Communication Vol. 267, No . 33, Issue of November 25, pp. 23447-23450,1992 0 1992 by Th e American Society for Biochemistry and Molecular Biology, Ine. Printed i n U. S.A. THEJOURNAL OF BIOLOGICAL CHEMISTRY Protein Tyrosine Phosphatase- 1 C Is Rapidly Phosphorylated in Tyro sine i n Macrophages in Response to Colony Stimulating Factor-1" (Received for publication , Aug ust 27, 1992) Yee-GuideYeung, Karen L. Berg$, Fiona J. Pixleys, Rut h Hogue Angeletti, and E. Richard StanleyT From the Department of Deuelopmental Biology and Cancer, Albert E instein College o f Medicine, Bronx, New York 10461 An -64-kDa cytoplasmic protein is rapidly phos- phorylated in tyro sine in the response of macrophages to colony stimulating actor- 1. To identify this protein, BAC 1.2F6 macrophages were incubated with or with - out colony st imulating factor- 1, the phosp hotyrosine- containing portion of their cyt osolic fractio ns subj ected to s ize exclusion chromatography, and the 46-70-kDa fraction further fractionated by reverse phase hig h pressu re liquid chromatography (RP-HPLC). Try ptic peptides of pooled RP-HPLC fractions from stimulated cells (containing the -64- kD a protein and an -64-kDa protein ) and from unstimulate d cells (contai ning the -64-kDa protein alone) , were sequenced directly . All seven readable sequences of 8 sequenceable peptides present uniquely in the stimulated fraction were pres- ent in the sequence of the 81% homology 2 domain- containing protei n tyrosine phosphatase-lC (PT P- 1C). The identity of the -64-kDa protei n was confirmed by Western blotting with a n antibody raised to a PTP-1C peptide. The rapid, growth factor-induced tyrosin e phosphorylation of PTP -1C sugge sts that it may be involved in very early events in growth factor signal transduction. The action of the mononuclear phagocyte growth factor, colony stimu lating factor- 1 (CSF -1)l is mediated by a specific * This work was supported by National Institutes o f Health Gran t CA 26504, Albert Ei nstei n Core Cancer Gr ant P3 O- CA 1330, an d a grant from the Lucille P. Markey Charitable Trust. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "aduer- tisernent" in accordance with 18U.S.C. Section 1734 solely to indicate this fact. 2 T3 2 CA09173 . $Supported by National Institutes of Health Training Grant Fel low of th e Leu kemia Soci ety of America. II To whom correspondence should be add ress ed Dept. of Devel- opmen tal Biology and Cancer, Albert Ein stein Colle ge o f Medicin e, 1300 Morris Pa rk Ave ., Bronx, Ne w York 10461. The abbreviations used are: CSF-1, colony stimulating factor-1; PY, phosphotyrosine; aPY, anti-phosphotyrosine; CSF-lR, colony stimulating factor-1 receptor; FPLC, fast protein liquid chromatog- raphy; PBS, phosphate-buffered saline; PTPase, protein tyrosine phosphatase; PT P- lC , prote in tyros ine phosphatas e 1C; RP-HPLC, reverse phase high pressure liquid chromatogra phy; PAGE, polyacryl- amide gel electrophoresis; SH2, src homology 2 ; TBS, Tris-buff ered saline. rec eptor ( I ) , the c-fm s proto-oncogene product (2), which possess es tyrosine kinase activity (3) . Incubation of rn acro - phages with CSF- 1 causes non-covalent CSF-1 receptor (CSF- 1R) dimerization, activation, and tyrosine phosphorylation followed b y the tyrosine phosphorylation of several primarily cytop lasmic proteins (4-7). The identity of these prot eins has not been establi shed. Their appe aran ce is maximally stimu- lated by 60 s at 37 "C, but difference s in the kinetics of their appearance can be resol ved at 4 "C (6 ). They ma y be phos- phorylated directly by the activated CSF-IR or indirectly by non-receptor tyrosine kin ases t hat are activat ed as part of a signal transduction process, or their tyros ine phosphoryla tion may increase due to growth factor-induced inhibitio n of a protei n tyrosine phosphatase. As few of the tyrosine-phosphorylated proteins app ear to stably associ ate with the CSF -1R : receptor-based purifica- tion/cl oning methods ar e no t gener ally applicable. Therefore we h ave adopted the approach of identifying them directly by purification and microsequencing. Because a -64-kDa protein is markedly tyrosine-phosphorylated in macrophages in re- sponse to CSF-1, w e have focused our initial purification effort s on the CSF-1-induced tyrosine-phosphorylated pro- teins of 45-70 kDa. In this communication, we briefly describe methods fo r the purification of these proteins in sequenceable amounts and show that the -64-kDa cytoplasmic protein which is rapidly tyrosine-phosphorylated in response to CSF- 1 is protein tyros ine phosphat ase 1C ( PTP -I C). EXPERIMENTAL PROCEDURES Cel l Culture, Protein Purification, and Sequencing-Cells of the BAC1.2F5 macrophage line were cultured in 100-mm tissue culture dishes and stimu lated with CSF-1 (human rec ombinant macrophage colony stimulating factor, a gift from Chiron Corp.) at 4 "C in the presence of 2 m M io doacetic acid to increase th e yield of phosphoty- rosine-containing proteins as described previously (4, 8). The cells were then washed once with ice-c old phosphate-buffered saline (136 m M NaCI, 3 m M KCI, 8 mM NaZHPO,, 1.5 mM KH zP 04 , pH 7.4) (PB S), s craped in col d PBS containing 100pM sodium orthovanadate (Fisher Scientific), 100 p~ phenylmethylsulfonyl fluoride (Sigma) and 2 m M iodoacetic acid (Flu ka), collected in centrifuge bottles, and pelleted at 400 X g for 4 min at 4 "C. Homogenization and the subcellular fractionation were performed as described b y Yeung and Stanley (9). Phosphotyrosine (PY)-containing proteins f rom the cy- tosol of two thousand subconfluent cultures wer e prepared by affinity column chromatography using anti-phosphotyrosin e (aPY) antibody (10) coupled to Sepharose 4B (Oncogene Science). The affinity chro- matograph y was carri ed ou t wi th t he buffer s ystem of Li et al. (8 ) except that 0.8% octyl glucoside (Boehringer Mannheim) was used instead of 0.5% Nonidet P-40 in th e last 3 washes o f th e column prior to elution an d in the elution buffer. Proteins eluted with 5 mM phenyl phosphate (Sigma) were concentrated to approximately 500 pl by Centriprep 30 (Amicon) and then to 100pl by Centricon 30 (Amicon). Crystall ine guani dine-HCI (110 mg ) (P ierce Chemical Co.), 94 p1 of 2 M Tris-HC1 (Sigma), pH 8.5, and 1.3 pl of P-mercaptoethanol (Pier ce) were added to yield 190 p l of a 6 M guani dine-H C1,lOO m M Tris- HCI , 100 m M mercaptoethan ol so luti on, p H 8.5. The pro teins were reduced and denatured by incubation for 2 h at 20 "C and then overnight at 4 "C. The denatured phosphotyrosyl proteins were separated by size exclusion chromatography on a Superose S- 6 c lumn (10 X 300 mm, Pharmacia) in an FPLC system (Pharmacia) in 6 M guanidine-HC1, 50 m M Tris-HC1,0.5%dodecyltrimethylammonium bromide (Sigma), and 100 m M mercapto ethanol, pH 6.5 , a t room temperat ure with a flow rate of 0.25 ml/min. Fractions (0.3 ml) wer e collected and the protein in each fraction detected by silver staining (11) of gradient K. L. Berg, unpublished results. 23447

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

8/8/2019 J. Biol. Chem.-1992-Yeung-23447-50
http://slidepdf.com/reader/full/j-biol-chem-1992-yeung-23447-50 1/4
Communication Vol. 267, No . 33, Issue of November 25, pp. 23447-23450,19920 1992 by Th e American Society for Biochemistry and Molecular Biology, Ine.
Printed in U.S.A.
THEJOURNALOF BIOLOGICALCHEMISTRY
Protein Tyrosine Phosphatase-1 CIs Rapidly Phosphorylatedin
Tyrosine in Macrophages inResponse to Colony StimulatingFactor- 1"
(Received for publication, August 27, 1992)
Yee-GuideYeung, Karen L . Berg$,FionaJ. Pixleys, Ruth Hogue Angeletti, andE. Richard StanleyT
From the Departm ent ofDeuelopmental Biology and
Cancer, Albert E instein College of Medicine,Bronx, New York10461
An -64-kDa cytoplasmic protein is rapidly phos-phorylated in tyrosine in the response of macrophages
to colony stimulatingfactor- 1. To identify this protein,BAC1.2F6 macrophages were incubated with or with-out colony stimulating factor- 1, the phosphotyrosine-containing portion oftheir cytosolic fractions subjectedto size exclusion chromatography, and the46-70-kDafraction further fractionated by reverse phase highpressure liquid chromatography (RP-HPLC). Trypticpeptides of pooled RP-HPLCfractions from stimulatedcells (containing the-64-kDa protein and an-64-kDaprotein) and from unstimulated cells (containing the-64-kDa protein alone), were sequenced directly. Allseven readable sequences of8 sequenceable peptidespresent uniquely in the stimulatedfraction were pres-ent in the sequence of the 81% homology 2 domain-containing protein tyrosine phosphatase-lC (PTP- 1C).
The identityof the -64-kDa protein was confirmed byWestern blottingwith an antibody raised to aPTP-1Cpeptide. The rapid, growth factor-induced tyrosinephosphorylation of PTP-1C suggests that it may beinvolved in very early events in growth factor signaltransduction.
The action of the mononuclear phagocyte growth factor,colony stimulating factor-1 (CSF-1)l is mediated by aspecific
* This work was supported by National Institutes of Health GrantCA 26504, Albert Einstein Core Cancer Grant P3O-CA 1330, and agrant from the Lucille P. Markey Charitable Trust. The costs ofpublication of this article were defrayed in part by the payment of
page charges. This article must therefore be hereby marked "aduer-tisernent" in accordance with 18U.S.C. Section 1734 solelyto indicatethis fact.
2 T32 CA09173.$Supported by National Institutes of Health Training Grant
Fellow of the Leukemia Society of America.
II To whom correspondence should be addressed Dept. of Devel-opmental Biology and Cancer, Albert Einstein College of Medicine,1300 Morris Park Ave., Bronx, New York 10461.
The abbreviations used are: CSF-1, colony stimulating factor-1;PY, phosphotyrosine; aPY, anti-phosphotyrosine; CSF-lR, colonystimulating factor-1 receptor; FPLC, fast protein liquid chromatog-raphy; PBS, phosphate-buffered saline; PTPase, protein tyrosinephosphatase; PT P- lC , protein tyrosine phosphatase 1C; RP-HPLC,reverse phase high pressure liquid chromatography; PAGE, polyacryl-amide gel electrophoresis; SH2, src homology 2;TBS, Tris-bufferedsaline.
receptor (I), the c-fms proto-oncogene product (2) , whichpossesses tyrosine kinase activity (3). Incubation of rnacro-phages with CSF-1 causes non-covalent CSF-1 receptor (CSF-
1R) dimerization, activation, and tyrosine phosphorylationfollowed by the tyrosine phosphorylationof several primarilycytoplasmic proteins (4-7). Theidentity of these proteins hasnot been established. Their appearance is maximally stimu-lated by 60 s at 37 "C,but differences in the kinetics of theirappearance can be resolved at4 "C (6). They may be phos-phorylated directly bythe activated CSF-IR or indirectly bynon-receptor tyrosine kinases that are activated as part of asignal transduction process, ortheir tyrosine phosphorylationmay increase due togrowth factor-induced inhibition of aprotein tyrosine phosphatase.
As few of the tyrosine-phosphorylated proteins appear tostably associate with the CSF-1R: receptor-based purifica-tion/cloning methods are not generally applicable. Thereforewe have adopted the approach of identifyingthem directly bypurification and microsequencing. Because a-64-kDa proteinis markedly tyrosine-phosphorylated in macrophages in re-sponse to CSF-1, we have focused our initial purificationefforts on the CSF-1-induced tyrosine-phosphorylated pro-teins of 45-70 kDa. In this communication, we briefly describemethods for the purification ofthese proteins in sequenceableamounts and show that the -64-kDa cytoplasmic proteinwhich is rapidly tyrosine-phosphorylated in responseto CSF-1 is protein tyrosine phosphatase 1C (PTP-IC).
EXPERIMENTAL PROCEDURES
Cell Culture , Protein Purifica tion,and Sequencing-Cells of theBAC1.2F5 macrophage line were cultured in 100-mm tissue culturedishes and stimulated with CSF-1 (human recombinant macrophage
colony stimulating factor, a gift from Chiron Corp.) at 4 "C in thepresence of 2 mM iodoacetic acid to increase the yield of phosphoty-rosine-containing proteins as described previously (4, 8). The cellswere then washed once with ice-cold phosphate-buffered saline (136mM NaCI, 3 mM KCI, 8 mM NaZHPO,, 1.5 mM KH zP 04 ,pH 7.4)(PBS), scraped in cold PBS containing 100pM sodium orthovanadate(Fisher Scientific), 100 p~ phenylmethylsulfonyl fluoride (Sigma)and 2 mM iodoacetic acid (Fluka), collected in centrifuge bottles, andpelleted at 400 X g for 4 min at 4 "C. Homogenization and thesubcellular fractionation were performed as described by Yeung andStanley (9). Phosphotyrosine (PY)-containing proteins from the cy-tosol of two thousand subconfluent cultures were prepared by affinitycolumn chromatography using anti-phosphotyrosine (aPY)antibody(10) coupled to Sepharose 4B (Oncogene Science). The affinity chro-matography was carried out with the buffer system of Li et al. (8)except that 0.8% octyl glucoside (Boehringer Mannheim) was usedinstead of 0.5% Nonidet P-40 inthe last 3 washes of the column prior
to elution and in the elution buffer. Proteins eluted with 5 mM phenylphosphate (Sigma) were concentrated to approximately 500pl byCentriprep 30 (Amicon) and then to100pl by Centricon 30 (Amicon).Crystalline guanidine-HCI (110 mg) (Pierce Chemical Co.), 94 p1 of 2M Tris-HC1 (Sigma), pH8.5, and 1.3 pl of P-mercaptoethanol (Pierce)
were added to yield 190p l of a 6M guanidine-HC1,lOO mM Tris-HCI,100 mM mercaptoethanol solution, p H 8.5. The proteins were reducedand denatured by incubation for 2 h at 20 "C and then overnight at4 "C. The denatured phosphotyrosyl proteins were separated by sizeexclusion chromatography on a Superose S-6 column (10 X 300 mm,Pharmacia) in an FPLC system (Pharmacia) in 6M guanidine-HC1,50 mM Tris-HC1,0.5%dodecyltrimethylammonium bromide (Sigma),and 100 mM mercaptoethanol, pH 6.5, a t room temperature with aflow rate of 0.25 ml/min. Fractions (0.3 ml) were collected and theprotein in each fraction detected by silver staining (11) of gradient
K. L. Berg, unpublished results.
23447

8/8/2019 J. Biol. Chem.-1992-Yeung-23447-50
http://slidepdf.com/reader/full/j-biol-chem-1992-yeung-23447-50 2/4
23448 CSF-1 Induces P TP -I C Tyrosine Phosphorylation in Macrophages
(7.5-17.5% acrylamide) sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) gels (12). Fractions containing proteinsof interest were pooled and concentrated to100 pl (Centricon 10,Amicon) at 18"C. The concentrate was diluted with 1 ml of 6 M
guanidine-HCI, containing 0.8% octyl glucoside and concentratedagain to 100 pl twice more at 18"C. It was then acidified with 20%trifluoroacetic acid (Pierce) to <pH 2, diluted with an equal volume
of 0.1% trifluoroacetic acid, centrifuged to removed debris, and in-jected into a C, reverse phase high performance liquid chromatogra-
phy (RP-HPLC) column (2.1X
30 mm, BU300, Brownlee). Thecolumn was washed and developed with a gradient of 35-50% aceto-nitrile (Burdick and Jackson) in 0.1% trifluoroacetic acid at roomtemperature for 30 min with a flow rate of 0.2 ml/min in aHP1090M
system (Hewlett Packard) equipped with a diode array detector.
Fractions (100 pl) were collected, and 2 p1 from each fraction were
used for SDS-PAGE and silver staining. Fractions containing theproteins of interest were pooled and either digested with trypsin
(sequencing-grade, Boehringer Mannheim) immediately after dilu-
tion or pyridylethylated and purified once more on the same column
(13). The tryptic digest was separated on aCS RP-HPLC column (2.1X 30 mm, RP300, Brownlee) using a gradient of5 4 0 % acetonitrile
in 0.1% trifluoroacetic acid in 100 min with a flow rate of 0.05 ml/min a t room temperature. Absorption peaks a t 214 nm were collected
manually, and sequences of the peptides in these fractions weredetermined using an Applied Biosystems 477A protein sequenator. Adata base search was carried out to compare these sequences with
those stored in the GenEMBL data bank using the Wisconsin Ge-
netics Computer Group (WGCG) sequence analysis package (14).
Western Blotting-Western blotting was performed as described(X) , with the following modifications: Immobilon membrane (poly-vinylidene difluoride, Millipore) was used and the transfer buffercontained 0.05% SDS and10% methanol. The transfer was carried
out at30 V for 2 h, then overnight a t 80V a t 5 "C. For the detection
of PY-containing proteins the blot was blocked with 5%bovine serumalbumin (ICN), in 10 mM Tris-HCI, 140 mM NaCI, pH 7.2 (Tris-
buffered saline, TBS) at4 "C overnight. After washing the blot with
TB S containing 0.05% Nonidet P-40 (Sigma), it was incubated with
the peroxidase-conjugated anti- PY antibody (ICN) in T BS contain-ing 0.05% Nonidet P-40 for 2 h a t room temperature and developedwith enhanced chemiluminescence (ECL) reagents(Amersham). The
tyPTP-1C Western blotting was carried out as above except t ha t5%nonfat dry milk replaced bovine serumalbumin, and peroxidase-
conjugated goat anti-rabbit antibody (Amersham) was used to detectth e bound rabbit LuPTP-1C antibody. The LuPTP-IC antiserum (a giftfrom Drs. Jiao-Ti Huang and Joseph Schlessinger, Department of
Pharmacology, New York University Medical Center, New York) wasraised to a unique carboxyl-terminal peptide (KREEKVKKQRS-
ADKEKS).
RESULTS
To identify the proteinswhich are tyrosine-phosphorylated
in the response of BAC1.2F5 macrophages to CSF-1, affinity-
purified aPY-reactive proteins from CSF-1-stimulated (Fig.
1,lane 2 )and unstimulated (Fig.1,lane 1) cells wereseparated
under reducing and denaturing conditions by size exclusionchromatography.The fractions containing a highly stimulated
-64-kDa protein of interest (Fig. 1, lane 13 )and correspond-ing fractions from the unstimulated cells (Fig.1,lane 12 )were
separately pooled and further fractionated by RP-HPLC.Asshown in Fig. 1, the -64-kDa protein was recovered exclu-
sively inthe fractions from stimulated cells (lanes20-23) and
was almost completely resolved from a protein of -54-kDa
found in the fractions from unstimulated cells (lanes 4-9).Fractions run in lanes 20-23 (Fig. l),containing the -64 kDa
protein, were pooled and pyridylethylated prior to rechroma-
tography on RP-HPLC. While relatively pure when examined
by SDS-PAGE and silver staining, the amountof pyridyleth-
ylated protein recovered (approximately 7 pmol) was not
sufficient to obtain a definitive amino acid sequence from
tryptic peptides. A second preparation was obtained in which
th e separation of the -64-kDa from the -54-kDa protein(approximately 30 pmol of each) was not achieved. However,
as the major difference betweenthe stimulated and unstimu-
1
10-
5 - ,
0 J J
.,
L -
1 3 5 7 9 11 I 3 IS 17 19 21 23
205 ' a
116 4
98
m
4!
31
,+-64
"54
-+""+-+-
FIG.1. Cc RP-HPLC fractionation of the 45-70-kDapro-teins from the aPY-reactive fraction of cytosols fromBAC1.2F5 cells. Cells were incubated for 2 h at 4"C with (+) orwithout (-) 4.4 nM human recombinant CSF-1 priorto fractionation
as described under "Experimental Procedures." The upper panelshows portions of the RP-HPLC absorption profiles together withthe acetonitrile gradient(- - -) in the range 37.25-39.50%. The lowerpanel shows the silver-stained SDS-PAGEof the RP-HPLCfractionsaligned with the absorption profiles shown in the upper panel. Lanes1 and 2, LuPY-reactive fractions; lanes 12and 13,45-70-kDa proteinsfrom the Superose 6 FPLC separations of the aPY-reactive fractions
that were applied to the RP-HPLCcolumn.
lated cell fractions was the presence of the -64-kDa protein
in the fraction from stimulated cells, we sequenced the tryptic
fragments that were unique to the latterfraction. Both stim-ulated (Fig. 2 A , inset) and unstimulated (Fig. 2B,inset) cellfractions were individually digested with trypsin and the
resulting tryptic peptides separated by RP-HPLC (Fig. 2).
Eleven peaks, apparently unique to the stimulated cell frac-
tion (Fig. 2 A , 1-11) were sequenced. Peaks 1-4 and 6-8
contained sequences that were highly homologous to se-quences within PTP-1C (TableI), peaks 9-11 failed to yield
a sequence, and thesequence from peak 5 was uninterpretable.Three peaks present in both unstimulated and stimulated
fractions (peaks A , B, and C,Fig. 2,panels A and B ) were
sequenced from each fraction. Sequences obtained from peaks
A, B, and C from the stimulated fraction were identical tothose from the unstimulated fraction. Peak B from the stim-
ulated fraction also contained a PTP-1Csequence not presentin the unstimulated fraction (data not shown).
To confirm the identity of the -64-kDa protein, the aPY-
8/8/2019 J. Biol. Chem.-1992-Yeung-23447-50
http://slidepdf.com/reader/full/j-biol-chem-1992-yeung-23447-50 3/4
CSF-1 Induces PTP -1C Tyrosine
A 907 li”3
zc5:
38 4
28 30 40 50 60 70Tlmc ( m l n . )
B I
20 30 48 5 6 60 7 0 8 0
T lm e (min. 1
FIG. 2. C. RP-HPLC chromatography of tryptic digests ofpooled fractions from theC4 RP-HPLC purification step. A,CSF-1-stimulated;€3, unstimulated. Insets show silver-stained SD S-PAG E lan es for sam ples that were subjected to tryptic digestion.
TABLEI
Comparison of the sequencesof trypticpeptidesfrom the -64-kDaprotein with those predicted for murinePTP-IC
SourceAmino acid
SeauencePTP-1C-64-kDa, peak 1”
-64-kDa, peak 2
-64-kDa, peak 3
-64-kDa, peak 4
-64-kDa, peak 6
-64-kDa, peak I
-64-kDa. Deak 8
PTP-1C
PTP-IC
PTP-1C
PTP-1C
PTP-1C
PTP-1C
WGLYSVTNSR
WGLYSVTNS
NILPFDHSR
NILPFXXS~
DSNIPGSDYINANYW
XSNIPGXDYXNANYW
IQNSGDFYDLYGGEK
IQNXXFYXLYGGEK
GEPWTFLVR
GXPXIFLV
AGFWEEFESLQK
AGFXEXF’XXLQ
ESLSQPGDFVLSVLNDQPK
ESLXIPGXFVLSVLN
a Peak number, Fig.lA.* Amino acids whoseidentities were ambiguous are m arkedX.
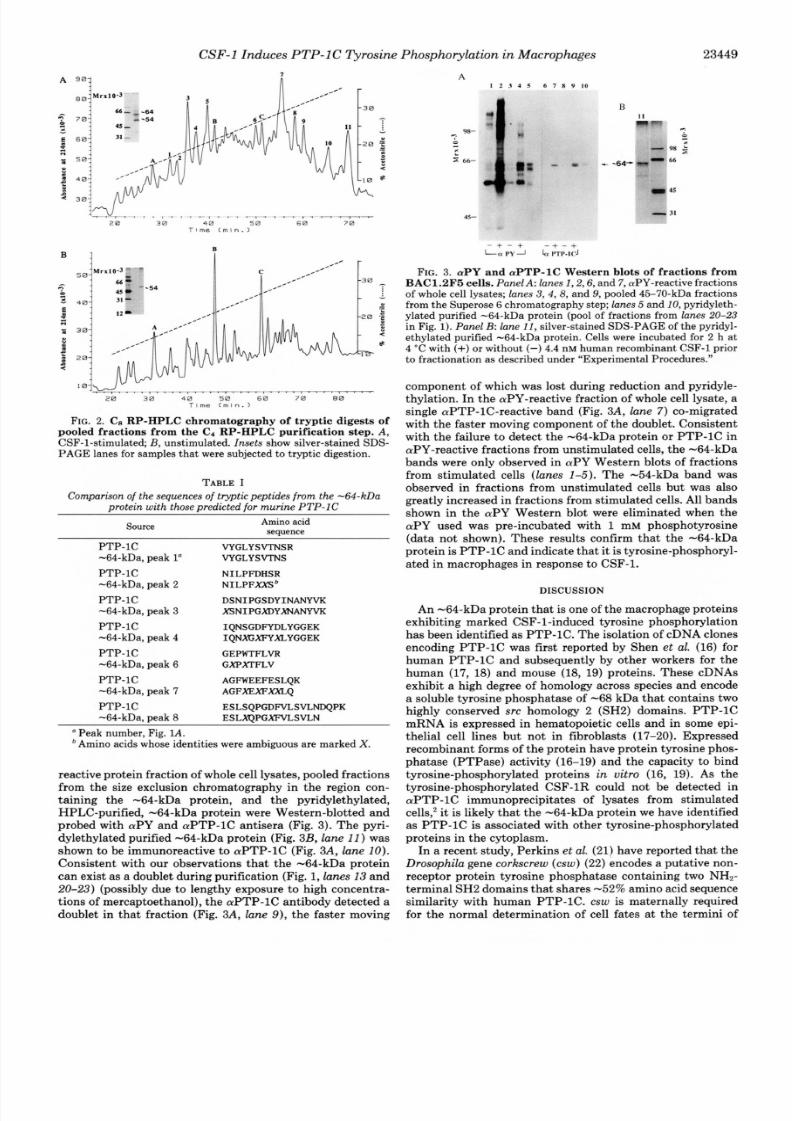
reactive protein fraction of whole cell lysates, pooled fractionsfrom the size exclusion chromatography in the region con-taining the -64-kDa protein, and the pyridylethylated,HPLC-purified, -64-kDa protein were Western-blotted andprobed with aPY and aPTP-1C antisera (Fig.3 ) . The pyri-dylethylated purified -64-kDa protein (Fig. 3B, lane 11)was
shown to be immunoreactive to aPTP-1C(Fig. 3A, lane 10).
Consistent with our observations that the -64-kDa protein
can exist as a doublet during purification (Fig. 1,lanes 13 and
20-23) (possibly due to lengthy exposure to high concentra-tions of mercaptoethanol), the aPTP-1Cantibody detected adoublet in that fraction (Fig. 3A, lane 9) , the faster moving
Phosphorylation in M acrophages 23449
A1 2 3 4 5 6 7 8 9 1 0
45-
15
- + - + - + - +Lrz PY J h ITP-ICJ
FIG. 3. aPY and aPTP-1C Western blots of fractions fromBAC1.2F5 cells. PanelA: lanes I , 2,6, and 7,aPY-reactive fractionsof whole cell lysates; lanes 3 ,4 ,8 , and 9, pooled 45-70-kDa fractionsfrom the Superose 6 chromatographystep; lanes 5 and 10,pyridyleth-ylated purified -64-kDa protein (pool of fractions from lanes20-23
in Fig. 1). Panel B: lane 11,silver-stained SDS-P AGE of the pyridyl-ethylated purified -64-kDa protein. Cells were incubated for 2 h at4 “C with (+) or without (-) 4.4 nM human recombinantCSF-1 prior
to fractionation as described under “Experimental Procedures.”
component of which was lost during reduction and pyridyle-thylation. In theaPY-reactive fraction of whole cell lysate, asingle aPTP-1C-reactive band (Fig. 3A, lane 7 ) co-migratedwith the faster moving component of the doublet. Consistentwith the failure to detect the -64-kDa protein or PTP-1C inaPY-reactive fractions from unstimulated cells, the -64-kDabands were only observed in aPY Western blots of fractionsfrom stimulated cells (lanes 1-5). The -54-kDa band wasobserved in fractions from unstimulated cells but was alsogreatly increased in fractions from stimulated cells. All bandsshown in the aPY Western blot were eliminated when theaPY used was pre-incubated with 1 mM phosphotyrosine
(data not shown). These results confirm that the -64-kDaprotein is PTP-1C andindicate that itis tyrosine-phosphoryl-ated in macrophages in response to CSF-1.
DISCUSSION
An -64-kDa protein that is one of the macrophage proteinsexhibiting marked CSF-1-induced tyrosine phosphorylationhas been identified as PTP-1C. The isolation of cDNA clonesencoding PTP-1C was first reported by Shen et al. (16) forhuman PTP-1C and subsequently by other workers for thehuman (17, 18) and mouse (18, 19) proteins. These cDNAsexhibit a high degree of homology across species and encodea soluble tyrosine phosphatase of -68 kDa that contains two
highly conserved SI % homology 2 (SH2) domains. PTP-1CmRNA is expressed in hematopoietic cells and in some epi-thelial cell lines but not in fibroblasts (17-20). Expressedrecombinant forms of the protein have protein tyrosine phos-phatase (PTPase) activity (16-19) and the capacity to bindtyrosine-phosphorylated proteins in vitro (16, 19). As thetyrosine-phosphorylated CSF-1R could not be detected inaPTP-1C immunoprecipitates of lysates from stimulatedcells: it is likely that the-64-kDa protein we have identifiedas PTP-1C is associated with other tyrosine-phosphorylatedproteins in the cytoplasm.
In a recent study, Perkins et al. (21) have reported that theDrosophila gene corkscrew (csw) (22) encodes a putative non-
receptor protein tyrosine phosphatase containing two NH2-
terminal SH2domains that shares -52% amino acid sequencesimilarity with human PTP-1C. csw is maternally requiredfor the normal determination of cell fates at the termini of

8/8/2019 J. Biol. Chem.-1992-Yeung-23447-50
http://slidepdf.com/reader/full/j-biol-chem-1992-yeung-23447-50 4/4
23450 CSF-1 Induces PT P-1 C Tyrosine Phosphorylation in Macrophages
the embryo. The determination of terminal cell fates is regu-lated by a receptor tyrosine kinase, torso. Genetic evidence
indicates that csw acts downstream of torso, in concert with
the serine/threonine kinase D- raf to regulate the downstream
terminal gene encoding the transcription factor, tailless (re-
viewed in Ref. 23) .
Analogous to the regulation of D-raf by torso, the CSF-1R
has been shown to regulate RAF-1 in BAC1.2F5 cells (24).
CSF-1 stimulation causes maximum protein tyrosine phos-phorylation of proteins, including the -64-kDa protein (PTP-lC), within 30 s at 37 "C (6, 8),' followed by an increase in
the serine phosphorylation of RAF-1 that is apparent at 1.5
min but of maximum intensity by 5-15 min after CSF-1
addition (24). RAF-1 serine phosphorylation is temporally
correlated with activation of a RAF-1associated serine kinase
activity (24). These experiments indicate tha t the tyrosine
phosphorylation of PTP-1C precedes the serine phosphoryl-
ation andactivation of the RAF-1 kinase. We have also shown
that epidermal growth factor receptor phosphotyrosine de-
phosphorylation does not affect the increase in RAF-1 serine
phosphorylation during the response to epidermal growth
factor (25), suggesting that there is atleast one step between
growth factor receptor activation and phosphorylation andactivation of RAF-1. If tyrosine phosphorylation of PTP-1C
has a role in regulating its enzyme activity, either by intra-
molecular interaction of the SH 2 domain with a tyrosine
phosphorylation site, as suggested for the src-related family
of nonreceptor tyrosine kinases (reviewed in Ref. 26) or by
modulating the interaction of PTP-1C with other proteins,
then these observations would favor the firstof three models
suggested by Perkins et al.( X ) , in which csw acts upstream
of D-rat. Future studies on the associationof PTP-1C with
other tyrosine-phosphorylated proteins, its role in mitogenic
signaling, and the effect of tyrosine phosphorylation on i ts
catalytic activity should greatly increase our understanding
of how this enzyme is involved in the early events regulated
by growth factors.This report also briefly describes methods that can be used
to identify other proteinsthat aretyrosine-phosphorylated in
the growth factor response. The procedures described are
suitable for proteins possessing a molecular mass of less tha n
70 kDa. For proteins with molecular masses significantly
above 70 kDa, significant losses are encountered in the HPLCstep.
Acknowledgments-We thank Drs. Jiao-Ti Huang and JosephSchlessinger for kindly providing the aPTP-1C antiserum, Dr.R a y -mond Frackleton for advice, and the various members of the labora-tory (past andpresent) who assisted in the preparation of cell extracts.The sequencing was performed at theLaboratory for Macromolecular
Analysis of the Albert Einstein College of Medicine. We thank YuanShi for careful handling of the samples and interpretation of the data.
REFERENCES
2. Sherr , C. J., Rhtenmier, C. W., Sacca, R., Roussel, M. F., Look, A.T.,and1. Guilbert, L. J. and Stanley, E.R. (1980)J.Cell Biol. 85 , 153-159
3. Rettenmier, C. W., Chen,J. H., Roussel, M. F., and Sherr, C.J. (1985)Stanley, E.R. (1985) Cell 41 , 665-676
Science 2 2 8 . 3 2 0 - 3 2 24. Li, W., a nds tan ley ,E.R. (1991) E M B O J. 10 , 277-2885. Bacearini, M., Li, W., Dello Sbarba , P., and Stanley,E. R. (1991) Receptor
I ~~~ ~~~
6. Sengupta, A,, Liu, W.-K., Yeung, Y.-G., Yeung,D. C.-Y., Frackelton, A. R. ,
7. Downing,J. R., Rettenmier, C. W. , and Sherr, C.J. (1988) Mol. Cell. Biol.
8. Li, W., Yeung, Y. G., and Stanley, E. R.(1991)J.Biol. Chem. 2 6 6 , 6 8 0 8 -
1,243-259
and Stanley, E.R. (1988) Proc. Natl. Acad. Sei.U .S. A . 85,8062-8066
8,1795-1799
&A14
9.
10 .
11 .12 .13 .
14 .
15 .
16.
Ye&-& Y.G., and Stanley, E.R. (1990) in Receptor Purification(Litwack,
Huhn, R. D., Posner, M. R., Rayten, S. I., Foulkes, J. G., and Frackelton,
Morrissey, J. H. (1981) Anal. Biochem. 117 , 307-310Laemmli, U . K. (1970) Nature 227 , 680-685Renlund, S., Klintrot, I. , Nunn, M., Schrimsher, J. L., Wernstedt, C., and
Devereux, J., Herbert, M., and Smithies,0.(1984) Nucleic Acids Res.1 2 ,
Towbin, H., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sci.
Shen, S.-H., Bastien, L., Posner, B. I., and Chrhtien, P. (1991) Nature 362,
G. , ed) pp. 315-328, Humana Press, Clifton, NJ
A. R., Jr. (1987) Proc. Natl. Acad. Sei. U.S. A . 84,4408-4412
Hellmann, U. (1990)J.C h r o m t o g r . 512 , 325-335
387-395
U.S. A. 76 , 4350-4354
7 R f i " n Q
17 . Plutzky, J., Neel, B. G., and Rosenberg, R. D. (1992) Proc. Natl.Acad. Sei.
18 . Yi, T.,Cleveland, J. L., and Ihle, J. N. (1992) Mol. Cell. Biol.12 , 836-84619 . Matthews, R. J., Bowne, D. B., Flores, E., and Thomas,M.L. (1992) Mol.
20 . Yi, T.,Cleveland, J. L., and Ihle, J. N. (1992) Blood 78,2222-222821 . Perkins, L. A,, Larsen, I., and Perrimon, N.(1992) Cell 70 , 225-23622 . Perrimon, N., Engstrom, L., and Mahowald, A. P. (1985) Genetics 1 1 1 ,
."" .""U.S.A. 89 , 1123-1127
Cell. Bid . 12 , 2396-2405
23-41
23 . St. Johnston,D. , and Nusslein-Volhard, C. (1992) Cell 68 , 201-21924 . Baccarini, M., Sabat ini, D. M., App, H., Rapp,U . R., and Stanley, E. R.
25 . Baccarini, M. , Gill, G. N., and Stanley, E. R.(1991) J. Biol. Chem. 2 6 6 ,(1990) EM BO J.9,3649-3657
26. Cantley, L. C., Auger, K. R., Carpenter, C., Duckworth, B., Graziani, A.,10941-10945
Kappeller, R., and Soltoff, S. (1991) Cell 6 4 , 281-302
Related Documents











