que promoveram a reabsorção das células
trifoblásticas de camundongos gestantes, com um efeito pró-inflamatório
contrário ao aqui observado.
4.5.4.2 Inibição da produção de oxido nítrico in vitro
A atividade inibitória do extrato hidroetanólico de S. alata na produção
de NO por macrófagos estimulados com lipopolissacarídeos (LPS) foi
estatisticamente significante quando comparado aos resultados obtidos em
macrófagos controles, não tratados. Não foram observadas diferenças
significativas entre as concentrações de 12,5 e 25 µg/mL de S. alata, em
comparação com o controle LPS. Com a concentração de 50 µg/mL de S.
alata, a produção de NO foi estatisticamente menor (p < 0,05) quando
comparado ao grupo controle LPS, com o acesso Ca inibindo 44,21 ± 3,04 % e
o extrato aquoso do acesso RE inibindo 16,11 ± 3,22 da produção de NO
induzida por LPS (figura 18).
Ambos os acessos foram ativos sobre a inibição da produção de NO,
apenas na concentração 50 μg/mL, sugerindo um efeito concentração-
dependente. Esses resultados demonstram uma eficiência na redução do
Figure 18 Produção de óxido nítrico nos grupos tratados com Senna alata (L.) Roxb. acesso caatinga e
acesso restinga. Valores expressos pela média das triplicatas. Legenda: C. controle positivo - macrófagos estimulado por LPS. (*) Diferença significativa em comparação ao grupo controle (p˂0,05).

80
processo inflamatório induzido por LPS. Esse resultado já era esperado, pois
espécies do mesmo gênero, como S. angustifolia e outras espécies da família
Fabaceae apresentaram resultados similares (Almeida, 2005; Rodrigues, et al.
2009; Severo, 2013). O NO é um importante mensageiro intercelular nos
mamíferos, exercendo sinalização intracelular, sem receptores
transmembranosos, o que torna o NO um eficiente e rápido elemento de
resposta não só do sistema imunológico, mas também em outros importantes
sistemas do organismo. Portanto, o NO é um importante alvo terapêutico de
substâncias antiinflamatórias (Alemayehu et al. 1998; Jesus, 2016; Silva,
2015).
4.5.5 Atividade anti-inflamatória in vitro com Zymosan
No estudo com o Zymosan, assim como o realizado com o LPS, foi
verificada a produção de NO e TNF-α após o estímulo em macrófagos. Os
efeitos anti-inflamatórios dos extratos das folhas de S.alata estimulados com Zy
apresentaram similaridade na redução de NO nos extratos dos dois acessos
caatinga e restinga na concentração de 50 μg/mL (figura 19), contudo, houve
diferenciação na dosagem de TNF-α que apresentou maior inibição no extrato
da restinga, com redução da produção a partir de 25 μg/mL. (figura 20).
4.5.5.1 Inibição da produção de TNF-α
A citocina TNF-α, foi quantificada no sobrenadante obtido da cultura de
macrófagos, após tratamento com extratos hidroetanólicos de S alata do
acesso caatinga e restinga e estimulados com Zy (1 µg/mL) por 24 horas. No
acesso caatinga, a inibição da liberação do TNF-α foi significativa (p < 0,05)
após tratamento com o extrato etanólico nas concentrações 100 µg/mL (684,12
± 38,21 pg/mL) e 50 µg/mL (678,12 ± 39,46 pg/mL) quando comparado ao
controle positivo (984 ± 97,0 pg/mL). A concentração de 25 µg/mL não
apresentou diferença significativa quando comparado ao controle positivo
indicando um efeito inibitório concentração-dependente, conforme figura 19.
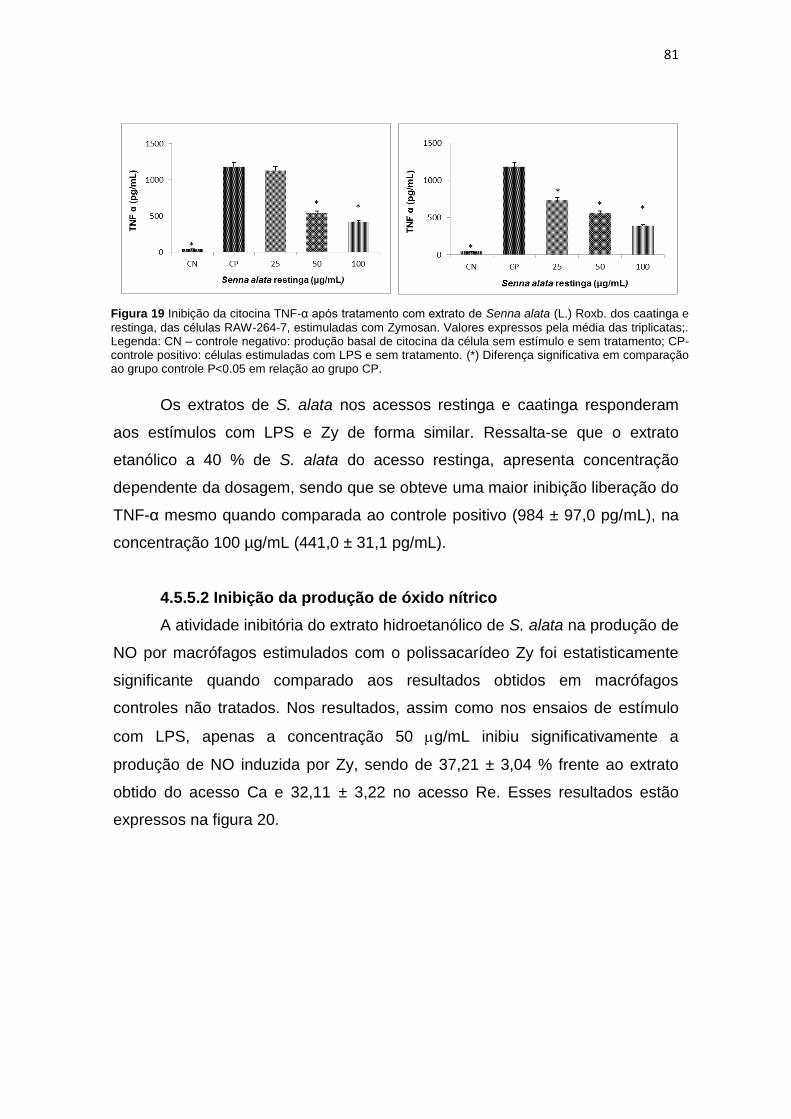
81
Os extratos de S. alata nos acessos restinga e caatinga responderam
aos estímulos com LPS e Zy de forma similar. Ressalta-se que o extrato
etanólico a 40 % de S. alata do acesso restinga, apresenta concentração
dependente da dosagem, sendo que se obteve uma maior inibição liberação do
TNF-α mesmo quando comparada ao controle positivo (984 ± 97,0 pg/mL), na
concentração 100 µg/mL (441,0 ± 31,1 pg/mL).
4.5.5.2 Inibição da produção de óxido nítrico
A atividade inibitória do extrato hidroetanólico de S. alata na produção de
NO por macrófagos estimulados com o polissacarídeo Zy foi estatisticamente
significante quando comparado aos resultados obtidos em macrófagos
controles não tratados. Nos resultados, assim como nos ensaios de estímulo
com LPS, apenas a concentração 50 g/mL inibiu significativamente a
produção de NO induzida por Zy, sendo de 37,21 ± 3,04 % frente ao extrato
obtido do acesso Ca e 32,11 ± 3,22 no acesso Re. Esses resultados estão
expressos na figura 20.
Figura 19 Inibição da citocina TNF-α após tratamento com extrato de Senna alata (L.) Roxb. dos caatinga e
restinga, das células RAW-264-7, estimuladas com Zymosan. Valores expressos pela média das triplicatas;. Legenda: CN – controle negativo: produção basal de citocina da célula sem estímulo e sem tratamento; CP- controle positivo: células estimuladas com LPS e sem tratamento. (*) Diferença significativa em comparação ao grupo controle P<0.05 em relação ao grupo CP.
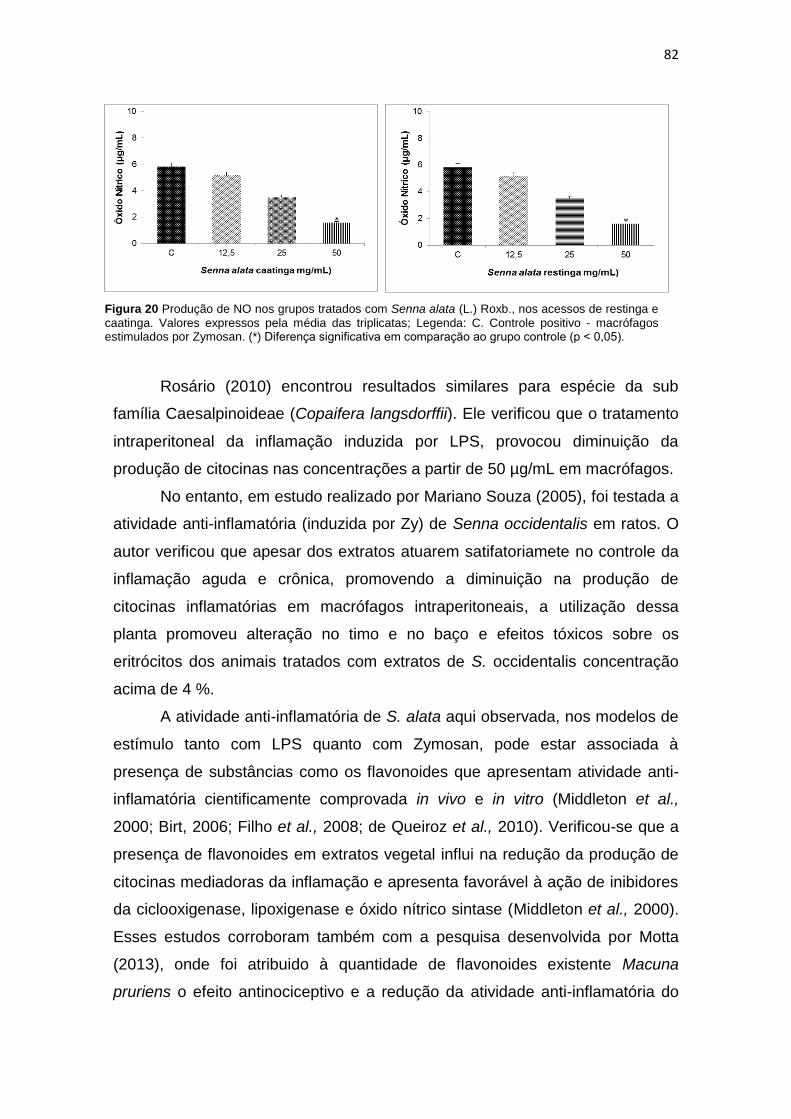
82
Rosário (2010) encontrou resultados similares para espécie da sub
família Caesalpinoideae (Copaifera langsdorffii). Ele verificou que o tratamento
intraperitoneal da inflamação induzida por LPS, provocou diminuição da
produção de citocinas nas concentrações a partir de 50 µg/mL em macrófagos.
No entanto, em estudo realizado por Mariano Souza (2005), foi testada a
atividade anti-inflamatória (induzida por Zy) de Senna occidentalis em ratos. O
autor verificou que apesar dos extratos atuarem satifatoriamete no controle da
inflamação aguda e crônica, promovendo a diminuição na produção de
citocinas inflamatórias em macrófagos intraperitoneais, a utilização dessa
planta promoveu alteração no timo e no baço e efeitos tóxicos sobre os
eritrócitos dos animais tratados com extratos de S. occidentalis concentração
acima de 4 %.
A atividade anti-inflamatória de S. alata aqui observada, nos modelos de
estímulo tanto com LPS quanto com Zymosan, pode estar associada à
presença de substâncias como os flavonoides que apresentam atividade anti-
inflamatória cientificamente comprovada in vivo e in vitro (Middleton et al.,
2000; Birt, 2006; Filho et al., 2008; de Queiroz et al., 2010). Verificou-se que a
presença de flavonoides em extratos vegetal influi na redução da produção de
citocinas mediadoras da inflamação e apresenta favorável à ação de inibidores
da ciclooxigenase, lipoxigenase e óxido nítrico sintase (Middleton et al., 2000).
Esses estudos corroboram também com a pesquisa desenvolvida por Motta
(2013), onde foi atribuido à quantidade de flavonoides existente Macuna
pruriens o efeito antinociceptivo e a redução da atividade anti-inflamatória do
Figura 20 Produção de NO nos grupos tratados com Senna alata (L.) Roxb., nos acessos de restinga e
caatinga. Valores expressos pela média das triplicatas; Legenda: C. Controle positivo - macrófagos estimulados por Zymosan. (*) Diferença significativa em comparação ao grupo controle (p ˂ 0,05).
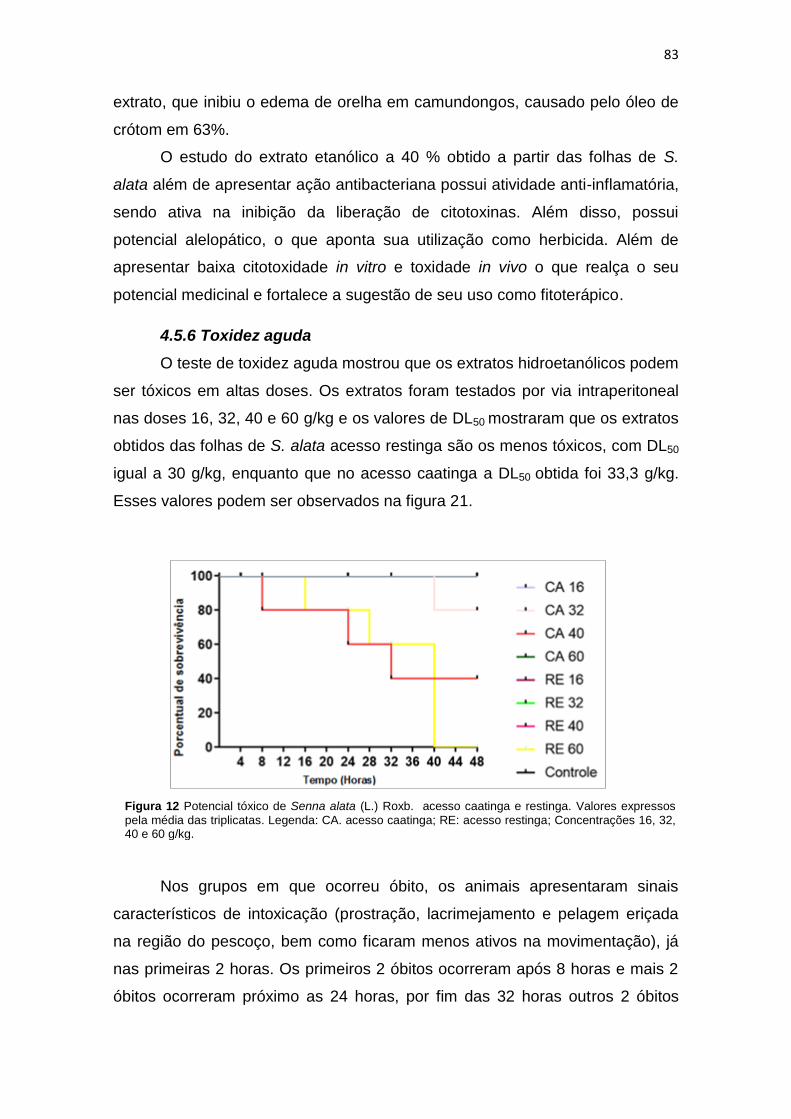
83
extrato, que inibiu o edema de orelha em camundongos, causado pelo óleo de
crótom em 63%.
O estudo do extrato etanólico a 40 % obtido a partir das folhas de S.
alata além de apresentar ação antibacteriana possui atividade anti-inflamatória,
sendo ativa na inibição da liberação de citotoxinas. Além disso, possui
potencial alelopático, o que aponta sua utilização como herbicida. Além de
apresentar baixa citotoxidade in vitro e toxidade in vivo o que realça o seu
potencial medicinal e fortalece a sugestão de seu uso como fitoterápico.
4.5.6 Toxidez aguda
O teste de toxidez aguda mostrou que os extratos hidroetanólicos podem
ser tóxicos em altas doses. Os extratos foram testados por via intraperitoneal
nas doses 16, 32, 40 e 60 g/kg e os valores de DL50 mostraram que os extratos
obtidos das folhas de S. alata acesso restinga são os menos tóxicos, com DL50
igual a 30 g/kg, enquanto que no acesso caatinga a DL50 obtida foi 33,3 g/kg.
Esses valores podem ser observados na figura 21.
Nos grupos em que ocorreu óbito, os animais apresentaram sinais
característicos de intoxicação (prostração, lacrimejamento e pelagem eriçada
na região do pescoço, bem como ficaram menos ativos na movimentação), já
nas primeiras 2 horas. Os primeiros 2 óbitos ocorreram após 8 horas e mais 2
óbitos ocorreram próximo as 24 horas, por fim das 32 horas outros 2 óbitos
Figura 12 Potencial tóxico de Senna alata (L.) Roxb. acesso caatinga e restinga. Valores expressos
pela média das triplicatas. Legenda: CA. acesso caatinga; RE: acesso restinga; Concentrações 16, 32, 40 e 60 g/kg.

84
foram registrados. Nos grupos das doses 40 g/kg Ca e 60 g/kg Re foi
observado um óbito por horário registrado. Esses resultados, conforme
Schorderet (1992) e outros autores, não representam toxidez considerável, pois
relatam que doses letais médias acima de 5 g/kg são consideradas não tóxicas.
A atividade tóxica dos extratos de S. alata obtidos na caatinga e restinga
apresentou-se menos tóxico do que os resultados encontrados por Pieme
(2006), para a mesma espécie obtida no centro sul da África, onde foi
observada uma dose letal média de 18,50 g/kg, em ratos, com ausência de
danos histopatológico do fígado mesmo em períodos prologados de
administração. Em que pese as variantes do experimento realizado neste
trabalho com a de Pieme (2006) como o local de coleta que deve imprimir um
diferencial na composição química do extrato e mesmo o tipo de animal
testado, vale ressaltar que, segundo este autor, neste trabalho em ratos, o
extrato etanólico de S. alata sugeriu proteção de hepatócitos e promoveu uma
melhora na arquitetura do fígado.
4.5.6.1 Liberação de enzima citoplasmática lactato desidrogenase
(LDH)
Neste ensaio constatou-se que ocorreu baixa liberação de LDH no soro
de camundongos swiss, em todas as concentrações testadas com os extratos
de S. alata (acesso caatinga e restinga). Os resultados desse ensaio
evidenciaram dose dependente, ou seja, a quantidade de LDH no soro dos
animais aumentou proporcionalmente ao aumento das concentrações testadas
dos extratos de S. alata (figura 22).
Figura 22 Liberação da LDH em soro sanguíneo de camundongos Swiss tratados com extratos de Senna
alata (L.) Roxb. (n=5). Legenda: A. Extratos de Senna alata acesso caatinga; B. Extratos de Senna alata acesso restinga; CN. Controle negativo: Soro sanguineo dos camundongos Swiss sem tratamento com os extratos; CP. Controle positivo: Soro sanguíneo dos camundongos swiss com DMSO a 1 %.
A B

85
Os resultados obtidos neste ensaio sugerem que a toxidez acometida
nos camundongos swiss, até mesmo a seguida por óbito (Ca 40 e Re 60 g/kg)
deve não ter ocasionado alterações renais, infarto do miocárdio ou qualquer
lesões hepáticas, uma vez que o valor da LDL liberada no soro desses animais
foi baixa, próximo ao controle negativo. Segundo Balls e colaboraores (1992)
os valores para a LDH liberada no soro de sangue de camundongos, conforme
o tempo de vida desses animais apresenta quantidade referencia entre 74 a
253 unidade por litro (U/L), para não ter hemólise nas amostras testadas. Como
os valores obtidos neste ensaio estão todos dentro da faixa referencia proposta
pelo autor citado, pode-se afirmar que não houve hemólise neste teste e que os
valores obtidos da LDH no soro dos animais nesta faixa etária são normais,
não podendo ser tomado como um indicativo de que houve morte celular
seguida de rompimento de membrana, o que é classificado como necrose
(Balls et al., 1992; Lantto et al., 2009).
Pieme (2006), utilizando extrato etanólico a 80 % obtido a partir das
folhas de S. alata, coletadas no centro sul da África, determinou capacidade
protetora de hepatócitos de ratos para este extrato. Em outro trabalho com
Senna, de Rajan e colaboradores (2009) com Cassia tora, corroboram com os
resultados obtidos neste ensaio, pois trabalho este autor verificou a capacidade
hepatoprotetora de Cassia tora, que apresenou efeitos satisfatórios contra
danos ao fígado de ratos albinos. Os resultados obtidos destes autores
corroboram com o ensaio que se realizou e apontam para o uso seguro de S.
alata como medicinal em doses e formulações conhecidas.

86
5. CONCLUSÕES
As características macro e microscópicas dos acessos de S. alata na
restinga e na caatinga são semelhantes e apresentam a forma, o ápice e
a presença de estômatos nas duas faces epidérmicas e tricomas de
cutícula lisa como caracteres úteis na autenticação das folhas como
insumos farmacêuticos.
Os acessos de S. alata, apresentaram similaridade de teor extrativo
sendo maior no extrato etanólico a 40 %;
As amostras apresentaram índices de pureza compatível com as
requeridas na V edição da Farmacopéia Brasileira. A diferença básica
entre os acessos reside no fato do acesso caatinga apresentar taninos e
extrato com maior pH. O teor total de polifenois e de flavonoides foi
semelhante nos dois acessos estudados;
O perfil cromatográfico em CCD apresenta 2 bandas semelhantes para
os acessos da restinga e caatinga;
Detectou-se atividade antibacteriana por bioautografia nos acessos
obtidos na caatinga e na restinga, indicando atividade de amplo
espectro, tanto para bactérias gram positivas como para gram negativas;
Ficou determinada atividade alelopática dos extratos de S. alata, com
diminuição de germinabilidade de diásporos de L. sativa e
descaracterização das radículas, principalmente no acesso da caatinga,
denotando possível atividade inseticida e bio-herbicida para estes
extratos;

87
Os extratos etanólico a 40 % oriundos das folhas de S. alata acesso
caatinga e restinga não apresentaram citotoxidez pelo método MTT, pois
não houve interferência desses extratos na viabilidade de macrófagos,
sugerindo a ausência de morte celular por necrose.
Os testes in vitro com LPS e Zy sugeriram que os extratos são eficazes
no controle de TNF e NO em macrófagos, principalmente nas
concentrações 100 µg/mL e 50 µg/mL, sem diferença entre os dois
acessos.
A toxidez aguda da droga testada pode ser considerada como baixa,
uma vez que em camundongos Swiss, determinou-se DL50 apenas nas
doses acima de 30 g/kg, sem elevação dos níveis séricos de LDH.

88
REFERENCIAS
A3P – Agenda ambiental na administração pública. Ministério do Meio Ambiente. Brasília – DF. 5ª Edição | Revista e atualizada, 2009. ABBAS, A. K.; Lichtman, A. H. Imunologia Celular e Molecular. 5ed. Rio de Janeiro: Elsevier, p.580, 2005. ABIFITO – Associação Brasileira da Indústria de Fitoterápicos. Uma legislação justa para os produtos de origem natural (2002). DisponÌvel em <http://www.abifito.com.br>. Acesso em 12 de Junho de 2017. AGBAGWA, I. O.; Onofeghara, F. A.; Mensah, S. I. Stimulation of growth and development of Celosia argentea L. by crude extracts of Senna alata (L.) Roxb. J. Appl. Sci. Environ. Manag., v. 7, n. 1, p. 9-13, 2003. AGERBIRK, N.; Olsen, C.E. and Neilsen, J.K. Seasonal variation in leaf glucosinolate and insect resistance in two types of Barbarea vulgaris spp. arcuata. Phytochemistry, 58, 91-100, 2001. AKERELE, O. Medicinal plants and primary health care: a agent for action. Fitoterapia, v.LIX, n.5, p.355-63, 1988. ALBERTON, J.R.I; Ribeiro, A.; Sacramento, L.V.S.; Franco, S.L.; Lima, M.A.P. Caracterização farmacognóstica do jambolão (Syzypiam cumini (L.) Skeels). Rev Bras Farmacogn 11: 37-5, 2001. ALEMAYEHU, G.; Abegaz, B. M. Bianthraquinones from the seeds of Senna multiglandulosa. Phytochemistry, v. 41, n. 3, p. 919-921, 1996. ALEMAYEHU, G., Abegaz, B., Kraus W. A. Anthraquinone-dihydroantrhacenone dimer from Senna sophera. Phytochemistry, v. 48, n. 4, p. 699-702, 1998. ALMEIDA, A. A. de.; Ming, T. D.; Corrêa, C. L.; Lavinas, T. Verificação da qualidade dos fitoterápicos Sene e Boldo-do-chile comercializados na região de Campinas. Faculdade e ciências farmacêuticas, Centro de ciências da vida. Pontifícia Universidade Católica de Campinas. Revista de ciências médicas de Campinas, 14 (3), p 279-285, 2005. AMARAL P., Amaral Neto, M; Krämer, F. Oficina de manejo comunitário e certificação na América Latina: resultados e propostas. Belém: IMAZON,GTZ,IEB. 44p, 2005. AMARAL, D. D; Prot, M. T; Bastos, M. N. C; Costa Neto, S. V. C; Santos, J. U. M. Restingas do litoral amazônico, estado do Pará e Amapá. Boletim do Museu Paraense Emílio Goeldi Ciências Naturais, v. 3, n. 1, p. 35-67, 2008. ANVISA - Agencia Nacional de Vigilância Sanitária. Guia para a condução de estudos não clínicos de toxicologia e segurança farmacológica necessários ao

89
desenvolvimento de medicamentos. Gerência de Avaliação de Segurança e Eficácia – GESEF. Versão 2. Brasília, 31 de janeiro de 2013. ALONSO, R. R. P.; Mendonça, M. S.; Alonso, A. A. anatomia, histoquímica e prospecção fitoquímica de folhas e raiz de Senna occidentalis (l.) link e Senna reticulata (willd.) h. s. Irwin e barneby usadas no tratamento de malária na Amazônia. Revista eletrônica de educação Dda faculdade Araguaia, 7: 337-357, 2015. APEZZATO-DA-GLORIA, B. e Carmello-Guerreiro, S.M. Anatomia Vegetal. UFV: Viçosa. 438p., 2003. ARTURO, A.; Wilkins-Rodriguez, A. A.; Escalona-Montaño, A. R; Aguirre-García M.; Becker, I.; Gutiérrez-Kobeh, L. Regulation of the expression of nitric oxide synthase by Leishmania Mexicana amastigotes in murine dendritic cells. Experimental Parasitology, v. 126, p. 426-434, 2010. AQUINO, D. S. da; Por que o uso racional de medicamentos deve ser uma prioridade? Ciência e Saúde Coletiva, v.13, p.733–736, 2008. ARAUJO, D. S. D. e Henriques, R. P. B. Análise florística das restingas do Estado do Rio de Janeiro. In: Lacerda, L. D. de et al. (orgs.). Restingas: Origem, Estrutura e Processos. Niterói, CEUFF. p. 159-194, 1984. ARAÚJO, C. S e Sobrinho, J. F. O Bioma Caatinga no Entendimento dos alunos da rede pública de ensino da cidade de Sobral, Ceará. Revista Homem. Espaço e Tempo. Universidade Estadual Vale do Acaraú –UVA. Centro de Ciências Humanas. 34-51p, 2009. AWAL, M. A.; Ainun Nahar, Shamim, M.; Bari, M. A.; Rahman, M. and Haque, M. E. Brine shrimp toxicity of leaf and seed extracts of Cassia alata Linn. and their antibacterial potency. J. Med. Sci., v. 4, n. 3, p. 188-193, 2004. ARAÚJO, N. K.; Pagnoncelli, M .G. B.; Pimentel, V. C.; Xavier, M. L. O.; Padilha, C. E. A.; Macedo, G. R.; Santos, E. S. Single-step purification of chitosanases from Bacillus cereus expanded bed chromatography, v.82, p. 291-298, 2016. BALBINO, C. A. Anti-inflamatórios: uma compreensão total. Pharmacia Brasileira nº 81, 2007. Disponível em http://www.cff.org.br/sistemas/geral/revista/pdf/131/030a045_entrevista_dr_balbino.pdf. Acesso em 04 de Janeiro de 2017. BALLOU, Ronald H.; Logística Empresarial - Transportes, Administração de Materiais, Distribuição Física. - São Paulo: Editora Atlas, 1993. BALLS, M. and Clothier, R.H. Cytotoxicity assays for intrinsic toxicity and irritancy. In: In Vitro Methods for Toxicology (ed. R.R. Watson), p. 37-52, 1992.

90
BANNERMAN, D.D. Pathogen-dependent induction of cytokines and other soluble inflammatory mediators during intramammary infection of dairy cows. J. Anim. Sci. 87:10-25, 2009. BANSAL, G. L.; Bhan, V. M. Status of research on allelopathy and future scope of work in Indian. Ind. J. Agric. Sci., v. 63, n. 12, p. 769-776, 1993. BARRESE, P., Y.; Hernández, J. M. E.; Pulpeiro, O. G. Caracterización y estudio fitoquímico de Cassia alata L. R. Cub. Plant. Med., v. 10, n. 2, p. 1-5, 2005. BARROSO, G. M. Sistemática e Angiospermas do Brasil. Edt. Ltc livros técnicos Cie. Vol 1, p. 255, 1978. BAUMANN H, Gauldie J. The acute phase response. Immunol Today.15:74–80, 1994. BECHARA, G. H. and Szabó, M.P.J. Processo inflamatório: Alterações Vasculares e Mediação Química, 2006. Disponível em: http://www.fcav.unesp.br/Home/departamentos/patologia/GERVASIOHENRIQUEBECHARA/inflam_aspectosvasculares2006.pdf. Acesso em 04 de Janeiro de 2017. BEYAERT, R and Fiers, W. Tumor necrossis factor na lymphotoxin. In: Mire-Sluis, A. R. e Thorpe, R. Cytokines, 1 ed. San Diego: Academic Press, p. 335-360, 2015. BIRT, D.F. Phytochemicals and cancer prevetion: from epidemiology to mechanism of action. J. of the American Dietetic Association, V. 106, p. 20-21, 2006. BLUMENTHAL, M.; Busse, W.R.; Goldberg, A. The complete German Commission E monographs: therapeutic guide to herbal medicines. Austin, TX: American Botanical Council, 1998. BOTZ, L.; Kocsis, B.; Nagy, S. Bioautography In: Worsfold, P.; Townshend, A.; Poole, C. Encyclopedia of Analytical Science. 2 ed. Oxford: Elsevier, 2005. BRANDÃO, M.G.L.; Cosenza, G.P., Moreira R.A.; Monte-Mór, R.L.M. Medicinal plants and other products from the Brazilian Official Pharmacopoeia. Rev Bras Farmacogn 16: 408-420, 2006. BRANDÃO, M.G.L.; Cosenza, G.P., Moreira R.A.; Monte-Mór, R.L.M. Traditional uses of American Plants from the 1st Edition of Brazilian Official Pharmacopoeia. Revista Brasileira de Farmacognosia, 19(2A): 478-487, 2009a. BRASIL. Ministério da Saúde. Política nacional de medicina natural e práticas complementares-PMNPC. Brasília, DF, 2005

91
BRASIL. Farmacopeia Brasileira, volume V. Agência Nacional de Vigilância Sanitária. Brasília: Anvisa, 524 p, 2010. BRASIL. Ministério da Saúde. Secretaria de Assistência à Saúde. Departamento de Atenção Básica. Política Nacional de Práticas Integrativas e Complementares no SUS: PNPIC-SUS. Brasília: Ministério da Saúde, 2006. BRASIL. Agência Nacional de Vigilância Sanitária. Formulário de Fitoterápicos da Farmacopeia Brasileira. Brasília: Anvisa, 2011. Disponível em <www.anvisa.gov.br>. Acesso em: 30 de Fevereiro de 2016. BRASIL. Agencia Nacional de Vigilância Sanitária, Resolução-RDC nº 26, de 13 de maio de 2014. Dispõe sobre o registro de medicamentos fitoterápicos e o registro e a notificação de produtos tradicionais fitoterápicos. Diário Oficial da União. Brasília, DF, 2014. BRASIL, Ministério da Saúde. Secretaria de Vigilância em Saúde. Boletim Epidemiológico. Monitoramento dos casos de dengue, febre de chikungunya e febre pelo vírus Zika até a Semana Epidemiológica 3, v. 47, n. 6, p. 1-7. 2016. ISSN: 2358-9450. 2016. BRASILEIRO FILHO, G., (2011). Bogliolo patologia. 8. Ed Rio de Janeiro: Guanabara Koogan. BRITO, A.S. Manual de Ensaios Toxicológicos In Vivo. Campinas: Unicamp. 122p., 1994. BRITO, F. Farmacocinética: Via de administração de medicamentos. Material didático, disciplina farmacologia. Universidade federal fluminense, s. d. Disponível em http://www.proac.uff.br/farmacomed/sites/default/files/farmacocinetica_vias_admin_f.pdf. Acesso em 20 de Setembro de 2017. BRUNETON, J. Triterpenes and Steroids. In: BRUNETON, J. Pharmacognosy, Phytochemistry, Medicinal Plants. Londres: Intercept Ltd, 1999. v.2, cap. X, p. 661- 719, 2003. BRUTON, B. D. Soilborne diseases in Cucurbitaceae: pathogen virulence and host resistence. In: CUCURBITACEAE 98, 1998. Proceedings [S.l.:s.n.]. p. 143-166, 1998. CAVALHO, L. B. Plantas daninhas. 1° edição, Lages –SC, 2013. CHANDRA, S.; MEJIA, E. G.; Polyphenolic compounds, antioxidant capacity and quinone reductase activity of an aqueous extract of Ardisia compressa in comparision to Mate (Ilex paraguaiensis) and Green (Camellia sinensis) Teas.; Journal of Agricultural and Food Chemistry; 52; 3583-3589, 2004.

92
CHAVES, M. C.; Gouveia, J.P.G.; Leite, C.A. Caracterização físico-química do suco de acerola. Rev. de Biologia e Ciências da Terra, v.4, n.2, set, p. 23-31, 2004. CHEN, L.F., Green, W.C. Shapping the nuclear action of NF-kB. Mol Cell Biol, 5, 392-401, 2004. CHOM, S. U.; Kim, Y. M. Herbicidal potencial and quantification of suspected allelochemichals from four grass crop extrats. J. Agron. Crop Sci., v. 190, p. 145-150, 2004. CHOMA, I. The use of thin-layer chromatography with direct bioautography for antimicrobial analysis. LC-GC Europe, v.18, n. 9, p. 482-488, 2005. CLIMA TEMPO. Previsão do tempo. Disponível em https://www.climatempo.com.br/. Acesso em 20 de Setembro de 2017. CLIMATE. Clima do Brasil. Disponível em https://pt.climate-data.org/country/114/. Acesso em 20 de Setembro de 2017. COLORADO, J. R.; Galeano, E. J.; Martínez, A. M. Desarrolo de La bioautografia directa como método de referencia para evaluar la actividad antimicrobiana de la gentamicina contra Escherichia coli. Vitae, Revista de la Facultad de Química Farmacéutica, v.14, n.1, p.67-71, 2007. CONAMA, Conselho nacional de meio ambiente. Resolução 07 de 23 de julho de 2002. CORDEIRO, C. H. G. Análise farmacognóstica e atividade antibacteriana de extratos vegetais empregados em formulação para a higiene bucal. Revista Brasileira de Ciências Farmacêuticas. v. 42, n. 3, p. 395-404, 2006 CORREIA, W. R. Prospecção de substâncias bioativas em Pfaffia townssendii e Pfaffia tuberosa (Gomphereneae, Amaranthaceae).Tese de doutorado. Instituto de Biologia. Jornal da Unicamp. Campinas – SP. http://www.unicamp.br/unicamp/sites/default/files/jornal/paginas/ju_620_paginacor_11_web.pdf. Acesso em 28 de Setembro de 2015. COSTA, A. F. Farmacognosia. 5.ed. Lisboa: Fundação Calouste Gulbenkia, v.3, 1994. COSTA, V.C., Triane, B.P. and Costa, N.M.C. Impactos ambientais em trilhas: agricultura × Ecoturismo - um estudo de caso na Trilha do Quilombo (PEPB - RJ). Revista Brasileira de Ecoturismo 1: 84-113, 2008. COWAN, C. Curious affection of language. Phren.J. 291p, 1981. CUTTER, E.G. Anatomia vegetal: órgãos, experimentos e interpretações. São Paulo: Roca, 336p, 1987.

93
DAMODARAN, S.; Venkataraman, S. A study on the therapeutic efficacy of Cassia alata, Linn. Leaf extract against Pityriasis versicolor. J. Ethnopharmacol., v. 42, n. 1, p. 19-23,1994. DE QUEIROZ, A.C.; De Lira, D.P.; Dias, T.D.E.L.; De Souza, E.T.; Da Matta, C.B.; De Aquino, A.B.; Silva, L.H.; DA Silva, D.J.; Mella, E.A.; Agra, MDE, F.; Filho, J.M.; De Araujo-Junior, J.X.; Santos, B.V.; Alexandre-Moreira, M.S. The antinociceptive and anti-inflammatory activities of Piptadenia stipulacea Benth. (Fabaceae). J. of Etnopharm., V.128, p. 377-83, 2010. DICKISON, W.C. Integrative Plant Anatomy. Harcourt Academic Press, Sant Diego, 2000. ENGEL, I. C.; Ferreira, R. A.; Cechinel-Filho, V.; Meyre-Silva, C. 2008. Controle de qualidade de drogas vegetais a base de Bauhinia forficata Link (Fabaceae). Rev. Bras. de Farm. 18(2), p. 258-264.
EMBRAPA. Preservação e uso da Caatinga / Embrapa Informação Tecnológica; Embrapa Semi- Árido. – Brasília, DF : Embrapa Informação Tecnológica. 39 p. : il. – (ABC da Agricultura Familiar, 16), 2012. ESAU, K. Anatomia Vegetal. Omega. Barcelona, 1985. ESTEVES, B. Avaliação da atividade anti-inflamatória e antitumoral dos extratos de Solidago chilensis, Aristochia cymbifera E Piper umbellatum. Dissertação de mestrado do curso de pós graduação em ciências biológica, Juís de Fora, MG, 2017. ESTEVES, B. dos S. Efeito da salinidade sobre as plantas. Programa de Pós-graduação em ecologia e recursos naturais. Universidade n=do Norte fluminense. Revista de ecologia brasileira, 12 (4): pag. 662-679, 2008. FAHN, A. Plant anatomy. 4.ed. Oxford: Pergamon Press, 588 p., 1990. FERREIRA, M.C.; Souza, J.R.P.; Faria, T.J. Potenciação alelopática de extratos vegetais na germinação e no crescimento inicial de picão-preto e alface. Ciência e Agrotecnologia, v. 31, n. 4, p. 1054-1060, 2007. FERREIRA, W.A.; Silva, M.E.S.T.; Paula, A.C.C.F.F.; Resende CAMB, Avaliação de Farmácia Caseira no Município de Divinópolis (MG) por Estudantes do Curso de Farmácia da Unifenas. Rev. Infarma, v.17, nº 7/9, 2005. FIGUEREDO, N.M.A. Ensinando o cuidar em Saúde Pública. São Paulo: Yendis, 2005. FILHO, A.W.; Filho, V.C.; Olinger, L.; De Souza, M.M. Quercetin: further investigation of its antinociceptive properties and mechanism of action. Archives of Pharmacal Research, V.31, p. 713-21, 2008.

94
FOLASHADE, K. O.; Omoregie, E. H. and Ochogu, A. P. Standardization of herbal medicines: a review. International Journal of Biodiversity and Conservation, v. 4, n. 3, p. 101-112, 2012. FORMAGIO, A. S. N.; Masetto, T. E.; Baldivia, D. S.; Vieira, M. C.; Zárate, N. A. H.; E Pereira, Z. V. Potencial alelopático de cinco espécies da família Annonaceae. Revista Brasileira de Biociências, v.8, p.349-354, 2010. FOYE, W.O.; Lemke, T.L. and Williams, D.A. Principles of medicinal chemistry. Baltimore: Lea e Febiger, 1995. FRAGA, C. A. M. Razões da Atividade biológica: Interações Micro-e-Biomacro-moléculas. Atividade Biológica. Cadernos Temáticos de Química Nova na Escola. Faculdade de Farmácia da Universidade Federal do Rio de Janeiro, caderno n° 3, 2011. FREIRE, A.F.M. A seqüência holocênica na plataforma continental central do Estado da Bahia – costa do cacau. Dissertação de Mestrado. Instituto de Geociências, Universidade Federal da Bahia Brasil. 172 p., 2006. FUKUSHIMA, A. R.; Azevedo, F. A. de. História da Toxicologia. Parte I – breve panorama brasileiro. Revista Intertox de Toxicologia, Risco Ambiental e Sociedade, vol.1, nº1, out, 2008. GALDINO, D. Efeito do extrato hidroalcóolico de Cassia angustifolia na gestação de camundongos. Dissertação de mestrado. Universidade Federal de Afenas – UNIFAL. Afenas, MG. 103 f, 2012. GALINA, K. J. Guazuma ulmifolia Lam. Sterculiacedae: Estudo botânico, químico e microbiológico. USP, Araraquara, 2003. GATTUSO, M. A.; Gattuso, S. J. Manual de procedimentos para analisis de drogas em polvo. Rosário: Universidade Nacional da Rosário. 48 p, 1999. GIULIETTI, A. M. Diagnóstico da vegetação nativa do bioma Caatinga. In: SILVA, J. M. C.; Tabarelli, M.; Fonseca, M. T.; Lins, L. V. (Org.). Biodiversidade da caatinga: áreas e ações prioritárias para a conservação. Brasília, DF: Ministério do Meio Ambiente, Universidade Federal de Pernambuco. 382 p, 2003. GHOSH, S., Hayden, M.S. New regulators of NFkappaB in inflammation. Nat Rev Immunol, 11, 837-848, 2008. GOBBO-NETO, L; Lopes, N. P. Plantas Medicinais: Fatores de Influência no Conteúdo de Metabólitos Secundários. Química. Nova, v.30, n.2, p.374-381, 2007. GOTTLIEB, O. R., Kaplan, M. A. Das plantas medicinais aos fármacos naturais. Ciência Hoje v.15, p.51-54, 1993.

95
GRACE, S. C.; Logan, B. A.; Adams III, W. W. Seasonal differences in foliar content of chlorogenic acid, a phenylpropanoid antioxidant, in Mahonia repens. Plant, Cell and Environment, v. 21, p. 513-521, 1998. GRANUCCI, F.; Feau S.; Angeli V.; Trottein F., and Ricciardi-Castagnoli P. Early IL-2 production by mouse dendritic cells is the result of microbial-induced priming. J. Immunol. 170, 5075–5081, 2003. GUPTA, D.; Singh, J. Flavonoid glycosides from Cassia alata. Phytochemistry, v. 30, n. 8, p. 2761- 2763, 1991. GUSMAN GS; Bittencourt AHC; Vestena S. Alelopatia de Baccharis dracunculifolia DC. sobre a germinação e desenvolvimento de espécies cultivadas. Acta Scientiarum 30: 119-125, 2008 HART, B. E.; Tapping, R. I. Genetic Diversity of Toll-Like Receptors and Immunity to M. leprae Infection, Journal of Tropical Medicine. 2012. HARTMANN T. H.; Kester DE; Davies J.R.F.T; Geneve R. Plant propagation: principles and practices. 7 ed. New York: Prentice Hall, 880p, 2001. HAYES, W. A. Principles and Methods of Toxicology 3º edição: Raven Press, p.579- 600, (1994). HEBER, D. Physicians Desk Reference for Herbal Medicines. 3oed. Los Angeles: Thomson, 2004. HENRIQUE, A. B. Botânica 1.v.1 / Anaíze Borges Henrique. – Rio de Janeiro : Fundação CECIERJ, 2008. HICKEY, L. J. Classification de la arquitetura de las hojas de dicotiledoneas. Boletin Sociedad Argentina de Botánica, Buenos Aires, v. 16, n. 1-2, p. 1-26, 1974. HOGEDAL, B. D.; Molgaard, P. HPLC analysis os seasonal and diurnal variation of iridoids in cultivars of Antirrhinum majus. Biochemical Systematics andEcology, Oxford, v,28, p.929-962, 2000. HOLETZ F.B; Pessini, G.L; Sanches, N.R; Cortez, D.A and Nakamura, C.V. Screening of some plants used in the Brazilian folk medicine for the treayment of infectious diseases. Mem I Oswaldo Cruz 97: 1027-1031, 2002. HORVÁTH, G.; Jámbor, N; Végh, A.; Boszorményi, A.; Lemberkovics, É. Héthelyi, É.; Kovács, K.; Kocsis, B., (2010). Antimicrobial activity of essential oils: the possibilities of TLC-bioautography. Flavour Fragrance Journal, v.25, p. 178-182. HOSTETTMANN, K.; Gupta, M.P.;Marston, A.; Queiroz, E. F. Manual de estratégias para el aislamiento de produtos naturals bioactivos. Bogotá:Quebecor, 2008.

96
IBRAHIM, D.; Osman, H. Antimicrobial activity of Cassia alata from Malaysia. Journal of Ethnopharmacology, Lausanne, v. 45, n. 3, p. 51-156, 1995. ICCVAM. Peer review panel report: The use of in vitro basal cytotoxicity test methods for estimating starting doses for acute oral systemic toxicity testing. Research Triangle Park: National Toxicology Program, n. 07-4519, novembro. 2006. IRWIN, H.S. e BARNEBY, R.C. The American Cassiinae. Memoirs of the New York Botanical Garden 35:1-918, 1982. IWALEWA, E.; McGaw, L. J; Naidoo, V.; Eloff, J. N; Inflammation: the foundation of diseases and disorders. A review of phytomedicines of South African origin used to treat pain and inflammatory conditions. Afr J Biotechnol. 6:2868–2885, 2007. JANEWAY, C.A; Travers P., Walport, M.; Shlomchik, M. Imunobiologia – O sistema imune na saúde e na doença, 5ª ed, Editora Artmed, 2002. JESUS, D. Atividade antimicrobiana, anti-inflamatória, citotoxicidade e genotoxicidade do extrato glicólico de Betula pendula Roth (bétula). Dissertação (Mestrado em Biopatologia Bucal) - Pós-graduação em Biopatologia Bucal - Instituto de Ciência e Tecnologia de São José dos Campos, UNESP - Univ Estadual Paulista, 2016. JUDD, W. S. et al. Pant systematics: a phylogenetic a approach. Sunderland, Mass., U.S.A. ; Sinauer Associates, 2002. JUNI, P.; Dieppe, P. Older people should NOT be prescribed “coxibs” in place of conventional NSAIDs. Age Ageing. v.33, p.100-104, 2004. KAYANO, A. C. A., Lopes, S. C., Bueno, F. G., Cabral, E. C., Souza-Neiras, W. C., Yamauchi, L. M. e Costa, F. T. In vitro and in vivo assessment of the anti-malarial activity of Caesalpinia pluviosa. Malaria Journal, 10(112): 1-11, 2011. KHAN,J., Wei,J.S., Ringner,M., Saal,L.H., Ladanyi,M., Westermann,F., Berthold,F., Schwab,M., Antonescu,C.R., Peterson,C. Classification and diagnostic prediction of cancers using gene expression profiling and artificial neural networks. Nat. Med., 7, 673-679, 2001. KHURRAM, M.; Khan, M.A.; Hameed, A.; Abbas, N.; Qayum, A.; Inayat, H., (2009). Antibacterial Activities of Dodonaea viscosa using Contact Bioautography Techinique. Molecules, v.14, p. 1332-1341. KLAASSEN, C. D.; Watkins III, I. B. Fundamentos em toxicologia. Porto Alegre: AMGH, 2012. KOES, R.E.; Quattrocchio , F. and Mol, J.N.M. The flavonoid biosynthetic pathway in plants: Function and evolution. BioEssays 16:123-132, 1994.

97
KREMER, J.M.; Dougados, M.; Emery, P.; Durez, P.; Sibilia, J.; Shergy, W.; Steinfeld. S.; Tindall. E.; Becker, J.C.; Li, T.; Nuamah, I.F.; Aranda, R.; Moreland, L.W.; Treatment of rheumatoid arthritis with the selective costimulation modulator abatacept: twelve-month results of a phase iib, doubleblind, randomized, placebo-controlled trial. Arthritis Rheum; 52(8):2263-71, 2005. KROES, B. H. The legal framework governing the quality of (traditional) herbal medicinal products in the European Union. Journal of Ethnopharmacology, p. 1-5, 2014. LACAILLE, D. M.A.; Wagner, H. A review of the biological and pharmacological activities of saponins. Phytomedicine, v.2, p. 363-386, 1996. LANTTO, T. A.; Dorman, H. J. D.; Shikov, A. N.; Pozharitskaya, O. N.; Makarov, V. G.; Tikhonov, V. P.; Hiltunen, R.; Raasmaja, A. Chemical composition, antioxidative activity and cell viability effects of a Siberian pine (Pinus sibirica Du Tour) extract. Food Chemistry, v. 112, p. 936–943, 2009. LARCHER, W. Ecofisiologia Vegetal. São Carlos: RIMA. 531 p, 2006. MURO, J.; Ramis, A.; Pastor, J.; Velarde, R.; Tarres, J.; Lavin, S. Chronic rhinitis associated with herpesviral infection in captive spur-thighed tortoises from Spain. J Wildl Dis. 34(3):487-95, 1998. LAUAND, C. A. As plantas mágicas. Composição baseada nasteorias de Paracelso, Philippus Aureolus Theophrastus Bombastus von Hohenheim, 1976. LAZZARINI, J. de A. Avaliação fitoquimica, potencial antifúngico, antioxidante e citotóxico de Pimenta pseudocaryophillus (Gomes) Landrum (Myrtaceae). Dissertação de Mestrado. Universidade Federal de Juiz de fora. Faculdade de Farmacia. Mestrado em Ciências farmacêuticas, 2015. LEAL, I.R.; Tabarelli, M. e Silva, J.M.C. Ecologia e conservação da Caatinga. Editora Universitária, Universidade Federal de Pernambuco, Recife, Brasil, 2003. LEMMENS, K. Protective and adaptive responses by antioxidant flavonoids, dissertação apresentada à escola de Nutrição, toxologia e metabolismo (NUTRIM) da Univesidade de Maastricht, 2015. Citação de HLADOVEC, J. The effect of antithrombotics in a new model of arterial thrombosis. Thromb Res, 41: 665-670, 1986b. LEWIS, G.P. Legumes of Bahia. Kew, Royal Botanic Gardens, 1987. LEWIS, G.P. Owen, P.E. Legumes of the Ilha de Maracá. Kew, Royal Botanic Gardens, 1989.

98
LEWIS, G.P. e Polhill, R.M. A situação atual da sistemática de Leguminosae neotropicais. Monographs in Systematic Botany from the Missouri Botanical Garden 68:113-129, 1998. LEWIS, G.P.; Schrire, B.D.; Mackinder, B.A. e Lock, J.M. Legumes of the World. Kew, Royal Botanic Gardens, 2005. LIMA, G.; Ippolito, A.; Nigro, F.; Salerno, M. Atempts in the biological Control of citrus mal secco (Phoma tracheiphila) using endophytic bacteria. Difesa Delle Piante, v.17, p.43-49, 1994. LIMA, A.L.; Pauletti, P.; Susin, I.; Machado-Neto, R. Flutuação das variáveis séricas em cabras e estudo comparativo da absorção de anticorpos em cabritos recém-nascidos utilizando colostro bovino e caprino. Rev. Bras. Zootec., v.38, p.2211-2217, 2009. LIMA, M. de F. Formação em preparação e administração de medicamentos. Curso de formação continua em farmácia. Farmácia Marques. Junho de 2008. Disponível em http://www.farmaciamarques.com/Imgs/content/page_87/formacao%20em%20administracao%20de%20medicamentos.pdf. Acesso em 20 de Setembro de 2017. LIU, S.; LO, C.; Shibu, M. A.; Leu, Y., Jen, B., Peng, K., Study on the Anthraquinones Separated from the Cultivation of Trichoderma harzianum Strain Th-R16 and Their Biological Activity, Journal of Agricultural and Food Chemistry., v.57, p. 7288–7292, 2009. LOMBARDO, M., Kiyota, S., Kaneko, T.M. Aspectos étnicos, biológicos e químicos de Senna (Fabaceae). Universidade de São Paulo, USP, SP – Brasil. Revista de Ciências Farmacêuticas Básica e Aplicada, pg. 9-17, SP, 2009. LOPES, C. E Amaral, F. Explorando o sistema imunológico. Faculdade PUC, Minas Gerais, 2013. Disponível em http://www1.pucminas.br/imagedb/documento/DOC_DSC_NOME_ARQUI20130912164902.pdf?PHPSESSID=79cc7fe47e3e88d4676e77be2f4224e2. Acesso em 01 de Janeiro de 2017. LORENZI, H.; Souza, H. M.; Torres, M. A. V.; Bacher, L. B. Árvores Brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 3. ed.Vol. 1. Nova Odessa/SP: Instituto Plantarum, 2000. LORENZI, H.; Matos, F.J.A. lantas Medicinais no Brasil: Nativas e exóticas cultivadas. 1ª Ed. Nova Odessa, SP: Instituto Plantarum de estudos da flora Ltda, p 544, 2002. LU, J.J.; Bao, J.L.; Wu, G.S.; Xu, W.S.; Huang, M.Q.; Chen, X.P.; Wang, Y.T. Quinones Derived from Plant Secondary Metabolites as Anti-cancer Agents. Anti-Cancer Agents in Medicinal Chemistry, v. 13, n. 3, p. 456–463, 2013.

99
LULINSKI, S.; Serwatowski, J. Bromine as the ortho-directing group in the aromatic metalation/silylation of substituted bromobenzenes. Journal of Organic Chemistry, v. 68, n. 24, p. 9384-9388, nov. 2003. LUNA, V. da S. Padronização química e botânica de Senna occidentalis Link. Dissertação de mestrado. Universidade Federal de Pernambuco. Programa de Pós graduação em Ciencias farmacêuticas, 2013. MACHADO P. R. L.; Araújo, M. I.; Carvalho, A. S.; Carvalho, L. E. M. Mecanismos de resposta imune às infecções. Educação média continuada. Anais brasileiro de dermatologia, Rio de Janeiro, p. 647-664, 2004. MACHADO, A. A.; Nakashima, T.; Silva, W. A.; Krüger , E. Contribuição ao estudo fitoquímico e citotóxico de Synadenium carinatum Boiss (EUPHORBIACEAE).Revista Eletrônica de Farmácia Vol VIII (2), 1 – 24, 2011. MACÍAS, F. A., Molinillo, J. M. G., Varela, R. M., Galindo, J. C. G. Allelopathy – a natural alternative for weed control. Pest Managemente Science, 63: 327-348, 2007. MACIEL, M. A. M.; Pinto, A. C. and Veiga, V.E. Plantas medicinais:a necessidade de estudos multidisciplinares. Química Nova 23: 429-438, 2002. MACIEL, G. K. F.; Silva, F. M. Uso Sustentável da Caatinga - Guia prático para um manejo mais sustentável na caatinga. Projeto Comunidades Rurais Sustentáveis: Manejo Florestal e Silvicultura Sustentável em General Sampaio/ Ce, 2010. Disponível em: http://www.aprece.org.br/site/downloads/CartilhaUsoSustentavelversoimpresso_1989130226 4917.pdf, acesso em 15 Agosto de 2015. MALHEIROS A.; Peres M. T.L.P. Alelopatia: Interações químicas entre espécies. In: YNES, R. A., Calixto, J. B. Plantas medicinais sob a ótica da química medicinal moderna. Chapecó: Argos. p. 503-523, 2001. MARABESI, M. A. Efeito do alto CO2 no crescimento inicial e na fisiologia da fotossíntese em plântulas Senna alata (L.) Roxb. 78 f. Tese (Mestrado em Biodiversidade Vegetal e Meio Ambiente) – Instituto de Botânica da Secretaria do Meio Ambiente, São Paulo, 2007. MARKHAN, K.R. Techniques of flavonoid identification. London: Academic Press, 1982. MARIANO SOUZA, D. P. Avaliação dos efeitos tóxicos da Senna ocidentalis em ratos. Parâmetros: bioquimicos, hematológicos, anatomapotológicos e inflamatórios. Universidade de São Paulo. Faculdade de medicina veterinária e zootecnia, 2005. MAZUR, B. AI, Kolaczkowska E, Plytycz B. Modulation of zymosan-induced peritonitis by riboflavin co-injection, pre-injection or post-injection in male Swiss mice. Life Sci. 91:1351–7, 2012.

100
MEDEIROS, E. V.; Viana, M. G.; Albuquerque, C. A.; Viana, F. A.; Silva, K. M. B. Extrato etanólico de partes de Senna alata no controle de Myrothecium roridum, agente causal do cancro-de-mirotécio. Revista Planta Daninha, v.29, p.1-7, 2011. MEFFERT, M.K.; Baltimore, D. Physiological functions for brain NF-κB. trends in Neurosciences. 28: 27–43, 2005. MELO, J.G.; Martins, J. D. G. R.; Amorim, E. L. C.; Albuquerque, U.P. Qualidade de produtos a base de plantas medicinais comercializados no Brasil: castanha-da-índia (Aesculus hippocastanum L.), capim-limão (Cymbopogon citratus (DC.) Stapf ) e centela (Centella asiatica (L.) Urban). Acta Bot Bras 21: 27-36, 2007. METCALFE, C. F.; Chalk, L. Anatomy of the Dicotyledons: Leaves, stem and wood in relation to taxonomy with notes on economic uses. Oxford: Clarendon Press. v. 1. 724 p, 1950. MIDDLETON, E.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease and cancer. Pharmacologycal Reviews, V.52, p.673-751, 2000. MOFFAT, A. C., Osselton, M. D., Widdop, B. Clarke’s Analysis of Drugs and Poisons: in pharmaceutics, body fluids and postmortem material, London, 3rd edn., 2004, 1463 – 1464. MOGILSKAI, S.; Kubacka, M.; Lażewska, D.; Więcek, M.; Lutwin, M. G.; Czochara, M. T.; Strakova, K. B.; Filipek, B. and Kieć, K. K. Aryl-1,3,5-triazine ligands of histamine H4 receptor attenuate inflammatory and nociceptive response to carrageen, zymosan and lipopolysaccharide. Inflammation Research. Received. Original research paper, 2016. MOLISCH, H. Der Einfluss einer Pflanze auf die andere-Allelopathie. Fischer, Jena, 1937. MONTANHA, F. P. e Azevedo, M. G. P. de. Administração medicamentosa: vantagens e desvantagens das diferentes vias. Revista Científica Eletrônica de Medicina Veterinária. Periódicos semestral. Ano XI – Número 20 – Janeiro de 2013. MONTEIRO, W. R.; Giulietti, A. M. and Castro M. M. Aspects of leaf structure of some species of Eriocaulon L. (Eriocaulaceae), from Serra do Cipó (Minas Gerais, Brasil). Revista. Brasileira de Botânica, 7(2): 137-147, 1985. MORAES, M.; Carmim: corante do sorvete vem de inseto, disponível em http://www.sebrae-sc.com.br/novos_destaques/oportunidade/mostrarmateria. asp?cd_noticia=9369, acesso em 10 fev. 2010.

101
MORI, H.; Isono, K.; Horiuchi ,T; Miki, T.Functional genomics of Escherichia coli in Japan. Res Microbiol 151: 121–128, 2000. MORIYAMA, H.; Iizuka, T.; Nagai, M. A stabilized flavonoid glycoside in heat-treated Cassia alata leaves and its structural elucidation. J. Pharm. Soc. Japan, v. 121, n. 11, p. 817-820, 2001. MORIYAMA, H.; Iizuka, T.; Nagai, M. HPLC quantification of kaempferol- 3-O-gentiobioside in Cassia alata. Fitoterapia, v. 74, p. 425-430, 2003. MOTA, F. S. Meteorologia Agrícola. Livraria Nobel S. A. São Paulo. 376 p., 1975
MOTTA, E. V.; Pinto, N.C.C.; Duque, A.P.N.; Mendes, R. F.; Bellozi, P.M.Q.; Scio, E. Atividades antioxidante, antinociceptiva e anti-inflamatória das folhas de Mucuna pruriens (L.) DC. Revista brasileira de plantas medicinais. vol.15 no.2, Botucatu, SP, 2013 MULLER, C.H. Allelopathy as a factor in ecological process. Vegetatio, 18:348-357, 1969. NAMBA, T.; Morita, O.; Hattori, M.; Kakiuchi, N. Studies on cardio-active crude drugs. I. Effect of coumarins on cultured myocardial cells. Planta Medica, v. 54, p. 277-282. 1988. NASCIMENTO, J. S.; Queiroz, A. A.; Queiroz, S. S. S.; Couto, A. P. Quebra de dormência e Germinação de sementes de S. alata (L.) Link em mata ciliar do sudoeste baiano. Sociedade Botânica do Brasil, p 61, 2012. NIH, Publication. Guidance document on using in vitro data to estimate in vivo starting doses for acute toxicity, n. 01-4500, ago. 2001. NOGUEIRA, L. G. Senna macranthera: constituição quimica e atividades biológicas. Dissertação apresentada ao programa de pós-graduação em Ciências biológicas. Universidade Federal de Juíz de Fora, 2009. OGUNKUNLE, A.T.J.; Ladejobi, T. A. Ethnobotanical and phytochemical studies on some species of Senna in Nigeria. African Journal of Biotechnology, v.5, n. , p.2020-2023, 2006. OKWU, D.E. Evaluation of the chemical composition of indigenous spices and flavouring agents. Global Journal Pure and Applied Science, v.7, n. , p.455-459, 2001. OLIVEIRA, F.; Akisue, G.; Akisue, M.K.; Farmacognosia. São Paulo: Ateneu, 1991. OLIVEIRA, F. C.; Albuquerque, U. P.; Fonseca K. V. S. e Hanazaki, N.; Avanços nas pesquisas etnobotânicas no Brasil. Acta Bot. Bras., 23(2): 590-605, 2009.

102
OLIVEIRA, J.D.; Silva, J.B.; Dapont, E.C.; Souza, L.M.S.; Ribeiro, A.A.L. Métodos para detecção de fungos e assepsia de sementes de Schizoobium amazonicum (Caesalpinioideae). Bioscience Journal, 28(6): 945-953, 2012. OMS - Organización Mundial De La Salud. Métodos básicos de laboratorio en parasitologia medica. Genebra, OMS. 76p, Suiza, 1992. ORDOÑEZ, M. G.; Govín, E. S.; Blanco, M. A. Actividad antimicrobiana de Senna alata L. Revista Cubana de Plantas Medicinais, v. 9, n. 1, 2004. OSTROSKY, E.A.; Mizumoto, M.K.; Lima, M.E.L.; Kaneko, T.M.; Nishikawa, S.O.; Freitas, B.R. Métodos para avaliação da atividade antimicrobiana e determinação da concentração mínima inibitória (CMI) de plantas medicinais. Brazilian Journal of Pharmacognosy, v.18, n.2, p.301- 307, 2008. PETERSON, J.; Dwyer, J. Flavonoids: Dietary occurrence and biochemical activity. Nutrition Research, v. 18, n. 12, p. 1995-2018, 1998. PLANTAMED – Plantas e Ervas Medicinais e Fitoterápicos. Disponível em: http://www.plantamed.com.br/plantaservas/especies/Senna_alata.htm Acesso em: 18 Out. 2016. PIEME, C. K. Evaluation of acute and subacute toxicities of aqueous ethanolic extract of leaves of Senna alata (L.) Roxb. (Ceasalpiniaceae). African Journal of Biotechnology Vol. 5 (3) pp. 283-289, 2006. POLHILL, R.M., Raven, P.H. e Stirton, C.H. Evolution and systematics of the Leguminosae. In Advances in legume systematics (R.M. Polhill e P.H. Raven, eds.). Royal Botanic Gardens, Kew, p.1-26. 1981. POLITI, F. A. S. Estudos farmacognósticos e avaliação de atividades biológicas de extratos obtidos das cascas pulverizadas de Endopleura uchi (huber) cuatrec. (humiriaceae). UNESP – Campus de Araraquara. 143 f., 2009. QUEIROZ, A. A.; Nascimento, J. S.; Queiroz, S. S. S.; Couto, A. P. Conhecimento popular a cerca de Senna ocidentallis (L.) Link (fedegosos) no distrito de catolezinho no sudoeste baiano. Sociedade botânica do Brasil, p 299, 2012. RAJAM, A. V.; Shanmugavalli, N.; Sunitha, C. G.; Umashankar, V. Hepatoprotective effects of Cassia tora on CCl4 induced liver damage in albino rats. Indian Journal of Science and Technology. Vol.2 no 3, 2009. RANG, H., Dale, M. Farmacologia. Rio de Janeiro, Elsevier, 2008. RANGEL, M.; Malpezzi, E. L. A; Susini, S. M.M.; Freitas, J. C. Hemolytic activity em extracts of the diatom Nitzschia. Toxicon, v. 35, p. 305-30, 1997.

103
RAO, Y.K.; Fang, S.H.; Tzeng, Y.M. Anti-inflammatory activities of flavonoids isolated from Caesalpinia pulcherrima. J. of Ethnopharm., V.100, p. 249-53, 2005. RASUL, A.; Khan, M.; Yu, B.; Ma, T.; Yang, H. Xanthoxyletin , a Coumarin Induces S Phase Arrest and Apoptosis in Human Gastric Adenocarcinoma SGC- 81 7901 Cells. Asian Pacific Journal of Cancer Prevention, v. 12, p. 1219–1223, 2011. RIBEIRO, R. A.; Vale, M. L.; Thomazzi, S. M.; Paschoalato, A. B.; Poole, S.; Ferreira, S. H.; Cunha, F. Q. Involvement of resident macrophages and mast cells in the writhing nociceptive response induced by zymosan and acetic acid in mice. Eur. J. Pharmacol. 387: 111-118, 2000 RICE, E.L. Allelopathy. 2nd ed., New York, Academic Press, 1984. RIETSCHEL, E.T.; Kirikae,T.; Schade, F. U.; Mamat, U.; Schmidt, G.; Loppnow, H. Bacterial endotoxin: molecular relationships of structure to activity and function. FASEB J. 8:217-225, 1994. RIO, R.G.W. Métodos de controle químico de amostras de própolis. Dissertação de mestrado, Universidade de São Paulo, São Paulo, 1996. RIZVI, S.J.H.; Haque, H.; Singh, U.K. and Rizvi, V. A discipline called allelopathy. In: RIZVI, S.J.H. e RIZVI, H. (Eds.) Allelopathy: Basic and applied aspects. London, Chapman e Hall, p.1-10, 1992. RIZZINI, C. T. Tratado de Fitogeografia do Brasil. v. 2. São Paulo. HUCITEC EDUSP. 374 p, 1979. ROCHA F. A. C.; Rocha, J. C. da S.; Peixoto, M. E. B.; Jancar S.; Cunha F. de Q. e Ribeiro R.de A. Efeito de inibidores da sintase de óxido nítrico na dor inflamatória articular e influxo celular da artrite induzida por zymosan em ratos. Revista Brasileira Reumatologia, v. 43, n. 4, p. 206-17, 2003. RODRIGUES, B. N.; Passini, T.; Ferreira, A. G. Research on allelopathy in Brazil. In: NARWAL, S. S. (ed.) Allelopathy update. New Hampshire: Science Publishers. p. 307-323, 1999. RODRIGUES, R. S.; Flores, A. S.; Miotto, S. T. S.; Baptista, L. M. O gênero Senna (Leguminosae, Caesalpinoideae) no Rio Grande do Sul, Brasil. Acta Botanica Brasílica, Porto Alegre, v. 19, n. 1, p. 1-16, 2005. RODRIGUES, I. M. C., Souza F., Ferreira, F.A. Estudo fitoquímico de Senna alata por duas metodologias. Planta Daninha. Viçosa-MG, v. 27, n. 3, p. 507-513, 2009. RODRIGUES, C.; Santos, A. G. dos; Garcia, B. H.; Silva, A. L. B. R.; Cortez, L. E. R. Avaliação preliminar fitoquímica da espécie Equisetum arvense

104
(cavalinha). Centro Universitário de Maringá. Anais e eventos do VII Encontro Internacional de produção científica Cesumar, Maringá, Paraná, Brasil, 2011. ROSÁRIO, M. M. T. Caracterização de xiloglucanas nativas e modificadas e efeitos sobre vias de ativação em macrófagos peritoniais de camundongos. Tese de Doutorado apresentado ao programa de pós graduação e bioquímica da Universidade Federal do Paraná, Curitiba, 2010. RUPPENTHAL, J. E. Toxicologia. Universidade Federal de Santa Maria. Colégio técnico industrial de Santa Maria. Rede e-tec Brasil, 2013. RUSSELL, W.M.S. and Burch ,R.L. Wilhelmus The Principle of Humane Experimental Technique”. 1959. Disponível em http://altweb.jhsph.edu/pubs/books/humane_exp/chap4d. Acesso em 15 de Abril de 2016. RUTALA, W.A.; Weber , D.J. Disinfection of endoscopes: reviw of new chemical sterilizants used for high-level disinfection. Infect. Control. Hosp. Epidemiol.; Thorofare, NJ, 20(1):69-76, 1999. SAHOO, N.; Manchikanti, P.; Dey, S. Herbal drugs: Standards and regulation. Fitoterapia. 81(6):462–71, 2010. SALEMME, A.; Togna, A. R.; Mastrofrancesco, A.; Cammisotto, V.; Ottaviani, M.; Bianco, A.; Venditti, A. Anti-inflammatory effects and antioxidant activity of dihydroasparagusic acid in lipopolysaccharide-activated microglial cells. Brain Res Bulletin, v.120, p.151-158, 2016. SANTOS, S.C.; Mello, J.C.P. Taninos. In: Simões, C.M.O.; Schenkel, E.P.; Gosmann, G.; Mello, J.C.P.; Mentz, L.A.; Petrovick, P.R. Farmacognosia: da planta ao medicamento. Florianópolis: UFRGS/UFSC. p.615-656, 2004. SHARAPIN, N. Fundamentos de tecnologia de Produtos Fitoterápicos. Santafé de Bogotá: Cyted, 2000. p 145-157. SCHEFFER, M.C.; Ming, L.C.; Araújo, A.J. Conservação de recursos genéticos de plantas medicinais. In: QUEIROZ, M.A.; GOEDERT, C.O.; RAMOS, S.R.R. Recursos genéticos e melhoramento de plantas para o nordeste Brasileiro. Brasília: Editora da Embrapa - Sede / DF. 24p, 1999. SCHORDERET, M. Pharmacologie. Des concepts fondamentaux aux applications theurapeutiques. Editions Slatkine Geneve, Edition Frison-Roche paris pp. 33-34, 1992. SCHULZ V.; Hansel R.; Tyler V. E. “Fitoterapia racional: um guia para as ciências da saúde”, Manole, Barueri, SP, 2002. SCORZONI, L.; Benaducci, T.; Almeida, A.M.F.; Silva, D.H.S; Bolzani, V.S.; Gianinni, M.J.S.M. The use of Standard methodology for determination of

105
antifungal activity of Natural Products against medical yeasts Candida sp. and Cryptococcus sp. Brazilian Journal of Microbiology, v. 38, p. 391-397, 2007. SEVERO, A. de A. L.; Souza, T. D. de; Rolim, L. A.; Sobrinho, J. L. S.; Medeiros, F. P M. de; Neto, P. J. R. Otimização das condições de extração de senosídeos por soluções hidroetanólicas das folhas de Senna alexandrina MILL empregando planejamento fatorial. Universidade Federal de Pernambuco/UFPE. Revista de ciências farmacêuticas Básica e aplicada. 34 (4): 603-609, 2013. SHARAPIN, N. Fundamentos de tecnologia de Produtos Fitoterápicos. Santafé de Bogotá: Cyted, p 145-157, 2000. SILVA, C. T. B da; Ishida, A. K. N.; Lemos, W. P. L.; Freire, A. de N. R. Atividade antibacteriana de extratos de Senna alata L. Roxb. Sobre a mancha bacteriana do maracujazeiro. Enciclopédia Biosfera, Centro Científico Conhecer - Goiânia, v.14 n.25; p. 182, 2017. SILVA, J. I. F.; Oliveira, R. G.; Soares, I. M.; Alvim, T. C.; Ascênio, S. D.; Martins, D. T. O. Evaluation of acute toxicity, antibacterial activity, and mode of the hidroethanolic extract of Piper umbellataum L. Journal of Ethnopharmacology, v. 151, n. 1, p. 137-43, 2014. SILVA, F. D. B. da; Sales, M. A. G.; Sá, O. R. M.; Santana, G. M.; Deus, M. do S. M.; Sousa, J. M. de C.; Ferreira, P. M. P.; Peron, A. P. Potencial citotóxico, genotóxico e citoprotetor de extratos aquosos de Caesalpinia pyramidalis Tul., Caesalpinia ferrea Mart. e Caesalpinia pulcherrima Sw. Revista Brasileira de Biociências. Porto Alegre, v. 13, n. 2, p. 101-109, 2015. SILVA, S. M. Composição florística e fitossociologia de um trecho de floresta de restinga na Ilha do Mel, Município de Paranaguá, PR. Dissertação de Mestrado. Campinas. Universidade Estadual de Campinas. 146 p., 1990. SIMÕES, C. M. O.; Schenkel, E. P.; Gosmann, G.; Mello, J. C. P.; Mentz, L. A.; Petrovick, P.R. (org.) Farmacognosia: da planta ao medicamento. 3.ed. Porto Alegre/Florianópolis: Editora da Universidade UFRGS / Editora da UFSC, 2001. SIMÕES, C. M. S.; Spitzer, V. Óleos voláteis. In: SIMÕES, C. M O.; Schenkel, E. P.; Gosmann, G.; De Mello, J. C. P.; Mentz, L. A.; Petrovick, P. R. (Orgs). Farmacognosia: da planta ao medicamento. 5ª ed, revisada e ampliada: Porto Alegre/Florianópolis. Editora da UFSC/Editora da UFRGS, 2003. SIMÕES, C. M. O.; Sptizer, V. Óleos essenciais. In: SIMÕES, C. M. O. et al. Farmacognosia da Planta ao medicamento. 5ª ed. Porto Alegre/ Florianópolis: Ed. Universidade UFRGS/EdUFSCar. p. 467- 469, 2004. SOARES, L. A. L.; Ortega, G. G.; Bassani, V. L.; Petrovick, P. R. Desenvolvimento tecnológico de soluções extrativas aquosas de Phyllanthus niruri L. (quebra-pedra) empregando planejamento fatorial. Caderno de Farmácia, v. 14, n. 1/2, p. 21-26, 1998.

106
SOLEREDER, H. Systematic anatomy of the dicotyledons. Oxford: Clarendon Press, v. 2. 1182 p., 1908. SOMCHIT, M. N.; Reezal, I.; Nur, I. E.; Mutalib, A. R. In vitro antimicrobial activity of ethanol and water extracts of Cassia alata. Journal of Ethnopharmacology, Lausanne, v. 84, p. 1-4, 2003. SORDILLO, L. M.; Redmond, M.J.; Campos, M.; Warren, L.; Babiuk, L.A. Cytokine activity in bovine mammary gland secretions during the periparturient period. Canadian Journal of Veterinary Research, v.55, p.298-301, 1991. SOUZA, S.M.; Aquino, L.C.; Milach J. A.C.; Bandeira, M.A.; Nobre, M.E.; Viana, G.S. Antiinflammatory and antiulcer properties of tannins from Myracroduon urundeuva Allemao (Anacardiaceae) in rodents. Phytotherapy Res., V. 21, p. 220-25, 2007. SOUZA, V.C. e Bortoluzzi, R.L.C. Chamaecrista.In: Forzza, R.C. et al. (eds.). Lista de espécies da flora do Brasil. Jardim Botânico do Rio de Janeiro, 2012. Disponível em <http://floradobrasil.jbrj.gov.br/2012/FB022876>. Acesso em 10 Jul, 2016. SOUZA S. A. M.; Vesz, L C.; Peixoto, D. V.; Piana, C. F. de B.; Bobrowski, V. L.; Rocha, B. H.G. Efeito de extratos aquosos de plantas medicinais nativas do Rio Grande do Sul sobre a germinação de sementes de alface. Publ. UEPG Ci. Biol. Saúde 11: 29-38, 2005. SOUZA FILHO, A.P.S. Potencialidades alelopáticas envolvendo gramíneas e leguminosas forrageiras e plantas invasoras de pastagens. Jaboticabal: UNESP, 1995. 137p. Tese (Doutorado em Zootecnia) – Universidade Estadual de São Paulo, 1995. SOUZA F, A.P.S. Alelopatia e as plantas. Belém: Embrapa. 159pp, 2006, SOUZA F, A. P. da S.; GUILHON, G. M. S. P.; SANTOS, L. S. Metodologias empregadas em estudos de avaliação da atividade alelopática em condições de laboratório – revisão crítica. Planta Daninha, Viçosa, v. 28, n. 3, p. 689-697, 2010. STAMP, N. Out of the quagmire of plant defense hypotheses. The quarterly review of biology. Department of Biological Sciences, Binghamton University Binghamton, New York 13902 US, Volume 78, No 1, 2003. SZCZEPANSKI, A. J. Allelopathy as a mean of biological controlo f water weeds. Aquatic Botany, Amsterdam, V. 3, p. 193-197, 1977. THAIZ, L. e Zeiger, E. Fisiologia vegetal. 3 ed. São Paulo: ARTMED. 792 p., 2002.

107
TATAGIBA, S. D.; Pezzopane, J. E. M.; Reis, E. F.; Dardengo, M. C. J. D.; Efgen, T. A. M. Comportamento fisiológico de dois clones de Eucalyptus na época seca e chuvosa. Cerne, v. 13, n. 2, p. 149 - 159, 2007. TATTINI, M.; Galardi, C.; Pinelli, P.; Massai, R.; Remorini, D.; Agati, G. Differential accumulation of flavonoids and hydroxycinnamates in leaves of Ligustrum vulgare under excess light and drought stress. New Phytologist.;163:547–561, 2004. TILLEY, S.L.; Coffman, T.M. and Koller, B.H. Mixed messages modulation of inflammation and immune responses by prostaglandins and thromboxanes. J Clin Invest, 2001;108:15-23. TIMOTHY, S. Y.; Lamu, F. W.; Rhoda, A.S.; Adati, R. G.; Maspalma, I. D.; Askira,M. Acute toxicity, phytochemistry and antibacterial activity of aqueous and ethanolic leaf extracts of Cassia alata Linn. International research journal of pharmacy. P. 73-76, 2012. TINCANI, A., Branch, W., Levy, R.A., Piette, J.C., Carp, H., Rai, R.S., Khamashta, M., Shoenfeld, Y. Treatment of pregnant patients with antiphospholipid syndrome, p. 524-9, 2003. TUIN, A.; Vlag, A .H. V. D., Voenen, W. A. M. M. A. V., Meijer, D. K. F., Poelstra, K. On the role and fate of LPS-dephosphorylating activity in the rat liver. American Journal of Physiology-Gastrointestinal and Liver Physiology, v. 290, p. 377-385, 2006. TUROLLA, F. A. e Lima, M. F. F. Aché: deveria um laboratório nacional fazer diferente? ESPM, Central de cases, 2007. Disponível em https://casesdesucesso.files.wordpress.com/2008/01/ache.pdf. Acesso em 15 de Junho de 2017. UNDERHILL, D. M.; Ozinsky, A.; Hajjar, A.M.; Stevens, A.; Wilson, C.B.; Bassetti, M.; Aderem, A. The Toll-like receptor 2 is recruited to macrophage phagosomes and discriminates between pathogens. Nature 401: 811-815, 1999. VALÊNCIA, A.; Bustillo, A. E.; Ossa, G. E.;Chrispeels, M.J. α-amylases of the coffee berry borer (Hypothenemus hamoei) and their inhibition by two plant amylase inhibition. Insect Biochem. Molecular. v. 30, p. 207-213, 2000. VALGAS, C.; Souza, M.; Smania, S.; Smanis, E.F.A.; Screening methods to determine antibacterial activity of natural products. Braz. J. Microbial; 38: 369-380, 2007. VEIGA J. R.; Pinto, A. C. Plantas medicinais: cura segura? Química Nova. v. 28, n. 3, p. 519-528, 2005.

108
VEIGA JUNIOR, V. F. Estudo do consumo de plantas medicinais na Região Centro-Norte do Estado do Rio de Janeiro: aceitação pelos profissionais de saúde e modo de uso pela população. Rev Bras Farmacogn 18: 308-313, 2008. VIANA, M. G.; Albuquerque, C. C.; Medeiros, E. V.; Viana, F. A.; Silva, K. M. B. Avaliação do potencial fungicida de extratos etanólicos de Senna alata contra Monosparacus cannonballus. Ciência e Agrotecnologia, v.32, p.1387-1393, 2008. VIGO, C.L.S.; Narita, E.; Gutierrez, M.M.A.; Ribeiro, P.G.F.; Marques, L.C. Caracterização farmacognóstica comparativa de Pfaffia glomerata (Spreng.) Pedersen e Hebanthe paniculata Martius - Amaranthaceae Kuntze. Revista Brasileira de Plantas Medicinais, Botucatu, v.6, n.2, p.7-19, 2004. VITORINO, R.; Fernandes, O.; Lima, L.R.; Cavalcante, R.R.L.; Martins, M.C.C.; Parente, D.M.; Cavalcante, A. A. M. C., Cavalcante, R. R. L.; Martins, M. C. C.; Parente, D. M.; Cavalcante, A. A. M. C. Avaliação da atividade antiedematogênica, antimicrobiana e mutagênica das sementes de Amburana cearensis (A. C. Smith) (Imburana-decheiro). Revista Brasileira de Plantas Medicinais, v. 15, n. 3, p. 415–422, 2013. VOLTOLINO, C. H.; Santos, M. Variações na morfoanatomia foliar de Aechmea lindenii (E. Morren) Baker var. lindenii (Bromeliaceae) sob distintas condições ambientais. Acta Botanica Brasilica, v. 25, p. 2-10, 2011. WANG, X.; Hoekstra, M.F.; DeMaggio, A.J.; Dhillon. N.; Vancura, A.; Kuret, J.; Johnston, G.C.; Singer, R.A. Prenylated isoforms of yeast casein kinase I, including the novel Yck3p, suppress the gcs1 blockage of cell proliferation from stationary phase. Saccharomyces genome database – SGD. Disponível em https://www.yeastgenome.org/reference/S000051920. Acesso em 08 Abril de 2016. WAGNER, H.; Bladt, S. Plant drug analysis - A thin layer cromatography atlas. 2.ed. Berlim: Springer, 384 p, 1996. WAKSMUNDZKA, H., Sherma, M. J., Kowalska, T. Thin Layer Chromatography in Phytochemistry. Chromatographic science series, volume 99, 2008. WANNMACHER, L. Inibidores seletivos de cicloxigenase-2 revisitados um ano depois. Uso racional de medicamentos. v.2, n.2, p. 1-6, 2005. WARDLAW, G.M.; Insel, P.M.; Seyler, M.F. Contemporary Nutrition: Issues and Insights, 2ª. ed., Mcgraw-Hill Book, 602p., 1994. WATSON, L. An automated system of generic description for Caesalpinioideae, and its application to classification and key-making. In: POLHILL, R. M.; RAVEN, P. H. Adv. Leg. Syst., v. 1, p. 65-80, 1981. WHO - World Health Organization. Quality control methods for medicinal plant material. WHO: Geneva. 115p., 1992.

109
WHO - World Health Organization. Regional office for the western pacific research guidelines for evaluating the safety and efficacy of herbal medicines. Manila, 1998. ZACCHINO, S. Estratégia para a descoberta de novos agentes antifúngicos. In: YUNES, R.A. E Calixto, J.B. Plantas medicinais sob a ótica da química medicinal moderna. Chapecó: Argos. p. 435-479, 2001. ZANIN, J. B., Carvalho, B. A., Martineli, P. S., Santos, M. H., Lago, J. H., Sartorelli, P., Viegas JR, C. e Soares, M. G. The genus Caesalpinia (Caesalpiniaceae): phytochemical and pharmalogical characteristics. Molecules, 17: 7887-7902, 2012. ZHAO, L.; Li, T.; Wang, Y.; Pan, Y.; Ning, H. Elevated plasma osteopontin level is predictive of cirrhosis in patients with hepatitis B infection. Int J Clin Pract 62: 1056–1062, 2008. ZHOU, J.; Liao, M.; Ueda, M.; Gong, H.; Xuan, X. and Fujisaki, K. Sequence characterization and expression patterns of two defensin-like antimicrobial peptides from the tick Haemaphysalis longicornis. Peptides 28(6): 1304-10, 2007. ZUANAZZI, J. A.; Montanha, J. A. Flavonoides. In: SIMÕES, et al. (org.). Farmacognosia: da planta ao medicamento. 6.ed. Porto Alegre: Editora da Universidade UFRGS, 2010.
Related Documents











