176 BBA 5 1068 INTRAVASCULAR METABOLISM OF AN ARTIFICIAL TRANSPORTER OF TRIACYLGLYCEROLS ALTERATIONS OF SERUM LIPOPROTEINS RESULTING FROM TOTAL PARENTERAL NUTRITION WITH INTRALIPID * STEVEN H. UNTRACHT ** (Received November 18th. 1981) As a model for transport in the bloodstream of exogenous triacylglycerols, we have studied the intravascular metabolism of Intralipid, an artificial, cholesterol- and protein-free ‘chylomicron’. Two patients with Crohn’s disease and one patient with intermittent abdominal pain received total parenteral nutrition including between 50 and 100 g of Intralipid per day. Samples of serum were analyzed chemically for lipids, assayed for lecithin:cholesterol acyltransferase (EC 2.3.1.43), and analyzed by isopycnic density-gradient ultracentrifuga- tion and electron microscopy. After 3 weeks of infusion, single-bilayer vesicles, 300-600 A in diameter, were present in the serum of each patient. Comprising equimolar amounts of phospholipids and unesterified cholesterol, and approximately 5% by weight of protein, these vesicles accumulated to levels of approximately 100,300 and 50 mg/dl for the three patients indicated above, causing hyperlipidemia in the first two cases. In conjunction, the levels of HDL and lecithin:cholesterol acyltransferase had decreased by 40% in each patient. Upon stopping the infusions, vesicles disappeared with monophasic kinetics with a half-life of 2 days, and were not converted to HDL or other lipoproteins; HDL and lecithin: cholesterol acyltransferase returned to normal levels after a lag period of at least 4 days. Two normal subjects were studied during and after a single 4-h infusion of 50 g of Intralipid. Triacylglycerols were cleared with a half-life of 1 h, but at least 40% of the phospholipids remained in the serum as vesicles. Although Intralipid contains no steroids, unesterified cholesterol progressively entered the serum after the vesicles appeared. During these short-term studies, the levels of HDL and lecithin:cholesterol acyltransferase did not change. The observations indicate that vesicles, derived from a variable fraction of the Intralipid phospholipids, extract unesterified cholesterol into plasma. Inert and slowly cleared, the resulting mixed vesicles accumulate in the blood. The levels of HDL and 1ecithin:cholesterol acyltransferase decrease only when the intestine is bypassed for long periods of time, probably because the lipoprotein and enzyme are synthesized more slowly than usual. Our results suggest that in chylomicrons other mechanisms prevent phospholipids from being released into the bloodstream as vesicles. * Accounts of this work have appeared in abstract form (( 1979) Clin. Res. 27, 444A; Clin. Res. 27, 504A). ** Present address: Department of Surgery; Massachusetts General Hospital; Boston, MA 021 14. U.S.A OOOS-2760/82/OOOC-OooO/$O2.75 Q I982 Elsevier Biomedical Press Abbreviations: HDL, high-density lipoproteins, i.c.. serum lipoproteins with density between 1.063 and 1.21 g/ml: LDL, low-density lipoproteins, i.e.. serum lipoproteins with density between I.019 and I.063 g/ml; VLDL. very low density lipo- proteins, i.e., serum lipoproteins with density less than 1.006 g/ml; LpX. lipoprotein X.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

176
BBA 5 1068
INTRAVASCULAR METABOLISM OF AN ARTIFICIAL TRANSPORTER OF TRIACYLGLYCEROLS
ALTERATIONS OF SERUM LIPOPROTEINS RESULTING FROM TOTAL PARENTERAL NUTRITION WITH INTRALIPID *
STEVEN H. UNTRACHT **
(Received November 18th. 1981)
As a model for transport in the bloodstream of exogenous triacylglycerols, we have studied the intravascular metabolism of Intralipid, an artificial, cholesterol- and protein-free ‘chylomicron’. Two patients with Crohn’s disease and one patient with intermittent abdominal pain received total parenteral nutrition including between 50 and 100 g of Intralipid per day. Samples of serum were analyzed chemically for lipids, assayed for lecithin:cholesterol acyltransferase (EC 2.3.1.43), and analyzed by isopycnic density-gradient ultracentrifuga- tion and electron microscopy. After 3 weeks of infusion, single-bilayer vesicles, 300-600 A in diameter, were present in the serum of each patient. Comprising equimolar amounts of phospholipids and unesterified cholesterol, and approximately 5% by weight of protein, these vesicles accumulated to levels of approximately 100,300 and 50 mg/dl for the three patients indicated above, causing hyperlipidemia in the first two cases. In conjunction, the levels of HDL and lecithin:cholesterol acyltransferase had decreased by 40% in each patient. Upon stopping the infusions, vesicles disappeared with monophasic kinetics with a half-life of 2 days, and were not converted to HDL or other lipoproteins; HDL and lecithin: cholesterol acyltransferase returned to normal levels after a lag period of at least 4 days. Two normal subjects were studied during and after a single 4-h infusion of 50 g of Intralipid. Triacylglycerols were cleared with a half-life of 1 h, but at least 40% of the phospholipids remained in the serum as vesicles. Although Intralipid contains no steroids, unesterified cholesterol progressively entered the serum after the vesicles appeared. During these short-term studies, the levels of HDL and lecithin:cholesterol acyltransferase did not change. The observations indicate that vesicles, derived from a variable fraction of the Intralipid phospholipids, extract unesterified cholesterol into plasma. Inert and slowly cleared, the resulting mixed vesicles accumulate in the blood. The levels of HDL and 1ecithin:cholesterol acyltransferase decrease only when the intestine is bypassed for long periods of time, probably because the lipoprotein and enzyme are synthesized more slowly than usual. Our results suggest that in chylomicrons other mechanisms prevent phospholipids from being released into the bloodstream as vesicles.
* Accounts of this work have appeared in abstract form
(( 1979) Clin. Res. 27, 444A; Clin. Res. 27, 504A).
** Present address: Department of Surgery; Massachusetts General Hospital; Boston, MA 021 14. U.S.A
OOOS-2760/82/OOOC-OooO/$O2.75 Q I982 Elsevier Biomedical Press
Abbreviations: HDL, high-density lipoproteins, i.c.. serum
lipoproteins with density between 1.063 and 1.21 g/ml: LDL,
low-density lipoproteins, i.e.. serum lipoproteins with density
between I.019 and I.063 g/ml; VLDL. very low density lipo-
proteins, i.e., serum lipoproteins with density less than 1.006
g/ml; LpX. lipoprotein X.

Introduction
As they transport lipids through the bloodstream, the four major classes of serum lipo-
proteins - chylomicrons, VLDL, LDL, and
HDL-are interconverted through a variety of
pathways [l-3]. Chylomicrons and VLDL prim-
arily serve to deliver absorbed and endogenously
synthesized triacylglycerols, respectively, to
adipose tissue, skeletal muscle and lactating
mammary glands [2,4]. In the case of chylomicrons,
it has long been known [5-71 that when the tri-
acylglycerols are removed from the circulation,
new phospholipids appear in the HDL fraction-
presumably through the action of lecithin : cholesterol acyltransferase [ 1,8- 111. Recent ra- dioisotope tracer studies in rats [ 12,131 have dem-
onstrated that these HDL-bound phospholipids are derived directly from chylomicrons. Radioac-
tive-labeling studies in humans [14] and in rats
[ 12,131 have also shown that apolipoproteins are
transferred simultaneously with phospholipids
from the chylomicrons to HDL.
Such interconversions seem to be important in
maintaining homeostasis of serum lipids and lipo-
proteins, and may be governed in part by the
complicated chemical make-up of these particles.
We were, therefore, prompted to study the in-
travascular metabolism of Intralipid, and artificial
transporter of triacylglycerols whose chemical
composition is much simpler than that of
chylomicrons. Intralipid is an aqueous dispersion
of egg-yolk lecithin (12% by weight), soybean-oil
triacylglycerols (88% by weight), and trace amounts of cholesterol. The lipids are emulsified into a
heterogeneous group of particles that range in diameter from 200 to 1000 A [ 151. This phos-
pholipid-stabilized triacylglycerol emulsion is in- fused intravenously as a source of calories in pa-
tients who suffer from inflammatory bowel dis-
ease, malnutrition, gastro-intestinal pain or ob-
struction, or any other disoder in which the gastro-
intestinal tract is not functional [15]. In many
cases, patients receive balanced diets in which all of the fat is supplied as Intralipid, carbohydrates are infused as glucose, and amino acids, vitamins and minerals are administered as such [ 15- 171. In these ‘total parenteral nutrition’ regimens, all of the calorie sources enter the circulation directly
and are not processed by the small intestine.
Therefore, during total parenteral nutrition
chylomicrons do not enter the bloodstream; under
these conditions the Intralipid particle carries exo-
geous triacylglycerols and functions as a
chylomicron.
A variety of studies suggest that Intralipid and
chylomicrons are metabolized similarly. For exam-
ple, Hallberg [ 181 has shown that normal adults
clear Intralipid triacylglycerols from the blood at a
rapid, dose-dependent, and saturable rate. Subse-
quently, Have1 et al. [7] demonstrated that In-
tralipid acquires C-peptides from HDL, and
thereby becomes a suitable substrate for lipopro-
tein lipase. More recently, Robinson and Quar-
fordt [19] have found that Intralipid particles can also obtain apolipoprotein A-I, the arginine-rich
peptide, and other proteins from HDL in vitro.
Moreover, ultra-histochemical studies have shown
that, as is the case with chylomicrons, the tri-
acylglycerols of Intralipid are hydrolyzed at the
surface of endothelial cells in capillaries [20,21].
For the above-mentioned reasons, total
parenteral nutrition with Intralipid is a good model
system to study the intravascular transport of ex-
ogenous fats. With this purpose in mind, we now
present our studies of five human subjects who
have been infused with the triacylglycerol emul- sion.
Materials and Methods
Subjects. Three of the five subjects were pa-
tients from the Gastroenterology Service at the
University of Chicago.
Subject 1 (case l), a lCyear-old white male with a 7-year history of Crohn’s disease, was admitted
to the hospital for correction of secondary malnutrition and to be evaluated for arrested
growth and development. Initially, he was in-
travenously hyperalimented with 50% glucose.
However, this therapy affected his liver adversely:
serum glutamic-oxaloacetic transaminase, glutamic -pyruvic transaminase, lactate dehydrogenase and alkaline phosphatase all rose to abnormal levels, and he developed hepatomegaly. The treatment, therefore, was changed to a 5-week total parenteral nutrition regimen comprising 1000 ml/day of 10% Intralipid (Cutter Laboratories: Berkeley, CA),

17x
Fig I. Levels of total cholesterol (unesterified and esterified) in
the serum of subject 1 during total parenteral nutrition with
Intralipid. This patient received 1000 ml of Intralipid per week
prior to day 0. On day 0. he began to receive 1000 ml of
Intralipid per day for approximately 6 weeks. These data were
obtained from the patient’s clinical records.
2000 ml/day of 20% glucose in water, and 1000
ml/day 10% glucose in water containing 3% Freamine II (total calories: 3130/day). Under the
second treatment, his hepatic function returned to
normal. However, serum-cholesterol levels began
to rise immediately, and they reached a total value
of nearly 500 mg/dl by the end of 2 weeks, as
shown in Fig. 1. The cholesterol level remained
elevated for the duration of the infusion period *.
Subject 2 (case2), a 35-year-old white female,
required total parenteral nutrition because of in-
termittent abdominal pain. She was given 1000
ml/day of 10% Intralipid, and 3000 ml/day 5%
glucose in aqueous 0.45% sodium chloride contain-
ing 3% Frearnine II (total calories: 2350/day). In
this patient, total parenteral nutrition lasted for 3 weeks. Her serum-cholesterol level was 168 ml/d1
on the day that parenteral nutrition was started,
but this value rose to 276 mg/dl by the end of the
infusion period. Subject 3 (case 3) was a 16-year-old black female
with a 3-year history of Crohn’s disease. She was hospitalized for treatment of an anal fistula and for ‘bowel rest’. The latter was achieved with a
* In three patients receiving chronic total parenteral nutrition
in which 60% of the non-protein calories were supplied as
hypertonic glucose and the remaining 40% were supplied as
Intralipid. Broviac et al. [ 161 found that the steady-state
concentration of total serum cholesterol was 315-’ 100
mg/dl. When the same patients chronically received all of their non-protein calories as glucose, their serum cholesterol
levels averaged I56 i 15 mg/dl.
5-week course of total parenteral nutrition with
10% Intralipid (500 ml/day) and 3000 ml/day of
20% glucose in water containing a 4.5% Freamine
II (total calories: 3058/day).
During their hospitalizations, none of the three
patients had icterus, elevated levels of serum bi-
lirubin, or other clinical signs of cholestasis. No
other known causes of hyperlipidemia were pre-
sent in any of these cases.
Two healthy, male, student volunteers served as
subjects 4 and 5 (ages 23 and 22 years, respec-
tively). Neither subject thad received Intralipid
before this study or had a history of liver disease
or serum-lipid abnormalities.
The clinical details of the five subjects are listed
in Table I. In no case did the dose of tri-
acylglycerols exceed that in a normal diet. For
example, after total parenteral nutrition was dis-
continued, subjects 1 and 2 continued to consume
approximately the same amount of fat and total
calories. Therefore, in these two cases, the dose of
Intralipid was equivalent to the normal daily flux
of chylomicrons. Each of the other three subjects
received much smaller relative doses (Table I).
Thus, none of the results can be attributed to ‘fat
overloading’.
Sampling schedule. Approximately 8 ml of blood
was obtained from subjects 1 and 2 at 48-h or, in
some cases, 24-h intervals, for a least 2-weeks after
the last infusion. Subject 3 was studied 4 weeks after the first infusion, as well as 6 and 11 weeks
after parenteral nutrition was discontinued. The
blood was allowed to clot for 1 h at room tempera-
ture. The formed elements were then sedimented
by low-speed centrifugation for 10 min, and the
serum was analyzed by the procedures outlined
below.
The two normal subjects were studied after an
overnight fast and, except for the intravenous fat,
both subjects fasted during the 8-h experiment. An Angiocath, inserted into a forearm vein for pur- poses of obtaining blood, was maintained patent by infusing sterile physiological saline at a rate of 250 ml/h. After a control blood sample was ob- tained, 500 ml of 10% Intralipid was infused in- travenously through the opposite arm at a con- stant rate of 125 ml/h, using an Imed (San Diego, CA) Model 922 Volumetric Infusion Pump. Fur- ther samples of blood were obtained at 0.5, 1, 2, 3,
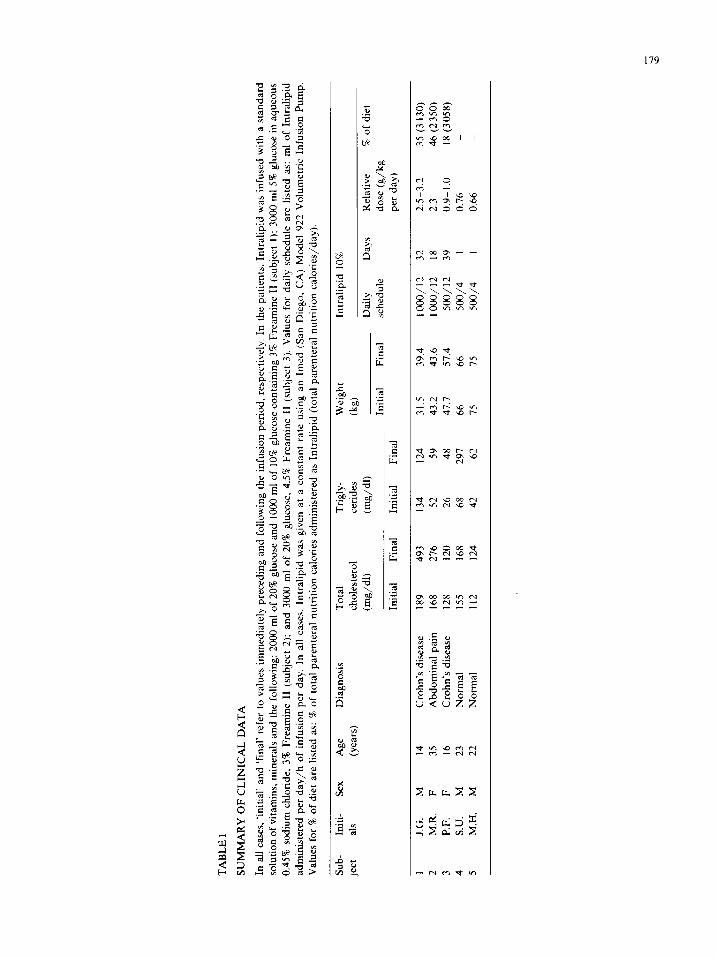
TA
BL
E
I
SUM
MA
RY
O
F C
LIN
ICA
L
DA
TA
In
all
case
s,
‘ini
tial’
an
d ‘f
inal
’ re
fer
to
valu
es
imm
edia
tely
pr
eced
ing
and
follo
win
g th
e in
fusi
on
peri
od.
resp
ectiv
ely.
In
th
e pa
tient
s,
Intr
alip
id
was
in
fuse
d w
ith
a st
anda
rd
solu
tion
of v
itam
ins,
m
iner
als
and
the
follo
win
g:
2000
m
l of
20
% g
luco
se
and
loo0
m
l of
10
% g
luco
se
cont
aini
ng
3% F
ream
ine
II (
subj
ect
I):
3000
m
l 5%
glu
cose
in
aqu
eous
0.45
%
sodi
um
chlo
ride
, 3%
Fr
eam
ine
II
(sub
ject
2)
; an
d 30
00
ml
of
20%
gl
ucos
e,
4.5%
Fr
eam
ine
II
(sub
ject
3)
. V
alue
s fo
r da
ily
sche
dule
ar
e lis
ted
as:
ml
of
Intr
alip
id
adm
inis
tere
d pe
r da
y/h
of
infu
sion
pe
r da
y.
In
all
case
s,
Intr
alip
id
was
gi
ven
at
a co
nsta
nt
rate
us
ing
an
Imed
(S
an
Die
go,
CA
) M
odel
92
2 V
olum
etri
c In
fusi
on
Pum
p.
Val
ues
for
S of
di
et
are
liste
d as
: %
of
tota
l pa
rent
eral
nu
triti
on
calo
ries
ad
min
iste
red
as
Intr
alip
id
(tot
al
pare
nter
al
nutr
ition
ca
lori
es/d
ay).
Sub-
ject
Initi
- Se
x A
ge
Dia
gnos
is
Tot
al
Tri
gly-
W
eigh
t In
trah
pid
10%
als
(yea
rs)
chol
este
rol
ceri
des
(kg)
(mg/
dB
(mg/
dB
Dai
ly
Day
s R
elat
ive
% o
f di
et
Initi
al
Fina
l sc
hedu
le
dose
(g
/kg
Initi
al
Fina
l In
itial
Fi
nal
per
day)
I J.
G.
M
14
Cro
hn’s
di
seas
e 18
9 49
3 13
4 12
4 31
.5
39.4
10
00/1
2 32
2.
5-3.
2 35
(3
130)
2 M
.R.
F 35
A
bdom
inal
pa
in
168
276
52
59
43.2
43
.6
1000
/12
I8
2.3
46
(235
0)
3 P.
F.
F I6
C
rohn
’s
dise
ase
I28
120
26
48
41.7
57
.4
500/
12
39
0.9-
I .o
I8
(30
58)
4 S.
U.
M
23
Nor
mal
15
5 I6
8 68
29
7 66
66
50
0/4
I 0.
76
5 M
.H.
M
22
Nor
mal
11
2 12
4 42
62
75
75
50
0/4
I 0.
66
_

1 x0
and 4 h-during the infusion, and at various inter-
vals after the infusion was completed. Each of the
resulting sera were analyzed as outlined below. In
addition, parameters of hepatic and renal func-
tion, which remained normal in both subjects,
were measured periodically by the Clinical
Laboratories of the University of Chicago.
Informed consent was obtained from all par-
ticipants before studies were undertaken. The
single-infusion protocol (i.e., the studies on sub-
jects 4 and 5) was approved by the Clinical Inves-
tigation Committee of the University of Chicago (Protocol No. 2779).
Analytical methods. Density-gradient ultra- centrifugation was performed on l-ml aliquots of
whole serum, according to the method of Foreman
et al. [22]. The resulting gradient was fractionated
and monitored at 280 nm, using an Isco density-
gradient flow cell. A typical analysis is shown in
Fig. 2.
Triacylglycerols and total cholesterol were mea-
sured with a Technicon AutoAnalyzer II [23]; un-
esterified cholesterol was measured as the dig-
itonide [24,25]. Phospholipids were extracted from serum into chloroform/methanol (2: 1 by vol.),
and then measured in the extract, as described by
Bartlett [26].
Free fatty acids were determined by the method
of Mikac-Devic et al. [27]; sodium palmitate was used as the standard. HDL was quantitated chemi-
cally through its cholesterol after precipitating
other lipoproteins with dextran sulfate [28]. Cholesterol weight was multiplied by 8.4 (based on
chemical composition), to obtain the weight of
HDL. I .o ,. , ,-I.25
0.0 - 1.20
u x
U 0.6 -
Y
1.15 H
::
0.4 - 1.10 p
z =
Q 0.2 - “LDL I .05
0 ! ! 1.00 0 2 4 6 6 IO I2 I4 16 top VOLUME (ml)
Fig. 2. Representative isopycnic density-gradient pattern of I ml
of normal human serum (total cholesterol. 180 mg/dl; tri-
acylglycerols, SO mg/dl). The gradients were prepared and
analyzed as described in Materials and Methods.
Lecithin: cholesterol acyltransferase activity was
assayed as previously reported [29]. With this
method, normal activity of lecithin : cholesterol
acyltransferase ranges from 700 to 1400 dpm/h.
For electron microscopy, specimens of nega-
tively stained lipoproteins were prepared as de-
scribed by Ohtsuki et al. [30], and viewed with a
Philips EM 300 electron microscope operated at a
beam-acceleration voltage of 80 kV and a primary
magnification of 70000 X Gel-filtration chromatography was performed
at 23’C, using a column (1.5 X 90 cm; Pharmacia;
Uppsala, Sweden) packed with 4% agarose (Biogel
A 15 m; Biorad; Richmond, CA). Samples were
eluted with 0.15 M NaCl/O.OS% EDTA/O.OS%
NaN,, pH 7.4, at a downward flow rate of 15
ml/h. Absorbance of the effluent was monitored
at 280 nm.
Protein was measured by the procedure of
Lowry et al. [31], with bovine serum albumin as
the standard. Radioimmunoassays for
apolipoproteins A-I, A-II and B were performed
as previously described [32-341.
Results
Long-term infusions of Intrulipid in patients.
After the last infusion of Intralipid, the serum of
subjects I and 2 contained abnormally large con-
centrations of unesterified cholesterol (2 13 and
180 mg/dl, respectively) and phospholipids (550 and 450 mg/dl, respectively), whereas cholesteryl
esters and triacylglycerols were present in normal
amounts. Subject 3, who received a relatively small
dose of Intralipid, was not hyperlipidemic. After
each patient was infused for at least 3 weeks,
lecithin : cholesterol acyltransferase activity and
HDL concentrations were reduced by approxi-
mately 40%. Upon stopping the treatment in subjects 1 and
2, the concentrations of phospholipids and un- esterified cholesterol decreased immediately and reached normal levels approximately 2 weeks later (Figs. 3A and 4A) *. After terminating total
-~ * Between days 5 and 8. the concentration of cholesteryl
esters increased sharply in subject 2 (Fig. 4A). This second
hypercholesterolemia resolved slowly over the ensuing
3 weeks. Density-gradient ultracentrifugation revealed that
this second hyperlipidemia was caused by increased lcvcls of LDL.

TABLE II
SERUM CHEMISTRIES IN CASE 3
During total parenteral refers to day 26 of the 39.day infusion
period. After total parenteral refers to I I weeks after the last
infusion. Values are mg/dl, except for lecithin : cholesterol
acyltransferase activity where results are given as meanirange
of duplicate determinations. Values for HDL were determined
from dextran-sulfate-soluble cholesteroi.
During After
total total
parenteral parenteral
nutrition nutrition
Cholesterol
Total 120 141
Unesterified 51 39
Esterified 69 102
Phospholipids 185 186
Triacylglycerols 59 62
HDL 134 294
Lecithin : cholesterol
acyltransferase activity 464% 32 631263
parenteral nutrition in subject 3, levels of
cholesteryl esters increased by SO%, whereas the level of unesterified cholesterol decreased by 25%
(Table 11). Lecithin: cholesterol acyltransferase ac-
tivity and HDL concentrations simultaneously
returned to normal values, following a lag period
ranging from 4 days in subject 1 (Fig. 3B) to 6 weeks in subject 2 (Fig. 4B). In all cases, as
illustrated in Fig. 5, HDL concentrations corre-
lated with lecithin: cholesterol acyltransferase ac- tivity.
To obtain complementary information of the
manner in which the lipids were distributed among
the serum lipoproteins, the post-infusion sera were
analyzed by isopycnic density-gradient ultra- centrifugation. The results are shown in Figs. 6 and 7. Initially, the profiles were very unusual, but after approximately 2 weeks, they were similar to
those of most healthy donors (cf., Fig. 2, lower
tracings in Fig. 6, and thin-line tracings in Fig. 7). In conjunction with the elevated levels of serum phospholipids, a large, asymmetric peak was pre- sent over the density range between 1.020 and 1.034 g/ml (3-5 ml, arrow A in the top panel of Figs. 6 and 7). This component, which is absent from normal sera (e.g., Fig. 2), disappeared rapidly
in subjects 1 and 2 and was not detectable 10 or 11
days after the last infusion (Fig. 6 and upper panel
of Fig. 7). Moreover, although subject 3 was not hyperlipidemic, the abnormal peak was present
during the infusion period in this case as well
(Fig. 7, lower panel, arrow A).
The density-gradient profiles, in addition, con-
firmed the reduced levels of HDL (Figs. 6 and 7;
8-12 ml). The areas of the HDL peaks increased in parallel with the serum levels of dextran-sulfate-
soluble cholesterol (e.g., Fig. 3B) *. Despite the
reduced levels of HDL, however, the contours of the HDL peaks were normal in all cases. There-
fore, it is likely that the HDL remained normal
qualitatively.
The lipoproteins corresponding to the two major
peaks (Fig. 6 and 7, arrows A and B) were
examined by electron microscopy. Representative
micrographs are displayed in Fig. 8. The denser,
minor component (arrow B, Figs. 6 and ‘7) con-
sisted of homogeneous spherical particles with a
mean diameter of 249 k 1.6 A (mean t S.E. of 100
measurements for Subject 1) and appeared identi- cal to normal LDL (Figs. 8B, D, and F). The
major component, which banded over the density range between 1.020 and 1.034 g/ml (arrow A,
Figs 6 and 7), contained a few particles that may
be LDL, but consisted mostly of vesicular com- plexes 300-600 A in diameter (Figs. 8A, C, and E).
These latter complexes tended to stack; when
viewed in cross-section they each had a hollow interior and a 50-A-thick limiting membrane (e.g.,
Fig. SA, arrow).
From 4% agarose columns the vesicular lipo-
protein eluted near the void volume, although it
penetrated the gel considerably. These characteris-
tics are consistent with particle sizes that range
from 300 to 6OOA. Only a small amount of
material, approximately 2% of the total mass, eluted in the position that corresponds to normal LDL.
* Case 3 was an exception to this general rule. Although
chemical analyses demonstrated the rise of serum cholestetyl
ester and HDL levels (Table II), the density-gradient peaks corresponding to HDL did not increase in size after total
parenteral nutrition was stopped. The reasons for this dis-
crepancy are uncertain; however, the ultracentrifugal
method measures protein, whereas the chemical method
measures cholesterol, and the latter is probably more rele-
vant in this case.
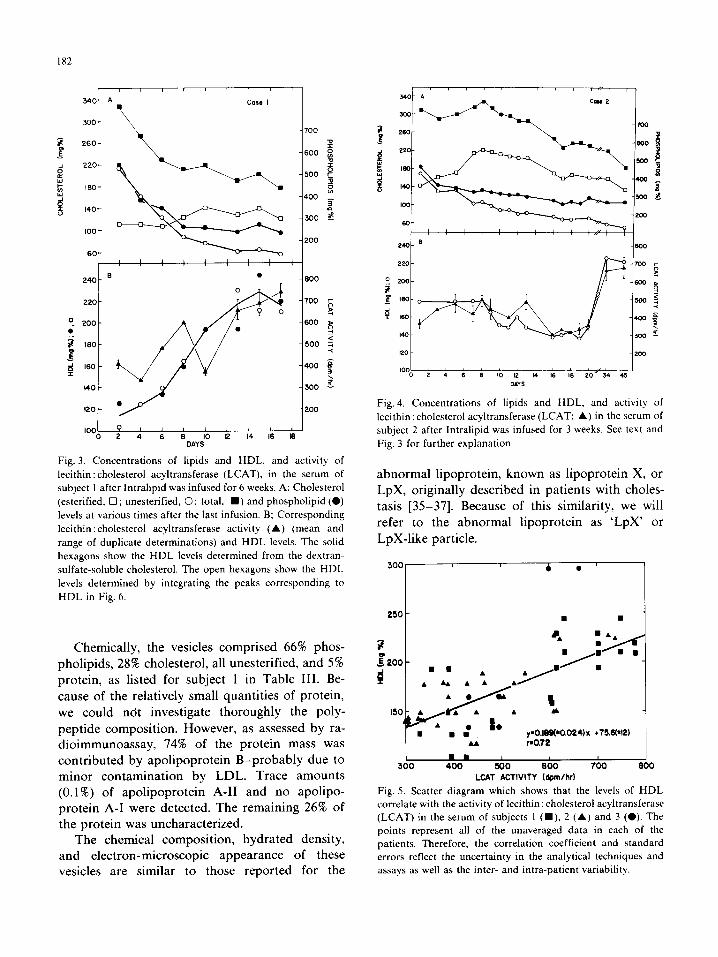
182
340. A cats I
3oo-
240 CS .
-I 800
9 zoo- .
s ISO-
s
$ ‘SO-
140
t 120
t
-700 6
3 -600 P
2 -500 z
-400 E 5 2
-300 =
10000 DAYS
Fig. 3. Concentrations of lipids and HDL. and activity of
lecithin: cholesterol acyltransferase (LCAT), in the serum of
subject I after Intralipid was infused for 6 weeks. A: Cholesterol
(esterified, 0 ; unesterified, 0; total, n ) and phospholipid (0)
levels at various times after the last infusion. B: Corresponding
lecithin : cholesterol acyltransferase activity (A) (mean and
range of duplicate determinations) and HDL levels. The solid
hexagons show the HDL levels determined from the dextran-
sulfate-soluble cholesterol. The open hexagons show the HDL
levels determined by integrating the peaks corresponding to
HDL in Fig. 6.
Chemically, the vesicles comprised 66% phos- pholipids, 28% cholesterol, all unesterified, and 5%
protein, as listed for subject 1 in Table III. Be-
cause of the relatively small quantities of protein,
we could not investigate thoroughly the poly-
peptide composition. However, as assessed by ra- dioimmunoassay, 74% of the protein mass was contributed by apolipoprotein B-probably due to minor contamination by LDL. Trace amounts (0.1%) of apolipoprotein A-II and no apolipo- protein A-I were detected. The remaining 26% of the protein was uncharacterized.
The chemical composition, hydrated density, and electron-microscopic appearance of these vesicles are similar to those reported for the
Fig. 4. Concentrations of lipids and HDL, and activity of
lecithin: cholesterol acyltransferase (LCAT; A) in the serum of
subject 2 after Intralipid was infused for 3 weeks. See text and
Fig. 3 for further explanation.
abnormal lipoprotein, known as lipoprotein X, or LpX, originally described in patients with choles- tasis [35-371. Because of this similarity, we will
refer to the abnormal lipoprotein as ‘LpX’ or
LpX-like particle.
300 I 6 . ’
250 - n 8
y.O.IW~0.024)~ +75.6(‘12) . . PO.72
m . 1 400 500 800 700 800
LCAT ACTIVITY (dpmlhr)
Fig. 5. Scatter diagram which shows that the levels of HDL
correlate with the activity of lecithin: cholesterol acyltransferase
(LCAT) in the serum of subjects I (m), 2 (A) and 3 (0). The
points represent all of the unaveraged data in each of the
patients. Therefore, the correlation coefficient and standard
errors reflect the uncertainty in the analytical techniques and
assays as well as the inter- and intra-patient variability.
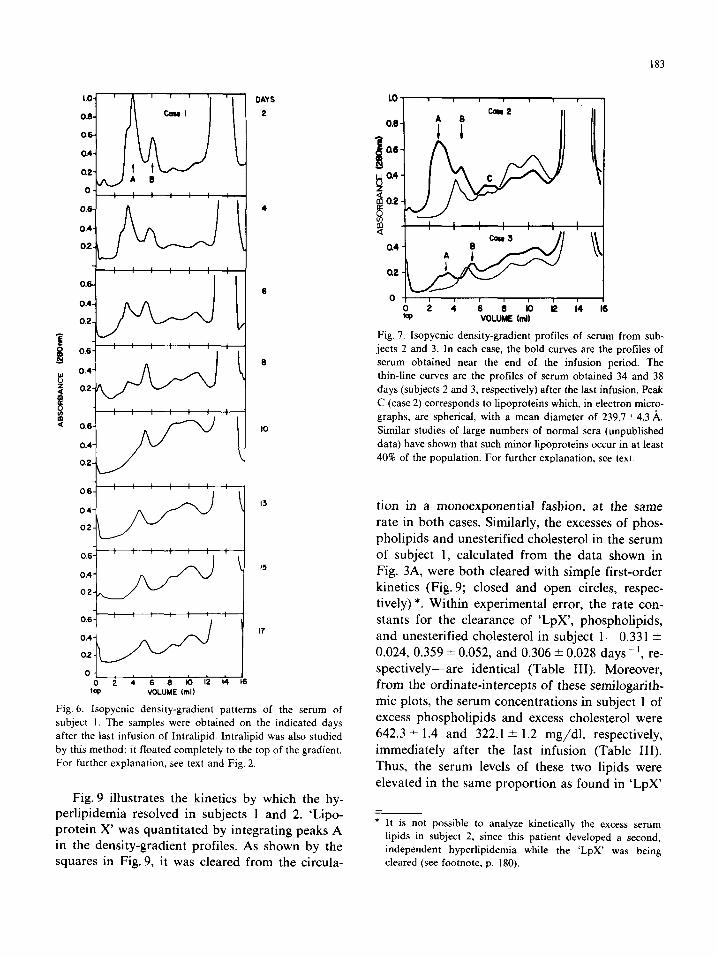
183
0.6-
0.4-
0.2-
0.6
0.4
0.2
06
0.6
0.4
02
0.6
4
6
6
Fig. 6. Isopycnic density-gradient patterns of the serum of
subject I. The samples were obtained on the indicated days
after the last infusion of Intralipid. Intralipid was also studied
by this method; it floated completely to the top of the gradient.
For further explanation, see text and Fig. 2.
Fig. 9 illustrates the kinetics by which the hy- perlipidemia resolved in subjects 1 and 2. ‘Lipo- protein X’ was quantitated by integrating peaks A in the density-gradient profiles. As shown by the squares in Fig. 9, it was cleared from the circula-
a6
w a4 Y 8 a2
pi $
a4
a2
Fig. 7. Isopycnic density-gradient profiles of serum from sub-
jects 2 and 3. In each case, the bold curves are the profiles of
serum obtained near the end of the infusion period. The
thin-line curves are the profiles of serum obtained 34 and 38
days (subjects 2 and 3, respectively) after the last infusion. Peak
C (case 2) corresponds to lipoproteins which, in electron micro-
graphs, are spherical, with a mean diameter of 239.7*4.3 A.
Similar studies of large numbers of normal sera (unpublished
data) have shown that such minor lipoproteins occur in at least
40% of the population. For further explanation, see text
tion in a monoexponential fashion, at the same
rate in both cases. Similarly, the excesses of phos-
pholipids and unesterified cholesterol in the serum
of subject 1, calculated from the data shown in
Fig. 3A, were both cleared with simple first-order
kinetics (Fig. 9; closed and open circles, respec-
tively) *. Within experimental error, the rate con-
stants for the clearance of ‘LpX’, phospholipids,
and unesterified cholesterol in subject l-0.331 *
0.024, 0.359 * 0.052, and 0.306 -C 0.028 days-‘, re-
spectively-are identical (Table III). Moreover,
from the ordinate-intercepts of these semilogarith-
mic plots, the serum concentrations in subject 1 of excess phospholipids and excess cholesterol were
642.3 i 1.4 and 322.1 + 1.2 mg/dl, respectively,
immediately after the last infusion (Table III).
Thus, the serum levels of these two lipids were
elevated in the same proportion as found in ‘LpX’
* It is not possible to analyze kinetically the excess serum
lipids in subject 2, since this patient developed a second.
independent hyperlipidemia while the ‘LpX’ was being
cleared (see footnote, p. 180).
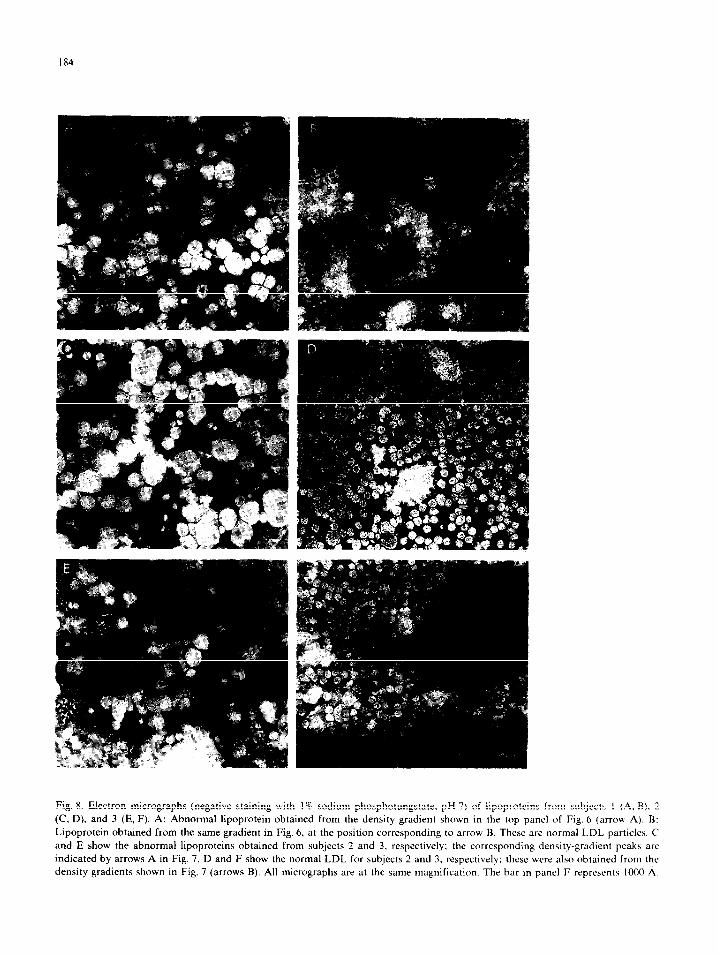
Fig. 8. Electron micrographs (negative staining with 1% sodium phosphotungstate, pH 7) of lipoproteins from subJccta I (A, B), 2
(C, D). and 3 (E, F). A: Abnormal lipoprotein obtained from the density gradient shown in the top panel of Fig. 6 (arrow A). B:
Lipoprotein obtained from the same gradient in Fig. 6, at the position corresponding to arrow B. These are normal LDL particles. C
and E show the abnormal lipoproteins obtained from subjects 2 and 3. respectively; the corresponding density-gradient peaks arc
indicated by arrows A in Fig. 7. D and F show the normal LDL for subjects 2 and 3. respectively: these were also obtained from the
density gradients shown in Fig. 7 (arrows B). All micrographs arc at the same magnification. The bar in pane1 F reprcscnts 1000 A.

185
TABLE III
ABNORMALITIES OF SERUM LIPIDS IN CASE I
Values for abnormal lipoprotein were obtained from the pooled fractions of the density gradients (Fig. 6) that correspond to peak A.
Excess serum level is the concentration in serum, in excess of the normal values for this patient, at the end of the last infusion. The
values were determined by analyzing the data according to the equation: In X(t)= -X(+lnX(O), where X(f) is the excess
concentration in the serum at time t after the last infusion, and k is the first-order rate constant for clearance of X (k is expressed in
units of I/time). X(0) is the value given in the table. Results are expressed as meancS.E. r,,2 =0.693/k. where k is as defined above.
Abnormal lipoprotein Excess serum level Clearance half-life
percent composition (mg/dl) (t,,r) (days)
(by wt.)
Density-gradient peak A
Phospholipids
Unesterified cholesterol
Cholesteryl esters
Protein
Weight ratio
Phospholipids/unesterified cholesterol
_ 2.09”0.15 a
66.2 642.3 * I .4 1.93-co.33
28.1 322.1 * 1.2 2.21-0:23
0 0 _
5.7
2.36 to.24 1.99-cO.012 _
a In case 2, ‘LpX’ was cleared with a half-life of 2. I8 to.06 days.
(Table III). These results confirm that the hyper-
lipidemia resulted exclusively because ‘LpX’ accu-
mulated. This abnormal lipoprotein is removed
directly from the circulation as intact units, with
3-
0 2 4 6 6 IO 12 OI\ys
Fig. 9. Semi-logarithmic-plots showing monoexponential first-
order clearance of ‘LpX’ and excess lipids from the serum of
subjects 1 and 2. w, 0: Serum level of ‘LpX’ in subjects I and 2, respectively, determined by integrating peak A in the ap-
propriate density-gradient profiles (i.e., Fig. 6 in the case of
subject 1, upper panel of Fig. 7 and other data (not shown) in the case of subject 2. The peak areas were normalized so that
for both subjects the semi-logarithmic plots would have the
same ordinate-intercept); 0: excess phospholipids in subject 1:
0 : excess unesterified cholesterol in subject I.
monoexponential first-order kinetics and a half-life
of 2 days.
Single infusions of Intralipid in normal subjects.
Two normal subjects were studied during and immediately following a 4-h infusion of Intralipid.
Results of chemical analyses are displayed in Fig.
10. Serum lecithin : cholesterol acyltransferase ac-
tivity, HDL concentrations, and cholesteryl levels
remained constant, except for some minor dilution
effects during the 1st h. Venous blood pH was
measured in subject 5; it remained constant at 7.34
despite the large elevation of serum free-fatty-acid
levels (see below). Wright-stained smears of pe-
ripheral blood were normal at the beginning and
end of the experiment.
In both cases, levels of serum triacylglycerols
rose to their maximum values by the end of the
infusion and then rapidly declined thereafter (Fig. 10, top panels). In subject 5, triacylglycerol con-
centrations quickly reached a steady-state level of
300 mg/dl. After the infusion was terminated, this excess lipid was completely eliminated within 4 h,
with first-order kinetics and a half-life of 1 h. In subject 4, triacylglycerol concentrations rose at -a nearly constant rate to 600 mg/dl by the end of infusion, with no indication that a steady-state level had been attained. In this case, approxi- mately 50% of the excess lipid was eliminated within 2 h after the end of the infusion, whereas

0 2 4 6 6 0 2 4 6 a
Fig. 10. Concentrations of lipids and HDL, and activity of lecithin : cholesterol acyltransferase (LCAT) in the serum of subjects 4 and
5. These subjects were each studied during and immediately after the intravenous infusion of 500 ml of Intralipid. Results for subject 4
are shown on the left; those for subject 5 are on the right, The upper panels show the levels of triacylglycerola, free fatty acids, and
cholesteryl esters. The lower panels show the corresponding lecithin: cholesterol acyltransferase activity (mean and range of duplicate
determinations) and the concentrations of HDL, phospholipids, and unesterified cholesterol. The infusion period is indicated by the
solid black bar along the abscissa.
the remainder cleared slowly, with a half-life greater than 18 h.
Serum levels of free fatty acids also increased to their maximum values by the end of the infusion, and subsequently became normal within 3 h (Fig. 10, upper panels). Because Intralipid does not contain free fatty acids, they were presumably produced by hydrolysis of the triacylglycerols.
The clearance kinetics of the triacylglycerols and free fatty acids are not directly relevant to this
article and will be described elsewhere (Ref. 38 and unpublished data).
As shown in the lower portion of Fig. 10, the serum concentration of phospholipids increased by
100 mg/dl in each subject. Approximately 50% of this excess lipid was eliminated during the follow- ing 4 h. Furthermore, despite the fact that Intrali- pid contains no sterols, in both subjects the level of serum cholesterol rose significantly during the infusion, and this elevation was sustained during
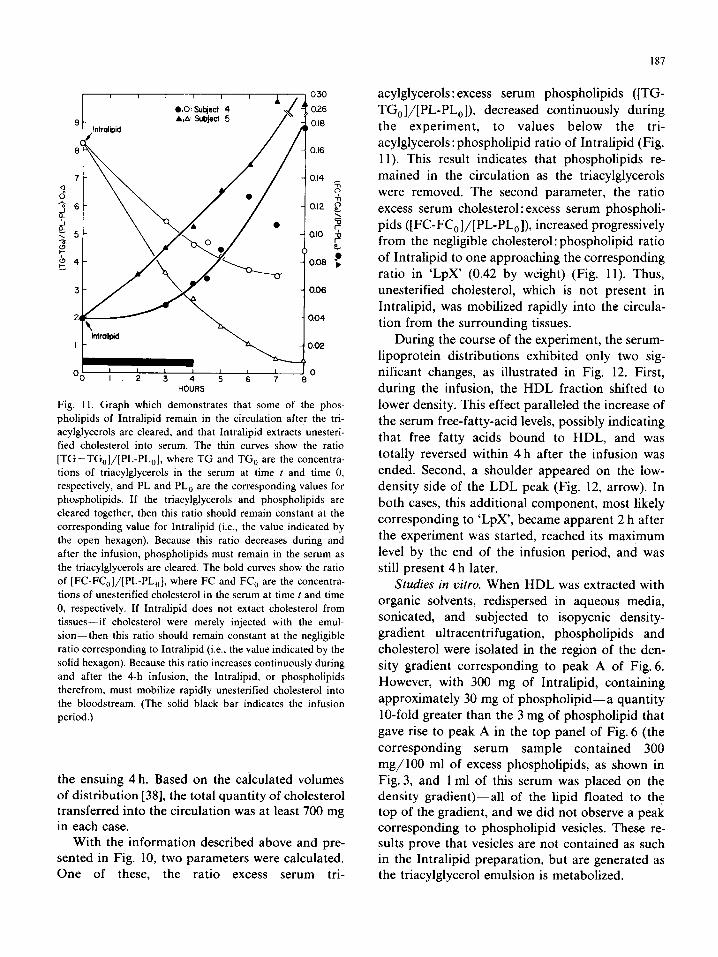
187
HOURS
Fig. I I. Graph which demonstrates that some of the phos-
pholipids of Intralipid remain in the circulation after the tri-
acylglycerols are cleared, and that Intralipid extracts unesteri-
fied cholesterol into serum. The thin curves show the ratio
[TG-TG,]/[PL-PL,], where TG and TG, are the concentra-
tions of triacylglycerols in the serum at time t and time 0.
respectively, and PL and PL, are the corresponding values for
phospholipids. If the triacylglycerols and phospholipids are
cleared together, then this ratio should remain constant at the
corresponding value for Intralipid (i.e., the value indicated by
the open hexagon). Because this ratio decreases during and
after the infusion, phospholipids must remain in the serum as
the triacylglycerols are cleared. The bold curves show the ratio
of [FC-FC,]/[PL-PL,], where FC and FC, are the concentra-
tions of unesterified cholesterol in the serum at time t and time
0, respectively. If Intralipid does not extact cholesterol from
tissues-if cholesterol were merely injected with the emul-
sion-then this ratio should remain constant at the negligible
ratio corresponding to Intralipid (i.e., the value indicated by the
solid hexagon). Because this ratio increases continuously during
and after the 4-h infusion, the Intralipid. or phospholipids
therefrom, must mobilize rapidly unesterified cholesterol into
the bloodstream. (The solid black bar indicates the infusion
period.)
the ensuing 4 h. Based on the calculated volumes of distribution [38], the total quantity of cholesterol transferred into the circulation was at least 700 mg in each case.
With the information described above and pre- sented in Fig. 10, two parameters were calculated. One of these, the ratio excess serum tri-
acylglycerols: excess serum phospholipids ([TG-
TG,]/[PL-PL,]), decreased continuously during
the experiment, to values below the tri-
acylglycerols: phospholipid ratio of Intralipid (Fig.
11). This result indicates that phospholipids re-
mained in the circulation as the triacylglycerols
were removed. The second parameter, the ratio
excess serum cholesterol : excess serum phospholi-
pids ([FC-FC,]/[PL-PL,]), increased progressively
from the negligible cholesterol: phospholipid ratio
of Intralipid to one approaching the corresponding
ratio in ‘LpX’ (0.42 by weight) (Fig. 11). Thus,
unesterified cholesterol, which is not present in
Intralipid, was mobilized rapidly into the circula-
tion from the surrounding tissues.
During the course of the experiment, the serum-
lipoprotein distributions exhibited only two sig- nificant changes, as illustrated in Fig. 12. First,
during the infusion, the HDL fraction shifted to
lower density. This effect paralleled the increase of
the serum free-fatty-acid levels, possibly indicating
that free fatty acids bound to HDL, and was
totally reversed within 4 h after the infusion was
ended. Second, a shoulder appeared on the low-
density side of the LDL peak (Fig. 12, arrow). In
both cases, this additional component, most likely
corresponding to ‘LpX’, became apparent 2 h after
the experiment was started, reached its maximum
level by the end of the infusion period, and was
still present 4 h later.
Studies in vitro. When HDL was extracted with
organic solvents, redispersed in aqueous media,
sonicated, and subjected to isopycnic density- gradient ultracentrifugation, phospholipids and
cholesterol were isolated in the region of the den-
sity gradient corresponding to peak A of Fig. 6.
However, with 300 mg of Intralipid, containing
approximately 30 mg of phospholipid-a quantity
IO-fold greater than the 3 mg of phospholipid that
gave rise to peak A in the top panel of Fig. 6 (the
corresponding serum sample contained 300
mg/lOO ml of excess phospholipids, as shown in
Fig. 3, and 1 ml of this serum was placed on the density gradient)-all of the lipid floated to the top of the gradient, and we did not observe a peak corresponding to phospholipid vesicles. These re- sults prove that vesicles are not contained as such in the Intralipid preparation, but are generated as the triacylglycerol emulsion is metabolized.
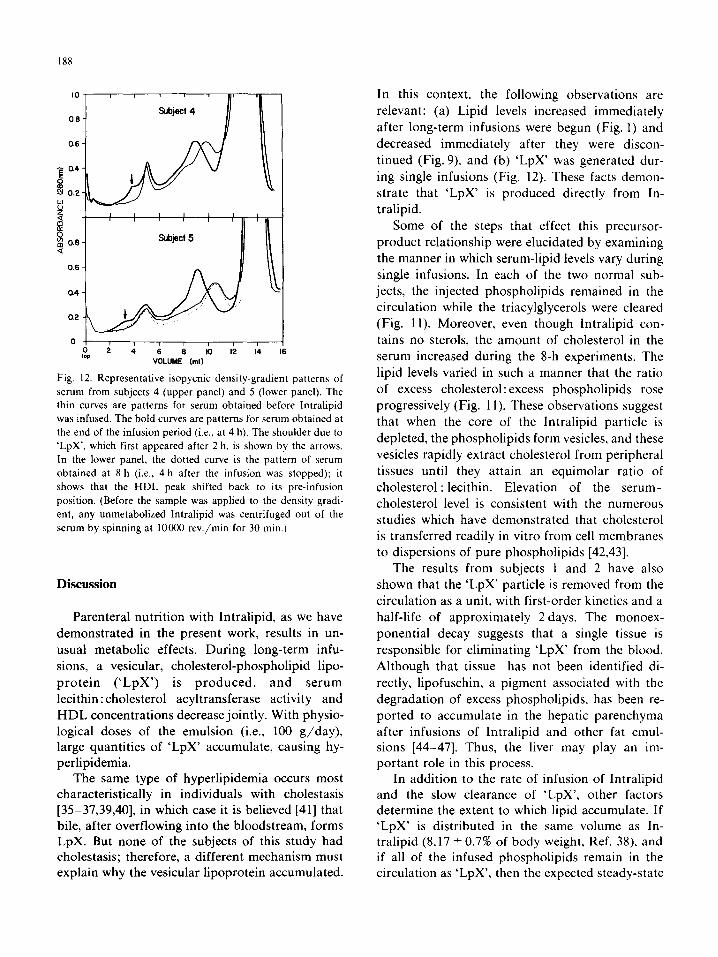
188
0.6 -
3 I I I I 1 I I
!$ 0.6- S&ct 5 Q
0.6 -
‘Q w
VOLUME (ml)
Fig. 12. Representative isopycnic density-gradient patterns of
serum from subjects 4 (upper panel) and 5 (lower panel). The
thin curves are patterns for serum obtained before Intralipid
was infused. The bold curves are patterns for serum obtained at
the end of the infusion period (i.e.. at 4 h). The shoulder due to
‘LpX’, which first appeared after 2 h. is shown by the arrows.
In the lower panel, the dotted curve is the pattern of serum
obtained at R h (i.e., 4 h after the infusion was stopped); it
shows that the HDL peak shifted back to its pre-infusion
position. (Before the sample was applied to the density gradi-
ent, any unmetabolized Intralipid was centrifuged out of the
serum by spinning at 10000 rev./min for 30 min.)
Discussion
Parenteral nutrition with Intralipid, as we have
demonstrated in the present work, results in un-
usual metabolic effects. During long-term infu-
sions, a vesicular, cholesterol-phospholipid lipo-
protein (‘LpX’) is produced, and serum lecithin : cholesterol acyltransferase activity and
HDL concentrations decrease jointly. With physio- logical doses of the emulsion (i.e., 100 g/day), large quantities of ‘LpX’ accumulate, causing hy- perlipidemia.
The same type of hyperlipidemia occurs most characteristically in individuals with cholestasis [35-37,39,40], in which case it is believed [41] that bile, after overflowing into the bloodstream, forms LpX. But none of the subjects of this study had cholestasis; therefore, a different mechanism must explain why the vesicular lipoprotein accumulated.
In this context, the following observations are
relevant: (a) Lipid levels increased immediately
after long-term infusions were begun (Fig. 1) and
decreased immediately after they were discon-
tinued (Fig. 9), and (b) ‘LpX’ was generated dur-
ing single infusions (Fig. 12). These facts demon-
strate that ‘LpX’ is produced directly from In- tralipid.
Some of the steps that effect this precursor-
product relationship were elucidated by examining
the manner in which serum-lipid levels vary during
single infusions. In each of the two normal sub-
jects, the injected phospholipids remained in the
circulation while the triacylglycerols were cleared
(Fig. 11). Moreover, even though Intralipid con-
tains no sterols, the amount of cholesterol in the
serum increased during the 8-h experiments. The
lipid levels varied in such a manner that the ratio
of excess cholesterol : excess phospholipids rose
progressively (Fig. 11). These observations suggest that when the core of the Intralipid particle is
depleted, the phospholipids form vesicles, and these
vesicles rapidly extract cholesterol from peripheral
tissues until they attain an equimolar ratio of
cholesterol : lecithin. Elevation of the serum-
cholesterol level is consistent with the numerous
studies which have demonstrated that cholesterol is transferred readily in vitro from cell membranes
to dispersions of pure phospholipids [42,43].
The results from subjects 1 and 2 have also shown that the ‘LpX’ particle is removed from the
circulation as a unit, with first-order kinetics and a
half-life of approximately 2 days. The monoex-
ponential decay suggests that a single tissue is
responsible for eliminating ‘LpX’ from the blood.
Although that tissue has not been identified di-
rectly, lipofuschin, a pigment associated with the
degradation of excess phospholipids, has been re-
ported to accumulate in the hepatic parenchyma after infusions of Intralipid and other fat emul- sions [44-471. Thus, the liver may play an im- portant role in this process.
In addition to the rate of infusion of Intralipid and the slow clearance of ‘LpX’, other factors determine the extent to which lipid accumulate. If ‘LpX’ is distributed in the same volume as In- tralipid (8.17 * 0.7% of body weight, Ref. 38), and if all of the infused phospholipids remain in the circulation as ‘LpX’, then the expected steady-state

Fig. 13. Suggested scheme for the metabolism of Intralipid. Shortly after Intralipid enters the circulation, the triacylglycerols are
hydrolyzed to free fatty acids and glycerol, which are in turn cleared rapidly (cf., Fig. I I). Between 60 and 90% of the phospholipids
are also removed from the plasma within several hours; the pathway for this process is at unknown present. The remainder of the
phospholipids form single-bilayer vesicles, which rapidly extract unesterified cholesterol from tissues until the molar ratio of
cholesterol : phospholipids reaches I : I These mixed vesicles, ‘LpX’ are cleared slowly from the circulation with monoexponential
first-order kinetics-the half-life is approximately 2 days. In particular, these mixed vesicles are not converted to other lipoproteins.
Evidence from the literature suggests that the hepatocyte may clear the ‘LpX’ from the blood, although there is no direct evidence to
support this possibility. For further explanation, see text. LCAT, lecithin : cholesterol acyltransferase: triglycerides, triacylglycerols.
level of excess phospholipids, PL,,, can be calcu-
lated with the formula [38,48]:
PL,, =gg, d
where Q is the rate of infusion of phospholipids
(in mg/day), [l/z is the clearance half-life of ‘LpX’
(in days), Vd is the volume of distribution (in dl), and PL,, is expressed in mg/dl. Although the ‘LpX’ was cleared at the same rate in subjects 1 and 2, the steady-state phospholipid levels were not proportional to the rate at which Intralipid was administered (Table1 and Figs. 3A and 4A), even after correcting for differences in V,. Morever, the serum-lipid levels were, in all cases, elevated by
less than 50% of the amount that would be ex-
pected on the basis of Eqn. 1. These observations,
together with the simple first-order clearance of all
of the excess lipids (Fig. 9), indicate that a signifi-
cant fraction of the phospholipids are cleared from
the circulation with a half-life of approximately
5 h. Using Eqn. 1 and the data given in Tables I, II
and III, the fraction of rapidly cleared phospholi-
pids was estimated to be 52, 80 and 85% in sub-
jects 1, 2 and 3, respectively. During the single infusions as well, between 40 and 60% of the excess phospholipids was cleared at a rate com- parable to, although slower than, that of the tri- acylglycerols (Fig. 10, lower panels). Thus, the degree of hyperlipidemia depends on the rate at which Intralipid is administered, on the slow rate

190
at which ‘LpX’ is cleared, and also on the fraction
of infused phospholipids that is transformed into
‘LpX’.
At present, we cannot draw firm conclusions
about the factors that determine the fraction of
slowly cleared phospholipids. Future studies must
elucidate whether that fraction as well as the half-
life of ‘LpX’ vary-for example, increase-during
the course of long-term parenteral nutrition, or
whether they are predetermined and constant in
each individual; these are the only alternatives
with which the current data are consistent. In the
first case, hyperlipidemia should develop during
the later stages of total parenteral nutrition; in the
latter case, hyperlipidemia, if it occurs, should
begin to develop immediately. The various possi-
bilities can only be resolved by studying serum-
lipid levels during the course of long-term infusion
periods. The data shown in Fig. 1 favor the latter
alternative, although more precise measurements
should be made in a larger number of subjects.
On the basis of the above-mentioned evidence,
the metabolism of Intralipid can be represented
schematically, as shown in Fig. 13. A principal
feature of this scheme is that ‘LpX’ does not serve
as a precursor for other lipoproteins. In subject 1,
for example, the excess phospholipids and
cholesterol were removed from the serum at the
same rate as the ‘LpX’ (Fig. 9) thereby suggesting
that net transfer of lipids, from ‘LpX’ to other
lipoproteins, did not occur. Moreover, as ‘LpX’ (peak A, Fig. 6) was cleared, additional peaks did
not appear in the density-gradient profiles, further
demonstrating that new complexes were not formed. Although, in subject 1, the concentrations
of HDL increased as ‘LpX’ was removed (Fig. 6)
these two processes did not occur in parallel
(Fig. 3). And in subject 2, the ‘LpX’ was also
cleared with a half-life of 2 days, but the HDL
levels remained low for at least 3 weeks after all of the abnormal lipoprotein was eliminated from the serum (Fig. 4). Thus, the HDL is resynthesized independently of the clearance of ‘LpX’.
The levels of HDL and the activity of
lecithin : cholesterol acyltransferase decline only during long-term infusions. Moreover, these quan- tities are not affected by incubating Intralipid with serum under various conditions in vitro (Ref. 38 and unpublished data). These findings demon-
strate that the enzyme is not directly inhibited,
and the lipoprotein particles are not disrupted
(e.g., by losing apolipoprotein A-I), by either In-
tralipid or its metabolites. Most likely, HDL and
lecithin : cholesterol acyltransferase are synthe-
sized, during total parenteral nutrition, more slowly
than usual. This explanation is consistent with the
notion that whereas chylomicrons donate lipids
and proteins to HDL, Intralipid does not. How-
ever, we cannot presently rule out other long-term effects on the small intestine and liver-organs
which are also involved in synthesizing HDL and
lecithin : cholesterol acyltransferase [ 12,13,49,50].
It is important to emphasize that the failure of
‘LpX’ to be converted to other lipoproteins, the slow rate at which ‘LpX’ was cleared in subjects 1
and 2, and the consequent hyperlipidemia in sub- jects 1 and 2, cannot reasonably be ascribed to the
modest reduction of lecithin : cholesterol
acyltransferase activity. In each of the three pa-
tients the activity was not sufficiently low to cause
any of the classical disturbances-such as virtual
absence of cholesteryl esters from serum, or accu-
mulation of large amounts of grossly abnormal,
discoidal HDL-that invariably ensue when the
enzyme activity falls below acceptable physiologi-
cal levels [51]. In subject 3, moreover,
lecithin : cholesterol acyltransferase activity was
moderately reduced during total parenteral nutri- tion (Table II), and hyperlipidemia did not occur.
Also, as demonstrated by Wengeler and Seidel [52] and Patsch [53], LpX (from patients with cholesta-
sis) is a poor substrate for lecithin: cholesterol
acyltransferase. It therefore seems reasonable to
conclude that the slow clearance of ‘LpX’ reflects
the properties of the vesicles themselves and is not
related to extrinsic factors such as the reduced
levels of HDL or lowered activity of
lecithin: cholesterol acyltransferase.
Because ‘LpX’ accumulates as an abnormal product when Intralipid is metabolized (Figs. 6 and 7), vesicles could not be similarly released from chylomicrons under physiological conditions. On the basis of other evidence, it has been sug- gested [54,55] that single-bilayer vesicles, such as ‘LpX’, may be one intermediate through which phospholipids are transferred post-prandially from chylomicrons to HDL: In vitro, single-bilayer vesicles can serve directly as substrates for

lecithin: cholesterol acyltransferase [56-581, or they
can first be converted to small protein-
phospholipid discs after acquiring apolipoprotein
A-l from HDL [59]. These processes can occur,
however, only if the molar ratio of choles-
terol : phospholipid is not larger than 1 : 2 [57,60].
‘LpX’ cannot undergo such physico-chemical
transformations and yield other lipoproteins,
probably because the initial phospholipid vesicles
rapidly acquire, in vivo, equimolar amounts of
unesterified cholesterol from a virtually unlimited
tissue pool (Figs. 11 and 13). This explanation is
supported by recent studies [61,62] on the fate in
vivo of cholesterol-containing vesicles. Future research efforts in this clinically and
theoretically important field must be directed to-
ward four general goals: establishing the precise
physical form in which surface components are
transferred from chylomicrons to HDL; finding
the long-term cause(s) for the joint reduction of
HDL levels and lecithin: cholesterol acyltransferase
activity; determining the kinetic parameters of the
phospholipids during the course of long-term total
parenteral nutrition with Intralipid; and preparing
other triacylglycerol emulsions in which the
surfactant is metabolizable rapidly and which could
therefore be administered without causing hyper-
lipidemia. The studies described in this article
have illustrated that excess surface is produced during the intravascular metabolism of at least
some lipid-laden particles. And the disposition of
those moieties, which solubilize and transport neu- tral lipids, is one important function fulfilled by
the intricate system of serum lipoproteins.
Acknowledgments
I wish to express my sincere appreciation to the
many colleagues who made this work possible.
Drs. Patricia McElroy and Joseph B. Kirsner sup-
plied most of the blood samples from the patients.
Mr. Roger Franz, Mrs. Rita Grammas and Mr.
Lance Lusk generously provided expert technical assistance in determining the serum lipid levels. Dr. Jiwhey Chung and Mrs. Darla Abano analyzed numerous samples for lecithin : cholesterol acyl- transferase activity. Mrs. Judy Swanson, R.N., as- sisted with the single infusions. I thank Drs. Ron Goldberg and Arthur Rubenstein for their interest
191
in this work and for carrying out radioimmunoas-
says on ‘LpX’. Drs. Ferenc J. KCzdy, Jayme Borensztajn, Godfrey S. Getz, Robert Josephs,
Marvin W. Makinen, Irwin H. Rosenberg, Gunther
M. Fless and Mrs. Celina Edelstein offered many
thoughtful comments and suggestions on various
versions of the manuscript. In addition, Mrs. Rose
Scott, Miss Sharron Riley, Mrs. Delores Walker
and Mrs. Julia Crawford provided excellent secre-
tarial assistance. Finally, I would like to thank the
patients as well as the normal subjects for their
genuine enthusiasm, limitless cooperation, and
profound interest in this work. These studies were
supported by Grants HL 18577 and HL 15026
from the United States Public Health Service, and
partially by Grant 78-09815 from the National
Science Foundation. The author is recipient of Medical Scientist Training Program Grant 5 T32
GM07281, from the National Institute of General
Medical Sciences, United States Public Health
Service.
References
I Schumaker, V.N. and Adams, G.H. (1979) Annu. Rev.
Biochem. 38, I 13- I36
2 Eisenberg, S. and Levy, R.I. (1975) Adv. Lipid Res. 13,
l-89
3 Schaefer, E.J., Eisenberg, S. and Levy, R.I. (I 978) J. Lipid
Res. 19,667-687
4 Robinson, D.S. (1970) in Comprehensive Biochemistry
(Florkin, M. and Stotz, E.H., eds.), Vol. 18, pp. 51-116,
5
6
7
8
9
IO
II
I2
13
14
I5
Elsevier, Amsterdam
Havel, R.J. (1957) J. Clin. Invest. 36, 848-854
Nichols, A.V., Rehnborg, C.S., Lindgren, F.T. and Wills,
R.D. (1962) J. Lipid Res. 3, 320-326
Havel, R.J., Kane, J.P. and Kashyap, M.L. (I 973) J. Clin.
Invest. 52, 32-38
Schumaker, V.N. and Adams, G.H. (I 970) J. Theoret. Biol.
26, 89-91
Marcel, Y.L. and Vezina, C. (1973) J. Biol. Chem. 248,
8254-8259
Wallentin, L. and Vikrot, 0. (1976) Stand. J. Clin. Lab.
Invest. 36, 473-479
Rose, H.G. and Juliano, J. ( 1977) J. Lab. Clin. Med. 89,
524-532
Tall, A.. Green, P., Abreu. E. and Glickman, P. (1978)
Circulation 58, (Suppl. II), II- I5 Redgrave, T.G. and Small, D.M. (1978) Circulation 58
(Suppl. II), II-14
Schaefer, E.J., Jenkins, L.L. and Brewer, H.B., Jr. (1978)
Biochem. Biophys. Res. Commun. 80, 405-412
Hansen, L.M., Hardie, W.R. and Hidalgo, J. ( 1976) Ann.
Surg. 184, 80-88

192
16 Broviac, J.W., Riella, M.C. and Scribner, B.H. ( 1976) Am.
J. Clin. Nutr. 29, 255-257
17 Jeejeebhoy, K.N., Anderson, G.H., Nakhooda, A.F.. Green-
berg, G.R.. Sanderson, I. and Marl&, E.B. (1976) J. Clin.
Invest. 57, 125-136
I8 Hallberg, D. (1965) Acta Physiol. Stand. Suppl. 65. 254
I9 Robinson, D.S. and Quarfordt. S.H. (1979) Lipids 14. 343-
349
20 Scow, R.O.. Hamosch, M., Blanchette-Mackie, E.J. and
Evans, A.J. (I 972) Lipids 7, 497-505
21 Blanchette-Mackie, E.J. and Scow, R.O. (1971) J. Cell Biol.
51, l-25
22 Foreman, J.R.. Karlin, J.B., Edelstein, C., Juhn. D.J..
Rubenstein. A.H. and Scanu, A.M. (1977) J. Lipid Res. IX,
759-767
23 Manual of Laboratory Operations, Lipid Research Clinics,
Vol I (Bethesda, MD), National Heart and Lung Institute,
1974)
24 Sperry, W.M. and Webb, M. (1950) J. Biol. Chem. 187,
97- 106
25 Franz, R.L. (1978) Clin. Chem. 24, 725-726
26 Bartlett, G.R. (1959) J. Biol. Chem. 234, 466-468
27 Mikac-Devic. D., Stankovic, H. and Boskovic. K. (1973)
Clin. Chem. Acta 45, 55-59
2X Sakagami, T. and Zilversmit. D.B. (1962) J. Lipid Res. 3,
111-116
29 Chung, J., Abano, D.A., Fless, G.M. and Scanu, A.M.
( 1979) J. Biol. Chem. 254, 7456-7464
30 Ohtsuki, M., Edelstein, C., Sogard, M. and Scanu, A.M.
(1977) Proc. Natl. Acad. Sci. U.S.A. 74. 5001-5005
31 Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall,
R.J. (1951) J. Biol. Chem. 193. 265-275
32 Karlin, J.B.. Juhn, D.J., Starr, J.I., Scanu. A.M. and Ruben-
stein, A.H. (1976) J. Lipid Res. 17, 30-37
33 Karlin, J.B., Juhn, D.J., Acanu, A.M. and Rubenstein, A.H.
(1978) Eur. J. Clin. Invest. 8, 19-26
34 Mao, S.J.T.. Gotto, A.M.. Jr. and Jackson, R.L. (1975)
Biochemistry 14, 4127-4131
35 Hamilton, R.L., Havel, R.J., Kane, J.P., Blaurock, A.E. and
Sata, T. (I 971) Science 172, 475-47X
36 Seidel, D.. Agostini, B. and Muller, P. (1972) Biochim.
Biophys. Acta 260. l46- I52
37 Patsch. J.R., Patsch. W., Sailer, S. and Braunsteiner. H.
( 1975) Biochim. Biophys. Acta 434, 4 19-427
38 Untracht, S.H. (1980) Ph.D. Thesis, The University of
Chicago, pp. 205-22X
39 Eder, H.A., Russ, E.M., Pritchett, R.A.R.. Wilber, M.M.
and Barr, D. (1955) J. Clin. Invest. 34. 1147-I 162
40 Russ, E.M., Raymont, J. and Barr. D.P. (1956) J. Clin.
Invest. 35. 133-144
41 Felker. T.E., Hamilton, R.L. and Havel, R.J. (197X) Proc.
Natl. Acad. Sci. U.S.A. 75, 3459-3463
42 Bruckdorfer K.R. and Graham, J.M. (1976) in Biological
Membranes (Chapman, D. and Wallach. D.F.H.. eds.) Vol.
3, pp. 103- 152, Academic Press, New York
43 Bruckdorfer, J.R., Edwards, P.A. and Green, C. (1968) Eur.
J. Biochem. 4, 506-5 I I
44 Koga, Y., Ikeda, K. and Inokuchi, K. (1975) Ann. Surg.
181, 186-190
45 Passwell, J.H., David, R., Katznelson, D. and Chone, B.E.
(1976) Arch. Dis. Child. 51, 366-36X
46 Van Haelst, U.J.G.M. and Sengers, R.C.A. (1976) Virchow
Arch. B. Cell Path. 22, 323-332
47 Friedman, Z.. Marks, K.H., Maisels, M.J.. Thorson, R. and
Naeye, R. (I 978) Pediatrics 6 I, 694-698
4X Wagner, J.G. (1975) Fundamentals of Clinical Pharmaco-
kinetics. Drug Intelligence Publications, Inc., Hamilton, IL
49 Hamilton, R.L.. Williams, M.C.. Fielding, C.S. and Havel,
R.J. (1976) J. Clin. Invest. 58, 667-680
50 Green, P.H.R.. Tall, A.R. and Glickman. R.M. (197X) J.
Clin. Invest. 6 I, 52X-534
5 I GJone, E. (1974) Stand. J. Clin. Lab. Invest. 33, Suppl. 137,
73-82
52 Wengeler. H. and Seidel. D. (1973) Clin. Chim. Acta 45.
429-432
53 Patsch, J.R. (1977) Eur. J. Clin. Invest. 7. 213-217
54 Tall, A.R. and Small, D.M. (1978) New Engl. J. Med. 299.
1232-1236
55 Patsch, W., Patsch, J.R., Kunz, F., Sailer, S. and Braun-
Steiner. H. (1977) Eur. J. Clin. Invest. 7, 523-530
56 Nakagawa, M. and Nishida. T. (1973) Biochim. Biophys.
Acta 296, 577-585
57 Fielding, C.J., Shore, V.G. and Fielding. P.E. Biochim.
Biophys. Acta 270, 513-518
5X Nichols, A.V. and Gong, E.L. (1971) Biochim. Biophys.
Acta 231, 175-184
59 Tall, A.R. and Small, D.M. (1977) Nature 265, 163-164
60 Tall, A.R. and Lange. Y. (1978) Biochem. Biophys. Rca.
Commun. 80. 206-212
61 Mauk. M.R. and Gamble, R.C. (I 979) Proc. Natl. Acad.
Sci. U.S.A. 76, 765-769
62 Gregoriadis, G. and Davis, C. (1979) Biochem. Biophys.
Res. Commun. 89. 12X7- I293
Related Documents











