Development/Plasticity/Repair Mature Purkinje Cells Require the Retinoic Acid-Related Orphan Receptor- (ROR) to Maintain Climbing Fiber Mono-Innervation and Other Adult Characteristics Xiao Ru Chen, 1,2 Nicolas Heck, 3 Ann M. Lohof, 1,2 Christelle Rochefort, 1,2 Marie-Pierre Morel, 1,2 Rosine Wehrle ´, 1,2 Mohamed Doulazmi, 1,2 Serge Marty, 4 Vidjeacoumary Cannaya, 1,2 Hasan X. Avci, 1,2 Jean Mariani, 1,2,5 Laure Rondi-Reig, 1,2 Guilan Vodjdani, 6 Rachel M. Sherrard, 1,2 Constantino Sotelo, 7 and Isabelle Dusart 1,2 1 UPMC Universite ´ Paris 06, UMR 7102, 75005 Paris, France, 2 CNRS, UMR 7102, 75005 Paris, France, 3 UPMC Universite ´ Paris 06, CNRS UMR7224/Institut National de la Sante ´ et de la Recherche Me ´dicale UMRS952, 75005 Paris, France, 4 Institute of Biology of the Ecole Normale Supe ´rieure, Inserm U1024, CNRS UMR8197, 75005 Paris, France, 5 Institut de la Longe ´vite ´, Ho ˆpital Charles Foix, 94205 Ivry sur Seine, France, 6 Centre de Recherche du Cerveau et de la Moelle Epiniere, UPMC/CNRS UMR7225/Institut National de la Sante ´ et de la Recherche Me ´dicale UMRS975, 75013 Paris, France, and 7 Ca ´tedra de Neurobiología del Desarrollo “Remedios Caro Almela,” Instituto de Neurociencias, Universidad Miguel Herna ´ndez–Consejo Superior de Investigaciones Científicas, 03550 San Juan de Alicante, Spain Neuronal maturation during development is a multistep process regulated by transcription factors. The transcription factor ROR (retinoic acid-related orphan receptor ) is necessary for early Purkinje cell (PC) maturation but is also expressed throughout adulthood. To identify the role of ROR in mature PCs, we used Cre-lox mouse genetic tools in vivo that delete it specifically from PCs between postnatal days 10 –21. Up to 14 d of age, differences between mutant and control PCs were not detectable: both were mono-innervated by climbing fibers (CFs) extending along their well-developed dendrites with spiny branchlets. By week 4, mutant mice were ataxic, some PCs had died, and remaining PC soma and dendrites were atrophic, with almost complete disappearance of spiny branchlets. The innervation pattern of surviving ROR-deleted PCs was abnormal with several immature characteristics. Notably, multiple functional CF innervation was reestablished on these mature PCs, simultaneously with the relocation of CF contacts to the PC soma and their stem dendrite. This morphological modification of CF contacts could be induced even later, using lentivirus-mediated depletion of rora from adult PCs. These data show that the late postnatal expression of ROR cell-autonomously regulates the maintenance of PC dendritic complexity, and the CF innervation status of the PC (dendritic vs somatic contacts, and mono-innervation vs multi-innervation). Thus, the differentiation state of adult neurons is under the control of transcription factors; and in their absence, adult neurons lose their mature characteristics and acquire some characteristics of an earlier developmental stage. Introduction Transcription factors drive the developmental acquisition of ma- ture neuronal features (Polleux et al., 2007; Hobert, 2011). Some of these transcription factors maintain their expression into adulthood. The exact role of this adult expression is not clear, but in some cases they are required to maintain adult neuronal characteristics. For example, the removal of Pet-1 or Nurr1 in the adult mouse nervous system results in a progressive loss of serotonergic or dopaminergic neuron function, respectively (Kadkhodaei et al., 2009; Liu et al., 2010). The present study was undertaken to investigate whether the transcription factor retinoic acid-related orphan receptor (rora, ROR, NR1D1), a member of the nuclear hormone-receptor superfamily, is necessary for the maintenance of adult Purkinje cell (PC) characteristics. The expression of ROR in PCs starts very early in develop- ment and continues during adulthood (Ino, 2004). In spontane- ous mutant (staggerer) or rora knock-out mice, most of the Purkinje cells die within the first month of life (Herrup and Mul- len, 1979; Dussault et al., 1998; Vogel et al., 2000; Doulazmi et al., 2001). In addition, surviving PCs do not develop spiny branch- lets, remaining in an immature morphological state (Landis and Sidman, 1978; Sotelo, 1978; Boukhtouche et al., 2006b). ROR is necessary for the retraction of transient PC dendrites early in development to allow the establishment of a mature dendritic tree (Boukhtouche et al., 2006b). Furthermore, studies of rora- Received June 22, 2012; revised April 20, 2013; accepted April 26, 2013. Author contributions: X.R.C., A.M.L., J.M., L.R.-R., C.S., and I.D. designed research; X.R.C., N.H., A.M.L., M.-P.M., R.W., V.C., S.M., A.X.H., R.M.S., and I.D. performed research; G.V. contributed unpublished reagents/analytic tools; X.R.C., N.H., A.M.L., C.R., M.-P.M., R.W., M.D., S.M., R.M.S., C.S., and I.D. analyzed data; X.R.C., A.M.L., R.M.S., C.S., and I.D. wrote the paper. This work was supported by the CNRS, UPMC, and ANR-07-NEURO-043– 01. C.S. was supported in part by the Spanish Ministry of Science and Innovation Grant BFU2010 –27326. We thank Richard Schwartzmann (Plateform Imagerie, IFR83) and David Godefroy (Cellular Imaging Facility of the Institut de la Vision, Paris) for help with confocal and Nanozoomer slides scanning, respectively. The authors declare no competing financial interests. Correspondence should be addressed to Dr. Isabelle Dusart, Equipe Diffe ´renciation Neuronale et Gliale, NPA UMR7102, Universite ´ Pierre et Marie Curie, Bat B, 6eme, Case 12, 9 Quai Saint Bernard, 75005 Paris, France. E-mail: [email protected]. DOI:10.1523/JNEUROSCI.2977-12.2013 Copyright © 2013 the authors 0270-6474/13/339546-17$15.00/0 9546 • The Journal of Neuroscience, May 29, 2013 • 33(22):9546 –9562

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Development/Plasticity/Repair
Mature Purkinje Cells Require the Retinoic Acid-RelatedOrphan Receptor-� (ROR�) to Maintain Climbing FiberMono-Innervation and Other Adult Characteristics
Xiao Ru Chen,1,2 Nicolas Heck,3 Ann M. Lohof,1,2 Christelle Rochefort,1,2 Marie-Pierre Morel,1,2 Rosine Wehrle,1,2
Mohamed Doulazmi,1,2 Serge Marty,4 Vidjeacoumary Cannaya,1,2 Hasan X. Avci,1,2 Jean Mariani,1,2,5
Laure Rondi-Reig,1,2 Guilan Vodjdani,6 Rachel M. Sherrard,1,2 Constantino Sotelo,7 and Isabelle Dusart1,2
1UPMC Universite Paris 06, UMR 7102, 75005 Paris, France, 2CNRS, UMR 7102, 75005 Paris, France, 3UPMC Universite Paris 06, CNRS UMR7224/InstitutNational de la Sante et de la Recherche Medicale UMRS952, 75005 Paris, France, 4Institute of Biology of the Ecole Normale Superieure, Inserm U1024,CNRS UMR8197, 75005 Paris, France, 5Institut de la Longevite, Hopital Charles Foix, 94205 Ivry sur Seine, France, 6Centre de Recherche du Cerveau et de laMoelle Epiniere, UPMC/CNRS UMR7225/Institut National de la Sante et de la Recherche Medicale UMRS975, 75013 Paris, France, and 7Catedra deNeurobiología del Desarrollo “Remedios Caro Almela,” Instituto de Neurociencias, Universidad Miguel Hernandez–Consejo Superior de InvestigacionesCientíficas, 03550 San Juan de Alicante, Spain
Neuronal maturation during development is a multistep process regulated by transcription factors. The transcription factor ROR�(retinoic acid-related orphan receptor �) is necessary for early Purkinje cell (PC) maturation but is also expressed throughout adulthood.To identify the role of ROR� in mature PCs, we used Cre-lox mouse genetic tools in vivo that delete it specifically from PCs betweenpostnatal days 10 –21. Up to 14 d of age, differences between mutant and control PCs were not detectable: both were mono-innervated byclimbing fibers (CFs) extending along their well-developed dendrites with spiny branchlets. By week 4, mutant mice were ataxic, somePCs had died, and remaining PC soma and dendrites were atrophic, with almost complete disappearance of spiny branchlets. Theinnervation pattern of surviving ROR�-deleted PCs was abnormal with several immature characteristics. Notably, multiple functional CFinnervation was reestablished on these mature PCs, simultaneously with the relocation of CF contacts to the PC soma and their stemdendrite. This morphological modification of CF contacts could be induced even later, using lentivirus-mediated depletion of rora fromadult PCs. These data show that the late postnatal expression of ROR� cell-autonomously regulates the maintenance of PC dendriticcomplexity, and the CF innervation status of the PC (dendritic vs somatic contacts, and mono-innervation vs multi-innervation). Thus,the differentiation state of adult neurons is under the control of transcription factors; and in their absence, adult neurons lose theirmature characteristics and acquire some characteristics of an earlier developmental stage.
IntroductionTranscription factors drive the developmental acquisition of ma-ture neuronal features (Polleux et al., 2007; Hobert, 2011). Someof these transcription factors maintain their expression intoadulthood. The exact role of this adult expression is not clear,but in some cases they are required to maintain adult neuronal
characteristics. For example, the removal of Pet-1 or Nurr1 inthe adult mouse nervous system results in a progressive loss ofserotonergic or dopaminergic neuron function, respectively(Kadkhodaei et al., 2009; Liu et al., 2010). The present studywas undertaken to investigate whether the transcription factorretinoic acid-related orphan receptor � (rora, ROR�, NR1D1), amember of the nuclear hormone-receptor superfamily, isnecessary for the maintenance of adult Purkinje cell (PC)characteristics.
The expression of ROR� in PCs starts very early in develop-ment and continues during adulthood (Ino, 2004). In spontane-ous mutant (staggerer) or rora knock-out mice, most of thePurkinje cells die within the first month of life (Herrup and Mul-len, 1979; Dussault et al., 1998; Vogel et al., 2000; Doulazmi et al.,2001). In addition, surviving PCs do not develop spiny branch-lets, remaining in an immature morphological state (Landis andSidman, 1978; Sotelo, 1978; Boukhtouche et al., 2006b). ROR� isnecessary for the retraction of transient PC dendrites early indevelopment to allow the establishment of a mature dendritictree (Boukhtouche et al., 2006b). Furthermore, studies of rora-
Received June 22, 2012; revised April 20, 2013; accepted April 26, 2013.Author contributions: X.R.C., A.M.L., J.M., L.R.-R., C.S., and I.D. designed research; X.R.C., N.H., A.M.L., M.-P.M.,
R.W., V.C., S.M., A.X.H., R.M.S., and I.D. performed research; G.V. contributed unpublished reagents/analytic tools;X.R.C., N.H., A.M.L., C.R., M.-P.M., R.W., M.D., S.M., R.M.S., C.S., and I.D. analyzed data; X.R.C., A.M.L., R.M.S., C.S.,and I.D. wrote the paper.
This work was supported by the CNRS, UPMC, and ANR-07-NEURO-043– 01. C.S. was supported in part by theSpanish Ministry of Science and Innovation Grant BFU2010 –27326. We thank Richard Schwartzmann (PlateformImagerie, IFR83) and David Godefroy (Cellular Imaging Facility of the Institut de la Vision, Paris) for help withconfocal and Nanozoomer slides scanning, respectively.
The authors declare no competing financial interests.Correspondence should be addressed to Dr. Isabelle Dusart, Equipe Differenciation Neuronale et Gliale, NPA
UMR7102, Universite Pierre et Marie Curie, Bat B, 6eme, Case 12, 9 Quai Saint Bernard, 75005 Paris, France. E-mail:[email protected].
DOI:10.1523/JNEUROSCI.2977-12.2013Copyright © 2013 the authors 0270-6474/13/339546-17$15.00/0
9546 • The Journal of Neuroscience, May 29, 2013 • 33(22):9546 –9562

deficient models have suggested some roles for ROR� in the adultPC: adult rora haploinsufficient mice show premature dendriticatrophy compared with wild-type mice and also show earlier PCloss (Zanjani et al., 1992; Hadj-Sahraoui et al., 1997; Doulazmi etal., 1999; Janmaat et al., 2011). However, it is unknown whetherthese effects are the result of ROR� deficiency in the adult or thecumulative effect of a reduced amount of ROR� throughoutdevelopment.
To address the role of ROR� in mature PCs, we developed amouse model in which the rora gene is deleted specifically in PCsbetween the 10th and 21st postnatal day. At 2 weeks of age, PCs inmutant and control mice have acquired most of their adult char-acteristics, including a complex dendritic tree with many tertiarybranchlets, parallel fiber (PF) synapses, and climbing fiber (CF)mono-innervation as well described previously (for review, seeSotelo and Dusart, 2009; Kano and Hashimoto, 2009; van Welieet al., 2011). The loss of ROR� from PCs at this relatively late stageof their maturation was associated with dendritic retraction, almostcomplete loss of spiny branchlets, new CF multi-innervation, andthe appearance of perisomatic spines, suggesting regressive changeswith a reversal of developmental processes. The effects of deleting PCrora even later in adulthood corroborated these observations. Ourfindings indicate that continued expression of the transcription fac-tor ROR� is necessary after neuronal maturation to maintain ma-ture morphological and innervation characteristics in the adult PC.In the absence of ROR�, PCs develop immature characteristics, in-cluding CF multi-innervation, suggesting that maintaining the stateof differentiation of adult neurons is under the control of transcrip-tion factors.
Materials and MethodsAnimals. All procedures were submitted and approved by the Regional Eth-ics Committee in Animal Experiment 3 of Ile-de-France region (p3/2009/020). Animals had ad libitum access to food and water with 12 h light-darkcycle. C57BL/6 mice homozygous for the floxed Rora allele (Rorafl/fl) (Clin-ique de la Souris; see Fig. 1A) were crossed with C57BL/6 heterozygous(Pcp2::Cre�/�) mice, expressing Cre under the control of the Pcp2 (L7)promoter (B6.129-Tg(Pcp2::Cre)2Mpin, The Jackson Laboratory; see Fig.1A) to generate Pcp2::Cre�/�;Rorafl/� and Pcp2::Cre�/�;Rorafl/� mice.Mating these mice yielded the parent mice used in our study:Pcp2::Cre�/�;Rorafl/�, Pcp2::Cre�/�;Rorafl/�, Pcp2::Cre�/�;Rorafl/fl, andPcp2::Cre�/�;Rorafl/fl. A YFPstopfl/fl reporter mouse in which a stop signal isflanked by two loxP sites was also used (Srinivas et al., 2001). By crossingYFPstopfl/fl mice with Rorafl/fl, we generated another line of parent miceYFPstopfl/fl;Rorafl/fl.
For the behavior experiments, we used male littermates derived fromcrosses between Pcp2::Cre�/�;Rorafl/� and Pcp2::Cre�/�;Rorafl/� to obtainwithin the same litters the mutant Pcp2::Cre�/�;Rorafl/fl (mutant) and thethree types of control: Pcp2::Cre�/�;Rora�/� (wild-type control: WT),Pcp2::Cre�/�;Rora�/� (L7-Cre control: L7CTRL), and Pcp2::Cre�/�;Rorafl/fl (floxed Rora control: RORCTRL).
For the immunohistochemistry studies, Pcp2::Cre�/�;Rorafl/fl andPcp2::Cre�/�;Rorafl/fl were mated to obtain the male mutant mice and theirmale littermate controls.
For the electrophysiology experiments, to visualize the mutant Purkinjeneurons, the parent mice YFPstopfl/fl;Rorafl/fl and Pcp2::Cre�/�;Rorafl/fl werecrossed to obtain Pcp2::Cre�/�;YFPstopfl/fl;Rorafl/fl as control andPcp2::Cre�/�;YFPstopfl/fl;Rorafl/fl as mutant. Mice of both sexes (P15–P16)and males of 1 month of age were used for these experiments.
For the Cre-expressing lentiviral vector injection studies, Rorafl/fl orYFPstopfl/fl; Rorafl/fl male mice of 2 months of age were used.
Mice were genotyped by PCR using genomic DNA prepared frommouse tails by incubating in 60 �l of 25 mM NaOH/0.2 mM EDTA at 95°Cfor 30 min followed by 5 min at 4°C, then neutralizing the mixture byadding 60 �l of 40 mM Tris, pH 8.1, or 5 �l of the sample was used in each
genotyping PCR according to the final volume (20 or 25 �l). Differentprimer pairs were used for PCR to identify the different alleles.
Two pairs of primers were used for identifying floxed rora allele: 5�-AGAGCAATGCCACCTACTCCTGTCC-3� and 5�-AGTACAGGACACTTCGGTGTC-3� for one loxP site, and 5�-TTGTGTATACCACCACAAGTGCACC-3� and 5�-CTAATCCTCCATCCCTTACAC-3� for the otherloxP site; one for L7Cre allele 5�-CGATGCAACGAGTGATGAGG-3�and 5�-GCATTGCTGTCACTTGGTCGT-3�, and 3 primers for YFP al-lele 5�-AAAAGTCGCTCTGAGTTGTTAT-3�, 5�-GGAGCGGGAGAAATGGATATG-3�, and 5�-GCCAAGAGTTTGTCCTCAACC-3�.
PCR cycling was initiated by a denaturation at 95°C for 5 min, then anamplification through 35 cycles of 30 s at 95°C, 30 s at Tm of primers, and30 s at 72°C, followed by10 min at 72°C. The Tm of floxed rora, Pcp2::Cre,and YFP primers are 60°C, 58°C, and 62°C respectively.
Behavior study. All behavioral studies were performed blind to thegenotype of mice. All animals were tested according to the SHIRPA pro-tocol (Rondi-Reig et al., 2001; Rondi-Reig and Mariani, 2002). Animalswere kept isolated throughout behavioral experiments and from 7 dbefore, to limit the variability resulting from social interaction. The testsaimed to detect potential differences in sensorimotor control perfor-mances. Mice were first positioned at the center of an arena made of grayperspex (45 � 45 cm) surrounded by red Plexiglas walls (30 cm height)and were allowed free exploration for 10 min, during which the numberof rearings were quantified. Footprint characteristics were measured bymeans of ink deposited on the paws of mice; the animals were then placedat the entrance of a corridor (60 cm long and 7.5 cm wide) with a floorcovered with paper. Dynamic balance was evaluated using the horizontalrod test (Rondi-Reig et al., 1999). The aim of this test was to estimate themouse’s ability to maintain its balance while in motion. The apparatusconsisted of a horizontal rod (50 cm long, 5 cm in diameter) covered withsticking plaster providing a good gripping surface. It was located 80 cmabove a soft carpet to cushion the possible fall of the animals. Both endsof the beam were limited by white altuglass disk (50 cm in diameter). Themouse was placed on the middle of the rod, its body axis perpendicular tothe rod axis. During the test, the time before falling, the distance traveled,and the walking time were recorded. The test ended when the animal fellor after 180 s.
To assess motor coordination, an automated hole-board was used(license #DI01873– 01) (Rondi-Reig et al., 2008). It consisted of an ex-perimental box made of transparent altuglass (32 � 32 � 25 cm), with awhite altuglass floor board, which has 36 holes (2 cm in diameter, 2 cmdeep) arranged in a 6 � 6 grid. The mouse was placed in the middle of theboard, and its behavior was recorded during 10 min. The walking timeand the frequency of stumbles, a measure of motor coordination, werecalculated (Rondi-Reig and Mariani, 2002).
The accelerating rotarod (LE-8200; Bioseb) consists of a horizontalrod (3 cm in diameter), turning on its longitudinal axis. Mice were placedon a 5 cm section of the rod facing in the direction opposite to thedirection of the rod rotation, such that the animal had to walk forward toavoid a fall. The training phase consisted of walking on the rod turning ata constant speed of 4 rotations per minute (rpm) for successive 30 s trials.When mice successfully walked on the rotating rod during three consec-utive 30 s trials, four trials were conducted in which the speed of therotation increased gradually from 4 rpm to 40 rpm over 5 min. Theanimal had to coordinate its walk with the rotation speed. This test re-quires strength, efficient balance, and motor coordination. Time spenton the rotarod was recorded and averaged for the 4 trials.
The muscular strength of the animal’s forepaws was measured using agrip test (Bioseb). The mouse was held by the base of its tail and allowedto firmly grab the grid of the device with its forepaws. The mouse wasthen pulled gently backwards until it released its grip. The peak force (N)of each trial was considered as the grip strength. Four successive mea-surements were averaged.
Tissue preparation. For light microscopy, mice were anesthetized withsodium pentobarbital (50 mg/kg i.p.) and perfused through the aortawith 0.12 M phosphate-buffer, pH 7.4, containing 4% paraformaldehyde.The cerebella were removed and weighed, then postfixed for 2 h. Somecerebella were cryoprotected in 20% sucrose, frozen in isopentane, andstored at �80°C until cryostat sectioning. Other fixed cerebella were kept
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9547

in phosphate buffer at 4°C until vibratome sectioning. Cryostat sec-tions were stored at �80°C and vibratome sections at 4°C untilimmunostaining.
For immunostaining, sections were blocked for 1 h in PBS containing0.25% Triton-X, 0.2% gelatin, 0.1% sodium azide and lysine (0.1 M)before applying overnight primary antibody in PBS containing 0.25%Triton-X, 0.2% gelatin, 0.1% sodium azide. Primary antibodies were asfollows: goat anti-FOXP2 (1/500; Abcam); goat anti-ROR� (1/500; SantaCruz Biotechnology); mouse or rabbit anti-calbindin-D28K (Calb1)(both 1/5000; Swant); guinea pig anti-vesicular glutamate transporter 1(VGLUT1, 1/3000; Millipore); guinea pig anti-vesicular glutamate trans-porter 2 (VGLUT2; 1/3000 Millipore); and chicken anti-GFP (1/200;Aveslab). After washes in PBS containing 0.25% Triton-X and 2 h incu-bation with a combination of appropriate species-specific secondary an-tibodies in PBS containing 0.25% Triton-X, 0.2% gelatin, 0.1% sodiumazide, the sections were washed several times in PBS and mounted inmowiol (Calbiochem). All the secondary antibodies were from donkey toallow triple or quadruple immunostainings. We used anti-mouse, anti-rabbit, and anti-goat linked to Alexa-488 (1:400; Invitrogen), anti-mouse, anti-rabbit, and anti-goat linked to aminomethylcoumarine (1/50), anti-rabbit and anti-goat linked to Cy3 (indocarbocyanine, 1/500),anti-guinea-pig linked to Cy3 (1/200), anti-mouse, anti-rabbit, and anti-goat linked to Cy5 (indodicarbocyanine, 1/200), and anti-chicken linkedto FITC (1/200, Jackson ImmunoResearch Laboratories).
For electron microscopy, three 2-month-old mutant (Pcp2::Cre �/�;Rora fl/fl) and three littermate control (Pcp2::Cre �/�; Rora fl/fl) male micewere anesthetized with sodium pentobarbital (50 mg/kg i.p.) and per-fused with 400 ml of a freshly prepared solution of 2% paraformaldehydeand 2% glutaraldehyde in 0.12 M phosphate buffer, pH 7.3, for 20 min atroom temperature. After 1 h at 4°C, the cerebella were carefully dissectedout and left in the same fixative 4 h at 4°C. Vermal slices 200 �m thickwere cut in the parasagittal plane. The lobule 3 of vermal sections weretransferred into a solution containing 2% osmium for 2 h at 4°C. Afterwashes, they were stained “en bloc” with a solution containing 2% uranylacetate for 1 h at 4°C. After washes, samples were dehydrated in gradedethanol followed by acetone and incubated in 50% acetone-50% Aralditefor 1 h, followed by 10% acetone-90% Araldite for 2 h. They were thenincubated in Araldite for 2 h, before being embedded. Ultrathin (70 nm)sections were cut using an EM UC6 (Leica Microsystems) and collectedon 400 mesh copper grids. The sections were stained by incubation with5% uranyl acetate in 70% methanol for 5 min and then with lead citrate(0.08 M lead nitrate, 0.12 M sodium citrate in CO2-free dH2O) for 5 min.The sections were observed with a Philips TECNAI 12 (FEI).
Analysis of ROR� expression and PC survival. The time course of ROR�expression was analyzed by immunohistochemistry at P10, P14, P21, andP60. Three mutant mice (Pcp2::Cre�/�;Rorafl/fl) and their control litter-mates (Pcp2::Cre�/�;Rorafl/fl) were used for each time point. Cerebellawere cut on a cryostat into six series of 16-�m-thick sagittal sections. Oneseries was stained with rabbit anti-Calb1 (to label the PCs), guinea piganti-VGLUT2, and goat anti-ROR� antibodies. Another series wasstained with goat anti-FOXP2. All the images were acquired using ananozoomer (Hamamatsu Nanozoomer Digital Pathology, 2.0 HT,its fluorescence unit option, L11600-05, and the NanoZoomer’s3-CCD TDI camera, Hamamatsu Photonics; Cellular Imaging Facilityof the Institut de la Vision, Paris) and analyzed with a 20� objective.The numbers of ROR�-positive and FOXP2-positive PCs werecounted in the anterior lobe (lobules I-V), the posterior lobe (lobulesVI-IX), and the lobule X. The means were calculated for each animal,and then the percentage of ROR�-positive PCs in mutant animals wasnormalized to the number of ROR�-positive PCs in control animals.PC survival was evaluated by calculating the percentage of FOXP2-positive mutant PCs normalized to the number of FOXP2-positivePCs in the control animals.
Fluorescence quantification of FOXP2 PCs. As PC numbers were ob-tained by counting FOXP2-positive PC nuclei, we verified that deletionof rora did not reduce PC expression of FOXP2 and potentially confoundour quantitative analysis. At 2 months, the mean fluorescence signalintensity of FOXP2 immunolabeling was measured in single PCs. Weperformed the experiment using three different mice per genotype, and
for each of them we analyzed at least 30 PCs, in lobule III, from threevermal sections. Pictures were captured at 40� magnification with anexposure time of 500 ms. The mean fluorescence intensity per pixel wasmeasured on FOXP2-positive PC nuclei using MetaMorph.
Analysis of VGLUT2 innervation on the PCs during development. Thesections stained with Calb1, VGLUT2, and ROR� were also used tomeasure the height of molecular layer and VGLUT2 terminal distribu-tion in lobule III. Photomicrographs of lobule III from three vermalsections were taken with a 40� objective (DMR microscope, Leica). Onthese pictures, the thickness of the molecular layer in lobule III wasmeasured at every fifth PC. In parallel, on every fifth PC from threesections of the vermis, we also performed a semiquantification by classi-fying the somatodendritic distribution of the VGLUT2 puncta on the PC:(1) present only on the soma, (2) present on both the soma and thedendritic tree (soma and dendrites), or (3) present only on the dendritictree and absent from the soma of the PCs (dendrites only). The distribu-tion among the three different groups was calculated.
Analysis of VGLUT1 and VGLUT2 distribution in the adult mutantmouse. Six series of 50-�m-thick cerebellar vibratome sections from2-month-old mice, three controls (Pcp2::Cre�/�;Rorafl/fl), and three mu-tants (Pcp2::Cre�/�;Rorafl/fl) were analyzed. Sections were stained withanti-Calb1, anti-VGLUT1, or anti-VGLUT2, and anti-ROR�. Images oflobule III in the vermis were taken with a Leica SP5 confocal microscope.The 16-bit confocal images were acquired with a 63� objective, a framesize of 1024 � 1024 pixels, and a scan speed of 700 Hz (lines scanned/s),2-line averaging, and 4-zoom mode. Three regions were analyzed in thelobule III, and three vermis sections were analyzed per animal. The den-sity of VGLUT1 or VGLUT2 immunostaining was measured using Meta-Morph 5.0 image analysis system (Universal Imaging). For VGLUT1immunostaining, the density was measured within a rectangle (730�m 2) located on the middle part of the molecular layer. For VGLUT2,the density was measured in four different areas. The area termed “PCsoma” is a 400 �m 2 circle centered on a PC soma. The area termed “PCstem dendrite” is a 515 �m 2 rectangle containing a PC’s stem dendrite.The “middle part of molecular layer” is a 730 �m 2 rectangle positionedtwo-thirds up the molecular layer. The “superficial part of molecularlayer” is a 730 �m 2 rectangle located at the most superficial third of themolecular layer. For appropriate comparison, all the density measureswere normalized to 100 �m 2. The absence of ROR� immunostainingallows confirmation that the PCs were deleted for rora.
Analysis of axon torpedoes. One easily identified morphological changewas selected to study the PC axons: the torpedoes (Dusart and Sotelo,1994). Torpedoes are axonal varicosities in the granular layer with diam-eters �7 �m. On Calb1-immunostained lobule III, vermal sections from2-month-old animals (three controls Pcp2::Cre�/�;Rorafl/fl and threemutants Pcp2::Cre�/�;Rorafl/fl), we counted the number of torpedoes onat least 100 PCs with clearly visible axons.
Electron microscopic quantification of PF synapses. To count the num-ber of PF synaptic profiles, photomicrographs were taken on one ultra-thin section from each of three control and three rora-depleted mice.Photomicrographs of 7.74 �m 2 were taken at 20,500-fold magnificationevery two fields in one square of the 400 mesh grid, located just above thePC layer in the middle of lobule III. Synaptic profiles were then countedin a total of 308 photomicrographs (153 from control and 155 fromrora-depleted mice). Synaptic profiles crossing two sides of a picture wereexcluded, whereas those intersecting the two other sides were included.PF synaptic profiles were identified as presynaptic varicosities containingat least three synaptic vesicles and facing a postsynaptic density, and wereeasily differentiated from CF profiles that contain a higher density ofsynaptic vesicles in a darker matrix (Palay and Chan-Palay, 1974). Thelength of postsynaptic densities was measured using ImageJ software(Rasband, 2008).
Anterograde tracer labeling. After behavioral testing, eight mutant(Pcp2::Cre�/�;Rorafl/fl) and three control (Pcp2::Cre�/�;Rorafl/fl) micereceived an injection of anterograde tracer into the inferior olive, aspreviously described (Sugihara et al., 2003; Dixon and Sherrard, 2006).
Using the obex as a landmark, a micropipette (tip diameter �40 �m)was inserted at 50° from the vertical to a depth determined from aweight– depth curve (Sherrard et al., 1986). The anterograde tracer,
9548 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells

Figure 1. Loss of ROR� in mutant Purkinje cells. A, Strategy for the generation of conditional rora knock-out mice. The Cre-deleter line (B6.129-Tg(Pcp2::Cre)2Mpin) expresses a cre gene insertedinto exon 4 of the Pcp2/L7. LoxP sites were introduced flanking exon 4 of the rora gene. These floxed mice were bred to the Cre-deleter to generate null alleles. B–E, Photomicrographs of parasagittalcerebellar sections (B,C) and of lobule III (D,E) immunostained to reveal ROR� from Pcp2::Cre�/�;Rorafl/fl (Ctrl, B,D) and Pcp2::Cre�/�;Rorafl/fl (Mut, C,E) mice at (Figure legend continues.)
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9549

lysine fixable dextran-fluorescein solution (4% in distilled water; Fluo-roemerald, D-1820 10,000 molecular weight; Invitrogen), was injectedinto the right inferior olive. In each mouse, there was a single injection of40 nl placed either medially or mid-laterally in the caudal or mid-rostralregion of the inferior olive. The wound was cleaned, the muscles returnedto their natural position, the skin sutured, and the animals maintained ina warm box until fully recovered. At 72 h after tracer injections, animalswere perfused as described above (tissue preparation). The brainstemand cerebellum were removed, and the inferior olive and cerebellumwere cut on a vibratome. The 30 �m sections were taken; the brainstemwas always cut coronally and the cerebellum either coronally or parasag-ittally. Vibratome sections were immunostained for Calb1, ROR�, andVGLUT2. Sections were analyzed with a Nikon E800 microscope, and thelocation of olivary injections and CF terminals was mapped onto outlinesof the inferior olive and cerebellar cortex as previously described (Dixonand Sherrard, 2006). The distribution of labeled CF terminals was com-pared with the areas of labeling, which would be anticipated from thelocation of the olivary injection according to known olivocerebellar to-pography (Sugihara and Shinoda, 2004). Images were taken with a Con-focal Laser Scanning Microscope (SP5, Leica).
Electrophysiological and morphological analysis. Mice with the genotypePcp2::Cre�/�;YFPstopfl/fl;Rorafl/fl were used as controls; these were com-pared with Pcp2::Cre�/�;YFPstopfl/fl;Rorafl/fl mutant mice, which lackROR� and express YFP. Fluorescent PCs were considered to be mutantfor the purposes of electrophysiological recording.
Mice were deeply anesthetized with inhaled isoflurane and decapi-tated. Cerebellar slices (250 �m) were prepared and whole-cell patch-clamp recordings from fluorescent PCs performed as previouslydescribed (Letellier et al., 2007). Extracellular medium contained (inmM) as follows: 125 NaCl, 2.5 KCl, 1.25 NaH2PO4, 1 MgCl2, 2 CaCl2, 26NaHCO3, 25 D-glucose, saturated with 95% O2/5% CO2. Picrotoxin (100�M) was added to the extracellular recording solution to block inhibitorycurrents. Patch pipettes were filled with a solution containing the follow-ing (in mM): 120 CsD-gluconate, 13 biocytin, 10 HEPES, 10 BAPTA, 3TEACl, 2 Na2ATP, 2 MgATP, 0.2 NaGTP, pH 7.3, 290 –300 mOsm.Recordings were performed using an upright microscope (BX50WI,Olympus) at 20°C. EPSCs resulting from activation of PF-EPSCs or CF-EPSCs were elicited by stimulation with a saline-filled glass pipette in thearea surrounding the PC. CF-EPSCs were distinguished from PF-EPSCsby their all-or-none character and by the demonstration of paired-pulsedepression (Konnerth et al., 1990). For PCs in which CF-EPSCs weresuccessfully recorded at �80 mV, we subsequently depolarized the PC to�20 mV to avoid voltage-clamp escape of the CF current.
To determine the number of CFs innervating a PC, we counted thenumber of discrete CF-EPSC steps that appeared when the stimulationintensity was gradually increased and/or when the position of the stim-ulation electrode was changed.
Synaptic currents were further analyzed using Igor Pro (Wavemetrics)and the NeuroMatic program developed by Jason Rothman (UniversityCollege London). Amplitudes and rise times for the CF-EPSCs weremeasured. Several parameters were also evaluated for PF-EPSCs re-corded from control and mutant PCs, including the increase in PF-EPSC
amplitude with increased stimulus intensity (input– output relation-ship), and short-term paired-pulse facilitation.
After electrophysiological recordings, slices were fixed in 4% parafor-maldehyde in PB 0.1 M and processed for immunohistochemistry. Sliceswere incubated with streptavidin-Cy3 (1:800, Sigma) for 4 h, and thenwith anti-ROR� overnight to further confirm that the recorded PCs,filled with biocytin, lacked ROR�.
Analysis of PC dendritic trees and spines: image acquisition and morpho-logical analysis. The biocytin-filled PCs from the electrophysiologicalstudy were used for morphological analysis. Image stacks were obtainedwith a Confocal Laser Scanning Microscope (SP5, Leica) equipped with a1.4 NA objective (oil-immersion, 63�, Leica) and with the pinhole ap-erture set to 1 Airy unit. Images coded in 16 bits depth were acquired inthe 570 – 620 nm emission range and 2 images were taken and averagedfor each z-step. For whole neuron observation, the pixel size was 240 nmand z-step of 1 �m was used. Images stacks were deconvolved using aMaximum Likelihood Estimation algorithm performed with Huygens3.6 software (Scientific Volume Imaging) with 50 iterations using theo-retical PSF. For dendritic spine analysis, the pixel size was set to 60 nmand z-step to 0.2 �m. The images were deconvolved with 100 iterationsusing an experimental PSF, obtained from images of 170 nm diameterfluorescent latex beads (PS-Speck, Invitrogen) (Heck et al., 2012).
For analysis of PC morphology, a 3D model of the dendritic tree wasreconstructed using Neuronstudio (version 9.92; Rodriguez et al., 2008)(http://research.mssm.edu/cnic/tools.html) and saved in swc file format.First, the ImageJ plugin tubeness (� value of 0.8; http://rsb.info.nih.gov/ij/index.html) was applied to the image stack to reinforce tubularstructures (i.e., dendrites), and the tree was semiautomatically recon-structed with Neuronstudio; then the 3D model obtained was refined onthe original image stack. Dendrite branches were labeled according toStrahler order with Neuronstudio. Order 1 corresponds to terminalbranches, and the order number increases toward the soma. Quantitativemorphological parameters of the whole neuron were extracted usingLmeasure software (Scorcioni et al., 2008) (http://cng.gmu.edu:8080/Lm/). Sholl analysis was performed with the plugin Simple NeuriteTracer in Fiji program (http://pacific.mpi-cbg.de). For dendritic spineanalysis, segments from the upper part of the dendrites were recon-structed so a total of 150 –200 �m dendrite length per neuron was ana-lyzed. Automated spine detection followed by manual correction wasperformed using Neuronstudio. Spine density is defined as the numberof spines for 10 �m of dendrite length.
Recombinant lentiviral vectors and production. Recombinant lentiviralvectors expressing Cre or GFP under the control of the CMV promoterwere used to prepare stocks of LV-CMV-Cre and LV-CMV-GFP viralparticles as previously described (Zennou et al., 2001). Briefly, HEK 293Tcells were transiently cotransfected with the p8.91 encapsidation plasmid(Zufferey et al., 1997), the pHCMV-G (vesicular stomatitis virus pseu-dotype) envelope plasmid, and the pFlap recombinant vectors. The su-pernatants were collected 48 h after transfection, treated with DNaseI(Roche Diagnostics), and filtered before ultracentrifugation. The viralpellet was then resuspended in PBS, aliquoted, and stored at �80°C untiluse. The amount of p24 capsid protein was determined by the HIV-1 p24ELISA antigen assay (Beckman Coulter). Virus from different produc-tions averaged 175 ng/�l of p24 antigen.
Intracerebellar injections of Cre-expressing lentiviral vector into matureRorafl/fl mice and analysis of VGLUT2 distribution. Two-month-oldRorafl/fl or YFPstopfl/fl;rorafl/fl male mice were anesthetized by intraperi-toneal injection with ketamine (146 mg/kg) and xylazine (7.4 mg/kg) andplaced on a Kopf stereotaxic apparatus (Harvard Apparatus). One injec-tion (2 �l, over 8 min) per animal of either LV-CMV-Cre or LV-CMV-GFP was performed on the midline at the suture between the parietal andthe occipital bones, and 0.5 mm from pial surface to target lobule VI ofthe vermis. Two to 4 weeks after the viral injection (N � 7 for LV-CMV-Cre, N � 3 for LV-CMV-GFP) or 6 weeks (N � 6 for LV-CMV-Cre, N �5 for LV-CMV-GFP), the mice were anesthetized and perfused asdescribed above. Sagittal cerebellar vibratome sections (30 �m) wereimmunostained with Calb1, ROR�, GFP, and VGLUT2. Transduced PCswere identified by the presence of both CaBP and GFP immunostaining,and the absence of ROR� immunostaining was systematically verified.
4
(Figure legend continued.) postnatal day 10 (P10, B,C) and P21 (D,E). Asterisks indicateROR�-immunoreactive Purkinje cells. The interneurons of the molecular layer (arrows) are stillROR�-immunoreactive in P21 mutant (E). Scale bars: (in B) B, C, 340 �m; D, E, 120 �m. F, G,Number of mutant PCs expressing ROR� in different lobules. The mutant values are expressedas a percentage of their respective control values. Percentage of ROR�-immunoreactive Pur-kinje cells in the vermis (F) and in the different lobes of the vermis (lobules I-V, lobules VI-IX, andlobule X; G) over time. N � 3 mutants and 3 controls at the different ages: P10, P14, P21, and 2months. Error bars indicate SEM. F, Two-way ANOVA was applied to analyze the effects ofgenotype (F(1,19) �52.63, p �0.0001) and age (F(3,19) �8.34, p �0.001), followed by a PLSDFisher post hoc analysis. G, Three-way ANOVA was applied to analyze the effects of genotype(F(1,210) � 6604.17, p � 0.0001), age (F(3,210) � 729.43, p � 0.0001), and lobules (F(2,210) �3198.44, p � 0.0001) followed by a PLSD Fisher post hoc analysis to compare control andmutant values at the different ages and the different lobules: **p � 0.01; ***p � 0.001.
9550 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells

z-stack confocal images (acquired as described above) of the soma andthe stem dendrite of these transduced PCs were obtained with Z step of 1�m. Using ImageJ software, the number of VGLUT2 puncta directlyapposed on the soma or on the stem dendrite (first 20 �m) were countedthrough the stack of images. For the soma, we assigned PCs to threegroups: PCs with (1) 0 –2, (2) 3– 6, and (3) �6 VGLUT2 puncta on theirsoma. For the stem dendrite, we assigned PCs to four groups as follows: PCswith (1) 0–5, (2) 6–10, (3) 11–15, and (4) �16 VGLUT2 puncta on the first
20 �m of dendritic shaft. The distribution in thedifferent groups of the PCs transduced with LV-CMV-GFP (2–6 weeks), LV-CMV-Cre (2–4weeks), and LV-CMV-Cre (6 weeks) was calcu-lated.
Statistical analysis. All variables were initiallyanalyzed with the Shapiro–Wilk test to deter-mine whether the data varies significantly fromthe pattern expected if the data were drawnfrom a population with a normal distribution.Furthermore, the Levene test was performed toprobe the homogeneity of variances acrossgroups. Variables that failed the Shapiro–Wilkor the Levene test were analyzed with nonpara-metric statistics using Mann–Whitney ranksum tests for comparing two groups. Variablesthat passed the normality test were analyzed bymeans of ANOVA followed by a post hoc tests(PLSD Fisher, Scheffe, or Dunnett) analysis formultiple comparisons or by Student’s t test forcomparing two groups. Categorical variableswere compared using the Fisher’s exact test. A pvalue of � 0.05 was used as a cutoff for statisticalsignificance. Data are presented as mean � SEM.The statistical tests are described in each figurelegend.
ResultsTime course of the loss of ROR�expression in Pcp2::Cre �/�;Rorafl/fl PCsTo deplete ROR� specifically from PCs,mice with a floxed rora gene were crossedwith mice expressing Cre recombinase un-der control of the PC-specific Pcp2 genepromoter to obtain control (Pcp2::Cre�/�;Rorafl/fl) mice in which ROR� is expressed,and mutant (Pcp2::Cre�/�;Rorafl/fl) mice inwhich rora has been deleted from the PCs(Fig. 1A).
In control mice, ROR� was detected im-munohistochemically in PCs and molecularlayer interneurons at all ages tested as previ-ously described (Fig. 1B,D) (Ino, 2004). Bycontrast, in mutant mice, ROR� immuno-reactivity was present in PCs at P10 (Fig.1C), but almost entirely absent at P21 (Fig.1E), whereas the molecular layer interneu-rons continued to express ROR� (Fig. 1E),confirming the specificity of the deletion.
To determine the time course of ROR�deletion, we quantified the number ofROR�-immunoreactive PCs in the vermisof control and mutant mice. In mutants atP10, 93% of PCs expressed ROR� protein.The percentage of these ROR�-immuno-reactive PCs decreased rapidly between P10and P21, falling to only 5% at 2 months (Fig.1F). The loss of ROR� expression was not
homogeneous throughout the cerebellar cortex (Fig. 1G), occurringearlier in the anterior lobe (lobules I-V) than in the posterior lobe(lobules VI-IX), and rora deletion in lobule X was much reducedcompared with other lobules (Fig. 1G).
These data show that PC-specific ROR� depletion occurs inthis model during a relatively late stage of PC development,mainly between P10 and P21. We thus used this mutant mouse
Figure 2. Loss of ROR� in Purkinje cells strongly impaired sensorimotor abilities. Different sensorimotor tasks assessed inPcp2::Cre�/�;Rorafl/fl (Mutant) and the three types of control: Pcp2::Cre�/�;Rora�/� (wild-type control; WT),Pcp2::Cre�/�;Rora�/� (L7Cre control, L7CTRL), and Pcp2::Cre�/�;Rorafl/fl (floxed Rora control, RORCTRL). A, Examples of foot-print traces in a WT and a mutant mouse. B–G, Analyses of motor performances revealed that the absence of ROR� affectedmuscular strength (B), rearing frequency (C), dynamic balance (D,E), and motor coordination (F,G). Error bars indicate SEM.N values are indicated in the bars for each experiment. One-way ANOVA was applied to test the effect of genotype for muscularstrength (F(3,55) � 9.7, p � 0.0001), rearing frequency (F(3,39) � 9.4, p � 0.0001), walking time on the rod (F(3,55) � 3.67, p �0.05), distance traveled on the rod (F(3,55) � 9.4, p � 0.0001), the hole-board task (F(3,51) � 6.2, p � 0.005), and the time spenton the accelerating rotarod (F(3,55) � 25.1, p � 0.0001) followed by Scheffe’s post hoc analysis. Significant differences betweenmutant and all control groups, except for D, were as follows: *p � 0.05; **p � 0.005; ***p � 0.0001.
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9551

(Pcp2::Cre�/�;Rorafl/fl) to study the role played by ROR� in PCsafter their early developmental phases.
Rora deletion in PCs after the first postnatal week impairsmotor coordinationBecause defects in PC function are generally associated with im-paired motor behavior, we tested motor behavior in adult rora-deleted mice. The first signs of motor dysfunction were detected4 –5 weeks after birth, with very slight tremors during locomo-tion. The symptoms became more evident at 7– 8 weeks of age,with the development of an ataxic gait. We therefore performed acomplete motor behavior study on 2-month-old mice. A foot-print assay revealed that the mutant mice had an irregular gaitcompared with controls, with an abnormally large hindpaw anglerelative to the direction of walking, indicating reduced hindlimb
motor coordination (Fig. 2A). In addition, forepaw muscularstrength was also reduced (Fig. 2B). Analysis of locomotor activ-ity in the open field revealed that rearing frequency was drasti-cally reduced in the mutant mice compared with the three controlgroups (described in Materials and Methods; Fig. 2C). Dynamicbalance was also affected in the mutant mice as revealed by thesignificant reduction in both walking time (Fig. 2D) and dis-tance traveled on the rod (Fig. 2E). Finally, motor coordina-tion was also strongly affected by the deletion of rora fromPCs, with the mutant mice showing significantly more stum-bles than controls in the hole-board task (Fig. 2F ) and im-paired performance on the accelerating rotarod (Fig. 2G).These data indicate that mice with PC-specific rora deletionhad a strong impairment of motor abilities, presumably re-sulting from PC dysfunction and/or loss.
Figure 3. Loss of ROR� in Purkinje cells leads to cerebellar atrophy and Purkinje cell loss. A, Changes in cerebellar weight during late development; two-way ANOVA was applied to analyze theeffects of genotype (F(1,33) � 35.80, p � 0.0001) and age (F(4,33) � 11.52, p � 0.001), followed by a PLSD Fisher post hoc analysis to compare control and mutant at each age: **p � 0.01; ***p �0.001. B, C, Images of Pcp2::Cre�/�;Rorafl/fl (Ctrl, B) and Pcp2::Cre�/�;Rorafl/fl (Mut, C) brains and their cerebella at 2 months showed that rora deletion changes only the cerebellar size. D, Graphicrepresentation of the mean fluorescence intensity of FOXP2 immunostaining in Purkinje cell nuclei. Error bars indicate SEM. **p � 0.01 (Student’s t test). E–H, Photomicrographs of FOXP2-immunostained lobule III in vermis parasagittal cerebellar sections from Pcp2::Cre�/�;Rorafl/fl (Ctrl, E,G), and Pcp2::Cre�/�;Rorafl/fl (Mut, F,H) at P21 (E,F) or 2-month-old (G,H). Scale bar, 120 �m.I, J, Percentage of mutant FOXP2-immunoreactive PC in the lobules I-IV, lobules VI-IX, and lobule X of the vermis at P21 (I) and 2 months (J). Error bars indicate SEM. I, Two-way ANOVA was appliedto analyze the effects of genotype and lobules; no difference was detected between the two genotypes. J, Two-way ANOVA was applied to analyze the effects of genotype (F(1,12) � 373.12, p �0.0001) and lobules (F(2,12) � 455.37, p � 0.0001) followed by a PLSD Fisher post hoc analysis: ***p � 0.001.
9552 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells

Rora deletion after the first postnatal week reduces thenumber of PCs in the adultTo evaluate the cause of the motor dysfunction, we first examinedthe survival of rora-deficient PCs. It is known that mutant ortransgenic mice that lack functional ROR� throughout develop-ment have greatly reduced numbers of cerebellar PCs (Dussaultet al., 1998; Vogel et al., 2000; Doulazmi et al., 2001). In addition,mice haploinsufficient for rora show earlier aging-related PCdeath (Zanjani et al., 1992; Hadj-Sahraoui et al., 1997; Doulazmiet al., 1999; Janmaat et al., 2011). We thus asked whether deletionof rora after the initial period of cerebellar development also af-fected PC survival.
From P10 to P17, the weight of cerebella from mutant mice wascomparable to control littermates at the same age (Fig. 3A), but fromP21 the cerebellar weight of mutant mice remained stable, whereasthat of controls continued to increase (Fig. 3A–C). No defects infoliation were observed. When PC nuclei were labeled using FOXP2immunostaining, no differences were observed between mutant andcontrol cerebella at P21 (Fig. 3E,F), but there were many gaps in thePC layer of mutants at 2 months (Fig. 3G,H). To determine whetherthese gaps were the result of decreased FOXP2 expression in mutantPCs and thus faulty detection, we measured the fluorescent intensityof FOXP2 immunoreactivity in mutant and control PC nuclei. Infact, we actually observed increased intensity of FOXP2 immunore-activity in mutant PCs compared with controls (Fig. 3D). We there-fore used FOXP2 immunostaining to determine PC number. ThePC counts in the vermis of mutant and control animals were similarat P21 but 30% lower in the mutant at the age of 2 months (Fig. 3I).The PC loss was uniform throughout the different lobules (Fig. 3J).At P21, although ROR� protein was detectable in only 8% of the PCs(Fig. 1F), PC number in the mutant mice was similar to that ofcontrols (Fig. 3I). In addition, although loss of ROR� expressionoccurred earlier in the anterior lobe (Fig. 1G), rates of PC loss were
similar in the anterior and posteriorlobes (Fig. 3J ), indicating that PC mat-uration at the time of rora deletion didnot affect survival.
PC-specific rora deletion after the firstpostnatal week induces atrophicchanges in mature PCsThe motor dysfunction we observedcould also be the result of abnormal mor-phologic maturation of the surviving PCs.ROR� is well known to be essential fordendritic development in PCs, and specif-ically the transition from early transientbipolar dendrites to the later permanentdendritic tree (Boukhtouche et al.,2006b). We evaluated the potential role ofrora in the later stages of PC morphologi-cal maintenance. The quantification ofCalb1-immunostained cells revealed nodifference either in the size of the PC body(Fig. 4G) or the height of the PC dendritictree (Fig. 4H) between P10 mutant andcontrol mice, an age at which most PCsstill express ROR� (Fig. 4A,B). The samephenotype was found at P14 (Fig. 4G,H),an age at which most of the mutant PCsare devoid of ROR� (compare Fig. 4C,D).However, by P21, mutant mice showedsmaller PC cell bodies (Fig. 4E–G) and a
considerable reduction in the height of the PC dendritic tree (Fig.4E,F,H) associated with a reduction of the height of the molec-ular layer. Then, we evaluated the axons and dendritic trees ofadult mutant ROR�-deleted PCs.
An analysis of PC axonal morphology was performed in mu-tant and control mice at the age of 2 months. Axons in the mutantwere not entirely normal, with a higher incidence of torpedoescompared with controls (14% and 0.5%, respectively). These re-sults indicate that some PC axons were modified in the absence ofrora.
To study dendritic morphology of individual PCs, we ana-lyzed dendritic trees of PCs filled with biocytin. At P15, no dif-ferences were detected between control PCs and PCs lackingROR� (Fig. 5A), but the effects of the rora deletion were obviousat later ages. In 1-month-old animals (Fig. 5B), 30% (4 of 14) ofthe mutant PCs had a tree complexity similar to control, but witha reduced size (“small PCs”), whereas most mutant cells (70%, 10of 14) had a highly atrophic dendritic tree (“atrophied PCs”).Because these two morphological mutant phenotypes were char-acterized by different overall shape, they were separated for quan-titative 3D analysis (Fig. 5C–E). Whereas the height of thedendritic tree and the total dendritic length were highly reducedboth in small and atrophied PCs, the width of the dendritic treewas only reduced in the atrophied PCs (Fig. 5C). Sholl analysisrepresents the density of branch points at different distances fromthe soma. Compared with control PCs, small PCs had similarnumbers of branch points, but the tree was shorter, whereas theatrophied PCs had very low dendritric complexity on a very smalltree (Fig. 5D). Moreover, in agreement with a Sholl analysis, amajor reduction in the Strahler orders was observed for atro-phied PCs, whereas only one order was lost for the small PCs (Fig.5C). Our results indicate that the atrophy process in these smallPCs does not affect all parts of the dendritic complexity equally:
Figure 4. Dendritic tree of mutant Purkinje cells is reduced in height from P21. A–F, Photomicrographs of lobule III from vermisparasagittal cerebellar sections double immunostained for Calb1 (green) and ROR� (red) from Pcp2::Cre�/�;Rorafl/fl (Ctrl, A,C,E)and Pcp2::Cre�/�;Rorafl/fl (Mut, B,D,F) at P10 (A,B), P14 (C,D), and P21 (E,F). Asterisks label ROR�-immunoreactive Purkinje cellnuclei. There is the disappearance of ROR� immunostaining in mutant PCs from P14, whereas it is still present in the nucleus ofmolecular interneurons (arrowheads). Scale bar, 30 �m. G, H, Graphic representations of the surface of PC soma (G) and the PCdendritic height (H). Error bars indicate SEM. Two-way ANOVA was applied to test the effects of genotype (F(1,12) � 5.048, p �0.05); and age (F(2,12) �18.55, p�0.001) for the soma (G) and genotype (F(1,12) �36.38, p�0.0001) and age (F(2,12) �135.96,p � 0.0001) for dendritic height (H), followed by a PLSD Fisher post hoc analysis to compare between genotypes: *p � 0.05;**p � 0.01.
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9553

loss of entire proximal dendritic branches occurred rather than auniform centripetal dendritic regression.
Finally, the spine density measured in the upper dendriticbranches was highly reduced in both small and atrophied mutantPCs (Fig. 5E).
It is important to note that the general morphology of themutant PC dendritic trees appeared more mature at P15 than at 1month of age. This indicates that the deletion of rora in our modeldoes not interfere with early PC development but rather inducesthe regression of the mature dendritic morphology.
Rora deletion in PCs after the first postnatal week induces adecrease of VGLUT1 terminal density and a redistribution ofVGLUT2 terminal density on mature PCsBecause the mutant PC dendritic trees are less elaborate, withfewer branches and distal spines, we asked whether rora deletionduring the second and third postnatal weeks influences the num-ber and location of their glutamatergic presynaptic partners (i.e.,PFs and CFs). PF terminals, identified using anti-VGLUT1 im-munohistochemistry, were observed in the molecular layer ofboth control (Fig. 6A) and mutant (Fig. 6B) cerebella in
Figure 5. Morphology of mutant Purkinje cells. A, Representative control and mutant Purkinje cells filled with biocytin at P15. Neither tree size nor spine density is affected at this stage, althoughthese PCs lacked ROR�. Error bars indicate SEM. No difference was detected (width or height: Student’s t test; and spine density: Mann–Whitney test). B, Representative control, small, and atrophiedPurkinje cells with their respective 3D dendritic tree model in 1-month-old animals. The Strahler orders are color-coded on the models. The small Purkinje phenotype exhibits a particular tree shapethat is different from the atrophied cells. C, Reconstruction of dendritic trees allows measurements and analysis of width, height, total dendrite length, and Strahler orders. All measurements areperformed in three dimensions on the whole dendritic tree. D, Sholl analysis of control and mutant dendritic trees. A different pattern is observed for the two mutant phenotypes. Small Purkinje cellshave similar branch complexity but reduced height compared with control. Atrophied cell trees have small size and major loss of branches. E, Spine densities measured on upper dendrites show asimilar decrease for both mutant cell populations compared with control. Scale bar, 20 �m for all Purkinje cell images and 2 �m for all dendritic spine images. Error bars indicate SEM. For statisticalanalysis, measurements from control neurons were compared with the mutant phenotypes using one-way ANOVA for the following parameters: tree width (F(2,17) �24.14, p�0.0001), tree height(F(2,17) � 78.62, p � 0.0001), total length (F(2,16) � 143.1, p � 0.0001), Strahler order (F(2,16) � 29.67, p � 0.0001), and spine density (F(2,16) � 50.83, p � 0.0001) followed by Dunnett posthoc test. NS, Nonsignificant. *p � 0.05. ***p � 0.001. N � 6 control, 4 small, and 10 atrophied PCs.
9554 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells

2-month-old animals. However, a 40% reduction in the densityof VGLUT1 immunostaining was observed in mutants comparedwith control littermates (Fig. 6C).
Using VGLUT2 immunohistochemistry, we then determinedthe location of CF terminals on control and mutant PCs at 2months of age. In control mice, VGLUT2 staining appeared aspreviously described (Miyazaki et al., 2003): rarely around thesoma, very little on stem dendrites, and mainly distributed on theother parts of the primary dendritic compartment (Fig. 7A–C; i.e., inthe middle part of the molecular layer; Fig. 7G). In contrast, in therora-deleted mutant mice, VGLUT2-immunopositive puncta wereobserved around the PC soma and the stem dendrite (Fig. 7E,F), butseldom on the middle and upper dendrites (Fig. 7D; i.e., in the mid-dle and superficial part of the molecular layer; Fig. 7G). Densitomet-ric analysis showed that, compared with the controls, the density ofVGLUT2 immunoreactivity was 42 times higher on the soma inmutants but was less dense (one-sixth) in the mid-molecular layer(Fig. 7G). Moreover, the increased density of VGLUT2-stained ter-minals on the soma, and stem dendrite was associated with the pres-ence of thorn spines (Fig. 7F), such as those seen during the
developmental “capuchon” stage of CF development (Ramon y Ca-jal, 1911; Chedotal and Sotelo, 1992, 1993; Sugihara, 2006).
To determine at what age the altered distribution of VGLUT2-immunopositive puncta began in control and mutant PCs, wequantified the localization of VGLUT2 staining at P10, P14, andP21, determining the relative distribution of VGLUT2 stainingon the soma and dendrites. During development, VGLUT2 labelsboth climbing and PF terminals; however, it is possible to distin-guish between the large CF puncta and the diffuse PF labeling(Fig. 8). We categorized three types of distribution of VGLUT2-positive CF puncta on PCs as follows: (1) CF puncta only on thePC soma, (2) CF puncta on both the soma and the dendrites, and(3) CF puncta on the dendrites only.
At P10, large VGLUT2 puncta were seen on the proximaldendrites of both control and mutant PCs (Fig. 8), indicating thatnormal developmental somatodendritic CF translocation waswell underway in both cases. In P14 controls, most puncta wereabsent from the PC soma and were localized on the dendrites. Inthe mutant, a considerable proportion of VGLUT2-positivepuncta (27%) still contacted both the soma and the dendrite. It isimportant to note that the percentage of mutant PCs with largeVGLUT2 puncta apposed to the dendrites was higher at P14(72%) than at P21 (54%), indicating that the deletion of rora doesnot simply block CF somatodendritic translocation: the deletionreverses this process and favors the reestablishment of CF inner-vation on the soma.
Electron microscopy of excitatory synaptic inputs to PCs in2-month-old mutant miceAt 2 months, electron microscopic observations suggest no on-going degenerative process in mutant mice. Necrotic debris wereseen only rarely, facing normal-looking PF varicosities, or inphagocytic cells along with lipid inclusions. The drastically re-duced molecular layer contained large numbers of astrocytic pro-files filled with gliofilaments, most likely the result of reactivegliosis after the death of some PCs (Fig. 9F). Surviving PCs wereatrophic, with smaller somata and dendritic trees than in thecontrols, as observed in light microscopy. In addition, and as isthe case in the staggerer mutant mouse (Sotelo, 1975), the hypo-lemmal cisterna (Palay and Chan-Palay, 1974) was very poorlydeveloped. However, accumulations of short tubular and vesicu-lar profiles consistent with smooth endoplasmic reticulum,hypolemmal cisterna, were frequently observed in spine-like pro-files (Fig. 9A) as well as in medium-sized dendrites (Fig. 9B). Thelatter did not resemble the stacks of flattened cisterna reportedafter a few minutes of anoxia, which correspond to fixation arti-facts (Takei et al., 1994).
The most notable characteristic of these dendritic trees wasthe scarceness of long-necked spines, which are the postsynapticelements of PF synapses onto PCs. Spiny branchlets were practi-cally absent, although some long filopodial-like processes occa-sional emerged from smooth medium-sized dendrites (Fig. 9D).Axo-spinous synapses on PCs were much less frequent (Fig. 9A) inmutants than in controls, and some occasional synapses were estab-lished with spines emerging from large proximal branches of PCdendrites (Fig. 9D); this is similar to what is seen in PCs that have losttheir CF innervation (Sotelo et al., 1975). In the molecular layer, thePF varicosities were often apposed to neuronal perikarya or coveringthe surface of dendritic segments (Fig. 9C,J). Furthermore, some ofthe remaining PC spines and filopodia were contacted by more thanone PF (Fig. 9B) and could even be shared by a PF and a CF varicosity(Fig. 9F). Nevertheless, the density of synaptic profiles established byPF varicosities remained at 50% of control value (Fig. 9I), in ac-
Figure 6. VGLUT1 immunostaining decreases in mutant molecular layer. A, B, Confocal im-ages of vermal lobule III from 2-month-old Pcp2::Cre�/�;Rorafl/fl (Ctrl, A) andPcp2::Cre�/�;Rorafl/fl (Mut, B) parasagittal cerebellar sections stained for VGLUT1. Scale bar, 7�m. C, Graphic representation of the density of VGLUT1 immunostaining in the molecular layer:area occupied by VGLUT1 immunofluorescence per 100 �m 2. Error bars indicate SEM. **p �0.01 (Student’s t test). Three animals were analyzed for each genotype and 9 fields per animal.
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9555

cordance with the results of VGLUT1 im-munohistochemistry. This reduction ofsynaptic profile density was likely the resultof a true loss of synapses rather than theirshrinkage because there was no difference inthe size of postsynaptic densities (Fig. 9K).To determine whether the remaining PFsynapses were functional, we measured theinput–output relationship and the maxi-mum PF-EPSC amplitudes recorded incontrol and mutant PCs. There were no sig-nificant differences in these parameters be-tween the two groups (data not shown),indicating that, although PF synaptic con-tacts are differently organized on the mutantPCs, they are still functional.
As reported above with light micros-copy, the CFs innervating the mutant PCsdid not reach the upper half of the thinnermolecular layer. They rather showedcharacteristics of the immature “capu-chon” stage (Chedotal and Sotelo, 1992),with CF contacts on the PC cell body. Atthe ultrastructural level, CF varicositieswere easily identified by several morpho-logical parameters (Larramendi and Vic-tor, 1967), including the tight packing ofspherical synaptic vesicles, filling almostthe entire surface of the varicosity, to-gether with the occurrence of large gran-ulated vesicles and the dense appearanceof the axoplasm (Fig. 9E–H). AlthoughCF varicosities were not very numerous,they were systematically observed in prox-imity to the PC perikarya and in the neu-ropil of the deeper part of the molecularlayer, close to the emergence of the stemdendrites, in parallel with ascending col-laterals of basket cell axons (Fig. 9H) as incontrol PCs (Chan-Palay and Palay,1970). They were occasionally apposed tothe smooth surface of the PC somata,linked to them through attachment plates,or contacting small somatic protrusions(Fig. 9G). In the molecular layer, CF estab-lished synaptic contacts with either shortthorns emerging form presumptive PC dendrites (Fig. 9F), withhypertrophic spines forming pseudo-glomerular arrangementswrapped in several layers of thin astrocytic processes (Fig. 9E), orsometimes in close vicinity to spines emerging from primary den-dritic stems. Together, these observations showed that PCs did notpresent signs of ongoing degenerative processes, but they did have analmost-complete loss of dendritic spiny branchlets. The PF and CFinnervations were reorganized and mixed, the PFs mainly contactingthe remaining spines and the smooth surface of PC dendrites andthe CFs contacting the PC soma and stem dendrite.
Rora deletion in PCs after the first postnatal week triggersCF multi-innervationThe developmental elimination of supernumerary CF synapsesdoes not occur in the absence of ROR� (Crepel et al., 1980; Mari-ani and Changeux, 1980). However, it was not clear whether thisis strictly a developmental effect or whether late deletion of rora
later in development would have any effect once mono-innervation is achieved. Is continued ROR� expression necessaryto maintain this synaptic specificity?
In mutant mice, recombination occurs mainly between P10and P21 (Fig. 1); thus, ROR� is present during the embryonic(E14, Hamilton et al., 1996) and early postnatal period and is thenno longer synthesized starting at an age when CF synapse elimi-nation is well underway or nearly complete (Kano andHashimoto, 2009). Our morphological studies revealed dendriticchanges and particularly a reversal of the CF somatodendritictranslocation, suggesting functional changes in the CF/PC syn-apses. We thus evaluated the maturation of CF innervation toPCs in adult mutant mice using both anterograde tract tracingand electrophysiology.
Consistent with the VGLUT2 labeling and ultrastructure de-scribed above, in 2-month-old mutants, anterogradely labeledCF terminals showed abnormal structure: specifically, they were
Figure 7. Distribution of VGLUT2 immunostaining is different in mutant compared with control. A–F, Confocal images ofvermal lobule III from 2-month-old Pcp2::Cre�/�;Rorafl/fl (Ctrl, A–C) and Pcp2::Cre�/�;Rorafl/fl (Mut, D–F) parasagittal cerebel-lar sections double stained for Calb1 (green) and VGLUT2 (red). There is the absence of VGLUT2 in the superficial part of themolecular layer both for control and mutant (A,D), the decrease of VGLUT2 immunostaining in the middle part of the mutantmolecular layer (D) compared with control (B), whereas there is an increase on the mutant soma and stem dendrite (E) comparedwith control (C). F, Enlarged from E, representing two serial confocal sections. There is the presence of varicosities on the soma ofthe Purkinje cells (arrowheads). Scale bars: A–E, 15 �m; F, 7.5 �m. G, Graphic representation of the density of VGLUT2 immuno-staining in the molecular layer: area occupied by VGLUT2 immunofluorescence per 100 �m 2 (see Materials and Methods fordescription). Error bars indicate SEM. Two-way ANOVA was applied to test the effects of genotype (F(1,64) � 9.55, p � 0.001) andlocation (F(3,64) � 10.97, p � 0.001) followed by a PLSD Fisher post hoc analysis: ***p � 0.001. Three animals were analyzed foreach genotype and 9 fields per animal.
9556 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells

shorter and concentrated around the PC soma and lower den-dritic branches. Importantly, these adult mutant PCs were con-tacted by two types of CF terminals: those that were single-labeledonly by VGLUT2 and others that were double-labeled by bothVGLUT2 and Fluoremald tracer, which is consistent with inner-vation by at least two separate CF axons (Fig. 10A) (Miyazaki andWatanabe, 2011). Electrophysiological recordings demonstratedthe degree of CF multi-innervation: of 33 mutant PCs recordedfrom 5 mutant animals (1-month-old), only 12 were mono-innervated (Fig. 10 B, C). In parallel experiments using controllittermates of the mutant animals, we found no multiple CF in-nervation (N � 21 cells, from 3 animals, Fig. 10C). These resultsthus indicate that the loss of ROR� expression has a strong effecton the CF innervation state of PCs in these mice.
In some previous reports of mutant mice, a return to multipleCF innervation of PCs in the mature cerebellum has been shownto involve new CFs contacting the most distal part of the dendritictree (Miyazaki et al., 2010). The EPSCs from these distal-contacting CFs have slow kinetics, with rise times of up to 6 ms insome cases, whereas CFs contacting the normal proximal dendriticcompartment have rise times �1 ms (Miyazaki et al., 2010, their Fig.S5). We thus compared the rise times of CF-EPSCs recorded incontrol and mutant PCs to determine whether the multiple CFs
contacting the mutant PCs were in the samedendritic compartment, as suggested bytracing studies and immunohistochemistry.We found no significant differences in risetimes of CF-EPSCs between mutant andcontrol PCs: CF-EPSC rise times variedfrom 0.71 to 1.18 ms in control PCs and0.53–2.68 ms in mutant PCs (Fig. 10D).More importantly, in the multi-innervatedmutant PCs, the rise times of CF-EPSCs onthe same PC did not vary substantially, thedifference between rise times never exceed-ing 0.77 ms. Thus the multiple CFs contact-ing a mutant PC in this model share thesame functional dendritic compartment.
We next asked whether the adult multi-innervation observed in the mutant micerepresented an early-stage arrest of the de-velopmental synapse-elimination processor abnormal reinnervation by additionalCFs. Because developmental synapse elimi-nation is necessary for the refinementof normal olivocerebellar topography(Fuhrman et al., 1994), we evaluated the dis-tribution of anterogradely labeled CF axonsin adult mutant mice. Despite the presenceof multi-innervation, the topographic dis-tribution of labeled CFs was consistent withthe location of the olivary injections (datanot shown) in accordance with known ol-ivocerebellar topography (Sugihara andShinoda, 2004), suggesting that olivocer-ebellar topography had undergone normaldevelopmental regression and that multi-innervation was the result of subsequent lo-cal CF sprouting.
To verify this, we recorded from PCs ofmutant animals and their control litter-mates at P15-P16, an age at which normaldevelopmental synapse elimination is
nearly complete (Fig. 10E). We found that 87.5% of mutant PCs(N � 16 cells, n � 3 animals) were mono-innervated at this age,compared with 75% of control littermate PCs (N � 16 cells, n �3 animals); this difference is not statistically significant. Thus,mutant PCs appear to undergo a normal process of synapse elim-ination during development, and are contacted later by addi-tional CFs once ROR� is no longer expressed.
Injection of Cre-expressing lentivirus in adultrorafl/fl cerebellumTo verify that rora deletion in our model did not alter some latecomponent of PC dendritic development, we examined the effectof rora deletion in adulthood. We injected LV-CMV-GFP (as acontrol) or LV-CMV-Cre lentiviral vectors into the cerebella of2-month-old YFPstopfl/fl;rorafl/fl. The PC morphology was studiedafter survival times of either 2–4 weeks or 6 weeks. In LV-CMV-GFP–injected rorafl/fl mice, transduced PCs (GFP-immunoreactive)displayed normal morphology and VGLUT2 distribution on theirdendrites (Fig. 11D). However, in LV-CMV-Cre–injected rorafl/fl
mice, the few transduced PCs, as identified by the lack of an ROR�-immunoreactive nucleus and either the expression of Calb1 (Fig.11A,B) or the expression of YFP revealed by GFP immunohisto-chemistry (Fig. 11C), had VGLUT2-stained puncta around the soma
Figure 8. Developmental distribution of VGLUT2 immunostaining. Photomicrographs of lobules III from vermis parasagittalcerebellar sections triple immunostained for Calb1 (green), VGLUT2 (red), and ROR� (blue) from Pcp2::Cre�/�;Rorafl/fl (Ctrl) andPcp2::Cre�/�;Rorafl/fl (Mut) at P10, P14, and P21. *Purkinje cell somata in contact with VGLUT2-immunoreactive puncta. Arrow-heads indicate VGLUT2 puncta on Purkinje cell dendrites; arrows indicate PC soma lacking ROR�. Scale bar, 30 �m. Graphicrepresentation of the distribution of the VGLUT2 puncta on Purkinje cells at P10, P14, and P21. ***p � 0.001 (Fisher’s exact test).
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9557
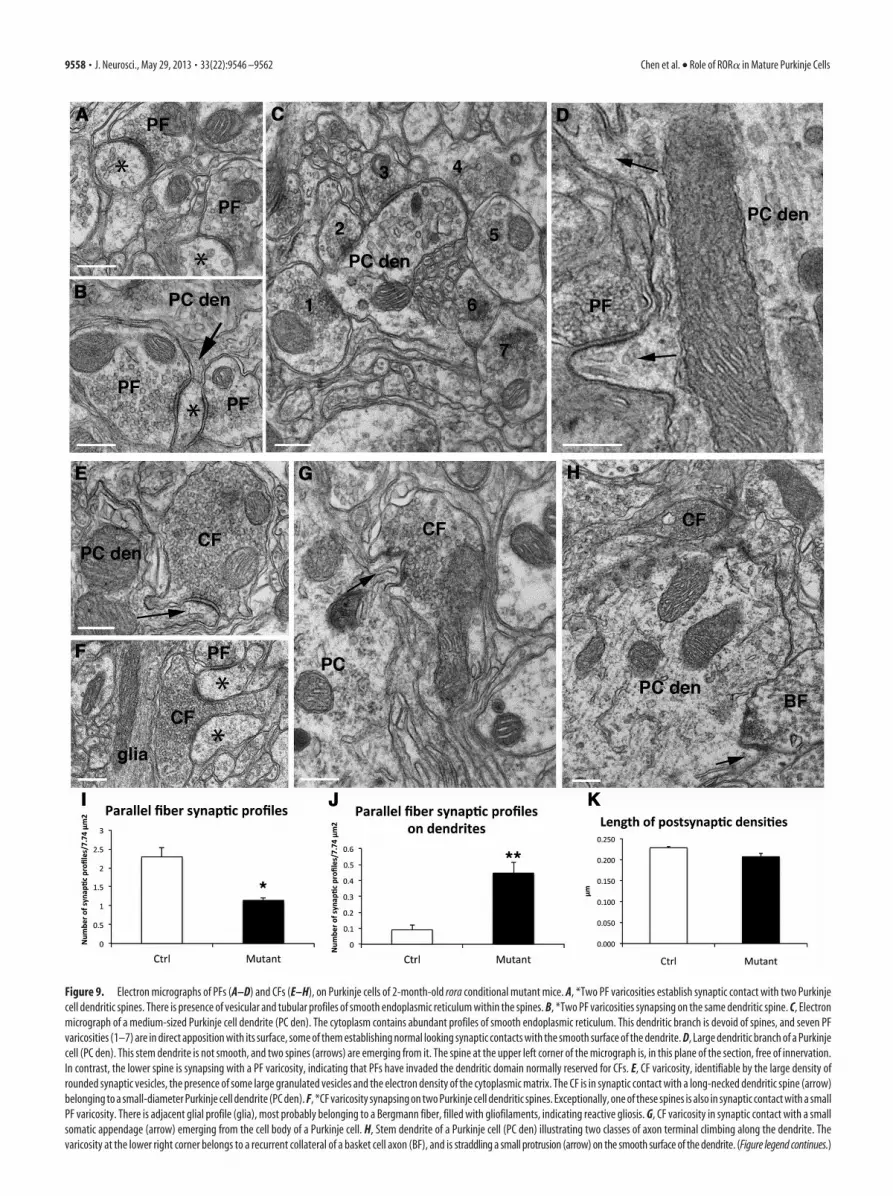
Figure 9. Electron micrographs of PFs (A–D) and CFs (E–H), on Purkinje cells of 2-month-old rora conditional mutant mice. A, *Two PF varicosities establish synaptic contact with two Purkinjecell dendritic spines. There is presence of vesicular and tubular profiles of smooth endoplasmic reticulum within the spines. B, *Two PF varicosities synapsing on the same dendritic spine. C, Electronmicrograph of a medium-sized Purkinje cell dendrite (PC den). The cytoplasm contains abundant profiles of smooth endoplasmic reticulum. This dendritic branch is devoid of spines, and seven PFvaricosities (1–7) are in direct apposition with its surface, some of them establishing normal looking synaptic contacts with the smooth surface of the dendrite. D, Large dendritic branch of a Purkinjecell (PC den). This stem dendrite is not smooth, and two spines (arrows) are emerging from it. The spine at the upper left corner of the micrograph is, in this plane of the section, free of innervation.In contrast, the lower spine is synapsing with a PF varicosity, indicating that PFs have invaded the dendritic domain normally reserved for CFs. E, CF varicosity, identifiable by the large density ofrounded synaptic vesicles, the presence of some large granulated vesicles and the electron density of the cytoplasmic matrix. The CF is in synaptic contact with a long-necked dendritic spine (arrow)belonging to a small-diameter Purkinje cell dendrite (PC den). F, *CF varicosity synapsing on two Purkinje cell dendritic spines. Exceptionally, one of these spines is also in synaptic contact with a smallPF varicosity. There is adjacent glial profile (glia), most probably belonging to a Bergmann fiber, filled with gliofilaments, indicating reactive gliosis. G, CF varicosity in synaptic contact with a smallsomatic appendage (arrow) emerging from the cell body of a Purkinje cell. H, Stem dendrite of a Purkinje cell (PC den) illustrating two classes of axon terminal climbing along the dendrite. Thevaricosity at the lower right corner belongs to a recurrent collateral of a basket cell axon (BF), and is straddling a small protrusion (arrow) on the smooth surface of the dendrite. (Figure legend continues.)
9558 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells
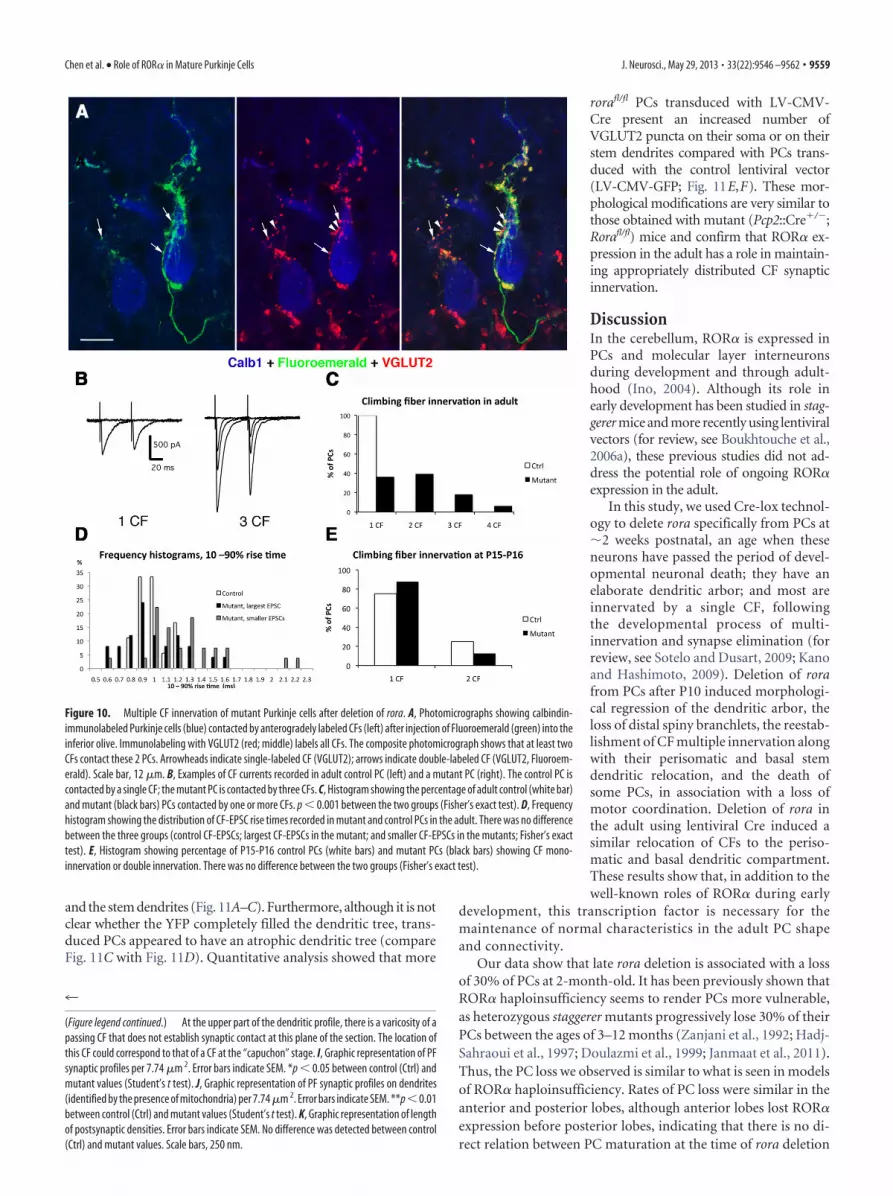
and the stem dendrites (Fig. 11A–C). Furthermore, although it is notclear whether the YFP completely filled the dendritic tree, trans-duced PCs appeared to have an atrophic dendritic tree (compareFig. 11C with Fig. 11D). Quantitative analysis showed that more
rorafl/fl PCs transduced with LV-CMV-Cre present an increased number ofVGLUT2 puncta on their soma or on theirstem dendrites compared with PCs trans-duced with the control lentiviral vector(LV-CMV-GFP; Fig. 11E,F). These mor-phological modifications are very similar tothose obtained with mutant (Pcp2::Cre�/�;Rorafl/fl) mice and confirm that ROR� ex-pression in the adult has a role in maintain-ing appropriately distributed CF synapticinnervation.
DiscussionIn the cerebellum, ROR� is expressed inPCs and molecular layer interneuronsduring development and through adult-hood (Ino, 2004). Although its role inearly development has been studied in stag-gerer mice and more recently using lentiviralvectors (for review, see Boukhtouche et al.,2006a), these previous studies did not ad-dress the potential role of ongoing ROR�expression in the adult.
In this study, we used Cre-lox technol-ogy to delete rora specifically from PCs at2 weeks postnatal, an age when theseneurons have passed the period of devel-opmental neuronal death; they have anelaborate dendritic arbor; and most areinnervated by a single CF, followingthe developmental process of multi-innervation and synapse elimination (forreview, see Sotelo and Dusart, 2009; Kanoand Hashimoto, 2009). Deletion of rorafrom PCs after P10 induced morphologi-cal regression of the dendritic arbor, theloss of distal spiny branchlets, the reestab-lishment of CF multiple innervation alongwith their perisomatic and basal stemdendritic relocation, and the death ofsome PCs, in association with a loss ofmotor coordination. Deletion of rora inthe adult using lentiviral Cre induced asimilar relocation of CFs to the periso-matic and basal dendritic compartment.These results show that, in addition to thewell-known roles of ROR� during early
development, this transcription factor is necessary for themaintenance of normal characteristics in the adult PC shapeand connectivity.
Our data show that late rora deletion is associated with a lossof 30% of PCs at 2-month-old. It has been previously shown thatROR� haploinsufficiency seems to render PCs more vulnerable,as heterozygous staggerer mutants progressively lose 30% of theirPCs between the ages of 3–12 months (Zanjani et al., 1992; Hadj-Sahraoui et al., 1997; Doulazmi et al., 1999; Janmaat et al., 2011).Thus, the PC loss we observed is similar to what is seen in modelsof ROR� haploinsufficiency. Rates of PC loss were similar in theanterior and posterior lobes, although anterior lobes lost ROR�
expression before posterior lobes, indicating that there is no di-rect relation between PC maturation at the time of rora deletion
Figure 10. Multiple CF innervation of mutant Purkinje cells after deletion of rora. A, Photomicrographs showing calbindin-immunolabeled Purkinje cells (blue) contacted by anterogradely labeled CFs (left) after injection of Fluoroemerald (green) into theinferior olive. Immunolabeling with VGLUT2 (red; middle) labels all CFs. The composite photomicrograph shows that at least twoCFs contact these 2 PCs. Arrowheads indicate single-labeled CF (VGLUT2); arrows indicate double-labeled CF (VGLUT2, Fluoroem-erald). Scale bar, 12 �m. B, Examples of CF currents recorded in adult control PC (left) and a mutant PC (right). The control PC iscontacted by a single CF; the mutant PC is contacted by three CFs. C, Histogram showing the percentage of adult control (white bar)and mutant (black bars) PCs contacted by one or more CFs. p � 0.001 between the two groups (Fisher’s exact test). D, Frequencyhistogram showing the distribution of CF-EPSC rise times recorded in mutant and control PCs in the adult. There was no differencebetween the three groups (control CF-EPSCs; largest CF-EPSCs in the mutant; and smaller CF-EPSCs in the mutants; Fisher’s exacttest). E, Histogram showing percentage of P15-P16 control PCs (white bars) and mutant PCs (black bars) showing CF mono-innervation or double innervation. There was no difference between the two groups (Fisher’s exact test).
4
(Figure legend continued.) At the upper part of the dendritic profile, there is a varicosity of apassing CF that does not establish synaptic contact at this plane of the section. The location ofthis CF could correspond to that of a CF at the “capuchon” stage. I, Graphic representation of PFsynaptic profiles per 7.74 �m 2. Error bars indicate SEM. *p � 0.05 between control (Ctrl) andmutant values (Student’s t test). J, Graphic representation of PF synaptic profiles on dendrites(identified by the presence of mitochondria) per 7.74 �m 2. Error bars indicate SEM. **p �0.01between control (Ctrl) and mutant values (Student’s t test). K, Graphic representation of lengthof postsynaptic densities. Error bars indicate SEM. No difference was detected between control(Ctrl) and mutant values. Scale bars, 250 nm.
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9559

and PC death. Furthermore, at the elec-tron microscopic level, we did not detectsigns of ongoing degeneration. Theseobservations support the proposal thatROR� is a protective agent rather than asurvival factor for PCs in adulthood,possibly by providing neuroprotectionagainst reactive oxygen species-inducedapoptosis (Boukhtouche et al., 2006c)and steroid hormone deficiency (Jan-maat et al., 2011). Our data indicate thatthe absence of ROR� will progressivelyincrease the risk of cell death over time,depending on the accumulated insultsreceived by PCs.
One clear result of the loss of ROR� inthis model is a striking reduction of thedendritic tree and torpedo axon modifica-tions, suggesting regressive events. Thenormal PC dendritic tree is compartmen-talized into a proximal compartment,composed of thick dendritic branches,giving rise to the distal compartment ofthe spiny branchlets. The proximal com-partment is the postsynaptic domain forCFs (on small rounded spines, thethorns); the distal compartment is con-tacted by PFs (on spines of the tertiarybranchlets) (for review, see Cesa andStrata, 2009). ROR� haploinsufficiency isassociated with premature dendritic atro-phy from 4 months of age (Hadj-Sahraouiet al., 2001). However, in that model, thehaploinsufficiency is present throughoutlife, and it is thus not possible to deter-mine whether this dendritic atrophy issimply a delayed effect of abnormal devel-opment. Here, with the removal of roraafter substantial development of the den-dritic tree, we observed a significant de-crease of the size of the arborization 6weeks after rora deletion at P30. This in-dicates that the atrophy in the previouslystudied haploinsufficient mice may wellbe the result of a need for ROR� in theadult to maintain the dendritic tree, ratherthan a developmental phenomenon. Thus,in addition to its role in the initial devel-opment of the dendritic tree (Boukhtou-che et al., 2006b), ROR� also seems to berequired for dendritic maintenancethroughout life.
Our results further show that the reduction of the dendritic tree isnot simply the result of nonspecific dendritic atrophy. In 30% of theanalyzed cells (the “small PCs”), mutant PCs were reduced in sizebut had a dendritic complexity near to control. These small PCs havereduced dendritic tree height but not width compared with controlPCs. It is interesting to recall that, during rat PC development, den-drite growth occurs first mainly in sagittal direction, reaching its fullwidth by P15, and then in height (Berry and Bradley, 1976). Thus, atleast for some PCs, the dendritic tree degeneration process appearsto be the reverse of the developmental pattern.
Adult mutant PCs had many fewer spiny branchlets than P15mutant PCs, suggesting that dendritic spiny branchlets are lostrather than failing to develop. In addition, spiny branchlets arealso absent from quite complex dendritic tree (small PCs), indi-cating that branchlet loss precedes, and might be independent of,branch retraction. It is known that, if ROR� is absent throughoutdevelopment, the surviving PCs lack spiny branchlets and PFsynapses (Sotelo and Changeux, 1974; Landis and Sidman, 1978).However, the multiple developmental roles of ROR� complicatethe analysis of its role in the formation and maintenance ofspines. Nevertheless, numerous observations from both normaland abnormally developed cerebella indicate that the formation
Figure 11. Effect of adult rora deletion. A, B, Photomicrograph (A) and confocal image (B) of vermis parasagittal cerebellarsections triple-immunostained for Calb1 (green), VGLUT2 (red), and ROR� (blue) from 2-month-old Pcp2::Cre�/�;Rorafl/fl in-jected with Cre lentiviral vector with a 2 week survival time. A�, The ROR� staining at the level of the PC nucleus is presented inblack and white. Double-head arrows point to the same PC nuclei between A and A�. Arrows indicate the presence of VGLUT2puncta. B, The transduced Purkinje cell (lacking an ROR�-immunoreactive nucleus, arrow) presents VGLUT2-immunoreactivepuncta (arrowhead) around its soma. C, D, Maximal projections from stacks of confocal images of vermis parasagittal cerebellarsections triple-immunostained for GFP (green), VGLUT2 (red), and ROR� (blue) from 2-month-old YFPstopfl/fl;Rorafl/fl injectedwith Cre lentiviral vector (C) or GFP lentiviral vector (D) with a 6 week survival time. C�, D�, Confocal images from the stack used tomake images in C and D, respectively. The magnification is the same in C and D, indicating that the rora-depleted PC is atrophic. Thetransduced Purkinje cells (lacking an ROR�-immunoreactive nucleus, arrows) present VGLUT2-immunoreactive puncta (arrow-heads) either around their soma or on the stem part of their main dendritic branch (C,C�), whereas a control PC (D,D�) does not.Scale bars: A, 100 �m; B, 10 �m; C, C�, D, D�, 20 �m. E, F, Histograms showing the percentage of PCs transduced withLV-CMV-GFP (white bar; control vector), or LV-CMV-Cre with 2 week to 4 week survival time (gray bars) or 6 week survival time(black bars) contacted by more or fewer VGLUT2 puncta on soma (E) and the stem dendrite (F). The number of VGLUT2 punctadirectly apposed to the soma or to the stem dendrite (first 20 �m from the cell body) were counted throughout the stack of images.In both cases, p � 0.001 (Fisher’s exact test).
9560 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells

of PC tertiary spines does not depend on presynaptic PF axonsbut are likely to be an intrinsic property of the neuron (Sotelo,1978; Yuste and Bonhoeffer, 2004). The experimental model de-scribed here allows us to delete rora in a relatively mature PC. Theobservation that these PCs then lose most of their distal spinesdespite the presence of PF terminals on PC dendrites leads us topropose that rora may be one of the important factors in themaintenance of PC distal spines throughout life.
In addition to these regressive events, late removal of rorafrom PCs induces three immature characteristics. First, these PCspresent a higher level of FOXP2 immunoreactivity comparedwith control. This is consistent with reacquisition of PC imma-turity because it is known that FOXP2 expression decreases inPCs as development proceeds (Ferland et al., 2003; Schuller et al.,2006). Second, in both mutant and virally transduced maturePCs, the location of CFs resembles the immature “capuchon”stage (Ramon y Cajal, 1911; Chedotal and Sotelo, 1993; Wa-tanabe and Kano, 2011). Similar patterns of abnormal CF loca-tion were described in several spontaneous mouse mutationsaffecting the cerebellum (Rossi et al., 1995); the CF morphologyreflects preceding interactions with their postsynaptic neuronsand is determined by the age of onset and progression of PCdegeneration.
Third, in addition to the abnormal CF localization, we ob-served a return to multi-innervation of PCs by CFs. To ourknowledge, the only other clear reestablishment of multiple CFinnervation (as opposed to a persistence of developmental multi-innervation) has been observed in the case of late deletion ofGluR�2 (Miyazaki et al., 2010). GluR�2 is involved in the main-tenance of PF/PC synapses (Takeuchi et al., 2005). In the mousemodel of GluR�2 ablation in adult PCs, Miyazaki et al. (2010)showed that this receptor is a key element for maintaining CFmono-innervation, likely by preventing the sprouting of thesefibers and their invasion of PF sites on nearby PCs. Indeed, con-siderable evidence suggests that the distribution of innervationterritory on the PC dendrites results from heterosynaptic compe-tition between PFs and CFs (Cesa and Strata, 2009). In the ab-sence of ROR�, the process of new CF multi-innervation is verydifferent: the CFs, instead of invading the PF territory, return tothe immature perisomatic net and “capuchon” stages (Ramon yCajal, 1911; Chedotal and Sotelo, 1992; Watanabe and Kano,2011). Nevertheless, the regressive morphological processes thatthese PC dendritic trees undergo, by suppressing the vast major-ity of tertiary spines, create an abnormal situation for adult PCs inwhich the partition of dendritic territory between PFs and CFs isdisturbed. This situation is similar to that described for PC-Cav2.1 KO mice, in which the extent of CF territory was limited tothe soma and basal dendrites, whereas PF territory was expanded(Miyazaki et al., 2012). However, in late rora-deleted PCs, we donot observe the hyperspiny transformation of the PC proximalsomatodendritic domain to the extent described in PC-Cav2.1mutants. Further work is necessary to understand the differentmechanisms involved in heterosynaptic competition betweenPFs and CFs and homosynaptic competition among multiple CFs(Watanabe and Kano, 2011).
Thus, the loss of ROR� in mature PCs induces the reacquisi-tion of several immature characteristics, suggesting that this tran-scription factor is important for the maintenance of the maturedifferentiation stage of PCs. We propose that ROR� is a terminaldifferentiation gene as defined by Hobert (2011), that is, part ofthe program that defines the functional properties of a mature PCthroughout its life.
ReferencesBerry M, Bradley P (1976) The growth of the dendritic trees of Purkinje cells
in the cerebellum of the rat. Brain Res 112:1–35. CrossRef MedlineBoukhtouche F, Doulazmi M, Frederic F, Dusart I, Brugg B, Mariani J
(2006a) ROR�, a pivotal nuclear receptor for Purkinje neuron survivaland differentiation: from development to ageing. Cerebellum 5:97–104.CrossRef Medline
Boukhtouche F, Janmaat S, Vodjdani G, Gautheron V, Mallet J, Dusart I,Mariani J (2006b) Retinoid-related orphan receptor � controls the earlysteps of Purkinje cell dendritic differentiation. J Neurosci 26:1531–1538.CrossRef Medline
Boukhtouche F, Vodjdani G, Jarvis CI, Bakouche J, Staels B, Mallet J, MarianiJ, Lemaigre-Dubreuil Y, Brugg B (2006c) Human retinoic acid receptor-related orphan receptor �1 overexpression protects neurones against ox-idative stress-induced apoptosis. J Neurochem 96:1778 –1789. CrossRefMedline
Cesa R, Strata P (2009) Axonal competition in the synaptic wiring of thecerebellar cortex during development and in the mature cerebellum. Neu-rosci 162:624 – 632. CrossRef Medline
Chan-Palay V, Palay SL (1970) Interrelations of basket cell axons and climb-ing fibers in the cerebellar cortex of the rat. Z Anat Entwicklungsgesch132:191–227. CrossRef Medline
Chedotal A, Sotelo C (1992) Early development of olivocerebellar projec-tions in the fetal rat using CGRP immunocytochemistry. Eur J Neurosci4:1159 –1179. CrossRef Medline
Chedotal A, Sotelo C (1993) The “creeper stage” in cerebellar climbing fibersynaptogenesis precedes the “pericellular nest”: ultrastructural evidencewith parvalbumin immunocytochemistry. Brain Res Dev Brain Res 76:207–220. CrossRef Medline
Crepel F, Delhaye-Bouchaud N, Guastavino JM, Sampaio I (1980) Multipleinnervation of cerebellar Purkinje cells by climbing fibres in staggerermutant mouse. Nature 283:483– 484. CrossRef Medline
Dixon KJ, Sherrard RM (2006) Brain-derived neurotrophic factor inducespost-lesion transcommissural growth of olivary axons that develop nor-mal climbing fibers on mature Purkinje cells. Exp Neurol 202:44 –56.CrossRef Medline
Doulazmi M, Frederic F, Lemaigre-Dubreuil Y, Hadj-Sahraoui N, Delhaye-Bouchaud N, Mariani J (1999) Cerebellar Purkinje cell loss during lifespan of the heterozygous staggerer mouse (Rora(�)/Rora(sg)) is gender-related. J Comp Neurol 411:267–273. CrossRef Medline
Doulazmi M, Frederic F, Capone F, Becker-Andre M, Delhaye-Bouchaud N,Mariani J (2001) A comparative study of Purkinje cells in two ROR�gene mutant mice: staggerer and ROR�(�/�). Dev Brain Res 127:165–174. CrossRef Medline
Dusart I, Sotelo C (1994) Lack of Purkinje cell loss in adult rat cerebellumfollowing protracted axotomy: degenerative changes and regenerative at-tempts of the severed axons. J Comp Neurol 347:211–232. CrossRefMedline
Dussault I, Fawcett D, Matthyssen A, Bader JA, Giguere V (1998) Orphannuclear receptor ROR �-deficient mice display the cerebellar defects ofstaggerer. Mech Dev 70:147–153. CrossRef Medline
Ferland RJ, Cherry TJ, Preware PO, Morrisey EE, Walsh CA (2003) Charac-terization of Foxp2 and Foxp1 mRNA and protein in the developing andmature brain. J Comp Neurol 460:266 –279. CrossRef Medline
Fuhrman Y, Thomson MA, Piat G, Mariani J, Delhaye-Bouchaud N (1994)Enlargement of olivo-cerebellar microzones in the agranular cerebellumof adult rats. Brain Res 638:277–284. CrossRef Medline
Hadj-Sahraoui N, Frederic F, Zanjani H, Herrup K, Delhaye-Bouchaud N,Mariani J (1997) Purkinje cell loss in heterozygous staggerer mutantmice during aging. Dev Brain Res 98:1– 8. CrossRef Medline
Hadj-Sahraoui N, Frederic F, Zanjani H, Delhaye-Bouchaud N, Herrup K,Mariani J (2001) Progressive atrophy of cerebellar Purkinje cell den-drites during aging of the heterozygous staggerer mouse (Rora(�/sg)).Dev Brain Res 126:201–209. CrossRef Medline
Hamilton BA, Frankel WN, Kerrebrock AW, Hawkins TL, FitzHugh W, Ku-sumi K, Russell LB, Mueller KL, van Berkel V, Birren BW, Kruglyak L,Lander ES (1996) Disruption of the nuclear hormone receptor ROR� instaggerer mice. Nature 379:736 –739. CrossRef Medline
Heck N, Betuing S, Vanhoutte P, Caboche J (2012) A deconvolutionmethod to improve automated 3D-analysis of dendritic spines: applica-tion to a mouse model of Huntington’s disease. Brain Struct Funct 217:421– 434. CrossRef Medline
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9561

Herrup K, Mullen RJ (1979) Regional variation and absence of large neu-rons in the cerebellum of the staggerer mouse. Brain Res 172:1–12.CrossRef Medline
Hobert O (2011) Regulation of terminal differentiation programs in thenervous system. Annu Rev Cell Dev Biol 27:681– 696. CrossRef Medline
Ino H (2004) Immunohistochemical characterization of the orphan nuclearreceptor ROR� in the mouse nervous system. J Histochem Cytochem52:311–323. CrossRef Medline
Janmaat S, Akwa Y, Doulazmi M, Bakouche J, Gautheron V, Liere P, Ey-chenne B, Pianos A, Luiten P, Groothuis T, Baulieu EE, Mariani J, Sher-rard RM, Frederic F (2011) Age-related Purkinje cell death is steroiddependent: ROR� haplo-insufficiency impairs plasma and cerebellar ste-roids and Purkinje cell survival. Age (Dordr) 33:565–578. CrossRefMedline
Kadkhodaei B, Ito T, Joodmardi E, Mattsson B, Rouillard C, Carta M, Mura-matsu S, Sumi-Ichinose C, Nomura T, Metzger D, Chambon P, LindqvistE, Larsson NG, Olson L, Bjorklund A, Ichinose H, Perlmann T (2009)Nurr1 is required for maintenance of maturing and adult midbrain do-pamine neurons. J Neurosci 29:15923–15932. CrossRef Medline
Kano M, Hashimoto K (2009) Synapse elimination in the central nervoussystem. Curr Opin Neurobiol 19:154 –161. CrossRef Medline
Konnerth A, Llano I, Armstrong CM (1990) Synaptic currents in cerebellarPurkinje cells. Proc Natl Acad Sci U S A 87:2662–2665. CrossRef Medline
Landis DM, Sidman RL (1978) Electron microscopic analysis of postnatalhistogenesis in the cerebellar cortex of staggerer mutant mice. J CompNeurol 179:831– 863. CrossRef Medline
Larramendi EM, Victor T (1967) Synapses on Purkinje cell spines in themouse: an electron microscopic study. Brain Res 5:15–30. CrossRefMedline
Letellier M, Bailly Y, Demais V, Sherrard RM, Mariani J, Lohof AM (2007)Reinnervation of late postnatal Purkinje cells by climbing fibers: neosynap-togenesis without transient multi-innervation. J Neurosci 27:5373–5383.CrossRef Medline
Liu C, Maejima T, Wyler SC, Casadesus G, Herlitze S, Deneris ES (2010)Pet-1 is required across different stages of life to regulate serotonergicfunction. Nat Neurosci 13:1190 –1198. CrossRef Medline
Mariani J, Changeux JP (1980) Multiple innervation of Purkinje cells byclimbing fibers in the cerebellum of the adult staggerer mutant mouse.J Neurobiol 11:41–50. CrossRef Medline
Miyazaki T, Watanabe M (2011) Development of an anatomical techniquefor visualizing the mode of climbing fiber innervation in Purkinje cellsand its application to mutant mice lacking GluR�2 and Ca(v)2.1. Anat SciInt 86:10 –18. CrossRef Medline
Miyazaki T, Fukaya M, Shimizu H, Watanabe M (2003) Subtype switchingof vesicular glutamate transporters at parallel fibre–Purkinje cell synapsesin developing mouse cerebellum. Eur J Neurosci 17:2563–2572. CrossRefMedline
Miyazaki T, Yamasaki M, Takeuchi T, Sakimura K, Mishina M, Watanabe M(2010) Ablation of glutamate receptor GluR�2 in adult Purkinje cellscauses multiple innervation of climbing fibers by inducing aberrant inva-sion to parallel fiber innervation territory. J Neurosci 30:15196 –15209.CrossRef Medline
Miyazaki T, Yamasaki M, Hashimoto K, Yamazaki M, Abe M, Usui H, KanoM, Sakimura K, Watanabe M (2012) Cav2.1 in cerebellar Purkinje cellsregulates competitive excitatory synaptic wiring, cell survival, and cere-bellar biochemical compartmentalization. J Neurosci 32:1311–1328.CrossRef Medline
Palay SL, Chan-Palay V (1974) Cerebellar cortex: cytology and organiza-tion. New York: Springer.
Polleux F, Ince-Dunn G, Ghosh A (2007) Transcriptional regulation of ver-tebrate axon guidance and synapse formation. Nat Rev Neurosci 8:331–340. CrossRef Medline
Ramon y Cajal S (1911) Histologie du Systeme Nerveux de l’Homme et desVertebres, Vol. 2. Maloine: Paris.
Rasband WS (2008) ImageJ, 1997–2008. Bethesda, MD: U.S. National Insti-tutes of Health. http://rsb.info.nih.gov/ij/.
Rodriguez A, Ehlenberger DB, Dickstein DL, Hof PR, Wearne SL (2008)Automated three-dimensional detection and shape classification of den-dritic spines from fluorescence microscopy images. PloS One 3:e1997.CrossRef Medline
Rondi-Reig L, Mariani J (2002) To die or not to die, does it change the
function? Behavior of transgenic mice reveals a role for developmental celldeath. Brain Res Bull 57:85–91. CrossRef Medline
Rondi-Reig L, Lohof A, Dubreuil YL, Delhaye-Bouchaud N, Martinou JC,Caston J, Mariani J (1999) Hu-Bcl-2 transgenic mice with supernumer-ary neurons exhibit timing impairment in a complex motor task. EurJ Neurosci 11:2285–2290. CrossRef Medline
Rondi-Reig L, Lemaigre-Dubreuil Y, Montecot C, Muller D, Martinou JC,Caston J, Mariani J (2001) Transgenic mice with neuronal overexpres-sion of bcl-2 gene present navigation disabilities in a water task. Neurosci104:207–215. CrossRef Medline
Rondi-Reig L, Arabo A, Leboucher P, Rateau S, Dupraz Y (2008) Evaluationautomatique des capacites de coordination motrices chez la souris. No. DI01873– 01. France.
Rossi F, Jankovski A, Sotelo C (1995) Target neuron controls the integrity ofafferent axon phenotype: a study on the Purkinje cell-climbing fiber sys-tem in cerebellar mutant mice. J Neurosci 15:2040 –2056. Medline
Schuller U, Kho AT, Zhao Q, Ma Q, Rowitch DH (2006) Cerebellar “tran-scriptome” reveals cell-type and stage-specific expression during postna-tal development and tumorigenesis. Mol Cell Neurosci 33:247–259.CrossRef Medline
Scorcioni R, Polavaram S, Ascoli GA (2008) L-Measure: a web-accessibletool for the analysis, comparison and search of digital reconstructions ofneuronal morphologies. Nat Protoc 3:866 – 876. CrossRef Medline
Sherrard RM, Bower AJ, Payne JN (1986) Innervation of the adult rat cere-bellar hemisphere by fibres from the ipsilateral inferior olive followingunilateral neonatal pedunculotomy: an autoradiographic and retrogradefluorescent double-labelling study. Exp Brain Res 62:411– 421. Medline
Sotelo C (1975) Dendritic abnormalities of Purkinje cells in the cerebellumof the neurological mutant mice (weaver and staggerer). In: Advances inneurology, Vol 12: Physiology and pathology of dendrites (GW Kreutz-berg, ed), 335–351. New York: Raven.
Sotelo C (1978) Purkinje cell ontogeny: formation and maintenance ofspines. Prog Brain Res 48:149 –170. CrossRef Medline
Sotelo C, Changeux JP (1974) Transsynaptic degeneration “en cascade” inthe cerebellar cortex of staggerer mutant mice. Brain Res 67:519 –526.CrossRef Medline
Sotelo C, Dusart I (2009) Intrinsic versus extrinsic determinants during thedevelopment of Purkinje cell dendrites. Neuroscience 162:589 – 600.CrossRef Medline
Sotelo C, Hillman DE, Zamora AJ, Llinas R (1975) Climbing fiber deaffer-entation: its action on Purkinje cell dendritic spines. Brain Res 98:574 –581. CrossRef Medline
Srinivas S, Watanabe T, Lin CS, William CM, Tanabe Y, Jessell TM, Costan-tini F (2001) Cre reporter strains produced by targeted insertion ofEYFP and ECFP into the ROSA26 locus. BMC Dev Biol 1:4. CrossRefMedline
Sugihara I (2006) Organization and remodeling of the olivocerebellarclimbing fiber projection. Cerebellum 5:15–22. CrossRef Medline
Sugihara I, Shinoda Y (2004) Molecular, topographic, and functional orga-nization of the cerebellar cortex: a study with combined aldolase C andolivocerebellar labeling. J Neurosci 24:8771– 8785. CrossRef Medline
Sugihara I, Lohof AM, Letellier M, Mariani J, Sherrard RM (2003) Post-lesion transcommissural growth of olivary climbing fibres creates func-tional synaptic microzones. Eur J Neurosci 18:3027–3036. CrossRefMedline
Takei K, Mignery GA, Mugnaini E, Sudhof TC, De Camilli P (1994) Inositol1,4,5-trisphosphate receptor causes formation of ER cisternal stacks intransfected fibroblasts and in cerebellar Purkinje cells. Neuron 12:327–342. CrossRef Medline
Takeuchi T, Miyazaki T, Watanabe M, Mori H, Sakimura K, Mishina M(2005) Control of synaptic connection by glutamate receptor �2 in theadult cerebellum. J Neurosci 25:2146 –2156. CrossRef Medline
van Welie I, Smith IT, Watt AJ (2011) The metamorphosis of the developingcerebellar microcircuit. Curr Opin Neurobiol 21:245–253. CrossRefMedline
Vogel MW, Sinclair M, Qiu D, Fan H (2000) Purkinje cell fate in staggerermutants: agenesis versus cell death. J Neurobiol 42:323–337. CrossRefMedline
Watanabe M, Kano M (2011) Climbing fiber synapse elimination in cere-bellar Purkinje cells. Eur J Neurosci 34:1697–1710. CrossRef Medline
Yuste R, Bonhoeffer T (2004) Genesis of dendritic spines: insights from ul-
9562 • J. Neurosci., May 29, 2013 • 33(22):9546 –9562 Chen et al. • Role of ROR� in Mature Purkinje Cells

trastructural and imaging studies. Nat Rev Neurosci 5:24 –34. CrossRefMedline
Zanjani HS, Mariani J, Delhaye-Bouchaud N, Herrup K (1992) Neuronalcell loss in heterozygous staggerer mutant mice: a model for genetic con-tributions to the aging process. Dev Brain Res 67:153–160. CrossRefMedline
Zennou V, Serguera C, Sarkis C, Colin P, Perret E, Mallet J, Charneau P(2001) The HIV-1 DNA flap stimulates HIV vector-mediated cell trans-duction in the brain. Nat Biotechnol 19:446 – 450. CrossRef Medline
Zufferey R, Nagy D, Mandel RJ, Naldini L, Trono D (1997) Multiply atten-uated lentiviral vector achieves efficient gene delivery in vivo. Nat Bio-technol 15:871– 875. CrossRef Medline
Chen et al. • Role of ROR� in Mature Purkinje Cells J. Neurosci., May 29, 2013 • 33(22):9546 –9562 • 9562a
Related Documents











