Involvement of ryanodine receptors in neurotrophin- induced hippocampal synaptic plasticity and spatial memory formation Tatiana Adasme a,1 , Paola Haeger a,1 , Andrea C. Paula-Lima a , Italo Espinoza a , M. Mercedes Casas-Alarcón a , M. Angélica Carrasco a,b,2 , and Cecilia Hidalgo a,b,3 a Centro de Estudios Moleculares de la Célula, Facultad de Medicina, and b Programa de Fisiología y Biofísica, Instituto de Ciencias Biomédicas, Universidad de Chile, 838-0453 Santiago, Chile Edited* by Ramón Latorre, Centro de Neurociencias, Universidad de Valparaíso, Valparaíso, Chile, and approved January 5, 2011 (received for review September 11, 2010) Ryanodine receptors (RyR) amplify activity-dependent calcium inux via calcium-induced calcium release. Calcium signals trigger post- synaptic pathways in hippocampal neurons that underlie synaptic plasticity, learning, and memory. Recent evidence supports a role of the RyR2 and RyR3 isoforms in these processes. Along with calcium signals, brain-derived neurotrophic factor (BDNF) is a key signaling molecule for hippocampal synaptic plasticity and spatial memory. Upon binding to specic TrkB receptors, BDNF initiates complex signaling pathways that modify synaptic structure and function. Here, we show that BDNF-induced remodeling of hippocampal dendritic spines required functional RyR. Additionally, incubation with BDNF enhanced the expression of RyR2, RyR3, and PKMζ, an atypical protein kinase C isoform with key roles in hippocampal mem- ory consolidation. Consistent with their increased RyR protein con- tent, BDNF-treated neurons generated larger RyR-mediated calcium signals than controls. Selective inhibition of RyR-mediated calcium release with inhibitory ryanodine concentrations prevented the PKMζ, RyR2, and RyR3 protein content enhancement induced by BDNF. Intrahippocampal injection of BDNF or training rats in a spatial memory task enhanced PKMζ, RyR2, RyR3, and BDNF hippocampal protein content, while injection of ryanodine at concentrations that stimulate RyR-mediated calcium release improved spatial memory learning and enhanced memory consolidation. We propose that RyR-generated calcium signals are key features of the complex neu- ronal plasticity processes induced by BDNF, which include increased expression of RyR2, RyR3, and PKMζ and the spine remodeling re- quired for spatial memory formation. dendritic spine remodeling | Morris water maze | endoplasmic reticulum | protein expression A ctivity-generated neuronal Ca +2 signals promote gene tran- scription and protein synthesis, two essential events of long- lasting synaptic plasticity and memory formation (1). Hippo- campal neuronal plasticity requires de novo protein synthesis for the generation and growth of dendritic spines (2). Brain-derived neurotrophic factor (BDNF) is a key signaling factor implicated in activity-dependent neuronal changes underlying memory ac- quisition and persistence of long-term memory storage (3). At the cellular level, BDNF enhances synaptic transmission in rat hip- pocampus by inducing local protein synthesis in postsynaptic dendrites (4). A central feature of the responses elicited by BDNF is cyclic AMP-responsive element binding (CREB) activation (5), which occurs via BDNF-induced signaling pathways that promote the expression of genes that prompt the formation, elongation and stretching of dendritic spines (4, 6). BDNF binding to specic TrkB receptors induces Ca 2+ release through inositol 1,4,5- trisphosphate receptors (InsP3R) and stimulates calcium- dependent kinases in hippocampal neurons (6), which contribute to induce and maintain long-term potentiation (LTP) (7). Besides BDNF, other signaling molecules are essential for late- LTP and long-term memory formation. Among them, protein kinase Mζ (PKMζ), an atypical protein kinase C isoform with constitutive activity (8), is both necessary and sufcient for hip- pocampal late-LTP maintenance (9, 10) and long-term memory retention (11). Translation of PKMζ is rapidly up-regulated during LTP induction (12), whereas selective PKMζ inhibition reverses hippocampal LTP maintenance in vivo and disrupts long- term memory retention elaborated in the hippocampus (13). The essential role of Ca +2 signals in long-term neuronal plasticity processes is well established (14). Ca +2 release through ryanodine receptors (RyR) and InsP3R present in the endoplasmic reticulum of axons and dendrites produces Ca +2 signals (15) that make an important contribution to synaptic plasticity processes in central neurons (16). In particular, RyR-mediated Ca +2 -induced Ca +2 re- lease (CICR) provides a major fraction of the Ca +2 increase induced in hippocampal CA1 spines by synaptic activity (17). High-frequency eld stimulation of primary hippocampal neurons produces Ca +2 signals primarily derived from RyR-mediated Ca +2 release (18), while caffeine-induced RyR-mediated Ca 2+ release promotes den- dritic spine elongation in primary hippocampal neurons (19). Sev- eral reports support RyR participation in hippocampal LTP and memory processes (20–22). Thus, RyR inhibition before tetanic stimulation suppresses hippocampal long-lasting LTP, and RyR activation converts early LTP into late-LTP (23). Hippocampal expression of the RyR2 isoform increases after spatial memory training (24), but selective knockdown of RyR2 and RyR3, but not of RyR1, impairs avoidance memory processes (20). Inhibition of overall brain RyR activity with dantrolene decreases associative memory in chicken (21), whereas the RyR agonist 4-chloro-m-cresol (4-CMC) administered to whole brain 30 min before a passive avoidance task enhances memory retention in mice (20) and pro- motes long-term memory consolidation in young chicks (22). The present results show that BDNF increased RyR2, RyR3, and PKMζ protein expression and promoted spine remodeling in primary hippocampal neurons; these effects required functional RyR. Intrahippocampus BDNF injection and memory training also enhanced RyR2, RyR3, and PKMζ expression, while intra- hippocampus ryanodine injection at concentrations that stimu- late RyR-mediated Ca 2+ release facilitated memory acquisition and consolidation. Results BDNF Stimulates Hippocampal Expression of the Ca 2+ Release Channel Isoforms RyR2 and RyR3 and of the Plasticity-Related Protein PKMζ. Throughout this work, we used primary hippocampal cultures at 14 Author contributions: T.A., P.H., M.A.C., and C.H. designed research; T.A., P.H., A.C.P.-L., I.E., and M.M.C.-A. performed research; T.A., P.H., A.C.P.-L., I.E., M.M.C.-A., M.A.C., and C.H. analyzed data; and T.A., P.H., A.C.P.-L., and C.H. wrote the paper. The authors declare no conict of interest. *This Direct Submission article had a prearranged editor. 1 T.A. and P.H. contributed equally to this work. 2 Deceased November 19, 2010. 3 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1013580108/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1013580108 PNAS | February 15, 2011 | vol. 108 | no. 7 | 3029–3034 NEUROSCIENCE

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Involvement of ryanodine receptors in neurotrophin-induced hippocampal synaptic plasticity and spatialmemory formationTatiana Adasmea,1, Paola Haegera,1, Andrea C. Paula-Limaa, Italo Espinozaa, M. Mercedes Casas-Alarcóna,M. Angélica Carrascoa,b,2, and Cecilia Hidalgoa,b,3
aCentro de Estudios Moleculares de la Célula, Facultad de Medicina, and bPrograma de Fisiología y Biofísica, Instituto de Ciencias Biomédicas, Universidad deChile, 838-0453 Santiago, Chile
Edited* by Ramón Latorre, Centro de Neurociencias, Universidad de Valparaíso, Valparaíso, Chile, and approved January 5, 2011 (received for reviewSeptember 11, 2010)
Ryanodine receptors (RyR) amplify activity-dependent calcium in!uxvia calcium-induced calcium release. Calcium signals trigger post-synaptic pathways in hippocampal neurons that underlie synapticplasticity, learning, and memory. Recent evidence supports a role ofthe RyR2 and RyR3 isoforms in these processes. Along with calciumsignals, brain-derived neurotrophic factor (BDNF) is a key signalingmolecule for hippocampal synaptic plasticity and spatial memory.Upon binding to speci"c TrkB receptors, BDNF initiates complexsignaling pathways that modify synaptic structure and function.Here, we show that BDNF-induced remodeling of hippocampaldendritic spines required functional RyR. Additionally, incubationwith BDNF enhanced the expression of RyR2, RyR3, and PKM!, anatypicalproteinkinaseC isoformwithkey roles inhippocampalmem-ory consolidation. Consistent with their increased RyR protein con-tent, BDNF-treated neurons generated larger RyR-mediated calciumsignals than controls. Selective inhibition of RyR-mediated calciumrelease with inhibitory ryanodine concentrations prevented thePKM!, RyR2, and RyR3 protein content enhancement induced byBDNF. Intrahippocampal injectionof BDNFor training rats in a spatialmemory task enhanced PKM!, RyR2, RyR3, and BDNF hippocampalprotein content, while injection of ryanodine at concentrations thatstimulate RyR-mediated calcium release improved spatial memorylearning and enhanced memory consolidation. We propose thatRyR-generated calcium signals are key features of the complex neu-ronal plasticity processes induced by BDNF, which include increasedexpression of RyR2, RyR3, and PKM! and the spine remodeling re-quired for spatial memory formation.
dendritic spine remodeling | Morris water maze | endoplasmic reticulum |protein expression
Activity-generated neuronal Ca+2 signals promote gene tran-scription and protein synthesis, two essential events of long-
lasting synaptic plasticity and memory formation (1). Hippo-campal neuronal plasticity requires de novo protein synthesis forthe generation and growth of dendritic spines (2). Brain-derivedneurotrophic factor (BDNF) is a key signaling factor implicatedin activity-dependent neuronal changes underlying memory ac-quisition and persistence of long-term memory storage (3). At thecellular level, BDNF enhances synaptic transmission in rat hip-pocampus by inducing local protein synthesis in postsynapticdendrites (4). A central feature of the responses elicited by BDNFis cyclic AMP-responsive element binding (CREB) activation (5),which occurs via BDNF-induced signaling pathways that promotethe expression of genes that prompt the formation, elongationand stretching of dendritic spines (4, 6). BDNF binding to speci!cTrkB receptors induces Ca2+ release through inositol 1,4,5-trisphosphate receptors (InsP3R) and stimulates calcium-dependent kinases in hippocampal neurons (6), which contributeto induce and maintain long-term potentiation (LTP) (7).Besides BDNF, other signaling molecules are essential for late-
LTP and long-term memory formation. Among them, proteinkinase M! (PKM!), an atypical protein kinase C isoform with
constitutive activity (8), is both necessary and suf!cient for hip-pocampal late-LTP maintenance (9, 10) and long-term memoryretention (11). Translation of PKM! is rapidly up-regulatedduring LTP induction (12), whereas selective PKM! inhibitionreverses hippocampal LTPmaintenance in vivo and disrupts long-term memory retention elaborated in the hippocampus (13).The essential role of Ca+2 signals in long-term neuronal plasticity
processes is well established (14). Ca+2 release through ryanodinereceptors (RyR) and InsP3R present in the endoplasmic reticulumof axons and dendrites produces Ca+2 signals (15) that make animportant contribution to synaptic plasticity processes in centralneurons (16). In particular, RyR-mediated Ca+2-induced Ca+2 re-lease (CICR) provides amajor fraction of theCa+2 increase inducedin hippocampalCA1 spines by synaptic activity (17).High-frequency!eld stimulation of primary hippocampal neurons produces Ca+2
signals primarily derived from RyR-mediated Ca+2 release (18),while caffeine-induced RyR-mediated Ca2+ release promotes den-dritic spine elongation in primary hippocampal neurons (19). Sev-eral reports support RyR participation in hippocampal LTP andmemory processes (20–22). Thus, RyR inhibition before tetanicstimulation suppresses hippocampal long-lasting LTP, and RyRactivation converts early LTP into late-LTP (23). Hippocampalexpression of the RyR2 isoform increases after spatial memorytraining (24), but selective knockdown of RyR2 and RyR3, but notof RyR1, impairs avoidance memory processes (20). Inhibition ofoverall brain RyR activity with dantrolene decreases associativememory in chicken (21), whereas theRyRagonist 4-chloro-m-cresol(4-CMC) administered to whole brain 30 min before a passiveavoidance task enhances memory retention in mice (20) and pro-motes long-term memory consolidation in young chicks (22).The present results show that BDNF increased RyR2, RyR3,
and PKM! protein expression and promoted spine remodeling inprimary hippocampal neurons; these effects required functionalRyR. Intrahippocampus BDNF injection and memory trainingalso enhanced RyR2, RyR3, and PKM! expression, while intra-hippocampus ryanodine injection at concentrations that stimu-late RyR-mediated Ca2+ release facilitated memory acquisitionand consolidation.
ResultsBDNF Stimulates Hippocampal Expression of the Ca2+ Release ChannelIsoforms RyR2 and RyR3 and of the Plasticity-Related Protein PKM!.Throughout this work, we used primary hippocampal cultures at 14
Author contributions: T.A., P.H., M.A.C., and C.H. designed research; T.A., P.H., A.C.P.-L.,I.E., and M.M.C.-A. performed research; T.A., P.H., A.C.P.-L., I.E., M.M.C.-A., M.A.C., andC.H. analyzed data; and T.A., P.H., A.C.P.-L., and C.H. wrote the paper.
The authors declare no con!ict of interest.
*This Direct Submission article had a prearranged editor.1T.A. and P.H. contributed equally to this work.2Deceased November 19, 2010.3To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1013580108/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1013580108 PNAS | February 15, 2011 | vol. 108 | no. 7 | 3029–3034
NEU
ROSC
IENCE
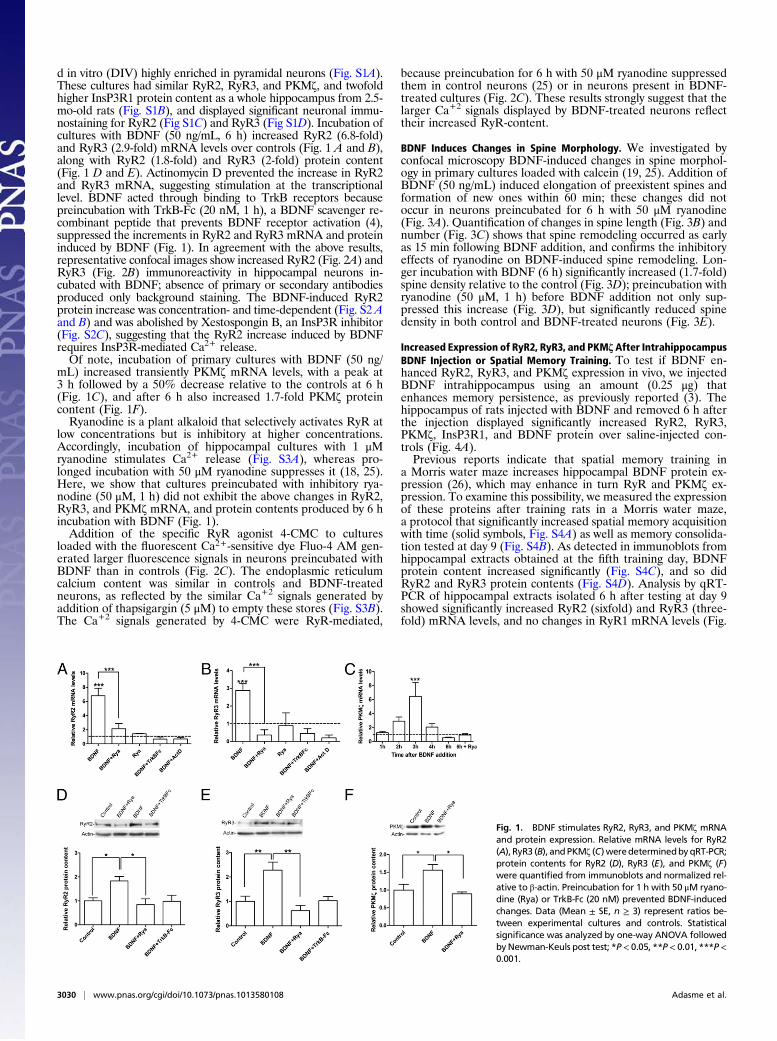
d in vitro (DIV) highly enriched in pyramidal neurons (Fig. S1A).These cultures had similar RyR2, RyR3, and PKM!, and twofoldhigher InsP3R1 protein content as a whole hippocampus from 2.5-mo-old rats (Fig. S1B), and displayed signi!cant neuronal immu-nostaining for RyR2 (Fig S1C) and RyR3 (Fig S1D). Incubation ofcultures with BDNF (50 ng/mL, 6 h) increased RyR2 (6.8-fold)and RyR3 (2.9-fold) mRNA levels over controls (Fig. 1 A and B),along with RyR2 (1.8-fold) and RyR3 (2-fold) protein content(Fig. 1 D and E). Actinomycin D prevented the increase in RyR2and RyR3 mRNA, suggesting stimulation at the transcriptionallevel. BDNF acted through binding to TrkB receptors becausepreincubation with TrkB-Fc (20 nM, 1 h), a BDNF scavenger re-combinant peptide that prevents BDNF receptor activation (4),suppressed the increments in RyR2 and RyR3 mRNA and proteininduced by BDNF (Fig. 1). In agreement with the above results,representative confocal images show increased RyR2 (Fig. 2A) andRyR3 (Fig. 2B) immunoreactivity in hippocampal neurons in-cubated with BDNF; absence of primary or secondary antibodiesproduced only background staining. The BDNF-induced RyR2protein increase was concentration- and time-dependent (Fig. S2 Aand B) and was abolished by Xestospongin B, an InsP3R inhibitor(Fig. S2C), suggesting that the RyR2 increase induced by BDNFrequires InsP3R-mediated Ca2+ release.Of note, incubation of primary cultures with BDNF (50 ng/
mL) increased transiently PKM! mRNA levels, with a peak at3 h followed by a 50% decrease relative to the controls at 6 h(Fig. 1C), and after 6 h also increased 1.7-fold PKM! proteincontent (Fig. 1F).Ryanodine is a plant alkaloid that selectively activates RyR at
low concentrations but is inhibitory at higher concentrations.Accordingly, incubation of hippocampal cultures with 1 "Mryanodine stimulates Ca2+ release (Fig. S3A), whereas pro-longed incubation with 50 "M ryanodine suppresses it (18, 25).Here, we show that cultures preincubated with inhibitory rya-nodine (50 "M, 1 h) did not exhibit the above changes in RyR2,RyR3, and PKM! mRNA, and protein contents produced by 6 hincubation with BDNF (Fig. 1).Addition of the speci!c RyR agonist 4-CMC to cultures
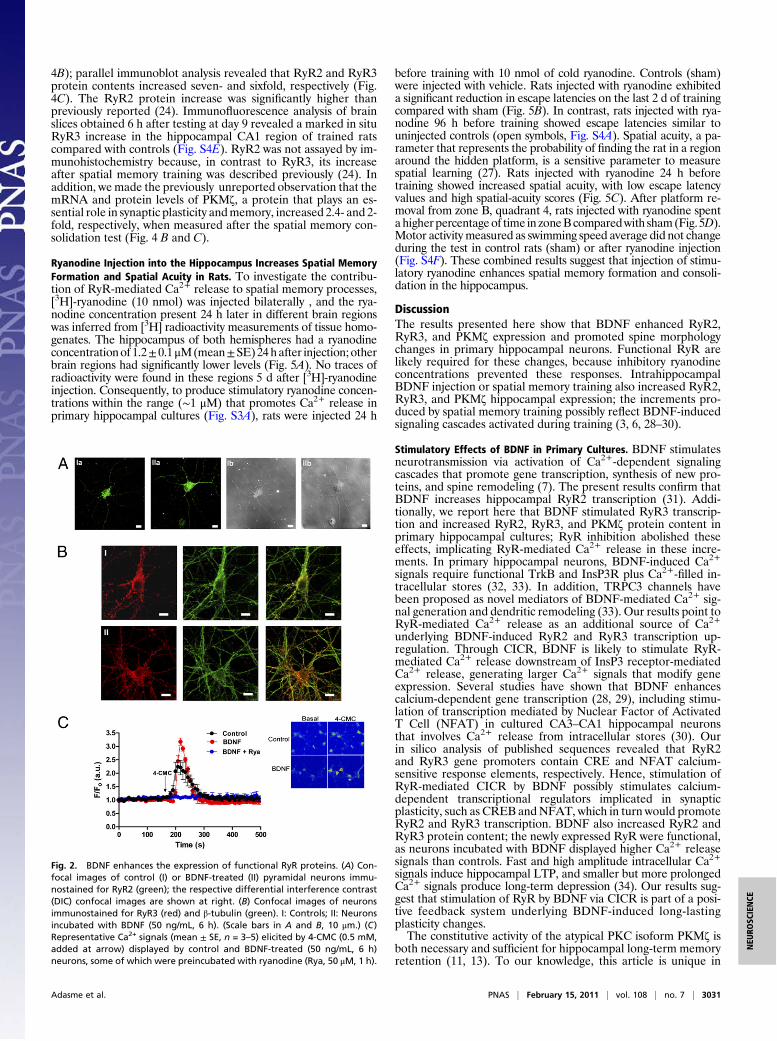
loaded with the "uorescent Ca2+-sensitive dye Fluo-4 AM gen-erated larger "uorescence signals in neurons preincubated withBDNF than in controls (Fig. 2C). The endoplasmic reticulumcalcium content was similar in controls and BDNF-treatedneurons, as re"ected by the similar Ca+2 signals generated byaddition of thapsigargin (5 "M) to empty these stores (Fig. S3B).The Ca+2 signals generated by 4-CMC were RyR-mediated,
because preincubation for 6 h with 50 "M ryanodine suppressedthem in control neurons (25) or in neurons present in BDNF-treated cultures (Fig. 2C). These results strongly suggest that thelarger Ca+2 signals displayed by BDNF-treated neurons re"ecttheir increased RyR-content.
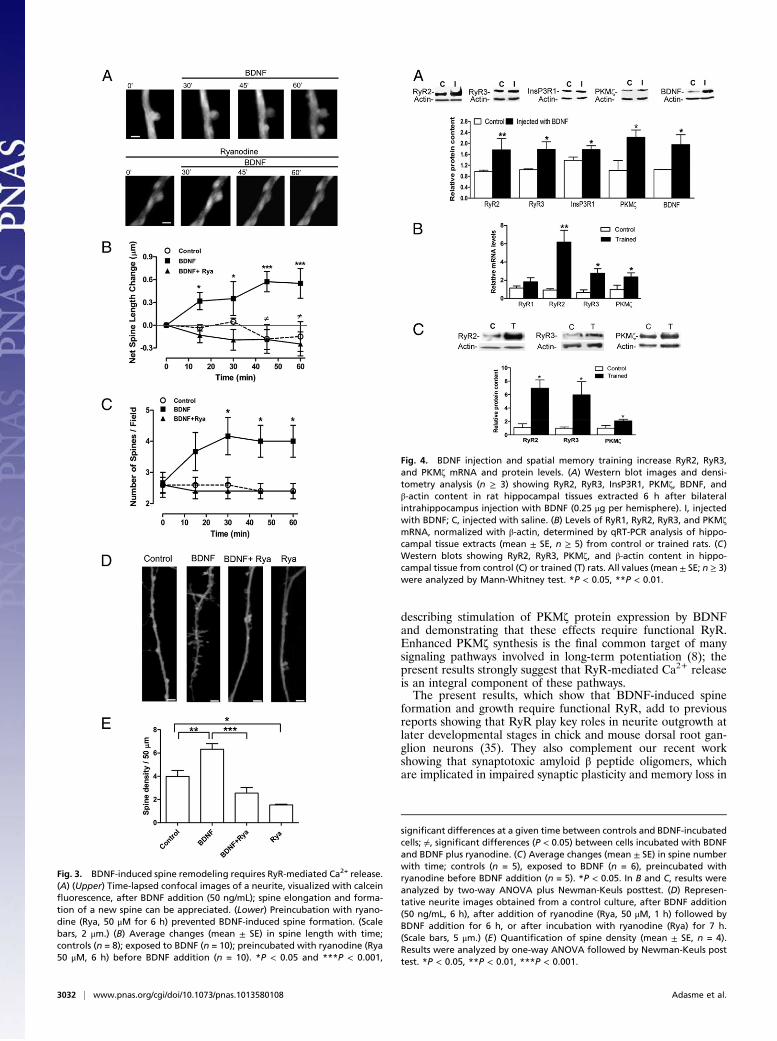
BDNF Induces Changes in Spine Morphology. We investigated byconfocal microscopy BDNF-induced changes in spine morphol-ogy in primary cultures loaded with calcein (19, 25). Addition ofBDNF (50 ng/mL) induced elongation of preexistent spines andformation of new ones within 60 min; these changes did notoccur in neurons preincubated for 6 h with 50 "M ryanodine(Fig. 3A). Quanti!cation of changes in spine length (Fig. 3B) andnumber (Fig. 3C) shows that spine remodeling occurred as earlyas 15 min following BDNF addition, and con!rms the inhibitoryeffects of ryanodine on BDNF-induced spine remodeling. Lon-ger incubation with BDNF (6 h) signi!cantly increased (1.7-fold)spine density relative to the control (Fig. 3D); preincubation withryanodine (50 "M, 1 h) before BDNF addition not only sup-pressed this increase (Fig. 3D), but signi!cantly reduced spinedensity in both control and BDNF-treated neurons (Fig. 3E).
Increased Expression of RyR2, RyR3, and PKM! After IntrahippocampusBDNF Injection or Spatial Memory Training. To test if BDNF en-hanced RyR2, RyR3, and PKM! expression in vivo, we injectedBDNF intrahippocampus using an amount (0.25 "g) thatenhances memory persistence, as previously reported (3). Thehippocampus of rats injected with BDNF and removed 6 h afterthe injection displayed signi!cantly increased RyR2, RyR3,PKM!, InsP3R1, and BDNF protein over saline-injected con-trols (Fig. 4A).Previous reports indicate that spatial memory training in
a Morris water maze increases hippocampal BDNF protein ex-pression (26), which may enhance in turn RyR and PKM! ex-pression. To examine this possibility, we measured the expressionof these proteins after training rats in a Morris water maze,a protocol that signi!cantly increased spatial memory acquisitionwith time (solid symbols, Fig. S4A) as well as memory consolida-tion tested at day 9 (Fig. S4B). As detected in immunoblots fromhippocampal extracts obtained at the !fth training day, BDNFprotein content increased signi!cantly (Fig. S4C), and so didRyR2 and RyR3 protein contents (Fig. S4D). Analysis by qRT-PCR of hippocampal extracts isolated 6 h after testing at day 9showed signi!cantly increased RyR2 (sixfold) and RyR3 (three-fold) mRNA levels, and no changes in RyR1 mRNA levels (Fig.
Fig. 1. BDNF stimulates RyR2, RyR3, and PKM! mRNAand protein expression. Relative mRNA levels for RyR2(A), RyR3 (B), andPKM! (C)weredeterminedbyqRT-PCR;protein contents for RyR2 (D), RyR3 (E), and PKM! (F)were quanti"ed from immunoblots and normalized rel-ative to #-actin. Preincubation for 1 h with 50 "M ryano-dine (Rya) or TrkB-Fc (20 nM) prevented BDNF-inducedchanges. Data (Mean ± SE, n ! 3) represent ratios be-tween experimental cultures and controls. Statisticalsigni"cance was analyzed by one-way ANOVA followedbyNewman-Keuls post test; *P< 0.05, **P< 0.01, ***P<0.001.
3030 | www.pnas.org/cgi/doi/10.1073/pnas.1013580108 Adasme et al.
4B); parallel immunoblot analysis revealed that RyR2 and RyR3protein contents increased seven- and sixfold, respectively (Fig.4C). The RyR2 protein increase was signi!cantly higher thanpreviously reported (24). Immuno"uorescence analysis of brainslices obtained 6 h after testing at day 9 revealed a marked in situRyR3 increase in the hippocampal CA1 region of trained ratscompared with controls (Fig. S4E). RyR2 was not assayed by im-munohistochemistry because, in contrast to RyR3, its increaseafter spatial memory training was described previously (24). Inaddition, wemade the previously unreported observation that themRNA and protein levels of PKM!, a protein that plays an es-sential role in synaptic plasticity andmemory, increased 2.4- and 2-fold, respectively, when measured after the spatial memory con-solidation test (Fig. 4 B and C).
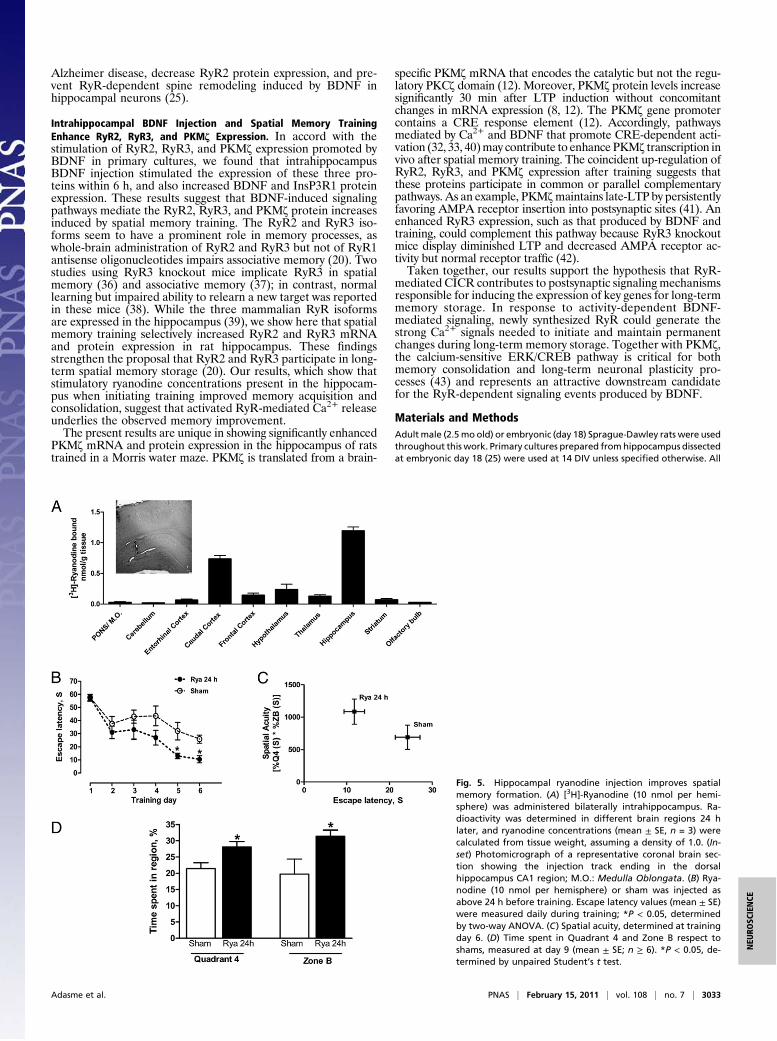
Ryanodine Injection into the Hippocampus Increases Spatial MemoryFormation and Spatial Acuity in Rats. To investigate the contribu-tion of RyR-mediated Ca2+ release to spatial memory processes,[3H]-ryanodine (10 nmol) was injected bilaterally , and the rya-nodine concentration present 24 h later in different brain regionswas inferred from [3H] radioactivity measurements of tissue homo-genates. The hippocampus of both hemispheres had a ryanodineconcentrationof 1.2±0.1"M(mean±SE)24hafter injection;otherbrain regions had signi!cantly lower levels (Fig. 5A). No traces ofradioactivity were found in these regions 5 d after [3H]-ryanodineinjection. Consequently, to produce stimulatory ryanodine concen-trations within the range ("1 "M) that promotes Ca2+ release inprimary hippocampal cultures (Fig. S3A), rats were injected 24 h
before training with 10 nmol of cold ryanodine. Controls (sham)were injected with vehicle. Rats injected with ryanodine exhibiteda signi!cant reduction in escape latencies on the last 2 d of trainingcompared with sham (Fig. 5B). In contrast, rats injected with rya-nodine 96 h before training showed escape latencies similar touninjected controls (open symbols, Fig. S4A). Spatial acuity, a pa-rameter that represents the probability of !nding the rat in a regionaround the hidden platform, is a sensitive parameter to measurespatial learning (27). Rats injected with ryanodine 24 h beforetraining showed increased spatial acuity, with low escape latencyvalues and high spatial-acuity scores (Fig. 5C). After platform re-moval from zone B, quadrant 4, rats injected with ryanodine spentahigherpercentageof time in zoneBcomparedwith sham(Fig. 5D).Motor activity measured as swimming speed average did not changeduring the test in control rats (sham) or after ryanodine injection(Fig. S4F). These combined results suggest that injection of stimu-latory ryanodine enhances spatial memory formation and consoli-dation in the hippocampus.
DiscussionThe results presented here show that BDNF enhanced RyR2,RyR3, and PKM! expression and promoted spine morphologychanges in primary hippocampal neurons. Functional RyR arelikely required for these changes, because inhibitory ryanodineconcentrations prevented these responses. IntrahippocampalBDNF injection or spatial memory training also increased RyR2,RyR3, and PKM! hippocampal expression; the increments pro-duced by spatial memory training possibly re"ect BDNF-inducedsignaling cascades activated during training (3, 6, 28–30).
Stimulatory Effects of BDNF in Primary Cultures. BDNF stimulatesneurotransmission via activation of Ca2+-dependent signalingcascades that promote gene transcription, synthesis of new pro-teins, and spine remodeling (7). The present results con!rm thatBDNF increases hippocampal RyR2 transcription (31). Addi-tionally, we report here that BDNF stimulated RyR3 transcrip-tion and increased RyR2, RyR3, and PKM! protein content inprimary hippocampal cultures; RyR inhibition abolished theseeffects, implicating RyR-mediated Ca2+ release in these incre-ments. In primary hippocampal neurons, BDNF-induced Ca2+signals require functional TrkB and InsP3R plus Ca2+-!lled in-tracellular stores (32, 33). In addition, TRPC3 channels havebeen proposed as novel mediators of BDNF-mediated Ca2+ sig-nal generation and dendritic remodeling (33). Our results point toRyR-mediated Ca2+ release as an additional source of Ca2+underlying BDNF-induced RyR2 and RyR3 transcription up-regulation. Through CICR, BDNF is likely to stimulate RyR-mediated Ca2+ release downstream of InsP3 receptor-mediatedCa2+ release, generating larger Ca2+ signals that modify geneexpression. Several studies have shown that BDNF enhancescalcium-dependent gene transcription (28, 29), including stimu-lation of transcription mediated by Nuclear Factor of ActivatedT Cell (NFAT) in cultured CA3–CA1 hippocampal neuronsthat involves Ca2+ release from intracellular stores (30). Ourin silico analysis of published sequences revealed that RyR2and RyR3 gene promoters contain CRE and NFAT calcium-sensitive response elements, respectively. Hence, stimulation ofRyR-mediated CICR by BDNF possibly stimulates calcium-dependent transcriptional regulators implicated in synapticplasticity, such as CREB andNFAT, which in turn would promoteRyR2 and RyR3 transcription. BDNF also increased RyR2 andRyR3 protein content; the newly expressed RyR were functional,as neurons incubated with BDNF displayed higher Ca2+ releasesignals than controls. Fast and high amplitude intracellular Ca2+signals induce hippocampal LTP, and smaller but more prolongedCa2+ signals produce long-term depression (34). Our results sug-gest that stimulation of RyR by BDNF via CICR is part of a posi-tive feedback system underlying BDNF-induced long-lastingplasticity changes.The constitutive activity of the atypical PKC isoform PKM! is
both necessary and suf!cient for hippocampal long-term memoryretention (11, 13). To our knowledge, this article is unique in
Fig. 2. BDNF enhances the expression of functional RyR proteins. (A) Con-focal images of control (I) or BDNF-treated (II) pyramidal neurons immu-nostained for RyR2 (green); the respective differential interference contrast(DIC) confocal images are shown at right. (B) Confocal images of neuronsimmunostained for RyR3 (red) and #-tubulin (green). I: Controls; II: Neuronsincubated with BDNF (50 ng/mL, 6 h). (Scale bars in A and B, 10 "m.) (C)Representative Ca2+ signals (mean ± SE, n = 3–5) elicited by 4-CMC (0.5 mM,added at arrow) displayed by control and BDNF-treated (50 ng/mL, 6 h)neurons, some of which were preincubated with ryanodine (Rya, 50 "M, 1 h).
Adasme et al. PNAS | February 15, 2011 | vol. 108 | no. 7 | 3031
NEU
ROSC
IENCE
describing stimulation of PKM! protein expression by BDNFand demonstrating that these effects require functional RyR.Enhanced PKM! synthesis is the !nal common target of manysignaling pathways involved in long-term potentiation (8); thepresent results strongly suggest that RyR-mediated Ca2+ releaseis an integral component of these pathways.The present results, which show that BDNF-induced spine
formation and growth require functional RyR, add to previousreports showing that RyR play key roles in neurite outgrowth atlater developmental stages in chick and mouse dorsal root gan-glion neurons (35). They also complement our recent workshowing that synaptotoxic amyloid # peptide oligomers, whichare implicated in impaired synaptic plasticity and memory loss in
Fig. 3. BDNF-induced spine remodeling requires RyR-mediated Ca2+ release.(A) (Upper) Time-lapsed confocal images of a neurite, visualized with calcein!uorescence, after BDNF addition (50 ng/mL); spine elongation and forma-tion of a new spine can be appreciated. (Lower) Preincubation with ryano-dine (Rya, 50 "M for 6 h) prevented BDNF-induced spine formation. (Scalebars, 2 "m.) (B) Average changes (mean ± SE) in spine length with time;controls (n = 8); exposed to BDNF (n = 10); preincubated with ryanodine (Rya50 "M, 6 h) before BDNF addition (n = 10). *P < 0.05 and ***P < 0.001,
signi"cant differences at a given time between controls and BDNF-incubatedcells; #, signi"cant differences (P < 0.05) between cells incubated with BDNFand BDNF plus ryanodine. (C) Average changes (mean ± SE) in spine numberwith time; controls (n = 5), exposed to BDNF (n = 6), preincubated withryanodine before BDNF addition (n = 5). *P < 0.05. In B and C, results wereanalyzed by two-way ANOVA plus Newman-Keuls posttest. (D) Represen-tative neurite images obtained from a control culture, after BDNF addition(50 ng/mL, 6 h), after addition of ryanodine (Rya, 50 "M, 1 h) followed byBDNF addition for 6 h, or after incubation with ryanodine (Rya) for 7 h.(Scale bars, 5 "m.) (E) Quanti"cation of spine density (mean ± SE, n = 4).Results were analyzed by one-way ANOVA followed by Newman-Keuls posttest. *P < 0.05, **P < 0.01, ***P < 0.001.
Fig. 4. BDNF injection and spatial memory training increase RyR2, RyR3,and PKM! mRNA and protein levels. (A) Western blot images and densi-tometry analysis (n ! 3) showing RyR2, RyR3, InsP3R1, PKM!, BDNF, and#-actin content in rat hippocampal tissues extracted 6 h after bilateralintrahippocampus injection with BDNF (0.25 "g per hemisphere). I, injectedwith BDNF; C, injected with saline. (B) Levels of RyR1, RyR2, RyR3, and PKM!mRNA, normalized with #-actin, determined by qRT-PCR analysis of hippo-campal tissue extracts (mean ± SE, n ! 5) from control or trained rats. (C)Western blots showing RyR2, RyR3, PKM!, and #-actin content in hippo-campal tissue from control (C) or trained (T) rats. All values (mean ± SE; n ! 3)were analyzed by Mann-Whitney test. *P < 0.05, **P < 0.01.
3032 | www.pnas.org/cgi/doi/10.1073/pnas.1013580108 Adasme et al.
Alzheimer disease, decrease RyR2 protein expression, and pre-vent RyR-dependent spine remodeling induced by BDNF inhippocampal neurons (25).
Intrahippocampal BDNF Injection and Spatial Memory TrainingEnhance RyR2, RyR3, and PKM! Expression. In accord with thestimulation of RyR2, RyR3, and PKM! expression promoted byBDNF in primary cultures, we found that intrahippocampusBDNF injection stimulated the expression of these three pro-teins within 6 h, and also increased BDNF and InsP3R1 proteinexpression. These results suggest that BDNF-induced signalingpathways mediate the RyR2, RyR3, and PKM! protein increasesinduced by spatial memory training. The RyR2 and RyR3 iso-forms seem to have a prominent role in memory processes, aswhole-brain administration of RyR2 and RyR3 but not of RyR1antisense oligonucleotides impairs associative memory (20). Twostudies using RyR3 knockout mice implicate RyR3 in spatialmemory (36) and associative memory (37); in contrast, normallearning but impaired ability to relearn a new target was reportedin these mice (38). While the three mammalian RyR isoformsare expressed in the hippocampus (39), we show here that spatialmemory training selectively increased RyR2 and RyR3 mRNAand protein expression in rat hippocampus. These !ndingsstrengthen the proposal that RyR2 and RyR3 participate in long-term spatial memory storage (20). Our results, which show thatstimulatory ryanodine concentrations present in the hippocam-pus when initiating training improved memory acquisition andconsolidation, suggest that activated RyR-mediated Ca2+ releaseunderlies the observed memory improvement.The present results are unique in showing signi!cantly enhanced
PKM! mRNA and protein expression in the hippocampus of ratstrained in a Morris water maze. PKM! is translated from a brain-
speci!c PKM! mRNA that encodes the catalytic but not the regu-latory PKC! domain (12). Moreover, PKM! protein levels increasesigni!cantly 30 min after LTP induction without concomitantchanges in mRNA expression (8, 12). The PKM! gene promotercontains a CRE response element (12). Accordingly, pathwaysmediated by Ca2+ and BDNF that promote CRE-dependent acti-vation (32, 33, 40)may contribute to enhance PKM! transcription invivo after spatial memory training. The coincident up-regulation ofRyR2, RyR3, and PKM! expression after training suggests thatthese proteins participate in common or parallel complementarypathways. As an example, PKM!maintains late-LTP by persistentlyfavoring AMPA receptor insertion into postsynaptic sites (41). Anenhanced RyR3 expression, such as that produced by BDNF andtraining, could complement this pathway because RyR3 knockoutmice display diminished LTP and decreased AMPA receptor ac-tivity but normal receptor traf!c (42).Taken together, our results support the hypothesis that RyR-
mediated CICR contributes to postsynaptic signaling mechanismsresponsible for inducing the expression of key genes for long-termmemory storage. In response to activity-dependent BDNF-mediated signaling, newly synthesized RyR could generate thestrong Ca2+ signals needed to initiate and maintain permanentchanges during long-term memory storage. Together with PKM!,the calcium-sensitive ERK/CREB pathway is critical for bothmemory consolidation and long-term neuronal plasticity pro-cesses (43) and represents an attractive downstream candidatefor the RyR-dependent signaling events produced by BDNF.
Materials and MethodsAdult male (2.5mo old) or embryonic (day 18) Sprague-Dawley rats were usedthroughout this work. Primary cultures prepared fromhippocampus dissectedat embryonic day 18 (25) were used at 14 DIV unless speci"ed otherwise. All
Fig. 5. Hippocampal ryanodine injection improves spatialmemory formation. (A) [3H]-Ryanodine (10 nmol per hemi-sphere) was administered bilaterally intrahippocampus. Ra-dioactivity was determined in different brain regions 24 hlater, and ryanodine concentrations (mean ± SE, n = 3) werecalculated from tissue weight, assuming a density of 1.0. (In-set) Photomicrograph of a representative coronal brain sec-tion showing the injection track ending in the dorsalhippocampus CA1 region; M.O.: Medulla Oblongata. (B) Rya-nodine (10 nmol per hemisphere) or sham was injected asabove 24 h before training. Escape latency values (mean ± SE)were measured daily during training; *P < 0.05, determinedby two-way ANOVA. (C) Spatial acuity, determined at trainingday 6. (D) Time spent in Quadrant 4 and Zone B respect toshams, measured at day 9 (mean ± SE; n ! 6). *P < 0.05, de-termined by unpaired Student’s t test.
Adasme et al. PNAS | February 15, 2011 | vol. 108 | no. 7 | 3033
NEU
ROSC
IENCE

protocols were approved by the Bioethics Committee, Faculty of Medicine,Universidad de Chile. Before BDNF (Millipore Corp.) addition, some cultureswere preincubated for 1 h with 20 pM recombinant human TrkB/Fc Chimera(TrkB-Fc; R&D Systems), 50 "Mryanodine (Alexis Corp.), or 1.5 "MActinomycinD (ActD; Sigma). Total RNA isolation, real-time quantitative PCR (qRT-PCR)and Western blot analysis using antibodies against RyR2 (Af"nity Biore-agents), RyR3 (Chemicon), or PKM! (Zymed) were performed as describedelsewhere (25). Ampli"cation was performed using the primers and con-ditions detailed in Table S1. Immunocytochemistry assays were carried out asdescribed in detail elsewhere (18, 25). Brain tissue "xation and immunohis-tochemistry was performed as described (44), except that coronal slices wereincubated at 4 °C overnight with anti-RyR3 antibodies. For further details onCa2+ signal detection as Fluo-4 !uorescence and morphological analysis ofdendritic spines, see SI Materials and Methods.
Water Maze Training. Rats were trained in a circular water maze for 1-minperiods three times daily for 6 consecutive days, followed by 2 d off; 1 ad-ditional day without platform was added to assess memory retention. Thehippocampus was removed 6 h after the last session at day 9. Control animalswere subjected for 4 consecutive days to three swimming trials daily with thecued platform; the platform location varied daily. At day 4, the whole hip-pocampus was removed 6 h after the last trial. To assess spatial acuity, thepool was subdivided by imaginary lines into four equal quadrants and threeequidistant concentric zones (A–C); the hidden platform was located inquadrant 4, zone B. The escape latency during each trial and the percentage
of permanence in speci"c regions of the pool was assessed with a software-coupled video camera (HVS Imagen). Further details are provided in SIMaterials and Methods.
Ryanodine and BDNF Injection. Rats under anesthesia were bilaterally injectedin the dorsal CA1 region of the hippocampus with 2 "L of 5 mM ryanodine or[3H]-ryanodine, or with 0.25 "g BDNF. Ryanodine injection was performed 96or 24 h before training; sham controls were injected with 2 "L of saline.Radioactivity was determined 24 h or 96 h after injection in different brainregions as detailed in SI Materials and Methods.
Statistical Analysis. Results are expressed as mean ± SE. Signi"cance was eval-uated with the GraphPad Software, using the unpaired Student’s t test orANOVAanalysis followedbyNewman-Keulsposttest,unless speci"edotherwise.
ACKNOWLEDGMENTS. We thank P. Fernández, N. Leal, and L. Montecinosfor their skilful professional assistance, K. Gysling and J. Hidalgo for their kindsupport and help, N. Inestrosa for providing access to a Morris water mazesystem, T. C. Sacktor and A. I. Hernández for providing PKM! antibodies, andE. Jaimovich for InsP3R1 antibodies. This work was supported by FondoNacional de Desarrollo Cientí"co y Tecnológico (FONDECYT)-Fondo de Inves-tigación Avanzada en Areas Prioritarias (FONDAP) 15010006, FONDECYT1060177 and 1100052, FONDECYT postdoctoral Grants 3070035 and3085025, Comisión Nacional de Investigación Cientí"ca y Tecnológica-FONDAP 79090021, and FONDECYT Doctoral Fellowship 24080073.
1. Alberini CM (2009) Transcription factors in long-term memory and synaptic plasticity.Physiol Rev 89:121–145.
2. Kitanishi T, Ikegaya Y, Matsuki N, Yamada MK (2009) Experience-dependent, rapidstructural changes in hippocampal pyramidal cell spines. Cereb Cortex 19:2572–2578.
3. Bekinschtein P, et al. (2008) BDNF is essential to promote persistence of long-termmemory storage. Proc Natl Acad Sci USA 105:2711–2716.
4. Tanaka J, et al. (2008) Protein synthesis and neurotrophin-dependent structuralplasticity of single dendritic spines. Science 319:1683–1687.
5. Finkbeiner S, et al. (1997) CREB: A major mediator of neuronal neurotrophinresponses. Neuron 19:1031–1047.
6. Alonso M, Medina JH, Pozzo-Miller L (2004) ERK1/2 activation is necessary for BDNF toincrease dendritic spine density in hippocampal CA1 pyramidal neurons. Learn Mem11:172–178.
7. Poo MM (2001) Neurotrophins as synaptic modulators. Nat Rev Neurosci 2:24–32.8. Kelly MT, Crary JF, Sacktor TC (2007) Regulation of protein kinase Mzeta synthesis by
multiple kinases in long-term potentiation. J Neurosci 27:3439–3444.9. Serrano P, Yao Y, Sacktor TC (2005) Persistent phosphorylation by protein kinase
Mzeta maintains late-phase long-term potentiation. J Neurosci 25:1979–1984.10. Yao Y, et al. (2008) PKM zeta maintains late long-term potentiation by N-
ethylmaleimide-sensitive factor/GluR2-dependent traf"cking of postsynaptic AMPAreceptors. J Neurosci 28:7820–7827.
11. Sacktor TC (2010) How does PKM!maintain long-term memory? Nat Rev Neurosci 12:9–15.
12. Hernandez AI, et al. (2003) Protein kinase M zeta synthesis from a brain mRNAencoding an independent protein kinase C zeta catalytic domain. Implications for themolecular mechanism of memory. J Biol Chem 278:40305–40316.
13. Pastalkova E, et al. (2006) Storage of spatial information by the maintenancemechanism of LTP. Science 313:1141–1144.
14. Greer PL, Greenberg ME (2008) From synapse to nucleus: Calcium-dependent genetranscription in the control of synapse development and function. Neuron 59:846–860.
15. Verkhratsky A (2005) Physiology and pathophysiology of the calcium store in theendoplasmic reticulum of neurons. Physiol Rev 85:201–279.
16. Bardo S, Cavazzini MG, Emptage N (2006) The role of the endoplasmic reticulum Ca2+store in the plasticity of central neurons. Trends Pharmacol Sci 27:78–84.
17. Emptage N, Bliss TV, Fine A (1999) Single synaptic events evoke NMDA receptor-mediated release of calcium from internal stores in hippocampal dendritic spines.Neuron 22:115–124.
18. Riquelme D, et al. (2011) High-frequency "eld stimulation of primary neuronsenhances ryanodine receptor-mediated Ca(2+) release and generates hydrogenperoxide, which jointly stimulate NF-$B activity. Antioxid Redox Signal, 10.1089/ars.2010.3238.
19. Korkotian E, Segal M (1999) Release of calcium from stores alters the morphology ofdendritic spines in cultured hippocampal neurons. Proc Natl Acad Sci USA 96:12068–12072.
20. Galeotti N, et al. (2008) Different involvement of type 1, 2, and 3 ryanodine receptorsin memory processes. Learn Mem 15:315–323.
21. Edwards TM, Rickard NS (2006) Pharmaco-behavioural evidence indicating a complexrole for ryanodine receptor calcium release channels in memory processing fora passive avoidance task. Neurobiol Learn Mem 86:1–8.
22. Baker KD, Edwards TM, Rickard NS (2010) A ryanodine receptor agonist promotes theconsolidation of long-term memory in young chicks. Behav Brain Res 206:143–146.
23. Lu YF, Hawkins RD (2002) Ryanodine receptors contribute to cGMP-induced late-phaseLTP and CREB phosphorylation in the hippocampus. J Neurophysiol 88:1270–1278.
24. Zhao W, et al. (2000) Spatial learning induced changes in expression of the ryanodinetype II receptor in the rat hippocampus. FASEB J 14:290–300.
25. Paula-Lima AC, et al. (2011) Amyloid beta peptide oligomers stimulate RyR-mediatedCa2+ release inducing mitochondrial fragmentation in hippocampal neurons andprevent RyR-mediated dendritic spine remodeling produced by BDNF. Antioxid RedoxSignal, 10.1089/ars.2010.3287.
26. Harvey DR, et al. (2008) Emergence of an egocentric cue guiding and allocentricinferring strategy that mirrors hippocampal brain-derived neurotrophic factor (BDNF)expression in the Morris water maze. Neurobiol Learn Mem 89:462–479.
27. Chacon MA, Barria MI, Lorca R, Huidobro-Toro JP, Inestrosa NC (2003) A human prionprotein peptide (PrP(59-91)) protects against copper neurotoxicity. Mol Psychiatry 8:853–862, 835.
28. Ying SW, et al. (2002) Brain-derived neurotrophic factor induces long-termpotentiation in intact adult hippocampus: Requirement for ERK activation coupled toCREB and upregulation of Arc synthesis. J Neurosci 22:1532–1540.
29. Zheng F, Luo Y, Wang H (2009) Regulation of brain-derived neurotrophic factor-mediated transcription of the immediate early gene Arc by intracellular calcium andcalmodulin. J Neurosci Res 87:380–392.
30. Groth RD, Mermelstein PG (2003) Brain-derived neurotrophic factor activation ofNFAT (nuclear factor of activated T-cells)-dependent transcription: A role for thetranscription factor NFATc4 in neurotrophin-mediated gene expression. J Neurosci 23:8125–8134.
31. Ring RH, et al. (2006) Transcriptional pro"ling of brain-derived-neurotrophic factor-induced neuronal plasticity: a novel role for nociceptin in hippocampal neuriteoutgrowth. J Neurobiol 66:361–377.
32. Berninger B, García DE, Inagaki N, Hahnel C, Lindholm D (1993) BDNF and NT-3 induceintracellular Ca2+ elevation in hippocampal neurones. Neuroreport 4:1303–1306.
33. Amaral MD, Pozzo-Miller L (2007) BDNF induces calcium elevations associatedwith IBDNF,a nonselective cationic current mediated by TRPC channels. J Neurophysiol 98:2476–2482.
34. Yang SN, Tang YG, Zucker RS (1999) Selective induction of LTP and LTD bypostsynaptic [Ca2+]i elevation. J Neurophysiol 81:781–787.
35. Arie Y, et al. (2009) Developmental changes in the regulation of calcium-dependentneurite outgrowth. Biochem Biophys Res Commun 379:11–15.
36. Futatsugi A, et al. (1999) Facilitation of NMDAR-independent LTP and spatial learningin mutant mice lacking ryanodine receptor type 3. Neuron 24:701–713.
37. Kouzu Y, Moriya T, Takeshima H, Yoshioka T, Shibata S (2000) Mutant mice lackingryanodine receptor type 3 exhibit de"cits of contextual fear conditioning andactivation of calcium/calmodulin-dependent protein kinase II in the hippocampus.Brain Res Mol Brain Res 76:142–150.
38. Balschun D, et al. (1999) Deletion of the ryanodine receptor type 3 (RyR3) impairsforms of synaptic plasticity and spatial learning. EMBO J 18:5264–5273.
39. Giannini G, Conti A, Mammarella S, Scrobogna M, Sorrentino V (1995) The ryanodinereceptor/calcium channel genes are widely and differentially expressed in murinebrain and peripheral tissues. J Cell Biol 128:893–904.
40. He J, Gong H, Luo Q (2005) BDNF acutely modulates synaptic transmission and calciumsignalling in developing cortical neurons. Cell Physiol Biochem 16:69–76.
41. Migues PV, et al. (2010) PKMzeta maintains memories by regulating GluR2-dependent AMPA receptor traf"cking. Nat Neurosci 13:630–634.
42. ShimutaM, et al. (2001) Postsynaptic modulation of AMPA receptor-mediated synapticresponses and LTP by the type 3 ryanodine receptor. Mol Cell Neurosci 17:921–930.
43. Sweatt JD (2004) Mitogen-activated protein kinases in synaptic plasticity andmemory. Curr Opin Neurobiol 14:311–317.
44. Haeger P, et al. (2006) Estrogen receptors alpha and beta differentially regulate thetranscriptional activity of the Urocortin gene. J Neurosci 26:4908–4916.
3034 | www.pnas.org/cgi/doi/10.1073/pnas.1013580108 Adasme et al.

Supporting InformationAdasme et al. 10.1073/pnas.1013580108SI Materials and MethodsDetermination of Intracellular Ca2+ Signals.Cells were transferred tomodi!ed Tyrode solution (in mM: 129 NaCl, 5 KCl, 2 CaCl2, 1MgCl2, 30 glucose, and 25 Hepes, pH 7.3), preloaded for 30 minat 37 °C with 5 !M Fluo-4 AM (Molecular Probes), and washedfor 10 min in modi!ed Tyrode solution to allow complete dye de-esteri!cation. Fluorescence images of intracellular Ca2+ signals inprimary hippocampal neurons [10 days in vitro (DIV)] were ob-tained every 15 s with an inverted confocal microscope [CarlZeiss; Axiovert 200, LSM 5 Pascal, Plan Apochromatic 63! Oildifferential interference contrast (DIC) objective, excitation 488nm, argon laser beam]. Image data were acquired in the cellbodies and neurites. Frame scans were averaged using theequipment data acquisition program. Ca2+ signals are presentedas F/Fo values, where Fo corresponds to the basal "uorescence, oras (F ! Fmin)/(Fmax ! F), where F corresponds to the experimental"uorescence, Fo to the basal "uorescence, Fmax to the "uores-cence of Ca+2-satured dye after addition of calcium ionophoreionomycin (100 !g/mL), and Fmin to the "uorescence signal ob-tained after Ca+2 chelation with EGTA. In all cases, the increasein intracellular [Ca2+] caused by 4-chloro-m-cresol (4-CMC),ryanodine, or tapsigargin did not saturate the probe, as indicatedby the larger "uorescence increase caused by ionomycin. All ex-periments were done at room temperature (20–22 °C).
Morphological Analysis of Dendritic Spines. Hippocampal cultures,incubated in culture medium for 6 h, were loaded for 20 min with1 !M calcein-AM in Tyrode solution. Cells in coverslips werestimulated by addition of 50 !g/mL brain-derived neurotrophicfactor (BDNF) (from a fresh stock solution), or vehicle (control).In some cases, 50 !M ryanodine was added during the 6-h in-cubation period and was maintained during calcein loading andsubsequent BDNF addition. Fluorescence and optical DIC im-ages were obtained at 15-min intervals in an inverted confocalmicroscope, as above. Randomly selected dendrites were analyzedindependently of thickness or spine density. The Image J softwareprogram (National Institutes of Health, Bethesda, MD) was usedfor image deconvolution images, to reconstruct "-projections from
7 to 15 stacks (0.4 !m each) and to measure spine length. Thenumber of spines was quanti!ed in 35-!m2
!elds.
Water Maze Training. Rats were trained in a circular water maze(1.6 m diameter, 75 cm deep). To force rats to locate the platformusing spatial memory, a hidden platform was placed in a !xedlocation in the pool during the course of training. Rats unable to!nd the platform within 1 min in the !rst trial were guided to it bythe experimenter. Rats were trained for 1-min periods three timesdaily. Training was performed during 6 consecutive days, followedby 2 d off; 1 additional day without platform was added to assessmemory retention. The hippocampus was removed, 6 h after thelast session at day 9. Control animals were subjected for 4 con-secutive days to three swimming trials daily with the cued plat-form; the platform location varied daily. At day 4, the wholehippocampus was removed 6 h after the last trial. To assess spatialacuity, the pool was subdivided by imaginary lines into four equalquadrants (1–4) and three equidistant concentric zones (A–C);the hidden platform was located in quadrant 4, zone B. Theescape latency during each trial and the percentage of perma-nence in speci!c regions of the pool was assessed with a soft-ware-coupled video camera (HVS Imagen). Space acuity wascalculated as the product of the percentage of permanence inquadrant 4 by the percentage of permanence in zone B.
Ryanodine Injection. Rats under deep chloral hydrate anesthesia(400 mg/kg body weight, i.p.) were bilaterally injected with 2 !L of5 mM ryanodine (Alexis Biochemical, dissolved in saline), usinga Hamilton syringe; sham controls were injected with 2 !L ofsaline. Injections were done in the dorsal CA1 region of thehippocampus at the coordinates: anterior, !4.3, lateral, ± 3.0;ventral 2.4 (1), 96 h or 24 h before training. Alternatively, 2 !L of5 mM [3H]-ryanodine (1.4 ! 108 cpm/pmol; PerkinElmer) wereinjected bilaterally intrahippocampus; 24 h after injection, dif-ferent brain regions were dissected, weighed, and incubated in10% SDS for 24 h. Radioactivity was determined with a PackardTri-Carb 2100TR Liquid Scintillation Analyzer.
1. Paxinos G, Watson C (1986) The Rat Brain in Stereotaxic Coordinates (Academic Press,New York), second edition.
Adasme et al. www.pnas.org/cgi/content/short/1013580108 1 of 5
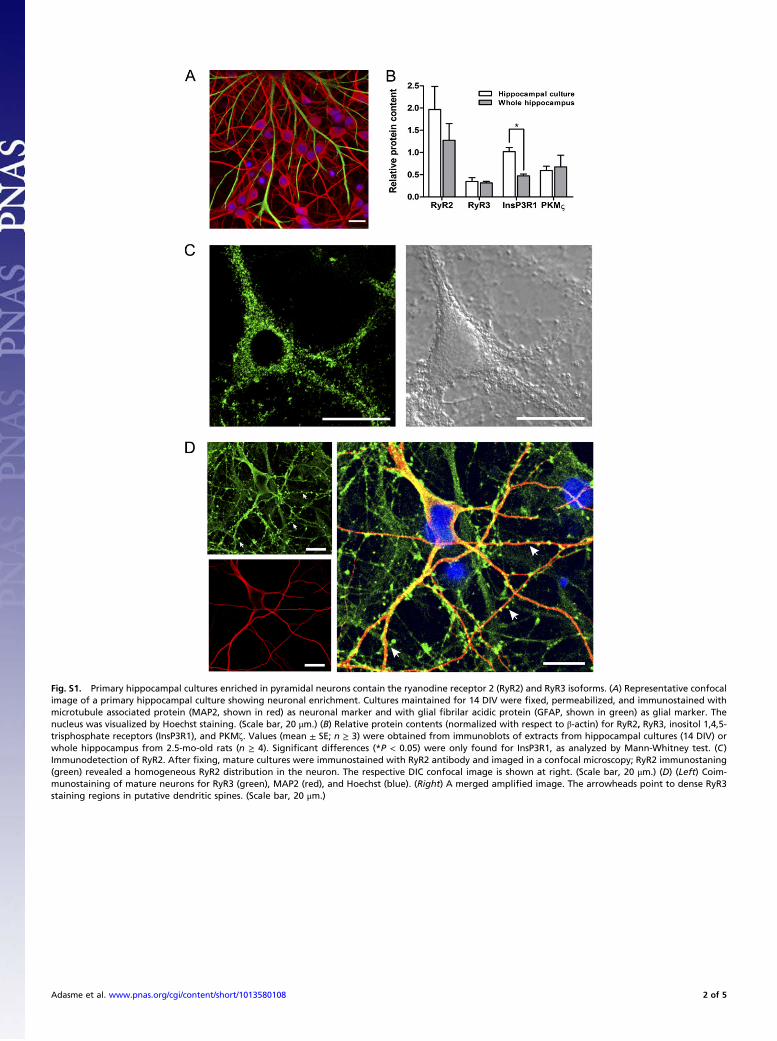
Fig. S1. Primary hippocampal cultures enriched in pyramidal neurons contain the ryanodine receptor 2 (RyR2) and RyR3 isoforms. (A) Representative confocalimage of a primary hippocampal culture showing neuronal enrichment. Cultures maintained for 14 DIV were !xed, permeabilized, and immunostained withmicrotubule associated protein (MAP2, shown in red) as neuronal marker and with glial !brilar acidic protein (GFAP, shown in green) as glial marker. Thenucleus was visualized by Hoechst staining. (Scale bar, 20 !m.) (B) Relative protein contents (normalized with respect to "-actin) for RyR2, RyR3, inositol 1,4,5-trisphosphate receptors (InsP3R1), and PKM#. Values (mean ± SE; n ! 3) were obtained from immunoblots of extracts from hippocampal cultures (14 DIV) orwhole hippocampus from 2.5-mo-old rats (n ! 4). Signi!cant differences (*P < 0.05) were only found for InsP3R1, as analyzed by Mann-Whitney test. (C)Immunodetection of RyR2. After !xing, mature cultures were immunostained with RyR2 antibody and imaged in a confocal microscopy; RyR2 immunostaning(green) revealed a homogeneous RyR2 distribution in the neuron. The respective DIC confocal image is shown at right. (Scale bar, 20 !m.) (D) (Left) Coim-munostaining of mature neurons for RyR3 (green), MAP2 (red), and Hoechst (blue). (Right) A merged ampli!ed image. The arrowheads point to dense RyR3staining regions in putative dendritic spines. (Scale bar, 20 !m.)
Adasme et al. www.pnas.org/cgi/content/short/1013580108 2 of 5
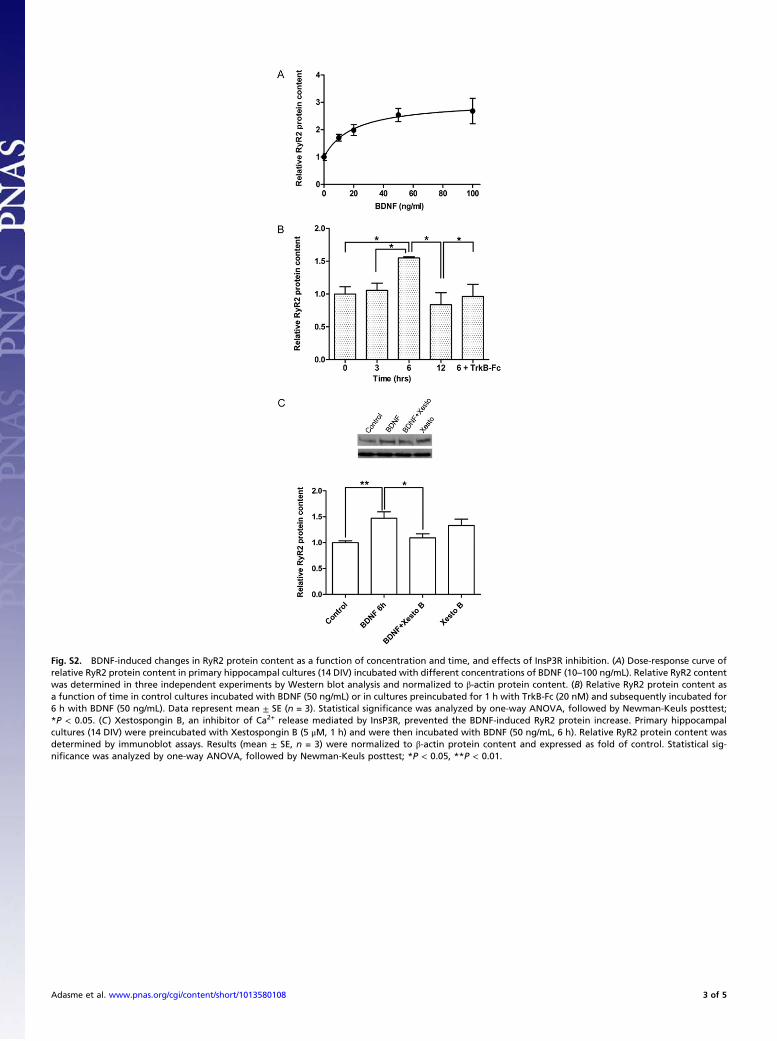
Fig. S2. BDNF-induced changes in RyR2 protein content as a function of concentration and time, and effects of InsP3R inhibition. (A) Dose-response curve ofrelative RyR2 protein content in primary hippocampal cultures (14 DIV) incubated with different concentrations of BDNF (10–100 ng/mL). Relative RyR2 contentwas determined in three independent experiments by Western blot analysis and normalized to !-actin protein content. (B) Relative RyR2 protein content asa function of time in control cultures incubated with BDNF (50 ng/mL) or in cultures preincubated for 1 h with TrkB-Fc (20 nM) and subsequently incubated for6 h with BDNF (50 ng/mL). Data represent mean ± SE (n = 3). Statistical signi!cance was analyzed by one-way ANOVA, followed by Newman-Keuls posttest;*P < 0.05. (C) Xestospongin B, an inhibitor of Ca2+ release mediated by InsP3R, prevented the BDNF-induced RyR2 protein increase. Primary hippocampalcultures (14 DIV) were preincubated with Xestospongin B (5 "M, 1 h) and were then incubated with BDNF (50 ng/mL, 6 h). Relative RyR2 protein content wasdetermined by immunoblot assays. Results (mean ± SE, n = 3) were normalized to !-actin protein content and expressed as fold of control. Statistical sig-ni!cance was analyzed by one-way ANOVA, followed by Newman-Keuls posttest; *P < 0.05, **P < 0.01.
Adasme et al. www.pnas.org/cgi/content/short/1013580108 3 of 5
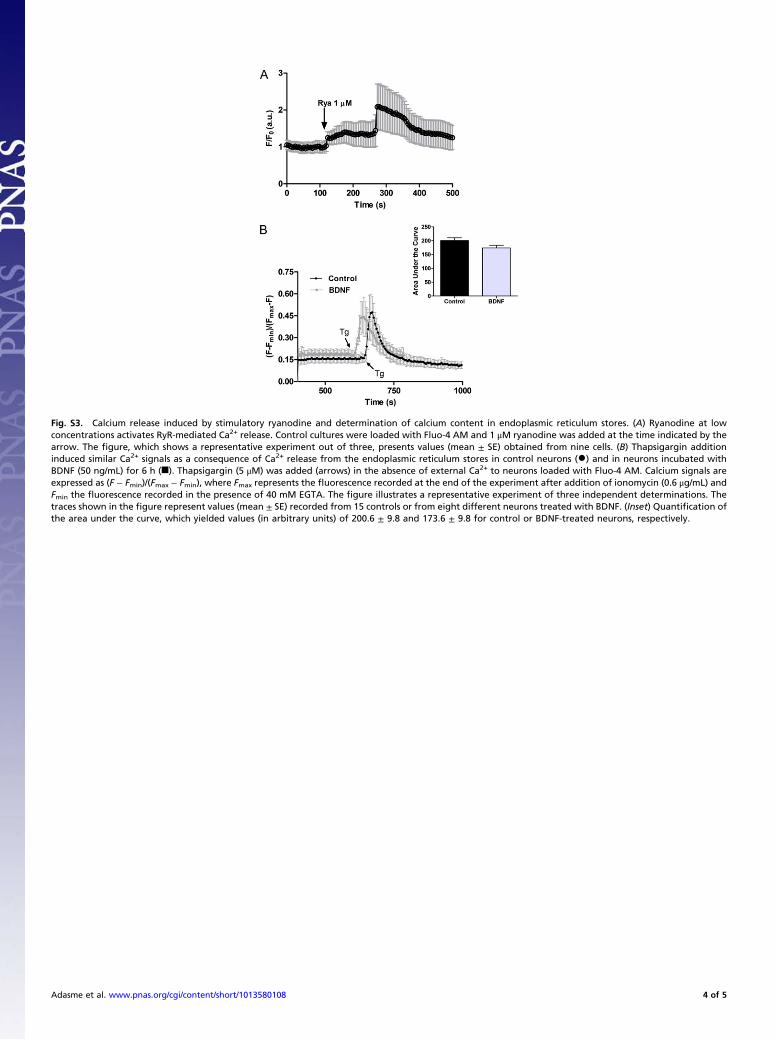
Fig. S3. Calcium release induced by stimulatory ryanodine and determination of calcium content in endoplasmic reticulum stores. (A) Ryanodine at lowconcentrations activates RyR-mediated Ca2+ release. Control cultures were loaded with Fluo-4 AM and 1 !M ryanodine was added at the time indicated by thearrow. The !gure, which shows a representative experiment out of three, presents values (mean ± SE) obtained from nine cells. (B) Thapsigargin additioninduced similar Ca2+ signals as a consequence of Ca2+ release from the endoplasmic reticulum stores in control neurons (•) and in neurons incubated withBDNF (50 ng/mL) for 6 h (!). Thapsigargin (5 !M) was added (arrows) in the absence of external Ca2+ to neurons loaded with Fluo-4 AM. Calcium signals areexpressed as (F ! Fmin)/(Fmax ! Fmin), where Fmax represents the "uorescence recorded at the end of the experiment after addition of ionomycin (0.6 !g/mL) andFmin the "uorescence recorded in the presence of 40 mM EGTA. The !gure illustrates a representative experiment of three independent determinations. Thetraces shown in the !gure represent values (mean ± SE) recorded from 15 controls or from eight different neurons treated with BDNF. (Inset) Quanti!cation ofthe area under the curve, which yielded values (in arbitrary units) of 200.6 ± 9.8 and 173.6 ± 9.8 for control or BDNF-treated neurons, respectively.
Adasme et al. www.pnas.org/cgi/content/short/1013580108 4 of 5
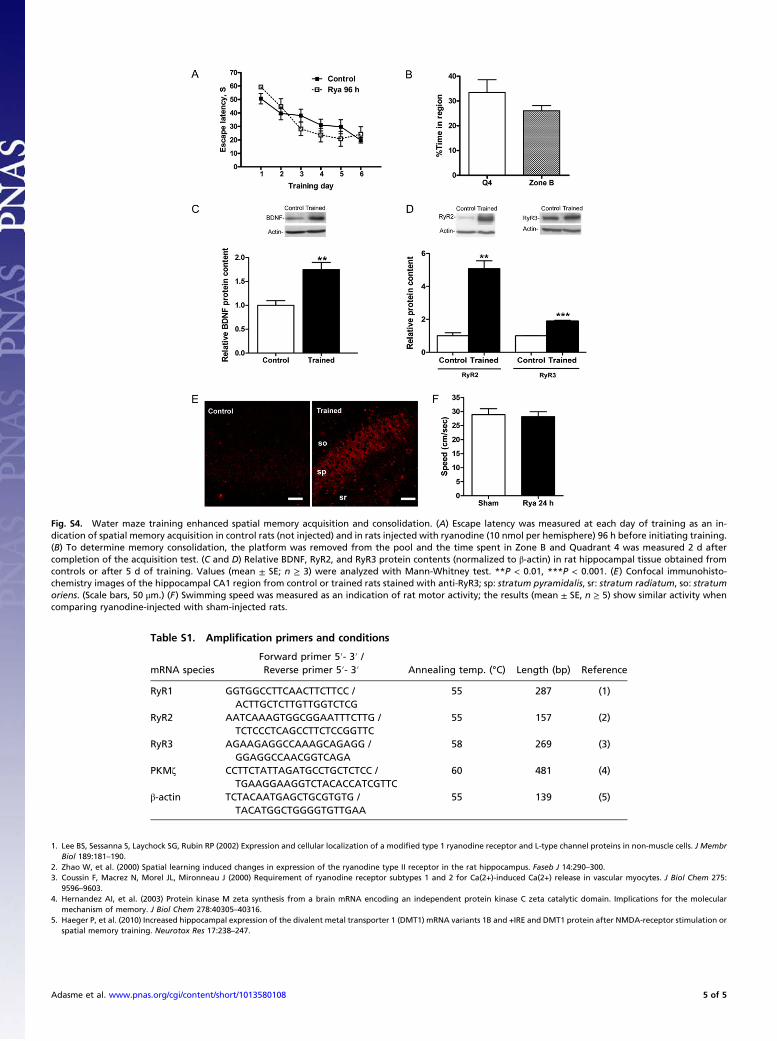
Fig. S4. Water maze training enhanced spatial memory acquisition and consolidation. (A) Escape latency was measured at each day of training as an in-dication of spatial memory acquisition in control rats (not injected) and in rats injected with ryanodine (10 nmol per hemisphere) 96 h before initiating training.(B) To determine memory consolidation, the platform was removed from the pool and the time spent in Zone B and Quadrant 4 was measured 2 d aftercompletion of the acquisition test. (C and D) Relative BDNF, RyR2, and RyR3 protein contents (normalized to !-actin) in rat hippocampal tissue obtained fromcontrols or after 5 d of training. Values (mean ± SE; n ! 3) were analyzed with Mann-Whitney test. **P < 0.01, ***P < 0.001. (E) Confocal immunohisto-chemistry images of the hippocampal CA1 region from control or trained rats stained with anti-RyR3; sp: stratum pyramidalis, sr: stratum radiatum, so: stratumoriens. (Scale bars, 50 "m.) (F) Swimming speed was measured as an indication of rat motor activity; the results (mean ± SE, n ! 5) show similar activity whencomparing ryanodine-injected with sham-injected rats.
Table S1. Ampli!cation primers and conditions
mRNA speciesForward primer 5!- 3! /
Annealing temp. (°C) Length (bp) ReferenceReverse primer 5!- 3!
RyR1 GGTGGCCTTCAACTTCTTCC / 55 287 (1)ACTTGCTCTTGTTGGTCTCG
RyR2 AATCAAAGTGGCGGAATTTCTTG / 55 157 (2)TCTCCCTCAGCCTTCTCCGGTTC
RyR3 AGAAGAGGCCAAAGCAGAGG / 58 269 (3)GGAGGCCAACGGTCAGA
PKM# CCTTCTATTAGATGCCTGCTCTCC / 60 481 (4)TGAAGGAAGGTCTACACCATCGTTC
!-actin TCTACAATGAGCTGCGTGTG / 55 139 (5)TACATGGCTGGGGTGTTGAA
1. Lee BS, Sessanna S, Laychock SG, Rubin RP (2002) Expression and cellular localization of a modi!ed type 1 ryanodine receptor and L-type channel proteins in non-muscle cells. J MembrBiol 189:181–190.
2. Zhao W, et al. (2000) Spatial learning induced changes in expression of the ryanodine type II receptor in the rat hippocampus. Faseb J 14:290–300.3. Coussin F, Macrez N, Morel JL, Mironneau J (2000) Requirement of ryanodine receptor subtypes 1 and 2 for Ca(2+)-induced Ca(2+) release in vascular myocytes. J Biol Chem 275:
9596–9603.4. Hernandez AI, et al. (2003) Protein kinase M zeta synthesis from a brain mRNA encoding an independent protein kinase C zeta catalytic domain. Implications for the molecular
mechanism of memory. J Biol Chem 278:40305–40316.5. Haeger P, et al. (2010) Increased hippocampal expression of the divalent metal transporter 1 (DMT1) mRNA variants 1B and +IRE and DMT1 protein after NMDA-receptor stimulation or
spatial memory training. Neurotox Res 17:238–247.
Adasme et al. www.pnas.org/cgi/content/short/1013580108 5 of 5
Related Documents











