Œil et Physiologie de la Vision - IV IV- LES VOIES VISUELLES : RAPPELS ANATOMO- FONCTIONNELS Florence Rigaudière Jean-François Le Gargasson Eliane Delouvrier Pour citer ce document Florence Rigaudière, Jean-François Le Gargasson et Eliane Delouvrier, «IV- LES VOIES VISUELLES : RAPPELS ANATOMO-FONCTIONNELS», Oeil et physiologie de la vision [En ligne], IV-Les voies visuelles, mis à jour le 18/06/2013, URL : http://lodel.irevues.inist.fr/oeiletphysiologiedelavision/index.php?id=209, doi:10.4267/oeiletphysiologiedelavision.209 Plan Spécificités du pôle sensoriel L’œil Les milieux antérieurs Le pôle postérieur Les voies visuelles de conduction Zone rétrobulbaire : les nerfs optiques Le canal optique : un passage périlleux Zone de décussation des voies visuelles Les tractus optiques Les corps géniculés latéraux Les radiations optiques vers les aires visuelles primaires Les aires visuelles primaires : 80 % de la projection fovéale Pour information : les aires extrastriées… Spécificité du pôle moteur Systèmes oculomoteurs Coordinations des deux yeux Retentissement sur l’EOG Retentissement sur l’ERG flash En pratique… Conclusion 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Œil et Physiologie de la Vision - IV
IV- LES VOIES VISUELLES : RAPPELS ANATOMO-FONCTIONNELS
Florence Rigaudière
Jean-François Le Gargasson
Eliane Delouvrier
Pour citer ce document
Florence Rigaudière, Jean-François Le Gargasson et Eliane Delouvrier, «IV- LES VOIES
VISUELLES : RAPPELS ANATOMO-FONCTIONNELS», Oeil et physiologie de la vision [En
ligne], IV-Les voies visuelles, mis à jour le 18/06/2013, URL :
http://lodel.irevues.inist.fr/oeiletphysiologiedelavision/index.php?id=209,
doi:10.4267/oeiletphysiologiedelavision.209
Plan
Spécificités du pôle sensoriel
L’œil Les milieux antérieurs Le pôle postérieur
Les voies visuelles de conduction Zone rétrobulbaire : les nerfs optiques Le canal optique : un passage périlleux Zone de décussation des voies visuelles Les tractus optiques Les corps géniculés latéraux Les radiations optiques vers les aires visuelles primaires Les aires visuelles primaires : 80 % de la projection fovéale Pour information : les aires extrastriées…
Spécificité du pôle moteur
Systèmes oculomoteurs Coordinations des deux yeux Retentissement sur l’EOG Retentissement sur l’ERG flash En pratique…
Conclusion
1

Œil et Physiologie de la Vision - IV
Texte intégral
Remerciements : Pr Patrick Barbet, Marie-Pierre Beaunoir, Dr S-Yves Cohen, Dr Philippe
Lanthony, Nikola Markov, Dr Th NGuyen, Jean-Philippe Onolfo, François Vital-Durand, Dr
Samir Zeki pour leurs contributions aux illustrations et Dr Philippe Cayrol, Dr Robert
Verdet et Dr Catherine Vignal pour leurs précieuses informations.
Ce chapitre diffère d’une description classique d’anatomie visuelle. Il rappelle quelques
points particuliers de notre système visuel qui peuvent moduler les signaux
électrophysiologiques décrits dans les chapitres suivants : Electro-oculogramme : EOG
(V-2), Electrorétinogrammes : ERG-flash (V-3), ERG multifocal (V-4-A), ERG Pattern (V-
4-B) ou Potentiels évoqués visuels corticaux : PEV flash et damier (V-5).
Spécificités du pôle sensoriel
L’œil
Les milieux antérieurs
Avant d'atteindre la rétine, les rayons lumineux traversent les milieux antérieurs
composés °d’un certain nombre de dioptres liés à la cornée -film lacrymal, dioptre
cornéen antérieur et postérieur- et au cristallin -ensemble des dioptres antérieurs et
postérieurs- et °de milieux transparents avasculaires : l’humeur aqueuse dans la
chambre antérieure de composition proche de celle du plasma et le corps vitré, tissu
conjonctif gélatineux, dans la chambre postérieure (figure IV-1).
Les rayons lumineux sont diaphragmés par l’iris qui règle le diamètre de la pupille
assurant ainsi la régulation et le contrôle de l’éclairement rétinien.
Les dioptres Ils assurent la réfraction des rayons lumineux et permettent la formation de l’image de
l’objet regardé. Pour que cette image soit perçue nette, elle doit se former au centre de
la surface rétinienne, sur la zone fovéale.
Dioptres sphériques Les rayons lumineux qui pénètrent dans l’oeil sont limités par la surface pupillaire. La
portion des dioptres alors rencontrée peut être considérée comme sphérique, ce qui
permet l’application de l’approximation paraxiale ; en conséquence, un point objet a pour
image, un point (figure IV-2).
Amétropies sphériques En considérant un objet à l’infini -c'est-à-dire à 5 mètres-, l’image formée par l’ensemble
des dioptres se forme dans leur plan focal image qui doit être confondu avec le plan
rétinien : c’est la situation de l’emmétropie. L’image peut aussi se former en avant ou en
arrière du plan rétinien ce qui correspond aux amétropies sphériques, respectivement la
myopie et l’hypermétropie (figure IV-3, figure IV-4).
Dioptres astigmates réguliers ou irréguliers L’ensemble des dioptres mis à contribution pour la réfraction des faisceaux lumineux
incidents, peut ne pas être sphérique. Cependant s’ils sont réguliers et que le dioptre
équivalent présente deux rayons de courbure différents dans deux directions
perpendiculaires, le système est astigmate : c’est la situation de l’astigmatisme régulier.
Tous les points et lignes objet ont une image composée de deux parties dites focales, qui
ne sont pas situées dans le même plan (figure IV-5).
2

Œil et Physiologie de la Vision - IV
L’astigmatisme est le plus souvent dû à la modification de la forme de la cornée qui reste
cependant régulière. Il est classiquement de quelques dioptries et peut être compensé
par des verres. Il est beaucoup plus rarement lié à une modification de forme du cristallin
[el-Hage, Berny, 1973].
La déformation de la cornée peut être importante jusqu'à en devenir irrégulière comme
dans le cas du kératocône qui est à l’origine d’un fort astigmatisme myopique irrégulier
(figure IV-6).
Le kératocône est une maladie cornéenne dégénérative avec atteinte bilatérale et perte de la sphéricité de la cornée [Grunauer-Kloevekorn, Duncker, 2006]. Il y a protrusion excentrique de la cornée en forme de cône qui est, le plus souvent, asymétrique entre les deux yeux [Burns et al., 2004]. Elle s’accompagne fréquemment d’une diminution de l’acuité visuelle avec sensation de brouillard ou d’éblouissement.
La déformation de la cornée peut être irrégulière après un traumatisme cornéen. Le
rétablissement de la régularité de la surface cornéenne est indispensable pour qu’une
image puisse se former ; elle se fait grâce au port d’une lentille de contact rigide (figure
IV-7).
L’iris et la pupille L'énergie lumineuse d’une stimulation L –luminance exprimée en cd/m2- est délivrée
dans une portion de l'espace large par rapport à la surface pupillaire. Environ 2% de
cette énergie est réfléchie par la cornée, le reste est transmis à travers la pupille pour
l'éclairement rétinien.
La taille de la pupille limite le flux lumineux entrant La limitation du faisceau lumineux entrant permet l'application des approximations
paraxiales pour les calculs optiques (figure IV-2) et une meilleure mise au point de
l’image sur le plan rétinien par augmentation de la profondeur de champ due à la
diminution des aberrations géométriques.
La taille de la pupille contrôle l’éclairement rétinien L’éclairement rétinien dépend de la luminance incidente de la stimulation L, du facteur de
transmittance des milieux antérieurs (voir ci-dessous) et de la surface pupillaire S.
Il est défini par la formule empirique suivante : E rét= 0,36 d S L
est sans dimension, S : en cm2, L : en cd/m2. L'éclairement rétinien s'exprime alors en Lux, ce qui sous-
entend que la surface rétinienne éclairée est normale à la direction de la stimulation incidente et évaluée en m2 (!).
La valeur de la transmittance des milieux antérieurs étant difficile à préciser à cause de
ses variations en fonction de la longueur d’onde, on utilise pour l’éclairement rétinien,
une formule simplifiée : E rét= S L, exprimé en troland.
Un troland correspond à un éclairement rétinien de 1 cd/m2 reçu sur toute la surface rétinienne, à travers une surface pupillaire de 1 mm2
Quelle que soit l'expression utilisée pour l'éclairement rétinien, il varie avec la surface
pupillaire ce qui est fréquent.
La taille de la pupille varie entre deux sujets de même âge Pour des sujets de même âge, placés depuis un certain temps dans une ambiance
lumineuse donnée, le diamètre pupillaire peut varier entre 3 et 8 mm. Les surfaces
pupillaires correspondantes varient de 7 mm2 à 50 mm2, ce qui correspond à une
variation d'éclairement rétinien de 1 à 7.
3

Œil et Physiologie de la Vision - IV
Cette variation interindividuelle d’éclairement rétinien est une des causes des variations
interindividuelles observées pour les amplitudes des réponses électrorétinographiques
(ERG flash) par exemple.
La taille de la pupille varie en fonction de l’âge Pour une ambiance lumineuse donnée, le diamètre pupillaire varie en fonction de l'âge du
sujet.
Celui du nouveau-né peut être de petite taille (Ø d’environ 1 mm) [Abramov, Hainline,
1991] ; il est en moyenne de 4 mm de diamètre [Vaughan, Asbury, 1986], il augmente
progressivement avec l'âge, passe par une valeur maximale à l'adolescence puis diminue
avec l’âge (figure IV-8).
La taille de la pupille varie en fonction des caractéristiques des
stimulations Pour des stimulations structurées, même si leur niveau lumineux moyen est constant, la
modification périodique de la répartition spatiale de leur luminance entraîne des
modifications de tailles pupillaires comme par exemple pour les damiers onset-offset
L'apparition de mires de différentes fréquences spatiales, durant 200 ms -phase onset- suivie de leur disparition -phase offset- avec leur substitution par un champ lumineux homogène de même niveau lumineux moyen, entraîne une diminution des diamètres pupillaires, avec des temps de latence allant de 300 à 500 ms [Barbur et al., 1989] ; ceci revient à dire que, dans cette configuration, si les pupilles sont de tailles naturelles, elles fluctuent tout au cours de la stimulation.
La taille de la pupille varie en fonction de l’ambiance lumineuse Le diamètre pupillaire met plusieurs secondes avant d’atteindre sa taille d’équilibre lors
d’un changement d’ambiance lumineuse.
° Variation d’ambiance de longue durée. Pour un passage de la lumière –ambiance
photopique- à une obscurité de longue durée, la taille pupillaire croît progressivement
pour n’atteindre un diamètre stable qu’au bout de 2 minutes environ (figure IV-9).
Réciproquement, lors du passage de l’obscurité à une ambiance photopique de longue
durée, après un bref temps de latence, on observe une contraction pupillaire puis une
relaxation avec de petites oscillations régulières dites « hippus » qui apparaissent à la
fréquence de 2 Hz [Loewenfeld, 1999]. Au bout de 1 à 2 minutes, le diamètre final
moyen est inférieur au diamètre initial comme attendu.
° Variation pas à pas du niveau lumineux de l’ambiance. Certaines procédures
d’enregistrement de l’ERG flash par exemple -qui sortent du protocole clinique classique-
passent de l’obscurité où le sujet a séjourné environ 30 mn, à la lumière, avec une
augmentation pas à pas, pour atteindre des valeurs photopiques élevées. En moyenne, le
diamètre pupillaire diminue progressivement de 8 mm à 2,5 mm avec des valeurs
extrêmes allant de 9 à 1 mm, modifiant l’éclairement rétinien.
Non seulement les cellules rétiniennes qui répondent à une stimulation flash par exemple, varient au cours des modifications d’ambiances lumineuses, mais également le diamètre pupillaire du sujet. Dans ces cas spécifiques, les pupilles du sujet sont dilatées par mydriatique pour garantir des pupilles de taille stable au
cours des enregistrements.
° Incrément, décrément lumineux durant environ 200 ms. C’est sensiblement le cas pour
l’enregistrement de l’ERG ON-OFF. S’il y a augmentation -incrément durant 200 ms- ou
diminution –décrément durant 300 ms- du niveau lumineux d’une stimulation ou d’une
ambiance donnée, il se produit d’abord une constriction pupillaire dont la latence est de
l’ordre de 150 à 200 ms pour l’incrément et de l’ordre de 500 ms pour le décrément…
4

Œil et Physiologie de la Vision - IV
° Cas des flashs. Dans une ambiance lumineuse donnée, des flashs répétés provoquent
des constrictions et dilatations successives ou oscillations rapides du diamètre pupillaire
qui se stabilise rapidement autour d’une valeur moyenne fonction du niveau lumineux
moyen du fond [Milton, Longtin, 1990].
C’est le principe des flashs délivrées par les appareils photos juste avant la prise, pour que les pupilles des sujets se contractent ; ceci évite l’aspect « d’yeux rouges » sur les photos ; les « yeux rouges » ne sont que la photo du fond d’œil de chaque sujet…
Les oscillations du diamètre pupillaire suivent la fréquence temporelle des flashs. Si elle
est supérieure à 3 Hz, les oscillations pupillaires individuelles fusionnent quasiment
instantanément, le diamètre pupillaire prend alors une valeur moyenne stable qui ne
dépend que de l’intensité du flash [Loewenfeld, 1999].
Cinétique de la dilatation pupillaire par mydriatique La dilatation pupillaire par mydriatique s’effectue avec un temps de latence de 5 à 10
minutes. Elle n’est complète et stable qu’au bout de 25 à 30 minutes après l’instillation
du topique [Loewenfeld, 1999]. Si elle est mise en oeuvre en vue d’explorations
électrophysiologiques, les enregistrements ne doivent pas débuter avant que leur action
ne soit complète, soit 30 minutes.
Rappel. La dilatation pupillaire par mydriatique s’associe à la paralysie de
l’accommodation et le diamètre pupillaire final varie d’un sujet à l’autre.
En pratique Au cours d’une exploration fonctionnelle par électrophysiologie, il peut être nécessaire
dans un premier temps, de laisser au sujet la possibilité d’accommoder, puis, dans un
deuxième temps, de procéder à une dilatation pupillaire complète.
Dans ce cas, il faut attendre au moins 25 minutes après instillation du mydriatique pour
débuter l’installation du sujet. Ce temps est relativement long ; il peut être délicat à
supporter pour des patients fatigués ou difficiles.
A notre avis A moins que le sujet ne présente des pupilles naturelles de très petites tailles -ce qui
entraînerait une diminution importante de l'éclairement rétinien lors de la stimulation- la
dilatation pupillaire systématique ne nous paraît pas nécessaire.
En respectant les précautions d’adaptation rappelées ci-dessus, une pupille de taille
naturelle peut être considérée comme un bon compromis. Dans les conditions de
protocoles standard d’ERG, elle permet que l’éclairement rétinien individuel reste
sensiblement constant et que les réponses enregistrées chez un sujet donné puissent
être comparée d’un examen à l’autre.
Cristallin, accommodation et presbytie Lorsqu’un objet se rapproche du sujet, le plan sur lequel se forme son image doit rester
confondu avec le plan rétinien. Ceci est possible grâce à une augmentation de puissance
du cristallin, c’est l’accommodation [Krag, Andreassen, 2003].
La capacité accommodative s'émousse avec l'âge ; lorsque l’objet est rapproché, il n’y a
plus coïncidence entre le plan image et le plan rétinien : c'est la presbytie [Strenk et al.,
2005], [Charman, 2008].
5

Œil et Physiologie de la Vision - IV
Au cours des explorations, il est important d'en tenir compte et d’adapter au mieux les
compensations optiques des sujets essentiellement lorsque des stimulations structurées
sont présentées de près, entre 30 et 50 cm, comme pour l’enregistrement de l’ERG
multifocal ou de l’ERG pattern ou entre 100 à 150 cm, pour l’enregistrement des PEV
damier.
En pratique… Que le sujet soit presbyte, porteur d’une amétropie sphérique ou d’un astigmatisme, il
est indispensable de compenser son défaut de réfraction lors de l’utilisation de
stimulations structurées pour le rendre emmétrope, c'est-à-dire pour rétablir la
coïncidence entre le plan focal image de l’ensemble de ses dioptres antérieurs et son plan
rétinien.
Les milieux transparents Les milieux antérieurs doivent être transparents, c'est-à-dire ne pas modifier la
composition de la lumière incidente issue de l’objet, pour l’image -énergie, composition
spectrale, structure…
Variations physiologiques Les milieux antérieurs absorbent naturellement les ultraviolets, davantage les courtes
longueurs d’onde que les grandes longueurs d’onde et laissent passer les infrarouges
allant de 700 à 1400 nm.
Bien que ne participant pas à la vision, les infrarouges provoquent des échauffements des milieux oculaires pouvant entraîner des opacifications précoces du cristallin pour les sujets qui y sont exposés soit régulièrement
-réverbération solaire-, soit occasionnellement pour traitement rétinien –thermothérapie- [Van den Berg, Spekreijse, 1997].
Ainsi, une stimulation de lumière blanche comme la lumière naturelle contient-elle,
après passage des milieux antérieurs et lors de son impact sur la rétine, moins de
courtes longueurs d’onde que de grandes longueurs d’onde [Polo et al., 1996] (figure IV-
10). La lumière qui arrive sur la rétine ne pourra donc jamais être une « lumière
blanche » au sens physique du terme.
Variations avec l’âge La transparence des milieux antérieurs varie avec l’âge et de façon significative après 50
ans en relation avec les remaniements du cristallin [Said, Weale, 1959], [Sample et al.,
1988], [Polo, et al., 1996]. L’épaisseur du cristallin croît [Cook et al., 1994], [Richdale et
al., 2008], sa charge en pigment augmente ce qui entraîne une augmentation de sa
densité optique [Said, Weale, 1959], [Savage et al., 2001].
Il en résulte une absorption plus importante de l’énergie lumineuse globale mais aussi,
plus particulièrement, des courtes longueurs d'onde.
Pour voir « comme avant », le sujet a besoin de davantage de lumière ; les couleurs
perdent une partie de leur composante « bleue » allant jusqu’à donner une sensation
orangeâtre comme l’a bien décrit et montré dans ses tableaux le célèbre peintre Claude
Monet (figure IV-11).
Après intervention de sa cataracte à droite, Claude Monet confie ses impressions au Dr Jacques Mawas ophtalmologiste : « Je vois bleu, je ne vois plus le rouge, je ne vois plus le jaune ; ça m'embête terriblement parce que je sais que ces couleurs existent; parce que je sais que sur ma palette il y a du rouge, du jaune, il y a un vert spécial, il y a un certain violet ; je ne les vois plus comme je les voyais dans le temps, et pourtant je me rappelle très bien les couleurs que ça donnait »… (in http://www.snof.org/art/monet.html).
L’extraction du cristallin de Claude Monet a supprimé le filtre pathologique qui ne laissait passer que les grandes longueurs d’onde (orange et rouge) rendant alors de nouveau possible la perception des courtes longueurs d’onde c'est-à-dire des bleus…
6

Œil et Physiologie de la Vision - IV
Variations pathologiques Elles se situent classiquement au niveau de la cornée comme au cours de taies de cornée
[Ciralsky, Colby, 2007], du cristallin avec la survenue d’inhomogénéité ou cataracte
qu’elle soit congénitale [Roche et al., 2006], traumatique [Baklouti et al., 2005], liée à la
myopie [Praveen et al., 2008] ou à associée à une pathologie rétinienne comme au cours
de la rétinopathie pigmentaire [Hamel, 2006], des troubles du corps vitré, qu’il s’agisse
de décollement du vitré et de sa densification ou de rares hémorragies.
Ces troubles des milieux transparents vont contribuer non seulement à l’absorption d’une
partie de l’énergie lumineuse incidente mais aussi à l’absorption sélective d’une partie
des longueurs d’onde émises (figure IV-10).
Dans ce cas, la stimulation lumineuse émise et celle transmise à la rétine sont de
caractéristiques différentes. En particulier, lorsque la stimulation est d’un niveau
lumineux faible, l’énergie qui arrive effectivement sur la rétine peut être très faible par
absorption par les milieux antérieurs. La réponse électrophysiologique enregistrée peut
alors ne pas être discernable. Dans ce cas, avant d’attribuer cette absence de réponse à
une déficience rétinienne, il faut envisager que la stimulation rétinienne a pu être
d’énergie insuffisante et donc inefficace.
De même, chez un sujet âgé présentant une cataracte, une stimulation achromatique a,
au niveau rétinien, non seulement une énergie inférieure à celle arrivant sur la rétine
d’un sujet plus jeune, mais aussi une composition spectrale différente puisque les courtes
longueurs d’onde sont absorbées en grande partie.
Tout se passe alors comme si la stimulation était essentiellement composée de moyennes
et grandes longueurs d’onde ; leur efficacité lumineuse est faible pour les bâtonnets tout
en restant correcte pour les cônes (figure I-2-8).
En pratique… Tout trouble de transparence des milieux antérieurs doit être considéré avec attention. Il
faut ajuster les caractéristiques de la stimulation aux modifications de milieux
transparents. S’il s’agit de flashs, il convient d’en augmenter l’énergie lumineuse
incidente pour qu’ils conservent leur efficacité. Si ce n’est pas possible compte tenu des
contraintes des appareillages, il faut penser que ces stimulations peuvent générer des
réponses d’amplitudes moindres.
S’il s’agit de stimulations structurées, leur image peut être totalement déstructurée à
l’origine d’une stimulation inefficace qui peut ne pas générer de réponse.
Ces situations fonctionnelles doivent être bien analysées avant de conclure que les
réponses atypiques ou l’absence de réponse résultent d’un dysfonctionnement organique
de la rétine ou des voies visuelles.
Le pôle postérieur
Il est le siège de la rétine, tunique la plus interne des membranes de l’œil ; c’est un tissu
vasculo-nerveux inhomogène. On décrit classiquement, d’une part, sa composante
neurologique : la rétine neurosensorielle ou neurorétine et l'épithélium pigmentaire avec
lequel elle est en contiguïté et, d’autre part, ses apports vasculaires rétiniens et
choroïdiens.
7

Œil et Physiologie de la Vision - IV
La neurorétine
Surface totale La surface totale de la neurorétine est proche de 1200 mm2. Son épaisseur est variable,
minimale à l'ora serrata et à la fovéola : 0,1 mm, maximale au voisinage de la papille :
0,5 mm et, en moyenne, de l'ordre de 0,23 mm au pôle postérieur.
L’utilisation courante de l’OCT (Optical Coherence Tomography) (www.oct-ophtalmo.fr/) permet une évaluation précise de l’épaisseur de la rétine [Kim et al., 2009] qui est utile dans le diagnostic précoce de son altération, par exemple, les oedèmes maculaires [Lang, 2007], le glaucome [Schuman, 2008]…
Zone centrale ou maculaire et la fovéola La zone centrale du pôle postérieur ou macula est une ellipse à grand axe horizontal de 2
à 3 mm, vue sous un angle de 6 à 10 degrés environ. Sa surface est de l’ordre de 28
mm2 ce qui correspond à 2,5 % environ de la surface rétinienne totale.
En son centre se trouve une dépression, la fovéola, vue sous un angle de 1° 20' (figure
IV-12) ; son épaisseur est d’environ 0,1mm. Elle est avasculaire (figure IV-13) et ne
contient que des cônes alors que partout ailleurs, on trouve des cônes intercalés entre les
bâtonnets.
C’est la zone où se focalise l’image de l’objet fixé. Elle assure la vision précise, quantifiée
sous forme d’acuité visuelle.
Eclairée par une lumière anérythre, c'est-à-dire ne contenant pas de grandes longueurs
d’onde (rouge), elle apparaît sous la forme d'une ellipse colorée dite tache jaune, liée à la
présence des pigments maculaires.
Pigments maculaires ° Situation. Ils sont situés au niveau des segments internes des photorécepteurs et en
quantité plus importante, dans la strate des fibres de Henlé (zone synaptique entre les
photorécepteurs et les cellules bipolaires) [Snodderly et al., 1984b], [Delori et al., 2001],
[van der Veen et al., 2009]. Ils s’interposent donc entre la lumière et les photopigments
des photorécepteurs qui sont contenus dans leurs articles externes [Snodderly et al.,
1984a] (figure IV-14).
° Composition. Ils sont composés de deux pigments, la lutéïne et la zéaxanthine, cette
dernière étant deux fois plus abondante que la première [Bone et al., 1988].
° Densité. La densité des pigments maculaires est maximale au niveau de la fovéola et
de la couche plexiforme interne [Snodderly, et al., 1984a] ; elle varie de 0,41 à 0,81
[van de Kraats et al., 1996], [Snodderly, et al., 1984a], avec une moyenne de 0,33
[Bone, et al., 1988]. L’évaluation de la densité des pigments maculaires in vivo est
possible par différentes méthodes (photométrie hétérochromatique à papillotement,
mesure de l’autofluorescence et de la réflectance du fond d’œil, spectroscopie Raman…)
[Delori, et al., 2001], [Gellermann, Bernstein, 2004], [Rougier et al., 2008].
Les caractéristiques des pigments maculaires semblent stables avec l’âge [Bone, et al.,
1988], [Delori, et al., 2001] ou subissent des modifications mineures [Hammond,
Caruso-Avery, 2000].
° Absorption. Les pigments maculaires absorbent essentiellement les courtes longueurs
d'onde comprises entre 400 et 550 nm, avec un pic d'absorption maximale vers 455 nm
[Sharpe et al., 1998] (figure IV-15).
8

Œil et Physiologie de la Vision - IV
° Rôle. Les pigments maculaires évitent les effets nocifs des courtes longueurs d'onde sur
la rétine [Werner et al., 2000]. Ils sont anti-oxydants et préviennent la dégénérescence
maculaire liée à l’âge (DMLA) [Seddon et al., 1994], [Bone et al., 2000], [Davies,
Morland, 2004], [Whitehead et al., 2006], [Nolan et al., 2007].
Plus accessoirement, les pigments maculaires améliorent la qualité de l’image fovéale par
l’atténuation de l’aberration chromatique [Le Grand, 1972], [Wooten, Hammond, 2002],
[Davies, Morland, 2004] ou par atténuation de la diffraction atmosphérique [Wooten,
Hammond, 2002].
Subdivision fonctionnelle de la macula La macula est subdivisée en trois zones fonctionnelles différentes, la fovéola vu
schématiquement sous un angle de 2° et deux anneaux qui lui sont concentriques : un
1er anneau vu sous un angle compris entre 2° et 4° environ et un 2ième anneau vu sous
un angle compris entre 4° et 10° environ.
Cette subdivision fonctionnelle est liée à la possibilité d’utiliser des stimulations
structurées organisées en damier avec des cases de tailles différentes qui testent
séparément la fovéola et ces deux anneaux maculaires (figure IV-16), leurs réponses
respectives sont recueillies séparément sous forme de potentiels évoqués visuels : PEV
damier (voir ci-dessous)
Toujours d’un point de vu fonctionnel, la rétine située en dehors des 10 degrés centraux
est considérée comme une zone périphérique.
Zone périphérique fonctionnelle ° Image de l’espace visuel. La surface focale image, donnée par l'ensemble des dioptres
de l'œil, devrait coïncider avec la surface de la neurorétine, assimilée à une portion de
sphère, pour que l'image de tout l’espace visuel soit perçue nettement.
Or, dès que l'on considère l’image rétinienne projetée en dehors des dix degrés centraux,
elle est entachée d’aberrations -géométriques, chromatiques…- et perçue floue.
° Stratégie visuelle. Cette zone extramaculaire a cependant un rôle important dans la
vision. Toute image qui s’y projette -pourtant perçue floue- retient l’attention alors
qu’elle n’est pas regardée. Elle déclenche la mobilisation des yeux et/ou de la tête [Land,
2006] pour la ramener dans la zone centrale de vision précise.
Ainsi toute altération fonctionnelle de la rétine périphérique peut-elle être pénalisante
pour la qualité de la vision, même si la rétine centrale est normalement fonctionnelle et
que le sujet présente une acuité visuelle normale.
° Exploration fonctionnelle globale. Le fonctionnement de l’ensemble de la neurorétine
périphérique est possible grâce au recueil de l’ERG flash ; il représente une réponse
globale ne permettant pas de distinguer les zones saines de possibles zones déficientes.
Pour ce faire, il faut stimuler séparément les différents secteurs du pôle postérieur.
° Exploration fonctionnelle en secteurs. Les différents secteurs du pôle postérieur
peuvent être testés à l’aide de stimulations en hexagones réparties sur les 40 à 50
degrés centraux (figure V-4-1), l’ensemble de la réponse de chaque secteur étant
présenté sous forme d’un ERG multifocal.
9

Œil et Physiologie de la Vision - IV
Ce test permet la mise en évidence des zones localisées fonctionnellement déficientes
(figure V-4-3).
Figure IV-17. Exemple de S. 27 ans, sujet atteint de rétinopathie pigmentaire avec une acuité visuelle de 10/10ième à chaque oeil. Le relevé du champ visuel Goldmann montre une sensibilité normale de la zone centrale, associée à un déficit de sensibilité des zones périphériques.
Figure IV-18. Sujet S : Correspondance entre l’aspect du fond d’œil droit et la sensibilité relevée au champ visuel Goldmann. Seule la zone centrale et un ilôt périphérique gardent une sensibilité normale à la stimulation lumineuse. Toute la zone périphérique sur laquelle des pigments sont visibles, présente un déficit majeur de sensibilité à la lumière.
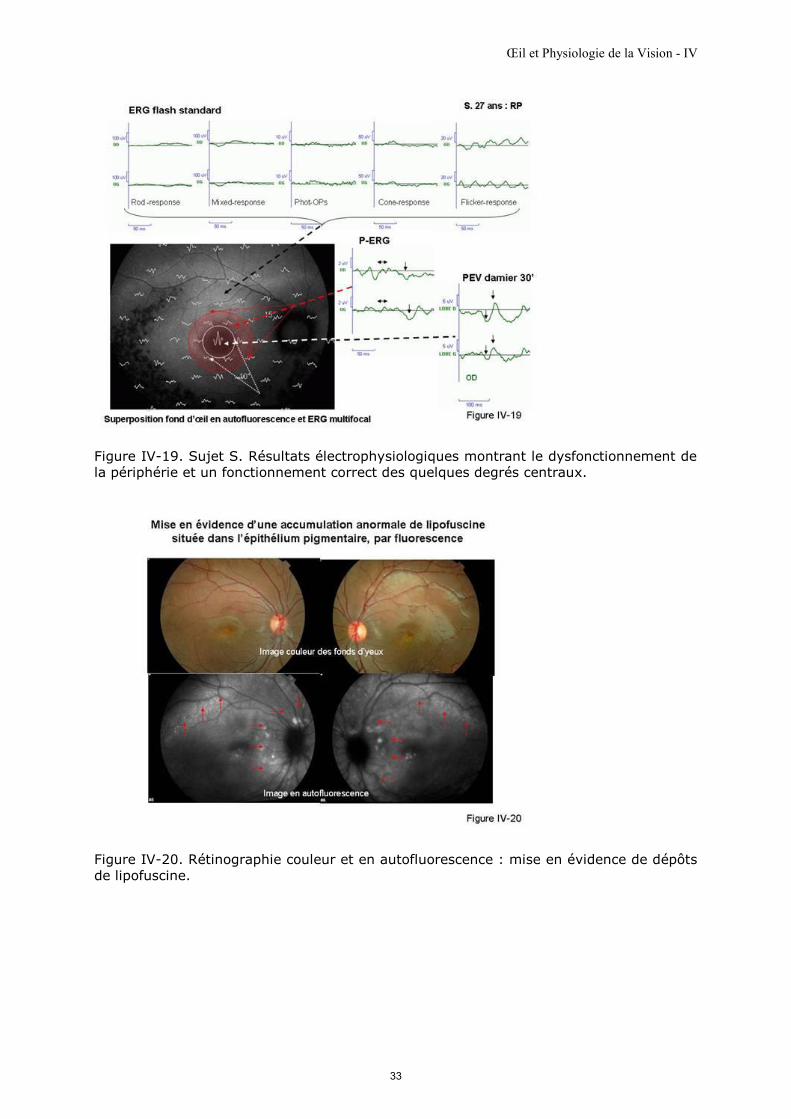
Figure IV-19. Résultat de l’ensemble du bilan électrophysiologique. Il met en évidence un dysfonctionnement rétinien global important : l’ERG flash protocole standard présente des réponses discernables, mais d’amplitudes très diminuées. Le dysfonctionnement du pôle postérieur est mis en évidence par l’ERG multifocal superposé ici, à la photo du fond d’œil prise en autofluorescence ; ses amplitudes sont très diminuées sauf celles correspondant aux quelques degrés centraux. Le pic fovéolaire est d’amplitude pratiquement normale attestant que les quelques degrés centraux sont fonctionnels. Les PERG sont d’amplitude diminuée montrant que les quinze degrés centraux (zone maculaire) dysfonctionnent ; PEV damier 30’ sont discernables et sensiblement normaux ; ils confirment que le fonctionnement des 4 degrés centraux est sensiblement normal.
Zone périphérique du clinicien La zone périphérique fonctionnelle décrite ci-dessus est plus restreinte que la périphérie
des cliniciens qui s’étend au-delà des arcs vasculaires avec une moyenne périphérie puis
une extrême périphérie [Behar-Cohen F. et al., 2009].
Sens de propagation de la lumière La majorité des photons lumineux doit traverser toute la rétine pour atteindre les
photorécepteurs situés à l'étage le plus externe, sauf au niveau de la fovéola où les
articles externes des photorécepteurs peuvent être atteints pratiquement directement.
Absorption des différents photons par la rétine Les différentes couches de la rétine absorbent de façon variable les rayons lumineux de
différentes longueurs d’onde. Les proches infrarouges transmis par les milieux antérieurs
pénètrent jusqu’à la couche de l’épithélium pigmentaire.
Seules les longueurs d’onde comprises entre 400 et 700 nm sont absorbées par les
photorécepteurs avec une probabilité d’absorption dépendant de son type (figure I-2-8).
Elles sont à l'origine de la sensation lumineuse.
L’épithélium pigmentaire et son autofluorescence La neurorétine est en rapport étroit avec l'épithélium pigmentaire.
Si la lipofuscine est contenue en proportion anormalement importante dans l'épithélium
pigmentaire, elle peut être visualisée grâce à ses propriétés de fluorescence.
Une lumière de moyennes longueurs d’onde (comprises entre 470 et 500 nm), délivrée
sur un fond d’œil, excite la fluorescence de la lipofuscine ; le retour à l’état de base se
fait avec une émission de photons fluorescents visibles, de longueurs d’onde comprises
entre 550 et 600 nm, permettant ainsi la localisation de son accumulation par
« autofluorescence » [Delori et al., 1995] (figure IV-20).
Systématisation neuronique et vasculaire Les altérations de la rétine ont des conséquences différentes selon qu’elles sont d’origine
neuronique ou vasculaire. Ces altérations s’organisent selon une systématisation
différente qui répond à leur anatomie.
10

Œil et Physiologie de la Vision - IV
Répartition neuronique Les axones amyéliniques des cellules ganglionnaires rétiniennes courent à la surface de
la neurorétine après avoir fait synapse avec les cellules bipolaires et, en amont, les
photorécepteurs. On peut les mettre en évidence en éclairant le fond d’œil par une
lumière bleue (figure IV-21).
Ces axones non myélinisés forment une succession d’arcs ovalaires avec regroupement
systématisé des fibres à la papille (figure IV-21 et figure IV-22).
Les fibres maculaires occupent le centre supérieur et inférieur du secteur temporal, celles issues de la rétine temporale supérieure et inférieure se regroupent respectivement sur son secteur temporal supérieur et inférieur ; celles provenant de la rétine nasale supérieure et inférieure occupent respectivement son secteur nasal supérieur et inférieur.
Les fibres provenant des secteurs rétiniens les plus centraux sont les plus proches du centre de la papille, tandis que celles provenant des secteurs rétiniens les plus périphériques se situent davantage à la périphérie papillaire.
On peut observer des déficits fasciculaires systématisés comme ici dans le cas d’un
glaucome (figure IV-23).
Répartition vasculaire La vascularisation rétinienne est double, indirecte assurée par le réseau choroïdien et
directe due au réseau capillaire terminal d'artères intrarétiniennes [Ducasse, Segal,
1985], [Behar-Cohen F et al., 2009].
° Réseau choroïdien. Il est composé de capillaires en maille (figure IV-24), issus des
artères ciliaires ; ils assurent la vascularisation indirecte de toute la surface du premier
étage rétinien (couche des photorécepteurs), y compris celle de la zone fovéolaire, à
travers le tapis monocellulaire de l'épithélium pigmentaire.
° Réseau rétinien. Il est issu de l'artère centrale de la rétine et se distribue en un plan
profond pour la vascularisation du deuxième étage rétinien (couche des cellules
bipolaires) et en un plan superficiel, pour la vascularisation du troisième étage rétinien,
au sein de la couche des cellules ganglionnaires.
La zone centrale incluant la fovéola, vue sous un angle d’environ 2°, n'est pas
vascularisée par ces branches terminales de l'artère centrale de la rétine [Provis,
Hendrickson, 2008] (figure IV-13).
Il existe, de façon inconstante des artères rétiniennes d'origine ciliaire, les artères ciliorétiniennes qui vascularisent la région fovéolaire. Elles sont à l'origine de l'épargne maculaire en cas d'oblitération du tronc de l'artère centrale de la rétine.
° Réseau veineux. Il se draine principalement vers la veine centrale de la rétine située au niveau de la papille, pour gagner ensuite le sinus caverneux, soit isolément, soit en se jetant dans la veine ophtalmique supérieure ou moyenne.
Dysfonctionnement rétinien d’origine vasculaire
Occlusions des artères et veines centrales : lésion de toute la rétine On peut considérer que les lésions vasculaires des artères et veines centrales de la rétine
touchent la totalité du tissu rétinien et en particulier celui du pôle postérieur…/…
Occlusions de branches : lésions altudinales …alors que les occlusions de branches vont entraîner, selon leur localisation supérieure
ou inférieure, des altérations du champ visuel ayant une dominante altitudinale.
11

Œil et Physiologie de la Vision - IV
On peut aussi remarquer que l’organisation vasculaire temporale et nasale a pour plan de
clivage la papille (figure IV-25).
Occlusions de branches temporales : lésion maculaire Le pôle postérieur de la rétine est vascularisé par les branches temporales de l’artère et
de la veine centrale de la rétine. Toute altération de l’une de ces branches aura donc des
répercutions sur la fonction maculaire et en particulier sur les examens
électrophysiologiques testant le fonctionnement maculaire (ERG multifocal, P-ERG, PEV).
Occlusions de branches nasales : lésions périphériques Les altérations vasculaires nasales ne touchent que la rétine périphérique et n’ont donc
pas nécessairement de conséquence sur l’acuité visuelle.
C’est la raison pour laquelle elles sont souvent détectées très tardivement. Ces lésions ne
sont pas ou peu mises en évidence par des examens électrophysiologiques.
Réseau microvasculaire Le réseau microvasculaire centré sur la fovéola est une région rétinienne particulière
puisqu’elle correspond à la zone procurant les meilleures acuités visuelles et la meilleure
sensibilité neurosensorielle.
Son dysfonctionnement va entraîner des signes cliniques et fonctionnels importants. En
effet, autour de la région avasculaire centrale s’organise un riche réseau de mailles
capillaires formant une ligne en bordure de la zone avasculaire centrale appelée maille
capillaire (figure IV-13).
° Altération de la maille capillaire. Les altérations de cette maille capillaire font partie des
signes des maladies touchant la microvascularisation comme le diabète. Les occlusions
de cette structure entraînent souvent l’apparition d’oedèmes ou d’épaississement du tissu
rétinien qui s’accompagne de modifications fonctionnelles de la neurorétine avec un
retentissement important sur la genèse des signaux électrophysiologiques qui testent le
fonctionnement maculaire.
° Altération périfovéolaire. La zone périfovéolaire concentrant les cellules de la rétine
interne de la région centrofovéolaire et les limites de la maille capillaire forment une
surface annulaire souvent incriminée dans les signes primitifs de nombreuses pathologies
du pôle postérieur.
Cette zone est fondamentale pour l’examen électrophysiologique, car elle est souvent
impliquée dans les atteintes de type hérédo-dégénératif ou celles dues à certains
toxiques comme la chloroquine ou l’hydroxychloroquine.
Dysfonctionnement rétinien d’origine neuronique La segmentation neurologique basée sur l’organisation du nerf optique présente une
double symétrie droite-gauche et haut-bas, centrée sur la fovéola (figure IV-26).
Altération des voies de conduction Les altérations mettant en évidence cette organisation n’ont généralement pas pour
origine la rétine, mais les voies de la conduction. Ceci est lié au fait que la
systématisation latérale et altitudinale est conservée le long des voies visuelles.
Les altérations rétiniennes liées au système neurologique sont en rapport avec un
dysfonctionnement des axones issus des cellules ganglionnaires. Il peut s’agir, par
exemple, d’un déficit du transport axoplasmique qu’il soit rétrograde ou antérograde.
12

Œil et Physiologie de la Vision - IV
Ce type de dysfonctionnement entraîne, s’il est chronique, une perte de certains
faisceaux de fibre du nerf optique, visible à l’imagerie du fond d’œil (figure IV-23).
Si l’altération est rapide, on note souvent l’apparition de dépôts nodulaires blancs qui
traduisent un dysfonctionnement aigu du transport axoplasmique.
Retentissement sur la rétine adjacente Cette altération de la rétine interne a des conséquences sur le fonctionnement de la
rétine adjacente, ce qui entraîne des déficits de type arciforme ayant pour origine la
papille et se développant dans la rétine temporale selon une surface rétinienne appelée
aire de Bjerum.
Retentissement sur les examens électrophysiologiques Les altérations d’origine neurologique ont, en général, des conséquences sur les
potentiels évoqués visuels plus que sur l’électrorétinogramme.
Il existe une exception à cette règle qui correspond à une atteinte mixte vasculaire et
rétinienne d’origine papillaire. Dans ce cas, on note à la fois une altération des voies de
la conduction et de la rétine interne. Cette altération est globale et touche tous les types
cellulaires.
La systématisation anatomique des lésions a des conséquences fonctionnelles avec
retentissements en partie prévisibles, sur les examens électrophysiologiques.
C’est pourquoi la confrontation entre l’imagerie de fond d’œil par rétinographie,
angiographie et OCT est une nécessité, si l’on désire tirer le maximum d’informations
d’une exploration fonctionnelle par électrophysiologie.
Papille optique
Description Elle se présente sous forme d’un disque ovalaire vu sous un angle de 10° environ. Son
centre est situé à 15° en nasal et 2° en inférieur par rapport à la fovéola (figure IV-27).
Elle est aussi le lieu d'émergence de la vascularisation rétinienne à travers la lame
criblée, structure relativement rigide. Sa coloration normale est beige rosée.
Systématisation La papille est le lieu de regroupement des axones des cellules ganglionnaires qui sont
amyéliniques jusqu’à la lame criblée. Ils formeront les fibres du nerf optique, la papille en
constituant la tête avec une répartition systématisée (figure IV-22).
Vascularisation La papille possède une vascularisation propre, indépendante des artères et veines
centrales de la rétine qui la traversent. Cette vascularisation est assurée principalement
par des capillaires provenant des artères ciliaires courtes postérieures.
Ces capillaires sont particulièrement fragiles, sensibles notamment à l'équilibre entre pression intraoculaire et pression artérielle. Ceci explique la fréquence des pathologies ischémiques de la tête du nerf optique, secondaires à des maladies vasculaires ou au glaucome [Flammer, Orgul, 1998].
Absence de tissu sensoriel Dépourvue de tissu sensoriel, la papille est à l’origine d’un scotome (absence de vision)
bien mis en évidence sur les relevés des champs visuels (figure IV-17).
13

Œil et Physiologie de la Vision - IV
Cette absence de tissu sensoriel est de surface trop faible par rapport au reste de la
surface rétinienne pour modifier les résultats électrorétinographiques, qu’ils soient
générés par flash (ERG flash) ou par hexagones (ERG multifocal).
Rôle fonctionnel majeur : passage obligé de l’influx visuel La papille est le passage obligé de l’influx visuel, codé par les corps des cellules
ganglionnaires après variations de polarisation des photorécepteurs puis des cellules
bipolaires. Il se propage le long de l’ensemble des voies visuelles avant d’atteindre les
centres visuels.
Toute atteinte de la papille peut modifier le mode de conduction de l’influx visuel et être
à l’origine d’anomalies des signaux électrophysiologiques recueillis au niveau des aires
occipitales sous forme de potentiels évoqués visuels corticaux (PEV).
Exemples ° Oedèmes papillaires. Leurs origines sont multiples. Si l’influx visuel est généré
normalement au niveau des neurorétines, y compris par les corps des cellules
ganglionnaires, il peut être désorganisé par la présence d’un œdème papillaire.
Les influx unitaires (ou potentiels d’action) émis par les différents groupes d’axones peuvent ne plus être synchrones à cause de l’œdème. Les différences de potentiels générées au niveau cortical ne sont alors plus synchrones et les réponses visuelles ne sont pas discernables du bruit de fond (PEV V-5). Dans ce cas, l’absence de PEV discernables ne permet pas de trancher entre un œdème de stase et une papillite.
Cependant, un œdème peut être cliniquement important et ne pas avoir (encore) de retentissement sur la synchronisation des influx unitaires ; les PEV peuvent être normaux. Ce résultat oriente vers un oedème papillaire de stase [Sureda et al., 1991]. Un bilan neuro-ophtalmologique complet est indispensable à la recherche d'une hypertension intracranienne avec IRM et, si l'IRM est normale, une ponction lombaire avec mesure de la pression du liquide céphalo-rachidien à l'ouverture, à la recherche d’une hypertension intracranienne bénigne [Biousse, Newman, 2009].
Exemple de K. 20 ans (figure IV-28) ; elle présente au fond d’œil un œdème de stase découvert à la suite de maux de tête persistants, sans autre signe par ailleurs. Son bilan électrophysiologique est normal (ERG flash normaux, P-ERG normaux, PEV damier normaux) en faveur d’un fonctionnement normal des voies de conduction et donc d’un œdème de stase.
° Pâleurs papillaires globales. Elles sont, le plus souvent, signe d’une atrophie avancée de
l’ensemble des fibres optiques. Selon leur ancienneté, les influx générés au niveau
rétinien peuvent être soit ralentis, à l’origine de PEV d’amplitudes diminuées et de temps
de culmination augmentés, soit ne plus être transmis aboutissant à des PEV non
discernables.
° Pâleurs papillaires en secteur. C’est un cas intéressant car les résultats des PEV sont en
partie expliqués par ces atteintes systématisées.
Si on observe une pâleur du secteur temporal correspondant à la perte des fibres
axonales maculaires comme au cours de l’atrophie optique autosomique dominante, il y a
une atteinte prépondérante des macula. Dans ce cas, les PEV sont difficilement
discernables voire non discernables [Votruba et al., 2003] puisqu’ils sont majoritairement
le résultat de la réponse générée au niveau des zones maculaires.
A contrario, par exemple au cours d’un processus d’ischémie restreinte de la tête du nerf
optique, il est possible d’observer une atteinte localisée de la tête du nerf optique en
zone nasale ou temporale, supérieure ou inférieure avec conservation du faisceau
maculaire. Dans ce cas, particulier certes, l’acuité visuelle est le plus souvent conservée
avec des PEV sensiblement normaux.
Ces exemples montrent qu’il y a, souvent, une bonne corrélation anatomo-fonctionnelle.
14

Œil et Physiologie de la Vision - IV
Les voies visuelles de conduction
Zone rétrobulbaire : les nerfs optiques
Myélinisation des axones Après passage de la lame criblée, les axones des cellules ganglionnaires qui forment le
nerf optique se myélinisent. Chaque nerf optique regroupe de 700 000 à 1 700 000 fibres
selon les auteurs [Vital-Durand, 1986].
90% environ des fibres sont de petits calibres. 80% sont les axones des cellules ganglionnaires naines et bistratifiées -voie P et voie K- à destinée géniculo-corticale et 10% sont les axones des cellules ganglionnaires de la voie rétino-tectale ou colliculaire, de la voie rétino-prétectale, du système optique accessoire et de la voie rétino-hypthalamique [Tovée, 1996] ; les 10% restant sont de fibres de gros calibres, issues des cellules ganglionnaires parasols qui forment la voie M à destinées géniculo-corticale (figure IV-29).
La conduction de l’influx nerveux devient de type saltatoire, d’un nœud de Ranvier à
l’autre, permettant une augmentation de la vitesse de propagation jusqu’aux aires
visuelles.
Pathologie de la myéline Dans le cadre de pathologie de la myéline -sclérose en plaque, leucodystrophie...- c’est à
partir de ce point qu’il y a désynchronisation possible des influx unitaires, pouvant être à
l’origine de réponses évoquées visuelles atypiques (PEV de morphologies anormales
et/ou de temps de culmination augmentés…).
Trajet sinueux Les deux nerfs optiques se dirigent vers les trous optiques dans un cône graisseux selon
un trajet sinueux de 25 à 30 mm ce qui évite leur étirement lors de la mobilisation
normale des globes.
Vascularisation intra-orbitaire du nerf optique Elle est divisée en deux parties par le point de pénétration de l'artère centrale de la
rétine, à 10 mm environ en arrière du globe (figure IV-30).
En avant de ce point, la vascularisation est axiale et périphérique, essentiellement centripète, assurant à cette portion du nerf optique, une riche vascularisation. En arrière de ce point, la vascularisation est principalement périphérique et centripète.
La papille et la portion postérieure du nerf optique sont ainsi moins richement
vascularisées que la portion antérieure ; cette configuration anatomique les rend plus
sensibles aux processus ischémiques qui pourront alors modifier la conduction des influx
nerveux.
Le canal optique : un passage périlleux
Les nerfs optiques traversent la boîte crânienne par les trous optiques dont la portion
intracanalaire est d’environ 5-7 mm.
Ce passage est un goulet rigide, étroit, siège possible de compressions temporaires ou
durables comme lors d’inflammations rétrobulbaires ou d’étirements anormaux,
fréquents lors de traumatismes crâniens frontaux ou latéraux qui se traduisent par une
douleur à la mobilisation des globes.
Ces compressions peuvent s’accompagner ou non d’une baisse de l’acuité visuelle et se
manifester par un ralentissement ou une désorganisation de l’influx nerveux qui entraîne
des modifications significatives des caractéristiques des potentiels évoqués visuels (PEV).
15

Œil et Physiologie de la Vision - IV
Si la compression dure, l’acuité visuelle baisse, l’influx nerveux peut être pratiquement
interrompu, les PEV ne sont plus discernables alors qu’anatomiquement le nerf optique
peut paraître normal à l’IRM.
Si la compression est levée rapidement, l’influx nerveux peut se restaurer avec
récupération de l’acuité visuelle ; dans le cas contraire, malgré l’intégrité anatomique, il y
a interruption de la transmission, c’est la section dite physiologique du nerf optique.
Exemple d’un patient J. âgé de 46 ans. Il a été victime d’un accident de la voie publique avec plaie de la paupière droite. Il se plaint rapidement d’une baisse importante de l’acuité visuelle à droite. L’examen clinique ophtalmologique met en évidence une pupille droite aréactive.
Figure IV-31. Le champ visuel Goldmann relevé à droite montre un scotome central englobant la tache aveugle. L’IRM des voies optiques est normale.
Figure IV-32. Le bilan électrophysiologique montre pour l’ERG à droite, une diminution d’amplitude des réponses du système scotopique et photopique, pour le P-ERG et les PEV damier à droite, une absence de réponse.
Dans le contexte de traumatisme facial et oculaire droit, avec IRM normale, ces résultats
reflètent l’absence de conduction de l’influx nerveux le long du nerf optique droit pouvant
entrer dans le cadre d’une section physiologique du nerf optique droit avec un discret
retentissement sur le fonctionnement neurorétinien droit.
Ces résultats expliquent l’absence de sensibilité de la zone centrale droite relevée au champ visuel.
Zone de décussation des voies visuelles
Chiasma Le chiasma est la zone de réunion des deux nerfs optiques sous forme d’une lame
épaisse d’où émergent, en arrière, les tractus optiques (figure IV-33, figure IV-34, figure
IV-35).
Décussation normale Chez un sujet normal, 50% environ des fibres issues de chaque œil décussent [Kupfer et
al., 1967]. Les fibres qui proviennent des zones temporales sont directes tandis que
celles issues des zones nasales croisent. Seules les fibres issues des maculas se
projettent sur les deux pôles [Victor et al., 1991].
Cette configuration permet le regroupement des informations venant de l’hémichamp nasal droit et de l’hémichamp temporal gauche dans le tractus optique droit autorisant, en vision binoculaire, la perception de tout l’hémichamp visuel.
La figure IV-34 est une représentation du chiasma par tractographie. La tractographie est une reconstitution mathématique au cours d’une IRM cérébrale de la trajectoire de molécules d’eau. Leur déplacement peut coïncider avec l’ensemble des axones neuronaux et de leur gaine de myéline. Cette méthode permet une visualisation indirecte, dans l’espace et in vivo, de l’architecture des voies optiques, des corps géniculés latéraux et des radiations optiques qui, par des méthodes traditionnelles, ne sont pas discernables de la substance blanche [Nguyen et al., 2003].
Décussation anormale Les enfants albinos présentent une décussation anormale de leurs fibres temporales qui
sont en grande partie croisées au lieu d’être directes [Guillery et al., 1975]. Cette
particularité anatomique simule un syndrome chiasmatique avec atteinte des voies
directes. Il ne s’agit pas en réalité, d’une véritable atteinte des voies directes, mais d’une
diminution significative du nombre de fibres directes avec augmentation du nombre de
fibres croisées (voir : figure V-5-24 et chapitre VII-1).
16

Œil et Physiologie de la Vision - IV
Les asymétries d’amplitudes des PEV recueillis en regard de chacun des lobes, droit et
gauche, après stimulation de chacun des deux yeux, permettent de localiser les niveaux
d’atteintes des voies visuelles par rapport au chiasma (figure V-5-22, figure V-5-23,
figure V-5-24, figure V-5-25).
Zone de rapports anatomiques multiples Le chiasma est en rapport étroit avec la paroi du 3ième ventricule, l’hypophyse et le
polygone de Willis.
Toutes anomalies de proximité avec leurs manifestations générales et/ou
ophtalmologiques -hydrocéphalie, tumeur du 3ième ventricule, pathologie hypophysaire ou
du polygone de Willis- déclenchent des examens neuroradiologiques -scanner, IRM, voire
scintigraphie- et le relevé d’un champ visuel. Un bilan électrophysiologique avec
potentiels évoqués visuels n’est que rarement demandé.
En effet, par exemple pour le suivi d’une tumeur de l’hypophyse, on préfère effectuer une
IRM associée, à des rythmes variables, à des relevés de champs visuels par périmétrie
automatisée (24 ou 30 degrés), à la place d’enregistrement des PEV [Pakzaban et al.,
2000], [Baglin et al., 2009].
La majorité des fibres se dirige vers les tractus optiques.
Quelques unes quittent la face supérieure du chiasma pour former la voie rétino-thalamique qui joue un rôle dans le rythme circadien (figure IV-29).
Les tractus optiques
Ils se terminent en deux racines :
° l'une, importante est latérale ; elle véhicule toutes les fibres visuelles jusqu'aux corps
géniculés latéraux. Elle constitue la voie rétino-géniculo-corticale, avec des fibres de
petits et gros calibres : voies P ou parvo, voie K ou konio et voie M ou magno (figure IV-
35 et figure IV-36)
° l'autre, peu importante est médiane. Elle constitue les voies optiques secondaires, avec quelques fibres de gros calibres et surtout des fibres de très petits calibres qui ont plusieurs destinées : colliculus, voie optique accessoire… (figure IV-29). Elle pourrait être responsable du « blind-sight » perception inconsciente d’un objet en mouvement dans le champ visuel aveugle. Elle est responsable de la persistance d’un réflexe photomoteur en cas de cécité corticale [Stoerig, 2010], [Overgaard et al., 2008], [Stoerig, 2006].
Les corps géniculés latéraux
Aucun signal électrophysiologique clinique ne permet actuellement d’en isoler le
fonctionnement.
Zone d’amplification des fibres maculaires Les corps géniculés latéraux sont des relais synaptiques organisés en six couches (figure
IV-37), avec une amplification importante des fibres issues des zones maculaires et
d’autant plus qu’elles sont proches de la fovéola.
Zones d’afférence des voies P, M et K et de regroupement des fibres issues de l’œil droit et gauche
Les axones des cellules ganglionnaires naines de la voie P se terminent sur les quatre
couches dorsales dites parvocellulaires (3 à 6) [Ahmad, Spear, 1993] ; ceux des cellules
ganglionnaires parasols de la voie M, sur les deux couches ventrales dites
magnocellulaires (1, 2) et ceux des cellules ganglionnaires bistratifiées de la voie K, sur
deux intercouches dites koniocellulaires [Hendry, Reid, 2000] (figure IV-38).
17

Œil et Physiologie de la Vision - IV
Chaque corps géniculé latéral regroupe les fibres issues de chaque hémichamp ce qui en
permet la représentation complète.
Les radiations optiques vers les aires visuelles primaires
Les radiations optiques assurent la connexion entre les corps géniculés latéraux et les
deux aires visuelles primaires -ou cortex strié- l’une à droite et l’autre à gauche, situées
en zone occipitale.
Projections de part et d’autre de la scissure calcarine A leur terminaison, les faisceaux supérieur et inférieur se projettent sur une surface
restreinte, respectivement au-dessus et au-dessous de la scissure calcarine de l'aire
visuelle primaire en respectant l'excentricité rétinienne, les projections les plus
antérieures correspondant aux zones rétiniennes les plus périphériques (figure IV-39).
Le faisceau maculaire se termine à la partie postérieure de la scissure calcarine sur une
large surface du pôle postérieur du cortex visuel en zone occipitale (figure IV-40).
Projections de l’aire maculaire et fovéale Chez l’homme, les 30° centraux se projettent sur 83% de la surface du cortex strié, les
10° centraux ou macula, sur 50 à 60% de la surface du cortex strié réalisant une
amplification maculaire considérable [Horton, Hoyt, 1991].
Les projections fovéales -les 2° centraux environ- occupent seules au moins 25% de la
surface du cortex visuel [Dow, 1990], ce qui souligne la prépondérance de la
représentation fovéale dans l’amplification maculaire avec une large représentation au
niveau des deux pôles occipitaux.
Ce phénomène d'amplification maculaire cortical semble proportionnel à la densité des
cônes rétiniens [Dow et al., 1981] et se poursuit tout le long de la chaîne rétino-corticale
[Popovic, Sjostrand, 2001].
Mais l’amplification corticale aurait, d’après Wässle [Wassle et al., 1989], davantage pour
origine un mécanisme de niveau rétinien que de niveau géniculé ou cortical. Il est en
effet bien corrélé à la densité des cellules ganglionnaires rétiniennes qui est 1 000 à
2 000 fois plus importante au centre qu'à la périphérie rétinienne.
L'amplification maculaire n'aurait donc pas besoin, pour être expliquée, de présupposer
l'existence d'un relais amplificateur géniculé ou cortical…
Vitesse de conduction des voies P, K et M La vitesse de conduction le long de l’ensemble des voies visuelles dépend du diamètre de
leurs axones.
Les voies P et K sont respectivement de petits et de très petits calibres ; les vitesses de
conduction de leurs potentiels d’action sont respectivement lentes et très lentes. Le
diamètre des axones de la voie M est plus important, la vitesse de conduction de ses
potentiels d’action est beaucoup plus rapide.
Les aires visuelles primaires : 80 % de la projection fovéale
Les projections géniculées se font sur des couches et sous-couches distinctes avec une
large représentation des projections maculaires et surtout fovéales.
18

Œil et Physiologie de la Vision - IV
Projections géniculées essentiellement sur la couche 4C Le cortex visuel primaire est divisé en 6 couches numérotées de 1 à 6 (figure IV-41) ; la
couche 4C est le lieu privilégié des afférences géniculées (figure IV-38).
Très schématiquement, la voie P se termine sur la couche 4C béta, la voie M sur la
couche 4C alpha et la voie K sur les couches 2 et 3 (figure IV-38) en des zones
fonctionnelles bien identifiées dites blobs -spécifiquement dévolues au traitement des
paramètres spectraux de la stimulation- (figure IV-42) [Ding, Casagrande, 1998] sans
faire la différence entre les informations véhiculées par les voies ON ou OFF dont les
projections s’effectuent en un même lieu.
La couche 4 est organisée en colonnes : par exemple, les informations provenant de la
mise en activité d’un œil ou de l’autre alternent régulièrement (figure IV-43) et même en
hypercolonne [Hubel, Wiesel, 1979], [Hubel, 1988], [Bressloff, Cowan, 2003], [Alonso,
2009], [Ts'o et al., 2009].
Cellules binoculaires des couches 2 et 3 Certains groupes de cellules corticales situées dans les couches 2 et 3 sont appelés
cellules binoculaires. Elles sont de deux types 1- les cellules qui répondent
indifféremment à la stimulation d’un œil ou de l’autre ; elles seront dites, à titre
pédagogique, cellules binoculaires OU (réponse aussi bien à la stimulation d’un œil OU de
l’autre) et 2- les cellules qui ne répondent qu’à la stimulation conjointe des deux yeux,
qui sont dites cellules binoculaires ET (réponse uniquement lors de la stimulation des
deux yeux).
Elles deviennent progressivement fonctionnelles avec la mise en place de la fusion
binoculaire, qui apparaît normalement à partir de l’âge de trois mois.
Lors de stimulations binoculaires, si les cellules binoculaires ET sont normalement
fonctionnelles, elles génèrent une activité mesurable sur la réponse évoquée visuelle. En
effet, les amplitudes des PEV flash essentiellement, sont d’environ 30% supérieures
après stimulation binoculaire, à celles enregistrées après stimulations monoculaires
(figure VII-1-28).
Variations anatomiques Les aires visuelles primaires sont réparties sur les faces externe et inférieure du pôle
postérieur occipital. Elles se poursuivent à la face interne ou face interhémisphérique du
lobe occipital, sur le fond et les deux bords de la scissure calcarine.
Selon les individus, l'ensemble des deux aires visuelles primaires droite et gauche occupe
une surface variable à la face externe et inférieure des pôles occipitaux, l'enfouissement
à l'intérieur de leur partie interhémisphérique pouvant être plus ou moins important
[Brindley, 1972].
Ces variations anatomiques peuvent expliquer les variations de répartitions de potentiel
observées entre différents individus lors du recueil des PEV et en particulier lors de
l’utilisation d’une électrode active de recueil médiane en Oz (figure IV-44).
Enregistrement de l’activité des aires visuelles primaires L’activité des aires visuelles primaires droite et gauche est accessible à l’enregistrement
électrophysiologique clinique grâce à des électrodes posées en surface sur le cuir chevelu
-en O2 et O1 ou en 04 et O3- puisque les lobes occipitaux sont proches de la surface
(figure IV-40).
19

Œil et Physiologie de la Vision - IV
Le signal global émis, enregistré puis traité par des méthodes électrophysiologiques
cliniques (chapitre V-1) aboutit à l’émergence des potentiels évoqués visuels corticaux
(PEV).
Caractéristiques des PEV et propriétés des aires visuelles Les propriétés anatomiques et celles des réponses cellulaires des aires visuelles
occipitales se reflètent dans les caractéristiques des PEV.
PEV binoculaire : reflet de la maturation corticale Si le fonctionnement des cellules binoculaires ET est normal, il génère un supplément
d’activité visible à partir de 6 à 7 mois, sur les réponses évoquées qui sont plus amples
après stimulation binoculaire que monoculaire (figure VII-1-28).
Ainsi, surtout chez les petits enfants, la comparaison entre les amplitudes des PEV flash
monoculaire et binoculaire est-elle un moyen de juger du degré de maturation des aires
corticales primaires en fonction du jeune âge.
PEV : reflet du fonctionnement des aires maculaires Les PEV sont des réponses qui résultent du recueil et traitement de signaux issus des
zones postérieures des aires visuelles occipitales. Ils reflètent essentiellement l’activité
des aires maculaires avec une participation importante des réponses des zones
fovéolaires.
Ainsi, toute altération du seul fonctionnement des zones maculaires aura-t-elle pour
résultat des PEV difficilement discernables, alors que ce dysfonctionnement peut n’avoir
qu’un retentissement nul ou minime sur les caractéristiques de l’ERG flash, qui, lui,
résulte du fonctionnement global neurorétinien.
A contrario, toute altération du fonctionnement global des neurorétines, attestée par un
ERG flash difficilement discernable, peut laisser les zones maculaires correctement
fonctionnelles. Dans ce cas les PEV, réponses de l’activité maculaire, seront discernables
et peuvent même être normaux (figure IV-19).
PEV : reflet des propriétés des champs récepteurs des cellules corticales Le champ récepteur d’une cellule correspond à la surface de rétine qui, lorsqu’elle est
stimulée, entraîne une modification de son activité [Angelucci, Bressloff, 2006].
Schématiquement, les champs récepteurs des cellules corticales sont rectangulaires.
Leurs tailles varient selon que les cellules répondent à une stimulation présentée au
centre de la rétine ou à sa périphérie. Leur largeur est d'environ 15’ pour les champs
récepteurs des cellules correspondant aux projections fovéales et de 1° pour les champs
récepteurs correspondant à une zone rétinienne située à quelques degrés d’excentricité.
Dow et Bauer ont même établi une relation entre la taille du champ récepteur d'une
cellule corticale correspondant à la projection fovéale et un nombre défini de cônes qui
serait de 240 pour une excentricité rétinienne allant de 0° à 2° et même au-delà,
compatible avec les diamètres croissant des cônes avec l'excentricité [Dow, Bauer,
1984].
Pour les stimulations en damier, le choix des tailles des cases suit donc la physiologie. La
taille des cases de 15’ correspond bien à la taille des champs récepteurs des cellules
corticales de projection fovéale et la taille des cases de 1° correspond à celle des champs
récepteurs des cellules corticales de projection plus périphérique… (figure IV-16).
20

Œil et Physiologie de la Vision - IV
Les réponses évoquées ou PEV au damier dit « 15’ » correspondent donc bien ainsi aux
réponses des zones fovéales et les réponses évoquées par des cases plus grandes sont le
reflet des réponses de zones situées en dehors des fovéolas.
Pour information : les aires extrastriées…
Indispensables à l’aboutissement du processus visuel, les aires extrastriées sont
actuellement exclues de l’exploration fonctionnelle visuelle par électrophysiologie. Une
lésion localisée peut entraîner des perturbations de perception comme par exemple celle
de la couleur d’une scène visuelle lors de lésions de l’aire V4 et V4 alpha (figure IV-45)
alors que le bilan fonctionnel visuel par électrophysiologie peut être normal.
Les aires extrastriées ont initialement été mises en évidence dans le cortex visuel du
singe (figure IV-46) Leurs dénominations ont été reprises chez l’homme : aires V2, V3,
V4 et V4 alpha, V5 (ou MT) [Zeki, 1992]. Elles ne reçoivent pratiquement pas d'afférents
géniculés directs. Elles présentent des organisations fonctionnelles régulières révélées
par la cytochrome oxydase qui colore des cellules ou des groupes de cellules lorsque
l’activité de ces cellules est intense ou spécifique lors de la présentation de certains
paramètres de la stimulation (figure IV-42). Les aires extrastriées entretiennent de
nombreuses interconnexions avec d’autres aires corticales : temporales, pariétales
[Henry, 1991]…
Leur existence a été confirmée chez l'homme par des techniques de tomographie à
émission de positons [Zeki, 1990] puis par celles d'imageries fonctionnelles par
résonance magnétique nucléaire [Sereno et al., 1995] qui démontrent un parallélisme
fonctionnel certain entre les cortex visuels de l’homme et ceux du primate.
Les aires visuelles extrastriées de l’homme semblent plus larges et plus variables que
celles du primate [Van Essen et al., 2001] lorsqu’on en établit une cartographie ou un
atlas probabiliste à l’aide des variations interindividuelles [Van Essen, et al., 2001].
Leur exploration se poursuit en combinant plusieurs techniques d’exploration cérébrales
[Grill-Spector, Malach, 2004], [McKeefry et al., 2009].
Spécificité du pôle moteur
La vision n’est pas une fonction exclusivement sensorielle. La mobilité et coordination des
deux yeux et de la tête sont indispensables à son accomplissement [Proudlock, Gottlob,
2007].
Systèmes oculomoteurs
La motricité de chaque œil est assurée par 6 muscles extraoculaires, sous la dépendance
des 3 nerfs oculomoteurs dont les noyaux sont situés dans le tronc cérébral. Entre ces
noyaux et en amont, on trouve six systèmes supranucléaires qui interagissent pour
optimiser la fonction visuelle. Chacun de ces six systèmes a un substrat anatomique et
une organisation physiologique distincte qui fait intervenir un grand nombre de structures
en interrelation, réparties dans tout le système nerveux central.
Trois d’entre eux permettent la stabilisation de l’image sur la rétine, ce sont la fixation qui maintient l’image d’une cible immobile sur la fovéola, le système vestibulo-oculaire et le système opto-cinétique qui permettent de compenser les mouvements de la tête. Les trois autres sont à l’origine de la poursuite, des saccades et des vergences et contribuent à diriger la fovéola vers un objet d’intérêt.
21

Œil et Physiologie de la Vision - IV
Coordinations des deux yeux
Lorsque l’objet est au loin (ou « à l’infini »), les axes des deux yeux, ligne reliant l’objet
fixé à la fovéola, doivent être alignés. Lors des saccades et des mouvements de
poursuite, les mouvements des deux yeux sont normalement parfaitement coordonnés,
de même direction, ils sont dits « conjugués ». Un défaut de leur alignement entraîne un
strabisme, avec ou sans diplopie selon l’âge où il survient.
Lorsque l’objet est proche, les axes des deux yeux doivent se croiser au niveau de l’objet
(figure IV-47). Cette capacité de convergence est indispensable à la vision
stéréoscopique et la défaillance de la convergence entraîne une diplopie.
Un flou visuel avec parfois oscillopsie (sensation erronée d'une instabilité de la scène
visuelle) peut être due °à un nystagmus, °à des intrusions ou oscillations saccadiques qui
viennent rompre la fixation normale, °à un dysfonctionnement vestibulaire central ou
périphérique ou °à des lésions centrales qui perturbent la fixation normale [Leigh, Zee,
2006], [Tilikete et al., 2007], [Wong, 2008].
Ces mouvements anormaux vont perturber les résultats des examens
électrophysiologiques et en particulier ceux qui sont effectués en conditions binoculaires.
Retentissement sur l’EOG
Si le recueil des variations de potentiels à l’origine des signaux EOG est bien
monoculaire, l’enregistrement s’effectue lorsqu’un point cible est présenté
alternativement à droite et à gauche, la vision étant binoculaire (figure V-2-3). Un
strabisme, une paralysie oculomotrice - limitation de l’excursion d’un œil – une anomalie
des saccades oculaires, une fixation instable, rendent l’enregistrement de l’EOG difficile
voire impossible ; les résultats sont entachés d’artéfacts, donc non fiables ou
ininterprétables (figure IV-48).
Retentissement sur l’ERG flash
La stimulation par flash est délivrée aux deux yeux simultanément, le recueil du signal
est monoculaire (ERG V-3-A).
Au cours d’un strabisme par exemple, les axes oculaires ne sont plus parallèles ; dans le
plan frontal, les deux surfaces pupillaires apparentes stimulées sont différentes. Tout se
passe comme s’il y avait diminution de la surface stimulante pour l’œil strabique, par
rapport à l’œil normal, pouvant entraîner une diminution d'éclairement rétinien de l’ordre
de 25 % par exemple, pour un degré d'obliquité de 50 degrés [Le Grand, 1956] (figure
IV-49).
Il peut en résulter une asymétrie d’amplitudes des ERG recueillis pour chacun des yeux ;
dans ce contexte, il faut faire la part entre l’asymétrie due à la diminution de
l’éclairement rétinien d’un oeil par rapport à l’autre et celle éventuellement liée à une
asymétrie de fonctionnement entre les yeux…
Exemple figure IV-50. L’ERG flash est enregistré chez un sujet strabique dont l’œil droit est convergent ; la stimulation est binoculaire. Pour les trois séquences enregistrées, on observe une asymétrie d’amplitude des
réponses, moins amples à droite qu’à gauche, avec même une augmentation des temps de culmination pour la rod- et mixed-response. Ce résultat peut être dû à une diminution de l’éclairement rétinien. La stimulation a ensuite été effectuée œil par œil ; les résultats -non montrés ici- sont symétriques. On peut donc en conclure que l’asymétrie initiale des réponses était bien liée à la différence d’éclairement rétinien due au strabisme et non pas à une différence de fonctionnement entre les deux rétines.
22

Œil et Physiologie de la Vision - IV
En pratique…
Dans le cas de strabisme qu’il soit convergent ou divergent, il est indispensable de
stimuler chaque œil séparément ; la surface stimulante qu’elle soit homogène comme
pour les flashs ou structurée comme pour les damiers, doit être présentée dans un plan
perpendiculaire à l’axe visuel de chaque œil. La stimulation en hexagone -pour l’ERG
multifocal- a souvent beaucoup de mal à être fixée de façon stable par l’œil strabique ce
qui en limite la mise en œuvre chez ces sujets.
Dans le cas de nystagmus ou de dyskinésie des saccades, l’image de toute surface
structurée -qu’elle soit en damier ou en hexagone- perd sa cohérence spatiale du fait de
l’instabilité de la fixation. Il n’est donc pas possible de recueillir une réponse valable pour
les ERG multifocal, ERG pattern ou PEV damier ; pour les PEV damier, il est
éventuellement possible de se limiter à une présentation brève du damier pour
l’enregistrement des PEV onset-offset.
Conclusion
De la stimulation aux réponses visuelles, toute une chaîne anatomo-fonctionnelle
complexe s’interpose, organisée, stratifiée.
La stimulation physique en est le premier maillon mais seule la stimulation rétinienne est
efficace. C’est elle qui est à l’origine de la « mise en activité » du système visuel.
A l’électrophysiologiste de jouer entre les paramètres rétiniens et les écueils anatomo-
physiologiques dont beaucoup ont été rappelés ici, pour que les réponses enregistrées
soient les plus spécifiques possibles et témoignent des dysfonctionnements recherchés.
23

Œil et Physiologie de la Vision - IV
Figures
Figure IV-1. Succession schématique des différentes structures des milieux antérieurs.
Figure IV-2. Dioptre sphérique : formule de conjugaison.
24

Œil et Physiologie de la Vision - IV
Figure IV-3. Myopie. Le plan focal image est en avant du plan rétinien.
Figure IV-4. Hypermétropie. Le plan focal image est en arrière du plan rétinien.
25

Œil et Physiologie de la Vision - IV
Figure IV-5. Astigmatisme régulier.
Figure IV-6. Kératocône. Le sujet est porteur d’un astigmatisme myopique irrégulier.
26

Œil et Physiologie de la Vision - IV
Figure IV-7. Astigmatisme irrégulier compensé par lentille de contact.
Figure IV-8.
27

Œil et Physiologie de la Vision - IV
Figure IV-9.
Figure IV-10. En pointillé rouge, transmittance à travers la cataracte du sujet âgé avec
absorption pratiquement totale des courtes longueurs d’onde (d’après Boettner A, Wolter
JR, 1962. Invest Ophthalmol, 1, 776-783).
28

Œil et Physiologie de la Vision - IV
Figure IV-11. Variations des perceptions de Claude Monet avant et après extraction de
son cristallin (Clichés Dr Ph Lanthony).
Figure IV-12. Dix degrés centraux ou rétine centrale de l’électrophysiologiste.
29

Œil et Physiologie de la Vision - IV
Figure IV-13. Le creux fovéolaire avasculaire (Autorisation : Provis JM, Hendrickson AE,
2008. Arch Ophthalmol, 126 (4), 507-511).
Figure IV-14. Répartition des pigments maculaires dans l’épaisseur de la neurorétine
(D’après Delori et al. 2001. JOSA 18:1212-30).
30

Œil et Physiologie de la Vision - IV
Figure IV-15. Densité des pigments maculaires maximale vers 455 nm (D’après Sharpe
et al. 1998. Vis Res 38:3233-39).
Figure IV-16. Secteurs maculaires fonctionnels.
31

Œil et Physiologie de la Vision - IV
Figure IV-17. Champ visuel Goldmann ; à gauche : sujet atteint de rétinopathie
pigmentaire avec un ilôt de vision centrale ; à droite : sujet normal.
Figure IV-18. Sujet S. Correspondance entre le fond d’œil et la sensibilité relevée au
champ visuel Goldmann.
32
Œil et Physiologie de la Vision - IV
Figure IV-19. Sujet S. Résultats électrophysiologiques montrant le dysfonctionnement de
la périphérie et un fonctionnement correct des quelques degrés centraux.
Figure IV-20. Rétinographie couleur et en autofluorescence : mise en évidence de dépôts
de lipofuscine.
33

Œil et Physiologie de la Vision - IV
Figure IV-21. Axones non myélinisés des cellules ganglionnaires visibles en lumière bleue
(Cliché Dr SY Cohen).
Figure IV-22. Répartition schématique de la provenance des axones des cellules
ganglionnaires arrivant à la papille.
34

Œil et Physiologie de la Vision - IV
Figure IV-23. Déficit fasciculaire chez un patient glaucomateux (Cliché Dr SY Cohen).
Figure IV-24. Réseau choroïdien de capillaires en maille : rétine humaine (Coupe
histologique J-Ph Onolfo - Cliché Pr P Barbet).
35

Œil et Physiologie de la Vision - IV
Figure IV-25. Le pôle postérieur. Secteurs rétiniens temporal et nasal : ligne de partage
vasculaire passant par la papille.
Figure IV-26. Le pôle postérieur. Ligne de partage neuronique passant par la fovéola.
36

Œil et Physiologie de la Vision - IV
Figure IV-27
Figure IV-28. A gauche : papille avec oedème de stase à bords flous. A droite : papille
normale à bord net.
37

Œil et Physiologie de la Vision - IV
Figure IV-29 Schéma de la voie optique primaire et des voies secondaires (Schéma
François Vital-Durand).
Figure IV-30. Nerf optique. Portion rétrobulbaire et intracanalaire et vascularisation.
38

Œil et Physiologie de la Vision - IV
Figure IV-31. J. 46 ans. Champ visuel Goldmann de l’œil gauche : normal ; de l’œil
droit : large scotome central.
Figure IV-32. J. 46 ans. ERG flash, P-ERG et PEV damier en faveur d’une section
physiologique du nerf optique droit.
39

Œil et Physiologie de la Vision - IV
Figure IV-33. Voies visuelles préchiasmatiques à l’IRM (cliché Dr TH Nguyen).
Figure IV-34. Chiasma et ses possibles orientations axonales (cliché Dr TH Nguyen).
40

Œil et Physiologie de la Vision - IV
Figure IV-35
Figure IV-36
41

Œil et Physiologie de la Vision - IV
Figure IV-37. Corps géniculés latéral d’Erythrocébus patas organisé en six couches
(Préparation et cliché François Vital-Durand)
Figure IV-38. Voies visuelles schématiques : projections homo et controlatérales sur les
corps géniculés latéraux et le cortex visuel primaire.
42

Œil et Physiologie de la Vision - IV
Figure IV-39. Scissure calcarine (D’après HM Duvernoy. The human brain, surface, blood
supply and three dimensional sectional anatomy. 2nd ed. Springer V. Wien, New York,
1999).
Figure IV-40
43

Œil et Physiologie de la Vision - IV
Figure IV-41. Cortex visuel primaire d’un singe ; organisation en six couches. La couche
4 est lieu important d’afférences géniculées (Préparation et cliché François Vital-Durand).
Figure IV-42. Montage à plat, à cheval sur les aires V1 et V2 d’un singe. Coloration à la
cytochrome oxydase. Mise en évidence de l’organisation des couches 2 et 3 de l’aire V1
en blobs (Préparation et cliché François Vital-Durand).
44

Œil et Physiologie de la Vision - IV
Figure IV-43. Zones blanches : sites discontinus de projection de l’œil injecté, l’œil
adelphe se projette dans les intervalles (Préparation et cliché François Vital-Durand).
Figure IV-44. American Clinical Neurophysiology Society. Guideline 5: Guidelines for
standard electrode position nomenclature. J Clin Neurophysiol. 2006; 23:107-10.
Disponible à https://www.acns.org/.
45
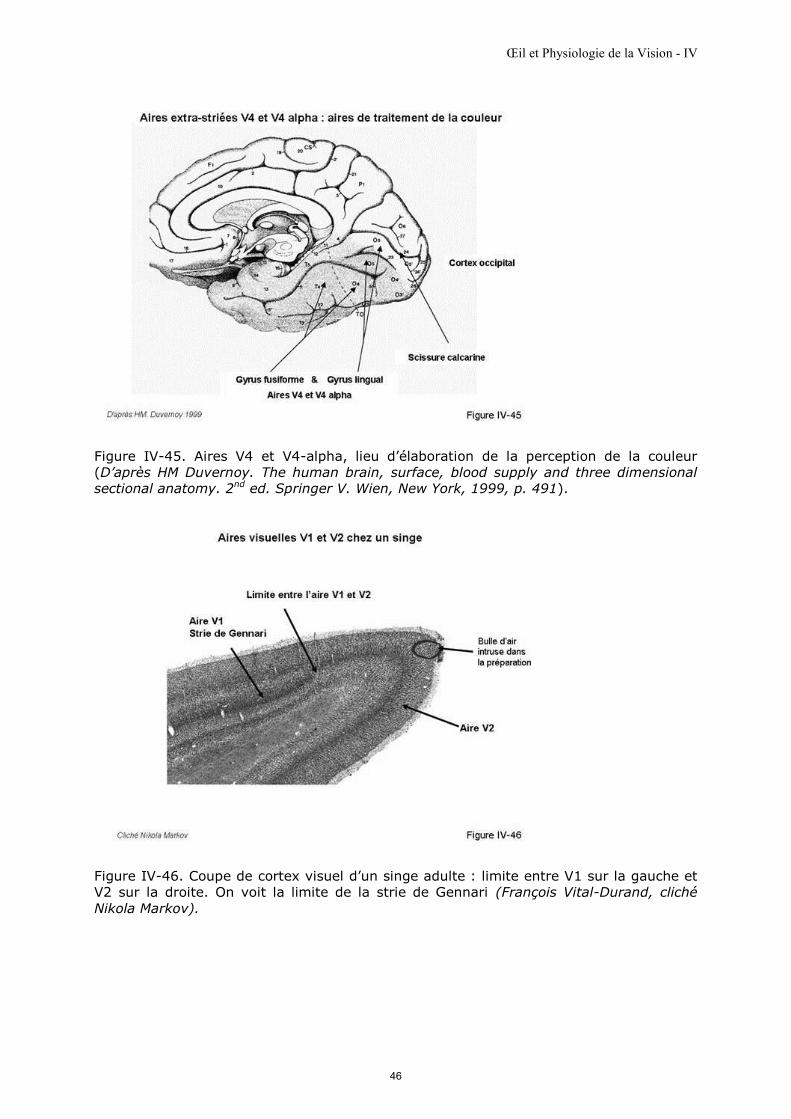
Œil et Physiologie de la Vision - IV
Figure IV-45. Aires V4 et V4-alpha, lieu d’élaboration de la perception de la couleur
(D’après HM Duvernoy. The human brain, surface, blood supply and three dimensional
sectional anatomy. 2nd ed. Springer V. Wien, New York, 1999, p. 491).
Figure IV-46. Coupe de cortex visuel d’un singe adulte : limite entre V1 sur la gauche et
V2 sur la droite. On voit la limite de la strie de Gennari (François Vital-Durand, cliché
Nikola Markov).
46

Œil et Physiologie de la Vision - IV
Figure IV-47. Vision binoculaire.
Figure IV-48. EOG. A gauche : sujet nystagmique. A droite : sujet normal.
47

Œil et Physiologie de la Vision - IV
Figure IV-49. Variation de la surface stimulante apparente d’une source en fonction de
l’obliquité de l’œil.
Figure IV-50. Variation de l’ERG flash chez un sujet strabique.
48

Œil et Physiologie de la Vision - IV
Bibliographie
Abramov, I., & Hainline, L. (1991). Light and the developing visual system. In Vision and
Visual Dysfunction: The Susceptible Visual Apparatus (pp. 104-133). Houndmills:
MacMillan Press.
Ahmad, A., & Spear, P.D. (1993). Effects of aging on the size, density, and number of
rhesus monkey lateral geniculate neurons. J Comp Neurol, 334 (4), 631-643. [Abstract]
Alonso, J.M. (2009). My recollections of Hubel and Wiesel and a brief review of functional
circuitry in the visual pathway. J Physiol, 587 (Pt 12), 2783-2790. [Abstract]
Angelucci, A., & Bressloff, P.C. (2006). Contribution of feedforward, lateral and feedback
connections to the classical receptive field center and extra-classical receptive field
surround of primate V1 neurons. Prog Brain Res, 154, 93-120. [Abstract]
Baglin, G., Betermiez, P., Bertout, A., Toussaint, P., Bremond-Gignac, D., & Milazzo, S.
(2009). [Pituitary apoplexy and severe bilateral visual loss: a case report]. J Fr
Ophtalmol, 32 (8), 572-576. [Abstract]
Baklouti, K., Mhiri, N., Mghaieth, F., & El Matri, L. (2005). [Traumatic cataract: clinical
and therapeutic aspects]. Bull Soc Belge Ophtalmol, (298), 13-17. [Abstract]
Barbur, J.L., Keenleyside, M.S., & Thomson, W.D. (1989). Investigation of central visual
processing by means of pupillometry. In Seeing contour and colour (pp. 431-451).
Oxford: Pergamon Press.
Behar-Cohen F, Kowalczuk L, Keller N, Savoldelli M, Azan F, & Jeanny J-C (2009).
Anatomie de la rétine. EMC Elsevier Masson, SAS, Paris, Ophtalmologie, 21-003-C-40, 1-
14.
Behar-Cohen F., Kowalczuk L., Keller N., Savoldelli M., Azan F., & Jeanny J-C. (2009).
Anatomie de la rétine. EMC Elsevier Masson, SAS, Paris, Ophtalmologie, 21-003-C-40, 1-
14.
Biousse, V., & Newman, N. (2009). Neuro-ophthalmology illustrated. Thieme Medical
Publishers (p. cf 249).
Bone, R.A., Landrum, J.T., Dixon, Z., Chen, Y., & Llerena, C.M. (2000). Lutein and
zeaxanthin in the eyes, serum and diet of human subjects. Exp Eye Res, 71 (3), 239-
245. [Abstract]
Bone, R.A., Landrum, J.T., Fernandez, L., & Tarsis, S.L. (1988). Analysis of the macular
pigment by HPLC: retinal distribution and age study. Invest Ophthalmol Vis Sci, 29 (6),
843-849. [Abstract]
Bressloff, P.C., & Cowan, J.D. (2003). A spherical model for orientation and spatial-
frequency tuning in a cortical hypercolumn. Philos Trans R Soc Lond B Biol Sci, 358
(1438), 1643-1667. [Abstract]
Brindley, G.S. (1972). The variability of the human striate cortex. J Physiol, 225 (2), 1-
3P. [Abstract]
Burns, D.M., Johnston, F.M., Frazer, D.G., Patterson, C., & Jackson, A.J. (2004).
Keratoconus: an analysis of corneal asymmetry. Br J Ophthalmol, 88 (10), 1252-1255.
[Abstract]
49

Œil et Physiologie de la Vision - IV
Charman, W.N. (2008). The eye in focus: accommodation and presbyopia. Clin Exp
Optom, 91 (3), 207-225. [Abstract]
Ciralsky, J., & Colby, K. (2007). Congenital corneal opacities: a review with a focus on
genetics. Semin Ophthalmol, 22 (4), 241-246. [Abstract]
Cook, C.A., Koretz, J.F., Pfahnl, A., Hyun, J., & Kaufman, P.L. (1994). Aging of the
human crystalline lens and anterior segment. Vision Res, 34 (22), 2945-2954. [Abstract]
Davies, N.P., & Morland, A.B. (2004). Macular pigments: their characteristics and
putative role. Prog Retin Eye Res, 23 (5), 533-559. [Abstract]
Delori, F.C., Dorey, C.K., Staurenghi, G., Arend, O., Goger, D.G., & Weiter, J.J. (1995).
In vivo fluorescence of the ocular fundus exhibits retinal pigment epithelium lipofuscin
characteristics. Invest Ophthalmol Vis Sci, 36 (3), 718-729. [Abstract]
Delori, F.C., Goger, D.G., Hammond, B.R., Snodderly, D.M., & Burns, S.A. (2001).
Macular pigment density measured by autofluorescence spectrometry: comparison with
reflectometry and heterochromatic flicker photometry. J Opt Soc Am A Opt Image Sci
Vis, 18 (6), 1212-1230. [Abstract]
Ding, Y., & Casagrande, V.A. (1998). Synaptic and neurochemical characterization of
parallel pathways to the cytochrome oxidase blobs of primate visual cortex. J Comp
Neurol, 391 (4), 429-443. [Abstract]
Dow, B.M. (1990). Nested maps in Macaque monkey visual cortex. In Science of Vision
(p. 87). New York: Springer Verlag.
Dow, B.M., & Bauer, R. (1984). Retinotopy and orientation columns in the monkey: a
new model. Biol Cybern, 49 (3), 189-200. [Abstract]
Dow, B.M., Snyder, A.Z., Vautin, R.G., & Bauer, R. (1981). Magnification factor and
receptive field size in foveal striate cortex of the monkey. Exp Brain Res, 44 (2), 213-
228. [Abstract]
Ducasse, A., & Segal, A. (1985). Anatomie de la rétine. Encycl. Médico-chirurgicale:
Ophtalmologie (pp. 1-8). Paris: Elsevier.
el-Hage, S.G., & Berny, F. (1973). Contribution of the crystalline lens to the spherical
aberration of the eye. J Opt Soc Am, 63 (2), 205-211. [Abstract]
Flammer, J., & Orgul, S. (1998). Optic nerve blood-flow abnormalities in glaucoma. Prog
Retin Eye Res, 17 (2), 267-289. [Abstract]
Gellermann, W., & Bernstein, P.S. (2004). Noninvasive detection of macular pigments in
the human eye. J Biomed Opt, 9 (1), 75-85. [Abstract]
Grill-Spector, K., & Malach, R. (2004). The human visual cortex. Annu Rev Neurosci, 27,
649-677. [Abstract]
Grunauer-Kloevekorn, C., & Duncker, G.I. (2006). [Keratoconus: epidemiology, risk
factors and diagnosis]. Klin Monatsbl Augenheilkd, 223 (6), 493-502. [Abstract]
Guillery, R.W., Okoro, A.N., & Witkop, C.J., Jr. (1975). Abnormal visual pathways in the
brain of a human albino. Brain Res, 96 (2), 373-377. [Abstract]
Hamel, C. (2006). Retinitis pigmentosa. Orphanet J Rare Dis, 1, 40. [Abstract]
50

Œil et Physiologie de la Vision - IV
Hammond, B.R., & Caruso-Avery, M. (2000). Macular pigment optical density in a
Southwestern sample. Invest Ophthalmol Vis Sci, 41 (6), 1492-1497. [Abstract]
Hendry, S.H., & Reid, R.C. (2000). The koniocellular pathway in primate vision. Annu Rev
Neurosci, 23, 127-153. [Abstract]
Henry, G.H. (1991). Afferent inputs, receptive field properties and morphological cell
types in different laminae of the striate cortex. In Vision and visual dysfunction: The
Neural Basis of Visual Function (p. 239). London: MacMillan Press.
Horton, J.C., & Hoyt, W.F. (1991). The representation of the visual field in human striate
cortex. A revision of the classic Holmes map. Arch Ophthalmol, 109 (6), 816-824.
[Abstract]
Hubel, D.H. (1988). Eye, Brain and Vision. (pp. 1-245). New York: Scientific American
Library.
Hubel, D.H., & Wiesel, T. (1979). Les mécanismes cérébraux de la vision. In Les
mécanismes cérébraux de la vision (pp. 79-93).
Kim, J.S., Ishikawa, H., Gabriele, M.L., Wollstein, G., Bilonick, R.A., Kagemann, L.,
Fujimoto, J.G., & Schuman, J.S. (2009). Retinal Nerve Fiber Layer Thickness
Measurement Comparability between Time Domain Optical Coherence Tomography (OCT)
and Spectral Domain OCT. Invest Ophthalmol Vis Sci, 2010 Feb ; 51(2):896-902.
[Abstract]
Krag, S., & Andreassen, T.T. (2003). Mechanical properties of the human lens capsule.
Prog Retin Eye Res, 22 (6), 749-767. [Abstract]
Kupfer, C., Chumbley, L., & Downer, J.C. (1967). Quantitative histology of optic nerve,
optic tract and lateral geniculate nucleus of man. J Anat, 101 (3), 393-401. [Abstract]
Land, M.F. (2006). Eye movements and the control of actions in everyday life. Prog Retin
Eye Res, 25 (3), 296-324. [Abstract]
Lang, G.E. (2007). Optical coherence tomography findings in diabetic retinopathy. Dev
Ophthalmol, 39, 31-47. [Abstract]
Le Grand, Y. (1956). L'Espace Visuel. In Optique Physiologique (p. 140). Paris: Revue
d'Optique.
Le Grand, Y. (1972). Lumière et Couleurs. In Optique Physiologique (p. 229). Paris:
Masson.
Leigh, R.J., & Zee, D.S. (2006). The neurology of Eye Movements. Contemporary
Neurology Series, 4° ed (p. 763 ). New York: Oxford University Press.
Loewenfeld, I.E. (1999). The pupil: anatomy, physiology and clinical applications. (p.
2223). Boston, Oxford Butterworth Heinenmann
McKeefry, D.J., Gouws, A., Burton, M.P., & Morland, A.B. (2009). The noninvasive
dissection of the human visual cortex: using FMRI and TMS to study the organization of
the visual brain. Neuroscientist, 15 (5), 489-506. [Abstract]
Milton, J.G., & Longtin, A. (1990). Evaluation of pupil constriction and dilation from
cycling measurements. Vision Res, 30 (4), 515-525. [Abstract]
51

Œil et Physiologie de la Vision - IV
Nguyen, T.H., Stievenart, J.L., Yoshida, M., Iba-Zizen, M.T., Bellinger, L., Abanou, A., &
Cabanis, E.A. (2003). [Tractography of the visual pathways: routine examination in
magnetic resonance imaging]. J Fr Ophtalmol, 26 (9), 941-951. [Abstract]
Nolan, J.M., Stack, J., O, O.D., Loane, E., & Beatty, S. (2007). Risk factors for age-
related maculopathy are associated with a relative lack of macular pigment. Exp Eye Res,
84 (1), 61-74. [Abstract]
Overgaard, M., Fehl, K., Mouridsen, K., Bergholt, B., & Cleeremans, A. (2008). Seeing
without Seeing? Degraded Conscious Vision in a Blindsight Patient. PLoS One, 3 (8),
e3028. [Abstract]
Pakzaban, P., Westmark, K., & Westmark, R. (2000). Chiasmal apoplexy due to
hemorrhage from a pituitary adenoma into the optic chiasm: case report. Neurosurgery,
46 (6), 1511-1513; discussion 1513-1514. [Abstract]
Polo, V., Pinilla, I., Abecia, E., Larrosa, J.M., Pablo, L.E., & Honrubia, F.M. (1996).
Assessment of the ocular media absorption index. Int Ophthalmol, 20 (1-3), 7-9.
[Abstract]
Popovic, Z., & Sjostrand, J. (2001). Resolution, separation of retinal ganglion cells, and
cortical magnification in humans. Vision Res, 41 (10-11), 1313-1319. [Abstract]
Praveen, M.R., Vasavada, A.R., Jani, U.D., Trivedi, R.H., & Choudhary, P.K. (2008).
Prevalence of cataract type in relation to axial length in subjects with high myopia and
emmetropia in an Indian population. Am J Ophthalmol, 145 (1), 176-181. [Abstract]
Proudlock, F.A., & Gottlob, I. (2007). Physiology and pathology of eye-head coordination.
Prog Retin Eye Res, 26 (5), 486-515. [Abstract]
Provis, J.M., & Hendrickson, A.E. (2008). The foveal avascular region of developing
human retina. Arch Ophthalmol, 126 (4), 507-511. [Abstract]
Richdale, K., Bullimore, M.A., & Zadnik, K. (2008). Lens thickness with age and
accommodation by optical coherence tomography. Ophthalmic Physiol Opt, 28 (5), 441-
447. [Abstract]
Roche, O., Beby, F., Orssaud, C., Dupont Monod, S., & Dufier, J.L. (2006). [Congenital
cataract: general review]. J Fr Ophtalmol, 29 (4), 443-455. [Abstract]
Rougier, M.B., Delyfer, M.N., & Korobelnik, J.F. (2008). [Measuring macular pigment in
vivo]. J Fr Ophtalmol, 31 (4), 445-453. [Abstract]
Said, F.S., & Weale, R.A. (1959). The variation with age of the spectral transmissivity of
the living human crystalline lens. Gerontologia, 3, 213-231. [Abstract]
Sample, P.A., Esterson, F.D., Weinreb, R.N., & Boynton, R.M. (1988). The aging lens: in
vivo assessment of light absorption in 84 human eyes. Invest Ophthalmol Vis Sci, 29 (8),
1306-1311. [Abstract]
Savage, G.L., Johnson, C.A., & Howard, D.L. (2001). A comparison of noninvasive
objective and subjective measurements of the optical density of human ocular media.
Optom Vis Sci, 78 (6), 386-395. [Abstract]
Schuman, J.S. (2008). Spectral domain optical coherence tomography for glaucoma (an
AOS thesis). Trans Am Ophthalmol Soc, 106, 426-458. [Abstract]
52

Œil et Physiologie de la Vision - IV
Seddon, J.M., Ajani, U.A., Sperduto, R.D., Hiller, R., Blair, N., Burton, T.C., Farber, M.D.,
Gragoudas, E.S., Haller, J., Miller, D., & et al. (1994). Dietary carotenoids, vitamins A, C,
and E, and advanced age-related macular degeneration. Eye Disease Case-Control Study
Group. Jama, 272 (18), 1413-1420. [Abstract]
Sereno, M.I., Dale, A.M., Reppas, J.B., Kwong, K.K., Belliveau, J.W., Brady, T.J., Rosen,
B.R., & Tootell, R.B. (1995). Borders of multiple visual areas in humans revealed by
functional magnetic resonance imaging. Science, 268 (5212), 889-893. [Abstract]
Sharpe, L.T., Stockman, A., Knau, H., & Jagle, H. (1998). Macular pigment densities
derived from central and peripheral spectral sensitivity differences. Vision Res, 38 (21),
3233-3239. [Abstract]
Snodderly, D.M., Auran, J.D., & Delori, F.C. (1984). The macular pigment. II. Spatial
distribution in primate retinas. Invest Ophthalmol Vis Sci, 25 (6), 674-685. [Abstract]
Snodderly, D.M., Brown, P.K., Delori, F.C., & Auran, J.D. (1984). The macular pigment. I.
Absorbance spectra, localization, and discrimination from other yellow pigments in
primate retinas. Invest Ophthalmol Vis Sci, 25 (6), 660-673. [Abstract]
Stoerig, P. (2006). Blindsight, conscious vision, and the role of primary visual cortex.
Prog Brain Res, 155, 217-234. [Abstract]
Stoerig, P. (2010). Cueless blindsight. Front Hum Neurosci, Vol 3 - Art 74, 1-8.
[Abstract]
Strenk, S.A., Strenk, L.M., & Koretz, J.F. (2005). The mechanism of presbyopia. Prog
Retin Eye Res, 24 (3), 379-393. [Abstract]
Sureda, B., Alberca, R., & Boza, F. (1991). [Visual evoked potentials in benign
intracranial hypertension]. Neurologia, 6 (7), 242-246. [Abstract]
Tilikete, C., Pisella, L., Pelisson, D., & Vighetto, A. (2007). [Oscillopsia:
pathophysiological mechanisms and treatment]. Rev Neurol (Paris), 163 (4), 421-439.
[Abstract]
Tovée, M.J. (1996). An introduction to the visual system. (pp. 1-202): Cambridge
University Press.
Ts'o, D.Y., Zarella, M., & Burkitt, G. (2009). Whither the hypercolumn? J Physiol, 587 (Pt
12), 2791-2805. [Abstract]
van de Kraats, J., Berendschot, T.T., & van Norren, D. (1996). The pathways of light
measured in fundus reflectometry. Vision Res, 36 (pp. 2229-2247). [Abstract]
Van den Berg, T.J., & Spekreijse, H. (1997). Near infrared light absorption in the human
eye media. Vision Res, 37 (2), 249-253. [Abstract]
van der Veen, R.L., Ostendorf, S., Hendrikse, F., & Berendschot, T.T. (2009). Macular
pigment optical density relates to foveal thickness. Eur J Ophthalmol, 19 (5), 836-841.
[Abstract]
Van Essen, D.C., Lewis, J.W., Drury, H.A., Hadjikhani, N., Tootell, R.B., Bakircioglu, M., &
Miller, M.I. (2001). Mapping visual cortex in monkeys and humans using surface-based
atlases. Vision Res, 41 (10-11), 1359-1378. [Abstract]
53

Œil et Physiologie de la Vision - IV
Vaughan, D., & Asbury, T. (1986). Ophtalmologie générale. (pp. 297-302). Padova:
Piccin.
Victor, J.D., Conte, M.M., & Adecola, C. (1991). Ocular dependence of hemifield visual
evoked potentials : relevance to bilateral cortical representation of central vision. Clin
Vision Sci, 6 (4), 261-276.
Vital-Durand, F. (1986). Anatomie fonctionnelle des voies optiques intracrâniennes.
Encyclopédie Médico-Chirurgicale: Ophtalmologie, 21-008-A-40 (pp. 1-10). Paris:
Elsevier.
Votruba, M., Aijaz, S., & Moore, A.T. (2003). A review of primary hereditary optic
neuropathies. J Inherit Metab Dis, 26 (2-3), 209-227. [Abstract]
Wassle, H., Grunert, U., Rohrenbeck, J., & Boycott, B.B. (1989). Cortical magnification
factor and the ganglion cell density of the primate retina. Nature, 341 (6243), 643-646.
[Abstract]
Werner, J.S., Bieber, M.L., & Schefrin, B.E. (2000). Senescence of foveal and parafoveal
cone sensitivities and their relations to macular pigment density. J Opt Soc Am A Opt
Image Sci Vis, 17 (11), 1918-1932. [Abstract]
Whitehead, A.J., Mares, J.A., & Danis, R.P. (2006). Macular pigment: a review of current
knowledge. Arch Ophthalmol, 124 (7), 1038-1045. [Abstract]
Wong, A.M.F. (2008). Eye Movements Disorders. (p. 295). New York: Oxford University
Press.
Wooten, B.R., & Hammond, B.R. (2002). Macular pigment: influences on visual acuity
and visibility. Prog Retin Eye Res, 21 (2), 225-240. [Abstract]
Zeki, S. (1990). Colour vision and functional specialisation in the visual cortex. In
Discussions in Neuroscience (pp. VI-n°2): Elsevier
Zeki, S. (1992). Les images visuelles. In Pour la Science (pp. 60-68). Paris: Belin.
54
Related Documents











