Journal of the Science of Food and Agriculture J Sci Food Agric 87:1197–1206 (2007) Isoflavone profiles, phenol content, and antioxidant activity of soybean seeds as influenced by cultivar and growing location in Ohio Ken M Riedl, 1 Jae Hwan Lee, 1 Marjory Renita, 1 Steven K St Martin, 2 Steven J Schwartz 1 and Yael Vodovotz 1∗ 1 Department of Food Science & Technology, 110 Parker Food Science and Technology Bldg, 2015 Fyffe Rd., Columbus, OH 43210-1007, USA 2 Department of Horticulture and Crop Science, The Ohio State University, USA Abstract: Isoflavone levels and isoflavone chemical composition in soybeans vary between planting locations although the exact factors which control isoflavone biosynthesis are unclear. We compared levels of 12 isoflavones in soybean seeds of six cultivars grown in four different locations in Ohio in 2002 as determined by high- performance liquid chromatography. Antioxidant activity contained in plant-based foods can improve food oxidative stability and phenolics and isoflavones have proven active in food systems. Radical scavenging activity was assessed using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical. Total phenolics (TPCs) were determined by using Folin–Ciocalteu reagent. Total isoflavones (TIs) varied five-fold (1573–7710 nmol g −1 ) between seeds from the various location–cultivar combinations. One location (Wooster, Ohio) produced seeds containing half the isoflavones as the other locations tested apparently due to poor growing conditions. The cultivars could be divided into two groups based on TI, one having approximately 50% more isoflavones. Surprisingly, across the entire data set, with increasing TI, the proportion of isoflavones accounted for by the daidzein family increased due primarily to malonyl daidzin. DPPH scavenging did not differ significantly by location or cultivar (P > 0.05) and did not correlate with TPC or TI. Profiling soybean isoflavones could help elucidate how isoflavone biosynthesis is regulated and lead to better disease resistance of soybean crops and soy foods with greater health benefits. 2007 Society of Chemical Industry Keywords: isoflavones; antioxidant; metabolic profile; planting location INTRODUCTION Epidemiological studies have shown that consumption of soybeans and soy products may reduce the risk of cardiovascular disease, improve bone health, and inhibit the growth of human cancers. 1–3 Isoflavones are one of the bioactive low molecular weight components of soy. They are structurally similar to naturally occurring estrogens and show some promise in protecting against hormone-dependent cancers. 4–6 Therefore, increased consumption of soybeans with enhanced levels of isoflavones may be beneficial to human health. The isoflavones in soybeans occur primarily in their storage form as 6-O -malonyl-β -glucosides of the respective aglycones genistein, daidzein, and glycitein. In soybean seeds the concentration of isoflavone aglycones is low while significant levels of their simple β -glucoside (i.e. glucose not conjugated with acetyl or malonyl groups) are present. The 6-O - acetyl-β -glucosides are present in some soy foods that have been processed with high temperature and low water content, causing decarboxylation of the malonyl conjugate. Isoflavone content and distribution in soybeans is influenced by many factors, including cultivar, crop year, planting location, soil nutrition, and storage duration. 7–11 Tsukamoto et al . 9 reported that of the total isoflavones in seeds 80–90% were located in the soybean cotyledon, and planting dates and temperatures during seed development affected the total isoflavone content. Low temperatures and high precipitation during seed development have been shown to result in higher isoflavone contents. 9,12,13 Wang and Murphy 11 compared the isoflavone composition and content in eight American and three Japanese soybean cultivars at different locations and crop years with the latter affecting isoflavone content more than planting location. 11 Hoeck et al . 7 tested six soybean cultivars grown in eight locations in Iowa and showed that genotype, genotype year, genotype location, and genotype year location interactions all influenced the isoflavone concentration in soybeans. These authors ∗ Correspondence to: Yael Vodovotz, Department of Food Science & Technology, 110 Parker Food Science and Technology Bldg, 2015 Fyffe Rd., Columbus, OH 43210-1007, USA E-mail: [email protected] Contract/grant sponsor: Ohio Agricultural and Research Development Center. (Received 23 February 2006; revised version received 21 July 2006; accepted 21 July 2006) Published online 11 April 2007; DOI: 10.1002/jsfa.2795 2007 Society of Chemical Industry. J Sci Food Agric 0022–5142/2007/$30.00

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of the Science of Food and Agriculture J Sci Food Agric 87:1197–1206 (2007)
Isoflavone profiles, phenol content, andantioxidant activity of soybean seeds asinfluenced by cultivar and growing location inOhioKen M Riedl,1 Jae Hwan Lee,1 Marjory Renita,1 Steven K St Martin,2
Steven J Schwartz1 and Yael Vodovotz1∗1Department of Food Science & Technology, 110 Parker Food Science and Technology Bldg, 2015 Fyffe Rd., Columbus, OH 43210-1007,USA2Department of Horticulture and Crop Science, The Ohio State University, USA
Abstract: Isoflavone levels and isoflavone chemical composition in soybeans vary between planting locationsalthough the exact factors which control isoflavone biosynthesis are unclear. We compared levels of 12 isoflavonesin soybean seeds of six cultivars grown in four different locations in Ohio in 2002 as determined by high-performance liquid chromatography. Antioxidant activity contained in plant-based foods can improve foodoxidative stability and phenolics and isoflavones have proven active in food systems. Radical scavenging activitywas assessed using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical. Total phenolics (TPCs) were determinedby using Folin–Ciocalteu reagent. Total isoflavones (TIs) varied five-fold (1573–7710 nmol g−1) between seedsfrom the various location–cultivar combinations. One location (Wooster, Ohio) produced seeds containing halfthe isoflavones as the other locations tested apparently due to poor growing conditions. The cultivars could bedivided into two groups based on TI, one having approximately 50% more isoflavones. Surprisingly, across theentire data set, with increasing TI, the proportion of isoflavones accounted for by the daidzein family increased dueprimarily to malonyl daidzin. DPPH scavenging did not differ significantly by location or cultivar (P > 0.05) anddid not correlate with TPC or TI. Profiling soybean isoflavones could help elucidate how isoflavone biosynthesis isregulated and lead to better disease resistance of soybean crops and soy foods with greater health benefits. 2007 Society of Chemical Industry
Keywords: isoflavones; antioxidant; metabolic profile; planting location
INTRODUCTIONEpidemiological studies have shown that consumptionof soybeans and soy products may reduce the riskof cardiovascular disease, improve bone health, andinhibit the growth of human cancers.1–3 Isoflavonesare one of the bioactive low molecular weightcomponents of soy. They are structurally similar tonaturally occurring estrogens and show some promisein protecting against hormone-dependent cancers.4–6
Therefore, increased consumption of soybeans withenhanced levels of isoflavones may be beneficial tohuman health.
The isoflavones in soybeans occur primarily intheir storage form as 6-O′′-malonyl-β-glucosides of therespective aglycones genistein, daidzein, and glycitein.In soybean seeds the concentration of isoflavoneaglycones is low while significant levels of theirsimple β-glucoside (i.e. glucose not conjugated withacetyl or malonyl groups) are present. The 6-O′′-acetyl-β-glucosides are present in some soy foodsthat have been processed with high temperature
and low water content, causing decarboxylationof the malonyl conjugate. Isoflavone content anddistribution in soybeans is influenced by manyfactors, including cultivar, crop year, planting location,soil nutrition, and storage duration.7–11 Tsukamotoet al.9 reported that of the total isoflavones in seeds80–90% were located in the soybean cotyledon,and planting dates and temperatures during seeddevelopment affected the total isoflavone content.Low temperatures and high precipitation duringseed development have been shown to result inhigher isoflavone contents.9,12,13 Wang and Murphy11
compared the isoflavone composition and contentin eight American and three Japanese soybeancultivars at different locations and crop years withthe latter affecting isoflavone content more thanplanting location.11 Hoeck et al.7 tested six soybeancultivars grown in eight locations in Iowa and showedthat genotype, genotype year, genotype location, andgenotype year location interactions all influenced theisoflavone concentration in soybeans. These authors
∗ Correspondence to: Yael Vodovotz, Department of Food Science & Technology, 110 Parker Food Science and Technology Bldg, 2015 Fyffe Rd., Columbus,OH 43210-1007, USAE-mail: [email protected]/grant sponsor: Ohio Agricultural and Research Development Center.(Received 23 February 2006; revised version received 21 July 2006; accepted 21 July 2006)Published online 11 April 2007; DOI: 10.1002/jsfa.2795
2007 Society of Chemical Industry. J Sci Food Agric 0022–5142/2007/$30.00

KM Riedl et al.
also suggested that the isoflavone content can beincreased in soybeans as a quantitative cultivar traitthrough breeding. Lee and co-workers8,14 analyzedisoflavone content in 15 soybean cultivars storedfor 3 years at three different locations and reporteda significant effect of environmental factors on theisoflavone content in soybeans. Vyn et al.10 reportedminerals in soil such as potassium may promote theformation of isoflavones in soybeans.
Free-radical scavenging activity of food componentsmay be beneficial since free radicals are deleteriousto food quality and function in biological systems.Excessive formation of free radicals accelerates theoxidation of lipid in foods and decreases foodquality and consumer acceptance.15 High levels offree radicals in humans are associated with theaging process and age-related diseases.16Antioxidantactivities of isoflavones have been reported in vivoand in vitro17 in simple (liposomes) as well as morecomplex (lipoproteins) systems.18 Lee et al.19 reportedthe free-radical scavenging activities of 80% methanolextracts of soybeans varied depending on plantinglocation and storage year. However, the bases for theeffect of planting location on scavenging activity werenot fully understood.
The environmental effects and factors that influenceisoflavone contents in soybeans are unclear. Theobjective of this study was to determine the effects ofenvironment, as varied with four planting locations, onthe isoflavone content, free radical scavenging activity,and total phenolic content of six soybean cultivarsgrown in Ohio in 2002.
EXPERIMENTALMaterialsSoybean cultivars and experimental lines, includingTiffin, IA 2053, Ohio FG3, HS96-3850, HS98-3407,and HS96-3145 were obtained from the Depart-ment of Horticulture and Crop Science at the OhioState University. Their respective pedigrees are Haro-son × Chapman, unknown, HS89-8843 × OhioFG1, HS89-2966 × HS89-8843, HS89-3078 × OhioFG2Y, HS89-8843 × Ohio FG1. These cultivarswere selected because they cover a broad range ofpedigrees and include one grain-type cultivar (Tif-fin), the rest being food-grade cultivars. The soybeanswere grown at four different locations in 2002 in thestate of Ohio, including Wooster (WO), Manchesterfarm in Lakeview (WC), South Charleston (WE), andHoytville (NW) (Fig. 1). The latitude and longitudeof WO, WC, WE, and NW are 40◦783′N 81◦917′W,40◦507′N 83◦907′W, 39◦840′N 83◦658′W, and41◦190′N 83◦782′W, respectively, and the plantingdates of soybeans in WO, WC, WE, and NW were2 May, 26 April, 1 May, and 1 June, respectively(Fig. 1). Soil types were Wooster silt loam at WO,Milford silt loam at WC, Kokomo silt loam at WE andHoytville silty clay loam at NW. Herbicide treatmentlevel varied depending on the needs at a particular
NW, Hoytville
WC, Lakeview
WE, S. Charleston
WO, Wooster
Figure 1. Map of Ohio indicating the locations of the soybean cropsstudied. Latitude–longitude and planting dates for the respectivelocations were: WO, 40◦783′N 81◦917′W, 2 May 2002; NW,41◦190′N 83◦782′W, 1 June 2002; WC, 40◦507′N 83◦907′W, 26 April2002; and WE, 39◦840′N 83◦658′ W, 1 May 2002.
location. Since all soybean cultivars were Roundup-ready isoflavone metabolism should not be directlyaffected by herbicide treatment. This does not pre-clude the possibility that the prevalence of weeds inplots could indirectly affect isoflavone biosynthesis.The cultivars and experimental lines were planted aspart of replicated tests for yield and agronomic traits.A single sample of seed was taken as representativeof each variety or experimental line at each site. Thefour sites exhibited a range of yields, with yields atWE (3.4 Mg ha−1) approximately 50% higher thanthose at WC (2.2 Mg ha−1). Yields averaged 2.3 Mgha−1 at NW and 2.6 Mg ha−1 at WO. The checkcultivar (‘Kottman’, maturity group III), maturedapproximately 12 September for WO, 20 Septem-ber at WC and WE and 1 October at NW. Plots wereharvested about 2 weeks after maturity. The corre-sponding growth periods from planting to maturity forWO, WC, WE and NW were therefore 133, 147, 141,and 121 days, respectively.
All 12 standard isoflavones, including daidzein,glycitein, genistein, daidzin, glycitin, genistin, mal-onyl daidzin, malonyl glycitin, malonyl genistin,acetyl daidzin, acetyl glycitin, and acetyl genistinwere purchased from LC Laboratories (Woburn,MA). 2,2-Diphenyl-1-picrylhydrazyl (DPPH), buty-lated hydroxytoluene (BHT), formononetin andFolin–Ciocalteu reagent were purchased from Aldrich(St Louis, MO). HPLC-grade methanol, acetonitrile,HCl, water, and acetic acid were purchased fromFisher Scientific (Fairlawn, NJ).
Soy flour preparationSoybeans were stored for approximately 8 monthsat 23 ◦C without humidity control and moisture
1198 J Sci Food Agric 87:1197–1206 (2007)DOI: 10.1002/jsfa

Isoflavone profiles of soybeans
equilibrated to about 7% (w/w) before analysis. Houand Chang have shown that little change in isoflavonestakes place in seeds stored at this temperatureand low humidity.20 Soybeans were dehulled witha scarifier and those seed coats that resisted thescarifier were manually dehulled. Seeds with brownor black seed coats (common at WO) were notincluded (not suitable for food use) but greenish seeds(<5%) were included in the analysis. Dehulled seeds(approximately 3 g, 25 seeds) were ground using acoffee grinder (Black & Decker, Trumbull, CO) tomake the soy flour with a particle size less than 0.6 mm(through 30 mesh). Soy meals were stored in the darkat −20 ◦C until analyzed.
Isoflavone extractionTwo samples of soybean flour (each 0.5 g) fromeach cultivar and location as prepared above wasmixed with 100 mmol L−1 HCl (2 mL), acetonitrile(7 mL), and deionized water (3 mL) in a 50 mLcentrifuge bottle (Nalge company, Rochester, NY).Next, formononetin was added as internal standard toconfirm full recovery through drying, redissolution andfiltration. It is a 4-O-methylated version of daidzeinand thus has similar chemical and physical propertiesas the other isoflavone aglycones. Sample bottleswere vortexed for 1 min and shaken with a multi-wrist shaker (Lab-line Instruments, Inc., MelrosePark, IL) on setting 9 out of 10 scale for 2 h atroom temperature before placing in a Sorvall SS-34 rotor (Sorvall, Kendro Laboratory, CT) andcentrifuging at 2425 × g (4500 rpm) for 30 min. A1 mL aliquot of supernatant was transferred to a10 mL glass bottle and dried under nitrogen at roomtemperature. Dried samples were stored at −20 ◦C inthe dark until HPLC analysis.21 The extracts werere-dissolved in 1 mL methanol and the mixture wasvortexed and passed through a 0.2 µm syringe filter(Nylon, 13 mm diameter, Alltech Associates Inc.,Deerfield, IL) prior to HPLC injection. The reliabilityof the drying and re-dissolution steps was assessedby quantifying the recovery of a known concentrationof formononetin (internal standard) and glycitein (in80% methanol) added to the first extraction mixture.Glycitein was chosen among all isoflavones due to itslow concentration in the samples. Recovery percentagefor formononetin and glycitein were 98.3 ± 5.3 and97.4 ± 5.9%, respectively (n = 6).
HPLC analysis of isoflavonesA Waters model 2690 high performance liquidchromatograph (HPLC) equipped with a Waters 2996photodiode array detector (PDA) (Waters Associated,Milford, MA) was used to separate, identify, andquantify isoflavones.22 Separation of isoflavones wasachieved using a 4 µm Waters Novapak C18 reversed-phase HPLC column (150 mm × 3.9 mm i.d.) witha Novapak C18 stationary phase guard column anda 0.5 µm pre-column filter from Vydac (Hesperia,CA). The sample injection volume was 10 µL and
components were eluted using a mobile phase gradientconsisting of 1% (v/v) acetic acid in water (solvent A)and 100% acetonitrile (solvent B) at a flow rate of0.6 mL min−1. The solvent gradient was: solvent Aat 85% from 0 to 5 min; linear decrease in solvent Afrom 85 to 65% from 5 to 44 min; linear increase insolvent A from 65 to 85% from 44 to 45 min; and thenthe column was re-equilibrated with solvent A at 85%for 5 min. Between each injection, a mixture of 85%solvent A and 15% solvent B was run for 20 minto ensure column equilibration.23 The UV–visiblespectra of the eluate were monitored from 240 to400 nm by the PDA.
Isoflavone identification and quantificationAll 12 standard isoflavones, including daidzein,glycitein, genistein, daidzin, glycitin, genistin, malonyldaidzin, malonyl glycitin, malonyl genistin, acetyldaidzin, acetyl glycitin, and acetyl genistin were used toidentify unknowns by comparing the retention times inHPLC chromatograms and UV–visible spectra.11,24,25
The stock solutions of standards were preparedby weighing 3 mg (weighed on a Mettler-ToledoAB204-S balance, Greifensee, Switzerland, repeata-bility 0.1 mg) or 1.0 mg (if exact amount sup-plied in ampoules) of crystalline standard com-pounds: daidzein, glycitein, genistein, daidzin, glyc-itin, genistin, malonyl daidzin, malonyl genistin,malonyl glycitin, acetyl daidzin, acetyl glycitin, andacetyl genistin, and dissolving them in the appropri-ate amount of 80% methanol in water to give 1.0 mg100 mL−1. The solutions were placed in the refriger-ator overnight to ensure complete dissolution. Eachstock solution of isoflavones was serially diluted with80% methanol in water to make the working solu-tions for the standard curves. The concentration ofworking solutions was determined according to theBeer–Lambert law using the peak UV absorbancereading (Hewlett Packard Spectrophotometer 8453)in the range 240–360 nm and their molar extinctioncoefficients in 80% methanol.24,26 Aliquots of work-ing solutions of each isoflavone were injected into theHPLC and the peak area of each isoflavone in the chro-matogram was determined. The relationship betweenHPLC peak area and concentration of isoflavonesfrom UV–visible spectroscopy was used to quantifyeach isoflavone peak. Since acetyl glucosides and mal-onyl glucosides are known to degrade (hydrolyze) inaqueous solution, chromatograms of the fresh stan-dard solutions were first scrutinized to ensure absenceof degradation products and once cocktails of sev-eral standards were prepared, their chromatogramswere validated daily against results for the fresh solu-tions. The correlation coefficient (R2) of all calibrationcurves for isoflavone standards was greater than 0.95.
Free-radical scavenging activityThe free radical scavenging activities of soybeancultivars were determined using the DPPH scavengingmethod.27 Dried sample extracts as prepared above
J Sci Food Agric 87:1197–1206 (2007) 1199DOI: 10.1002/jsfa

KM Riedl et al.
were re-dissolved in 1 mL 100% methanol andfiltered through a 0.2 µm disk syringe filter (Nylon,13 mm diameter, Alltech Associates Inc, Deerfield,IL). DPPH was dissolved in 100% methanol to aconcentration of 0.5 mmol L−1. DPPH (3.75 mL of0.5 mmol L−1) and mixed with 0.25 mL of sampleextract in methanol. The initial absorbance of theDPPH mixtures were measured at 517 nm and againafter 30 min to determine the amount of DPPHscavenged. The free-radical scavenging activity ofthese samples was expressed relative to that of BHT,a well-known radical scavenging compound. Theadvantage of using this BHT equivalent expressionis to minimize variations in analysis conditionssuch as the volume ratio of DPPH and sample orconcentration of DPPH. It also facilitates comparisonwith other reports in the literature. A standard curveof the free-radical scavenging activity of BHT onDPPH was obtained by measuring the absorbanceat 517 nm 30 min after mixing 3.75 mL of 0.5 mmolL−1 DPPH with 0.25 mL of 0.05, 0.075, 0.1, 0.25,and 0.5 mmol L−1 BHT in 100% methanol. Thefree-radical scavenging activity was expressed as mMBHT equivalents. Two replicate samples for eachcombination of cultivar–location were analyzed.
Determination of total phenolic compoundsTotal phenolic compounds (TPCs) of soybeanextracts were determined according to the method ofZielinski and Kozlowska28 with minor modification.A 0.25 mL methanolic soybean extract was mixedwith 4 mL water and 0.25 mL Folin–Ciocalteu reagent(previously diluted with water 1:1 v/v) in a 10 mL vial.
The mixture was vortexed for 30 s and left to standfor 5 min at room temperature. Next, 0.5 mL saturatedsodium carbonate was added and the mixture wasallowed to stand for 30 min at room temperature inthe dark and then centrifuged at 3000 ×g (5000 rpm)for 10 min (Sorvall, Kendro Laboratory, CT). Theabsorbance of supernatant was measured at 725 nmusing a UV–visible spectrophotometer (Shimadzu,Wood Dale, IL) and compared to a standard curve ofcatechin (0, 3, 15, 30, 75, and 150 µg (±) catechinmL−1) to calculate total phenolics on a catechinequivalents basis. Two replicate samples for eachcombination of cultivar–location were analyzed.
Statistical analysisisoflavone, DPPH scavenging and TPC data fromduplicate determinations of each cultivar–locationcombination were averaged over six cultivars or fourlocations, respectively, in order to have sufficientstatistical power to make reliable comparisons. Thesepooled values were analyzed by MANOVA (multipleanalysis of variance) using SPSS software, v. 12.0(SPSS Inc., Chicago, IL) and a P-value ≤0.05was considered significant. The data sets werenormally distributed about the respective means. Inthe statistical analysis, cultivars and locations wereregarded as fixed factors. A Bonferroni post hoc test
was used to evaluate the significance of differencesbetween means grouped by location or cultivar. Forthe correlation analyses, the square of the Pearson’scorrelation coefficient (R2) is reported along with its P-value. Correlations with P-values less than 0.10 weredeemed significant.
RESULTS AND DISCUSSIONChromatographyA representative HPLC chromatogram of soybeanisoflavones is shown in Fig. 2 with genistein anddaidzein as well as their β-glucosides, the acetyl-β-glucoside of genistin, malonyl-β-glucosides, andformononetin (internal standard) successfully sepa-rated and identified. Although peaks corresponding tothe standards for glycitein, acetyl glycitein and acetyldaidzein were observed, the absorbances were too lowto confirm by UV spectrum. The coefficient of varia-tion for analyses of the remaining nine isoflavones wastypically less than 5% (n = 4).
Of the three types of acetyl glucoside only acetylgenistin could be reliably identified and it was presentat low levels. This finding is consistent with previousreports that found significant levels of acetylatedglucosides only after heat-induced decarboxylation ofmalonyl glucosides under low moisture conditions.23
Isoflavone content and profilesIsoflavone contents of six soybean cultivars from fourdifferent planting locations are shown in Table 1. Inthis study the range of total isoflavones (TIs) was1573 to 7110 nmol g−1 (451 to 2107 µg g−1 soy asaglycone equivalents). This range is similar to thatfound in previous reports9,7,14,29 where TI in soybeansof one cultivar were between 460 and 1950 µg g−1 soyamong four locations,29 and from 794 to 1763 µg g−1
soy among eight Iowa locations.7 Growing year,location, and cultivar have each been shown to play
Mal
onyl
dai
dzin
Mal
onyl
gen
isti
n
Ab
sorb
ance
, 260
nm
Elution time, minutes
Gen
isti
n
Dai
dzei
n
Dai
dzin
Ace
tyl g
enis
tin
Gen
iste
in
For
mon
onet
in
0.00 10.00 20.00 30.00 40.00 50.00
0.00
0.02
0.04
0.06
0.08
0.10
0.12
Gly
citi
n
Mal
onyl
gly
citi
n
Figure 2. Representative HPLC chromatogram of soybean isoflavoneanalysis. Isoflavones were separated using a C18 column with anacetic acid/acetonitrile gradient and photodiode array (200–400 nm)for detection. Formononetin was used as an internal standard.
1200 J Sci Food Agric 87:1197–1206 (2007)DOI: 10.1002/jsfa
Isoflavone profiles of soybeans
Table 1. Isoflavone contents (nmol g−1) of extracts from seeds of six soybean cultivars grown at four different locations in Ohio in 2002†
Isoflavone, nmol g−1
Cultivar‡ Location DE DI MDI TDE GE GI AGI MGI TGE GLI MGLI TGL TI
Tiffin WO 120 285 550 955 54 454 19 1593 2120 50 44 94 31695839a NW 127 478 1535 2140 36 785 24 3412 4256 40 50 90 6486
WE 214 749 1250 2213 117 1350 24 3074 4565 76 49 125 6903WC 231 921 1353 2505 123 1211 26 2825 4184 71 39 110 6799
IA2053 WO 96 96 199 391 29 240 19 768 1055 69 58 127 15733301b NW 118 346 743 1207 45 728 26 2148 2948 88 86 174 4329
WE 124 279 680 1083 72 729 26 2037 2865 86 71 157 4105WC 150 335 352 837 104 880 25 1220 2230 78 50 128 3195
Ohio FG3 WO 94 205 480 779 25 454 17 1690 2185 32 59 91 30555355a NW 127 515 1208 1850 69 1239 27 3124 4459 48 69 117 6426
WE 148 690 1153 1991 104 1209 31 3312 4656 50 73 123 6770WC 135 370 945 1450 78 788 26 2727 3619 37 61 98 5167
HS96-3850 WO 95 260 399 754 29 490 18 1453 1990 38 78 116 28604105b NW 119 341 723 1183 70 822 26 2583 3501 50 115 165 4849
WE 117 322 632 1071 83 804 23 2308 3218 47 90 137 4426WC 132 342 510 984 103 856 25 2188 3171 43 87 130 4285
HS98-3407 WO 106 264 511 881 51 569 18 1575 2213 94 136 230 33245490a NW 107 596 1724 2427 33 1063 29 3981 5106 64 113 177 7710
WE 120 467 814 1401 59 963 30 2275 3327 54 77 131 4859WC 188 644 1108 1940 107 1218 27 2638 3991 59 76 135 6066
HS96-3145 WO 105 240 372 717 49 788 18 1345 2200 49 57 106 30234113b NW 112 448 877 1437 38 881 24 2399 3342 54 74 128 4907
WE 134 525 750 1409 76 1094 24 2054 3248 50 66 116 4773WC 123 475 673 1271 49 950 25 1345 2369 48 59 107 3747
† Isoflavone values (wet basis) are averages of extractions from two soybean meals where two injections from each extract were analyzed by HPLC.Acetyl daidzin, glycitein and acetyl glycitin were either present at trace levels or not detected. WO: Wooster, NW: Hoytville, WE: South Charleston,WC: Manchester farm in Lakeview. DE: daidzein, DI: daidzin, MDI: malonyl daidzin, TDE: total daidzeins, GE: genistein, GI: genistin, AGI: acetylgenistin, MGI: malonyl genistin, TGE: total genisteins, GLI: glycitin, MGLI: malonyl glycitin, TGL: total glyciteins, TI: total isoflavones.‡ Values below cultivar names are TI for each cultivar averaged over four locations where same superscript indicates not significantly different at95% CI using Bonferroni as post hoc test.
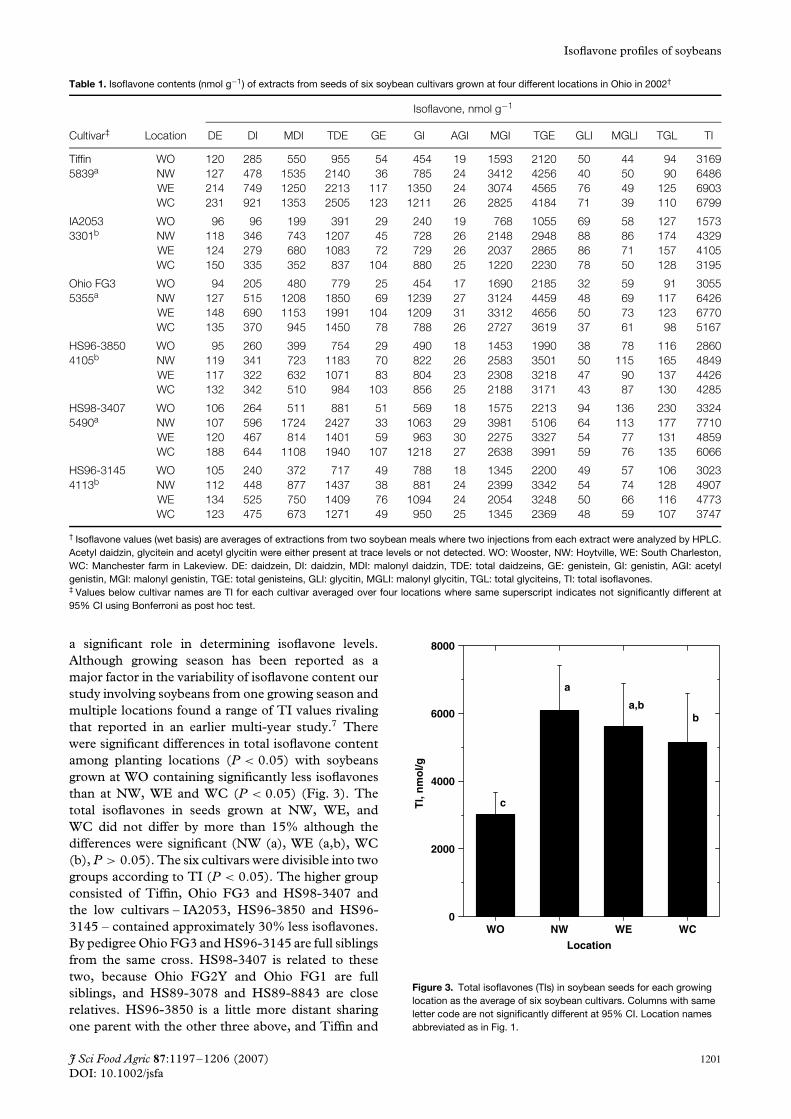
a significant role in determining isoflavone levels.Although growing season has been reported as amajor factor in the variability of isoflavone content ourstudy involving soybeans from one growing season andmultiple locations found a range of TI values rivalingthat reported in an earlier multi-year study.7 Therewere significant differences in total isoflavone contentamong planting locations (P < 0.05) with soybeansgrown at WO containing significantly less isoflavonesthan at NW, WE and WC (P < 0.05) (Fig. 3). Thetotal isoflavones in seeds grown at NW, WE, andWC did not differ by more than 15% although thedifferences were significant (NW (a), WE (a,b), WC(b), P > 0.05). The six cultivars were divisible into twogroups according to TI (P < 0.05). The higher groupconsisted of Tiffin, Ohio FG3 and HS98-3407 andthe low cultivars – IA2053, HS96-3850 and HS96-3145 – contained approximately 30% less isoflavones.By pedigree Ohio FG3 and HS96-3145 are full siblingsfrom the same cross. HS98-3407 is related to thesetwo, because Ohio FG2Y and Ohio FG1 are fullsiblings, and HS89-3078 and HS89-8843 are closerelatives. HS96-3850 is a little more distant sharingone parent with the other three above, and Tiffin and
WO NW WE WC0
2000
4000
6000
8000
TI,
nm
ol/g
Location
a
a,bb
c
Figure 3. Total isoflavones (TIs) in soybean seeds for each growinglocation as the average of six soybean cultivars. Columns with sameletter code are not significantly different at 95% CI. Location namesabbreviated as in Fig. 1.
J Sci Food Agric 87:1197–1206 (2007) 1201DOI: 10.1002/jsfa
KM Riedl et al.
IA2053 are not closely related to the others. Thus thegenetic background of the soybeans did not have aclear effect on the seed isoflavone content.
The amounts of particular genisteins and daidzeinsfollowed that of total isoflavones such that thoseseeds with higher TIs also had more of each typeof isoflavone. The four most abundant isoflavoneswere malonyl genistin, genistin, malonyl daidzin, anddaidzin, accounting for approximately 45%, 18%,15%, and 8% of TIs, respectively. It is typical formalonyl glucosides to dominate the isoflavone profilesin soybean seeds since they are the main chemicalform for storage.30
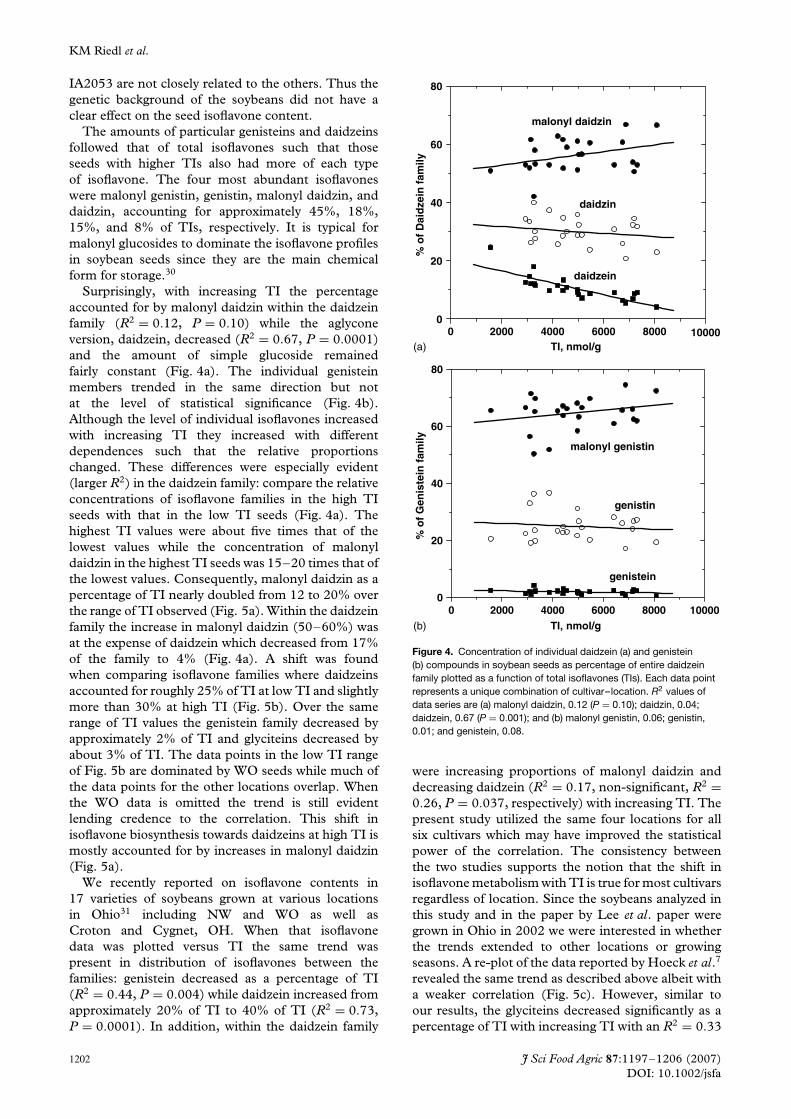
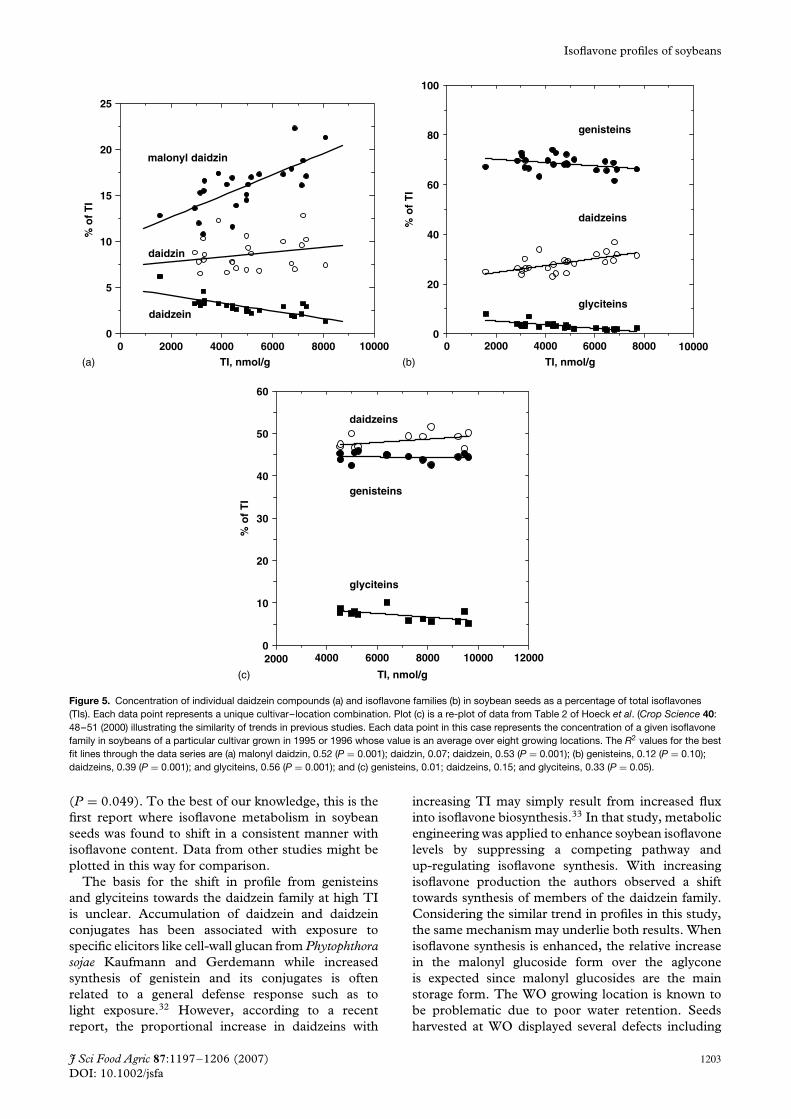
Surprisingly, with increasing TI the percentageaccounted for by malonyl daidzin within the daidzeinfamily (R2 = 0.12, P = 0.10) while the aglyconeversion, daidzein, decreased (R2 = 0.67, P = 0.0001)and the amount of simple glucoside remainedfairly constant (Fig. 4a). The individual genisteinmembers trended in the same direction but notat the level of statistical significance (Fig. 4b).Although the level of individual isoflavones increasedwith increasing TI they increased with differentdependences such that the relative proportionschanged. These differences were especially evident(larger R2) in the daidzein family: compare the relativeconcentrations of isoflavone families in the high TIseeds with that in the low TI seeds (Fig. 4a). Thehighest TI values were about five times that of thelowest values while the concentration of malonyldaidzin in the highest TI seeds was 15–20 times that ofthe lowest values. Consequently, malonyl daidzin as apercentage of TI nearly doubled from 12 to 20% overthe range of TI observed (Fig. 5a). Within the daidzeinfamily the increase in malonyl daidzin (50–60%) wasat the expense of daidzein which decreased from 17%of the family to 4% (Fig. 4a). A shift was foundwhen comparing isoflavone families where daidzeinsaccounted for roughly 25% of TI at low TI and slightlymore than 30% at high TI (Fig. 5b). Over the samerange of TI values the genistein family decreased byapproximately 2% of TI and glyciteins decreased byabout 3% of TI. The data points in the low TI rangeof Fig. 5b are dominated by WO seeds while much ofthe data points for the other locations overlap. Whenthe WO data is omitted the trend is still evidentlending credence to the correlation. This shift inisoflavone biosynthesis towards daidzeins at high TI ismostly accounted for by increases in malonyl daidzin(Fig. 5a).
We recently reported on isoflavone contents in17 varieties of soybeans grown at various locationsin Ohio31 including NW and WO as well asCroton and Cygnet, OH. When that isoflavonedata was plotted versus TI the same trend waspresent in distribution of isoflavones between thefamilies: genistein decreased as a percentage of TI(R2 = 0.44, P = 0.004) while daidzein increased fromapproximately 20% of TI to 40% of TI (R2 = 0.73,P = 0.0001). In addition, within the daidzein family
0 2000 4000 6000 8000 100000
20
40
60
80
% o
f D
aid
zein
fam
ily
TI, nmol/g
malonyl daidzin
daidzin
daidzein
0 2000 4000 6000 8000 100000
20
40
60
80
% o
f G
enis
tein
fam
ily
TI, nmol/g
malonyl genistin
genistin
genistein
(a)
(b)
Figure 4. Concentration of individual daidzein (a) and genistein(b) compounds in soybean seeds as percentage of entire daidzeinfamily plotted as a function of total isoflavones (TIs). Each data pointrepresents a unique combination of cultivar–location. R2 values ofdata series are (a) malonyl daidzin, 0.12 (P = 0.10); daidzin, 0.04;daidzein, 0.67 (P = 0.001); and (b) malonyl genistin, 0.06; genistin,0.01; and genistein, 0.08.
were increasing proportions of malonyl daidzin anddecreasing daidzein (R2 = 0.17, non-significant, R2 =0.26, P = 0.037, respectively) with increasing TI. Thepresent study utilized the same four locations for allsix cultivars which may have improved the statisticalpower of the correlation. The consistency betweenthe two studies supports the notion that the shift inisoflavone metabolism with TI is true for most cultivarsregardless of location. Since the soybeans analyzed inthis study and in the paper by Lee et al. paper weregrown in Ohio in 2002 we were interested in whetherthe trends extended to other locations or growingseasons. A re-plot of the data reported by Hoeck et al.7
revealed the same trend as described above albeit witha weaker correlation (Fig. 5c). However, similar toour results, the glyciteins decreased significantly as apercentage of TI with increasing TI with an R2 = 0.33
1202 J Sci Food Agric 87:1197–1206 (2007)DOI: 10.1002/jsfa
Isoflavone profiles of soybeans
0 2000 4000 6000 8000 100000
5
10
15
20
25
% o
f T
I
TI, nmol/g
malonyl daidzin
daidzin
daidzein
TI, nmol/g0 2000 4000 6000 8000 10000
% o
f T
I
0
20
40
60
80
100
genisteins
daidzeins
glyciteins
TI, nmol/g
2000 4000 6000 8000 10000 12000
% o
f T
I
0
10
20
30
40
50
60
genisteins
daidzeins
glyciteins
(a) (b)
(c)
Figure 5. Concentration of individual daidzein compounds (a) and isoflavone families (b) in soybean seeds as a percentage of total isoflavones(TIs). Each data point represents a unique cultivar–location combination. Plot (c) is a re-plot of data from Table 2 of Hoeck et al. (Crop Science 40:48–51 (2000) illustrating the similarity of trends in previous studies. Each data point in this case represents the concentration of a given isoflavonefamily in soybeans of a particular cultivar grown in 1995 or 1996 whose value is an average over eight growing locations. The R2 values for the bestfit lines through the data series are (a) malonyl daidzin, 0.52 (P = 0.001); daidzin, 0.07; daidzein, 0.53 (P = 0.001); (b) genisteins, 0.12 (P = 0.10);daidzeins, 0.39 (P = 0.001); and glyciteins, 0.56 (P = 0.001); and (c) genisteins, 0.01; daidzeins, 0.15; and glyciteins, 0.33 (P = 0.05).
(P = 0.049). To the best of our knowledge, this is thefirst report where isoflavone metabolism in soybeanseeds was found to shift in a consistent manner withisoflavone content. Data from other studies might beplotted in this way for comparison.
The basis for the shift in profile from genisteinsand glyciteins towards the daidzein family at high TIis unclear. Accumulation of daidzein and daidzeinconjugates has been associated with exposure tospecific elicitors like cell-wall glucan from Phytophthorasojae Kaufmann and Gerdemann while increasedsynthesis of genistein and its conjugates is oftenrelated to a general defense response such as tolight exposure.32 However, according to a recentreport, the proportional increase in daidzeins with
increasing TI may simply result from increased fluxinto isoflavone biosynthesis.33 In that study, metabolicengineering was applied to enhance soybean isoflavonelevels by suppressing a competing pathway andup-regulating isoflavone synthesis. With increasingisoflavone production the authors observed a shifttowards synthesis of members of the daidzein family.Considering the similar trend in profiles in this study,the same mechanism may underlie both results. Whenisoflavone synthesis is enhanced, the relative increasein the malonyl glucoside form over the aglyconeis expected since malonyl glucosides are the mainstorage form. The WO growing location is known tobe problematic due to poor water retention. Seedsharvested at WO displayed several defects including
J Sci Food Agric 87:1197–1206 (2007) 1203DOI: 10.1002/jsfa
KM Riedl et al.
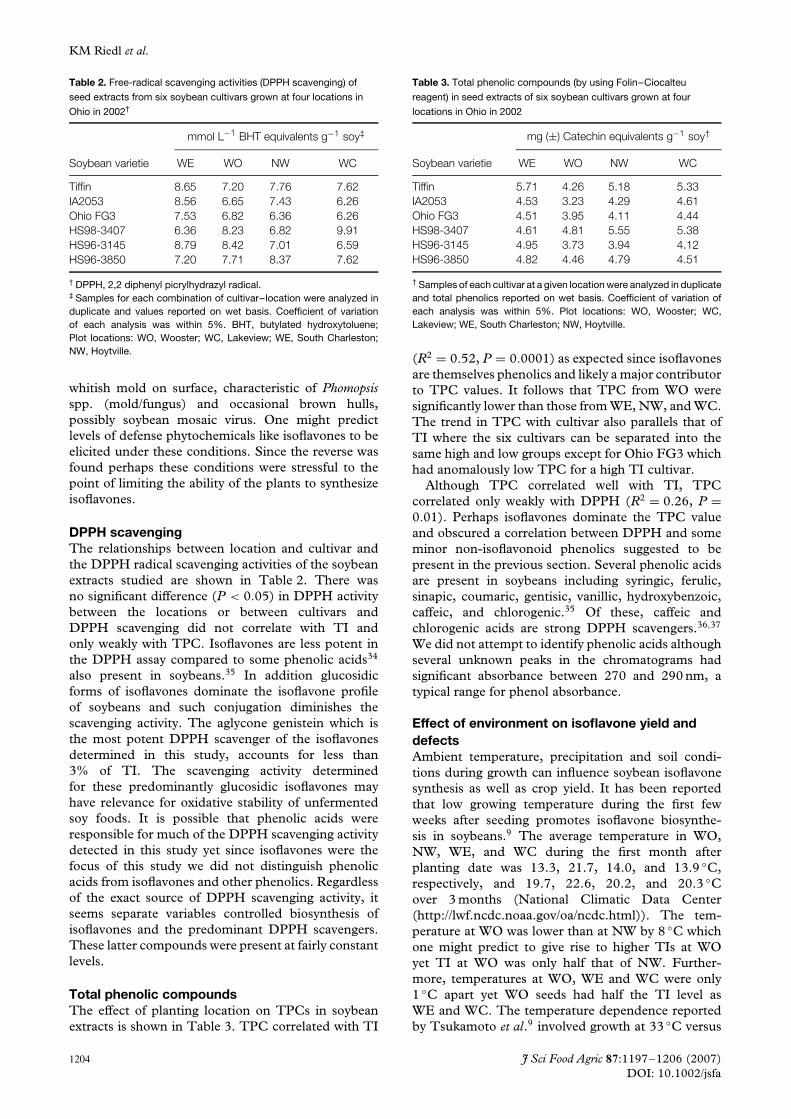
Table 2. Free-radical scavenging activities (DPPH scavenging) of
seed extracts from six soybean cultivars grown at four locations in
Ohio in 2002†
mmol L−1 BHT equivalents g−1 soy‡
Soybean varietie WE WO NW WC
Tiffin 8.65 7.20 7.76 7.62IA2053 8.56 6.65 7.43 6.26Ohio FG3 7.53 6.82 6.36 6.26HS98-3407 6.36 8.23 6.82 9.91HS96-3145 8.79 8.42 7.01 6.59HS96-3850 7.20 7.71 8.37 7.62
† DPPH, 2,2 diphenyl picrylhydrazyl radical.‡ Samples for each combination of cultivar–location were analyzed induplicate and values reported on wet basis. Coefficient of variationof each analysis was within 5%. BHT, butylated hydroxytoluene;Plot locations: WO, Wooster; WC, Lakeview; WE, South Charleston;NW, Hoytville.
whitish mold on surface, characteristic of Phomopsisspp. (mold/fungus) and occasional brown hulls,possibly soybean mosaic virus. One might predictlevels of defense phytochemicals like isoflavones to beelicited under these conditions. Since the reverse wasfound perhaps these conditions were stressful to thepoint of limiting the ability of the plants to synthesizeisoflavones.
DPPH scavengingThe relationships between location and cultivar andthe DPPH radical scavenging activities of the soybeanextracts studied are shown in Table 2. There wasno significant difference (P < 0.05) in DPPH activitybetween the locations or between cultivars andDPPH scavenging did not correlate with TI andonly weakly with TPC. Isoflavones are less potent inthe DPPH assay compared to some phenolic acids34
also present in soybeans.35 In addition glucosidicforms of isoflavones dominate the isoflavone profileof soybeans and such conjugation diminishes thescavenging activity. The aglycone genistein which isthe most potent DPPH scavenger of the isoflavonesdetermined in this study, accounts for less than3% of TI. The scavenging activity determinedfor these predominantly glucosidic isoflavones mayhave relevance for oxidative stability of unfermentedsoy foods. It is possible that phenolic acids wereresponsible for much of the DPPH scavenging activitydetected in this study yet since isoflavones were thefocus of this study we did not distinguish phenolicacids from isoflavones and other phenolics. Regardlessof the exact source of DPPH scavenging activity, itseems separate variables controlled biosynthesis ofisoflavones and the predominant DPPH scavengers.These latter compounds were present at fairly constantlevels.
Total phenolic compoundsThe effect of planting location on TPCs in soybeanextracts is shown in Table 3. TPC correlated with TI
Table 3. Total phenolic compounds (by using Folin–Ciocalteu
reagent) in seed extracts of six soybean cultivars grown at four
locations in Ohio in 2002
mg (±) Catechin equivalents g−1 soy†
Soybean varietie WE WO NW WC
Tiffin 5.71 4.26 5.18 5.33IA2053 4.53 3.23 4.29 4.61Ohio FG3 4.51 3.95 4.11 4.44HS98-3407 4.61 4.81 5.55 5.38HS96-3145 4.95 3.73 3.94 4.12HS96-3850 4.82 4.46 4.79 4.51
† Samples of each cultivar at a given location were analyzed in duplicateand total phenolics reported on wet basis. Coefficient of variation ofeach analysis was within 5%. Plot locations: WO, Wooster; WC,Lakeview; WE, South Charleston; NW, Hoytville.
(R2 = 0.52, P = 0.0001) as expected since isoflavonesare themselves phenolics and likely a major contributorto TPC values. It follows that TPC from WO weresignificantly lower than those from WE, NW, and WC.The trend in TPC with cultivar also parallels that ofTI where the six cultivars can be separated into thesame high and low groups except for Ohio FG3 whichhad anomalously low TPC for a high TI cultivar.
Although TPC correlated well with TI, TPCcorrelated only weakly with DPPH (R2 = 0.26, P =0.01). Perhaps isoflavones dominate the TPC valueand obscured a correlation between DPPH and someminor non-isoflavonoid phenolics suggested to bepresent in the previous section. Several phenolic acidsare present in soybeans including syringic, ferulic,sinapic, coumaric, gentisic, vanillic, hydroxybenzoic,caffeic, and chlorogenic.35 Of these, caffeic andchlorogenic acids are strong DPPH scavengers.36,37
We did not attempt to identify phenolic acids althoughseveral unknown peaks in the chromatograms hadsignificant absorbance between 270 and 290 nm, atypical range for phenol absorbance.
Effect of environment on isoflavone yield anddefectsAmbient temperature, precipitation and soil condi-tions during growth can influence soybean isoflavonesynthesis as well as crop yield. It has been reportedthat low growing temperature during the first fewweeks after seeding promotes isoflavone biosynthe-sis in soybeans.9 The average temperature in WO,NW, WE, and WC during the first month afterplanting date was 13.3, 21.7, 14.0, and 13.9 ◦C,respectively, and 19.7, 22.6, 20.2, and 20.3 ◦Cover 3 months (National Climatic Data Center(http://lwf.ncdc.noaa.gov/oa/ncdc.html)). The tem-perature at WO was lower than at NW by 8 ◦C whichone might predict to give rise to higher TIs at WOyet TI at WO was only half that of NW. Further-more, temperatures at WO, WE and WC were only1 ◦C apart yet WO seeds had half the TI level asWE and WC. The temperature dependence reportedby Tsukamoto et al.9 involved growth at 33 ◦C versus
1204 J Sci Food Agric 87:1197–1206 (2007)DOI: 10.1002/jsfa

Isoflavone profiles of soybeans
17 ◦C, i.e. a 16 ◦C difference. Something other thangrowing temperature appears to have been the dom-inant factor affecting TIs in this study. The majorityof isoflavones in seeds are synthesized during seedfill.38 Seed fill follows pod development continuingfor ∼2 weeks before seed maturation commences. Arecent report involving a controlled irrigation studyalerted us to the possibility that rainfall during seedfill might determine isoflavone content.39 Indeed, TIcontent of seeds at each location followed increasingorder of precipitation during seed fill where precipita-tion at WO was 26 mm, WC was 48 mm, WE 52 mmand NW 57 mm but did not correspond with totalrainfall over the entire growing season. The soil atWO also has low water-holding capacity and this mayhave exacerbated the precipitation effect and furtherdepressed isoflavone synthesis in WO seeds. Precip-itation during the rapid growth phase of soybeansis a reliable predictor of crop yield. Soybean plantsconsume the most water during the first 3 monthsafter planting; this period of development includesflowering and pod development. High isoflavone con-tent would not be advantageous unless married withhigh soybean yields. Records of rainfall over 3 monthsafter planting showed that WE received much morerain than the other three locations: an average of76 mm per 2 week period at WE versus 33 mm at theother three locations (National Climatic Data Cen-ter (http://lwf.ncdc.noaa.gov/oa/ncdc.html)). Averagecrop yield at WE (3.4 Mg ha−1 = 51 bushels acre−1)was approximately 50% higher than at WO, NW, andWC which gave 2.6 Mg ha−1 (39 bushels acre−1),2.3 Mg ha−1 (34 bushels acre−1) and 2.2 Mg ha−1 (33bushels acre−1), respectively. Between cultivars theyield averaged over the four locations varied by only∼15% with Tiffin and HS98-3407 having the highestyields. Yield did not correlate with isoflavones, DPPHor TPC (R2 < 0.1, data not shown). This result is incontrast with a report by Yin and Vyn40 which foundisoflavones increased proportional to seed yield.
Several combinations of cultivar and locationresulted in both high isoflavone content and goodyield. Since WE was among the top three TI-producing locations and gave the highest yield it wasclearly the superior location in this study. Growthat the other three locations resulted in relatively lowyields and since WO also had low isoflavone contentit was clearly the most inferior location. It follows thatTiffin and HS98-3407 which had the highest yieldsaveraged over the four locations and also gave two ofthe highest TI values are the most promising cultivarsfor food use. HS98-3407 is already classified as a food-grade soybean and Tiffin might be considered for fooduse or breeding even though at present it is considereda commodity-type cultivar.
CONCLUSIONSTI content in soybean seeds was influenced by plantinglocation and cultivar giving TI values that varied
five-fold. DPPH scavenging activity was relativelyconstant regardless of cultivar or location and TPCsimply correlated with TI. With increasing TI therewas a distinct change in isoflavone profile shiftingfrom genisteins as the predominant family to similaramounts of the daidzein and genistein families andwithin the daidzein family towards malonyl daidzin.Rainfall during seed fill was the only environmentalvariable identified that correlated with TI. The bestgrowing locations and soybean cultivars in this studybased on a combination of good yield and TI wereWE and the cultivars Tiffin and HS98-3407.
This is the first study of soybean isoflavones toour knowledge that includes several Ohio soybeancultivars grown at several locations in the samegrowing season. The performance of the variouscultivars and growing locations can help select cultivarsbased on various parameters. The shift in isoflavonemetabolism towards daidzeins from genistein andglycitein is a novel finding. Correlations betweencertain isoflavones and total isoflavones found inthis study may help identify environmental variablesassociated with isoflavone production and lead toimproved disease resistance of soybean crops.
ACKNOWLEDGEMENTThis research was supported by a research grantprovided by the Ohio Agricultural and ResearchDevelopment Center. Special thanks to Dr TerrenceGraham (Dept. Plant Pathology, The Ohio StateUniversity) for discussions on isoflavone metabolism,plant defense and help identifying seed defects.Thanks also to Torsten Bohn for advice on statisticalanalysis.
REFERENCES1 U.S. Food and Drug Administration, Food Labeling: Health
claims; soy protein and coronary heart disease. Federal Register64:57699–57733 (1999).
2 Messina MJ, Persky V, Setchell KDR and Barnes S, Soy intakeand cancer risk – a review of the in-vitro and in-vivo data.Nutr Cancer 21:113–131 (1994).
3 Potter SM, Baum JA, Teng HY, Stillman RJ, Shay NF andErdman JW, Soy protein and isoflavones: their effects onblood lipids and bone density in postmenopausal women. AmJ Clin Nutr 68:1375S–13759S (1998).
4 Hendrich S, Wang GJ, Lin HK, Xu X, Twe BY, Wang HJ,et al, Isoflavone metabolism and bioavailability, in AntioxidantStatus, Diet, Nutrition and Health, ed. by Papas AM. CRC,Boca Raton, FL, pp. 211–230 (1999).
5 Hendrich S, Wang GJ, Xu X, Twe BY, Wang HJ and Mur-phy PA, Human bioavailability of soybean isoflavones: influ-ences of diet, dose, time and gut microflora, in FunctionalFoods for Disease Prevention, vol. 701, ed. by Shibamoto T,Terao J and Osawa T. American Chemical Society, Washing-ton, DC, pp. 150–156 (1998).
6 Kulling SE, Honig DM, Simat TJ and Metzler M, Oxidativein vitro metabolism of the soy phytoestrogens daidzein andgenistein. J Agric Food Chem 48:4963–4972 (2000).
7 Hoeck JA, Fehr WR, Murphy PA and Welke GA, Influence ofgenotype and environment on isoflavone contents of soybean.Crop Sci 40:48–51 (2000).
J Sci Food Agric 87:1197–1206 (2007) 1205DOI: 10.1002/jsfa

KM Riedl et al.
8 Lee SJ, Yan WK, Ahn JK and Chung IM, Effects of year,site, genotype and their interactions on various soybeanisoflavones. Field Crop Res 81:181–192 (2003).
9 Tsukamoto C, Shimada S, Igita K, Kudou S, Kokubun M,Okubo K, et al, Factors affecting isoflavone content in soy-bean seeds – changes in isoflavones, saponins, and compo-sition of fatty-acids at different temperatures during seeddevelopment. J Agric Food Chem 43:1184–1192 (1995).
10 Vyn TJ, Yin XH, Bruulsema TW, Jackson CJC, Rajcan I andBrouder SM, Potassium fertilization effects on isoflavoneconcentrations in soybean [Glycine max (L.) Merr.]. J AgricFood Chem 50:3501–3506 (2002).
11 Wang HJ and Murphy PA, Isoflavone content in commercialsoybean foods. J Agric Food Chem 42:1666–1673 (1994).
12 Caldwell CR, Britz SJ and Mirecki RM, Effect of temperature,elevated carbon dioxide, and drought during seed develop-ment on the isoflavone content of dwarf soybean [Glycinemax (L.) Merrill] grown in controlled environments. J AgricFood Chem 53:1125–1129 (2005).
13 Lozovaya VV, Lygin AV, Ulanov AV, Nelson RL, Dayde Jand Widhohn JM, Effect of temperature and soil moisturestatus during seed development on soybean seed isoflavoneconcentration and composition. Crop Sci 45:1934–1940(2005).
14 Lee SJ, Ahn JK, Kim SH, Kim JT, Han SJ, Jung MY, et al,Variation in isoflavone of soybean cultivars with locationand storage duration. J Agric Food Chem 51:3382–3389(2003).
15 Min DB, Lipid oxidation of edible oil, in Food Lipids: Chemistry,Nutrition and Biotechnology, ed. by Akoh CC and Min DB.Marcel Dekker, Inc., New York, pp. 283–296 (1998).
16 Wickens AP, Ageing and the free radical theory. Respir Physiol128:379–391 (2001).
17 Naim M, Gestetner B, Bondi A and Birk Y, Antioxidative andantihemolytic activities of soybean isoflavones. J Agric FoodChem 24:1174–1177 (1976).
18 Patel RP, Boersma BJ, Crawford JH, Hogg N, Kirk M, Kalya-naraman B, et al, Antioxidant mechanisms of isoflavones inlipid systems: Paradoxical effects of peroxyl radical scaveng-ing. Free Radic Biol Med 31:1570–1581 (2001).
19 Lee S-J, Chung I-M, Ahn J-K, Lee S-K, Kim SH and Yoo N-H, Variation in antioxidative activity of soybean (Glycine maxL.) varieties with crop year and duration of storage time. FoodSci Biotech 11:649–653 (2002).
20 Hou HJ and Chang KC, Interconversions of isoflavones insoybeans as affected by storage. J Food Sci 67:2083–2089(2002).
21 Nguyenle T, Wang E and Cheung AP, An investigationon the extraction and concentration of isoflavones insoy-based products. J Pharm Biomed Anal 14:221–232(1995).
22 Walsh KR, Zhang YC, Vodovotz Y, Schwartz SJ and Failla ML,Stability and bioaccessibility of isoflavones from soy breadduring in vitro digestion. J Agric Food Chem 51:4603–9(2003).
23 Griffith AP and Collison MW, Improved methods for theextraction and analysis of isoflavones from soy-containingfoods and nutritional supplements by reversed-phase high-performance liquid chromatography and liquid chromatog-raphy–mass spectrometry. J Chromatogr A 913:397–413(2001).
24 Murphy PA, Barua K and Hauck CC, Solvent extractionselection in the determination of isoflavones in soy foods.J Chromatogr B 777:129–138 (2002).
25 Wang HJ and Murphy PA, Isoflavone composition of Americanand Japanese soybeans in Iowa – effects of variety, crop year,and location. J Agric Food Chem 42:1674–1677 (1994).
26 Franke AA, Custer LJ, Cerna CM and Narala KK, Quantitationof phytoestrogens in legumes by HPLC. J Agric Food Chem42:1905–1913 (1994).
27 Ozcelik B, Lee JH and Min DB, Effects of light, oxygen, and pHon the absorbance of 2,2-diphenyl-1-picrylhydrazyl. J Food Sci68:487–490 (2003).
28 Zielinski H and Kozlowska H, Antioxidant activity and totalphenolics in selected cereal grains and their differentmorphological fractions. J Agric Food Chem 48:2008–2016(2000).
29 Eldridge AC and Kwolek WF, Soybean isoflavones – effect ofenvironment and variety on composition. J Agric Food Chem31:394–396 (1983).
30 Graham TL, Kim JE and Graham MY, Role of constitutiveisoflavone conjugates in the accumulation of glyceollinin soybean infected with Phytophthora–Megasperma. MolPlant–Microbe Interact 3:157–166 (1990).
31 Lee J, Renita M, Fioritto RJ, St Martin SK, Schwartz SJ andVodovotz Y, Isoflavone characterization and antioxidantactivity of Ohio soybeans. J Agric Food Chem 52:2647–2651(2004).
32 Graham TL and Graham MY, Signaling in soybean phenyl-propanoid responses – Dissection of primary, secondary, andconditioning effects of light, wounding, and elicitor treat-ments. Plant Physiol 110:1123–1133 (1996).
33 Yu O, Shi J, Hession AO, Maxwell CA, McGonigle B andOdell JT, Metabolic engineering to increase isoflavonebiosynthesis in soybean seed. Phytochemistry 63:753–763(2003).
34 Cos P, Hermans N, Calomme M, Maes L, De Bruyne T,Pieters L, et al, Comparative study of eight well-knownpolyphenolic antioxidants. J Pharm Pharmacol 55:1291–1297(2003).
35 Kozlowska H, Zadernowski R and Sosulski FW, Phenolic acidsin oilseed flours. Nahrung 27:449–453 (1983).
36 Chen JH and Ho CT, Antioxidant activities of caffeic acid andits related hydroxycinnamic acid compounds. J Agric FoodChem 45:2374–2378 (1997).
37 Sroka Z and Cisowski W, Hydrogen peroxide scavenging,antioxidant and anti-radical activity of some phenolic acids.Food Chem Toxicol 41:753–758 (2003).
38 Dhaubhadel S, McGarvey BD, Williams R and Gijzen M,Isoflavonoid biosynthesis and accumulation in developingsoybean seeds. Plant Mol Biol 53:733–743 (2003).
39 Bennett JO, Yu O, Heatherly LG and Krishnan HB, Accumula-tion of genistein and daidzein, soybean isoflavones implicatedin promoting human health, is significantly elevated by irriga-tion. J Agric Food Chem 52:7574–7579 (2004).
40 Yin XH and Vyn TJ, Relationships of isoflavone, oil, and proteinin seed with yield of soybean. Agronomy J 97:1314–1321(2005).
1206 J Sci Food Agric 87:1197–1206 (2007)DOI: 10.1002/jsfa
Related Documents











