Biomaterials 25 (2004) 757–768 Isolation, proliferation and differentiation of osteoblastic cells to study cell/biomaterial interactions: comparison of different isolation techniques and source Heidi Declercq a , Natasja Van den Vreken b , Erna De Maeyer b,c , Ronald Verbeeck b , Etienne Schacht d , Leo De Ridder a , Maria Cornelissen a, * a Department of Anatomy, Embryology, Histology and Medical Physics, Ghent University, Louis Pasteurlaan 2, B-9000 Ghent, Belgium b Department of Dental Materials Science, De Pintelaan 185 (P8), Ghent University, B-9000 Ghent, Belgium c Senior Research Assistant of the FSR-Flanders, Belgium d Polymer Material Research Group, Krijgslaan 281, Ghent University, B-9000 Ghent, Belgium Received 20 December 2002; accepted 14 July 2003 Abstract A sufficient amount of easily obtained and well-characterized osteoblastic cells is a useful tool to study biomaterial/cell interactions essential for bone tissue engineering. Osteoblastic cells were derived from adult and fetal rat via different isolation techniques. The isolation and in vitro proliferation of primary cultures were compared. The osteogenic potential of subcultures was studied by culturing them in osteogenic medium and compared with respect to alkaline phosphatase activity, nodule formation and mineralization potential. Calvaria cells were easier to obtain and the amount of cells released by enzymatic isolation was higher than for the long bone cells. The expansion of the cells in primary culture was highest for fetal calvaria cells compared to fetal and adult long bone cells. All cultures expressed high alkaline phosphatase activity except for calvaria cells obtained by spontaneous outgrowth. Enzymatic isolation of fetal calvaria and long bone cells favoured the osteogenic differentiation. Enzymatically isolated calvaria cells formed well-defined three-dimensional nodules which mineralized restricted to this area. On the contrary, cultures derived from fetal as well as adult long bones mineralized in ill-defined deposits throughout the culture and only formed occasionally nodular-like structures. The mineral phase of all osteoblastic cultures was identified as a carbonate-containing apatite. The present study demonstrates that considering the isolation method, proliferation capacity and the osteogenic potential, the enzymatically released fetal calvaria cells are most satisfactory to study cell/biomaterial interactions. r 2003 Elsevier Ltd. All rights reserved. Keywords: Cell culture; Cell proliferation; Osteogenesis; Osteoblast; Alkaline phosphatase; Calcification 1. Introduction Osteoblastic cells are used to study bone metabolism and biomaterial/cell interactions essential for bone tissue engineering. A sufficient amount of bone cells should be easily available. To study interactions of osteoblastic cells with biomaterials, osteoblast-like cell lines from rat, mouse or human osteosarcoma (UMR-106, MC3T3-E1, ROS 17/ 2.8, SAOS-2, HOS) are often used [1–4]. Although cell lines can be used for biocompatibility testing (cytotoxi- city), it is more convenient to use freshly isolated osteoblasts to study cell adhesion and the osteoconduc- tive and osteoinductive properties of the biomaterials. Osteogenic sarcoma cells retain many markers of the osteoblast phenotype but they differ from normal bone cells in their functional activities, responsivity to osteogenic agents and cell surface molecules [5,6]. Tissues often used to obtain osteoblastic cells for a large variety of experimental set-ups include cancellous bone specimens derived from long bones (femora and tibiae) of a wide range of adult and young animals (sheep, rat, mouse, etc.) as well as calvarial bones from fetal or neonatal animals. A number of methods are described to isolate osteoblastic cells from bone tissue. Bone cells can be obtained from bone by enzymatic digestion or from outgrowth of bone explants depending on the age of the animal. The use of proteolytic enzymes ARTICLE IN PRESS *Corresponding author. Tel.: +32-9-2649241; fax: +32-9-2649498. E-mail address: [email protected] (M. Cornelissen). 0142-9612/$ - see front matter r 2003 Elsevier Ltd. All rights reserved. doi:10.1016/S0142-9612(03)00580-5

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biomaterials 25 (2004) 757–768
ARTICLE IN PRESS
*Correspondin
E-mail addres
0142-9612/$ - see
doi:10.1016/S014
Isolation, proliferation and differentiation of osteoblastic cells tostudy cell/biomaterial interactions: comparison of different
isolation techniques and source
Heidi Declercqa, Natasja Van den Vrekenb, Erna De Maeyerb,c, Ronald Verbeeckb,Etienne Schachtd, Leo De Riddera, Maria Cornelissena,*
a Department of Anatomy, Embryology, Histology and Medical Physics, Ghent University, Louis Pasteurlaan 2, B-9000 Ghent, Belgiumb Department of Dental Materials Science, De Pintelaan 185 (P8), Ghent University, B-9000 Ghent, Belgium
c Senior Research Assistant of the FSR-Flanders, Belgiumd Polymer Material Research Group, Krijgslaan 281, Ghent University, B-9000 Ghent, Belgium
Received 20 December 2002; accepted 14 July 2003
Abstract
A sufficient amount of easily obtained and well-characterized osteoblastic cells is a useful tool to study biomaterial/cell
interactions essential for bone tissue engineering. Osteoblastic cells were derived from adult and fetal rat via different isolation
techniques. The isolation and in vitro proliferation of primary cultures were compared. The osteogenic potential of subcultures was
studied by culturing them in osteogenic medium and compared with respect to alkaline phosphatase activity, nodule formation and
mineralization potential. Calvaria cells were easier to obtain and the amount of cells released by enzymatic isolation was higher than
for the long bone cells. The expansion of the cells in primary culture was highest for fetal calvaria cells compared to fetal and adult
long bone cells. All cultures expressed high alkaline phosphatase activity except for calvaria cells obtained by spontaneous
outgrowth. Enzymatic isolation of fetal calvaria and long bone cells favoured the osteogenic differentiation. Enzymatically isolated
calvaria cells formed well-defined three-dimensional nodules which mineralized restricted to this area. On the contrary, cultures
derived from fetal as well as adult long bones mineralized in ill-defined deposits throughout the culture and only formed occasionally
nodular-like structures. The mineral phase of all osteoblastic cultures was identified as a carbonate-containing apatite. The present
study demonstrates that considering the isolation method, proliferation capacity and the osteogenic potential, the enzymatically
released fetal calvaria cells are most satisfactory to study cell/biomaterial interactions.
r 2003 Elsevier Ltd. All rights reserved.
Keywords: Cell culture; Cell proliferation; Osteogenesis; Osteoblast; Alkaline phosphatase; Calcification
1. Introduction
Osteoblastic cells are used to study bone metabolismand biomaterial/cell interactions essential for bonetissue engineering.
A sufficient amount of bone cells should be easilyavailable. To study interactions of osteoblastic cells withbiomaterials, osteoblast-like cell lines from rat, mouse orhuman osteosarcoma (UMR-106, MC3T3-E1, ROS 17/2.8, SAOS-2, HOS) are often used [1–4]. Although celllines can be used for biocompatibility testing (cytotoxi-city), it is more convenient to use freshly isolated
g author. Tel.: +32-9-2649241; fax: +32-9-2649498.
s: [email protected] (M. Cornelissen).
front matter r 2003 Elsevier Ltd. All rights reserved.
2-9612(03)00580-5
osteoblasts to study cell adhesion and the osteoconduc-tive and osteoinductive properties of the biomaterials.
Osteogenic sarcoma cells retain many markers of theosteoblast phenotype but they differ from normal bonecells in their functional activities, responsivity toosteogenic agents and cell surface molecules [5,6].
Tissues often used to obtain osteoblastic cells for alarge variety of experimental set-ups include cancellousbone specimens derived from long bones (femora andtibiae) of a wide range of adult and young animals(sheep, rat, mouse, etc.) as well as calvarial bones fromfetal or neonatal animals. A number of methods aredescribed to isolate osteoblastic cells from bone tissue.Bone cells can be obtained from bone by enzymaticdigestion or from outgrowth of bone explants dependingon the age of the animal. The use of proteolytic enzymes

ARTICLE IN PRESSH. Declercq et al. / Biomaterials 25 (2004) 757–768758
(collagenase and trypsin) works well with the calvarialbones of fetal or neonatal rodents and fetal chicks. Forheavily mineralized bones of the adult, however, the useof explants, with or without a collagenase pretreatment,is the method of choice [7].
Successful bone tissue engineering may rely not onlyon the differentiation of cells in contact with the implantmaterials but also on the proliferation capacity of thecells to obtain a proper colonization of the three-dimensional matrix.
The aim of the present work is to obtain osteoblastcultures that can be used to evaluate cell/biomaterialinteractions for bone tissue engineering. For thispurpose, we compared the in vitro cell expansion andthe osteogenic potential of osteoblastic cells derivedfrom adult 2 and fetal rat via different isolationtechniques in parallel assays. Cellular alkaline phospha-tase activity (early stage) and the formation of anextracellular matrix containing calcium phosphate (finalstage) were used as markers of the osteoblast phenotype.
2. Materials and methods
2.1. Materials
2.1.1. Cell culture
DMEM glutamax-1 (low glucose) (Cat No. 21885-025), fetal bovine serum (FBS heat inactivated, E.C.approved), a -MEM supplemented with nucleotides (a-MEM DNA/RNA) (Cat No. 22571-020), l-glutamine,penicillin–streptomycin (10,000 U/ml–10,000 mg/ml) andFungizones were purchased from Gibco BRL (LifeTechnologies, Merelbeke, Belgium). l-ascorbic acid 2-phosphate, b-glycerophosphate, dexamethasone-watersoluble (Cat No. D-2915), trypsin II-S and collagenaseIA were supplemented from Sigma (Sigma-Aldrich NV/SA, Bornem, Belgium). Tissue culture dishes andThermanox coverslips were purchased from Nunc (LifeTechnologies, Merelbeke, Belgium).
Two different culture media were used in theseexperiments. Standard culture medium was used toincrease the number of cells in primary culture. Fordifferentiation assays, the cells were transferred to anosteogenic culture medium.
Standard culture medium, which consists of DMEMglutamax-1 (low glucose)+10 vol% fetal bovine ser-um+penicillin–streptomycin (50 U/ml–50 mg/ml)+1 vol% Fungizones and 100 mm l-ascorbic acid 2-phosphate, was used for osteoblastic cells in primaryculture until subculture.
Osteogenic culture medium, which consists of a-MEMDNA/RNA+10 vol% fetal bovine serum+4 mm
l-glutamine+penicillin–streptomycin (50 U/ml–50 mg/ml)+1 vol% Fungizones+100 mm l-ascorbic acid
2-phosphate+10 mm b-glycerophosphate, was used foralkaline phosphatase and mineralization assays.
In an additional experiment, 250 mm l-ascorbic acid2-phosphate or 100 nm dexamethasone were usedthroughout secondary culture.
2.1.2. Assays
P-nitrophenylphosphate, p-nitrophenol, glycine andsilver nitrate were purchased from ICN Biomedicals Inc.(Asse-Relegem, Belgium). 5-bromo-4-chloro-3-indolylphosphate/nitro blue tetrazolium (BCIP/NBT) liquidsubstrate system, non-enzymatic dissociation reagent(C-5789), Alizarin Red S and collagen (C-8886) werepurchased from Sigma (Sigma-Aldrich NV/SA, Bor-nem, Belgium). The protein assay kit and bovine gammaglobulin standard was supplemented from Bio-RadLaboratories (Nazareth-Eke, Belgium). Sodiumthiosul-phate was purchased from Merck (VWR International,Leuven, Belgium). K4Fe(CN)6 � 3H2O and Canadabalsam were purchased from Vel (VWR International,Leuven, Belgium).
2.2. Cell culture methodology
2.2.1. Isolation and culture of osteoblastic cells
Osteoblastic cells were isolated either enzymatically orobtained from explant cultures from fetal (21 day)Wistar rat calvaria and long bones (tibiae and femorafrom the hind limbs of 12 fetal rats) and from adult(3-month old) Wistar rat long bones (tibiae and femorafrom the hind limbs of one adult rat).
Explant culture. Osteoblastic cells were isolated fromfetal Wistar rat calvaria, long bones and adult longbones. Briefly, skin, soft connective tissue and perios-teum were removed. Diaphyses were cut free ofepiphyseal cartilage. Bone was cut into pieces ofapproximately 2� 2 mm2 and washed. Bone pieces fromadult long bones were repeatedly vortexed in 10 mlRinger solution to remove blood and bone marrow.
Bone pieces (for both fetal and adult rat) wereincubated for 15 min at 37�C on the bottom of astanding T25 tissue culture dish to let them adhere to theculture dish before standard culture medium was added[8]. The bone pieces were divided over T25 tissue culturedishes with approximately 20 bone pieces/T25 tissueculture dish. They were cultured until confluence in acontrolled atmosphere (5% CO2/95% air, 37�C).
Enzymatic isolation. Osteoblastic cells were enzyma-tically isolated from fetal Wistar rat calvaria and longbones by the method of Bellows et al. [9–11] with someminor modifications. The bone pieces (isolated asdescribed above) were digested sequentially in a trypsinII-S (25 mg)-collagenase IA (70 mg) in 50 ml Ringersolution according to Schwartz [8] at 37�C for 10, 20, 30,50 and 70 min.

ARTICLE IN PRESSH. Declercq et al. / Biomaterials 25 (2004) 757–768 759
Rat calvaria cells obtained from the last threedigestion steps were pooled and plated together in aT25 tissue culture dish at a concentration of 20,000cells/cm2. Cells from fetal rat long bones obtained fromthe last four digestion steps were pooled and platedtogether in a T25 4.tissue culture dish also at aconcentration of 20,000 cells/cm2. The cells werecultured until confluence in standard culture mediumor in standard culture medium supplemented with10 mm b-glycerophosphate in a controlled atmosphere(5% CO2/95% air, 37�C). Two independent experimentswere performed. In each experiment 12 fetal rats or oneadult rat was used.
Differentiation assay. After confluence, cells isolatedeither enzymatically or by explant culture were trypsi-nized and plated into 24-well tissue culture dishes (2–4replicates for biochemical detection of the alkalinephosphatase activity) or on Thermanoxs coverslips(4–6 replicates for histochemical assays, TEM andinfrared spectrometry) at a concentration of 40,000cells/well (20,000 cells/cm2) in 0.5 ml osteogenic culturemedium. The cells were lysed or fixed after 4, 14 and 21days in their culture medium.
2.3. Alkaline phosphatase detection and mineralization of
the extracellular matrix
2.3.1. Cellular alkaline phosphatase activity
Alkaline phosphatase activity in the cultures wasdetermined biochemically as well as histochemically.
Biochemical detection of the alkaline phosphatase
activity. After rinsing the monolayers with Ringersolution, the cellular material was removed after lysisinto 250 ml of a buffer containing 10 mm Tris HCl pH7.5, 0.5 mm MgCl2 and 0.1% Triton X-100. The cellularmaterial was homogenized by two freeze-and-thawcycles. Alkaline phosphatase activity was determinedwith p-nitrophenylphosphate as the substrate. Samplevolumes of 50 ml were added to 50 ml p-nitrophenylpho-sphate (4.34 mm) in 100 mm glycine, pH 10.3, 1 mm
MgCl2 and incubated at 37�C for 30 min on a benchshaker. The enzymatic reaction was stopped byadding 50 ml of 1m NaOH. Enzyme activity wasquantified by absorbance measurements at 405 nm(Universal Microplate Reader EL 800, BIO-TEKinstruments Inc.) and calculated according to a seriesof p-nitrophenol standards [12]. Total protein contentwas determined with the Bradford method fromaliquots of the same samples with the Bio-Rad proteinassay kit, read at 595 nm (BioPhotometer, eppendorf)and calculated according to a series of bovine gammaglobulin standards. Alkaline phosphatase activity wasexpressed as mm p-nitrophenol/mg protein. Cultures oftwo independent experiments in 2–4 replicates wereanalysed.
Histochemical staining of alkaline phosphatase positive
cells. After rinsing the cell monolayers on the Therma-noxs coverslips with cold (4�C) Ringer solution, thecells were fixed with acetone (�20�C) during 5 min. Thefixed monolayers were washed with cold distilled waterand let to dry for 30 min. The monolayers wereincubated for 10 min with BCIP/NBT liquid substratesystem at room temperature. The reaction was stoppedby removing the substrate solution and washing withdistilled water. The Thermanox coverslips were dehy-drated in a graded alcohol series and mounted withCanada balsam.
2.3.2. Mineralization of the extracellular matrix
Different methods were used to analyse the mineraldeposits in the osteoblast cultures. The presence ofphosphate and calcium deposits were analysed histo-chemically by respectively Von Kossa and Alizarin RedS staining. The crystalline structure of the calciumphosphate deposits was analysed qualitatively bytransmission electron microscopy and infrared spectro-metry.
2.3.3. Phosphate staining (Von Kossa)
Calcium phosphate deposits can be detected by theVon Kossa technique in which phosphate deposits arestained black.
The monolayers were rinsed with cold (4�C) Ringersolution and fixed with 4% formaldehyde buffered with10 mm phosphate (pH 6, 9) (4�C, 10 min). After washingwith cold distilled water, the fixed cultures were coveredwith a 5% silver nitrate solution and kept for 30 min in adark room. The cultures were rinsed with distilled waterand a sodium-carbonate-formaldehyde solution (5 gNa2CO3 in 75 ml distilled water with added 25 ml of36% formaldehyde) was added for 2 min. The cultureswere rinsed in distilled water for 10 min and incubatedfor 20 min with a staining solution, according toFarmer, containing 1 vol% 10% K4Fe(CN)6 � 3H2Oand 9 vol% 10% sodiumthiosulphate [13]. After rinsingwith distilled water, the Thermanox coverslips weredehydrated in a graded alcohol series and mounted withCanada balsam.
2.3.4. Calcium staining (Alizarin Red S)
Staining of calcium with Alizarin Red S reveals thepresence of calcium deposits. After rinsing the mono-layers with cold (4�C) Ringer solution, the cells werefixed with 4% formaldehyde buffered with 10 mm
phosphate (pH 6, 9) (4�C, 10 min). The fixed mono-layers were washed with cold distilled water. Thecultures were covered with a 2% Alizarin Red S solution(adjusted to pH 4.2 with 10% ammoniumhydroxide) for5 min. The Thermanoxs coverslips were rinsed for 30 sin acetone, followed by 15 s in an acetone/toluenesolution (50/50 vol%). After rinsing in toluene, the
ARTICLE IN PRESSH. Declercq et al. / Biomaterials 25 (2004) 757–768760
Thermanoxs coverslips were embedded in Canadabalsam.
2.3.5. Transmission electron microscopy
Ultrastructural analysis was carried out to visualizethe collagen I fibers and the mineral deposits in theextracellular matrix.
Thermanoxs monolayers on which three-dimensionalnodules were grown, were incubated in a non-enzymaticdissociation reagent while cutting the nodule away fromthe surrounding monolayer. The nodules were fixed at4�C for 1 h in 1% glutaraldehyde buffered with 0.1msodium cacodylate (pH 7.2), and then washed in thesame buffer.
The nodules were post-fixed with osmium tetroxide,dehydrated in graded concentrations of alcohol andembedded in epoxy resin. Thin sections (60 nm) were cutwith a diamond knife, mounted on cupper grids, stainedwith uranyl acetate and lead citrate and examined usinga JEOL 1200 EX II transmission electron microscopeoperating at 80 KeV.
2.3.6. Infrared spectrometry
The physical and chemical composition of the mineralphase was determined using infrared spectrometry.
The monolayers were rinsed with Ringer solution andlysed and scraped into 250 ml of a buffer containing10 mm Tris HCl pH 7.5, 0.5 mm MgCl2 and 0.1% TritonX-100. After centrifugation at 9000 rpm for 3 min, thesupernatant was removed and the cell pellet was dried.FTIR spectra of the samples dispersed in KBr wererecorded in the range 400–4000 cm�1 with a resolutionof 1 cm�1 using a Mattson Galaxy 6030 FTIR spectro-photometer. The spectra were compared with those of acommercial collagen sample (C-8886, Sigma, St Louis,MO, USA) and calcium phosphate standards preparedin our laboratory [14,15].
Osteoblastic cells isolated by explant cu
4 d
ays
14 d
ays0
10
20
30
Osteoblastic cells after respectively 4, 14 andin osteogenic medium
Alk
alin
e p
ho
sph
atas
e ac
tivi
ty:
mM
pN
P/m
g p
rote
in
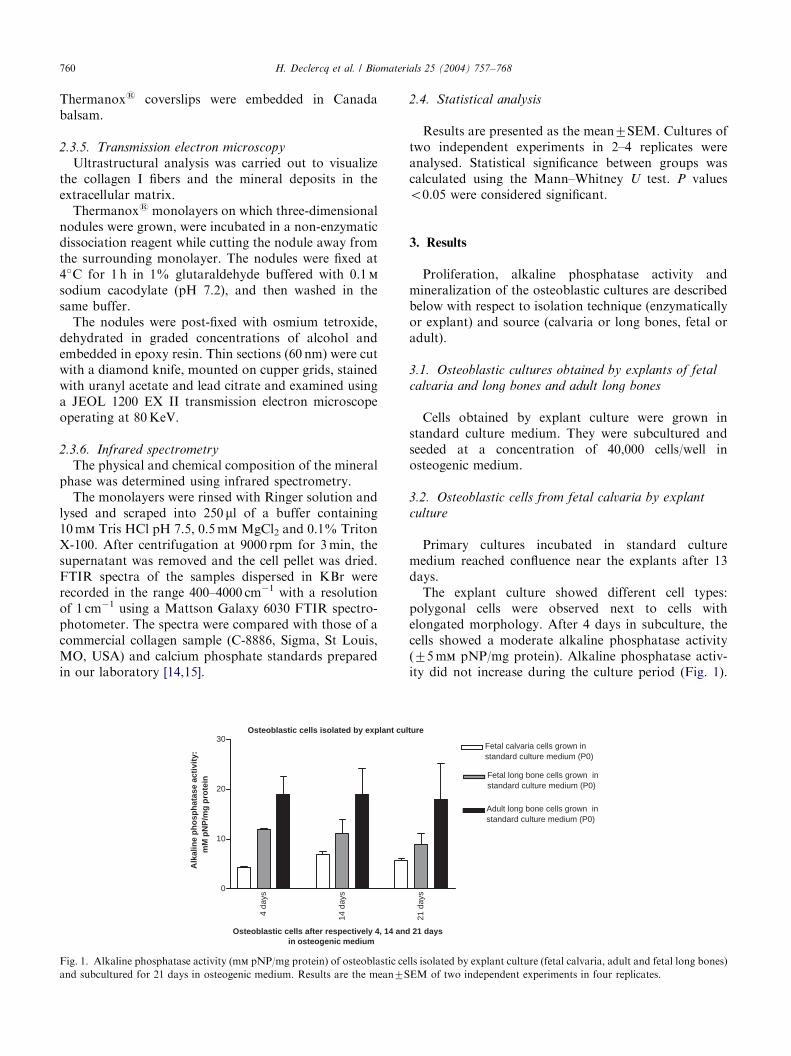
Fig. 1. Alkaline phosphatase activity (mm pNP/mg protein) of osteoblastic ce
and subcultured for 21 days in osteogenic medium. Results are the mean7S
2.4. Statistical analysis
Results are presented as the mean7SEM. Cultures oftwo independent experiments in 2–4 replicates wereanalysed. Statistical significance between groups wascalculated using the Mann–Whitney U test. P valueso0.05 were considered significant.
3. Results
Proliferation, alkaline phosphatase activity andmineralization of the osteoblastic cultures are describedbelow with respect to isolation technique (enzymaticallyor explant) and source (calvaria or long bones, fetal oradult).
3.1. Osteoblastic cultures obtained by explants of fetal
calvaria and long bones and adult long bones
Cells obtained by explant culture were grown instandard culture medium. They were subcultured andseeded at a concentration of 40,000 cells/well inosteogenic medium.
3.2. Osteoblastic cells from fetal calvaria by explant
culture
Primary cultures incubated in standard culturemedium reached confluence near the explants after 13days.
The explant culture showed different cell types:polygonal cells were observed next to cells withelongated morphology. After 4 days in subculture, thecells showed a moderate alkaline phosphatase activity(75 mm pNP/mg protein). Alkaline phosphatase activ-ity did not increase during the culture period (Fig. 1).
lture
21 d
ays
Fetal calvaria cells grown instandard culture medium (P0)
Fetal long bone cells grown instandard culture medium (P0)
Adult long bone cells grown instandard culture medium (P0)
21 days
lls isolated by explant culture (fetal calvaria, adult and fetal long bones)
EM of two independent experiments in four replicates.

ARTICLE IN PRESS
Fig. 2. Osteoblastic cells isolated by explant culture and cultured for 4 days (a,c,e) and 21 days (b,d,f) in osteogenic medium. (a,b) fetal rat calvaria
cells; (c,d) fetal long bone cells; (e,f) adult rat long bone cells. Phase contrast microscopy.
H. Declercq et al. / Biomaterials 25 (2004) 757–768 761
No nodule formation was observed (Figs. 2a and b) andVon Kossa staining was negative throughout theexperiment.
3.3. Osteoblastic cells from fetal long bones by explant
culture
The osteoblastic cells were grown for 20 days instandard culture medium until confluence near theexplants.
Cultures at low density showed elongated morphol-ogy. Alkaline phosphatase activities of subcultured cells(4 days in subculture) was higher than 10 mm pNP/mgprotein and remained steady state throughout theculture period (Fig. 1). Explant cultures showed nonodule formation (Figs. 2c and d). Mineralization wasvisible at day 21 (Figs. 2c and d) and showed a diffusepattern.
3.4. Osteoblastic cells from adult long bones by explant
culture
Primary cultures of osteoblastic cells reached con-fluence near the explants after 18 days. The cultures
showed high alkaline phosphatase activity (>15 mm
pNP/mg protein) after 4 days of subculture in osteo-genic medium and remained steady state (Fig. 1). Thecells showed elongated morphology at low density (Fig.2e). At higher densities, cells are grown in network-likestructures and sporadically nodules were evident.
Mineralization was visible at day 14 and increasedwithin the following days (Fig. 2f). The calciumphosphate deposits were diffuse.
Taking into account the relative intensities of theabsorptions in the IR spectrum of protein (Fig. 3d) andcarbonate-containing apatite (Fig. 3e), the presence ofthe mineral phase generated by the osteoblastic cells isclearly seen in the IR spectrum (Fig. 3a). The intenseband in the region 950–1300 cm�1 with shouldersaround 962, 1040 and 1095 cm�1 and the band around600 cm�1 with absorptions at 571 and 601 cm�1 arisefrom PO4
3� vibrations in an apatitic lattice [14,15]. Inaddition, a small peak around 872 cm�1 is typical for thebending vibration of CO3
2� substituting for PO43� in an
apatite lattice [14]. The typical stretching vibrations ofthis CO3
2� around 1420 and 1455 cm�1, however, arebarely detectable due to an overlap with the C–Hbending vibrations of the methyl groups of the protein.
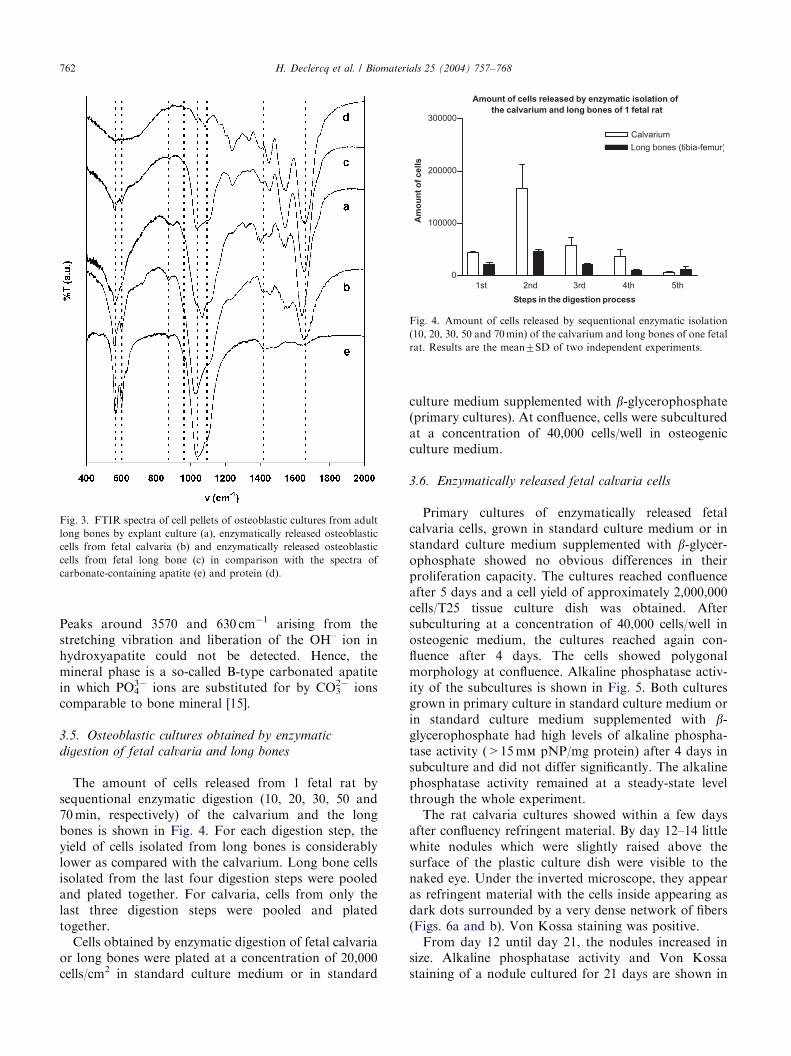
ARTICLE IN PRESS
Fig. 3. FTIR spectra of cell pellets of osteoblastic cultures from adult
long bones by explant culture (a), enzymatically released osteoblastic
cells from fetal calvaria (b) and enzymatically released osteoblastic
cells from fetal long bone (c) in comparison with the spectra of
carbonate-containing apatite (e) and protein (d).
Amount of cells released by enzymatic isolation of
the calvarium and long bones of 1 fetal rat
1st 2nd 3rd 4th 5th
0
100000
200000
300000
Calvarium
Long bones (tibia-femur)
Steps in the digestion process
Am
ou
nt
of
cells
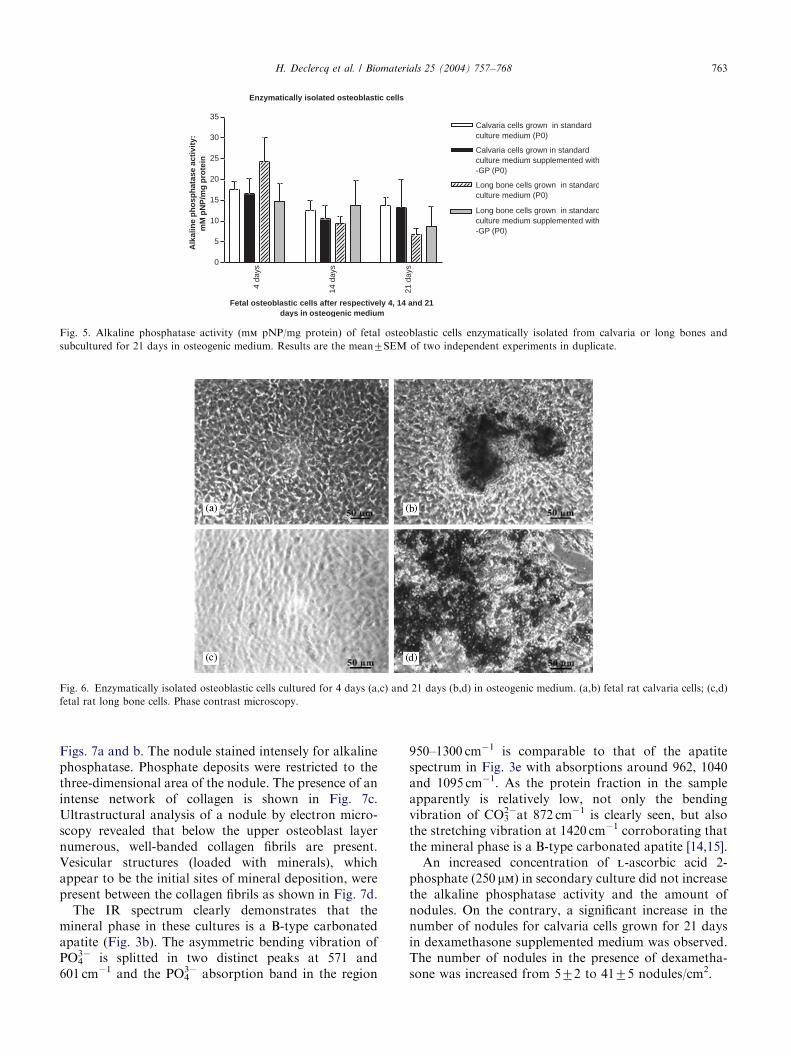
Fig. 4. Amount of cells released by sequentional enzymatic isolation
(10, 20, 30, 50 and 70 min) of the calvarium and long bones of one fetal
rat. Results are the mean7SD of two independent experiments.
H. Declercq et al. / Biomaterials 25 (2004) 757–768762
Peaks around 3570 and 630 cm�1 arising from thestretching vibration and liberation of the OH– ion inhydroxyapatite could not be detected. Hence, themineral phase is a so-called B-type carbonated apatitein which PO4
3� ions are substituted for by CO32� ions
comparable to bone mineral [15].
3.5. Osteoblastic cultures obtained by enzymatic
digestion of fetal calvaria and long bones
The amount of cells released from 1 fetal rat bysequentional enzymatic digestion (10, 20, 30, 50 and70 min, respectively) of the calvarium and the longbones is shown in Fig. 4. For each digestion step, theyield of cells isolated from long bones is considerablylower as compared with the calvarium. Long bone cellsisolated from the last four digestion steps were pooledand plated together. For calvaria, cells from only thelast three digestion steps were pooled and platedtogether.
Cells obtained by enzymatic digestion of fetal calvariaor long bones were plated at a concentration of 20,000cells/cm2 in standard culture medium or in standard
culture medium supplemented with b-glycerophosphate(primary cultures). At confluence, cells were subculturedat a concentration of 40,000 cells/well in osteogenicculture medium.
3.6. Enzymatically released fetal calvaria cells
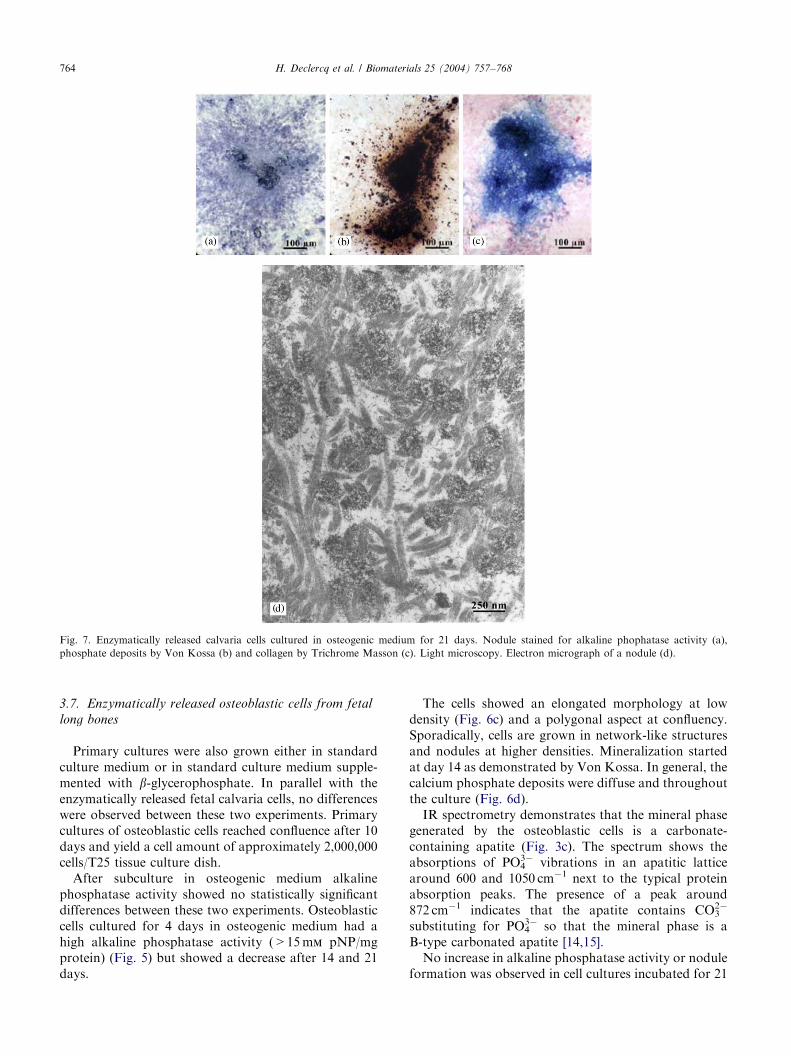
Primary cultures of enzymatically released fetalcalvaria cells, grown in standard culture medium or instandard culture medium supplemented with b-glycer-ophosphate showed no obvious differences in theirproliferation capacity. The cultures reached confluenceafter 5 days and a cell yield of approximately 2,000,000cells/T25 tissue culture dish was obtained. Aftersubculturing at a concentration of 40,000 cells/well inosteogenic medium, the cultures reached again con-fluence after 4 days. The cells showed polygonalmorphology at confluence. Alkaline phosphatase activ-ity of the subcultures is shown in Fig. 5. Both culturesgrown in primary culture in standard culture medium orin standard culture medium supplemented with b-glycerophosphate had high levels of alkaline phospha-tase activity (>15 mm pNP/mg protein) after 4 days insubculture and did not differ significantly. The alkalinephosphatase activity remained at a steady-state levelthrough the whole experiment.
The rat calvaria cultures showed within a few daysafter confluency refringent material. By day 12–14 littlewhite nodules which were slightly raised above thesurface of the plastic culture dish were visible to thenaked eye. Under the inverted microscope, they appearas refringent material with the cells inside appearing asdark dots surrounded by a very dense network of fibers(Figs. 6a and b). Von Kossa staining was positive.
From day 12 until day 21, the nodules increased insize. Alkaline phosphatase activity and Von Kossastaining of a nodule cultured for 21 days are shown in
ARTICLE IN PRESS
Enzymatically isolated osteoblastic cells
4 da
ys
14 d
ays
21 d
ays0
5
10
15
20
25
30
35Calvaria cells grown in standardculture medium (P0)
Calvaria cells grown in standardculture medium supplemented with-GP (P0)
Long bone cells grown in standardculture medium (P0)
Long bone cells grown in standardculture medium supplemented with-GP (P0)
Fetal osteoblastic cells after respectively 4, 14 and 21days in osteogenic medium
Alk
alin
e p
ho
sph
atas
e ac
tivi
ty:
mM
pN
P/m
g p
rote
in
Fig. 5. Alkaline phosphatase activity (mm pNP/mg protein) of fetal osteoblastic cells enzymatically isolated from calvaria or long bones and
subcultured for 21 days in osteogenic medium. Results are the mean7SEM of two independent experiments in duplicate.
Fig. 6. Enzymatically isolated osteoblastic cells cultured for 4 days (a,c) and 21 days (b,d) in osteogenic medium. (a,b) fetal rat calvaria cells; (c,d)
fetal rat long bone cells. Phase contrast microscopy.
H. Declercq et al. / Biomaterials 25 (2004) 757–768 763
Figs. 7a and b. The nodule stained intensely for alkalinephosphatase. Phosphate deposits were restricted to thethree-dimensional area of the nodule. The presence of anintense network of collagen is shown in Fig. 7c.Ultrastructural analysis of a nodule by electron micro-scopy revealed that below the upper osteoblast layernumerous, well-banded collagen fibrils are present.Vesicular structures (loaded with minerals), whichappear to be the initial sites of mineral deposition, werepresent between the collagen fibrils as shown in Fig. 7d.
The IR spectrum clearly demonstrates that themineral phase in these cultures is a B-type carbonatedapatite (Fig. 3b). The asymmetric bending vibration ofPO4
3� is splitted in two distinct peaks at 571 and601 cm�1 and the PO4
3� absorption band in the region
950–1300 cm�1 is comparable to that of the apatitespectrum in Fig. 3e with absorptions around 962, 1040and 1095 cm�1. As the protein fraction in the sampleapparently is relatively low, not only the bendingvibration of CO3
2�at 872 cm�1 is clearly seen, but alsothe stretching vibration at 1420 cm�1 corroborating thatthe mineral phase is a B-type carbonated apatite [14,15].
An increased concentration of l-ascorbic acid 2-phosphate (250 mm) in secondary culture did not increasethe alkaline phosphatase activity and the amount ofnodules. On the contrary, a significant increase in thenumber of nodules for calvaria cells grown for 21 daysin dexamethasone supplemented medium was observed.The number of nodules in the presence of dexametha-sone was increased from 572 to 4175 nodules/cm2.
ARTICLE IN PRESS
Fig. 7. Enzymatically released calvaria cells cultured in osteogenic medium for 21 days. Nodule stained for alkaline phophatase activity (a),
phosphate deposits by Von Kossa (b) and collagen by Trichrome Masson (c). Light microscopy. Electron micrograph of a nodule (d).
H. Declercq et al. / Biomaterials 25 (2004) 757–768764
3.7. Enzymatically released osteoblastic cells from fetal
long bones
Primary cultures were also grown either in standardculture medium or in standard culture medium supple-mented with b-glycerophosphate. In parallel with theenzymatically released fetal calvaria cells, no differenceswere observed between these two experiments. Primarycultures of osteoblastic cells reached confluence after 10days and yield a cell amount of approximately 2,000,000cells/T25 tissue culture dish.
After subculture in osteogenic medium alkalinephosphatase activity showed no statistically significantdifferences between these two experiments. Osteoblasticcells cultured for 4 days in osteogenic medium had ahigh alkaline phosphatase activity (>15 mm pNP/mgprotein) (Fig. 5) but showed a decrease after 14 and 21days.
The cells showed an elongated morphology at lowdensity (Fig. 6c) and a polygonal aspect at confluency.Sporadically, cells are grown in network-like structuresand nodules at higher densities. Mineralization startedat day 14 as demonstrated by Von Kossa. In general, thecalcium phosphate deposits were diffuse and throughoutthe culture (Fig. 6d).
IR spectrometry demonstrates that the mineral phasegenerated by the osteoblastic cells is a carbonate-containing apatite (Fig. 3c). The spectrum shows theabsorptions of PO4
3� vibrations in an apatitic latticearound 600 and 1050 cm�1 next to the typical proteinabsorption peaks. The presence of a peak around872 cm�1 indicates that the apatite contains CO3
2�
substituting for PO43� so that the mineral phase is a
B-type carbonated apatite [14,15].No increase in alkaline phosphatase activity or nodule
formation was observed in cell cultures incubated for 21
ARTICLE IN PRESS
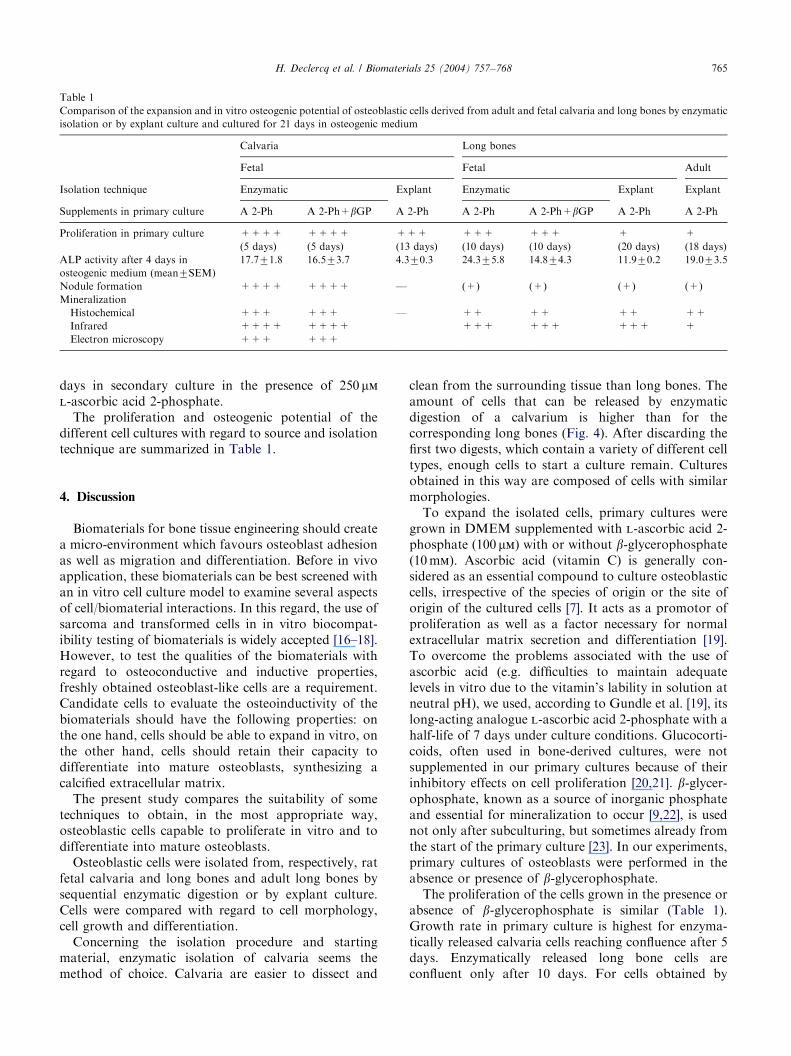
Table 1
Comparison of the expansion and in vitro osteogenic potential of osteoblastic cells derived from adult and fetal calvaria and long bones by enzymatic
isolation or by explant culture and cultured for 21 days in osteogenic medium
Calvaria Long bones
Fetal Fetal Adult
Isolation technique Enzymatic Explant Enzymatic Explant Explant
Supplements in primary culture A 2-Ph A 2-Ph+bGP A 2-Ph A 2-Ph A 2-Ph+bGP A 2-Ph A 2-Ph
Proliferation in primary culture ++++ ++++ +++ +++ +++ + +
(5 days) (5 days) (13 days) (10 days) (10 days) (20 days) (18 days)
ALP activity after 4 days in
osteogenic medium (mean7SEM)
17.771.8 16.573.7 4.370.3 24.375.8 14.874.3 11.970.2 19.073.5
Nodule formation ++++ ++++ — (+) (+) (+) (+)
Mineralization
Histochemical +++ +++ — ++ ++ ++ ++
Infrared ++++ ++++ +++ +++ +++ +
Electron microscopy +++ +++
H. Declercq et al. / Biomaterials 25 (2004) 757–768 765
days in secondary culture in the presence of 250 mml-ascorbic acid 2-phosphate.
The proliferation and osteogenic potential of thedifferent cell cultures with regard to source and isolationtechnique are summarized in Table 1.
4. Discussion
Biomaterials for bone tissue engineering should createa micro-environment which favours osteoblast adhesionas well as migration and differentiation. Before in vivoapplication, these biomaterials can be best screened withan in vitro cell culture model to examine several aspectsof cell/biomaterial interactions. In this regard, the use ofsarcoma and transformed cells in in vitro biocompat-ibility testing of biomaterials is widely accepted [16–18].However, to test the qualities of the biomaterials withregard to osteoconductive and inductive properties,freshly obtained osteoblast-like cells are a requirement.Candidate cells to evaluate the osteoinductivity of thebiomaterials should have the following properties: onthe one hand, cells should be able to expand in vitro, onthe other hand, cells should retain their capacity todifferentiate into mature osteoblasts, synthesizing acalcified extracellular matrix.
The present study compares the suitability of sometechniques to obtain, in the most appropriate way,osteoblastic cells capable to proliferate in vitro and todifferentiate into mature osteoblasts.
Osteoblastic cells were isolated from, respectively, ratfetal calvaria and long bones and adult long bones bysequential enzymatic digestion or by explant culture.Cells were compared with regard to cell morphology,cell growth and differentiation.
Concerning the isolation procedure and startingmaterial, enzymatic isolation of calvaria seems themethod of choice. Calvaria are easier to dissect and
clean from the surrounding tissue than long bones. Theamount of cells that can be released by enzymaticdigestion of a calvarium is higher than for thecorresponding long bones (Fig. 4). After discarding thefirst two digests, which contain a variety of different celltypes, enough cells to start a culture remain. Culturesobtained in this way are composed of cells with similarmorphologies.
To expand the isolated cells, primary cultures weregrown in DMEM supplemented with l-ascorbic acid 2-phosphate (100 mm) with or without b-glycerophosphate(10 mm). Ascorbic acid (vitamin C) is generally con-sidered as an essential compound to culture osteoblasticcells, irrespective of the species of origin or the site oforigin of the cultured cells [7]. It acts as a promotor ofproliferation as well as a factor necessary for normalextracellular matrix secretion and differentiation [19].To overcome the problems associated with the use ofascorbic acid (e.g. difficulties to maintain adequatelevels in vitro due to the vitamin’s lability in solution atneutral pH), we used, according to Gundle et al. [19], itslong-acting analogue l-ascorbic acid 2-phosphate with ahalf-life of 7 days under culture conditions. Glucocorti-coids, often used in bone-derived cultures, were notsupplemented in our primary cultures because of theirinhibitory effects on cell proliferation [20,21]. b-glycer-ophosphate, known as a source of inorganic phosphateand essential for mineralization to occur [9,22], is usednot only after subculturing, but sometimes already fromthe start of the primary culture [23]. In our experiments,primary cultures of osteoblasts were performed in theabsence or presence of b-glycerophosphate.
The proliferation of the cells grown in the presence orabsence of b-glycerophosphate is similar (Table 1).Growth rate in primary culture is highest for enzyma-tically released calvaria cells reaching confluence after 5days. Enzymatically released long bone cells areconfluent only after 10 days. For cells obtained by

ARTICLE IN PRESSH. Declercq et al. / Biomaterials 25 (2004) 757–768766
explant cultures, the time to obtain a sufficient amountof cells took between 13 and 20 days. The effect of theisolation technique correlates with the observations ofVoegele et al. [24] who reported a higher growth rate forosteoblastic cells, obtained by enzymatic treatment oflong bone fragments compared to cells obtained byspontaneous outgrowth.
To favour osteoblastic differentiation, cells weresubcultured in an osteogenic medium (a-MEM supple-mented with l-ascorbic acid 2-phosphate and b-glycer-ophosphate) at a concentration of 40,000 cells/well.Although DMEM as well as a-MEM are used inosteogenic media, a-MEM was preferred for thedifferentiation studies. According to Coelho et al. [23],ALP activity and mineralization capacity are higher ina-MEM compared to DMEM. Both parameters weredetermined after 4, 14 and 21 days in subculture.
The first measurements, performed 4 days post-subculture in the osteogenic medium, show that startingmaterial as well as isolation technique influence the ALPlevels of the subcultures (Figs. 1 and 5). Cells originatingfrom explants of fetal calvaria showed the lowest levels,significantly different from cultures originating fromexplants of fetal long bones and adult long bones.Although obtained from the same animals, ALP levelsin subcultures obtained by enzymatic treatment of fetalcalvaria and long bones were higher. Hence, theisolation procedure, explant versus enzyme treatment,apparently influences the ALP levels at the start of thesubcultures, especially for fetal calvaria compared tolong bones. An explanation could be that an enzymaticdigestion offers the possibility to work with morepurified ‘osteoblastic’ cultures by discarding the firstdigests. Especially in explant cultures of calvaria, a largeamount of contaminating cells could origin from notcompletely removed periosteum, rich in uncommittedmesenchymal cells and fibroblasts.
During the extended subculture period in osteogenicmedium (performed up to 21 days), markers of finalosteoblastic differentiation, like mineralization of acollagen-containing extracellular matrix, become ob-vious. According to Aronow et al. [25], the onset ofmineralization in rat osteoblastic cultures only startsafter sufficient upregulation of ALP activity. After‘extensive’ mineralization, a decline of ALP activity inturn is noted. Also in our cultures, a correlation wasobserved between levels of ALP activity and theability to produce a mineralized matrix. In thecultures obtained by subculturing cells from outgrowthsof calvaria, ALP levels may not be high enough toinitiate mineralization. The cultures from explants oflong bones showed mineralization towards the end ofthe culture period. In correlation with the elevated ALPlevels at the start of the subcultures, the mostpronounced calcification is found in the enzymaticallyobtained cultures.
With Von Kossa staining, striking differences inmineralization patterns between the different culturesare observed. Cultures obtained from long bones show amore dispersed mineralization pattern, in contrast to themineralization in the calvarial cultures, which isrestricted to foci of well-defined multilayered nodules.
Descriptions concerning mineralization patterns incorrelation to different cell cultures (with regard tosource of the cells, type and age of the animal) arevariable. The formation of mineralized nodules incalvaria-derived cell populations is commonly described[9,10,22,25,26]. However, for long bone cell populations,mineralization restricted to well-defined and sharplymarginated nodules [7,27–29] as well as mineralizationin ill-defined deposits throughout the whole culture[19,24,30] are described.
In our experiments, cell cultures originating fromdifferent sources from the same animal were comparedside by side for their osteogenic potential. For 21-dayold fetal rats, the presence of nodules in calvaria-derivedcultures in contrast to their absence in long bone-derivedcultures could be explained as follows. In calvaria of 21-day old fetal rats the bone pieces are still connected byfibrous sutures, containing preosteoblastic cells andpossible colony forming unit-osteogenic (CFU-O). Thelatter cells, absent in the long bones, are the potentialnodule forming cells. Nodules are the result of theclonogenic potential of cells. It is assumed that eachnodule in vitro arises from the clonogenic potential of 1cell, described as a CFU-O [7,26]. Since the formation ofnodules is absolutely dependent on the maintenance ofadequate levels of vitamin C, the effect of an elevatedlevel of l-ascorbic acid 2-phosphate (250 mm) onthe nodule formation was examined. No increase inthe number of nodules was observed compared to thecontrol conditions (100 mm). Dexamethasone on theother hand caused a significant increase in the numberof nodules in the calvaria-derived cultures. As describedearlier, the degree of susceptibility to glucocorticoidtreatment depends on the developmental stage of thecells and is also species dependent. In agreement to ourobservations, some authors [7,20,31] describe a stimu-latory effect of dexamethasone on the amount ofnodules in cell cultures obtained from fetal rat calvaria.It must be emphasized however that infrared spectro-metry revealed that despite the differences in themineralization patterns, the mineral deposits in bothsystems appear to correspond to B-type carbonatedapatite (Fig. 3). However, this does not mean that thechemical composition of the apatitic phase is the same.In fact, differences in the chemical composition of thecarbonated apatite due to differences in the Na, CO3
and HPO4 content and caused by differences inmineralization, could occur [14,15]. Concerning themineralization pattern, the deposition of mineral depos-its which remains restricted to the nodules is again

ARTICLE IN PRESSH. Declercq et al. / Biomaterials 25 (2004) 757–768 767
indicative for the nature of these nodules, arising fromosteoprogenitor cells from the osteogenic lineage and beable to differentiate in mature osteoblasts under appro-priate culture conditions (osteogenic medium). On thecontrary, long bone-derived osteoblasts have lost thiscapability of forming nodules but still have the capacityto deposit minerals in a matrix, resulting in the dispersedmineralization pattern observed in the cultures.
The present study demonstrates that, in our experi-mental set-ups, cells obtained enzymatically from fetalrat calvaria are the most appropriate to study interac-tions with biomaterials in vitro. Calvaria cells have ahigh proliferation capacity, being able to expand in vitro.With regard to differentiation, they have a moderate tohigh alkaline phosphatase activity and the highestability to from well-defined three-dimensional noduleswhich can mineralize under conditions which favourosteogenesis. They will be used to study the osteocon-ductive and inductive capacities of candidate materialsfor bone tissue engineering.
Acknowledgements
The work was supported by a fund of the GhentUniversity (GOA project 2001, No 12050701). Theauthors thank N. Fran@ois, L. Pieters, J. Aernoudt,R. De Vos and C. Vercruysse for technical assistance.
References
[1] Elgendy HM, Norman ME, Keaton AR, Laurencin CT. Osteo-
blast-like cell (MC3T3-E1) proliferation on bioerodible polymers:
an approach towards the development of a bone-bioerodible
polymer composite material. Biomaterials 1993;14(4):263–9.
[2] Ahmad M, Mc Carthy MB, Gronowicz G. An in vitro model for
mineralization of human osteblast-like cells on implant materials.
Biomaterials 1999;20:211–20.
[3] Lo H, Kadiyala S, Guggino SE, Leong KW. Poly(l-lactic acid)
foams with cell seeding and controlled-release capacity. J Biomed
Mater Res 1996;30:475–84.
[4] Matsuura T, Hosokawa R, Okamoto K, Kimoto T, Akagawa Y.
Diverse mechanisms of osteoblast spreading on hydroxyapatite
and titanium. Biomaterials 2000;21:1121–7.
[5] Miller MD, Puzas JE. Differential growth factor effects on
normal and transformed rat bone cells. J Bone Miner Res
1988;3(1):S201.
[6] Rifas L, Halstead LR, Scott MJ, Fedarko NS, Avioli LV, Gehron
Robey P. Comparison of secretory products of normal rat
osteoblasts and osteogenic sarcoma cells in vitro. J Bone Miner
Res 1989;4(1):S409.
[7] Aubin JE, Herbertson A. Osteoblast lineage in experimental
animals. In: Beresford JN, Owen ME, editors. Marrow stromal
cell culture. Cambridge: Cambridge University Press; 1998.
[8] Freshney RI. Culture of specific cell types, mesenchymal
cell, bone. In: Schwartz E, editor. Culture of animal cells. A
manual of basic technique, 2nd ed. New York: Alan R. Liss, Inc.;
p. 273–5.
[9] Bellows CG, Aubin JE, Heersche JNM, Antosz ME.
Mineralized bone nodules formed in vitro from enzymatically
released rat calvaria cell populations. Calcif Tissue Int
1986;38:143–54.
[10] Bellows CG, Aubin JE, Heersche JNM. Initiation and progression of
mineralization of bone nodules formed in vitro: the role of alkaline
phosphatase and organic phosphate. Bone Miner 1991;14:27–40.
[11] Ishida H, Bellows CG, Aubin JE, Heersche JNM. Characteriza-
tion of the 1,25-(OH)2D3-induced inhibition of bone nodule
formation in long-term cultures of fetal rat calvaria cells.
Endocrinology 1993;132(1):61–6.
[12] Furuya K, Nifuji A, Rosen V, Noda M. Effects of GDF7/BMP12
on proliferation and alkaline phosphatase expression in rat
osteoblastic osteosarcoma ROS 17/2.8 cells. J Cell Biochem
1999;72:177–80.
[13] B .ock P. Der Semid .unnschnitt. M .unchen: J.F. Bergmann Verlag;
1984. p. 98–9.
[14] Pieters IY, De Maeyer EAP, Verbeeck RMH. Influence of Na+
on the stoichiometry of carbonated hydroxyapatite obtained by
the hydrolysis of octacalcium phosphate. Inorg Chem
1998;37:6392–5.
[15] Driessens FCM, Verbeeck RMH. Biominerals. Boca Raton, FL:
CRC Press; 1990.
[16] Morrison C, Macnair R, MacDonald C, Wijkman A, Goldie I,
Grant MH. In vitro biocompatibility testing of polymers for
orthopaedic implants using cultured fibroblasts and osteoblasts.
Biomaterials 1995;16:987–92.
[17] Ignatius AA, Claes LE. In vitro biocompatibility of bioresorbable
polymers: poly(l, dl-lactide) and poly(l-lactide-co-glycolide).
Biomaterials 1996;17:831–9.
[18] Moreau MF, Chappard D, Lesourd M, Month!eard JP, Basl!e MF.
Free radicals and side products released during methylmethacry-
late polymerization are cytotoxic for osteoblastic cells. J Biomed
Mater Res 1998;40:124–31.
[19] Gundle G, Stewart K, Screen J, Beresford JN. Isolation and
culture of human bone-derived cells. In: Beresford JN, Owen ME,
editors. Marrow stromal cell culture. Cambridge: Cambridge
University Press; 1998.
[20] Beresford JN, Graves SE, Smoothy CA. Formation of miner-
alised nodules by bone derived cells in vitro: a model of bone
formation? Am J Med Gen 1993;45:163–78.
[21] Wong MM, Rao LG, Ly H, Hamilton L, Tong J, Sturtridge W,
Mc Broom R, Aubin JE, Murray TM. Long-term effects of
physiologic concentrations of dexamethasone on human bone-
derived cells. J Bone Miner Res 1990;5:803–13.
[22] Bellows CG, Heersche JNM, Aubin JE. Inorganic phosphate added
exogenously or released from b-glycerophosphate initiates miner-
alization of osteoid nodules in vitro. Bone Miner 1992;17:15–29.
[23] Coelho MJ, Cabral AT, Fernandes MH. Human bone cell
cultures in biocompatibility testing. Part I: osteoblastic differ-
entiation of serially passaged human bone marrow cells cultured
in a-MEM and in DMEM. Biomaterials 2000;21:1087–94.
[24] Voegele TJ, Voegele-Kadletz M, Esposito V, Macfelda K,
Oberndorfer U, Vecsei V, Schabus R. The effect of different
isolation techniques on human osteoblast-like cell growth. Antic-
ancer Res 2000;20:3575–82.
[25] Aronow MA, Gerstenfeld LC, Owen TA, Tassinari MS, Stein GS,
Lian JB. Factors that promote progressive development of the
osteoblast phenotype in cultured fetal rat calvaria cells. J Cell
Physiol 1990;143:213–21.
[26] Malaval L, Liu F, Roche P, Aubin JE. Kinetics of osteopro-
genitor proliferation and osteoblast differentiation in vitro. J Cell
Biochem 1999;74:616–27.
[27] Halstead LR, Scott MJ, Rifas L, Avioli LC. Characterization of
osteoblast-like cells from normal adult rat femoral trabecular
bone. Calcif Tissue Int 1992;50:93–5.
[28] Manduca P, Palermo C, Caruso C, Brizzolara A, Sanguineti C,
Filanti C, Zicca A. Rat tibial osteoblasts III: propagation in vitro

ARTICLE IN PRESSH. Declercq et al. / Biomaterials 25 (2004) 757–768768
is accompanied by enhancement of osteoblast phenotype. Bone
1997;21(1):31–9.
[29] Stringa E, Filanti C, Giunciuglio D, Albini A, Manduca P.
Osteoblastic cells from rat long bone. I. Characterization of their
differentiation in culture. Bone 1995;16(6):663–70.
[30] Siggelkow H, Rebenstorff K, Kurre W, Niedhart C, Engel I,
Schulz H, Atkinson MJ, H .ufner M. Development of the
osteoblast phenotype in primary human osteoblasts in culture:
comparison with rat calvarial cells in osteoblast differentiation.
J Cell Biochem 1999;75:22–35.
[31] Bellows CG, Aubin JE, Heersche JNM. Physiological con-
centrations of glucocorticoids stimulate formation of bone
nodules from isolated rat calvaria cells in vitro. Endocrinology
1986;121:1985–92.
Related Documents











