Isotope reconstruction of plant palaeoecology. Case study of Cenomanian £oras from Bohemia Thanh Thuy Nguyen Tu a; , Jir ›i Kvac ›ek b , David Ulic ›ny ¤ c , Herve ¤ Bocherens a , Andre ¤ Mariotti a , Jean Broutin d a Laboratoire de Bioge ¤ochimie Isotopique, Universite ¤ Paris VI, C.C. 120, 4 Place Jussieu, 75 252 Paris Cedex 05, France b Department of Palaeontology, National Museum, Va ¤clavske ¤ na ¤m 68, 115 79 Prague 1, Czech Republic c Geophysical Institute, Czech Academy of Sciences, Bocni II/1401, 14131 Prague 4, Czech Republic d Laboratoire de Pale ¤obotanique et Pale ¤oe ¤cologie, Universite ¤ Paris VI, 12 Rue Cuvier, 75 005 Paris, France Received 23 February 2001; accepted 25 October 2001 Abstract Palaeoenvironments inferred from stable carbon isotope ratios of fossil plants collected from various environments of the Cenomanian of Bohemia were compared with palaeobotanical and sedimentological data to test the use of isotope ratios as a record of local palaeoenvironments and fossil-plant ecology. A number of isotope and biogeochemical patterns suggested that stable carbon isotope ratios in the Cenomanian plants from Bohemia have not been significantly affected by diagenesis. Stable carbon isotope ratios of the palaeoflora from one of the sections studied were used as a reference for a non-stressed habitat since both sedimentological and palaeobotanical evidence suggested that the fossil flora underwent no environmental stresses that could have influenced its N 13 C values. Comparisons of N 13 C values of the other palaeofloras with that of the reference section, allowed inference of the palaeoenvironmental stresses undergone by fossil plants. These palaeoenvironmental patterns deduced from 13 C/ 12 C ratios are in agreement with the palaeoenvironments previously reconstructed by sedimentological and palaeobotanical studies. Combining evidence from those studies with isotope data provided detailed insights into the palaeoecology of the plants studied. Finally, stable carbon isotope ratios allowed precise characterisation of the ecology of the best-represented species of the deposits studied, the ginkgoalean plant Eretmophyllum obtusum and the conifer Frenelopsis alata, which were quite common in the salt-marsh environments in Europe during the Cenomanian. Hence, stable carbon isotope ratios can help in evaluating the environmental stresses undergone by fossil plants and the combination of these results with palaeobotanical and sedimentological data can provide detailed insights into fossil-plant ecology. ȣ 2002 Elsevier Science B.V. All rights reserved. Keywords: 13 C/ 12 C; fossil leaves; palaeoecology; Cenomanian; Eretmophyllum; Frenelopsis 1. Introduction The reconstruction of terrestrial palaeoenviron- ments in general and plant palaeoecology in par- ticular is usually based on sedimentological and palaeobotanical data (Batten, 1974; Pelzer et al., 0031-0182 / 02 / $ ^ see front matter ȣ 2002 Elsevier Science B.V. All rights reserved. PII:S0031-0182(01)00447-3 * Corresponding author. Present address: Laboratoire de Pale ¤obotanique et Pale ¤oe ¤cologie, Universite ¤ Paris VI, 12 Rue Cuvier, 75005 Paris, France. Fax: +33-1-4427-4164. E-mail address: [email protected] (T.T. Nguyen Tu). Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 www.elsevier.com/locate/palaeo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Isotope reconstruction of plant palaeoecology. Case study ofCenomanian £oras from Bohemia
Thanh Thuy Nguyen Tu a;�, Jir›i Kvac›ek b, David Ulic›ny¤ c, Herve¤ Bocherens a,Andre¤ Mariotti a, Jean Broutin d
a Laboratoire de Bioge¤ochimie Isotopique, Universite¤ Paris VI, C.C. 120, 4 Place Jussieu, 75 252 Paris Cedex 05, Franceb Department of Palaeontology, National Museum, Va¤clavske¤ na¤m 68, 115 79 Prague 1, Czech Republic
c Geophysical Institute, Czech Academy of Sciences, Bocni II/1401, 14131 Prague 4, Czech Republicd Laboratoire de Pale¤obotanique et Pale¤oe¤cologie, Universite¤ Paris VI, 12 Rue Cuvier, 75 005 Paris, France
Received 23 February 2001; accepted 25 October 2001
Abstract
Palaeoenvironments inferred from stable carbon isotope ratios of fossil plants collected from variousenvironments of the Cenomanian of Bohemia were compared with palaeobotanical and sedimentological data totest the use of isotope ratios as a record of local palaeoenvironments and fossil-plant ecology. A number of isotopeand biogeochemical patterns suggested that stable carbon isotope ratios in the Cenomanian plants from Bohemiahave not been significantly affected by diagenesis. Stable carbon isotope ratios of the palaeoflora from one of thesections studied were used as a reference for a non-stressed habitat since both sedimentological and palaeobotanicalevidence suggested that the fossil flora underwent no environmental stresses that could have influenced its N
13Cvalues. Comparisons of N13C values of the other palaeofloras with that of the reference section, allowed inference ofthe palaeoenvironmental stresses undergone by fossil plants. These palaeoenvironmental patterns deduced from13C/12C ratios are in agreement with the palaeoenvironments previously reconstructed by sedimentological andpalaeobotanical studies. Combining evidence from those studies with isotope data provided detailed insights into thepalaeoecology of the plants studied. Finally, stable carbon isotope ratios allowed precise characterisation of theecology of the best-represented species of the deposits studied, the ginkgoalean plant Eretmophyllum obtusum and theconifer Frenelopsis alata, which were quite common in the salt-marsh environments in Europe during theCenomanian. Hence, stable carbon isotope ratios can help in evaluating the environmental stresses undergone byfossil plants and the combination of these results with palaeobotanical and sedimentological data can provide detailedinsights into fossil-plant ecology. 7 2002 Elsevier Science B.V. All rights reserved.
Keywords: 13C/12C; fossil leaves; palaeoecology; Cenomanian; Eretmophyllum ; Frenelopsis
1. Introduction
The reconstruction of terrestrial palaeoenviron-ments in general and plant palaeoecology in par-ticular is usually based on sedimentological andpalaeobotanical data (Batten, 1974; Pelzer et al.,
0031-0182 / 02 / $ ^ see front matter 7 2002 Elsevier Science B.V. All rights reserved.PII: S 0 0 3 1 - 0 1 8 2 ( 0 1 ) 0 0 4 4 7 - 3
* Corresponding author. Present address: Laboratoire dePale¤obotanique et Pale¤oe¤cologie, Universite¤ Paris VI, 12 RueCuvier, 75005 Paris, France. Fax: +33-1-4427-4164.
E-mail address: [email protected](T.T. Nguyen Tu).
PALAEO 2802 30-5-02
Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70
www.elsevier.com/locate/palaeo

1992; Ulic›ny¤ et al., 1997). Palaeovegetation re-constructions are generally restricted to informa-tion on the taxonomic composition, relative abun-dance of individual taxa, or on tree density.Detailed reconstructions of the ecological distri-bution of fossil-plant communities usually needexceptional tapho£oras, such as fossil forestswith stumps preserved in situ (Mosbrugger etal., 1994). However, geochemical tools and espe-cially stable carbon isotope ratios1 generally needless fossil material and can yield valuable infor-mation on palaeoenvironments, provided that theisotope composition has not been signi¢cantly al-tered during diagenesis. For example, in the ma-rine realm, stable carbon isotope ratios are oftenused to infer palaeoatmospheric CO2 levels (e.g.Lasaga et al., 1985; Berner, 1990) or to recon-struct oceanic palaeoproductivity (e.g. Altabet etal., 1991; Knoll and Walter, 1992). In the conti-nental realm, stable carbon isotope ratios of ter-restrial plants have a great potential as palaeoen-vironmental and/or palaeoecological indicators.Indeed, during the photosynthetic uptake of at-mospheric CO2 in plants, carbon isotope fractio-nations take place, which di¡er fundamentally ac-cording to the di¡erent carbon ¢xation pathwaysof C3 and C4 plants (Smith and Epstein, 1971).Carbon isotope ratios of sediment or soil organicmatter were thus used to reconstruct recentchanges in vegetation composition (e.g. Dzurecet al., 1985; De Laune, 1986). Stable carbon iso-tope ratios in plants are also dependent on theisotope composition of atmospheric CO2 (e.g.Park and Epstein, 1960). N13C values of ancientplants were then studied to investigate the isotopiccomposition of past CO2 (e.g. Marino and MacElroy, 1991). However, it should be noted thatwhile marine phytoplankton N
13C values can rec-ord long-term variation in atmospheric CO2 lev-els, higher plant N13C values tend to remain con-stant with varying CO2 levels (e.g. Popp et al.,1989; Arens et al., 2000).Moreover, stable carbon isotope ratios in living
plants are also dependent on local environmental
parameters. Indeed, N13C values in terrestrial
plants are inversely proportional to the intercellu-lar CO2 concentration inside the leaf, Pi (Farqu-har, 1980, 1983; Francey and Farquhar, 1982)and Pi is related to environmental parameters,such as irradiance, temperature, soil-water con-tent or soil-water salinity (e.g. Farquhar, 1980;Farquhar et al., 1982a; Guy et al., 1986). Pi isinversely proportional to the e⁄ciency of intercel-lular CO2 utilisation, which depends on environ-mental parameters. Water availability plays a ma-jor role in inducing variations of N
13C values inplants (e.g. Farquhar et al., 1982b). Hydric stress(decreasing water availability) or osmotic stress(increasing salinity) induces a stomatal closureto limit water loss, which leads to increasedCO2-use e⁄ciency inside the leaf. When wateravailability decreases, isotope fractionation inplants thus tends to decrease, which ¢nally leadsto an increase in N
13C values (Guy et al., 1986;Farquhar et al., 1982b). Plants from water-stressed or salt-stressed environments are thus13C-enriched (e.g. Guy et al., 1980; Ehleringer,1993; Lipp et al., 1996). Otherwise, a decreaseof temperature or irradiance induces a decreaseof the enzymatic activity of plants, which leadsto a decrease of the CO2-use e⁄ciency inside theleaf (Troughton, 1972; Smith et al., 1976; Fran-cey et al., 1985; Schleser, 1990). Carbon isotopefractionation thus increases, ¢nally leading to adecrease of plant N13C value. Sun plants thus ex-hibit N
13C signi¢cantly higher than shade plants(e.g. Francey et al., 1985; Zimmerman and Ehler-inger, 1990).Variations of stable carbon isotope ratios in
response to local environmental parameters aretherefore well-documented for living plants (e.g.O’Leary, 1981; Ehleringer, 1991). N13C values intree rings were used to reconstruct recent climaticand/or environmental variations (e.g. Stuiver andBraziunas, 1987; Lipp et al., 1996; Sakata andSuzuki, 1998). Stable carbon isotope ratios inconifer needles were also recently studied to inves-tigate climatic characteristics of the last 40 000years (Pendall et al., 1999). Up to present, how-ever, only a few studies looked at the isotopecomposition of fossil plants to reconstruct global(Gro«cke, 1997; Hasegawa, 1997) or local (Lu«cke
1 Generally expressed as N13C values: N13C= [(13Rsample/13Rstandard)31]U1000, where 13R stands for the ratio 13C/12C.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7044

et al., 1999) palaeoenvironments. All the afore-mentioned studies were undertaken on fossilwoods. Nevertheless, fossil leaves represent an im-portant part of the plant fossil record and theiridenti¢cation is often easier than that of woods.In a study on Cenomanian leaves originatingfrom a single deposit, we recently demonstratedthat the intra-species and inter-species variationsof the N
13C values of the samples re£ected thesalinity of the soil on which the plants grew(Nguyen Tu et al., 1999). Stable carbon isotoperatios of fossil leaves thus have a great potentialto help in reconstructing local palaeoenviron-ments or plant palaeoecology. To fully validatethe use of carbon isotopes as a tool to character-ise plant palaeoecosystems, it is necessary to testthe variations of fossil plants N
13C values in var-ious deposits in which the environments are al-ready well documented by independent methods(e.g. sedimentology, palaeobotany). In this paper,we propose an isotope characterisation of severalfossil-leaf deposits corresponding to di¡erent en-vironments to (1) test the use of carbon isotopes
as local palaeoenvironment indicators by compar-ing the environments inferred from isotopes tothose inferred from other methods, and (2) inves-tigate whether isotopes can provide additional in-sights into fossil-plant ecology.The Peruc^Korycany Formation of the Bohe-
mian Cretaceous Basin o¡ers a number of fossil-iferous localities (e.g. Brn|¤k, Horous›any, Hlou-be›t|¤n, Pec|¤nov; Fig. 1) that yielded very richpalaeo£oras (Kvac›ek, 1995, 1997, 2000; Kvac›ekand Knobloch, 1997; Knobloch, 1999; Figs. 2and 3). The Cenomanian £oras from the Bohe-mian Basin have been known for their diversityand excellent preservation since the nineteenthcentury (Sternberg, 1820; Velenovsky¤, 1882,1883, 1884, 1885a,b, 1888a,b, 1889; Velenovsky¤and Vinikla¤r, 1926, 1927, 1929, 1931). Most ofthe Cenomanian Bohemian plants have alreadybeen described (Hlus›t|¤k and Konza¤lova, 1976;Kvac›ek, 1983, 1995, 1999; Hlus›t|¤k, 1986a; Kva-c›ek and Knobloch, 1997). Sedimentology andpalynology of the Cenomanian deposits havebeen studied in detail in several localities (Paclto-
Fig. 1. Schematic palaeogeographic map of the study area during the early Late Cenomanian with location of the sampling sites.F.Z., Fault zone; H, palaeohigh/palaeohigh axis; V, palaeovalleys and coastal plains; D, valley axis and direction of stream sys-tems; F, major fault zone/sense of movement; S, sampling sites. Partly modi¢ed after Ulic›ny¤ and SVpic›a¤kova¤ (1996) and SVpic›a¤ko-va¤ (1999).
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 45

PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7046

va¤, 1965, 1971, 1977, 1990; Svobodova¤, 1995;Ulic›ny¤ and SVpic›a¤kova¤, 1996; Svobodova¤ et al.,1998; SVpic›a¤kova¤, 1999), allowing a reasonably de-tailed environmental interpretation of the planthabitats. The palaeoenvironments span a widerange of £uvial and coastal settings (Ulic›ny¤ etal., 1997). Moreover, the Cenomanian Bohemian£oras are of considerable interest because: (1) allthe plants probably use the C3 photosyntheticpathway since there is no evidence of the C4 pho-tosynthetic pathway before Cenozoic, and (2) thefossil-rich sites occupy a relatively small region(Fig. 1) and occur within a relatively narrowstratigraphic interval of the Middle^early LateCenomanian (Fig. 4) indicating that global envi-ronmental parameters such as the mean isotopecomposition of atmospheric CO2 and temperaturedid not vary signi¢cantly among the sectionsstudied. Therefore, we assume the stable carbonisotope ratios of the Bohemian plants to havebeen primarily controlled by local environmentalconditions such as soil-water content, soil-watersalinity or irradiance. These plants therefore pro-vide exceptional opportunities to test the ability ofstable carbon isotopes in plants to record palae-oenvironments and palaeoecology.In this study, the environmental and ecological
signi¢cance of stable carbon isotope ratios inplants is tested in four localities of the Cenoma-nian of Bohemia exhibiting di¡erent environmen-tal conditions (Fig. 4). The combination of evi-dence from palaeobotany, sedimentology andisotope techniques should lead in future to a bet-ter utilisation of isotopes in palaeoenvironmentalsciences.
2. Material and methods
2.1. Regional framework
The sections sampled are located at the south-
western margin of the Bohemian Cretaceous Ba-sin, in the broader surroundings of Prague (Fig.1). The basin formed by reactivation of fault sys-tems in the Variscan basement of the BohemianMassif caused intra-plate stresses transmittedfrom the Alpine-Carpathian orogenic front (e.g.Ziegler, 1990). The south-western basin marginwas characterised by (1) low subsidence rate(not more than 35 m/Myr, as inferred from quan-titative analysis ; Ulic›ny¤, 1997), (2) a generally£at, ramp-type physiography, and (3) low sedi-ment input from the £at drainage area of thesouth-western part of the Bohemian Massif. Thetectonic setting and regional aspects are discussedin detail by Ulic›ny¤ and SVpic›a¤kova¤ (1996) andUlic›ny¤ (1997). The initial transgression duringthe Middle through Late Cenomanian created a£uvial^estuarine depositional setting by gradually£ooding a system of broad, northeast-directed pa-laeovalleys (Fig. 1), separated by £at palaeohighs.The trends of the valleys and highs were inheritedfrom structural characteristics of the Variscanbasement (Fig. 1). The valley ¢lls and contempo-raneous clastic deposits of Cenomanian age con-stitute the Peruc^Korycany Formation (CV ech etal., 1980), the oldest unit of the Bohemian Creta-ceous Basin ¢ll.The Peruc^Korycany Formation is formed by
£uvial, paralic (Peruc Member), shallow marine(Korycany Member) clastic deposits, and o¡shoreshales to marls (Pec|¤nov Member; Ulic›ny¤ et al.,1997; Fig. 4). In the region of interest, the oldestCretaceous deposits are of Middle Cenomanianage, based on palynologic data (Svobodova¤,1999, personal communication). During the lateMiddle Cenomanian (Acanthoceras jukesbrowneizone), a rapid sea-level rise caused the £oodingof most of the basin and a widespread depositionof mostly tide-in£uenced estuarine facies that ei-ther overlie older £uvial deposits or onlap the pre-Cretaceous basement. Individual units recognisedwithin the Peruc and Korycany Members (Fig. 4)
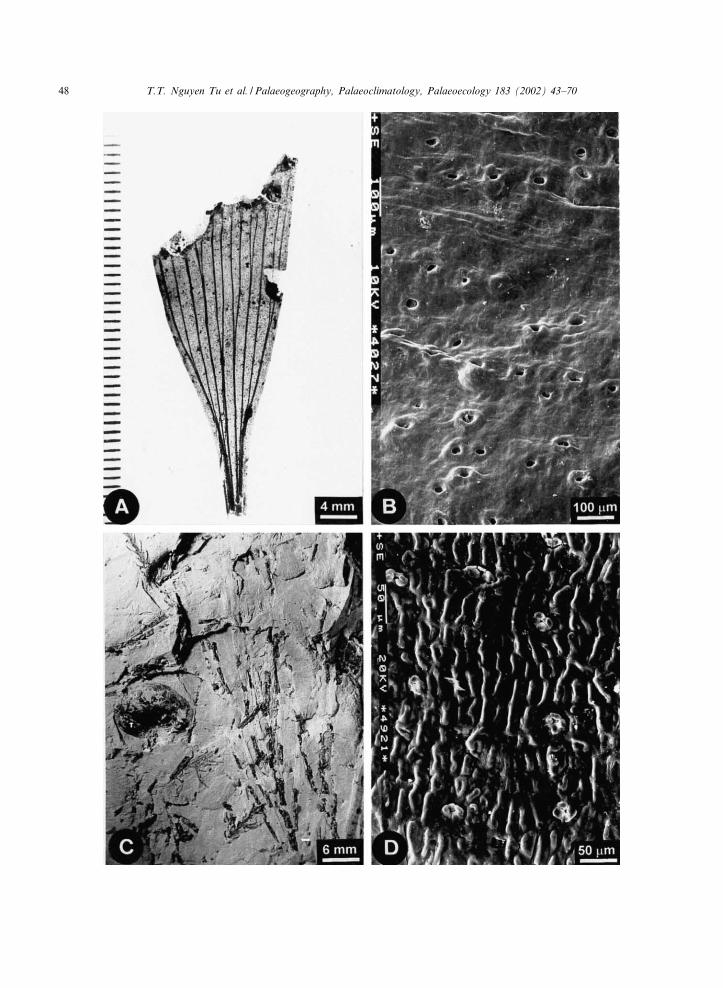
Fig. 2. Morphology of the main conifers and angiosperms sampled. (A) Leafy shoot of Sphenolepis pecinovensis, Pec|¤nov, unit 5,F 2110. (B) Leafy shoot of Cunninghamites lignitum, Pec|¤nov, unit 5, F 2113. (C) Leaf of Grevilleophyllum constans, Pec|¤nov, unit2, F 2867. (D) Leaf of Myrtophyllum geinitzii Heer, Pec|¤nov, unit 2, F 2123. (E) Leafy shoots of Ceratostrobus sequoiaphyllusand leaf of ‘Diospyros’ cretacea, Pec|¤nov, unit 3, F 2292.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 47
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7048

represent a response to high-frequency (ca. 100^120 Kyr) relative sea-level £uctuations, surim-posed on the long-term rise in sea level through-out the late Middle and early Late Cenomanian(Ulic›ny¤ and SVpic›a¤kova¤, 1996). After another ma-jor sea-level rise (Metoicoceras geslinianum zone),dark o¡shore shales of the Pec|¤nov Member weredeposited on the top of the Korycany Member.The Peruc^Korycany Formation is overlain byhemipelagic marls and limestones of the B|¤la¤Hora Formation (early Turonian, Fig. 4). Pa-laeoclimatic conditions for the Peruc^KorycanyFormation were recently calculated as subtropicalmesothermal semi-humid. CLAMP analysis (Kva-c›ek et al., 2000) based on 31 morphological char-acters of 68 angiosperm leaves gave the followingdata for the Peruc^Korycany £ora: mean annualtemperature 17^20‡C, mean annual precipitation1400 mm, length of growing season 9.5^10.5months.
2.2. Local geological setting
In the paper we concentrate on the deposits ofthe Peruc and Korycany Members. Each of thefour sections sampled (Fig. 4) comes from a sep-arate palaeovalley, and therefore each represents aslightly di¡erent palaeogeographic setting. Directcorrelation of individual units between the sec-tions in Fig. 4 is di⁄cult. However, a major £ood-ing surface, used as a datum in Fig. 4, can be usedfor rough physical correlation with a relativelyhigh degree of con¢dence (SVpic›a¤kova¤, 1999).Although the sections in Brn|¤k and Horous›anyquarries come from the deepest palaeovalleys¢lled with Cenomanian strata in Central Bohe-mia, the quarries do not provide access to thelowermost parts of the valley ¢lls which areknown only from boreholes. However, palynolog-ical evidence from the above quarries (Svobodo-va¤, 1999, personal communication) indicate aMiddle Cenomanian age of the £uvial depositsin the Brn|¤k and Horous›any sections. The over-
lying tide-dominated estuarine and storm-domi-nated shore-face deposits are of late Middle toLate Cenomanian age. The section of Hloube›t|¤ncontains only a thin portion of £uvial origin atthe base, unconformably overlying Ordovicianshales, while the upper part of the Hloube›t|¤n sec-tion is mostly analogous to the upper part of theHorous›any¤ section (with the exception of thestorm-dominated shore-face sandstones presentin Horous›any¤). The Pec|¤nov quarry (Fig. 1) islocated at the southwestern erosional limit ofthe Bohemian Cretaceous Basin. The Cenomanianstrata exposed in this quarry were deposited nearthe axis of the broad, shallow palaeovalley, andrepresent the most landward part of the preservedbasin ¢ll. Therefore, the £uvial deposits at thebase of the valley are approximately time-equiva-lent to the marsh or intertidal estuarine depositsof Hloube›t|¤n and Horous›any¤. Details on sedimen-tology and sequence stratigraphy of this sectionare provided by Ulic›ny¤ and SVpic›a¤kova¤ (1996)and Ulic›ny¤ et al. (1997).
2.3. Palaeobotanical context
The Peruc^Korycany £ora is very rich in taxaand represents the richest Cretaceous £ora in Eu-rope. Consequently, it is the ¢rst Cretaceous £orain Europe to exhibit a high diversi¢cation of an-giosperms. Their occurrence together with numer-ous Lower Cretaceous archaic gymnosperms(Tempskya, Sagenopsteris, Eretmophyllum, Frene-lopsis) makes the £ora even more interesting.Some of these have their last occurrence here(e.g. Tempskya, Nilssoniopteris, Eretmophyllum).Fossil plants in the Bohemian Cretaceous typi-cally occur in £uvial or sea-in£uenced sediments,particularly mudstones. Nineteen species originat-ing from various plant assemblages were exam-ined for isotope composition (Table 1).Fluvial sediments in the Peruc^Korycany For-
mation are generally of two types: braided river£uvial sediments bearing a fossil assemblage that
Fig. 3. Morphology of Eretmophyllum obtusum, Frenelopsis alata and Alvinia bohemica. (A) Basal part of naturally translucentleaf of E. obtusum, Prague, Hloube›t|¤n, F 116. (B) Scanning electron micrograph (SEM) of adaxial leaf surface of E. obtusumwith stomata, Pec|¤nov, unit 3, F 2481a. (C) Leafy shoot of F. alata and bract of cone scale of A. bohemica, Pec|¤nov, unit 3, F2288. (D) SEM of stem surface of F. alata, Podles›|¤n, F 16b.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 49

PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7050

is characterised by angiosperms associated withthe family Lauraceae (Myrtophyllum, Fig. 2D;Grevilleophyllum, Fig. 2C; Mauldinia), and mean-dering river sediments characterised by a morediverse plant assemblage with Myrtophyllum an-gustum (Velenovsky¤) Knobloch, Myrtophyllumgeinitzii Heer, Platanus sp., cycads (Jirusia, Nil-sonia, Microzamia) and pteridophytes (Anemia,Schizeopsis, Adianthium).Fossil plants from sea-in£uenced sediments are
divided into assemblages of back-swamp and saltmarsh. The back-swamp environment is charac-terised by numerous taxodiaceous conifers (‘Se-quoia’ ; Quasisequoia ; Sphenolepis, Fig. 2A; Cun-ninghamites, Fig. 2B), pteridophytes (Gleichenia),and scarce angiosperms (Cocculophyllum, Grevil-leophyllum). Environmentally the most interestingplant assemblages occupied salt marshes. This as-semblage, known as the Frenelopsis assemblage, isof low diversity and represented by the cheirole-pidiaceous conifer Frenelopsis alata (K. Feistman-tel) Knobloch and ginkgoalean plant Eretmophyl-lum obtusum (Velenovsky¤) J. Kvac›ek (Fig. 3).There are further specialised plants such as theenigmatic Dammarites albens Presl in Sternbergand Pseudoasterophyllites cretaceus O. Feistman-tel and the angiosperm ‘Diospyros’ cretacea Vele-novsky¤ and Vinikla¤r› (Fig. 2E). The plant physi-ognomy of this halophyte assemblage is char-acterised by thick leathery cuticles with stomatasunken in stomatal pits (Frenelopsis, Eretmophyl-lum), specialised stem morphology (Dammarites),reduced leaves (Frenelopsis, Pseudoasterophyl-lites), etc. The plants are semi-autochthonous, oc-curring in peat sediment composed mainly ofplant material (branches of F. alata and leavesof E. obtusum).
2.4. Analyses
The material analysed for this study includesabout 500 fossil-plant specimens. Within each de-
posit, more than 40 isotope measurements wereundertaken on Frenelopsis alata and on Eretmo-phyllum obtusum, while 10 samples or less havebeen analysed for the less represented taxa. Apreliminary series of isotope measurements wasundertaken to investigate the preservation of theisotope signal. A number of isotope features char-acteristic of modern plants were examined: (1)small variability (i.e. standard deviation) of N13Cvalues within a single species in a given £ora (i.e.less than 1.0x), (2) small variability within asame leaf (i.e. less than 0.5x), (3) 13C-enrich-ment of petiole when compared with the wholeleaf, and (4) 13C depletion of the cuticle and ofthe lipids when compared with the whole leaf.Therefore, as often as possible, various measure-ments were taken from di¡erent parts of the lam-ina and isotope measurements were performedseparately on the petiole, on the cuticle and onthe lipids. Carbon and nitrogen elemental abun-dances were also measured simultaneously to N
13Cvalues to investigate the possible pattern of dia-genetic alteration of the isotope signal. Moreover,for each deposit, the isotope composition of thesurrounding sediment was also measured to exam-ine possible homogenisation of the isotope signalby that of the sediment.Fossil leaves were collected in the ¢eld in July
1998. Blocks of sediments containing leaves weretaken from the ¢eld. Leaves were removed in thelaboratory and dusted free of any adhering par-ticle by two successive ultrasonications in distilledwater (10 min). Possible exogenous carbonateswere removed by acid treatment (HCl, pH 3; Bo-cherens and Mariotti, 1999); however, most of thesamples appeared devoid of exogenous carbo-nates. The cuticles were isolated from the rest ofthe leaves by 45 min ultrasonication in distilledwater. Lipids were extracted by stirring thecrushed leaves in CH2Cl2/CH3OH (2/1, v/v) over-night at room temperature. Since De Niro andHastorf (1985) have shown on prehistoric leaves
Fig. 4. Measured sections of the four localities sampled for this study. The vertical logs contain sedimentological informationand basic palaeoenvironmental interpretation for each of the units recognised in the sections. Numbers 1^5 for individual unitsin the Pec|¤nov section apply to that section only. A regionally correlated £ooding surface is used as a datum in this ¢gure; corre-lation between Pec|¤nov and Hloube›t|¤n is tentative. Sections from Ulic›ny¤ et al. (1997), SVpic›a¤kova¤ (1999) and unpublished data ofUlic›ny¤.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 51
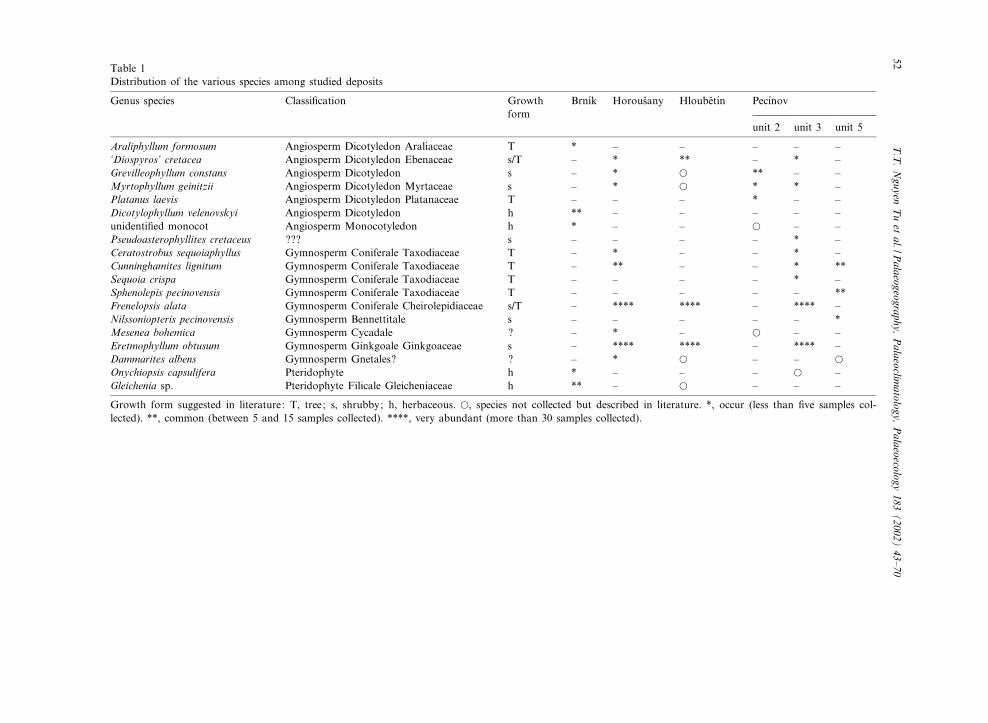
Table 1Distribution of the various species among studied deposits
Genus species Classi¢cation Growthform
Brn|¤k Horous›any Hloube›t|¤n Pec|¤nov
unit 2 unit 3 unit 5
Araliphyllum formosum Angiosperm Dicotyledon Araliaceae T * ^ ^ ^ ^ ^‘Diospyros’ cretacea Angiosperm Dicotyledon Ebenaceae s/T ^ * ** ^ * ^Grevilleophyllum constans Angiosperm Dicotyledon s ^ * a ** ^ ^Myrtophyllum geinitzii Angiosperm Dicotyledon Myrtaceae s ^ * a * * ^Platanus laevis Angiosperm Dicotyledon Platanaceae T ^ ^ ^ * ^ ^Dicotylophyllum velenovskyi Angiosperm Dicotyledon h ** ^ ^ ^ ^ ^unidenti¢ed monocot Angiosperm Monocotyledon h * ^ ^ a ^ ^Pseudoasterophyllites cretaceus ??? s ^ ^ ^ ^ * ^Ceratostrobus sequoiaphyllus Gymnosperm Coniferale Taxodiaceae T ^ * ^ ^ * ^Cunninghamites lignitum Gymnosperm Coniferale Taxodiaceae T ^ ** ^ ^ * **Sequoia crispa Gymnosperm Coniferale Taxodiaceae T ^ ^ ^ ^ * ^Sphenolepis pecinovensis Gymnosperm Coniferale Taxodiaceae T ^ ^ ^ ^ ^ **Frenelopsis alata Gymnosperm Coniferale Cheirolepidiaceae s/T ^ **** **** ^ **** ^Nilssoniopteris pecinovensis Gymnosperm Bennettitale s ^ ^ ^ ^ ^ *Mesenea bohemica Gymnosperm Cycadale ? ^ * ^ a ^ ^Eretmophyllum obtusum Gymnosperm Ginkgoale Ginkgoaceae s ^ **** **** ^ **** ^Dammarites albens Gymnosperm Gnetales? ? ^ * a ^ ^ a
Onychiopsis capsulifera Pteridophyte h * ^ ^ ^ a ^Gleichenia sp. Pteridophyte Filicale Gleicheniaceae h ** ^ a ^ ^ ^
Growth form suggested in literature: T, tree; s, shrubby; h, herbaceous. a, species not collected but described in literature. *, occur (less than ¢ve samples col-lected). **, common (between 5 and 15 samples collected). ****, very abundant (more than 30 samples collected).
PALAEO
280230-5-02
T.T.Nguyen
Tuet
al./Palaeogeography,
Palaeoclim
atology,Palaeoecology
183(2002)
43^7052
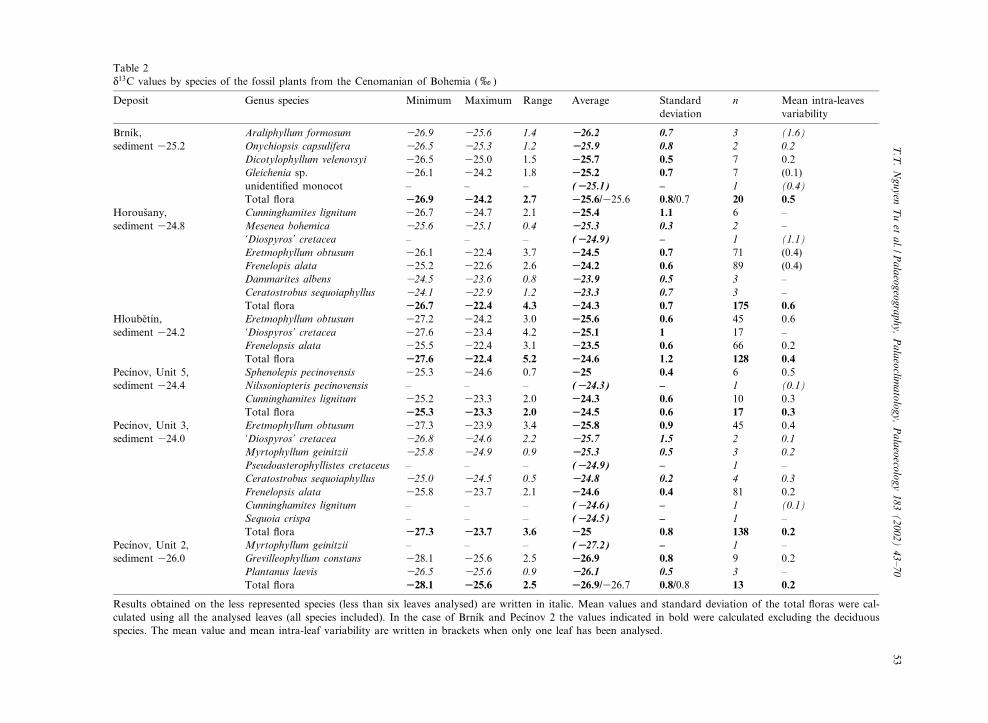
Table 2N13C values by species of the fossil plants from the Cenomanian of Bohemia (x)
Deposit Genus species Minimum Maximum Range Average Standarddeviation
n Mean intra-leavesvariability
Brn|¤k, Araliphyllum formosum 326.9 325.6 1.4 3326.2 0.7 3 (1.6)sediment 325.2 Onychiopsis capsulifera 326.5 325.3 1.2 3325.9 0.8 2 0.2
Dicotylophyllum velenovsyi 326.5 325.0 1.5 3325.7 0.5 7 0.2Gleichenia sp. 326.1 324.2 1.8 3325.2 0.7 7 (0.1)unidenti¢ed monocot ^ ^ ^ (3325.1) ^ 1 (0.4)Total £ora 3326.9 3324.2 2.7 3325.6/325.6 0.8/0.7 20 0.5
Horous›any, Cunninghamites lignitum 326.7 324.7 2.1 3325.4 1.1 6 ^sediment 324.8 Mesenea bohemica 325.6 325.1 0.4 3325.3 0.3 2 ^
‘Diospyros’ cretacea ^ ^ ^ (3324.9) ^ 1 (1.1)Eretmophyllum obtusum 326.1 322.4 3.7 3324.5 0.7 71 (0.4)Frenelopis alata 325.2 322.6 2.6 3324.2 0.6 89 (0.4)Dammarites albens 324.5 323.6 0.8 3323.9 0.5 3 ^Ceratostrobus sequoiaphyllus 324.1 322.9 1.2 3323.3 0.7 3 ^Total £ora 3326.7 3322.4 4.3 3324.3 0.7 175 0.6
Hloube›t|¤n, Eretmophyllum obtusum 327.2 324.2 3.0 3325.6 0.6 45 0.6sediment 324.2 ‘Diospyros’ cretacea 327.6 323.4 4.2 3325.1 1 17 ^
Frenelopsis alata 325.5 322.4 3.1 3323.5 0.6 66 0.2Total £ora 3327.6 3322.4 5.2 3324.6 1.2 128 0.4
Pec|¤nov, Unit 5, Sphenolepis pecinovensis 325.3 324.6 0.7 3325 0.4 6 0.5sediment 324.4 Nilssoniopteris pecinovensis ^ ^ ^ (3324.3) ^ 1 (0.1)
Cunninghamites lignitum 325.2 323.3 2.0 3324.3 0.6 10 0.3Total £ora 3325.3 3323.3 2.0 3324.5 0.6 17 0.3
Pec|¤nov, Unit 3, Eretmophyllum obtusum 327.3 323.9 3.4 3325.8 0.9 45 0.4sediment 324.0 ‘Diospyros’ cretacea 326.8 324.6 2.2 3325.7 1.5 2 0.1
Myrtophyllum geinitzii 325.8 324.9 0.9 3325.3 0.5 3 0.2Pseudoasterophyllistes cretaceus ^ ^ ^ (3324.9) ^ 1 ^Ceratostrobus sequoiaphyllus 325.0 324.5 0.5 3324.8 0.2 4 0.3Frenelopsis alata 325.8 323.7 2.1 3324.6 0.4 81 0.2Cunninghamites lignitum ^ ^ ^ (3324.6) ^ 1 (0.1)Sequoia crispa ^ ^ ^ (3324.5) ^ 1 ^Total £ora 3327.3 3323.7 3.6 3325 0.8 138 0.2
Pec|¤nov, Unit 2, Myrtophyllum geinitzii ^ ^ ^ (3327.2) ^ 1 ^sediment 326.0 Grevilleophyllum constans 328.1 325.6 2.5 3326.9 0.8 9 0.2
Plantanus laevis 326.5 325.6 0.9 3326.1 0.5 3 ^Total £ora 3328.1 3325.6 2.5 3326.9/326.7 0.8/0.8 13 0.2
Results obtained on the less represented species (less than six leaves analysed) are written in italic. Mean values and standard deviation of the total £oras were cal-culated using all the analysed leaves (all species included). In the case of Brn|¤k and Pec|¤nov 2 the values indicated in bold were calculated excluding the deciduousspecies. The mean value and mean intra-leaf variability are written in brackets when only one leaf has been analysed.
PALAEO
280230-5-02
T.T.Nguyen
Tuet
al./Palaeogeography,
Palaeoclim
atology,Palaeoecology
183(2002)
43^7053

that the adsorption of particulate soil matter (i.e.humic and fulvic acids) could shift the N13C valuesby 10^20x, 20 samples were treated with NaOHto remove any humic or fulvic acids. The absenceof signi¢cant variation of N13C values before andafter this treatment demonstrated that exogenoushumic or fulvic acid content of these fossil plantsdid not a¡ect the ¢nal measurements. Crushedsediment samples were prepared with the sameprotocol as for the fossil samples except the twoinitial rinses in distilled water. Stable isotope ra-tios were measured using an automatic unit thatcombined an elemental analyser with an isotoperatio mass spectrometer. The samples were com-busted in a Carlo-Erba CHN elemental analyserconnected to a VG-SIRA 10, isotope ratio massspectrometer. After £ash combustion at 1000‡Cwith copper oxide, CO2 was cryogenically dis-tilled, puri¢ed and introduced on-line into themass spectrometer to determine carbon isotoperatios (Girardin and Mariotti, 1991; Bocherensand Mariotti, 1999). The N
13C values were mea-sured with instrumental standard deviation of0.02x and repeated measurements on the samesample yielded a precision (1c) of 0.1x. Com-parisons of N
13C values between two batches ofsamples were undertaken by Mann^Whitney tests(Statview) and species-related e¡ect on N
13C val-ues was investigated by variance analyses (AN-OVA, Systat). The probabilities given in the textcorrespond to those of the samples studied be-longing to the same population.
3. Results and discussion
3.1. Preservation of the isotope signal in the fossilplants
The N13C values for the fossils studied range
from 328.1 to 322.4x, with an average valueof 324.7x. The results of isotope measurementsare summarised for each species in Table 2. Aver-age values for each species range from 327.2; to323.3x. As expected for Mesozoic times, theN13C values of fossil plants studied fall in therange of living C3 plants in which N
13C valuesvary between 335.0 and 321.0x (e.g. Ehler-
inger, 1991). Several experimental studies on mod-ern plants (Balesdent and Mariotti, 1996; Lallier-Verge's et al., 1998; Huang et al., 1997) and inves-tigations on fossil ones (Degens, 1969; Nambudiriet al., 1978; Rigby et al., 1981; Aucourt and Hill-aire-Marcel, 1993; Bocherens et al., 1993; NguyenTu et al., 1999) suggested that fossil plants typi-cally have retained their original isotope compo-sition. However, some studies on fossil-plant ma-terial reported diagenetic alteration of the isotopesignal. For example, Jones (1994) noticed unusu-ally high N
13C values for woods originating from aCarboniferous high-ranked coal.
3.1.1. Biogeochemical patternsAll the samples studied were collected in sedi-
ments of similar lithology (i.e. clays) to avoid dif-ferential diagenetic alteration due to the compo-sition of the surrounding sediment (van Bergen etal., 1994; Gro«cke, 1998). Moreover, the clays ofthe Peruc^Korycany Formation were depositedunder anoxic conditions, which provided favour-able conditions for a good preservation of thefossil plants. Furthermore, within each deposit,all the fossil species studied generally exhibitN13C values that are di¡erent from each otherand di¡erent from the one of the surroundingsediment (Table 2). These results show that theisotope signal of the fossil plants has not beenobliterated by that of the surrounding sediment.Diagenesis can lead to an alteration of the N13C
values of an entire plant tissue by selective loss ofcarbon with a N
13C value that is signi¢cantly dif-ferent from that of the carbon which is preserved(e.g. Benner et al., 1987). The various constituentsof plant tissues exhibit di¡erent C/N ratios andRundgren et al. (2000) have recently suggested,on Holocene subfossil leaves, that C/N ratioscan provide useful indicators for the degree ofdecomposition of fossil leaves. Moreover, claysand sands are known to lead to di¡erent degreesof preservation of fossil plants. In a study onCretaceous woods collected from sand and claylevels, Gro«cke (1998) demonstrated that the di¡er-ence of N13C values between samples from sandsand samples from clays was strongly correlated totheir C/N ratios. Therefore, a correlation betweenC/N ratios and N
13C values appears to be a good
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7054

indicator of diagenesis of the isotope signal. Nosigni¢cant correlation between the C/N ratios andthe N
13C values could be obtained for the fossilplants of the Cenomanian of Bohemia either atthe leaf level, nor at the species or at the depositlevel. Correlation coe⁄cients obtained for linearor exponential regression were all signi¢cantlylower than the Pearson critical value at the 5%or 1% con¢dence level. These lack of correlationsbetween N
13C values and C/N ratios tend to dem-onstrate that diagenesis has not signi¢cantly af-fected the isotope signal of the Cenomanianplants of Bohemia.
3.1.2. Isotope patternsThe speci¢c range of variation of N13C values of
the fossil plants studied vary between 0.7 and4.2x. These ranges of variation are similar tothose observed for living plants (e.g. Ehleringer,1991). Moreover, living plants generally exhibit,within a given population, mean standard devia-tion of N
13C values of 0.2 and 0.4x within agiven leaf and within a given species, respectively(Ehleringer, 1991). The mean standard deviationswithin a given leaf and within a given speciescalculated for the fossil plants of Bohemia are0.3 and 0.7x, respectively. These standard devi-ations are slightly higher than these reported byEhleringer (1991). However, they are of the samemagnitude and the intra-leaf deviation is system-atically inferior to the intra-species deviation.Additionally, isotope analyses of speci¢c parts
of the fossil plants revealed that: (1) cuticles ofEretmophyllum obtusum and Frenelopsis alata are13C-depleted by about 1x when compared withthe bulk leaves, (2) total lipids of E. obtusum andF. alata are 13C-depleted by about 5x whencompared with the bulk leaves, (3) petioles from‘Diospyros’ cretacea, Platanus sp. and the gneta-lean leaves are 13C-enriched by about 1x whencompared with the lamina and (4) the 13C-enrich-ment of the petiole when compared with the lam-ina could not be detected for E. obtusum. Isotopedistribution patterns (1), (2) and (3) are similar tothese generally observed for living plants. More-over, the absence of any 13C-enrichment of thepetiole of E. obtusum when compared with thelamina is in agreement with the fact that this spe-
cies does not exhibit a petiole clearly di¡erenti-ated from the lamina.All these isotope distribution patterns suggest
that diagenesis has not signi¢cantly a¡ected theisotope signal in the fossil plants of the Cenoma-nian of Bohemia.
3.1.3. N13C values of the Bohemia fossils when
compared with living plantsThe mean N
13C value of the samples (324.7x)are signi¢cantly higher than the mean value ofextant C3 plants living in an optimal environment(328.0x). Such a 13C-enrichment of fossil plantshas already been noticed by several authors (e.g.Popp et al., 1989; Bocherens et al., 1993). Gro«cke(1998) has reviewed the possible causes of this13C-enrichment of fossil plants :(1) A 13C-enrichment of atmospheric CO2 dur-
ing the Cenomanian when compared with nowa-days could explain the observed enrichment. Pastisotope compositions of atmospheric CO2 arenot well-documented. However, it was probablyabout 36.5x during the Cretaceous (Cerling,1991) when compared with 38.0x at present(Globalview-CO2, 1988).(2) On a short time scale, a decrease of atmo-
spheric CO2 levels can induce an increase in theN13C values of plants (e.g. Park and Epstein,1960; Pen‹uelas and Estiarte, 1997). However,higher plant N13C values tend to remain constantunder long-term CO2 level variations (e.g. Popp etal., 1989; Arens et al., 2000). Moreover, even ifpast CO2 levels are not well known they wereprobably higher during the Cretaceous than atpresent (e.g. Berner, 1990; Ehleringer and Cerl-ing, 1995; Raven, 1998).(3) An increase in the atmospheric O2 level can
induce an increase of plant N13C value (Smith etal., 1976; Deines, 1980). Atmospheric O2 levelswere indeed probably higher during the Creta-ceous (Berner and Landis, 1988).(4) According to some authors an increase in
the mean annual temperature could induce a 13C-enrichment in plants (e.g. Pearman et al., 1976;Nielsen and Shari¢, 1998). Temperature wasprobably higher during the Cenomanian whencompared with nowadays (Barron, 1983; Kvac›ek,2000).
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 55

(5) Hydrically stressed environmental condi-tions (i.e. limited water availability) can inducean increase in plants N
13C value (e.g. Guy et al.,1980) and the climate was probably semi-arid inBohemia during the Cenomanian (Ulic›ny¤ et al.,1997).Obviously, all the aforementioned hypotheses
can partially explain the 13C-enrichment observedon the Cenomanian plants but none of them canbe favoured. It is therefore preferable to under-take the palaeoecological reconstruction of thesamples studied using comparison of isotopedata within the Bohemian £ora, rather than di-rectly comparing the isotope ratios of fossil plantswith these of living plants.
3.2. Stable carbon isotope ratios and localpalaeoenvironments in the Bohemian £oras
As previously demonstrated, stable carbon iso-tope ratios have not been signi¢cantly a¡ected bydiagenesis in the fossil plants from the Cenoma-nian of Bohemia. The aim of the present study isthen to investigate the record of a palaeoenviron-mental/palaeoecological signal by carbon isotoperatios in these fossil plants. Since all the sampleswere collected in the same formation, within ashort time-period and a restricted area, it can beconsidered that global environmental parameters(i.e. temperature, atmospheric O2 level or 13Ccontent of atmospheric CO2) did not vary signi¢-cantly among the various deposits studied.A cultivar-level variation in carbon isotope
composition is known to exist within crop species.While such a variation may represent the productof agricultural breeding, this variation probablyre£ects the level of genetic variation to be ex-pected in native species (Ehleringer, 1993). Never-theless, N13C values in plants are believed to beprimarily controlled by environmental parametersrather than genetics (e.g. Comstock and Ehler-inger, 1992; Rice, 1999). Signi¢cant variations ofleaf N
13C values can occur within a given plantpresenting an arborescent bearing. Indeed, di¡er-ences in microenvironmental parameters such asirradiance, 13C/12C ratio of atmospheric CO2 andbranch length and orientation (e.g. Medina andMinchin, 1980; Schleser, 1990; Waring and
Silvester, 1994), can induce di¡erences up to6x in the 13C content of leaves collected fromdi¡erent parts of a large tree. Nevertheless, biaslinked to intra-individual variations of the isotopesignal can be avoided by working with the meanN13C value of a given species of which the numberof samples investigated is signi¢cant. In thisstudy, the palaeoenvironmental/palaeoecologicalinterpretations are based mainly on species meanN13C values, although only a third of the speciesstudied might have constituted trees (Table 1) andthe interpretation of species represented by lessthan six specimens is cautiously expressed.Additionally, studies on extant and fossil
woods (Stuiver and Braziunas, 1987; Lu«cke etal., 1999) revealed that gymnosperms can exhibitsigni¢cantly heavier isotope compositions thanangiosperms, which is probably due to the di¡er-ence in chemical and structural composition thatis well known between angiosperm and gymno-sperm woods (e.g. Sarkanen and Ludwig, 1971).However, this di¡erence did not appear to hide anenvironmental signal since Stuiver and Braziunas(1987) found a signi¢cant relationship betweentemperature and N
13C values combining both an-giosperm and gymnosperm data. Moreover, it isthe di¡erence between N
13C values of deciduousand evergreen leaves that is signi¢cant, ratherthan the di¡erence between N
13C values of gym-nosperm and angiosperm leaves (e.g. Buchmannet al., 1997; Hanba et al., 1997; Kloeppel et al.,1998). Except for Platanus and Dicotylophyllum,all the genera investigated in the present study(1) have been diagnosed as evergreen (i.e. Grevil-leophyllum, Myrtophyllum, Pseudoasterophyllites ;Ulic›ny¤ et al., 1997), or (2) exhibit xerophytic char-acteristics (i.e. other angiosperms, Eretmophyllum,Dammarites ; Hlus›t|¤k, 1977; Kvac›ek, 1983; Ulic›ny¤et al., 1997; Kvac›ek and Dilcher, 2000) that aretypical of evergreen plants (e.g. Tomlinson, 1994),or else (3) belong to families that are now onlyrepresented by evergreen genera (i.e. other gym-nosperms, pteridophytes). Therefore, the palaeo-ecological interpretations suggested in the presentstudy are based on N
13C values of evergreen spe-cies.Finally, considering (1) the high abundance of
plant macrofossils, (2) their excellent morpholog-
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7056

ical preservation and (3) the absence of any evi-dence of abrupt burial of living vegetation in situ,we can conclude that the fossil leaves fell at ma-turity which allow exclusion of the possibility thatthe di¡erences in N
13C observed between the var-ious plants are due to di¡erence of maturity. Inconclusion, it appears that N
13C values of thefossil plants studied can be interpreted in termsof local environments. As previously suggested,these palaeoenvironmental interpretations willbe undertaken by comparing the N
13C values ofthe fossil plants within a same deposit or betweendeposits. However, to interpret N
13C values interm of palaeoenvironments, that is to say interms of stresses, it is ¢rstly necessary to have areference deposit in which sedimentological or pa-laeobotanical data show that the fossil plantunderwent no stress.
3.2.1. Reference sample: meandering £uvialsystem, Brn|¤k sectionTwo main lithological units occur in the Brn|¤k
quarry (Figs. 1 and 4). The lower portion of thesection is dominated by muddy £oodplain depos-its interbedded with channel systems and compris-ing several palaeosoils (SVpic›a¤kova¤ and Ulic›ny¤,1996). The overlying succession is dominated bysandstones and conglomerates overlying a majorerosional unconformity and is interpreted as adeposit of a braided river ¢lling an incised valleysystem (Fig. 4; SVpic›a¤kova¤ and Ulic›ny¤, 1996). Theplant macrofossils sampled come from a part ofthe section interpreted as a back-swamp of a me-andering stream £oodplain. Because unequivocalmarine indicators are absent in the facies sampledwe conclude that the fossil plants from this local-ity underwent no osmotic stress.The clay beds from Brn|¤k quarry yielded a fos-
sil macro£ora essentially composed of angio-sperms (Myrtophyllum angustum, Dicotylophyllumvelenovskyi Knobloch, Araliphyllum formosum(Heer) Velenovsky¤, ‘Magnolia’ sp., ‘Bignonia’ sp.etc), but also including some cycads and severalferns (Anemia sp. and Gleichenia sp.; Kvac›ek,1998). A marked absence of gymnosperm was no-ticed for this palaeo£ora. This assemblage is sug-gested to grow in the £ood plain of meanderingriver, where ferns vegetated the most wet areas
and angiosperms perhaps occupied levees andslopes. Five species were sampled for isotopeanalyses: two dicotyledonous angiosperms (A.formosum and the water plant D. velenovskyi),one unidenti¢ed monocotyledonous angiospermand two ferns (Gleichenia sp. and Animia sp.).The presence of a water plant with large leavesand its possible £oating system should be noted(i.e. D. velenovskyi). Indeed, it provides evidencefor large water availability in the environment.The other plants of the deposit exhibit a degreeof morphological preservation similar to that ofthe water plant. Hence, it can be concluded thatall the plants were transported over similar dis-tances prior to deposition and we assume that allthese plants grew in the same environment. As aresult, it can be considered that the fossil plantsfrom the level sampled at Brn|¤k underwent nohydric stress. Hence, it appears that the palaeo-£ora from Brn|¤k did not grow in an environmentwith any requirement for the stomatal conduc-tance (salinity, drought) that could have led toan increase in their N
13C values.The two other parameters that can in£uence
plant N13C values are temperature and irradiance(e.g. O’Leary, 1981). A decrease of these parame-ters induces a decrease of plant N13C values. Localrecycling of respired CO2 may also lead to a de-crease of plant 13C content (e.g. Vogel, 1978;Medina and Minchin, 1980). The most obviousenvironment that can induce a signi¢cant decreaseof plant N
13C values is canopy forest. Indeed,under canopy, recycling of 13C-depleted CO2 re-leased by soil micro-organisms during respirationand limited irradiance are known to induce a 13Cdepletion of the leaves growing near the soil whencompared with canopy leaves (e.g. Vogel, 1978;Medina and Minchin, 1980). No unequivocal pa-laeobotanical data provides evidence for the pres-ence or absence of canopy forest vegetation.However, we will assume, as a working hypothe-sis, that the plants from the Brn|¤k section did notbelong to a canopy forest vegetation that wouldhave led to a decrease of their N13C values. More-over, the hypothesis of any variation, among thedeposits studied, of global environmental param-eters such as temperature and N
13CCO2 has beenruled out previously. Therefore, it will be assumed
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 57
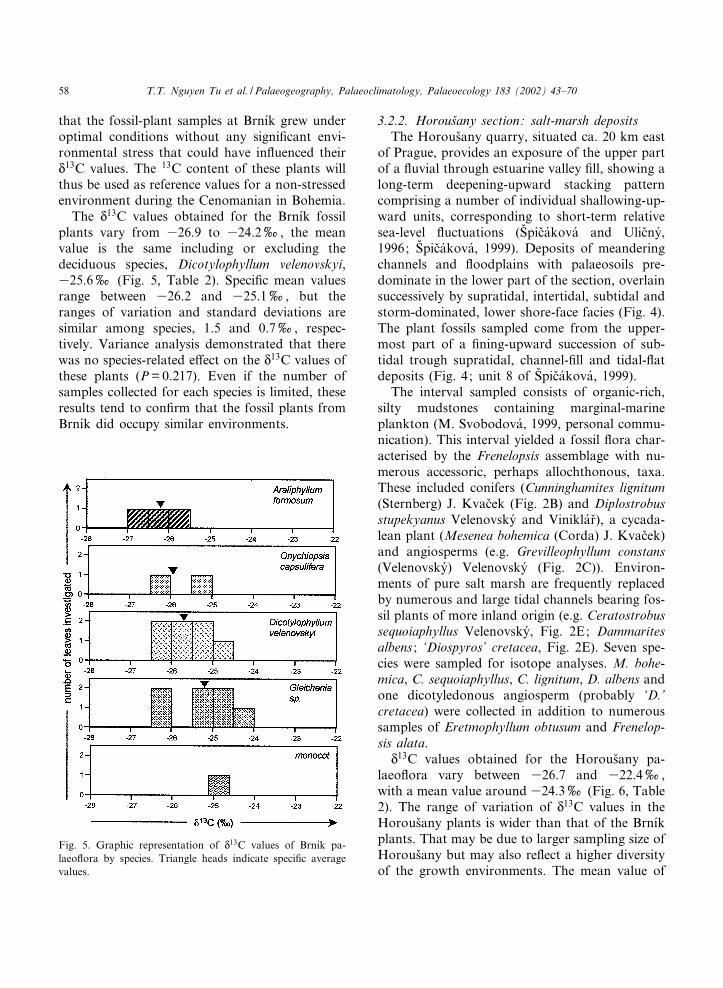
that the fossil-plant samples at Brn|¤k grew underoptimal conditions without any signi¢cant envi-ronmental stress that could have in£uenced theirN13C values. The 13C content of these plants willthus be used as reference values for a non-stressedenvironment during the Cenomanian in Bohemia.The N
13C values obtained for the Brn|¤k fossilplants vary from 326.9 to 324.2x, the meanvalue is the same including or excluding thedeciduous species, Dicotylophyllum velenovskyi,325.6x (Fig. 5, Table 2). Speci¢c mean valuesrange between 326.2 and 325.1x, but theranges of variation and standard deviations aresimilar among species, 1.5 and 0.7x, respec-tively. Variance analysis demonstrated that therewas no species-related e¡ect on the N13C values ofthese plants (P=0.217). Even if the number ofsamples collected for each species is limited, theseresults tend to con¢rm that the fossil plants fromBrn|¤k did occupy similar environments.
3.2.2. Horous›any section: salt-marsh depositsThe Horous›any quarry, situated ca. 20 km east
of Prague, provides an exposure of the upper partof a £uvial through estuarine valley ¢ll, showing along-term deepening-upward stacking patterncomprising a number of individual shallowing-up-ward units, corresponding to short-term relativesea-level £uctuations (SVpic›a¤kova¤ and Ulic›ny¤,1996; SVpic›a¤kova¤, 1999). Deposits of meanderingchannels and £oodplains with palaeosoils pre-dominate in the lower part of the section, overlainsuccessively by supratidal, intertidal, subtidal andstorm-dominated, lower shore-face facies (Fig. 4).The plant fossils sampled come from the upper-most part of a ¢ning-upward succession of sub-tidal trough supratidal, channel-¢ll and tidal-£atdeposits (Fig. 4; unit 8 of SVpic›a¤kova¤, 1999).The interval sampled consists of organic-rich,
silty mudstones containing marginal-marineplankton (M. Svobodova¤, 1999, personal commu-nication). This interval yielded a fossil £ora char-acterised by the Frenelopsis assemblage with nu-merous accessoric, perhaps allochthonous, taxa.These included conifers (Cunninghamites lignitum(Sternberg) J. Kvac›ek (Fig. 2B) and Diplostrobusstupekyanus Velenovsky¤ and Vinikla¤r›), a cycada-lean plant (Mesenea bohemica (Corda) J. Kvac›ek)and angiosperms (e.g. Grevilleophyllum constans(Velenovsky¤) Velenovsky¤ (Fig. 2C)). Environ-ments of pure salt marsh are frequently replacedby numerous and large tidal channels bearing fos-sil plants of more inland origin (e.g. Ceratostrobussequoiaphyllus Velenovsky¤, Fig. 2E; Dammaritesalbens ; ‘Diospyros’ cretacea, Fig. 2E). Seven spe-cies were sampled for isotope analyses. M. bohe-mica, C. sequoiaphyllus, C. lignitum, D. albens andone dicotyledonous angiosperm (probably ‘D.’cretacea) were collected in addition to numeroussamples of Eretmophyllum obtusum and Frenelop-sis alata.
N13C values obtained for the Horous›any pa-
laeo£ora vary between 326.7 and 322.4x,with a mean value around 324.3x (Fig. 6, Table2). The range of variation of N
13C values in theHorous›any plants is wider than that of the Brn|¤kplants. That may be due to larger sampling size ofHorous›any but may also re£ect a higher diversityof the growth environments. The mean value of
Fig. 5. Graphic representation of N13C values of Brn|¤k pa-
laeo£ora by species. Triangle heads indicate speci¢c averagevalues.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7058
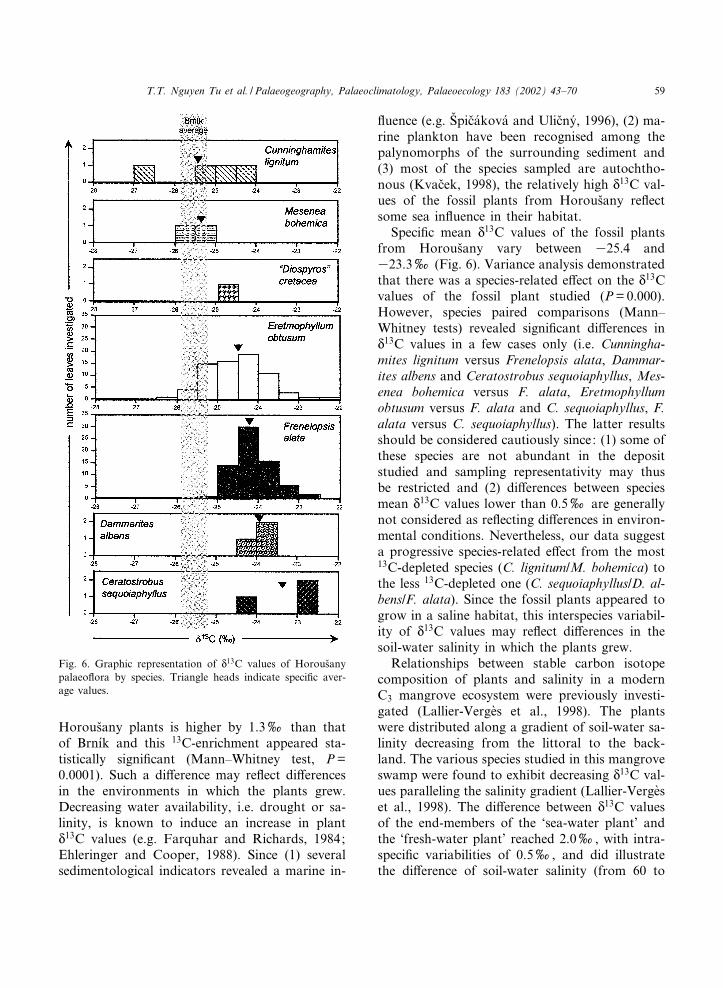
Horous›any plants is higher by 1.3x than thatof Brn|¤k and this 13C-enrichment appeared sta-tistically signi¢cant (Mann^Whitney test, P=0.0001). Such a di¡erence may re£ect di¡erencesin the environments in which the plants grew.Decreasing water availability, i.e. drought or sa-linity, is known to induce an increase in plantN13C values (e.g. Farquhar and Richards, 1984;Ehleringer and Cooper, 1988). Since (1) severalsedimentological indicators revealed a marine in-
£uence (e.g. SVpic›a¤kova¤ and Ulic›ny¤, 1996), (2) ma-rine plankton have been recognised among thepalynomorphs of the surrounding sediment and(3) most of the species sampled are autochtho-nous (Kvac›ek, 1998), the relatively high N
13C val-ues of the fossil plants from Horous›any re£ectsome sea in£uence in their habitat.Speci¢c mean N
13C values of the fossil plantsfrom Horous›any vary between 325.4 and323.3x (Fig. 6). Variance analysis demonstratedthat there was a species-related e¡ect on the N
13Cvalues of the fossil plant studied (P=0.000).However, species paired comparisons (Mann^Whitney tests) revealed signi¢cant di¡erences inN13C values in a few cases only (i.e. Cunningha-mites lignitum versus Frenelopsis alata, Dammar-ites albens and Ceratostrobus sequoiaphyllus, Mes-enea bohemica versus F. alata, Eretmophyllumobtusum versus F. alata and C. sequoiaphyllus, F.alata versus C. sequoiaphyllus). The latter resultsshould be considered cautiously since: (1) some ofthese species are not abundant in the depositstudied and sampling representativity may thusbe restricted and (2) di¡erences between speciesmean N
13C values lower than 0.5x are generallynot considered as re£ecting di¡erences in environ-mental conditions. Nevertheless, our data suggesta progressive species-related e¡ect from the most13C-depleted species (C. lignitum/M. bohemica) tothe less 13C-depleted one (C. sequoiaphyllus/D. al-bens/F. alata). Since the fossil plants appeared togrow in a saline habitat, this interspecies variabil-ity of N
13C values may re£ect di¡erences in thesoil-water salinity in which the plants grew.Relationships between stable carbon isotope
composition of plants and salinity in a modernC3 mangrove ecosystem were previously investi-gated (Lallier-Verge's et al., 1998). The plantswere distributed along a gradient of soil-water sa-linity decreasing from the littoral to the back-land. The various species studied in this mangroveswamp were found to exhibit decreasing N
13C val-ues paralleling the salinity gradient (Lallier-Verge'set al., 1998). The di¡erence between N
13C valuesof the end-members of the ‘sea-water plant’ andthe ‘fresh-water plant’ reached 2.0x, with intra-speci¢c variabilities of 0.5x, and did illustratethe di¡erence of soil-water salinity (from 60 to
Fig. 6. Graphic representation of N13C values of Horous›any
palaeo£ora by species. Triangle heads indicate speci¢c aver-age values.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 59
1 mg l31). The N13C values of the species studied
in Horous›any deposit exhibit the same pattern.Therefore, it could be inferred that the plant spe-cies of Horous›any £ora were distributed from themost sea-in£uenced part of the salt marsh to themost-inland part of the marsh, along a decreasingsalinity gradient according to their N
13C values;the less 13C-depleted species occupying the mostsaline habitat. Such a variability of plant habitatcould, at least partially, explain the wide range ofplant N
13C values observed in Horous›any whencompared with Brn|¤k.The salinity gradient distribution of the plants
from Horous›any inferred from stable carbon iso-tope ratios thus helped in precising the taphonom-ic/palaeoecological conclusions previously estab-lished from palaeobotany and sedimentology. Ifthe few isotope values measured for the otherless abundant species are representative of theirpalaeoecology, then Ceratostrobus sequoiaphyllusand Dammarites albens might have been aswater-stressed as Frenelopsis alata and Eretmo-phyllum obtusum since they exhibit similar N
13Cvalues (Fig. 6). Therefore, even if C. sequoiaphyl-lus and D. albens are considered as rather al-lochthonous, they might have grown in salinehabitats.
3.2.3. Hloube›t|¤n section: salt-marsh depositsThe Hloube›t|¤n section, located at the northeast-
ern margin of Prague (Fig. 1), exposes a part of asuccession in¢lling a minor palaeovalley essen-tially parallel to that containing the Horous›anysection. The vertical succession of facies is gener-ally similar to that of Horous›any, but is thinnerand the £uvial deposits at the bottom representonly a minor portion of the whole succession (Fig.4). Several dark-grey mudstones levels, interpretedas supratidal salt marsh were recognised amongthe section (Hlus›t|¤k, 1986b). They yielded well-preserved, autochthonous fossil plants of purelyFrenelopsis assemblage including Alvinia bohemica(Velenovsky¤) J. Kvac›ek (Fig. 3C) and possibleovuliferous cone of Frenelopsis alata. In one ofthese supratidal marsh levels (Fig. 4), three specieswere sampled for isotope analyses: the coniferF. alata, the ginkgoalean plant Eretmophyllum ob-tusum and one dicotyledonous angiosperm, ‘Dio-
spyros’ cretacea. No channel sediments weresampled.
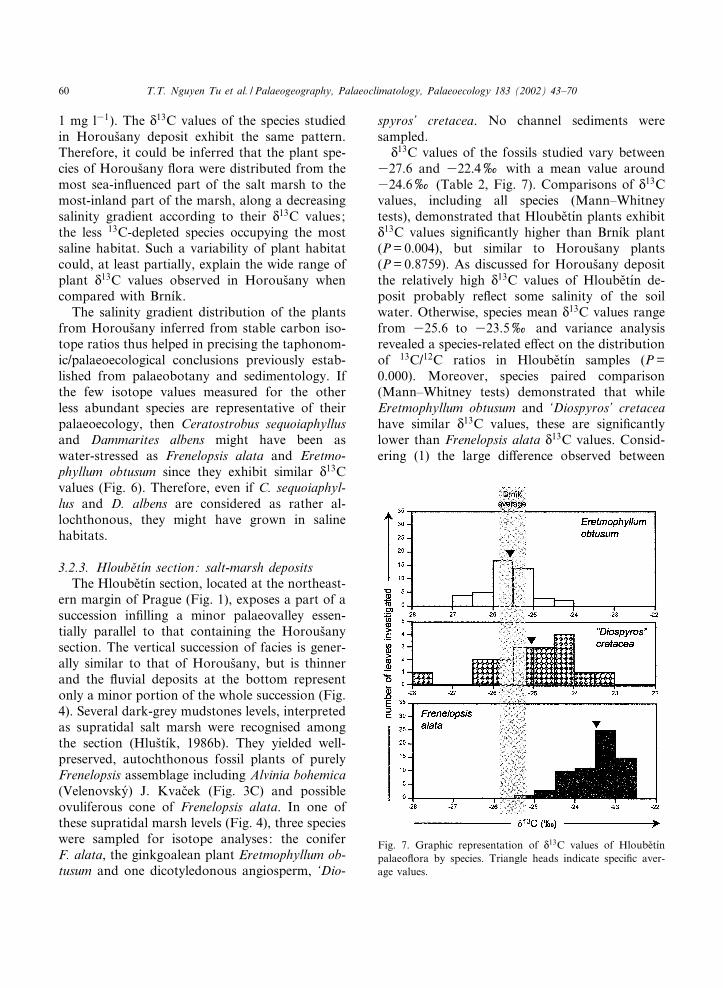
N13C values of the fossils studied vary between
327.6 and 322.4x with a mean value around324.6x (Table 2, Fig. 7). Comparisons of N13Cvalues, including all species (Mann^Whitneytests), demonstrated that Hloube›t|¤n plants exhibitN13C values signi¢cantly higher than Brn|¤k plant(P=0.004), but similar to Horous›any plants(P=0.8759). As discussed for Horous›any depositthe relatively high N
13C values of Hloube›t|¤n de-posit probably re£ect some salinity of the soilwater. Otherwise, species mean N
13C values rangefrom 325.6 to 323.5x and variance analysisrevealed a species-related e¡ect on the distributionof 13C/12C ratios in Hloube›t|¤n samples (P=0.000). Moreover, species paired comparison(Mann^Whitney tests) demonstrated that whileEretmophyllum obtusum and ‘Diospyros’ cretaceahave similar N
13C values, these are signi¢cantlylower than Frenelopsis alata N
13C values. Consid-ering (1) the large di¡erence observed between
Fig. 7. Graphic representation of N13C values of Hloube›t|¤n
palaeo£ora by species. Triangle heads indicate speci¢c aver-age values.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7060
these two sets of data (2x) and (2) the large sizeof samplings, it can be deduced from the afore-mentioned di¡erence that F. alata probably grewin more saline habitats than E. obtusum and ‘D.’cretacea. Otherwise, as suggested for Horous›any,the variability in salinity of plant habitats atHloube›t|¤n could, at least partially, explain thewide range of plant N
13C values (5.2x) com-pared with that of Brn|¤k (2.7x).
3.2.4. Deposit of Pec|¤novThe sedimentology and palaeontology of Pec|¤-
nov quarry, located ca. 40 km NW of Prague(Fig. 1), have been previously studied in detailand the locality serves as an informal referencesection for the Cenomanian succession in centralBohemia (e.g. Ulic›ny¤ and SVpic›a¤kova¤, 1996;Ulic›ny¤ et al., 1997). The lower part of the expo-sure, corresponding to the Peruc and KorycanyMembers, shows an overall deepening-upward fa-cies succession, composed of ¢ve lithological units(Fig. 4): (1) gravelly braided river, (2) tide-in£u-enced braided river, (3) supratidal marsh, (4) tidal£at and channel and (5) estuary mouth in¢ll(Ulic›ny¤ et al., 1997). Isotope analyses were per-formed on plant fossil samples taken from units 2,3 and 5.
3.2.4.1. Unit 2: tide-in£uence braided riverUnit 2, dominated by sandstones with muddy
interbeds, represents deposits of a large braidedriver with tidal in£uence (Ulic›ny¤ et al., 1997).Mudstone-dominated in¢lls of minor channelsyielded an allochtonous fossil £ora containinghigh number of leaves of lauraceous angiospermsMyrtophyllum geinitzii (Fig. 2D) and Grevilleo-phyllum constans (Velenovsky¤) Velenovsky¤ (Fig.2C). They are accompanied by Pseudoctenis sp.,bennettit Cycadolepsis sp. and in£orescence/in-fructescences Mauldinia bohemica Eklund and J.Kvac›ek; angiosperms occur more rarely (Platanuslaevis (Velenovsky¤) Velenovsky¤, Araliphyllumdaphnophyllum (Velenovsky¤) Velenovsky¤, ‘Myrica’zenkerii Ettingshausen, Monocotylophyllum sp.).Three angiosperms have been sampled for isotopeanalyses: P. laevis, G. constans and M. geinitzii. Itis suggested that the Myrtophyllum assemblagewas growing in a £ood plain of a braided river.
Myrtophyllum and Grevilleophyllum, because oftheir abundance, perhaps occupied niches nearthe river (unstable coasts, sandy bars, etc.), envi-ronments recently occupied by Salix sp. Platanusand other angiosperms, conifers and cycads couldbe con¢ned to river levees forming perhaps galleryforests (Ulic›ny¤ et al., 1997).
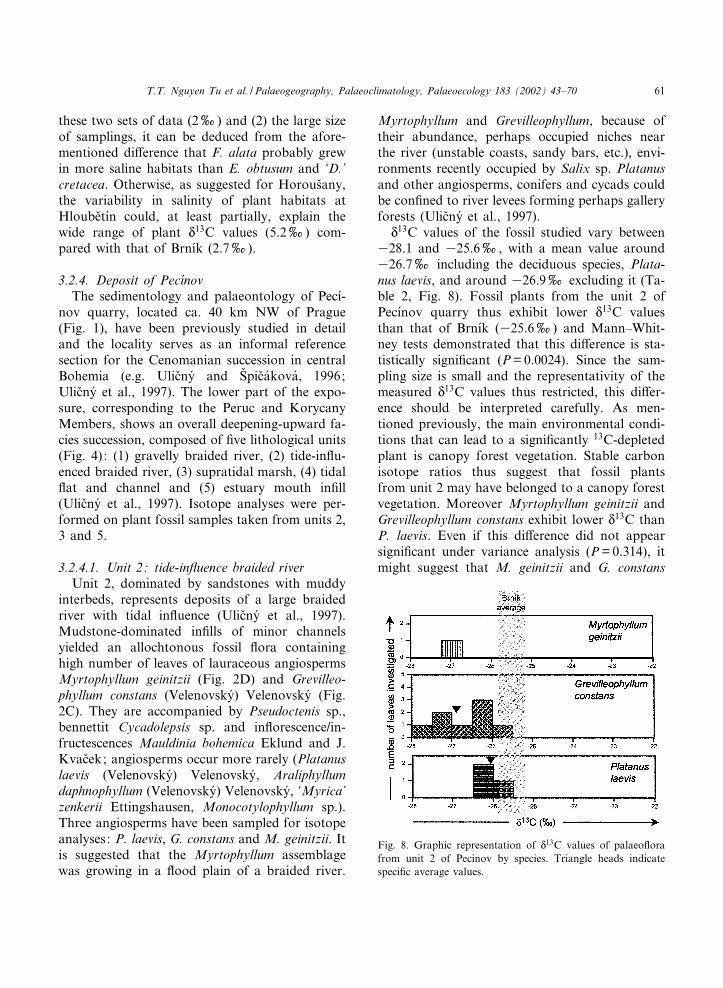
N13C values of the fossil studied vary between
328.1 and 325.6x, with a mean value around326.7x including the deciduous species, Plata-nus laevis, and around 326.9x excluding it (Ta-ble 2, Fig. 8). Fossil plants from the unit 2 ofPec|¤nov quarry thus exhibit lower N
13C valuesthan that of Brn|¤k (325.6x) and Mann^Whit-ney tests demonstrated that this di¡erence is sta-tistically signi¢cant (P=0.0024). Since the sam-pling size is small and the representativity of themeasured N
13C values thus restricted, this di¡er-ence should be interpreted carefully. As men-tioned previously, the main environmental condi-tions that can lead to a signi¢cantly 13C-depletedplant is canopy forest vegetation. Stable carbonisotope ratios thus suggest that fossil plantsfrom unit 2 may have belonged to a canopy forestvegetation. Moreover Myrtophyllum geinitzii andGrevilleophyllum constans exhibit lower N13C thanP. laevis. Even if this di¡erence did not appearsigni¢cant under variance analysis (P=0.314), itmight suggest that M. geinitzii and G. constans
Fig. 8. Graphic representation of N13C values of palaeo£ora
from unit 2 of Pec|¤nov by species. Triangle heads indicatespeci¢c average values.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 61
have grown near the soil, under a canopy formed,among others, by P. laevis. Indeed, in extant can-opy forest, the plants growing near the soil are13C-depleted when compared with canopy leaves(e.g. Medina and Minchin, 1980). Such a canopyvegetation is in agreement with previous palaeo-ecological interpretation (Ulic›ny¤ et al., 1997).
3.2.4.2. Unit 3: salt marshThis unit is mainly made up of grey to black
mudstones, occasionally including thin sand lam-inae, rich in pyrite concretions and locally con-taining broken bivalve shells. The environmentof unit 3, which contains abundant marine plank-ton, was interpreted as an intertidal to supratidalmarsh fringing an estuarine mouth (Ulic›ny¤ et al.,1997). Unit 3 of Pec|¤nov quarry yielded a fossil£ora similar to that described at the locality ofHorous›any: (1) an autochthonous Frenelopsis as-semblage almost exclusively constituted by Frene-lopsis alata and Eretmophyllum obtusum (Fig. 3)and occasionally yielding the angiosperm ‘Dio-spyros’ cretacea (Fig. 2E) and the enigmatic plantsDammarites albens and Pseudoasterophyllites cre-taceus and (2) a semi-autochthonous assemblagedominated by gymnosperms (e.g. Ceratostrobussequoiaphyllus, Fig. 2E; Cunninghamites lignitum,Fig. 2B; Sequoia crispa) but also containing someangiosperms (e.g. Cocculophyllum cinnamoneum(Velenovsky¤) ; ‘D.’ cretacea, Fig. 2E; Myrtophyl-lum genitizii, Fig. 2D) and pteridophytes (Ulic›ny¤et al., 1997). Eight species were sampled for iso-tope analyses: numerous specimens of F. alataand E. obtusum, as well as a few leaves ofS. crispa, C. lignitum, C. sequoiaphyllus, ‘D.’ cre-tacea, M. geinitzii and P. cretaceus. According toUlic›ny¤ et al. (1997), the Taxodiaceaes probablyconstituted a forest in the back-swamp with theangiosperms growing in the understorey whileF. alata and E. obtusum probably grew in thesalt marsh.
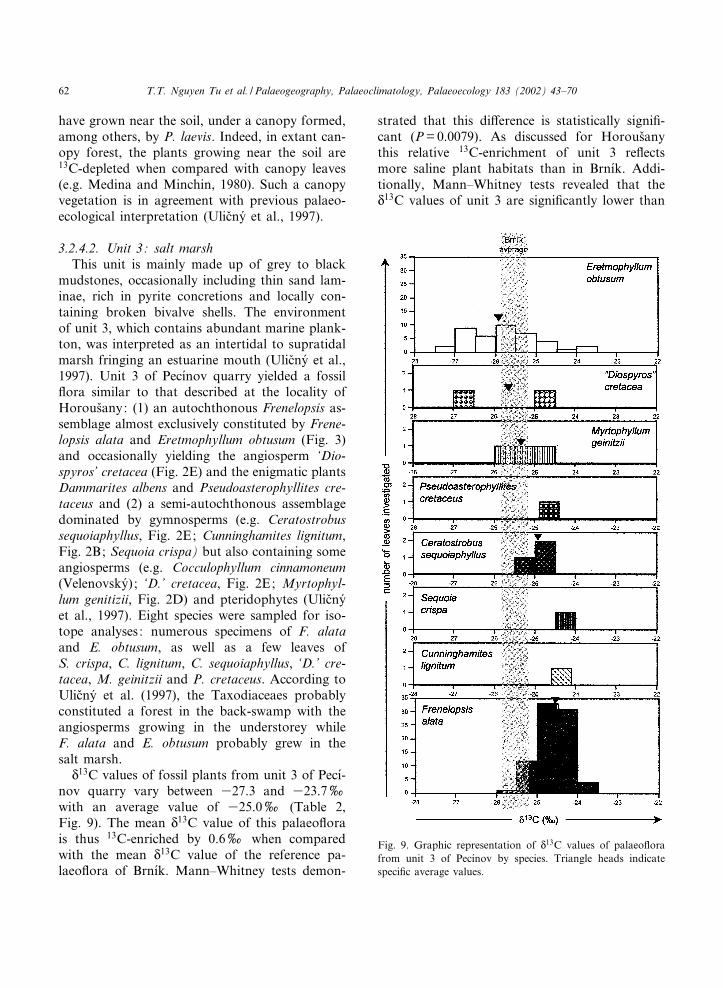
N13C values of fossil plants from unit 3 of Pec|¤-
nov quarry vary between 327.3 and 323.7xwith an average value of 325.0x (Table 2,Fig. 9). The mean N
13C value of this palaeo£orais thus 13C-enriched by 0.6x when comparedwith the mean N
13C value of the reference pa-laeo£ora of Brn|¤k. Mann^Whitney tests demon-
strated that this di¡erence is statistically signi¢-cant (P=0.0079). As discussed for Horous›anythis relative 13C-enrichment of unit 3 re£ectsmore saline plant habitats than in Brn|¤k. Addi-tionally, Mann^Whitney tests revealed that theN13C values of unit 3 are signi¢cantly lower than
Fig. 9. Graphic representation of N13C values of palaeo£ora
from unit 3 of Pec|¤nov by species. Triangle heads indicatespeci¢c average values.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7062
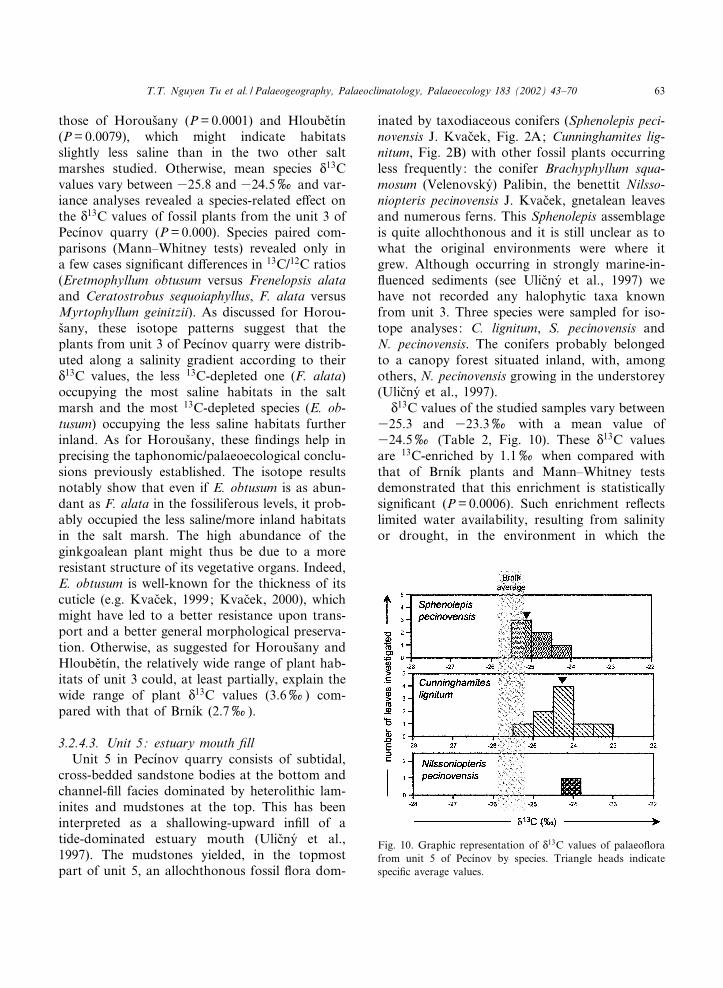
those of Horous›any (P=0.0001) and Hloube›t|¤n(P=0.0079), which might indicate habitatsslightly less saline than in the two other saltmarshes studied. Otherwise, mean species N
13Cvalues vary between 325.8 and 324.5x and var-iance analyses revealed a species-related e¡ect onthe N
13C values of fossil plants from the unit 3 ofPec|¤nov quarry (P=0.000). Species paired com-parisons (Mann^Whitney tests) revealed only ina few cases signi¢cant di¡erences in 13C/12C ratios(Eretmophyllum obtusum versus Frenelopsis alataand Ceratostrobus sequoiaphyllus, F. alata versusMyrtophyllum geinitzii). As discussed for Horou-s›any, these isotope patterns suggest that theplants from unit 3 of Pec|¤nov quarry were distrib-uted along a salinity gradient according to theirN13C values, the less 13C-depleted one (F. alata)occupying the most saline habitats in the saltmarsh and the most 13C-depleted species (E. ob-tusum) occupying the less saline habitats furtherinland. As for Horous›any, these ¢ndings help inprecising the taphonomic/palaeoecological conclu-sions previously established. The isotope resultsnotably show that even if E. obtusum is as abun-dant as F. alata in the fossiliferous levels, it prob-ably occupied the less saline/more inland habitatsin the salt marsh. The high abundance of theginkgoalean plant might thus be due to a moreresistant structure of its vegetative organs. Indeed,E. obtusum is well-known for the thickness of itscuticle (e.g. Kvac›ek, 1999; Kvac›ek, 2000), whichmight have led to a better resistance upon trans-port and a better general morphological preserva-tion. Otherwise, as suggested for Horous›any andHloube›t|¤n, the relatively wide range of plant hab-itats of unit 3 could, at least partially, explain thewide range of plant N
13C values (3.6x) com-pared with that of Brn|¤k (2.7x).
3.2.4.3. Unit 5: estuary mouth ¢llUnit 5 in Pec|¤nov quarry consists of subtidal,
cross-bedded sandstone bodies at the bottom andchannel-¢ll facies dominated by heterolithic lam-inites and mudstones at the top. This has beeninterpreted as a shallowing-upward in¢ll of atide-dominated estuary mouth (Ulic›ny¤ et al.,1997). The mudstones yielded, in the topmostpart of unit 5, an allochthonous fossil £ora dom-
inated by taxodiaceous conifers (Sphenolepis peci-novensis J. Kvac›ek, Fig. 2A; Cunninghamites lig-nitum, Fig. 2B) with other fossil plants occurringless frequently: the conifer Brachyphyllum squa-mosum (Velenovsky¤) Palibin, the benettit Nilsso-niopteris pecinovensis J. Kvac›ek, gnetalean leavesand numerous ferns. This Sphenolepis assemblageis quite allochthonous and it is still unclear as towhat the original environments were where itgrew. Although occurring in strongly marine-in-£uenced sediments (see Ulic›ny¤ et al., 1997) wehave not recorded any halophytic taxa knownfrom unit 3. Three species were sampled for iso-tope analyses: C. lignitum, S. pecinovensis andN. pecinovensis. The conifers probably belongedto a canopy forest situated inland, with, amongothers, N. pecinovensis growing in the understorey(Ulic›ny¤ et al., 1997).
N13C values of the studied samples vary between
325.3 and 323.3x with a mean value of324.5x (Table 2, Fig. 10). These N
13C valuesare 13C-enriched by 1.1x when compared withthat of Brn|¤k plants and Mann^Whitney testsdemonstrated that this enrichment is statisticallysigni¢cant (P=0.0006). Such enrichment re£ectslimited water availability, resulting from salinityor drought, in the environment in which the
Fig. 10. Graphic representation of N13C values of palaeo£orafrom unit 5 of Pec|¤nov by species. Triangle heads indicatespeci¢c average values.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 63

plants grew. Ulic›ny¤ et al. (1997) suggested thatthe deposition of the upper part of unit 5 markedthe onset of a long-term relative sea-level fall andthat the plant assemblages grew under falling con-ditions of groundwater table. However, no sedi-mentological argument can help in decidingwhether the 13C-enrichment of the plants is dueto salinity or drought. Otherwise, variance analy-ses revealed no species-related e¡ect (P=0.057),which tends to demonstrate that all the plantssampled grew in similar environments.
3.3. Implications
3.3.1. Isotope palaeoecology of Frenelopsis alataand Eretmophyllum obtusumContrary to the other species sampled, the
ginkgoalean plant Eretmophyllum obtusum andthe conifer Frenelopsis alata are very well repre-sented in three of the four deposits studied. Thesize of their sampling, at least 45 specimens col-lected for each species in each deposit, is largeenough allowing their N
13C values to be consid-ered as a meaningful representation of detailedenvironmental conditions.
Frenelopsis alata exhibits mean N13C values of
324.2 ] 0.6, 323.5 ] 0.6 and 324.6 ] 0.4x in thedeposits of Horous›any, Hloube›t|¤n and Pec|¤nov(unit 3), respectively. Indeed, Frenelopsis andPseudofrenelopsis genera are generally consideredas being adapted to various hydrically stressedenvironments (Upchurch and Doyle, 1981). Sev-eral studies tended to show that Frenelopsis andPseudofrenelopsis almost systematically occupycoastal environments, one of them being recon-structed as the modern halophyte Salicornia (e.g.Batten and McLennan, 1984; Watson, 1988).However, the ability of these genera to systemati-cally occupy saline environments can be discussed(e.g. Plaziat, 1995; Gomez et al., 2001). Neverthe-less, as shown in 3.2. Stable carbon isotope ratiosand local palaeoenvironments in the Bohemian£oras of the present paper, the relatively highN13C values con¢rm the ability of the species F.alata to occupy saline environments. Otherwise,F. alata mean N
13C values are di¡erent in thethree samples studied (Fig. 11) and sample-pairedcomparisons (Mann^Whitney tests) demonstrated
that these di¡erences are statistically signi¢cant inthe three cases. These di¡erences may thus re£ectsome di¡erences in the ecology of F. alata amongthe sections studied, especially for the Hloube›t|¤nsample where the Cheirolepidiaceae exhibit N
13Cvalues enriched by 1x when compared withHorous›any and Pec|¤nov (unit 3). Such a 13C-en-richment may re£ect more saline habitats for F.alata in Hloube›t|¤n than in the two other deposits.This last ¢nding tends to show that this coniferwas able to grow in soils of di¡erent salinities.However, within a given deposit, the intra-speci¢cvariability of F. alata N
13C values is narrower thanthe overall (i.e. including all the specimens of thisconifer from the three localities studied) intra-spe-ci¢c variability of F. alata N
13C values (Fig. 11).Therefore, the ability of F. alata to occupy anenvironment of di¡erent salinities does not seemto be linked to a tolerance to varying salinitieswithin a given deposit.
Eretmophyllum obtusum exhibits mean N13C val-
ues of 324.5 ] 0.7, 325.6 ] 0.6 and 325.8 ] 0.9xin the deposits of Horous›any, Hloube›t|¤n and Pec|¤-nov (unit 3), respectively. N13C values of E. obtu-sum are lower than those of Frenelopsis alata inthe three sections considered (Table 2). Althoughthis di¡erence remains small in the Horous›anysamples (0.3x), it may re£ect, at least for Hlou-be›t|¤n (2.1x) and Pec|¤nov (1.2x), less salinehabitats for E. obtusum when compared with F.alata. Otherwise, Mann^Whitney tests demon-strated that while E. obtusum N
13C values are sim-ilar in Hloube›t|¤n and in Pec|¤nov, they are signi¢-cantly lower than those of Horous›any deposit,which may re£ect a more saline habitat for theginkgoalean plant at Horous›any. Moreover, thewidth of E. obtusum leaves is signi¢cantly reducedat Horous›any (V9.8 mm) when compared withHloube›t|¤n (V12.5 mm) and Pec|¤nov (V15.8 mm)which tends to con¢rm that the Ginkgoaleanplant grew in more stressed conditions in Horou-s›any. As for F. alata these ¢ndings suggest that E.obtusum could be adapted to various soil-watersalinities. Here again, such an adaptation doesnot seem to be linked to an ability to occupy soilsof contrasted salinities within a given depositsince the intra-speci¢c variability of E. obtusumN13C values is narrower than the overall (i.e. all
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7064
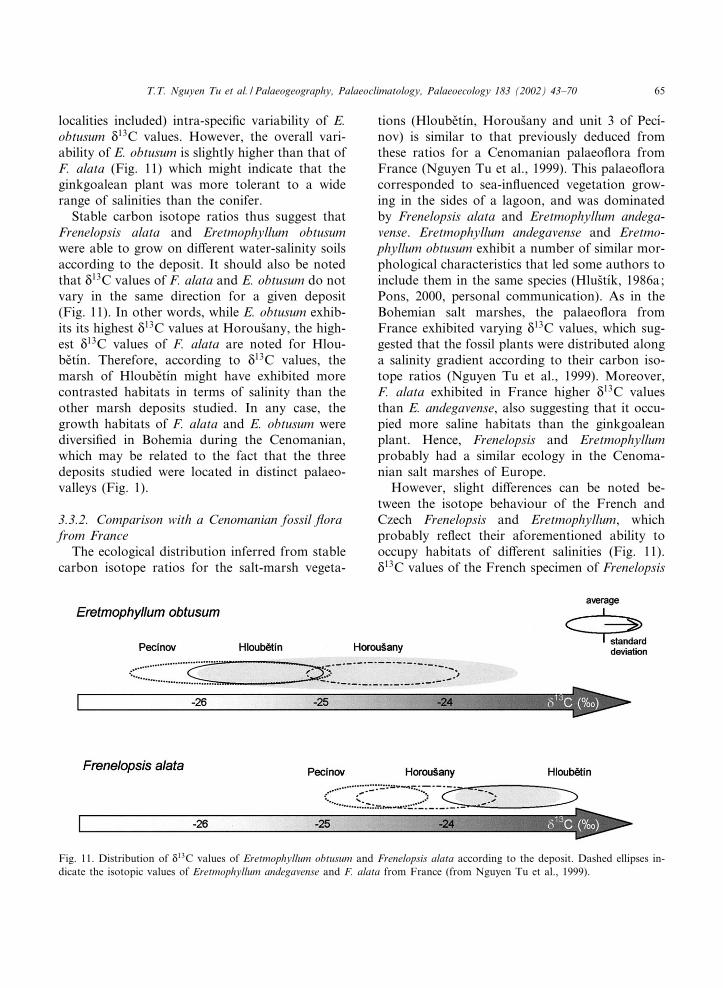
localities included) intra-speci¢c variability of E.obtusum N
13C values. However, the overall vari-ability of E. obtusum is slightly higher than that ofF. alata (Fig. 11) which might indicate that theginkgoalean plant was more tolerant to a widerange of salinities than the conifer.Stable carbon isotope ratios thus suggest that
Frenelopsis alata and Eretmophyllum obtusumwere able to grow on di¡erent water-salinity soilsaccording to the deposit. It should also be notedthat N13C values of F. alata and E. obtusum do notvary in the same direction for a given deposit(Fig. 11). In other words, while E. obtusum exhib-its its highest N13C values at Horous›any, the high-est N
13C values of F. alata are noted for Hlou-be›t|¤n. Therefore, according to N
13C values, themarsh of Hloube›t|¤n might have exhibited morecontrasted habitats in terms of salinity than theother marsh deposits studied. In any case, thegrowth habitats of F. alata and E. obtusum werediversi¢ed in Bohemia during the Cenomanian,which may be related to the fact that the threedeposits studied were located in distinct palaeo-valleys (Fig. 1).
3.3.2. Comparison with a Cenomanian fossil £orafrom FranceThe ecological distribution inferred from stable
carbon isotope ratios for the salt-marsh vegeta-
tions (Hloube›t|¤n, Horous›any and unit 3 of Pec|¤-nov) is similar to that previously deduced fromthese ratios for a Cenomanian palaeo£ora fromFrance (Nguyen Tu et al., 1999). This palaeo£oracorresponded to sea-in£uenced vegetation grow-ing in the sides of a lagoon, and was dominatedby Frenelopsis alata and Eretmophyllum andega-vense. Eretmophyllum andegavense and Eretmo-phyllum obtusum exhibit a number of similar mor-phological characteristics that led some authors toinclude them in the same species (Hlus›t|¤k, 1986a;Pons, 2000, personal communication). As in theBohemian salt marshes, the palaeo£ora fromFrance exhibited varying N
13C values, which sug-gested that the fossil plants were distributed alonga salinity gradient according to their carbon iso-tope ratios (Nguyen Tu et al., 1999). Moreover,F. alata exhibited in France higher N
13C valuesthan E. andegavense, also suggesting that it occu-pied more saline habitats than the ginkgoaleanplant. Hence, Frenelopsis and Eretmophyllumprobably had a similar ecology in the Cenoma-nian salt marshes of Europe.However, slight di¡erences can be noted be-
tween the isotope behaviour of the French andCzech Frenelopsis and Eretmophyllum, whichprobably re£ect their aforementioned ability tooccupy habitats of di¡erent salinities (Fig. 11).N13C values of the French specimen of Frenelopsis
Fig. 11. Distribution of N13C values of Eretmophyllum obtusum and Frenelopsis alata according to the deposit. Dashed ellipses in-dicate the isotopic values of Eretmophyllum andegavense and F. alata from France (from Nguyen Tu et al., 1999).
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 65

alata exhibit standard deviation and range of var-iation similar to that observed for F. alata withineach Bohemian section, with a mean value closeto that of Hloube›t|¤n specimens (Fig. 11). WhileEretmophyllum andegavense, the French fossilginkgo, was collected in a single deposit, it exhib-its a wider range of variation and a wider stan-dard deviation of its N13C values than Eretmophyl-lum obtusum within each Bohemian section. Thewide range of variation of isotope ratios in theFrench Ginkgo (V6.0x) was attributed to anability to occupy habitats of contrasted salinitysince (1) the other environmental parametersthat can induce such an intra-speci¢c variabilityimply large-size trees and (2) E. andegavense prob-ably did not constitute trees (Nguyen Tu et al.,1999). The standard deviation of E. andegavenseis similar to the overall standard deviation ofE. obtusum within all the Bohemian sectionstudied (Fig. 11). This ¢nding may (1) be due towider size and/or more diversi¢ed salinities of thecatchment basin collected by the deposition zoneor (2) re£ect slight di¡erences between the ecologyof E. andegavense and E. obtusum.
4. Conclusions
A number of isotope and biogeochemical pat-terns suggested that stable carbon isotope ratiosin the Cenomanian plants originating from Bohe-mia have not been signi¢cantly a¡ected by dia-genesis. Palaeoenvironments and palaeoecologyof the fossil plants studied could thus be inferredfrom N
13C values. These fossil plants originatedfrom four di¡erent deposits in the Bohemian Cre-taceous Basin (Brn|¤k, Horous›any, Hloube›t|¤n andPec|¤nov), in one of which three distinct strati-graphic levels could be sampled (Pec|¤nov). Inthe deposit of Brn|¤k, several sedimentologicaland palaeobotanical indicators suggested thatthe fossil £ora underwent no environmental stresswhich could have in£uenced its N
13C values. Sta-ble carbon isotope ratios of the palaeo£ora fromthe Brn|¤k deposit were thus used as reference val-ues for a non-stressed environment. Comparisonsof N13C values from the other deposits with thoseof Brn|¤k, allowed inference of palaeoenvironmen-
tal stresses undergone by fossil plants. The rela-tive 13C-enrichment of the plants from Hloube›t|¤n,Horous›any, unit 3 and unit 5 of Pec|¤nov revealedthat they grew in environments where water avail-ability was limited. The relative 13C depletion ofthe plants from the unit 2 of Pec|¤nov suggestedthat they grew under limited irradiance or re-cycled 13C-depleted respired CO2. These broadpalaeoenvironmental patterns deduced from13C/12C ratios are in agreement with the palaeoen-vironments previously reconstructed by sedimen-tological and palaeobotanical studies (e.g. Ulic›ny¤et al., 1997).Additionally, combination of isotope data and
palaeobotanical or sedimentological data pro-vided detailed insights into the palaeoecology ofthe fossil plants studied. The salt-marsh vegeta-tion of the deposits of Hloube›t|¤n, Horous›anyand unit 3 of Pec|¤nov exhibited, within each de-posit, a relatively wide range of N13C values. Theplants from these deposits may have been distrib-uted from the most sea-in£uenced part of themarsh to the more inland part of the marsh alonga decreasing salinity gradient according to theirN13C values, the most 13C-enriched plants occupy-ing the most saline habitats. Moreover the13C-depleted values of the back-land vegetationfrom unit 2 of Pec|¤nov deposit suggested thatthey belonged to a canopy forest vegetation.However, a combination of isotope data and pa-laeobotany could not provide additional informa-tion on the ecology of the fossil plants from unit 5of Pec|¤nov since no indicators allowed the deter-mination of whether their 13C-enrichment waslinked to drought or salinity. Nevertheless, stablecarbon isotope ratios also allowed re¢nement ofthe knowledge of ecology of the best-representedspecies of the salt-marsh deposits, the ginkgoaleanplant Eretmophyllum obtusum and the conifer Fre-nelopsis alata. Since F. alata systematically exhib-its higher N13C values than E. obtusum it was con-cluded that it occupied more saline environments.The wide range of 13C/12C ratios in F. alata andE. obtusum suggested that the marsh palaeoenvi-ronments of individual localities might have dif-fered in salinity. Finally, the palaeoecological¢ndings inferred from stable carbon isotope ratiosof the Bohemian fossil plants are in agreement
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7066

with that previously obtained with a similar ap-proach from marine-in£uence £ora in the Ceno-manian of France, also dominated by Eretmophyl-lum and Frenelopsis (Nguyen Tu et al., 1999).This study thus demonstrated that if (1) the
isotope signal has not been signi¢cantly a¡ectedby diagenesis and (2) a reference deposit of non-environmentally stressed plants is available, stablecarbon isotope ratios can allow determination ofthe kind of environmental stress undergone byfossil plant. Moreover, the combination of isotopedata with palaeobotanical or sedimentologicaldata can provide detailed insight into the palaeo-ecology of fossil plants.
Acknowledgements
We are grateful to M. Grably, C. Girardin andG. Bardoux for technical assistance. We would liketo sincerely thank J. Sakala for his assistance in thefieldwork. In particular we are grateful to thecompanies owning the quarries in which weundertook our research: CV eske¤ lupkove¤ za¤vodya.s., for granting access to the Pec|¤nov quarry, andKeramost a.s., for allowing us to work in the Brn|¤kand Horous›any quarries. We are also indebted toP.F. van Bergen, D. Pons, V. Millien, S. Derenne,C. Largeau, T. Bariac and J.-C. Plaziat for veryhelpful discussions. Many thanks to L. Guitard,G. Daniel and M. Roland for language correctionsof the manuscript. Thanks to the two anonymousreviewers who provided constructive reviews of thepaper. The study was supported by a grant fromCNRS (Fe¤de¤ration de Recherche en EcologieFondamentale et Applique¤e) and NEB.
References
Altabet, M.A., Deuser, W.G., Honjo, S., Stienen, C., 1991.Seasonal and depth-related changes in the source of sinkingparticles in the North Atlantic. Nature 354, 136^139.
Arens, N.C., Jahrens, A.H., Amundson, R., 2000. Can C3plants faithfully record the carbon isotopic composition ofatmospheric carbon dioxide? Palaeobiology 26, 137^164.
Aucourt, A.-M., Hillaire-Marcel, C., 1993. A 30 000 year rec-ord of 13C and 18O changes in organic matter from anequatorial peatbog. Geophys. Monogr. 78, 343^351.
Balesdent, J., Mariotti, A., 1996. Measurement of soil organicmatter turnover using 13C natural abundance. In: Boutton,T.W., Yamasaki, S.-I. (Eds.), Mass Spectrometry of Soils.Marcel Dekker, New York, pp. 83^111.
Barron, E.J., 1983. A warm, equable Cretaceous: the nature ofthe problem. Earth Sci. Rev. 19, 305^338.
Batten, D.J., 1974. Wealden palaeoecology from the distribu-tion of plant fossils. Proc. Geol. Assoc. 85, 433^458.
Batten, D.J., McLennan, A.M., 1984. The palaeoenvironmen-tal signi¢cance of the conifer family Cheirolepidiaceae in theCretaceous of Portugal. In: Reif, W.-E., Westphal, F.,(Eds.), Third Symposium of Mesozoic Terrestrial Ecosys-tems. S.P. Tubingen University Press, Tubingen, pp. 7^12.
Benner, R., Fogel, M.L., Sprague, E.K., Hodson, R.E., 1987.Depletion in 13C of lignin and its applications for stablecarbon isotope studies. Nature 329, 708^710.
van Bergen, P.F., Collinson, M.E., Hatcher, P.G., de Leeuw,J.W., 1994. Lithological control on the state of preservationof fossil seed coat of water plants. Org. Geochem. 22, 683^702.
Berner, R.A., 1990. Atmospheric carbon dioxide levels overphanerozoic times. Science 249, 1382^1386.
Berner, R.A., Landis, G.P., 1988. Gas bubbles in fossil amberas possible indicators of the major gas composition of an-cient air. Science 239, 1406^1409.
Bocherens, H., Friis, E.M., Mariotti, A., Pedersen, K.R., 1993.Carbon isotope abundances in Mesozoic and Cenozoicplants: palaeoecological implications. Lethaia 26, 347^358.
Bocherens, H., Mariotti, A., 1999. Carbon stable isotope anal-ysis of fossil plants. In: Jones, T.P., Rowe, N.P. (Eds.),Fossil Plants and Spores: Modern Techniques. The Geolog-ical Society, London, pp. 139^142.
Buchmann, N., Kao, W.-Y., Ehleringer, J., 1997. In£uence ofstand structure on carbon-13 of vegetation, soils and canopyair within deciduous and evergreen forests in Utah, UnitedStates. Oecologia 110, 109^119.
CV ech, S., Klein, V., Kr|¤z›, J., Valer›ka, J., 1980. Revision of theUpper Cretaceous stratigraphy of the Bohemian CretaceousBasin. Ve›stn. UŁ str›ed. UŁ stavu Geol. 55, 277^296.
Cerling, T.E., 1991. Carbon dioxide in the atmosphere: evi-dence from Cenozoic and Mesozoic paleosols. Am. J. Sci.29, 619^627.
Comstock, J.P., Ehleringer, J.R., 1992. Correlating genetic var-iations in carbon isotope composition with complex climaticchanges. Proc. Natl. Acad. Sci. USA 89, 7747^7751.
De Laune, R.D., 1986. The use of N13C signature of C-3 andC-4 plants in determining past depositional environments inrapidly accreting marshes of Mississippi river deltaic plain,Louisiana, USA. Chem. Geol. 59, 315^320.
De Niro, M.J., Hastorf, C.A., 1985. Alteration of 15N/14N and13C/12C ratios of plant matter during the initial stages ofdiagenesis: studies utilizing archeological specimens fromPeru. Geochim. Cosmochim. Acta 49, 97^115.
Degens, E.T., 1969. Biogeochemistry of stable carbon isotopes.In: Eglinton, G., Murphy, M.T. (Eds.), Organic Geochem-istry: Methods and Research. Springer, Berlin, pp. 304^329.
Deines, P., 1980. The isotope composition of reduced organic
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 67

carbon. In: Fritz, P., Fontes, J.C. (Eds.), Handbook of En-vironmental Isotope Geochemistry I. Elsevier, Amsterdam,pp. 329^406.
Dzurec, R.S., Boutton, T.W., Caldwell, M.M., Smith, B.N.,1985. Carbon isotope ratios of soil organic matter and theiruse in assessing community composition changes in CurlewValley, Utah. Oecologia 66, 17^24.
Ehleringer, J.R., 1991. 13C/12C fractionation studies and itsutility in terrestrial plant fractionation. In: Coleman, D.C.,Fry, B. (Eds.), Carbon Isotope Techniques. Academic Press,San Diego, CA, pp. 187^200.
Ehleringer, J.R., 1993. Carbon and water relation in desertplants: an isotope perspective. In: Ehleringer, J.R., Hall,A.E., Farquhar, G.D. (Eds.), Stable Isotopes and Plant Car-bon^Water Relations. Academic Press, San Diego, CA, pp.157^172.
Ehleringer, J.R., Cerling, T.E., 1995. Atmospheric CO2 andthe ratio of intercellular to ambient CO2 concentration inplants. Tree Physiol. 15, 105^111.
Ehleringer, J.R., Cooper, T.A., 1988. Correlations betweencarbon isotope ratios and microhabitats in desert plants.Oecologia 76, 562^566.
Farquhar, G.D., 1980. Carbon isotope discrimination byplants: E¡ects of carbon dioxide concentration and temper-ature via the ratio of intercellular and atmospheric CO2
concentrations. In: Pearman, G.I. (Ed.), Carbon Dioxideand Climate: Australian Research. Australian Academy ofScience, Canberra, pp. 105^110.
Farquhar, G.D., 1983. On the nature of isotope discriminationin C4 species. Aust. J. Plant Physiol. 9, 205^226.
Farquhar, G.D., O’Leary, M.H., Berry, J.A., 1982a. On therelationship between carbon isotope discrimination and theintercellular carbon dioxide concentration in leaves. Aust. J.Plant Physiol. 9, 121^137.
Farquhar, G.D., Ball, M.C., von Caemmerer, S., Roksandic,Z., 1982b. E¡ect of salinity and humidity on N
13C values ofHalophytes. Evidence for di¡usional isotope fractionationdetermined by the ratio of intercellular/atmospheric partialpressure of CO2 under di¡erent environmental conditions.Oecologia 52, 121^124.
Farquhar, G.D., Richards, R.A., 1984. Isotope composition ofplant carbon correlates with water-use e⁄ciency of wheatgenotypes. Aust. J. Plant Physiol. 11, 539^552.
Francey, R.D., Farquhar, G.D., 1982. An explanation of 13C/12C variation in tree rings. Nature 297, 28^31.
Francey, R.D., Gi¡ord, R.M., Sharkey, T.D., Weir, B., 1985.Physiological in£uences on carbon isotope discriminationand the intercellular carbon dioxide concentration in leaves.Aust. J. Plant Physiol. 9, 121^137.
Girardin, C., Mariotti, A., 1991. Analyse isotopique du 13C enabondance naturelle dans le carbone organique: un syste'meautomatique avec robot pre¤parateur. Cah. Orstom Se¤r. Pe¤-dol. XXVI, 371^380.
Globalview-CO2, 1988. Cooperative Atmospheric Data Inte-gration Project: Carbon Dioxide. CD-ROM, NOAA/CMDL, Boulder, CO.
Gomez, B., Mart|¤n-Closas, C., Me¤on, H., The¤venard, F., Bar-
ale, G., 2001. Plant taphonomy and palaeoecology in thelacustrine Un‹a delta (Late Barremian, Iberian Ranges,Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 170, 133^148.
Gro«cke, D.R., 1997. Carbon-isotope stratigraphy of terrestrialplant fragments in the early Cretaceous from south-easternAustralia. In: Wolberg, D.L., Stump, E., Rosenberg, G.D.(Eds.), Dinofest International. Academy of Natural Scien-ces, Philadelphia, PA, pp. 457^461.
Gro«cke, D.R., 1998. Carbon-isotope analyses of fossil plantsas a chemostratigraphic and palaeoenvironmental tool.Lethaia 31, 1^13.
Guy, R.D., Reid, D.M., Krouse, H.R., 1980. Shifts in carbonisotope ratios of two C3 halophytes under natural and arti-¢cial conditions. Oecologia 44, 241^247.
Guy, R.D., Reid, D.M., Krouse, H.R., 1986. Factors a¡ecting13C/12C ratios of inland halophytes. II. Ecophysiological in-terpretation in the ¢eld. Can. J. Bot. 64, 2700^2707.
Hanba, Y.T., Mori, S., Lei, T.L., Koike, T., Wada, E., 1997.Variations in leaf N13C along a vertical pro¢le of irradiancein a temperate Japanese forest. Oecologia 110, 253^261.
Hasegawa, T., 1997. Cenomanian^Turonian carbon isotopeevents recorded in terrestrial organic matter from northernJapan. Palaeogeogr. Palaeoclimatol. Palaeoecol. 130, 251^273.
Hlus›t|¤k, A., Konzalova¤, M., 1976. Frenelopsis alata (K.Feistm.) Knobloch (Cupressaceae) from the Cenomanianof Bohemia, a new plant producing Classopollis pollen.Evol. Biol. 5, 125^131.
Hlus›t|¤k, A., 1977. Remarks on Dammarites albens Presl. Ve›stn.UŁ str›ed. UŁ stavu Geol. 52, 359^366.
Hlus›t|¤k, A., 1986a. Eretmophyllous Ginkgoalean plants fromthe Cenomanian. Sborn. Na¤rod. Muz. Praze B 42, 99^114.
Hlus›t|¤k, A., 1986b. A disappearing palaeobotanical locality inPrague-Hloube›t|¤n. Cas. Na¤rod. Muz. Praze 154, 129^141.
Huang, Y., Eglinton, G., Ineson, P., Latter, P.M., Bol, R.,Harkness, D., 1997. Absence of carbon isotope fractionationof individual n-alkanes in a 23-years decomposition experi-ment with Calluna vulgaris. Org. Geochem. 26, 497^501.
Jones, T.P., 1994. 12C-enriched Lower Carboniferous fossilplant from Donegal, Ireland: carbon isotope constraintson taphonomy, diagenesis and palaeoenvironments. Rev.Palaeobot. Palynol. 81, 53^64.
Kloeppel, B.D., Gower, S.T., Treichel, I.W., Kharuk, S., 1998.Foliar carbon isotope discrimination in Larix species andsympatric evergreen conifers: a global comparison. Oecolo-gia 114, 153^159.
Knobloch, E., 1999. Neue oder wenig bekannte Planzenartenaus den Perucer Schichten (Cenoman) der Bo«hmichenMasse. Acta Mus. Nat. Pragae B Hist. Nat. 55, 25^60.
Knoll, A.H., Walter, M.R., 1992. Latest Proterozoic stratigra-phy and Earth history. Nature 356, 673^678.
Kvac›ek, J., 1995. Cycadales and Bennetitales leaf compressionsof the Bohemian Cenomanian, Central Europe. Rev. Paleo-bot. Palynol. 84, 389^412.
Kvac›ek, J., 1997. Sphenolepis pecinovensis so. nov., a new tax-odiaceous conifer from the Bohemian Cenomanian, Central
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7068

Europe. Meded. Neth. Inst. Toegep. Geowet. TNO 58, 121^129.
Kvac›ek, J., 1998. Cuticles Analysis of the Gymnosperms of theBohemian Cenomanian. Ph.D. thesis, National Museum ofPrague.
Kvac›ek, J., 1999. New data and revision of three gymnospermsfrom the Cenomanian of Bohemia: Segenopteris variabilis(Velenovsky¤) Velenovsky¤, Mesenea bohemica (Corda)comb. n. and Eretmophyllum obtusum (Velenovsky¤) comb.n.. Acta Mus. Natl. Pragae B 55, 15^24.
Kvac›ek, J., 2000. Frenelopsis alata and its microsporangiateand ovuliferous reproductive structures from the Cenoma-nian of Bohemia (Czech Republic, Central Europe). Rev.Palaeobot. Palynol. 112, 51^78.
Kvac›ek, J., Dilcher, D.L., 2000. Comparison of CenomanianFloras from Western Interior North America and CentralEurope. Acta Univ. Carol. Geol. 44, 17^38.
Kvac›ek, J., Knobloch, E., 1997. Representatives of the genusNilsonia Brongniart from the Cenomanian of the BohemianMassif (Czech Republic, Central Europe). Rev. Palaeobot.Palynol. 97, 41^52.
Kvac›ek, J., Spicer, R.A., Herman, A.B., 2000. Palaeoclimaticparameters based on the Peruc^Korycany Flora and its re-lationship to other Laurasian Cenomanian £oras. 6th Conf.Int. Org. Palaeobot., Qinhuangdao, China. Abstract, pp.69^70.
Kvac›ek, Z., 1983. Cuticular studies in Angiosperms of theBohemian Cenomanian. Acta Palaeontol. Polon. 28, 159^170.
Lallier-Verge's, E., Perrussel, B.P., Disnar, J.-R., Baltzer, F.,1998. Relationships between environmental conditions andthe diagenetic evolution of organic matter derived fromhigher plants in a modern mangrove swamp system (Gua-deloupe, French West Indies). Org. Geochem. 29, 1663^1686.
Lasaga, A.C., Berner, R.A., Garrels, R.M., 1985. An improvedmodel of atmospheric CO2 £uctuations over the past 100million years. In: Sundquist, E.T., Broecker, W.S. (Eds.),The Carbon Cycle and Atmospheric CO2 : Natural Varia-tions Archean to Present. American Geophysical Union,Washington, DC, pp. 397^411.
Lipp, J., Trimborn, P., Edwards, T., Waisel, Y., Yakir, D.,1996. Climatic e¡ects on the N
18O and N13C of cellulose in
the desert tree Tamarix jordanis. Geochim. Cosmochim.Acta 60, 3305^3309.
Lu«cke, A., Helle, G., Schleser, G.H., Figueiral, I., Mosbrug-ger, V., Jones, T.P., Rowe, N., 1999. Environmental historyof the German Lower Rhine Embayment during the MiddleMiocene as re£ected by carbon isotopes in brown coal. Pa-laeogeogr. Palaeoclimatol. Palaeoecol. 154, 339^352.
Marino, B.D., Mac Elroy, M.B., 1991. Isotope composition ofatmospheric CO2 inferred from carbon in C4 plant cellulose.Nature 343, 127^131.
Medina, E., Minchin, P., 1980. Strati¢cation of N13C values ofleaves in Amazonian rain forest. Oecologia 45, 377^378.
Mosbrugger, V., Gee, C.T., Belz, G., Ashraf, A.R., 1994.Three-dimensional reconstruction of an in-situ Miocene
peat forest from the Lower Rhine Embayment, northwestGermany-new methods in palaeovegetation analysis. Palae-ogeogr. Palaeoclimatol. Palaeoecol. 110, 295^317.
Nambudiri, E.M.V., Tidwell, W.D., Smith, B.N., Hebber,N.P., 1978. A C4 plant from the Pliocene. Nature 276,816^817.
Nguyen Tu, T.T., Bocherens, H., Mariotti, A., Baudin, F.,Pons, D., Broutin, J., Derenne, S., Largeau, C., 1999. Eco-logical distribution of Cenomanian terrestrial plants basedon 13C/12C ratios. Palaeogeogr. Palaeoclimatol. Palaeoecol.145, 79^93.
Nielsen, E.K., Shari¢, M.R., 1998. Carbon isotope composi-tion of legumes with photosynthetic stems from Mediterra-nean and desert habitats. Am. J. Bot. 84, 1707^1713.
O’Leary, M.H., 1981. Carbon isotope fractionation in plants.Phytochemistry 20, 553^567.
Pacltova¤, B., 1965. Uº ber den Nachweis mariner Beein£ussunfdes oberen Teils der Perucer Schiten (Cenoman) in Nord-bo«hmen. Geologie 14, 892.
Pacltova¤, B., 1971. Palynological study of angiospermae fromPeruc Formation (?Albian - Lower Cenomanian) of Bohe-mia. Sborn. UŁ str›. UŁ st. Geol. 13, 105^141.
Pacltova¤, B., 1977. Cretaceous angiosperms of Bohemia. Bot.Rev. 43, 128^142.
Pacltova¤, B., 1990. Marginal facies of the Bohemian UpperCretaceous (Palynological study). Proc. Symp. Palaeo£oris-tic Changes in the Cretaceous and Tertiary, Prague, 1989.pp. 47^52.
Park, R., Epstein, S., 1960. Carbon isotope fractionation dur-ing photosynthesis. Geochim. Cosmochim. Acta 21, 110^126.
Pearman, G.I., Francey, R.J., Fraser, P.J.B., 1976. Climaticimplications of stable isotopes in tree rings. Nature 260,771^772.
Pelzer, G., Riegel, W., Wilde, V., 1992. Depositional controlson the Lower Cretaceous Wealden coals of the northernGermany. Geol. Soc. Am. 267, 227^244.
Pendall, E., Betancourt, J.L., Leavitt, S.W., 1999. Paleocli-matic signi¢cance of ND and N
13C values in pin‹on needlesfrom packrat middens spanning the last 40 000 years. Pa-laeogeogr. Palaeoclimatol. Palaeoecol. 147, 53^72.
Pen‹uelas, J., Estiarte, M., 1997. Trends in plant carbon con-centration and plant demand for N throughout this century.Oecologia 109, 69^73.
Plaziat, J.-C., 1995. Modern and fossil mangrove and mangals:their climatic and biogegraphic variability. In: Bosence,D.W., Allison, A. (Eds.), Marine PalaeoenvironmentalAnalysis from Fossils. Geological Society Special Publica-tions, London, pp. 73^96.
Popp, B.N., Takigiku, R., Hayes, J.M., Louda, J.W., Baker,E.W., 1989. The post-Paleozoic chronology and mechanismof 13C-depletion in primary marine organic matter. Am. J.Sci. 289, 436^454.
Raven, J.A., 1998. Phylogeny, palaeoatmospheres and the evo-lution of phototrophy. In: Gri⁄ths, G. (Ed.), Stable Iso-topes, Integration of Biological, Ecological and GeochemicalProcesses. Bios Scienti¢c Publisher, Oxford, pp. 323^346.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^70 69

Rice, S., 1999. Variability in carbon isotope discrimination inSphagnum. XVIth International Botanical Congress, August,St. Louis, MO. Abstract No. 5180.
Rigby, D., Batts, B.D., Smith, J.W., 1981. The e¡ect of mat-uration on the isotope composition of fossil fuels. Org. Geo-chem. 3, 29^36.
Rundgren, N., Loader, N.J., Beerling, D.J., 2000. Variation inthe carbon isotope composition of late-Holocene plant mac-rofossils: a comparison of whole-leaf and cellulose trends.Holocene 10, 149^154.
Sakata, M., Suzuki, K., 1998. Assessment method for environ-mental stresses in tree using N
13C records of annual growthrings. Geochem. J. 32, 331^338.
Sarkanen, K.V., Ludwig, C.H., 1971. Lignins. Wiley Inter-science, New York.
Schleser, G.H., 1990. Investigation of the N13C pattern in
leaves of Fagus sylvatica L.. J. Exp. Bot. 41, 565^572.Smith, B.N., Epstein, S., 1971. Two categories of 13C/12C ra-tios for higher plants. Plant Physiol. 47, 380^384.
Smith, B.N., Oliver, J., McMillan, C., 1976. In£uence of car-bon source, oxygen concentration, light intensity, and tem-perature on 13C/12C ratios in plant tissues. Bot. Gaz. 137,99^104.
SVpic›a¤kova¤, L., 1999. Fluvial deposits of the southwestern mar-gin of the Bohemian Cretaceous Basin (Cenomanian, Czechrepublic), their spatial and temporal evolution. Ph.D. thesis,Charles University, Prague (in Czech).
SVpic›a¤kova¤, L., Ulic›ny¤, D., 1996. Eustatic and tectonic controlson £uvial sedimentation, Cenomanian of central Bohemia.In: Abstracts, 5th International Cretaceous Symposium and2nd Workshop on Inoceramids, Freiberg, Sept. 16^24, 1996.
Sternberg, K.M., 1820. Versuch einer geognotisch-botanischenDarstellung der Flora der Vorwelt, vol. I. Fleischer, Leipzig,pp. 1^24.
Stuiver, M., Braziunas, T.F., 1987. Tree cellulose 13C/12C ra-tios and climatic changes. Nature 328, 58^60.
Svobodova¤, M., 1995. Palynology and stratigraphy of Peruc^Korycany Formation of Pec|¤nov-Bab|¤n, Cenomanian,Czechoslovakia. Acta Univ. Carol. Geol. 1992, 89^95.
Svobodova¤, M., Me¤on, H., Pacltova¤, B., 1998. Characteristicsof palynospectra of the Upper Cenomanian-Lower Turonian(anoxic facies) of the Bohemian and Vocontian Basins. Ve›st.CV es. Geol. UŁ st. 73, 229^251.
Tomlinson, P.B., 1994. The Botany of Mangroves. CambridgeUniversity Press, Cambridge.
Troughton, J.H., 1972. Carbon isotope fractionation by plants.Proceedings of the 8th International Congress on Radiocar-bon. The Royal Society of New-Zealand, Wellington, pp.420^457.
Ulic›ny¤, D., 1997. Sedimentation in a reactivated intra-conti-nental strike-slip fault zone: the Bohemian Cretaceous Ba-sin, Central Europe. Abstracts, 18th IAS Regional EuropeanMeeting, September 2^4, 1997, Heidelberg. Gaea heidelber-gensis 3, 347.
Ulic›ny¤, D., SVpic›a¤kova¤, L., 1996. Response to high frequency
sea-level change in £uvial to estuarine succession: Cenoma-nian palaeovalleys ¢ll, Bohemian Cretaceous basin. In: Ho-well, J.A., Aiken, J.F. (Eds.), High Resolution SequenceStratigraphy: Innovations and Applications. Geological So-ciety, London, pp. 247^268.
Ulic›ny¤, D., Kvac›ek, J., Svobodova¤, M., SVpic›a¤kova¤, L., 1997.High-frequency sea-level £uctuations and plant habitats inCenomanian £uvial to estuarine succession: Pec|¤nov quarry,Bohemia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 136,165^197.
Upchurch, G.R., Doyle, J.A., 1981. Paleoecology of the coni-fers Frenelopsis and Pseudofrenelopsis (Cheirolepidiaceae)from the Cretaceous Potomac Group of Maryland and Vir-ginia. In: Romans, R.C. (Ed.), Geobotany II. Plenum, NewYork, pp. 192^196.
Velenovsky¤, J., 1882. Die Flora der bo«hmischen Kreideforma-tion. I. Beitr. Pala«ontol. Geol. Oº ster-Ung. Orient. 2, 8^32.
Velenovsky¤, J., 1883. Die Flora der bo«hmischen Kreideforma-tion. II. Beitr. Pala«ontol. Geol. Oº ster-Ung. Orient. 3, 1^22.
Velenovsky¤, J., 1884. Die Flora der bo«hmischen Kreideforma-tion. III. Beitr. Pala«ontol. Geol. Oº ster-Ung. Orient. 4, 1^14.
Velenovsky¤, J., 1885a. Die Flora der bo«hmischen Kreidefor-mation. IV. Beitr. Pala«ontol. Geol. Oº ster-Ung. Orient. 5, 1^14.
Velenovsky¤, J., 1885b. Die Gymnospermen der bo“hmischenKreideformation. Greger, Prague.
Velenovsky¤, J., 1888a. Die Fa«rne der bo«hmischen Kreidefor-mation. Abh. Kgl. Bo«hm. Ges. Wiss. 2, 1^32.
Velenovsky¤, J., 1888b. Uº ber einige neue P£anzenformen derbo«mischen Kreideformation. Sistzungsb. Ber. K. Bo«hm.Ges. Wiss. Math.-Nat. Kl. 1887, 590^593.
Velenovsky¤, J., 1889. Kve›tena CV eske¤ho Cenomanu. Abh. Kgl.Bo«hm. Ges. Wiss. 3, 1^75.
Velenovsky¤, J., Vinikla¤r, L., 1926. Flora Cretacea Bohemiae I.Rozpr. Sta¤tn. Geol. UŁ stav. CV eske¤ Rep. 1, 1^57.
Velenovsky¤, J., Vinikla¤r, L., 1927. Flora Cretacea Bohemiae II.Rozpr. Sta¤tn. Geol. UŁ stav. CV eske¤ Rep. 2, 1^54.
Velenovsky¤, J., Vinikla¤r, L., 1929. Flora Cretacea BohemiaeIII. Rozpr. Sta¤tn. Geol. UŁ stav. CV eske¤ Rep 1^52.
Velenovsky¤, J., Vinikla¤r, L., 1931. Flora Cretacea BohemiaeIV. Rozpr. Sta¤tn. Geol. UŁ stav. CV eske¤ Rep 1^111.
Vogel, J.C., 1978. Recycling of carbon in a forest environment.Oecol. Plant. 13, 89^94.
Waring, R.H., Silvester, W.B., 1994. Variation in foliar N13C
values within the crowns of Pinus radiata trees. Tree Physiol.14, 1203^1213.
Watson, J., 1988. The Cheirolepidiaceae. In: Beck, C.B. (Ed.),Origin and Evolution of Gymnosperms. University Press,New York, pp. 382^447.
Ziegler, P.A., 1990. Geological Atlas of Western and CentralEurope. Shell Internationale Petroleum Maatschappij, TheHague.
Zimmerman, J.K., Ehleringer, J.R., 1990. Carbon isotope ra-tios are correlated with irradiance levels in the Panamianorchid Catasetum viridi£avum. Oecologia 83, 247^249.
PALAEO 2802 30-5-02
T.T. Nguyen Tu et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 183 (2002) 43^7070
Related Documents











