Re-evaluation of physical interaction between plant peroxisomes and other organelles using live-cell imaging techniques FA Kazusato Oikawa 1† , Makoto Hayashi 2† , Yasuko Hayashi 3 and Mikio Nishimura 4 * 1. Biomacromolecules Research Team, RIKEN Center for Sustainable Resource Science, 2-1 Hirosawa, Wako, Saitama 351-0198, Japan 2. Department of Bioscience, Nagahama Institute of Bio-Science and Technology, 1266 Tamura-Cho, Nagahama 526-0829, Japan 3. Department of Biology, Faculty of science, Niigata University, Niigata 950-2181, Japan 4. Department of Cell Biology, National Institute for Basic Biology, Okazaki 444-8585, Japan † These authors contributed equally to the study. doi: 10.1111/jipb.12805 Mikio Nishimura *Correspondence: [email protected] Abstract The dynamic behavior of organelles is essen- tial for plant survival under various environmental conditions. Plant organelles, with various functions, migrate along actin filaments and contact other types of organelles, leading to physical interactions at a specific site called the membrane contact site. Recent studies have revealed the importance of physical interactions in maintaining efficient metabolite flow between organelles. In this review, we first summarize peroxisome function under different environmental conditions and growth stages to understand organelle interactions. We then discuss current knowledge regarding the interactions between peroxisome and other organelles, i.e., the oil bodies, chloroplast, and mitochondria from the perspective of metabolic and physiological regulation, with reference to various organelle interactions and techniques for estimating organelle interactions occurring in plant cells. Edited by: Jianping Hu, Michigan State University, USA Received Nov. 1, 2018; Accepted Mar. 18, 2019; Online on Mar. 27, 2019 FA: Free Access INTRODUCTION Organelles contain a series of enzymes for metabolic pathways to perform their unique function. However, some metabolic pathways are known to have metab- olites in common that exist in two or more organelles. To maintain such metabolic pathways, the metabolic intermediates produced should be transferred between organelles, and the associated mechanisms have been frequently discussed in relation to subcellular organelle positioning (Schrader et al. 2015; Eisenberg-Bord et al. 2016). For example, electron microscopy (EM) has revealed that organelles participating in the same metabolic pathway often stick together (Trelease et al. 1971; Frederick et al. 1975; Tolbert and Essner 1981; Nishimura et al. 1986; Hayashi et al. 2001). In live Arabidopsis thaliana cells, peroxisomes, chloroplasts, and mitochondria often interact directly (Figure 1). Recent advances in biochemical, biophysical, and bioimaging techniques have provided evidence of direct interaction between two organelles at the membrane contact site (MCS) in yeast, plant, and mammalian cells (Prinz 2014; Schrader et al. 2015; Eisenberg-Bord et al. 2016; Perez-Sancho et al. 2016; Rodriguez-Serrano et al. 2016). These findings imply that metabolites are delivered to organelles via the MCS. Plant cells contain organelles that differ from those of yeast and mammalian cells, such as chloroplasts and a large vacuole (see The Plant Organelles World (http://podb.nibb.ac.jp/Organellome/PODBworld/en/) © 2019 Institute of Botany, Chinese Academy of Sciences JIPB Journal of Integrative Plant Biology www.jipb.net July 2019 | Volume 61 | Issue 7 | 836-852 Free Access Invited Expert Review

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Re-evaluation of physical interaction betweenplant peroxisomes and other organelles usinglive-cell imaging techniquesFA
Kazusato Oikawa1†, Makoto Hayashi2†, Yasuko Hayashi3 and Mikio Nishimura4*
1. Biomacromolecules Research Team, RIKEN Center for Sustainable Resource Science, 2-1 Hirosawa, Wako, Saitama 351-0198, Japan2. Department of Bioscience, Nagahama Institute of Bio-Science and Technology, 1266 Tamura-Cho, Nagahama 526-0829, Japan3. Department of Biology, Faculty of science, Niigata University, Niigata 950-2181, Japan4. Department of Cell Biology, National Institute for Basic Biology, Okazaki 444-8585, Japan†These authors contributed equally to the study.doi: 10.1111/jipb.12805
Mikio Nishimura
*Correspondence:[email protected]
Abstract The dynamic behavior of organelles is essen-tial for plant survival under various environmentalconditions. Plant organelles, with various functions,migrate along actin filaments and contact other typesof organelles, leading to physical interactions at a specific
site called the membrane contact site. Recent studieshave revealed the importance of physical interactions inmaintaining efficient metabolite flow between organelles.In this review, we first summarize peroxisome functionunder different environmental conditions and growthstages to understand organelle interactions. We thendiscuss current knowledge regarding the interactionsbetween peroxisome and other organelles, i.e., the oilbodies, chloroplast, andmitochondria from the perspectiveofmetabolic and physiological regulation, with reference tovariousorganelle interactions and techniques for estimatingorganelle interactions occurring in plant cells.
Edited by: Jianping Hu, Michigan State University, USAReceived Nov. 1, 2018; AcceptedMar. 18, 2019; Online onMar. 27,2019
FA: Free Access
INTRODUCTION
Organelles contain a series of enzymes for metabolicpathways to perform their unique function. However,some metabolic pathways are known to have metab-olites in common that exist in two or more organelles.To maintain such metabolic pathways, the metabolicintermediates produced should be transferred betweenorganelles, and the associated mechanisms have beenfrequently discussed in relation to subcellular organellepositioning (Schrader et al. 2015; Eisenberg-Bord et al.2016). For example, electron microscopy (EM) hasrevealed that organelles participating in the samemetabolic pathway often stick together (Trelease et al.1971; Frederick et al. 1975; Tolbert and Essner 1981;
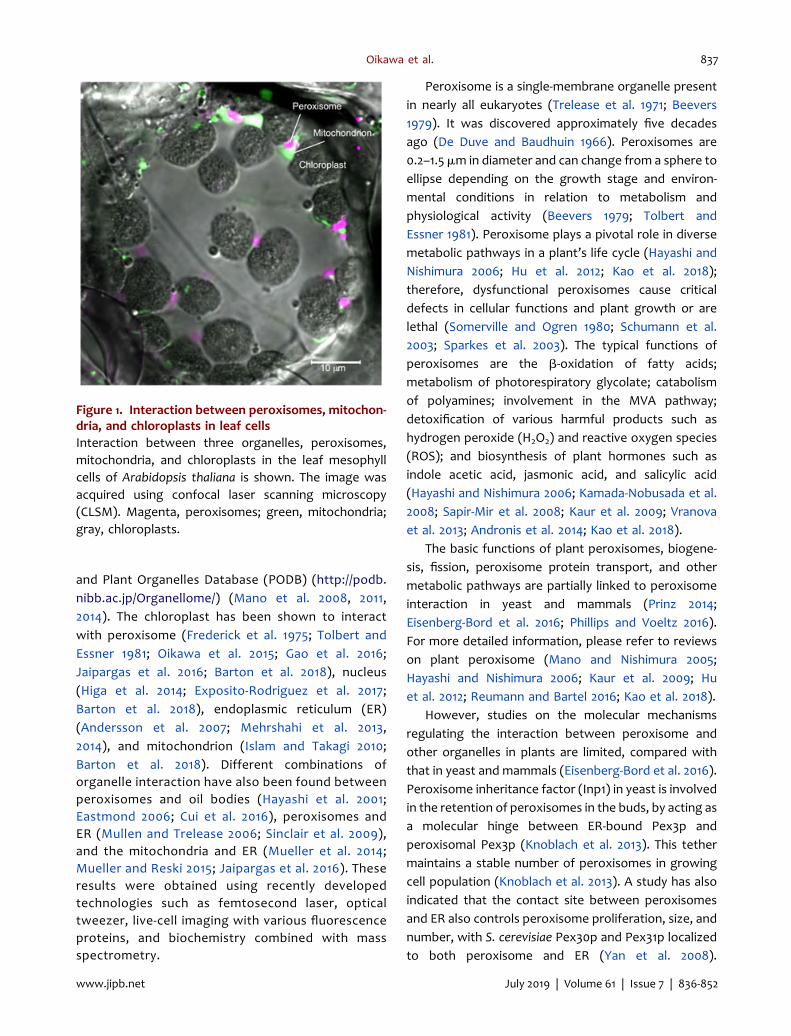
Nishimura et al. 1986; Hayashi et al. 2001). In liveArabidopsis thaliana cells, peroxisomes, chloroplasts,and mitochondria often interact directly (Figure 1).
Recent advances in biochemical, biophysical, andbioimaging techniques have provided evidence of directinteraction between two organelles at the membranecontact site (MCS) in yeast, plant, and mammalian cells(Prinz 2014; Schrader et al. 2015; Eisenberg-Bord et al.2016; Perez-Sancho et al. 2016; Rodriguez-Serrano et al.2016). These findings imply that metabolites aredelivered to organelles via the MCS.
Plant cells contain organelles that differ from thoseof yeast and mammalian cells, such as chloroplastsand a large vacuole (see The Plant Organelles World(http://podb.nibb.ac.jp/Organellome/PODBworld/en/)
© 2019 Institute of Botany, Chinese Academy of Sciences
JIPB Journal of IntegrativePlant Biology
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852
Free
Access
InvitedEx
pert
Rev
iew
and Plant Organelles Database (PODB) (http://podb.nibb.ac.jp/Organellome/) (Mano et al. 2008, 2011,2014). The chloroplast has been shown to interactwith peroxisome (Frederick et al. 1975; Tolbert andEssner 1981; Oikawa et al. 2015; Gao et al. 2016;Jaipargas et al. 2016; Barton et al. 2018), nucleus(Higa et al. 2014; Exposito-Rodriguez et al. 2017;Barton et al. 2018), endoplasmic reticulum (ER)(Andersson et al. 2007; Mehrshahi et al. 2013,2014), and mitochondrion (Islam and Takagi 2010;Barton et al. 2018). Different combinations oforganelle interaction have also been found betweenperoxisomes and oil bodies (Hayashi et al. 2001;Eastmond 2006; Cui et al. 2016), peroxisomes andER (Mullen and Trelease 2006; Sinclair et al. 2009),and the mitochondria and ER (Mueller et al. 2014;Mueller and Reski 2015; Jaipargas et al. 2016). Theseresults were obtained using recently developedtechnologies such as femtosecond laser, opticaltweezer, live-cell imaging with various fluorescenceproteins, and biochemistry combined with massspectrometry.
Peroxisome is a single-membrane organelle presentin nearly all eukaryotes (Trelease et al. 1971; Beevers1979). It was discovered approximately five decadesago (De Duve and Baudhuin 1966). Peroxisomes are0.2–1.5mm in diameter and can change from a sphere toellipse depending on the growth stage and environ-mental conditions in relation to metabolism andphysiological activity (Beevers 1979; Tolbert andEssner 1981). Peroxisome plays a pivotal role in diversemetabolic pathways in a plant’s life cycle (Hayashi andNishimura 2006; Hu et al. 2012; Kao et al. 2018);therefore, dysfunctional peroxisomes cause criticaldefects in cellular functions and plant growth or arelethal (Somerville and Ogren 1980; Schumann et al.2003; Sparkes et al. 2003). The typical functions ofperoxisomes are the b-oxidation of fatty acids;metabolism of photorespiratory glycolate; catabolismof polyamines; involvement in the MVA pathway;detoxification of various harmful products such ashydrogen peroxide (H2O2) and reactive oxygen species(ROS); and biosynthesis of plant hormones such asindole acetic acid, jasmonic acid, and salicylic acid(Hayashi and Nishimura 2006; Kamada-Nobusada et al.2008; Sapir-Mir et al. 2008; Kaur et al. 2009; Vranovaet al. 2013; Andronis et al. 2014; Kao et al. 2018).
The basic functions of plant peroxisomes, biogene-sis, fission, peroxisome protein transport, and othermetabolic pathways are partially linked to peroxisomeinteraction in yeast and mammals (Prinz 2014;Eisenberg-Bord et al. 2016; Phillips and Voeltz 2016).For more detailed information, please refer to reviewson plant peroxisome (Mano and Nishimura 2005;Hayashi and Nishimura 2006; Kaur et al. 2009; Huet al. 2012; Reumann and Bartel 2016; Kao et al. 2018).
However, studies on the molecular mechanismsregulating the interaction between peroxisome andother organelles in plants are limited, compared withthat in yeast and mammals (Eisenberg-Bord et al. 2016).Peroxisome inheritance factor (Inp1) in yeast is involvedin the retention of peroxisomes in the buds, by acting asa molecular hinge between ER-bound Pex3p andperoxisomal Pex3p (Knoblach et al. 2013). This tethermaintains a stable number of peroxisomes in growingcell population (Knoblach et al. 2013). A study has alsoindicated that the contact site between peroxisomesand ER also controls peroxisome proliferation, size, andnumber, with S. cerevisiae Pex30p and Pex31p localizedto both peroxisome and ER (Yan et al. 2008).
Figure 1. Interaction between peroxisomes, mitochon-dria, and chloroplasts in leaf cellsInteraction between three organelles, peroxisomes,mitochondria, and chloroplasts in the leaf mesophyllcells of Arabidopsis thaliana is shown. The image wasacquired using confocal laser scanning microscopy(CLSM). Magenta, peroxisomes; green, mitochondria;gray, chloroplasts.
2 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
837

Peroxisomes have been known to interact strongly withthe mitochondria for division and biogenesis, and forlipid degradation (metabolic cooperation) (Shai et al.2016). Recently, two peroxisome-mitochondria tether-ing factors, Fzo1 and Pex34, have been identified by thesplit Venus complementation assay in S. cerevisiae (Shaiet al. 2018). They showed that Pex34 is a potentialtether for peroxisome and mitochondrion responsiblefor the ß-oxidation of fatty acids (Shai et al. 2018). Atethering complex mediating membrane associationsbetween peroxisome and ER in human cells has beenrecently identified in the two groups using a combina-tion of pull-down studies and mass spectrometry(Costello et al. 2017; Hua et al. 2017). The authorsdemonstrated that the tether, peroxisome membraneacyl-CoA binding domain containing five (ACBD5) andresident ER protein vesicle-associated membraneproteins A and B (VAPA and VAPB), is necessary forthe growth of peroxisomes, synthesis of plasmalogenphospholipids, and maintenance of cellular cholesterollevels (Hua et al. 2017).
In this review, we highlight current knowledge on
the interaction between peroxisome and other organ-
elles in plant cells, focusing on the functions of
peroxisomes and implications of the interaction in
different environments and at different growth stages.
FUNCTIONAL DIFFERENTIATION OFPLANT PEROXISOMES
Plant peroxisomes are remarkably plastic, and they areknown to differentiate based on function depending onthe tissue, organ, growth stage, and environmentalcondition (Nishimura et al. 1996; Kaur et al. 2009). Thereare at least four different classes of peroxisomes,namely, glyoxysomes, and leaf, root, and unspecializedperoxisomes (Johnson and Olsen 2001; Kamada et al.2003; Hayashi and Nishimura 2006).
These different peroxisomes are known to intercon-vert during certain cellular processes such as greeningof cotyledons and senescence of leaves (Hayashi andNishimura 2006; Kaur et al. 2009; Kao et al. 2018). Forexample, glyoxysome and leaf peroxisome flexiblyinterconvert their functions in response to environmen-tal condition, light, and senescence in cotyledonary cells(Tolbert 1981; Titus and Becker 1985; Nishimura et al.1986, 1993; De Bellis et al. 1991a, 1991b; Hayashi et al.2000; Goto-Yamada et al. 2015). Furthermore,
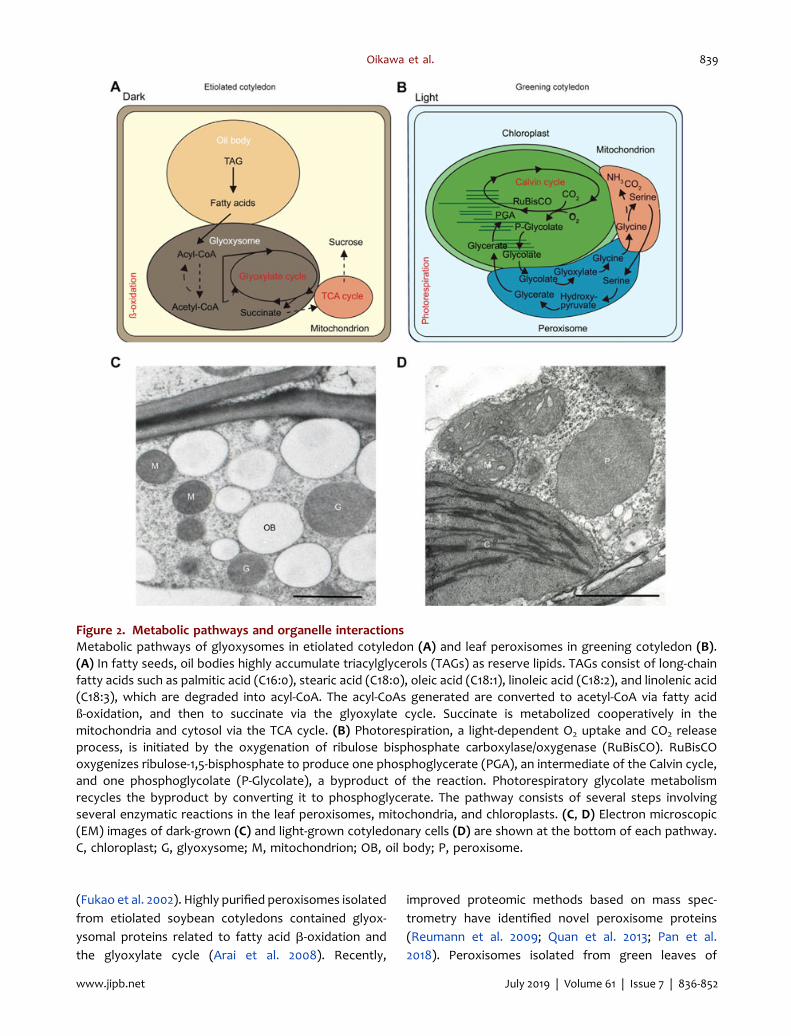
glyoxysomes first appear in the cells of etiolatedcotyledon, and then convert into leaf peroxisomesduring the greening process of seedlings. The transitionchanges the energy source necessary for growth fromseed-reserved lipids to sugars produced by photosyn-thesis. Glyoxysome is involved in lipid metabolism viafatty acid ß-oxidation and the glyoxylate cycle (Trelease1984; Baker et al. 2006; Hayashi and Nishimura 2006;Graham 2008) (Figure 2A, C). Succinate is metabolizedcooperatively in the mitochondria and cytosol(Figure 2A, C). The leaf peroxisomes are found in thecells of photosynthetic organs such as the greencotyledon and leaf, and they play a pivotal role inphotorespiration (Ogren and Bowes 1971; Tolbert 1982;Somerville 2001; Hayashi and Nishimura 2006; Kaur et al.2009; Peterhansel et al. 2010; Reumann and Bartel 2016;Kao et al. 2018) (Figure 2B, D). The pathway consists ofseveral steps with several enzymatic reactions inthe leaf peroxisome, mitochondrion, and chloroplast(Figure 2B, D). Reverse transition from leaf peroxisometo glyoxysome also occurs during the senescence ofcotyledonary cells, leaves, and petals (De Bellis et al.1991b; Pistelli et al. 1996). These phenomena are definedas functional transitions (Tolbert 1981; Titus and Becker1985; Nishimura et al. 1986; Hayashi et al. 2000; Goto-Yamada et al. 2015).
During the transition, enzymes required only for theformer organelles are degraded and those necessary forthe subsequent organelles are newly synthesized(Nishimura et al. 1986; Hayashi et al. 2000; Goto-Yamada et al. 2015). Furthermore, transcription, trans-port, and degradation of the matrix proteins ofperoxisomes are well organized (Nishimura et al.1986; Goto-Yamada et al. 2015). Indeed, after lightirradiation of etiolated seedlings, the mRNA level of theglyoxysomal enzymes, 3-ketoacyl-CoA thiolase (Katoet al. 1996) and isocitrate lyase (Mano et al. 1996),decreases and that of leaf peroxisomal enzymes,hydroxypyruvate reductase, serine-glyoxylate amino-transferases (Hondred et al. 1987), and glycolateoxidase (Tsugeki et al. 1993), increases.
Proteomic approaches have been used to identifyundefined-peroxisome proteins (Fukao et al. 2002,2003; Arai et al. 2008; Reumann et al. 2009; Pan et al.2018). Peroxisomes isolated from the greening cotyle-dons of Arabidopsis contained the leaf peroxisomeproteins related to the glycolate pathway, hydrogenperoxide scavenging pathway, and protein kinases
Physical interaction between plant organelles 3
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
838
(Fukao et al. 2002). Highly purified peroxisomes isolatedfrom etiolated soybean cotyledons contained glyox-ysomal proteins related to fatty acid b-oxidation andthe glyoxylate cycle (Arai et al. 2008). Recently,
improved proteomic methods based on mass spec-trometry have identified novel peroxisome proteins(Reumann et al. 2009; Quan et al. 2013; Pan et al.2018). Peroxisomes isolated from green leaves of
Figure 2. Metabolic pathways and organelle interactionsMetabolic pathways of glyoxysomes in etiolated cotyledon (A) and leaf peroxisomes in greening cotyledon (B).(A) In fatty seeds, oil bodies highly accumulate triacylglycerols (TAGs) as reserve lipids. TAGs consist of long-chainfatty acids such as palmitic acid (C16:0), stearic acid (C18:0), oleic acid (C18:1), linoleic acid (C18:2), and linolenic acid(C18:3), which are degraded into acyl-CoA. The acyl-CoAs generated are converted to acetyl-CoA via fatty acidß-oxidation, and then to succinate via the glyoxylate cycle. Succinate is metabolized cooperatively in themitochondria and cytosol via the TCA cycle. (B) Photorespiration, a light-dependent O2 uptake and CO2 releaseprocess, is initiated by the oxygenation of ribulose bisphosphate carboxylase/oxygenase (RuBisCO). RuBisCOoxygenizes ribulose-1,5-bisphosphate to produce one phosphoglycerate (PGA), an intermediate of the Calvin cycle,and one phosphoglycolate (P-Glycolate), a byproduct of the reaction. Photorespiratory glycolate metabolismrecycles the byproduct by converting it to phosphoglycerate. The pathway consists of several steps involvingseveral enzymatic reactions in the leaf peroxisomes, mitochondria, and chloroplasts. (C, D) Electron microscopic(EM) images of dark-grown (C) and light-grown cotyledonary cells (D) are shown at the bottom of each pathway.C, chloroplast; G, glyoxysome; M, mitochondrion; OB, oil body; P, peroxisome.
4 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
839

four-week-old Arabidopsis contained a high proportionof photorespiratory proteins (Reumann et al. 2009).Furthermore, peroxisomes from dark-treated senes-cent leaf contained higher amount of glyoxysomalproteins and lower amount of leaf peroxisome proteins,such as photorespiration-related proteins (Pan et al.2018). These studies have shown different patternsof protein accumulation among glyoxysome, leafperoxisome, and peroxisome in senescence leaf;however, the proteins identified by proteome analysisof peroxisomes from etiolated seedling are largelyoverlapping in glyoxysomes and leaf peroxisomes(Quan et al. 2013).
The molecular mechanisms of functional transitionhave been studied extensively; however, the detailedmechanisms are still unclear. The biochemical approachand immunoelectronmicroscopyusingantibodiesagainstglyoxysomal and leaf peroxisomal enzymes duringtransition have been used. Two opposing hypotheseshave been proposed (Kagawa et al. 1975; Beever 1979),and one of them is the “one population model”. In thismodel, glyoxysomes are directly transformed into leafperoxisomes during greening of cotyledons. The processis accompanied by the import of newly synthesized leafperoxisomal enzymes and elimination of pre-existingglyoxysomal enzymes in glyoxysomes at the transitionstage. Another hypothesis is the “twopopulationmodel”inwhichglyoxysomes aredegradedand leafperoxisomesare synthesized as new organelles.
To demonstrate the specific hypothesis operatingduring the transition of peroxisomes, immunoelectronmicroscopic analysis using antibodies against variousglyoxysomal and leaf peroxisomal enzymes has beenperformed (Titus and Becker 1985; Nishimura et al. 1986;Nishimura et al. 1993). Both glyoxysomal and leafperoxisomal enzymes coexist in all peroxisomes atthe transition stage. The results provide strong evidencethat the one population model is likely involved in theprocess.
However, it has been shown that major glyoxysomalmembrane proteins, initially decrease in content, andthen disappear during the transition in greeningpumpkin cotyledons as glyoxysome-specific enzymesdo (Yamaguchi et al. 1995; Fukao et al. 2001). Thisfinding is apparently against the one population modeland suggests that glyoxysome-specific enzymes andglyoxysommal membrane proteins degrade as a wholeduring the transition.
The degradation of the matrix proteins by proteasessuch as LON protease 2 (Lon2), DEG15, and M16
protease is involved in peroxisome transition accompa-nied by oxidative damage (Lingard and Bartel 2009;
Lingard et al. 2009; Goto-Yamada et al. 2014, 2015).
Against the one and two population models, acompromised model has been proposed. It has been
recently shown that a double mutant of Lon2 and ATG2,but not single mutants, blocked the degradation of
glyoxysomes during the transition (Goto-Yamada et al.
2014, 2015). This indicates that both degradation ofglyoxysome-specific enzymes by Lon2 and degradation
of whole glyoxysomes by autophagy are necessary and
that they mediate the degradation of glyoxysomesunder transition conditions (Farmer et al. 2013; Goto-
Yamada et al. 2014, 2015). Two degradation systems
interact with each other, and the chaperone domain ofLON2 interferes with and represses autophagy during
the transition (Goto-Yamada et al. 2014). Elucidation of
the mechanism by which peroxisome transition isorganized and regulated with these two degradation
systems at the molecular level is encouraged.
PLANT PEROXISOME INTERACTION WITHOTHER ORGANELLES
Glyoxysome-oil body interaction during post-germinative growthIn etiolated seedlings of oil seeds, direct contact
between glyoxysome and oil body has been demon-
strated based on EM observations (Trelease et al. 1971;
Frederick et al. 1975; Nishimura et al. 1986; Hayashi et al.
2001). The contact was thought to promote efficient
flux of metabolic intermediates between glyoxysomes,
oil bodies, and mitochondria, which are the organellesinvolved in the same metabolic pathway, i.e., lipiddegradation (Figure 2A, C). Isolation of mutants with adefect in lipid degradation clearly demonstrated theimportance of organelle interaction (Cui et al. 2016).
Furthermore, ped1 is a mutant with a defective3-ketoacyl-CoA thiolase gene encoding the enzyme thatcatalyzes the last step of fatty acid b-oxidation (Hayashiet al. 1998); therefore, it cannot degrade seed-reservedlipid during post-germinative seedling growth. Thedefect enlarged glyoxysomes and invaginated itsmembrane to form dendritic tubules containing smallvesicles at the MCS between glyoxysomes and oil
Physical interaction between plant organelles 5
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
840

bodies (Hayashi et al. 2001). It implied that directphysical interaction between glyoxysome and oil bodyplays an important role in the flux of metabolicintermediates produced during lipid degradation.
The degradation of seed reserved lipid is initiated by
the triacylglycerol lipase, sugar-dependent 1 (SDP1),
which hydrolyzes triacylglycerol into diacylglycerol and
one fatty acid in the oil body membrane (Eastmond
2006; Kelly et al. 2011; Cui et al. 2016). SDP1 in the oil
body is transported to glyoxysome, extending the
tubular structure to the oil body with a retromer
complex (Thazar-Poulot et al. 2015). Sucrose-dependentinteractions between glyoxysomes and oil bodies havebeen observed in seedlings with an sdp1-deficientbackground (Cui et al. 2016). In the seedling, multipleglyoxysomes and oil bodies combined to form aggre-gates under low sucrose concentrations, whereasglyoxysomes could not interact with the oil bodiesunder high sucrose concentrations (Cui et al. 2016). Theresult suggests that sucrose negatively regulates lipiddegradation by dissociating the interaction betweenglyoxysomes and oil bodies.
Accumulating evidence has emphasized the impor-tance of glyoxysome-oil body interaction in lipidmetabolism; however, factors involved in the interac-tion should be determined. SPD1 and monoacylglycerollipase 8 (MAGL8) are located in the oil body membrane,and PED3/CTS/PXA1, a member of the ABC transporterfamily, is located in the glyoxysomemembrane (Zolmanet al. 2001; Footitt et al. 2002; Hayashi et al. 2002),indicating that fatty acid transport occurs at contactsites, where these components accumulate. Furtherstudies should elucidate the involvement of SPD1,MAGL8, and PED3/CTS/PXA1, in the direct physicalinteraction between the oil bodies and peroxisomes.
Leaf peroxisome-mitochondrion-chloroplastinteraction in the photosynthetic tissueIn the cells of photosynthetic tissues, direct contact
between the chloroplast, leaf peroxisome, and mito-chondrion has been detected by EM (Trelease et al. 1971;
Frederick et al. 1975; Titus and Becker 1985; Nishimuraet al. 1986). These three organelles are involved in
the same metabolic pathway, i.e., photorespiratoryglycolate metabolism (Figure 2B, D). This observation
suggests that the contact between these organelles isinvolved in efficient metabolite flow during photorespi-
ration (Trelease et al. 1971; Beevers 1979). However, this
is controversial, because fixed samples i.e., dead cells,were used for EM.
Recent studies have confirmed that the contact isindeed operational in live A. thaliana cells (Oikawa et al.2015; Eisenberg-Bord et al. 2016; Gao et al. 2016), usingvarious microscopic techniques such as fluorescenceand CLSM as live imaging tools (Jedd and Chua 2002;Mano et al. 2002; Mathur et al. 2002). Under lightcondition, peroxisomes extend to an elliptical shapealong the chloroplast, whereas they are spherical underdark condition (Oikawa et al. 2015). Similarly, this light-dependent morphological change is also applicable tothe mitochondrion (Oikawa et al. 2015). Coincidentally,the photorespiration enzymes, catalase and glycolateoxidase, are activated after light perception, indicatinga connection with enhanced organelle interactions(Oikawa et al. 2015). Similar peroxisome elongationunder light condition has been reported to occur beforeperoxisome proliferation via phytochrome A signaling(Desai and Hu 2008). However, it might be differentfrom photosynthesis-dependent peroxisome elonga-tion for enhancing the interaction with chloroplast(Oikawa et al. 2015). In the light-dependent interaction,peroxisomes and mitochondria accumulate onthe chloroplasts via transportation by actin filaments(F-actin), which connect chloroplasts (Mano et al.2002; Oikawa et al. 2015). Mobile peroxisomes onthe chloroplast, with actively changing morphology,capture the mitochondria at high frequency under lightcondition (Oikawa et al. 2015).
The femtosecond laser and optical tweezer analyseshave suggested the existence of tethering factorsbetween peroxisome and chloroplast at the MCS(Oikawa et al. 2015; Gao et al. 2016; Sparkes 2016).However, the components that are involved in organ-elle interactions remain to be elucidated. One possiblecandidate is PEX10, a C3HC4 zinc RING-finger peroxi-somal membrane protein (Schumann et al. 2007). It isessential for peroxisome biogenesis, and the loss-of-function mutants were embryo lethal at the heart stage(Schumann et al. 2003; Sparkes et al. 2003; Kamigakiet al. 2009). The zinc finger motif is related to protein-protein interaction (Gamsjaeger et al. 2007). Arabidop-sis overexpressing PEX10 with a dysfunctional zincfinger in a wild-type background, used as a dominantnegative, showed small-clustered peroxisomes that lostthe interaction with chloroplast (Schumann et al. 2007),showing defects in plant growth similar to that of
6 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
841

photorespiratory mutants. However, further analysesare necessary to examine whether PEX10 is atethering factor that directly connects peroxisomeand chloroplast.
Differentiation of plant organelle interaction duringperoxisome transitionAs described in above the section, glyoxysomes interactwith the oil bodies, whereas the leaf peroxisomespreferentially interact with chloroplasts and mitochon-dria (Trelease et al. 1971; Frederick et al. 1975; Nishimuraet al. 1986; Hayashi et al. 2001) (Figure 2). Here, we raisethe question of when does the transition from the
peroxisome-oil body interaction to the peroxisome-
chloroplast interaction occur under light-dependent
conditions (Figure 3). We have provided EM images of
the oil body and peroxisome in cells of 5-d-old dark-
grown etiolated cotyledon, and peroxisome and
chloroplast in cells of 5-d-old light-grown green
cotyledon (Figure 4A). There are several oil bodies in
cells of etiolated cotyledon (Figure 4A, left column), as
well as peroxisomes. These peroxisomes are close to
the oil body. On the contrary, the chloroplasts develop
together with peroxisomes and mitochondria closely in
cells of greening cotyledon (Figure 4A, right column).
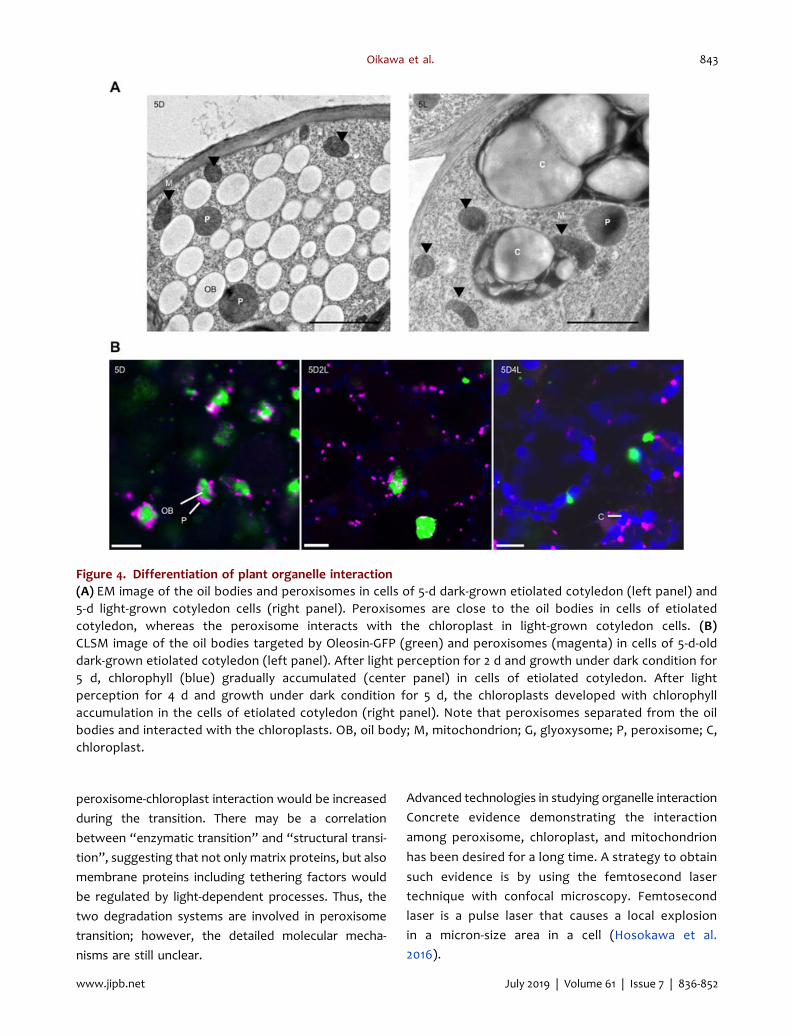
We have provided CLSM images of organelle interactionduring transition from 5-d-old dark-grown cotyledonarycells to an additional 2 and 4 d of continuous lightirradiation (Figure 4B). The image showed that the shiftin organelle interaction actually occurred from the oilbody-peroxisome interaction in 5-d-old dark-growncotyledonary cells to the chloroplast-peroxisome inter-action in the process of peroxisomal transition accom-panied by chloroplast development after 4 d of lightirradiation to 5 d of growth under dark condition.
Double-labeled EM analysis using light-grown coty-ledonary cells during transition demonstrated the co-existence of glyoxysomal and leaf peroxisomal proteinsin individual peroxisomes regardless of their interactionwith other organelles, whereas the glyoxoysomal andleaf peroxisomal proteins are important in dark-grownand light-grown cotyledonary cells, respectively (Titusand Becker 1985; Nishimura et al. 1986). Asmentioned inthe section “Functional differentiation of plant perox-isomes”, glyoxysomal-membrane proteins, peroxisomalascorbate peroxidase (PMP31), PMP28, and PMP38 areeliminated during the transition (Yamaguchi et al. 1995;Fukao et al. 2001). These results suggest that thetethering factors responsible for the glyoxysome-lipidbody interaction would be eliminated and those for leaf
Figure 3. Model for interaction between peroxisome and distinct organelles in the transition of cotyledon cellsAmodel of interaction between glyoxysome and oil body in cells of etiolated cotyledon (left panel), and interactionamong the leaf peroxisomes, mitochondria, and chloroplasts in a greening-cotyledonary cell (right panel). Themechanisms underlying interactions in cells of etiolated cotyledon shift to the interaction in greening-cotyledonarycell during peroxisomal transition from glyoxysome to leaf peroxisome (center panel) are questioned. OB, oil body;M, mitochondrion; G, glyoxysome; P, peroxisome; E, etioplast; C, chloroplast.
Physical interaction between plant organelles 7
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
842
peroxisome-chloroplast interaction would be increased
during the transition. There may be a correlation
between “enzymatic transition” and “structural transi-
tion”, suggesting that not only matrix proteins, but also
membrane proteins including tethering factors would
be regulated by light-dependent processes. Thus, the
two degradation systems are involved in peroxisome
transition; however, the detailed molecular mecha-
nisms are still unclear.
Advanced technologies in studying organelle interaction
Concrete evidence demonstrating the interaction
among peroxisome, chloroplast, and mitochondrion
has been desired for a long time. A strategy to obtain
such evidence is by using the femtosecond laser
technique with confocal microscopy. Femtosecond
laser is a pulse laser that causes a local explosion
in a micron-size area in a cell (Hosokawa et al.
2016).
Figure 4. Differentiation of plant organelle interaction(A) EM image of the oil bodies and peroxisomes in cells of 5-d dark-grown etiolated cotyledon (left panel) and5-d light-grown cotyledon cells (right panel). Peroxisomes are close to the oil bodies in cells of etiolatedcotyledon, whereas the peroxisome interacts with the chloroplast in light-grown cotyledon cells. (B)CLSM image of the oil bodies targeted by Oleosin-GFP (green) and peroxisomes (magenta) in cells of 5-d-olddark-grown etiolated cotyledon (left panel). After light perception for 2 d and growth under dark condition for5 d, chlorophyll (blue) gradually accumulated (center panel) in cells of etiolated cotyledon. After lightperception for 4 d and growth under dark condition for 5 d, the chloroplasts developed with chlorophyllaccumulation in the cells of etiolated cotyledon (right panel). Note that peroxisomes separated from the oilbodies and interacted with the chloroplasts. OB, oil body; M, mitochondrion; G, glyoxysome; P, peroxisome; C,chloroplast.
8 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
843

An impulsive force is produced by subsequent
propagation of the shock wave from the laser focal
point formed by a regeneratively amplified Ti:sapphire
laser (Kaji et al. 2007). This force can detach individual
adherent cells without causing considerable cell
damage, and it is used to measure local adhesion
forces between animal cells (Iino et al. 2010;
Hosokawa et al. 2011). This force is reflected by the
vibratory movement of the atomic force microscopy
(AFM) cantilever and calculated as a unit of impulse
[N-s] (Iino et al. 2010; Hosokawa et al. 2011). The
impulses needed to break leukocyte-endothelial and
interepithelial interactions are calculated at ordersof 10�13 and 10�12 N-s, respectively (Hosokawa et al.2011).
The femtosecond laser technology was usedto measure the intensity of adhesion between peroxi-some and chloroplast under different light conditions(Figure 5) (Oikawa et al. 2015; Hosokawa et al. 2016). Theinteraction strength of plant organelle was successfullyestimated for the first time, and it certainly revealed acorrelation between light-dependent morphologicalchanges (elongation and expansion of surface area)of peroxisome along the chloroplast and interactionstrength between peroxisome and chloroplast (Oikawa
Figure 5. Measuring interactive force between the peroxisome and chloroplast using the femtosecond lasersystemSchematic diagram of microscope with femtosecond laser equipment. Femtosecond laser was focused at 5mmaway from the membrane contact site (MCS) between the peroxisome and chloroplast (left box). The forcegenerated at focus point was calculated from the measurement using atomic force microscopy (AFM) cantilever(right box). Femtosecond laser pulses from an amplified Ti:Sapphire femtosecond laser system (wavelength:780 nm, pulse duration: 230 fs, pulse energy: < 1 mj/pulse, repetition rate: 125 Hz) are introduced to a confocalmicroscope (Oikawa et al. 2015; Hosokawa et al. 2016). A single shot laser pulse was shot by opening themechanicalshutter between the laser and the microscope, and the laser-induced impulsive force was loaded to the targetedperoxisome. The images were obtained using a confocal imaging system and CCD camera.
Physical interaction between plant organelles 9
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
844

et al. 2015, 2016). Under dark condition, the intensitywas 23.4� 2.0 fNnm�2, whereas it was 60.9� 4.5fNnm�2 under light condition (Oikawa et al. 2015). Ithas been reported that the intensity of light is 2.5 timesstronger than that of dark. In addition, the disruption ofF-actin with the inhibitor, latrunculin B, furtherstrengthened the interaction between peroxisomeand chloroplast accompanied by excess elongation ofperoxisome along chloroplast (Oikawa et al. 2015). The
intensity was 161.7� 4.9 fNnm�2, which was 2.5 times
stronger than that in the absence of latrunculin B. This
implies that plant cells regulate the balance between
peroxisome movement and its interaction with
chloroplast.Since the first demonstration of trapping biological
objects with infrared laser light (Ashkin et al. 1987),optical tweezers have been used in biological studies(Bayoudh et al. 2001). Previous studies have adequatelydescribed the principle of optical tweezer (Bayoudhet al. 2001; Moffitt et al. 2008). A single beam highlyfocused using a high-numerical-aperture objective lensform laser trap to a diffraction limited spot that createsa three-dimensional trapping zone becoming the tip ofthe tweezers (Bayoudh et al. 2001). The optical forces,and scattering and gradient forces exert on the trappeddielectric particle a refractive index slightly higher thanthat of its surrounding medium, and three-dimensionaltrapping is barred by domination of gradient force fromthe scattering force (Bayoudh et al. 2001). Opticaltweezers have been used for measuring the interactionforce between organelles such as the ER and chlor-oplasts. The ER fragments, released from rupturedprotoplasts by a laser scalpel, remained attached to thereleased chloroplasts and could be stretched usingoptical tweezers at �425 pN (Andersson et al. 2007).However, after treatment with trypsin, the putativeER-derived membrane could be pulled free from achloroplast using an optical force of 130 pN (Anderssonet al. 2007). The results reflect protein-proteininteractions at the ER–chloroplast MCSs possiblyinvolved in lipid synthesis and transport (Anderssonet al. 2007; Block et al. 2015). Optical tweezerswere also used to trap and move Golgi bodies inArabidopsis leaf epidermal cells (Sparkes et al. 2009),highlighting that the movement of trapped Golgibodies remodeled the ER, indicating a physicalassociation between the two organelles (Sparkes2016).
Optical tweezers have been recently appliedto estimate the interaction between peroxisomes andchloroplasts (Gao et al. 2016). Chloroplasts and perox-isomes are shown to be physically tethered throughperoxules in vivo in epidermal cells of tobacco leaf bytrapping and the organelles were moved using near-infrared optical laser as a tweezer by total internalreflection fluorescence (TIRF) microscopy (Gao et al.2016). Theperoxule formation implied thepresenceof anunknown structure connecting peroxisomes and chlor-oplasts. Femtosecond laser and optical tweezer are alsouseful tools for estimating and identifying structuresof tethering factor in the MCS between two organelles.
FORMATION OF ORGANELLEINTERACTION
Movement of plant peroxisomesIt has been a fundamental question how organelles
interact under a circumstance where numerous organ-
elles actively move with cytoplasmic streaming in plant
cells. To resolve this, EM techniques have been used for
a long time with fixed cells, which have provided
significant information on organelle interaction with
high-resolution images (Trelease et al. 1971; Frederick
et al. 1975; Titus and Becker 1985; Nishimura et al. 1986).
However, recent live-cell imaging techniques can be
good tools for revealing hidden mechanisms of organ-
elle interaction. CLSM using green fluorescence protein
(GFP) (Tsien 1998) has allowed the visualization of
organelles in live plant cells. For example, peroxisomes
have been visualized by constitutively expressing GFP
fused with PTS1 (GFP-PTS1), at the carboxyl-terminus in
Arabidopsis (Jedd and Chua 2002; Mano et al. 2002;
Mathur et al. 2002), and the mitochondria by cyto-
chrome c oxidase IV (coxIV)-GFP (Kohler et al. 1997) and
the ATPase (amino-terminal extra peptide of ATPase d-
subunit)-GFP (Arimura et al. 2002).The visualization of organelles has allowed the
analysis of their movement in live plant cells (Jedd andChua 2002; Mano et al. 2002; Mathur et al. 2002; Wadaet al. 2003; Wada and Suetsugu 2004; Iwabuchi et al.2010; Ueda et al. 2010). The role of organelle movement,e.g., light-dependent chloroplast movement for pre-venting photoinhibition (Kasahara et al. 2002) andnuclear positioning for preventing ultraviolet-inducedDNA damage and cell death (Iwabuchi et al. 2016) has
10 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
845

been related to cell function. It has been shown thatloss-of-functionmutation ofmyosin and overexpressionof the dominant-negative form and the isoform isolatedfrom other species, modulated plant growth bystopping organelle movements (Tominaga et al. 2013;Ryan and Nebenfuhr 2018). This phenomenon indicateda significant role of organelle movement in plantgrowth. Plant organelles preferentially use F-actin forsuitable subcellular distribution (Jedd and Chua 2002;Mano et al. 2002; Mathur et al. 2002; Wada et al. 2003,Wada and Suetsugu 2004; Iwabuchi et al. 2010; Uedaet al. 2010). As a motility force, peroxisomes andmitochondria use multiple myosin XIs (Reisen andHanson 2007; Avisar et al. 2008; Peremyslov et al. 2008),whereas the chloroplasts use unique system-termedchloroplast-actin (CP-actin) (Kadota et al. 2009) regu-lated by blue light receptors called phototropins(Kagawa et al. 2001; Christie 2007). Peroxisomes haveshown a directional movement along F-actin; however,the average speed of peroxisomes in leaf mesophyll cellis lower than that in the epidermis of Arabidopsis (Jeddand Chua 2002; Mano et al. 2002; Mathur et al. 2002;Oikawa et al. 2015). The maximal velocity of perox-isomes (1.36mm/s) (Oikawa et al. 2015) is approximately100 times higher than that of chloroplast (1mm/min)(Wada and Kong 2011). We have previously shown thatperoxisomes in leaf mesophyll cells move from onechloroplast to another to form interactions; however,
most peroxisomes maintain interaction with chloro-
plasts for a long time (Oikawa et al. 2015). Furthermore,
in F-actin-disrupted leaf mesophyll cells, peroxisomes
showed strong interaction with chloroplast instead of
lacking movement (Oikawa et al. 2015). These results
suggest that plant cells balance between organelle
movement and interaction of peroxisome and chloro-
plast. The F-actin-dependent organelle movement
might have a role in the development of interactions
between peroxisomes, mitochondria and chloroplasts.
ROS is a candidate for inducing peroxisomeinteraction with chloroplastUnder high-light conditions, peroxisomes further elon-gated to connect with spherical mitochondria with theER (Jaipargas et al. 2016), and they showed highaccumulation of ROS in anisotropy1 (any1) (Fujita et al.2013) mutants, and it was found to increase thefrequency of peroxules (Jaipargas et al. 2016). Thesefindings suggest that ROS affect organelle shape and
interaction. The finding that UV rays and H2O2 inducedperoxisome elongation (peroxule) along the ER close tothe chloroplasts (Sinclair et al. 2009) supports this idea.The ROS generated from Cd-imposed cell increased theperoxisomemovement (Rodriguez-Serrano et al. 2009).
Stroma-filled tubular structures (stromules) gener-ated from chloroplast comprise the stroma surroundedby the envelope membrane, through which proteins,and undoubtedly other molecules, but not DNA orribosomes, flow (Hanson and Hines 2018). Recently,ROS has been highlighted as an inducer of stromuleformation (Natesan et al. 2005; Hanson and Sattarzadeh2008; Brunkard et al. 2015; Caplan et al. 2015). Stromuleswere formed in response to light-sensitive redoxcomponents (Brunkard et al. 2015) and pathogensthrough H2O2 production in a light-dependent manner(Caplan et al. 2015). It has been shown that H2O2 in thechloroplasts produced under high light condition aretransferred to the nucleus through chloroplast interac-tion for gene expression (Exposito-Rodriguez et al.2017). Collectively, these observations indicate thatorganelle interaction is partially induced by ROS withmorphological changes in the organelles.
Reactive oxygen species accumulate in both glyox-ysomes and leaf peroxisomes through metabolic path-ways (Osmond 1981; Foyer et al. 2009; Del R�ıo andL�opez-Huertas 2016; Corpas et al. 2017). ROS areharmful to cellular functions and therefore shouldbe removed. Recently, it has been reported thatoxidative peroxisomes are preferentially degraded byautophagy (Kim et al. 2013; Shibata et al. 2013;Yoshimoto et al. 2014). In autophagy-related gene(atg) mutants, atg2, atg5, atg7, and atg18a, undegradedperoxisomes are highly accumulated with dysfunctionalcatalase aggregation (Shibata et al. 2013; Yoshimotoet al. 2014). Meanwhile, lower amounts of ROSgenerated in peroxisomes also play a role in cellsignaling (Del R�ıo and L�opez-Huertas 2016).
It is still unclear how ROS induces organelleinteraction and autophagy; however, ROS might beinvolved in these two systems as one of the key playersduring the transition of peroxisomes.
CONCLUSIONS AND PROSPECTIVE
Cell organelles have unique functions and containspecific enzymes and metabolites, which are
Physical interaction between plant organelles 11
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
846

occasionally shared with other organelles. In thisreview, we provided various lines of evidence tosupport that metabolites are exchanged betweenorganelles by direct physical interactions. However,the mechanism involved in the interaction betweenperoxisome and chloroplast remains to be elucidated. Itmight involve unknown factors, which are essential forcell function, as mutants are lethal without thesefactors like photorespiratory mutants in ambient air.From current knowledge of other organelle interac-tions, we hypothesized that the following mechanismsmay mediate the interaction between peroxisomes,mitochondria, and chloroplast: (i) accidental contactbetween distinct organelles following cytoplasmicstreaming. The cytoskeletal regulation of F-actinorganization with the activity of myosin motors andmembrane flexibility of organelles would also beinvolved in this mechanism; (ii) association betweendistinct organelle membrane structures such as thestromule, peroxule, and matrixule induced by ROS(Noctor and Foyer 2007; Hanson and Sattarzadeh 2008;Mathur et al. 2012); (iii) known peroxisome andchloroplast membrane proteins or undefined-tetheringfactors connect both the interacting organelles (Prinz2014; Eisenberg-Bord et al. 2016; Shai et al. 2018); (iv)membrane hemifusion similar to the interaction be-tween the ER and mitochondria (Mehrshahi et al. 2014);and (v) a combination of these four mechanisms.Considerable effort has been devoted to identifyingMCSs using resourceful approaches such as interac-tome based on biochemistry combined with massspectrometry (Hua et al. 2017), artificial tethering(Kornmann et al. 2009), splitting of fluorescenceproteins (Shai et al. 2018), and mutant screening usingRNAi libraries (Chu et al. 2015). Recent studies haveshown that ROS are involved in modulating theinteractions between peroxisomes and chloroplasts(Barton et al. 2018). Recent intriguing findings usingadvanced techniques indicate the existence of tether-ing factors between peroxisome and chloroplast(Oikawa et al. 2015; Gao et al. 2016). Therefore, thenext challenge is discovering the tethering factors andunderstanding the role of ROS in the interaction.Artificial tethering and splitting of fluorescence proteinscombined with mutant screening would help gainfurther insights.
In this review, we focused on plant peroxisomesduring functional transition of interactions for the
transfer of metabolites from the oil bodies to the
chloroplast after light perception of etiolated seed-
lings. During this process, the degradation of matrix
proteins of glyoxysome and importation of newly
synthesized matrix proteins for the leaf peroxisomes
coincidentally occurs in cooperation with peroxins and
Lon protease. Studies on plant autophagy have
revealed that they degrade oxidatively damaged
peroxisomes (Kim et al. 2013; Shibata et al. 2013;
Yoshimoto et al. 2014; Goto-Yamada et al. 2015). The
production and accumulation of ROS depends on the
metabolic process involving fatty acid b-oxidation in
glyoxysomes and photorespiration in leaf peroxi-
somes (Osmond 1981; Kozaki and Takeba 1996;
Foyer et al. 2009; Del R�ıo and L�opez-Huertas 2016;
Corpas et al. 2017). During the transition, peroxisomes
transiently involve two types of metabolic pathways
originating from glyoxysome and leaf peroxisome. In
this situation, the disruption of the metabolic pathway
would produce ROS; therefore, we predicted that
plant pexophagy might be involved in peroxisome
transition by sensing these ROS-accumulated perox-
isomes. Investigating this hypothesis with tethering
factors for two types of organelle interaction during
transition would be a remarkable research focus for
plant scientists in the near future.
ACKNOWLEDGEMENTS
We would like to thank Professor Yoichiro Hosokawa
and Dr. Takanori Iino for their technical support and
insightful discussions on the femtosecond laser
analysis (NIST). This work was supported by the
Ministry of Education, Culture, Sports, Science and
Technology (MEXT, KAKENHI Grant-in-Aid for Scien-
tific Research on Innovative Areas to M. N. [No.
22120007]) and by a fund to M. N. from Wyeth
Foundation.
AUTHOR CONTRIBUTIONS
K.O., M.H., and M.N. conceived this study. K.O. and Y.H.
provided CLSM and EM images. K.O., M.H., Y.H., and
M.N. wrote this manuscript. All authors discussed and
revised the manuscript.
12 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
847

REFERENCES
Andersson MX, Goksor M, Sandelius AS (2007) Opticalmanipulation reveals strong attracting forces at mem-brane contact sites between endoplasmic reticulum andchloroplasts. J Biol Chem 282: 1170–1174
Andronis EA, Moschou PN, Toumi I, Roubelakis-Angelakis KA(2014) Peroxisomal polyamine oxidase and NADPH-oxidase cross-talk for ROS homeostasis which affectsrespiration rate in Arabidopsis thaliana. Front Plant Sci5: 132
Arai Y, Hayashi M, Nishimura M (2008) Proteomic analysis ofhighly purified peroxisomes from etiolated soybeancotyledons. Plant Cell Physiol 49: 526–539
Arimura S, Tsutsumi N (2002) A dynamin-like protein (ADL2b),rather than FtsZ, is involved in Arabidopsis mitochondrialdivision. Proc Natl Acad Sci USA 99: 5727–5731
Ashkin A, Dziedzic JM, Yamane T (1987) Optical trapping andmanipulation of single cells using infrared laser beams.Nature 330: 769–771
Avisar D, Prokhnevsky AI, Makarova KS, Koonin EV, Dolja VV(2008) Myosin XI-K is required for rapid trafficking of Golgistacks, peroxisomes, and mitochondria in leaf cells ofNicotiana benthamiana. Plant Physiol 146: 1098–1108
Baker A, Graham IA, Holdsworth M, Smith SM, Theodoulou FL(2006) Chewing the fat: b-oxidation in signalling anddevelopment. Trends Plant Sci 11: 124–132
Barton KA, Wozny MR, Mathur N, Jaipargas EA, Mathur J(2018) Chloroplast behaviour and interactions with otherorganelles in Arabidopsis thaliana pavement cells. J Cell Sci131: doi: 10.1242/jcs.202275
Bayoudh S, Mehta M, Rubinsztein-Dunlop H, Heckenberg NR,Critchley C (2001) Micromanipulation of chloroplasts usingoptical tweezers. J Microsc 203: 214–222
Beevers H (1979)Microbodies in higher plants.Annu Rev PlantPhysiol 30: 159–193
Block MA, Jouhet J (2015) Lipid trafficking at endoplasmicreticulum-chloroplast membrane contact sites. Curr OpinCell Biol 35: 21–29
Brunkard JO, Runkel AM, Zambryski PC (2015) Chloroplastsextend stromules independently and in response tointernal redox signals. Proc Natl Acad Sci USA 112:10044–10049
Caplan JL, Kumar AS, Park E, Padmanabhan MS, Hoban K,Modla S, Czymmek K, Dinesh-Kumar SP (2015) Chloroplaststromules function during innate immunity. Dev Cell 34:45–57
Christie JM (2007) Phototropin blue-light receptors. Annu RevPlant Biol 58: 21–45
Chu BB, Liao YC, Qi W, Xie C, Du X, Wang J, Yang H, Miao HH,Li BL, Song BL (2015) Cholesterol transport throughlysosome-peroxisomemembranecontacts.Cell 161: 291–306
Corpas FJ, Barroso JB, Palma JM, Rodriguez-Ruiz M (2017)Plant peroxisomes: A nitro-oxidative cocktail. Redox Biol11:535–542
Costello JL, Castro IG, Hacker C, Schrader TA, Metz J,Zeuschner D, Azadi AS, Godinho LF, Costina V, FindeisenP, Manner A, Islinger M, Schrader M (2017) ACBD5 andVAPB mediate membrane associations between perox-isomes and the ER. J Cell Biol 216: 331–342
Cui S, Hayashi Y, Otomo M, Mano S, Oikawa K, Hayashi M,Nishimura M (2016) Sucrose production mediated by lipidmetabolism suppresses the physical interaction of perox-isomes and oil bodies during germination of Arabidopsisthaliana. J Biol Chem 291: 19734–19745
Desai M, Hu J (2008) Light induces peroxisome proliferation inArabidopsis seedlings through the photoreceptor phyto-chrome A, the transcription factor HY5 HOMOLOG, andthe peroxisomal protein PEROXIN11b. Plant Physiol 146:1117–1127
De Bellis L, Nishimura M (1991a) Development of enzymes ofthe glyoxylate cycle during senescence of pumpkincotyledons. Plant Cell Physiol 32: 555–561
De Bellis L, Tsugeki R, Nishimura M (1991b) Glyoxylate cycleenzymes in peroxisomes isolated from petals of pumpkin(Cucurbita sp.) during senescence. Plant Cell Physiol 32:1227–1235
De Duve C, Baudhuin P (1966) Peroxisomes (microbodies andrelated particles). Physiol Rev 46: 323–357
Del R�ıo LA, L�opez-Huertas E (2016) ROS generation inperoxisomes and its role in cell signaling. Plant CellPhysiol 57: 1364–1376
Eastmond PJ (2006) SUGAR-DEPENDENT1 encodes a patatindomain triacylglycerol lipase that initiates storage oilbreakdown in germinatingArabidopsis seeds. Plant Cell 18:665–675
Eisenberg-Bord M, Shai N, Schuldiner M, Bohnert M (2016)A tether is a tether is a tether: Tethering at membranecontact sites. Dev Cell 39: 395–409
Exposito-Rodriguez M, Laissue PP, Yvon-Durocher G, SmirnoffN, Mullineaux PM (2017) Photosynthesis-dependent H2O2
transfer from chloroplasts to nuclei provides a high-lightsignalling mechanism. Nat Commun 8: 49
Farmer LM, Rinaldi MA, Young PG, Danan CH, Burkhart SE,Bartel B (2013) Disrupting autophagy restores peroxisomefunction to an Arabidopsis lon2 mutant and reveals a rolefor the LON2 protease in peroxisomal matrix proteindegradation. Plant Cell 25: 4085–4100
Footitt S, Slocombe SP, Larner V, Kurup S, Wu Y, Larson T,Graham I, Baker A, Holdsworth M (2002) Control ofgermination and lipid mobilization by COMATOSE, theArabidopsis homologue of human ALDP. EMBO J 21:2912–2922
Foyer CH, Bloom AJ, Queval G, Noctor G (2009) Photo-respiratory metabolism: Genes, mutants, energetics, andredox signaling. Annu Rev Plant Biol 60: 455–484
Frederick SE, Gruber PJ, Newcomb EH (1975) Plant micro-bodies. Protoplasma 84: 1–29
FujitaM, Himmelspach R,Ward J,Whittington A, Hasenbein N,Liu C, Truong TT, Galway ME, Mansfield SD, Hocart CH,Wasteneys GO (2013) The anisotropy1 D604N mutation in
Physical interaction between plant organelles 13
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
848

the Arabidopsis cellulose synthase1 catalytic domainreduces cell wall crystallinity and the velocity of cellulosesynthase complexes. Plant Physiol 162: 74–85
Fukao Y, Hayashi Y, Mano S, Hayashi M, Nishimura M (2001)Developmental analysis of a putative ATP/ADP carrierprotein localized on glyoxysomal membranes during theperoxisome transition in pumpkin cotyledons. Plant CellPhysiol 42: 835–841
Fukao Y, Hayashi M, Nishimura M (2002) Proteomic analysis ofleaf peroxisomal proteins in greening cotyledons ofArabidopsis thaliana. Plant Cell Physiol 43: 689–696
Fukao Y, Hayashi M, Hara-Nishimura I, Nishimura M (2003)Novel glyoxysomal protein kinase, GPK1, identified byproteomic analysis of glyoxysomes in etiolated cotyledonsof Arabidopsis thaliana. Plant Cell Physiol 44: 1002–1012
Gamsjaeger R, Liew CK, Loughlin FE, Crossley M, Mackay JP(2007) Sticky fingers: Zinc-fingers as protein-recognitionmotifs. Trends Biochem Sci 32: 63–70
Gao H, Metz J, Teanby NA, Ward AD, Botchway SW, Coles B,Pollard MR, Sparkes I (2016) In vivo quantification ofperoxisome tethering to chloroplasts in Tobacco epider-mal cells using optical tweezers. Plant Physiol 170: 263–272
Goto-Yamada S, Mano S, Nakamori C, Kondo M, Yamawaki R,Kato A, Nishimura M (2014) Chaperone and proteasefunctions of LON protease 2 modulate the peroxisomaltransition and degradation with autophagy. Plant CellPhysiol 55: 482–496
Goto-Yamada S, Mano S, Yamada K, Oikawa K, Hosokawa Y,Hara-Nishimura I, Nishimura M (2015) Dynamics of thelight-dependent transition of plant peroxisomes. PlantCell Physiol 56: 1264–1271
Graham IA (2008) Seed storage oil mobilization. Annu RevPlant Biol 59: 115–142
Hanson MR, Hines KM (2018) Stromules: Probing formationand function. Plant Physiol 176: 128–137
Hanson MR, Sattarzadeh A (2008) Dynamic morphology ofplastids and stromules in angiosperm plants. Plant CellEnviron 31: 646–657
Hayashi M, Toriyama K, Kondo M, Nishimura M (1998) 2,4-Dichlorophenoxybutyric acid-resistant mutants of Arabi-dopsis have defects in glyoxysomal fatty acid β-oxidation.Plant Cell 10: 183–195
Hayashi M, Toriyama K, Kondo M, Kato A, Mano S, De Bellis L,Hayashi-Ishimaru Y, Yamaguchi K, Hayashi H, Nishimura M(2000) Functional transformation of plant peroxisomes.Cell Biochem Biophys 32: 295–304
HayashiM, Nito K, Takei-Hoshi R, YagiM, KondoM, Suenaga A,Yamaya T, NishimuraM (2002) Ped3p is a peroxisomal ATP-binding cassette transporter that might supply substratesfor fatty acid β-oxidation. Plant Cell Physiol 43: 1–11
Hayashi M, Nishimura M (2006) Arabidopsis thaliana: A modelorganism to study plant peroxisomes. Biochim BiophysActa 1763: 1382–1391
Hayashi Y, Hayashi M, Hayashi H, Hara-Nishimura I,Nishimura M (2001) Direct interaction between
glyoxysomes and lipid bodies in cotyledons of theArabidopsis thaliana ped1 mutant. Protoplasma 218: 83–94
Higa T, Suetsugu N, Kong SG, Wada M (2014) Actin-dependentplastid movement is required for motive force generationin directional nuclear movement in plants. Proc Natl AcadSci USA 111: 4327–4331
Hondred D, Wadle DM, Titus DE, Becker WM (1987) Light-stimulated accumulation of the peroxisomal enzymeshydroxypyruvate reductase and serine: Glyoxylate amino-transferase and their translatable messenger-RNAs incotyledons of cucumber seedlings. Plant Mol Biol 9:259–275
Hosokawa Y, Hagiyama M, Iino T, Murakami Y, Ito A (2011)Noncontact estimation of intercellular breaking forceusing a femtosecond laser impulse quantified by atomicforce microscopy. Proc Natl Acad Sci USA 108:1777–1782
Hosokawa Y, Iino T, Oikawa K, Mano S, Yamada K, NishimuraM (2016) Quantification of the adhesion strength betweenperoxisomes and chloroplasts by femtosecond lasertechnology. Bio‐protocol: doi: 10.21769/BioProtoc.1834
Hu J, Baker A, Bartel B, Linka N, Mullen RT, Reumann S,Zolman BK (2012) Plant peroxisomes: Biogenesis andfunction. Plant Cell 24: 2279–2303
Hua R, Cheng D, Coyaud E, Freeman S, Di Pietro E, Wang Y,Vissa A, Yip CM, Fairn GD, Braverman N, Brumell JH,Trimble WS, Raught B, Kim PK (2017) VAPs and ACBD5tether peroxisomes to the ER for peroxisome mainte-nance and lipid homeostasis. J Cell Biol 216: 367–377
Iino T, Hosokawa Y (2010) Direct measurement of femtosec-ond laser impulse in water by atomic force microscopy.Appl Phys Express 3: 107002
IslamMS, Takagi S (2010) Co-localization of mitochondria withchloroplasts is a light-dependent reversible response.Plant Signal Behav 5: 146–147
Iwabuchi K, Minamino R, Takagi S (2010) Actin reorganizationunderlies phototropin-dependent positioning of nuclei inArabidopsis leaf cells. Plant Physiol 152: 1309–1319
Iwabuchi K, Hidema J, Tamura K, Takagi S, Hara-Nishimura I(2016) Plant nuclei move to escape ultraviolet-inducedDNA damage and cell death. Plant Physiol 170: 678–685
Jaipargas EA,Mathur N, Bou Daher F,Wasteneys GO,Mathur J(2016) High light intensity leads to increased peroxule-mitochondria interactions in plants. Front Cell Dev Biol4: 6
Jedd G, Chua NH (2002) Visualization of peroxisomes in livingplant cells reveals acto-myosin-dependent cytoplasmicstreaming and peroxisome budding. Plant Cell Physiol 43:384–392
Johnson TL, Olsen LJ (2001) Building new models forperoxisome biogenesis. Plant Physiol 127: 731–739
Kadota A, Yamada N, Suetsugu N, Hirose M, Saito C, Shoda K,Ichikawa S, Kagawa T, Nakano A, Wada M (2009) Shortactin-based mechanism for light-directed chloroplastmovement in Arabidopsis. Proc Natl Acad Sci USA 106:13106–13111
14 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
849

Kagawa T, Beevers H (1975) The development of microbodies(glyoxysomes and leaf peroxisomes) in cotyledons ofgerminating watermelon seedlings. Plant Physiol 55:258–264
Kagawa T, Sakai T, Suetsugu N, Oikawa K, Ishiguro S, Kato T,Tabata S, Okada K, Wada M (2001) Arabidopsis NPL1: Aphototropin homolog controlling the chloroplast high-light avoidance response. Science 291: 2138–2141
Kaji T, Ito S, Miyasaka H, Hosokawa Y, Masuhara H, ShukunamiC, Hiraki Y (2007) Nondestructive micropatterning of livinganimal cells using focused femtosecond laser-inducedimpulsive force. Appl Phys Lett 91: 023904
Kamada T, Nito K, Hayashi H, Mano S, Hayashi M, Nishimura M(2003) Functional differentiation of peroxisomes revealedby expression profiles of peroxisomal genes in Arabidopsisthaliana. Plant Cell Physiol 44: 1275–1289
Kamada-Nobusada T, Hayashi M, Fukazawa M, Sakakibara H,Nishimura M (2008) A putative peroxisomal polyamineoxidase, AtPAO4, is involved in polyamine catabolism inArabidopsis thaliana. Plant Cell Physiol 49: 1272–1282
Kamigaki A, KondoM,Mano S, HayashiM, NishimuraM (2009)Suppression of peroxisome biogenesis factor 10 reducescuticular wax accumulation by disrupting the ER networkin Arabidopsis thaliana. Plant Cell Physiol 50: 2034–2046
Kao YT, Gonzalez KL, Bartel B (2018) Peroxisome function,biogenesis, and dynamics in plants. Plant Physiol 176:162–177
Kasahara M, Kagawa T, Oikawa K, Suetsugu N, Miyao M,Wada M (2002) Chloroplast avoidance movement reducesphotodamage in plants. Nature 420: 829–832
Kato A, Hayashi M, Takeuchi Y, Nishimura M (1996) cDNAcloning and expression of a gene for 3-ketoacyl-CoAthiolase in pumpkin cotyledons. Plant Mol Biol 31:843–852
Kaur N, Reumann S, Hu J (2009) Peroxisome biogenesisand function. In: Somerville CR, Meyerowitz EM, eds.The Arabidopsis Book. American Society of Plant Biolo-gists: Rockville, MD, http://www.aspb.org/publications/arabidopsis/
Kelly AA, Quettier AL, Shaw E, Eastmond PJ (2011) Seedstorage oil mobilization is important but not essential forgermination or seedling establishment in Arabidopsis.Plant Physiol 157: 866–875
Kim J, Lee H, Lee HN, Kim SH, Shin KD, Chung T (2013)Autophagy-related proteins are required for degradationof peroxisomes in Arabidopsis hypocotyls during seedlinggrowth. Plant Cell 25: 4956–4966
Knoblach B, Sun X, Coquelle N, Fagarasanu A, Poirier RL,Rachubinski RA (2013) An ER-peroxisome tether exertsperoxisome population control in yeast. EMBO J 32:2439–2453
Kohler RH, Zipfel WR, Webb WW, Hanson MR (1997) Thegreen fluorescent protein as a marker to visualize plantmitochondria in vivo. Plant J 11: 613–621
Kornmann B, Currie E, Collins SR, Schuldiner M, Nunnari J,Weissman JS, Walter P (2009) An ER-mitochondria
tethering complex revealed by a synthetic biology screen.Science 325: 477–481
Kozaki K, Takeba G. (1996) Photorespiration protects C3plants from photooxidation. Nature 384: 557–560
Pistelli L, Nieri B, Smith SM, Alpi A, DeBellis L (1996) Glyoxylatecycle enzyme activities are induced in senescent pumpkinfruits. Plant Sci 119: 23–29
Lingard MJ, Bartel B (2009) Arabidopsis LON2 is necessary forperoxisomal function and sustained matrix proteinimport. Plant Physiol 151: 1354–1365
LingardMJ, Monroe-Augustus M, Bartel B (2009) Peroxisome-associatedmatrix protein degradation in Arabidopsis. ProcNatl Acad Sci USA 106: 4561–4566
Mano S, Hayashi M, Kondo M, Nishimura M (1996) cDNAcloning and expression of a gene for isocitrate lyase inpumpkin cotyledons. Plant Cell Physiol 37: 941–948
Mano S, Nakamori C, Hayashi M, Kato A, Kondo M, NishimuraM (2002) Distribution and characterization of peroxisomesin Arabidopsis by visualization with GFP: Dynamic mor-phology and actin-dependent movement. Plant CellPhysiol 43: 331–341
Mano S, Nishimura M (2005) Plant peroxisomes. Vitam Horm72: 111–154
Mano S, Miwa T, Nishikawa S, Mimura T, Nishimura M (2008)The plant organelles database (PODB): A collection ofvisualized plant organelles and protocols for plantorganelle research. Nucleic Acids Res 36: D929–937
Mano S, Miwa T, Nishikawa S, Mimura T, Nishimura M (2011)The Plant Organelles Database 2 (PODB2): An updatedresource containing movie data of plant organelledynamics. Plant Cell Physiol 52: 244–253
Mano S, Nakamura T, Kondo M, Miwa T, Nishikawa S,Mimura T, Nagatani A, Nishimura M (2014) The PlantOrganelles Database 3 (PODB3) update 2014: Integratingelectron micrographs and new options for plant organelleresearch. Plant Cell Physiol 55: e1
Mathur J, Mathur N, Hulskamp M (2002) Simultaneousvisualization of peroxisomes and cytoskeletal elementsreveals actin and not microtubule-based peroxisomemotility in plants. Plant Physiol 128: 1031–1045
Mathur J, Mammone A, Barton KA (2012) Organelle exten-sions in plant cells. J Integr Plant Biol 54: 851–867
Mehrshahi P, Stefano G, Andaloro JM, Brandizzi F, FroehlichJE, DellaPenna D (2013) Transorganellar complementationredefines the biochemical continuity of endoplasmicreticulum and chloroplasts. Proc Natl Acad Sci USA 110:12126–12131
Mehrshahi P, Johnny C, DellaPenna D (2014) Redefining themetabolic continuity of chloroplasts and ER. Trends PlantSci 19: 501–507
Moffitt JR, Chemla YR, Smith SB, Bustamante C (2008) Recentadvances in optical tweezers. Annu Rev Biochem 77:205–228
Mullen RT, Trelease RN (2006) The ER-peroxisome connectionin plants: development of the “ER semi-autonomous
Physical interaction between plant organelles 15
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
850

peroxisome maturation and replication” model for plantperoxisome biogenesis. Biochim Biophys Acta 1763:1655–1668
Mueller SJ, Lang D, Hoernstein SN, Lang EG, Schuessele C,Schmidt A, Fluck M, Leisibach D, Niegl C, Zimmer AD,Schlosser A, Reski R (2014) Quantitative analysis of themitochondrial and plastid proteomes of the mossPhyscomitrella patens reveals protein macrocompartmen-tation and microcompartmentation. Plant Physiol 164:2081–2095
Mueller SJ, Reski R (2015) Mitochondrial dynamics and the ER:The plant perspective. Front Cell Dev Biol 3: 78
Natesan SK, Sullivan JA, Gray JC (2005) Stromules: Acharacteristic cell-specific feature of plastid morphology.J Exp Bot 56: 787–797
Nishimura M, Yamaguchi J, Mori H, Akazawa T, Yokota S(1986) Immunocytochemical analysis shows that glyox-ysomes are directly transformed to leaf peroxisomesduring greening of pumpkin cotyledons. Plant Physiol 81:313–316
Nishimura M, Takeuchi Y, Debellis L, Hara-Nishimura I (1993)Leaf peroxisomes are directly transformed to glyoxy-somes during senescence of pumpkin cotyledons. Proto-plasma 175: 131–137
Nishimura M, Hayashi M, Kato A, Yamaguchi K, Mano S (1996)Functional transformation of microbodies in higher plantcells. Cell Struct Funct 21: 387–393
Noctor G, Foyer CH (2007) Shape-shifters building bridges?Stromules, matrixules and metabolite channeling inphotorespiration. Trends Plant Sci 12: 381–383
Oikawa K,Matsunaga S, Mano S, KondoM, Yamada K, HayashiM, Kagawa T, Kadota A, SakamotoW, Higashi S,WatanabeM,Mitsui T, Shigemasa A, Iino T, Hosokawa Y, NishimuraM(2015) Physical interaction between peroxisomes andchloroplasts elucidated by in situ laser analysis. Nat Plants1: 15035
Oikawa K, Mano S, Yamada K, Hosokawa Y, Nishimura M(2016) Measuring the interactions between peroxisomesand chloroplasts by in situ laser analysis. Bio‐protocol 6:doi 10.21769/BioProtoc.1790
Ogren WL, Bowes G (1971) Ribulose diphosphate carboxylaseregulates soybean photorespiration. Nat New Biol 230:159–160
Osmond CB (1981) Photorespiration and photoinhibition:Some implications for the energetics of photosynthesis.BBA Biomembranes 639: 77–98
Pan R, Reumann S, Lisik P, Tietz S, Olsen LJ, Hu J (2018)Proteome analysis of peroxisomes from dark-treatedsenescent Arabidopsis leaves. J Integr Plant Biol 60:1028–1050
Peremyslov VV, Prokhnevsky AI, Avisar D, Dolja VV (2008) Twoclass XI myosins function in organelle trafficking and roothair development in Arabidopsis. Plant Physiol 146:1109–1116
Perez-Sancho J, Tilsner J, Samuels AL, Botella MA, Bayer EM,Rosado A (2016) Stitching organelles: Organization and
function of specialized membrane contact sites in plants.Trends Cell Biol 26: 705–717
Peterhansel C, Horst I, Niessen M, Blume C, Kebeish R,Kurkcuoglu S, Kreuzaler F (2010) Photorespiration.Arabidopsis Book 8: e0130
Phillips MJ, Voeltz GK (2016) Structure and function of ERmembrane contact sites with other organelles. Nat RevMol Cell Biol 17: 69–82
Prinz WA (2014) Bridging the gap: Membrane contact sites insignaling, metabolism, and organelle dynamics. J Cell Biol205: 759–769
Quan S, Yang P, Cassin-Ross G, Kaur N, Switzenberg R, Aung K,Li J, Hu J (2013) Proteome analysis of peroxisomes frometiolated Arabidopsis seedlings identifies a peroxisomalprotease involved in β-oxidation and development. PlantPhysiol 163: 1518–1538
Reisen D, Hanson MR (2007) Association of six YFP-myosinXI-tail fusions with mobile plant cell organelles. BMC PlantBiol 7: 6
Reumann S, Quan S, Aung K, Yang P, Manandhar-Shrestha K,Holbrook D, Linka N, Switzenberg R, Wilkerson CG, WeberAP, Olsen LJ, Hu J (2009) In-depth proteome analysis ofArabidopsis leaf peroxisomes combined with in vivosubcellular targeting verification indicates novel metabolicand regulatory functions of peroxisomes. Plant Physiol150: 125–143
Reumann S, Bartel B (2016) Plant peroxisomes: Recentdiscoveries in functional complexity, organelle homeosta-sis, and morphological dynamics. Curr Opin Plant Biol 34:17–26
Rodriguez-Serrano M, Romero-Puertas MC, Sparkes I, HawesC, del Rio LA, Sandalio LM (2009) Peroxisome dynamics inArabidopsis plants under oxidative stress induced bycadmium. Free Radical Bio Med 47: 1632–1639
Rodriguez-Serrano M, Romero-Puertas MC, Sanz-FernandezM, Hu J, Sandalio LM (2016) Peroxisomes extendperoxules in a fast response to stress via a reactiveoxygen species-mediated induction of the peroxin PEX11a.Plant Physiol 171: 1665–1674
Ryan JM, Nebenfuhr A (2018) Update on myosin motors:Molecular mechanisms and physiological functions. PlantPhysiol 176: 119–127
Schrader M, Godinho LF, Costello JL, Islinger M (2015) Thedifferent facets of organelle interplay-an overview oforganelle interactions. Front Cell Dev Biol 3: 56
Schumann U, Wanner G, Veenhuis M, Schmid M, Gietl C(2003) AthPEX10, a nuclear gene essential for peroxi-some and storage organelle formation duringArabidopsis embryogenesis. Proc Natl Acad Sci USA100: 9626–9631
Schumann U, Prestele J, O’Geen H, Brueggeman R, Wanner G,Gietl C (2007) Requirement of the C3HC4 zinc RING fingerof the Arabidopsis PEX10 for photorespiration and leafperoxisome contact with chloroplasts. Proc Natl Acad SciUSA 104: 1069–1074
16 Oikawa et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netJuly 2019 | Volume 61 | Issue 7 | 836-852www.jipb.net
851

Shai N, Schuldiner M, Zalckvar E (2016) No peroxisome is anisland: Peroxisome contact sites. Biochim Biophys Acta1863: 1061–1069
Shai N, Yifrach E, van Roermund CWT, Cohen N, Bibi C, IJ L,Cavellini L, Meurisse J, Schuster R, Zada L, Mari MC,Reggiori FM, Hughes AL, Escobar-Henriques M, CohenMM, Waterham HR, Wanders RJA, Schuldiner M, ZalckvarE (2018) Systematic mapping of contact sites revealstethers and a function for the peroxisome-mitochondriacontact. Nat Commun 9: 1761
Shibata M, Oikawa K, Yoshimoto K, Kondo M, Mano S,Yamada K, Hayashi M, Sakamoto W, Ohsumi Y,Nishimura M (2013) Highly oxidized peroxisomes areselectively degraded via autophagy in Arabidopsis. PlantCell 25: 4967–4983
Sinclair AM, Trobacher CP, Mathur N, Greenwood JS, Mathur J(2009) Peroxule extension over ER-defined paths con-stitutes a rapid subcellular response to hydroxyl stress.Plant J 59: 231–242
Somerville CR, Ogren WL (1980) Photorespiration mutants ofArabidopsis thaliana deficient in serine-glyoxylate amino-transferase activity. Proc Natl Acad Sci USA 77: 2684–2687
Somerville CR (2001) An early Arabidopsis demonstration:Resolving a few issues concerning photorespiration. PlantPhysiol 125: 20–24
Sapir-Mir M, Mett A, Belausov E, Tal-Meshulam S, Frydman A,Gidoni D, Eyal Y (2008) Peroxisomal localization ofArabidopsis isopentenyl diphosphate isomerases suggeststhat part of the plant isoprenoidmevalonic acid pathway iscompartmentalized to peroxisomes. Plant Physiol 148:1219–1228
Sparkes IA, Brandizzi F, Slocombe SP, El-Shami M, Hawes C,Baker A (2003) AnArabidopsis pex10 null mutant is embryolethal, implicating peroxisomes in an essential role duringplant embryogenesis. Plant Physiol 133: 1809–1819
Sparkes I, Runions J, Hawes C, Griffing L (2009) Movementand remodeling of the endoplasmic reticulum in nondivid-ing cells of tobacco leaves. Plant Cell 21: 3937–3949
Sparkes I (2016) Using optical tweezers to characterizephysical tethers at membrane contact sites: Grab it, pullit, set it free? Front Cell Dev Biol 4: 22
Thazar-Poulot N, Miquel M, Fobis-Loisy I, Gaude T (2015)Peroxisome extensions deliver the Arabidopsis SDP1 lipaseto oil bodies. Proc Natl Acad Sci USA 112: 4158–4163
Titus DE, Becker WM (1985) Investigation of the glyoxysome-peroxisome transition in germinating cucumber cotyle-dons using double-label immunoelectron microscopy.J Cell Biol 101: 1288–1299
Tolbert NE (1981) Metabolic pathways in peroxisomes andglyoxysomes. Annu Rev Biochem 50: 133–157
Tolbert NE, Essner E (1981) Microbodies: Peroxisomes andglyoxysomes. J Cell Biol 91: 271s–283s
Tolbert NE (1982) Leaf peroxisomes. Ann NY Acad Sci 386:254–268
Tominaga M, Kimura A, Yokota E, Haraguchi T, Shimmen T,Yamamoto K, Nakano A, Ito K (2013) Cytoplasmicstreaming velocity as a plant size determinant. DevCell 27: 345–352
Trelease RN, Becker WM, Gruber PJ, Newcomb EH (1971)Microbodies (glyoxysomes and peroxisomes) in cucumbercotyledons: Correlative biochemical and ultrastructuralstudy in light- and dark-grown seedlings. Plant Physiol 48:461–475
Trelease RN (1984) Biogenesis of glyoxysomes. Annu RevPlant Phys 35:321–347
Tsien RY (1998) The green fluorescent protein. Annu RevBiochem 67: 509–544
Tsugeki R, Hara-Nishimura I, Mori H, Nishimura M (1993)Cloning and sequencing of cDNA for glycolate oxidasefrom pumpkin cotyledons and northern blot analysis.Plant Cell Physiol 34: 51–57
Ueda H, Yokota E, Kutsuna N, Shimada T, Tamura K, ShimmenT, Hasezawa S, Dolja VV, Hara-Nishimura I (2010) Myosin-dependent endoplasmic reticulum motility and F-actinorganization in plant cells. Proc Natl Acad Sci USA 107:6894–6899
Vranova E, Coman D, Gruissem W (2013) Network analysis ofthe MVA and MEP pathways for isoprenoid synthesis.Annu Rev Plant Biol 64: 665–700
Wada M, Kagawa T, Sato Y (2003) Chloroplast movement.Annu Rev Plant Biol 54: 455–468
Wada M, Suetsugu N (2004) Plant organelle positioning. CurrOpin Plant Biol 7: 626–631
Wada M, Kong SG (2011) Analysis of chloroplast movementand relocation in Arabidopsis. Methods Mol Biol 774:87–102
Yamaguchi K, Takeuchi Y, Mori H, Nishimura M (1995)Development of microbody membrane proteins duringthe transformation of glyoxysomes to leaf peroxisomes inpumpkin cotyledons. Plant Cell Physiol 36: 455–464
Yan M, Rachubinski DA, Joshi S, Rachubinski RA, Subramani S(2008) Dysferlin domain-containing proteins, Pex30p andPex31p, localized to two compartments, control thenumber and size of oleate-induced peroxisomes in Pichiapastoris. Mol Biol Cell 19: 885–898
Yoshimoto K, Shibata M, Kondo M, Oikawa K, Sato M,Toyooka K, Shirasu K, Nishimura M, Ohsumi Y (2014)Organ-specific quality control of plant peroxisomes ismediated by autophagy. J Cell Sci 127: 1161–1168
Zolman BK, Silva ID, Bartel B (2001) The Arabidopsis pxa1mutant is defective in an ATP-binding cassette trans-porter-like protein required for peroxisomal fatty acidβ-oxidation. Plant Physiol 127: 1266–1278
Physical interaction between plant organelles 17
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXJuly 2019 | Volume 61 | Issue 7 | 836-852 www.jipb.net
852
Related Documents




![arXiv:1811.10804v1 [cs.IR] 27 Nov 2018 - Scinapse](https://static.cupdf.com/doc/110x72/633457de4e43a4bcd80d2edc/arxiv181110804v1-csir-27-nov-2018-scinapse.jpg)






