System. App!. Microbio!. 21, 13S-143 (1998) SYSTEMATIC AND _©_G_us_ta_v F_is_ch_er_V_erl_a g ________________ APPLIED MICROBIOLOGY Isolation and Characterisation of Obligately Anaerobic, Lipolytic Bacteria from the Rumen of Red Deer GRAEME N. JARVIS l , CARSTEN STROMPL 1 , EDWARD R. B. MOORE 2 , and JORGEN H. THlELE1 1 Waste Technology Research Group, Department of Microbiology, University of Otago, Dunedin, New Zealand 2 Division of Microbiology, National Research Centre for Biotechnology, Braunschweig, Germany Received November 11, 1997 Summary Two Gram-positive, obligately anaerobic, lipolytic bacteria, isolates LIP4 and LIPS, were obtained from the rumen contents of juvenile red deer. These mesophilic bacterial strains were capable of hydrolysing the neutral lipids, tallow, tripalmitin and olive oil, into their constituent free long-chain fatty acid and glycerol moieties. The latter compound was dissimilated by both isolates, with isolate LIP4 producing propionate as the predominant product, while isolate LIPS produced acetate, ethanol and succinate. The lactate-utilising isolate LIP4 grew on a limited range of saccharide substrates including glucose, fructose and ribose, and exhibited an unusual cell wall structure and morphology. The isolate LIPS grew upon a wider range of saccharides, but was unable to use lactate as a substrate. Based upon pheno- typic and 16S rRNA gene sequence analyses, isolate LIP4 clusters with species in the genus Propionibacterium, while isolate LIPS is a member of clostridial cluster XIVa. Key words: Deer, rumen, lipolytic bacteria, lipase, tallow, Clostridium, Propionibacterium. Introduction Anaerobic lipid hydrolysis has been studied more ex- tensively in the rumen than in any other anaerobic ecosystem. However, to date, only two obligately anaero- bic rumen bacteria occupying the lipid-hydrolysing eco- logical niche have been isolated. Lipids comprise 2-10% of the total dry weight of foraged material ingested by ruminant animals (HARFOOT, 1978; HARFOOT and HA- ZLEWOOD, 1988; MACKIE et al., 1991). While this lipid content may be considered to be relatively low, it should be noted that calculations indicate that as much as 1500 g of lipid (dry weight) may be ingested per day by lactat- ing dairy cows (HARFOOT, 1978). It is within the rumen that dietary lipids derived from plant material are metabolised upon entering the animal, and it is microbial activity which is responsible for this process occurring (HARFOOT, 1978; HARFOOT and HAZLEWOOD, 1988; MACKIE et al., 1991). The lipids are hydrolysed by fer- mentative bacteria generating long chain- and medium chain-length free fatty acids, and glycerol and galactose. The glycerol and galactose are then readily fermented to volatile fatty acids (VFA), CO2 and H2 (HARFOOT, 1978; HAZLEWOOD and DAWSON, 1979; LOUGH and GARTON, 1968). The two obligately anaerobic rumen bacteria known to occupy the lipid-hydrolysing ecological niche, Anaerovibrio lipolytica and Butyrivibrio fibrisolvens strains 52, were isolated from the rumen of sheep, and their lipid-hydrolysing activities have been studied exten- sively (HOBSON and MANN, 1961; HAZLEWOOD and DAW- SON, 1979; HENDERSON, 1971; PRINS et al., 1975). A. lipolytica has been reported to be present in the rumen of sheep at population densities in the range of 0.5-1.1x1 0 7 cells ml- 1 and demonstrated a rate of trilinolein hydroly- sis which was of a magnitude sufficient to account for the total rates of hydrolysis of esterified fatty acids with- in the sheep rumen (HARFOOT, 1978). Here, we report the isolation, physiological descrip- tion, and phylogenetic classification of two obligately anaerobic, fermentative, mesophilic, lipolytic bacteria that were capable of growth in minimal media contain- ing no rumen fluid. These organisms were isolated from the rumen contents of red deer and are representatives of bacterial genera which have not been previously isolated from this ecosystem. Materials and Methods Chemicals, media and cultivation techniques: All chemicals used in the current study were of analytical grade and were ob- tained from Sigma Chemical Co., (St. Louis, USA) unless other- wise stated. The basal minimal medium (BMM) used for the initial en- richment of the bacteria was carried out using a modified ver- sion of a phosphate-buffered basal medium described previous- ly (KENEALY and ZEIKlIS, 1981), containing NaHC01 (4.2 g 1-1),

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

System. App!. Microbio!. 21, 13S-143 (1998) SYSTEMATIC AND _©_G_us_ta_v F_is_ch_er_V_erl_ag ________________ APPLIED MICROBIOLOGY
Isolation and Characterisation of Obligately Anaerobic, Lipolytic Bacteria from the Rumen of Red Deer
GRAEME N. JARVIS l , CARSTEN STROMPL1, EDWARD R. B. MOORE2, and JORGEN H. THlELE1
1 Waste Technology Research Group, Department of Microbiology, University of Otago, Dunedin, New Zealand 2 Division of Microbiology, National Research Centre for Biotechnology, Braunschweig, Germany
Received November 11, 1997
Summary
Two Gram-positive, obligately anaerobic, lipolytic bacteria, isolates LIP4 and LIPS, were obtained from the rumen contents of juvenile red deer. These mesophilic bacterial strains were capable of hydrolysing the neutral lipids, tallow, tripalmitin and olive oil, into their constituent free long-chain fatty acid and glycerol moieties. The latter compound was dissimilated by both isolates, with isolate LIP4 producing propionate as the predominant product, while isolate LIPS produced acetate, ethanol and succinate. The lactate-utilising isolate LIP4 grew on a limited range of saccharide substrates including glucose, fructose and ribose, and exhibited an unusual cell wall structure and morphology. The isolate LIPS grew upon a wider range of saccharides, but was unable to use lactate as a substrate. Based upon phenotypic and 16S rRNA gene sequence analyses, isolate LIP4 clusters with species in the genus Propionibacterium, while isolate LIPS is a member of clostridial cluster XIVa.
Key words: Deer, rumen, lipolytic bacteria, lipase, tallow, Clostridium, Propionibacterium.
Introduction
Anaerobic lipid hydrolysis has been studied more extensively in the rumen than in any other anaerobic ecosystem. However, to date, only two obligately anaerobic rumen bacteria occupying the lipid-hydrolysing ecological niche have been isolated. Lipids comprise 2-10% of the total dry weight of foraged material ingested by ruminant animals (HARFOOT, 1978; HARFOOT and HAZLEWOOD, 1988; MACKIE et al., 1991). While this lipid content may be considered to be relatively low, it should be noted that calculations indicate that as much as 1500 g of lipid (dry weight) may be ingested per day by lactating dairy cows (HARFOOT, 1978). It is within the rumen that dietary lipids derived from plant material are metabolised upon entering the animal, and it is microbial activity which is responsible for this process occurring (HARFOOT, 1978; HARFOOT and HAZLEWOOD, 1988; MACKIE et al., 1991). The lipids are hydrolysed by fermentative bacteria generating long chain- and medium chain-length free fatty acids, and glycerol and galactose. The glycerol and galactose are then readily fermented to volatile fatty acids (VFA), CO2 and H2 (HARFOOT, 1978; HAZLEWOOD and DAWSON, 1979; LOUGH and GARTON, 1968). The two obligately anaerobic rumen bacteria known to occupy the lipid-hydrolysing ecological niche, Anaerovibrio lipolytica and Butyrivibrio fibrisolvens strains 52, were isolated from the rumen of sheep, and their lipid-hydrolysing activities have been studied exten-
sively (HOBSON and MANN, 1961; HAZLEWOOD and DAWSON, 1979; HENDERSON, 1971; PRINS et al., 1975). A. lipolytica has been reported to be present in the rumen of sheep at population densities in the range of 0.5-1.1x1 07
cells ml- 1 and demonstrated a rate of trilinolein hydrolysis which was of a magnitude sufficient to account for the total rates of hydrolysis of esterified fatty acids within the sheep rumen (HARFOOT, 1978).
Here, we report the isolation, physiological description, and phylogenetic classification of two obligately anaerobic, fermentative, mesophilic, lipolytic bacteria that were capable of growth in minimal media containing no rumen fluid. These organisms were isolated from the rumen contents of red deer and are representatives of bacterial genera which have not been previously isolated from this ecosystem.
Materials and Methods
Chemicals, media and cultivation techniques: All chemicals used in the current study were of analytical grade and were obtained from Sigma Chemical Co., (St. Louis, USA) unless otherwise stated.
The basal minimal medium (BMM) used for the initial enrichment of the bacteria was carried out using a modified version of a phosphate-buffered basal medium described previously (KENEALY and ZEIKlIS, 1981), containing NaHC01 (4.2 g 1-1),

136 G. N. JARVIS et al.
in place of NaCI, and yeast extract (0.01 %, w/v). Glycerol (100 or 150 mM) was added as the major growth substrate, and during the initial enrichment phase, sodium acetate (10 mM) was also included. Lipolytic activity in a complex medium was determined using Modified Brain Heart Infusion (MBHI) broth, composed of (g I-lor mll-1): Brain Heart Infusion (Difco, MI), 37.1; gelatin (Difco), 2.5; tripalmitin, 2.5. Hungate tubes (Bellco, NJ) containing 10 ml of media, were maintained with a headspace of a CO2:N2 (50:50, v/v). Stringent anaerobic cultivation, media preparation, and culture transfer techniques (HUNGATE, 1966) were maintained at all times. Solid media (agar, 2%, w/v) were prepared in an anaerobic chamber (Forma Scientific, 'USA) with a gas phase of N2:C02:H2 (85:5:10, v/v/v). All experimental incubations were carried out at 37°C, with broth cultures being incubated horizontally on an orbital shaker (120 rpm). All media had a final pH of 7.2-7.4 after autoclaving. Olive oil (triolein), tripalmitin and tallow were used as neutral lipid growth substrates. Tallow was obtained from a slaughterhouse offal rendering factory (Fortex, NZ). Elemental analysis (n = 3) of the tallow indicated that it was composed of carbon (76.4 ± 0.3%), oxygen (11.5 ± 0.3%) and hydrogen (12.0 ±
0.3%), and had a moisture content of 0.25 ± 0.7% (w/v), based on triplicate results obtained from weight percentage data. Sulfur, nitrogen, and phosphorous levels in the tallow were less than 0.1-0.2%. The cholesterol content of the tallow was found to be 223 ± 17 mg per 100 g tallow (n = 3), using an enzymatic test kit according to the manufacturers instructions (Boehringer-Mannheim, Germany). Analysis of the profile of long chain fatty acids (LCFA) indicated that the tallow was composed primarily of oleic (42.0 ± 0.9%), stearic (29.1 ±
0.8%) and palmitic (20.9 ± 0.1 %) acids (JARVIS and THIELE, 1997a). The tallow (10 g I-I) was emulsified by partial saponification under stringent anaerobic conditions (JARVIS, 1995), and autoclaved prior to use in experiments. The content of free LCFA in the tallow emulsion was 0.47 ± 0.12 g I-I (n = 3).
Source of isolates and isolation procedures: The isolates were obtained from red deer (Cervus elaphus) at the Invermay Agricultural Research Station in Dunedin, New Zealand. Three eight-month old deer were culled and the rumen contents were collected in pre-autoclaved 150 ml serum bottles (Alltech Associates, USA) containing an oxygen-free nitrogen gas atmosphere. Each bottle was filled to the top and sealed with 20 mm butyl rubber stoppers (Belleo, NJ) to minimise oxygen contamination during transport to the laboratory. The three rumen samples were pooled and transferred to medium in Hungate tubes at a 10% (v/v) inoculum level. The enrichment involved a serial dilution (10-1-10-10) in Hungate tubes containing glycerol-BMM with an initial 10% (v/v) inoculum of the pooled deer rumen contents, and subsequent 10% (v/v) serial transfers. After three consecutive passages in liquid medium, samples from the cultures were streak-plated onto rhodamine B-olive oil BMM agar (JARVIS and THIELE, 1997b). Individual lipolytic colonies were selected based on their expression of qualitative lipase activity as indicated by the formation of a rhodamine BLCFA conjugate (KOUKER and JAEGER, 1987).
Growth experiments, biochemical and physiological characterisation: Growth in liquid culture was followed by spectrophotometric measurement of optical density at 600 nm. Utilisation of various organic substrates as carbon and energy sources was determined using the bioMerieux API 20A test kit according to the manufacturer's instructions (bioMerieux, France). Both isolates were tested for their ability to reduce sulfate, by adding FeS04. 7 H20 (2 g 1-1) to glycerol-BMM and examination of the tubes for blackening of the medium (due to H2S formation) during incubation at 37°C. Determination of growth rates, on different carbon sources was undertaken (in duplicate), by the ad-
dition of organic substrates (glucose, fructose, glycerol, ribose) at equimolar concentrations (100 mM) to BMM broths. The growth rates of the isolates on lipid substrates were determined in both MBHI broths (containing tripalmitin), and BMM broths containing tallow (10 g I-I). The effect of yeast extract upon growth and metabolism of the isolates was determined in the glycerol-BMM medium (without the sodium acetate supplement added) and the tallow media incubated at 37°C, in the presence or absence of 0.01 % (w/v) yeast extract.
Analytical procedures: Culture sub-samples (1 ml) were centrifuged (15000xg, 15 minutes) and the pellets were assayed for bacterial cell protein using a modified version of the Lowry protein assay (HARTREE, 1972). Glycerol concentration in culture supernatants was determined using a previously published method (JARVIS et aI., 1997). Volatile fatty acid and alcohol concentrations in the culture supernatants, and culture headspace compositions were analysed using established techniques (JARVIS et aI., 1997). Long chain fatty acids in culture sub-samples were quantified after derivatisation to nitrophenylhydrazides, and separation by HPLC (JARVIS and THIELE, 1997a).
Quantitative esterase assays were performed at 37°C, using the method of WINKLER and STUCK MANN (1979). The quantitative, total lipase activities were determined using the pure bacterial isolates grown anaerobically, at 37°C, in BMM broth containing the emulsified tallow (10 g I-I), and the MBHI broth with the tripalmitin (2.5 g I-I) as the lipid substrate. The liberation of LCFA from the neutral lipid substrate and bacterial protein levels were assayed to determine specific enzyme activities. Both esterase and lipase assays used exponential-phase cells from glycerol-grown cultures as the inoculum. For esterase and lipase assays, one enzyme unit (U) was defined as the amount of enzyme required to release 1 nmol of p-nitrophenol (pNPP) per minute or 1 IImol of LCFA per minute, respectively. The cells were washed anaerobically in sterile saline (0.9% w/v) to avoid glycerol carry-over with the inocula for each assay.
Bacterial morphology and cell characteristics: Cell morphology was examined by light microscopy using an Olympus Vanox microscope, and utilising Gram-, spore- and capsule stains as described elsewhere (DOETSCH, 1981). Motility of cultures grown in glycerol-BMM was determined using the hanging drop technique (DOETSCH, 1981). Culture purity was assessed microscopically by growing broth cultures in glycerol- or glucose-BMM and, after growth in MBHI broth, subsequent streak-plating onto BHI agar (Difco, MI). Thin section electron microscopy was undertaken on cells obtained from liquid cultures in the late log phase of growth. Samples were prepared using standard techniques (JARVIS, 1995), and were examined using an Akashi EM-002A transmission electron microscope.
Determination and analysis of 16S rRNA gene sequences: Isolation of genomic DNA from late-exponential phase cultures of isolates LIP4 and LIP5 was carried out using a protocol described previously (WILSON, 1987). Nearly complete PCR-amplified 16D rDNA was generated using a forward primer, hybridizing at the complement of positions 8-27 (E. coli 16S rRNA gene sequence numbering), and a reverse primer hybridizing at positions 1541-1525. PCR was carried out using a GeneAmp 9600 (Perkin-Elmer Corp., Conn. USA), and conditions as reported previously (KARLSON et aI., 1993). PCR-generated products were purified using Centricon-l00 microconcentrators (Amicon GmbH, Witten, FRG) and sequenced directly using a "Taq cycle-sequencing" reaction with fluorescent dyelabelled dideoxynucleotides, as recommended by the manufacturer (Perkin-Elmer Applied Biosystems GmbH, Weiterstadt, FRG) for the ABI 373A Nucleic Acid Sequencer. Sequence data was aligned with reference rRNA (and rRNA gene sequences) (MAIDAK et aI., 1997; STOESSER et aI., 1997), using the evolu-

tionary conserved primary and secondary structure as references (NEEFS et al., 1990). Evolutionary distances (JUKES and CANTOR, 1969) were calculated from sequence dissimilarities, using only unambiguously-determined nucleotide positions. The dendrogram was generated using the FITCH programme in the PHYLIP (Phylogeny Inference Package, Version 3.Sc) software package (FELSENSTEIN, 1989).
Nucleotide sequence accession numbers: The 16S rDNA sequences for isolates LIP4 and LIPS have been deposited with the EMBL nucleic acid sequence database under the accession numbers Y12288 and Y12289 respectively.
Results and Discussion
Isolation and characterisation
Using glycerol as the carbon source, two strains of lipolytic anaerobic bacteria were isolated from the pooled rumen contents of three red deer. The isolates, LIP4 and LIPS, originated from the 10-5 and 10-3 dilutions of the rumen contents, respectively. When streakplated onto the rhodamine B-olive-oil based agar, isolates LIP4 and LIPS exhibited qualitative lipase activity. The isolate LIP4 was pleomorphic (0.7-1.5 flm in length by 0.5-.8 flm in width) and formed cell aggregates in latelog phase. Thin section electron micrographs of isolate LIP4 demonstrated an unusual cell wall structure (Fig. 1). No endospores or motility were observed and cells always stained Gram-positive. Small (0.5 mm diameter), white circular colonies with a convex elevation and entire margin were formed on glycerol-BMM agar. The spore-forming isolate LIPS stained Gram-positive and
Lipolytic Bacteria from the Red Deer Rumen 137
formed non-motile rods of size 2.0-3.0 flm by 0.5-0.7 11m when grown with glycerol as the carbon source. Large areas of granulose-like storage material were observed in thin section preparations of this isolate (Fig. 2). The colonies were 1 mm in diameter, colourless, and exhibited an irregular form, flat elevation, and entire margin when grown on GAYE agar.
Both LIP4 and LIPS were strict anaerobes, based upon their inability to grow on GAYE agar incubated aerobically at 37°C, and the failure of the isolates to grow in glycerol-based broth cultures containing a headspace (2 atm.) of sterilised air. The isolates were able to grow with glycerol as the sole carbon and energy source. Both isolates were catalase-negative and did not reduce sulfate to H 2S or form indole. The optimum growth temperature was found to be 37°C for both isolates. Isolate LIP4 utilised a narrower range of saccharides as growth substrate compared to isolate LIPS (Table 1). The isolate LIP4 produced propionate (24 mM) and CO2 (6 mM) as the major products from the dissimilation of glycerol (57 mM). Formate, ethanol, acetate, 1,3-propanediol, lactate, succinate, butyrate and butanol were below detection limits (<1 mM) for the analytical techniques. LIP4 exhibited slow growth rates in defined medium containing glycerol (0.0008 h- i ) and saccharide (glucose, fructose, ribose) substrates (0.002-0.0007 h-1), compared to the tallow-BMM and MBHI media (0.01 and 0.05 h- i respectively). Neither the addition of yeast extract (0.01 %, w/v), nor the addition of 5, 15, or 30% (w(v) clarified rumen fluid stimulated the growth of the isolate LIP4 in the glycerol-BMM.
Fig. 1. Thin section electron microscopy of exponentially growing culture of isolate LIP4 in glycerol-based medium. Note the pleomorphic nature of the cells, and unusual cell wall structure. (Bar = 111m).
Fig. 2. Thin section electron microscopy of a late-log phase culture of isolate LIPS in glycerol-based medium. Note the presence of granular storage material in the cell cytoplasm. (Bar = 1 pm).

138 G. N. JARVIS et al.
Table 1. Comparison of substrate fermentation by the deer rumen isolates LIP4 and LIPS with phenotypically similar species from the genera Clostridium, Anaerovibrio and Propionibacteriuma
Substrate LIP4 LIPS Clostridium Anaerovibrio Propionibacterium celerecrescens lipolytica acnres
Arabinose + Cellobiose + + Cellulose + ND Esculin (hydrolysis) + Fructose + + + + + Gelatin + + Glucose + + + + + Glycerol + + + + Lactate + ND + + Lactose + + Maltose + a a + + Mannitol + + Mannose + + + + Melezitose + + Olive Oilb + + ND + + Raffinose + + Ribose + + + + + Saccharose + + + Salicin + + Tallowh + + ND ND ND Trehalose + + + TripalmitinC + + ND ND ND Xylose +
a Based on duplicate assays for each tested carbon source using API 20A test kit (bioMerieux, France) for isolates LIP4 and LIPS. The data for C. celerecrescens, P. acnes and A. lipolytica adapted from PALOP et al. (1989), CUMMINGS and JOHNSON (1986) and OUATTARA et al. (1992) respectively. All specieslisolates do not ferment sorbitol. b Based upon on bacterial colony growth and fluorescence of colonies on agar for isolates LIP4 and LIPS. Cultures incubated for a minimum time of 7 days at 37°C. C Based upon quantitative measurements of bacterial growth in liquid media cultures and liberation of free LCFA. ND: Not determined.
Table 2. Levels of sequence similarity based on alignment of the full16S rRNA gene sequence of deer rumen isolates LIP4 and LIPS and related organisms from the EBI database.
Organism % Sequence similarity
2 3 4 5 6 7 8 9 10 11 12 13 14
1 LIP4 75.6 75.3 75.3 74.3 75.4 76.6 74.7 76.4 99.6 91.9 87.9 85.7 87.4 2 LIPS 93.5 98.5 98.1 97.7 92.3 88.2 88.2 75.9 75.3 75.6 76.5 77.5 3 Clostridium clostridiiforme 93.5 92.4 93.4 92.1 88.5 87.3 75.7 75.4 75.4 76.9 77.1 4 C. celerecrescens (DSM 5628T ) 98.8 97.7 91.9 88.2 88.3 75.7 75.0 75.7 76.6 77.6 5 C. sphenoides (DSM) 632T ) 97.0 91.0 87.2 87.3 74.7 74.0 74.7 75.6 77.0 6 C. aerotolerans (DSM 5434T ) 92.3 88.2 87.9 75.7 75.1 75.9 76.6 77.8 7 C. nexile (DSM 1787T ) 86.6 87.2 77.0 76.1 75.2 77.4 77.3 8 Butyrivivibrio fibrisolvens g5.9 75.1 75.3 74.8 75.6 75.9
(ATCC 19171T )
9 Acetitomaculum ruminis 76.8 76.7 77.0 77.9 77.9 (ATCC 43876T )
10 Propionibacterium acnes 92.4 88.1 86.0 87.8 11 Propionibacterium freudenreichii 88.6 86.7 87.3 12 Nocardioides simplex 88.7 89.0 13 Rhodococcus erythropolis 88.7
(DSM 43066T )
14 Micrococcus luteus
T Indicates that the sequence data are obtained from the type culture.

Fig. 3. Phylogenetic dendrogram showing the positions of isolates LIP4 and LIPS within the radiation of the two genera Clostridium and Propionibacterium. The scale bar represents 1 nucleotide substitution per 100 nucleotides. E. coli has been
.-
,
included as the outgroup. 1.0 %
Isolate LIPS exhibited a faster growth rate when grown in glycerol- (0.08 h- I ) and MBHI-containing media (0.12 h-1), compared to tallow-based medium (0.02 h-1) or ribose (0.007 h-1). However, it exhibited similar growth rates with glucose and fructose (0.05 and 0.07 b-1 respectively), that were faster compared to those established for LIP4. Interestingly, in comparisOll to LIP4 and LIPS, A. lipolytica demonstrated a reported growth rate of 0.2 h-1 when grown on glycerol in a continuous culture chemostat study (HOBSON and SUMMERS, 1966). However, in that study, both casitone (0.1 %, w/v) and yeast extract (0.1 %, w/v) were present in the glycerol medium and it has been established that both of these components are known to improve the growth of A. lipolytica (PRINS et al., 1975). In contrast to LIP4, isolate LIPS produced ethanol (56 mM), acetate (40 mM), and succinate (28 mM) as the major fermentation products from glycerol (125 mM), with CO2 (12 mM) and butyrate (4 mM) also being produced.
Phenotypic data on isolate LIP4 indicated that it was similar to species of the Anaerovibrio and Propionibacterium genera. These genera are believed to occupy the ecological niches of lactate utilisation (both genera) and lipid hydrolysis (Anaerovibrio only) in the rumen (STEWART and BRYANT, 1988). Interestingly, A. lipolytica, P. acnes and LIP4 grew on similar carbod substrates (Table 1). Isolate LIP4 was able to utilise mannose and trehalose, as well as hydrolyse esculin, in contrast to
-
r-
1
Lipolytic Bacteria from the Red Deer Rumen 139
ostn lum Cl 'd' sphenoides
Clostridium celerecrescens
-LIPS
~ Clostridiu m aerotolerans
Clostridiu m clostridiiforme
nexile Clostridium
Butyri
Acetitomac
vibrio jlbrisolvens
ulum ruminis
Clostridium but vricum
Clostridium hasti iforme
Bacillus subtilis
I I
IP ropionibacterium acnes
LIP 4
Noca
Propionibacterium freudenreichii
rdioides simplex
Rhodoc occus erythropolis
Cory nehacterium glutamicum
Micrococ cus luteus
Escherichia coli
A. lipolytica (OUATTARA et al., 1992). Isolate LIPS was assigned tentatively to the genus Clostridium based on endospore formation, exhibition of anaerobic energy metabolism, inability to carry out dissimilatory sulfate reduction and exhibition of a Gram-positive cell wall structure. The acidogenic-solventogenic metabolic endproducts detected from glycerol fermentation, and growth on a number of saccharide substrates (Table 1), further supported the assignment of this isolate to the genus Clostridium.
Phylogenetic analyses
Sequencing of the PCR-amplified 16S rDNA of the deer rumen isolates LIP4 and LIPS enabled 92% of the complete 16S rRNA gene to be determined (estimated by comparison with the rRNA gene sequence of E. coli). Sequence analysis of the 16S rRNA gene of isolate LIP4 indicated that this bacterial isolate clustered with members of the genus Propionibacterium (Fig. 3). The sequence similarity values (Table 2) indicated that isolate LIP4 was most similar to Propionibacterium acnes (99.6%). However the species P. acnes whilst being able to hydrolyse gelatin and produce catalase, is unable to hydrolyse esculin or ferment trehalose (CUMMINGS and JOHNSON, 1986), which are phenotypic properties of LIP4. Therefore, it would seem that isolate LIP4 represents a nongelatinolytic, lipid-hydrolysing bacterial strain similar, but not identical to p, acnes.

140 G. N. JARVIS et al.
Sequence comparisons (Table 2) indicated that isolate LIPS was positioned in clostridial cluster XIVa of the Clostridium (sensu lato) taxonomic group (Fig. 3), and was 98.9% similar to the type strain (DSM S628T ) of Clostridium celerecrescens, a cellulolytic bacterial species isolated from cow manure (PALOP et al., 1989). The 16S rDNA sequence similarity value (98.9%) between isolate LIPS and C. celerecrescens alone, does not allow a definitive statement to be made regarding whether LIPS should be considered a strain of C. celerecrescens. In addition, isolate LIPS exhibited no cellulolytic activity and was unable to utilise arabinose, gelatin or xylose, unlike C. celerecrescens (PALOP et al., 1989). LIPS was also capable of glycerol dissimilation and demonstrated measurable lipolytic activity, which are not phenotypic characteristics associated with C. celerecrescens (PALOP et al., 1989). Reports exist wherein organisms which have been shown to belong to different species, on the basis of DNA-DNA hybridisation, possessed identical, or nearly identical, 16S rDNA sequences (Fox et al., 1992; BENNASAR et al., 1996). Therefore, it is clear that high 16S rDNA sequence similarities cannot guarantee that two organisms belong to the same species. Although it is accepted that 16S rRNA sequence similarities below 97% indicate that the organisms definitely do not belong to the same species (STACKEBRANDT and GOEBEL, 1994), in general, observations show that organisms with sequence similarities below 99% often belong to different species. Therefore, the 16S rDNA sequence similarity between isolate LIPS and C. celerecrescens, as well as the phenotypic differences, noted, suggested that the deer rumen isolate LIPS many represent a phylogenetic ally closely related, yet distinct clostridial species.
The taxonomy of the genus Clostridium is currently undergoing significant revisions (COLLINS et al., 1994),
and the majority of species currently classified within this genus will ultimately be separated and reclassified as distinct genera. The isolate LIPS described in the current paper was observed to group with species of cluster XIVa (Fig. 3). This phylogenetically heterogeneous taxon comprises validly described species of Clostridium, as well as several other recognised genera (COLLINS et al., 1994). It is clear that all clostridial species within cluster XIVa will be reclassified in the future as species in newly recognised genera. Thus, it is prudent at this time, to recognise the deer rumen isolate of the current study simply as "Clostridium species" (strain LIPS), rather than attempting to describe it as a new species.
Growth on neural lipids
Growth, metabolism, and lipolytic activity in complex and defined medium indicated that lipid-hydrolysis occurred irrespective of whether neutral lipid were presented as the sole carbon and energy source, or as co-substrate (Table 3). In the defined media, strain LIP4 exhibited lipid hydrolysis in the absence of co-factor supplement (yeast extract). However, lipid hydrolysis was accelerated in the complex BHIItripalmitin (MBHI) medium (Table 3). The similar levels ofVFA and biomass production in the glycerol-, and tallow-containing media (± yeast extract) implied that the glycerol liberated by tallow hydrolysis, was used as a source of energy by strains LIP4 and LIPS when cultured in minimal medium. Interestingly, tallow hydrolysis and biomass production were also stimulated by the presence of co-factors supplied by yeast extract for isolate LIPS (Table 3), which exhibited better growth rates and yields in the complex medium (MBHI) compared to the tallow/yeast extract medium (Table 3). Isolate LIPS produced less alcohols in both the
Table 3. Summary of growth and metabolism of lipolytic strains LIP4 and LIPS in defined (glycerol and tallow-containing) and complex (BHI-based) liquid media
Isolate Growth substrate Measured parametersa Ratio of Lipid hydrolysis Alcohols: VFA (%)b
VFA Cell biomass Alcohols (mM) (fIg ml-1) (mM)
LIP4 Glycerol 24.0 12 0 NA NA Tallow 21.5 21 0 NA 53 TallowlYeast Extract 20.3 17 0 NA 24 BHIffripalmitin 49.3 (0.7) 70 (7) 0 NA 82
LIPS Glycerol 74 138 56 0.76 NA Tallow 19.2 14 13.8 0.72 30 TallowlYeast Extract 24.1 43 11.9 0.49 50 BHIITripalmitin 121.6 (13.5) 179 (7) 27.1 (0.8) 0.22 24
a Data for BHIffripalmitin-containing medium represents mean (n = 3) with standard deviation in parentheses. All other data represents the average (n = 2) with an average deviation from the mean less than 10%. All cultures were incubated at 37 °C for 902 h except glycerol cultures which were incubated for 1008 h. b Total hydrolysis of tallow and tripalmitin would liberate 38.1 mM and 9.3 mM LCFA respectively. VFA: Volative fatty acids. NA: Not applicable.
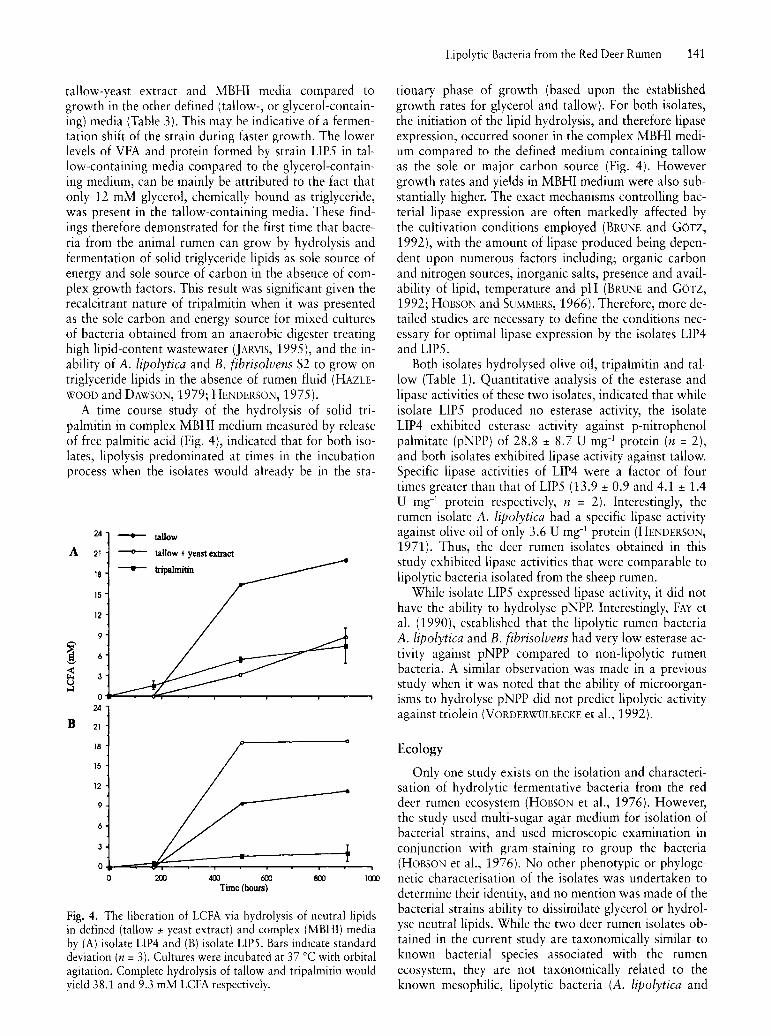
tallow-yeast extract and MBHI media compared to growth in the other defined (tallow-, or glycerol-containing) media (Table 3). This may be indicative of a fermentation shift of the strain during faster growth. The lower levels of VFA and protein formed by strain LIP5 in tallow-containing media compared to the glycerol-containing medium, can be mainly be attributed to the fact that only 12 mM glycerol, chemically bound as triglyceride, was present in the tallow-containing media. These findings therefore demonstrated for the first time that bacteria from the animal rumen can grow by hydrolysis and fermentation of solid triglyceride lipids as sole source of energy and sole source of carbon in the absence of complex growth factors. This result was significant given the recalcitrant nature of tripalmitin when it was presented as the sole carbon and energy source for mixed cultures of bacteria obtained from an anaerobic digester treating high lipid-content wastewater (JARVIS, 1995), and the inability of A. lipolytica and B. fibrisolvens S2 to grow on triglyceride lipids in the absence of rumen fluid (HAZLEWOOD and DAWSON, 1979; HENDERSON, 1975).
A time course study of the hydrolysis of solid tripalmitin in complex MBHI medium measured by release of free palmitic acid (Fig. 4), indicated that for both isolates, lipolysis predominated at times in the incubation process when the isolates would already be in the sta-
24
A 21
18
15
12
9
6
0 24
B 21
18
15
12
9
6
0 0
- tallow
- tallow + yeast extract
---- tripalmitin
200 400 600 800 1000 Time (hours)
Fig. 4. The liberation of LCFA via hydrolysis of neutral lipids in defined (tallow ± yeast extract) and complex (MBHI) media by (A) isolate LIP4 and (B) isolate LIPS. Bars indicate standard deviation (n = 3). Cultures were incubated at 37°C with orbital agitation. Complete hydrolysis of tallow and tripalmitin would yield 38.1 and 9.3 mM LCFA respectively.
Lipolytic Bacteria from the Red Deer Rumen 141
tionary phase of growth (based upon the established growth rates for glycerol and tallow). For both isolates, the initiation of the lipid hydrolysis, and therefore lipase expression, occurred sooner in the complex MBHI medium compared to the defined medium containing tallow as the sole or major carbon source (Fig. 4). However growth rates and yields in MBHI medium were also substantially higher. The exact mechanisms controlling bacterial lipase expression are often markedly affected by the cultivation conditions employed (BRUNE and GOTZ, 1992), with the amount of lipase produced being dependent upon numerous factors including; organic carbon and nitrogen sources, inorganic salts, presence and availability of lipid, temperature and pH (BRUNE and GOTZ, 1992; HOBSON and SUMMERS, 1966). Therefore, more detailed studies are necessary to define the conditions necessary for optimal lipase expression by the isolates LIP4 and LIP5.
Both isolates hydrolysed olive oil, tripalmitin and tallow (Table 1). Quantitative analysis of the esterase and lipase activities of these two isolates, indicated that while isolate LIP5 produced no esterase activity, the isolate LIP4 exhibited esterase activity against p-nitrophenol palmitate (pNPP) of 28.8 ± 8.7 U mg-1 protein (n = 2), and both isolates exhibited lipase activity against tallow. Specific lipase activities of LIP4 were a factor of four times greater than that of LIP5 (13.9 ± 0.9 and 4.1 ± 1.4 U mg-1 protein respectively, n = 2). Interestingly, the rumen isolate A. lipolytica had a specific lipase activity against olive oil of only 3.6 U mg-1 protein (HENDERSON, 1971). Thus, the deer rumen isolates obtained in this study exhibited lipase activities that were comparable to lipolytic bacteria isolated from the sheep rumen.
While isolate LIP5 expressed lipase activity, it did not have the ability to hydrolyse pNPP. Interestingly, FAY et al. (1990), established that the lipolytic rumen bacteria A. lipolytica and B. fibrisolvens had very low esterase activity against pNPP compared to non-lipolytic rumen bacteria. A similar observation was made in a previous study when it was noted that the ability of microorganisms to hydrolyse pNPP did not predict lipolytic activity against triolein (VORDERWOLBECKE et al., 1992).
Ecology
Only one study exists on the isolation and characterisation of hydrolytic fermentative bacteria from the red deer rumen ecosystem (HOBSON et al., 1976). However, the study used multi-sugar agar medium for isolation of bacterial strains, and used microscopic examination in conjunction with gram-staining to group the bacteria (HOBSON et al., 1976). No other phenotypic or phylogenetic characterisation of the isolates was undertaken to determine their identity, and no mention was made of the bacterial strains ability to dissimilate glycerol or hydrolyse neutral lipids. While the two deer rumen isolates obtained in the current study are taxonomically similar to known bacterial species associated with the rumen ecosystem, they are not taxonomically related to the known mesophilic, lipolytic bacteria (A. lipolytica and

142 G. N. JARVIS et al.
B. fibrisolvens strain 52) associated with the lipid-hydrolysing niche in the rumen ecosystem. Interestingly, unlike A. lipolytica and B. fibrisolvens strain 52, the rumen isolates LIP4 and LIPS were capable of growth with triglyceride lipids as sole carbon and energy source. This paper therefore represents the first report of the isolation and characterisation of obligately anaerobic lipolytic bacteria from the red deer rumen ecosystem.
The fact that both isolates obtained in the current study were highly lipolytic, was demonstrated qualitatively and quantitatively using three different neutral lipids as substrates. The rationale for the use of tallow, rather than olive oil, in defined medium for quantitative lipase assays, was consistent with the current practice of adding tallow as a feed supplement to concentrate-based diets in order to improve animal productivity, especially during lactation (KRONFELD et al., 1980). The use of tallow, in the lipase assay system was further justified, as ruminant animals fed a concentrate-based diet invariably have the lipid portion presented as neutral lipids rather than polar or complex lipids (HARFOOT and HAZLEWOOD, 1988).
Isolate LIPS was capable of growth on a wide range of saccharide substrates aside from the neutral lipids, and therefore may occupy both the lipid-hydrolysing and polysaccharide-degrading ecological niches within the red deer rumen ecosystems. The rapid growth in complex media and in defined minimal media with carbohydrates as energy source (0.02-0.053 h-1) would allow LIPS to colonise the deer rumen. Isolate LIP4, in contrast, was slow growing, lipolytic and capable of growth on lactate. Due to the slow growth, it would seem difficult for this isolate to occupy both the lipid-hydrolysing and lactateutilising ecological niches in the deer rumen ecosystem. P. acnes has been isolated previously from the rumen of cattied fed a hay-based diet (GUTIERREZ, 1953). However, the deer rumen isolate LIP4, while phylogenetically similar to the type strain of P. acnes differs from the bovine rumen strains (G UTIERREZ, 1953) due to its inability to
ferment sorbitol and has hydrolyse gelatin. Interestingly, the majority of bovine rumen strains were capable of fermenting mannitol and maltose (GUTIERREZ, 1953), while the deer rumen isolate LIP4 cannot. P. acnes was found in large numbers (1.3x1 09 cells ml-1) in the cattle rumen (GUTIERREZ, 1953). However, it was also isolated at high levels in soil (6x108 cells g-I), and in the hay-feed (0.6-2x109 cells g-l) (GUTIERREZ, 1953). Therefore it was felt that the bovine rumen strains of P. acnes were probably derived from the feed (HUNGATE, 1966). Therefore, it is conceivable that the hay-feed may have been the source of the isolate LIP4, allowing the isolate to be continuously introduced into the rumen of juvenile red deer used in the current study.
Glycerol was used as the carbon and energy source for the enrichment of lipolytic bacteria in the current study. The rationale being, that, the glycerol represents the only fermentable moiety of the triglyceride molecule. From a thermodynamic perspective, it is unlikely that any lipolytic microorganism could utilise the saturated LCFA portion of a triglyceride as a source of energy due to the
large positive Gibbs free energy value for oxidation of LCFA. For stearic and palmitic acids the value is +383.8 and +336.7 kJ/reaction respectively (THAUER et al., 1977). In contrast, the Gibbs free energy value for fermentation of glycerol to acetate (and bicarbonate) is -73.2 kJ/mol (THAUER et al., 1977), a thermodynamically much more favourable reaction.
It does however, seem highly probable that isolate LIP4 requires LCFA to stimulate growth, and future experiments will be directed at determining optimal growth conditions in lipid-containing media for this isolate. It is anticipated that more phylogenetic analyses (including DNA-DNA hybridization studies) and chemotaxonomic data will be gathered on the two isolates to determine exact species assignments in the near future. Another area of research may involve the purification and characterization of the lipase from these isolates, which may increase our knowledge in the area of lipolytic activity associated with obligately anaerobic rumen bacteria. these lipases may also have potential use in a wide range of biotechnological applications such as organic synthesising of optically pure compounds, detergent production, synthesis of flavour and fragrance compounds and use in cosmetics photographic processing and digestive aids (BRUNE and GOTZ, 1992).
Acknowledgements The authors thank R. EASINGWOOD and S. JOHNSTONE for
their invaluable assistance with the electron microscopy, and A. ARNSCHNEIDT for her technical support. We are indebted to Dr. COLIN MAcINTOSH of AgResearch (Invermay, New Zealand) for access to the red deer. This study was supported in part through the German-New Zealand Science and Technology Cooperation Project (STC 93.20 and NEU-015-96), sponsored by the German Research Centre for the Environment and Health and Professor K. N. TIMMIS (German National Research Centre for Biotechnology).
References
BENNASAR, A., ROSSELLO-MoRA, R., LALUCAT, J., MOORE, E. R. B.: 16S rRNA gene sequence analysis relative to genomovars of Pseudomonas stutzeri and proposal of Pseudomonas balearica sp. nov. Int. J. Syst. Bacteriol. 46, 200-206 (1996).
BRUNE, K. A., GOTZ, E: Degradation of lipids by bacteriallipases. pp. 243-266. In: Microbial degradation of natural products. (G. WINKELMANN, ed.) New York, VCH-Winheim 1992.
COLLINS, M. D., LAWSON, P. A., WILLEMS, A., CORDOBA, J. J., FERNANDEz-GARAYZABAL, J., GARCIA, P., CAl, J., HIPPE, H., FARROW, J. A. E.: The phylogeny of the genus Clostridium: proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 45,812-826 (1994).
CUMMINGS, C. S., JOHNSON, J. L.: Genus Propionibacterium, pp. 1346-1353. In: Bergey's manual of systematic bacteriology (P. H. A. SNEATH, N. S. MAIR, M. E. SHARPE, J. G. HOLT, eds.) Baltimore, Williams and Wilkins 1986.
DOETSCH, R. N.: Determinative methods of light microscopy. pp. 21-22. In: Manual of methods for general bacteriology (P. GERHARDT, R. G. E. MURRAY, R. N. COSTILOW, W. E. NESTER, W. A. WOOD, N. R. KRIEG, G. BRIGGS-PHILLIPS, eds.) Washington, American Society for Microbiology 1981.

FAY, J. P., JAKOBER, K. D., CHE"IG, K.-J., COSTERON, J. w.: Esterase activity of pure cultures of rumen bacteria as expressed by the hydrolysis of p-nitrophenylpalmitate. Can. J. Microbiol. 36,585-589 (1990):
FELSENSTEIN, J.: PHYLIP-phylogeny inference package (Version 3.2). Cladistics. 5, 164-166 (1989).
Fox, G. E., WISOTZKEY, j. E, jURTSHUK Jr., P.: How close is close: 16S rRNA sequence identity may nat be sufficient to guarantee species identity. Int. J. Syst. Bacteriol. 42, 166-170 (1992).
GUTIERREZ, J.: Numbers and characteristics of lactate utilizing organisms in the rumen of cattle. J. Bacteriol. 66: 123-128 (1953).
HARFOOT, C. G.: Lipid metabolism in the rumen. Progr. Lipid Res. 17,21-54 (1978).
HARFOOT, C. G., HAZLEWOOD, G. P.: Lipid metabolism in the rumen. pp. 285-321. In: The rumen microbial ecosystem (P. N. HOBSON, ed.) New York, Elsevier Science Publishing Co. Inc., 1988.
HARTREE, E. E: A modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 48, 422-427 (1972).
HAZLEWOOD, G. P., DAWSON, R. M. c.: Characteristics of a lipolytic and fatty acid requiring Butyrivibrio sp. isolated from the ovine rumen. J. Gen. Microbiol. 112, 15-27 (1979).
HENDERSON, C.: A study of the lipase produced by Anaerovibrio lipolytica, a rumen bacterium. J. Gen. Microbiol. 65, 81-89 (1971).
HENDERSON, C.: The isolation and characterisation of strains of lipolytic bacteria from the ovine rumen. J. Appl. Bacteriol. 39,101-109 (1975).
HOBSON, P. N., MANN, S. 0.: The isolation of glycerol fermenting and lipolytic bacteria from the rumen of the sheep. j. Gen. Microbiol. 25,227-240 (1961).
HOBSON, P. N., SUMMERS, R.: Effect of growth rate on the lipase activity of a rumen bacterium. Nature. 209, 736-737 (1966).
HOBSON, P. N., MANN, S. 0., SUMMERS, R.: Rumen micro-organisms in red deer, hill sheep and reindeer in the Scottish Highlands. Proc. R. Soc. Edinb. Sect. B. 75, 171-180 (1976).
HUNGATE, R. E.: The rumen and its microbes. pp. 533. New York, Academic Press 1966.
JARVIS, G. N.: The microbiology of anaerobic tallow hydrolysis. Ph.D. thesis. University of Otago, Dunedin, New Zealand, 1995.
JARVIS, G. N., THIELE, j. H.: High-performance liquid chromatographic analysis of free long chain fatty acids produced during lipolysis by anaerobic digestor sludge. J. Chromo A. 760,271-277 (1997a).
JARVIS, G. N., THIELE, J. H.: Qualitative rhodamine B assay which uses tallow as a substrate for lipolytic obligately anerobic bacteria. j. Microbiol. Methods. 760: 271-277 (1977b).
JARVIS, G. N., MOORE, E. R. B., THIELE, J. H.: Formate and ethanol are the major products of glycerol fermentation produced by a Klebsiella planticola strain isolated from red deer. J. Appl. Microbiol. 83: 166-174 (1997).
JUKES, T. H., CANTOR, C. R.: Evolution of protein molecules. p. 21-132. In: Mammalian protein metabolism. (H. N. MUNRO, ed.) New York, Academic Press Inc., 1969.
KARLSON, U., DWYER, D. E, HOOPER, S. W., MOORE, E. R. B., TIMMIS, K. N., ELTIS, L. D.: Two independently regulated cytochromes p-450 in a Rhodococcus rhodochrous strain that degrades 2-ethoxyphenol and 4-methoxybenzoate. J. Bacteri-01. 175, 1467-1474 (1993).
KENEALY, W., ZEIKUS, J. G.: Influence of corrinoid antagonists on methanogen metabolism. J. Bacterio!' 146, 133-140 (1981).
Lipolytic Bacteria from the Red Deer Rumen 143
KOUKER, G., JAEGER, K.-E.: Specific and sensitive plate assay for bacterial lipases. Appl. Environ. Microbiol. 53, 211-213 (1987).
KRONFELD, D. S., DONOGHUE, S., NAYLOR, J. M., JOHNSON, K., BRADLEY, C. A.: Metabolic effects of feeding protected tallow to dairy cows. J. Dairy Sci. 63, 545-552 (1980).
LOUGH, A. K., GARTON, G. A.: Digestion and metabolism of feed lipids in ruminants and non-ruminants. Symposiums of Zoological Society of London. 21,163-173 (1968).
MACKIE, R. ]., WHITE, B. A., BRYANT, M. P.: Lipid metabolism in anaerobic ecosystems. Crit. Rev. Microbiol. 17, 449-478 (1991 ).
MAIDAK, B., OLSEN, G. J., LARSEN, N., OVERBEEN, R., McCAUGHEY, M. J., WOESE, C. R.: The RDP (ribosomal database project). Nucl. Acids Res. 25, 109-110 (1997).
NEEFS, J.-M., VAN DE PEER, Y., HENDRICKS, L., DE WACHTER, R.: Compilation of small ribosomal subunit RNA sequences. Nucleid Acid Res. 18 (supplement), 2237-2317 (1990).
OUATTARA, A. S., TRAORE, A. S., GARCIA, J.-L.: Characterisation of Anaerovibrio burkinabensis sp. nov., a lactate-fermenting bacterium isolated from rice field soils. Int. J. Syst. Bacteriol. 42,390-397 (1992).
PALOP, M. L., VALLES, S., PINAGA, E, FLORS, A.: Isolation and characterization of an anaerobic, cellulolytic bacterium, Clostridium celerecrescens sp. nov. Int. J. Syst. Bacteriol. 39, 68-71 (1989).
PRINS, R. A., LANKHORST, A., VAN DER MEER, P., VAN NEVEL, C. J.: Some characteristics of Anaerovibrio lipolytica, a rumen lipolytic organism. Antonie Van Leeuwenhoek. 41, 1-11 (1975).
STACKEBRANDT, E., GOEBEL, B. M.: Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 44,846-849 (1994).
STEWART, C. S., BRYANT, M. P.: The rumen bacteria, pp. 21-75. In: The rumen microbial ecosystem. (P. N. HOBSON, ed.) New York, Elsevier Applied Science 1988.
STOESSER, G., STERK, P., TULl, M. A., STOEHR, P. J., CAMERON, G. N.: The EMBL nucleotide sequence database. Nucl. Acids Res. 25, 7-13 (1997).
THAUER, R. K., JUNGERMANN, K., DECKER, K.: Energy conservation in chemotrophic anaerobic bacteria. Bacterio!' Rev. 41, 100-180 (1977).
VORDERWOLBECKE, T., KIESLICH, K., ERDMANN, H.: Comparison of lipases by different assays. Enzyme Microb. Technol. 14, 631-639 (1992).
WILSON, K.: Preparation of genomic DNA from bacteria. p. 2.4.1.-2.4.2. In: Current protocols in molecular biology. (E M. AUSUBEL, R. BRENT, R. E. KINGSTON, D. D. MOORE, J. G. SEIDMAN, J. A. SMITH, K. STRUHL, eds.) New York, John Wiley & Sons Inc 1987.
WINKLER, U. K., STUCK MANN, M.: Glycogen, hyaluronate, and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. J. Bacteriol. 138, 663-670 (1979).
Corresponding author: Dr. GRAEME JARVIS, Rumen Microbiology Unit, AgResearch Grasslands Centre, Private Bag 11008, Palmerston North, New Zealand. Phone: +64-6-356-8019; Fax: +64-6-351-8003; e-mail: [email protected]
Related Documents











