51 REVIEW ISSN 1750-1911 10.2217/EPI.11.104 © 2012 Future Medicine Ltd Epigenomics (2012) 4(1), 51–65 Involvement of miRNA in erythroid differentiation miRNAs are a family of small ncRNAs that reg- ulate gene expression by targeting mRNAs in a sequence-specific manner, inducing translational repression or mRNA degradation [1–12] . miRNAs are first transcribed as primary transcripts or pri-miRNA and processed in the nucleus to short (approximately 70-nucleotides) stem–loop structures, known as pre-miRNAs. This process is carried out by a protein complex known as the microprocessor complex, includ- ing the nuclease Drosha and the dsRNA binding protein DGCR8 [1–5] . These pre-miRNAs are then processed to mature miRNAs in the cyto- plasm, following interaction with the endo- nuclease Dicer assisted by TRBP, which also initiates the formation of the RNA-induced silencing complex (RISC) [1–5] . This complex is responsible for the gene silencing. miRNA have so far been described to be involved in all the differentiation steps leading to the production of highly specialized cells. For example, miRNAs have been described to be involved in cell cycle and terminal differentiation [13] . Furthermore, miRNAs have been shown to play important roles in several other biological processes, including apoptosis and inflammation [14–16] . In this review, we analyze the available literature on the expression of miRNAs in erythroid cells. First, we describe the cellular systems employed by different groups to validate the hypothesis sup- porting the involvement of miRNAs in erythroid differentiation and helping in identifying possible mRNA targets. Second, we describe the miRNAs for which the involvement in erythroid differen- tiation has been firmly confirmed. Some methods employed by both miRNA identification and val- idation, as erythroid-specific miRNAs, will also be briefly reported. Finally, we discuss the pos- sible applications of these studies for treatment of hematological disorders. Experimental model systems to identify miRNAs involved in erythroid differentiation & their putative targets Erythroid differentiation is one of the most stud- ied pathway, and studies on this specific topic are based on a large variety of in vitro and in vivo experimental model systems. Whilst the in vitro systems are flexible and allow treatment with inducers/inhibitors of specific steps along the ery- throid program, in many cases they do not fully ricapitulate the erythroid program. Therefore, in vivo systems are appealing to sustain the conclusions reached by in vitro studies. Human embryonic stem cells cultured in experimental conditions promoting hematopoietic induction Human embryonic stem cells (hESCs) have been extensively used in the recent past to study the early phases of development [17] . The major steps in the miRNAs are a family of small ncRNAs that regulate gene expression by targeting mRNAs in a sequence-specific manner, inducing translational repression or mRNA degradation. In this review, we present and discuss the available literature on the expression of miRNAs in erythroid cells. There are several experimental systems that can be employed for studies focusing on the relationship between miRNAs and erythroid differentiation, including human embryonic stem cells forced to erythroid differentiation, K562 and UT-7 cells induced to hemoglobin production by chemical compounds, erythropoietin-treated erythroid precursor cells from normal subjects or patients affected by hematological disease and in vivo systems, such as zebrafish embryos. Several miRNAs were identified as deeply involved in the erythroid phenotype, including miR-15a, miR-16–1, miR-126, miR-144, miR-451 and miR-210. Several functions related with erythroid cells were demonstrated to be regulated by these miRNAs, including maturation and proliferation of early erythroid cells, expression of fetal g-globin genes and enucleation. These identified erythroid specific miRNAs represent the starting point to develop new protocols for miRNA therapeutics, based on both anti-miR molecules or miRNA replacement. KEYWORDS: b-thalassemia erythroid differentiation globin genes HbF miRNA Nicolea Bianchi 1 , Crisna Zuccato 1,2 , Alessia Fino 1 , Ilaria Lampron 1 , Monica Borga 1 & Roberto Gambari* 1,3 1 BioPharmaNet, Department of Biochemistry & Molecular Biology, University of Ferrara, via Fossato di Mortara, 74-44121 Ferrara, Italy 2 ThalLab, Associazione Veneta per la Loa alla Talassemia, Rovigo, Italy 3 Biotechnology Center, University of Ferrara, via Fossato di Mortara, 64-44121 Ferrara, Italy *Author for correspondence: [email protected] part of For reprint orders, please contact: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

51
Review
ISSN 1750-191110.2217/EPI.11.104 © 2012 Future Medicine Ltd Epigenomics (2012) 4(1), 51–65
Involvement of miRNA in erythroid differentiation
miRNAs are a family of small ncRNAs that reg-ulate gene expression by targeting mRNAs in a sequence-specific manner, inducing translational repression or mRNA degradation [1–12].
miRNAs are first transcribed as primary transcripts or pri-miRNA and processed in the nucleus to short (approximately 70-nucleotides) stem–loop structures, known as pre- miRNAs. This process is carried out by a protein complex known as the microprocessor complex, includ-ing the nuclease Drosha and the dsRNA binding protein DGCR8 [1–5]. These pre-miRNAs are then processed to mature miRNAs in the cyto-plasm, following inter action with the endo-nuclease Dicer assisted by TRBP, which also initiates the formation of the RNA-induced silencing complex (RISC) [1–5]. This complex is responsible for the gene silencing. miRNA have so far been described to be involved in all the differentiation steps leading to the production of highly specialized cells. For example, miRNAs have been described to be involved in cell cycle and terminal differentiation [13]. Furthermore, miRNAs have been shown to play important roles in several other biological processes, including apoptosis and inflammation [14–16].
In this review, we analyze the available literature on the expression of miRNAs in erythroid cells. First, we describe the cellular systems employed by different groups to validate the hypothesis sup-porting the involvement of m iRNAs in erythroid
differentiation and helping in identifying possible mRNA targets. Second, we describe the miRNAs for which the involvement in erythroid differen-tiation has been firmly confirmed. Some methods employed by both miRNA identification and val-idation, as erythroid-specific miRNAs, will also be briefly reported. Finally, we discuss the pos-sible applications of these studies for t reatment of hematological disorders.
Experimental model systems to identify miRNAs involved in erythroid differentiation & their putative targetsErythroid differentiation is one of the most stud-ied pathway, and studies on this specific topic are based on a large variety of in vitro and in vivo experimental model systems. Whilst the in vitro systems are flexible and allow treatment with inducers/inhibitors of specific steps along the ery-throid program, in many cases they do not fully ricapitulate the erythroid program. Therefore, in vivo systems are appealing to sustain the conclusions reached by in vitro studies.
� Human embryonic stem cells cultured in experimental conditions promoting hematopoietic inductionHuman embryonic stem cells (hESCs) have been extensively used in the recent past to study the early phases of development [17]. The major steps in the
miRNAs are a family of small ncRNAs that regulate gene expression by targeting mRNAs in a sequence-specific manner, inducing translational repression or mRNA degradation. In this review, we present and discuss the available literature on the expression of miRNAs in erythroid cells. There are several experimental systems that can be employed for studies focusing on the relationship between miRNAs and erythroid differentiation, including human embryonic stem cells forced to erythroid differentiation, K562 and UT-7 cells induced to hemoglobin production by chemical compounds, erythropoietin-treated erythroid precursor cells from normal subjects or patients affected by hematological disease and in vivo systems, such as zebrafish embryos. Several miRNAs were identified as deeply involved in the erythroid phenotype, including miR-15a, miR-16–1, miR-126, miR-144, miR-451 and miR-210. Several functions related with erythroid cells were demonstrated to be regulated by these miRNAs, including maturation and proliferation of early erythroid cells, expression of fetal g-globin genes and enucleation. These identified erythroid specific miRNAs represent the starting point to develop new protocols for miRNA therapeutics, based on both anti-miR molecules or miRNA replacement.
KEYWORDS: b-thalassemia � erythroid differentiation � globin genes � HbF � miRNA Nicoletta Bianchi1, Cristina Zuccato1,2, Alessia Finotti1, Ilaria Lampronti1, Monica Borgatti1 & Roberto Gambari*1,3
1BioPharmaNet, Department of Biochemistry & Molecular Biology, University of Ferrara, via Fossato di Mortara, 74-44121 Ferrara, Italy 2ThalLab, Associazione Veneta per la Lotta alla Talassemia, Rovigo, Italy 3Biotechnology Center, University of Ferrara, via Fossato di Mortara, 64-44121 Ferrara, Italy *Author for correspondence: [email protected]
part of
For reprint orders, please contact: [email protected]
Epigenomics (2012) 4(1)52 future science group
Review Bianchi, Zuccato, Finotti, Lampronti, Borgatti & Gambari
generation of pluripotent hESC are the following: after 5–7 days of in vitro cell culture, a blastocyst is generated showing a clearly visible and easily acces-sible inner cell mass (ICM). Pluripotent stem cells can be easily isolated from the ICM, giving rise to in vitro hESC lines; from these cell lines embryoid bodies (EBs) can be developed to be employed for further tissue-specific differentiation.
For erythroid induction, Huang et al. used hESC lines maintained as undifferentiated cells by weekly passage with collagenase IV onto g-irradiated mouse embryonic fibroblast feeder layers [18]. To generate EBs, hESCs were incu-bated with dispase, colonies were collected and plated in 6-well ultra-low attachment plates in EB differentiation medium. Using the embryonic stem cells system, the miRNA expression profile can be analyzed in undifferentiated hESCs, in EBs and in erythroid differentiated cells.
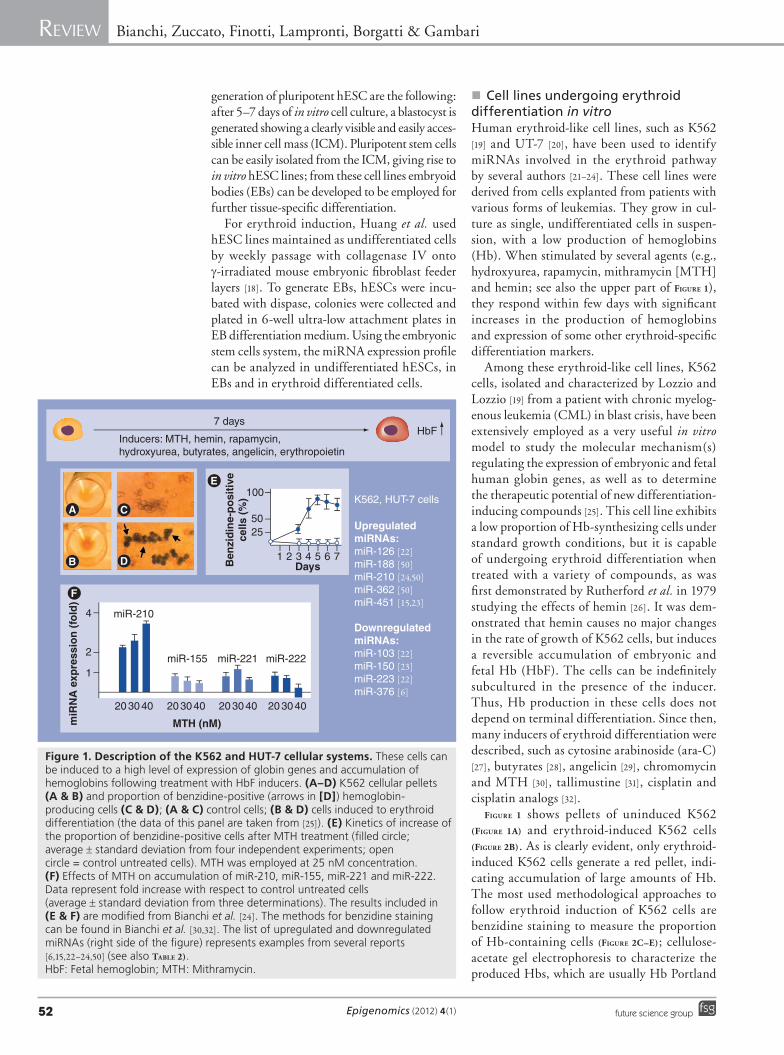
� Cell lines undergoing erythroid differentiation in vitroHuman erythroid-like cell lines, such as K562 [19] and UT-7 [20], have been used to identify miRNAs involved in the erythroid pathway by several authors [21–24]. These cell lines were derived from cells explanted from patients with various forms of leukemias. They grow in cul-ture as single, undifferentiated cells in suspen-sion, with a low production of hemoglobins (Hb). When stimulated by several agents (e.g., hydroxyurea, rapamycin, mithramycin [MTH] and hemin; see also the upper part of Figure 1), they respond within few days with significant increases in the production of hemoglobins and expression of some other erythroid-specific differentiation markers.
Among these erythroid-like cell lines, K562 cells, isolated and characterized by Lozzio and Lozzio [19] from a patient with chronic myelog-enous leukemia (CML) in blast crisis, have been extensively employed as a very useful in vitro model to study the molecular mechanism(s) regulating the expression of embryonic and fetal human globin genes, as well as to determine the therapeutic potential of new differentiation-inducing compounds [25]. This cell line exhibits a low proportion of Hb-synthesizing cells under standard growth conditions, but it is capable of undergoing erythroid differentiation when treated with a variety of compounds, as was first demonstrated by Rutherford et al. in 1979 studying the effects of hemin [26]. It was dem-onstrated that hemin causes no major changes in the rate of growth of K562 cells, but induces a reversible accumulation of embryonic and fetal Hb (HbF). The cells can be indefinitely subcultured in the presence of the inducer. Thus, Hb production in these cells does not depend on terminal differentiation. Since then, many inducers of erythroid differentiation were described, such as cytosine arabinoside (ara-C) [27], butyrates [28], angelicin [29], chromomycin and MTH [30], tallimustine [31], cisplatin and cisplatin analogs [32].
Figure 1 shows pellets of uninduced K562 (Figure 1A) and erythroid-induced K562 cells (Figure 2B). As is clearly evident, only erythroid-induced K562 cells generate a red pellet, indi-cating accumulation of large amounts of Hb. The most used methodological approaches to follow erythroid induction of K562 cells are benzidine staining to measure the proportion of Hb-containing cells (Figure 2C–e); cellulose-acetate gel electrophoresis to characterize the produced Hbs, which are usually Hb Portland
K562, HUT-7 cells
7 days
Inducers: MTH, hemin, rapamycin,hydroxyurea, butyrates, angelicin, erythropoietin
Upregulated miRNAs:miR-126 [22]miR-188 [50]miR-210 [24,50]miR-362 [50]miR-451 [15,23]
Downregulated miRNAs:miR-103 [22]miR-150 [23]miR-223 [22]miR-376 [6]
1
2550
100
2 3 4 5 6 7DaysB
enzi
din
e-p
osi
tive
cells
(%
)
miR-222miR-221miR-155
miR-210
20
1
2
4
miR
NA
exp
ress
ion
(fo
ld)
3040 203040 203040 203040
MTH (nM)
HbF
Figure 1. Description of the K562 and HUT-7 cellular systems. These cells can be induced to a high level of expression of globin genes and accumulation of hemoglobins following treatment with HbF inducers. (A–D) K562 cellular pellets (A & B) and proportion of benzidine-positive (arrows in [D]) hemoglobin-producing cells (C & D); (A & C) control cells; (B & D) cells induced to erythroid differentiation (the data of this panel are taken from [25]). (E) Kinetics of increase of the proportion of benzidine-positive cells after MTH treatment (filled circle; average ± standard deviation from four independent experiments; open circle = control untreated cells). MTH was employed at 25 nM concentration. (F) Effects of MTH on accumulation of miR-210, miR-155, miR-221 and miR-222. Data represent fold increase with respect to control untreated cells (average ± standard deviation from three determinations). The results included in (E & F) are modified from Bianchi et al. [24]. The methods for benzidine staining can be found in Bianchi et al. [30,32]. The list of upregulated and downregulated miRNAs (right side of the figure) represents examples from several reports [6,15,22–24,50] (see also TABle 2).HbF: Fetal hemoglobin; MTH: Mithramycin.

www.futuremedicine.com 53future science group
miRNA & erythroid differentiation Review
(z2g
2), Hb Gower 1 (z
2e
2) and HbF (a
2g
2);
northern blotting and real-time (RT)-PCR analyses to determine the effects of erythroid inducers on the expression of globin genes [25]. These data are taken from research and review articles recently published by our group [24,30–32]. These assays demonstrate high expression of the human g-globin genes following induction to erythroid differentiation. The low expression of HbF with respect to the embryonic-type hemo-globins Gower 1 and Portland is caused by an a-thalassemia-like phenotype [33].
The human pluripotent UT-7 cell line is growth factor-dependent for proliferation and differentiation, and able to undergo the expres-sion of the erythroid phenotype when cultured in the presence of erythropoietin (EPO) [20].
The use of K562 and HUT-7 cell systems allows the comparison of the miRNA pattern in two different conditions: in their undifferenti-ated state (low proportion of globin gene expres-sion and HbF synthesis and following induc-tion to erythroid differentiation with inducers (Figure 1).
� Erythroid progenitors from peripheral & cord bloodIt is possible to obtain large cultures of rela-tively pure and synchronized erythroid cell population and compounds can be added on different days when the culture consists of cells at specific stages of maturation. In a procedure developed by Fibach et al. [34,35], the culture is divided into two phases: an EPO-independent
Figure 2. The erythroid precursor cell experimental system. These cells are isolated from peripheral blood of normal subjects or patients affected by hematological diseases, following the procedure developed by Fibach et al. [35]. These cells (A), following treatment with EPO, undergo eythroid differentiation, (B) originating red pellets and (C–F) expressing different levels of HbF. (C–E) Represent the HPLC profile derived from differentiated ErPCs from (C) a normal subject, (D) a b-thalassemic patient expressing high HbF levels and (E) a b-thalassemic patient expressing low HbF levels. When treated with HbF, inducers of ErPCs increase HbF production (compare [F] with [E]). (A & B) Modified from Gambari and Fibach [25]; methods describing HPLC and HPLC profiles similar to the representative profiles included in (C–E) can be found in Lampronti et al. [29] and Fibach et al. [35]. (G) Microarray ana lysis performed using RNA from ErPCs isolated from a US, Th1 (producing low HbF levels) and Th17 (producing high HbF levels). Heatmap of 194 miRNA expression profiles found expressed in at least one sample is depicted using a color-bar approach. The pattern of Th17 is different from both US and Th1 miRNA profiles. (H) Restricted ana lysis of expression of 30 miRNAs with similar production in Th1 and US samples (Th1/US = 0.09–1.1). Arrow indicates miR-210, which is significantly much more represented in Th17 cells. Results depicted in (G & H) are modified from Bianchi et al. [24].AU: Absorbance units; EPO: Erythropoietin; ErPC: Erythroid precursor cell; HbA: Hemoglobin A; HbA2: Hemoglobin A2; HbF: Fetal hemoglobin; Th: Thalassemic patient; US: Unaffected subject.

Epigenomics (2012) 4(1)54 future science group
Review Bianchi, Zuccato, Finotti, Lampronti, Borgatti & Gambari
phase, in which peripheral blood cells are first cultured in the presence of a combination of growth factors, but in the absence of EPO. In the second phase, the culture (Figure 2A), supple-mented with EPO, generates orthochromatic normoblasts and enucleated erythrocytes [35]. As these cells differentiate, they decrease in size and accumulate Hb, the aggregates assume a reddish color and the cellular pellets are brown–red in color (Figure 2B).
Peripheral blood cells are employed in this procedure for the following reasons: the avail-ability of blood from normal individuals and patients [25]; the homogeneity of the periph-eral blood erythroid progenitors, namely early burst-forming unit-erythroid, as opposed to the bone marrow which contains progenitors at various developmental stages [34,35]. Good results can be obtained with cells derived from other sources, including CD34+ cells purified by immuno-magnetic bead technologies, but in these cases, some modifications of the pro-cedure are required [25,34]. Unlike K562 cells and other experimental model systems based on leukemic cell lines, this system faithfully recapitulates many aspects of in vivo erythro-poiesis including globin RNA metabolism, cell cycle kinetics activated during terminal stages of differentiation, expression of cell surface antigens, iron and ferritin metabolism, and production of erythroid-specific t ranscription factors (TFs) [35].
Therefore, several research groups have used this system to study the effects of hundreds of compounds, including butyroids, hemin, EPO and histone deacetylase inhibitors [36–39]. In order to study their potential to enhance HbF production, compounds can be added to phase I, phase II or both. Nontoxic drugs, such as cytokines and hemin, may be added to the cultures at any time [34]. With cytotoxic drugs, such as hydroxyurea (HU) and 5-aza-cytidine, because of their cytotoxic/cytostatic effects, they are u sually added on day 4–8 of phase II [25,34].
Since the erythroid cells in phase II are grown in suspension, samples of cells can be withdrawn at any time without disturbing the cultures and assayed for morphology, size, number, cell via-bility and apoptosis, cell cycle or expression of surface antigens.
Hemoglobinization can be easily followed by staining the cells with benzidine. The Hb content of the developing erythroid cells can be measured by a variety of techniques, such as alkaline denaturation, benzidine staining,
HPLC, cation-exchange HPLC for hemo globins and reverse-phase HPLC for globin chains.
Figure 2C–F shows representative HPLC profiles of erythroid precursor cells (ErPCs) lysates iso-lated from a normal subject (Figure 2C), ab-thal-assemia patient expressing high levels of HbF (Figure 2D) and a b-thalassemia patient expressing low levels of HbF (Figure 2e & F; Figure 2A shows uninduced ErPC cultures; Figure 2B shows ErPC cultures induced with MTH).
� Zebrafish as an in vivo model systemThe zebrafish experimental system offers several advantages over mice for the study of miRNA function, particularly in developmental pro-cesses. In fact, in zebrafish, loss- and gain-of-function studies can be undertaken quickly using transient genetic approaches: overexpres-sion by microinjecting a synthetic RNA duplex and miRNA knockdown by injecting antagonist oligonucleotides [40–49].
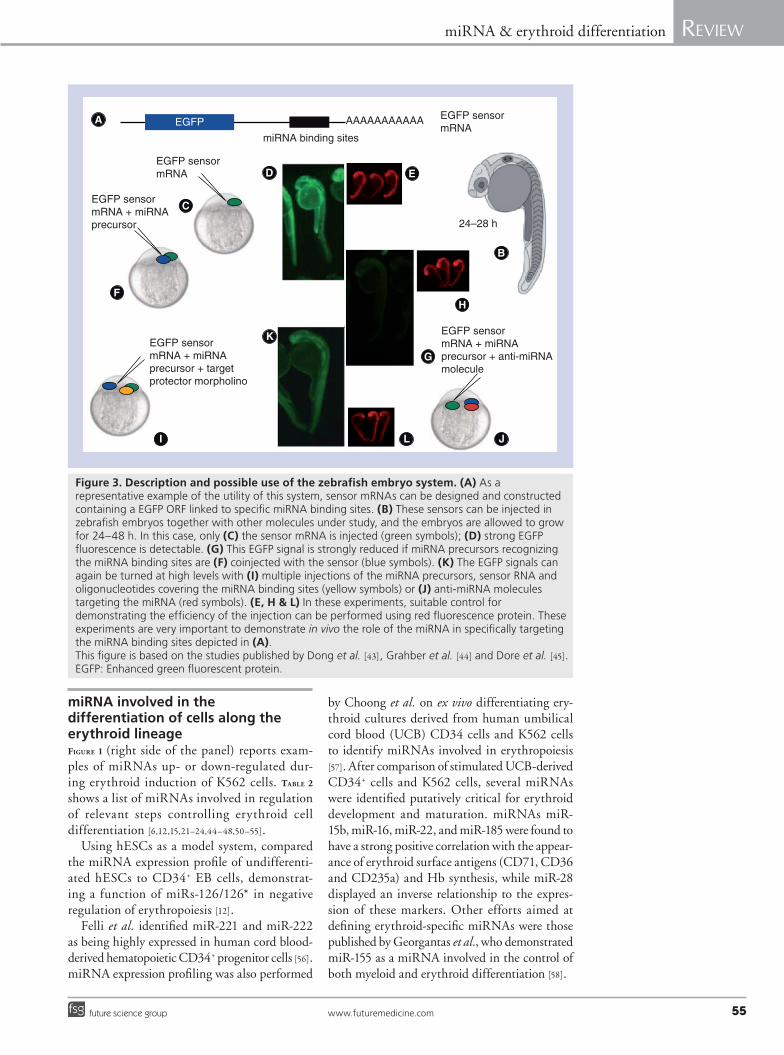
Conversely, predicted miRNA/transcript interactions can be evaluated using fluorescent reporter assays (Figure 3). For example, reporter constructs can be generated producing RNA sequences coding the enhanced green fluores-cence protein (EGFP) linked to stetches con-taining putative signals for miRNAs [43–45]. Microinjections of these contructs or correspond-ing RNAs into zebrafish embryos lead to the development of 24–48 embryos expressing a high level of EGFP (Figure 3A–e). Interactions between endogenous miRNAs and their targets can be specifically interrupted by target-blocking oligo-nucleotides competing with miRNA in binding to miRNA-binding sites (Figure 3i, K & l) [44] or anti-miRNA (Figure 3J–l) [45], allowing the reach-ment of important conclusions about the role of the specific miRNAs in targeting the putative miRNA stretches linked to the EGFP sequences [43–45]. Because these approaches are transient, the efficacy and specificity of reagents can quickly and efficiently be rigorously evaluated in vivo in a manner not possible in mice.
Moreover, miRNA biogenesis mutants can be produced and employed to study the role of miRNAs in development. This was the case of the zebrafish mutant Dicer, which was employed for elucidating the role of miR-430 in early development [41]. Finally, mutants selectively deficient in a particular miRNA but retaining the cell type in which it is expressed are particu-larly useful. TABle 1 shows reported examples on the usefulness of the zebrafish embryo system to evaluate the role of miRNAs in erythroid functions [43–49].
www.futuremedicine.com 55future science group
miRNA & erythroid differentiation Review
miRNA involved in the differentiation of cells along the erythroid lineageFigure 1 (right side of the panel) reports exam-ples of miRNAs up- or down-regulated dur-ing erythroid induction of K562 cells. TABle 2 shows a list of miRNAs involved in regulation of relevant steps controlling erythroid cell differentiation [6,12,15,21–24,44–48,50–55].
Using hESCs as a model system, compared the miRNA expression profile of undifferenti-ated hESCs to CD34+ EB cells, demonstrat-ing a function of miRs-126/126* in negative r egulation of erythropoiesis [12].
Felli et al. identified miR-221 and miR-222 as being highly expressed in human cord blood-derived hematopoietic CD34+ progenitor cells [56]. miRNA expression profiling was also performed
by Choong et al. on ex vivo differentiating ery-throid cultures derived from human umbilical cord blood (UCB) CD34 cells and K562 cells to identify miRNAs involved in erythropoiesis [57]. After comparison of stimulated UCB-derived CD34+ cells and K562 cells, several miRNAs were identified putatively critical for erythroid development and maturation. miRNAs miR-15b, miR-16, miR-22, and miR-185 were found to have a strong positive correlation with the appear-ance of erythroid surface antigens (CD71, CD36 and CD235a) and Hb synthesis, while miR-28 displayed an inverse relationship to the expres-sion of these markers. Other efforts aimed at defining erythroid-specific miRNAs were those published by Georgantas et al., who demonstrated miR-155 as a miRNA involved in the control of both myeloid and erythroid differentiation [58].
EGFP sensormRNA + miRNAprecursor
EGFP sensormRNA + miRNAprecursor + targetprotector morpholino
EGFP sensormRNA + miRNAprecursor + anti-miRNAmolecule
EGFP sensormRNA
EGFP sensormRNA
24–28 h
miRNA binding sites
I J
K
L
AAAAAAAAAAAEGFP
Figure 3. Description and possible use of the zebrafish embryo system. (A) As a representative example of the utility of this system, sensor mRNAs can be designed and constructed containing a EGFP ORF linked to specific miRNA binding sites. (B) These sensors can be injected in zebrafish embryos together with other molecules under study, and the embryos are allowed to grow for 24–48 h. In this case, only (C) the sensor mRNA is injected (green symbols); (D) strong EGFP fluorescence is detectable. (G) This EGFP signal is strongly reduced if miRNA precursors recognizing the miRNA binding sites are (F) coinjected with the sensor (blue symbols). (K) The EGFP signals can again be turned at high levels with (I) multiple injections of the miRNA precursors, sensor RNA and oligonucleotides covering the miRNA binding sites (yellow symbols) or (J) anti-miRNA molecules targeting the miRNA (red symbols). (E, H & L) In these experiments, suitable control for demonstrating the efficiency of the injection can be performed using red fluorescence protein. These experiments are very important to demonstrate in vivo the role of the miRNA in specifically targeting the miRNA binding sites depicted in (A). This figure is based on the studies published by Dong et al. [43], Grahber et al. [44] and Dore et al. [45].EGFP: Enhanced green fluorescent protein.

Epigenomics (2012) 4(1)56 future science group
Review Bianchi, Zuccato, Finotti, Lampronti, Borgatti & Gambari
Using zebraf ish embryos as experimen-tal system, Grabher et al. demonstrated that miR-126 is a novel physiological regulator of the proto-oncogene c-Myb during definitive
hematopoiesis. This conclusion was achieved following knockdown of miR-126, leading to increasing c-Myb levels and promoting erythropoiesis in vivo [44].
Table 1. Zebrafish and in vivo studies on miRNAs and erythroid differentiation.
Author (year) Experimental model system
miRNA Conclusion Possible applications Ref.
Dong et al. (2009)
Microinjection of miRNAs into zebrafish embryos
miR-30a The miRNA–shRNA, when microinjected into zebrafish embryos, induced an efficient knockdown of two developmentally essential genes, chordin and a-catenin, in a dose-controllable fashion
This methodology provides an invaluable tool to knockdown developmentally important genes in a tissue-specific manner or to establish animal models in which the gene dosage is critically important in the pathogenesis of human disorders
[43]
Grabher et al. (2011)
Knockdown of miRNA following injections of anti-miRNA
miR-126 miR-126 knockdown leads to increased c-Myb levels and promotes erythropoiesis in vivo
These results support the notion that miRNAs not only function to provide precision to developmental programs but are also essential determinants in the control of variable potential functions of a single gene during hematopoiesis
[44]
Yu et al. (2010) miRNA-/- zebrafish erythroblasts
miR-144miR-451
In miR-144/451-/- erythroblasts, 14-3-3 z accumulates, causing partial relocalization of FOXO3 from the nucleus to the cytoplasm with dampening of its transcriptional program, including antioxidant-encoding genes Cat and Gpx1
This strategy allows for defining novel miRNA-regulated pathways that protect erythrocytes against oxidant stress
[48]
Du et al. (2009) Transient reporter assay in vivo
miR-144 Although hmbsb, miR-451, KLFd and miR-144 are all coexpressed in the developing erythroid progenitors during zebrafish erythropoiesis, only KLFd can be validated as a bona fide physiological target of miR-144
The results address the importance of using multiple approaches combined with western blot ana lysis to validate the physiological target of a given miRNA
[46]
Fu et al. (2009) Zebrafish embryogenesis
miR-144 miR-144 negatively regulates the embryonic a-globin, but not embryonic b-globin gene expression, through physiologically targeting KLFd
The selective effect of the miR-144–KLFd pathway on globin gene regulation may constitute a novel therapeutic target for improving the clinical outcome of patients with thalassemia
[49]
Pase et al. (2009)
mnr miR-144 miR-451
Although erythropoiesis initiated normally in mnr, erythrocyte maturation was morphologically retarded. Morpholino knockdown of miR-451 increased erythrocyte immaturity in wild-type embryos, and miR-451 RNA duplexes partially rescued erythroid maturation in mnr, demonstrating a requirement and role for miR-451 in erythrocyte maturation
These studies define a new genetic pathway promoting erythroid maturation (mnr/miR-451/ GATA-2) and provide a rare example of partial rescue of a mutant phenotype solely by miRNA overexpression
[47]
Dore et al. (2008)
Gene complementation strategy combined with microarray screening
miR-144 miR-451
Zebrafish embryos depleted of miR-451 by using antisense morpholinos form erythroid precursors, but their development into mature circulating red blood cells is strongly and specifically impaired
These results reveal a miRNA locus that is required for erythropoiesis and uncover a new regulatory axis through which GATA-1 controls this process
[45]
Myb: v-myb myeloblastosis viral oncogene homolog transcription factor.

www.futuremedicine.com 57future science group
miRNA & erythroid differentiation Review
Table 2. miRNAs involved in regulating gene expression in erythroid cells.
Author (year)
miRNA Target Cellular system
Biological effects Possible applications Ref.
Sankaran et al. (2011)
miR-15a, miR-16-1
Myb Human trisomy 13
Increase of HbF production Therapy of sickle cell disease and b-thalassemia
[51]
Yang et al. (2009)
miR-103 FOXJ2 K562 cell line Inhibition of hemin-induced K562 erythroid differentiation
Studies on molecular mechanism regulating human erythropoiesis
[22]
Huang et al. (2010)
miR-126/126* PTPN9 hESCs Negative regulator of erythroid development
Studies on hematopoietic lineage
[12]
Grabher et al. (2011)
miR-126/126* Myb Zebrafish Control of the cell fate decision between erythroid and megakaryocytic lineages during definitive hematopoiesis
Studies on developmental programs during hematopoiesis
[44]
Sangokoya et al. (2010)
miR-144 NRF2 K562 cell line and primary erythroid progenitor cells
Interference with antioxidant capacity; susceptibility to oxidative stress, hemolysis and severe anemia
Studies on clinicalheterogeneity and pathobiology ofsickle cell disease
[21]
Kosaka et al. (2008)
miR-188, miR-210, miR-362
To be identified
K562 cell line Increased following erythroid differentiation
Studies on molecular mechanisms regulating erythroid differentiation
[50]
Zhang et al. (2011)
miR-191 Riok3, Mxi1
Mouse CFU-E erythroid progenitors
Block of erythroblast enucleation Studies on terminal erythroid differentiation
[54]
Bianchi et al. (2009)
miR-210 To be identified
K562 cell line Increased following erythroid differentiation
Studies on molecular mechanism linking hypoxia to differentiation
[24]
Gabbianelli et al. (2010)
miR-221,miR-222
c-kit CD34+ cells from pre-, mid- and full-term cord blood
Decrease of erythroblast proliferation and HbF content
Studies on peri/post-natal Hb switch with implications for therapy of b-thalassemia based on HbF reactivation
[52]
Felli et al. (2009)
miR-223 LMO2 Cord blood human progenitor cells
Decline of miR-223 induces expansion of erythroblast cells
Studies on mechanism involved in adult erythropoiesis
[53]
Wang et al. (2011)
miR-376 To be identified
K562 cell line Decreased following erythroid differentiation
Induction of differentiation using anti-miRNA
[6]
Bruchova-Votavova et al. (2010)
miR-451 UBE2H, ARPP-19
K562 cell line Induction of erythroid maturation and differentiation
Studies on abnormal erythropoiesis [23]
Svasti et al. (2010)
miR-451 GATA-1/GATA-2
Erythroid progenitors
Alteration of erythropoiesis Studies on dysregulation of expression program in b-thalassemia
[55]
Dore et al. (2008)
miR-451 GATA-1 Erythroblast line G1E-ER4
Essential for erythropoiesis Studies on red blood cell production [45]
Du et al. (2009)
miR-451 hmbsb Zebrafish Maturation of erythroid progenitors during embryogenesis and definitive erythropoiesis
Studies on identification of miRNA targets during primitive erythropoiesis
[46]
Pase et al. (2009)
miR-451 GATA-2 Zebrafish Modulation of rate of erythrocyte maturation
Studies on erythrocyte maturation [47]
Yu et al. (2010)
miR-451 14-3-3 z miR-144/451−/− mice
Protection against erythroid oxidant stress
Studies on functions related to oxidative stress in erythroid and nonerythroid tissues
[48]
CFU-E: Colony-forming unit-erythroid; Hb: Hemoglobin; HbF: Fetal hemoglobin; hESC: Human embryonic stem cell; hmbsb: Hydroxymethylbilane synthase b; LMO2: LIM domain only 2 (rhombotin-like 1); Myb: v-myb myeloblastosis viral oncogene homolog transcription factor.

Epigenomics (2012) 4(1)58 future science group
Review Bianchi, Zuccato, Finotti, Lampronti, Borgatti & Gambari
Our research group has recently analyzed the miRNA profile in uninduced K562 cells and in K562 cells induced to erythroid differ-entiation by MTH; the identified upregulated miRNAs were then assayed by RT-PCR in order to confirm the microarray ana lysis. Following this approach, we demonstrated that miR-210 is induced in time-dependent and dose-depen-dent fashion during MTH-mediated erythroid differentiation of K562 cells, allowing to pro-pose this miRNA as associated with erythroid differentiation [24]. It is very interesting to note that miR-210 has been recently associated with hypoxia [59,60]. This has been first demonstrated by Kulshreshtha et al., who described for the first time the miRNA signature of hypoxia, which includes high expression of miR-210 [59]. Interestingly, hypoxia has been recently dem-onstrated to alter progression of the erythroid program [61]. Low oxygen pressure might indeed modulate the relative amounts and types of Hb produced by erythroid cells, leading to increased HbF during stress erythropoiesis [62].
The involvement of miR-210 in erythroid differention was also reported by Kosaka et al. using UT-7 cells as a model system [50]. This group used a human miRNA microar-ray to analyze miRNA expression in the EPO-dependent cell line UT-7/EPO. Among 324 human miRNAs, miR-188, miR-362 and miR-210 levels were significantly elevated in UT-7/EPO cells, and stimulation with EPO in UT-7 cells increased the level of these three miRNAs. Notably, knockdown of miR-210 in UT-7/EPO cells led to apoptosis [50]. The expression of miR-210 was elevated during ery-throid maturation in vitro. These data suggest miR-210 as a member a new class of regulatory miRNAs that might play an important role in erythroid maturation and, possibly, g-globin gene expression. Accordingly, in consideration of the importance of controlling the fetal–adult switching in globin gene expression, several studies have been focused on characterizing possible relatioships between g-globin gene expression and miRNAs.
miRNA involved in the control of the expression of fetal globin genesThe control of the expression of fetal globin genes is of great interest for theoretical as well as practical points of view. Induced increase of HbF production, in fact, has been firmly dem-onstrated to be clinically beneficial. Clinical trials aimed at increasing HbF synthesis in b-thalassemia have included administration of
cell-cycle-specific agents, hematopoietic growth factors and short-chain fatty acids, all of which stimulate g-globin synthesis by different mech-anisms. Several clinical trials employing HbF inducers have been undertaken in the last decade [25,63,64], most of them are based on the use of hydroxyurea.
Several conclusions should be drawn follow-ing the comparative ana lysis of the data found in the literature on HbF inducers as potential drugs for pharmacological treatment of b-thalas-semia: the approach is reasonable, on the basis of the clinical parameters exhibited by patients with hereditary persistence of fetal hemoglobin (HPFH) phenotype; clinical trials employing HbF inducers were effective in ameliorating in vivo clinical parameters of b-thalassemia patients; and a combined use of HbF induc-ers might be useful to maximize HbF induc-tion, both in vitro and in vivo. However, several considerations introduce cautions, including the fact that the response to HbF inducers, evaluated in vitro and in vivo is variable, and some b-thalassemia patients (and the erythroid cells derived from them) might be refractory to induction; the reasons for this phenomenon are largely unknown.
Accordingly, the studies on the possible effects of miRNAs on HbF production by erythroid cells are important. In our laboratory, we have analyzed the miRNA profile in erythroid precur-sor cells from normal and thalassemic patients expressing different levels of HbF (some of them displayed hereditary persistence of fetal hemoglobin [HPFH] phenotype) by micro-array (Figure 2g & H). For the miRNA expression study two approaches were followed: miRNA profiling and quantitative RT-PCR. RNA was isolated from unaffected subjects, b-thalassemia patients expressing low HbF levels and b-thalas-semia-HPFH patients, labeled and hybridized on miRNA microarray chips [24], using 2 µg of RNA from each sample, biotin labeled during reverse transcription using random hexamers. Raw data were normalized and analyzed by GeneSpring GX software version 7.3 (Agilent Technologies, Palo Alto, CA, USA). The microarray data (Figure 2g) allowed us to identify miR-210 (Figure 2H) as a highly expressed miRNA in the erythroid precursor cells from a HPFH patient [24]. This miRNA is also expressed fol-lowing treatment of erythroid precursors with MTH, in association with a sharp increase of HbF production (see the representative results shown in Figure 2C–F) [24]. This is one among several observations, suggesting a link between

www.futuremedicine.com 59future science group
miRNA & erythroid differentiation Review
miRNA expression and HbF production in erythroid cells.
In this respect, a very intriguing observation was reported by Sankaran et al., who noted that in human trisomy 13, there is delayed switching and persistence of HbF and elevation of embry-onic Hb in newborns [51]. In partial trisomy cases, this trait maps to chromosomal band 13q14; by examining the genes in this region, two miR-NAs, miR-15a and miR-16-1, appear to be top candidates for the elevated HbF levels. Indeed, increased expression of these miRNAs in primary human erythroid progenitor cells results in ele-vated fetal and embryonic Hb gene expression. Moreover, this group showed that a direct target of these miRNAs, myb, plays an important role in silencing the fetal and embryonic Hb genes. myb may be important therapeutic targets to increase HbF levels in patients with sickle cell disease and b-thalassemia [51].
This research article is in line with a general concept that was followed using DNA binding drugs (DBD) or oligonucleotide decoys for TFs (decoy-ODN) able to inhibit TF–DNA inter-actions: the human g-globin genes are under negative transcriptional control caused by TFs exerting repressor activities. Examples are reported in the literature are Oct-1 [65], Myb [66] and BCL11A [67]. Interestingly BCL11A expres-sion appears to be regulated by KLF [68–70]. Notably, miRNAs targeting some of these TFs have been described and are responsible for driving induction of g-globin gene expression, in some cases by inhibiting the production of transcription repressor factors. For example, the already described paper by Sankaran et al. iden-tified miR-15a, miR-16-1 as important thera-peutic targets to increase HbF levels in patients with sickle cell disease and b-thalassemia, via i nhibition of Myb [67].
Erythroid-specific TFs can regulate tran-scription of both globin and miRNA genes. Fu et al. found that the erythroid lineage-specific miRNA gene, miR-144, expressed at specific developmental stages during zebrafish embryo-genesis, negatively regulates the embryonic a-globin, but not embryonic b-globin gene expression, through physiologically targeting Klfd, an erythroid-specific Krüppel-like TF [49]. Klfd selectively binds to the CACCC boxes in the promoters of both a-globin and miR-144 genes to activate their transcriptions, thus form-ing a negative feedback circuitry to fine-tune the expression of embryonic a-globin gene. The selective effect of the miR-144–Klfd pathway on globin gene regulation may thereby constitute
a novel therapeutic target for improving the c linical outcome of patients with t halassemia [49].
Interestingly, a recent study has shown the murine miR-144/451 locus is transcriptionally regulated by GATA-1 [45], an inducer of g-globin gene transcription [71,72]. In agreement, miRNAs are able to target mRNAs coding erythroid TFs and, conversely, erythroid TFs can modulate transcription of genes for miRNA involved in erythroid differentiation. The evidence sup-porting this conclusion are that at least two TFs, GATA-1 and KLF, bind to the promoter of a miRNA (miR-144) targeting repressor of g-globin gene trascription (myb) [45,46,49]. Figure 4
tentatively outlines a working hypothesis on the interplay between the transcription of miR-NAs, miRNA-regulated levels of TF mRNAs, production of TFs and transcription of g-globin genes. This working hypothesis is based on the assumption that miRNA sequences and miR-binding sites present in target mRNA stetches are conseved through molecular evolution. The GATA-1 dependent upregulation of miR-144 and the reported loop miR-144–KLF leading to miR-144-dependent downregulation of KLF has a secondary effect, that is, the downregula-tion of BLC11A, a strong repressor of g-globin gene transcription [67–70,73,74]. More specifically, Borg et al. [70] reported that the promoter area of BCL11A gene contains several putative KLF1 binding sites affecting its transcription. When the KLF1 binding activity is low, either through mutations of KLF1 alleles in HPFH or experi-mentally thorugh shRNA-mediated knockdown in erythroid cells, transcription of BCL11A decreases, leading to high transcription of g-globin genes [70]. According with the working hypothesis outlined in Figure 4, GATA-1 might retain a direct effect in upregulating g-globin gene expression at transcriptional level; further-more, the same TF is able to upregulate miRNAs affecting expression of g-globin gene repressors, leading to an indirect positive effect of g-globin gene transcription. Despite this model still hav-ing to be confirmed on different erythroid cel-lular systems, the available data strongly suggest that the interplay between TFs and miRNAs is a fashinating possibility occurring during erythroid-specific development.
Involvement of miRNA in terminal stages of erythoid differentiation: enucleationAlthough most of the available data on miRNA and erythroid differentiation are focused on the onset of this process and on expression of the

Epigenomics (2012) 4(1)60 future science group
Review Bianchi, Zuccato, Finotti, Lampronti, Borgatti & Gambari
major erythroid-specific genes (such as the glo-bin genes), miRNAs have been associated also to late stages of erythropoiesis. For instance, recent reports suggest that miRNA might be involved in enucleation, a very important step of terminal erythroid differentiation [54].
In all vertebrates, the cell nucleus becomes highly condensed and transcriptionally inactive during the final stages of red cell biogenesis. Enucleation, the process by which the nucleus is extruded by budding off from the erythro-blast, is unique to mammals. Enucleation has
miRNA gene
TF-rsynthesis
miRNA gene
TF-aTF-r mRNA
AAAAAA
AAAAAA
High mRNAtranscription
DownregulatedTF-r synthesis
TF-r gene
TF-r gene
TF-r mRNA
No/low mRNAtranscription
γ-globin gene
γ-globin gene
AAAAAA
RISC
Figure 4. Crosstalk between transcription factors regulating the promoter of g-globin genes and the transcription of miRNAs: a working hypothesis. (A) In this example, a TF responsible for activating the human g-globin gene (TF-a, such as GATA-1) is also an activator of a miRNA (miR-144) gene, which produces a mature miRNA targeting a mRNA coding a TF (such as KFL1) activator of a gene coding one of the major repressors of the g-globin gene transcription (TF-r, such as BCL11A). (B) The activation of the g-globin gene, therefore, may be due to two parallel converging effects: the miRNA-dependent inhibition of a transcription repressor and a direct enhancing effect of TFs targeting the human g-globin gene promoter. Recently described miRNA-regulated TFs are v-myb myeloblastosis viral oncogene homolog transcription factors, (downregulated by miR-15a and miR-16-1) and the KLFd, homologous to the key erythroid regulator KLF1 (downregulated by miR-144). This scheme has been designed on the basis of studies published by Dore et al. [45], demonstrating binding of GATA-1 (a TF known to activate human g-globin genes) to the miR-144 promoter, by Fu et al. [49]., demonstrating targeting of KLFd mRNA by miR-144, by Borg et al. [70], demonstrating interaction of KFL1 to the BCL11A promoter, and by Zhou et al. [69], demonstrating that downregulation of KLF1 leads to downregulation of BCL-11A and, subsequently, to increased transcription of g-globin genes [69,70].RISC: RNA-induced silencing complex TF: Transcription factor.

www.futuremedicine.com 61future science group
miRNA & erythroid differentiation Review
critical physiological and evolutionary signifi-cance, allowing elevation of Hb levels in the blood and the flexible biconcave shape of red cells [75]. Recent experiments reveal that enucle-ation involves multiple molecular and cellular pathways that include histone deacetylation, actin polymerization, cytokinesis, cell–matrix interactions, specific miRNAs and vesicle traf-ficking; many evolutionarily conserved proteins and genes have been recruited to participate in this unique mammalian process [75].
Using RNA-seq technology, Zhang et al. found that the majority of miRNAs present in colony-forming unit-erythroid (CFU-E) pro-genitors are downregulated during terminal erythroid differentiation [54]. Of the develop-mentally downregulated miRNAs, ectopic over-expression of miR-191 blocks erythroid enucle-ation but has minor effects on proliferation and differentiation. The mRNA targets of miR-191 were two erythroid-enriched and developmen-tally upregulated genes, Riok3 and Mxi1. As expected, knockdown of either Riok3 or Mxi1 inhibits enucleation, and either physiological overexpression of miR-191 or knockdown of Riok3 or Mxi1 blocks chromatin condensation. The conclusion of this study is that downregula-tion of miR-191 is essential for erythroid chro-matin condensation and enucleation by allowing upregulation of Riok3 and Mxi1 [54].
miRNA therapeutics for hematological diseasesThe discovery of miRNAs involved in hemato-logical diseases has two important implications: the development of diagnostic and prognostic tools; and the development of novel therapeu-tic approaches [76]. As far as therapeutics, the most interesting finding about the studies on the involvement of miRNAs in erythroid differen-tiation is that some of them are downregulated, while others are upregulated in associations with HbF production. This clearly has clinical appli-cations, since induction of HbF is very important for the treatment of b-thalassemia and sickle cell anemia [25,35–39,63–65,77–79]. Accordingly, pharma-cologically-induced downregulation of miRNAs can be obtained using antisense molecules tar-geting miRNAs which are found downregulated in association with HbF induction; conversely, the employment of miRNA mimics might be used in the cases miRNAs are upregulated to stimulate HbF production.
A further possible interest for development of clinical relevant protocols based on miRNAs is the recent finding that oxidative stress can
be modulated by miRNAs. An example of this research field is the very interesting study recently published by Sangokoya et al., who found that the miRNA expression in normal and erythrocytes containing sickle cell anemia hemoglobin (HbSS) exhibit dramatic differences. In this study, they identify a subset of HbSS patients with higher erythrocytic miR-144 expression and more severe anemia [21]. HbSS erythrocytes are known to have reduced tolerance for oxidative stress, yet the basis for this phenotype remains unknown. This study reveals that miR-144 directly regulates NRF2, a central regulator of cellular response to oxidative stress, and modulates the oxidative stress response in K562 cell line and primary erythroid progeni-tor cells. Increased miR-144 is associated with reduced NRF2 levels in HbSS reticulocytes, with decreased glutathione regeneration and attenu-ated antioxidant capacity in HbSS erythrocytes, thereby providing a possible mechanism for the reduced oxidative stress tolerance and increased anemia severity seen in HbSS patients [21].
Similar activities related to the bicistronic miRNA locus miR-144/451 have been reported by Yu et al., who demonstrated that at least some protective activities of miR-451 are related to its ability to directly suppress production of 14–3-3z, a phospho-serine/threonine-binding protein that inhibits nuclear accumulation of TF FoxO3, a positive regulator of erythroid a ntioxidant genes [48].
These two reports define novel miRNA-regu-lated pathway that protects erythrocytes against oxidant stress and should be considered for pos-sible development of novel therapeutic proto-cols aimed at reducing oxidative stress, a major problem in thalassemia and sickle cell anemia.
ConclusionmiRNAs are deeply involved in gene expres-sion of erythroid cells and on the onset of early stages of differentiation, modulating transcrip-tion of globin genes and the maturation of the erythroid cells during the terminal stages of differentiation.
Before considering the evidence reviewed here to be conclusive, we must accept the fact that several observations were performed based on leukemia cell lines (e.g., the K562 and UT-7 cell lines). As miRNA expression pattern changes in cancer [80–82], miRNA expression in a leukemia cell line may not represent normal physiologic conditions of erythroid cells. Furthermore, as shown in Figure 1, several data on these systems are based on chemical treatment to force erythroid induction [25]. Changes in miRNA expression in

Epigenomics (2012) 4(1)62 future science group
Review Bianchi, Zuccato, Finotti, Lampronti, Borgatti & Gambari
erythroid induced K562-like cells may be due to chemical treatment, rather than activation of ery-throid pathways. For these reasons, all the impor-tant information obtained using these leukemia cell lines should be confirmed in other in vitro experimental systems resembling normal ery-throipoiesis (including erythroid precursor cells from normal donors and thalassemic patients) as well as in vivo studies.
These studies will allow for the percise identi-fication of these miRNA/target mRNA networks and are expected to allow the design of novel diag-nostic/prognostic tools as well as novel therapeutic approaches based of miRNA antisense technology or miRNA mimics. For instance, anti-miRNA molecules targeting miRNAs, which are down-regulated in association with high HbF levels, might be considered for inducing HbF in ery-throid precursor; this effects might be reached also with molecules mimicking the function of miRNAs upregulated in HbF-producing cells. As already pointed out, HbF induction is of interest as a possible therapeutic interevention in b-thalassemia and sickle cell anemia.
Future perspectiveThe studies that demonstrate that miRNAs are promising candidates for drug targeting with the aim of developing possible reagents for therapy and diagnostics have supported activities to bring this research to industrial exploitation and a clini-cal setting [74]. Several commercial companies have already launched their activities, by including research activities miRNA-based diagnostics and therapeutics including Mirna Therapeutics [201], Regulus Therapeutics [202], Sirna Therapeutics [203], Alnylam Pharmaceuticals [204], Santaris Pharma [205], Rosetta Genomics [206], ActiGenics [207], Isis Pharmaceuticals [208] and Miragen Therapeutics [209]. The interest of these compa-nies in miRNA is based on the involvement of miRNAs in human pathologies and on the fact that alterations of differentiated functions, pos-sibly associated with disregulation of miRNA activity, may be corrected by interfering with miRNA activity (antisense approach) or mim-icking miRNA functions (miRNA replacement therapy). This appealing approach is expected to bring important innovative strategies in hema-tological diseases, despite the fact that this field of application of miRNA-based technologies is just starting.
By looking at FreshPatents [210] and Espacenet [211] data banks, very few examples of patents related to the employment of approaches target-ing miRNAs are available. An example of a general
patent is the US2010003674-A1 (Adult stem cells, molecular signatures and applications in the evalu-ation, diagnosis, and therapy of mammalian con-ditions) [101] which relates to the identification of a stem cell-specific signature or signatures composed of protein and/or nucleic acid markers expressed by virtue of the position of a cell or cells in the time line of its/their development and the impact of the cells’ environment on this signature as it relates to the cells’ stem cell potential. The composition and combination of these signatures provides a means of identifying, manipulating and differentiating adult stem cells and thus, their acquisition and utilization in research, diagnosis and therapy of normal and pathological conditions. An example of a more specific patent is the USPTO Patent Application 20100296630 (Increasing EPO using nucleic acids hybridizable to miRNA and precur-sors thereof) (Amgen Inc., Law Department, Seattle, WA, USA; inventors: Sumedha D Jayasena, Susan Swift) [102] is focused on the pos-sible objective of increasing EPO levels following targeting of pri-miR or mature miRNAs.
With regard to miRNA-based clinical tri-als [212], only one is applied in the hematological field (NCT01108159) [213] to identify altered miRNA/mRNA expression involved in the initia-tion, progression or treatment response of hemato-logic disorders. The majority of the other miRNA-based clinical trials (most of which focused on diagnosis rather than miRNA-based therapy) are related to cancer (41 trials), infectious diseases (six clinical trials) and inflammation (five clinical trials).
In conclusion, all the scientific and trans-lational-oriented data available suggest that miRNA technology will be applied to clinical issues in the near future (both diagnostic and therapeutic) and that this activity will also include hematologicals diseases, such as thalassemia and sickle cell anemia.
Financial & competing interests disclosureR Gambari is granted by Fondazione Cariparo (Cassa di Risparmio di Padova e Rovigo), CIB, by UE ITHANET Project (eInfrastructure for the Thalassaemia Research Network), by Telethon (contract GGP10214) and by COFIN-2007. This research is also supported by Associazione Veneta per la Lotta alla Talassemia (AVLT), Rovigo. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.

www.futuremedicine.com 63future science group
miRNA & erythroid differentiation Review
Executive summary
Background � miRNAs regulate gene expression by targeting mRNAs, inducing translational repression or mRNA degradation.
� miRNA are involved in cell cycle, apoptosis, inflammation and terminal differentiation.
Methods � The miRNA profiling has been conducted using both in vitro and in vivo experimental model systems.
� The involvement of miRNA is gene expression has been validated using antisense molecules for selective interference and forced expression of miRNA under ana lysis.
Results � miRNAs are deeply involved in erythroid differentiation, including enucleation occurring during the terminal stages of differentiation.
� miRNAs control the expression of g-globin genes, targeting mRNAs coding repressor transcription factors.
� miRNAs expression regulates erythroid oxidant stress.
Conclusion � The study of miRNAs involved in erythroid differentiation allows further ana lysis on the molecular mechanisms regulating human erythropoiesis.
� The precise identification of the miRNA/target mRNA networks allows the design of novel diagnostic/prognostic tools and therapeutic approaches.
Future perspective � Several companies involved in miRNA antisense technology or miRNA mimics are expected to include hematological disorders in their future pipelines.
References1 He L, Hannon GJ. MicroRNAs: small RNAs
with a big role in gene regulation. Nat. Rev. Genet. 5, 522–531 (2010).
2 Kozomara A, Griffiths-Jones S. miRBase: integrating microRNA annotation and deep-sequencing data. Nucl. Acids Res. 39, 152–157 (2010).
3 Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 11, 597–610 (2010).
4 Sontheimer EJ, Carthew RW. Silence from within: endogenous siRNAs and miRNAs. Cell 122, 9–12 (2005).
5 Subramanian S, Steer CJ. microRNAs as gatekeepers of apoptosis. J. Cell Physiol. 223, 89–98 (2010).
6 Wang F, Yu J, Yang GH, Wang XS, Zhang JW. Regulation of erythroid differentiation by miR-376a and its targets. Cell Res. 21(8), 1196–1209 (2011).
7 Alvarez-Garcia I, Miska EA. microRNA functions in animal development and human disease. Development 132, 4653–4662 (2005).
8 Taccioli C, Fabbri E, Visone R et al. UCbase & miRfunc: a database of ultraconserved sequences and microRNA function. Nucl. Acids Res. 37(Suppl. 1), D41–D48 (2009).
9 Bracken CP, Szubert JM, Mercer TR et al. Global ana lysis of the mammalian RNA degradome reveals widespread miRNA-dependent and miRNA-independent
endonucleolytic cleavage. Nucl. Acids Res. 39(13), 5658–5668 (2011).
10 Huntzinger E, Izaurralde E. Gene silencing by microRNAs: contributions of translational repression and mRNA decay. Nat. Rev. Genet. 12, 99–110 (2011).
11 van Rooij E. The art of microRNA research. Circ. Res. Rev. 108(2), 219–234 (2011).
12 Huang Y, Zou Q, Song H et al. A study of miRNAs targets prediction and experimental validation. Prot. Cell 1(11), 979–986 (2010).
13 Bueno MJ, Malumbres M. microRNAs and the cell cycle. Biochim. Biophys. Acta 1812(5), 592–601 (2011).
14 Bayarsaihan D. Epigenetic mechanisms in inflammation. J. Dent. Res. Rev. 90(1), 9–17 (2011).
15 Zhang W, Dolan ME. The emerging role of microRNAs in drug responses. Curr. Opin. Mol. Ther. Rev. 12(6), 695–702 (2010).
16 Salminen A, Ojala J, Kaarniranta K. Apoptosis and aging: increased resistance to apoptosis enhances the aging process. Cell Mol. Life Sci. Rev. (6), 1021–1031 (2011).
17 Metallo CM, Azarin SM, Ji L, de Pablo JJ, Palecek SP. Engineering tissue from human embryonic stem cells. J. Cell Mol. Med. 12(3), 709–729 (2008).
18 Huang X, Gschweng E, Van Handel B, Cheng D, Mikkola HK, Witte ON. Regulated expression of microRNAs-126/126* inhibits erythropoiesis from human embryonic stem cells. Blood 117(7), 2157–2165 (2011).
19 Lozzio CB, Lozzio BB. Human chronic myelogenous leukemia cell-line with positive Philadelphia chromosome. Blood 45, 321–334 (1975).
20 Mikami M, Sadahira Y, Haga A, Otsuki T, Wada H, Sugihara T. Hypoxia-inducible factor-1 drives the motility of the erythroid progenitor cell line, UT-7/Epo, via autocrine motility factor. Exp. Hematol. 33(5), 531–541 (2005).
21 Sangokoya C, Telen MJ, Chi JT. microRNA miR-144 modulates oxidative stress tolerance and associates with anemia severity in sickle cell disease. Blood 116(20), 4338–4348 (2010).
22 Yang GH, Wang F, Yu J, Wang XS, Yuan JY, Zhang JW. microRNAs are involved in erythroid differentiation control. J. Cell Biochem. 107(3), 548–556 (2009).
23 Bruchova-Votavova H, Yoon D, Prchal JT. miR-451 enhances erythroid differentiation in K562 cells. Leuk. Lymphoma 51(4), 686–693 (2010).
24 Bianchi N, Zuccato C, Lampronti I, Borgatti M, Gambari R. Expression of miR-210 during erythroid differentiation and induction of g-globin gene expression. BMB Rep. 42(8), 493–499 (2009).
25 Gambari R, Fibach E. Medicinal chemistry of fetal hemoglobin inducers for treatment of b-thalassemia. Curr. Med. Chem. Rev. 14(2), 199–212 (2007).
26 Rutherford TR, Clegg JB, Weatherall DJ. K562 human leukaemic cells synthesise embryonic haemoglobin in response to haemin. Nature 280(5718), 164–165 (1979).

Epigenomics (2012) 4(1)64 future science group
Review Bianchi, Zuccato, Finotti, Lampronti, Borgatti & Gambari
27 Wanda PE, Walker MM. Hemoglobin induction by Ara-C in human erythroleukemic cells (K562) is cell-cycle dependent. Leuk. Res. 13(8), 683–688 (1989).
28 Witt O, Sand K, Pekrun A. Butyrate-induced erythroid differentiation of human K562 leukemia cells involves inhibition of ERK and activation of p38 MAP kinase pathways. Blood 95(7), 2391–2396 (2000).
29 Lampronti I, Bianchi N, Borgatti M, Fibach E, Prus E, Gambari R. Accumulation of g-globin mRNA in human erythroid cells treated with angelicin. Eur. J. Haematol. 71(3), 189–195 (2003).
30 Bianchi N, Osti F, Rutigliano C et al. The DNA-binding drugs mithramycin and chromomycin are powerful inducers of erythroid differentiation of human K562 cells. Br. J. Haematol. 104(2), 258–265 (1999).
31 Bianchi N, Chiarabelli C, Borgatti M, Mischiati C, Fibach E, Gambari R. Accumulation of g-globin mRNA and induction of erythroid differentiation after treatment of human leukaemic K562 cells with tallimustine. Br. J. Haematol. 113(4), 951–961 (2001).
32 Bianchi N, Ongaro F, Chiarabelli C et al. Induction of erythroid differentiation of human K562 cells by cisplatin analogs. Biochem. Pharmacol. 60(1), 31–40 (2000).
33 Alter BP, Goff SC. Electrophoretic separation of human embryonic globin demonstrates 'a-thalassemia’ in human leukemia cell line K562. Biochem. Biophys. Res. Commun. 94(3), 843–848 (1980).
34 Pope SH, Fibach E, Sun J, Chin K, Rodgers GP. Two-phase liquid culture system models normal human adult erythropoiesis at the molecular level. Eur. J. Haematol. 64(5), 292–303 (2000).
35 Fibach E, Bianchi N, Borgatti M, Prus E, Gambari R. Mithramycin induces fetal hemoglobin production in normal and thalassemic human erythroid precursor cells. Blood 102, 1276–1281 (2003).
36 Liakopoulou E, Li Q, Stamatoyannopoulos G. Induction of fetal hemoglobin by propionic and butyric acid derivatives: correlations between chemical structure and potency of Hb F induction. Blood Cells Mol. Dis. 29(1), 48–56 (2002).
37 Fibach E, Kollia P, Schechter AN, Noguchi CT, Rodgers GP. Hemin-induced acceleration of hemoglobin production in immature cultured erythroid cells: preferential enhancement of fetal hemoglobin. Blood 85(10), 2967–2974 (2005).
38 Amoyal I, Fibach E. Hemoglobin switch in the newborn: a flow cytometry ana lysis Neonatology 1, 61–68 (2007).
39 Johnson J, Hunter R, McElveen R, Qian XH, Baliga BS, Pace BS. Fetal hemoglobin induction by the histone deacetylase inhibitor, scriptaid. Cell Mol. Biol. 51(2), 229–238 (2005).
40 Rhodes J, Hagen A, Hsu K et al. Interplay of pu.1 and GATA1 determines myelo-erythroid progenitor cell fate in zebrafish. Dev. Cell 8(1), 97–108 (2005).
41 Galloway JL, Wingert RA, Thisse C, Thisse B, Zon LI. Loss of GATA1 but not GATA2 converts erythropoiesis to myelopoiesis in zebrafish embryos. Dev. Cell 8(1), 109–116 (2005).
42 Schier AF, Giraldez AJ. microRNA function and mechanism: insights from zebra fish. Cold Spring Harb. Symp. Quant. Biol. 71, 195–203 (2006).
43 Dong M, Fu YF, Du TT et al. Heritable and lineage-specific gene knockdown in zebrafish embryo. PLoS ONE 4(7), e6125 (2009).
44 Grabher C, Payne EM, Johnston AB et al. Zebrafish microRNA-126 determines hematopoietic cell fate through c-Myb. Leukemia 25(3), 506–514 (2011).
45 Dore LC, Amigo JD, Dos Santos CO et al. A GATA-1-regulated microRNA locus essential for erythropoiesis. Proc. Natl Acad. Sci. USA 105(9), 3333–3338 (2008).
46 Du TT, Fu YF, Dong M et al. Experimental validation and complexity of miRNA–mRNA target interaction during zebrafish primitive erythropoiesis. Biochem. Biophys. Res. Commun. 381(4), 688–693 (2009).
47 Pase L, Layton JE, Kloosterman WP, Carradice D, Waterhouse PM, Lieschke GJ. miR-451 regulates zebrafish erythroid maturation in vivo via its target GATA2. Blood 113(8), 1794–1804 (2009).
48 Yu D, dos Santos CO, Zhao G et al. miR-451 protects against erythroid oxidant stress by repressing 14–3–3z. Genes Dev. 24(15), 1620–1633 (2010).
49 Fu YF, Du TT, Dong M et al. Mir-144 selectively regulates embryonic a-hemoglobin synthesis during primitive erythropoiesis. Blood 113(6), 1340–1349 (2009).
50 Kosaka N, Sugiura K, Yamamoto Y et al. Identification of erythropoietin-induced microRNAs in haematopoietic cells during erythroid differentiation. Br. J. Haematol. 142, 293–300 (2008).
51 Sankaran VG, Menne TF, Šćepanović D, et al. microRNA-15a and -16–1 act via Myb to elevate fetal hemoglobin expression in human trisomy 13. Proc. Natl Acad. Sci. USA 108(4), 1519–1524 (2011).
52 Gabbianelli M, Testa U, Morsilli O et al. Mechanism of human Hb switching: a possible role of the kit receptor/miR 221–222
complex. Haematologica 95(8), 1253–1260 (2010).
53 Felli N, Pedini F, Romania P et al. microRNA 223-dependent expression of LMO2 regulates normal erythropoiesis. Haematologica 94(4), 479–486 (2009).
54 Zhang L, Flygare J, Wong P, Lim B, Lodish HF. miR-191 regulates mouse erythroblast enucleation by down-regulating Riok3 and Mxi1. Genes Dev. 25(2), 119–124 (2011).
55 Svasti S, Masaki S, Penglong T et al. Expression of microRNA-451 in normal and thalassemic erythropoiesis. Ann. Hematol. 89(10), 953–958 (2010).
56 Felli N, Fontana L, Pelosi E et al. microRNAs 221 and 222 inhibit normal erythropoiesis and erythroleukemic cell growth via kit receptor down-modulation. Proc. Natl Acad. Sci. USA 102, 8081–8086 (2005).
57 Choong ML, Yang HH, McNiece I. microRNA expression profiling during human cord blood-derived CD34 cell erythropoiesis. Exp. Hematol. 35, 551–564 (2007).
58 Georgantas RW 3rd, Hildreth R, Morisot S et al. CD34+ hematopoietic stem-progenitor cell microRNA expression and function: a circuit diagram of differentiation control. Proc. Natl Acad. Sci. USA 104, 2750–2755 (2007).
59 Kulshreshtha R, Davuluri RV, Calin GA, Ivan M. A microRNA component of the hypoxic response. Cell Death Differ. 15, 667–671 (2008).
60 Fasanaro P, D’Alessandra Y, Di Stefano V et al. microRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J. Biol. Chem. 283, 15878–15883 (2008).
61 Rogers HM, Yu X, Wen J, Smith R, Fibach E, Noguch CT. Hypoxia alters progression of the erythroid program. Exp. Hematol. 36(1), 17–27 (2008).
62 Alter BP. Fetal erythropoiesis in stress hematopoiesis. Exp. Hematol. 7, 200–209 (1979).
63 Dixit A, Chatterjee TC, Mishra P et al. Hydroxyurea in thalassemia intermedia – a promising therapy. Ann. Hematol. 84(7), 441–446 (2005).
64 Watanapokasin Y, Chuncharunee S, Sanmund D et al. In vivo and in vitro studies of fetal hemoglobin induction by hydroxyurea in b-thalassemia/hemoglobin E patients. Exp. Hematol. 33(12), 1486–1492 (2005).
65 Xu XS, Hong X, Wang G. Induction of endogenous g-globin gene expression with decoy oligonucleotide targeting Oct-1 transcription factor consensus sequence. J. Hematol. Oncol. 2, 15 (2009).

www.futuremedicine.com 65future science group
miRNA & erythroid differentiation Review
65www.futuremedicine.com
66 Jiang J, Best S, Menzel S et al. cMyb is involved in the regulation of fetal hemoglobin production in adults. Blood 108(3), 1077–1083 (2006).
67 Sankaran VG, Xu J, Orkin SH. Transcriptional silencing of fetal hemoglobin by BCL11A. Ann. NY Acad. Sci. 1202, 64–68 (2010).
68 Satta S, Perseu L, Moi P et al. Compound heterozygosity for KLF1 mutations associated with remarkable increase of fetal hemoglobin and red cell protoporphyrin. Haematologica 96(5), 767–770 (2011).
69 Zhou D, Liu K, Sun CW, Pawlik KM, Townes TM. KLF1 regulates BCL11A expression and g- to b-globin gene switching. Nat. Genet. 42(9), 742–744 (2010).
70 Borg J, Papadopoulos P, Georgitsi M et al. Haploinsufficiency for the erythroid transcription factor KLF1 causes hereditary persistence of fetal hemoglobin. Nat. Genet. 42(9), 801–805 (2010).
71 Woon Kim Y, Kim S, Geun Kim C, Kim A. The distinctive roles of erythroid specific activator GATA-1 and NF-E2 in transcription of the human fetal g-globin genes. Nucl. Acids Res. 39(16), 6944–6955 (2011).
72 Yao X, Kodeboyina S, Liu L et al. Role of STAT3 and GATA-1 interactions in g-globin gene expression. Exp. Hematol. 37(8), 889–900 (2009).
73 Bauer DE, Orkin SH. Update on fetal hemoglobin gene regulation in hemoglobinopathies. Curr. Opin. Pediatr. 23(1), 1–8 (2011).
74 Sankaran VG, Nathan DG. Reversing the hemoglobin switch. N. Engl. J. Med. 363(23), 2258–2260 (2010).
75 Ji P, Yeh V, Ramirez T, Murata-Hori M, Lodish HF. Histone deacetylase 2 is required for chromatin condensation and subsequent enucleation of cultured mouse fetal erythroblasts. Haematologica 95(12), 2013–2021 (2010).
76 Mack GS. MicroRNA gets down to business. Nat. Biotechnol. 25(6), 631–638 (2007).
77 Wilber A, Hargrove PW, Kim YS et al. Therapeutic levels of fetal hemoglobin in erythroid progeny of b-thalassemic CD34+ cells after lentiviral vector-mediated gene transfer. Blood 117(10), 2817–2826 (2011).
78 Watanapokasin R, Sanmund D, Winichagoon P, Muta K, Fucharoen S. Hydroxyurea responses and fetal hemoglobin induction in b-thalassemia/HbE patients’ peripheral blood erythroid cell culture. Ann. Hematol. 85(3), 164–169 (2006).
79 Cao H. Pharmacological induction of fetal hemoglobin synthesis using histone deacetylase inhibitors. Hematology 9(3), 223–233 (2004).
80 Zuo Z, Calin GA, de Paula HM et al. Circulating microRNAs let-7a and miR-16 predict progression-free survival and overall survival in patients with myelodysplastic syndrome. Blood 118(2), 413–415 (2011).
81 Corthals SL, Jongen-Lavrencic M, de Knegt Y et al. micro-RNA-15a and micro-RNA-16 expression and chromosome 13 deletions in multiple myeloma. Leuk. Res. 34(5), 677–681 (2010).
82 Bhattacharya R, Nicoloso M, Arvizo R et al. miR-15a and miR-16 control Bmi-1 expression in ovarian cancer. Cancer Res. 69(23), 9090–9095 (2009).
� Patents101 Slukvin II, Thomson JA, Vodyanyk MA,
Gumenyuk ME. US0003674 (2008).
102 AMGEN, INC.: US0296630 (2008).
� Websites201 Mirna Therapeutics.
www.mirnatherapeutics.com
202 Regulus Therapeutics. www.regulusrx.com
203 Sirna Therapeutics. www.sirna.com
204 Alnylam Pharmaceuticals. www.alnylam.com
205 Santaris Pharma A/S. www.santaris.com
206 Rosetta Genomics. www.rosettagenomics.com
207 Actigenics. www.actigenics.com
208 Isis Pharmaceuticals. www.isispharm.com
209 Miragen Therapeutics. www.miragentherapeutics.com
210 FreshPatents. http://freshpatents.com
211 Espacenet. http://worldwide.espacenet.com
212 Clinical Trials. www.clinicaltrials.gov
213 Integrated whole-genome analysis of hematologic disorders. http://clinicaltrials.gov/ct2/show/NCT01108159
Related Documents











