Involvement of μ-opioid receptors in antinociception and inhibition of gastrointestinal transit induced by 7-hydroxymitragynine, isolated from Thai herbal medicine Mitragyna speciosa Kenjiro Matsumoto a, ⁎ , Yoshio Hatori b , Toshihiko Murayama b , Kimihito Tashima a , Sumphan Wongseripipatana c , Kaori Misawa d , Mariko Kitajima d , Hiromitsu Takayama d , Syunji Horie a a Laboratory of Pharmacology, Faculty of Pharmaceutical Sciences, Josai International University, 1 Gumyo, Togane, Chiba 283-8555, Japan b Department of Chemical Pharmacology, Graduate School of Pharmaceutical Sciences, Chiba University, 1-8-1 Inohana, Chuo-ku, Chiba 260-8675, Japan c Department of Pharmacognosy, Faculty of Pharmaceutical Sciences, Chulalongkorn University, Bangkok 10330, Thailand d Department of Molecular Structure and Biological Function, Graduate School of Pharmaceutical Sciences, Chiba University, 1-33 Yayoi-cho, Inage-ku, Chiba 263-8522, Japan Received 28 February 2006; received in revised form 31 July 2006; accepted 4 August 2006 Available online 16 August 2006 Abstract 7-Hydroxymitragynine, a constituent of the Thai herbal medicine Mitragyna speciosa, has been found to have a potent opioid antinociceptive effect. In the present study, we investigated the mechanism of antinociception and the inhibitory effect on gastrointestinal transit of 7- hydroxymitragynine, and compared its effects with those of morphine. When administered subcutaneously to mice, 7-hydroxymitragynine produced antinociceptive effects about 5.7 and 4.4 times more potent than those of morphine in the tail-flick (ED 50 = 0.80 mg/kg) and hot-plate (ED 50 = 0.93 mg/kg) tests, respectively. The antinociceptive effect of 7-hydroxymitragynine was significantly blocked by the μ 1 /μ 2 -opioid receptor antagonist β-funaltrexamine hydrochloride (β-FNA) and the μ 1 -opioid receptor-selective antagonist naloxonazine in both tests. Thus, 7- hydroxymitragynine acts predominantly on μ-opioid receptors, especially on μ 1 -opioid receptors. Isolated tissue studies further supported its specificity for the μ-opioid receptors. Further, 7-hydroxymintragynine dose-dependently (ED 50 = 1.19 mg/kg, s.c.) and significantly inhibited gastrointestinal transit in mice, as morphine does. The inhibitory effect was significantly antagonized by β-FNA pretreatment, but slightly antagonized by naloxonazine. The ED 50 value of 7-hydroxymitragynine on gastrointestinal transit was larger than its antinociceptive ED 50 value. On the other hand, morphine significantly inhibits gastrointestinal transit at a much smaller dose than its antinociceptive dose. These results suggest that μ-opioid receptor mechanisms mediate the antinociceptive effect and inhibition of gastrointestinal transit. This compound induced more potent antinociceptive effects and was less constipating than morphine. © 2006 Elsevier B.V. All rights reserved. Keywords: 7-Hydroxymitragynine; Antinociception; Gastrointestinal transit; μ-Opioid receptor; Morphine 1. Introduction 7-Hydroxymitragynine (Fig. 1) is a minor constituent of Mi- tragyna speciosa (Ponglux et al., 1994). This herb has long been used in Thailand for its opium- (Burkill, 1935) and coca-like effects, and as a replacement for opium (Grewal, 1932; Suwanlert, 1975). We have studied the pharmacological activities of mitragynine, a major alkaloid of this herb (Watanabe et al., 1997; Matsumoto et al., 2005), and related alkaloids (Yamamoto et al., 1999; Takayama et al., 2002; Takayama, 2004; Matsumoto et al., 2006) and found that these compounds have opioid activities. Recently, we studied the opioid agonistic effects of the constituents of M. speciosa using in vitro assays. Among them, 7- hydroxymitragynine, which has a hydroxyl group at the C7 position of mitragynine, produced the most potent effect, which European Journal of Pharmacology 549 (2006) 63 – 70 www.elsevier.com/locate/ejphar ⁎ Corresponding author. Tel./fax: +81 475 53 4585. E-mail address: [email protected] (K. Matsumoto). 0014-2999/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2006.08.013

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

logy 549 (2006) 63–70www.elsevier.com/locate/ejphar
European Journal of Pharmaco
Involvement of μ-opioid receptors in antinociception and inhibition ofgastrointestinal transit induced by 7-hydroxymitragynine,isolated from Thai herbal medicine Mitragyna speciosa
Kenjiro Matsumoto a,⁎, Yoshio Hatori b, Toshihiko Murayama b,Kimihito Tashima a, Sumphan Wongseripipatana c, Kaori Misawa d,
Mariko Kitajima d, Hiromitsu Takayama d, Syunji Horie a
a Laboratory of Pharmacology, Faculty of Pharmaceutical Sciences, Josai International University, 1 Gumyo, Togane, Chiba 283-8555, Japanb Department of Chemical Pharmacology, Graduate School of Pharmaceutical Sciences,
Chiba University, 1-8-1 Inohana, Chuo-ku, Chiba 260-8675, Japanc Department of Pharmacognosy, Faculty of Pharmaceutical Sciences, Chulalongkorn University, Bangkok 10330, Thailand
d Department of Molecular Structure and Biological Function, Graduate School of Pharmaceutical Sciences,Chiba University, 1-33 Yayoi-cho, Inage-ku, Chiba 263-8522, Japan
Received 28 February 2006; received in revised form 31 July 2006; accepted 4 August 2006Available online 16 August 2006
Abstract
7-Hydroxymitragynine, a constituent of the Thai herbal medicine Mitragyna speciosa, has been found to have a potent opioid antinociceptiveeffect. In the present study, we investigated the mechanism of antinociception and the inhibitory effect on gastrointestinal transit of 7-hydroxymitragynine, and compared its effects with those of morphine. When administered subcutaneously to mice, 7-hydroxymitragynineproduced antinociceptive effects about 5.7 and 4.4 times more potent than those of morphine in the tail-flick (ED50=0.80 mg/kg) and hot-plate(ED50=0.93 mg/kg) tests, respectively. The antinociceptive effect of 7-hydroxymitragynine was significantly blocked by the μ1/μ2-opioid receptorantagonist β-funaltrexamine hydrochloride (β-FNA) and the μ1-opioid receptor-selective antagonist naloxonazine in both tests. Thus, 7-hydroxymitragynine acts predominantly on μ-opioid receptors, especially on μ1-opioid receptors. Isolated tissue studies further supported itsspecificity for the μ-opioid receptors. Further, 7-hydroxymintragynine dose-dependently (ED50=1.19 mg/kg, s.c.) and significantly inhibitedgastrointestinal transit in mice, as morphine does. The inhibitory effect was significantly antagonized by β-FNA pretreatment, but slightlyantagonized by naloxonazine. The ED50 value of 7-hydroxymitragynine on gastrointestinal transit was larger than its antinociceptive ED50 value.On the other hand, morphine significantly inhibits gastrointestinal transit at a much smaller dose than its antinociceptive dose. These resultssuggest that μ-opioid receptor mechanisms mediate the antinociceptive effect and inhibition of gastrointestinal transit. This compound inducedmore potent antinociceptive effects and was less constipating than morphine.© 2006 Elsevier B.V. All rights reserved.
Keywords: 7-Hydroxymitragynine; Antinociception; Gastrointestinal transit; μ-Opioid receptor; Morphine
1. Introduction
7-Hydroxymitragynine (Fig. 1) is a minor constituent of Mi-tragyna speciosa (Ponglux et al., 1994). This herb has long beenused in Thailand for its opium- (Burkill, 1935) and coca-likeeffects, and as a replacement for opium (Grewal, 1932; Suwanlert,
⁎ Corresponding author. Tel./fax: +81 475 53 4585.E-mail address: [email protected] (K. Matsumoto).
0014-2999/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2006.08.013
1975). We have studied the pharmacological activities ofmitragynine, a major alkaloid of this herb (Watanabe et al.,1997; Matsumoto et al., 2005), and related alkaloids (Yamamotoet al., 1999; Takayama et al., 2002; Takayama, 2004; Matsumotoet al., 2006) and found that these compounds have opioidactivities. Recently, we studied the opioid agonistic effects of theconstituents ofM. speciosa using in vitro assays. Among them, 7-hydroxymitragynine, which has a hydroxyl group at the C7position of mitragynine, produced the most potent effect, which

Fig. 1. Chemical structure of 7-hydroxymitragynine.
64 K. Matsumoto et al. / European Journal of Pharmacology 549 (2006) 63–70
suggests that the opioid effect ofM. speciosa is mostly based onthe activity of 7-hydroxymitragynine (Horie et al., 2005). 7-Hydroxymitragynine induced potent antinociceptive effects inmouse tail-flick and hot-plate tests, and its effects were morepotent than those of morphine when subcutaneously or orallyadministered (Matsumoto et al., 2004). Receptor-binding assaysrevealed that 7-hydroxymitragynine has a higher affinity for μ-opioid receptors than for the other opioid receptor types(Takayama et al., 2002; Matsumoto et al., 2004) (Fig. 1).
μ-Opioids represent the major class of strong analgesics, suchasmorphine, used clinically.Morphine plays an important role as apain-relieving agent, but it has a number of adverse effects, e.g.,respiratory depression, nausea, vomiting, constipation, tolerance,and dependence. Constipation can become amajor problemduringchronic opioid administration (Schug et al., 1992; McQuay, 1999;Portenoy, 1996), and relief from the adverse gastrointestinal effectsmarkedly enhances the quality of life for patients. In the case ofmorphine, the dose required for its analgesic effect is much higherthan that required for its constipating effect; thus, when morphineis used for analgesia, constipation is not a negligible issue (Megenset al., 1998).
Opioid receptors are widely distributed throughout thecentral and peripheral nervous system and play a fundamentalrole in pain and its adverse effects (Quock et al., 1999). In thepresent study, we studied the opioid receptor mechanisms of 7-hydroxymitragynine by using selective opioid antagonists in invivo and in vitro assays to clarify the mechanism by which 7-hydroxymitragynine produces antinociceptive effects in mice.In addition, we investigated the inhibition of gastrointestinaltransit to evaluate the constipating effect of 7-hydroxymitrgy-nine in comparison with morphine.
2. Materials and methods
2.1. Experimental animals
Male ddY-strain mice (Japan SLC, Hamamatsu, Japan)weighing 25–32 g and male albino guinea-pigs (Japan SLC)weighing 320–550 g were used. Animals were housed in atemperature-controlled room at 24 °C with lights on from 07:00–19:00 and had free access to food and water. All experiments wereperformed in compliance with the “Guiding Principles for the Careand Use of Laboratory Animals” approved by the JapanesePharmacological Society and the guidelines approved by theEthicalCommittee onAnimalCare andAnimal Experimentation ofJosai International University (#12). The number of animals used
was kept to the minimum necessary for a meaningful interpretationof the data, and animal discomfort was kept to the minimum.
2.2. Drugs
The drugs used in this study were morphine hydrochloride(Takeda Chemical Ind., Osaka, Japan), naloxone hydrochloride(MP Biomedicals, Irvine, CA), [D-Pen2, D-Pen5]-enkephalin(DPDPE; Bachem, Torrance, CA), naltrindole hydrochloride,cyprodime hydrobromide, nor-binaltorphimine dihydrochloride(norBNI), naloxonazine dihydrochloride, naloxone methiodide,[D-Ala2, N-MePhe4, Gly-ol5]-enkephalin (DAMGO), (5α,7α,8β)-(+)-N-Methyl-N-[7-(1-pyrrolidinyl)-1-oxaspiro[4.5]dec-8-yl]-ben-zeneacetamide (U69593) (Sigma Chemical Co., St. Louis, MO,USA), and β-funaltrexamine hydrochloride (β-FNA; Tocris-Cookson, Bristol, UK). 7-Hydroxymitragynine was synthesizedfrom mitragynine as described previously (Takayama et al., 2002).The purity (N99%) of these compounds was checked by high-performance liquid chromatography and 1H-nuclear magneticresonance (500 MHz) analysis (Takayama et al., 2002).
For the in vivo assays, 7-hydroxymitragyninewas dissolved inphosphate-buffered saline (pH 5.5). The other drugs weredissolved in saline. 7-Hydroxymitragynine was administeredsubcutaneously (s.c.) using a volume of 0.1 ml/10 g body weight.The opiate antagonists, naloxonemethiodide (3mg/kg), naloxone(2 mg/kg), naltrindole (3 mg/kg), norBNI (20 mg/kg), nalox-onazine (35 mg/kg), and β-FNA (40 mg/kg), were administereds.c. 15 min, 30 min, 30 min, 3 h, 24 h, and 24 h, respectively,before 7-hydroxymitragynine or morphine injection (s.c.). Theseprotocols were described by Paul et al. (1989) and Jinsmaa et al.(2004). High doses of naloxone and naloxone methiodide wereused to ensure maximum antagonism. For the in vitro assays, 7-hydroxymitragynine and cyprodime hydrobromide were firstdissolved in 100% dimethylsulfoxide to yield a 5 mM solution,and then subsequently diluted with distilled water. The otherdrugs were dissolved in distilled water.
2.3. Antinociceptive activity
2.3.1. Tail-flick testThe method was adapted from that of D'Amour and Smith
(1941). Mice respond to a focused heat stimulus by flicking ormoving their tail from the path of the stimulus, thereby exposing aphotocell located in the tail-flick analgesia meter (Ugo Basile Tail-flick Unit 7360, Ugo Basile, Comerio, Italy) immediately belowthe tail. The reaction time is automatically recorded. Prior totreatmentwith 7-hydroxymitragynine,morphine, vehicle, or saline,the nociceptive threshold was measured three times, and the meanof the reaction time was used as the pre-drug latency for eachmouse. A cut-off time of 10 s was used to prevent tissue damage.
2.3.2. Hot-plate testAnimals were placed on an electrically heated plate at 55±
0.2 °C, and the latency period until the occurrence of nociceptiveresponses such as licking, shaking the legs, or jumping wasmeasured. Prior to treatment with 7-hydroxymitragynine, mor-phine, vehicle, or saline, the nociceptive threshold was measured
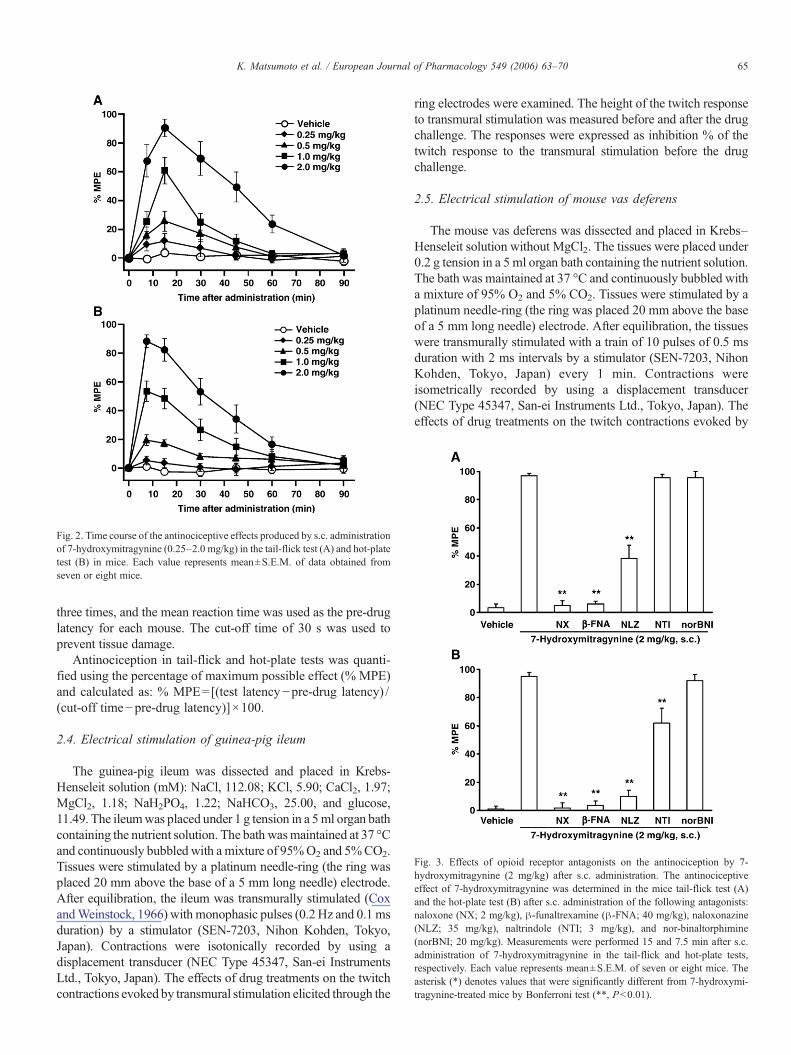
Fig. 3. Effects of opioid receptor antagonists on the antinociception by 7-hydroxymitragynine (2 mg/kg) after s.c. administration. The antinociceptiveeffect of 7-hydroxymitragynine was determined in the mice tail-flick test (A)and the hot-plate test (B) after s.c. administration of the following antagonists:naloxone (NX; 2 mg/kg), β-funaltrexamine (β-FNA; 40 mg/kg), naloxonazine(NLZ; 35 mg/kg), naltrindole (NTI; 3 mg/kg), and nor-binaltorphimine(norBNI; 20 mg/kg). Measurements were performed 15 and 7.5 min after s.c.administration of 7-hydroxymitragynine in the tail-flick and hot-plate tests,respectively. Each value represents mean±S.E.M. of seven or eight mice. Theasterisk (*) denotes values that were significantly different from 7-hydroxymi-tragynine-treated mice by Bonferroni test (**, Pb0.01).
Fig. 2. Time course of the antinociceptive effects produced by s.c. administrationof 7-hydroxymitragynine (0.25–2.0 mg/kg) in the tail-flick test (A) and hot-platetest (B) in mice. Each value represents mean±S.E.M. of data obtained fromseven or eight mice.
65K. Matsumoto et al. / European Journal of Pharmacology 549 (2006) 63–70
three times, and the mean reaction time was used as the pre-druglatency for each mouse. The cut-off time of 30 s was used toprevent tissue damage.
Antinociception in tail-flick and hot-plate tests was quanti-fied using the percentage of maximum possible effect (% MPE)and calculated as: % MPE=[(test latency−pre-drug latency) /(cut-off time−pre-drug latency)]×100.
2.4. Electrical stimulation of guinea-pig ileum
The guinea-pig ileum was dissected and placed in Krebs-Henseleit solution (mM): NaCl, 112.08; KCl, 5.90; CaCl2, 1.97;MgCl2, 1.18; NaH2PO4, 1.22; NaHCO3, 25.00, and glucose,11.49. The ileumwas placed under 1 g tension in a 5ml organ bathcontaining the nutrient solution. The bath wasmaintained at 37 °Cand continuously bubbled with a mixture of 95%O2 and 5%CO2.Tissues were stimulated by a platinum needle-ring (the ring wasplaced 20 mm above the base of a 5 mm long needle) electrode.After equilibration, the ileum was transmurally stimulated (CoxandWeinstock, 1966) with monophasic pulses (0.2 Hz and 0.1 msduration) by a stimulator (SEN-7203, Nihon Kohden, Tokyo,Japan). Contractions were isotonically recorded by using adisplacement transducer (NEC Type 45347, San-ei InstrumentsLtd., Tokyo, Japan). The effects of drug treatments on the twitchcontractions evoked by transmural stimulation elicited through the
ring electrodes were examined. The height of the twitch responseto transmural stimulation was measured before and after the drugchallenge. The responses were expressed as inhibition % of thetwitch response to the transmural stimulation before the drugchallenge.
2.5. Electrical stimulation of mouse vas deferens
The mouse vas deferens was dissected and placed in Krebs–Henseleit solution without MgCl2. The tissues were placed under0.2 g tension in a 5 ml organ bath containing the nutrient solution.The bath was maintained at 37 °C and continuously bubbled witha mixture of 95% O2 and 5% CO2. Tissues were stimulated by aplatinum needle-ring (the ring was placed 20 mm above the baseof a 5 mm long needle) electrode. After equilibration, the tissueswere transmurally stimulated with a train of 10 pulses of 0.5 msduration with 2 ms intervals by a stimulator (SEN-7203, NihonKohden, Tokyo, Japan) every 1 min. Contractions wereisometrically recorded by using a displacement transducer(NEC Type 45347, San-ei Instruments Ltd., Tokyo, Japan). Theeffects of drug treatments on the twitch contractions evoked by

Table 1Effect of cyprodime on twitch contraction inhibition by 7-hydroxymitragynine,DAMGO, and U69593 in guinea-pig ileum
Compound (concentration) Contraction(%)inhibitedbycompound
Contraction (%) reversedby cyprodime
30 nM 1 μM
7-Hydroxymitragynine (100 nM) 19.5±1.6 59.9±5.0 a 108.2±9.2 a
DAMGO (100 nM) 12.5±3.8 41.7±10.8 104.5±9.2 a
U69593 (1 μM) 11.7±3.2 11.4±2.6 15.7±1.4
Each value represents the mean±S.E.M. of five animals.a Pb0.01 significantly different from the values before the addition of
cyprodime (Bonferroni multiple comparison test).
Table 3Effect of naltrindole on twitch contraction inhibited by 7-hydroxymitragynine,DPDPE, DAMGO, and U69593 in mouse vas deferens
Compound (concentration) Contraction(%)inhibitedbycompound
Contraction (%) reversedby naltrindole
3 nM 30 nM
7-Hydroxymitragynine (300 nM) 7.8±1.5 8.4±1.9 18.9±2.9 a
DPDPE (100 nM) 12.1±3.2 42.5±8.0 b 83.8±2.9 b
DAMGO (300 nM) 11.9±2.1 13.8±3.3 19.4±3.6U69593 (1 μM) 19.1±4.8 21.4±5.2 24.0±6.9
Each value represents the mean±S.E.M. of five animals.a Pb0.05.b Pb0.01 significantly different from the values before the addition of
naltrindole (Bonferroni multiple comparison test).
66 K. Matsumoto et al. / European Journal of Pharmacology 549 (2006) 63–70
transmural stimulation elicited through the ring electrodes wereexamined. The height of the twitch response to transmuralstimulation was measured before and after the drug challenge.The responses were expressed as % inhibition of the twitchresponse to the transmural stimulation before the drug challenge.
2.6. Gastrointestinal transit
Mice were fasted, with water available ad libitum, for 18 hbefore the experiments. Fifteen minutes after s.c. injection of 7-hydroxymitragynine, morphine, vehicle, or saline, a charcoalmeal (an aqueous suspension of 10% charcoal and 5% gumarabic) was orally administered at a volume of 0.25 ml. Thirtyminutes after administration of the charcoal meal, the animalwas sacrificed by cervical dislocation, and the small intestinefrom the pylorus to the cecum was carefully removed. Both thelength of the small intestine from the pylorus to the cecum andthe farthest distance to which the charcoal meal had traveledwere measured. For each animal, the gastrointestinal transit(GIT) was calculated as the percentage of distance traveled bythe charcoal meal relative to the total length of the smallintestine. The inhibition of gastrointestinal transit (%) wascalculated as: Inhibition of gastrointestinal transit (%)=[(salineor vehicle GIT−drug GIT) / (saline or vehicle GIT)]×100.
2.7. Statistical analysis
The data are expressed as the mean±S.E.M. Statisticalanalyses were performed with two-tailed Student's t-test for
Table 2Effect of nor-binaltrophimine (norBNI) on twitch contraction inhibited by 7-hydroxymitragynine, DAMGO, and U69593 in guinea-pig ileum
Compound (concentration) Contraction(%)inhibitedbycompound
Contraction (%) reversedby norBNI
1 nM 30 nM
7-Hydroxymitragynine (100 nM) 15.6±3.8 18.6±3.7 29.2±3.6DAMGO (100 nM) 8.7±2.9 9.4±3.4 10.3±3.2U69593 (1 μM) 10.5±5.3 13.8±6.9 118.1±8.5 a
Each value represents the mean±S.E.M. of five animals.a Pb0.01 significantly different from the values before the addition of
norBNI (Bonferroni multiple comparison test).
comparison of two groups, and by a one-way analysis ofvariance, followed by a Bonferroni multiple comparison test forcomparison of more than two groups. A P valueb0.05 wasconsidered statistically significant. ED50 values and 95%confidence limits were determined using the Litchfield–Wilcoxon method (Litchfield and Wilcoxon, 1949).
3. Results
3.1. Antinociceptive effect of 7-hydroxymitragynine in mice
7-Hydroxymitragynine (0.25–2 mg/kg, s.c.) induced dose-related antinociceptive responses in the tail-flick and hot-platetests (Fig. 2). The effect peaked at 15 and 7.5 min after injectionin the tail-flick and hot-plate tests, respectively. The ED50
values (95% confidence limits) for 7-hydroxymitragynine were0.80 mg/kg (0.48–1.33) and 0.93 mg/kg (0.59–1.45) in the tail-flick and the hot-plate tests, respectively. The vehicle did notshow any antinociceptive activity in either test.
Morphine (1.25–8 mg/kg, s.c.) produced a dose-relatedantinociceptive response with a peak effect at 30 min in bothtests (data not shown). The ED50 values (95% confidence limits)for morphine were 4.57 mg/kg (3.12–6.69) and 4.08 mg/kg(2.75–6.06) in the tail-flick and hot-plate tests, respectively.Compared to morphine on a mg/kg (μmol/kg) basis, 7-hydroxymitragynine was 5.7 (6.3) and 4.4 (4.9) times morepotent in the tail-flick and hot-plate tests, respectively (Fig. 4A, B,Table 5). Antinociception elicited by 7-hydroxymitragynineaffected behavioral responses: 2 mg/kg of 7-hydroxymitragynine
Table 4pD2 values for inhibition of electrically stimulated contraction by 7-hydroxymitragynine, DAMGO, and U69593 in guinea-pig ileum, and pA2
values of naloxone inhibition of 7-hydroxymitragynine, DAMGO, and U69593
pD2 pA2 Slope
7-Hydroxymitragynine 7.78±0.08 8.95±0.30 0.91±0.20DAMGO 7.83±0.07 8.77±0.35 1.18±0.18U69593 9.01±0.12 7.50±0.36 1.19±0.09
pD2 values are the negative logarithm of the IC50 values. The pA2 values arecalculated from parallel shifts of the curves for the agonists. Data are expressedas the mean±S.E.M. of five animals.
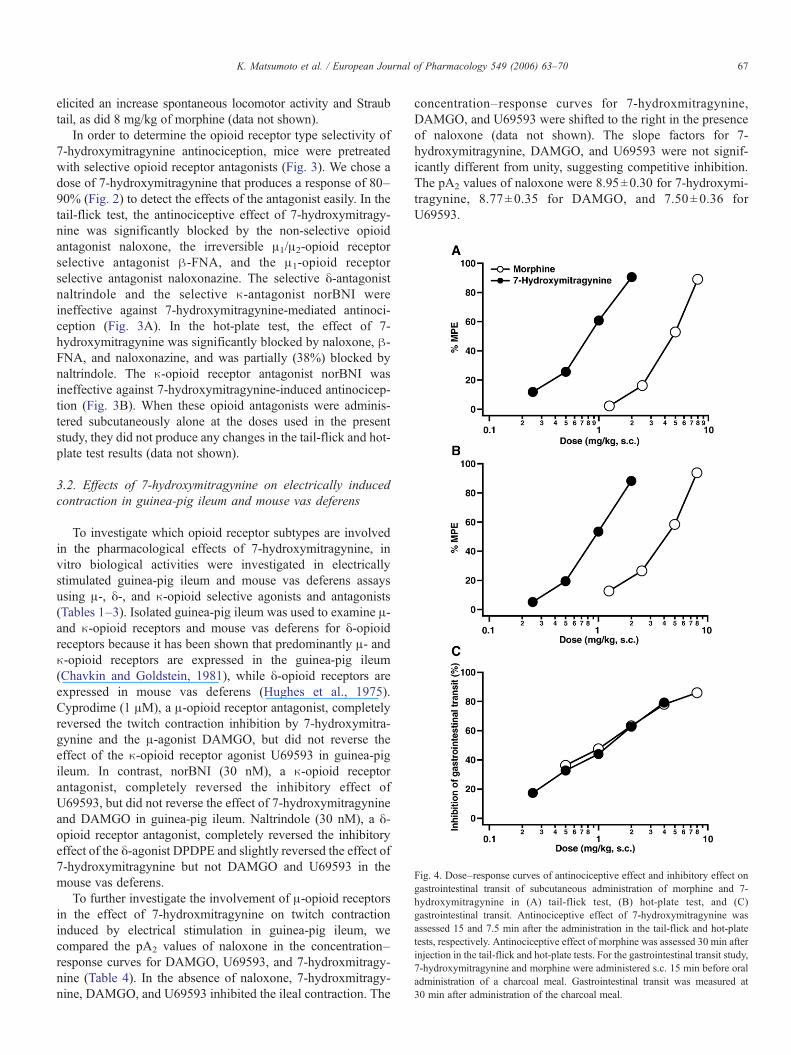
Fig. 4. Dose–response curves of antinociceptive effect and inhibitory effect ongastrointestinal transit of subcutaneous administration of morphine and 7-hydroxymitragynine in (A) tail-flick test, (B) hot-plate test, and (C)gastrointestinal transit. Antinociceptive effect of 7-hydroxymitragynine wasassessed 15 and 7.5 min after the administration in the tail-flick and hot-platetests, respectively. Antinociceptive effect of morphine was assessed 30 min afterinjection in the tail-flick and hot-plate tests. For the gastrointestinal transit study,7-hydroxymitragynine and morphine were administered s.c. 15 min before oraladministration of a charcoal meal. Gastrointestinal transit was measured at30 min after administration of the charcoal meal.
67K. Matsumoto et al. / European Journal of Pharmacology 549 (2006) 63–70
elicited an increase spontaneous locomotor activity and Straubtail, as did 8 mg/kg of morphine (data not shown).
In order to determine the opioid receptor type selectivity of7-hydroxymitragynine antinociception, mice were pretreatedwith selective opioid receptor antagonists (Fig. 3). We chose adose of 7-hydroxymitragynine that produces a response of 80–90% (Fig. 2) to detect the effects of the antagonist easily. In thetail-flick test, the antinociceptive effect of 7-hydroxymitragy-nine was significantly blocked by the non-selective opioidantagonist naloxone, the irreversible μ1/μ2-opioid receptorselective antagonist β-FNA, and the μ1-opioid receptorselective antagonist naloxonazine. The selective δ-antagonistnaltrindole and the selective κ-antagonist norBNI wereineffective against 7-hydroxymitragynine-mediated antinoci-ception (Fig. 3A). In the hot-plate test, the effect of 7-hydroxymitragynine was significantly blocked by naloxone, β-FNA, and naloxonazine, and was partially (38%) blocked bynaltrindole. The κ-opioid receptor antagonist norBNI wasineffective against 7-hydroxymitragynine-induced antinocicep-tion (Fig. 3B). When these opioid antagonists were adminis-tered subcutaneously alone at the doses used in the presentstudy, they did not produce any changes in the tail-flick and hot-plate test results (data not shown).
3.2. Effects of 7-hydroxymitragynine on electrically inducedcontraction in guinea-pig ileum and mouse vas deferens
To investigate which opioid receptor subtypes are involvedin the pharmacological effects of 7-hydroxymitragynine, invitro biological activities were investigated in electricallystimulated guinea-pig ileum and mouse vas deferens assaysusing μ-, δ-, and κ-opioid selective agonists and antagonists(Tables 1–3). Isolated guinea-pig ileum was used to examine μ-and κ-opioid receptors and mouse vas deferens for δ-opioidreceptors because it has been shown that predominantly μ- andκ-opioid receptors are expressed in the guinea-pig ileum(Chavkin and Goldstein, 1981), while δ-opioid receptors areexpressed in mouse vas deferens (Hughes et al., 1975).Cyprodime (1 μM), a μ-opioid receptor antagonist, completelyreversed the twitch contraction inhibition by 7-hydroxymitra-gynine and the μ-agonist DAMGO, but did not reverse theeffect of the κ-opioid receptor agonist U69593 in guinea-pigileum. In contrast, norBNI (30 nM), a κ-opioid receptorantagonist, completely reversed the inhibitory effect ofU69593, but did not reverse the effect of 7-hydroxymitragynineand DAMGO in guinea-pig ileum. Naltrindole (30 nM), a δ-opioid receptor antagonist, completely reversed the inhibitoryeffect of the δ-agonist DPDPE and slightly reversed the effect of7-hydroxymitragynine but not DAMGO and U69593 in themouse vas deferens.
To further investigate the involvement of μ-opioid receptorsin the effect of 7-hydroxmitragynine on twitch contractioninduced by electrical stimulation in guinea-pig ileum, wecompared the pA2 values of naloxone in the concentration–response curves for DAMGO, U69593, and 7-hydroxmitragy-nine (Table 4). In the absence of naloxone, 7-hydroxmitragy-nine, DAMGO, and U69593 inhibited the ileal contraction. The
concentration–response curves for 7-hydroxmitragynine,DAMGO, and U69593 were shifted to the right in the presenceof naloxone (data not shown). The slope factors for 7-hydroxymitragynine, DAMGO, and U69593 were not signif-icantly different from unity, suggesting competitive inhibition.The pA2 values of naloxone were 8.95±0.30 for 7-hydroxymi-tragynine, 8.77±0.35 for DAMGO, and 7.50±0.36 forU69593.
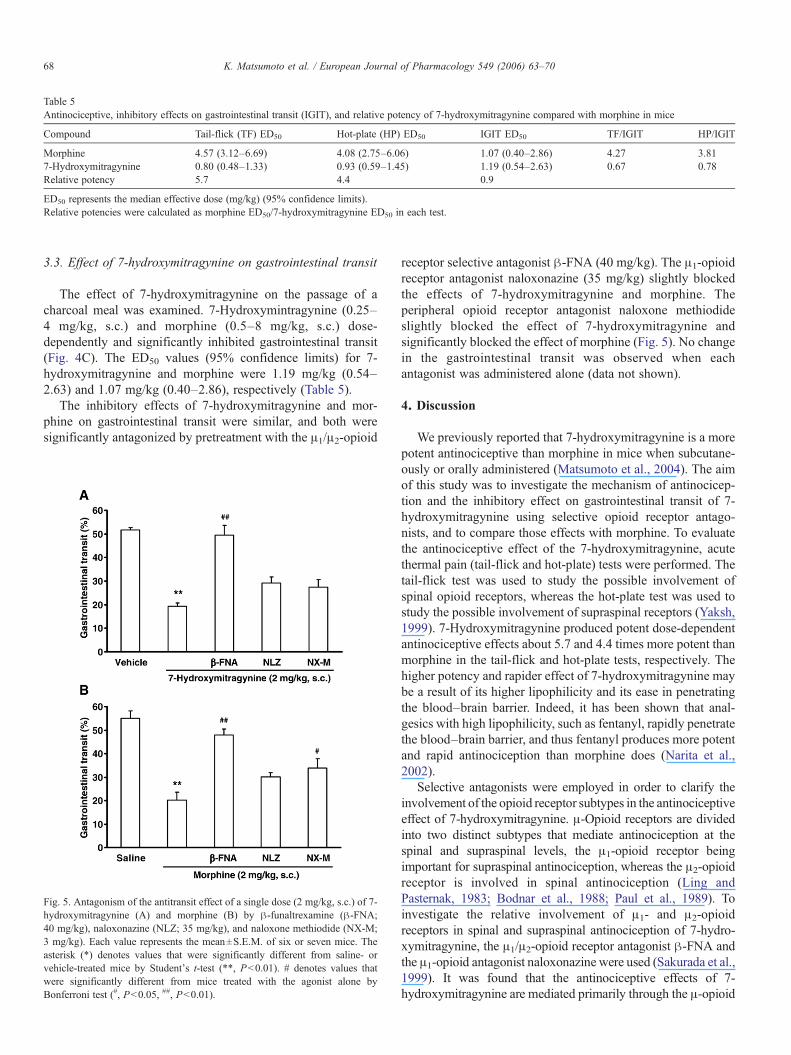
Table 5Antinociceptive, inhibitory effects on gastrointestinal transit (IGIT), and relative potency of 7-hydroxymitragynine compared with morphine in mice
Compound Tail-flick (TF) ED50 Hot-plate (HP) ED50 IGIT ED50 TF/IGIT HP/IGIT
Morphine 4.57 (3.12–6.69) 4.08 (2.75–6.06) 1.07 (0.40–2.86) 4.27 3.817-Hydroxymitragynine 0.80 (0.48–1.33) 0.93 (0.59–1.45) 1.19 (0.54–2.63) 0.67 0.78Relative potency 5.7 4.4 0.9
ED50 represents the median effective dose (mg/kg) (95% confidence limits).Relative potencies were calculated as morphine ED50/7-hydroxymitragynine ED50 in each test.
68 K. Matsumoto et al. / European Journal of Pharmacology 549 (2006) 63–70
3.3. Effect of 7-hydroxymitragynine on gastrointestinal transit
The effect of 7-hydroxymitragynine on the passage of acharcoal meal was examined. 7-Hydroxymintragynine (0.25–4 mg/kg, s.c.) and morphine (0.5–8 mg/kg, s.c.) dose-dependently and significantly inhibited gastrointestinal transit(Fig. 4C). The ED50 values (95% confidence limits) for 7-hydroxymitragynine and morphine were 1.19 mg/kg (0.54–2.63) and 1.07 mg/kg (0.40–2.86), respectively (Table 5).
The inhibitory effects of 7-hydroxymitragynine and mor-phine on gastrointestinal transit were similar, and both weresignificantly antagonized by pretreatment with the μ1/μ2-opioid
Fig. 5. Antagonism of the antitransit effect of a single dose (2 mg/kg, s.c.) of 7-hydroxymitragynine (A) and morphine (B) by β-funaltrexamine (β-FNA;40 mg/kg), naloxonazine (NLZ; 35 mg/kg), and naloxone methiodide (NX-M;3 mg/kg). Each value represents the mean±S.E.M. of six or seven mice. Theasterisk (*) denotes values that were significantly different from saline- orvehicle-treated mice by Student's t-test (**, Pb0.01). # denotes values thatwere significantly different from mice treated with the agonist alone byBonferroni test (#, Pb0.05, ##, Pb0.01).
receptor selective antagonist β-FNA (40 mg/kg). The μ1-opioidreceptor antagonist naloxonazine (35 mg/kg) slightly blockedthe effects of 7-hydroxymitragynine and morphine. Theperipheral opioid receptor antagonist naloxone methiodideslightly blocked the effect of 7-hydroxymitragynine andsignificantly blocked the effect of morphine (Fig. 5). No changein the gastrointestinal transit was observed when eachantagonist was administered alone (data not shown).
4. Discussion
We previously reported that 7-hydroxymitragynine is a morepotent antinociceptive than morphine in mice when subcutane-ously or orally administered (Matsumoto et al., 2004). The aimof this study was to investigate the mechanism of antinocicep-tion and the inhibitory effect on gastrointestinal transit of 7-hydroxymitragynine using selective opioid receptor antago-nists, and to compare those effects with morphine. To evaluatethe antinociceptive effect of the 7-hydroxymitragynine, acutethermal pain (tail-flick and hot-plate) tests were performed. Thetail-flick test was used to study the possible involvement ofspinal opioid receptors, whereas the hot-plate test was used tostudy the possible involvement of supraspinal receptors (Yaksh,1999). 7-Hydroxymitragynine produced potent dose-dependentantinociceptive effects about 5.7 and 4.4 times more potent thanmorphine in the tail-flick and hot-plate tests, respectively. Thehigher potency and rapider effect of 7-hydroxymitragynine maybe a result of its higher lipophilicity and its ease in penetratingthe blood–brain barrier. Indeed, it has been shown that anal-gesics with high lipophilicity, such as fentanyl, rapidly penetratethe blood–brain barrier, and thus fentanyl produces more potentand rapid antinociception than morphine does (Narita et al.,2002).
Selective antagonists were employed in order to clarify theinvolvement of the opioid receptor subtypes in the antinociceptiveeffect of 7-hydroxymitragynine. μ-Opioid receptors are dividedinto two distinct subtypes that mediate antinociception at thespinal and supraspinal levels, the μ1-opioid receptor beingimportant for supraspinal antinociception, whereas the μ2-opioidreceptor is involved in spinal antinociception (Ling andPasternak, 1983; Bodnar et al., 1988; Paul et al., 1989). Toinvestigate the relative involvement of μ1- and μ2-opioidreceptors in spinal and supraspinal antinociception of 7-hydro-xymitragynine, the μ1/μ2-opioid receptor antagonist β-FNA andthe μ1-opioid antagonist naloxonazine were used (Sakurada et al.,1999). It was found that the antinociceptive effects of 7-hydroxymitragynine are mediated primarily through the μ-opioid

69K. Matsumoto et al. / European Journal of Pharmacology 549 (2006) 63–70
receptors because the μ1/μ2-opioid receptor antagonist β-FNAalmost completely blocked the effect in the tail-flick and hot-platetests. In addition, naloxonazine has been shown to preferentiallyblock μ1-opioid receptors rather than μ2-opioid receptors(Sakurada et al., 1999). Naloxonazine significantly blocked theantinociceptive effect of 7-hydroxymitragynine in the tail-flickand hot-plate tests, suggesting that the antinociception induced by7-hydroxymitragynine is highly involved with the μ1-opioidreceptors. However, it was also found that the effect of 7-hydroxymitragynine was partially blocked by the δ-selectiveantagonist naltrindole in the hot-plate test, suggesting partialinvolvement of the supraspinal δ-opioid receptors. In addition,Thongpradichote et al. (1998) revealed that mitragynine, which isa main constituent ofM. speciosa and has structural similarities to7-hydroxymitragynine, has an antinociceptive activity throughthe supraspinal μ- and δ-opioid receptors. At this moment, we canonly speculate that the supraspinal δ-opioid receptors are involvedin the antinociceptive effect of 7-hydroxymitragynine, and furtherstudies are in progress to try to clarify the mechanism of itsantinociception.
Studies of the isolated guinea-pig ileum and mouse vasdeferens in electrical field stimulation preparations furtherestablished the opioid receptor specificity of the effect of 7-hydroxymitragynine. The μ-opioid selective antagonist cypro-dime completely reversed the inhibitory effect of 7-hydro-xymitragynine, confirming a mechanism of action through theμ-opioid receptors. The pA2 values of the opioid antagonistnaloxone against the inhibitory action of the μ opioid agonistDAMGO and κ opioid agonist U69593 represent the affinity ofnaloxone for μ- and κ-opioid receptors, respectively. The pA2
value of naloxone against 7-hydroxymitragyninewas very similarto that against DAMGO, but clearly different from that againstU69593. These results support the hypothesis that 7-hydroxymi-tragynine inhibited the electrically stimulated ileum contractionthrough the μ-opioid receptors. Previous radioligand bindingstudies (Takayama et al., 2002; Matsumoto et al., 2004) alsosupport the involvement of the μ-opioid receptors in the effect of7-hydroxymtragynine. Taken together, the results obtained invitro assay systems confirm that the antinociceptive effect of 7-hydroxymitragynine is mediated by μ-opioid receptors.
Opioids arewell known to inhibit gastrointestinal transit. In thecase ofmorphine, the dose required for its analgesic effect ismuchhigher than required for its constipating effects. As previouslyreported, mitragynine has a minimal effect on gastric motility atits analgesic levels (Macko et al., 1972). We investigated theinhibition of gastrointestinal transit to evaluate the constipatingeffect of 7-hydroxymitragynine and its antinociceptive effect incomparison to morphine. 7-Hydroxymitragynine inhibited gas-trointestinal transit in a dose-dependent manner, as morphine did.The ratios of ED50 values for the antinociceptive effect in the tail-flick or hot-plate test and inhibitory effect on gastrointestinaltransit (IGIT) are shown in Table 5. The IGIT ED50 value of 7-hydroxymitragynine was larger than that of its antinociceptiveED50. On the other hand, morphine significantly inhibited gastro-intestinal transit at much smaller doses than its antinociceptivedoses. The IGIT ED50 of morphine was about 4.3 and 3.8 timessmaller than those of its tail-flick ED50 and hot-plate ED50 values,
respectively. These results suggest that 7-hydroxymitragynineinduces constipation 4.9–6.4 times less potently thanmorphine atantinociceptive doses.
It appears that among opiate receptors the μ-opioid receptorsplay a prominent role in morphine-induced constipation (Royet al., 1998). We investigated the pharmacological effects of 7-hydroxymitragynine on gastrointestinal transit. The inhibitoryeffects of 7-hydroxymitragynine and morphine were markedlyblocked by β-FNA, indicating that their effects are mediated byμ-opioid receptors. It is well known that the inhibitory effectson the gut of systemic morphine administration are mediated byopioid receptors located at central and peripheral sites (Gold-berg et al., 1998; Shook et al., 1987). We investigated the effectof 7-hydroxymitragynine using centrally and peripherallyacting antagonists. The inhibitory effects of 7-hydroxymitragy-nine and morphine were slightly blocked by the centrally actingμ1-opioid antagonist naloxonazine. We also investigated theperipheral component using naloxone methiodide, which hasrestricted access to the central nervous system (Lewanowitschand Irvine, 2002). Naloxone methiodide slightly blocked theeffects of 7-hydroxymitragynine, although it moderately andsignificantly blocked the effects of morphine. These resultssuggest that 7-hydroxymitragynine inhibits gastrointestinalpropulsive activity through central and peripheral action ofthe opioid receptors. These findings allow us to speculate that 7-hydroxymitragynine interacts less with the peripheral opioidreceptors than morphine in the inhibition of gastrointestinaltransit.
Opioid analgesia has been suggested to be mediated by μ1-opioid receptors, whereas the μ2-receptors seemed to beinvolved in the inhibition of gastrointestinal transit (Pasternak,1993). The μ1/μ2-opioid receptor antagonist β-FNA signifi-cantly blocked the antinociceptive and anti-transit effect of 7-hydroxymitragynine. The μ1-opioid antagonist naloxonazinesignificantly blocked the antinociceptive effect but slightlyblocked the anti-transit effect. Taken together, it is suggestedthat antinociception and inhibition of gastrointestinal transit by7-hydroxymitragynine are mediated predominantly by μ1- andμ2-opioid receptors, respectively.
7-Hydroxymitragynine appears to be a potent μ-opioid agonistwith interesting pharmacological properties. The antinociceptiveeffect of 7-hydroxymitragynine was about 4.4–5.7 times morepotent than that of morphine. Furthermore, 7-hydroxymitragy-nine is about 4.9–6.4 times less constipating than morphine atequi-antinociceptive doses. In conclusion, 7-hydroxymitragynineshows promising characteristics as an analgesic.
Acknowledgements
This work was supported in part by Grants-in-Aid forScientific Research from the Ministry of Education, Science,Sports, and Culture of Japan and by Pfizer Nagoya Laborato-ries, Pfizer Inc, Japan. We appreciate the technical assistance ingastrointestinal transit studies by Dr. Shingo Yano and Dr.Shizuko Tsuchiya (Department of Molecular Pharmacology andPharmacotherapeutics, Graduate School of PharmaceuticalSciences, Chiba University).

70 K. Matsumoto et al. / European Journal of Pharmacology 549 (2006) 63–70
References
Bodnar, R.J., Williams, C.L., Lee, S.J., Pasternak, G.W., 1988. Role of mu 1-opiate receptors in supraspinal opiate analgesia: a microinjection study.Brain Res. 447, 25–34.
Burkill, I.H., 1935. A Dictionary of the Economic Products of the MalayPeninsula, vol. II. Crown Agents for the Colonies, London, pp. 1480–1483.
Chavkin, C., Goldstein, A., 1981. Demonstration of a specific dynorphinreceptor in guinea pig ileum myenteric plexus. Nature 291, 591–593.
Cox, B.M., Weinstock, M., 1966. The effect of analgesic drugs on the release ofacetylcholine from electrically stimulated guinea-pig ileum. Br. J.Pharmacol. Chemother. 27, 81–92.
D'Amour, F.E., Smith, D.L., 1941. A method for determining loss of painsensation. J. Pharmacol. Exp. Ther. 72, 74–79.
Goldberg, I.E., Rossi, G.C., Letchworth, S.R., Mathis, J.P., Ryan-Moro, J.,Leventhal, L., Emmel, W., Su, D., Bolan, E.A., Pasternak, G.W., 1998.Pharmacological characterization of endomorphin-1 and endomorphin-2 inmouse brain. J. Pharmacol. Exp. Ther. 286, 1007–1013.
Grewal, K.S., 1932. Observations on the pharmacology of mitragynine. J.Pharmacol. Exp. Ther. 46, 251–271.
Horie, S., Koyama, F., Takayama, H., Ishikawa, H., Aimi, N., Ponglux, D.,Matsumoto, K., Murayama, T., 2005. Indole alkaloids of a Thai medicinalherb, Mitragyna speciosa, that has opioid agonistic effect in guinea-pigileum. Planta Med. 71, 231–236.
Hughes, J., Kosterlitz, H.W., Leslie, F.M., 1975. Effect of morphine onadrenergic transmission in the mouse vas deferens. Assessment of agonistand antagonist potencies of narcotic analgesics. Br. J. Pharmacol. 53,371–381.
Jinsmaa, Y., Okada, Y., Tsuda, Y., Shiotani, K., Sasaki, Y., Ambo, A., Bryant,S.D., Lazarus, L.H., 2004. Novel 2′,6′-dimethyl-L-tyrosine-containingpyrazinone opioid mimetic mu-agonists with potent antinociceptive activityin mice. J. Pharmacol. Exp. Ther. 309, 432–438.
Lewanowitsch, T., Irvine, R.J., 2002. Naloxone methiodide reverses opioid-induced respiratory depression and analgesia without withdrawal. Eur. J.Pharmacol. 445, 61–67.
Ling, G.S., Pasternak, G.W., 1983. Spinal and supraspinal opioid analgesia inthe mouse: the role of subpopulations of opioid binding sites. Brain Res.271, 152–156.
Litchfield, J.T., Wilcoxon, F., 1949. A simplified method of evaluating dose–effect experiments. J. Pharmacol. Exp. Ther. 96, 99–113.
Macko, E., Weisbach, J.A., Douglas, B., 1972. Some observations on thepharmacology of mitragynine. Arch. Int. Pharmacodyn. Ther. 198, 145–161.
Matsumoto, K., Horie, S., Ishikawa, H., Takayama, H., Aimi, N., Ponglux, D.,Watanabe, K., 2004. Antinociceptive effect of 7-hydroxymitragynine inmice: discovery of an orally active opioid analgesic from the Thai medicinalherb Mitragyna speciosa. Life Sci. 74, 2143–2155.
Matsumoto, K., Yamamoto, L.T., Watanabe, K., Yano, S., Shan, J., Pang, P.K.,Ponglux, D., Takayama, H., Horie, S., 2005. Inhibitory effect ofmitragynine, an analgesic alkaloid from Thai herbal medicine, onneurogenic contraction of the vas deferens. Life Sci. 78, 187–194.
Matsumoto, K., Takayama, H., Ishikawa, H., Aimi, N., Ponglux, D., Watanabe,K., Horie, S., 2006. Partial agonistic effect of 9-hydroxycorynantheidine onmu-opioid receptor in the guinea-pig ileum. Life Sci. 78, 2265–2271.
McQuay, H., 1999. Opioids in pain management. Lancet 353, 2229–2232.Megens, A.A., Artois, K., Vermeire, J., Meert, T., Awouters, F.H., 1998.
Comparison of the analgesic and intestinal effects of fentanyl and morphinein rats. J. Pain Symptom Manage. 15, 253–258.
Narita, M., Imai, S., Itou, Y., Yajima, Y., Suzuki, T., 2002. Possible involvementof mu1-opioid receptors in the fentanyl- or morphine-induced antinocicep-tion at supraspinal and spinal sites. Life Sci. 70, 2341–2354.
Pasternak, G.W., 1993. Pharmacological mechanisms of opioid analgesics. Clin.Neuropharmacol. 16, 1–18.
Paul, D., Bodnar, R.J., Gistrak, M.A., Pasternak, G.W., 1989. Different mureceptor subtypes mediate spinal and supraspinal analgesia in mice. Eur. J.Pharmacol. 168, 307–314.
Ponglux, D., Wongseripipatana, S., Takayama, H., Kikuchi, M., Kurihara, M.,Kitajima, M., Aimi, N., Sakai, S., 1994. A new indole alkaloid, 7–a–hydroxy–7H–mitragynine, from Mitragyna speciosa in Thailand. PlantaMed. 60, 580–581.
Portenoy, R.K., 1996. Report from the International Association for the Study ofPain Task Force on cancer pain. J. Pain Symptom Manage. 12, 93–96.
Quock, R.M., Burkey, T.H., Varga, E., Hosohata, Y., Hosohata, K., Cowell, S.M.,Slate, C.A., Ehlert, F.J., Roeske, W.R., Yamamura, H.I., 1999. The delta-opioid receptor: molecular pharmacology, signal transduction, and thedetermination of drug efficacy. Pharmacol. Rev. 51, 503–532.
Roy, S., Liu, H.C., Loh, H.H., 1998. Mu-Opioid receptor-knockout mice: therole of mu-opioid receptor in gastrointestinal transit. Brain Res. Mol. BrainRes. 56, 281–283.
Sakurada, S., Zadina, J.E., Kastin, A.J., Katsuyama, S., Fujimura, T.,Murayama, K., Yuki, M., Ueda, H., Sakurada, T., 1999. Differentialinvolvement of mu-opioid receptor subtypes in endomorphin-1- and -2-induced antinociception. Eur. J. Pharmacol. 372, 25–30.
Schug, S.A., Zech, D., Grond, S., 1992. Adverse effects of systemic opioidanalgesics. Drug Safety 7, 200–213.
Shook, J.E., Pelton, J.T., Hruby, V.J., Burks, T.F., 1987. Peptide opioidantagonist separates peripheral and central opioid antitransit effects. J.Pharmacol. Exp. Ther. 243, 492–500.
Suwanlert, S., 1975. A study of kratom eaters in Thailand. Bull. Narc. 27,21–27.
Takayama, H., 2004. Chemistry and pharmacology of analgesic indole alkaloidsfrom the rubiaceous plant, Mitragyna speciosa. Chem. Pharm. Bull. 52,916–928.
Takayama, H., Ishikawa, H., Kurihara, M., Kitajima, M., Aimi, N., Ponglux, D.,Koyama, F., Matsumoto, K., Moriyama, T., Yamamoto, L.T., Watanabe, K.,Murayama, T., Horie, S., 2002. Studies on the synthesis and opioid agonisticactivities of mitragynine-related indole alkaloids: discovery of opioidagonists structurally different from other opioid ligands. J. Med. Chem. 45,1949–1956.
Thongpradichote, S., Matsumoto, K., Thoda, M., Takayama, H., Aimi, N.,Sakai, S., Watanabe, H., 1998. Identification of opioid receptor subtypes inantinociceptive actions of supraspinally-administered mitragynine in mice.Life Sci. 62, 1371–1378.
Watanabe, K., Yano, S., Horie, S., Yamamoto, L.T., 1997. Inhibitory effect ofmitragynine, an alkaloid with analgesic effect from Thai medicinal plantMitragyna speciosa, on electrically stimulated contraction of isolatedguinea-pig ileum through the opioid receptor. Life Sci. 60, 933–942.
Yaksh, T.L., 1999. Spinal systems and pain processing: development of novelanalgesic drugs with mechanistically defined models. Trends Pharmacol.Sci. 20, 329–337.
Yamamoto, L.T., Horie, S., Takayama, H., Aimi, N., Sakai, S., Yano, S., Shan,J., Pang, P.K.T., Ponglux, D., Watanabe, K., 1999. Opioid receptor agonisticcharacteristics of mitragynine pseudoindoxyl in comparison with mitragy-nine derived from Thai medicinal plant Mitragyna speciosa. Gen.Pharmacol. 33, 73–81.
Related Documents











