ISSN 1018-5593 ECOSYSTEMS RESEARCH REPORT 8 Introduced Species in European Coastal Waters lia doryphora, Sargassum mutic um aponica, Sphaerotrichia dir pinnatifida, Eucheilota p¿ franciscana, Gonionem us lineata, Oculin a ricolor, Cra gigas, %della inopina EUROPEAN COMMISSION EUR 15309 EN

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ISSN 1018-5593
ECOSYSTEMS RESEARCH REPORT 8
Introduced Species in
European Coastal Waters
lia doryphora, Sargassum mutic um aponica, Sphaerotrichia dir pinnatifida, Eucheilota p¿ franciscana, G o n i o n e m u s lineata, O c u l i n a ricolor, Cra gigas,
%della inopina
EUROPEAN COMMISSION EUR 15309 EN


s European Commission Directorate-General XII for Science, Research and Development
INTRODUCED SPECIES IN
EUROPEAN COASTAL WATERS
Report on an international Workshop organised jointly by the
ENVIRONMENT PROGRAMME of
DGXÏÏ of the European Commission
and the
Commission Internationale pour l'Exploration Scientifique de la Mer Méditerranée (CIESM) a <¿j>t* J( l/\ Qu *î
(Monaco, 2-3 March 19Ô9Í
Edited by
PARL EUROP. Biblioth.
N.C.&ΪΟΪΙόβ CI.
CF. Boudouresque, F. Briand and C. Nolan
ENVIRONMENT Commission Internationale pour
Γ Exploration Scientifique de la Mer Méditerranée

Published by the EUROPEAN COMMISSION
Directorate-General XIII Telecommunications, Information Market and Exploitation of Research
L-2920 Luxembourg
Introduced species in European Coastal Waters
Edited by: CF. Boudouresque, Faculté des Sciences de Luminy Lab. de Biologie Marine et d'Ecologie du Benthos Route Léon Lachamps, 70/case 901 F - 13288 Marseille Cedex 9
F. Briand, Commission Internationale pour l'Exploration Scientifique de la Méditerranée 16, bd de Suisse MC - 98000 Monte-Carlo
C. Nolan Commission Européenne DG XII - Science, Recherche et Développement Β - 1049 Bruxelles
This is Report 8 in the Ecosystems Research Report Series of the Environmental Research Programme of the Commission of the European Communities, Directorate-General for Science, Research and Development.
For more information concerning this series, please contact: C. NOLAN CEC- DGXII/D-1 200, rue de la Loi Β - 1049 Brussels
No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without indicating the abovementioned references.
LEGAL NOTICE
Neither the Commission of the European Communities nor any person acting on behalf of the Commission is responsible for the use which might be made of
the following information
Cataloguing data can be found at the end of this publication
Luxembourg: Office for Official Publication of the European Communities, 1994
ISBN 92-826-6727-8
© ECSC-EC-EAEC, Brussels· Luxembourg, 1994
Printed in Belgium

— IH —
Report on EEC/DG ΧΠ - CIESM Workshop on Introduced Species in European Coastal Waters (Monaco, 2-3 March 1993)
C.F. Boudouresque, F. Briand and C. Nolan (eds.)
Summary
This report, with scientific articles in English and French, is based on an expert workshop jointly organized by EEC/DGXII and CIESM. It illustrates a spectacular increase of accidental species introductions in European coastal waters in the last 20 years. Some of these are clearly linked with recent developments in aquaculture, marine transport (ballast waters), or aquariology, others with anthropic and / or climatic changes, others yet with the deepening of the Suez Canal. Numerous examples taken from various compartments of the biota - algae and invertebrates in particular - are provided : they illustrate the fragmentary nature of existing knowledge on this issue, as well as its complex and broad implications in the technical, economic and legal domains.
Compared to the large body of knowledge gathered on terrestrial species introductions, data on the dynamics of coastal species introductions remain far too scarce for the development of general predictive models. The participants called for the launch of an European research programme in order to provide reliable tools for the proper management and monitoring of this problem. Beyond the constitution of an European data bank on introduced species and receptive habitats, identified research priorities include the dynamics of these introductions, their history, their mode of introduction and dispersal, their biological and socio-economic impact (both negative and positive) on local ecosystems. Specific attention is drawn to the need (a) for systematic investigations and monitoring of ballast waters, suspected to play a major role in transporting marine and brackish species across wide distances, (b) for the genetic characterisation of aquaculture yş wild populations, and (c) for investigating possible relations between pollution level and vulnerability to biological invasions.
The report stresses that existing legal international instruments, such as the EC Habitats Directive, the Helsinki Convention or the Barcelona Convention, remain largely silent on this issue, and must be rapidly strengthened. In particular, the participants concluded that further legislation, based on the "precautionary principle", should be drafted to control and limit the transport of live species, which may be introduced deliberately or not into the frontiers of the European Community.

— IV —
Publications to date in the Ecosystems Research Reports Series include
1. Decomposition and accumulation of organic matter in terrestrial ecosystems : research priorities and approaches (N. Van Breeman, Ed.)
2. The N1TREX Project : Nitrogen saturation experiments (N.B. Dise & R.F. Wright, Eds.)
3. The CORE Project : Regulation of organic matter and nutrient turnover in the soil comportment of European Forests (M. Raubuch, Ed.)
4. Experimental manipulations of biota and biogeochemical cycling in ecosystems approach and methodologies - Findings (L. Rasmussen, T. Brydges & P. Mathy, Eds.)
5. Effects of forest fires on Mediterranean Ecosystems (L. Trabaud, R. Prodam & P. Mathy, Eds.)
6. Design and Execution of experiments on COj enrichment (E.D. Schulze and H.A. Mooney, Eds.)
7. The EXMAN Project : Experimental manipulation of forest ecosystems in Europe (C. Beier & L. Rasmussen, Eds.)
8. Introduced species in European coastal waters (CF. Boudouresque, F. Briand & C. Nolan, Eds.)
9. ALPE-1 : Acidification of Mountain Lakes. Paleolimnology and ecology (Wathne, B.M. & Patrick, S.T., Eds.)

— ν—
CONTENTS
Summary iii
Introduced species in European coastal waters 1 Canice Nolan
Species Introductions in coastal waters of Europe : A call for action 4 Frédéric Briand
Les espèces introduites dans les eaux côtières d'Europe et de Méditerranée : 8 Etat de la question et conséquences
Charles F. Boudouresque
The introduction of non-indigenous marine species to Europe : 25 Planktonic species
Gerald T. Boaich
Introduction of marine benthic algae into Atlantic European waters 28 William F. Farnham
Les macrophytes marins introduits en Méditerranée : biogéographie 37 Maria A. Ribera
Introduced invertebrates : examples of success and nuisance in the 44 European Atlantic and in the Mediterranean
Helmut Zibrowius
Réflexions sur les problèmes d'introduction de mollusques 50 Henri Grizel
Caractéristiques génétiques et capacité d'invasion chez les invertébrés 56 dans les eaux littorales et les lagunes méditerranéennes
Anna Occhipinti Ambrogi
Lessepsian migration - Biological invasion of the Mediterranean 57 Bella S. Galii
Non-indigenous species in the Baltic Sea 60 Erkki Leppükoski
Concerns and activities of the ICES Working Group on introductions 67 and transfers of marine organisms
Inger Wallentinus
The introduction of exotic species and the law 75 Cyrille De Klemm
Bibliography 82


Introduced species in European coastal waters
Canice NOLAN
Commission of the European Communities DG XII Science, Research and Development
Environment Research Programme 200, rue de la Loi - 1049 BRUSSELS (B)
Rates of introductions of non-native species to European coastal waters has been increasing dramatically in recent years. It is estimated that only a small fraction of all introductions are successful in establishing new populations and that only a small fraction of these successes are harmful. Unfortunately the consequences can then be devastating. Introductions do give rise to concern on a global level, witness :
a) the creation of a special working group in ICES to examine the problem, to formulate codes of practice and to review their effectiveness;
b) the implementation of legislation in Australia to protect coastal waters against introductions;
c) the introduction of legislation in the U.S. to protect against further introductions of the shellfish Dreissena polymorpha and other species;
d) the activation of the EC Emergency Response Task Force by Italy, France and Spain following the introduction of Caulerpa taxifolia into the Mediterranean;
e) the recent severe impact of the recently introduced ctenophore Mnemiopsis leidyi on Black Sea ecosystems and fisheries.
In 1991, the European Commission (EC) was requested to help finance the introduction of a non-native edible kelp (Undaria spp.) to the French Channel coast for commercial exploitation. One of the justifications used was that this would provide employment to the local communities, partly in compensation for the continued decline of the local fishing industries.
The following year, the EC received formal requests from the French, Spanish, and Italian governments, for support in combatting the spread of tropical alga, Caulerpa taxifolia, along their coasts. The non-native alga had apparently been accidentally introduced to Mediterranean coastal waters and fears has been expressed that it would proliferate at the expense of the valuable Posidonia oceanica seagrass ecosystems. An international
. programme was launched in response to these latter requests, with the support of the LIFE Programme of DG XI (responsible for environmental policy etc.) and the ENVIRONMENT Programme of DG XII (responsible for research and development) of the Commission.
These apparently contradictory actions do not imply any inconsistency in EC policy but they illustrate the complex issues involved in relation to the introduction of non-native species into European coastal waters. They also illustrate the need for a sound scientific basis in environmental policy-making and the necessity of integrating environmental considerations into other aspects of policy-making e.g. agriculture, fisheries, transport.

Many examples of introduced species exist, from the Mediterranean, Black and Baltic Seas to the Atlantic coasts. There are instances of accidental, deliberate, and natural introductions. Some are benign, others are clearly detrimental to ecosystems. In many ways, non-native species are just as foreign to an ecosystem as are genetically-modified organisms. It could even be argued that the latter are merely a special class of the former. Yet, whereas very strict regulations exist to control introductions of genetically-modified organisms into the environment, with the exception of some ICES guidelines (page xxx) and two EC Directives', there are few general principles, at European-level, applied in governing the introduction of non-native species.
At the 33rd CIESM (International Commission for the Scientific Exploration of the Mediterranean) Congress held in Trieste in 1992, the question of introduced species in the Mediterranean Sea was the subject of a well-attended session. This session concluded that, except for the relatively well documented lessepsian species, our knowledge remains quite fragmentary about the modes of introduction, the rate and patterns of dispersal of introduced species in the Mediterranean marine realm. Little is known about their impact on indigenous species and ecosystems and it was recommended that CIESM quickly establish a task force on this subject with a view to explore priority areas and provide impartial advice on scientific and legal issues to the decision-making level.
The need for action was further highlighted by the Science Panel of the EC concerted action "Change in Coastal Ecosystems" during their first meeting in Brussels in November 1992, who recommended that an EC working group be established on this theme.
Because of this mutual concern and the recognition of a need for action, both the ENVIRONMENT Programme of DG XII (EC) and CIESM, organised a workshop on the theme of introduced species in European coastal waters in Monaco on 2-3 March 1993.
The meeting was attended by xxx European experts, including representatives of the EC, CIESM and ICES. Although unable to participate actively, others provided additional material (de Klemm, Galii, Leppäkoski). The objective of the workshop was to summarise the state-of-the-science in this area, to define, in a report, the critical priority issues on this question and to formulate an agenda for action at both theoretical and practical levels. It was intended that the report of the meeting could form a basis for research to be considered as eligible for EC support in the future. This report was produced as a result of the meeting. It documents the history, rates, modes and consequences of introductions into the seas around Europe. It also reviews the legislative tools available to control introductions and identifies research needed to underpin a more effective implementation of policy in relation to both accidental and deliberate introductions.
Among the research needs identified during the meeting are :
a) Testing the hypothesis that stressed ecosystems (e.g. through pollution or physical disturbance) with low biodiversity are particularly vulnerable to invasions by introduced species.
b) Development of quarantine/sterilisation techniques to destroy viable organisms in ballast and bilge water, and laboratory and aquaria outflows.
c) Improving knowledge of the ecology and dynamics of populations of introduced species including aspects such as competition for resources such as space and nutrients.
d) Continuing work on the genetics of populations in aquaculture e.g. development of sterile polyploid strains.

- 3 -
e) Elucidating the pathology of accidentally introduced parasites etc. f) Development of techniques to evaluate the economic case for and against introducing
species which have commercial benefits but which carry a risk of disrupting or even destroying existing ecosystems.
In the text, the authors show that there are also needs to improve our knowledge of taxonomy and the mapping of existing species distributions. The rates of introductions of exotic species to European coastal waters have been increasing dramatically over the last three decades. Most reports of new sightings came from the vicinity of marine laboratories, often conveniently located near towns and ports. Whether this is a "real" phenomenon (because ecosystems near towns and ports are doubly vulnerable, being stressed through pollution or disturbance and being subject to increased traffic and opportunity) or not is not known. More work is needed on monitoring remote areas to (i) test the hypothesis above and (ii) to help measure the dispersal of existing introduced organisms.
As illustrated in the article by Cyrille de Klemm, many international agreements and conventions exist which could be used and, many would argue, should be used to protect our coastal resources from the risks posed by accidental and deliberate introductions. This requires a concerted effort at all levels, not just in education and research but also in policy formulation and implementation. This effort should not be restricted to Europe and its neighbours - it is a global issue, recognised in many international agreements, and with implications on a global scale.
Although the report is focused primarily on research needs in this area, I feel that its scope is of sufficiently broad interest to encourage a wider reflection on the subject of introduced species; on the need for stimulating more cooperation among scientists in fields as diverse as ecology and genetics and also between the scientific and the policy-making communities. The contents of this report reflect the views of the authors and of the meeting; they do not necessarily reflect the views of the Commission of the European Communities.
Commission of the European Communities
Council Directive 91/67/EEC of 28 January 1991 concerning the health conditions governing the placing on the market of aquaculture animals and products
Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora.

- 4 -
Species introductions in the coastal waters of Europe A call for action
Frédéric BRIAND
Director General, Commission Internationale pour l'Exploration Scientifique de la mer Méditerranée
As this report makes plain, the problem of species introductions into coastal and offshore waters is not new. Developed centuries ago, often deliberately, along with the expansion of shipping trade and the movements of commercial fishery products, it was viewed generally as a welcome enrichment of the local fauna and flora. It is only in recent years that our perception did shift significantly as species introductions became more frequent, more widespread, and their unwanted impact on ecosystem stability, genetic diversity, fisheries and aquaculture more widely recognised.
The papers assembled in this report well illustrate that in the marine /coastal realm, where changes are less immediately obvious than on the continent, the increased frequency and spread of introductions must be taken seriously. Two recent cases in particular, both linked with the accidental transport of marine invertebrate larvae in the ballast waters of vessels, have struck the imagination for the large socio-economic costs which they have generated.
The first instance concerns the ctenophore Mnemiopsis mccradyi. Introduced accidentally a few years ago from the atlantic coast of north-America, this effective predator quickly proliferated - reaching densities above 30 individuals/m3 " into the troubled Black Sea ecosystem where it is credited for the sudden collapse of the pelagic fisheries. Recent reports attest that this euryhaline / eurythermal species has now crossed the Bosphorus, and is currently expanding along the mediterranean coast of Turkey (KIDEYS & NIERMANN 1993).
The second instance refers to the zebra mussel Dreissena potymorpha. Around 1986 it was introduced accidentally from Europe (CARLTON 1993) into the Laurentian Great Lakes, where it caused enormous economic damages, estimated at 5 billion US dollars, due to its rapid colonization of water pipes and boat hulls. A special Act of the US Congress resulted, establishing a program to monitor nonindigenous aquatic nuisance species. These examples, among many others, easily demonstrate the potential threat of introducing exotic coastal species. Now that the dangers - of ballast water transport notably - are more clearly recognised (see CARLTON 1985, 1989, 1991), Europe would do well in setting up a properly adapted monitoring and prevention program.
Of course, as is well known, insurance looks costly only before the accident. In the case of species introductions, considering the poor state of systematic knowledge, the absence of monitoring on a regional scale and the weakness of existing legislation, the time has come indeed for concerned european agencies to take vigourous, concerted actions. The experts gathered for this workshop have identified clear priorities in this regard, which I summarize

- 5 -
briefly below. The interested reader is referred of course to the main papers for a detailed development.
A - Priorities for research:
Without a sound scientific grasp of the issue, there can be no proper management of species introductions. Unfortunately our knowledge of the biology, physiology, genetics and population dynamics of introduced species remains rudimentary. When they exist, reference lists on coastal introduced species are fragmentary, available only for certain taxa and for limited geographic areas. Further the record on the chronology, the geographic distribution, and the rates of species invasions in european coastal waters is clearly insufficient to allow the development of reliable predictive models on rates of dispersal. Finally there must be systematic investigations of vulnerable habitats, of ballast waters, of natural predators. It is only on this basis that Europe shall be able to build the future region-wide monitoring and prevention program which it deserves.
A clear priority is to fill those gaps quickly. Among the research objectives identified by the workshop participants, the following rank particularly high :
o launch systematic, comparative investigations on the physiology, ecology and dynamics of introduced species in their area of introduction vs. their area of origin;
o carry out genetic characterisation of aquaculture v§ wild populations (using genetic markers, and systematically testing for sexual incompatibility between introduced species and closely related natives);
o search for potential indicators of the vulnerability of coastal habitats to biological invasions;
o analyse, on the basis of chronological records and up-to-date water circulation models, the actual dispersal rates of introduced species; develop reliable predictive models;
o quantify the overall impact of species introductions, taking into account integrated environmental costs and socio-economic effects (both positive and negative);
o develop suitable programs for the valorization of introduced species.
Β - Priorities for action :
1. Monitoring
o initiate rapidly, on a pilot basis and a reduced geographic area in the first phase, an international collaborative program to survey the rate of invasion of certain potentially harmful species; it is recommended that this program focuses at first on highly recognisable species such as the ctenophore Mnemiopsis and the jellyfish Rhopilema in the Eastern Mediterranean, upon

2. Training
the model already supported by the LIFE program to survey the extension of Caulerpa taxifolia.
develop a European data bank on introduced species and receptive habitats; this data base shall be easily accessible, and include all relevant parameters such as distribution maps, chronology of invasion, known natural enemies in areas of origin, etc ...; further it shall be designed in a way that would allow its nesting within an enlarged European CORINE data base.
engage, in a uniform manner, regular surveys of the most vulnerable coastal habitats of Europe (areas of low diversity; highly degraded environments, simplified or highly fluctuating ecosystems ...)
set up, in collaboration with the International Maritime Organization, and with experienced foreign agencies (such as NOAA in the USA) an international monitoring programme of ballast waters; this will include the development of reliable technologies for the elimination of accompanying species.
generally rehabilitate the domains of taxonomy and systematics, as these are the fundamental building blocks to understand and properly manage ecosystems; this will involve, among other things, a revamping of the training and career opportunities of taxonomists
include coverage of species introductions in high school and university curricula;
support and develop adequate exchange programs for coastal and aquaculture scientists on both sides of the ocean, so as to promote the standardisation of research, monitoring and prevention technologies.
3. Legislation
This aspect cannot be underestimated. Indeed, if not accompanied by a proper enforcement of existing legislation, the best research and monitoring programs shall have little chance to restrain the introduction of harmful species. Yet, legal enforcement and the application of stronger penalties to violators - while indispensable measures - still appear very insufficient. As pointed out in the paper by De Klemm, very few texts directly address the subject in the national and EC legislation; when they exist these are largely unadapted to the scope of the problem, and lack effective mechanisms of enforcement. What is very much needed is a revamped legislation. To this end we recommend that the following steps be taken without delay :
o strengthen the relevant existing legislation, beginning with the EC Habitat Directive, the Barcelona Convention, and the Helsinki Convention :
by building upon the excellent ICES Codes of aquaculture practices,

- 7 -
and introducing new provisions in the existing texts, to address explicitly the matters of accidental introductions (ballast waters, aquariology .. .).
To this end the formation of an advisory group, including both legal and biological experts, appears highly advisable;
o revise the EC Directive of 2 January 1991 on sanitary policy, which is wholly unadapted to the problem of introduced species.
4. Public awareness
Finally one must not neglect the importance of raising public awareness on the ecological risks associated with deliberate species introductions. Well adjusted sensibilisation programs must be developed on this issue :
in the public at large, with a particular focus on certain target groups such as aquariologists, economic actors in the domains of maritime transport, aquaculture, aquariology, ...;
in the scientific community, o by facilitating the exchange of information through the venue of
international conferences on the subject; o and to promote the adoption by laboratories of more precautionary
measures to prevent the accidental release into the environment of exotic species - and particularly unicellular organisms - acquired for experimental reasons.

Les espèces introduites dans les eaux côtières d'Europe et de Méditerranée : Etat de la question et conséquences
Charles F. BOUDOURESQUE
Laboratoire de Biologie marine et d'Ecologie du Benthos (LBMEB; EA 876), Faculté des Sciences de Luminy, 13288 Marseille Cedex 9, Fiance.
RESUME
La probabilité qu'une espèce nouvelle pour une région soit introduite se fonde sur un ensemble de critères qui ne sont que rarement réunis : discontinuité géographique, nouvelle station très ponctuelle, cohérence de la cinétique d'expansion, tendance à la pullulation initiale, proximité d'une source d'introduction, variabilité génétique faible. Certaines espèces apparaissent comme certainement introduites; d'autres sont perçues comme probablement introduites alors qu'elles ne sont pas encore connues dans leur région d'origine; quelques-unes enfin, d'abord considérées comme introduites, ne le sont plus aujourd'hui. Les causes d'introduction sont le fouling sur les coques des navires (dont l'importance a sans doute diminué de nos jours), le déballastage des navires, les appâts pour la pêche, la recherche scientifique, l'aquariologie et surtout l'aquaculture. En Méditerranée, il convient d'y ajouter le percement du canal de Suez, responsable de l'invasion biologique marine la plus massive à l'échelle mondiale. La faible biodiversité du milieu d'accueil constitue sans doute le principal facteur explicatif du succès initial d'une introduction; c'est le cas de la Méditerranée orientale dans son ensemble (migrants lessepsiens), des milieux lagunaires (où se pratique l'aquaculture), des ports et autres milieux pollués (ou arrivent les espèces du fouling). On assiste actuellement à une formidable accélération du rythme des introductions d'espèces : en Méditerranée, près de 60% des invertébrés introduits (non compris les espèces lessepsiennes) l'ont été postérieurement à 1970.
Il n'est pas rare que les espèces introduites présentent une phase de prolifération initiale spectaculaire, puis que leur abondance décline; il ne s'agit pas toutefois d'une règle générale. La plupart des espèces introduites ne semblent pas avoir eu d'impact sur les espèces ou les communautés indigènes; il s'agit toutefois d'une conclusion en grande partie subjective en l'absence de recherches précises sur la question; en outre, l'impact des espèces introduites ne doit plus être abordé au cas par cas, mais dans sa globalité : de nombreuses espèces introduites coexistent en effet dans certains biotopes, et leur impact peut être synergique. Dans un certain nombre de cas, un impact des espèces introduites a été mis en évidence : compétition avec une espèce indigène dont la niche est similaire, glissement de la niche d'une espèce indigène, élimination totale des espèces concurrentes et modification profonde de l'ensemble de l'écosystème. Par ailleurs, l'introduction de gènes, à partir de populations de la même espèce qu'une espèce indigène, mais génétiquement différentes, ou d'espèces voisines susceptibles de s'hybrider avec l'espèce indigène, est très probable, bien qu'encore peu documentée. Les espèces introduites génèrent en outre des nuisances : compétition avec des espèces exploitées, gêne à la pêche, à l'aquaculture, au tourisme, à la navigation et fouling.

Les introductions d'espèces, que l'on désigne parfois sous le nom de pollution biologique, constituent peut-être l'un des problèmes écologiques majeurs du prochain siècle : il s'agit en effet d'un phénomène irréversible (contrairement à d'autres types de pollution), n'épargnant aucun sanctuaire (Parcs Nationaux par exemple) et susceptible à long terme d'homogénéiser les peuplements et les paysages à l'échelle planétaire. Un effort de recherche considérable est nécessaire, d'une part pour mieux cerner l'impact réel des introductions, et d'autre part pour établir les bases scientifiques d'une limitation de ces introductions.
INTRODUCTION
Les introductions d'espèces, c'est-à-dire l'apparition d'une espèce dans une région où elle n'était pas présente auparavant, peuvent se faire par dispersion marginale (extension d'aire de proche en proche : "marginal dispersal"; CRISP, 1958) ou par saut (une discontinuité spatiale existe entre son aire d'origine et sa nouvelle station : "remote dispersal"). L'extension d'aire de proche en proche n'implique pas l'action de l'homme; elle ne fera donc pas l'objet de la présente revue; en outre, chaque espèce occupe normalement depuis longtemps la totalité de l'aire qui lui est accessible compte-tenu de ses exigences écologiques et des barrières géographiques existantes; tout au plus peut-on assister à des fluctuations périodiques (avancée ou recul) aux frontières de son aire de répartition, en liaison par exemple avec de petits épisodes climatiques. FISCHER-PIETTE (1959, 1963) signale de telles fluctuations d'aire chez Fucus spiralis (Fucophyceae) etLittorina saxatilis (gastrópode), au Sud du Portugal; l'arrivée de Laminaria ochroleuca (Fucophyceae) en Grande-Bretagne, à partir des côtes continentales, pourrait être due à une légère élévation de la température de l'eau (PARKE, 1948; FARNHAM, 1980).
A l'échelle des temps géologiques, il s'agit d'un phénomène naturel qui a joué un rôle important dans l'histoire du peuplement de chaque région et dans la spéciation. Un archipel comme les Açores, isolé dès l'origine, a ainsi été peuplé progressivement par des espèces arrivant plus ou moins accidentellement, la plupart à partir des côtes européennes (FELDMANN, 1946). Quelques-unes ont déjà évolué (spéciation) pour donner naissance à une espèce endémique, par exemple le Codium elisabethae (Chlorophyta) à partir de son ancêtre européen probable, C. bursa (SCHMIDT, 1931; FELDMANN, 1946). De même, après les crises messiniennes (5 à 6 millions d'années), qui ont partiellement détruit la faune et la flore méditerranéennes, la Méditerranée a été repeuplée par des espèces en provenance de l'Atlantique (par le détroit de Gibraltar) et de l'Indo-Pacifique (POR, 1990). Chez Cladophora albida (Chlorophyta), espèce à répartition actuelle très large, l'hybridation DNA-DNA (HOEK et al., 1990) montre des divergences relativement récentes entre les populations des deux côtés de l'Atlantique (2-4 Ma), qui peuvent être interprétées soit comme la date de mise en place d'une barrière au flux génique, soit comme la date de franchissement de l'Atlantique, dans un sens ou dans l'autre. Plus près de nous, les glaciations du Quaternaire ont contribué à étendre en Méditerranée l'aire de répartition de certaines espèces nordiques. C'est peut-être le cas par exemple de Fucus virsoides (Fucophyceae), très proche (FORTI, 1931), sinon identique (SAUVAGEAU, 1908a) de Fucus spiralis de l'Atlantique, et cantonné à la mer Adriatique (BOUDOURESQUE et al., 1990) et de Littorina saxatilis (gastropode), présent en Haute Adriatique et dans le golfe de Gabès (Tunisie). Le gastropode Littorina littorea, natif des côtes atlantiques d'Europe, a pu traverser l'Atlantique et coloniser le nord-est de l'Amérique du Nord au cours de "l'optimum climatique" de l'Holocène (il y a environ 5000 ans), avec les navigateurs danois (il y a environ 1000 ans), ou plus récemment encore (KRAEUTER, 1976; CARLTON et al., 1982; ZIBROWIUS, 1991).

- 10-
POR (1978) n'exclut pas que des espèces de Mer Rouge aient pu entrer en Méditerranée il y a environ deux millénaires, par les canaux percés à l'époque des pharaons entre la Mer Rouge et la Méditerranée : Acanthophora najadiformis (Rhodophyta) et Pinctada radiata (mollusque bivalve) pourraient être dans ce cas.
Des introductions plus récentes sont possibles. C'est ainsi que les Cladophora sericea des deux côtés de l'Atlantique sont identiques (hybridation DNA-DNA) (HOEK et al., 1990), de telle sorte que l'on peut penser qu'il y a eu soit brassage génétique (permanent ou intermittent), soit plus probablement introduction d'un des côtés de l'Atlantique, introduction antérieure à l'ère linnéenne. L'huître portugaise Crassostrea ungulata et l'huître japonaise C. gigas sont très proches morphologiquement et donnent des hybrides viables; certains auteurs considèrent qu'il s'agit de la même espèce, et que C. angulata correspond à une introduction d'extrême Orient au Portugal, au XVIème siècle (ZIBROWIUS, 1991); les différences morphologiques et physiologiques proviendraient du fait que les populations de C. angulata, fondées par un petit nombre d'individus, ne représenteraient qu'une petite partie de la variabilité génétique de C. gigas. De même, le scléractinaire Oculina patagónica, originaire semble-t-il de l'Atlantique tempéré sud-américain, pourrait avoir été introduit en Méditerranée (Sud-Est de l'Espagne) au 16ème ou au 17ème siècle (ZIBROWIUS, 1991).
La période linnéenne est évidemment mieux documentée. Il est clair toutefois qu'il faut attendre le XXème siècle pour que les connaissances sur la systématique et sur la biogéographie permettent d'avancer qu'une espèce est probablement introduite. Encore aujourd'hui, dans des genres dont la systématique est très complexe, ou bien quand il s'agit d'espèces de petite taille, passant facilement inaperçues, la découverte d'une station éloignée des plus proches stations connues est interprétée (faute de mieux) comme une contribution à la connaissance de l'aire de répartition plutôt que comme une introduction ou une extension d'aire. La génétique moléculaire permettra parfois, dans l'avenir, de résoudre ce genre de problèmes.
Les introductions naturelles, comme les extinctions naturelles d'espèces au cours des temps géologiques, se font à un rythme très lent. Pour les extinctions naturelles, on a avancé le chiffre d'une extinction tous les mille ans. Avec l'intervention de l'homme, le rythme change. Pour les extinctions d'espèces liées à l'homme, on a avancé le chiffre d'une espèce par jour.
Π convient de rattacher au phénomène général des introductions d'espèces le cas particulier des introductions de gènes. En introduisant dans une région où une espèce existe des souches de la même espèce, mais provenant d'une région éloignée, on peut y apporter des gènes qui normalement n'y existent pas; dans le cas d'espèces à large répartition, des barrières géographiques s'opposent en effet généralement au gene flow et au brassage génétique; un processus de spéciation peut ainsi s'amorcer. Les organismes génétiquement modifiés issus de l'aquaculture (y compris les polyploïdes) entrent dans cette catégorie.

11
Fig. 1 : Chronologie de l'expansion de Sargassum muticum (Fucophyceae) le long des côtes atlantiques d'Europe (d'après VERLAQUE, COMM. pers.). A partir de la date (hypothétique) d'introduction, les courbes enveloppent successivement les années 60, 70, 80 et 90.
Critères de reconnaissance d'une espèce introduite .
Les critères qui permettent de considérer qu'une espèce est probablement introduite sont les suivants :
(i) L'espèce est nouvelle pour l'aire considérée.
(ii) Il existe une discontinuité géographique entre son aire connue et sa nouvelle station (WILLIAMSON et al., 1986).
(iii) La nouvelle station est très ponctuelle; des biotopes similaires à celui qui est colonisé, situés au voisinage de cette station, ne sont pas (encore ?) colonisés. C'est le cas de Tricellaria inopinata (Bryozoaire) qui, lors de sa découverte, était cantonné à un petit secteur de la lagune de Venise, proche du centre historique de la cité (d'HONDT et OCCHIPINTI-AMBROGI (1985); ultérieurement, il a envahi la totalité des canaux de l'ensemble de la lagune.
(iv) A partir d'une station initiale ponctuelle, la cinétique d'extension de l'aire est cohérente. Le cas de Sargassum muticum (Fucophyceae) le long des côtes atlantiques

1 2
d'Europe est exemplaire de ce point de vue (Fig. 1). Le polychète Marenzelleria viridis, originaire d'Amérique du Nord, semble avoir été introduit de façon indépendante en deux sites distincts : l'estuaire de l'Ems (Nord des Pays-Bas) en 1983 et l'estuaire de la Tay (Ecosse) en 1984 (ESSINK et KLEEF, 1988, 1992); son expansion ultérieure est chronologiquement cohérente (Fig. 2).
(ν) L'espèce introduite a tendance à pulluler, tout au moins pendant un certain temps. C'est le cas de l'algue brune Colpomenia sinuosa (SAUVAGEAU, 1906) et de la méduse Bhopilema nomadica (SPANIER et GALIL, 1991).
(vi) Il existe à proximité une source d'introduction potentielle : installations aquacoles, port, laboratoire, aquariums.
(vii) La nouvelle population ne possède qu'une partie de la variabilité génétique de l'espèce dans son aire d'origine; on parle d'"effet de fondation". Dans le cas des algues, le cycle peut être incomplet; c'est le cas de Codium fragile (Chlorophyta) en Méditerranée, qui n'est représenté que par des individus femelles parthénogénétiques (FELDMANN, 1956).
Ces critères, qui ne sont généralement pas tous réunis, permettent de juger de la probabilité qu'une espèce soit introduite. Le fait qu'une espèce soit de grande taille, et ne passe donc pas facilement inaperçue, et que son statut taxonomique soit clair, facilitent le diagnostic. Ce n'est sans doute pas un hasard si des groupes dont la systématique est complexe et plus ou moins confuse, tels que les Ulva, Enteromorpha, Cladophora (Chlorophyta) et Ectocarpaceae (Fucophyceae), ne comportent pas d'espèces présumées introduites.
Au total, certaines espèces peuvent être considérées de façon certaine comme des espèces introduites : c'est le cas par exemple de deux algues de grande taille, connues comme endémiques du Japon, Undaria pinnatifida et Laminaria japónica; il est impossible qu'elles aient pu exister antérieurement en Europe et y passer inaperçues.
Des espèces ont été considérées dès leur découverte comme très probablement ou certainement introduites, alors que leur région d'origine est incertaine ou même inconnue. Le bryozoaire Tricellaria inopinata, découvert dans la lagune de Venise, décrit comme une espèce nouvelle, a été aussitôt interprété comme une espèce introduite dont la région d'origine est inconnue (OCCHIPINTI-AMBROGI, 1991). De même, la méduse Rhopilema nomadica, bien que décrite comme une espèce nouvelle lors de sa découverte sur les côtes d'Israël, a été interprétée comme un immigrant lessepsien; ce n'est qu'ultérieurement que sa présence en Mer Rouge a été confirmée (SPANIER et GALIL, 1991). C'est encore le cas de Colpomenia peregrina (Fucophyceae), perçu dès sa découverte sur les côtes atlantiques (Bretagne et Grande Bretagne) comme une espèce introduite; il est d'abord attribué à l'espèce méditerranéenne C. sinuosa (SAUVAGEAU, 1906, 1908b, 1912); ultérieurement, il devient clair que c'est un taxon distinct, non encore décrit dans sa région d'origine, et SAUVAGEAU (1927) le décrit sous le nom de C. sinuosa var. peregrina; c'est finalement BLACKLER (1963) qui l'identifie dans sa région d'origine, le Pacifique. Le scléractiniaire Oculina patagónica, lors de sa découverte en Méditerranée, a été considéré comme une espèce introduite; il n'est pourtant connu qu'à l'état fossile du Pleistocène des côtes atlantiques d'Amérique du Sud (ZIBROWIUS, 1974, 1991; ZIBROWIUS et RAMOS, 1983). En revanche, certaines espèces n'ont pas été perçues comme introduites lors de leur découverte en Europe. C'est le cas d'Antithamnionella sarniensis (Rhodophyta), décrit de Guernesey comme une espèce nouvelle par LYLE (1922); ce n'est qu'ultérieurement que l'hypothèse d'une introduction sera envisagée (WESTBROOK, 1930; FARNHAM, 1980). Son origine reste toutefois imprécise : sans doute l'hémisphère Sud (FARNHAM, 1980).

- 1 3 -
Enfin, le statut de certaines espèces s'est modifié au cours du temps. C'est le cas de Polyphysa párvula (Chlorophyta). Lors de sa découverte en Méditerranée, il a été considéré comme une espèce endémique, sous le nom d'Acetabularia wettsteinii (SCHUSSNIG, 1930). Ultérieurement, mis en synonymie avec Acetabularia moebii puis A. párvula (= Polyphysa párvula), il a été considéré comme une espèce indo-pacifique entrée en Méditerranée par le canal de Suez (POR, 1978). Sa présence aux Antilles, puis sa découverte aux Canaries et à Madeira (VALET, 1969; PRUD'HOMME VAN REINE et al, 1984) conduisent toutefois à se demander si ce n'est pas une espèce pantropicale dont la présence en Méditerranée est ancienne; on note du reste qu'elle a été signalée en Méditerranée occidentale (SCHUSSNIG, 1930) avant de l'être en Méditerranée orientale (ALEEM, 1948), ce qui n'est pas le cas des espèces lessepsiennes.
Fig. 2 : Distribution de Marenzelleria viridis (polychète). Four chaque localité, l'année de la première observation est indiquée. Les flèches indiquent les routes possibles de la progression de l'espèce (d'après ESSINK et KLEEF, 1992).
Les vecteurs des introductions d'espèces
En milieu marin, les introductions liées à l'homme ont plusieurs origines :
Le transport sur les coques de navires d'espèces fixées (fouling) ou vagiles (clinging) sur les coques des navires. Le déballastage des navires (eau ou matières solides). Les appâts pour la pêche et les espèces vivantes dans les marchés. La recherche scientifique.

- 1 4 -
L'aquaculture (introduction délibérée d'espèces aquacoles, introduction accidentelle d'espèces accompagnatrices). L'aquariologie. En Méditerranée, il convient d'ajouter le percement du canal de Suez.
Historiquement, les introductions liées au fouling ont été les plus anciennes. Elles semblent s'être faites sur un rythme assez lent. On peut penser que la plupart des espèces qui avaient vocation à être introduites ainsi l'ont déjà été, et que cette source d'introduction s'est aujourd'hui ralentie, d'autant plus que les peintures anti-fouling limitent maintenant le fouling sur les coques des bateaux. Il est d'ailleurs probable que certaines espèces qui sont des composantes régulières du fouling et qui ont aujourd'hui une répartition mondiale (espèces cosmopolites) correspondent à des introductions anciennes, bien qu'elles ne soient pas perçues ainsi. La génétique moléculaire pourra le démontrer. Chez les polychètes, on peut penser à certains Serpulidae, chez les crustacés aux isopodes foreurs de bois du genre Limnoria, chez les algues aux Acrochaetiales, aux Ectocarpaceae et aux Cladophora. Le type des introductions liées au fouling est constitué par le crustacé cirripède Elminius modestus, arrivé en Grande Bretagne au cours de la seconde guerre mondiale (CRISP, 1958).
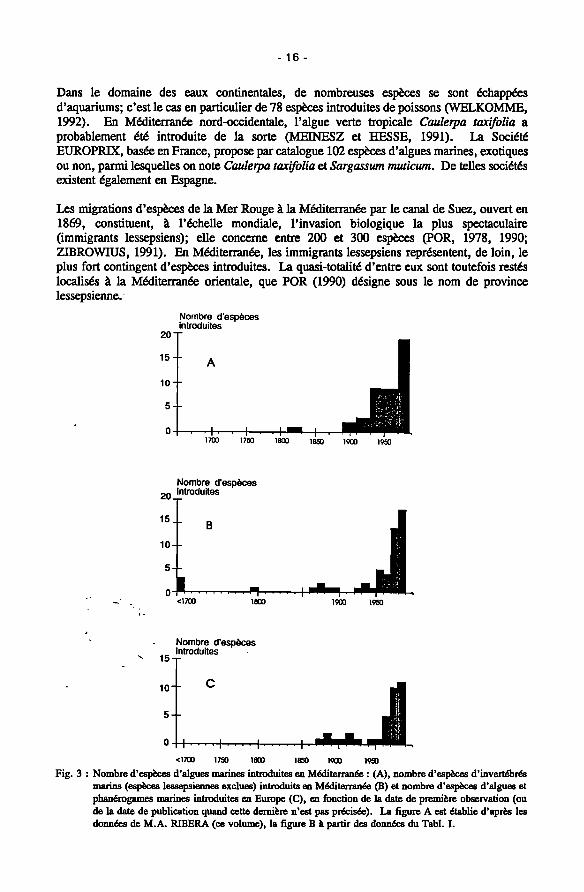
Mais, à partir des années 60, on note une formidable accélération des introductions, les autres causes d'introduction prenant le relais. On remarque que plus de 70% de toutes les introductions d'algues marines en Europe sont postérieures à 1960 (Fig. 3C). Par ailleurs, plus de 50% ont (dans un premier temps !) concerné la France (Fig. 4). En Méditerranée, près de 50% des algues introduites (y compris les espèces lessepsiennes) et près de 60% des invertébrés introduits (non compris les lessepsiens) y ont été observées postérieurement à 1970 (Fig. 3A et B). Naturellement, ces chiffres sont à considérer avec prudence dans la mesure où la première observation d'une espèce introduite est presque toujours postérieure à sa date réelle d'introduction; en outre, ces chiffres peuvent traduire un accroissement de la pression d'observation et une amélioration des connaissances dans le domaine de la biogéographie.
Le rejet des eaux de ballast des navires est à l'origine de l'introduction de la diatomée planctonique Odontella (= Biddulphia) sinensis (CARLTON, 1985; BOALCH, 1987). Les larves du mollusque nord-américain Ensis directus sont probablement arrivées en Europe du Nord de la même façon (ESSINK, 1986).
Les appâts pour la pêche constituent une cause mineure d'introduction. On peut citer le cas de Fucus spiralis (Fucophyceae), qui sert à les emballer et qui provient de Bretagne, introduit dans l'étang de Gruissan (Aude, côtes méditerranéennes de France) (SANCHOLLE, 1988).
L'algue rouge atlantique Mastocarpus stellatus a été introduite en Allemagne (Helgoland, Mer du Nord) à la fin des années 70 par un scientifique de passage pour les besoins de ses recherches. Elle a depuis colonisé toute la côte Ouest de l'île d'Helgoland (K. LUNING in WALLENTINUS, sous pressé).
En 1981, des algues supposées appartenir à Eucheuma spinosum (Rhodophyta) en provenance des Philippines ont été cultivées en mer ouverte aux Antilles françaises (Guadeloupe), pour comparer leur vitesse de croissance avec celle des Eucheuma spinosum indigènes. En deux mois, ils sont morts, victimes de nécroses que les auteurs de l'expérience (BARBAROUX et al., 1984) attribuent à la maladie ice-ice, connue de l'Asie du Sud-Ouest. Les auteurs constatent que cette maladie ne s'est pas transmise aux Eucheuma indigènes, qu'ils considèrent donc comme immunisés. Ils ont donc, en toute connaissance de cause, pris le risque d'introduire une maladie dans une région du monde d'où elle était absente ... Il est

- 1 5 -
surprenant qu'une telle expérience ait été autorisée par l'ISTPM, administration à laquelle appartenaient les auteurs.
L'huître japonaise Crassostrea gigas a été introduite en France à partir de 1966 pour compenser la baisse de productivité de l'huître portugaise C. ungulata, puis pour la remplacer quand cette dernière a été décimée (en 1967 et en 1970-72) par une maladie d'origine virale (COMPS et al., 1976; GRIZEL et HERAL, 1991). Entre 1971 et 1977, plus de 10 000 t de naissain ont été importées du Japon (baie de Sandai). A son arrivée en France, le naissain subissait un contrôle sanitaire et était immergé une heure dans l'eau douce (instruction ISTPM du 30 Avril 1971), afin de détruire les organismes fixés (fouling) et les prédateurs, principalement le turbellarié Pseudostylochus (GRIZEL et HERAL, 1991) ; ce temps de séjour dans l'eau douce ne semble pas résulter d'une étude scientifique approfondie, et son efficacité est sérieusement mise en doute par GRUET et al. (1976) qui relèvent, sur du naissain ainsi traité, une trentaine d'espèces encore vivantes : algues, actinies, turbellariés, polychètes, bryozoaires, mollusques, crustacés, ascidies; en outre, sa mise en oeuvre est confiée aux ostréiculteurs, qui ne semblent pas l'appliquer avec toute la rigueur requise (ce qui est un euphémisme). On comprend dans ces conditions pourquoi les introductions on été si nombreuses. GRUET et al. (1976) citent, sur le littoral atlantique français, l'annélide Hydroides ezoensis, le cnidaire Aiptasia pulchella, le mollusque Anomia chinensis, et les crustacés cirripèdes Balanus amphitrite amphitrite et B. albicostatus auxquels d'HONDT (1984) ajoute le bryozoaire Watersipora aterrima. Dans l'étang de Thau (France, Méditerranée), ce sont au moins 9 espèces d'algues dont l'introduction est liée à celle du naissain de Crassostrea gigas (PEREZ et al, 1981; RIOUALL et al, 1985; BEN MAIZ et al, 1987; VERLAQUE et RIOUALL, 1989) : Laminaria japónica, Undaria pinnatifida, Sargassum muticum, Sphaerotrichia divaricata (Fucophyceae), Antithamnion nipponicum, Chrysymenia wrightii, Grateloupia doryphora, Lomentaria hakodatensis, et Porphyra yesoensis (Rhodophyta). On a longtemps affirmé que C. gigas avait été introduit pour pallier au déclin, puis à la disparition de C. ungulata (GRIZEL, 1983). Une autre thèse est que ce soit exactement le contraire : les C. gigas importés en France en 1966 pour des essais d'aquaculture auraient apporté les Bonamia et le virus (dont ils étaient "porteurs sains") qui ont décimé les C. ungulata; une troisième possibilité est que le virus soit arrivé du Portugal avec des C. ungulata adultes (GRIZEL et HERAL, 1991).
L'algue géante Macrocystispyrifera (Fucophyceae), qui peut dépasser 40 m de longueur, est exploitée pour la production d'alginates, en particulier en Californie. En 1950, la Scottish Research Association a proposé son introduction en Ecosse, mais l'autorisation lui a été refusée par décision gouvernementale (BOALCH, 1981). Au début des années 70, elle a été cultivée par l'ISTPM en mer ouverte pendant 7 mois à Roscoff (Bretagne, France), en vue de tester la faisabilité de son introduction. Les souches provenaient du Chili. De très vigoureuses protestations, en France et dans les autres pays d'Europe (BOALCH, 1981), ont obligé l'ISTPM à mettre fin à cette expérience. Par chance, l'espèce n'a pas été introduite, mais les expériences avaient toutefois confirmé que cette algue géante était parfaitement capable de vivre en Bretagne. Ultérieurement, elle a été délibérément introduite en Chine (NEUSHUL, 1983).
La Fucophyceae Undaria pinnatifida a été délibérément introduite en Bretagne, à partir de l'étang de Thau (Méditerranée), dans un but aquacole (PEREZ et al, 1988, 1991). Sur la base des travaux d'AKIYAMA (1965), il avait été établi qu'elle ne pourrait pas se reproduire en mer ouverte, dans les conditions thermiques de la Bretagne. Elle s'est pourtant échappée des élevages (FLOC'H et al, 1991).
16
Dans le domaine des eaux continentales, de nombreuses espèces se sont échappées d'aquariums; c'est le cas en particulier de 78 espèces introduites de poissons (WELKOMME, 1992). En Méditerranée nord-occidentale, l'algue verte tropicale Caulerpa taxifolia a probablement été introduite de la sorte (MEINESZ et HESSE, 1991). La Société EUROPRIX, basée en France, propose par catalogue 102 espèces d'algues marines, exotiques ou non, parmi lesquelles on note Caulerpa taxifolia et Sargassum muticum. De telles sociétés existent également en Espagne.
Les migrations d'espèces de la Mer Rouge à la Méditerranée par le canal de Suez, ouvert en 1869, constituent, à l'échelle mondiale, l'invasion biologique la plus spectaculaire (immigrants lessepsiens); elle concerne entre 200 et 300 espèces (POR, 1978, 1990; ZIBROWIUS, 1991). En Méditerranée, les immigrants lessepsiens représentent, de loin, le plus fort contingent d'espèces introduites. La quasi-totalité d'entre eux sont toutefois restés localisés à la Méditerranée orientale, que POR (1990) désigne sous le nom de province lessepsienne.
Nombre d'espèces introduites
20 Τ
15
10 +
f—
Nombre d'espèces „Q introduites
1 5 . .
1 0
5
Λ 1900 1950
Nombre d'espèces introduites
10
5 +
+ <1700 1750 IMI) 1850 1900 1950
Fig. 3 : Nombre d'espèces d'algues marines introduites en Méditerranée : (A), nombre d'espèces d'invertébrés murine (espèces lessepsiennes exclues) introduits en Méditerranée (B) et nombre d'espèces d'algues et phanérogames marines introduites en Europe (C), en fonction de la date de première observation (ou de la date de publication quand cette dernière n'est pas précisée). La figure A est établie d'après les données de M.A. RIBERA (ce volume), la figure Β à partir des données du Tabi. I.

17
Les facteurs de succes des introductions d'espèces
Selon WILLIAMSON et BROWN (1986), les caractéristiques biologiques d'une espèce introduite (ou susceptible de l'être) sont : (i) l'absence d'ennemis naturels, (ii) une grande capacité de dispersion, (iii) une stratégie de type r-sélection et (iv) la présence d'une niche écologique vide. Ces auteurs soulignent toutefois qu'aucune de ces conditions n'est nécessaire, ou suffisante, pour expliquer le succès des introductions observées. Par exemple, dans l'étang de Thau, Undaria pinnatifida (Fucophyceae) est une espèce très appréciée par l'oursin Paracetamols lividus (SAN MARTIN, 1986). Dans la lagune de Venise, Ruditapes phitippinarum (mollusque) occupe une niche écologique qui n'était pas vide (CESARI et PELLIZZATO, 1985).
L'arrivée de quelques individus d'une espèce exotique n'implique pas automatiquement sa naturalisation; c'est même très certainement le contraire, les introductions "réussies" ne représentant qu'un faible pourcentage des introductions potentielles. La plupart des espèces japonaises observées par GRUET et al. (1976) sur du naissain de l'huître Crassostrea gigas, importé du Japon et immergé au Croisic et à Bourgneuf (région de Nantes, France) ne semblent pas s'être naturalisées. La taille de l'inoculum, et donc la variabilité génétique de la population introduite, constitue certainement un facteur important du succès d'une introduction (CRISP, 1958).
En fait, le facteur essentiel du succès d'une espèce introduite nous semble être la faible biodiversité de la communauté (ou de la région) d'accueil. La faible biodiversité de la Méditerranée orientale, résultat de son histoire, de la fermeture de l'isthme de Suez et des glaciations quaternaires, explique sans doute le succès exceptionnel que les immigrants lessepsiens y ont connu (SPANIER et GALIL, 1991). En dehors de la Méditerranée orientale, ce sont également des biotopes à faible biodiversité qui ont accueilli la grande majorité des espèces introduites avec succès : lagunes, ports, estuaires et milieux pollués (Tabi. I et Π).
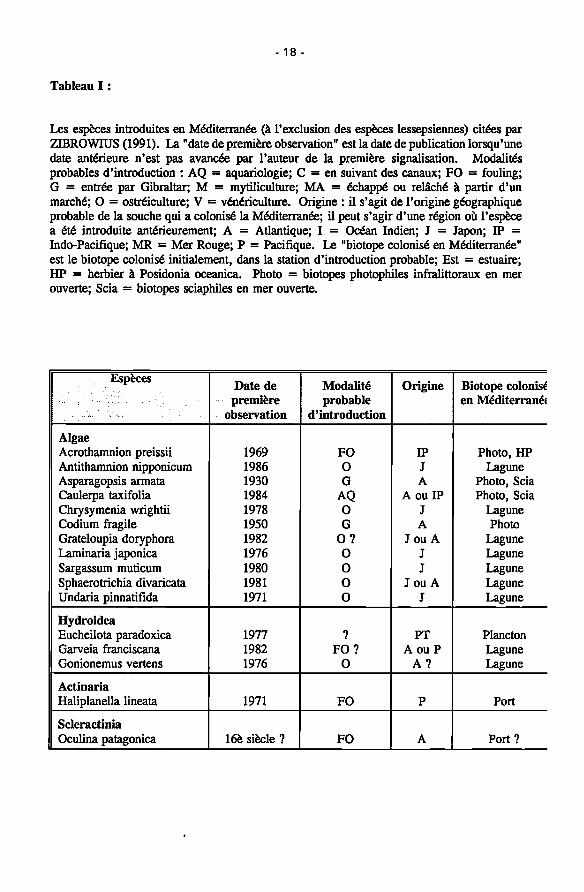
18
Tableau I :
Les espèces introduites en Méditerranée (à l'exclusion des espèces lessepsiennes) citées par ZIBROWIUS (1991). La "date de première observation" est la date de publication lorsqu'une date antérieure n'est pas avancée par l'auteur de la première signalisation. Modalités probables d'introduction : AQ = aquariologie; C = en suivant des canaux; FO = fouling; G = entrée par Gibraltar; M = mytiliculture; MA = échappé ou relâché à partir d'un marché; O = ostréiculture; V = vénériculture. Origine : il s'agit de l'origine géographique probable de la souche qui a colonisé la Méditerranée; il peut s'agir d'une région où l'espèce a été introduite antérieurement; A = Atlantique; I = Océan Indien; J = Japon; IP = Indo-Pacifique; MR = Mer Rouge; Ρ = Pacifique. Le "biotope colonisé en Méditerranée" est le biotope colonisé initialement, dans la station d'introduction probable; Est = estuaire; HP = herbier à Posidonia oceanica. Photo = biotopes photophiles infralittoraux en mer ouverte; Scia = biotopes sciaphiles en mer ouverte.
Espèces
. . . · . . · · - ■ ■ · . . · . " ; : " · .
Algae Acrothamnion preissii Antithamnion nipponicum Asparagopsis armata Caulerpa taxifolia Chrysymenia wrightii Codium fragile Grateloupia doryphore Laminaria japónica Sargassum muticum Sphaerotrichia divaricata Undaria pinnatifida
Hydroidea Eucheilota paradoxica Garveia franciscana Gonionemus vertens
Actinaria Haliplanella lineata
Scleractinia Oculina patagónica
Date de première
observation
1969 1986 1930 1984 1978 1950 1982 1976 1980 1981 1971
1977 1982 1976
1971
16è siècle ?
Modalité probable
d'introduction
FO 0 G
AQ 0 G
0 ? 0 0 O 0
? FO?
O
FO
FO
Origine
IP J A
A ou IP J A
J ou A J J
J ou A J
PT A ou Ρ
A?
Ρ
A
Biotope colonisé en Mediterranei
Photo, HP Lagune
Photo, Scia Photo, Scia
Lagune Photo
Lagune Lagune Lagune Lagune Lagune
Plancton Lagune Lagune
Port
Port?

19
■'·. "s Espèces
. · v . > ; ν · , · : , ■
Mollusca Chromodoris quadricolor Crassostrea angulata Crassostrea gigas Crepidula calyptraeformis Crepidula fornicata Gibbula cineraria Littorina littorea Littorina saxatilis Mercenaria mercenaria Musculista senhousia Petricola pholadiformis Rapana venosa Ruditapes philippinarum Saccostrea commercialis Scapharca inaequivalvis Siphonaria pectinata
Polychaeta Ficopomatus enigmaticus Hydroides dianthus Hydroides dirampha Hydroides elegans Pileolaria berkeleyana Spirorbis marioni
Crustacea Ammothea hilgendorfi Callinectes sapidus Cancer pagurus Elasmopus pectenicrus Eriocheir sinensis Myicola ostreae Mytilicola orientalis Necora puber Pachygrapsus transversus Paracerceis sculpta Paradella dianae Sphaeroma walkeri
Bryozoa Tricellaria inopinata Ascidia Microcosmus exasperatus Polyandrocarpa zorritensis
Vate de première
Observation
1986 fìn 19è siècle
1970 16è siècle ?
1957 1979 1982 1792 1965 1982 1963 1947 1981 1985 1973 1980
années 20 1865 1870 1883 1977 1977
1982 1955 1792 1936 1959 1980 1979 1958 1876 1978 1991 1977
1982
1963 1981
Modalité . probable
d'introduction
? O 0
FO? M, 0
0 M ? V 0 7
FOouO V ?
FO ?
FO FO FO FO FO FO
FO ?.
MA FO C 0 O M FO FO FO FO
FO
FO FO
Origine
MR, IP A J A A A A A A
J ou A A Ρ A Ρ IP A
A A A ? Ρ Ρ
IP A A
MR, I A J J A A Ρ
PT I
?
PT Ρ, A
Biotope colonisé en Méditerranée
7 Lagune Lagune
Port Pollué ? Photo
Lagune Lagune Lagune Lagune
? Pollué ? Lagune Lagune
? ?
Lagune, Est Port Port Port Port Port
Lagune 7
Port ?
Lagune Lagune Lagune Pollué Port
Lagune Port Port
Lagune
Port Port

2 0
Dans le Sud de l'Angleterre, le Soient, où de nombreuses espèces ont été introduites, est considéré par FARNHAM comme un secteur relativement favorable aux introductions en raison de la présence de communautés "ouvertes" et de la relative rareté des communautés "fermées" à Fucales et Laminariales. FARNHAM (1980) suggère que le fouling, comme cause d'introduction, a été surestimé, dans la mesure où des milieux aussi fortement pollués que les ports lui paraissent des milieux défavorables au succès d'une introduction; nous considérons au contraire que la faible biodiversité des ports en fait des milieux très favorables aux introductions. Les communautés à forte biodiversité offrent probablement une plus grande résistance aux introductions; ce n'est peut-être pas un hasard si, en Méditerranée, l'herbier à Posidonia oceanica et le coralligène ne sont presque jamais cités comme biotopes d'espèces introduites (Tabi. I et Π). Il peut toutefois s'agir d'un artefact, au moins en partie : en effet, il est logique que les introductions dues à l'homme concernent d'abord des milieux anthropisés, quelle que soit leur biodiversité.
Tableau Π :
Nombre d'espèces introduites en fonction du biotope ou du milieu colonisé initialement. D'après les données du Tabi. I.
Biotopes et milieux
Lagunes Estuaires Ports Biotopes pollués Biotopes photophiles infralittoraux Biotopes sciaphiles Herbier à Posidonia oceanica Plancton Inconnu
Nombre d'espèces
23 à 24 O à l
13 à 14 l à 3 2 à 5 0 à 2 O à l
1 6 à 9
Modalités de l'expansion des espèces introduites
Dans le domaine continental comme en milieu marin, une espèce introduite peut avoir un développement initial spectaculaire. Par la suite, l'exubérance de l'espèce diminue souvent, mais ce n'est pas toujours le cas. Le gastropode lessepsien Bhinoclavis kochii est devenu l'une des espèces dominantes, à la fin des années 70, le long des côtes israéliennes; il a depuis considérablement décliné (SPANIER et GALIL, 1991). Au début du siècle, en Bretagne, Colpomenia peregrina a connu un développement initial spectaculaire, couvrant plus ou moins complètement les parcs à huîtres (SAUVAGEAU, 1906); quelques années plus tard, C. peregrina était "rentré dans le rang", ne tenant plus qu'une place discrète dans les peuplements littoraux (SAUVAGEAU, 1908b). Dans l'étang de Thau (Méditerranée, France), Sargassum muticum a décliné à la fin des années 80, à la suite de l'expansion de l'oursin Paracentrotus lividus qui broute ses germinations (LAURET in BELSHER, 1991). En Méditerranée, Codium fragile (Chlorophyta) a connu une phase d'exubérance dans la région marseillaise au début des années 60; il était alors rejeté en grande quantité sur les

21
plages, lors des tempêtes, constituant des laisses suffisamment importantes pour que les autorités municipales les fassent enlever avec des engins motorisés; il a depuis régressé. C'est également le cas, en Grande Bretagne, des diatomées planctoniques Pleurosigma planctonicum et Thalassiosira angstii (BOALCH, 1987). Les causes du déclin de certaines espèces introduites, après leur phase de prolifération initiale, sont mal connues; parmi les causes possibles, on peut citer : (i) l'accoutumance à sa consommation des prédateurs indigènes; (ii) la faiblesse de la variabilité génétique (due à la taille insuffisante de l'inoculum), qui rend difficile l'adaptation aux changements ou aux fluctuations de l'environnement; (iii) la réduction de la vigueur génétique due à la consanguinité (dépression de consanguinité) et/ou à l'élimination de certains alleles.
Pour d'autres espèces, la phase de développement semble avoir atteint un plateau, mais aucun déclin ne s'est produit (ou ne s'est encore produit); c'est le cas, en Israël, du crabe Charybdis longicollis et de la méduse Rhopilema nomadica (SPANIER et GALIL, 1991). Il n'est pas possible de prévoir quelle est la durée de la phase d'expansion, puis de la phase de plateau, chaque espèce introduite constituant un cas particulier. Dans le cas à'Eichornia crassipes (jacynthe d'eau), qui a colonisé l'ensemble des eaux douces intertropicales de la planète, à partir de l'Amazonie, aucun signe de déclin n'est perceptible, plus d'un siècle après le début de son introduction et de sa prolifération.
La vitesse d'expansion d'une espèce introduite est très variable. En 20 ans, Sargassum miuicum a colonisé, sur les côtes atlantiques d'Europe, plusieurs milliers de km de côtes, de la Norvège au Portugal, et cette expansion se poursuit (RUENESS, 1989). A vol d'oiseau, 2500 km séparent ses stations les plus éloignées (Fig. 1). En Méditerranée, il a fallu plus d'un siècle à la Phanérogame Halophila stipulacea pour occuper une aire s'étendant sur 2000 km, de la Syrie à Malte et à la Sicile (BILIOTTI et ABDELAHAD, 1990). Dans la Mer du Nord, le mollusque Ensis directus, signalé initialement à Cuxhaven (Allemagne) en 1978, a progressé de près de 400 km vers le Nord, entre 1978 et 1982 (ESSINK, 1986), soit une vitesse moyenne de 100 km/an. Dans la lagune de Venise, Tricellaria inopinata (bryozoaire) a progressé, entre 1983 et 1988, à la vitesse moyenne de 6 km/an (OCCHIPINTIAMBROGI, 1991).
TE3
Kg. 4 : Répartition par pays du nombre de premières signalisations d'algues marines introduites en Europe (l'apparition dans un pays d'une espèce introduite signalée antérieurement dans un autre pays d'Europe n'est donc pas prise en compte).

- 2 2 -
L'impact écologique des invasions biologiques
De très nombreux travaux ont été consacrés à l'impact des invasions biologiques sur les milieux continentaux et les eaux douces : publications, colloques, ouvrages de synthèse. Cet impact est en revanche très mal connu en milieu marin (POSEY, 1988). D'une façon générale, 80% des espèces introduites n'auraient pas d'effet sur les communautés indigènes (SIMBERLOFF, 1981). Pour les eaux côüères européennes, c'est le cas par exemple de Bonnemaisonia hamifera (Rhodophyta) et (actuellement) de Colpomenia peregrina (FARNHAM, 1980). De même, dans la lagune de Venise, Tricellaria inopinata (bryozoaire) n'a pas sérieusement modifié la structure des communautés de bryozoaires; tout au plus peut on noter des modifications dans l'abondance de certaines espèces, par exemple le déclin de Bugula stolonifera (OCCfflPIOTI AMBROGI, 1991). L'introduction du polychète Marenzelleria viridis n'a pas interféré significaüvement avec la communauté des vases estuariennes en Europe du Nord (ESSINK et KLEEF, 1992).
D'autres espèces introduites ont un impact sur les espèces ou les communautés indigènes : on parle alors de pollution biologique (SINDERMANN et al., 1992). Dans certaines régions (côtes méridionales de l'Angleterre et de l'Irlande), l'algue verte introduite Codium fragile est entrée en compétition avec les espèces indigènes C. vermilara et C. tomentosum, et l'abondance de cette dernière, dont le port est semblable et les niches écologiques voisines, a diminué (PARKES, 1975; FARNHAM, 1980). A Hawaii, les Rhodophyta introduites Hypnea musciformis et Acanthophora spicifera occupent la même niche écologique que les espèces indigènes Laurencia nidifica et Hypnea cervicornis; seule cette dernière a vu son abondance réduite (de moitié) du fait de la compétition (RUSSELL, 1992). En Méditerranée orientale, la crevette Penaeidae Penaeus japonicus a remplacé plus ou moins complètement, selon les régions, l'espèce indigène Penaeus kerathurus; de même, l'étoile de mer Asterina wega a remplacé en Israël l'espèce indigène A. gibbosa. Les poissons introduits Upeneus moluccensis et Saurida undosquamis ont provoqué un glissement de la niche écologique de Mullus barbants et de Merluccius merluccius, repoussés en profondeur (SPANIER et GALIL, 1991).
Les écosystèmes méditerranéens sont caractérisés par la faiblesse du compartiment herbivores. L'arrivée de plusieurs poissons herbivores (Siganus rivulatus, S. luridus) en Méditerranée orientale (POR, 1978), et leur abondance, a probablement modifié profondément le fonctionnement des écosystèmes infralittoraux, d'autant plus que ces poissons ont des préférences alimentaires très précises (LUNDBERG et LIPKIN, 1992), ce qui favorise les espèces évitées au détriment des espèces préférées.
Les algues de grande taille telles que les Laminariales et les Fucales, qui sont souvent des espèces-clé (key-stone species), ont un impact potentiel important. En éliminant des espèces indigènes, elles peuvent modifier le fonctionnement de l'écosystème. En Californie, l'installation de Sargassum muticum en 1976, favorisée par l'élévation momentanée de la température (El Niño), a ultérieurement empêché le recrutement de l'algue géante Macrocystis pyrifera, en diminuant de façon drastique l'éclairement au niveau de la sous-strate (AMBROSE et NELSON, 1982). En Colombie britannique (Canada), Sargassum muticum a réduit le recouvrement de Rhodomela larix (Rhodophyta); toutefois, en cas d'élimination expérimentale ou naturelle (tempêtes) de S. muticum, Rhodomela larix est capable de se réinstaller à sa place, puis d'empêcher la recolonisation ultérieure par S. muticum (DE WREDDE, 1983). Dans l'étang de Thau (Méditerranée, France), S. muticum a éliminé l'espèce indigène Cystoseira barbata (Fucophyceae) de certaines stations; en constituant une canopée très dense, ne laissant pas filtrer la lumière, à l'époque où Cystoseira barbata est fertile (février à avril), S. muticum empêche son recrutement; par ailleurs, les indices de

- 2 3 -
diversité et d'équitabilité sont exceptionnellement faibles dans les peuplements à S. muticum (GERBAL et al., 1985). Dans la Manche, Sargassum muticum entre en compétition avec diverses espèces, en particulier Laminaria sp. plur., Cystoseira sp. plur., Scytosiphon lamentaria, (Fucophyceae), Gracilaria verrucosa, Chondrus crispus (Rhodophyta). Lorsque le peuplement de S. muticum est particulièrement dense, il y a élimination totale des espèces concurrentes (FLETCHER et FLETCHER, 1975; CRITCHLEY, 1983).
Dans les peuplements à Caulerpa taxifolia (Chlorophyta) des Alpes-Maritimes (Méditerranée, France), sur substrat dur, le nombre d'espèces d'algues est quatre fois plus faible, la biomasse des algues indigènes 9 fois plus faible et le recouvrement des algues indigènes près de 500 fois plus faible que dans les peuplements d'algues photophiles qu'ils remplacent (BOUDOURESQUE et al., 1992).
En fait, on connaît peu de choses sur l'impact des espèces introduites sur les communautés indigènes, y compris lorsqu'il s'agit d'espèces dominantes et de grande taille : Asparagopsis armata (Rhodophyta) couvre parfois à 100% le substrat en hiver, en Méditerranée nord-occidentale, sur 20 à 30 cm de hauteur; associé à Codium fragile (Chlorophyta), il constitue l'espèce dominante de certains peuplements infralittoraux dans la région de Marseille; Polysiphonia setacea (Rhodophyta) recouvre d'une couche épaisse de 5 cm, pratiquement monospécifique, les peuplements coralligènes de la réserve de Scandola (Corse) : quel est son impact sur les algues calcaires édificatrices du coralligène ?
Les espèces introduites occupent parfois une niche écologique plus large que dans leur région d'origine; en outre, leurs caractéristiques morphologiques peuvent être différentes. C'est ainsi par exemple que Sargassum muticum est, au Japon, une espèce plutôt discrète parmi une trentaine d'espèces du genre Sargassum, cantonnée à la frange infralittorale et ne dépassant pas 1.0-1.5 m de longueur (RUENESS, 1989); en Europe, sa niche écologique s'est élargie, elle se rencontre depuis les flaques médiolittorales jusqu'à 20 m de profondeur (NORTON, 1977) et peut dépasser 10 m de longueur (BELSHER et BOYEN, 1983). La croissance d'Undaria pinnatiflda s'avère meilleure en Bretagne, où elle peut atteindre 2 à 3 m de longueur, que dans sa région d'origine, le Japon, où elle ne dépasse pas 1.5 m de longueur (PEREZ et al., 1988). U en va de même pour Caulerpa taxifolia (Chlorophyta), qui constitue en Méditerranée des peuplements extrêmement denses (plus de 8000 feuilles/m) et où les feuilles peuvent dépasser 60 cm de longueur (MEINESZ et HESSE, 1991), alors que, dans les mers chaudes, ses peuplements sont généralement clairsemés à très clairsemés, et ses feuilles ne dépassent guère 2-15 (25 cm).
Plus de 15 à 25 années après son installation (la date précise est sujette à discussions), Sargassum muticum a pu provoquer une diminution de l'abondance de certaines espèces indigènes (voir plus haut), mais rien ne permet de dire qu'il ait éliminé des espèces indigènes, tout au moins des espèces de grande taille telles que les Laminaria ou les Fucus (RUENESS, 1989). Toutefois, ce dernier auteur remarque que 15 années représentent une période de temps relativement courte dans ce domaine; en outre, l'arrivée continuelle de nouvelles espèces introduites (Undaria pinnatiflda, Laminaria japónica, etc) peut déboucher sur un effet cumulatif ou sur des synergies dépassant largement l'impact de chacune des espèces considérées isolément.
L'introduction de gènes est très probable, même si elle n'est que très peu documentée. En Corse, des souches à'Hypnea musciformis (Rhodophyta) provenant du Sénégal ont été cultivées (MOLLION, 1984); l'espèce y existe, mais donnait de mauvais résultats en culture, contrairement aux souches du Sénégal, ce qui traduit des différences génétiques; ces gènes ont pu être introduits. L'introduction des Fucophyceae Chorda filum et Sphaerotrichia

24
divarícata (RIOUALL, 1985) dans l'étang de Thau (France, Méditerranée) peut avoir été faite aussi bien à partir de l'Atlantique Nord que du Japon (avec le naissain de l'huître Crassostrea gigas), puisque l'espèce existe dans les deux régions. Dans le cas de Sphaerotrichia divaricata, PETERS et al. (1992) ont mis en évidence une barrière de stérilité entre les exemplaires de l'étang de Thau et ceux de l'Atlantique Nord, et concluent que l'introduction s'est faite à partir du Japon; une telle barrière de stérilité intraspécifique n'existe sans doute pas chez toutes les espèces à large répartition mondiale; si des souches japonaises de Sphaerotrichia divaricata et (éventuellement) de Chorda filum sont arrivées dans l'étang de Thau, elles ont pu tout aussi bien arriver sur les côtes atlantiques d'Europe, bien que leur détection y soit moins aisée, avec pour conséquence la contamination génétique des souches indigènes (quand elles ne sont pas protégées par une barrière de stérilité). La Fucophyceae Laminaria japónica, introduite dans l'étang de Thau, peut s'hybrider avec une espèce de l'Atlantique Nord, L. saccharina (BOLTON et al., 1983); pour le moment, les deux espèces ne sont pas en contact, mais le mélange de leur génome est tout à fait probable si cela se produisait (RUENESS, 1989).
Les nuisances dues aux invasions biologiques
Les nuisances font référence à l'utilisation de la mer par l'homme. Les nuisances dues à des espèces introduites sont beaucoup mieux documentées que l'impact sur les milieux naturels.
(i) Compétition avec des espèces exploitées. Crepidula fornicata (mollusque) est une nuisance pour les élevages de Mytilus et d'huîtres, avec lesquels il entre en compétition pour la nourriture et l'occupation de l'espace. Π en est de même pour Rapano venosa (mollusque) en Mer Noire.
(ii) Gêne à la pêche. Le long des côtes d'Israël, la méduse Rhopilema nomadica peut atteindre, lors des pullulations estivales, la densité de 25 individus/m3; les filets sont endommagés et il devient difficile de trier le contenu des filets : durant l'été 1990, la pêche a dû être interrompue pendant plusieurs semaines (SPANIER et GALIL, 1991). Dans le golfe de Giens, Pofysiphonia setacea (Rhodophyta) constitue une gêne pour la pêche en colmatant les filets (VERLAQUE, 1989). En Grande-Bretagne, les mucilages produits par la diatomée Coscinodiscus wailesii colmataient les chaluts, à la fin des années 70; un chalut a même été perdu à cause de la surcharge en poids (BOALCH, 1987).
(iii) Gêne à l'aquaculture. Dans les premières années qui ont suivi son introduction, Colpomenia peregrina (Fucophyceae) se développait en abondance sur les huîtres; à marée basse, le thalle, qui est creux, se vide de son eau; lorsque la marée remonte, il ne se remplit d'eau que lentement et constitue ainsi une sorte de bouée pour l'huître qui flotte quelque temps et est emportée hors du parc; SAUVAGEAU (1906) écrivait : "on conçoit quel énorme préjudice peut causer à l'ostréiculture l'invasion de ce nouveau commensal" que les ostréiculteurs ont nommé "la voleuse d'huîtres". Sargassum muticum, lorsqu'il se fixe sur les huîtres, peut également les entraîner grâce à ses flotteurs; en outre, sa prolifération dans les parcs conchylicoles, ou à leurs alentours, a constitué une gêne considérable et a nécessité le dégagement des parcs (BELSHER, 1991). Dans l'Atlantique Nord, le Dinophyte toxique Gyrodinium aureolum a causé des mortalités dans des fermes piscicoles (BOALCH, 1987).
(iv) Gêne au tourisme. Le long des côtes d'Israël, on a compté jusqu'à 135 individus de méduses Rhopilema nomadica par 100 m de plage. Les filets destinés à protéger les plages se sont révélés inopérants, dans la mesure où des fragments de méduses passent

2 5
au travers; les symptômes ressentis par les baigneurs vont d'une simple piqûre à une brûlure avec cloque laissant une cicatrice rémanente. Dans quelques cas, la brûlure s'accompagne de fièvre et de douleurs musculaires. Dans ces conditions, il n'est pas étonnant que la fréquentation des stations balnéaires diminue (SPANIER et GALIL, 1991). A Chypre, une Cladophora (Chlorophyta) introduite (peut-être Cladophora patentiramosa, Marc VERLAQUE, comm. verb.) a proliféré devant les plages, et ses épaves se sont accumulées sur les plages elles-mêmes, gênant ainsi les activités balnéaires (DEMETROPOULOS et HADJICHRISTOPHOROU, comm. verb.).
(ν) Gêne à la navigation. L'introduction de Macrocystis pyrifera (Fucophyceae) en Europe aurait gêné la navigation littorale (BOALCH, 1981). Sargassum muticum a effectivement constitué une gêne à la navigation, par exemple dans le golfe du Morbihan (BELSHER, 1991); un accident a été signalé à Jersey, où un ferry dont les hélices étaient bloquées par S. muticum a violemment heurté le quai.
(vi) Fouling. Le polychète introduit Hydroides elegans, extrêmement résistant au chlore, s'installe dans les installations de pompage de l'eau de mer (fouling), et est considéré en Italie comme une nuisance sérieuse (PAOLETTI et SEBASTIO, 1973). En Amérique du Nord, la moule zébrée Dreissena pofymorpha envahit également les canalisations de prise d'eau.
Les effets bénéfiques de l'introduction d'espèces
L'introduction volontaire de certaines espèces a permis de générer, ou de maintenir, des activités économiques. C'est le cas de l'huître Crassostrea gigas, si l'on suppose qu'elle a été introduite après la disparition de C. ungulata (GRIZEL et HERAL, 1991), et non que son introduction est la cause de la disparition de cette dernière. En Israël, les poissons introduits via le Canal de Suez représentent actuellement le tiers des prises; la crevette lessepsienne Penaeus japonicus est aujourd'hui largement pêchée en Israël et en Turquie (SPANIER et GALIL, 1991). A Hawaii, l'introduction des Rhodophyta .¿caniAopftora spicifera et Hypnea musciformis a eu pour résultat un accroissement de la biomasse algale; on note toutefois qu'il s'agit de deux espèces non consommées par les herbivores (RUSSELL, 1992).
Toutefois, pour qu'une espèce introduite ait réellement un effet économique bénéfique, il faut démontrer que le total des prises (toutes espèces confondues) est supérieur à ce qu'il était avant son introduction : les espèces introduites exploitables peuvent simplement remplacer une espèce indigène, également exploitée, comme c'est le cas des crevettes P. japonicus et P. kerathurus, respectivement. L'interprétation des statistiques de pêche est toujours difficile, et il ne semble pas qu'un tel accroissement soit démontré le long des côtes d'Israël. Des recherches ont été effectuées en vue de valoriser Sargassum muticum (alginates, pharmacologie, horticulture), mais elles ne semblent pas avoir donné de résultats concluants (BELSHER et POMMELEC, 1988; BELSHER, 1991).
Les tentatives d'éradication d'espèces introduites
L'éradication (complète ou partielle) d'une espèce marine introduite génératrice de nuisances est certainement très difficile. Par ailleurs, peu de recherches ont été effectuées dans ce domaine. Elles concernent principalement la Fucophyceae Sargassum muticum. L'éradication manuelle, tentée en Grande-Bretagne, a été un échec (GRAY et JONES, 1977 ; FARNHAM, 1980). En zone intertidale, on a utilisé un tracteur muni d'une herse pour dégager les parcs mytilicoles du Cotentin (France), mais cette technique entraîne des

2 6
bouleversements considérables du substrat. En zone sublittorale, l'utilisation d'une drague a permis de dégager les chenaux du golfe du Morbihan (France) et de rétablir la circulation des bateaux. Les parcs conchylicoles de l'île de Ré (France) ont été dégagés au moyen d'une chaîne traînée par deux engins amphibies; mais ce système a provoqué la fragmentation et la dispersion d'une grande partie des algues (1000 t), pour 250 t seulement ramenées à terre. La lutte chimique, au moyen d'herbicides, du sulfate de cuivre, de Γ hypochlorite de sodium et de phytohormones, a été tentée en Grande-Bretagne et en France; si certains produits, tels le sulfate de cuivre, se sont avérés efficaces, leur manque de spécificité et leur rémanence dans le milieu conduisent à n'envisager leur utilisation que dans des cas exceptionnels (JUPIN, 1989; BELSHER, 1991).
Thèmes prioritaires de recherche
(i) Le fait que la plupart des espèces introduites n'aient pas d'impact majeur sur les espèces et les communautés indigènes n'est pas basé sur des recherches spécifiques mais sur des impressions subjectives, (ii) Qu'il y ait ou non un impact évident, l'importance de l'impact et le temps nécessaire pour que s'établisse un nouvel équilibre ne sont pas connus, (iii) Par ailleurs, il convient d'analyser, dans des régions ou des écosystèmes où les espèces introduites sont nombreuses (Soient, côtes levantines, étang de Thau par exemple), l'effet d'un ensemble d'espèces introduites, et non pas seulement l'effet de chacune d'entre elles, prise isolément, (iv) L'impact des introductions de gènes, soit en provenance de populations éloignées d'une espèce déjà présente dans la zone considérée, soit par hybridation entre des espèces indigènes et des espèces introduites, doit être étudié, (v) Des méthodes destinées à prévenir l'introduction accidentelle des espèces, ou tout au moins à minimiser les risques d'introduction, doivent être mises au point; on déterminera en particulier les traitements ou les conditions de quarantaine qui permettraient d'éliminer les espèces accompagnatrices (fouling en particulier) lors des transferts, d'une région à une autre, d'espèces aquacoles. (vi) Les législations nationales ou internationales, en Europe, sont inexistantes, noues ou non contraignantes pour ce qui concerne la prévention des introductions en milieu marin; l'article 22 de la "Directive Habitat" (CEE) en est l'illustration; il conviendra donc de proposer l'amélioration des législations en vigueur (voir article de C. de KLEMM dans ce rapport), (vii) Les méthodes d'éradication n'ont guère fait l'objet d'études approfondies; les échecs enregistrés jusqu'à présent, à la suite de tentatives parfois artisanales, souvent naïves, ne doivent pas conduire à les rejeter; la connaissance de la biologie et de la génétique des espèces introduites est susceptible de déboucher sur des méthodes plus élégantes, plus efficaces et moins coûteuses que les méthodes mécaniques ou chimiques expérimentées jusqu'à maintenant.
Conclusions
Au siècle dernier, l'introduction d'espèces exotiques était systématiquement recherchée. Π existait par exemple, en France, une "Société nationale d'Acclimatation" dont l'objectif était d'acclimater en Europe des espèces d'intérêt économique, mais aussi, éventuellement, esthétique. LAMIRAL (1863) rapporte par exemple une tentative (infructueuse) d'acclimatation des éponges de Syrie dans ce qui sera, un siècle plus tard, le Parc National de Port-Cros (Var, France). La lecture des comptes-rendus montre que l'aspect ludique tenait en fait une part non négligeable dans ces activités : une sorte de jeu couronné par la réussite d'un élevage ou d'une acclimatation. La prise en compte de l'impact des introductions sur les espèces et les écosystèmes indigènes, tout comme la préservation des milieux naturels, ne faisaient pas partie, à l'évidence, de la culture de l'époque.

-27 -
Aujourd'hui, l'accélération des introductions d'espèces, le risque (à long terme) d'une homogénéisation planétaire des communautés pour une latitude ou un biotope considérés, constituent un problème majeur pour la communauté scientifique (SINDERMANN, 1991). Une réglementation appropriée permettrait sans doute de réduire le flux des introductions, mais certainement pas de l'arrêter. Comment par exemple arrêter le flux d'espèces indo-pacifiques via le canal de Suez ? serait-ce d'ailleurs souhaitable dans ce cas précis ?
Des intérêts économiques (introduction d'espèces aquacoles) s'opposent à d'autres intérêts économiques (introduction de maladies et de parasites, compétition avec des espèces indigènes exploitées, génération de nuisances) mais aussi à des valeurs éthiques difficilement chiffrables en termes économiques. Même si un taillis de figuier de barbarie produisait plus de biomasse et d'oxygène que la lande à Collima et bruyère de Bretagne, faudrait-il accepter son remplacement ? Peut-on accepter qu'une forêt d'Eucalyptus remplace les sansouires à Salicornia de Camargue ou la hêtraie de Fontainebleau (par ailleurs déjà plus ou moins artificialisée) ? Même s'il était démontré (par hypothèse) qu'un MacDonald's était plus rentable que Westminster Abbey, faudrait-il laisser détruire cette dernière ?
Les introductions d'espèces conduisent à poser le problème de la responsabilité des états et de sa limitation. Une fois introduite, que ce soit délibérément ou accidentellement, une espèce ne connaît pas les frontières administratives et politiques. Si Macrocystis pyrifera avait été introduit en France, il aurait colonisé les côtes européennes de l'Espagne.à la Norvège (BOALCH, 1981), avec des conséquences difficilement prévisibles sur les courants, la navigation côtière, la pêche et bien sûr les écosystèmes littoraux.
Les introductions d'espèces constituent peut-être l'un des problèmes écologiques majeurs du prochain siècle : en effet, contrairement à d'autres problèmes tels que la pollution ou les incendies de forêts, que l'on commence à savoir contrôler (plus ou moins) et qui sont réversibles, à court ou à moyen terme, les introductions d'espèces apparaissent, en l'état actuel des connaissances et des techniques, comme irréversibles. En outre, contrairement à l'aménagement du littoral, nous ne saurions pas geler la situation en l'état, même si nous le voulions. Enfin, aucun sanctuaire n'est à l'abri des introductions d'espèces : réserves naturelles, Parcs Régionaux, Parcs Nationaux, monuments naturels et patrimoine de l'UNESCO peuvent un jour protéger des écosystèmes imprévisibles dominés par des espèces introduites.

-28
The introduction of non-indigenous marine species to Europe Planktonic species
Gerald T. BOALCH
Plymouth Marine Laboratory, Citadel Hill, Plymouth PLI 2PB, U.K.
Introductions of non-indigenous species are of a number of types:
1) natural introductions which may be : a) temporary b) permanent
2) accidental introductions by man : a) brought in on, or with, other organisms b) brought in by ships or other vectors c) brought in for research or commerce and subsequently escaping
3) purposeful introductions
Natural introductions are frequently the result of local changes in environmental conditions, often due to slight climatic variations, so that a species normally occurring outside the area under consideration can extend its range and move into that area. The spread may be reversed when the environmental conditions revert to the more normal pattern.
Perhaps the most elusive planktonic alga in this group is Trichodesmium (Oscillatoria) erythraeum which originates in the warm waters of the Caribbean and is carried across the Atlantic on the Gulf Stream. The actual path of the Gulf Stream is to some extent controlled by meteorological conditions in the Atlantic and, under some conditions, water from the Gulf Stream is pushed into the English Channel (RUSSELL et ál., 1971) and then Trichodesmium occurs off Plymouth (BOALCH in PARKE and DIXON, 1976). Although undoubtedly still alive this alga does not appear to be dividing and certainly we have never been able to get it to grow in culture. Once the flow of Gulf Stream water changes and moves to its more usual course further north the alga disappears from the Channel. Maybe this is not really an introduction of this alga but its presence is enough for it to be included in check-lists, etc (PARKE and DIXON, 1976). Less ephemeral are the phytoplankton species which are brought into the Channel when water flows from the south-west.
Then species such as the diatoms Corethron criophilum (BOALCH, 1987) and Bacteriastrum ficrcatum, (BOALCH, 1974) the dinoflagellates Ceratium candelabrum and Ceratium hexacanthum (BOALCH, 1987) and the prasinophyte Halosphaera viridis (PARKE and DEN HARTOG-ADAMS, 1965) all come into the English Channel and may persist there for long periods after the flow of water from the south-west has ceased (BOALCH, 1987). In fact some of them may persist for at least a year and hence must be reproducing in the area and must be listed as species which have emigrated into the area. A similar pattern exists with

- 2 9 -
the movement of Thálassiothrix longíssima into the North Sea. In this case it is carried around the north of the British Isles (REID et al., 1992).
Similar patterns exist for seaweeds. Codium decortication was found once on the Isles of Scilly, presumably brought by warm conditions from the Bay of Biscay but it did not persist (unpublished observation). On the other hand, Laminaria ochroleuca (PARKE, 1948) and Cystoseira myriophylloides (DIXON, 1961) both came across to the south-west of England in the warm period prior to 1940 and have persisted, although after cold winters their distribution is restricted. It seems likely that Solieria chordalis and S. tenera (FARNHAM and JEPHSON, 1977; FARNHAM and IRVINE, 1979) are other such species. It is of interest that there is a species of hermit crab (Clibanarius erythropus) which appears to have come over from France some time before 1960 (SOUTHWARD and SOUTHWARD, 1977) and has now disappeared. Again it was holding on very precariously in Cornwall and seems to have disappeared from most of its sites in Cornwall as a result of the 'clean-up' after the Torrey Canyon oil spill (SOUTHWARD and SOUTHWARD, 1988).
These types of introductions do not appear to do any harm, after all they are only the extensions of the distribution of species that occurred not far away. Even if the new arrivals become established permanently their existence is very precarious and they are not likely to be aggressive enough to disturb the native species or to be abundant enough to have any marked toxic effects.
Accidental introductions are much more numerous. We have no evidence of planktonic species being brought in with other organisms although this could well have occurred. There are however a considerable number of seaweeds and invertebrates which have been brought in with Japanese oysters. The best known seaweeds are Sargassum muticum (FARNHAM et al. 1973) and Undariapinnatifida (PEREZ et al., 1981) but no doubt other speakers will list many more. The brown seaweed Colpomenia peregrina probably came to Europe with other oysters and this has caused problems for oyster farmers (COTTON, 1908). It was probably also the importation of oysters that introduced the oyster drill (Urosalpinx cinerea) (ORTON and WTNCKWORTH, 1928) and the slipper limpet (Crepidula fornicata) (DODD, 1894) to British waters.
Organisms brought in by ships or other vectors are probably very numerous. In the phytoplankton it is likely that ballast water or bilge water was the source of the introduction of the diatoms Biddulphia sinensis (OSTENFELD, 1908), Thalassiosira angstii (BOALCH, 1987), Coscinodiscus wailesii ßOALCH and HARBOUR, 1977b), and Pleurosigma planktonicum (BOALCH and HARBOUR, 1977a). All of these appeared in European waters when their previous known distribution did not include the North Atlantic. The same is probably the case for the dinoflagellate Gyrodinium aureolum (HANSEN and SARMA, 1969, BOALCH, 1979,1987), although this was known from the western side of the North Atlantic (HULBURT, 1957). In each case soon after the first appearance of each of these species it showed a period of phenominally rapid growth and the species rapidly spread from the area where it was first reported. The spread of Biddulphia sinensis was well documented by OSTENFELD (1908) using samples that were collected on cruises originally organised by ICEM. In fact, apart from the more recent case of Sargassum muticum in Europe (CRITCHLEY et al., 1983) this is probably the most thoroughly documented spread of a marine alga. Biddulphia sinensis did not have any harmful effect (OSTENFELD, 1908) and by the 1930s (LEBOUR, 1930) it had settled down to be a respected member of the phytoplankton. In more recent years it has been found to be a convenient species to culture and use in feeding experiments, etc. Thalassiosira angstii and Pleurosigma planktonicum

- 3 0 -
have also become important members of the phytoplankton and do not appear to have caused any problems (BOALCH, 1987). However, Coscinodiscus wailesii did cause some problems when it first arrived as it produced copious amounts of mucilage which blocked fishing nets (BOALCH and HARBOUR, 1977b). Now it seems to have settled down as a respectable member of the phytoplankton and does not become numerous enough to cause problems with its mucilage. On the other hand, Gyrodinium aureolum has caused many problems since it first appeared in European waters in 1968. It still produces blooms, and fish kills are often associated with these blooms (BOALCH, 1987). It is interesting that this species was first described from the Woods Hole area on the eastern Atlantic seaboard of the U.S.A. where it was not known to bloom and was never associated with fish kills (HULBURT, 1957). Yet when it arrives in a new environment it produces extensive blooms and is associated with kills of both vertebrates and invertebrates. The evidence is that it produces a chemical which attacks cell membranes, especially gill membranes, and the victims lose a lot of blood and die (FORSTER, 1979; GRIFFITHS et al., 1979). These last two algae are certainly cases of introductions which have had harmful effects.
It is likely that some seaweeds have been introduced into Europe on boats, the most probable ones being Asparagopsis armata (DEVALERA, 1942) and Codium fragile (SILVA, 1955). Both of these have spread widely and have not caused any noticeable harmful effects. The two best known invertebrates that have been brought to Europe, probably by ships, are the barnacles (Elminius modestus and Balanus amphitrite var. deruiculata) (CRISP and CHIPPERFEELD, 1948; CRISP and MOLESWORTH, 1950; CRISP, 1958; SOUTHWARD and CRISP, 1963 ), the spreads of which have been well documented.
Again there are no records of phytoplankton species that have been brought in for research and have escaped, although with all the research with toxic species that is now going on at Marine Stations it will not be long before something does escape and develop into a toxic bloom. The best recorded cases of seaweeds that have escaped from experimental conditions are Caulerpa taxifolia (BOUDOURESQUE et al., 1992) and Undariapinnatifida (IRVINE et al., 1988) There are also several molluscs which have escaped from artificial culture experiments. All these are now well established and spreading fast.
Purposeful introductions amongst the algae are relatively few and none are known for planktonic species. Documented cases for seaweeds are the successful introduction of Undaria (PEREZ et al., 1984) to the Atlantic and Channel coasts of France and the unsuccessful attempt to introduce Macrocystis to British coasts in the 1950s (BOALCH and POWELL, 1975) and to the Channel coast of France in the 1970s (PEREZ, 1972).
What lessons should we learn? The main conclusion is that it is impossible to predict exactly how a species will behave when it is introduced into a new environment. Sargassum muticum is a rather insignificant species not exceeding half a meter on the coast of China, but in the English Channel it is completely dominating some shores and may exceed 8 m in length. Gyrodinium aureolum does not bloom on the east coast of North America and hence was not known as a toxic species. Coscinodicus wailesii on the north west coast of North America never forms a bloom and nothing was known about the large amounts of mucilage it secretes. Because of this unpredictability every care should be taken to prevent the introduction of species from one ecosystem into another. In these days of rapid means of transport, including fast ships, it is quite easy for marine organisms to be carried across the world either purposely or inadvertently. How to prevent this continual transfer of species from one ecosystem to another is a big problem. It also appears that there is inevitably 'leakage' from experiments with introduced species that eventually leads to the establishment of that species in a new environment.

31
Acknowledgements
The author expresses his grateful thanks to many of his colleagues, especially Professor A.J. SOUTHWARD, for help in tracing the records of immigrant species.

32
Introduction of marine benthic algae into Atlantic European waters
William F. FARNHAM
Marine Laboratory, University of Portsmouth, Hayling Islands, Hants. POH ODG, England
Additions to the British and mainland European marine floras may consist of new species being described e.g. Schmitzia hiscockiana and Pneophyllum lobescens, discovery of already known species (usually inconspicuous or microscopic forms in those areas where the flora is well known e.g. Streblonema oligosporum and Titanoderma laminariae) or the discovery of non-native seaweeds. Often, the latter finds have been reported from well-worked regions and so, presumably, represent quite recent arrivals.
Indeed, many species of exotic algae have been reported in the past for British Isles and elsewhere in Europe (FARNHAM, 1980). WALLENTINUS (in press) produced an updated list of European algal introductions. WALFORD and WICKLUND (1973) had earlier published a world-wide inventory of aliens but this was limited to marine animals only. HUTCHINGS et al. (1987) discussed the arrival of non-indigenous species in Australian waters. It is worth noting that only a few European algae seem to have been reported as aliens in other parts of the world e.g. Striarla attenuata into Australia (WOMERSLEY, 1987). RUSSELL (1981) reported on the introduction of two red algae into Hawaï. Quite likely, there are still other algal aliens waiting to be recognized in Europe and elsewhere, and, undoubtedly, introductions will continue to occur in the future. Thus, we are dealing with an on-going historical process. Table 1 lists seaweed introductions into Europe, excluding the Mediterranean. Localities and dates are not definitive!
This phenomenon of alien species pose various questions and problems, which have not been fully resolved for any one putative alien. The issues involved include :
1. Recognition, 2. Place of arrival, 3. Date of arrival, 4. Country of origin, 5. Mode of arrival (carrying agent ?), 6. Further spread (how far and how fast ?), 7. Any ecological effects on indigenous species and communities, 8. Detrimental effects on Man's activities, 9. "Success", 10. Predictability, 11. Control of "pest" introductions.
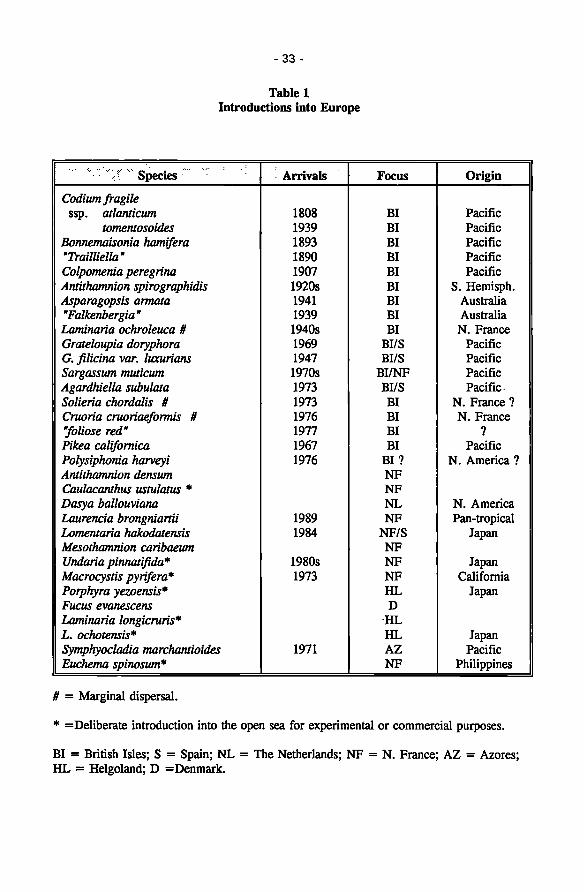
- 3 3 -
Table 1 Introductions into Europe
"A Species
Codium fragile ssp. atlanticum
tomentosoides Bonnemaisonia hamifera "Trailliella" Colpomenia peregrina Antithamnion spirographidis Asparagopsis armata "Falkenbergia" Laminaria ochroleuca # Grateloupia doryphora G. filicina var. luxurians Sargassum muticum Agardhiella subulata Solieria chordalis # Cruoria cruoriaeformis # "folióse red" Pikea californica Pofysiphonia harveyi Antithamnion densum Caulacanthus ustulatus * Dasya ballouviana Laurencia brongniartii Lomentaria hakodatensis Mesothamnion caribaeum Undaria pinnatifida* Macrocystis pyrifera* Porphyra yezoensis* Fucus evanescens Laminaria longicruris* L. ochotensis* Symphyocladia marchantioides Euchema spinosum*
Arrivals
1808 1939 1893 1890 1907 1920s 1941 1939 1940s 1969 1947 1970s 1973 1973 1976 1977 1967 1976
1989 1984
1980s 1973
1971
Focus
BI BI BI BI BI BI BI BI BI
BI/S BI/S
BI/NF BI/S BI BI BI BI
BI? NF NF NL NF
NF/S NF NF NF HL D
HL HL AZ NF
Origin
Pacific Pacific Pacific Pacific Pacific
S. Hemisph. Australia Australia N. France
Pacific Pacific Pacific Pacific
N. France ? N. France
? Pacific
N. America ?
N. America Pan-tropical
Japan
Japan California
Japan
Japan Pacific
Philippines
# = Marginal dispersal.
* =Deliberate introduction into the open sea for experimental or commercial purposes.
BI = British Isles; S = Spain; NL = The Netherlands; NF = N. France; AZ = Azores; HL = Helgoland; D =Denmark.

- 3 4 -
Often, adventitious species have not straightway been recognized as such because of taxonomic problems, e.g. the separation of Colpomenia peregrina from C. sinuosa, and Codium fragile from C. tomentosum. These may also apply to suggesting the geographical origin of alien. Concern has also been expressed as to whether young ecologiste now have the necessary systematic expertise for recognizing "strangers".
"Success" (from the point of view of the intruder) is a vague and subjective term related to 6 and 7. It can be defined by the overall impact of an invasive species in a new biotope and its final distribution. Some adventives do not spread much beyond their initial sites of colonization e.g. Grateloupia filicina var. luxurians in the central, southern coast of England. Others, in time, spread further but remain inconspicuous components of the ecosystem e.g. Colpomenia. Presumably, many potential introductions are transported to new regions but fail to become established. This could be due to a number of factors : poor survival en route; insufficient inoculum; lack of suitable habitats; unfavourable environmental conditions for survival (growth, reproduction), etc. A folióse red was found some years ago in Falmouth, which could not be identified, but was a new alga, for at least, the British Isles. Subsequent searches have failed to refind it. I conclude that it failed to survive and establish a population there because conditions were insuitable. In contrast, other algae have spread considerable distances and have become ecologically significant members of marine communities. The outstanding instance in European waters is, of course, Sargassum muticum.
In some 20 years since its initial discovery in European waters, it has become dominant in many lower littoral areas with standing water and in the sublittoral fringe, covering a range of conditions and habitats. It has spread extensively in European waters from Portugal to Scandinavia, and even in the Mediterranean - its latest sighting there being in Venice Lagoon. Sargassum has generally performed as an opportunist "gap-grabber", without significantly displacing or outcompeting native benthic plants, as had originally been feared. It can even be argued that S. muticum has increased productivity and species-diversity (rich epibionta). However, many would consider that its prolific growth has reduced the amenity value of some coastal areas (see 8 above). CRITCHLEY et al. (1990) have compiled a bibliography on this "weed".
Looking ahead, what predictions can be made concerning future introductions? What site features determine whether an ecological system will be prone, or resistant to an invasion? Are there particular species which may be introduced (deliberately or by chance) to look out for ? What successful invader ? Past experience tells us that organisms invading new places may perform ecologically quite differently from when in their native habitat.
From the above, it may then be appropriate for consideration to be given as to what control or legislation (both national and international) is required or feasible, particularly in the case of those aliens being regarded as potential "pests".
Many European exotic algae seem to have originated from as far as the Pacific e.g. Codium fragile. Table 2 lists various carrying-agents and dissemination mechanisms which may be responsible for the arrival and further spread of exotic species.

- 3 5 -
Table 2 Dissemination and vectors
Remote
Marginal
Vector
International
Fortuitous
Mariculture
Fouling Ballast : solid
water
Epibionts on introduced hosts
Shellfish packing Aquarium escapes
Currents : spores detached fronds
Stone-rafting
"Alien" structures e.g. oil-rigs
Examples
Macrocystis Undaria
? Solierìa Β. sinensis
Spirorbids on S. muticum
S. muticum Caulerpa
S. muticum
Solierìa
?
CRIPS (1958) used the term "remote dispersal" for the long-distance transportation of organisms, which would seem to implicate carrying-agents or "vectors" (sensu CRIPS, 1958; JONES, 1974). Even in the case of unintended or accidental introductions over long distances, human (synanthropic) involvement is implied. For most introductions, the vector is suggested by circumstantial evidence related to the initial localities. It is unusual for a vector to be caught in flagrante. Thus, the role of shipping is suggested by the occurence of many aliens in the vicinity of ports and harbours. These have often been foci for newly introduced species e.g. FALMOUTH (Bonnemaisonia hamifera) and GALWAY (Asparagopsis). In addition to such fortuitous events, deliberate introductions may be instigated for commercial exploitation e.g. Undaria into Brittany for the alginate industry.
"Marginal dispersal" refers to minor expansions in range-limits of populations of a species, which are brought about by natural methods of dissemination. Some workers do not consider such cases as constituting true "introductions" (CARLTON, 1975) but, as to how species arrived usually can only be surmised, it seems advisable to regard any alien species which has arrived in a new territory as constituting a (potential) introduction. The red alga Solieria chordalis may, like Laminaria ochroleuca, have spread across the Channel from N. France. Alternatively, its scattered British records could represent previously undiscovered relict populations.

- 3 6 -
Some seaweeds have probably spread by various mechanisms of both remote and marginal dispersal, as in the case of S. muticum - international transportation of shellfish (weed used for packing, germlings attached to shells), fronds wrapped round propellors and floating fronds dispersed by wind-induced surface currents.
Many of the earlier introductions have become so well-established ("naturalized") that they are now regarded as part of our flora e.g. Colpomenia peregrina (brown alga), Bonnemaisonia hamifera (red alga) and Codium fragile (green alga). The latter may have outcompeted in some areas of the British Isles a similar but indigenous species, C. tomentosum. Perhaps, some species, seemingly autochthonous, may have been originally of foreign origin.This might be indicated by disjunct, biogeographical distributions.
The "success" of an introduced species may be defined its distribution (how far and how fast) and what effect it has had on natural communities. Some aliens are insignificant by these criteria e.g. Grateloupia doryphora in the Solent (although recently discovered in Jersey and N. Spain), while others have been quite remarkable. Probably the best and most topical example is the "Japweed", Sargassum muticum.
I consider that the early recognition and surveillance of introduced species are important to detect and monitor any consequrent changes in the ecosystems invaded. Great caution should be exercised before species are deliberately introduced for exploitation, as with various proposals for the giant kelp Macrocystis. The "invasive potential" of introduced organisms is unpredictable. Unintended adventives may also be transferred as contaminants (parasites or epibionts) if quarantine procedures are lacking or ineffectual. Perhaps evaluation of such procedures should be initiated, including monitoring of discharged ballast waters.
It is worth recalling ELTON'S warning in 1958 with respect to unintended introductions that "the greatest agency of all that spreads marine animals to new quarters of the world must be the business of oyster culture". DRUEHL (1973) even predicted the occurence of 5. muticum in Europe resulting from importation of Pacific oysters.
Thoughts
1. Is the rate of introductions changing ? Same, more or fewer ? 2. How often do we recognize them ? Poor taxonomy or time lag in their discovery ? 3. How well do we know the native European flora - ecologically and taxonomically ? 4. How to assess whether an alien is detrimental or beneficai ? Who should be responsible
for this ? 5. If a "pest", what control measures are feasible or desirable ? See CRtTCHLEY et al.
(1986) for an account of attempted control of Sargassum.

-37 -
Les macrophytes marins introduits en Méditerranée : biogéographie
Maria A. RIBERA
Laboratori de Botànica, Facilitât de Farmacia, Universität de Barcelona, Espagne.
La flore marine de Méditerranée commence à être relativement bien connue, de même que la distribution géographique des espèces qui la constituent (RIBERA et ál., 1992); il devient alors possible de considérer qu'un certain nombre d'espèces sont probablement introduites.
Nous avons recensé 45 espèces (28 Rhodophyta, 10 Fucophyceae, 6 Chlorophyta et 1 Phanérogame) de macrophytes probablement introduites en Méditerranée (Tabi. I). Nous en avons exclu des espèces telles que Polyphysa párvula et Acetabularia calycuius, qui ont souvent été considérées comme des immigrants lessepsiens (POR, 1978), mais dont la présence en Méditerranée est probablement ancienne, compte tenu de leur présence dans l'Atlantique tropical et de leur découverte aux Acores et à Madère (VALET, 1969). Inversement, nous y avons inclus une espèce qui n'est pas connue en dehors de la Méditerranée, Amithamnion algerierne; en effet, cette espèce a été décrite d'Algérie en 1991 (VERLAQUE et SERIDJI, 1991); depuis lors, elle a été trouvée en mer d'Alboran (RIBERA-SIGUAN et SOTO-MORENO, 1992), puis en abondance aux îles Mèdes (Espagne) (E. BALLESTEROS, comm. verb.), dont la flore a été bien étudiée (ROS ARAGONÉS et al., 1984; BALLESTEROS, 1990), de telle sorte qu'il est peu probable que cette espèce ait pu y passer inaperçue; il semble donc qu'A, algeriense soit une espèce introduite, bien que sa région d'origine ne soit pas encore connue.
L'ouverture du canal de Suez (19 à 22 espèces) et l'ostréiculture (9 à 10 espèces), avec en particulier l'importation de naissains de l'huître Crassostrea gigas du Japon, constituent les deux principales causes d'introductions d'espèces en Méditerranée (Tabi. I et II). Il en résulte que l'Indo-Pacifique (mer Rouge et Japon compris) constitue le réservoir d'où proviennent la plupart (31 à 33) des espèces introduites (Tabi. Ill); on peut d'ailleurs y ajouter quatre espèces qui ont colonisé la Méditerranée par Gibraltar, à partir de populations préalablement introduites en Atlantique, mais provenant en réalité de l'Indo-Pacifique : Bonnemaisonia hamifera, Asparagopsis armata, Colpomenia peregrina et Codium fragile. En fait, la part prise par les espèces indo-pacifiques constitue peut-être un artefact, par sous-estimation des introductions d'origine atlantique; la plus grande partie de la flore méditerranéenne est constituée par des espèces dont l'aire de distribution est centrée sur l'Atlantique (FELDMANN, 1938, 1968); il en résulte que, lorsqu'une espèce, déjà connue de l'Atlantique, est rencontrée pour la première fois en Méditerranée, on ne l'interprète généralement pas comme une introduction mais comme une meilleure connaissance de son aire de répartition.
Les espèces introduites ne se répartissent pas de façon aléatoire entre les différents biotopes. Les espèces lessepsiennes se sont concentrées dans un petit nombre de biotope; elles ont surtout colonisé le biotope le plus répandu le photophile infralittoral, dans la mesure où la flore de Méditerranée orientale est très pauvre (faible biodiversité) et où la compétition y est relativement faible (Tabi. IV). Si l'on ne considère que les espèces non lessepsiennes (surtout

- 3 8 -
présentes en Méditerranée occidentale), on constate que, au contraire des espèces lessepsiennes, elles ont colonisé une assez grande variété de biotopes. Par ailleurs, la quantité d'espèces introduites est inversement proportionnelle à la biodiversité des biotopes : ce sont les milieux lagunaires, à faible biodiversité, qui en ont accueilli le plus (11), devant les biotopes photophiles (4 à 7) et l'herbier à Posidonia oceanica (1) (Tabi. IV). On pourrait penser qu'il y a là un artefact dû au fait que l'aquaculture se pratique en étang; en réalité, il existe également de nombreux sites aquacoles en mer ouverte, comme par exemple la rade de Toulon, le golfe de Fos et Séte pour ce qui concerne les côtes françaises, et ils ne semblent pas avoir donné lieu à des introductions.
Tableau I
Macrophytes introduits en Méditerranée. Probabilité d'introduction : H = haute, M = moyenne, F = faible. Modalités d'introduction : S = par le canal de Suez, G = par Gibraltar, FO = fouling sur coque de navire, O = ostréiculture, AP = appâts de pêche, AQ = aquariums. Origine : A = Atlantique, MR = mer Rouge, IP = Indo-Pacifique, J = Japon, PT = pantropicale, MA = mers australes. Par biotope colonisé en Méditerranée, nous entendons le biotope des premières signalisations. HP = herbier à Posidonia oceanica, Photo = biotopes photophiles infralittoraux en mer ouverte, Scia = biotopes sciaphiles en mer ouverte, Médio = médiolittoral. Données d'après ALEEM (1948, 1950), ANONYME (1982), BELSHER et al. (1984), BEN MAIZ et al. (1987), BOUDOURESQUE et al. (1985), CINELLI et SARTONI (1969), CORMACI et al. (1991,1992), DIAPOULIS et al. (1985), FELDMANN (1931, 1939, 1956), FELDMANN et FELDMANN (1938), FORTI (1928), FRITSCH (1895), HAMEL (1926, 1930a, 1930b), LIPKIN (1972), MEINESZ et HESSE (1991), MÉNDEZ DOMINGO (1957), PEREZ et al. (1981), PETERSEN (1918), POR (1978), RAMON et FRIEDMANN (1965), RAYSS (1941), RIOUALL et al. (1985), SANCHOLLE (1988), VERLAQUE (1981, 1989), VERLAQUE et BOUDOURESQUE (1991), VERLAQUE et RIOUALL (1989), VERLAQUE et SERIDJI (1991).
(1) Il s'agit soit d'une relique de la Téthys (CORMACI et al., 1982), soit d'un immigrant prélessepsien, entré en Méditerranée par les canaux creusés par les anciens égyptiens (ALEEM, 1948). (2) RAYSS (1963) a avancé qu'il pourrait s'agir d'une relique de la Téthys. (3) La possibilité qu'il s'agisse d'une espèce prélessepsienne a été envisagée (PERES, 1967) mais est très improbable. (4) Inclut Hypnea valiantiae qui, selon MAYHOUB (1976), est synonyme d'H. hamulosa. (5) Les signalisations de Sarconema scinaioides et de S. filiforme en Méditerranée correspondent peut-être à une même espèce. (6) Les signalisations de Sarconema furcellatum sont inclues sous S. filiforme, les deux espèces ayant été mises en synonymie (PAPENFUSS et EDELSTEIN, 1974). (7) Selon MAYHOUB (1976), synonyme de H. hamulosa. (8) Présent dans l'étang de Thau (France) selon BEN MAIZ et BOUDOURESQUE (inédit).
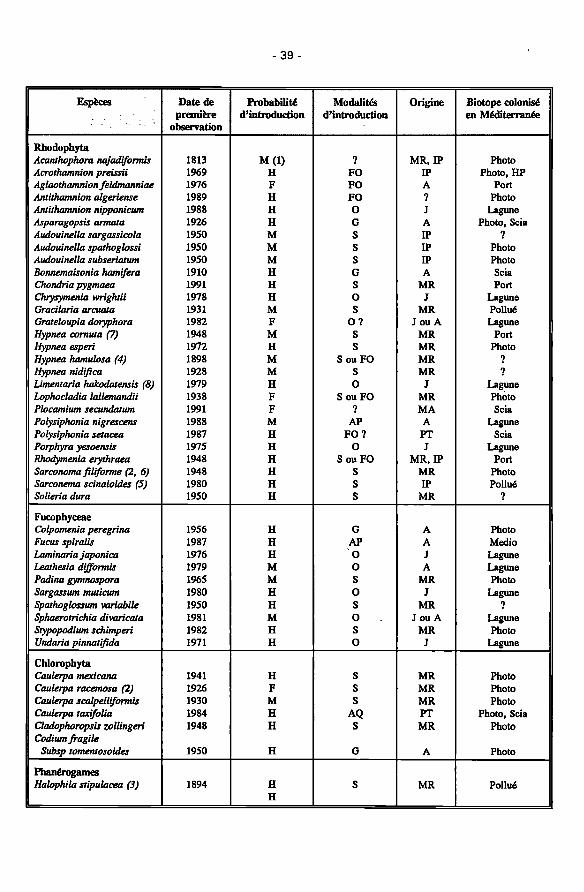
-39
Espèces
Rhodophyta Acanthophora najadiformis Acrothamnion preissii Aglaothamnion feldmanniae Antithamnion algeriense Antithamnion nipponicum Asparagopsis armata Audouinella sargassicola Audouinella spathoglossi Audouinella subseriatum Bonnemaisonia hamifera Chondria pygmaea Chrysymenia wrightii Gracilaria arcuala Grateloupia doryphora Hypnea cornuta (7) Hypnea esperi Hypnea hamulosa (4) Hypnea nidifica Limemaria hakodatensis (δ) Lophocladia lallemandii Plocamium secundatum Polysiphonia nigrescens Polysiphonia setacea Porphyra yesoensis Rhodymenia erythraea Sarconoma filiforme (2, 6) Sarconema scinaioides (5) Solieria dura
Fucophyceae Colpomenia peregrina Fucus spiralis Laminaria japónica Leathesia difformis Padina gymnospora Sargassum muticum Spathoglossum variabile Sphaerotrichia divaricata Stypopodium schimpen Undaria pinnatifida
Chlorophyta Caulerpa mexicana Caulerpa racemosa (2) Caulerpa scalpelliformis Caulerpa taxi/olia Cladophoropsis zollingeri Codium fragile
Subsp tomentosoides
Phanérogames Halophila stipulacea (3)
Date de première
observation
1813 1969 1976 1989 1988 1926 1950 1950 1950 1910 1991 1978 1931 1982 1948 1972 1898 1928 1979 1938 1991 1988 1987 1975 1948 1948 1980 1950
1956 1987 1976 1979 1965 1980 1950 1981 1982 1971
1941 1926 1930 1984 1948
1950
1894
Probabilité d'introduction
M ( l ) H F H H H M M M H H H M F M H M M H F F M H H H H H H
H H H M M H H M H H
H F M H H
H
H H
Modalités d'introduction
? FO FO FO O G S S S G S O s
o? s s
S o u FO S O
S o u FO 7
AP FO?
O S o u FO
S S S
G AP Ό O S O
s o s o
s s s
AQ S
G
S
Origine
MR, IP Π» A 7 J A IP IP IP A
MR J
MR J ou A
MR MR MR MR
J MR MA A
PT J
MR, IP MR IP
MR
A A J A
MR J
MR J ou A
MR J
MR MR MR PT MR
A
MR
Biotope colonisé en Méditerranée
Photo Photo, HP
Port Photo
Lagune Photo, Scia
? Photo Photo Scia Port
Lagune Pollué Lagune
Port Photo
? 7
Lagune Photo Scia
Lagune Scia
Lagune Port
Photo Pollué
?
Photo Medio Lagune Lagune Photo
Lagune 7
Lagune Photo
Lagune
Photo Photo Photo
Photo, Scia Photo
Photo
Pollué

40
Tableau Π : Modalités d'introduction des macrophytes marins en Méditerranée.
Modalités d'introduction
Canal de Suez Gibraltar Ostréiculture Fouling Appâts de pêche Aquariums Inconnues
Nombre d'espèces
19 à 22 4
9 à 10 3 à 7
2 1
2 à 3
Tableau ΙΠ : Origine géographique des macrophytes marins introduits en Méditerranée.
Zone géographique
Indo-Pacifique (Mer Rouge et Japon inclus) Zone pantropicale Atlantique Mers australes Inconnue
Nombre d'espèces
31 à 33 2
8 à 10 1 1
Tableau IV : Biotopes colonisés par les macrophytes introduits en Méditerranée (nombre d'espèces).
Biotope
Photophile infralittoral
Sciaphile Médiolittoral Herbier à Posidonia Lagunes Pollué Inconnu
Espèces lessepsiennes
10 à 11
— — — —
5 à 6 4 à 5
Espèces non lessepsiennes
4 à 7 f
3 à 5 1 1 11
l à 2 O à l
La progression des espèces introduites peut se faire de façon naturelle ou bien avec l'aide de l'homme. Le cas le plus simple de progression naturelle est l'expansion dans le sens du courant dominant; c'est ainsi qu'en Méditerranée orientale Halophila stipulacea a atteint les côtes levantines, turques, grecques, albanaises, maltaises et italiennes (BILIOTTI et ABDELAHAD, 1990; HARTOG, 1972), se propageant dans le sens du courant (sens

-41
contraire aux aiguilles d'une montre), alors que, dans l'autre sens, il n'a pas dépassé Marsah-Matrouh, dans l'Ouest égyptien (ALEEM, 1962). A une échelle moindre, Acrothamnion preissii, parti de Livorno (Italie) (CINELLI et SARTONI, 1969), a progressé vers Nice (BOILLOT et al., 1984), puis la côte varoise (France) (THELIN, 1984); en sens contraire, vers le Sud, il n'a pas dépassé la latitude de l'île d'Elbe, ce qui représente une progression de moins de 100 km (CINELLI et ál., 1984) Fig. 1). De même, Undaria pinnatifida (BOUDOURESQUE et al., 1985) et Sargassum muticum (KNOEPFFLER-PEGUY et al., 1985; BELSHER et POMMELEC, 1988), à partir de l'étang de Thau (France), ont étendu leur aire en direction de l'Espagne (qui n'est pas encore atteinte) (Fig. 1). Un autre cas de progression naturelle est la diffusion : à partir de son point d'introduction, l'espèce progresse dans toutes les directions, sans qu'une direction paraisse privilégiée; c'est le cas de Stypopodium shimperi qui, à partir du canal de Suez, s'est autant étendu dans un sens (Israël, Syrie, Chypre et Turquie) que dans l'autre (Egypte, Libye) (VERLAQUE et BOUDOURESQUE, 1991) (Fig. 2). La progression locale (dans les Alpes-Maritimes, France) de Caulerpa taxifolia est du même type (MEINESZ et HESSE, 1991). Un troisième type de progression est la progression irrégulière; ce cas est illustré par Codium fragile subsp. tomentosoides : signalé pour la première fois dans les Pyrénées-Orientales (France) (FELDMANN, 1956), il apparaît successivement en des lieux proches ou éloignés, sans relation apparente avec le sens des courants et la distance (Fig. 3). D'une façon générale, la progression par diffusion est réalisée par des espèces dont la niche écologique (tout au moins en Méditerranée) est relativement large, de telle sorte que la progression peut se faire de proche en proche et ne dépend pas du hasard de la rencontre de biotopes favorables. Inversement, la progression irrégulière, et dans une moindre mesure la progression dans le sens du courant, concernent des espèces à niche écologique étroite.
Montpellier mo.¿#£i985,
Perpignan 3 \
Fig. 1 : Chronologie de l'expansion d'Acrothamnion preissii (Rhodophyta; trait plein) et de Sargassum muticum (Fucophyceae; trait discontinu) en Méditerranée occidentale.

42
La dissémination due à l'homme prolonge les causes de l'introduction d'une espèce : aquaculture, aquariologie, mouillages. Il s'agit en fait d'introductions multiples. Il en résulte une progression par bonds, sans stations intermédiaires. C'est le cas à'Undaria pinnatifida, introduite en Méditerranée dans l'étang de Thau (France) avec le naissain de l'huître japonaise Crassostrea gigas, au début des années 70 (BOUDOURESQUE et al, 1985), et qui vient d'être signalée dans la lagune de Venise (Italie), où cette huître est également élevée (S. VOLPI, coram, verb.). C'est le cas également de Caulerpa taxifolia, qui a été introduite accidentellement, sans doute, à partir de rejets distincts d'aquariums, dans la région de Monaco (MEINESZ et HESSE, 1990) et à Saint-Cyr (J. LABOREL, comm. verb.), et signalée depuis à Mallorca (Baléares) et Imperia (Ligurie, Italie) disséminée, peut-être, de mouillage en mouillage (MEINESZ et al., 1993).
Les modalités de dissémination propres à chaque espèce jouent un rôle important, d'une part dans son mode d'introduction et d'autre part dans les modalités de son expansion une fois introduite. Les espèces telles que Sargassum muticum, produisant des épaves flottantes, peuvent progresser par des bonds importants. Par contre, les espèces dont les épaves (boutures) ne flottent pas, telles que Caulerpa taxifolia, effectuent des sauts sur de plus courtes distances (si l'on met à part l'action de l'homme). De même, la longévité et le poids des spores et zygotes peuvent influer sur les distances de dissémination des espèces. La présence dans le cycle d'une génération (gamétophyte) ou d'un stade végétatif rampant (nématothalle) microscopiques constitue un élément très favorable à l'introduction : en effet, ils passent facilement inaperçus lorsqu'ils sont fixés sur des espèces aquacoles, et ils peuvent être transportés sur de longues distances sur des coques de bateaux (fouling).
L'apport de gènes nouveaux à des espèces -déjà présentes en Méditerranée, à partir de populations extérieures, est probable, bien que peu documenté. On sait que des Hypnea musciformis en provenance du Sénégal ont été cultivés en Corse au début des années 80 (MOLLION, 1984); l'appel à des souches exotiques s'expliquait par le fait que, paradoxalement, on n'arrivait pas à cultiver les souches indigènes (ce qui indique des différences génétiques). De même, les importations de naissains de l'huître Crassostrea gigas à partir de bassins ostréicoles des côtes atlantiques ont probablement introduit en Méditerranée des souches atlantiques d'espèces déjà présentes en Méditerranée.
Fig. 2 : Chronologie de l'expansion de Stypopodium schimpen (Fucophyceae)en Méditerranée orientale (d'après VERLAQUE et BOUDOURESQUE, 1991).

-43
Propositions de recherches à effectuer : (i) Les principales sources d'espèces introduites étant le canal de Suez et l'aquaculture (tout particulièrement en lagunes littorales), il conviendrait d'explorer systématiquement ces zones; il est certain par exemple que l'étang de Thau renferme plusieurs espèces introduites non encore inventoriées, (ii) Vérifier la plus grande résistance des écosystèmes à forte biodiversité en démontrant que les sites aquacoles de mer ouverte sont contaminés, mais que les introductions y sont généralement un échec, (iii) Rechercher les différences morphologiques, physiologiques, biologiques entre les populations des régions d'origine et la population introduite, supposée ne posséder qu'une partie de leur variabilité génétique, (iv) Dans le domaine de la recherche appliquée, il est indispensable de déterminer les modalités de décontamination des produits aquacoles transportés d'une région à l'autre; en France et pour les naissains d'huîtres, il est prévu un séjour d' une heure en eau douce, mais cette règle (d'ailleurs non appliquée) n'a aucune base expérimentale.
Recommandations aux décideurs : (i) En fonction des connaissances sur les caractéristiques biologiques des espèces introduites, de leurs modalités de dissémination, ainsi que des caractéristiques des sites d'introduction, on formulera des prédictions sur la direction et la vitesse de leur expansion. Ces prédictions pourront être exploitées par les décideurs dans le cas des espèces considérées comme des nuisances (exemple : Sargassum muticum). (ii) Les côtes de la Communauté européenne appartiennent à des provinces biogéographiques très diverses; c'est également le cas à l'intérieur de plusieurs de ses pays : Espagne (Canaries et Baléares par exemple), France (côtes atlantique et méditerranéenne), Italie (Sardaigne et Nord-Adriatique). Des mouvements de matériel vivant entre ces provinces (aquaculture, aquariologie, appâts de pêche) sont donc susceptibles de constituer un risque d'introduction. On délimitera donc les provinces biogéographiques entre lesquelles tout transport de matériel biologique devrait être soumis à autorisation, contrôle et/ou à décontamination.
Fig. 3 : Chronologie de l'expansion de Codium fragile (Chlorophyta) en Méditerranée.

- 4 4 -
Introduced invertebrates : examples of success and nuisance in the European Atlantic and in the Mediterranean
Helmut ZIBROWIUS
Centre d'Océanologie de Marseille (CNRS URA 41), Station Marine d'Endoume, Rue Batterie des Lions, 13007 Marseille, France
Worldwide, both accidentally and intentionally, man has been responsible for the introduction of a great number of marine species that subsequently established reproducing populations far away from their native areas. Nonindigenous marine species are now a serious and increasing problem for all geographic areas, especially in near-shore environments characterized by an instability of abiotic parameters and a lower species diversity. Although examples of recognized marine introductions date back to the 19th century, concern about the impact of terrestrial and freshwater aliens widely preceded awareness that nonindigenous marine species may be harmful to fisheries, aquaculture and to man's other marine activities.
As a result of introductions, near-shore ecosystems in the European Atlantic and in the Mediterranean were locally modified and now permanently include non-indigenous species, some of which have a strong impact as consumers in the food chain. While some of the newcomers became essential for commercial practices, others acquired considerable negative economic importance by fouling, competing with or predating on commercially used species, and even by endangering human health (see SPANIER & GALIL, 1991, for the invasion of the Levant by the scyphomedusa Rhopilema nomadica).
The discovery of previously unrecorded species suspected to be introductions always needs careful analysis of the circumstances and cautious interpretation. In spite of a long tradition of marine biological research carried on in Western European and Mediterranean waters, reliable monographic studies are not available for all zoological groups. As in the past, persisting taxonomie difficulties and lack of specialists may still lead to confusions and obscure introductions. In the case of confirmed aliens, one has to be aware that arrival and firm establishment may precede considerably actual detection.
The first (accidental) marine introductions to the European Atlantic and to the Mediterranean may have occurred with the beginning of transoceanic navigation. Means of arrival for exotic species subsequently diversified and now include : transport on ship hulls, drilling platforms and other towed man-made structures; transport with ballast water (in former times transport with solid ballast such as sand and stones); intentional introduction of species in view of commercial use; accidental introduction of species accompanying intentionally introduced species; discarding from the market and back to the sea of imported live specimens, eventually together with an unnoticed accompanying fauna (this may include species used as bait and the fauna of algae used as packing material); and, potentially increasing in importance, escape from aquaria and laboratory cultures when these are run without a treatment of the effluent waters. These possible avenues of introducing alien species operate worldwide and are commonly discussed in the specialized literature (e.g. CARLTON, 1987, 1989; FOSTER & WILLAN, 1979; MILLER 1969; WILLIAMS et al., 1988).

- 4 5 -
In the case of the Mediterranean with its well defined landlocked area, migration of Red Sea species through the Suez canal (opened in 1869) has added by far the greatest number of newcomers, including beneficial, indifferent and noxious species. Lessepsian migration has a still increasing impact on the eastern basin and undoubtedly is the best documented type of marine biological invasion modifying the Mediterranean fauna (POR, 1978, 1990; SPANIER & GALEL, 1991). Syntheses for the main lessepsian migrant groups (molluscs, decapod crustaceans, fishes) are frequently updated. Accordingly, this compartment will not be discussed further in this contribution.
Invaders arriving otherwise, of other origins, and not being part of a spectacular canalized flow, have not been the subject of inventories as representative as those of lessepsian migrants. In the following, some examples are given of successful nonindigenous species, accidentally introduced by means of oyster movements and navigation. For an abundant literature introducing to general topics and to case histories the reader is referred to CARLTON (1987, 1989), ZIBROWIUS (1983, 1992) and ZIBROWIUS & THORP (1990).
Accidental introductions by oyster movements
Oysters are by far the most transferred commercial shellfish. Large scale movements started in the 19th century, by ship and rail. Air freight started to be used in the 1960s. Various alien oyster species have thus been brought to the Atlantic coasts of Europe and to the Mediterranean, together with the accompanying fauna and specific oyster nuisances. It is likely that not all these introductions left traces in official records.
From 1861 to 1939, the American Atlantic oyster (Gulf of St. Lawrence to Panama) Crassostrea virginica (Gmelin, 1791) was repeatedly imported into British and Irish waters and relaid on the oyster beds to supplement the larger summer market demand. It never became established in Britain and attempts to establish it in France and in Denmark were similarly unsuccessful. Nevertheless, C. virginica acted as the vector of introduction of various species from Eastern North America, including : an ostracod, a maldanid polychaete, possibly a serpulid polychaete that also travelled as a ship fouler (Hydroides dianthus), and the two undesirable prosobranch gastropods Crepidula fornicata Linnaeus, 1758, and Urosalpinx cinerea (Say, 1822). The former is a filter feeder that competes with oysters for both food and space whereas the latter is a predator (oyster drill) that kills large numbers of young oysters. C. fornicata arrived as early as 1870, approximately, whereas the first specimens of U. cinerea were collected in 1920. Both species of imported oyster pests spread from Southern England to other areas in Western Europe. C. fornicata has very successfully colonized the coasts of Sweden, Denmark, Germany, the Netherlands, Belgium and France where, by the mid-1970s, it had extended its southern limit South of the Arcachon area. Less successful, U. cinerea has been identified in the French oyster centres of Brest and Arcachon. The corresponding literature is abundant and frequently refers to their well confirmed accidental arrival with the American oyster. In the Mediterranean C. fornicata was repeatedly found in localities where Atlantic oysters and/or mussels were commonly imported, e.g. at La Seyne-sur-Mer (near Toulon, France) and in the lagoon of Faro near Messina (Sicily, Italy). In 1989 C. fornicata appeared in the lagoon of Salses-Leucate (France), where extensive aquaculture implies transfers of Crassostrea gigas from the Atlantic. Given this practice, C. fornicata may now have established permanently in the Languedoc lagoons and compete with the oysters.
Native of northeastern Asia, Crassostrea gigas (Thunberg, 1793) became the most widely distributed oyster, introduced world-wide in temperate and tropical seas. Introduction to

- 4 6 -
Europe started in France in the late 1960s (first to the Atlantic, then to the Mediterranean coast), on large scale from Japan and later also from the Pacific coast of North America (GRUET et al., 1976; GRIZEL & HERAL, 1991). Nonconformity to quarantine conditions leading to the joint introduction of seaweeds, various invertebrates, parasites and diseases has widely been commented. The well documented experience of previous joint introductions with the American Crassostrea virginica had not been taken into consideration. Example : in its native area, C. gigas is parasitized by the copepods Mytilicola orientalis Mori, 1935, and Myicola ostreae Hoshina & Sugiura, 1953. Both have negligently been introduced to the Atlantic coast and to Etang de Thau on the Mediterranean coast of France.
The mytilid Musculista senhousia (Benson in Cantor, 1842) native of East Asia, was part of the alien fauna imported to the Atlantic coast of France together with Crassostrea gigas (GRUET et al., 1976). It was then reported from lagoons on the Mediterranean coast of France (first record in 1982). Given the extensive shellfish culture in these lagoons and the related invasion by various other aliens, M. senhousia undoubtedly was brought in together with shipments of C. gigas. M. senhousia is highly opportunistic and adapts to both soft and hard bottoms. By establishing dense colonies (mat building), it may radically alter the biota and stabilize fine sediments; colonies may be so thick as to exclude infaunal species. Accordingly, its accidental arrival in New Zealand and Australia caused deep concern.
Accidental introductions by navigation
Serpulid tube worms are worldwide among the worst fouling nuisances on harbour structures and ship hulls, in cooling systems and oyster parks worldwide. Damage may be caused by indigenous species, but some main foulers have been spread widely by navigation.
The serpulid Ficopomatus enigmaticus (Fauvel, 1923) was probably brought to Europe among ship fouling during the first world war. Dense brackish water populations were first noticed at London harbour and in a canal of northern France. It then spread to brackish waters along the Atlantic coasts of Europe as far as Denmark, Ireland and Portugal, often causing heavy fouling, e.g. blocking sluice gates in an English site. A few years after its description it also invaded the Mediterranean where it is now widespread in coastal lagoons and estuaries, frequently building characteristic globular aggregations. Its apparition in San Francisco Bay, California, at the close of the first world war, was about contemporary with its apparition in Europe. Fauvel's hypothesis of an Indian/Indonesian origin proved to be inconsistent : he had not been able to distinguish his F. enigmaticus from congeneric tropical species. The origin from a subtropical to temperate area, possibly southern Australia, appears more likely.
Hydroides ezoensis Okuda, 1934, is well known throughout the Japanese islands. It was recognized in Europe for the first time in 1973 as part of the epifauna associated with oysters imported from Japan and relaid on the Atlantic coast of France (GRUET et al., 1976). When large encrustations of H. ezoensis were identified in 1982 from ship hulls (tugs) and man-made structures in Southampton Water (southern England), it was initially assumed that they could have originated from established populations in France. However, an extensive investigation in 1986 on the French coasts failed to reveal any specimens of H. ezoensis. Accordingly, and given an isolated earlier capture in 1976 in Southampton Water, it is more likely that the large number of Japanese ship movements into the port of Southampton comprised the effective vector of introduction of H. ezoensis, either on ship hulls or within ballast tanks.

-47 -
The earliest alien serpulid polychaetes recognized in the Mediterranean also belong to the worldwide and specious genus Hydroides : H. dianthus, H. dirampha and H. elegans. The areas of origin could be, respectively (with more or less substantial evidence), the Atlantic coast of North America, the tropical American Atlantic, and Australia (?). Collected together at Naples as early as 1888 from harbour fouling, two of them could even be traced back some years earlier in the Mediterranean. Their arrival as fouling organisms possibly antedates their first Mediterranean records considerably. All three are now widely distributed in Mediterranean harbours and coastal lagoons, but this distribution is typically disjunct, leaving out natural habitats of full marine salinity, except in areas adjacent to harbours. H. elegans is a main fouler on harbour structures and ship hulls, and has also been identified as a nuisance in industrial cooling systems.
Likewise, tiny spirorbid tubeworms may be common in fouling communities but generally have only a minor impact. Originally described from the eastern Pacific (Panama and Mexico), Spirorbis marioni and Pileolaria berkeleyana were both discovered in 1979 in a harbour at Marseille (Mediterranean coast of France), where they did not exist some years before. Investigation of harbours and adjacent areas in the following years showed a difference of distribution : P. berkeleyana was limited to the Marseille area whereas S. marioni was already widespread, from Morocco through Spain and France to Italy. S. marioni appears to have better spreading abilities and in 1987 was found in Izmir Bay and other localities on the Aegean coast of Turkey, always in harbour areas. Whereever they occur, these two alien spirorbids tend to build up dense populations. Balanomorph barnacles are usually well represented in communities that foul ship hulls and harbour structures and thereby can be a major nuisance. The corresponding literature contains many indications on wide to cosmopolitan or disjunct distribution and on the recent dissemination of various species. But the native area often remains conjectural. The case of Elminius modestus Darwin, 1854, of temperate Australian and New Zealand origin, is noteworthy because its arrival in southern England during the second world war and its subsequent spreading are well documented. This alien proved most successful along the western European coasts, colonizing the British Isles and extending as far as the Shetland Islands, Denmark and Portugal. But apparently it did not invade the Mediterranean. The exotic Balanus reticulums Utinomi, 1967, sampled in 1977 from a fouling community at Toulon harbour (Mediterranean coast of France) on a ship arriving from the Indian Ocean via the Suez Canal, also illustrates that navigation may be a factor of introduction.
Navigation has contributed to the wide distribution that now characterizes various species of ascidians known also, or mainly, from harbour and ship fouling communities. In the 1950s the far-eastern Styela clava Herdman, 1882, was introduced to southern England and was described from Plymouth as a new species, S. mammilata Carlisle, 1954. Subsequent records are from elsewhere in southern England, and also from Ireland, Denmark, the Netherlands, France (Channel and Arcachon) and northwestern Spain. In the Mediterranean the circumtropical ascidian Microcosmus exaspérants Heller, 1878, has been known since 1963 as a species related to navigation, occurring in harbours and neighbouring areas where it may contribute to the fouling communities by forming big clusters.
Typically, scleractinian corals are not part of the fouling communities that ships may contact in harbours. The first and up to now unique case of a scleractinian accidentally introduced to a distant area, is that reported from the Mediterranean as Oculina patagónica De Angelis, 1908. First discovered (in 1966 and 1978) in two localities near Savona (Liguria, Italy), the coral was subsequently found in various localities of southeastern Spain, including Alicante harbour. A colonial shallow water species possessing zooxanthellae, it thrives under a variety of conditions otherwise adverse for scleractinians : harbours and nearby polluted areas; heavy

- 4 8 -
metal pollution by mining discharge; exposure to silting and, elsewhere, to abrasion by sand. Being successful under such unusual conditions, as well as in natural algal communities, suggests a high degree of opportunism, a feature common among nonindigenous invaders. In Liguria, the coral's arrival had tentatively been explained by the release of larvae from a colony encrusting an old ship imported for the local steel industry. The colonization of diversiñed environments along southeastern Spain suggests an earlier arrival, possibly as early as the colonial period of South America. The temperate Atlantic coast of South America is suggested as the area of origin. Significant coincidence : the gastropod Crepidula caiytraeiformis Deshayes, 1830, known from Uruguay, also thrives in Alicante harbour.
Large decapod crustaceans arriving in new areas easily attract attention. Crabs as vagile organisms are quite often found clinging to ship hulls among the fouling overgrowth. The portunid crab Callinectes sapidus Rathbun, 1896, is native of the western Atlantic where it is commercially fished along the coast of North America (blue crab). In 1901 its first record from Europe was reported from Rochefort harbour, Bay of Biscay (France), but only starting with the 1950s additional records became known from the European Atlantic (Denmark, Germany, the Netherlands, France, Portugal), the Mediterranean (Italy, Malta, Greece, Turkey, Cyprus, Lebanon, Israel, Egypt), the Sea of Marmara and the Black Sea. Locally, it even acquired some commercial importance in the eastern Mediterranean. It is noteworthy that C. sapidus also settled in Japan.
The solenid bivalve Ensis directus (Conrad, 1843) is assumed to have been transported in its larval stage by a ship containing ballast water. Native of eastern North America, it was discovered in 1979 in the German Bight off the Elbe river and its arrival was extrapolated to 1978. Within a few years after its introduction populations were well established, including on intertidal flats of the Wadden Sea where it lives deeply burrowed in the sand. E. directus now occurs as far away from its first settling as northeastern Denmark and the Dunkerque area on the French coast (LUCZAK et al., 1993).
Starting with the 1970s, bivalves of the genus Scapharca, of Indo-Pacific origin, have been reported from the Mediterranean under various names of which S. inaequivalvis (Brugière, 1789) appears to be retained. First noticed near Ravenna in the northern Adriatic, where an accidental introduction by navigation is most likely, Scapharca rapidly invaded the Adriatic to the north and to the south. Other records are from Calabria, Sicily, Naples and Genova. Its capacity to survive in extreme and nearly anoxic conditions has been noted and the literature is now abundant on this successful invader. At about the same time Scapharca also settled in the harbour of Izmir (Aegean coast of Turkey), well known for its heavy pollution. In the Black Sea it was first reported from Romania in the 1980s.
The large muricid gastropod Rapano venosa (Valenciennes, 1847) is native of the northwestern Pacific and is also known from the Pacific coast of North America, where it accidentally arrived with Crassostrea gigas from Japan. In 1947 it was first noticed on the Ukrainian coast of the Black Sea, probably brought to the new area on ships hulls, although obscure oyster transfers have also been suggested. R. venosa spread throughout the Black Sea and, as a predator, became a considerable nuisance in the oyster and mussel beds. Its penetration into the Mediterranean has been predicted in 1964. Expectedly, it passed the Bosphorus and is currently fished in the Marmara Sea. In the 1970s it appeared in the northern Adriatic where it became common in the lagoon of Venice. First records from the western Mediterranean in the 1980s were from the island of Elba and from south of Livorno (Italy). Further dissemination of R. venosa can be expected, especially since it easily accomodates to brackish conditions in oyster growing areas.

-49 -
The arrival of additional alien species in the Mediterranean and western European waters is a fatality. No law can impose sterilization of vessels before entering an exclusive economic zone, and the Suez canal will not be filled in in order to stop the lessepsian migration. But a lot can be done by adequate measures in order to reduce the number of potential accidental introductions. These should comprise international conventions for, e.g., admitting exceptional deliberate introductions only after detailed impact evaluations and under absolute quarantine conditions; imposing to vessels the exchange of ballast water before entering the exclusive economic zone; and strictly controlling the commerce of exotic species for marine aquaria.

50-
Réflexions sur les problèmes d'introduction de mollusques
Henri GRIZEL
IFREMER, B.P. 133, 17390 - La Tremblade, France
Introduction
Les introductions d'espèces non indigènes dans le milieu sont des opérations courantes qui ont toujours été pratiquées par l'homme. Certaines ont eu des répercussions bénéfiques, d'autres ont entraîné des répercussions négatives à la fois pour le biotope mais aussi pour les activités humaines. Compte tenu des répercussions qu'ont eu ou que peuvent avoir ces introductions, il paraît irréaliste de vouloir s'opposer à toutes tentatives d'introduction en partant du postulat que les biotopes sont stables ou ne bougent naturellement que sur de longues périodes. En fait, lorsqu'il s'agit d'introduire à des fins économiques des espèces, il est beaucoup plus réaliste d'essayer d'encadrer ces opérations d'introductions qui comportent effectivement au moins trois types de risques (biologique, pathologique, génétique). Des groupes de travail internationaux ont déjà réfléchi sur ces sujets et ont fait des propositions constructives portant sur une procédure d'introduction et sur des recommandations scientifiques et techniques à suivre. Celles-ci doivent être préconisées, et ces informations doivent être largement diffusées auprès des décideurs et des acteurs des introductions. Enfin, sans préjuger des modifications que peuvent engendrer l'introduction de telles ou telles espèces, dans un biotope donné, il ne faut pas focaliser sur l'état de celui-ci à un moment, mais plutôt s'attacher à mieux comprendre ses fluctuations, sa dynamique dans l'espace et le temps et à conserver la variabilité des espèces même si celles-ci doivent changer.
L'exposé ci-après illustre différents types d'introductions concernant les mollusques, souligne les problèmes auxquels les activités humaines peuvent être confrontées et identifie certains risques pernicieux liés à des activités non encore prises en compte par des réglementations.
I. - Introductions accidentelles
Parmi les introductions accidentelles de mollusques, le cas récent de l'implantation de Dreissena polymorpha (Zebra mussel) est intéressant à rapporter. Identifiée pour la première fois en 1988, dans le lac Erié, elle a très rapidement colonisé les lacs Ontario et Michigan, puis s'est propagée dans la majorité des grands lacs Américano-Canadiens couplés au fleuve St-Laurent. L'explosion de cette espèce a considérablement perturbé l'écosystème en modifiant la chaîne alimentaire des larves de poissons mais a également entraîné de graves nuisances dans les économies locales qui ont dû accroître les charges d'entretien des pipe-lines obstrués par les moules. Le coût estimé pour réduire l'impact de cette espèce serait de 28 milliards de francs (S milliards de $US). Selon Carlton, cette espèce aurait été introduite via les rejets d'eaux des ballasts effectués par les navires qui remontent jusqu'aux grands lacs.

- 51 -
Les eaux rejetées, dont le volume peut atteindre jusqu'à 189.000m3, contiennent de nombreuses espèces pélagiques, zooplancton, phytoplancton et larves de plusieurs espèces. Les nombreuses études récentes démontrant le rôle des bateaux pour ces transferts accidentels devraient permettre l'adoption de réglementation. Des premières mesures ont d'ailleurs été prises par les Canadiens et les Américains. Sur un plan plus général, ces mesures sont très importantes car elles concernent des espèces généralement sans intérêt économique, donc plus difficilement contrôlables car non exploitées, mais également des espèces microscopiques dont l'impact sur le biotope et sur les économies locales sont parfois difficiles à apprécier. Pour illustrer ce point, je terminerai sur l'exemple du transport de spores de phytoplancton toxique dans les eaux de ballast, tel Gymnodinium catenation qui a été transféré dans les eaux du sud-est de l'Australie et a entraîné la fermeture de la vente des coquillages d'élevage et de pêche.
2. - Introductions intentionnelles d'une nouvelle espèce
A - Cas de Crassostrea gigas
L'huître creuse Crassostrea gigas est originaire du Japon où plusieurs populations ont été identifiées (Hokkaido, Miyagi, Hiroshima, Kumamoto) (QUAYLE, 1988). Cette espèce, très rustique, a progressivement été implantée dans plusieurs pays, en particulier au Canada (1926), aux USA (Etat de Washington), en Tasmanie, en Nouvelle-Zélande, en Europe (1970) et plus récemment en Chine (1980). Sa remarquable adaptation aux eaux tempérées a permis l'essor de son élevage et en fait la première espèce mondiale des bivalves de culture. Trois pays, la Corée, le Japon et la France mettent sur le marché 92,7% du total de la production mondiale qui avoisine 750.000 MT (Données F.A.O.).
L'introduction en France a eu lieu suite à l'installation d'une virose qui a décimé l'huître portugaise, Crassostrea ungulata. Les mortalités massives qui sont survenues entre 1970 et 1972 ont contraint les autorités à envisager le remplacement de C. ungulata par C. gigas d'autant que les premières introductions de cette huître avaient permis de constater sa résistance à la maladie et de bonnes performances de croissance (COMPS, 1972). En conséquence, au cours d'une opération "Résur" il a été décidé de recréer des gisements naturels en important des géniteurs de Colombie Britannique (Canada) et, afin d'ensemencer les parcs, d'importer du naissain de la région de Sendai (Japon). Ces opérations ont été précédées de missions de terrain, et des séries de contrôles zoosanitaires ont eu lieu chez les cheptels à importer. Les quantités d'huîtres et les lieux d'implantation sont récapitulés dans les tableaux, les totaux étant de 562 t pour les géniteurs et de 10.000 t pour les collecteurs de naissain soit, environ, plus de 5 milliards d'individus. Chaque lot de naissain importé en France a été contrôlé pour apprécier la qualité des huîtres et pour déceler la présence d'anomalies et de prédateurs. De plus, chaque lot devait être immergé en eau douce pendant une heure, afin de détruire d'éventuels prédateurs, notamment les vers plats du genre Pseudostylochus et les organismes fixés sur les coquilles et les collecteurs.
Cette opération a effectivement permis de recréer des gisements naturels et d'obtenir un auto-approvisionnement en naissain pour l'ostréiculture française, dans les régions atteintes par la virose, mais également à partir de 1975, et, à la suite de deux Protozooses de l'huître plate, Ostrea edulis, en Bretagne dans des bassins non producteurs traditionnellement d'huîtres creuses.
Malgré les précautions prises, les introductions massives ont permis l'implantation de quelques espèces originaires du Japon. De fait, très peu d'entre-elles se sont établies et

- 52-
aucune n'a, ni modifié fondamentalement les écosystèmes, ni grevé les activités humaines dans les secteurs géographiques où elles se sont développées. Les principales espèces sont résumées dans ce tableau. En parallèle, il est important de noter que l'opération Résur a permis de maintenir un tissu professionnel en place dans les régions côtières où les activités annuelles sont plutôt rares. Ainsi, malgré la perte de 60.0001 d'huîtres creuses portugaises, estimée à 556 millions de francs, plus de 5 000 ostréiculteurs concernés par cette crise ont pu continuer leur activité en pratiquant l'élevage de cette nouvelle espèce. La production actuelle d'environ 150.0001 dégage un chiffre d'affaire à l'élevage d'environ 1,35 milliards de francs (9F/kg).
Enfin, depuis l'introduction de C. gigas, originaire de la région de Sendai (Race Myagi), aussi bien pour le naissain que pour les géniteurs du Canada, les études conduites sur le polymorphisme enzymatique de plusieurs populations de cette espèce, établies dans plusieurs pays, ont montré qu'elle conservait une grande variabilité génétique. La comparaison des taux de polymorphisme, des taux d'hétérozygotie et du nombre moyen d'allèles entre les populations japonaises et les populations transférées en France ne révèle pas de différences notoires pour ces paramètres.
En conclusion, l'opération "Résur" peut être considérée comme un succès car elle a apporté une solution à une grave crise économique sans entraîner de perturbations notoires des écosystèmes conchylicoles et environnants et sans apparemment introduire de nouvelles maladies. Vingt ans après son implantation, C. gigas est devenue la première espèce élevée en Europe, et reste une des rares espèces d'élevage non sujette à de graves maladies contagieuses.
B - Cas de la palourde japonaise, Ruditapes philippinarum
Les premières introductions de la palourde en France ont été effectuées en 1973 puis 1974, en important du naissain et des adultes (tableau) originaires du Puget Sound. Quelques années plus tard (1980), un lot de 50 reproducteurs a également été importé d'Orégon vers l'Angleterre, au laboratoire de Conwy. La production issue de ces géniteurs a été expédiée à des écloseries anglaise et écossaise à partir desquelles ont eu lieu dès 1982 des envois de naissain dans plusieurs sites d'élevage en Angleterre, Ecosse mais aussi en Irlande et en Norvège.
A partir de ce stock européen de palourdes, des transferts ont également été effectués vers l'Italie (C. BREBER 1985) et l'Espagne.
Il est à noter que la production actuelle de palourde japonaise en Europe qui se situe autour de 18 0001, provient d'un stock initial évalué au maximum à 1 000 animaux. Ce dernier issu de la côte ouest du Canada et des USA, résulte lui-même de l'importation d'une faible quantité de palourdes originaires du Japon.
Toutes ces importations, dont l'objectif était l'implantation de l'espèce et le développement d'une activité d'élevage ont été réalisées en respectant les règles zoosanitaires de base (contrôles histologiques, mises en quarantaine, production d'une première génération destinée à l'élevage en milieu ouvert).
La mise au point de zootechnies précises (FLASSCH 1987-1989) et les bonnes performances de croissance et de survie obtenues avec la palourde japonaise ont permis la création d'une activité vénéricole, la production en France en 1991 étant évaluée à 600 t.

- 5 3 -
L'implantation de cette espèce dans le milieu, qui s'est faite à partir de naissain produit en écloserie, n'a pas été accompagnée de l'introduction d'autres espèces non indigènes. Par contre, les palourdes japonaises ensemencées sur parcs, ont proliféré engendrant la création de gisements naturels importants notamment en Italie, dans la lagune de Venise, et en France, dans le Morbihan.
Les captures, provenant de la pêche, évaluées à 23 000 t, ont provoqué une chute des cours de la palourde d'élevage, le prix étant passé de 50-55 F/kg à 35-45 F/kg selon la taille.
Aucune maladie nouvelle n'a été constatée lors des essais d'acclimatation de la palourde. Par contre, deux bactérioses, l'une engendrant des mortalités larvaires, l'autre étant à l'origine du syndrome de l'anneau brun ont été rapportées en 1987 et 1989. La dernière a entraîné une baisse de la production. Bien que l'origine de cette bactériose soit méconnue, plusieurs chercheurs s'accordent à penser que le nursage et l'élevage intensif pratiqués sans respecter les nonnes zootechniques établies devaient être une des causes favorisant son implantation. La régression de la maladie concommittament à l'application de mesures de prophylaxie et, à un retour aux normes »»techniques, conforte l'hypothèse précédente.
L'absence d'étude de référence sur la variabilité génétique des palourdes japonaises ne permet pas de suivre l'évolution du taux d'hétérozygoue. En 1986, MORAGA lors de recherche sur les populations cultivées de R. philippinarum soulignait cette absence et, par comparaison avec les données obtenues pour R. decussates, concluait au maintien d'une forte variabilité génétique, en dépit du fait que les populations étudiées soient issues d'écloserie. Enfin, et malgré l'absence de confirmation de terrain, il est intéressant de rappeler que ces deux espèces de palourdes peuvent s'hybrider (GERARD, 1971). Si tel est le cas dans la nature, il sera important de suivre l'évolution des nouveaux gisements.
3. - Transferts d'espèces déjà implantées
Le transfert d'espèces indigènes est devenue une pratique courante entre pays. Trop banalisés sous l'influence des sociétés d'import-export, ces transferts sont à traiter aussi sérieusement que les introductions intentionnelles. En effet, si les- risques d'ordre écologique sont minimes, les risques d'ordre pathologiques sont très élevés à cause de la spécificité de nombreuses maladies. Enfin, les risques d'ordre génétique apparaissent comme non négligeables.
En Europe, le cas de l'huître plate, Ostrea edulis, illustre bien cette situation. Cette huître indigène est présente en Scandinavie, en Angleterre, en Irlande, en Hollande, en France et dans les pays méditerranéens tels l'Espagne, l'Italie, la Grèce et le Maroc. Elle a également été introduite aux USA d'abord sur la côte est (état du Maine), puis sur la côte pacifique. Elle est présente au Canada.
En Europe, les principaux échanges pour l'élevage se faisaient entre la France, l'Espagne et la Hollande, les huîtres plates des autres pays étant importées essentiellement pour la consommation, mais souvent après un reparcage. En 1978-79, l'importation non agréée, en France, de naissain et juvéniles de la côte ouest des USA a permis l'introduction d'une maladie exotique dont l'étiologie était mal connue aux USA. Du fait des échanges précités, le parasite, Bonomia ostreae, s'est rapidement propagé dans les élevages du littoral français mais également en Hollande, Espagne, Angleterre, Irlande et Italie. L'impact de cette maladie sur l'économie ostréiocle a été très important, surtout en Hollande où l'élevage de

- 5 4 -
l'huître plate a quasiment disparu et en France (Bretagne) où la production de 2 000 t est obtenue à partir de quelques exploitations localisées dans les grandes baies. Cet exemple révèle bien l'importance des règles zoosanitaires et souligne la nécessité de s'y plier. Bien que paraissant contraignantes, elles sont indispensables à la protection du cheptel et au maintien des activités conchylicoles.
Concernant la génétique, l'étude réalisée par BLANC (1987) montre une divergence inter populationnelle entre les populations de Méditerranée provenant des étangs de St-Cyprien, Nador et Leucate, caractérisée par de faibles flux d'échanges, et des populations de la Manche et de l'Atlantique au sein desquelles se trouve la population de l'étang de Thau. Toutes ces dernières populations sont également très proches des populations irlandaises. La comparaison des indices d'identité génétique plaide en faveur d'un flux génique de fond important entre l'Atlantique, la Manche et la Méditerranée. Ces observations sont largement corroborées par l'histoire et les pratiques ostréicoles. En effet, au cours de la période 1920-1929 de nombreux gisements décimés par une maladie ont été recréés grâce à l'importation massive d'huîtres plates. Tel est le cas des gisements de la rivière d'Auray et de l'étang de Thau reconstitués en immergeant des huîtres originaires de gisements anglais et irlandais.
Conclusions
Ces quelques exemples illustrent bien les problèmes liés aux transferts et introductions. Dans le cas des mollusques, comme pour beaucoup d'autres espèces, les introductions accidentelles peuvent engendrer d'importantes répercussions pour les biotopes et pour les activités littorales. Les espèces introduites suivent des lois générales d'évolution au sein d'un écosystème. Lorsque l'une d'entre-elles trouve un biotope favorable elle passe par une phase d'installation, au cours de laquelle elle échappe souvent à l'observateur, suivie d'une phase explosive et enfin se stabilise dans le nouveau biotope en engendrant un nouvel équilibre. Elle sera alors, soit dominante, soit une espèce parmi d'autres car elle aura subi des régulations. Les répercussions sur le biotope et sur les activités pourront être importantes pendant la phase explosive et pourront pérenniser selon l'équilibre auquel aboutit cette espèce. La gestion des animaux ou végétaux transférés est, en général, difficile à maîtriser. En effet lorsque l'espèce ne présente aucun intérêt pour l'économie, elle n'est pas régulée par une exploitation, ou lorsqu'elle n'entrave pas le bon fonctionnement des activités littorales aucune mesure n'est prise pour tenter de la réguler. Pour les eaux de ballast, mises en cause dans plusieurs cas, les premières mesures prises par les Américains et les Canadiens devraient être l'objet de discussion au plan mondial afin d'aboutir à une réglementation internationale.
Dans les cas des mollusques, les introductions intentionnelles permettent d'établir un bilan global généralement positif. Quels que soient les exemples, les répercussions sur les économies conchylicoles ont toujours été positivement importantes sans qu'il y ait pour autant création de graves désordres écologiques ou encore création d'importantes perturbations sur la génétique des espèces élevées et environnantes. Ces faits tiennent probablement, pour partie, aux possibilités de contrôle des stocks de mollusques qui sont sessiles.
Toutefois, et en particulier pour les risques d'ordre pathologique, il est important de suivre les recommandations formulées par le Conseil International pour l'Exploration de la Mer (In Cooperative Research Report n°130).
Enfin, les transferts qui sont malheureusement trop banalisés présentent des risques élevés pour l'introduction de maladies. En Europe, l'ouverture des frontières avec le marché
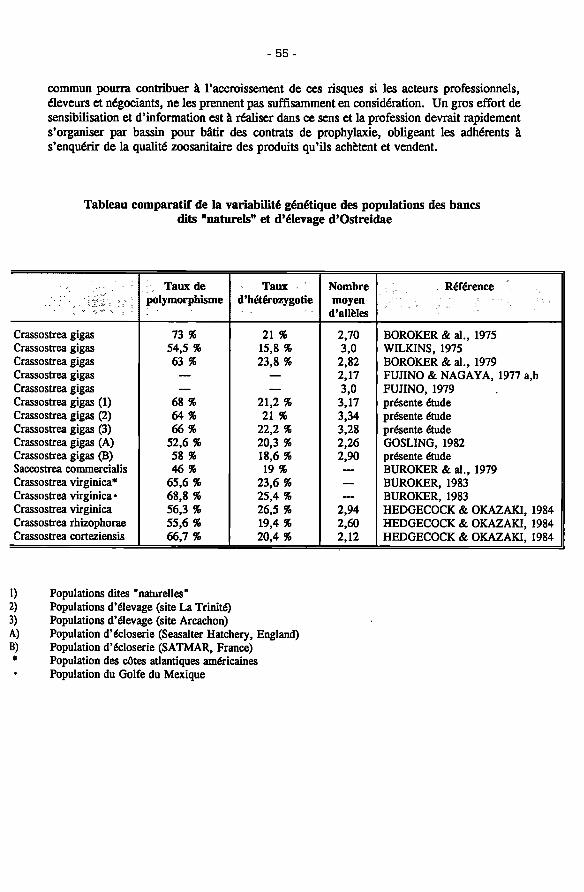
55
commun pourra contribuer à l'accroissement de ces risques si les acteurs professionnels, éleveurs et négociants, ne les prennent pas suffisamment en considération. Un gros effort de sensibilisation et d'information est à réaliser dans ce sens et la profession devrait rapidement s'organiser par bassin pour bâtir des contrats de prophylaxie, obligeant les adhérents à s'enquérir de la qualité zoosanitaire des produits qu'ils achètent et vendent.
Tableau comparatif de la variabilité génétique des populations des bancs dits "naturels" et d'élevage d'Ostreidae
[.':' V . ■ : ' · *
^ *■ - · , ··'■ \ . -
Crassostrea gigas Crassostrea gigas Crassostrea gigas Crassostrea gigas Crassostrea gigas Crassostrea gigas (1) Crassostrea gigas (2) Crassostrea gigas (3) Crassostrea gigas (A) Crassostrea gigas (Β) Saccostrea commercialis Crassostrea virginica* Crassostrea virginica · Crassostrea virginica Crassostrea rhizophorae Crassostrea corteziensis
Taux de polymorphisme
73 % 54,5 % 63 %
— —
68% 64% 66%
52,6% 58 % 46 %
65,6% 68,8% 56,3 % 55,6 % 66,7%
Taux d'hétérozygotie
21 % 15,8 % 23,8 %
— —
21,2% 21 %
22,2% 20,3 % 18,6% 19 %
23,6% 25,4% 26,5 % 19,4% 20,4%
Nombre moyen d'allèles
2,70 3,0
2,82 2,17 3,0
3,17 3,34 3,28 2,26 2,90 — — —
2,94 2,60 2,12
. Référence
BOROKER & al., 1975 WILKINS, 1975 BOROKER & al., 1979 FUJINO & NAGAYA, 1977 a FUJINO, 1979 présente étude présente étude présente étude GOSLING, 1982 présente étude BUROKER & al., 1979 BUROKER, 1983 BUROKER, 1983 HEDGECOCK & OKAZAKI, HEDGECOCK & OKAZAKI, HEDGECOCK & OKAZAKI,
,b
1984 1984 1984
1) 2) 3) A) B) *
Populations dites "naturelles" Populations d'élevage (site La Trinité) Populations d'élevage (site Arcachon) Population d'écloserie (Seasalter Hatchery, England) Population d'écloserie (SATMAR, France) Population des côtes atlantiques américaines Population du Golfe du Mexique

56
Caractéristiques génétiques et capacité d'invasion chez les invertébrés dans les eaux littorales et les lagunes méditerranéennes
Anna OCCfflPÜSm AMBROGI
Università degli Studi di Venezia, Dip. di Scienze Ambientali, Calle Larga Santa Marta, Dorsoduro 2137 30123 Venezia, Italia.
Résumé
On ne dispose pas à l'heure actuelle d'un cadre d'ensemble permettant d'interpréter synthétiquement les caractéristiques génétiques des espèces invasives. Celles-ci constituent en effet autant de problèmes quant à leur origine, à leur moyen de transport et à leur capacité d'adaptation. On envisage cependant, en prenant comme élément de comparaison des espèces volontairement introduites par l'homme, quelques traits qui expliquent le succès des envahisseurs, en soulignant en même temps le danger que représente pour la faune autochtone l'introduction d'espèces étrangères dans des écosystèmes sélectifs et à dynamique peu prévisible. La recherche génétique, doublée d'une analyse écologique précise sur la dynamique des peuplements, peut contribuer à des solutions efficaces de ces problèmes. On souligne à ce propos l'intérêt que revêtent les écosystèmes estuariens et lagunaires, théâtre depuis longtemps d'invasions suivies d'un succès durable.
Introduction
La génétique des invertébrés marins connaît depuis une dizaine d'années un regain d'actualité par l'application de méthodes d'analyse électrophorétique.Ainsi BATTAGLIA & BEARDMORE (1979) ont organisé un symposium à Padoue ayant pour thème les problèmes de génétique, d'écologie et d'évolution des organismes marins. Plusieurs contributions concernant les Mollusques ont été présentées lors d'une rencontre internationale à la Nouvelle-Orléans en 1982 (SINGH & GREEN, 1984; ZOUROS & FOLTZ, 1984); ainsi que dans le cadre de Congrès malacologiques successifs. Toutefois ces travaux ont surtout porté sur la génétique d'espèces cultivées en raison de leur importance alimentaire et économique. L'intérêt de ces recherches pour des espèces involontairement importées ne peut donc être que largement comparatif.
Cependant les considérations de FAVA & FONSATTI (1992) peuvent s'appliquer, dans une certaine mesure, aux immigrants occasionnels. Ces auteurs proposent, une fois connue la structure génétique de l'espèce étudiée dans son territoire d'origine, d'en évaluer la capacité d'adaptation à de nouveaux biotopes : en fait, pour des introductions involontaires, d'analyser quelle a été la base génétique de leur succès en des biocénoses étrangères. On comparera ensuite les populations d'origine à celles émigrées pour évaluer la signification des éventuelles différences génétiques rencontrées. On constate que les considérations de FAVA & FONSATTI, tout-à-fait valables pour des introductions voulues et entretenues par l'homme, ne peuvent s'appliquer aux immigrations "spontanées" que dans une certaine mesure. Il y

- 57
a, en effet, une différence notable entre les deux cas. Pour des espèces utiles à l'homme, il s'agit de trouver les structures génétiques les plus résistantes et productives, les plus faciles à défendre et à développer; pour des espèces que l'homme n'a pas voulues, et qui ne l'intéressent pratiquement que comme éventuelles compétitrices ou interferentes des peuplements autochtones, il s'agit plutôt de les décourager, de les limiter, sinon de les éliminer.
Pour l'écologiste, autant de solutions opposées; pour le généticien un problème substantiellement unitaire : l'identification de la base héréditaire régissant la capacité de sortir d'une niche limitée pour atteindre une valence écologique plus étendue. Ecologiste, jusqu'en 1992, au Département de Génétique de l'Université de Pavie, je me pencherai donc avec plus d'intérêt sur ces problématiques. Mon exposé se limitera à quelques points qui intéressent surtout l'écologie des populations et l'évolution des espèces accidentellement introduites.
Fig. 1 : Distribution de Tricellaria inopinata dans la lagune de Venise (situation 1992). (d'après SACCHI & OCCfflPIOTl AMBROGI, 1992).
Exemples et hypothèses
Les caractéristiques génétiques des espèces envahissantes font l'objet d'analyses plus ou moins détaillées et d'importants essais de synthèse. Ces derniers, malheureusement, se basent souvent sur des connaissances encore très incomplètes, concernant non seulement la génétique, mais également la répartition géographique et la valence écologique des envahisseurs.

- 5 8 -
Parmi les essais, qui, bien que traitant de végétaux, sont susceptibles par l'ampleur des perspectives, d'éclairer nos problèmes, celui que GRAY (1986) propose semble particulièrement intéressant, puisqu'il souligne le concept de "renouvellement génétique" chez des pionniers parvenant, par les possibilités offertes par la structure même des pools géniques d'origine, à s'échapper des constrictions subies dans leur patrie. GRAY rejoint pourtant d'autres auteurs (MOLLISON, 1986; CRAWLEY, 1986) pour recommander la prudence dans un domaine où des aspects spéculatifs priment parfois sur les données concrètes disponibles.
Nous ne pouvons que partager entièrement ce point de vue quant aux animaux invertébrés marins, si peu connus tant dans leurs aspects génétiques, que dans leur biologie générale.
On ne saurait trop insister sur les lacunes que présentent les données même de terrain, car l'on sait que, en dehors de quelques districts où se trouvent d'anciennes et glorieuses stations de biologie marine, telles Naples ou Marseille, la faune invertébrée est encore, somme toute, en grande partie à découvrir.
Une autre impasse à éviter est l'équivalence : espèce à répartition très limitée, ponctiforme, égale espèce récemment parvenue sur place. Il pourrait, au contraire, s'agir de reliques présentes dans un territoire restreint à cause des prémices conservatrices de l'environnement. Ces reliques peuvent rencontrer de nouveaux arrivants qui en partagent momentanément les limitations topographiques et écologiques. Les uns et les autres auront un stock génique limité, mais cette limitation peut avoir une signification opposée. Elle sera le résultat parallèle, ici d'une réduction, d'une spécialisation imposée par la sélection progressivement exercée au cours des temps, là d'un bottle-neck réalisé au départ, ou resserré lors du voyage ou au moment de l'arrivée. Cette limitation serait donc le symbole, soit d'une résistance dans une place assiégée, soit d'un début de conquête d'espace vital.
Un exemple tiré de la lagune de Venise, environnement imprévisible comme le sont les milieux lagunaires et estuartene (COGNETTI, 1992 ) illustre le résultat analogue de ces deux processus, si différents par leur signification historique et évolutive. L'Adriatique du Nord est une mer dont les caractéristiques "sub-atlantiques" (SACCHI et al. 1989), encore plus marquées dans les lagunes, sont l'issue d'un ensemble de facteurs inconnus ailleurs dans le domaine méditerranéen : températures modérées, marées non indifférentes, salinité assez faible, remarquable richesse trophique. Ce "sub-atlantisme" se traduit par la présence localisée d'espèces et de formes inconnues en d'autres zones de Méditerranée, mais reliées aux peuplements de l'Atlantique septentrional.
Dans l'horizon intertidal moyen-supérieur de la Lagune de Venise nous avons rencontré un Bryozoaire Chéilostome, espèce apparemment nouvelle pour la science, que nous avions baptisé Tricellaria inopinata (D'HONDT & OCCHTPINTIAMBROGI, 1985), voisine d'une espèce à distribution surtout indo-pacifique (Fig.l). Nous sommes actuellement en train d'analyser, en collaboration avec M. D'HONDT du Muséum de Paris, le polymorphisme enzymatique de cette "nouveauté" en établissant en même temps une série de comparaisons avec d'autres Tricellaires, car il est nécessaire d'en contrôler le statut taxonomique et les variations micro-systématiques, avant de tirer des conclusions d'ordre génétique. En tout cas, il s'agit sûrement d'une apparition récente, suivie d'une rapide diffusion, car dans les stations que notre Tricellaire peuple actuellement nous ne l'avions pas trouvée il y a quelques années seulement (OCCHIPINTI AMBROGI, 1991).
Depuis 1982, année de sa découverte dans le canal de la Giudecca, elle a colonisé tous les substrats durs lagunaires, surtout dans les grappes de moules des balises en bois le long des canaux navigables. En occupant les interstices entre les coquilles, ce Bryozoaire se fixe

59-
même sur les filaments de byssus. Tricellaria inopinata se constitue ainsi un abri contre un environnement hostile (par exemple lors de l'émersion des moules par basse-mer) et dispose d'un espace libre de compétiteurs, où l'on ne rencontre aucun autre Bryozoaire. Pour le moment, elle n'est absente que des stations très dessalées, ou soustraites, en pleine ville, au jeu de la marée (Fig. 2).
Cependant, dans les mêmes biotopes, se trouve une autre espèce qui n'est que lagunaire dans l'Adriatique, tout en étant largement répandue (répartition amphi-atlantique) sur les grèves de l'océan : le Gasteropode Prosobranche ovovivipare Littorina saxatilis . OLIVI (1792) l'a décrite pour les environs de Chioggia il y a deux siècles, mais récemment JANSON (1985) a formulé l'hypothèse d'une introduction, assez ancienne évidemment, mais due à l'homme. Elle base cette affirmation sur un polymorphisme enzymaüque réduit, d'après un faible lot en provenance d'une station périphérique de la lagune, monomorphe même en ce qui concerne les variations polychromatiques de la coquille. En réalité, dans la partie centrale et septentrionale de la lagune de Venise, L. saxatilis montre un large spectre de variations quant aux couleurs de la coquille, la taille, l'habitat (SACCHI & TORELLI, 1979; SMITH, 1981), peuplant non seulement des substrats cohérents artificiels, comme dans la station de JANSON, mais toute une série de substrats naturels variés, même sur fond meuble (barene) correspondant au schorre de la Manche et de la mer du Nord.
On ne saurait y voir un phénomène d'introduction récente, d'autant que des carottages pleistocenes dans l'autre grande lagune nord-adriatique (Grado-Marano) ont mis au jour des coquilles fossiles aux caractères de L. saxatilis (BRAMBATI, comm. pers.). Si les L. saxatilis des lagunes nord-adriatiques sont moins polymorphes que celles des grèves atlantiques, cela est vraisemblablement dû au fait que ces dernières sont plus riches en possibilités écologiques et en diversité biotique.
^~~A
Fig. 2 : Distribution de Tricellaria inopinata (Venise) et du groupe d'espèces les plus pioches (d'après OCCHIPINTI AMBROGI & d'HONDT, sous presse).

- 6 0 -
Discussion
Essayons de résumer quelques points de vue d'auteurs qui ont étudié les caractéristiques génétiques des espèces dont l'invasion a été couronnée de succès :
a. de telles espèces devraient appartenir à des stocks dotés d'une variabilité génétique assez riche, constituant les prémices d'une adaptation à des conditions variées, et même potentiellement différentes des conditions d'origine. Il doit s'agir, en somme, d'espèces "banales" dans leurs perspectives écologiques.
b. puisque, en tout cas, les envahisseurs emportent une portion plus ou moins limitée de cette puissance polymorphe, celle-ci devra être doublée d'une plasticité phénotypique élevée, tant individuelle qu'au niveau des populations, us doivent tolérer un environnement variable, telles les eaux littorales et surtout les lagunes, en survivant à des stress d'une certaine gravité; leur dynamique démographique devra être articulée sur des différences locales en taux de croissance, maturité sexuelle, durée des périodes de vie ralentie, production de disseminules, densité de population.
c. il est cependant possible que les envahisseurs possèdent un patrimoine génétique marginal dans leur patrie, où il seraient probablement destinés à une position évolutive de secondaire importance, et que leur succès dans les écosystèmes nouvellement colonisés se doive essentiellement à l'absence des compétiteurs et des prédateurs qui, à l'origine, contrôlent et limitent la niche écologique de l'espèce (FOX & FOX, 1986).
On pourrait ainsi y reconnaître le jeu de la "révolution génétique" de GRAY (1986). Moins probablement, au sens statistique, il se pourrait enfin que la portion restreinte de variabilité que l'émigrant porte en lui, soit la "bonne" dans le nouvel environnement, pour lequel elle serait en un certain sens "présélectionnée" (SACCHI & OCCHIPINTI AMBROGI, 1992).
L'un et l'autre modèle pourraient convenir pour expliquer l'affirmation en milieu lagunaire de certaines morphes de Crustacés Péracarides, présentes en eau très dessalée et "continentale" avec des morphes absentes ou rarissimes sur la côte de la mer ouverte : on connaît quelques cas chez Sphaeroma (LEJUEZ, 1974; KERAMBRUN, 1965, 1966; KERAMBRUN et al., 1964) et chez d'autres invertébrés marins (BOCQUET, 1974). On y retrouvera des analogies avec les Copépodes Harpacticoïdes, dont l'école de Padoue a étudié la génétique formelle (BATTAGLIA, 1967). Ces recherches intéressent, il est vrai, des espèces autochtones en Méditerranée, mais peuvent constituer un modèle important pour une recherche future concernant les immigrants.
Pour le moment nous ne pouvons que reconnaître qu'il n'y a ni hypothèse interprétative, ni solution synthétique "bonne à tout faire" : c'est l'idée des auteurs australiens vivant et travaillant dans un pays qui est un grand laboratoire d'introductions volontaires et involontaires (NEWSOME& NOBLE, 1986; BARRETT & RICHARDSON, 1986). D'autre part, comme SAFRIEL & RITTE (1983) l'affirment dans leur essai général sur les caractéristiques propres aux espèces colonisatrices, l'apport des connaissances génétiques ne peut que constituer l'un des nombreux outils que l'écologiste emploie lorsqu'il analyse des phénomènes de colonisation, surtout s'il veut faire des prévisions sur le succès d'une invasion.
On reconnaîtra d'ailleurs qu'il n'est même pas facile d'établir un éventuel avantage de l'hermaphroditisme ou de la parthénogenèse sur la reproduction amphigonique, car la

61
possibilité que chaque individu devienne un fondateur de colonies aurait une lourde contrepartie dans la réduction du patrimoine génétique lors des "effets de fondation" (GRAY, 1986; BARRETT & RICHARDSON, 1986).
De plus, les introductions en milieu marin peuvent être ultérieurement compliquées par des conditions particulières. Celles-ci comprennent les possibilités d'immigrations répétées, par exemple avec des échanges commerciaux réguliers, d'espèces destinées à l'alimentation ou à l'élevage, η est ainsi possible que des échecs momentanés finissent par se solder par un succès, dès que le rapport environnement/structure génétique atteint un niveau favorable; il est aussi probable que les arrivées successives élargissent le patrimoine génétique des immigrants par des contributions géniques variées, offrant des solutions positives, qu'une seule arrivée ne suffirait pas à proposer.
Le problème se pose enfin du contrôle par l'homme de ces invasions, qui intéressent l'écologiste, comme le systématicien, le spécialiste de l'environnement, comme le biologiste des pêcheries et des élevages.
FAVA & FONSAlll (1992) trouvent chez Tapes philippinarum, palourde d'origine indo-pacifique introduite en Europe à cause de sa productivité particulièrement élevée, un niveau d'hétérozygose remarquable pour des bivalves marins, où l'hétérozygose est plutôt rare (SINGH & GREEN, 1984; ZOUROS & FOLTZ, 1984). Les stocks volontairement emportés possédaient donc un patrimoine génétique assez étendu; il en est probablement de même pour les espèces qui en constituent un cortège difficile à éliminer par des moyens, tant mécaniques que chimiques, dans les parcs et les élevages : épibiontes, phorétiques, commensales.
Sous l'aspect synécologique, l'arrivée d'immigrants non désirés pose aujourd'hui à l'aquaculture des problèmes analogues à ceux que l'agriculture affronte depuis des siècles. Les intrus peuvent souvent être considérés comme des "maladies" des équilibres biocénotiques locaux, bien que parfois ils constituent des facteurs d'enrichissement de la diversité biotique (BERMAN & CARLTON, 1991) surtout si celle-ci est relativement réduite en espèces, comme dans les milieux "difficiles" que nous avons cités.
Comme exemple d'invertébrés récemment introduits dans deux environnements très "perturbés", tels la mer Noire et la Haute Adriatique, il faut citer les cas de Scapharca inaequivalvis, un bivalve à faible valeur commerciale, mais doué d'une extraordinaire résistance à l'anoxie, et de Rapana venosa, redoutable prédateur de bivalves, sur lequel on ne possède malheureusement aucune information génétique. Π est donc important de contrôler ces intrus par exemple par une lutte biologique intégrée, en favorisant, si possible, des ennemis et des compétiteurs naturels présents dans leur patrie d'origine (CRAWLEY, 1987) non nuisibles pour nous. Dans ce domaine la génétique aura son mot essentiel à dire, non moins que dans l'étude d'une résistance différentielle des espèces nuisibles à l'agression chimique et micro-biologique, suggérée par les considérations exposées au point (c).
La vulnérabilité et l'imprévisibilité des équilibres biotiques des littoraux et surtout de leurs secteurs les plus sensibles, tels encore une fois les estuaires, les lagunes, les ports, les zones péri-insulaires, conseillent précisément de pousser l'analyse au niveau des structures héréditaires plutôt que d'insister uniquement sur des remèdes mécaniques ou chimiques. Mais toute lutte biologique intégrée, synthèse appliquée des connaissances écologiques et génétiques, serait vouée à l'échec sans l'action prudente (car elle empiète forcément sur des intérêts locaux tout à fait respectables) mais bien définie et décidée, d'une collaboration législative et opérationnelle dans la sphère internationale.

- 6 2 -
Conclusion
En conclusion, les principaux besoins de la recherche dans le domaine des invasions nous semblent liés d'une part à la possibilité de prévoir la susceptibilité d'environnements et de communautés déjà reconnus comme instables à l'invasion d'espèces étrangères, et d'autre part à la capacité d'envisager le succès (en termes de vitesse, étendue et durée) des envahisseurs.
Dans ce processus d'accumulation de connaissances, la génétique peut apporter une contribution substantielle, surtout en s'appuyant sur les techniques de mesure de la variabilité génique, comme le taux d'hétérozygose, mais elle doit être étroitement liée à l'interprétation écologique, par l'analyse de la dynamique de population, des niches écologiques et des mécanismes de compétition avec les espèces indigènes.
Il faudra donc diriger les efforts sur des environnements bien connus dans leurs caractéristiques abiotiques et biotiques et riches de cas récents d'invasions quantitativement remarquables, comme la mer Noire et la Haute Adriatique (et en particulier ses lagunes). Il ne faudra cependant pas se limiter aux cas les plus frappants par leur intérêt économique, positif ou négatif, mais l'on devra également s'occuper de cas apparemment secondaires, qui, faute d'interventions humaines directes, peuvent de ce même fait mieux éclaircir le rôle des mécanismes écologiques et évolutifs.

- 6 3 -
Lessepsian migration - Biological invasion of the Mediterranean
Bella S. GALIL
Israel Océanographie and Limnologica! Research P.O.B. 8030 Haifa 31080, Israel
The lessepsian invasion is in effect an environmental roulette where the stakes are high and the chances of winning exceedingly low. The opening of the Suez Canal did, undeliberately, initiate an important event : the adjoining of two biogeographical provinces. Over 120 years Red Sea species migrating through the canal have been colonizing the Mediterranean. The recent proliferation of erythrean jellyfish along the Levant coast illustrates the profound changes in the indigenous biota caused by lessepsian migration.
Human intervention increasingly erodes the effectiveness of biogeographical boundaries. Here we examine the ecological consequences of the greatest marine engineering enterprise - the Suez Canal. The construction of a sea-level waterway between the Eastern Mediterranean and the northernmost tip of the Gulf of Suez linked the Atlantic-Mediterranean province with the Indo-Pacific. The deep-water communication between the Mediterranean and the Indian Ocean was breached during the early Miocene (ROBBA, 1987), though intermittent marine contacts between the respective basins continued well into the Messinian (SONNENFELD, 1985) and species of Indo-Pacific origin still inhabited the Pliocene Mediterranean (SORBINI, 1988). Early in the Pliocene the isthmus of Suez was elevated and the two seas separated completely. Thus, the present day Suez Canal is at once an iteration of an earlier connection and a repeat of a palaeobiogeographic process.
In 1869 the Suez Canal was inaugurated, inadvertently launching a remarkable faunal movement. Despite physical and hydrologicai impediments, hundreds of erythrean species traversed the canal and settled in the Mediterranean in a process termed "Lessepsian migration" - after Ferdinand Marie de LESSEPS, the frenen diplomat and engineer who built the canal (POR, 1989). It is, in the opinion of F.D. POR, its foremost scholar, "the most important biogeographic phenomenon witnessed in the contemporary oceans" (POR, 1989).
In 1977 a specimen of an unknown species of jellyfish was collected off the coast of Israel (LEWINSOHN, 1977). Preserved but unidentified, it remained unnoticed until the mid-1980s, when large aggregations of jellyfish along the Israeli coast prompted efforts to study the phenomenon. The jellyfish was identified as a new species - Rhopilema nomadica - and a lessepsian migrant (GALIL et ál., 1990). The largest specimen of this nomadic jellyfish measured more than a meter across its umbrella, though most specimens surveyed measured between 20 and 60 cm. The fresh weight of a specimen with bell diameter of 50 cm was over 9 kg. Mass swarming of R. nomadica occurs in the summer, but a sizable presence has been witnessed during the winter too. Marine and coastal surveys point to a "jellyfish" belt at a distance of 1 and 5 km offshore, where the maximal density was estimated at 25 specimens per m3. Similar phenomena were reported from Egypt and Lebanon. The mass aggregations of the jellyfish exert a significant impact on fisheries, coastal installations and tourism.

- 6 4 -
The presence of this jellyfish forced the reappraisal of some long established views concerning lessepsian migration, and led to réévaluation of its precepts as regards population dynamics patterns, competition between migrant and indigenous species and its course.
The population dynamics pattern of the lessepsian migrants was considered to be one of unremitting expansion. GHAT (1969) believed that "There is evidence that there is no regress in the building up of populations of the new immigrants" and POR (1978) reiterated "There are no known examples of reversals - i.e. of a disappearance or radical depletion of the lessepsian population". Certainly, hundreds of lessepsian migrants became established and founded thriving populations along the Levantine coasts. Some species are even of economic importance, being exploited commercially.
Examination of fishery statistics underscores the migrants' prominence in the local communities, migrant fish constituting a third of the total trawl catches off the Israeli coast (BEN TUVIA, 1985). Benthic lessepsian fish contribute nearly half of the total fish abundance and biomass on the hard bottom habitats of the southeastern Mediterranean shelf, though they constitute only 11 per cent of the species (SPANIER et ál., 1989). The rabbit fish, Siganus lundus and S. rivulatus, and the red soldier fish, Adioryx ruber, are the most common erythrean fish on hard substrata in the Levant Basin.
The immigrant penaeid prawns Penaeus japonicus and P. monoceros, make up most of the shrimp catches along both the Egyptian and Israeli coasts (GALIL, 1986). A massive "build up" of populations is found also in the swimming crab, Charybdis longicollis. First recorded from the Mediterranean in the mid-1950s, it has since dominated the macrobenthic fauna on silty sand bottoms, forming up to 70 per cent of the biomass at places (GALIL, 1986). A more recent case in point is that of Strombus decorus persicus, first discovered in Mersin Bay, Turkey, in 1978 (NICOLAY and ROMAGNA MANOJA, 1983) and then in quick succession off southern Turkey (RAYBAUDI-MASSILIA, 1983), Rhodes (VERHECKEN, 1984; BARASH and DANIN, 1989), Cyprus (BAZZOCCHI, 1985; ROBINSON, 1989) and Israel. The mass appearance of the lessepsian jellyfish, R. nomadica, unlike the seasonal proliferation of cyclic fluctuation of the indigenous Mediterranean jellyfish (VUCETIC, 1983), is again the exponential phase of an intruder.
However, among the lessepsian colonists there are several well documented cases of "explosive growth" followed by recession. In the late 1940s the migrant goldband goatfish, Upeneus moluccensis, made up 10-15 per cent of the total mullid catches. Following the exceptionally warm winter of 1954-1955, its percentage in the catch increased to 83 per cent (OREN, 1957). Its share has since been reduced to 30 per cent of the catch (BEN TUVIA, 1973). Following that same winter the brushtooth lizardfish, Saurida undosquamis, became a commercially important fish and its proportion in trawl fisheries catches rose to 20 per cent of the volume in the late 1950s. The population then diminished and catches stabilized at about 5 per cent of the total trawl catch (BEN-YAMI and GLASER, 1974). Leiognathus klunzingen was "extremely common on trawling grounds in depths of 20 and 100 m in the early fifties", but its populations were decimated during the sixties (BEN TUVIA, 1985). A more dramatic case is that of the gastropod Rhinoclavis kochi. It was first reported in Haifa Bay in the mid-1960s and has since spread rapidly and became one of the dominant species between 20 and 60 m, peaking in the late 1970s (GALIL and LEWINSOHN, 1981; TOM and GALIL, 1990). Collections made a decade later brought up mostly empty shells.
The precipitate merger of disjunct faunas may ensue in competition for different resources or direct interference between the newcomers and the autochthonous species, the latter sometime outcompeted in a part of their habitat space.Lessepsian migration is in effect a test

- 6 5 -
of the checks and balances within the host ecosystem. POR (1978) maintained that "other than the case of Asterina gibbosa there is no known case in which a lessepsian migrant species has completely replaced a local one". Indeed, the disappearance of the indigenous sea star, A. gibbosa, from the Israeli coast paralleled the rapid advent of its lessepsian congener A. wega (ACHlTUV, 1973). Though only rarely are sufficient data available about the life histories and the ecological relationships of both indigenous and lessepsian species to determine which are the competing species, there are documented instances providing evidence of a drastic change of abundance that can be attributed to the new competition. A native penaeid prawn, Penaeus kerathurus, was "very commonly caught by trawlers on Israel coastal shelf especially on a sandy or sandy mud bottom" (HOLTHUIS and GOTTLIEB, 1958) and supported a commercial fishery throughout the 1950s. It has since nearly disappeared and its habitat overrun by the lessepsian penaeid prawns. GELDIAY and KOCATAS (1972) reported similarly that off the southern coast of Turkey, the erythrean P. japonicus is replacing P. kerathurus in fishery catches. And as the migration proceeds we might perceive replacement of one immigrant by another - Trachypenaeus curvirostris is found now in smaller quantities where it is replaced by the similarly sized Metapenaeopsis aegyptia.
A less drastic competition might take the form of bathymétrie adjustment. Such competitive displacement probably occurs between the local red snapping shrimp, Alpheus glaber, and the intruding congener A. rapacida; the former appears mainly between 35 and 145 m and the latter between 15 and 50 m (GALIL, 1989). The local red mullet, Mullus barbatus, and the native hake, Merluccius merluccius, were both displaced into deeper, cooler waters by their respective erythrean competitors : the goldband goatfish and the toothbrush lizardfish (POR, 1978). But depth-shifting among lessepsian migrants could be ascribed also to the environment physical characteristics. Erythrean species that are rock and rubble dwellers are confined in the Mediterranean to infralittoral regions for want of hard substrate in deeper waters. Among erythrean species inhabiting soft bottoms, some occupy a depth range similar to that in the Red Sea, while others seem to shift their range of water 20-50 m deep. As temperature is considered the most important single factor for lessepsian migrants, it is assumed that the winter cooling of the shallow waters is the cause for the bathymétrie shifting (GALIL, 1989).
Most lessepsian migrants are concentrated off the southern Levant coast. A considerable number reaches the southern Turkish coast and Cyprus, whereas only a few species have been recorded as westward as Malta, Tunisia and Sicily (CHAKROUN, 1966; ARIANI and SARRA, 1969; KTARI-CHAKROUN and BOUHLAL, 1971; STEVCIC, 1979; BEN TUVIA, 1985). Some, evidently, reach further : CAMPOS and TURKAY (1989) report the occurrence of a lessepsian swimming crab, Charybdis hellen, off the Carribean coast of Colombia, surmising it was introduced by Israeli ships.
That the Levantine fauna evolves under Lessepsian influence is evident; the number of erythrean migrants grows continuously and modification of the composition and structure of the Levantine biota is already underway. The presence of lessepsian migrants augments the tropical affinities which quite noticeably distinguish the Levantine fauna. The infusion of Indo-Pacific elements and the concomitant change in the indigenous fauna enhance the tropical character of the Levant compared with the more temperate character of the sea as a whole, causing disruption of the Zoogeographie unity of the Mediterranean fauna. But, as with most cases of introduced species, we have little unequivocal evidence of the effects of the lessepsian migration on other species, primarily because we have no satisfactory baselines of pre-lessepsian conditions for evaluating the impact of the migration. The marine ecosystem is not well enough understood to determine the long-term effects of biological invasions,

. - 6 6 -
which potential may not be realized for decades following introduction. Especially vulnerable to alteration or domination by migrants are depauperate faunas.
Understanding the complexities of the interactions between the lessepsian and the indigenous components calls for a concentrated international effort to study the new additions to the marine biota, the ecological dynamics of the Levant Basin ecosystem, and the habitat, trophic and spatial alterations caused by lessepsian migration.

67-
Non-indigenous species in the Baltic Sea
Erkki Leppãkoski
Department of Biology and Husõ Biological Station Åbo Akademi University, SF-20500 Turku/Åbo, Finland
This report is based on papers presented at the 1st Conference on the Environmental Management of Enclosed Coastal Seas, Kobe, Japan,1990 (Leppãkoski 1991) and at the NOAA Workshop on Nonidigenous Estuarine Marine Organisms (NEMO), Seattle, Washington, 1993 (Leppãkoski 1993).
In the ecosystem of the Baltic Sea (5-8 %o salinity) alien species are numerous especially in habitats strongly modified by man's activities (harbours, underwater constructions, receiving waters of pollutants and cooling water discharges). Eutrophicated inlets and coastal lagoons and fouling communities on secondary hard bottoms are examples of sub-systems that have been invaded by nonindigenous species, whereas the open sea systems, both pelagial and benthal, are relatively free from them. The Baltic Sea is, in the majority of cases, a receiver area of introduced species; the donor areas are to be found in the adjacent coastal oceanic waters as well as in remote seas. More than 35 species of anihropochorous immigrants have been reported from the central and northern Baltic Sea, incl. its terrestrial coastal systems. Several introductions into the Baltic Sea have resulted in both structural and functional changes at ecosystem level. There are only a few harmful nonindigenous species in the Baltic Sea.
Species introductions into marine and brackish-water systems have not been studied systematically prior to the 1950's. Some comprehensive reviews on a regional or sea-wide basis appeared in the mid-80's (Carlton 1985; see Leppãkoski 1984 for the Baltic Sea).
Enclosed seas, such as the Baltic Sea and the Black Sea, can be regarded as disjunct "brackish-water islands", with an impoverished fauna and flora, locked in by an "ocean of land" (and thus presenting many semi-continental features) and isolated from other major brackish-water bodies by physical (ocean and land) barriers. In historic time many of these barriers have been weakened through man's activities: the opening up of interoceanic and intracontinental canals, the intentional or unintentional transportation of parts of the habitat itself or the biotic community living in it from one area to another (ballast water, dry ballast, fouling communities), the transplantation of commercial species of fish and shellfish with associated organisms, etc.
The Baltic Sea is a very young brackish-water sea. Practically all the marine fauna and flora have invaded the Baltic during the last 10,000 years, and it is obvious that this immigration still continues. After the latest glaciation period the Baltic as well as the Black Sea were

- 6 8 -
fresh-water lakes; presently they have meromictic conditions. The connection between the Baltic Sea and the NE Atlantic through the Danish Sounds was opened around 7,000 B.C.
The Baltic Sea basin is still isolated from the World Ocean by ecological barriers; one of the most important is the salinity factor. The surface salinity of the Baltic Proper is 6-8 %o salinity, i.e., approximaüvely a fourth or fifth of that of the adjacent North Sea. This results in an effective barrier to biological invasions between the brackish Baltic Sea and, e.g., East-Atlantic river mouths. In addition to its contacts by straits with the North Sea and the Atlantic Ocean, the Baltic Sea is connected by canals and rivers with the Black Sea and other adjacent bodies of water. Species introductions due to shipping operations via canals have resulted in major changes in some shore-near subsystems especially in estuaries in the Southern Baltic area.
The Baltic Sea - A sea of few species
The Baltic is a sea of few species of plants and animals. The sea is characterized by steep physical and chemical gradients, both horizontally and vertically (see Leppäkoski & Bonsdorff 1989). The water is too diluted for marine species, and too saline for freshwater species; e.g., of the more than 200 species of marine bivalve molluscs living in the North Sea, only 5 are found on the south coast of Finland, none in the northernmost part of it, the Bothnian Bay. Several major groups of marine animals, characteristic inhabitants of the oceans, are totally lacking in the Baltic Sea or found in its southwestern corner only (sea anemones, starfishes, sea urchins, brittle stars, crabs, lobsters, squids, sharks, etc.)
The number of animal species of marine origin decreases rapidly from the North Sea (> 1,000 species) and Kattegat (850?) to the Baltic Proper (about 80), the Bothnian Sea (>50) and the innermost Bothnian Bay (<10). This decrease is only to some extent compensated by an increase of the numbers of freshwater species. Thus the food webs become successively simplified and more easy to understand, from both structural and functional point of view.
The ecological importance of nonindigenous species must bee scrutinized against this background; a number of habitats and niches have obviously been unsaturated in the Baltic Sea. To take some examples, filter-feeding bivalves did formerly not occur on firm bottoms in the very diluted parts of coastal inlets and lagoons of the Baltic Sea. In the southern Baltic, the introduction of the zebra mussel (Dreissena polymorpha) filled this gap. In parts of the Szczecin Firth (Poland), Dreissena and the blue mussel (Mytilus edulis) co-occur but they do not basically overlap owing to isolation by the salinity gradient (Leppäkoski 1984,). At places the muskrat {Ondatra úbethicá) lives as the only shore-living rodent, the introduced prosobranchiate Potamopyrgus jenkinsi as the only gastropod species on sublittoral son bottoms, the Chinese crab (Eriocheir sinensis) as the only decapod crab, the cirripede Balanus improvisus as the only representative of barnacles, etc.
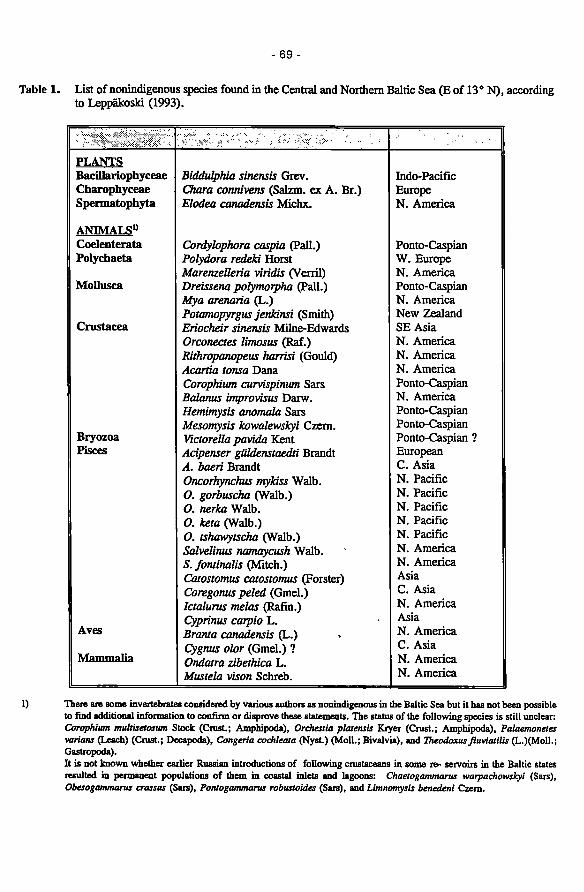
69-
Table 1. List of nonindigenous species found in the Central and Northern Baltic Sea (E of 13° N), according to Leppäkoski (1993).
;■ T^^r.^vf:!"i| \p^""' ■
PLANTS Bacillariophyceae Charophyceae Spermatophyta
ANIMALS" Coelenterata Polychaeta
Mollusca
Crustacea
Bryozoa Pisces
Aves
Mammalia
.·· ¿-».C Λ · . : ■ . , , · \ * -, ':' % ■ ,".. <... ; Λ ' ■'■ :
Ï-V ' \ ^ · - · y: < " '°'? ; *■ ''·· '■'.?'■'■' i ; i ? ' " ■ · ■< . ;
Biddulphia sinensis Grev. Chara connivens (Salzm. ex A. Br.) Elodea canadensis Michx.
Cordylophora caspia (Pall.) Potydora rédela Horst Marenzelleria viridis (Verril) Dreissena polymorpha (Pali.) Mya arenaria (L.) Potamopyrgus jenìdnsi (Smith) Eriocheir sinensis Milne-Edwards Orconectes limosus (Raf.) Rithropanopeus harrisi (Gould) Acartia tonsa Dana Corophium curvispinum Sars Balanus improvisus Darw. Hemimysis anomala Sars Mesomysis kowalewskyi Czem. Victorella pavida Kent Acipenser güldenstaedti Brandt A. baeri Brandt Oncorhynchus myldss Walb. O. gorbuscha (Walb.) O. nerka Walb. 0. keta (Walb.) O. tshawytscha (Walb.) Salvelinus namaycush Walb. S. fontinalis (Mitch.) Catostomus catostomus (Forster) Coregonus peled (Gmel.) Ictalurus melas (Rann.) Cyprinus carpio L. Branta canadensis (L.) Cygnus olor (Gmel.) ? Ondatra zibethica L. Mustela vison Schreb.
Indo-Pacific Europe N. America
Ponto-Caspian W. Europe N. America Ponto-Caspian N. America New Zealand SE Asia Ν. America Ν. America Ν. America Ponto-Caspian N. America Ponto-Caspian Ponto-Caspian Ponto-Caspian ? European C. Asia Ν. Pacific Ν. Pacific Ν. Pacific Ν. Pacific Ν. Pacific Ν. America Ν. America Asia C. Asia Ν. America Asia Ν. America C. Asia Ν. America Ν. America
There are some invertebrates considered by various authors as nonindigenous in the Baltic Sea but it has not been possible to find additional information to confirm or disprove these statements. The status of the following species is still unclear: Corophium multisetosum Stock (Crust.; Amphipoda), Orchestia platensis Kryer (Crust.; Amphipoda), Palaemonetes varians (Leach) (Crust.; Decapoda), Congeria cochleata (Nyst.) (Moll.; Bivalvia), and Theodoxus fluviatilis (L.)(Moll.; Gastropoda). It is not known whether earlier Russian introductions of following crustaceans in some re- servoirs in the Baltic states resulted in permanent populations of them in coastal inlets and lagoons: Chaetogammarus warpachowskyi (Sars), Obesogammarus crassus (Sars), Ponlogammarus robustoida (Sars), and Limnomysis benedeni Czern.

-70
Introduced species in the Baltic Sea
Both plant and animal species, not native to the Baltic, have been introduced, intentionally or unintentionally, into the Baltic during historic time. The Baltic is one of the most thoroughly investigated sea areas in the world. The appearance of a new species might therefore be expected to be observed and reported without delay. In fact, however, biological studies of the Baltic Sea are very unevenly distributed in space and time. It is not always easy to say whether a 'new' species has been overlooked in previous studies or whether it is a real newcomer.
More than 35 species of anthropochorous immigrants, mainly unintentionally introduced animals, have been reported from the central and northern Baltic (east of the Arkona Basin, approx. 13° E) or its terrestrial coastal systems (Leppãkoski 1984, 1991,1993, Rahkonen 1991); introductions of associated meio- and microorganisms, e.g., disease agents, are not included. Among this variety of species (Table 1) following mechanisms of dispersal can be identified: biofouling on ocean-going ships, seawater ballast, dry ballast, active or passive dispersal from adjacent freshwater systems along rivers or canals, intentional fisheries introductions, aquaculture, and escapees from fur farms.
Elodea canadensis
Eriocheir sinensis Potamopyrgus jen-kinsi
Cordylophora caspia
Chara connivens
Marenzelleria viridis
Corophium curvispinum
' Mesomysis kowalewskyi
Chaetogammarus warpa-chowskyi. Obesogamma-rus crassus. Pontogam-marus robustoides.Hemi-mysis anomala, Limno-mysis benedeni
'Rithropanopeus harrisi
Fig. 1. : Distribution of nonindigenous bentbic species in the Baltic Sea (E of 13 ° E) (From Leppãkoski 1984, 1991, and 1993; modified according to Nyman 1993).

71
Most of the species introduced with man's aid belong to the littoral or shallow sublittoral subsystems or shores. The hydrozoan Cordylophora caspia, the barnacle Balanus improvisas and the zebra mussel Dreissena pofymorpha (in the southern Baltic only) are, at places, common members of the biofouling community in shallow waters, especially at river mouths and in coastal inlets. Some exceptions can be mentioned. Mya can be found down to 30 m depth, at least. On sheltered sedimentation bottoms, Polydora rédela has been recorded at 29 m and Potamopyrgus jenMnsi at 23 m depth.
Of the 35-40 species listed (Table 1) 6 fish species have been intentionally introduced in the Baltic Sea. Of plants and invertebrates 15 species were unintentionally transferred from their primary or secondary donor areas, whereas 13 species have been intentionally planted in adjacent bodies of fresh water and from there spread to the Baltic coast.
The exact routes of dispersal are unknown in most cases; 21 species out of 35 may be considered transoceanic immigrants; not less than 18 of these originate from Ν America or the Ν Pacific area. The Ponto-Caspian complex, often considered an important source of Baltic Sea immigrants, is represented by 2-3 species only outside river mouths. There are examples of ballast species (Chara, Elodea, Eriocheir, Rithropanopeus, Acartia, Mya, Potamopyrgus) (see Table 1 for complete names of the species), species attached to the bottoms of ships (Cordylophora, Polydora, Marenzelleria (?), Dreissena, Balanus, Corophium, Victorella), and of species spread along river systems from adjacent lakes and reservoirs in which they have been acclimatized as economically valuable species (species of Mysidacea and Gammaridae listed in Table 1, Orconectes, Ictalurus, Cyprinus, Salvelinus, Ondatra).
All trophic levels are represented among the nonidigenous organisms in the Baltic Sea. The most successful species are members of detritus food chains (Dreissena, Potamopyrgus). In certain coastal waters, food chains and whole macrobenthic communities can be based upon introduced species. In the early 1960s, Dreissena made up no less than 88 % of the total benthic biomass in the Szczecin Firth, Poland (max. 20 kg m"2). Here the Ν American freshwater crayfish Orconectes limosus fed mainly upon the Ponto-Caspian coelenterate Cordylophora caspia, which uses planktonic larvae of the Ponto-Caspian bivalve Dreissena as food (Leppãkoski 1984, and references therein). ' In some estuarine habitats adult Rhitropanopeus fed mainly upon Dreissena whereas young individuals consumed Cordylophora; today most of the local Rhitropanopeus populations have strongly declined due to pollution.

72
Fig. 2. : Nonindigenous aquatic species in the southern Baltic Sea (from the left, Orconectes, Dreissena, Cordylophora, Rithropanopeus, Potamopyrgus, Chara, Corophium, Eriocheir. Poiydora, Mya, Coregonus, Acipenser, Salvelinus, Oncorhynchus; see Table 1 for complete names).
Sessile species, such as Balanus and Dreissena, change the physical structure of a habitat. They create secondary hard bottoms, thus multiplying the surface and volume available for associated species. Dreissena acts as a "sediment trap", turning huge amounts of seston into excreted pseudofaeces which create son substrates, rich in organic material, for deposit-feeding infaunal species.
The Baltic Sea seems to be quite well protected against introductions, the low salinity acting as an effective hindrance for most of them. There are only some few harmful introduced species in the Baltic Sea. In biofouling Cordylophora caspia, Balanus improvisus, and Dreissena polymorpha are of importance. Some exotics interfere even in the Baltic with boating. The common use of antifouling paints is due not only to the occurrence of annual, filamentous algae but mostly to the mass occurrence of Balanus improvisus attached to boat hulls. Biofouling also cause considerable losses in heath exchangers at industrial plants.
Some environmental problems have been caused by alien vertebrate species inhabiting major parts of the Baltic coastline. The aggressive Canada goose Branta canadensis, being the largest goose of Europe and the most common goose species living in the Baltic Sea, was firstly expected to became a severe competitor for the grey goose {Anser anser). The muskrat (Ondatra zibethica) but also the Chinese crab (Eriocheir sinensis) can cause some damage by their burrowing activities; the mink (Mustela vison) is a predator on water fowl.
In the Baltic the contribution of exotics to fisheries is almost zero. In aquaculture the North American rainbow trout is in practice the only species kept in fish-farming. This species can not, however, reproduce in the Baltic Sea area. Canada goose, muskrat and mink are hunted in Finland and Sweden.
Examples of the latest newcomers in NW European seas are the American jack-knife clam Ensis directus (E. americanas), first found in the German Bight in 1978, now recorded along

- 7 3 -
the NW European coast and on the Swedish west coast in NE (Essink 1988), Marenzelleria viridis, a spionid polychaete from N. America, first recorded in 1983 in the Netherlands, now found in the S and SE Baltic (Gruszka 1991) and, in 1990, on the S coast of Finland (Norkko et al. 1993), and the brown alga Sargassum muticum, introduced into Europe in the late 1960s, now found on the Swedish west coast (Rueness 1989). The Ponto-Caspian opossum shrimp Hemimysis anomala, earlier known from Russian introductions in a coastal lagoon in Lithuania, was found on the SW coast of Finland in 1992 (V. Hietalahti & H. Salemaa, unpublished).
Fig. 3. : Introduced mammals and birds in the SW archipelago of Finland (from the left, white-tailed deer (Odocoileus virginianus), pheasant (Phasianus colchicus), mute swan (Cygnus olor), mink (Mustela vison), Canada goose (Branta canadensis), raccoon-like dog (Nyclereutes procyonoides), and muskrat (Ondatra zibethica).
Conclusions
There is an inherent resistance to invasions in ecosystems (Mooney & Drake 1987). This capacity seems to have been reduced in enclosed coastal seas by the habitat modification caused by man's activities. This phenomenon is very well-known from several terrestrial systems. In brackish-water ecosystems the situation seems to be greatly the same. Nonidigenous species are numerous and, at places, predominant in strongly modified habitats (harbours, underwater constructions, receiving waters of pollutants and cooling water discharges). Such perturbations tend to alter not only the physical structure of the habitat (e.g., by offering secondary hard substrates for sessile species or extremely soft bottom material unsuitable for others than some specialized mud dwellers) but also energy flow patterns and interspecific relationships. Initial symptoms of ecosystem distress are often followed by major changes in species composition, including the invasion by exotic species (Rapport et al., 1985). In the Baltic Sea eutrophicated inlets and coastal lagoons and fouling communities, especially in the southern part of the area, serve as examples of sub-systems

- 7 4 -
that have been highly modified by introduced species, whereas the open coast and pelagial systems are relatively free from them.
When an entirely new growth form is introduced in a community, major ecological alterations can be expected. For example, sessile forms as cirripeds (Balanus improvisus) and the bivalve Dreissena polymorpha (in estuaries and lagoons on the southern Baltic coast) became space-dominant and influenced a great variety of accompanying species.
Brackish water regions in estuaries seem to be more open to immigration of exotic species than are the (in geological and evolutional sense older) marine and fresh-water parts of an estuary. In Northwest-European estuaries, some 3-5 % of all species are immigrants in the marine or fresh-water parts of the estuary but in the brackish water areas this percentage can be as high as 20-28 % (Wolff 1973; Vaas 1975). One reason to this has by several authors mentioned to be the fact that the number of indigenous species is low and there are many "empty", "vacant" or "free" niches in brackish-water systems. However, an empty niche is difficult to predict because an introduced species may develop a different realized niche in its novel environment, as a response to new ecological conditions.
Due to their ecological and evolutionary history, both the Baltic and the Black Sea seem to be predominately receiver areas of introduced species, donor areas of which are not to be found in the adjacent coastal oceanic waters only but also in distant seas. Several introductions into the Baltic and the Black Sea have resulted in both structural and functional changes and thus pose significant control mechanisms at ecosystem level; in fact, food chains and whole benthic communities can be based upon introduced species in both seas. Most exotics do, however, not have any influence on man's economic interests; their ecological importance has in most cases been more or less unobservable.
There are four successive stages of invasions : (1) arrival, (2) establishment, (3) spread, and (4) persistence (see Mollison (1986) for a comprehensive discussion). Perhaps hundreds of introduced species have arrived (most of them obviously by mere change) into the Baltic Sea or its coastal subsystems as planktonic larvae in ballast water tanks, as sessile organisms attached to hulls (Dreissena, Balanus), or as benthic stages living in (or on) the sediment settled on the bottom of the tanks (Mya, Potamopyrgus), or have been introduced intentionally (salmonid fishes, Canada goose). Most of them have been unable to maintain a self-sustaining population and thus only some tens of these species have established in the Baltic and still fewer have been able to spread in sea-wide scale. Their ultimate success may be evaluated in terms of persistence time. Mya arenaria has persisted in NW European waters in 300-400 or perhaps 700 years (Petersen et al., 1992), in the Black Sea not more than 30 years till now. Balanus improvisus has been very successful in the Baltic Sea since the late 1800s. There might be several unknown introductions from older times; as pointed out by Carlton (1989), the problem is not always how we know a species is introduced but how we know a species is native.
In conclusion, it is apparent that in the Baltic Sea only some few introduced species have been able to compete with native species, and in some cases displace them. This fact gives further evidence of the opportunistic nature and huge expansive capacity of introduced species when once transported with Man's aid over physical and ecological barriers into a new habitat.
All vessels destinated for any Baltic Sea harbour have to pass fully marine waters with oceanic salinities. Thus a relatively simple and cheap method of reducing the risk of unintentional introductions of estuarine (brackish-water) species into enclosed brackish-water

- 7 5 -
seas should be mid-ocean exchange of ballast water en route, as suggested by, e.g., Williams et al. (1988).
Man seems to be unable to intentionally manipulate the species composition of the Baltic Sea. The nonindigenous element has, generally speaking, occupied only marginal areas and in most cases only marginal niches (Leppãkoski, 1984). Biogeographically, however, this element signifies a great addition to the Baltic fauna and flora, with respect to the naturally low number of species present. For a conservationist, this addition indicates contamination of the biota by alien elements, reflecting the history of man's economic interests and activities. From an ecologist's point of view, nonindigenous species have contributed to species diversity and community structure, created new interspecific relationships, etc.
The enrichment of the Baltic Sea fauna and flora by introduced species can, from a biogeographer's point of view, be interpreted as a successive recovery from the last glaciation, anthropochorous dispersal being a spreading mechanism complementary to other manners of dispersal (Leppãkoski, 1984).

-76
Concerns and activities of the ICES Working Group on Introductions and Transfers of Marine organisms
Inger WALLENTINUS
Department of Marine Botany, University of Göteborg, Carl Skottsbergs Gata 22, S- 413 19 Göteborg, Sweden
Background
The International Council for the Exploration of the Sea (ICES) was already in the late 1960s concerned of the potential risks with introducing foreign marine species into new areas. Thus, in 1968 ICES stated the following : "During the discussion which arose the need for the establishment of principles to guide member countries [i.e. countries bordering the North Atlantic north of a line between Gibraltar and Cape Harteras, USA] contemplating the transplantation or acclimatization of alien species of marine animals and plants was mentioned. There was a general agreement on the advisability of giving attention to this subject, and it was agreed that the Committee would return to it in 1969. It was also mentioned that the Office International des Epizoo ties, Paris, was concerned with this matter."
In 1969 this concern resulted in a Council resolution and the establishment of a Working Group (WG) on Transplantation and Acclimatization of Alien Species into New Areas", which at its first meeting in 1970 took the name "WG on the Introduction of Non-Indigenous Marine Organisms". Four meetings were held between 1970 and 1974. From 1979 the WG has had regular annual meetings under the chair of Dr C. SINDERMANN (since 1981 under its present name), and since 1991 under the chair of Dr J. CARLTON. In 1986 the Council adjusted the previous terms of reference so that the WG would "assume the long-term responsibility for producing continuing advice to the Council on all matters relating to introductions and transfers".
Already during the first years one of the main goals of the WG was to establish an accepted International Code of Practice (see below) for procedures to be used to minimize the adverse risks with intentional introductions and transfers, as well as to collect and spread information on existing and proposed introductions. The main emphasis then was on shellfish and fish, mainly salmonids, and the concern to a large extent was focused on the spread of diseases and parasites.
In the early 1980s also the risks of ecological, genetic and behavioural impact was added to the Code (see below). Furthermore, attempts were made to keep a record of all applicable national laws and regulations concerning introductions of marine species.
The principles and policy of the WG never was to stop introductions of marine species totally, but based on scientific evidence to objectively evaluate the risks and impact on the marine environment of proposed introductions and to encourage the use of native species for mariculture, whenever feasible. Thus the goal is to cease poorly-considered introductions.

-77 -
Moreover the WG also has the mission to urge the member ţcrim^rj^tb guard ¿gainst accidental introductions into their waters.
Proposals of intentional introductions and transfers
As pointed out above the implementation of the Code of Practice has been a main issue of the WG. The first Code was adopted by the Council in 1973 and it has since been amended several times, the latest revised version (see Appendix) being adopted in 1990. Since 1985 the Code through co-operation with EIFAC (European Inland Fisheries Advisory Council) also applies to freshwater organisms. Guidelines and manual of procedures to the Code have been published several times (e.g. ANON. 1984, TURNER 1988, CARLTON 1992a) and some countries have translated the Code. In co-operation with EIFAC a new revised Code, including also Genetically Modified Organisms (GMOs), is now under preparation, and will be presented to ICES in 1993. Lately there has been great concern that the new EEC directives (91/67/EEC) permit- ting free transfers of shellfish between the EEC countries might counteract the intention of the ICES Code by increasing the risk of accidental introductions of parasites and accompanying species into waters in which they still do not occur, since no permission of the receiving country is needed for the import.
Several proposed introductions have been considered by the WG; only some examples will be given here. However, in some events the case was brought to the attention of the WG only after the first introduction already had taken place. Besides mariculture the intentional introductions and transfers also include scientific experiments, stock enhancements, use in hatcheries, import for storage prior to sale and for consumption, as well as for recreational purposes (see also CARLTON 1992b). Any project, independent of purpose, should be treated according to the Code of Prac- tice, although this can be a time-consuming procedure because of the amount of information that has to be assembled. To be able to achieve this
- the WG actively tries to spread in formation about the Code to as many groups as possible, including universities and the general public.
One of the earliest cases of deliberate introduction of seaweeds was the fanning of the kelp Macrocystis pyrifera in France, discussed at ICES meetings in 1973-74 (e.g. ICES 1974), when it was decided that the farming should be stopped. In 1984 the introduction to Brittany, France, of the Japanese brown alga Undaria pinnatifida ("wakame") using plants accidentally introduced with oysters to the French Mediterranean coast, was brought to the attention of the WG (ICES 1984), after the farming of the sporophytes in the sea already had started. The project was extensively discussed during several WG meetings until 1989. The proposers stated that it was not possible for the gametophytes to mature and produce sporophytes in situ at the water temperature regimes prevailing in Brittany, although evidence that it was possible was reviewed in the WG reports (for references see ICES 1986, FLOC'H et al, 1991). In 1987 FLOC'H and his co-worker showed that wild plants, originating from offspring of the cultured ones, did occur in the area (ICES 1987, FLOC'H et al., 1991) and that the full life cycle of the alga thus could be completed. In 1989 the WG was to give advice to ICES if the fanning of Undaria could be continued or should be stopped. After contact with several experts and reviews of the literature the final conclusion of the WG (ICES 1989) was that the cultivation could continue, also on a commercial scale, even though Undaria could reproduce naturally. However, it was pointed out in the advice of the WG that it seemed inevitable that Undaria would spread further, but that the detrimental effects on the ecosystem were considered not to be so large as to motivate stopping the project, which had been going on in a pilot scale since 1983. Annual reports on the farming sites and amounts were to be

- 7 8 -
given to the WG with a sum- mary of the ecological and environmental impact to be presented to ICES in 1994.
In 1991-92 a proposal to introduce a strain of the Japanese red alga Porphyra yezoensis to the coasts of Maine, USA, and New Brunswick, Canada, was examined by the WG (ICES 1991, 1992). The species is of great interest for producing high quality "nori" and free conchocoelis stages were to be brought from a hatchery in the State of Washington, to which it had earlier been imported from Japan. In the proposal it was stated that the water temperature in the suggested farming area was too low when the light : dark regime was suitable for the species to reproduce sexually and that no escapes have been seen on the Pacific American coast. However, the earlier proposals did not fully consider that the species had been accidentally introduced and had spread in the Mediterranean Sea, nor was any possible ecological/genetic impact discussed in case the plants/nets might be dislodged and spread to other waters. The final outcome of the proposal will be decided at the WG meeting in 1993, when additional information on the project and a more comprehensive literature survey of the species will be provided.
Several proposals of introducing molluscs have been discussed by the WG (for reviews see e.g. GRIZEL & HERAL 1991, UTTING & SPENCER 1992, GRIZEL this report, MUNRO et al., in press). The proposal in 1989 to introduce the Japanese scallop Patinopecten yessoensis to Ireland (ICES 1989, 1990) can be brought up as a good example of how a case should be treated according to the ICES Code of Practice. The proposal was accompanied by an extensive documentation and a very comprehensive literature survey over the last 10 years of the biology of the species in Japan (including known predators and competitors as well as parasites and pathogens), of possible inter- actions with native species in Ireland (predators and competitors as well as any possible risk of genetic impact). The introduced specimens were held in strict quarantine (a description of the hatchery facilities was included in the proposal) for studies of potential diseases in the imported brood stock, which was later destroyed. The project, which has been continuously reported to the WG, started on a pilot scale in 1990 when a limit amount of disease-free spat of the ̂ -generation were set out nets in the sea. However, the project has not yet developed into a financial success due to too low growth rates.
Status reports and overviews of ongoing introductions (intentional and accidental)
Besides giving advice on proposed introductions, a main issue of the WG has been to follow up reported introductions, intentional as well as accidental ones. National reports on introduced species are given at the annual meetings, generally based on national statistics and published literature as well as personal communications. Special emphasis is given to reports of species having gone through the procedures of the Code of Practice. These reports constitute the basis for the Status reports of introduced species, published for each decade since 1970 (ANON. 1972,1982, MUNRO et al., in press). Unfortunately, however, several countries not being able to participate in a meeting often neglect to deliver the reports, and also the reports are seldom complete for accidental introductions, which is especially true for algae. Thus when preparing the 1990 Status report it was decided to make a full cover of all introduced algae and their present status in the ICES member countries, based on literature surveys and questionnaires to phycologists in the different countries (WALLENTINUS 1992, MUNRO et al., in press).

- 7 9 -
Accidental introductions and identification of major risk vectors
During the last two decades there has been an increased awareness of the risk of accidental introductions and the interest of the WG in such events has risen considerably by the many cases presented, not the least due to the economic impact caused by many of them. Several vectors are responsible for these accidental introductions, the main ones being briefly described here.
Ballast introductions
Recently, there have been several episodes (see examples given below) focusing the WG's attention on ballast water introductions (by water and sediment) and the present chairman has for many years been involved in a study programme on the dimension of this problem, including sampling on board ships (e.g. CARLTON 1985, 1987, 1989, 1992c). Another vector scrutinized is introductions of fouling species on the ships' hulls. In 1991 the WG had a study group prepare recommendations for control of introductions through ballast discharges (ICES 1991). These included : Suggestions of guidelines for discharges of ballast water; definitions of geographic primary and secondary areas if exchanges were to take place at sea; encouragement of programmes for sampling and documentation of occurrences of non-indigenous species in ballast water and sediment; contacts with representatives of IMO (the International Maritime Organization) and educational programmes to inform the shipping industry and general public of the potential problem associated with release of ballast water.
The accidental introduction of the Zebra mussel (Dreissenia polymorpha) into the Laurentian Great Lakes in the mid 1980s (for references see e.g. ICES 1992) as larvae in ballast water, caused a population boom and has led to enormous clean-up operations of water pipes, jetties, buoys etc. for billions of dollars. This has resulted first, in voluntary guidelines in Canada and USA for the ships going into the Great Lakes, and then secondly, in the spring of 1993, in a new law being adopted in the USA). Also many other organisms brought by ballast water have been established in this area in the 1980s. The continuous spread of the Zebra mussel through rivers and canals and its impact is followed by a large number of scientists (e.g. NALEPA & SCHLOESSER 1992). Recently it has been found that a second closely related, but still not identified, Zebra mussel now occurs in the Great Lakes. Due to the different ecological performance of this species it might be a threat to new areas. The WG has also noticed with satisfaction how information on the introduction of the Zebra mussel is spread to the general public by various means.
The introduction of toxic phytoplankton species into Australia (e.g. HALLEGRAEFF et al., 1990, HALLEGRAEFF & BOALCH1991, HALLEGRAEFF 1993) and the threat these pose to aquaculture, have also illustrated the great significance of this problem, not the least through Australia's activity in the MEPC (Marine Environment Protection Committee) of IMO, resulting in voluntary guidelines for the shipping. The undertaking of the IMO is closely followed by the ICES WG, and the WG members are asked to maintain a liaison with the IMO's contact persons in their countries. At the Helsinki meeting (ICES 1991) the WG also emphasized the need for more taxonomic expertise on phytoplankton cysts (especially those of the toxic dinoflagellates) as well as research on which methods to use in order to destroy cysts occurring in the ballast tanks' sediment.
A third recent, severe case accentuating the ballast problem, is the accidental introduction of the American combjelly (ctenophore) Mnemiopsis leidyi in the Black Sea during the early 1980s. In the last ten years this has developed into a vast problem and eliminated the local fishery in the Azov Sea due to the huge filtering capacity of the introduced populations. The

- 8 0 -
species may now spread also into the Mediterranean Sea. The prospect of introducing predator species into the Black Sea is considered (J. CARLTON pen. comm.), but is a severe step which should be carefully judged by scientific expertise before being implemented.
Species accidentally introduced with deliberately introduced species
The early versions of the Code of Practice pointed to problems of introductions brought accidentally with target species and mentioned parasites and micro-organisms as well as epibiotic flora and fauna. Thus the need of quarantine facilities and the release into open waters of the first generation (F,) only, but not of the originally imported brood stock was strongly emphasized. The main concern of the WG is still to prevent such unnecessary introductions and it is highly important that the procedures of the Code are followed. Despite the existing Code, several new organisms have been introduced during the 1970s and 1980s, especially by the import of Japanese oysters. For instance the import of Japanese oysters has brought around a dozen of exotic macroalgal species to Europe (e.g. BOUDOURESQUE this report, WALLENTINUS 1992, MUNRO et al., in press). In many cases these algae have be- come dominant in the new areas (e.g. the brown algae Sargassum muticum, Laminaria japónica, Undaria pinnatifida) and some have later spread to other areas by natural means, while in some cases they have been dispersed to other European coasts through illegal imports of oysters. There are many examples of associated animals, symbionts and parasites being spread to new areas by this vector, too (e.g. CARLTON 1992b, MUNRO et ál., in press).
Release of organisms being kept in aquaria
There is hardly any control of release of organisms imported for omamental purposes including the aquarium trade. Although this trade may not reach the same volume as that of aquaculture, the literature reveals many examples of species which have become established as a result of escapes or through the "concern" of people about saving the lives of their former pets (for references see e.g. CARLTON 1992ft, MUNRO et al., in press). To inform the general public about the ecological consequences of these actions is thus an essential part of the task of the WG in co-operation with the national authorities. As described by BOUDOURESQUE in this report, the dispersal of the tropical green alga Caulerpa taxifolia in the Mediterranean may be linked with the aquarium trade. Although not strictly occurring within the area encompassed by ICES, this event has been of great interest to the WG, since the species might reach and possibly tolerate the winter temperatures on the Atlantic coasts of some of the ICES countries and thus, if it were to become established may have an impact also on these marine ecosystems. Therefore, the incident was thoroughly discussed at the WG meeting in 1992.
Fishing activities
In the 1990 Status report (MUNRO et al., in press), several introductions listed might have been caused by discarded packings of baits or by bait organisms themselves. Several others are listed by CARLTON (1992¿>), who also pointed out cases where species associated with seaweeds have been spread by this vector, a feature probably occurring more commonly than normally realized. Although this not one of the main issues, the WG still seeks to underscore that there is a risk in all unintentional introductions and that information to the general public is highly important to prevent this to happen.

-81 -
Stock enhancement
These events, which mostly are transfers offish or shellfish species, have been of interest to the WG both because the gene pool of the species might be affected and, as for the GMOs, there might be an ecological impact as well. The efficiencies and strategies of these programs have been debated and as a result of the WG's activity (cf. also CARLTON 1992Z») there will be an international theme session on these questions at an ICES Statutory Meeting, suggested to cover all types of organisms, including also invertebrates and seaweeds.
Genetically modified organisms (GMOs)
The WG has had a joint meeting with ICES WG on Genetics to discuss the possible impact of releases of GMOs. So far most attention has been focused on fish, but also other organisms have been discussed. The EEC Directives (90/220/EEC) have thoroughly elucidated these matters and many ICES member countries have adopted these directives and several of the countries outside EEC have similar directives or laws of their own. Besides any genetic impact the GMOs may have, they, including sterile specimens, may also affect the marine environment through their increased fitness. Thus, as mentioned above, the proposal for a new revised Code also will include GMOs. With the increased scientific interest in the development and achievements of these techniques this area may be an important field for the future activities of the WG.
International cooperation
The ICES member countries only comprise those of the North Atlantic shores. It is thus essential that the procedures such as those contained within the Code of Practice can be accepted also in other geographical areas, as has been the case with the European inland waters through EIFAC. Collaboration with other organisations such as EEC and CIESM, as well as with those working in the Pacific and in the tropical waters is thus most important to accomplish the overall goal of saving the marine ecosystems and natural resources of all oceans from being altered by unwanted introductions of alien species.

- 8 2 -
Annex 1 : Revised (ICES) 1990 code of practice to reduce the risks of adverse effects arising from introductions and transfers of marine species
As adopted by the International Council for the Exploration of the Sea, October 1990 (Council Resolution 1990/4:6; ICES Annual Report for the 78th Statutory Meeting 1990 (Copenhagen) January 1991, page 194 and ICES Document CM. 1990/F:37. A Report of the Working Group on Introductions and Transfers of Marine Organisms).
I Recommended procedure for all species prior to reaching of decision regarding new introductions. (A recommended procedure for introduced or transferred species which are part of current commercial practice is given in Section IV).
a) Member countries contemplating any new introduction should be requested to present to the Council at an early stage in the life cycle, area of origin, proposed plan of introduction and objectives, with such information on its habitat, epifauna, associated organisms, potential competition to species in the new environment, etc., as is available. The Council should then consider the possible outcome of the introduction, and offer advice on the acceptability of the choice.
b) Appropriate authorities of the importing country (including fishery management authorities) should examine each "candidate for admission" in its natural environment, to assess the justification for the introduction, its relationship with other members of the ecosystem, and the role played by parasites and diseases.
c) The probable effects of an introduced species in the new area should be assessed carefully, including examination of the effects of any previous introduction of this or similar species in other areas.
d) Results of b) and c) should be communicated to the Council evaluation and comment.
Π If the decision is taken to proceed with the introduction, the following action is recommended
a) A brood stock should be established in a quarantine situation approved by the country of receipt, in sufficient time to allow adequate evaluation of its health status.

83-
The first generation progeny of the introduced species can be transplanted to the natural environment if no diseases of parasites become evident in the F, progeny, but not the original import. In the case of fish, brood stock should be developed from stocks imported as eggs or juveniles, to allow sufficient time for observation in quarantine.
b) The F¡ progeny should be placed on a limited scale into open waters to assess ecological interactions with native species.
c) All effluent from hatcheries or establishments used for quarantine purposes in recipient countries should be sterilized in an approved manner (which should include the killing of all living organisms present in the effluent).
d) A continuing study should be made of the introduced species in its new environment, and progress reports submitted to the International Council for the Exploration of the Sea.
m Regulatory agencies of all member countries are encouraged to use the strongest possible measures to prevent unauthorized or unapproved introductions
IV Recommended procedure for introduced or transferred species which are part of current commercial practice
a) Periodic inspection (including microscopic examination) by the receiving country of material prior to mass transportation to confirm freedom from intraducibie pests and diseases. If inspection reveals any undesirable development, importation must be immediately discontinued. Findings and remedial actions should be reported to the International Council for the Exploration of the Sea.
b) Inspection and control of each consignment on arrival.
c) Quarantining or disinfection whenever possible and where appropriate.
d) Establishment of brood stock certified free of specific pathogens.
It is appreciated that countries will have different attitudes toward the selection of the place of inspection and control of the consignment either in the country of origin of in the country of receipt

84
Definition For the application of this code, the following definitions should be used :
Marine species. Any aquatic species that does not spend its entire life cycle in fresh water. Introduced species (= non-indigenous species, = exotic species). Any species intentionally or accidentally transported and released by humans into an environment outside its present range. Transferred species (= transplanted species). Any species intentionally or accidentally transported and released within its present range. Quarantined species. Any species held in a confined or enclosed system that is designed to prevent any possibility of the release of the species, or any of its diseases or any other associated organisms into the environment. Country of origin. The country where the species is native. Exporting country. The country from which a specific consignment of a species (regardless of its native region) is received. Brood stock. Specimens of a species, either as eggs, juveniles, or adults, from which a first or subsequent generation may be produced for possible introduction to the environment. Disease. For the purpose of the Code, "disease" is understood to mean all organisms, including parasites, that cause disease. (A list or prescribed disease agents, parasites, and other harmful agents is made for each introduced or transferred species in order that adequate methods for inspection are available. The discovery of other agents, etc., during such inspection should always be recorded and reported). Current commercial practice. Established and ongoing cultivation, rearing, or placement of an introduced or transferred species in the environment for economic or recreational purposes, which has been ongoing for a number of years. Established species. Species with existing reproductive populations. Maintained species. Species which are reproducing in aquaculture for several generations without artificial spawning.
Note
a) It is understood that an introduced species is what is also referred to herein as an introduction,; a transferred species as a transfer, and a quarantined species as a species in quarantine.
b) Introduced and transferred species, as defined above, include those species subject to the ICES Code of Practice, parts I to m, and IV, respectively.
c) Introduced species are understood to include exotic species, while transferred species include exotic individuals or populations of a species. It is, thus, understood that the general term "exotic" can include both introduced and transferred species.
d) It is understood for the purpose of the Code that introduced and transferred species may have the same potential to carry and transmit disease or any other associated organisms into a new locality where the disease or associated organisms does not presently occur.

- 8 5 -
The Introduction of Exotic Species and the Law
Cyrille DE KLEMM
21, rue de Dantzig, 75015 Paris
Introduction
The risks involved as a result of the introduction of alien species into the natural environment are well known to scientists world wide. For a long time however, public awareness of the potentially harmful effects of such introductions was mostly limited to pathogens of cultivated plants and domestic animals. The International Plant Protection Convention of December 1951 requires Contracting Parties to issue phytosanitary certificates for any export or reexport of plants. Importing countries may prohibit or restrict the entry of any plant into their territory for phytosanitary reasons. The Convention is of world-wide application.
Although there is no similar treaty with regard to animals, most countries have enacted animal quarantine regulations to prevent the introduction of diseases.
In contrast with these strict rules governing the import of domestic animals and plants, the introduction of wild species remained unregulated for a long time. Introductions were often considered to be a way of enriching the local fauna and flora for the benefit of hunters, fishermen and other groups. In countries such as Australia and New Zealand, acclimatisation societies flourished and hundreds of species were introduced. These countries now have the strictest introduction controls in the world.
The damage caused by ill-conceived introductions is now almost universally considered to be unacceptably harmful to biological diversity, and legislation has gradually been enacted in many countries to bring such introductions under control. As the problem is clearly international, specific provisions dealing with this subject have been included in most recent nature conservation conventions. However, the implementation and enforcement of legal provisions establishing controls on introductions is very difficult in view of the many practical problems which may arise.
1. A Review of existing Legislation
A. Treaties and other International Instruments
Provisions dealing with introductions appear in several global treaties as well as in an increasing number of regional conventions.
First and foremost, the new Convention on Biological Diversity of 5 June 1992 requires its Contracting Parties, as far as possible and appropriate,

- 8 6 -
"to prevent the introduction of, control or eradicate those alien species which threaten ecosystems, habitats or species", (article 8(h)).
In parallel and with particular regard to the marine environment, the Law of the Sea Convention of 1982, which will hopefully enter into force in the not-too-distant future, provides that
"States shall take all measures necessary to prevent, reduce and control ...the intentional or accidental introduction of species, alien or new, to a particular part of the marine environment, which may cause significant and harmful changes thereto" (article 196).
The term, "new species", is understood in this context as meaning genetically modified organisms.
Another global convention which applies both to the terrestrial and marine environments is the Convention of the Conservation of migratory Species of Wild Animals (Bonn, 1979). The Bonn Convention states that Agreements concluded under the Convention for the conservation of individual species or of groups of species should provide for
"the protection of (the) habitats (of migratory species) from disturbances, including stria control of the introduction of, or control of already introduced, exotic species detrimental to the migratory species" (article V (5) (e)).
Certain regional conservation instruments contain similar provisions. In Europe, the Berne Convention of 1979 on the Conservation of European Wildlife and Natural Habitats provides that
"each Contracting Party undertakes ...to strictly control the introduction of non-native species" (article 11).
The new EC Directive of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora requires member States to
"ensure that the deliberate introduction into the wild of any species which is not native to their territory is regulated so as not to prejudice natural habitats within their natural range or the wild native fauna and flora and, if they consider it necessary, prohibit such introduction" (article 22 (b)).
With regard more specifically to introductions into the marine environment in European seas, the only instrument which refers to the matter is the Protocol to the Barcelona Convention concerning Mediterranean Specially Protected Areas. This instrument, which was signed in Geneva in 1982, is limited in scope as it only addresses the problem in respect of protected areas. The Protocol obliges Parties to take measures progressively to protect these areas. These may include the prohibition of the introduction of exotic species, as well as "the regulation of the introduction of indigenous zoological or botanical species" in protected areas.
In contrast, the much more recent Protocol concerning Specially Protected Areas in the Wider Caribbean Region (Kingston, 17 January 1990) goes much further, as it applies to the whole of the geographical area concerned rather than just to protected areas. The Protocol lays down the obligation for Parties

- 8 7 -
" to prohibit intentional or accidental introduction of non-indigenous or genetically altered species to the wild that may cause harmful impacts to the natural flora, fauna or other features of the Wider Caribbean Region"
Strangely, the new Convention on the Protection of the Marine Environment of the Baltic Sea Area, signed at Helsinki on 9 April 1992, is silent on the matter of introductions. However, article IS of the Convention does require Parties to take all appropriate measures to conserve natural habitats and biological diversity and to protect ecological processes in the Baltic Sea and its coastal ecosystems. This provision could easily be interpreted as including the need to control introductions. Parties could therefore decide to adopt, as the Convention encourages them to do, subsequent instruments containing appropriate guidelines and criteria on the subject, or even an Annex to the Convention which would be binding in law.
B. National Legislation
An increasing number of countries have now adopted legislation to control introductions, but this is still far from being the case everywhere.
A distinction must firstly be made between three types of introduction, namely voluntary introductions, introductions resulting from escapes from captivity, or purely accidental introductions caused by the inadvertent transportation of harmful organisms by various means of transport, especially by ship.
Whereas the first two types of introduction may easily, at least in theory, be controlled by legislation, it is clear that the third type necessitates the taking of stringent measures over and above present phytosanitary and quarantine re-quirements in order to be effective. Few countries are probably prepared to accept such draconian measures. Where legislation does exist, it is therefore almost always limited to the first two aspects of the problem.
1. Voluntary Introductions
As introductions are often considered, rightly or wrongly, to be beneficial, there is no law which imposes a total prohibition in this field. The usual method instead consists of making introductions subject to a permit requirement. One such example is the United Kingdom's Wildlife and Countryside Act. 1981. Unless a licence is obtained in advance, this Act prohibits the release of any animal of a kind which is not an ordinary resident in, or a regular visitor to, Great Britain, as well as of any animal belonging to certain species which have already been introduced. The prohibition only applies to certain listed species of plants, which include a number of seaweeds.
Another example is the German Nature Conservation Act. 1976, which prohibits the release or plantation in the wild of non-indigenous animal or plant species, unless a permit has first been issued. The Act specifies that permits shall not be granted where it is impossible to exclude the risk of altering indigenous species or of jeopardizing the survival or range of indigenous species, or population of those species, as a result of the proposed introduction.
The Swiss Nature Conservation Act. 1966 requires a permit from the federal authorities for the introduction of non-indigenous species, sub-species or races of animals and plants. A permit is also required for the introduction into one region of the country of animals and plants which are not native to that region, even though they are indigenous to Switzerland as a whole.

- 8 8 -
A few laws provide praticai guidance for the avoidance of harmful voluntary introductions. For example, the Inland Fisheries Regulations 1973 of the Australian State of Tasmania prohibit the introduction of live fish which are able to live in waters the temperature of which is below 10CC. They also prohibit the possession, in the vicinity of river banks or lake shores, of live fish, amphibians or invertebrates belonging to species which are not already present in the river or lake concerned.
The precautionary principle is increasingly embodied into legislation, in the form of a provision that a permit for any introduction shall be granted only if the species introduced will cause no harm to indigenous species or ecosystems. This is indeed laudable. However, for this provision to be effectively implemented, the permit-issuing authority must be able to found its decision to grant or deny a permit on the best scientific evidence. Arbitrary or seemingly arbitrary decisions may be challenged in the courts in many countries. One possibility is to prohibit the authority concerned from issuing a permit unless some clear conditins are fulfilled.
By way of example, the Fish and Fisheries Act 1979 in Australia's Northern Territory only authorises the Director of Fisheries to grant permits where the proposed introduction concerns a native species or a species specifically listed as an authorised species by Regulations. When deciding whether to grant or deny the permit in respect of the latter species, the Director must also take into consideration the environment, the need to control disease and the applicant's character, experience and reasons for making the application. A permit may be granted subject to such limitations and conditions as the Director thinks fit.
In Europe, the Codes of pratice which were developed by the International Council for the Exploration of the Sea (ICES) and the European Inland Fisheries Advisory Commission (EIFAC), in 1984 and 1987 respectively, could constitute a useful basis for the issue of introduction permits for aquatic organisms. However, these documents have no legal status and are consequently not binding in law.
2. Escapes from Captivity
As the EIFAC Code of Practice rightly puts it, escapes are inevitable. The keeping of exotic species in captivity therefore presents the same potential problems as voluntary releases to the wild.
The risk may, however, be reduced by the imposition of strict standards for facilities where species are held captive, as exist inter alia for mink farms in the United Kingdom, and by the establishment of permit requirements for the possession, sale and transport of live exotic animals. In the Northern Territory of Australia, for instance, the sale of any live exotic aquatic animal is prohibited, except for those species which may be listed by Regulations. In the same jurisdiction, the transport of any live mamais, birds, amphibians and reptiles is also prohibited without a permit.
Furthermore, certain laws make it an offence to liberate exotic animals or to allow them to escape, whether negligently or otherwise. This is the legal position in the United Kingdom and the Northern Territory of Australia.
Although these measures may reduce the risk of escapes, they cannot eliminate it. There may be fires, earthquakes, floods and accidents, or there may be vandalism. There may even be do-gooders who want to set the animals free in spite of the penalties they may thus incur.

-89 -
The safest solution is consequently to prohibit the import of all live exotic animals, unless it is certain that such animals would not survive in the wild if they were to escape. This is the method which has been adopted in New Zealand and in the States of Australia. In the Northern Territory, once again, the bringing into the Territory of live mammals, birds, amphibians and reptiles is subject to a permit : there are also species which are listed as prohibited imports for which no permits may be granted. Similarly, the import of live aquatic animals is only authorised under a permit. As with the release of such organisms into the wild, the Director of Fisheries is bound by law to refuse import permits in respect of all species, other than those which are indigenous to the Territory and those expressly listed by Regulations as authorised imports.
In another Australian jurisdiction, Western Australia, the Agriculture and Related Resources Protection Act 1976 is entirely devoted to the problem of introductions. Animals are classified in several categories. Those listed in Category Al may not be brought into the State at all, except under a permit granted for scientific or educational purposes. Animals listed in Category A4 may only be imported under a permit. Those listed in Category A3 cannot be kept in captivity and those which belong to Category A6 can only be kept subject to specific conditions. Animals brought into the State or kept in contravention of these provisions may be seized and destroyed. Similar provisions apply to plants.
3. Accidental Imports
Apart from the usual phytosanitary and quarantine regulations few laws lay down specific rules for the prevention of accidental introductions. One exception is the law of Western Australia, which requires that inspections be carried out on all wool, hay, chaff, meal, grain, animal feed preparations, agricultural or earthmoving machines and empty used sacks introduced into the State, as a means to prevent the inadvertent introduction of seeds.
In 1990, following the introduction of the zebra mussel (Dreissena polymorpha) into the Great Lakes, presumably through ballast waters, the United States adopted the Non-Indigenous Aquatic Nuisance Prevention and Control Act which deals essentially with unintentional introductions. The Act establishes a national ballast water control programme and calls for the development of a programme for the prevention of the introduction of aquatic species and for the monitoring and control of species which have already been introduced.
Π. Discussion and proposals
Harmful introductions can be considered as a self-regenerating form of pollution which may have irreversible effects on natural ecosystems and other species. These effects may be impossible or extremely costly to control once the introduced species has become established. Prevention is consequently of the utmost importance.
Any legislation should be based on the precautionary principle which is gradually becoming an accepted basis for the development of environmental law.
This means that the rule for voluntary introductions should be that permits are in principle denied, unless it can be shown that there are at least good scientific reasons to believe that the proposed introduction will be harmless. The ICES and EBFAC Codes of Pratice should be given legal force and become binding upon permit-issuing authorities. This could be achieved within the European Community by the adoption of a Regulation. For those States

- 9 0 -
which are not members of the Community, it would be necessary to use other instruments, such as the Barcelona and Helsinki Conventions for the marine environment and the Berne Convention for terrestrial and freshwater species.
To prevent escapes from captivity or fraudulent releases, the implementation of the precautionary principle requires strict controls on imports of live specimens of those species which may survive in the European environment. Within the European Community, this would also require a Regulation. It might also be necessary to regulate intra-Community trade, although this could create difficulties as such restrictions would run counter to the basic objectives of the EC. Import controls should be accompanied by prohibitions or restrictions on the possession, sale and transport of exotic species.
Purely accidental introductions will be difficult to prevent, as effective precautionary measures may prove unacceptable to public opinion if they restrict the freedom of movement of persons and goods. Nonetheless, an effort should be made to identify the major pathways for such introductions. Ballast waters may be one route, but there are probably many more. Once such a pathway has been identified with certainty, control measures adapted to that specific case will be easier to develop and enforce.
The damage caused by the zebra mussel in the Great Lakes has been estimated at $ 5,000 million by the year 2000. This amply justifies the taking of regulatory measures which would probably have been impossible prior to that disastrous introduction. In any event, special precautions need to be taken to avoid accidental introductions into particularly sensitive areas, such as islands.
All such legislation should empower enforcement personnel to inspect premises and to seize and destroy specimens of exotic species which are illegally imported or possessed. This is hardly ever the case at present, although Western Australia is once again a notable exception.
Finally, it is also important to provide a clear definition of exotic species, which is not always the case with existing legislation. Moreover, efforts should be made to arrive at a common definition at international level.
All these rules should be applicable to all species, whether terrestrial, freshwater or marine. As matters stand, where national legislation on introductions does exist, it almost always applies only to terrestrial or freshwater species and marine species are therefore completely ignored. The reason for this is presumably that the national conservation authorities, which are generally aware of the risks involved as the result of introductions, usually have no jurisdiction over the marine environment. Fisheries departments are often more concerned by the development and regulation of fisheries than by the conservation of natural habitats and species of no economic value.
Where prevention has failed and a species thus introduced has become established, there is clearly a need to try to eradicate it as quickly as possible before it is too late. Once again, most laws are silent on this matter. The law of Western Australia outlines detailed rules relating to the control of "declared" animals and plants in public, municipal and private land.
A further legal issue relates to the criminal and civil liability of the author of the introduction, where he or she can be identified. Penalties in this field are generally very low, as legislators do not seem to consider that illegal introductions are serious offences.

-91 -
In addition, there is generally no provision for the recovery from the offender of civil damages to repair the harm caused to the environment and to compensate for the cost of eradication or control measures. One exception is the law of the northern Territory of Australia, which empowers the Court to order the offender, upon conviction, to pay any amount that it thinks fit the respect of any control measures which have been taken or any other loss damages.
Consideration should also be given to problems of international responsability and liability, where a species introduced into one country spreads to areas under the jurisdiction or control of another State. This problem may be particularly serious in the marine environment.
It is now an accepted principle of international law that States have the responsability to ensure that activities within their jurisdiction or control do not cause damage to the environment of other States or areas beyond the limits of national jurisdiction (Principle 21 of the Stockholm Declaration of 1972).
Under the Law of the Sea Convention of 1982, "States are responsible for the fulfilment of their international obligations concerning the protection and preservation of the marine environment" and "liable in accordance with international law" (article 235 (1)). As one of these obligations relates to the prevention, reduction and control of the introduction, whether intentional or accidental, of alien species, it follows that a State which has carried out an introduction or failed to take measures to control an introduction which has caused damage to another State may be required to compensate the victims of the damage.
The matter of liability is also dealt with in the new Convention on Biological Diversity which mandates the Conference of the Parties established thereunder to
'examine, on the basis of studies to be earned out, the issue of the liability and redress, including restoration and compensation, for damage to biological diversity, except where such liability is a purely internal matter" (article 12 (2)).
Compensation would only be payable, however, where prevention and early eradication had failed. It is therefore all the more important to develop international cooperation to control harmful introductions. This could be achieved on the basis of article 14 (1) of the new Convention on Biological Diversity, which requires its Parties "as far as possible and as appropriate" to take certain steps to minimise the adverse impacts of their actions on biological diversity, especially where these may affect other States or areas beyond the limits of national jurisdiction.
These measures include the making of environmental impact assessments and the introduction of "appropriate arrangements" to ensure that the environmental consequences of national programmes and policies that are likely to have significant adverse impacts on biological diversity are duly taken into account. These provisions clearly apply at both national and international level. Other provisions in the same article concern international cooperation where there is a risk of damage to the environment of another country or to areas beyond the limits of national jurisdiction. These seem to be particularly relevant to the issue of introductions.
Firstly, States must promote "notification, exchange of information, and consultations on activities under their jurisdiction or control which are likely to significantly affect adversely the biological diversity of other States" or of the high sea. This should be done by

- 9 2 -
"encouraging the conclusion of bilateral, regional or multilateral arragements, as appropriate".
Secondly, "in the case of imminent or grave danger or damage originating under its jurisdiction or control" to biological diversity in other States or in the high sea, a State must immediately notify the potentially affected States of such danger or damage and must initiate action to prevent or minimise such danger or damage.
Thirdly, States must
"promote national arrangements for emergency responses to activities or events which present a grave and imminent danger to biological diversity and encourage international cooperation to supplement such national efforts and, where appropriate and agreed by the States ... concerned, establish joint contingency plans"
With regard to the specific matter of introductions into the marine environment, these provisions should be interpreted as meaning that neighbouring States should be consulted on any proposed voluntary introductions, and notified immediately of any accidental introduction which would be likely to spread to areas under their jurisdiction. In addition, emergency procedures and contingency plans should be developed to control potentially harmful introductions before they have had time to spread widely. As in the case of oil spills, this may require special forms of international cooperation and assistance, if irreversible damage to species and ecosystems is to be avoided.
The legal basis for such cooperation is already provided for by various regional international instruments, including the Berne, Barcelona and Helsinki Conventions and the EC Habitats Directive, and there would seem to be no need to develop new instruments for that purpose, except perhaps for the North Sea.
However, there is a need to provide scientific advice to the regional bodies concerned. This could be the task of CD2SM for the Mediterranean and the Black Sea or of ICES for the other European seas.

93
BIBLIOGRAPHIC REFERENCES
ACHITUV, Y., 1973. - On the distribution and variability of the Indo-Pacific sea star Asterina wega (Echinodermata : Asteroidea) in the Meditenanean Sea. Mar. Biol., 18 : 333-336.
ΑΚΓΥΑΜΑ, K., 1965. - Studies of ecology and culture of Undaria pinnatifida (Harv.) Sur. Π. Environmental factors affecting the growth and maturation of gametophyte. Bull. Tohoku reg. Fish. Res. Lab., 25 : 143-170.
ALEEM, A.A., 1948. - The recent migration of certain Indo-Pacific algae from the Red Sea into the Mediterranean. New Phytol., 47 : 88-94.
ALEEM, G., 1950. - Some new records of marine algae from the Mediterranean sea, with reference to their geographical distribution. Med. Bot. Sverige, 18 : 275-288.
ALEEM, A.A., 1962. - The occurence of the sea-grass : Halophila stipulacea (Forsk.) Asch. on the west coast of Egypt. Bull. Fac. Sci. Univ. Alexandria, 4 : 79-84.
ANON., 1972. - Report of the Working Group on Introduction of Non-indigenous Marine Organisms. - Coop. Res. Rep., ICES, 32 : 1-59.
ANON., 1984. - Guidelines for implementing the ICES Code of Practice concerning introductions and transfers of marine organisms. Coop. Res. Rep., ICES, 130 : 1-20.
ANON., 1982. - Status (1980) of introductions of non-indigenous marine species to North Atlantic waters. - Coop. Res. Rep., ICES, 116 : 1-87.
AMBROSE, R.F., NELSON, B.V., 1982. - Inhibition of giant kelp recruitment by an introduced brown alga. Botanica marina, 25 : 265-267.
ARIANI, A.P. & S ARRA, V., 1969. - Sulla presenza del Portunus pelagicus (L.) in aque italiane, con osservazioni sulla morfologia della specie (Crustacea Decapoda). Arch. Botan. Biogeogr. hal., 45 (4) : 186-206.
BARASH, Α. & DANIN, Ζ., 1989. - Marine Mollusca at Rhodes. Isr. J. Zool., 35 : 1-74.
BALLESTEROS, E., 1990. - Check list of benthic marine algae from Catalonia (North-Western Mediterranean). Treb. Inst. bot. Barcelona, 13 : 1-52.
BARRETT, S.C.H. & RICHARDSON, B.J., 1986. - Genetic attributes of invading species. In : R.K. Groves & J.J. Burdon (Eds.) Ecology of biological invasions : an Australian perspective. Australian Academy of Science, Canberra : 21-33.

94
BARBAROUX, O., PEREZ, R., DRENO, J.P., 1984. - L'algue rouge Eucheuma spinosum. Possibilités d'exploitation et de culture aux Antilles. Sci. Pêche, Bull. Inst. Pêches marit., 348 : 2-9.
BATTAGLIA, Β., 1967. - Genetic aspects of benthic ecology in brackish waters. In : Lauff G.H. (Ed.) Estuaries. AAAS, 83: 574-577.
BATTAGLIA, Β. & BEARDMORE, J.Α., 1979 - Genetics, Ecology and Evolution. Plenum Press, London.
BAZZOCCHI, P., 1985. - Prima segnalazione di Strombus (Conomurex) decorus raybaudii Nicolay and Romagna Manoja, 1983 per l'Isola di Cipro. Boll. Malacologico, 21 (1-4) : 64.
BERMAN, J. & CARLTON, J.T., 1991. - Marine invasion processes : interactions between native and introduced marsh snails. J. Exp. Mar. Biol. Ecol., 150 : 267-281.
BELSHER, T., 1991. - Sargassum muticum (Yendo) Fensholt sur le littoral français. Synthèse des études 1983-1989. Rapport IFREMER, Brest : 1-99.
BELSHER, T., BOYEN, C , 1983. - Sargassum muticum (Yendo) Fensholt : une sargasse parmi d'autres. Biomasse Actualité, 12 (suppl.) : 13.
BELSHER, T., BAILLY DE BOIS, P. & SALOU, N., 1984. - Expansion de l'algue d'origine japonaise, Sargassum muticum (Yendo) Fensholt, sur les côtes françaises, de 1983 à 1984. Cah. Biol. mar., 25 (4) : 449-455.
BELSHER, T., POMMELEC, S., 1988. - Expansion de l'algue d'origine japonaise Sargassum muticum (Yendo) Fensholt sur les côtes françaises, de 1983 à 1987. Cah. Biol. mar., 29 (2) : 221-231.
BEN MAÍZ, Ν., BOUDOURESQUE, CF. , RIOUALL, R., LAURET, M., 1987. - Flore algale de l'étang de Thau (France, Méditerranée) : sur la présence d'une Rhodyméniale d'origine japonaise, Chrysymenia wrightii (Rhodophyta). Botanica marina, 30 : 357-364.
BEN TUVIA, Α., 1973. - Man-made changes in the Eastern Mediterranean Sea and their effect on the fishery resources. Mar. Biol. 19 : 197-203.
BEN TUVIA, Α., 1985. - The impact of the Lessepsian (Suez Canal) Fish Migration on the Eastern Mediterranean Ecosystem. In : Moraitou-Apostolopoulou and V. Kiortsis (eds.). Mediterranean Marine Ecosystems, Plenum Press, New-York : 367-375.
BEN YAMI, M. & GLASER, T., 1974. - The invasion of Saurida undosquamis (Richardson) into the Levant Basin. An example of biological effect of interoceanic canals. Fishery Bull., 72 : 359-373.
BILIOTTI, M., ABDELAHAD, N., 1990. - Halophila stipulacea (Forsk.) Ascher. (Hydro-charitaceae) : espèce nouvelle pour l'Italie. Posidonia Newsletter, 3 (2) : 23-26.
BLACKLER, H., 1963. - Some observations on the genus Colpomenia (Endlicher) Derbes et Solier 1851. Proc. internation. Seaweed Symp., 4 : 50-54.

9 5
BOALCH, G.T., 1974. - The type material of the diatom genus Bacteriastrum Shadbolt. Nova Hedwigia, Beih. 45 : 159-163.
BOALCH, G.T., 1979. - The dinoflagellate bloom on the coast of south-west England, August-September 1978. Journal of the Marine Biological Association of the U.K., 59 : 515-528.
BOALCH G.T., 1981. - Do we really need to grow Macrocystis in Europe ? Proc 10th internation. Seaweed Symp., T. Levring edit., Walter de Gruyter & co pubi., Berlin : 657-667.
BOALCH, G.T., 1987. - Changes in the phytoplankton of the Western English Channel in recent years. British Phycological Journal, 22 : 225-235.
BOALCH, G.T. & POWELL, H.T., 1975. - Introduction of giant kelp proposed for European waters. IUCN Bulletin, 6 (2) :5-6.
BOALCH, G.T. & HARBOUR, D.S., 1977a. - Observations on the structure of a planktonic Pleurosigma. Nova Hedwigia, Beih. 54 : 275-280.
BOALCH, G.T. & HARBOUR, D.S., 1977b. - Unusual diatom off the coast of south-west England and its effect on fishing. Nature, Lond., 269 : 687-688.
BOCQUET C , 1974. - Le polychromatisme des invertébrés marins : approche qualitative des problèmes. Mém. Soc. Zool. France, 37 : 15-40.
BOILLOT, Α., CARAM, B. et METNESZ, Α., 1982. - Sur l'Acrothamnion preissii, Rhodophycée nouvelle pour la flore française. Cryptog.-Algol., 3 (1) : 21-24.
BOLTON, J., GERMANN, J., LUNING, Κ., 1983. - Hybridization between Atlantic and Pacific representatives of the Simplices section of Laminaria (Phaeophyceae). Phycologia, 22 : 133-140.
BOUDOURESQUE, CF. , GERBAL, M. et KNOEPFFLER-PEGUY, M., 1985. -L'algue japonaise Undaria pinnatifida (Phaeophyceae, Laminariales) en Méditerranée. Phycologia, 24 : 364-366.
BOUDOURESQUE, CF. , BALLESTEROS, E., BEN MAÍZ, Ν., BOISSET, F., BOULADIER, E., CINELLI, F., CIRIK, S., CORMACI, M., JEUDY DE GRISSAC, Α., LABOREL, L, LANFRANCO, E., LUNDBERG, Β., MAYHOUB, M., MEINESZ, Α., PANAYOTIDIS, P., SEMROUD, R., SINNASSAMY, J.M., SPAN, Α., VUIGNIER, G., 1990. - Livre rouge "Gérard Vuignier" des végétaux, peuplements et paysages menacés de Méditerranée. Programme des Nations Unies pour l'Environnement pubi., Athènes, Grèce : 1-205.
BOUDOURESQUE, CF. , GOMEZ, Α., 1992. - El alga tropical Caulerpa taxifolia (Chlorophyta) en el Mediterraneo occidental. Algas, Espagne, 10 : 3-8.

- 9 6 -
BOUDOURESQUE, CF. , MEINESZ, Α., VERLAQUE, M., KNOEPFFLER-PEGUY, M., 1991. - The expansion of the tropical alga Caulerpa taxifolia (Chlorophyta) in the Mediterranean. Cryptog.-Algol., 13 (2) : 144-145.
CAMPOS, N.H. & TURKAY, M., 1989. - On a record of Charybdis hellen from the Caribbean Coast of Colombia. Senckenbergiana marit., 20 (3/4) : 119-123.
CARLTON, J.T., 1985. - Transoceanic and interoceanic dispersal of coastal marine organisms ; the biology of ballast water. Oceanogr. mar. Biol. arm. Rev., 23 : 313-373.
CARLTON, J.T., 1987. - Patterns of transoceanic marine biological invasions in the Pacific ocean. Bull. mar. Sci., 41 (2): 442-465.
CARLTON, J.T., 1989. - Man's role in changing the face of the ocean : biological invasions and implications for conservation of near-shore environments. Conserv. Biol., 3 (3) : 265-273.
CARLTON J.T., 1992α. - An international perspective on species introductions : The ICES protocol. In : DeVoe, M.R. (ed.) Introductions and transfers of marine species. Achieving a balance between economic development and resource protection. S. C. Sea Grant Consortium, pp. 31-33.
CARLTON J.T., 1992b. - Dispersal of living organisms into aquatic ecosystems as mediated by aquaculture and fisheries activities. In : Rosenfield, A. & R. Mann (eds). Dispersal of living organisms into ecosystems. Maryland Sea Grant Publications. College Park, Maryland, pp. 13-45.
CARLTON, J.T. 1992c. - Marine species introductions by ships' ballast water : An overview. In : DeVoe, M.R. (ed.) Introductions and transfers of marine species. Achieving a balance between economic development and resource protection. S. C. Sea Grant Consortium, pp. 23-25.
CARLTON J.T., 1993 - Dispersal mechanisms of the Zebra Mussel (Dreissena polymorpha). In : Zebra Mussels : Biology, impacts and control. Edited by Thomas F., Nalepa and Donald W. Schloesser. Lewis Pubi., Boca Raton, Florida, 810 pp.
CARLTON, J.T., CHENEY, D.P., VERMEIT, 1982. - Ecological effects and biogeography of an introduced marine species : the periwinckle, Littorina littorea. Malac. Rev., 15 (1-2) : 143-150.
CESARI, P., PELLIZZATO, M., 1985. - Molluschi pervenuti in laguna di Venezia per apporti volontari o casuali, acclimazione di Saccostrea commercialis (Iredale & Roughley, 1933) e di Tapes philippinarum (Adams & Reeve, 1950). Boll, malacologico, 21 (10-12) : 237-274.
CHAKROUN, F., 1986. - Captures d'animaux rares en Tunisie. Bull. Inst. Oceanogr. Pêche, Salammgo, 1 : 75-79.
CINELLI, F., SALGHETTI-DRIOLI, et SERENA, F., 1984. - Nota sull'areale di Acrothamnion preissii (Sonder) Wollaston nell'Alto Tirreno. Quad. Mus. Stor. nat. Livorno, 5 : 57-60.

9 7
CINELLI, F. et SARTONI, G., 1969. - Acrothamnion J.Ag. (Rhodophyta, Ceramiaceae) : genere algale nuovo per il mare mediterraneo. Pubbl. Staz. zool. Napoli, 37 : 567-574.
COGNETTI, G., 1992. - Colonization of stressed coastal environments. Mar. Poll. Bull., 24 : 12-24.
COMPS, M., BONAMI, J.R., VAGO, C , CAMPILLO, Α., 1976. - Une virose de l'huître portugaise (Crassostrea ungulata). CR. hebd. Séances Acad. Sci. Paris, 282 (D) : 1991-1993.
CORMACI, M., DURO, A. et FURNARI, G., 1982. - Considerazioni sugli elementi fitogeografici della flora algale della Sicilia. Nat. sicil., 4, 6 (suppl.) : 7-14.
CORMACI, M., FURNARI, G., ALONGI, G., DINARO, R. et PIZZUTO, F., 1992. - On the occurrence in Sicily of three Florideophyceae new to the Mediterranean Sea. Botanica marina, 35 : 447-449.
CORMACI, M., FURNAR.I G. et SERIO, D., 1991. - First record of the austral species Plocamium secundatum (Gigartinales, Rhodophyta) from the Mediterranean sea. Cryptog.-Algol., 12 (4) : 235-244.
COTTON, A.D., 1908. - Colpomenia sinuosa in Britain. Journal of Botany, Lond., 46 : 82-83.
CRAWLEY, M.J., 1986. - The population biology of invaders. Phil. Trans. R. Soc. Lond.,B3U : 711-731.
CRAWLEY, M.J., 1987. - What makes a community invasible ? In : A.J. Gray, M.J. Crawley & P.J. Edwards (Eds.) Colonization, succession and stability. Blackwell Sci. Pubi., Oxford, 429-453.
CRISP, D.J., 1958. - The spread of Elminius modestus Darwin in north-west Europe. Journal of the Marine Biological Association of the U.K., 37 : 483-520.
CRISP, D.J. & CHIPPERFIELD, D.N.J., 1948. - Occurrence of Elminius modestus (Darwin) in British waters. Nature, Lond., 161 : 64.
CRISP, D.J. & MOLESWORTH, W.H., 1950. - Habitat of Balanus amphitrite var. denticulata in Britain. Nature, Lond., 167 : 489-490.
CRITCHLEY, A.T., 1983. - The establishment and increase of Sargassum muticum (Yendo) Fensholt populations within the Solent area of southern Britain. Botanica marina, 26 (12) : 547-552.
CRITCHLEY, A.T., FARNHAM, W.F. & MORRELL, S.L., 1983. - A chronology of new European sites of attachment for the invasive brown alga Sargassum muticum. Journal of the Marine Biological Association of the U.K., 63 : 799-789.
CRITCHLEY, A.T., FARNHAM, W.F. & MORRELL, S.L., 1986. - An account of the attempted control of an introduced marine alga, Sargassum muticum, in southern England. Biol. Conserv. 35 : 313-332.

- 98-
CRITCHLEY, A.T., FARNHAM, W.F., YOSHIDA, T. & NORTON, T.A., 1990. -Bibliography of the invasive alga Sargassum mutìcum (Yendo) Fensholt (Fucales ; Sargassaceae). Bot. mar., 33 : 551-562.
DEVALERA, M., 1942. - A red alga new to Ireland : Asparagopsis armata Harv. on the west coast. Irish Naturalist Journal, 8:30.
DE WREDDE, R.E., 1983. - Sargassum mutìcum (Fucales, Phaeophyta) : regrowth and interaction with Rhodomela larix (Ceramiales, Rhodophyta). Phycologia, 22 (2) : 153-160.
DIAPOULIS, Α., VERLAQUE, M., HUVE, P. et BOUDOURESQUE, CF. , 1985. -Sur la présence d'une espèce dugenre Sarconema (Solieriaceae, Rhodophyta) en Grèce. Vie Milieu, 35 (1) : 57-59.
DIXON, P.S., 1961. - List of marine algae collected in the Channel Islands. British Phycological Bulletin, 2 : 71-80.
DODD, B.S., 1894. Proceedings of the Malacological Society, 1 : 31-32.
DRUEHL, L.D., 1973. - Marine transplantations. Science, N.Y. 179, 12.
ELTON, CS. , 1958. - The ecology of invasions of animals and plants. Methuen, London.
ESSINK, K., 1986. - Note on the distribution of the American jack-knife clam Ensis directus (Conrad, 1843) in N.W. Europe (Bivalvia, Cultellidae). Basteria, 50 (1-3) : 33-34.
ESSINK, K., 1988. Recent introductions of North American marine invertebrates in the North Sea and Baltic Sea. Notitie GWAO-88.2240. Rijkswaterstaat, The Netherlands. 8 pp.
ESSINK K., KLEEF, H.L., 1988. - Marenzelleria viridis (Verrill, 1873) (Polychaeta : Spionidae) : a new record from the Ems estuary (The Netherlands/Federal Republic of Germany). Zool. Bijdrag., Pays-Bas, 38 : 1-13.
ESSINK, K., KLEEF, H.L., 1992. - Distribution and life cycle of the north american spionid polychaete Marenzelleria viridis (Verrill, 1873) in the Ems estuary. J. aquatic Ecol, 26.
FARNHAM, W.F., 1980. - Studies on aliens in the marine flora of southern England. In : The shore environment. Volume 2 : Ecosystems. J.H. Price, D.E.G. Irvine & W.F. Farnham eds., Academic Press pubi., London, pp. 875-914.
FARNHAM, W.F., FLETCHER, R.L. & IRVINE, L.M., 1973. - Attached Sargassum muticum found in Britain. Nature, Lond., 243 : 231-232.
FARNHAM, W.F. & JEPHSON, N.A., 1977. - A survey of the maerl beds of Falmouth (Cornwall). British Phycological Journal, 12 :119.
FARNHAM, W.F. & IRVINE, L.M., 1979. - Discovery of members of the red algal family Solieriaceae in the British Isles. British Phycological Journal, 14 :123.

9 9
FAVA, G. & FONS ATTI, E., 1992. - Allozyme variability in the clam Ruditapes philippinarum from two stocks cultivated in different Adriatic lagoons. Rapp. Comm. int. Mer Médit., 33 : 360.
FELDMANN, J., 1931. - Note sur quelques algues marines de Tunisie. Stat. océanogr. Salammgo, Notes, 24 : 1-20
FELDMANN, J., 1938. - Recherches sur la végétation marine de la Méditerranée : la côte des Albères. Rev. algol, 10 (1-4) : 1-340.
FELDMANN, J., 1939. - Les algues marines de la côte des Albères. IV : Rhodophycées (Bangiales, Némalionales, Cryptonémiales). Rev. algol., 11 (3-4) : 247-330.
FELDMANN, J., 1946. - La flore marine des îles atlantides. Mém. Soc. Biogéogr., 8 : 395-435.
FELDMANN, J., 1956. - Sur la parthénogenèse du Codium fragile (Sur.) Hariot dans la Méditerranée. CR. hebd. Séances Acad. Sci., 243 : 305-307.
FELDMANN, J., 1968. - Origines et affinités du peuplement végétal benthique de la Méditerranée. Rapp. P.V. Réun. Commiss, intemation. Explor. sci. Médit., 14 : 515-518.
FELDMANN, J. et FELDMANN, G., 1938. - Présence de Lophocladia lallemandi (Mont.) Schmitz, aux environs d'Alger. Bull. Soc. Hist. nat. Afrique Nord, 29 : 479.
FISCHER-PIETTE, E., 1959. - Contribution à l'écologie intercotidale du Détroit de Gibraltar. Bull. Inst. océanogr., 56 (1145) : 1-32.
FISCHER-PIETTE, E., 1963. - Les progrès de Fucus spiralis combleront-ils la curieuse lacune de l'Algarve ? Bull. Inst. océanogr., 60 (1264) : 1-15.
FLETCHER, R.L., FLETCHER, S.M., 1975. - Studies on the recently introduced brown alga Sargassum muticum (Yendo) Fensholt. I. Ecology and reproduction. Botanica marina, 18 : 149-156.
FLOC'H, J.Y., PAJOT, R., WALLENITNUS, I., 1991. - The Japanese brown alga Undaria pinnatifida on the coast of France and its possible establishment in European waters. J. Cons, internation. Explor. Mer, 47 : 379-390.
FORSTER, G.R., 1979. - Mortality of the bottom fauna and fish in St Austell Bay and neighbouring areas. Journal of the Marine Biological Association of the U.K., 59 : 517-520.
FOSTER, B.A., WILLAN, R.C., 1979. - Foreign barnacles transported to New Zealand on an oil platform. N. Z. J. mar. freshw. Res., 13 (1) : 143-149.
FORTI, Α., 1928. - Nuove segnalazioni di alghe passivamente transportate a traverso al canale di Suez, poi naturalizzate. Nuovo G. bot. ital., N.S., 34 (2) : 1443-1451.
FORTI, Α., 1931. - Description de plusieurs formes de Fucus virsoides de l'Adriatique. Travaux cryptogamiques dédiés à Louis Manguin : 177-188 + 7 pi.

- 100 -
FOX, M.D. & FOX, B.J., 1986. - The susceptibility of natural communities to invasion. In : R.H. Groves & J.J. Burdon (Eds.). Ecology of biological invasions : an Australian perspective. Australian Academy of Science, Canberra : 57-66.
FRITSCH, C , 1895. - Ueber die Auffindung einer marinen Hydrocharidae im Mittelmeer. Verh. Zool.-Bot. Ges. Wien, 45 : 104-106.
GALIL, B.S., 1986. - Red Sea Decapods along the Mediterranean Coast of Israel, Ecology and distribution. In : Z. Dubinsky and Y. Steinberger (eds.) Environmental Quality and Ecosystem Stability, Vol. TWA., Bar Uan Univ. Press : 179-183.
GALIL, B.S., 1989. - Bathymétrie distribution and habitat preferences of Lessepsian migrant Decapoda along the Mediterranean coast of Israel, or : do decapods have cold feet ? In : E. Spanier, Y. Steinberger and M. Luna (eds.). Environmental Quality and Ecosystem Stability, Vol. IV/B. ISEEQS, Jerusalem : 147-153.
GALIL, B.S. & LEWINSOHN C , 1981. - Macrobenthic Communities of the Eastern Mediterranean Continental Shelf. PSZNI Mar. Ecol. 2 : 343-352.
GALIL B.S., SPANIER, E. & FERGUSON, W.W., 1990. - The Scyphomedusae of the Mediterranean coast of Israel, including two Lessepsian migrants new to the Mediterranean. Zool. Medea., 64 : 95-105.
GELDIAY, A. & KOCATAS, Α., 1972. - A report on the occurence of Penaeidae (Decapoda Crustacea) along the coast of Turkey from Eastern Mediterranean to the vicinity of Izmir, as a result of migration and its factors. 17 Congr. Int. Zool. (Monte-Carlo) 7 p.
GERBAL, M., BEN MAIZ, N., BOUDOURESQUE, CF. , 1985. - Les peuplements à Sargassum muticum de l'étang de Thau : données préliminaires sur la flore algale. Congr. nation. Soc. sav., 110 (2) : 241-254.
GELAT, E., 1969. - The macrobentic communities of the level bottom in the Eastern Mediterranean. In : The Hebrew University-Smithsonian Institution Joint Program : Biota of the Red Sea and the Eastern Mediterranean. Interim Report : 82-89.
GRAY, A.J., 1986. - Do invading species have definable genetic characteristics? Phil. Trans. R. Soc. Lond., B314 : 655-674.
GRAY, P.W.G. et JONES, E.B.G., 1977. - The attempted clearance of Sargassum muticum from Britain. Environm. Conserv., 4 : 303-308.
GRIFFITHS, A.B., DENNIS, R. & POTTS, G.W., 1979. - Mortality associated with a phytoplankton bloom off Penzance in Mounts Bay. Journal of the Marine Biological Association of the U.K., 59 : 520-521.
GRIZEL, H., 1983. - Introduction of the cupped oyster Crassostrea gigas in France. Report of the working group on introductions and transfers of marine organisms. International Council Exploration Sea, F27 : 37-39.
GRIZEL, H., HERAL, M., 1991. - Introduction into France of the Japanese oyster (Crassostrea gigas). J. Cons, internation. Explor. Mer, 47(3) : 399-403.

-101 -
GRUET, Y., HERAL, M., ROBERT, J.M., 1976. - Premières observations sur l'introduction de la faune associée au naissain d'huîtres japonaises Crassostrea gigas (Thunberg), importé sur la côte atlantique française. Cah. Biol. mar., 17 : 173-184.
GRUSZKA, P., 1991. Marenzelleria viridis (Verrill, 1873) (Polychaeta: Spionidae) - a new component of shallow water benthic community in the Southern Baltic. Acta Ichthyol. Pişcat. 21 SuppL, 57-65.
HALLEGRAEFF G.M., 1993. - A review of harmful algal blooms and their apparent global increase. Phycologia, 32 : 79-99.
HALLEGRAEFF G.M., BOLCH C.J., 1991. - Transport of toxic dinoflagellate cysts via ships' ballast water. Mar. Poll. Bull., 22 : 27-30.
HALLEGRAEFF G.M., BOLCH C.J., BRYAN J., KOERBIN B., 1990. - Microalgal spores in ships' ballast water : A danger to aquaculture. - In : Granali, Ε., Β. Sundström, L. Edler & D.M. Anderson (eds) : Toxic marine phytoplankton. Proc. Fourth Intern. Conf. Toxic Marine Phytoplankton, June 26-30, 1989, Lund, Sweden. Elsevier, New York, pp. 475-480.
HAMEL, G., 1926. - Quelques algues rares ou nouvelles pour la flore méditerranéenne. Bull. Mus. nation. Hist. nat., 32 (6) : 420.
HAMEL, G., 1930a. - Floridées de France VI. Rev. algol, 5 (1) : 61-109.
HAMEL, G., 1930b. - Les Caulerpes méditerranéennes. Rev. algol., 5 : 229.
HANSEN, V.K. & S ARMA, A.H. V., 1969. - On a Gymnodinium red water in the eastern North Sea during autumn 1968 and accompanying fish mortality with notes on océanographie conditions. ICES (Copenhagen) CM 21.
HARTOG, C. den, 1972. - Range extension of Halophila stipulacea (Hydrocharitaceae) in the Mediterranean. Blumea, 20 : 154.
HOEK, C. Van den, BREEMAN, A.M., STAM, W.T., 1990. - The geographic distribution of seaweed species in relation to temperature : present and past. Expected effects of climatic change on marine coastal ecosystems, Beukkema et al. edit. : 55-67.
HOLTHUIS, L.B. & GOTTLIEB, E., 1968. - An annotated list of the Decapod Crustacea of the Mediterranean coast of Israel, with an appendix listing the Decapoda of the Eastern Mediterranean. Bull. Res. Coun. Israel, 7B : 1-126.
d'HONDT, J.L., 1984. - Un nouvel immigrant dans le bassin d'Arcachon, Watersipora atterima (Ortmann, 1890) (Bryozoaire Cheilostome). Congr. nation. Soc. sav., 109 (2) : 237-245.
d'HONDT, J. J., OCCHIPINTI-AMBROGI, Α., 1985. - Tricellaria inopinata η. sp., un nouveau Bryozoaire Cheilostome de la faune méditerranéenne. P. S. Ζ. Ν. I Marine Ecology, 6 (1) : 35-46.

- 102 -
HULBURT, E.M., 1957. - The taxonomy of unarmoured Dinophyceae of shallow embayments on Cape Cod, Massachussets. Biological Bulletin, Woods Hole, 112 : 196-219.
HUTCHINGS, P.J., VAN DER VELDE & KEABLE, S.J., 1987. - Guidelines for the conduct of surveys for detecting introductions of non-indigenous marine species by ballast water and other vectors. Occ. Rep. Austr. Mus. N°3.
ICES, 1974. - Report of the Working Group on Introduction of Non-indigenous Marine Organisms, London, UK, 31 January-1 February 1974. - CM. 1974/E : 7.
ICES, 1984. - Report of the Working Group on Introductions and Transfers of Marine Organisms, Halifax, Canada, 17-19 May 1984. - CM. 1984/F : 35.
ICES, 1986. - Report of the Working Group on Introductions and Transfers of Marine Organisms, Gdynia, Poland, June 3-6, 1986. - CM. 1986/F : 51.
ICES, 1987. - Report of the Working Group on Introductions and Transfers of Marine Organisms, Brest, France, June 10-12, 1987. - CM. 1987/F : 35.
ICES, 1989. - Report of the Working Group on Introductions and Transfers of Marine Organisms, Dublin, Ireland, May 23-26, 1989. - CM. 1989/F : 16.
ICES, 1990. - Report of the Working Group on Introductions and Transfers of Marine Organisms, Halifax, Canada, June 6-8, 1990. - CM. 1990/F : 14.
ICES, 1991. - Report of the Working Group on Introductions and Transfers of Marine Organisms, Helsinki, Finland, June 4-7, 1991. - CM. 1991/F : 44.
ICES, 1992. - Report of the Working Group on Introductions and Transfers of Marine Organisms, Lisbon, Portugal, April 14-17, 1992. - CM. 1992/Poll : 3.
IRVINE, J.-Y., PAJOT, R. & WALLENTINUS, I., 1988. - The Japanese brown alga Undaria pinnatiflda on the coasts of France and the possibilities of its establishment in European waters. Journal du Conseil International pour l'Exploration de la Mer, 47 : 379-390.
JANSON, K., 1985. - A morphologie and genetic analysis of Littorina saxatilis (Prosobranchia) from Venice, and on the problem of saxatilis-rudis nomenclature. Biol. J.Linn. Soc, 24: 51-59.
JONES, W.E., 1974. - Changes in the seaweed flora of the British Isles. In : The changing flora and fauna of Britain, ed. D.L. Hawksworth, pp. 97-113. Academic Press, London.
JUPIN, Α., 1989. - Sargassum muticum (Yendo) Fensholt. Propositions de solutions de lutte mécanique. Rapport IFREMER DIT/EQE/89, Brest, 139 : 1-98.
KERAMBRUN, P., 1965. - Contribution à l'étude écologique et génétique de quelques populations de Sphaeroma hookeri Leach du bassin méditerranéen. Rapp. Comm.int. Mer Médit., 18 : 705-708

- 1 0 3 -
KERAMBRUN, P., 1966. - Contribution à l'étude écologique et génétique du polychromatisme de l'Isopode Sphaeroma hookeri Leach dans les eaux saumâtres méditerranéennes. Bull. Inst. océan. Monaco, 66 : 5-52.
KERAMBRUN, P., RAZAKANDISA R.N. & SCHACHTER, D., 1964. - Sur l'écologie de Sphaeroma hookeri Leach de la rivière La Durançole. CR. Acad. Sci. Paris, 259 : 1666-1668.
KIDEYS, A.E. & NIERMANN, Ulrich, 1993. - Intrusion of Mnemiopsis mccradyi (Ctenophora : Lobata) into the Mediterranean Sea. Senckenbergiana mărit., 23 (1/3) : 43-47.
KNOEPFFLER-PEGUY, M., BELSHER, T., BOUDOURESQUE, C F . & LAURET, M., 1985. - Sargassum muticum begins to invade the Mediterranean. Aquatic Botany, 23 : 291-295.
KRAEUTER, J.N., 1976. - Offshore currents, larval transport, and establishment of southern populations of Littorina littorea Linné along the U.S. Atlantic coast. Thalassia jugoslavica, 10 (1-2) : 159-170.
KTARI-CHAKROUN, F. & BOUHLAL, M., 1971. - Capture de Siganus luridus (Ruppell) dans le golfe de Tunis. Bull. Inst. Océanogr. Pêche, Salammgo, 2 : 49-52.
LAMIRAL, L., 1863. - Second rapport sur un essai d'acclimatation des éponges de Syrie dans les eaux françaises de la Méditerranée. Bull. Soc. impér. zool. Acclimat., 10 : 8-15.
LEBOUR, M.V., 1930. - The Planktonic Diatoms of Northern Seas. The Ray Society, London.
LEJUEZ, R., 1974. - Génétique comparée du polychromatisme chez les Sphéromes. Mém. Soc. zool. France, 37 : 41-59.
LEPPÄKOSKI, E., 1984. Introduced species in the Baltic Sea and its coastal ecosystems. Ophelia Suppl. 3, 123-135.
LEPPÄKOSKI, E., 1991. Introduced species - resource or threat in brackish-water seas? Examples from the Baltic and the Black Sea. Mar. Poll. Bull. 23: 219-223.
LEPPÄKOSKI, E., 1993. The Baltic and the Black Sea - seriously contaminated by nonindigenous species? Nonindigenous Estuarine and Marine Organisms (NEMO), Seattle, Washington, April 20-22, 1993 (in print).
LEPPÄKOSKI, E. & BONSDORFF, E., 1989. Ecosystem variability and gradients. -Examples from the Baltic Sea as a background for hazard assessment. In Chemicals in the Aquatic Environment. Advanced hazard assessment (L. Landner, Ed.), pp. 6-58. Springer, Berlin Heidelberg.
LEWINSOHN, C , 1977. - Injuries caused by marine animals. The Family Physician, 7(1-2) : 182-202.

- 104-
LIPKIN, Y., 1972. - Contributions to the knowledge of Suez canal migration. Marine algal and sea-grass flora of the Suez canal. Israel J. Zool., 21 : 405-446.
LUCZAK, C , DEWARUMEZ, J.M., ESSINK, K., 1993. - First record of the American jack knife clam Ensis directus on the French coast of the North Sea. J. mar. biol. Ass. U.K., 73(1): 233-235.
LUNDBERG, Β., LIPKIN, Y., 1992. - Seasonal, grazing site and fish size effects on patterns of algal consumption by the herbivorous fish, Siganus rivulatus, at Mikmoret (Mediterranean, Israel). Environmental quality and ecosystem stability, Gasith A., Adin Α., Steinberg Y., Garry J. edit., ISEEQS pubi., 5 (Β) : 577-583.
LYLE, L., 1922. - Antithamnionella, a new genus of algae. / . Bot. London, 60 : 346-350.
MAYHOUB, 'H., 1976. - Recherches sur la végétation marine de la côte syrienne. Etude expérimentale sur la morphogénèse et le développement de quelques espèces peu connues. Thèse Doct. Rtat Univ. Caen : 1-286 + 1 carte + 16 pi.
MEINESZ, Α., & HESSE, Β., 1991. - Introduction et invasion de l'algue tropicale Caulerpa taxifolia en Méditerranée nord-occidentale. Oceanologica Acta, 14 (4) : 415-426.
MEINESZ, Α., de VAUGELAS, L, BENICHOU, L., CAYE, G., COTTALORDA, J.M., DELAHAYE, L., FEBVRE, M., GARIN, S., KOMATSU, T., LEMEE, R., MARI, X., MOLENAAR, H., PERNEY, L. & VENTURINI, Α., 1993. - Suivi de l'invasion de l'algue tropicale Caulerpa taxifolia en Méditerranée. Situation au 31 décembre 1992. Laboratoire Environnement marin littoral et GIS Posidonie, France : 1-80.
MÉNDEZ DOMINGO, C , 1957. - Sur l'existence du Colpomenia peregrina (Sauv.) Hamel dans la Méditerranée. Vie Milieu, 8 (1) : 92-98.
MILLER, R.L., 1969. - Ascophyllum nodosum : a source of exotic invertebrates introduced into West coast near-shore marine waters. Veliger, 12 (2): 230-231.
MOLLION, J., 1984. - Seaweed cultivation of phycocolloid in the Mediterranean. Hydrobiologia, 116-117 : 288-291.
MOLLISON, D., 1986. - Modelling biological invasion : change explanation, prediction. Phil. Trans. R. Soc. Land., B314 : 675-693.
MOONEY, H. A. & DRAKE, J. Α., 1987. The ecology of biological invasions. Environment 29 (5): 10-37.
MUNRO Α., UTTING S., WALLENTINUS I., in press. 1990. - Status of introductions and transfers in ICES member countries. Coop. Res. Rep., ICES.
NALEPA T.F., SCHLOESSER D.W. (eds), 1992. - Zebra mussels : Biology, impacts, and control. Lewis Pubi. Inc. (CRC Press), Boca Raton.

- 105 -
NEUSHUL, M., 1983. - Morphology, structure, systematics and evolution of the giant kelp, Macrocystis. Proc. joint China-USphycol. Symp., TSENG edit. : 1-27.
NEWSOME, A.E. & NOBLE, LR., 1986. - Ecological and physiological character of invading species. In : R.H. Groves & J.J. Burdon (Eds.) Ecology of biological invasions : an Australian perspective. Australian Academy of Science, Canberra : 1-20.
NICOLAY, K. & ROMAGNA MANOJA, L., 1983. - Strombus (Conomurex) decorus raybaudii n. sp. La Conchiglia, 15 : 176-177 .
NORKKO Α., BONSDORFF, E. & BOSTRÖM, C , 1993. Observations on the polychaete Marenzelleria viridis (Verrill) on a shallow sandy bottom on the south coast of Finland. Mem. Soc. Fauna Flora Fennica (in print).
NORTON, T.A., 1977. - Ecological experiments with Sargassum muticum. J. mar. biol. Ass. U.K., 57 : 33-43.
NYMAN, M., 1993. Introduced species in the Gulf of Bothnia. Åbo Akademi University, Dept. of Biology, M. Sc. Thesis, 41 pp (in Swedish).
OCCHIPINTI-AMBROGI, 1991.- The spread of Tricellaria inopinata into the lagoon of Venice : an ecological hypothesis. In : F.P.Bigey (Ed.) Bryozoaires actuels et fossiles : Bryozoa living and fossil. Bull. Soc. Sci. Nat.Ouest Fr. Mém. H.S.I Nantes : 299-308.
OLIVI, G., 1792. - Zoologia Adriatica. Bassano, 334 p.
OREN, O.H. 1957. - Changes in the temperature of the Eastern Mediterranean Sea in relation to the catch of the Israel trawl fishery during the years 1954/55 and 1955/56. Bull. Inst. Océanogr. Monaco, 1102 :1-12.
ORTON, J.H. & WINCKWORTH, R., 1928. - The occurrence of the American oyster pest Urosalpinx cinerea (Say) on English oyster beds. Nature, Lond., 122 : 241.
OSTENFELD, C.H., 1908. - On the immigration of Biddulphia sinensis Grev. and its occurrence in the North Sea during 1903-1907. Meddelelser fra Kommissionen for Havundersogelser, Plankton 1 Nr 6 : 1-25.
PAOLETTI, Α., SEBASTIO, C , 1973. - Ricerche per eliminare il fouling e sopratutto i serpulidi dalle conduttore di acqua marina prelevata per usi industriali.. Atti Simp, noz. Conserv. Nat., 3 (1) : 73-108.
PAPENFUSS, G.F. et EDELSTEIN, T., 1974. - The morphology and taxonomy of the red alga Sarconema (Gigartinales : Solieriaceae). Phycologia, 13 (1) : 31-43.
PARKE, M., 1948. - Laminaria ochroleuca De la Pylaie growing on the coast of Britain. Nature, 162 : 295.
PARKES, H.M., 1975. - Records of Codium species in Ireland. Proc. R. Ir. Acad., 75 (B) : 125-134.

-106-
PARKE, M. & DEN HARTOG-ADAMS, I., 1965. - Three species of Halosphaera. Journal of the Marine Biological Association of the U.K., 45 : 537-557.
PARKE, M. & DKON, P.S., 1976. - Check-list of British marine algae-third revision. Journal of the Marine Biological Association of the U.K., 56 : 527-594.
PERES, J.M., 1967. - The Mediterranean benthos. Oceanogr. mar. Biol. ann. Rev., Barnes H. edit., 5 : 449-533.
PEREZ, R., 1972. - Opportunité de l'implantation de l'algue Macrocystis pirifera sur les côtes Bretonnes. Science et Pêche, 216 : 1-9
PEREZ, R., LEE, J.Y. & JUGE, C , 1981. - Observations sur la biologie de l'algue japonaise Undaria pinnatiflda (Harvey) Suringer introduite accidentellement dans l'Etang de Thau. Science et Pêche, 316 : 1-12.
PEREZ, R., KAAS, R. & BARBAROUX, O., 1984. - Culture expérimentale de l'algue Undaria pinnatifida sur les côtes de France. Science et Pêche, 343 : 1-15.
PEREZ, R., DURAND, P., KAAS, R., BARBAROUX, O., BARBIER, V., VINOT, C , BOURGEAY-CAUSSE, M., LECLERCQ, M., MOIGNE, J.Y., 1988. - Undaria pinnatifida on the French coasts. Cultivation method ; biochemical composition of the sporophyte and the gametophyte. Algal Biotechnology, Staedler T., Mollion J., Verdus M.C., Karamanos Y., Morvan H., Christiaen D. edit., Elsevier pubi., London : 315-328.
PEREZ, R., KAAS, R., BARBAROUX, O., ARBAULT, S., LE BAYON, N., MOIGNE, J.Y., 1991. - Undaria, une japonaise en Bretagne : nouvelle technique de culture d'une algue alimentaire. Equinoxe, 36 : 19-30.
PETERS, A.F., KAWAI, H., NOVACZEK, I., 1992. - Intraspecific sterility barrier provides evidence that introduction of Sphaerotrichia divaricata (Phaeophyceae, Chordariales) into the Mediterranean was from Japan. Proc. XlVth internation. Seaweed Symp., Brest, Abstract 249 : 115.
PETERSEN, K.S., RASMUSSEN, K.L., HEINEMEIER, J. & RUD, Ν., 1992. Clams before Columbus? Nature 359: 679.
PETERSEN, H.E., 1918. - Algae (excl. calcareous algae). Rep. Danish oceanogr. Expédition 1908-1910 Medit. adj. Seas, 2 (Biology) : 1-20.
POR, F.D., 1978. - Lessepsian migration - the influx of Red Sea biota into the Mediterranean by way of the Suez Canal. Berlin : Springer. VIU + 228 p.
POR, F.D., 1989. - The Legacy of Tethys - An Aquatic Biogeography of the Levant, Kluwer, 214 p.
POR, F.D., 1990. - Lessepsian migrations. An appraisal and new data. Bull. Inst, oceanogr., 7 (numéro spécial) : 1-10.
POSEY, M.H., 1988. - Community changes associated with the spread of an introduced seagrass, Zostera japónica. Ecology, 69 : 974-983.

-107 -
PRUD'HOMME VANREINE, W.F., GIL-RODRIGUEZ, M.C., HAROUN-TABRAUE, R.J., AFONSO-CARRILLO, J., WILDPRET DE LA TORRE, W., 1984. - Polyphysa párvula (Solms-Laubach) Schnetter & Bula Meyer (Dasycladaceae, Chlorophyta) en la región macaronesica. Vieraea, 13 (1-2) : 219-224.
RAHKONEN, R., 1991. Survey of introductions and transfers of marine organisms in Finland. Report of the Working Group on Introductions and Transfers of Marine Organisms, Helsinki, Finland, June 5-7, 1991, C. M. 1991/F:44, 8 pp.
RAMON, E. et FRIEDMANN, L, 1965. - The gametophyte of Padina in the Mediterranean. Proc. internation. Seaweed Symp., 5 : 183-195.
RAPPORT, D. J., REGIER, H.A. & HUTCHINSON, T.C., 1985. Ecosystem behavior under stress. Amer. Naturalist 125, 617-640.
RAYBAUDI-MASSILIA, L., 1983. - A Strombus from the Mediterranean. La Conchiglia, 15 : 174-175.
RAYSS, T., 1941. - Sur les Caulerpes de la côte palestinienne. Palest. J. Bot., 2 : 103-124.
RAYSS, T., 1963. - Sur la présence dans la Méditerranée orientale des algues tropicales de la famille des Soliériacées. Lucr. Gradimi Botan. Acta Bota. Hort. Bucaresti 1961-1962 : 91-106.
REÍD, P.C., SUREY-GENT, S.C., HUNT, H.G. & DURRANT, A.E., 1992. -Thalassiothrix longíssima, a possible oceanic indicator species in the North Sea. International Council for the Exploration of the Sea, Marine Science Symposia, 195 : 268-277.
RIBERA, M.A., GOMEZ-GARRETA, Α., GALLARDO, T., CORMACI, M., FURNAPJ, G. et GIACCONE, G., 1992. - Check list of Mediterranean seaweeds. I. Fucophyceae (Warming, 1884). Botanica marina, 35 : 109-130.
RIBERA-SIGUAN, M.A. et SOTO-MORENO, J., 1992. - Presencia de Anrithamnion algeriense (Rhodophyta, Ceramiales) en las costas españolas. Cryptog.-Algol., 13 (1) : 25-30.
RIOUALL, R., GUIRY, M.D., CODOMIER, L., 1985. - Introduction d'une espèce foliacée de Grateloupia dans la flore marine de l'étang de Thau (Hérault, France). Cryptog. Algol, 6 : 91-98.
ROBBA, E. 1987. - The final occlusion of the Tethys : Its bearing on Mediterranean benthic molluscs. In : K.G. McKenzie (ed.). Shallow Tethys 2, Balkema, Rotterdam, pp. 405-426.
ROBINSON, J., 1989. - Recent finds, Strombus decoruspersicus. Hawaiian Shell News, 37 : 10.
ROS ARAGONÉS, J.D., OLIVELLA, I., PRATS, I., GILÍ, J.M., SARDA, I. (Eds), 1984. - Els sistemes naturals de les liles Medes. Arx Sece. Ciències 73. Institut d'Estudis Catalans. : 828 p.

- 108 -
RUENESS, J., 1989. - Sargassum miUicum and other introduced Japanese macroalgae : biological pollution of European coasts. Mar. Poll. Bull., 20 (4) : 173-176.
RUSSELL, D.J., 1981. - The introduction and establishment of Acanthophora spicifera and Eucheuma striatum in Hawaii, Ph. D. Thesis.
RUSSELL, D.J., 1992. - The ecological invasion of Hawaiian reefs by two marine red algae, Acanthophora spicifera (Vahl) Boerg. and Hypnea musciformis (Wulfen) J. Ag., and their association with two native species, Laurencia nidifica J. Ag. and Hypnea cervicornis J. Ag. ICES mar. Sci. Symp., 194 : 110-125.
RUSSELL, F.S., SOUTHWARD, A.J., BOALCH, G.T. & BUTLER, E.I., 1971. -Changes in biological conditions in the English channel off Plymouth during the last half-century. Nature, Lond., 234 : 468-470.
SACCHI, C F . & OCCHIPINTI-AMBROGI, Α., 1992. - Confinement et biocénoses para-liques. Vie et Milieu, 42 (2) : 207-214.
SACCHI, C F . & TORELLI, Α., 1979. - La variabilité àeLittorina saxatilis (Olivi) dans la lagune de Venise. Rapp. Comm. int. Mer Médit., 25-26 (3) : 125-126.
SAFRIEL, U.N. & RITTE, U., 1983. - Universal correlates of colonizing ability. In : LR. Swingland & P.J. Grennwood (Eds.). The ecology of animal movement. Clarendon Press, Oxford, 215-239.
SANCHOLLE, M., 1988. - Présence de Fucus spiralis (Phaeophyceae) en Méditerranée occidentale. Cryptog. Algol, 9 (2) : 157-161.
SAN MARTIN, G., 1986. - Contribution à l'étude du comportement trophique de Péchinoïde Paracentrotus lividus dans l'étang de Thau (Hérault). Dipl. Etudes approf. Océanol., Univ. Aix-Marseille Π, France : 1-42.
SAUVAGEAU, C , 1906. - A propos du Colpomenia sinuosa signalé dans les huîtrières de la région de Vannes. Bull. Stat. biol. Arcachon, 9 : 1-14.
SAUVAGEAU, C , 1908a. - Sur deux Fucus récoltés à Arcachon (Fucusplatycarpus et F. lutarius). Bull. Stat. biol. Arcachon, 11 : 1-160.
SAUVAGEAU, C , 1908b. - Sur l'apparition, l'envahissement et la disparition du Colpomenia sinuosa. CR. Séances Soc. Biol., 67 : 751-753.
SAUVAGEAU, C , 1912. - Sur l'apparition du Colpomenia sinuosa dans le golfe de Gascogne. CR. Séances Soc. Biol., 72 : 478-480.
SAUVAGEAU, C , 1927. - Sur le Colpomenia sinuosa Derb, et Sol. Bull. Stat. biol. Arcachon, 24 : 309-356.
SCHMIDT, O.C, 1931. - Die marine vegetation der Azoren in ihre grund-zügen dargestellt. Bibliotheca botanica, 102 : 1-116.
SCHUSSNIG, B., 1930. - Acetabularia wettsteinii n.sp., im Mittelmeer. Bot. Z., 5468 : 1068-1071.

-109 -
SILVA, P.C., 1955. - The dichotomous species of Codium in Britain. Journal of the Marine Biological Association of the U.K., 34 : 565-577.
SIMBERLOFF, D., 1981. - Community effects of introduced species. Biotic crises in ecological and evolutionary time, Nitecki M. H. edit., Academic Press, London, pubi. : 53-81.
SINDERMANN, C.J., 1991. - Case histories of effects of transfers and introductions on marine resources. J. Cons, internation. Explor. Mer, 47 : 377-378.
SINDERMANN, C , STEINMETZ, R., HERSHBERGER, W., 1992. - Introduction and transfers of aquatic species. Introduction. ICES mar. Sci. Symp., 194 : 1-2
SINGH, S.M. & GREEN, R.H., 1984. - Excess of allozyme homozygosity in marine Molluscs and its possible biological significance. Malacologia, 25 (2) : 569-581.
SMITH, J.E., 1981. - The natural history and taxonomy of shell variation in the periwinkles Littorina saxatilis and Littorina rudis. J. Mar. Biol. Ass. U.K., 61 : 215-241.
SONNENFELD, P., 1985. - Models of Upper Miocene evaporite genesis in the Mediterranean region. In : D.J. Stanley and F.C. Wezel (eds.), Geological Evolution of the Mediterranean Basin, Springer, pp 323-346.
SORBINI, L., 1988. - Biogeography and climatology of Pliocene and Messinian fossil fish of Eastern Central Italy. Buoi. Mus. civ. St. nat.. Verona, 14 : 1-85.
SOUTHWARD, A.J. & CRISP, D.J., 1963. - Catalogue of Marine Fouling Organisms. 1. Barnacles. Paris, O.E.C.D.
SOUTHWARD, A.J. & SOUTHWARD, E.C., 1977. - Distribution and ecology of the hermit crab, Clibanarius erythropus in the Western Channel. Journal of the Marine Biological Association of the U.K., 57 : 441-452.
SOUTHWARD, A.J. & SOUTHWARD, E.C., 1988. - Disappearance of the warm-water hermit era, Clibanarius erythropus from South-west Britain. Journal of the Marine Biological Association of the U.K., 68 : 409-412.
SPANIER, E., 1989. - Swarming of Jellyfishes along the Mediterranean coast of Israel. Israel J. Zool, 36 : 55-56.
SPANIER, E., PISANTY, S., TOM, M., ALMOG-SHTAYER, G., 1989. - The fish assemblages of the coralligenous shallow shelf of the Mediterranean coast of Israel. J. Fish. Biol, 35 : 641-649.
SPANIER, E., GALIL, B.S., 1991. - Lessepsian migrations : a continuous biogeographical process. Endeavour, 16 (3) : 102-106.
STEVCIC, Z., 1979. - Contribution à la connaissance des crustacés décapodes de Malte. Rapp. Comm. int. mer Médit., 25/26 : 127-128.

- 1 1 0 -
THELIN, I., 1984. - Nouvelle signalisation d'Acrothamnionpreissii (Sonder) Wollaston sur les côtes françaises de la Méditerranée. Trav. sci. Parc nation. Port-Cros, 10 : 171-172.
TOM, M. & GALIL, B.S., 1990. - The Macrobenthic associations of Haifa Bay, Mediterranean coast of Israel. PSZNI Mar. Ecol. 12(1) : 75-86.
TURNER G., (ed.), 1988. - Codes of Practice and manual of procedures for consideration of the introductions and transfers of marine and freshwater organisms. Coop. Res. Rep., ICES, 159 : 1-44.
UTTING S.D., SPENCER B.E., 1992. - Introductions of marine bivalve molluscs into the United Kingdom for commercial culture - case histories. ICES Mar. Sci. Symp.. 194 : 84-91.
VAAS, K.F., 1975. Immigrants among the animals of the delta-area of the SW Netherlands. Hydrobiol. Bull. (Amsterdam) 9, 114-119.
VALET, G., 1969. - Contribution à l'étude des Dasycladales. 2 - Cytologie et reproduction. 3 - Révision systématique. Nova Hedwigia, 17 : 553-644 + pi. 133-162.
VERLAQUE, M., 1981. - Contribution à la flore des algues marines de Méditerranée : espèces nouvelles pour la Méditerranée occidentale. Bot. mar., 24 (11) : 559-568.
VERLAQUE, M., 1989. - Contribution à la flore des algues marines de Méditerranée : espèces rares ou nouvelles pour les côtes françaises. Botanica marina, 32 : 101-113.
VERLAQUE, M., RIOUALL, R., 1989. - Introduction de Polysiphonia nigrescens et d'Antithamnion nipponicum (Rhodophyta, Ceramiales) sur le littoral méditerranéen français. Cryptog. Algol., 10 : 313-323.
VERLAQUE, M. et BOUDOURESQUE, CF. , 1991. - Stypopodium schimpen (Buchinger ex Kützing) Verlaque et Boudouresque comb. nov. (Dictyotales, Fucophyceae), algue de Mer Rouge récemment apparue en Méditerranée. Cryptog.-AlgoL, 12 (3) : 195-211.
VERLAQUE, M. et SERIDI, H., 1991. - Antithamnion algeriensis nov. sp. (Ceramiaceae, Rhodophyta) from Algeria (Mediterranean sea). Bot. mar., 34 (2) : 153-160.
VERHECKEN, Α., 1984. - Strombus decorus raybaudii in the Middellandse Zee. Gloria Maris, 23 (4) : 79-88.
VUCETIC, T., 1983. - Fluctuation in the distribution of the scyphomedusa Pelagio noctiluca (Forskal) in the Adriatic. Oceanol. Acta., 6 : 207-211.
WALFORT, L. & WICKLUND, R., 1973. - Contribution to the worldwide inventory of exotic marine and andromous organisms. FAO Fish. Tech. Pap. 121, 1-49.
WALLENTINUS I., 1992. - Introduced algae in ICES member countries. 14th Intern. Seaweed Symp., Brittany, France, August 1992., p. 130 (abstract).

- 111 -
WALLENTTNUS, I., (sous presse). - Introduction and transfers of plants. Intern. Seaweed Symp., Brest, 1992.
WALLENTTNUS, I., (inpress). - Introductions and transfers. 4 Plants. Coop. Res. Rep.
WESTBROOK, Μ.Α., 1930. - Notes on the distribution of certain marine red algae. J. Bot., London, 68 : 257-264.
WELKOMME, R.L., 1992. - A history of international introductions of inland aquatic species. ICES mar. Sci. Symp., 194 : 3-14.
WILLIAMS, R.J., GRIFFITHS, F.B., VAN DER WAL, E.J., KELLY, J., 1988. -Cargo vessel ballast water as a factor for the transport of non indigenous marine species. Estuar, coast, shelf Sci., 26 (4) : 409-420.
WILLIAMSON, M.H., BROWN, K.C., 1986. - The analysis and modelling of British invasions. Phil. Trans, r. Soc. London, 314 (B) : 505-522.
WILLIAMSON, M.H., KORNBERG, Η., HOLDGATE, M.W., GRAY, A.J., CONWAY, G.R., 1986. - The British contribution to the SCOPE programme on the ecology of biological invasions. Phil. Trans, r. Soc. London, 314 (B) : 503-504.
WOLFF, W.J., 1973. The estuary as a habitat. - Zool. Verh. (Leiden) 126, 1-242.
WOMERSLEY, H.B.S., 1987. - The marine benthic flora of southern Australia. Pt. 2.S. Australian Govt.
ZIBROWIUS, H., 1974. - Oculina patagónica, scléractiniaire hermatypique introduit en Méditerranée. Helgol. Wiss. Meeresunters., 26 (2) : 153-173.
ZIBROWIUS, H., 1983. - Extension de l'aire de répartition favorisée par l'homme chez les invertébrés marins. Oceanis, 9 (4) : 337-353.
ZIBROWIUS, H., 1991. - Ongoing modification of the Mediterranean marine fauna and flora by the establishment of exotic species. Mésogée, 51 : 83-107.
ZIBROWIUS, H., 1992. - Ongoing modification of the Mediterranean fauna and flora by the establishment of exotic species. Mésogée, Bull. Mus. Hist. nat. Marseille, 51, 1991 : 83-107.
ZIBROWIUS, H., RAMOS, A.A., 1983. - Oculina patagónica, Scléractiniaire exotique en Méditerranée. Nouvelles observations dans le Sud-Est de l'Espagne. Rapp. P. V. Réun. Comm. Intemation. Explor. sci. Médit., 28 (3) : 297-301.
ZIBROWIUS, H., THORP, C.H., 1990. - A review of the alien serpulid and spirorbid polychaetes in the British Isles. Cah. Biol, mar., 30 (3), 1989 : 271-285.
ZOUROS, E. & FOLTZ, D.W., 1984. - Possible explanations of heterozygote deficiency in Bivalve Molluscs. Malacologia, 25 (2) : 583-591.


European Commission
EUR 15309 — Introduced species in European Coastal Waters
C.F. Boudouresque, F. Briand, C. Nolan
Luxembourg: Office for Official Publications of the European Union
1994 —VI . 112 p. —16,2 χ 22,9 cm
ISBN 92-826-6727-8
Price (excluding VAT) in Luxembourg: ECU 13.50


Venta y suscripciones · Salg og abonnement · Verkauf und Abonnement · Πωλήσεις και συνδρομές Sales and subscriptions · Vente et abonnements · Vendita e abbonamenti
Verkoop en abonnementen · Venda e assinaturas
BELGIQUE BELGIË Moniteur belge / Belgisch staatsblad Rue de Louvain 42 Leuvenseweg 42 B1000 Bruxelles Β100Ό Brussel Tel (02)512 00 26 Fax (02) 51101 84 Jean De Lannoy Avenue du RCM 202 / Koningslaan 202 B1060 Bruxelles B1060 Brussel Tél (02)538 51 69 Télex 63220 UNBOOK Β Fax (02) 538 08 41 Autres dstnbuteurs/ Ovenge verkooppunten: Ubrairle européenne/ Europese boekhandel Rue de la Loi 244/Wetstraat 244 B1040 Bruxelles B1040 Brussel Tél. (02) 231 04 35 Fax (02) 735 08 60 Document delivery: Credoc Hue de la Montagne 34 / Bergstraat 34 Ble 11 Bus 11 B1000 Bruxelles B1000 Brussel Tel (02)51169 41 Fax (02) 513 31 95
DANMARK J. H. Schultz Information A/S Herstedvang 1012 DK2620 Albertslund Ttf 43 63 23 00 Fax (Sales) 43 63 19 69 Fax (Management) 43 63 19 49
DEUTSCHLAND Bundesanzeloar Vertag Breite Straße 7880 Postfach 10 05 34 D50445 Koin Tel. (02 21 [20 290 Telex ANZEIGER BONN 8 882 595 Fax 202 92 78
GREECE/ΕΛΛΑΔΑ G.C. Ekiftheroudakls SA International Bookslore Nikis Street 4 G R10563 Athens Tel (01)322 63 23 Telex 219410 ELEF Fax 323 98 21
ESPANA Boletín Ondai del Estado Trafalgar, 2729 E28071 Madrid Tel (91)538 22 95 Fax (91 ) 538 23 49 MundlPrensa Ubres, SA Castello. 37 E28001 Madrid
1339" ; 1 32 22 (Suscripciones)
435 36 37 (Dirección) Télex 49370MPLIë Fax (91) 575 39 98 Sucursal: Libreria Internacional AEDOS Consejo de Ciento, 391 E08009 Barcelona Tel (93)488 34 92 Fax (93) 487 76 59 Librarla de la Generalität de Catalunya Rambla deis Estudie. 118 (Patau Moja) E08002 Barcelona Tel (93) 302 68 35 Tel (93 302 64 62 Fax (93) 302 12 99
FRANCE Journal officiel Service des publications des Communautés européennes 26, rue Desaix F75727 Pans Cedex 15 Tel (1)40 58 77 01/31 Fax 1| 40 58 77 00
IRELAND Government Supplies Agency 45 Ha/court Road Dubim 2 Tel (1)6613111 Fax (1 47 80 645
ITALIA
ÖSTERREICH"
SUOMI/FINLAND Akateemlnen Klrjakauppa Keskuskatu 1 PO Box 218 FIN00381 Helsinki Tel. (0) 121 41 Fax (0)121 44 41
NORGE o Center
Bertrand Narvesens vel 2 PO Box 6125 Etterstad N0602Osto6 Tel. (22) 57 33 00 Telex 79668 NIC Ν Fax (22) 68 19 01
SVEHIGE BTJAB Traktorvgert 13 S22100Lund Tel (046) 18 00 00 Fax (046) 18 01 25 30 7947
SCHWEIZ / SUISSE / SVIZZERA
OSEC Stampfenbachstra8e 85 CH6035 Zunch Tel. (01 (365 54 49 Fax (011365 54 11
Licosa SpA Via Duca di Calabria 1/1 Casella postale 552 150125 Frenze Tel (055) 64 54 15 Fax 64 12 57 Telex 570466 LICOSA I
GRANDDUCHÉ DE LUXEMBOURG Messageries du livre 5, rue Raifleisen L2411 Luxembourg
NEDERLAND SDU Overheidsinformatie Exteme Fondsen Postbus 20014 2500 EA 'sGravenhage Tel.(070)37 89 911 Fax (070) 34 75 778
PORTUGAL Imprensa Nacional Casa da Moeda, EP Rua D Francisco Manuel de Melo, 5 Ρ1092 Lisboa Codex Tel. (01)69 34 14 Fax (01) 69 31 66
Grupo Bertrand, SA Rua das Terras dos Vales, 4A Apartado 37 P2700 Amadora Codex Tel. (01 ) 49 59 050 Telex 15798 BERDIS Fax 49 60 255
UNITED KINGDOM HMSO Books (Agency section) HMSO Publications Centre 51 Nine Elms Lane London SW8 5DR Tel (071)873 9090 Fax 873 8463 Telex 29 71 138
BALGARUA"
Europress Klasslca BK Ltd 66. bd Vrtosha BG1463 Sofia Tel ./Fax 2 52 74 75
CESKA HE PUBLIKA
NIS ÒR Havelkova 22 CZ130 00Praha3 Tel. (2) 235 84 46 Fax 2) 235 97 88
MAGYARORSZAG
EurotntoServlce Europa Haz Margitsziget Η1138 Budapest TeL/Fax 1 111 60 61 1 111 6216
POLSKA
Business Foundation ul Krucza 38/42 PL00512 Warszawa Tel (22) 621 99 93. 6282882 International Fax&Phone (039)120077
ROMANIA
65. Strada Dionisio Lupu RO70184 Bucuresti TeL/Fax 0 12 96 46
Manz'sche Verlagsund Unlversitatsbuchhandlung Kohlmarkt 16 A1014 Wien Tel. (1)531 610 Telex 112 500 BOX A Fax (1)531 61181
9.604etiya Oktyabrya Avenue 117312 Moscow TeL/Fax (095) 135 5227
SLOVAKIA
Slovak Technical Library Nm. slobody 19 SO812 23 Bratislava 1 Tel. (7)220 452 Fax : (7) 295 785
CYPRUS
Cyprai Iridust
is Chamber of Commerce and tdustry
Chamber Building 38 Gnvas Dhigenis Ave 3 Deligiorgis Street PO Box 1455 Nicosia Tel (2)449500/462312 Fax (2) 458630
MALTA
Miler distributors Ltd PO Box 25 Marta International Airport LOA 05 Matta Tel 66 44 88 Fax 67 67 99
Pres Gazete Kltap Dergl Pazariama Dagltlm Tlcaret ve sanavi AS Narlibaçhe Sokak Ν. 15 IstanbulCagaloglu Tel (1)520 92 96528 55 68 Fax 520 64 57 Telex 23822 DSVOTR
ROY International PO Box 13056 41 Mishmar Hayarden Street Tel Aviv61130 Tel 3 496108 Fax 3 648 60 39
Middle East Observer 41 Shenf St. Cairo Tel/Fax 39 39 732
UNITED STATES OF AMERICA / CANADA
UNIPUB 4611 F Assembly Drive Lanham. MD 207064391 Tel Toll Free (800) 274 4888 Fax (301) 459 0056
Subscriptions only Uniquement abonnements
Renouf Publishing Co. Ltd 1294 Algoma Road Ottawa. Ontario K1B 3W8 Tel (613)741 43 33 Fax (613) 741 54 39 Telex 0534783
Hunter Publications 58A Gipps Street CoHingwood Victoria 3066 Tel. (3)417 5361 Fax (3) 419 7154
JAPAN
Klnokuniya Company Ltd 177 Shiniuku 3Chome Shinjukuku Tokyo 16091 Tel. (03) 34390121
Journal Department PO Box 55 Chitóse Tokyo 156 Tel. (03) 34390124
SOUTHEAST ASIA
Legal Library Services Ltd STK Agency Robinson Road PO Box 1817 Singapore 9036
SOUTH AFRICA
5th Floor, Export House Cnr Maude & West Streets Sandten 2146 Tel.(011)8833737 Fax (011)8836569
AUTRES PAYS OTHER COUNTRIES ANDERE LANDER
Office des publications officielles des Communautés européennes 2, rue Mercier L2985 Luxembourg Tel 499 281 TélexPUBOf LU1324b Fax 48 85 73/48 68 17
10066

Ol f.
NOTICE TO THE READER
All scientific and technical reports published by the European Commission are announced in the monthly periodical 'euro abstracts'. For subscription (1 year: ECU 60) please write to the address below.
o
αϊ ω o (O
O
Price (excluding VAT) ¡η Luxembourg: ECU 13,50 T ? R M R P - A P L - L 7 3 7 A
t* * * OFFICE FOR OFFICIAL PUBLICATIONS * « f r * OF THE EUROPEAN COMMUNITIES
* * * L-2985 Luxembourg
Related Documents











