Isomer-specific regulation of differentiating pig preadipocytes by conjugated linoleic acids 1 T. D. Brandebourg* and C. Y. Hu† 2 *Department of Animal Sciences and †College of Agricultural Sciences, Oregon State University, Corvallis 97331 ABSTRACT: Conjugated linoleic acids are a group of geometric and positional isomers of linoleic acid that decrease body fat in growing animals by a poorly under- stood mechanism. The objective of this study was to investigate the isomer-specific effect of CLA on the pro- liferation and differentiation of pig preadipocytes in primary culture. The effect of CLA on preadipocyte pro- liferation was determined using cleavage of the tetrazo- lium salt, WST-1, as a marker for proliferation. Preadi- pocyte number was decreased in a dose-dependent fash- ion by trans-12,cis-10 CLA (P < 0.05). No other fatty acid affected preadipocyte number. Differentiation was monitored on d 10 after induction morphologically, en- zymatically, and by measuring the mRNA abundance of key adipogenic transcription factors. Both a crude CLA preparation containing a mixture of CLA isomers (CLA-mix) and the pure trans-10,cis-12 CLA isomer inhibited glycerol-3-phosphate dehydrogenase (GPDH) activity in a dose-dependent fashion, with trans-10,cis- 12 CLA being more potent (P < 0.01) than the CLA-mix. Cis-9,trans-11 CLA failed to decrease GPDH activity; however, increasing concentrations of cis-9,trans-11 CLA tended to blunt the inhibitory effect of trans-10,cis- 12 CLA on GPDH activity (P < 0.09), suggesting that Key Words: Adipogenesis, Chicken Ovalbumin Upstream Promoter Transcription Factor 1, Conjugated Linoleic Acid, Primary Culture, Swine 2005 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2005. 83:2096–2105 Introduction Conjugated linoleic acids are naturally occurring iso- mers of linoleic acid that are associated with antiathero- genic, antidiabetic, and antitumorigenic action in mam- mals (reviewed by Pariza et al., 2000; Brown and McIn- 1 The authors thank Drahn Acres Farms for their assistance in support of this work and A. Menino for technical discussions. 2 Correspondence: Univ. of Hawaii at Manoa, 3050 Maile Way, Gilmore Hall 202, Honolulu 96822-2279 (phone: 808-956-8131; fax: 808-956-9105; e-mail: [email protected]). Received February 7, 2005. Accepted June 2, 2005. 2096 cis-9,trans-11 CLA may antagonize the action of trans- 10,cis-12 CLA in porcine adipocytes. Finally, the iso- mer-specific effect of CLA on adipogenic transcription factor gene expression was investigated. Trans-10,cis- 12 CLA decreased expression of peroxisome prolifera- tor-activated receptor γ (PPARγ; P < 0.01) and sterol regulatory element-binding protein-1c (SREBP-1c; P < 0.05) mRNA, while failing to alter the expression of CCAAT/enhancer binding protein α (C/EBPα) mRNA. Interestingly, both the CLA-mix and the trans-10,cis-12 CLA isomer increased the mRNA abundance of chicken ovalbumin upstream promoter transcription factor 1 (COUP-TF; P < 0.002). No other fatty acid affected COUP-TF mRNA levels. Collectively these data sup- port the concept that CLA decreases fat accretion in pigs, in part by inhibiting preadipocyte proliferation and differentiation, with trans-10,cis-12 CLA being an active isomer eliciting these effects. Furthermore, trans-10,cis-12 CLA inhibits porcine preadipocyte dif- ferentiation by a mechanism that involves the down- regulation of PPARγ and SREBP-1c mRNA. This mech- anism is independent of changes in C/EBPα mRNA abundance and may involve COUP-TF. tosh, 2003). Several trials suggest that pigs fed CLA deposit less fat and have improved body composition (Dugan et al., 1997; Azain, 2003; Dugan, 2004). Thus, CLA holds great promise as a feed additive in swine diets. Fat deposition in swine results from the additive con- tributions of an increase in adipocyte number and size. Although CLA can limit adipose tissue accretion in growing pigs, the underlying mechanisms are poorly understood. In vitro studies utilizing 3T3-L1 preadipo- cytes suggest that CLA may limit adipocyte number through inhibitory actions on both preadipocyte prolif- eration and differentiation (Brodie et al., 1999; Evans et al., 2001; Kang et al., 2003). Although hyperplasia of human primary preadipocytes is potently inhibited

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Isomer-specific regulation of differentiating pig preadipocytesby conjugated linoleic acids1
T. D. Brandebourg* and C. Y. Hu†2
*Department of Animal Sciences and †College of Agricultural Sciences,Oregon State University, Corvallis 97331
ABSTRACT: Conjugated linoleic acids are a group ofgeometric and positional isomers of linoleic acid thatdecrease body fat in growing animals by a poorly under-stood mechanism. The objective of this study was toinvestigate the isomer-specific effect of CLA on the pro-liferation and differentiation of pig preadipocytes inprimary culture. The effect of CLA on preadipocyte pro-liferation was determined using cleavage of the tetrazo-lium salt, WST-1, as a marker for proliferation. Preadi-pocyte number was decreased in a dose-dependent fash-ion by trans-12,cis-10 CLA (P < 0.05). No other fattyacid affected preadipocyte number. Differentiation wasmonitored on d 10 after induction morphologically, en-zymatically, and by measuring the mRNA abundanceof key adipogenic transcription factors. Both a crudeCLA preparation containing a mixture of CLA isomers(CLA-mix) and the pure trans-10,cis-12 CLA isomerinhibited glycerol-3-phosphate dehydrogenase (GPDH)activity in a dose-dependent fashion, with trans-10,cis-12 CLA being more potent (P < 0.01) than the CLA-mix.Cis-9,trans-11 CLA failed to decrease GPDH activity;however, increasing concentrations of cis-9,trans-11CLA tended to blunt the inhibitory effect of trans-10,cis-12 CLA on GPDH activity (P < 0.09), suggesting that
Key Words: Adipogenesis, Chicken Ovalbumin Upstream Promoter Transcription Factor 1,Conjugated Linoleic Acid, Primary Culture, Swine
2005 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2005. 83:2096–2105
Introduction
Conjugated linoleic acids are naturally occurring iso-mers of linoleic acid that are associated with antiathero-genic, antidiabetic, and antitumorigenic action in mam-mals (reviewed by Pariza et al., 2000; Brown and McIn-
1The authors thank Drahn Acres Farms for their assistance insupport of this work and A. Menino for technical discussions.
2Correspondence: Univ. of Hawaii at Manoa, 3050 Maile Way,Gilmore Hall 202, Honolulu 96822-2279 (phone: 808-956-8131; fax:808-956-9105; e-mail: [email protected]).
Received February 7, 2005.Accepted June 2, 2005.
2096
cis-9,trans-11 CLA may antagonize the action of trans-10,cis-12 CLA in porcine adipocytes. Finally, the iso-mer-specific effect of CLA on adipogenic transcriptionfactor gene expression was investigated. Trans-10,cis-12 CLA decreased expression of peroxisome prolifera-tor-activated receptor γ (PPARγ; P < 0.01) and sterolregulatory element-binding protein-1c (SREBP-1c; P <0.05) mRNA, while failing to alter the expression ofCCAAT/enhancer binding protein α (C/EBPα) mRNA.Interestingly, both the CLA-mix and the trans-10,cis-12CLA isomer increased the mRNA abundance of chickenovalbumin upstream promoter transcription factor 1(COUP-TF; P < 0.002). No other fatty acid affectedCOUP-TF mRNA levels. Collectively these data sup-port the concept that CLA decreases fat accretion inpigs, in part by inhibiting preadipocyte proliferationand differentiation, with trans-10,cis-12 CLA being anactive isomer eliciting these effects. Furthermore,trans-10,cis-12 CLA inhibits porcine preadipocyte dif-ferentiation by a mechanism that involves the down-regulation of PPARγ and SREBP-1c mRNA. This mech-anism is independent of changes in C/EBPα mRNAabundance and may involve COUP-TF.
tosh, 2003). Several trials suggest that pigs fed CLAdeposit less fat and have improved body composition(Dugan et al., 1997; Azain, 2003; Dugan, 2004). Thus,CLA holds great promise as a feed additive in swinediets.
Fat deposition in swine results from the additive con-tributions of an increase in adipocyte number and size.Although CLA can limit adipose tissue accretion ingrowing pigs, the underlying mechanisms are poorlyunderstood. In vitro studies utilizing 3T3-L1 preadipo-cytes suggest that CLA may limit adipocyte numberthrough inhibitory actions on both preadipocyte prolif-eration and differentiation (Brodie et al., 1999; Evanset al., 2001; Kang et al., 2003). Although hyperplasiaof human primary preadipocytes is potently inhibited

CLA in pig adipogenesis 2097
in vitro by the trans-10,cis-12 CLA isomer (Brown etal., 2003), it has been reported that CLA fails to inhibitthe proliferation and differentiation of pig preadipo-cytes, and it may even stimulate the differentiation ofthese cells (Ding et al., 2000; McNeel and Mersmann2003). Given that a stimulatory effect of CLA on adipo-genesis seems at odds with the ability of CLA to de-crease carcass adiposity, the effect of CLA on the hyper-plasia of adipocytes in pigs remains controversial. Ourobjective was to further characterize the isomer-specificeffect of CLA on the proliferation and differentiation ofpig preadipocytes in primary culture.
Materials and Methods
Materials
Linoleic acid was purchased from Sigma-Aldrich (St.Louis, MO). The 95% CLA mixture (CLA-mix; 40%cis-9,trans-11 CLA; 44% trans-10,cis-12 CLA; 11% cis-10,cis-12 CLA) was obtained from Nu-Chek Prep Inc.(Elysian, MN). The individual trans-10,cis-12 CLA andcis-9,trans-11 CLA isomers were purchased fromMatreya, Inc. (Pleasant Gap, PA). Dulbecco’s modifiedEagle’s medium, nutrient mixture F-12, dexametha-sone, dihydroxyacetone phosphate (DHAP), isobutyl-methylxanthine, reduced NADH, gentamicin sulfate,HEPES buffer, hydrocortisone, insulin, and transferrinwere purchased from Sigma Chemical (St. Louis, MO).Collagenase (type I) was purchased from WorthingtonBiochemical (Freehold, NJ), fetal bovine serum (FBS)from Intergen (Purchase, NY), and fungizone fromGibco BRL (Division of Life Technologies, Gaithers-burg, MD).
Animals and Primary Culture
Two-day-old crossbred pigs (York × Landrace) wereobtained from a commercial producer and killed by CO2
asphyxiation in a manner approved by the Animal Careand Use Committee at Oregon State University. Stro-mal-vascular (S-V) cells were harvested by a collagen-ase digestion procedure as previously described (Surya-wan and Hu, 1997). Aliquots of S-V cells were countedusing a hemacytometer and seeded in culture dishes ata density of 5 × 104 cells/cm2 and incubated at 37°Cin 5% CO2 in air (designated d −1). Plating mediumconsisted of DME/F12 (1:1, vol/vol) containing 15 mmol/L of NaHCO3, 15 mmol/L of HEPES buffer, and 50 mg/L of gentamicin sulfate supplemented with 10% FBS.Cells were plated on 96-well, six-well, or 10-cm platesto facilitate proliferation, differentiation, or gene ex-pression studies, respectively. After 24 h, attached cellswere washed three times with plating medium to re-move unattached cells and cellular debris (designatedd 0). After washing, cells were continually induced todifferentiate in medium containing 10% FBS, 580 ng/mL of insulin, 10 µg/mL of transferrin, 500 ng/mL ofhydrocortisone, and the indicated fatty acid treatments
until d 10. Fatty acids were prepared as indicated byBrodie et al. (1999) and were added to serum-containingdifferentiation medium before treating cells. The finalconcentration of dimethyl sulfoxide was less than 0.1%(vol/vol). Dimethyl sulfoxide alone did not affect mark-ers of proliferation or differentiation at the levels pres-ent in these experiments. Serum was not analyzed forfatty acid content. Culture media were changed every2 d until d 10 (except where stated otherwise), whencells were subjected to glycerol-3-phosphate dehydroge-nase (GPDH) assays, oil red O staining, or gene expres-sion analysis. By d 10 in differentiation medium,greater than 70% of the cells accumulated multilocularlipid droplets. Trypan blue exclusion tests were con-ducted to confirm cell viability, and no fatty acid treat-ment was associated with decreased viability at theconcentrations used in these experiments.
Experimental Design
Stromal-vascular cells were plated as describedabove. To determine the effect of CLA isomers on theproliferation of porcine preadipocytes, cultures werecontinuously treated from d 0 to d 2 with 0 to100 µMof a CLA-mix, cis-9,trans-11 CLA, trans-10,cis-12 CLA,or linoleic acid in plating medium. Cell number wasdetermined following 48 h of treatment (before conflu-ence) based on the formation of formazan after 4 h ofincubation with the tetrazolium salt WST-1. To deter-mine the effect of CLA isomers on enzymatic and mor-phological markers of adipogenesis, as well as themRNA transcript expression of adipocyte-relatedgenes, cultures were continuously treated from d 0 tod 10 with 0 to 100 µM of a CLA-mix, cis-9,trans-11 CLA,trans-10,cis-12 CLA, or linoleic acid in differentiationmedium. On d 10, treated cultures were either usedfor isolation of cell lysates to facilitate GPDH activitystudies, isolation of mRNA to facilitate gene expressionstudies, or were stained with oil red O (ORO) to mea-sure total lipid accumulation. For each experiment, in-dividual fatty acid treatments were compared withequivalent vehicle-treated cultures which served ascontrols. Linoleic acid was used to control for potentialeffects of the addition of PUFA. Three to six replicateswere performed for the described experiments, withcells harvested from a different pig for each replicate.
Oil Red O Staining
To qualitatively assess S-V cell differentiation by mi-croscopy, cells were exposed to differentiation mediafrom d 0 to 10 and then fixed in 10% formalin andstained with 0.3% ORO for lipid. Extractable ORO wasthen measured spectrophotometrically (570 nm) bymodifying the procedure of Suryawan and Hu (1993).Briefly, the wells were fixed with Baker’s formalin for 15min, rinsed with distilled water, equilibrated in 100%propylene glycol for 2 min, and then stained with OROfor 10 min. Wells were then treated with 60% propylene

Brandebourg and Hu2098
glycol (vol/vol) for 1 min to remove free ORO, and rinsedwith distilled water. The ORO was extracted with theaddition of isopropanol and ORO determined in aliquotsfrom wells following shaking the culture plates 30 minat room temperature.
Glycerol-3-Phosphate Dehydrogenase Activity
The Sn-glycerol-3-phosphate dehydrogenase (GPDH;EC 1.1.1.8) activity was determined by measuring spec-trophotometrically the disappearance of NADH duringthe GPDH-catalyzed reduction of DHAP under zero-order conditions by the method of Kozak and Jensen(1974), as modified by Wise and Green (1979). Briefly,differentiated cells were harvested in ice-cold lysatebuffer (0.25 M sucrose, 1 mM EDTA, 1 mM dithiothrei-tol, 5 mM Tris base, pH 7.4). Membranes were disruptedby sonication and supernatant fractions were collectedfollowing centrifugation at 13,000 × g for 10 min at 4°Cto remove cellular debris. The reaction was initiatedby the addition of supernatant fractions to a standardmixture containing 100 mM triethanolamine/HClbuffer (pH 7.4), 2.5 mM EDTA, 0.176 mM NADH, 0.37mM DHAP, and 0.1 mM β-mercaptoethanol. The reac-tion was linear for sample time and concentration. Glyc-erol-3-phosphate dehydrogenase activity was expressedas units per milligram of protein, where one unit ofactivity is defined as the oxidation of 1 nmol of NADH/min. Protein was measured according to Bradford(1976).
Cell Number Assay
The colorimetric assay for quantification of cell num-ber and cell viability, based on the cleavage of the tetra-zolium salt WST-1 (4-[3-{4-iodophenyl}-2-{4-nitrophe-nyl}-2H-5-tetrazolio]-1,3-benzene disulfonate) by mito-chondrial dehydrogenases, was performed according tothe manufacturer (catalog No. 1644 807; BoehringerMannheim, Indianapolis, IN) as detailed by Brodie etal. (1999). The WST-1 assay was validated for the pri-mary S-V cell system by verifying that increased S-Vcell plating density correlated with increasedformazan formation.
RNA Isolation
Cells were harvested with a cell scraper, and totalRNA was extracted using the guanidinium-phenol-chlo-roform method (Chomczynski and Sacchi, 1987). TotalRNA concentration was determined spectrophotometri-cally at 260 and 280 nm. The ratio of light absorbanceat 260 nm to that at 280 nm was between 1.7 and 2.1for all samples. Five micrograms of total RNA fromeach sample was separated on a 1.2% denaturing form-aldehyde gel and stained with ethidium bromide. TheRNA integrity was assessed visually by judging thequality of 18 and 28S rRNA bands.
Semiquantitative Reverse Transcription-PCR
Reverse transcription (RT) reaction solution (20 µL)consisted of 4 µg of total RNA, 50 U of SuperScriptII reverse transcriptase (Invitrogen/Life Technologies,Carlsbad, CA), 40 U of an RNAse inhibitor (Invitrogen/Life Technologies), 0.5 mmol/L of deoxyribonucleotidetriphosphate, and 100 ng of random hexamer primers.Polymerase chain reaction was performed in 50 µL con-taining 20 mmol/L Tris�HCl, pH 8.4, 50 mmol/L KCl,1.0 µL of RT reaction, 2.5 U of Platinum Taq DNApolymerase (Hot Start; Invitrogen/Life Technologies),0.2 mmol/L of deoxyribonucleotide triphosphate, 2mmol/L Mg2+ (Invitrogen/Life Technologies), 10 pmoleach of gene specific primers, and 10 pmol each of prim-ers specific for either β-actin or 36B4. Thermal cyclingparameters were as follows: one cycle at 94°C for 4 min,followed by 26 to 30 cycles at 94°C for 1 min, 56°C for2 min, and 72°C for 2 min, with a final extension at72°C for 8 min. Primers were synthesized at the Centerfor Gene Research at Oregon State University. Identityof PCR products was verified either by restriction digestanalysis or DNA sequencing. The cycle number for eachmultiplex PCR reaction was selected by experimentallydetermining the highest cycle number in which the am-plification of both cDNA products was within a linearrange. The optimal cycle number was then consideredto be two cycles lower than the highest cycle of linearity.The RT-PCR amplicons were visualized by separatingDNA on a 3% agarose gel and staining with SYBRGreen according to the manufacturer’s directions (Mo-lecular BioProbes, Eugene, OR), followed by detectionand quantification using the Kodak Digital Science(Rochester, NY) electrophoresis documentation andanalysis system 120. Primer sequences, amplicon sizeand cycle length are listed in Table 1. Data for eachreplicate represented the mean of three individualRT-PCR.
Statistical Analyses
Experimental animals were assigned to differenttreatments in a completely randomized design. Dataare expressed as the mean ± SEM. Each replicate con-sisted of a single batch of S-V cells harvested from thes.c. adipose tissue of an individual pig. Data were ana-lyzed by using one-way ANOVA, followed by multiplecomparisons of means with Fisher’s LSD using SAS(SAS Inst., Inc., Cary, NC). Differences were consideredsignificant at P < 0.05.
Results
Proliferation
To study the potential isomer-specific effect of CLAon the proliferation of porcine S-V cells, the cleavageof the tetrazolium salt, WST-1, by mitochondrial dehy-drogenases was measured on d 2 following treatmentwith 0 to 100 µmol/L of either CLA-mix, cis-9,trans-11

CLA in pig adipogenesis 2099
Table 1. Oligonucleotide polymerase chain reaction primers
Accession Primer Product MultiplexGenea No. source Primer sequence (5′→3′) Orientation size, bp PCR cycle
β-Actin AF054837 Pig CGTGGGCCGCCCTAGGCACCA Forward 210 —TTGGCCTTAGGGTTCAGGGGGG Reverse
36B4 BC011291 Mouse GCACTCTCGCTTTCTGGAGGGTGTC Forward 292 —TGACTTGGTTGCTTTGGCGGGATTAG Reverse
SREBP-1c U00968 Human TGTGACCTCGCAGATCCAGC Forward 433 29GCGAATGTAGTCGATGGCCT Reverse
C/EBPα U34070 Human GGTGGACAAGAACAGCAACG Forward 370 29AGGCACCGGAATCTCCTAGT Reverse
COUP-TF X16155.1 Human CCCACTTTGAGGCACTTCT Forward 201 30ACATCGAGTGCGTGGTGT Reverse
PPARγ L40904 Human CACAGGCCGAGAAGGAGAAG Forward 330 28ATCTCCTGCACAGCCTCCAC Reverse
aSterol regulatory element-binding protein-1c (SREBP-1c), CCAAT/enhancer binding protein α (C/EBPα), chicken ovalbumin upstreampromoter-transcription factor 1 (COUP-TF1), and peroxisome proliferator-activated receptor γ (PPARγ).
CLA, trans-10,cis-12 CLA, or linoleic acid. Proliferationwas decreased in a dose-dependent fashion by trans-10,cis-12 CLA (P < 0.05). The CLA-mix, cis-9,trans-11CLA, and linoleic acid did not affect cell number (Fig-ure 1).
Differentiation
Sn-Glycerol-3-phosphate dehydrogenase (EC 1.1.1.8)activity was used as an enzymatic marker of differentia-tion in our primary culture system because GPDH ac-tivity is expressed in terminally differentiated, maturefat cells but not in preadipocytes. The GPDH activitywas inhibited in a dose-dependent fashion by continu-ally treating porcine S-V cells with the CLA-mix (0 to100 µmol/L) from d 0 to 10 after induction of differentia-tion (P < 0.01; Figure 2). Linoleic acid increased GPDHactivity at all concentrations administered (P < 0.01).Next, the cis-9,trans-11 CLA and the trans-10,cis-12CLA isomers were tested for their effect on GPDH activ-ity to determine whether either isomer mimicked theantiadipogenic activity of the CLA-mix. cis-9,trans-11CLA failed to inhibit GPDH activity at any concentra-tion; however, trans-10,cis-12 CLA potently inhibitedGPDH activity in a dose-dependent fashion (P < 0.001),with inhibition of greater than 80% occurring at thehighest dose administered (Figure 2).
Differentiation also was monitored morphologically.In agreement with the GPDH activity data, both theCLA-mix and the trans-10,cis-12 CLA isomer decreasedthe number of lipid-filled cells present in culture (Figure3A and B), whereas the cis-9,trans-11 CLA isomer hadno discernible effect vs. the control.
Because it was suggested that cis-9,trans-11 CLAmay increase the differentiation of pig preadipocytes(Ding et al., 2000), it is possible that cis-9,trans-11 CLAmay antagonize the action of trans-10,cis-12 CLA in pigpreadipocytes. Thus, we measured the ability of 12.5µM trans-10,cis-12 CLA to inhibit GPDH activity in thepresence of increasing concentrations (0 to 100 µM) ofcis-9,trans-11 CLA (Figure 4). Although a dose-re-
sponse effect was not observed, the presence of cis-9,trans-11 CLA tended (P < 0.09) to blunt the ability oftrans-10,cis-12 CLA to inhibit GPDH activity of theadipose S-V cells by 35%.
Gene Expression
To further study the effect of CLA on the differentia-tion of porcine preadipocytes, the isomer-specific effectof CLA on the expression of key adipogenic transcrip-tion factors was examined. The expressions of mRNAtranscripts for these genes on d 10 after induction areshown in Figure 5. The abundance of peroxisome prolif-erator-activated receptor γ (PPARγ) mRNA was de-creased by both the CLA-mix and the trans-10,cis-12CLA isomer (P < 0.003; Figure 5A). No fatty acid testedaltered the expression of CCAAT/enhancer binding pro-tein α (C/EBPα) mRNA vs. controls (Figure 5B). Mean-while, both the CLA-mix and the trans-10,cis-12 CLAisomer markedly decreased sterol regulatory element-binding protein-1c (SREBP-1c) mRNA (P < 0.05; Fig-ure 5C).
Recent data from our laboratory suggest that a noveltranscription factor, chicken ovalbumin upstream pro-moter transcription factor (COUP-TF), may negativelyregulate adipogenesis (Brodie et al., 1996; Brandebourgand Hu, 2005). Thus, it was of interest to determinewhether COUP-TF expression was altered in pig pread-ipocytes treated with CLA. Both the CLA-mix andtrans-10,cis-12 CLA increased the mRNA abundance ofCOUP-TF (P < 0.002). No other fatty acid tested affectedCOUP-TF mRNA abundance.
Discussion
Several studies indicate that feeding CLA to growingpigs decreases carcass fat, although the effect of CLAhas been variable and occurs by an unknown mecha-nism (Azain et al., 2003; Dugan et al., 2004). The pres-ent study provides evidence that CLA inhibits fat celldifferentiation in the pig, as suggested by the ability of
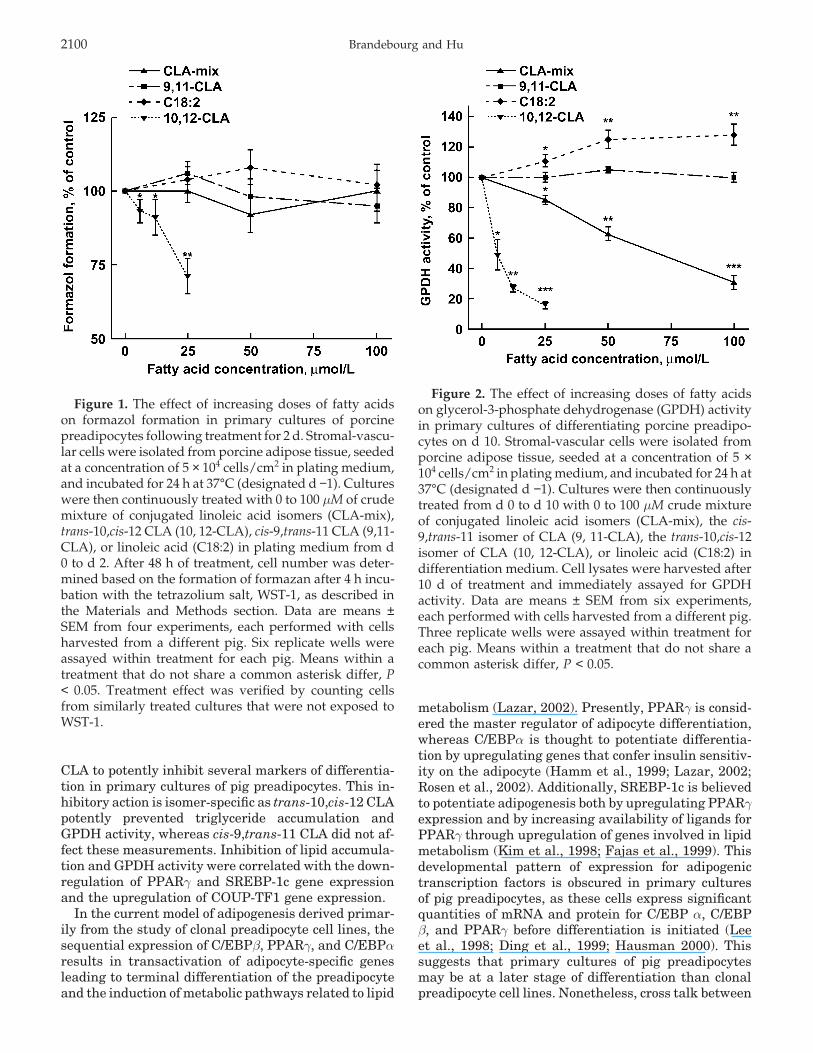
Brandebourg and Hu2100
Figure 1. The effect of increasing doses of fatty acidson formazol formation in primary cultures of porcinepreadipocytes following treatment for 2 d. Stromal-vascu-lar cells were isolated from porcine adipose tissue, seededat a concentration of 5 × 104 cells/cm2 in plating medium,and incubated for 24 h at 37°C (designated d −1). Cultureswere then continuously treated with 0 to 100 µM of crudemixture of conjugated linoleic acid isomers (CLA-mix),trans-10,cis-12 CLA (10, 12-CLA), cis-9,trans-11 CLA (9,11-CLA), or linoleic acid (C18:2) in plating medium from d0 to d 2. After 48 h of treatment, cell number was deter-mined based on the formation of formazan after 4 h incu-bation with the tetrazolium salt, WST-1, as described inthe Materials and Methods section. Data are means ±SEM from four experiments, each performed with cellsharvested from a different pig. Six replicate wells wereassayed within treatment for each pig. Means within atreatment that do not share a common asterisk differ, P< 0.05. Treatment effect was verified by counting cellsfrom similarly treated cultures that were not exposed toWST-1.
CLA to potently inhibit several markers of differentia-tion in primary cultures of pig preadipocytes. This in-hibitory action is isomer-specific as trans-10,cis-12 CLApotently prevented triglyceride accumulation andGPDH activity, whereas cis-9,trans-11 CLA did not af-fect these measurements. Inhibition of lipid accumula-tion and GPDH activity were correlated with the down-regulation of PPARγ and SREBP-1c gene expressionand the upregulation of COUP-TF1 gene expression.
In the current model of adipogenesis derived primar-ily from the study of clonal preadipocyte cell lines, thesequential expression of C/EBPβ, PPARγ, and C/EBPαresults in transactivation of adipocyte-specific genesleading to terminal differentiation of the preadipocyteand the induction of metabolic pathways related to lipid
Figure 2. The effect of increasing doses of fatty acidson glycerol-3-phosphate dehydrogenase (GPDH) activityin primary cultures of differentiating porcine preadipo-cytes on d 10. Stromal-vascular cells were isolated fromporcine adipose tissue, seeded at a concentration of 5 ×104 cells/cm2 in plating medium, and incubated for 24 h at37°C (designated d −1). Cultures were then continuouslytreated from d 0 to d 10 with 0 to 100 µM crude mixtureof conjugated linoleic acid isomers (CLA-mix), the cis-9,trans-11 isomer of CLA (9, 11-CLA), the trans-10,cis-12isomer of CLA (10, 12-CLA), or linoleic acid (C18:2) indifferentiation medium. Cell lysates were harvested after10 d of treatment and immediately assayed for GPDHactivity. Data are means ± SEM from six experiments,each performed with cells harvested from a different pig.Three replicate wells were assayed within treatment foreach pig. Means within a treatment that do not share acommon asterisk differ, P < 0.05.
metabolism (Lazar, 2002). Presently, PPARγ is consid-ered the master regulator of adipocyte differentiation,whereas C/EBPα is thought to potentiate differentia-tion by upregulating genes that confer insulin sensitiv-ity on the adipocyte (Hamm et al., 1999; Lazar, 2002;Rosen et al., 2002). Additionally, SREBP-1c is believedto potentiate adipogenesis both by upregulating PPARγexpression and by increasing availability of ligands forPPARγ through upregulation of genes involved in lipidmetabolism (Kim et al., 1998; Fajas et al., 1999). Thisdevelopmental pattern of expression for adipogenictranscription factors is obscured in primary culturesof pig preadipocytes, as these cells express significantquantities of mRNA and protein for C/EBP α, C/EBPβ, and PPARγ before differentiation is initiated (Leeet al., 1998; Ding et al., 1999; Hausman 2000). Thissuggests that primary cultures of pig preadipocytesmay be at a later stage of differentiation than clonalpreadipocyte cell lines. Nonetheless, cross talk between
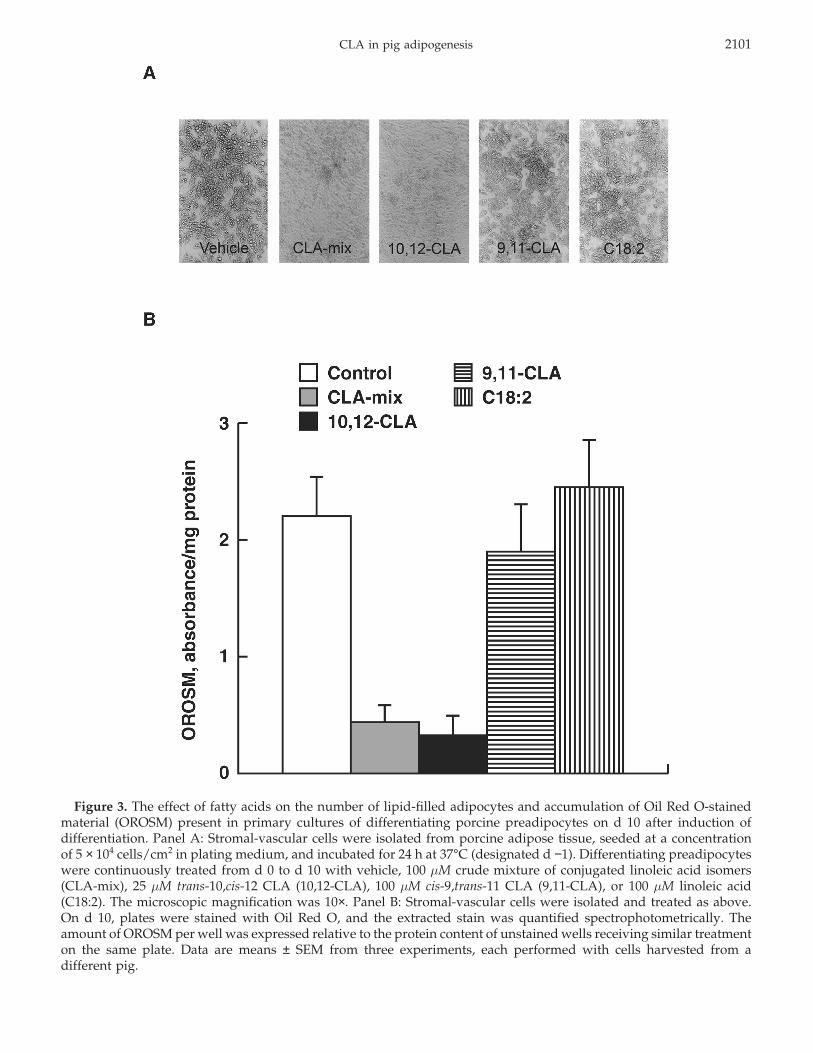
CLA in pig adipogenesis 2101
Figure 3. The effect of fatty acids on the number of lipid-filled adipocytes and accumulation of Oil Red O-stainedmaterial (OROSM) present in primary cultures of differentiating porcine preadipocytes on d 10 after induction ofdifferentiation. Panel A: Stromal-vascular cells were isolated from porcine adipose tissue, seeded at a concentrationof 5 × 104 cells/cm2 in plating medium, and incubated for 24 h at 37°C (designated d −1). Differentiating preadipocyteswere continuously treated from d 0 to d 10 with vehicle, 100 µM crude mixture of conjugated linoleic acid isomers(CLA-mix), 25 µM trans-10,cis-12 CLA (10,12-CLA), 100 µM cis-9,trans-11 CLA (9,11-CLA), or 100 µM linoleic acid(C18:2). The microscopic magnification was 10×. Panel B: Stromal-vascular cells were isolated and treated as above.On d 10, plates were stained with Oil Red O, and the extracted stain was quantified spectrophotometrically. Theamount of OROSM per well was expressed relative to the protein content of unstained wells receiving similar treatmenton the same plate. Data are means ± SEM from three experiments, each performed with cells harvested from adifferent pig.
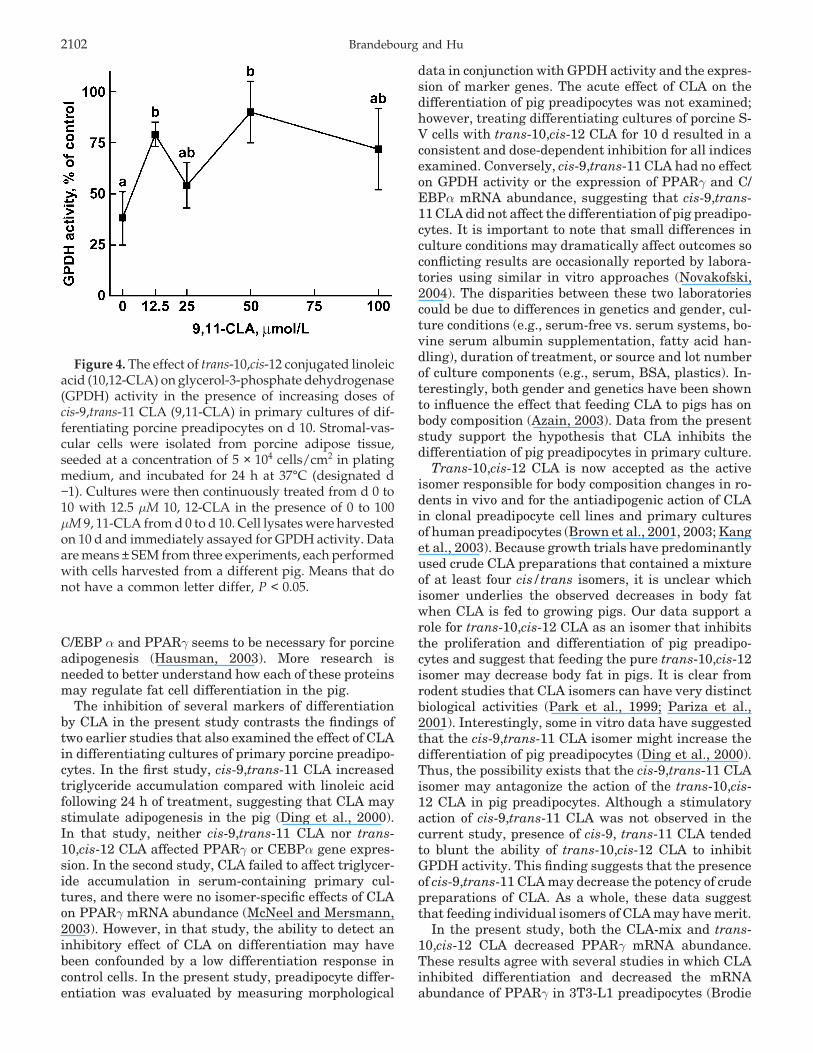
Brandebourg and Hu2102
Figure 4. The effect of trans-10,cis-12 conjugated linoleicacid (10,12-CLA) on glycerol-3-phosphate dehydrogenase(GPDH) activity in the presence of increasing doses ofcis-9,trans-11 CLA (9,11-CLA) in primary cultures of dif-ferentiating porcine preadipocytes on d 10. Stromal-vas-cular cells were isolated from porcine adipose tissue,seeded at a concentration of 5 × 104 cells/cm2 in platingmedium, and incubated for 24 h at 37°C (designated d−1). Cultures were then continuously treated from d 0 to10 with 12.5 µM 10, 12-CLA in the presence of 0 to 100µM 9, 11-CLA from d 0 to d 10. Cell lysates were harvestedon 10 d and immediately assayed for GPDH activity. Dataare means ± SEM from three experiments, each performedwith cells harvested from a different pig. Means that donot have a common letter differ, P < 0.05.
C/EBP α and PPARγ seems to be necessary for porcineadipogenesis (Hausman, 2003). More research isneeded to better understand how each of these proteinsmay regulate fat cell differentiation in the pig.
The inhibition of several markers of differentiationby CLA in the present study contrasts the findings oftwo earlier studies that also examined the effect of CLAin differentiating cultures of primary porcine preadipo-cytes. In the first study, cis-9,trans-11 CLA increasedtriglyceride accumulation compared with linoleic acidfollowing 24 h of treatment, suggesting that CLA maystimulate adipogenesis in the pig (Ding et al., 2000).In that study, neither cis-9,trans-11 CLA nor trans-10,cis-12 CLA affected PPARγ or CEBPα gene expres-sion. In the second study, CLA failed to affect triglycer-ide accumulation in serum-containing primary cul-tures, and there were no isomer-specific effects of CLAon PPARγ mRNA abundance (McNeel and Mersmann,2003). However, in that study, the ability to detect aninhibitory effect of CLA on differentiation may havebeen confounded by a low differentiation response incontrol cells. In the present study, preadipocyte differ-entiation was evaluated by measuring morphological
data in conjunction with GPDH activity and the expres-sion of marker genes. The acute effect of CLA on thedifferentiation of pig preadipocytes was not examined;however, treating differentiating cultures of porcine S-V cells with trans-10,cis-12 CLA for 10 d resulted in aconsistent and dose-dependent inhibition for all indicesexamined. Conversely, cis-9,trans-11 CLA had no effecton GPDH activity or the expression of PPARγ and C/EBPα mRNA abundance, suggesting that cis-9,trans-11 CLA did not affect the differentiation of pig preadipo-cytes. It is important to note that small differences inculture conditions may dramatically affect outcomes soconflicting results are occasionally reported by labora-tories using similar in vitro approaches (Novakofski,2004). The disparities between these two laboratoriescould be due to differences in genetics and gender, cul-ture conditions (e.g., serum-free vs. serum systems, bo-vine serum albumin supplementation, fatty acid han-dling), duration of treatment, or source and lot numberof culture components (e.g., serum, BSA, plastics). In-terestingly, both gender and genetics have been shownto influence the effect that feeding CLA to pigs has onbody composition (Azain, 2003). Data from the presentstudy support the hypothesis that CLA inhibits thedifferentiation of pig preadipocytes in primary culture.
Trans-10,cis-12 CLA is now accepted as the activeisomer responsible for body composition changes in ro-dents in vivo and for the antiadipogenic action of CLAin clonal preadipocyte cell lines and primary culturesof human preadipocytes (Brown et al., 2001, 2003; Kanget al., 2003). Because growth trials have predominantlyused crude CLA preparations that contained a mixtureof at least four cis/trans isomers, it is unclear whichisomer underlies the observed decreases in body fatwhen CLA is fed to growing pigs. Our data support arole for trans-10,cis-12 CLA as an isomer that inhibitsthe proliferation and differentiation of pig preadipo-cytes and suggest that feeding the pure trans-10,cis-12isomer may decrease body fat in pigs. It is clear fromrodent studies that CLA isomers can have very distinctbiological activities (Park et al., 1999; Pariza et al.,2001). Interestingly, some in vitro data have suggestedthat the cis-9,trans-11 CLA isomer might increase thedifferentiation of pig preadipocytes (Ding et al., 2000).Thus, the possibility exists that the cis-9,trans-11 CLAisomer may antagonize the action of the trans-10,cis-12 CLA in pig preadipocytes. Although a stimulatoryaction of cis-9,trans-11 CLA was not observed in thecurrent study, presence of cis-9, trans-11 CLA tendedto blunt the ability of trans-10,cis-12 CLA to inhibitGPDH activity. This finding suggests that the presenceof cis-9,trans-11 CLA may decrease the potency of crudepreparations of CLA. As a whole, these data suggestthat feeding individual isomers of CLA may have merit.
In the present study, both the CLA-mix and trans-10,cis-12 CLA decreased PPARγ mRNA abundance.These results agree with several studies in which CLAinhibited differentiation and decreased the mRNAabundance of PPARγ in 3T3-L1 preadipocytes (Brodie
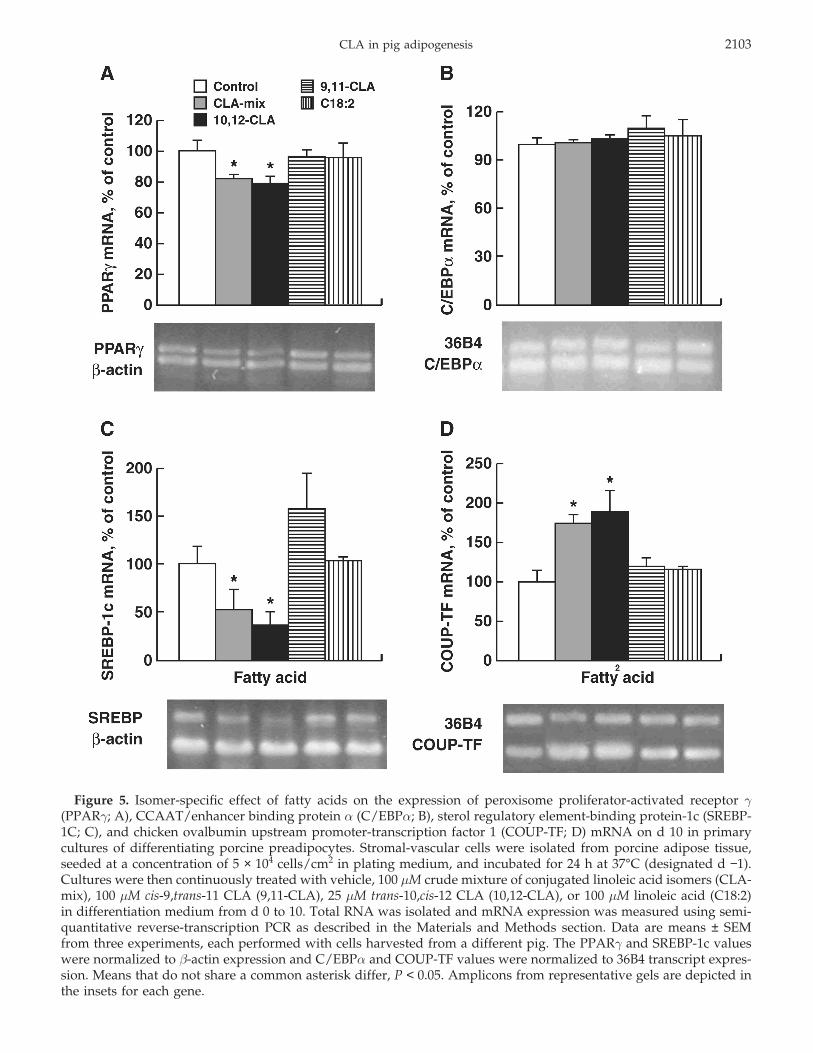
CLA in pig adipogenesis 2103
Figure 5. Isomer-specific effect of fatty acids on the expression of peroxisome proliferator-activated receptor γ(PPARγ; A), CCAAT/enhancer binding protein α (C/EBPα; B), sterol regulatory element-binding protein-1c (SREBP-1C; C), and chicken ovalbumin upstream promoter-transcription factor 1 (COUP-TF; D) mRNA on d 10 in primarycultures of differentiating porcine preadipocytes. Stromal-vascular cells were isolated from porcine adipose tissue,seeded at a concentration of 5 × 104 cells/cm2 in plating medium, and incubated for 24 h at 37°C (designated d −1).Cultures were then continuously treated with vehicle, 100 µM crude mixture of conjugated linoleic acid isomers (CLA-mix), 100 µM cis-9,trans-11 CLA (9,11-CLA), 25 µM trans-10,cis-12 CLA (10,12-CLA), or 100 µM linoleic acid (C18:2)in differentiation medium from d 0 to 10. Total RNA was isolated and mRNA expression was measured using semi-quantitative reverse-transcription PCR as described in the Materials and Methods section. Data are means ± SEMfrom three experiments, each performed with cells harvested from a different pig. The PPARγ and SREBP-1c valueswere normalized to β-actin expression and C/EBPα and COUP-TF values were normalized to 36B4 transcript expres-sion. Means that do not share a common asterisk differ, P < 0.05. Amplicons from representative gels are depicted inthe insets for each gene.

Brandebourg and Hu2104
et al., 1999; Evans et al., 2001, Kang et al., 2003). Brownet al. (2003) reported that CLA-induced inhibition ofhuman preadipocyte differentiation also was accompa-nied by decreased PPARγ mRNA abundance. Finally,feeding trans-10,cis-12 CLA to mice has consistentlydecreased PPARγ mRNA (Kang and Pariza, 2001; Ta-kahashi et al., 2002). Thus, there is a general consensusthat CLA decreases the expression of PPARγ mRNAabundance when inhibiting adipogenesis. Data fromour study is consistent with this conclusion.
Unexpectedly, CLA inhibited adipogenesis indepen-dent of effects on C/EBPα mRNA abundance in thepresent study; however, given the emerging role ofPPARγ as the master regulator of adipogenesis (Farmeret al., 2002; Rosen et al., 2002), effects on C/EBPα geneexpression may not be necessary to significantly inhibitfat cell differentiation. This idea is supported by recentwork, where retinoids also were shown to inhibit thedifferentiation of pig preadipocytes independent of aneffect on C/EBPα mRNA abundance (Brandebourg andHu, 2005). Furthermore, in clonal preadipocytes,SREBP-1c has been shown to regulate adipogenesisindependently of effects on C/EBPα through inductionof PPARγ expression and activity and through directregulation of lipogenic genes (Kim and Spiegelman,1996; Kim et al., 1998; Rosen et al., 2000). In the currentstudy, SREBP-1c mRNA was decreased in an isomer-specific pattern that mirrored the effect of trans-10,cis-12 CLA on GPDH activity and paralleled the expressionof PPARγ. The down-regulation of SREBP-1c by trans-10,cis-12 CLA is consistent with the current model ofadipogenesis. These data support a role for SREBP-1cin the mechanism by which CLA inhibits the differenti-ation of pig preadipocytes.
Chicken ovalbumin upstream promoter transcriptionfactor is an orphan nuclear receptor that has recentlybeen implicated as a potential negative regulator ofadipogenesis in the pig (Brandebourg and Hu, 2005).In the present study, both the CLA-mix and trans-10,cis-12 CLA increased the expression of COUP-TFmRNA concomitant with the down-regulation of mark-ers of preadipocyte differentiation, providing correla-tive evidence indicating that COUP-TF may play a rolein the antiadipogenic action of CLA in pig preadipo-cytes. It is known that COUP-TF can compete withPPARγ for both dimerization with RXR receptors andfor binding to the putative PPARγ direct repeat site inthe promoter regions of target genes (Tsai and Tsai,1997). Because binding of PPARγ to response elementsin the promoters of target genes represents a criticalpoint in the regulation of gene transcription by PPARγ,competition for DNA binding sites could be expectedto significantly decrease the transcriptional activity ofPPARγ. Thus, although we did not examine the effectof CLA on the transcriptional activity of PPARγ in thepresent study, a role for COUP-TF in the mechanismby which CLA inhibited the differentiation of porcinepreadipocytes is consistent with the current model ofadipogenesis.
Implications
Feeding conjugated linoleic acid mixtures to growingpigs decreases carcass fat, although this effect has beenvariable, and it occurs by an unknown mechanism.These data suggest that conjugated linoleic acid caninhibit the differentiation of pig preadipocytes in anisomer-specific manner, and this inhibition is correlatedwith the downregulation of peroxisome proliferator-ac-tivated receptor γ and sterol regulatory element-bind-ing protein-1c messenger RNA. This study is the firstto identify chicken ovalbumin upstream promoter-tran-scription factor 1 as a novel potential regulator of conju-gated linoleic acid action. Understanding the isomer-specific action of conjugated linoleic acid on the adipo-cyte will help us to devise designer conjugated linoleicacid mixtures that, when fed to growing pigs, will resultin higher quality pork products that are healthier toconsume.
Literature Cited
Azain, M. J. 2003. Conjugated linoleic acid and its effects on animalproducts and health in single-stomached animals. Proc. Nutr.Soc. 62:319–328.
Bradford, M. M. 1976. A rapid and sensitive method for the quantita-tion of microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal. Biochem. 72:248–254.
Brandebourg, T. D., and C. Y. Hu. 2005. Regulation of differentiatingpig preadipocytes by retinoic acid. J. Anim. Sci. 83:98–107.
Brodie, A. E., V. A. Manning, K. R. Ferguson, D. E. Jewell, and C.Y. Hu. 1999. Conjugated linoleic acid inhibits differentiation ofpre- and post- confluent 3t3-l1 preadipocytes but inhibits cellproliferation only in preconfluent cells. J. Nutr. 129:602–606.
Brodie, A. E., V. A. Manning, and C. Y. Hu. 1996. Inhibitors of preadi-pocyte differentiation induce COUP-TF binding to a PPAR/RXRbinding sequence. Biochem. Biophys. Res. Commun. 228:655–661.
Brown, J. M., M. S. Boysen, S. S. Jensen, R. F. Morrison, J. Storkson,R. Lea-Currie, M. Pariza, S. Mandrup, and M. K. McIntosh.2003. Isomer-specific regulation of metabolism and ppargammasignaling by cla in human preadipocytes. J. Lipid. Res.44:1287–1300.
Brown, J. M., Y. D. Halvorsen, Y. R. Lea-Currie, C. Geigerman, andM. McIntosh. 2001. Trans-10,cis-12, but not cis-9, trans-11, con-jugated linoleic acid attenuates lipogenesis in primary culturesof stromal vascular cells from human adipose tissue. J. Nutr.131:2316–2321.
Brown, J. M., and M. K. McIntosh. 2003. Conjugated linoleic acid inhumans: Regulation of adiposity and insulin sensitivity. J. Nutr.133:3041–3046.
Choi, Y., Y. C. Kim, Y. B. Han, Y. Park, M. W. Pariza, and J. M.Ntambi. 2000. The trans-10,cis-12 isomer of conjugated linoleicacid downregulates stearoyl-coa desaturase 1 gene expressionin 3t3-l1 adipocytes. J. Nutr. 130:1920–1924.
Chomczynski, P., and N. Sacchi. 1987. Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroform ex-traction. Anal. Biochem. 162:156–159.
Ding, S. T., R. L. McNeel, and H. J. Mersmann. 1999. Expression ofporcine adipocyte transcripts: Tissue distribution and differenti-ation in vitro and in vivo. Comp. Biochem. Physiol. B Biochem.Mol. Biol. 123:307–318.
Ding, S. T., R. L. McNeel, and H. J. Mersmann. 2000. Conjugatedlinoleic acid increases the differentiation of porcine adipocytesin vitro. Nutr. Res. 20:1569–1580.
Dugan, M. E., J. L. Aalhus, and J. K. Kramer. 2004. Conjugatedlinoleic acid pork research. Am. J. Clin. Nutr. 79:1212S–1216S.

CLA in pig adipogenesis 2105
Dugan, M. E. R., J. L. Aalhus, A. L. Schaefer, and J. K. G. Kramer.1997. The effect of conjugated linoleic acid on fat to lean reparti-tioning and feed conversion in pigs. Can. J. Anim. Sci.77:723–725.
Evans, M., Y. Park, M. Pariza, L. Curtis, B. Kuebler, and M. McIntosh.2001. Trans-10,cis-12 conjugated linoleic acid reduces triglycer-ide content, while differentially affecting peroxisome proliferatoractivated receptor gamma2 and AP2 expression in 3t3-l1 preadi-pocytes. Lipids 36:1223–1232.
Fajas, L., K. Schoonjans, L. Gelman, J. B. Kim, J. Najib, G. Martin,J. C. Fruchart, M. Briggs, B. M. Spiegelman, and J. Auwerx.1999. Regulation of peroxisome proliferator-activated receptorgamma expression by adipocyte differentiation and determina-tion factor 1/sterol regulatory element binding protein 1: Impli-cations for adipocyte differentiation and metabolism. Mol. Cell.Biol. 19:5495–5503.
Hamm, J. K., A. K. el Jack, P. F. Pilch, and S. R. Farmer. 1999. Roleof PPARγ in regulating adipocyte differentiation and insulin-responsive glucose uptake. Ann. N. Y. Acad. Sci. 892:134–145.
Hausman, G. J. 2000. The influence of dexamethasone and insulin onexpression of CCAAT/enhancer binding protein isoforms duringpreadipocyte differentiation in porcine stromal-vascular cell cul-tures: Evidence for very early expression of c/ebpα. J. Anim. Sci.78:1227–1235.
Hausman, G. J. 2003. Dexamethasone induced preadipocyte recruit-ment and expression of CCAAT/enhancing binding protein alphaand peroxisome proliferator activated receptor-gamma proteinsin porcine stromal-vascular (s-v) cell cultures obtained beforeand after the onset of fetal adipogenesis. Gen. Comp. Endocrinol.133:61–70.
Kang, K., W. Liu, K. J. Albright, Y. Park, and M. W. Pariza. 2003.Trans-10,cis-12 CLA inhibits differentiation of 3t3-l1 adipocytesand decreases PPARγ expression. Biochem. Biophys. Res. Com-mun. 303:795–799.
Kang, K., and M. W. Pariza. 2001. Trans-10,cis-12-conjugated linoleicacid reduces leptin secretion from 3t3-l1 adipocytes. Biochem.Biophys. Res. Commun. 287:377–382.
Kim, J. B., and B. M. Spiegelman. 1996. ADD1/SREBP1 promotesadipocyte differentiation and gene expression linked to fatty acidmetabolism. Genes Dev. 10:1096–1107.
Kim, J. B., H. M. Wright, M. Wright, and B. M. Spiegelman. 1998.ADD1/SREBP1 activates ppargamma through the production ofendogenous ligand. Proc. Natl. Acad. Sci. USA 95:4333–4337.
Kozak, L. P. 1974. Purification and characterization of two allelicforms of l-glycerol 3-phosphate dehydrogenase from inbredstrains of mice. Biochem. Genet. 12:69–79.
Lazar, M. A. 2002. Becoming fat. Genes Dev. 16:1–5.Lee, K., G. J. Hausman, and R. G. Dean. 1998. Expression of C/EBPα,
β and δ in fetal and postnatal subcutaneous adipose tissue. Mol.Cell. Biochem. 178:269–274.
McNeel, R. L., and H. J. Mersmann. 2003. Effects of isomers of conju-gated linoleic acid on porcine adipocyte growth and differentia-tion. J. Nutr. Biochem. 14:266–274.
Novakofski, J. 2004. Adipogenesis: Usefulness of in vitro and in vivoexperimental models. J. Anim. Sci. 82:905–915.
Pariza, M. W., Y. Park, and M. E. Cook. 2000. Mechanisms of actionof conjugated linoleic acid: Evidence and speculation. Proc. Soc.Exp. Biol. Med. 223:8–13.
Pariza, M. W., Y. Park, and M. E. Cook. 2001. The biologically activeisomers of conjugated linoleic acid. Prog. Lipid Res. 40:283–298.
Park, Y., J. M. Storkson, K. J. Albright, W. Liu, and M. W. Pariza.1999. Evidence that the trans-10,cis-12 isomer of conjugatedlinoleic acid induces body composition changes in mice. Lipids34:235–241.
Rosen, E. D., C. H. Hsu, X. Wang, S. Sakai, M. W. Freeman, F. J.Gonzalez, and B. M. Spiegelman. 2002. C/EBPα induces adipo-genesis through PPARγ: A unified pathway. Genes Dev.16:22–26.
Rosen, E. D., and B. M. Spiegelman. 2000. Molecular regulation ofadipogenesis. Ann. Rev. Cell. Dev. Biol. 16:266–285.
Suryawan, A., and C. Y. Hu. 1993. Effect of serum on differentiationof porcine adipose stromal-vascular cells in primary culture.Comp. Biochem. Physiol. Comp. Physiol. 105:485–492.
Suryawan, A., and C. Y. Hu. 1997. Effect of retinoic acid on differentia-tion of cultured pig preadipocytes. J. Anim. Sci. 75:112–117.
Takahashi, Y., M. Kushiro, K. Shinohara, and T. Ide. 2002. Dietaryconjugated linoleic acid reduces body fat mass and affects geneexpression of proteins regulating energy metabolism in mice.Comp. Biochem. Physiol. B Biochem. Mol. Biol. 133:395–404.
Tsai, S. Y., and M. J. Tsai. 1997. Chick ovalbumin upstream promoter-transcription factors (COUP-TFs): Coming of age. Endocrinol.Rev. 18:229–240.
Wise, L. S., and H. Green. 1979. Participation of one isozyme ofcytosolic glycerophosphate dehydrogenase in the adipose conver-sion of 3t3 cells. J. Biol. Chem. 254:273–275.
Related Documents











