Open Research Online The Open University’s repository of research publications and other research outputs Iterative evolution of digitate planktonic foraminifera Journal Article How to cite: Coxall, Helen K.; Wilson, Paul A.; Pearson, Paul N. and Sexton, Philip F. (2007). Iterative evolution of digitate planktonic foraminifera. Paleobiology, 33(4) pp. 495–516. For guidance on citations see FAQs . c 2007 The Paleontological Society Version: Not Set Link(s) to article on publisher’s website: http://paleobiol.geoscienceworld.org/ Copyright and Moral Rights for the articles on this site are retained by the individual authors and/or other copy- right owners. For more information on Open Research Online’s data policy on reuse of materials please consult the policies page. oro.open.ac.uk

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Open Research OnlineThe Open University’s repository of research publicationsand other research outputs
Iterative evolution of digitate planktonic foraminifera
Journal ArticleHow to cite:
Coxall, Helen K.; Wilson, Paul A.; Pearson, Paul N. and Sexton, Philip F. (2007). Iterative evolutionof digitate planktonic foraminifera. Paleobiology, 33(4) pp. 495–516.
For guidance on citations see FAQs.
c© 2007 The Paleontological Society
Version: Not Set
Link(s) to article on publisher’s website:http://paleobiol.geoscienceworld.org/
Copyright and Moral Rights for the articles on this site are retained by the individual authors and/or other copy-right owners. For more information on Open Research Online’s data policy on reuse of materials please consultthe policies page.
oro.open.ac.uk

� 2007 The Paleontological Society. All rights reserved. 0094-8373/07/3304-0001/$1.00
Paleobiology, 33(4), 2007, pp. 495–516
Iterative evolution of digitate planktonic foraminifera
Helen K. Coxall, Paul A. Wilson, Paul N. Pearson, and Philip F. Sexton
Abstract.—Digitate shell morphologies have evolved repeatedly in planktonic foraminiferathroughout the Cretaceous and Cenozoic. Digitate species are usually rare in fossil and modernassemblages but show increased abundance and diversity at times during the Cretaceous and mid-dle Eocene. In this paper we discuss the morphology and stratigraphic distribution of digitateplanktonic foraminifera and establish the isotopic depth ecology of fossil ones to draw parallelswith modern counterparts. �18O and �13C values of six extinct and two modern digitate species,from six time slices (Cenomanian, Turonian, Eocene, Miocene, Pleistocene and Holocene) have sim-ilar isotopic depth ecologies, consistently registering the most negative �13C and usually the mostpositive �18O compared to coexisting species. These results indicate a similar deep, subthermocline(�150 m) habitat, characterized by lower temperatures, reduced oxygen, and enrichment of dis-solved inorganic carbon. This is consistent with water-column plankton studies that provide in-sight into the depth preferences of the three modern digitate species; in over 70% of observationsdigitates occurred in nets below 150 m, and down to 2000 m. The correlation between digitatespecies and subsurface habitats across multiple epochs suggests that elongated chambers were ad-vantageous for survival in a deep mesopelagic habitat, where food is usually scarce. Increasedabundance and diversity of digitates in association with some early and mid-Cretaceous oceanicanoxic events, in middle Eocene regions of coastal and equatorial upwelling, and occasionally insome modern upwelling regions, suggests an additional link with episodes of enhanced ocean pro-ductivity associated with expansion of the oxygen minimum zone (OMZ). We suggest that the pri-mary function of digitate chambers was as a feeding specialization that increased effective shellsize and food gathering efficiency, for survival in a usually food-poor environment, close to theOMZ. Episodes of increased digitate abundance and diversity indicate expansion of the deep-waterecologic opportunity under conditions that were unfavorable to other planktonic species. Our re-sults provide evidence of iterative evolution reflecting common functional constraints on plank-tonic foraminifera shell morphology within similar subsurface habitats. They also highlight thepotential of digitate species to act as indicators of deep watermasses, especially where there wasexpansion of the OMZ.
Helen K. Coxall and P. N. Pearson.* School of Earth, Ocean and Planetary Sciences, Cardiff University,Main Building, Cardiff, CF10 3AT, United Kingdom. E-mail: [email protected]
P. A. Wilson and P. F. Sexton.* School of Ocean and Earth Science, National Oceanography Centre, South-ampton, European Way, Southampton, SO14 3ZH, United Kingdom
*Present address: Scripps Institution of Oceanography, University of California, San Diego, La Jolla, Cal-ifornia 92093-0244, and School of Earth, Ocean and Planetary Sciences, Cardiff University, Main Build-ing, Cardiff, CF10 3AT, United Kingdom
Accepted: 13 June 2007
Introduction
The fossil record of planktonic foraminiferareveals the repeated evolution of chambershapes and shell morphologies during inde-pendent Cretaceous and Cenozoic evolution-ary radiations (e.g., Berger 1969; Cifelli 1969;Frerichs 1971; Steineck and Fleischer 1978;Lipps 1979; Hart 1980; Caron and Homewood1983; Leckie 1989; Norris 1991; Pearson 1996;Moullade et al. 2002). Identifying biotic andabiotic factors that have played a role in selec-tion of morphologies across these radiations isa continuous goal for micropaleontologistsand an important area of research for pale-ontology in general. However, linking plank-
tonic ecology with shell morphology has beenproblematic, and isotopic data do not reveal aconsistent correlation with preferred depthhabitat among most common globular andcompressed shell morphologies (Berger 1969;Cifelli 1969; Frerichs 1971; Lipps 1979; Hart1980; Caron and Homewood 1983; reviews inCorfield and Cartlidge 1991, and Pearson1998).
A conspicuous but usually rare element ofplanktonic foraminifera assemblages through-out the Cretaceous and Cenozoic are ‘‘digitate’’planktonic foraminifera, i.e., forms that have oneor more chambers of the adult whorl radiallyelongated to form distinctive fingerlike exten-

496 HELEN K. COXALL ET AL.
sions. Little is known about the ecology of dig-itate species because of their scarcity and patchydistribution. Limited plankton-tow data (e.g.,Rhumbler 1911; Bradshaw 1959; Be 1977) havesuggested an unusually deep dwelling habitatfor modern digitates and a similar ecology hasbeen predicted for fossil species on the basis ofmorphological similarities (e.g., Berger 1969; Ci-felli 1969; Frerichs 1971; Steineck and Fleischer1978; Hart 1980; Caron and Homewood 1983;Leckie 1989; Norris 1991; Pearson 1996; Moul-lade et al. 2002). Several reports have also linkeddigitates from the Recent, Eocene, and Creta-ceous with enhanced surface ocean productivi-ty and/or low-oxygen conditions (e.g., Brad-shaw 1959; Magniez-Jannin 1998; Coccioni et al.2006; Coxall and Pearson 2006). These observa-tions suggest a correlation between digitateshell morphology and the environment but sup-porting data have been lacking. Here we test thehypothesis that digitate shell morphology is afunctional specialization to a deep-dwellingecology by comparing the isotopic paleodepthsignatures of digitate species from multiple timeslices. By integrating the geochemical data withstratigraphic and paleoenvironmental informa-tion, and with existing water-column planktonsampling data, we are able to explore parallelsbetween fossil and modern counterparts and as-sess the paleoceanographic significance of theseenigmatic forms.
Shell Function and Depth Ecology. Deter-mining the functional morphology of forami-nifera shells is a challenge because of theirsmall size (�1 mm), the difficulties in sustain-ing them under laboratory conditions, and thelimitation of their fossil remains and encasingsediment to provide environmental informa-tion. Oxygen and carbon stable isotopes (�13Cand �18O) measured on foraminiferal shell cal-cite can provide valuable insights into plank-tonic ecology that can be used to test aspectsof functional models for chamber design re-lated to depth habitat.
The method relies on isotope fractionationprocesses occurring in the water column. Ox-ygen isotope fractionation between ambientseawater and foraminiferal calcite during cal-cification is temperature dependent. Thismeans that depth-stratified foraminiferal as-semblages from open-ocean sites can be ex-
pected to exhibit a trend of increasing fora-miniferal �18O with depth that parallels thetrend of decreasing temperature (Fairbanks etal. 1980, 1982). Species living in the warmestwaters closest to the surface, therefore, regis-ter more negative �18O than deeper dwellersand bottom living (benthic) species. In con-trast, foraminiferal �13C decreases with depthbecause of the preferential uptake of 12CO2
during photosynthesis in the surface euphoticzone and its subsequent remineralization backinto the �CO2 pool by respiration at depth. Asa result, dissolved inorganic carbon (DIC) inthe modern surface ocean may be 1‰ to 2‰more positive than the �13C of DIC in the deepocean (Kroopnick 1985) and the shells of sur-face dwelling forms would be expected tohave more positive �13C values than deeperdwellers. However, �18O and �13C values canbe offset from ‘‘equilibrium’’ with ambientseawater by the effect on isotope fractionationof environmental parameters such as [CO3
2�]and pH (Spero et al. 1997; Zeebe 1999) orphysiological processes such as symbiont pho-tosynthesis or foraminiferal respiration (Spe-ro and Williams 1989; Spero et al. 1991; Speroand Lea 1993, 1996; Ortiz et al. 1996). Despitethese complications, which tend to affect �13Cmore than �18O, the basic pattern of forami-niferal �18O and �13C variation with water-col-umn depth facilitates the reconstruction offossil foraminiferal depth habitats based onthe relative stable isotope offsets between spe-cies in an assemblage (see Spero 1998; Pearson1998; Pearson et al. 2001; Sexton et al. 2006).This method is used widely to identify fora-miniferal tracers of shallow and deep water-masses on multiple geological scales.
Previous stable isotope and environmentalinterpretations have suggested some function-al fits between various planktonic foraminif-era groups but there are plenty of non-ana-logues. For example, the Paleogene morozo-vellids and Cretaceous globotruncanids hadcomparable ornate keeled shells and their iso-topic signatures suggest they were both sur-face dwelling and symbiotic (Houston andHuber 1998; Norris 1996). The living speciesGloborotalia menardii, which has a keeled un-ornamented shell, and its Paleogene analogueGlobanomalina pseudomenardii are or were both

497DIGITATE FORAM ITERATIVE EVOLUTION
deep dwellers (Hemleben et al. 1989; Olsson etal. 1999). This is where the similarities end, be-cause the Eocene G. menardii-analogue Turbo-rotalia cunialensis was a surface mixed layerform (Pearson et al. 2006b), whereas counter-parts from the Cretaceous occurred in bothshallow (Planomalina buxtorfi) and deeper (Ro-talipora spp.) habitats (Wilson and Norris2001). Among the ‘‘globigerine’’ morphotypesthere are also differences in depth habitat. Forexample, modern Globigerinoides sacculifer andG. ruber live in shallow surface waters whereasPaleogene analogues in the genus Subbotinaroutinely register deep, probably thermoclineisotopic signatures (Olsson et al. 2006a). Smalltriserial forms on the other hand, which occurin modern oceans and throughout the Creta-ceous and Paleogene, are or were all surfacedwellers, specialized to continental marginupwelling environments and unstable marineconditions generally (Kroon and Nederbragt1990).
In many of these examples the functionalcomparisons are less than perfect becausethere are usually fine-scale differences in mor-phology beyond the general form (e.g., keelsmay be double or single, ornamentation maybe built of muricae or pustules), indicatingthat there have been various structural con-vergences that may not always have similarfunctions in detail. The correlation betweenmorphology and depth habitat is also far fromsimple, because, as in the case of the moderncompressed globorotaliids, some species mi-grate from the surface to deeper parts of thewater column during ontogeny. Digitate mor-phologies, however, which involve simpleelongations of individual chambers, are morelikely to be homologous structures and, wesuppose, are more likely to have a similarfunction.
Materials and Methods
The stratigraphic distribution of digitateplanktonic foraminifera from the Cretaceousand Cenozoic was recorded from new micro-paleontological observations in deep-sea corematerial and the literature. Morphologies areconsidered digitate if one or more adult cham-bers are radially elongated and have an aspectratio (chamber length divided by width at the
midpoint) of 1.5 or greater. This arrangementresults in widely separated chambers anddeeply incised sutures giving individuals adistinctly lobate outline. We include formswith bifurcating digitate chambers, as some-times occurs in the genus Leupoldina and inHastigerinella digitata but exclude Globigerinoi-des fistulosus, which has multiple protuberanc-es on individual chambers. By our definition,tubulospines, as occur in Hantkenina andSchackoina, do not alone constitute a digitateform because they are slender non-perforateprojections, whereas digitate chambers are ex-tended chambers with continuous wall tex-ture. The exceptions to this are the earliesthantkeninids and some schackoinids that havetubulospines (or prototubulospines, e.g.,Hantkenina singanoae [Coxall and Pearson2006]) emerging from elongated chambers.We regard tubulospines as a different struc-tural modification, although they may haveoriginally served a similar function (i.e., in in-creasing the effective size of the foraminifera).
Evidence for the distribution and ecology ofliving digitate species was synthesized fromthe literature and unpublished observationsfrom plankton surveys. A reconstruction ofthe geographic distribution of digitate specieswas produced for the middle Eocene, a time ofhigh digitate diversity that is well representedin deep-sea and continental margin sedi-ments. �13C and �18O analysis was used to in-vestigate the paleoecologies of digitate hom-eomorphs from six time-slices in seven mid-to low-latitude deep-sea sites in the Atlanticand Pacific Oceans (Table 1). Preservation offoraminiferal specimens varied between sites.Cenomanian, Turonian, and Pleistocene sam-ples show excellent (glassy) preservation (seeWilson and Norris 2001; Pearson et al. 2004;Sexton et al. 2006). Scanning electron micro-scope (SEM) examination reveals neomorphicrecrystallization of foraminiferal calcite at themicron scale (see Sexton et al. 2006) in theMiocene and Eocene material, suggesting thatthe primary isotopic signal has been partiallyaltered through diagenesis. The effect of dia-genetic alteration on planktonic foraminiferalcalcite close to the seafloor will be to increaseoxygen isotope values, and this effect will begreatest at low latitudes (where the vertical

498 HELEN K. COXALL ET AL.
TA
BL
E1.
Tim
esl
ices
and
loca
liti
esin
the
isot
opic
pal
eo-d
epth
anal
ysi
s.
Ep
och
Bio
zon
eA
ge
Loc
alit
yL
at.
Lon
g.
Lit
hol
ogy
Dig
itat
eta
xon
Abu
nd
ance
Hol
ocen
e—
—C
ore
CD
154-
07-
7PK
,A
gu
lhas
Cu
rren
t
30�0
7.9�
S,31
�41.
8�E
Ora
ng
esi
lty
fo-
ram
ooze
Bee
lla
digi
tata
Rar
e
Lat
eP
leis
toce
ne
—14
.2K
a1B
oxC
ED
RC
92,
W.
Pac
ific
2�13
.5�S
,15
6�59
.9�E
Fora
m-n
ann
ooo
zeG
lobi
geri
nell
aad
amsi
Rar
e
Mid
dle
Mio
cen
eN
102
14.7
–15.
3M
aO
DP
Site
872,
N.E
.Pac
ific
10�6
�N,1
62�5
2�E
Fora
m-n
ann
ooo
zeC
lava
tore
lla
berm
udez
iR
are
Lat
est
earl
yE
ocen
eP
9349
–50.
4M
aK
ane-
9C
-42,
N.E
.Atl
anti
c21
�N,1
9�W
Fora
m-n
ann
ooo
zeC
lavi
geri
nell
aco
lom
bi-
ana,
C.
aker
si,C
.ja
r-vi
si
Few
-com
mon
Lat
est
earl
yE
ocen
eP
9349
–50.
4M
aO
DP
Site
865,
mid
Pac
ific
18�2
6.42
5�N
,179
�3.3
�WFo
ram
-nan
no
ooze
Cla
vige
rine
lla
eoca
nic
aFe
w
Lat
eT
uro
nia
nU
C9a
4,5
88–9
0M
aD
SDP
Site
144,
N.
Atl
anti
c9�
27�N
,54�
21�W
Bla
cksh
ales
,ca
rbon
aceo
us
mu
dst
ones
Cla
vihe
dber
gell
aw
ater
siR
are
Cen
oman
ian
KS1
7399
.1–9
6.8
Ma
DSD
PSi
te13
725
�55.
53�N
,27�
03.6
4�W
Nan
no
mar
l/ch
alk
‘‘Hed
berg
ella
’’si
mpl
exR
are
Dat
aso
urc
es:1 C
-14
dat
afr
omP
eng
etal
.(19
79)
con
vert
edto
cale
nd
arye
ars
usi
ng
cali
brat
ion
ofSt
uiv
eran
dR
eim
er(1
993)
;2 Pea
rson
1995
;3 Th
isst
ud
y,ti
mes
cale
and
zon
atio
nof
Ber
gg
ren
etal
.(19
95);
4 Wil
son
etal
.200
2;5 A
rth
ur
and
Nat
lan
d19
79.
temperature gradient in the water column isgreatest and where sedimentation rates arelow; Schrag et al. 1995; Rudnicki et al. 2001).Previous studies have demonstrated, however,that interspecies offsets, which we assume re-flect original differences in depth habitat, aremaintained despite such minor alteration, al-though the absolute �18O values are likely tobe artificially increased and the multispeciesgradient compressed (Corfield et al. 1990;Pearson et al. 2001; Sexton et al. 2006).
�13C and �18O was measured on monospe-cific samples of digitate morphotypes plusthree or more additional taxa and a benthicspecies (where possible) from each assem-blage representing a range of mixed-layer anddeeper-water ecologies. Samples were com-posed of 2–25 specimens, depending on spe-cies size and abundance (Appendix). Analyseswere performed at the National Oceanogra-phy Centre in the United Kingdom, using aEuropa Geo 20–20 mass spectrometerequipped with a ‘‘CAPS’’ automatic carbonatepreparation system and at Cardiff Universityon a MAT252 gas source mass spectrometerwith an automated KIEL device. Results arereported relative to Vienna Pee Dee Belemnitestandard (VPDB). Standard analytical preci-sion is better than 0.1‰ for �18O and �13C forall data.
Results
Digitate Species in the Cretaceous and Cenozo-ic. We recognize a minimum of 27 digitatespecies in 13 genera (Fig. 1), each havingchamber aspect ratios of 1.5 or greater in oneor more of the adult chambers (Table 2). Thenumber of chambers in the final whorl is con-sistent within species but varies from 3.5 to 7between taxa. The distal ends of the chambersalso vary between species, ranging from sim-ple and rounded (e.g., ‘‘Hedbergella’’ simplex,Clavigerinella eocanica, and Clavatorella bermu-dezi), to bulb-like (e.g., Leupoldina spp., Clavi-hedbergella watersi, Clavigerinella akersi) orpointed (Clavihedbergella alexanderi, Clavigeri-nella caucasica, and Globigerinella adamsi). Coil-ing is mostly low trochospiral or planispiralexcept in Beella digitata and Hastigerinella digi-tata, which tend to show more irregular coil-ing patterns. SEM imaging reveals differences

499DIGITATE FORAM ITERATIVE EVOLUTION
FIGURE 1. The stratigraphic distribution of living and fossil planktonic foraminifera with digitate morphologies,highlighting the iterative evolution of this morphology in successive evolutionary radiations. Foraminiferal diver-sity trends (right) from Norris (1991) and Leckie et al. (2002). Ranges are approximate because occurrences aresporadic. Note: The Pliocene to Holocene timescale is exaggerated. Data sources—Cretaceous: Longoria 1974; Mag-niez-Jannin 1998; Premoli Silva and Sliter 1999; Verga and Premoli Silva 2002; Coccioni et al. 2006; Mesozoic Plank-tonic Foraminifera Working Group. Paleogene: Coxall and Pearson 2006; Quilty 1976; de Klasz et al. 1987; Cicha etal. 1998. Neogene–Quaternary: Cicha et al. 1998; Kennett and Srinivasan 1983; Pearson 1995; Saito et al. 1976; Sri-nivasan and Kennett 1975. Generic abbreviations: B. Beella, C. Clavihedbergella, Cl. Clavigerinella, Clt. Cla-vatorella, E. Eohastigerinella, G. Globigerinella, Gn. Globigerinelloides, H. Hedbergella, Ha. Hastigerinella,Hastigerin. Hastigerinoides, Hk. Hantkenina, L. Leupoldina, P. Parasubbotina. Arrows on the right identifyCretaceous oceanic anoxic events (OAEs) of the Mediterranean Tethys (after Coccioni et al. 2006). W Weissert, F Faraoni. Ages of stage boundaries (left) are from Gradstein and Ogg (2004) for the Cretaceous and from Berggrenet al. (1995) for the Cenozoic.
500 HELEN K. COXALL ET AL.
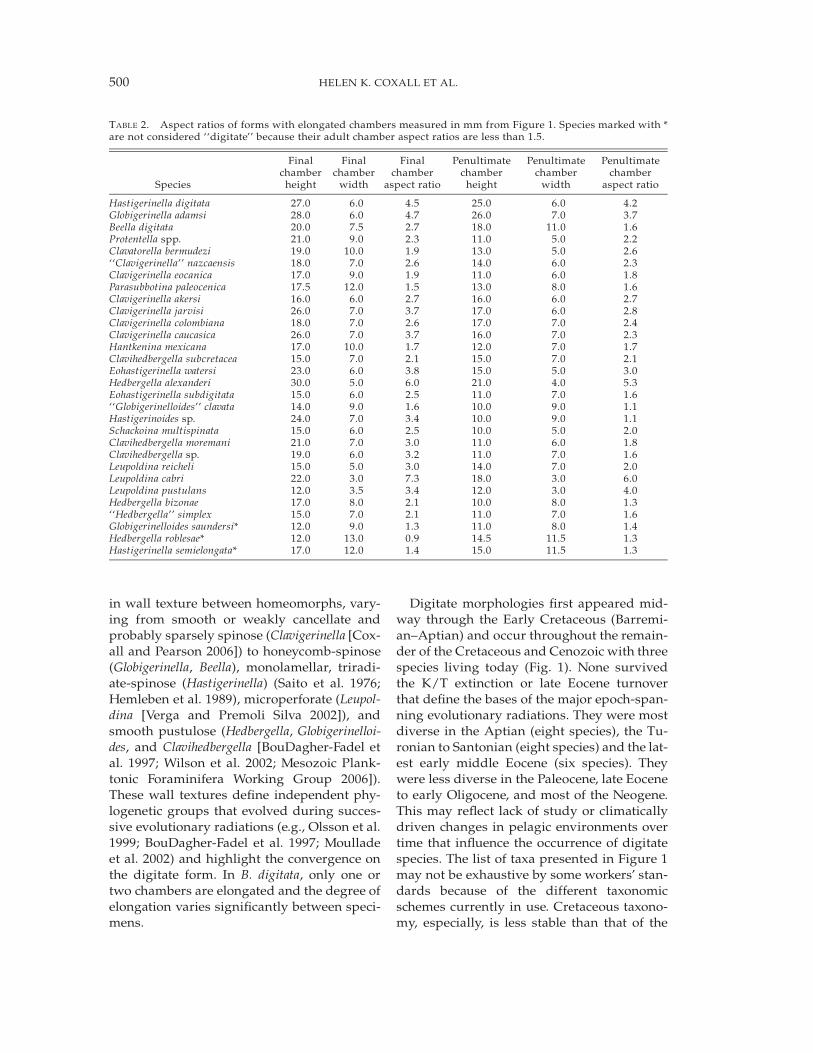
TABLE 2. Aspect ratios of forms with elongated chambers measured in mm from Figure 1. Species marked with *are not considered ‘‘digitate’’ because their adult chamber aspect ratios are less than 1.5.
Species
Finalchamber
height
Finalchamber
width
Finalchamber
aspect ratio
Penultimatechamber
height
Penultimatechamber
width
Penultimatechamber
aspect ratio
Hastigerinella digitata 27.0 6.0 4.5 25.0 6.0 4.2Globigerinella adamsi 28.0 6.0 4.7 26.0 7.0 3.7Beella digitata 20.0 7.5 2.7 18.0 11.0 1.6Protentella spp. 21.0 9.0 2.3 11.0 5.0 2.2Clavatorella bermudezi 19.0 10.0 1.9 13.0 5.0 2.6‘‘Clavigerinella’’ nazcaensis 18.0 7.0 2.6 14.0 6.0 2.3Clavigerinella eocanica 17.0 9.0 1.9 11.0 6.0 1.8Parasubbotina paleocenica 17.5 12.0 1.5 13.0 8.0 1.6Clavigerinella akersi 16.0 6.0 2.7 16.0 6.0 2.7Clavigerinella jarvisi 26.0 7.0 3.7 17.0 6.0 2.8Clavigerinella colombiana 18.0 7.0 2.6 17.0 7.0 2.4Clavigerinella caucasica 26.0 7.0 3.7 16.0 7.0 2.3Hantkenina mexicana 17.0 10.0 1.7 12.0 7.0 1.7Clavihedbergella subcretacea 15.0 7.0 2.1 15.0 7.0 2.1Eohastigerinella watersi 23.0 6.0 3.8 15.0 5.0 3.0Hedbergella alexanderi 30.0 5.0 6.0 21.0 4.0 5.3Eohastigerinella subdigitata 15.0 6.0 2.5 11.0 7.0 1.6‘‘Globigerinelloides’’ clavata 14.0 9.0 1.6 10.0 9.0 1.1Hastigerinoides sp. 24.0 7.0 3.4 10.0 9.0 1.1Schackoina multispinata 15.0 6.0 2.5 10.0 5.0 2.0Clavihedbergella moremani 21.0 7.0 3.0 11.0 6.0 1.8Clavihedbergella sp. 19.0 6.0 3.2 11.0 7.0 1.6Leupoldina reicheli 15.0 5.0 3.0 14.0 7.0 2.0Leupoldina cabri 22.0 3.0 7.3 18.0 3.0 6.0Leupoldina pustulans 12.0 3.5 3.4 12.0 3.0 4.0Hedbergella bizonae 17.0 8.0 2.1 10.0 8.0 1.3‘‘Hedbergella’’ simplex 15.0 7.0 2.1 11.0 7.0 1.6Globigerinelloides saundersi* 12.0 9.0 1.3 11.0 8.0 1.4Hedbergella roblesae* 12.0 13.0 0.9 14.5 11.5 1.3Hastigerinella semielongata* 17.0 12.0 1.4 15.0 11.5 1.3
in wall texture between homeomorphs, vary-ing from smooth or weakly cancellate andprobably sparsely spinose (Clavigerinella [Cox-all and Pearson 2006]) to honeycomb-spinose(Globigerinella, Beella), monolamellar, triradi-ate-spinose (Hastigerinella) (Saito et al. 1976;Hemleben et al. 1989), microperforate (Leupol-dina [Verga and Premoli Silva 2002]), andsmooth pustulose (Hedbergella, Globigerinelloi-des, and Clavihedbergella [BouDagher-Fadel etal. 1997; Wilson et al. 2002; Mesozoic Plank-tonic Foraminifera Working Group 2006]).These wall textures define independent phy-logenetic groups that evolved during succes-sive evolutionary radiations (e.g., Olsson et al.1999; BouDagher-Fadel et al. 1997; Moulladeet al. 2002) and highlight the convergence onthe digitate form. In B. digitata, only one ortwo chambers are elongated and the degree ofelongation varies significantly between speci-mens.
Digitate morphologies first appeared mid-way through the Early Cretaceous (Barremi-an–Aptian) and occur throughout the remain-der of the Cretaceous and Cenozoic with threespecies living today (Fig. 1). None survivedthe K/T extinction or late Eocene turnoverthat define the bases of the major epoch-span-ning evolutionary radiations. They were mostdiverse in the Aptian (eight species), the Tu-ronian to Santonian (eight species) and the lat-est early middle Eocene (six species). Theywere less diverse in the Paleocene, late Eoceneto early Oligocene, and most of the Neogene.This may reflect lack of study or climaticallydriven changes in pelagic environments overtime that influence the occurrence of digitatespecies. The list of taxa presented in Figure 1may not be exhaustive by some workers’ stan-dards because of the different taxonomicschemes currently in use. Cretaceous taxono-my, especially, is less stable than that of the

501DIGITATE FORAM ITERATIVE EVOLUTION
Cenozoic (see BouDagher-Fadel et al. 1997;Moullade 2002) but most workers accept thegeneric classification of Loeblich and Tappan(1988), separating planispiral clavate (Eohas-tigerinella), planispiral tapering (Hastigerinoi-des), and trochospiral clavate (Clavihedbergella)(B. Huber personal communication 2007), andFigure 1 has been constructed to reflect thisscheme. Despite differences in taxonomicopinion, we believe that the list is largely rep-resentative of the generic and specific diver-sity across epochs and should therefore cap-ture the essence of the stratigraphic distribu-tion of taxa exhibiting digitate morphologies.Moreover, our definition of digitate form (i.e.,elongate chamber(s) with an aspect ratiogreater than 1.5) will exclude some taxa thatother workers may consider digitate (e.g., Hed-bergella bollii, Claviblowiella sigali) (Table 2).There are also other genera, e.g., Protentella,that may meet our criteria for being digitatebut whose specific diversity is still unre-solved.
Ecology of Living Digitate Species. There arethree digitate species living today, Globigeri-nella adamsi (Banner and Blow 1959), Beella dig-itata (Brady 1879), and Hastigerinella digitata(Rhumbler 1911). None have been observedalive in the laboratory and only limited infor-mation is available on their ecology and en-vironmental preferences. All three species arerestricted to the low and mid latitudes (Table3) and are typically rare in open-ocean water-masses and underlying sediments. Beella dig-itata is more common than G. adamsi and H.digitata and occurs throughout the Atlanticand Mediterranean (Gross 2001; Hemleben etal. 1989), whereas the latter two species arethought to be mostly restricted to the Indianand Pacific Oceans (Bradshaw 1959; Be andTolderlund 1971). Because of its delicate mon-olamellar shell wall, H. digitata has an ex-tremely low preservation potential and is rare-ly seen in sediments (Hilbrecht 1996). Thereare several records, however, of modern digi-tate species occurring at unusually high abun-dance levels (5–10% of assemblage), e.g.,abundant G. adamsi in the South Pacific region(Brady 1884), west-central Equatorial Pacific,and off Japan (Bradshaw 1959). In addition,numerous H. digitata have been recorded on
video film by a remotely operated submers-ible vehicle (ROV) in Monterey Bay, California(S. Haddock, MBARI, unpublished), a regionthat is known for the strong seasonal upwell-ing and pronounced oxygen minimum con-ditions. None of these unusual occurrences,however, have been systematically studiedand presently they provide little informationabout the environmental preferences of mod-ern digitate species.
Rare observations of digitate species inplankton studies reveal deep habitats, usuallybelow 200 m; in over 70% of observations dig-itates occurred in nets below 150 m, and some-times as deep as 2000 m (Table 3). The Mon-terey Bay ROV observations also support adeep habitat for H. digitata, with concentra-tions observed at depths of approximately 200m, i.e., below the base of the seasonal ther-mocline (S. Haddock personal communication2002). Examination of ingested material inrare captured specimens suggests that H. dig-itata, like its close relative H. pelagica, is strictlycarnivorous (Rhumbler 1911; C. Hemlebenpersonal communication 2005), whereas B.digitata and G. adamsi are thought to be om-nivorous (C. Hemleben personal communica-tion 2005). Symbionts have not been observedin H. digitata or B. digitata (M. Kucera personalcommunication 2007). No information is avail-able on symbiotic associations of G. adamsi.
Although usually restricted to the low andmid latitudes, B. digitata has been reported inPleistocene sediments from the Rockall Chan-nel, 60�N (northeast Atlantic) (Holmes1984), together with a second species of Beella,B. megastoma Earland. However, the specimensof B. digitata illustrated by Holmes (1984: p.102, Pl. 1, Figs. 6, 7) do not possess the radiallyelongate chambers characteristic of B. digitatasensu stricto (i.e., with adult-chamber aspectratios of 1.5 or greater), suggesting that thesehigh-latitude forms represent morpho- andecotypes different from the extreme digitateforms found at lower latitudes. Interestingly,abundance spikes of B. megastoma in the Nor-wegian-Greenland Sea have been correlatedwith meltwater pulses associated with latePleistocene glacial terminations (Bauch 1994).Although these northerly occurrences of Beellaspp. are clearly of paleoceanographic interest,

502 HELEN K. COXALL ET AL.
TA
BL
E3.
Dep
thoc
curr
ence
ofm
oder
nd
igit
ate
pla
nk
ton
icfo
ram
inif
era
from
pla
nk
ton
tow
s.R
ecor
ds
are
from
the
lite
ratu
rean
du
np
ubl
ish
edin
form
atio
n.
Site
Lat
.L
ong
.
Pla
nk
ton
tow
bot
tom
dep
th(m
)
Pla
nk
ton
tow
top
dep
th(m
)Sp
ecie
sSo
urc
e
Equ
ator
ial
Pac
ific
??
5023
Bee
lla
digi
tata
Wat
kin
s20
03A
rabi
anSe
a,N
IOP-
310-
5-3
52.7
3 �N
16.0
2�E
4923
Bee
lla
digi
tata
Pee
ters
&B
rum
mer
2002
Ara
bian
Sea,
NIO
P-31
0-5-
152
.73 �
N16
.02�
E10
074
Bee
lla
digi
tata
Pee
ters
&B
rum
mer
2002
Ara
bian
Sea,
NIO
P-31
0-6-
452
.73 �
N16
.02�
E14
898
Bee
lla
digi
tata
Pee
ters
&B
rum
mer
2002
Ara
bian
Sea,
NIO
P-31
0-6-
352
.73 �
N16
.02�
E20
014
8B
eell
adi
gita
taP
eete
rs&
Bru
mm
er20
02A
rabi
anSe
a,N
IOP-
310-
6-1
52.7
3 �N
16.0
2�E
9849
Bee
lla
digi
tata
Pee
ters
&B
rum
mer
2002
Ara
bian
Sea,
NIO
P-91
7-1-
352
.92 �
N15
.89�
E20
315
3B
eell
adi
gita
taP
eete
rs&
Bru
mm
er20
02A
rabi
anSe
a,N
IOP-
917-
1-2
52.9
2 �N
15.8
9�E
302
203
Bee
lla
digi
tata
Pee
ters
&B
rum
mer
2002
Ara
bian
Sea,
NIO
P-92
0-1-
252
.64 �
N16
.08�
E30
320
2B
eell
adi
gita
taP
eete
rs&
Bru
mm
er20
02A
rabi
anSe
a,N
IOP-
920-
1-1
52.6
4 �N
16.0
8�E
501
303
Bee
lla
digi
tata
Pee
ters
&B
rum
mer
2002
Med
iter
ran
ean
??
200
100
Bee
lla
digi
tata
M.
Ku
cera
per
s.co
mm
.20
06M
edit
erra
nea
n?
?30
020
0B
eell
adi
gita
taM
.K
uce
rap
ers.
com
m.
2006
Med
iter
ran
ean
??
500
300
Bee
lla
digi
tata
M.
Ku
cera
per
s.co
mm
.20
06M
edit
erra
nea
n?
?70
050
0B
eell
adi
gita
taM
.K
uce
rap
ers.
com
m.
2006
N.W
.A
rabi
anSe
aSi
te30
8-1-
352
.5�N
16.1
4�E
7248
Glo
bige
rine
lla
adam
siP
eete
rs&
Bru
mm
er20
02N
.W.
Ara
bian
Sea
Site
922-
2-2
52.5
2 �N
16.1
7�E
7752
Glo
bige
rine
lla
adam
siP
eete
rs&
Bru
mm
er20
02E
.In
dia
nO
cean
05�5
8�S
90�0
0�E
200
—G
lobi
geri
nell
aad
amsi
Be
1977
US
Fis
h&
Wil
dli
feSe
rvic
e,P
acifi
cO
cean
icFi
sher
ies
Inve
st.S
tati
onD
33�0
2�N
118�
23�W
400
—G
lobi
geri
nell
aad
amsi
Bra
dsh
aw19
59
Ch
alle
ng
erSt
atio
n,K
iIs
lan
ds,
E.
Arc
hip
elag
o,P
acifi
c(n
otG
.di
gita
-ta
asst
ated
)
20
�N
168�
E10
61(5
80fa
thom
s)—
Glo
bige
rine
lla
adam
siB
rad
y18
84
Pac
ific
-Bio
traw
lV
18SB
T11
612
�13�
S16
0�58
�E21
3821
77G
lobi
geri
nell
aad
amsi
Sait
oet
al.1
976
Atl
anti
c13
�31�
N18
�03�
W10
10H
asti
geri
nell
adi
gita
taB
e19
77M
onte
rey
Bay
36.5
�N12
2.2�
W
200
—H
asti
geri
nell
adi
gita
taS.
Had
doc
kp
ers.
com
m.
Med
iter
ran
ean
??
200
100
Has
tige
rine
lla
digi
tata
M.
Ku
cera
per
s.co
mm
.20
06M
edit
erra
nea
n?
?30
020
0H
asti
geri
nell
adi
gita
taM
.K
uce
rap
ers.
com
m.
2006
Ind
ian
Oce
an04
�07�
N80
�44�
E36
0—
Has
tige
rine
lla
digi
tata
Be
1977
US
Fis
h&
Wil
dli
feSe
rvic
e,P
acifi
cO
cean
icF
ish
erie
sIn
vest
.St
atio
nD
33�0
2�N
118�
23�W
400
—H
asti
geri
nell
adi
gita
taB
rad
shaw
1959

503DIGITATE FORAM ITERATIVE EVOLUTION
TA
BL
E3.
Con
tin
ued
.
Site
Lat
.L
ong
.
Pla
nk
ton
tow
bot
tom
dep
th(m
)
Pla
nk
ton
tow
top
dep
th(m
)Sp
ecie
sSo
urc
e
Med
iter
ran
ean
??
500
300
Has
tige
rine
lla
digi
tata
M.
Ku
cera
per
s.co
mm
.20
06M
edit
erra
nea
n?
?70
050
0H
asti
geri
nell
adi
gita
taM
.K
uce
rap
ers.
com
m.
2006
‘‘H
um
bol
dt
Pla
nk
ton
Exp
edit
ion’
’?
?80
060
0H
asti
geri
nell
adi
gita
taR
hu
mbl
er19
11A
tlan
tic
05�0
8�N
42�2
4�W
800
—H
asti
geri
nell
adi
gita
taB
e19
77‘‘
Hu
mb
old
tP
lan
kto
nE
xped
itio
n’’
??
2000
—H
asti
geri
nell
adi
gita
taR
hu
mbl
er19
11B
eeb
eSt
atio
n12
00,
N.A
tlan
tic
??
1097
(600
fath
oms)
1097
Has
tige
rine
lla
digi
tata
Ban
ner
&B
low
1960
Pac
ific:
‘‘O
ffSa
nD
iego
’’d
eep
pla
nk
ton
hau
l32
�N11
7�W
‘‘D
eep
’’—
Has
tige
rine
lla
digi
tata
Bra
dsh
aw19
59
Dee
pto
w,j
un
ctio
nof
Ku
rosh
ioan
dO
yash
ioC
urr
ents
30
�N
135�
E‘‘
Dee
p’’
—H
asti
geri
nell
adi
gita
taB
rad
shaw
1959
we suggest they are unrelated to the distri-butional patterns of B. digitata s.s. and do notconsider them further in this paper.
This collection of observations providesonly limited insights into the ecology and dis-tribution of modern digitate species. However,the available evidence suggests a consistentlydeep dwelling habitat (�200 m) below thethermocline, for all living digitate species dur-ing some stage of the life cycle.
Stable Isotope Paleoecologies. Figure 2 shows�18O and �13C values of fossil digitate speciescompared to other planktonic and benthicspecies (unadjusted) in the same sample fromsix time slices at seven different deep-sea sites.Interspecies offsets are assumed to reflectoriginal differences in depth habitat, with thedeep dwellers registering higher �18O andlower �13C than surface dwellers. The differ-ences in absolute and relative �18O reflect re-gional differences in surface-water (plankton-ics) and deep-sea (benthics) temperature, de-gree of thermal stratification, and depth of thethermocline (Berger et al. 1978; Arthur andNatland 1979; Bralower et al. 1995). Minor dia-genetic alteration may also contribute to in-creased absolute �18O values and an artificiallycompressed multispecies gradient in materialfrom ODP Sites 865 and 872 and Kane-9 PistonCore. �13C differences reflect patterns of globalcarbon storage, local productivity, and deep-water circulation.
Despite site-to-site isotopic differences, therelative depth ranking of digitate species an-alyzed is very similar in each time slice. Cre-taceous, Eocene, Miocene, Pleistocene, andHolocene digitate forms consistently registerthe highest or close to the highest �18O values(implying the coolest calcification tempera-tures) and usually the lowest �13C of all plank-tonic foraminifera. This finding is consistentwith limited isotopic data from previous in-vestigations into Eocene and Miocene digitatespecies (Pearson et al. 1993; Coxall et al. 2000,2003 on Clavigerinella; Pearson and Shackleton1995 on Clavatorella bermudezi) that suggest adeep-dwelling habitat.
Distribution of Eocene Digitate Forms. Themiddle Eocene was a time of increased digi-tate diversity, with up to six species of Clavi-gerinella and several species of Hantkenina with

504 HELEN K. COXALL ET AL.
FIGURE 2. Multispecies stable isotope arrays from six time slices (seven sites) showing the relative isotopic depth-ranking of digitate homeomorphs (black and gray diamonds) compared with co-occurring planktonic and benthicspecies (where possible). A, DSDP Site 137-12-5, 4–6 cm. B, DSDP 144A-6R-1, 141–142.5 cm, data from Wilson etal. (2002). C, ODP 865C-8H-3, 70–110 cm. D, Kane-9 Core-42, 470 cm. E. ODP 872C-9H-3, 95–97 cm. F, Box ERDC-92-5, 22–23 cm, data from Berger et al. (1978) except G. adamsi, which are new. Note: Variation in �18O and �13Cbetween sites reflect inter-basin and temporal differences in seawater isotopic composition. Generic abbreviations:A. Acarinina,* B. Beella, C. Clavihedbergella, Cb. Cibicidoides, Cl. Clavigerinella, Clt. Clavatorella, G. Globigerinella, Gd. Globigerinoides,* Gl. Globorotalia, Glm. Globanomalina, Glq. Globoquadrina, Gn. Globi-gerinelloides, Gu. Guembelitrioides, H. Hedbergella,* Het. Heterohelix,* M. Morozovella,* N. Neogloboquadrina,P. Parasubbutina, Pr. Praeglobotruncana, Pu. Pulleniatina, R Rotalipora, S Subbotina. *Inferred or known surfacedwellers (Wilson et al. 2002; Boersma et al. 1987; Pearson 1998; Berger et al. 1978; Hemleben et al. 1989). Data arepresented in Appendix 1.

505DIGITATE FORAM ITERATIVE EVOLUTION
FIGURE 3. Distribution of Clavigerinella spp. during the middle Eocene (46 Ma base map). The map was constructedusing the method outlined by Ziegler et al. (1985). Stippled fill indicates records where Clavigerinella spp. were foundat unusually high abundance levels (�10% of assemblage). The sediments in other Clavigerinella localities are fo-raminiferal oozes with higher species diversity more typical of the low latitudes. Questions marks identify recordswhere other microfossil and host sediment characteristics are unknown. Clavigerinella-rich samples typically containonly a few other species of planktonic foraminifera (e.g., Parasubbotina spp.) but are usually rich in radiolaria. SeeTable 4 for key to localities and data sources.
elongate chambers recognized. This time in-terval is relatively well represented in deep-sea and land sections and we have been ableto produce a paleogeographic reconstructionof Clavigerinella spp. in order to explore thepossible environmental controls on its distri-bution. The reconstruction was produced us-ing 29 records compiled from new observa-tions and the literature (Fig. 3). Paleo-coordi-nates of the Eocene localities are presented inTable 4. The reconstruction shows that Clavi-gerinella spp., comprising C. eocanica, C. akersi,C. jarvisi, C. caucasica, and C. colombiana, is re-stricted to the mid and low latitudes. Recordsare concentrated along segments of continen-tal margin (mostly western boundaries) andthe equatorial Pacific, with fewer occurrencesin fully open ocean regions. In 12 localitiesClavigerinella sp. was recorded as being unusu-ally abundant (�10% planktonic assemblage)and occurred in association with radiolarian-rich sediments, suggesting high surface-waternutrient availability. Today these regions arecharacterized by high productivity due to up-welling; an atmospheric general circulationmodel suggests similar high-productivityconditions prevailed during Eocene time(Huber and Sloan 2000).
Discussion
Depth Ecology. Our �18O and �13C data sug-gest that all the fossil digitate species investi-gated calcified their shells in a relatively coolwatermass with high DIC content, as occursbelow the surface mixed layer. The occurrenceof modern digitates in deep-towed planktonnets supports this interpretation. The deepchlorophyll maximum, situated within or justbelow the thermocline, represents a possiblehabitat for digitate species that provides foodat depth. Deep chlorophyll maxima, however,are widespread features in much of theworld’s oceans and should support largerpopulations of digitates than are observed.Therefore, we suggest that digitate specieshave more-specialized deep water ecologiesdefined by physio-biological structures unfa-vorable to most planktonic foraminifera spe-cies. The typical scarcity of digitates in deep-sea records is consistent with this hypothesisbecause fewer pelagic organisms live belowthe mixed layer outside of chlorophyll maxi-ma, and even fewer in the mesopelagic zone(200–2000 m), where food is usually scarcecompared to the epipelagic zone (0–200 m)(Gage and Tyler 1991; Wishner et al. 1995;Gowing and Wishner 1998).

506 HELEN K. COXALL ET AL.
TABLE 4. Reconstructed middle Eocene coordinates of the Clavigerinella-localities mapped in Figure 3.
ID Locality Palaeolat.Palaeo-
long. Source
1 ODP Site 865, Alison Guyot 4 �143 This study2 ODP Site 1218, Equatorial Pacific 0 �107 Lyle et al. 20023 DSDP Site 42, Equatorial Pacific �3 �106 McManus et al. 19704 Oregon Southern Coast Range, Elkton/upper Tyee Fm. 49 �98 McKeel & Lipps 19755 Oregon Coast Range Tyee, Yamhill & Nestucca Fms. 42 �104 McKeel & Lipps 19726 Mount Diablo, California 35 �104 Church 1931; Clark & Camp-
bell 19427 Santa Cruz Mts., California, San Lorenzo Fm. 20 �94 Poore & Brabb 19778 Veracruz, Mexico, Aragon Fm. 20 �76 Nuttall 19309 Arroyo San Carlos, Carmen de Bolivar Pste., Colom-
bia, Chengue Fm.0 �74 This study
10 Guayaquil, Ecuador �13 �64 Stainforth et al. 194811 San Cristobal, Peru, Talara & Sechura Fms. �13 �66 Cruzado Castaneda 198512 Parinas, Peru, Talara & Sechura Fms. �13 �68 Cruzado Castaneda 198513 El Alto, Lobitos, Peru �15 �65 Cruzado Castaneda 198514 ODP Site 683, Peru Continental margin �18 �64 Suess et al. 198815 Basal Salina, Peru, Chacra & Talara Fms. �21 �61 Cruzado Castaneda 198516 DSDP Site 390, Blake Nose 24 �61 Benson et al. 197817 DSDP Site 150, Aruba Gap, Venezuela 3 �60 Premoli Silva & Bolli 197318 Dept. of Bolivar, Colombia �1 �60 Petters 195419 San Fernando, Trinidad 4 �49 Bolli 195720 ODP Site 1258, Demerara Rise �1 �40 Erbacher et al. 200421 Kane-9 Piston Core, Endeavor Seamount 21 �19 This study22 DSDP Site 21, Rio Grande Rise �31 �9 Saito et al. 197623 DSDP Site 523, S. Atlantic �33 �2 Hsu et al. 198424 ODP Site 960, Cote d’lvoire �1 �3 Mascle et al. 199625 Alicante, S. Spain 31 3 Cremedes Campos 197826 Mattsee, Austria, Helveticum Section 47 8 Gorbandt 1967; this study27 Northern Caucasus Mts., Russia 48 36 Subbotina 195828 Kilwa, Masoko, Tanzania �12 40 Pearson et al. 200429 Khasi Hills, Assam, S. India 7 91 Samanta 1973
Upwelling and Oxygen Minimum Conditions.Increased abundance and diversity of digitatespecies in the Cretaceous and middle Eocene,and in restricted regions today, indicate thatthe digitate ecologic opportunity can be ex-panded at times. Examination of these concen-trations of digitates should provide further in-sight into their ecology.
Cretaceous digitate species occurred inwarm epicontinental sea localities as well as inthe deep sea (e.g., Eicher and Worstell 1970;Longoria 1974; Masters 1977; Premoli Silva etal. 1999) (Fig. 1). Unusually diverse assem-blages have been found in association with or-ganic-rich sediments preserved on continentalmargins that are the hallmark of Cretaceousoceanic anoxic events (OAEs) (e.g., Magniez-Jannin 1998; Aguado et al. 1999; Cobianchi etal. 1999; Premoli Silva et al. 1999; Luciani et al.2001; Leckie et al. 2002; Verga and Premoli Sil-va 2002; Coccioni and Luciani 2004, 2005; Coc-cioni et al. 2006). OAEs involved severe per-
turbations to the marine carbon system andpelagic ecosystems and in some cases were ac-companied by extinction of many surface-liv-ing planktonic foraminifera (Leckie et al. 2002;Coccioni et al. 2006). Their cause is widely de-bated. One theory is that tectonic events andwidespread transgressions led to stagnationof deep waters and creation of a large numberof salinity-stratified marginal basins (Erbach-er et al. 2001). Another is that bottom-waterand water-column dysoxia resulted from in-tensified surface productivity related to activesubmarine volcanism (e.g., Schlanger and Jen-kyns 1976; Premoli Silva et al. 1999; Leckie etal. 2002; Jenkyns 2003). The most widespreadOAEs—the Faraoni, OA1a, and OA2—arethought to have involved extensive eutrophi-cation (Leckie et al. 2002; Coccioni et al. 2006).These are also the times when digitate specieswere most abundant and diverse (Coccioni etal. 2006). The association has led to the sug-gestion that digitate species were specialists of

507DIGITATE FORAM ITERATIVE EVOLUTION
low-oxygen environments (e.g., BouDagher-Fadel et al. 1997; Magniez-Jannin 1998; Agua-do et al. 1999; Cobianchi et al. 1999; PremoliSilva et al. 1999; Luciani et al. 2001; Coccioniand Luciani 2004, 2005; Coccioni et al. 2006).
Digitates were also unusually diverse in themiddle Eocene (six species). There were nosedimentary equivalents to the CretaceousOAEs during this time but Clavigerinella spp.occurrence shows a correlation with increasedmarine productivity in regions of coastal andequatorial upwelling (Fig. 3). There is no di-rect evidence for intensified oxygen-minimumconditions preserved in these regions (i.e., inthe form of increased sedimentary organiccarbon), but pronounced OMZs may havebeen features of these environments, as theyare in modern upwelling regions (e.g., Wish-ner et al. 1995), even if there is no record insediments. It seems logical that usually raredeep-dwelling digitate species would prolif-erate during times of high productivity be-cause of increased food sinking out of the sur-face layer. However, surface productivity can-not be the principal control on digitate distri-bution because, besides the Cretaceous,middle Eocene, and Monterey Bay occurrenc-es, digitate species have not been reportedwidely in typical modern or early Neogeneupwelling systems (e.g., Arabian Sea and Ben-guela Current) (Prell and Curry 1981; Sum-merhayes et al. 1992; Little et al. 1997; Reichartet al. 1998). Therefore, specialization to OMZconditions, which may be a consequence of in-creased surface productivity, and whichwould be unfavorable to most planktonic spe-cies, seems plausible. A closer look at the anat-omy of OMZs reveals the hostilities faced bypelagic organisms and provides clues to theirpotential attraction.
OMZs are ubiquitous and persistent fea-tures of midwater depths in much of theworld’s ocean but they may become expandedthrough heightened marine productivity orreduced water-column ventilation (Wishner etal. 1995). Although plankton biomass and di-versity are dramatically reduced withinOMZs because of the limitations on aerobicrespiration, plankton may become abundantat the ‘‘redoxcline,’’ the level at the base of theOMZ (400 to 1100 m) where oxygen levels be-
gin to increase. High concentrations of organicparticles and bacteria at the redoxcline pro-vide an abundant food source to deep-dwell-ing plankton (Wishner and Gowing 1992;Wishner et al. 1995; Gowing and Wishner1998). Although no records of planktonic fo-raminifera have been obtained so far, it is pos-sible that deep-dwelling species, such as thedigitates, can survive in these environments,feeding directly on bacteria and organic par-ticles or preying on the primary consumers ofmicrobes, e.g., copepods. This hypothesis isconsistent with distribution patterns of digi-tates associated with Cretaceous OAEs; digi-tate species were absent from the core of theOAEs where minimum oxygen levels wouldhave prevented aerobic respiration, but theydiversified during the recovery phase (Coc-cioni et al. 2006), perhaps as oxygen levels be-gan to increase but food availability remainedhigh. Digitates may have been able to respondquickly to this expanding deep-water ecolog-ical opportunity because of their existing tol-erance of hostile deep-water conditions.
The unusually low shell �13C values seen insome digitate species, e.g., Clavigerinella spp.G. adamsi, and B. digitata, which are alwayslower than in other planktonic species andsometimes lower than in co-occurring benth-ics, are consistent with this hypothesis be-cause water-column �13C values reach a mini-mum within the OMZ (Fig. 4). However, giventhat foraminiferal shell carbon isotope valuesusually reflect a mixed signal of seawater �13Cand physiological fractionation effects (e.g.,Zeebe 1999; Ortiz et al. 1996; Norris 1998; Spe-ro 1998), it is possible that the depleted �13Cvalues are the result of a vital effect. One wayof enriching 12C is incorporation of metabolicCO2 into test calcite, although this effect haspreviously been recorded only in surfaceforms such as Globigerina bulloides and Globi-gerinella siphonifera, which tend to have unusu-ally high feeding rates (e.g., Spero and Lea1996; Bijma et al. 1998). Shell calcite isotopicdepletion might also be achieved by feedingon an isotopically ‘‘light’’ food source, e.g.,bacteria; it is possible that, as has been dem-onstrated for other mesopelagic microplank-ton (Wishner et al. 1995; Gowing and Wishner1998), a large part of the forams’ diet at these

508 HELEN K. COXALL ET AL.
FIGURE 4. A, Typical vertical profile of seawater dis-solved inorganic carbon (�CO2), �13C, and dissolved O2
from the North Pacific (Kroopnick 1985) to aid interpre-tation of carbon isotopes. OMZ, oxygen minimum zone.
depths was derived from bacteria-dependentfood chains, whose biosynthetic pathways re-sult in 13C-depleted organic matter (e.g.,Hayes 2001). Because it is difficult to imaginethat 13C depletion through physiological frac-tionation effects would occur to a similar ex-tent in multiple independently evolved digi-tate homeomorphs, we suggest that the �13C isprimarily an environmental signal.
These various strands of evidence support aconnection between digitate species and oxy-gen minimum zones but this cannot be the fullstory because not all OMZ environments areassociated with digitate species. A possibleexplanation for why H. digitata is common inMonterey Bay but not other upwelling regionsis that the low-oxygen watermass off Califor-nia and the eastern Pacific is more or less per-manent, whereas the low-oxygen conditionsin the Arabian Sea and Benguela systems arehighly seasonal. If digitate species were slowin maturing, low population densities mighthave prevented their persistence where theOMZ is seasonal. The lack of evidence for dig-itate evolution during episodes of increasedorganic-matter accumulation associated withPlio-Pleistocene sapropels (e.g., Rohling 1994;
Rohling et al. 2004) may reflect differences inthe causal mechanisms of the oceanic dysoxiaand its effect on deep-dwelling plankton as-sociated with the late Neogene events as com-pared to the Cretaceous OAEs.
Data are lacking for the Oligocene and Mio-cene but available records suggest that digi-tate species were less common from the lateEocene onward and they have never been asabundant or diverse as they were during theearly Aptian, Albian, and late Cenomanian(Coccioni et al. 2006). This may be because pe-riods of sustained and expanded OMZ con-ditions, which occurred more frequently un-der greenhouse climates because of reducedwater-column stratification and increased vol-canic input of biologically limiting nutrients(Leckie et al. 2002), became less frequent ascrustal production slowed and thermal anddensity stratification of the surface ocean in-creased under the evolving icehouse climatesystem.
Functional Significance of Digitate Chambers.Our results suggest that digitate morpholo-gies were maintained in planktonic foraminif-era populations exposed to similar environ-mental constraints. One possibility is thatelongation of individual chambers is efficientfor survival in persistently low-oxygen envi-ronments by increasing shell surface area forimproved gas transport, as has been proposedfor Cretaceous species of the OAEs (e.g.,BouDagher-Fadel et al. 1997; Magniez-Jannin1998; Aguado et al. 1999; Cobianchi et al. 1999;Luciani et al. 2001; Coccioni and Luciani 2004,2005; Coccioni et al. 2006). The problem withthis hypothesis is that there is almost no ex-perimental evidence to support the idea thatgas exchange occurs across the shell surface,aside from the observation that pores are as-sociated with concentrations of mitochondria,which in turn are associated with gas trans-port. In addition, if gas transport is the issue,flattened discoidal shells might be expected toprovide a geometrically more efficient bodyplan. Moreover, not all digitate species showassociations with low-oxygen environments.Even among the Cretaceous species associatedwith OAEs, digitates are absent from the coreof the events and diversify in parallel withsome non-digitate species, suggesting that ad-

509DIGITATE FORAM ITERATIVE EVOLUTION
ditional factors control their distribution be-sides oxygen availability (Coccioni et al. 2006).
A more likely explanation is that radialchamber elongation was efficient for feedingin a subsurface habitat where the food supplywas sparse and/or irregular. Elongate cham-bers may function as ‘‘fishing rods,’’ support-ing long food-gathering rhizopods (strands ofcytoplasm) that increase the effective shellsize and volume of water that could besearched for food at minimum metabolic cost(R. D. Norris personal communication 2001).Radial elongation allows digitate species to at-tain unusually large shell sizes; the sizes ofClavigerinella spp. and Clavatorella bermudezi,commonly 500 mm, and of H. digitata, up to2 mm, are consistent with this hypothesis. Ef-fective shell size is further increased in somedigitate species by the presence of fine calciticspines; e.g., H. digitata has spines emergingfrom the tips of individual chambers, creatinga radiating net that more than triples the ra-dius. The purpose of these spines is very likelyto provide additional support for rhizopodsand to help secure large prey items. Beella dig-itata and G. adamsi are also spinose and mayhave had similar arrangements of spines, al-though live specimens have not been ob-served. Spines were absent in Cretaceous andpossibly most Paleogene digitate forms. Gasexchange efficiency under low-oxygen condi-tions may also have been improved as a con-sequence of having digitate chambers.
Although digitate species appear to be ex-clusively deep dwelling, digitate form is not aprerequisite for subsurface living becausethere are a number of other species with ob-served or inferred deep dwelling habitats thatdo not have elongated chambers e.g., Paleo-gene and Neogene Globorotaloides spp., includ-ing living G. hexagona (Ortiz et al. 1996), andPaleogene Parasubbotina spp. (Olsson et al.2006b). However, the evolution of some digi-tate species may have occurred within a sub-surface habitat in response to changing phys-ical chemical and nutrient structures. This hy-pothesis is consistent with phylogenetic re-constructions that suggest that acquisition ofdigitate chambers in some groups involvedparapatric speciation, with digitate speciesevolving from existing deep dwellers (e.g., C.
bermudezi from Globorotaloides hexagona (Fleish-er 1974) and C. eocanica from Parasubbotinaeoclava (Coxall et al. 2003)). Moreover, loss ofelongated chambers in the tubulospinose ge-nus Hantkenina was paralleled by a shift froma deep- to a shallow-water habitat (Coxall etal. 2000), although modified extensions of thechambers, in the form of narrow non-perfo-rate tubulospines, were retained in youngersurface-dwelling forms. Modern digitate spe-cies, on the other hand, appear to be moreclosely related to surface and intermediate-depth dwellers (e.g., H. pelagica and Globigeri-nella spp. [Hemleben et al. 1989; M. Kucerapersonal communication 2007]), suggestingthat different selection pressures were respon-sible for evolution of digitate morphologies inthe late Neogene.
The isotopic data and distribution patternspresented here provide an environmental cor-relate with digitate morphologies—life indeep, DIC-enriched, and possibly regularlyoxygen-depleted watermasses associated withincreased marine production. This iterativeevolution very likely reflects common func-tional constraints that could be regarded asrepresenting an adaptive process, eventhough we might not know their exact func-tion or be able to demonstrate the pathway ofspeciation. By this we mean that digitatechambers, arising through natural variation,are efficient for survival under certain envi-ronmental conditions and, therefore, are‘‘maintained’’ or ‘‘selected for’’ when theseconditions occur, while ineffective charactersare eliminated. This is a remarkable findingbecause few other structural convergencesamong planktonic foraminifera have beenfound to correlate with similar ecologies be-tween independently evolved genera andacross multiple epochs.
Summary and Conclusions
Digitate planktonic foraminifera morphol-ogies have evolved multiple times over thepast 130 million years. Previous studies havehinted that some digitate homeomorphs hadsimilar depth habitats, but our stable isotoperesults provide the first evidence of a clearcorrelation between this morphology and asubsurface habitat over multiple epochs.

510 HELEN K. COXALL ET AL.
These findings are consistent with planktontow data that suggest that all modern digitatespecies spend part of their life cycle living atmesopelagic or bathypelagic habitats. Geolog-ical and modern evidence also links some dig-itate species with increased marine productiv-ity and low-oxygen environments, but theyappear to like strong, quasi-permanent OMZconditions such as those that developed on theeastern sides of the major ocean basins and inequatorial upwelling systems. The need forthese persistent OMZ conditions may explainwhy these species do not occur in the temper-ate and high latitudes today but were commonduring Cretaceous OAEs. However, digitatespecies are not always associated with regionsand episodes of eutrophication and low oxy-gen concentration, suggesting there are addi-tional physio-chemical and biological controlson their distribution, such as temperature, sa-linity, nutrients, type of food, and trace ele-ments. We suggest that the function of elon-gate chambers is primarily a feeding adapta-tion that allows the foraminifera to search alarger volume of water for food at minimummetabolic cost in a usually food-poor meso-pelagic environment. There also appears to bean element of opportunism in the ecology ofdigitate species, allowing them to proliferateand diversify under certain environmentalconditions that are unfavorable to many otherplanktonic species. As has been suggested forCretaceous OAE digitates (Coccioni et al.2006), relative abundance of digitate speciesmay be proportional to the strength of the en-vironmental perturbation related to the OAEs.
Our findings suggest that digitate plank-tonic foraminifera may be used as a proxy forintervals of expansion of the OMZ, especiallyduring Mesozoic and early Cenozoic green-house climates when thermal and densitystratification may have been weaker and epi-sodes of widespread eutrophication weremore frequent. Further insight into digitateecologies and environmental preferences canbe obtained by studying the ecology of mod-ern digitate taxa, starting with the systematicassessment of Hastigerinella digitata distribu-tion in Monterey Bay, and building an ex-panded database on the isotopic paleoecolo-
gies and biogeographic distribution of digi-tate species over time.
Acknowledgments
We thank G. Miller and A. Henderson forloan of material from the Natural History Mu-seum London BP collection and S. Haddockfor information on Monterey Bay foraminif-era. Thanks to P. Markwick of Robinson Re-search for mapping Clavigerinella paleogeog-raphies. Thanks also to M. Bolshaw, M. Coo-per, and J. Becker for laboratory assistance, B.Huber, M. Hart, C. Hemleben, N. Norris, andan anonymous reviewer for constructive re-views of earlier manuscripts; and M. Leckiefor discussions on Cretaceous taxonomy. Weare also grateful to I. Hall for providing Ho-locene material, M. Kucera for sharing his wa-ter-column plankton-sampling data, and B.Huber for his data on Cretaceous taxonomythat were incorporated during the review pro-cess. This work was supported by the RoyalCommission for the Exhibition of 1851 UKODP, the Natural Environment ResearchCouncil and the Royal Society. Samples wereprovided by the Ocean Drilling Program (U.S.National Science Foundation under the man-agement of the Joint Oceanographic Institu-tions).
Literature CitedAguado, R., J. M. Castro, M. Company, and G. A. de Gea. 1999.
Aptian bio-events—an integrated biostratigraphic analysis ofthe Almadich Formation, Inner Prebetic Domain, SE Spain.Cretaceous Research 20:663–683.
Arthur, M. A., and J. H. Natland. 1979. Carbonaceous sedimentsin the North and South Atlantic: the role of salinity in stablestratification of Early Cretaceous basins. Pp. 375–401 in M.Talwani, W. Hay, and W. B. F. Ryan, eds. Deep drilling resultsin the Atlantic Ocean: continental margins and paleoenviron-ment. Maurice Ewing Series, Vol. 3. American GeophysicalUnion, Washington, D.C.
Banner, F. T., and W. H. Blow. 1959. The classification and strat-igraphical distribution of the Globigerinaceae. Palaeontology2:1–27.
———. 1960. The taxonomy, morphology and affinities of thegenera included in the subfamily Hastigerininae. Micropale-ontology 6:19–31.
Bauch, H. A. 1994. Beella megastoma (Earland) in late PleistoceneNorwegian-Greenland Sea sediments; stratigraphy and melt-water implication. Journal of Foraminiferal Research 24:171–177.
Be, A. W. H. 1977. An ecological, zoographic and taxonomic re-view of Recent planktonic foraminifera. Pp. 1–100 in A. T. S.Ramsay, ed. Oceanic micropalaeontology. Academic Press,London.
Be, A. W. H., and D. S. Tolderlund. 1971. Distribution and ecol-

511DIGITATE FORAM ITERATIVE EVOLUTION
ogy of living planktonic foraminifera in surface waters of theAtlantic and Indian Oceans. Pp. 105–149 in B. M. Funnel andR. W. Reidel, eds. The micropaleontology of oceans. Cam-bridge University Press, Cambridge.
Benson, W. E., R. E. Sheridan, et al., eds. 1984. Sites 389 and 390.Initial Reports of the Deep Sea Drilling Project 44:69–152. U.S.Government Printing Office, Washington, D.C.
Berger, W. H. 1969. Planktonic foraminifera: basic morphologyand ecological implications. Journal of Paleontology 43:1369–1384.
Berger, W. H., J. S. Killingley, and E. Vincent. 1978. Stable iso-topes in deep-sea carbonates: Box Core ERDC-92, WesternEquatorial Pacific. Oceanologica Acta 1:203–216.
Berggren, W. A., D. V. Kent, I. C. C. Swisher, and M. P. Aubry.1995. A Revised Cenozoic Geochronology and Chronostratig-raphy. Pp. 129–212 in W. A. Berggren, D. V. Kent, and J. Har-denbol, eds. Geochronology, time scales and global strati-graphic correlation: a unified temporal framework for an his-torical geology. Society for Sedimentary Geology Special Pub-lication 54:129–212.
Bijma, J., C. Hemleben, B. T. Huber, H. Erlenkeuser, and D.Kroon. 1998. Experimental determination of the ontogeneticstable isotope variability in two morphotypes of Globigerinellasiphonifera (d’Orbigny). Marine Micropaleontology 35:141–160.
Boersma, A., I. Premoli Silva, and N. J. Shackleton. 1987. Atlan-tic Eocene planktonic foraminiferal paleohydrographic indi-cators and stable isotope paleoceanography. Paleoceanogra-phy 2:287–331.
Bolli, H. M. 1957. Planktonic foraminifera from the Eocene Na-vet and San Fernando formations of Trinidad, B.W.I. U.S. Na-tional Museum Bulletin 215:155–172.
BouDagher-Fadel, M. K., F. T. Banner, and J. E. Whittaker. 1997.The early evolutionary history of planktonic foraminifera.Chapman and Hall, London.
Bradshaw, J. S. 1959. Ecology of living planktonic foraminiferain the North and Equatorial Pacific Ocean. Cushman Foun-dation of Foraminiferal Research Contributions 10:25–64.
Brady, H. B. 1879. Notes on some reticularian Rhizopoda of theChallenger Expedition. II. Additions to the knowledge of por-cellanous and hyaline types. Quarterly Journal of the Micro-scopical Society 19:261–299.
———. 1884. Report on the foraminifera dredge by H.M.S. Chal-lenger, during the years 1873–1876. Report on the ScientificResults of the Voyage of H.M.S. Challenger, Zoology, Vol. 9.Longmans, London.
Bralower, T. J., J. C. Zachos, E. Thomas, M. Parrow, C. K. Paull,D. C. Kelly, I. Premoli Silva, W. V. Sliter, and K. C. Lohman.1995. Late Paleocene to Eocene paleoceanography of theequatorial Pacific Ocean: stable isotopes record at Ocean Dril-ling Program Site 865, Allison Guyot. Paleoceanography 20:391–406.
Caron, M., and P. Homewood. 1983. Evolution of early foramin-ifers. Marine Micropaleontology 7:453–462.
Church, C. C. 1931. Foraminifera of the Kreyenhagen shale. Cal-ifornia Department of Natural Resources, Division of Mines,Report No. 27.
Cicha, I., F. Rogl, C. Rupp, and J. Ctyoka. 1998. Oligocene-Mio-cene foraminifera of the Central Paratethys. Abhandlungender Senckenbergischen Naturforschenden Gesellschaft 549:1–325.
Cifelli, R. 1969. Radiation of Cenozoic foraminifera. SystematicZoology 18:154–168.
Clark, B. L., and A. S. Campbell. 1942. Eocene radiolarian faunasfrom the Mount Diablo area, California. Geological Society ofAmerica Special Paper 39.
Cobianchi, M., V. Luciani, and A. Menegatti. 1999. The Selli Lev-el of the Gargano Promontory, Apulia, southern Italy: fora-
miniferal and calcareous nannofossil data. Cretaceous Re-search 20:255–269.
Coccioni, R., and V. Luciani. 2004. Planktonic foraminifera andenvironmental changes across the Bonarelli Event (OAE2, lat-est Cenomanian) in its type area: a high-resolution studyfrom the Tethyan reference Bottaccione section (Gubbio, Cen-tral Italy). Journal of Foraminiferal Research 34:109–129.
———. 2005. Planktonic foraminifers across the Bonarelli Event(OAE2, latest Cenomanian): the Italian record. Palaeogeog-raphy, Palaeoclimatology, Palaeoecology 224:167–185.
Coccioni, R., A. Marsili, and V. Luciani. 2006. Cretaceous oce-anic anoxic events and radially elongated chambered plank-tonic foraminifera: paleoecological and paleoceanographicimplications. Palaeogeography, Palaeoclimatology, Palaeoe-cology 235:66–92.
Corfield, R. M., and J. E. Cartlidge. 1991. Isotopic evidence forthe depth stratification of fossil and recent Globigerinina: areview. Historical Biology 5:37–63.
Corfield, R. M., M. A. Hall, and M. D. Brasier. 1990. Stable iso-tope evidence for foraminiferal habitats during the Cenoman-ian/Turonian ocean anoxic event. Geology 18:175–178.
Coxall, H. K., and P. N. Pearson. 2006. Taxonomy, biostratigra-phy and phylogeny of Hantkeninidae (Clavigerinella, Hantken-ina and Cribrohantkenina). Pp. 213–252 in Pearson et al. 2006a.
Coxall, H. K., P. N. Pearson, N. J. Shackleton, and M. A. Hall.2000. Hantkeninid depth adaptation: an evolving life strategyin a changing ocean. Geology 28:87–90.
Coxall, H. K., B. T. Huber, and P. N. Pearson. 2003. Origin andmorphology of the Eocene planktonic foraminifera Hantkeni-na. Journal of Foraminiferal Research 33:237–261.
Cremades Campos, J. 1978. Una nueva especie del genero Cla-vigerinella Bolli, Loeblich y Tappan. Cuadernos de Geologıa,Universidad de Granada 8–9:175–179.
Cruzado Castaneda, J. 1985. Foraminiferos planctonicos del no-roeste Peruano. Boletın de la Sociedad Geologica del Peru 74:1–45.
de Klasz, I., S. de Klasz, and J. Ausseil-Badie. 1987. Etude sys-tematique des foraminiferes du Danien de la Formation desMadeleines de Dakar (Senegal). Cahiers de Micropaleonto-logie 2:29–38.
Eicher, D. L., and P. Worstell. 1970. Cenomanian and Turonianforaminifera from the Great Plains, United States. Micropa-leontology 16:269–324.
Erbacher, J., B. T. Huber, R. D. Norris, and M. Markey. 2001. In-creased thermohaline stratification as a possible cause for anocean anoxic event in the Cretaceous Period. Nature 409:325–327.
Erbacher, J., D. C. Mosher, and M. J. Malone, eds. 2004. Site 1258.Proceedings of the Ocean Drilling Program, Initial Reports207. Ocean Drilling Program. College Station, Tex.. Availableonline at http://www-odp.tamu.edu/publications/207�IR/207ir.htm.
Fairbanks, R. G., P. H. Wiebe, and A. W. H. Be. 1980. Verticaldistribution and isotopic composition of living planktonic fo-raminifera in the western North Atlantic. Science 207:61–63.
Fairbanks, R. G., M. Sverdlove, R. Free, P. H. Wiebe, and A. W.H. Be. 1982. Vertical distribution and isotopic fractionation ofliving planktonic foraminifera from the Panama Basin. Na-ture 298:841–844.
Fleisher, R. S. 1974. Cenozoic planktonic foraminifera and bio-stratigraphy, Arabian sea (Deep Sea Drilling Project, leg 23A).Pp. 1001–1071 in R. B. Whitmarsh, D. A. Ross, et al., eds. Ini-tial Reports of Deep-Sea Drilling Project. Government Print-ing Office, Washington, D.C.
Frerichs, W. E. 1971. Evolution of planktonic foraminifera andpaleotemperatures. Journal of Paleontology 45:963–968.
Gage, J. D., and P. A. Tyler. 1991. Deep-sea biology: a natural

512 HELEN K. COXALL ET AL.
history of organisms at the deep-sea floor. Cambridge Uni-versity Press, Cambridge.
Gohrbandt, K. H. A. 1967. Some new planktonic foraminiferalspecies from the Austrian Eocene. Micropaleontology 13:319–326.
Gowing, M. M., and K. F. Wishner. 1998. Feeding ecology of thecopepod Lucicutia aff. L. grandis near the lower interface of theArabian Sea oxygen minimum zone. Deep-Sea Research PartII: Topical Studies in Oceanography 45:2433–2459.
Gradstein, F. M., and J. G. Ogg. 2004. A geologic time scale 2004.Cambridge University Press, Cambridge.
Gross, O. 2001. Foraminifera. Pp. 60–75 in M. J. Costello et al.,eds. European register of marine species: a check-list of themarine species in Europe and a bibliography of guides totheir identification. Collection Patrimoines Naturels 50:60–75.Museum National d’Histoire Naturelle, Paris.
Hart, M. B. 1980. A water depth model for the evolution of theplanktonic Foraminiferida. Nature 286:252–254.
Hayes, J. M. 2001. Fractionation of the isotopes of carbon andhydrogen in biosynthetic processes. In J. W. Valley and D. R.Cole, eds. Stable isotopic geochemistry. Reviews in Mineral-ogy and Geochemistry 43:225–278.
Hemleben, C., M. Spindler, and O. R. Anderson. 1989. Modernplanktonic foraminifera. Springer, New York.
Hilbrecht, H. 1996. Extant planktic foraminifera and the phys-ical environment in the Atlantic and Indian Oceans. Mittei-lungen aus dem Geologischen Institut der Eidgen. Technisch-en Hochschule und der Universitat Zurich, Neue Folge, No.300.
Holmes, N. A. 1984. An emendation of the genera Beella Bannerand Blow 1959, and Turborotalita Banner and Blow, 1962, withnotes on Orcadia Boltovosky and Watanabe, 1982. Journal ofForaminiferal Research 14:101–110.
Houston, R. M., and B. T. Huber. 1998. Evidence of photosym-biosis in fossil taxa? Ontogenetic stable isotope trends insome Late Cretaceous planktonic foraminifera. Marine Mi-cropaleontology 34:29–46.
Hsu, K. J., and J. L. La Breque, eds. 1984. Site 523. Initial Reportsof the Deep Sea Drilling Project 73:271–322. U.S. GovernmentPrinting Office, Washington, D.C.
Huber, M., and L. C. Sloan. 2000. Climatic responses to tropicalsea surface temperature changes on a ‘‘greenhouse’’ Earth.Paleoceanography 15:443–450.
Jenkyns, H. C. 2003. Evidence for rapid climate change in theMesozoic–Palaeogene greenhouse world. PhilosophicalTransactions of the Royal Society of London A 361:1885–1916.
Kennett, J. P., and M. S. Srinivasan. 1983. Neogene planktonicforaminifera: a phylogenetic atlas. Hutchinson Ross, Strouds-burg, Penn.
Kroon, D., and A. J. Nederbragt. 1990. Ecology and paleoecol-ogy of triserial planktic foraminifera. Marine Micropaleon-tology 16:25–38.
Kroopnick, P. M. 1985. The distribution of 13C in the worldoceans. Deep-Sea Research 32:57–84.
Leckie, R. M. 1989. A paleoceanographic model for the earlyevolutionary history of planktonic foraminifera. Palaeogeog-raphy, Palaeoclimatology, Palaeoecology 73:107–138.
Leckie, R. M., T. J. Bralower, and R. Cashman. 2002. Oceanic an-oxic events and plankton evolution: biotic response to tectonicforcing during the mid-Cretaceous. Paleoceanography 17:13–29.
Lipps, J. H. 1979. The ecology and paleoecology of planktic fo-raminifera. Pp. 62–104 in J. H. Lipps, W. H. Berger, M. A. Buz-as, R. G. Douglas, and C. A. Ross, eds. Foraminiferal ecologyand paleoecology. Society of Economic Paleontologists andMineralogists, Houston, Tex.
Little, M. G., R. R. Schneider, D. Kroon, B. Price, T. Bickert, andG. Wefer. 1997. Rapid paleoceanographic changes in the Ben-
guela Upwelling System for the last 160,000 years as indicatedby abundances of planktonic foraminifera. Palaeogeography,Palaeoclimatology, Palaeoecology 130:135–161.
Loeblich, A. R. J., and H. Tappan. 1988. Foraminiferal generaand their classification. Van Nostrand Reinhold, New York.
Longoria, J. F. 1974. Stratigraphic, morphologic and taxonomicstudies of Aptian planktonic foraminifera. Revista Espanolade Micropaleontologıa, numero extraordinario 107.
Luciani, V., M. Cobianchi, and H. C. Jenkyns. 2001. Biotic andgeochemical response to anoxic events: the Aptian pelagicsuccession of the Gargano Promontory (southern Italy). Geo-logical Magazine 138:277–298.
Lyle, M., P. A. Wilson, and T. R. Janecek, eds. 2002. Site 1218.Proceedings of the Ocean Drilling Program, Initial Reports199. Ocean Drilling Program, College Station, Tex.. Availableonline at http://www-odp.tamu.edu/publications/199�IR/199ir.htm.
Magniez-Jannin, F. 1998. L’elongation des loges chez les fora-miniferes planctoniques du Cretace inferieur: une adaptationa la sous-oxygenation des eaux? (Chamber elongation in EarlyCretaceous planktonic foraminifera: an adaptive response tooxygen depleted water?). Comptes Rendus de l’Academie desSciences, ser. II, Sciences de lat Terre et des Planetes 326:207–213.
Mascle, J., G. P. Lohmann, P. D. Clift, et al., eds. 1996. Site 960.Proceedings of the Ocean Drilling Program, Initial Reports159:151–215. Ocean Drilling Program, College Station, Tex.
Masters, B. A. 1977. Mesozoic planktonic foraminifera. Pp. 301–732 in A. T. S. Ramsay, ed. Oceanic micropalaeontology. Ac-ademic Press, London.
McKeel, D. R., and J. L. Lipps. 1972. Calcareous plankton fromthe Tertiary of Oregon. Palaeogeography, Palaeoclimatology,Palaeoecology 12:75–79.
———. 1975. Eocene and Oligocene planktonic foraminiferafrom the central and southern Oregon coast range. Journal ofForaminiferal Research 5:1–5.
McManus, D. A., et al., eds. 1970. Site 42. Initial Reports of theDeep Sea Drilling Project 5:367–410. U.S. Government Print-ing Office, Washington, D.C.
Mesozoic Planktonic Foraminiferal Working Group (B. T. Huber,coordinator). 2006. Mesozoic planktonic foraminiferal taxo-nomic dictionary. www.chronos.org.
Moullade, M., J.-P. Bellier, and G. Tronchetti. 2002. Hierarchy ofcriteria, evolutionary processes and taxonomic simplificationin the classification of Lower Cretaceous planktonic forami-nifera. Cretaceous Research 23:111–148.
Norris, R. 1991. Biased extinction and evolutionary trends. Pa-leobiology 17:388–399.
———. 1996. Symbiosis as an evolutionary innovation in the ra-diation of Paleocene planktic foraminifera. Paleobiology 22:461–480.
———. 1998. Recognition and macroevolutionary significance ofphotosymbiosis in molluscs, corals and foraminifera. Pp. 68–100 in Norris and Corfield 1998.
Norris, R. D., and R. M. Corfield, eds. 1998. Isotope paleobiol-ogy and paleoecology. Palaeontological Society Papers, Vol. 4.
Nuttall, W. L. F. 1930. Eocene foraminifera from Mexico. Journalof Paleontology 4:271–293.
Olsson, R. K., C. Hemleben, W. A. Berggren, and B. T. Huber.1999. Atlas of Paleocene planktonic foraminifera. Smithsoni-an Contributions to Paleobiology 85:252.
Olsson, R. K., C. Hemleben, B. T. Huber, and W. A. Berggren.2006a. Taxonomy, biostratigraphy and phylogeny of Globiger-ina, Globoturborotalita, Subbotina, and Turborotalita. Pp. 111–168 in Pearson et al. 2006a.
Olsson, R., P. N. Pearson, B. T. Huber, and I. Premoli Silva.2006b. Taxonomy, biostratigraphy, and phylogeny of EoceneCatapsydrax, Globorotaloides, Guembelitriodes, Paragloborotalia,

513DIGITATE FORAM ITERATIVE EVOLUTION
Parasubbotina, and Pseudoglobigerinella n.gen. Pp. 67–110 inPearson et al. 2006a.
Ortiz, J. D., A. C. Mix, W. Rugh, J. M. Watkins, and R. W. Collier.1996. Deep-dwelling planktonic foraminifera of the north-eastern Pacific Ocean reveal environmental control of oxygenand carbon isotopic disequilibria. Geochimica et Cosmochim-ica Acta 60:4509–4523.
Pearson, P. N. 1995. Planktonic foraminifera biostratigraphy andthe development of pelagic caps on guyots in the Marshall Is-lands group. In J. A. Haggerty, I. Premoli Silva, F. Rack, andM. K. McNutt, eds. Proceedings of the Ocean Drilling Pro-gram, Scientific Results 144:21–59. Ocean Drilling Program,College Station, Tex..
———. 1996. Cladogenetic, extinction and survivorship patternsfrom a lineage phylogeny: the Paleogene planktonic forami-nifera. Micropaleontology 42:179–188.
———. 1998. Stable isotopes and the study of evolution in plank-tonic foraminifera. Pp. 138–178 in Norris and Corfield 1998.
Pearson, P. N., and N. J. Shackleton. 1995. Neogene multispeciesplanktonic foraminifer stable isotope record, site 871, Lima-lok Guyot. In J. A. Haggerty, I. Premoli Silva, F. Rack, and M.K. McNutt, eds. Proceedings of the Ocean Drilling Program,Scientific Results 144:401–410. Ocean Drilling Program, Col-lege Station, Tex..
Pearson, P. N., N. J. Shackleton, and M. A. Hall. 1993. Stable iso-tope paleoecology of middle Eocene planktonic foraminiferaand multi-species isotope stratigraphy, DSDP Site 523, southAtlantic. Journal of Foraminiferal Research 23:123–140.
Pearson, P. N., P. W. Ditchfield, J. Singano, K. G. Harcourt-Brown, C. J. Nicholas, N. J. Shackleton, and M. A. Hall. 2001.Warm tropical sea surface temperatures in the Late Creta-ceous and Eocene epochs. Nature 413:481–487.
Pearson, P. N., C. J. Nicholas, J. Singano, P. R. Bown, H. K. Cox-all, B. E. van Dongen, B. T. Huber, A. Karega, J. A. Lees, E.Msaky, R. D. Pancost, M. Pearson, and A. P. Roberts. 2004. Pa-leogene and Cretaceous sediment cores from the Kilwa andLindi areas of coastal Tanzania: Tanzania Drilling ProjectSites 1 to 5. Journal of African Earth Sciences 39:25–62.
Pearson, P. N., R. K. Olsson, C. Hemleben, B. T. Huber, and W.A. Berggren, eds. 2006a. Atlas of Eocene planktonic forami-nifera. Cushman Foundation of Foraminiferal Research Spe-cial Publication 41. Allen Press, Lawrence, Kans.
Pearson, P. N. P., V. Premec-Fucek, and I. Premoli Silva. 2006b.Taxonomy, biostratigraphy and phylogeny of Eocene Turbo-rotalia. Pp. 433–460 in Pearson et al. 2006a.
Peeters, F. J. C., and G.-J. A. Brummer. 2002. The seasonal andvertical distribution of living planktic foraminifera in the NWArabian Sea. In P. D. Clift, D. Kroon, C. Gaedicke, and J. Craig,eds. The tectonic and climatic evolution of the Arabian Searegion. Geological Society of London Special Publication 195:463–497.
Peng, T. H., W. S. Broecker, and W. H. Berger. 1979. Rates of ben-thic mixing in deep-sea cores as determined by radioactivetracers. Quaternary Research 11:141–149.
Petters, V. 1954. Tertiary and Upper Cretaceous foraminiferafrom Colombia, South America. Contributions from the Cush-man Foundation of Foraminiferal Research 5:37–41.
Poore, R. Z., and E. E. Brabb. 1977. Eocene and Oligocene plank-tonic foraminifer form the Upper Butano sandstone and typeSan Lorenzo Formation, Santa Cruz Mountains, California.Journal of Foraminiferal Research 7:249–272.
Prell, W. L., and W. B. Curry. 1981. Faunal and isotopic indicesof monsoonal upwelling: Western Arabian Sea. OceanologicaActa 4:91–98.
Premoli Silva, I., and H. M. Bolli. 1973. Late Cretaceous to Eo-cene planktonic foraminifera and stratigraphy of the Leg 15Sites in the Caribbean Sea. In N. T. Edgar and J. B. Saunders,
eds. Initial Reports of the Ocean Drilling Program 15:449–549. U.S. Government Printing Office, Washington, D.C.
Premoli Silva, I., and W. V. Sliter. 1999. Cretaceous paleocean-ography: evidence from planktonic foraminiferal evolution.In E. Barrera and C. C. Johnson, eds. Evolution of the Creta-ceous ocean-climate system. Geological Society of AmericaSpecial Paper 332:301–328.
Premoli Silva, I., E. Erba, G. Salvini, C. Locatelli, and D. Verga.1999. Biotic changes in Cretaceous oceanic anoxic events ofthe Tethys. Journal of Foraminiferal Research 29:352–370.
Quilty, P. G. 1976. Planktonic foraminifera DSDP Leg 34—NazcaPlate. In R. D. Yeats, S. R. Hart, et al., eds. Initial Reports ofthe Deep Ocean Drilling Project 34:650–651. U.S. GovernmentPrinting Office, Washington, D.C.
Reichart, G. J., L. J. Lourens, and W. J. Zachariasse. 1998. Tem-poral variability in the northern Arabian Sea Oxygen Mini-mum Zone (OMZ) during the last 225,000 years. Paleocean-ography 13:607–621.
Rhumbler, L. 1911. Die Foraminiferen (Thalamophoren) derPlankton-Expedition, Teil 1. Die allgemeinen Organisations-verhaltnisse der Foraminiferen. Plankton Expedition Hum-bold-Stiftung, Ergeben, Vol. 3.
Rohling, E. J. 1994. Review and new aspects concerning the for-mation of eastern Mediterranean sapropels. Marine Geology122:1–28.
Rohling, E. J., M. Sprovieri, T. Cane, J. S. L. Casford, S. Cooke,I. Bouloubassi, K. C. Emeis, R. Schiebel, M. Rogerson, A.Hayes, F. J. Jorissen, and D. Kroon. 2004. Reconstructing pastplanktic foraminiferal habitats using stable isotope data: acase history for Mediterranean sapropel S5. Marine Micro-paleontology 50:89–123.
Rudnicki, M. D., P. A. W. Wilson, and W. T. Anderson. 2001. As-sessing the effects of diagenesis and sediment properties onpore fluid profiles, Blake Nose, ODP Leg 171. Paleoceanog-raphy 16:1–13.
Saito, T., P. R. Thompson, and D. Breger. 1976. Skeletal ultra-microstructure of some elongate-chambered planktonic fo-raminifera and related species. Pp. 278–304 in Y. Takayanagiand T. Saito, eds. Progress in micropaleontology; selected pa-pers in honor of Prof. Kiyoshi Asano. MicropaleontologyPress, New York.
Samanta, B. K. 1973. Planktonic foraminifera from the Paleo-cene- Eocene succession in the Rakhi Nala, Sulaiman Range,Pakistan. Bulletin of the British Museum (Natural History)Geology 22:423–482.
Schlanger, S. O., and H. C. Jenkyns. 1976. Cretaceous oceanicanoxic events: causes and consequences. Geologie en Mi-jnbouw 55:179–184.
Schrag, D. P., D. J. DePaolo, and F. M. Richter. 1995. Reconstruct-ing past sea-surface temperatures—correcting for diagenesisof bulk marine carbonate. Geochimica et Cosmochimica Acta59:2265–2278.
Sexton, P. F., P. A. Wilson, and P. N. Pearson. 2006. Microstruc-tural and geochemical perspectives on planktic foraminiferalpreservation—‘glassy’ versus ‘frosty.’ Geochemistry, Geo-physics, Geosystems 7:Q12P19, doi: 10.1029/2006GC001291.
Spero, H. J. 1998. Life history and stable isotope geochemistryof planktonic foraminifera. Pp. 7–36 in Norris and Corfield1998.
Spero, H. J., and D. W. Lea. 1993. Intraspecific variability in theplanktonic foraminifera Globigerinoides sacculifer: results fromlaboratory experiments. Marine Micropaleontology 22:221–234.
———. 1996. Experimental determination of stable isotope var-iability in Globigerina bulloides: implications for palaeoceano-graphic reconstructions. Marine Micropaleontology 28:231–246.
Spero, H. J., and D. F. Williams. 1989. Opening the carbon iso-

514 HELEN K. COXALL ET AL.
tope ‘vital effect’ black box. I. Seasonal temperatures in theeuphotic zone. Paleoceanography 4:593–601.
Spero, H. J., I. Lerche, and D. F. Williams. 1991. Opening the car-bon isotope ‘vital effect’ black box. 2. Quantitative model forinterpreting foraminiferal carbon isotope data. Paleoceanog-raphy 6:639–655.
Spero, H. J., J. Bijma, D. W. Lea, and B. E. Bemis. 1997. Effect ofseawater carbonate concentration on foraminiferal carbonand oxygen isotopes. Nature 390:497–500.
Srinivasan, M. S., and J. P. Kennett. 1975. The status of Bolliella,Beella, Prontentella and related planktonic foraminifera basedon the well ultra structure. Journal of Foraminiferal Research5:155–165.
Stainforth, R. M. 1948. Applied micropaleontology in coastalEcuador. Journal of Paleontology 22:113–151.
Steineck, P. L., and R. L. Fleisher. 1978. Towards the classicalevolutionary reclassification of Cenozoic Globigerinacea (For-aminiferida). Journal of Paleontology 52:618–635.
Stuiver, M., and P. J. Reimer. 1993. Extended 14C data base andrevised CALIB 3.0 14C age calibration program. Radiocarbon35:215–230.
Subbotina, N. N. 1958. New genera and species of foraminifera.In N. K. Bykova, et al., eds. Microfauna of the USSR, Vol. 9.Trudy, VNIGRI, new series 115:5–105 (All-Union PetroleumScientific Research Geological Prospecting Institute, Lenin-grad). [Russian.]
Suess, E., R. von Huene, et al. eds. 1988. Site 683. Proceedingsof the Ocean Drilling Program, Initial Reports 112:437–524.Ocean Drilling Program, College Station, Tex.
Summerhayes, C. P., W. L. Prell, and K. C. Emeis. 1992. Evolu-tion of upwelling systems since the Early Miocene. GeologicalSociety of London Special Publication 64.
Verga, D., and I. Premoli Silva. 2002. Early Cretaceous plank-tonic foraminifera from the Tethys: the genus Leupoldina. Cre-taceous Research 23:189–212.
Watkins, J. M. 2003. Foraminifera species abundances from towTT011�8-MOC68, PANGAEA, doi: 10.1594/PANGAEA.123041.
Wilson, P. A., and R. D. Norris. 2001. Warm tropical ocean sur-face and global anoxia during the mid-Cretaceous period. Na-ture 412:425–429.
Wilson, P. A., R. D. Norris, and M. J. Cooper. 2002. Testing themid-Cretaceous greenhouse hypothesis using glassy forami-niferal calcite from the core of the Turonian tropics on De-merara Rise. Geology 30:607–610.
Wishner, K. F., and M. M. Gowing. 1992. The role of deep-seazooplankton in carbon cycles. Pp. 29–43 in G. T. Rowe and V.Pariente, eds. Deep-sea food chains and the global carbon cy-cle. Kluwer Academic, Dordrecht.
Wishner, K. F., C. J. Ashjian, C. Gelfman, M. M. Gowing, L.Kann, L. A. Levin, L. S. Mullineaux, and J. Saltzman. 1995.Pelagic and benthic ecology of the lower interface of the East-ern Tropical Pacific oxygen minimum zone. Deep-Sea Re-search Part I, Oceanographic Research Papers 42:93–115.
Zeebe, R. E. 1999. An explanation of the effect of seawater car-bonate concentration on foraminiferal oxygen isotopes. Geo-chimica et Cosmochimica Acta 63:2001–2007.
Zeebe, R. E., and P. Westbroek. 2003. A simple model for theCaCO3 saturation state of the ocean: the ‘‘Strangelove,’’ the‘‘Neritan,’’ and the ‘‘Cretan’’ ocean. Geochemistry, Geophys-ics, Geosystems 4(12). doi: 10.1029/2003GC000538.
Ziegler, A. M., D. B. Rowley, A. L. Lottes, D. L. Sahagrian, M. L.Hulver, and A. L. Gierlowski. 1985. Palaeogeographic inter-pretation: with an example from the mid Cretaceous. AnnualReview of Earth and Planetary Sciences 113:385–425.

515DIGITATE FORAM ITERATIVE EVOLUTION
App
endi
x
Stab
leis
otop
ed
ata
and
sam
ple
info
rmat
ion
.*in
dic
ates
C14
ages
con
vert
edto
cale
nd
arye
ars.
See
Tabl
e1
for
age-
dat
eso
urc
es.
Sam
ple
Per
iod
Bio
zon
e/a
Spec
ies
�13C
�18O
Dat
aso
urc
eSi
zefr
acti
onN
o.
DSD
P-13
7-12
-5,4
–6cm
Cen
oman
ian
KS1
7‘‘H
edbe
rgel
la’’
sim
plex
1.45
�2.
99T
his
stu
dy
�11
2D
SDP-
137-
12-5
,4–6
cmC
enom
ania
nK
S17
Rot
alip
ora
brot
zeni
1.77
�2.
60T
his
stu
dy
250–
300
DSD
P-13
7-12
-5,4
–6cm
Cen
oman
ian
KS1
7R
otal
ipor
acf
.bro
tzen
i1.
61�
3.08
Th
isst
ud
y25
0–30
0D
SDP-
137-
12-5
,4–6
cmC
enom
ania
nK
S17
Pra
eglo
botr
unca
nast
epha
ni2.
05�
2.97
Th
isst
ud
y25
0–30
0D
SDP-
137-
12-5
,4–6
cmC
enom
ania
nK
S17
Pra
eglo
botr
unca
nade
lrio
ensi
s1.
95�
3.20
Th
isst
ud
y25
0–30
0D
SDP-
137-
12-5
,4–6
cmC
enom
ania
nK
S17
Hed
berg
ella
delr
ioen
sis
2.04
�3.
47T
his
stu
dy
250–
300
DSD
P-13
7-12
-5,4
–6cm
Cen
oman
ian
KS1
7H
edbe
rgel
lapl
anis
pira
1.93
�4.
00T
his
stu
dy
250–
300
DSD
P-13
7-12
-5,4
–6cm
Cen
oman
ian
KS1
7G
lobi
geri
nello
ides
ultr
amic
rus
1.63
�3.
57T
his
stu
dy
250–
300
DSD
P-13
7-12
-5,4
–6cm
Cen
oman
ian
KS1
7G
yroi
dino
ides
sp.
0.40
�1.
84T
his
stu
dy
250–
300
DSD
P-13
7-12
-5,4
–6cm
Cen
oman
ian
KS1
8G
avel
linel
lasp
.0.
50�
1.85
Th
isst
ud
y25
0–30
0D
SDP-
144A
-6-1
,141
.5–1
42.5
cmC
reta
ceou
s,T
uro
nia
nN
P-U
C9a
Het
eroh
elix
glob
ulos
a0.
21�
3.89
Wil
son
etal
.200
265
–125
25D
SDP-
144A
-6-1
,141
.5–1
42.5
cmC
reta
ceou
s,T
uro
nia
nN
P-U
C9a
Het
eroh
elix
glob
ulos
a0.
38�
4.14
Wil
son
etal
.200
265
–125
18D
SDP-
144A
-6-1
,141
.5–1
42.5
cmC
reta
ceou
s,T
uro
nia
nN
P-U
C9a
Hed
berg
ella
delr
ioen
sis
0.86
�4.
36W
ilso
net
al.2
002
65–1
2525
DSD
P-14
4A-6
-1,1
41.5
–142
.5cm
Cre
tace
ous,
Tu
ron
ian
NP-
UC
9aH
edbe
rgel
lade
lrio
ensi
s0.
63�
4.03
Wil
son
etal
.200
265
–125
16D
SDP-
144A
-6-1
,141
.5–1
42.5
cmC
reta
ceou
s,T
uro
nia
nN
P-U
C9a
Hed
berg
ella
amab
ilis
0.70
�4.
22W
ilso
net
al.2
002
65–1
2516
DSD
P-14
4A-6
-1,1
41.5
–142
.5cm
Cre
tace
ous,
Tu
ron
ian
NP-
UC
9aH
eter
ohel
ixm
orem
ani
�0.
15�
3.95
Wil
son
etal
.200
265
–125
14D
SDP-
144A
-6-1
,141
.5–1
42.5
cmC
reta
ceou
s,T
uro
nia
nN
P-U
C9a
Cla
vihe
dber
gella
wat
ersi
0.26
�3.
29W
ilso
net
al.2
002
65–1
2510
DSD
P-14
4A-6
-1,7
5–76
cmC
reta
ceou
s,T
uro
nia
nN
P-U
C9a
ben
thic
-1�
0.88
�0.
84W
ilso
net
al.2
002
65–1
2510
DSD
P-14
4A-6
-1,7
5–76
cmC
reta
ceou
s,T
uro
nia
nN
P-U
C9a
ben
thic
-2�
1.44
0.60
Wil
son
etal
.200
265
–125
9K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Mor
ozov
ella
lens
iform
is2.
91�
0.37
Th
isst
ud
y35
5–40
06
Kan
e-9,
Cor
e42
,42
cmTo
pm
ost
low
erE
ocen
eP
8-P
9A
cari
nina
aspe
nsis
2.90
�0.
82T
his
stu
dy
355–
400
5K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Aca
rini
nabu
llbro
oki
2.89
�0.
64T
his
stu
dy
355–
400
5K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Subb
otin
acf
.fro
ntos
a1.
060.
32T
his
stu
dy
250–
300
6K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Para
subb
otin
agr
iffina
e0.
490.
68T
his
stu
dy
250–
300
6K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Para
subb
otin
agr
iffina
e0.
500.
64T
his
stu
dy
250–
300
6K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Cla
vige
rine
llaak
ersi
0.45
�0.
05T
his
stu
dy
Bro
ken
cham
bers
3K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Glo
bige
rina
thek
ase
nni
1.61
�1.
21T
his
stu
dy
212–
250
12K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Glo
bano
mal
ina
chap
man
i1.
13�
0.24
Th
isst
ud
y12
5–25
016
Kan
e-9,
Cor
e42
,42
cmTo
pm
ost
low
erE
ocen
eP
8-P
9C
ibic
idoi
des
sp.
0.61
0.55
Th
isst
ud
y25
0–40
04
Kan
e-9,
Cor
e42
,42
cmTo
pm
ost
low
erE
ocen
eP
8-P
9M
oroz
ovel
laca
ucas
ica
3.20
�0.
97T
his
stu
dy
355–
400
6K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Aca
rini
nabu
llbro
oki
2.45
�1.
35T
his
stu
dy
300–
355
6K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Glo
bige
rina
thek
ase
nni
1.35
�1.
53T
his
stu
dy
212–
300
9K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Subb
otin
asp
.1.
79�
1.03
Th
isst
ud
y21
2–30
04
Kan
e-9,
Cor
e42
,42
cmTo
pm
ost
low
erE
ocen
eP
8-P
9C
lavi
geri
nella
colo
mbi
ana
0.56
�0.
13T
his
stu
dy
Bro
ken
cham
bers
6K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Cla
vige
rine
llaak
ersi
0.48
�0.
20T
his
stu
dy
Bro
ken
cham
bers
6K
ane-
9,C
ore
42,4
2cm
Top
mos
tlo
wer
Eoc
ene
P8-
P9
Cib
icid
oide
ssp
.0.
770.
41T
his
stu
dy
250–
400
3O
DP-
865C
-8H
-3,7
0–72
cmTo
pm
ost
low
erE
ocen
eP
9-P
10C
lavi
geri
nella
eoca
nica
1.11
0.20
Cox
all
etal
.200
3�
250
3O
DP-
865C
-8H
-3,7
0–72
cmTo
pm
ost
low
erE
ocen
eP
9-P
10A
cari
nina
pseu
doto
pile
nis
3.19
�1.
20C
oxal
let
al.2
003
250–
300
5O
DP-
865C
-8H
-3,7
0–72
cmTo
pm
ost
low
erE
ocen
eP
9-P
10Su
bbot
ina
fron
tosa
1.77
�0.
27C
oxal
let
al.2
003
250–
300
5O
DP-
865C
-8H
-3,7
0–72
cmL
ower
/m
idd
leE
ocen
eP
9-P
10C
ibic
idoi
des
sp.
0.49
0.40
Cox
all
etal
.200
325
0–30
02
OD
P-86
5C-8
H-3
,110
–112
cmL
ower
/m
idd
leE
ocen
eP
9-P
10C
lavi
geri
nella
eoca
nica
1.56
0.35
Cox
all
etal
.200
3�
250
2O
DP-
865C
-8H
-3,1
10–1
12cm
Low
er/
mid
dle
Eoc
ene
P9-
P10
Mor
ozov
ella
arag
onen
sis
3.56
�1.
20C
oxal
let
al.2
003
250–
300
4O
DP-
865C
-8H
-3,1
10–1
12cm
Low
er/
mid
dle
Eoc
ene
P9-
P10
Aca
rini
naps
eudo
topi
leni
s3.
02�
0.80
Cox
all
etal
.200
325
0–30
06
OD
P-86
5C-8
H-3
,110
–112
cmL
ower
/m
idd
leE
ocen
eP
9-P
10Su
bbot
ina
fron
tosa
1.71
�0.
30C
oxal
let
al.2
003
250–
300
6O
DP-
865C
-8H
-3,1
10–1
12cm
Low
er/
mid
dle
Eoc
ene
P9-
P10
Cib
icid
oide
ssp
.0.
83�
0.10
Cox
all
etal
.200
325
0–30
03
OD
P-86
5C-8
H-3
,110
–112
cmL
ower
/m
idd
leE
ocen
eP
9-P
10G
uem
belit
rioi
des
nutt
ali
2.39
�0.
40C
oxal
let
al.2
003
250–
300
5

516 HELEN K. COXALL ET AL.
App
endi
x.C
onti
nu
ed.
Sam
ple
Per
iod
Bio
zon
e/a
Spec
ies
�13C
�18O
Dat
aso
urc
eSi
zefr
acti
onN
o.
OD
P86
5B-8
H-4
,137
–139
cmL
ower
/m
idd
leE
ocen
eP
9-P
10Pa
rasu
bbot
ina
eocl
ava
1.19
�0.
77C
oxal
let
al.2
003
250–
300
3O
DP
865B
-8H
-4,1
37–1
39cm
Low
er/
mid
dle
Eoc
ene
P9-
P10
Mor
ozov
ella
spin
ulos
a2.
90�
1.93
Cox
all
etal
.200
325
0–30
07
OD
P86
5B-8
H-4
,137
–139
cmL
ower
/m
idd
leE
ocen
eP
9-P
10A
cari
nina
pseu
doto
pile
nsis
3.06
�1.
69C
oxal
let
al.2
003
250–
300
5O
DP
865B
-8H
-4,1
37–1
39cm
Low
er/
mid
dle
Eoc
ene
P9-
P10
Subb
otin
afr
onto
sa1.
34�
1.08
Cox
all
etal
.200
325
0–30
05
OD
P-87
2C-9
H-2
,95–
97cm
Mid
dle
Mio
cene
N10
Cla
vato
rella
berm
udez
i1.
600.
55T
his
stu
dy
�25
02
OD
P-87
2C-9
H-2
,95–
97cm
Mid
dle
Mio
cene
N10
Glo
boqu
adri
nave
nezu
elan
a1.
97�
0.71
Th
isst
ud
y�
250
7O
DP-
872C
-9H
-2,9
5–97
cmM
idd
leM
ioce
neN
10G
lobi
geri
noid
esru
ber
2.41
�2.
85T
his
stu
dy
�25
012
OD
P-87
2C-9
H-2
,95–
97cm
Mid
dle
Mio
cene
N10
Glo
boro
talia
peri
pher
oron
da1.
88�
2.64
Th
isst
ud
y�
250
22O
DP-
872C
-9H
-2,9
5–97
cmM
idd
leM
ioce
neN
10C
lava
tore
llabe
rmud
ezi
1.86
0.33
Th
isst
ud
y�
250
3O
DP-
872C
-9H
-2,9
5–97
cmM
idd
leM
ioce
neN
10G
lobo
quad
rina
vene
zuel
ana
2.15
�0.
35T
his
stu
dy
�25
011
OD
P-87
2C-9
H-2
,95–
97cm
Mid
dle
Mio
cene
N10
Glo
bige
rino
ides
rube
r2.
85�
2.07
Th
isst
ud
y�
250
17O
DP-
872C
-9H
-2,9
5–97
cmM
idd
leM
ioce
neN
10G
lobo
rota
liape
riph
eror
onda
1.62
�2.
73T
his
stu
dy
�25
014
OD
P-87
2C-9
H-2
,95–
97cm
Mid
dle
Mio
cene
N10
Cla
vato
rella
berm
udez
i1.
590.
08T
his
stu
dy
�25
03
OD
P-87
2C-9
H-2
,95–
97cm
Mid
dle
Mio
cene
N10
Glo
boqu
adri
nave
nezu
elan
a1.
71�
0.79
Th
isst
ud
y�
250
16O
DP-
872C
-9H
-2,9
5–97
cmM
idd
leM
ioce
neN
10G
lobi
geri
noid
esru
ber
2.87
�2.
20T
his
stu
dy
�25
014
OD
P-87
2C-9
H-2
,95–
97cm
Mid
dle
Mio
cene
N10
Glo
boro
talia
peri
pher
oron
da1.
84�
2.08
Th
isst
ud
y�
250
7B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*C
ibic
idoi
des
wue
llers
torfi
0.29
2.35
Ber
ger
etal
.197
840
0–60
03
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Cib
icid
oide
sw
uelle
rsto
rfi0.
282.
98B
erge
ret
al.1
978
400–
600
3B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobi
geri
niod
essa
ccul
ifer
1.44
�1.
62B
erge
ret
al.1
978
400–
600
5B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobi
geri
niod
essa
ccul
ifer
1.66
�1.
32B
erge
ret
al.1
978
400–
600
5B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobi
geri
niod
essa
ccul
ifer
1.94
�1.
47B
erge
ret
al.1
978
400–
600
5B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobo
rota
liam
enar
dii
0.69
�1.
02B
erge
ret
al.1
978
400–
600
3B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobo
rota
liam
enar
dii
1.51
�0.
05B
erge
ret
al.1
978
400–
600
3B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobo
rota
liam
enar
dii
1.73
�0.
46B
erge
ret
al.1
978
400–
600
4B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobo
rota
liatu
mid
a1.
940.
04B
erge
ret
al.1
978
400–
600
3B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobo
rota
liatu
mid
a2.
06�
0.18
Ber
ger
etal
.197
840
0–60
03
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Glo
boro
talia
tum
ida
2.02
0.02
Ber
ger
etal
.197
840
0–60
04
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Neo
glob
oqua
drin
apa
chyd
erm
a0.
78�
0.56
Ber
ger
etal
.197
840
0–60
04
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Neo
glob
oqua
drin
apa
chyd
erm
a1.
34�
0.41
Ber
ger
etal
.197
840
0–60
04
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Neo
glob
oqua
drin
apa
chyd
erm
a1.
26�
0.47
Ber
ger
etal
.197
840
0–60
04
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Pul
leni
atin
aob
liqui
locu
lata
1.11
�1.
00B
erge
ret
al.1
978
400–
600
5B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*P
ulle
niat
ina
obliq
uilo
cula
ta1.
02�
0.98
Ber
ger
etal
.197
840
0–60
05
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Pul
leni
atin
aob
liqui
locu
lata
1.14
�0.
77B
erge
ret
al.1
978
400–
600
4B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobi
geri
nella
adam
si�
0.12
�0.
41T
his
stu
dy
�40
01
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Glo
bige
rine
llaad
amsi
�0.
18�
1.05
Th
isst
ud
y�
400
2B
ox-c
ore
ED
RC
-92-
5,22
–23
cmU
pp
erP
leis
toce
ne14
.2ka
*G
lobi
geri
nella
cf.a
dam
si�
0.39
�0.
19T
his
stu
dy
�40
02
Box
-cor
eE
DR
C-9
2-5,
22–2
3cm
Up
per
Ple
isto
cene
14.2
ka*
Glo
bige
rine
llacf
.ada
msi
�0.
36�
1.15
Th
isst
ud
y�
400
2C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—B
eella
digi
tata
0.28
0.80
Th
isst
ud
y�
250
5C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—B
eella
digi
tata
0.15
0.87
Th
isst
ud
y�
250
5C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—G
lobo
rota
liatu
mid
a1.
33�
0.70
Th
isst
ud
y�
250
1C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—G
lobo
rota
liatr
unca
tulin
oide
s1.
270.
54T
his
stu
dy
�25
03
Cor
eC
D15
4-07
-7PK
core
top
Hol
ocen
e—
Glo
boro
talia
trun
catu
linoi
des
0.90
0.24
Th
isst
ud
y�
250
1C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—G
lobi
geri
niod
essa
ccul
ifer
1.63
�1.
78T
his
stu
dy
�25
02
Cor
eC
D15
4-07
-7PK
core
top
Hol
ocen
e—
Glo
bige
rino
ides
rube
r0.
96�
1.83
Th
isst
ud
y�
250
2C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—N
eogl
oboq
uadr
ina
deut
ertr
ei1.
51�
0.92
Th
isst
ud
y�
250
4C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—C
ibic
idoi
des
wue
llers
torfi
0.88
2.14
Th
isst
ud
y�
250
3C
ore
CD
154-
07-7
PKco
reto
pH
oloc
ene
—C
ibic
idoi
des
wue
llers
torfi
0.84
2.20
Th
isst
ud
y�
250
8
Related Documents











