J. Plant Res. 110: 195-207, 1997 Journal of Plant Research (~) by The Botanical Society of Japan 1997 Intraspecific Sequence Variation of Chloroplast DNA in Pedicularis chamissonis Steven (Scrophulariaceae) and Geographic Structuring of the Japanese "Alpine" Plants Noriyuki Fujii ~, Kunihiko Ueda 2, Yasuyuki Watano 2 and Tatemi Shimizu 2 1 Graduate School of Natural Science and Technology, Kanazawa University, Kakuma, Kanazawa, 920-11 Japan 2 Department of Biology, Faculty of Science, Kanazawa University, Kakuma, Kanazawa, 920-11 Japan In order to clarify evolutionary patterns and processes of intraspecific diversification of Pedicularis chamissonis Steven, we analyzed intraspaciflc variation of the nu- cleotide sequences of non-coding regions of chloroplast DNA: the intergenic spacers between trnT (UGU) and trnL (UAA) 5'exon, trnL (UAA) 3'exon and trnF (GAA), and atpB and rbcL. In 24 populations of P. chamissonis, 33 nucleotide substitutions and 12 insertions/deletions were inferred, and their genetic distances ranged from 0.001 to 0.014. Seventeen distinct cpDNA haplotypas could be recognized and each haplotype was found to be geographically structured. Two major cladas (the North- ern and Southern cladas) were revealed In phylogenetic analysas of cpDNA haplotypas. The heplotypas of the Northern clade had a wider distribution area in the popu- lations of Mts. Iide of central Honshu In Japan, northward to Unalaska Island in the Aleutians. Relationships among most haplotypes were unresolved polytomias. On the other hand, the haplotypas of the Southern clade occurred from the populations of Mt. Gassan southwards to Mt. Arakawa of central Honshu. Within this clade, three subclades were clearly recognized. From these results, we concluded that the haplotypas of the Northern and Southern cladas in P. charnissonis might have trav- eled down to Japanese Archipelago from the north in not a single glacial period. Key words: Chloroplast DNA-- Intraspecific variation m Molecular phylogeny-- Pedicularis chamissonis Phylogeography Soltis et al. (1992a) reviewed examples of intraspecific variation in chloroplast DNA (cpDNA) from a wide array of plant groups, and observed that the variability was often extensive. They discussed the utility of intraspecific cpDNA variation for studies of the multiple origins of polyploids, the origins of several crop species, the occur- rence of introgressive hybridizations, and so on. Such information, furthermore, allows the opportunity of phy- logeny reconstruction at the population level, while the geographic structuring of variation may provide insight into the historical biogeography of the species (Avise et al. 1987, Avise 1994, Soltis et al. 1992b). Many of the species constituting Japanese alpine flora are considered relics of those that had migrated, during the glacial periods, from the Asian or North American continents through land bridges to Japan. Although this scenario is often suggested in phytogeographical studies of the alpine flora in Japan (e.g., Koidzumi 1919, Kawano 1971, Toyokuni 1981), the geological evidence on which it is based (those from the Quaternary period) is too specu- lative and unorganized to promote more objective discus- sion. However, a phylogeographic study utilizing molec- ular data at the intraspecific level could greatly clarify the establishment of alpine flora in Japan. tn a previous study, we reported on the intraspecific sequence variation of Primula cuneifolia (Primulaceae) using the intergenic spacer between tmL (UAA) 3'exon and trnF (GAA) of cpDNA (Fujii et al. 1995). This species, an alpine meadow plant in Japan, occurs widely in the North Pacific coastal area (Fig. 1). Adding new sequence data on the previous study, we were able to recognize seven distinct cpDNA haplotypes in this species, each possessing geographic structure. Although two of these haplotypes from the northern regions had a wider distribu- tion area, the chloroplast genomes of the southern popu- lations from the central Honshu, Japan were further differ- entiated into five haplotypes. Pedicularis chamissonis Steven (Scrophulariaceae) is a perennial herb and a sunny meadow plant occurring in the subalpine and alpine areas in Japan (Fig. 2). This species has been classified into sect. Orthosiphonia H.L. Li, and is a unique taxon of the section in Japan. The distribution area of this species is similar to that of Primula cuneifolia (i.e., the North Pacific coastal area from the Japanese Archipelago northeastwards to southwest Alas- ka) (compare Figs. 1 and 2). Most populations are highly isolated into high mountains or islands, and several intraspecific taxa have been proposed. From their exter- nal morphology, and referring to the taxonomies of Ohwi (1978), Shimizu (1982), and Yamazaki (1993), we can rec- ognize five taxa: P. chamissonis vars. chamissonis, hok- kaidoansis T. Shimizu, rebunensis Yamazaki, japonica

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J. Plant Res. 110: 195-207, 1997 Journal of Plant Research (~) by The Botanical Society of Japan 1997
Intraspecific Sequence Variation of Chloroplast DNA in Pedicularis chamissonis Steven (Scrophulariaceae) and Geographic Structuring of the Japanese "Alpine" Plants
Noriyuki Fujii ~, Kunih iko Ueda 2, Yasuyuki Watano 2 and Tatemi Shimizu 2
1 Graduate School of Natural Science and Technology, Kanazawa University, Kakuma, Kanazawa, 920-11 Japan 2 Department of Biology, Faculty of Science, Kanazawa University, Kakuma, Kanazawa, 920-11 Japan
In order to clarify evolutionary patterns and processes of intraspecific diversification of Pedicularis chamissonis Steven, we analyzed intraspaciflc variation of the nu- cleotide sequences of non-coding regions of chloroplast DNA: the intergenic spacers between trnT (UGU) and trnL (UAA) 5'exon, trnL (UAA) 3'exon and trnF (GAA), and atpB and rbcL. In 24 populations of P. chamissonis, 33 nucleotide substitutions and 12 insertions/deletions were inferred, and their genetic distances ranged from 0.001 to 0.014. Seventeen distinct cpDNA haplotypas could be recognized and each haplotype was found to be geographically structured. Two major cladas (the North- ern and Southern cladas) were revealed In phylogenetic analysas of cpDNA haplotypas. The heplotypas of the Northern clade had a wider distribution area in the popu- lations of Mts. Iide of central Honshu In Japan, northward to Unalaska Island in the Aleutians. Relationships among most haplotypes were unresolved polytomias. On the other hand, the haplotypas of the Southern clade occurred from the populations of Mt. Gassan southwards to Mt. Arakawa of central Honshu. Within this clade, three subclades were clearly recognized. From these results, we concluded that the haplotypas of the Northern and Southern cladas in P. charnissonis might have trav- eled down to Japanese Archipelago from the north in not a single glacial period.
Key words: Chloroplast D N A - - Intraspecific variation m Molecular phylogeny-- Pedicularis chamissonis Phylogeography
Soltis et al. (1992a) reviewed examples of intraspecific variation in chloroplast DNA (cpDNA) from a wide array of plant groups, and observed that the variability was often extensive. They discussed the utility of intraspecific cpDNA variation for studies of the multiple origins of polyploids, the origins of several crop species, the occur- rence of introgressive hybridizations, and so on. Such information, furthermore, allows the opportunity of phy- logeny reconstruction at the population level, while the geographic structuring of variation may provide insight
into the historical biogeography of the species (Avise et al. 1987, Avise 1994, Soltis et al. 1992b).
Many of the species constituting Japanese alpine flora are considered relics of those that had migrated, during the glacial periods, from the Asian or North American continents through land bridges to Japan. Although this scenario is often suggested in phytogeographical studies of the alpine flora in Japan (e.g., Koidzumi 1919, Kawano 1971, Toyokuni 1981), the geological evidence on which it is based (those from the Quaternary period) is too specu- lative and unorganized to promote more objective discus- sion. However, a phylogeographic study utilizing molec- ular data at the intraspecific level could greatly clarify the establishment of alpine flora in Japan.
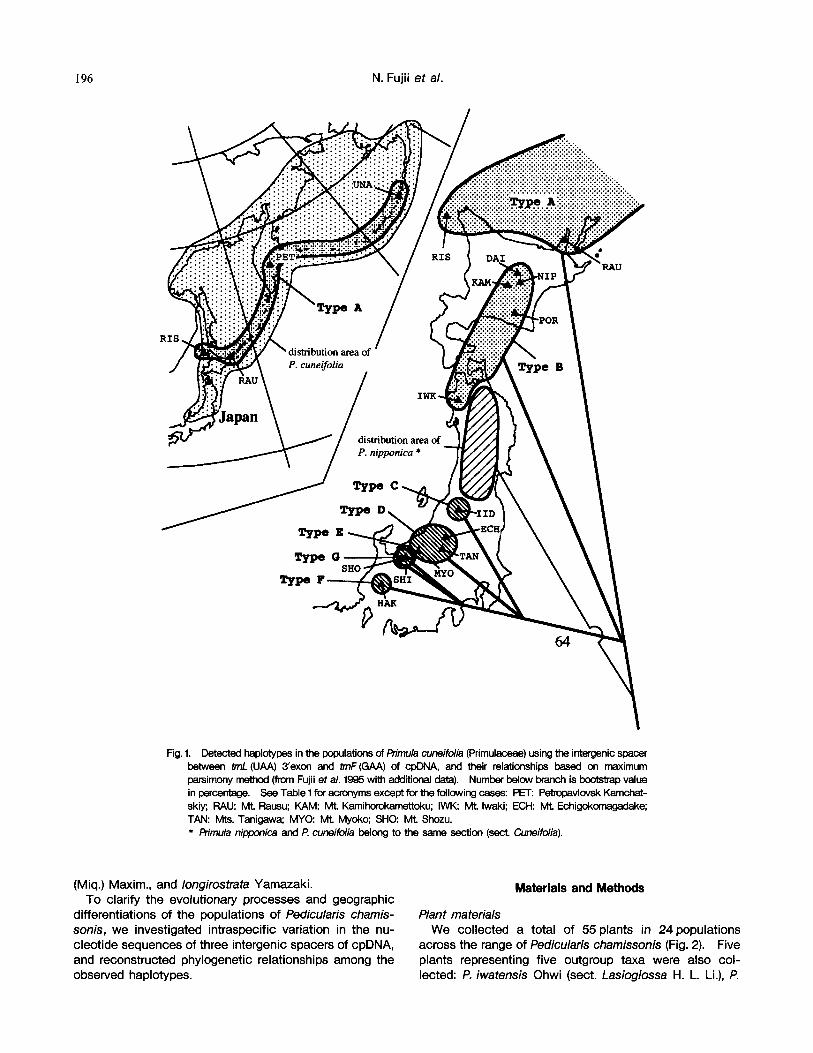
tn a previous study, we reported on the intraspecific sequence variation of Primula cuneifolia (Primulaceae) using the intergenic spacer between tmL (UAA) 3'exon and trnF (GAA) of cpDNA (Fujii et al. 1995). This species, an alpine meadow plant in Japan, occurs widely in the North Pacific coastal area (Fig. 1). Adding new sequence data on the previous study, we were able to recognize seven distinct cpDNA haplotypes in this species, each possessing geographic structure. Although two of these haplotypes from the northern regions had a wider distribu- tion area, the chloroplast genomes of the southern popu- lations from the central Honshu, Japan were further differ- entiated into five haplotypes.
Pedicularis chamissonis Steven (Scrophulariaceae) is a perennial herb and a sunny meadow plant occurring in the subalpine and alpine areas in Japan (Fig. 2). This species has been classified into sect. Orthosiphonia H.L. Li, and is a unique taxon of the section in Japan. The distribution area of this species is similar to that of Primula cuneifolia (i.e., the North Pacific coastal area from the Japanese Archipelago northeastwards to southwest Alas- ka) (compare Figs. 1 and 2). Most populations are highly isolated into high mountains or islands, and several intraspecific taxa have been proposed. From their exter- nal morphology, and referring to the taxonomies of Ohwi (1978), Shimizu (1982), and Yamazaki (1993), we can rec- ognize five taxa: P. chamissonis vars. chamissonis, hok- kaidoansis T. Shimizu, rebunensis Yamazaki, japonica
196 N. Fujii et al.
RIS
�9 . . . ~ .
: . : . : . : - : . : . : - :
A
distribution axca of P. cuneifolia
iiii!~iiiiii!iii!i!i!i!!i~'~ A iiiiiiiiiiii!ii!i!!!!!i!i!i!i!iiiiii::
DAI - ~ e
T y p e B
dis t r ibut ion a rea o f P. nipponica *
Type C ~ D \
Type Z ~ SHO"
64
Fig. 1. Detected haplotypes in the populations of Primula cuneifolia (Primulaceae) using the intergenic spacer between trnL (UAA) 3'exon and trnF (GAA) of cpDNA, and their relationships based on maximum parsimony method (from Fujii eta/. 1995 with additional data). Number below branch is beotstmp value in percentage. See Table 1 for acronyms except for the following cases: PET: Pebopavlovsk Kamchat- skiy; RAU: Mt. Rausu; KAM: Mt, K a m i ~ u ; IWK: Mt. Iwaki; ECH: Mt. Echigokomagadake; TAN: Mts, Tanigawa; MYO: Mt. Myoko; SHO: Mt. Shozu. * Pn'mula nipponica and P. cuneifolia belong to the same section (sect, Cuneifolia),
(Miq.) Maxim., and Iongirostrata Yamazaki . To clarify the evolut ionary processes and geographic
di f ferent iat ions of the populat ions of Pedicularis chamis- sonis, we investigated intraspeci f ic variat ion in the nu- c leot ide sequences of three intergenic spacers of cpDNA, and reconstructed phylogenet ic relat ionships among the observed haplotypes.
Materials and Methods
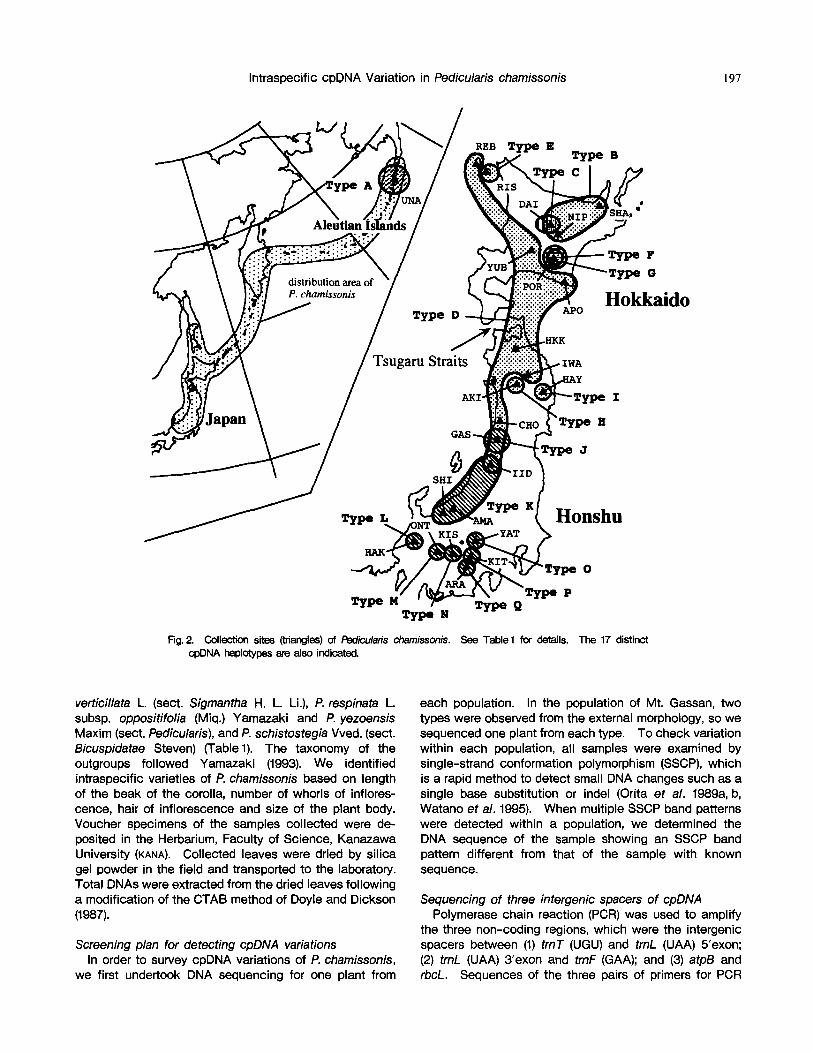
Plant materials We col lected a total of 55 plants in 24popu la t ions
across the range of Pedicularis chamissonis (Fig. 2). Five plants representing f ive outgroup taxa were also col- lected: P. iwatensis Ohwi (sect. Lasioglossa H. L. Li.), P.
Intraspecific cpDNA Variation in Pedicularis chamissonis 197
distribution area of P. chamissonis
T y p e D
~tpe B
~ B ~ L ~ e E [ ~ _ , . . x " Type s
iiiiiii! o Hokkaido .HKK
Fig. 2. Collection sites (triangles) of Pediculafis chamissonis. See Table1 for details. The 17 distinct cpDNA haplotypes are also indicated.
verticillata L. (sect. Sigmantha H. L. Li.), P. respinata L. subsp, oppositifolia (Miq.) Yamazaki and P. yezoensis Maxim (sect. Pedicularis), and P. schistostegia Vved. (sect. Bicuspidatae Steven) (Table1). The taxonomy of the outgroups followed Yamazaki (1993). We identified intraspecific varieties of P. chamissonis based on length of the beak of the corolla, number of whorls of inflores- cence, hair of inflorescence and size of the plant body. Voucher specimens of the samples collected were de- posited in the Herbarium, Faculty of Science, Kanazawa University (KANA). Collected leaves were dried by silica gel powder in the field and transported to the laboratory. Total DNAs were extracted from the dried leaves following a modification of the CTAB method of Doyle and Dickson (1987).
Screening plan for detecting cpDNA variations In order to survey cpDNA variations of P. chamissonis,
we first undertook DNA sequencing for one plant from
each population. In the population of Mt. Gassan, two types were observed from the external morphology, so we sequenced one plant from each type. To check variation within each population, all samples were examined by single-strand conformation polymorphism (SSCP), which is a rapid method to detect small DNA changes such as a single base substitution or indel (Orita et al. 1989a, b, Watano et al. 1995). When multiple SSCP band patterns were detected within a population, we determined the DNA sequence of the sample showing an SSCP band pattern different from that of the sample with known sequence.
Sequencing of three intergenic spacers of cpDNA Polymerase chain reaction (PCR) was used to amplify
the three non-coding regions, which were the intergenic spacers between (1) trnT (UGU) and trnL (UAA) 5'exon; (2) tmL (UAA) 3'exon and trnF (GAA); and (3) atpB and rbcL. Sequences of the three pairs of primers for PCR
198 N. Fujii e t al.
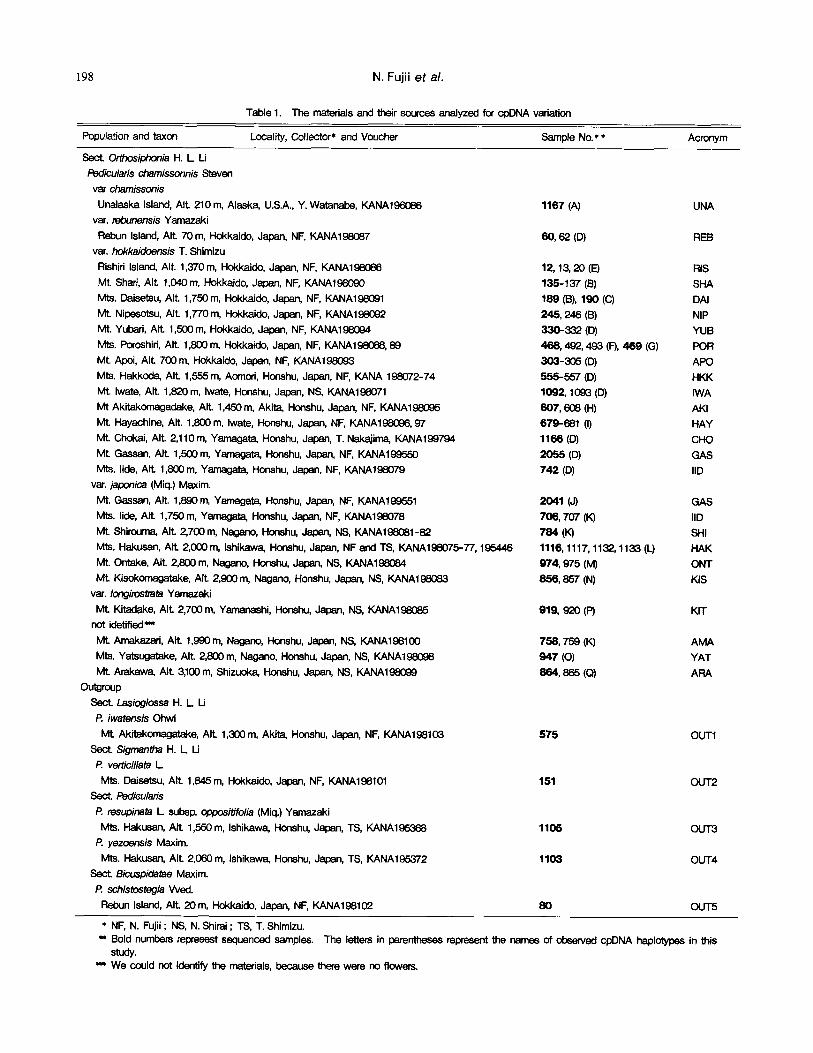
Table 1. The materials and their sources analyzed for cpDNA vanation
Population and taxon Locality, Collector* and Voucher Sample No. * * Acronym
Sect. Orttx~iphonia H. L U
Pedicularis chamissonnis Steven vat charnissonis
Unaleska Island, Air 210 m, Alaska, U.S.A., Y. Watanabe, KANA196066 1167 (A) var. rebunensis Yamazaki Rebun Island, AIt 70 m, Hokk~ido, Japan, NF, KANA196087
var. hokkaidoensis T. Shimizu Rishiri Island, AIt 1,370 m, Hokkaido, Japan, NF, KANA196086 Mt. Shad, Air. 1,040 m, Hokkaido, Japan, NF, KANA198090 Mrs. Dalsetsu, AIt 1,750 m, Hokkaldo, Japan, NF, KANA198091 IVE Nipesotsu, Air 1,770m, Hokkaldo, Japan, NF, KANA196092 Mt Yubari, AIt 1,500 m, Hokkaido, Japan, NF, KANA198094 Mls. Poroshiri, AIr 1,800 m, Hokkaido, Japan, NF, KANA198068, 89 Mr. Apoi, Air 700 m, Hokkaido, Japan, NF, KANA198093 Mrs. Hakkoda, AIt 1,555 m, Aornod, Honshu, Japan, NF, KANA 198072-74 Mt Iwate, Aft. 1,820 m, Iwate, Honshu, Japan, NS, KANA19(Y~)71 Mt Akitakomagadake, Aft. 1,450m, Akit~ I-kx~hu, Japan, NF, KANA198095 Iv~ Hayachine, Aft. 1,800 m, Iwate, Honshu, Japan, NF, KANA1960~6, 97 Mt ~ , AIt 2,110m, Yamagat~ Honshu, Japan, T. Nakajirr~ KANA199794 Mt Gessan, AIt 1,500 m, Yamagata, Honshu, Japan, NF, KANA199550 Mts. lide, AIt 1,800 m, Yamagata, Honshu, Japen, NF, KANA19807cj
var. j ~ i c a (Miq.) Maxim. Mt. Gassan, Air 1,890m, Yarnagata, Honshu, Japan, NF, KANA199551 Mrs. lide, AIt 1,750 m, Yamagata, Honshu, Japan, NF, KANA198078 Mt Shirourna, AIt 2,700 m, Nagano, Honshu, Japan, hiS, KANA196081-82 Mts. Hakusan, AIt 2,000 m, Ishikawa, Honshu, Japan, NFand TS, KANA196075-77, 195446 Mr. Ontake, AIt 2,800 m, Nagano, Hor~hu, Japan, NS, KANA196084 Mt. Kisokomagatake, AIt. 2,900 m, Nagano, Honshu, Japan, NS, KANA198083
var. Iongiros~'ata YarnazakJ
Mt Kitadake, AIt 2,700 m, Yamanashi, Honshu, Japan, hiS, KANA198085 not idetified =~
Mt Amakazad, AIt 1,990 m, Nagano, Hoc~hu, Japan, hiS, KANA198100 Mrs. Yatsugatake, Air 2,800 m, Nagano, Honshu, Japan, NS, KANA198(~8 Mt Arakawa, AIt 3,100 m, Shizuoka, Honshu, Japan, NS, KANA196099
Ou~oup Sect. Lasioglossa H. L U
P. iwatensis Ohwi
Ivlt Ak i takoma~e, AIt. 1,500 m, Akita, Honshu, J,~oan, NF, KANA198103 575 Sect. Sigmantha H. I l i
P. verticillata I
Mrs. Daisetsu, Aft. 1,845 m, Hokkaido, Japan, NF, KANA198101 151 Sect. Pedicu/aris
P. resupinata L subsp, opposilffolia (Miq.) Yamazaki Mrs. Hakusan, Aft. 1,550m, Ishikawa, Honshu, Japan, TS, KANA195368 1105
P. yezoensis Maxim. Mts. Hakusan, AlL 2,060 m, Ishikawa, Honshu, Japan, TS, KANA195372 1103
Sect. Bicuspidatae Maxim. P. schistostegia Vved.
Rebun Island, AIL 20 m, Hokkaido, Japan, NF, KANA19610~ 80
UNA
60, 62 (D) REB
12, 13, 20 (E) RIS 135-137 (B) SHA 189 (13), 190 (C) DAI 245, 246 (B) NIP 330-332 (D) YUB 468, 492, 493 (19, 459 (G) POR 303-305 (D) APO 555-567 (D) HKK 1092, 1093 (D) IWA 607, 606 (H) AKI 679-681 (I) HAY 1166 (D) CHO 2055 (D) GAS 742 (D) lid
2041 (J) GAS 706, 707 (K) liD 784 (K) SHI 1116, 1117, 1132, 1133 (L) HAK 974, 975 (M) ONT 856, 857 (N) KIS
919, 920 (P) KIT
758, 759 (19 AMA 947 (0) YAT 864,865 (Q) ARA
OUT1
OUT2
OUT3
OUT4
OUT5
* NF, N. Fujii ; hiS, N. Shirai ; TS, T. Shimizu. " Bold numbers represest sequenced samples. The letters in parentheses represent the names of obsewed cpDNA haplotypes in this
s~dy. m We could not identify the materials, because tt~re were no flowers.
Intraspecific cpDNA Variation in Pedicularis chamissonis
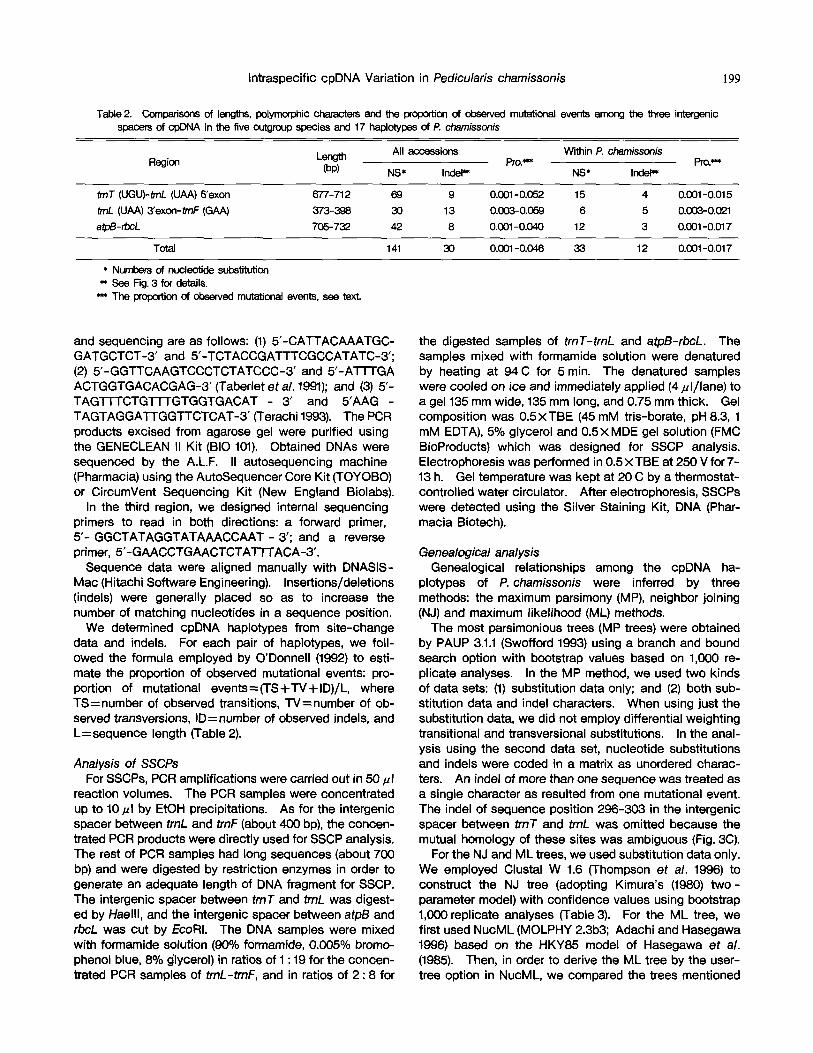
Table 2. Comparisons of lengths, polymorphic characters and the proportion of observed mutational events among the three intergenic spacers of cpDNA in the five outgroup species and 17 haplotypes of P. ~issonis
199
Region Length All accessions Within P. chamissonis Pro.*** Pro. =~
(bp) NS* IndeP* NS* In(let**
trnT (UGU)-trnL (UAA) 5'exon 677-712 69 9 0.001-0.052 15 4 0.001-0.015 trnL (UAA) 3'exon-trnF (GAA) 373-398 30 13 0.003-0.059 6 5 0.003-0.0~1 atpB-rbc.L 705-732 42 8 0.001-0.040 12 3 0.001-0.017
Total 141 30 0.001-0.046 33 12 0.001-0.017
* Numbers of nucleotide subs'dtution ** See Fig. 3 for details.
~'~ The proportion of obeefved mutational events, see text.
and sequencing are as follows: (1) 5'-CATTACAAATGC- GATGCTCT-3' and 5'-TCTACCGATTTCGCCATATC-3'; (2) 5'-GGTTCAAGTCCCTCTATCCC-3' and 5'-ATTTGA ACTGGTGACACGAG-3' (Taberlet et al. 1991); and (3) 5'- TAGTTTCTGTTTGTGGTGACAT - 3' and 5'AAG - TAGTAGGATTGGTTCTCAT-3' (Terachi 1993). The PCR products excised from agarose gel were purified using the GENECLEAN II Kit (BIO 101). Obtained DNAs were sequenced by the A.L.F. II autosequencing machine (Pharmacia) using the AutoSequencer Core Kit (TOYOBO) or CircumVent Sequencing Kit (New England Biolabs).
In the third region, we designed internal sequencing primers to read in both directions: a forward primer, 5'- GGCTATAGGTATAAACCAAT - 3'; and a reverse primer, 5'-GAACCTGAACTCTATTTACA-3'.
Sequence data were aligned manually with DNASIS- Mac (Hitachi Software Engineering). Insertions/deletions (indels) were generally placed so as to increase the number of matching nucleotides in a sequence position.
We determined cpDNA haplotypes from site-change data and indels. For each pair of haplotypes, we foll- owed the formula employed by O'Donnell (1992) to esti- mate the proportion of observed mutational events: pro- portion of mutational events=(TS+TV+lD)/L, where TS=number of observed transitions, TV=number of ob- served transversions, ID=number of observed indels, and L=sequence length (Table 2).
Analysis of SSCPs For SSCPs, PCR amplifications were carried out in 50 #1
reaction volumes. The PCR samples were concentrated up to 10/zl by EtOH precipitations. As for the intergenic spacer between tmL and trnF (about 400 bp), the concen- trated PCR products were directly used for SSCP analysis. The rest of PCR samples had long sequences (about 700 bp) and were digested by restriction enzymes in order to generate an adequate length of DNA fragment for SSCP. The intergenic spacer between troT and trnL was digest- ed by Haelll, and the intergenic spacer between atpB and rbcL was cut by EcoRl. The DNA samples were mixed with formamide solution (90% forrnamide, 0.005% bromo- phenol blue, 8% glycerol) in ratios of 1 : 19 for the concen- trated PCR samples of trnL-tmF, and in ratios of 2 : 8 for
the digested samples of trnT-trnL and atpB-rbcL. The samples mixed with formamide solution were denatured by heating at 94 C for 5 min. The denatured samples were cooled on ice and immediately applied (4 #l/lane) to a gel 135 mm wide, 135 mm long, and 0.75 mm thick. Gel composition was 0.5XTBE (45 mM tris-borate, pH 8.3, 1 mM EDTA), 5% glycerol and 0.5• MDE gel solution (FMC BioProducts) which was designed for SSCP analysis. Electrophoresis was performed in 0.5 X TBE at 250 V for 7- 13 h. Gel temperature was kept at 20 C by a thermostat- controlled water circulator. After electrophoresis, SSCPs were detected using the Silver Staining Kit, DNA (Phar- macia Biotech).
Genealogical analysis Genealogical relationships among the cpDNA ha-
plotypes of P. chamissonis were inferred by three methods: the maximum parsimony (MP), neighbor joining (N J) and maximum likelihood (ML) methods.
The most parsimonious trees (MP trees) were obtained by PAUP 3.1.1 (Swofford 1993) using a branch and bound search option with bootstrap values based on 1,000 re- plicate analyses. In the MP method, we used two kinds of data sets: (1) substitution data only; and (2) both sub- stitution data and indel characters. When using just the substitution data, we did not employ differential weighting transitional and transversional substitutions. In the anal- ysis using the second data set, nucleotide substitutions and indels were coded in a matrix as unordered charac- ters. An indel of more than one sequence was treated as a single character as resulted from one mutational event. The indel of sequence position 296-303 in the intergenic spacer between tmT and trnL was omitted because the mutual homology of these sites was ambiguous (Fig. 3C).
For the NJ and ML trees, we used substitution data only. We employed Clustal W 1.6 (Thompson et al. 1996) to construct the NJ tree (adopting Kimura's (1980) two- parameter model) with confidence values using bootstrap 1,000 replicate analyses (Table 3). For the ML tree, we first used NucML (MOLPHY 2.3b3; Adachi and Hasegawa 1996) based on the HKY85 model of Hasegawa et al. (1985). Then, in order to derive the ML tree by the user- tree option in NucML, we compared the trees mentioned

200 N. Fujii et al.
~Du~o E~ �9
�9 -i ~o o
..-i m r,~
~ m m
~ m o
,.-i e~l e.. �9 .-i N ~ ,.~ e , i m
~ r ~
..-4 ~ o
..-i o ~
~1, r.-
- o m
~ r n ~ m m m
m m ~ l
m o o ~
m o e - .
m o m m o , , 0
m , ~ , m
m m r ~ e ~ m ~
~ r ~ o N ~ o ~
e, i ~ e , i e , l ~ ..-i
N m m
: t
!!::
!ii!iii::::
: : : : : r r r : r
mm~)
ma)~ ,~ a) in
m m ~
mn1~
m m ~ m ~ o m ~ a ~ m e , ~ m m ~ i r-.
~ e m o
,.-t , - t o
, - i o ~ ~ o r - -
~ o ~ 1 ,
*..i 0 eli ~ o , - i , . 4 O O
O~ID
0 ~ 1 0 ~ , - t o ~ o
0 ~ m
iiiiii!
e ~ , e ~ e
gg~g~gg
it~o
~i,i ~.
I !
I I I
0
if'
I i I
~i::i!~ii i
H
iT.

Intraspecific cpDNA Variation in Pedicularis chamissonis
Table 3. Genetic distance me, ix*
201
A B C D E F H J K L 0 P Q OUT1 OUT2 OUT3 OUT4
Type A Type B .O05 Type C .005 .001 Type D .002 .003 ,003 Type E .002 .003 .004 .001 Fype F .001 .003 .004 .001 .001 Type H .002 .003 .004 .001 .001 .001 Type J .009 .012 .013 .009 .009 .009 .009 Type K .008 .012 .012 .009 .009 .008 .009 .001 Type L .039 .012 .013 .009 .009 .009 .010 .007 Type 0 .009 .012 .013 .009 .009 .009 .010 .007 Type P .009 .013 .014 .010 .010 .009 .010 .008 Type Q .009 .013 .014 .010 .010 .009 .010 .008 OUT1 .032 .036 .036 .033 .033 .032 .033 .033 OUT2 .031 .035 .035 .032 .032 .031 .033 .032 OUT3 .002 .027 .007 .004 .004 .003 .024 .024 OUT4 .006 .030 ,030 .027 .027 .026 .027 .027 OUT5 .022 .005 .026 .002 .022 .002 .003 .001
.006
.006 .002
.007 .003 .001
.007 .003 .001
.032 .032 .032
.031 .032 .032
.003 .023 .023
.006 .026 .026
.020 .000 .020
.001
.033 .033
.033 .033
.023 .023
.026 .026
.020 .020
.039
.008 .033
.009 .034 .010
.025 .030 .013 .014
For names of cpDNA haplotypes, see Fig. 1. For acronyms of outgroups, see Table 1. Type D includes Type G and I, end Type L includes Type M and N, because they did not show nucleotide substitution, respectively. * Genetic distances are based on Kimura's (1980) two-parameter model.
above, the NJ tree and 45 MP trees obtained from com- pletely dissolving the polychotomous branching of the trees into strict dichotomies. In this calculation, we used the weight between transition and transversion derived from the tree optimization option in NucML (~//~ rat io= 1.968).
Results
Intraspecific cpDNA variation of P. chamissonis We first determined the nucleotide sequences of three
non-coding regions of cpDNA for 25 plants from 24 popu- lations. In this manner, 15 cpDNA haplotypes were dis- tinguished. In the population of Mt. Gassan, we detected intra-populational cpDNA variation (Nos. 2041 and 2065). Furthermore, SSCP analysis for all samples revealed intra- populational variation in three populations: Mts. Daisetsu, Mts. Poroshiri and Mts. lide (Fig. 4). Additional DNA sequencing was undertaken for sample Nos. 190, 469 and 742, and showed SSCP band patterns different from those of the samples with known sequences, Finally, we could recognize 17 distinct cpDNA haplotypes in P. chamissonis (Fig. 2 and Table 1). The cpDNA haplotypes were named alphabetically, the most northerly distributed one as Type A, the second one as Type B, and so on. None of the haplotypes exhibited random distribution; all were found to be geographically structured (Fig. 2).
Nucleotide sequences of three intergenic spacers of cpDNA
In all, we sequenced 28 plants of P. chamissonis and
five outgroup taxa in Pedicularis. The determined parts of the sequences between troT and tmL, and between atpB and rbcL, were wholly non-coding region; however, the intergenic spacer between trnL and tmF included 24 bp of the coding region of trnF. The nucleotide sequence data reported in this paper will appear in the DDBJ, EMBL and GenBank DNA databases under accession numbers D88038 to D88105, and D88774 to D88776.
The length of the intergenic spacer between trnT and trnL varied from 677-712 bp, that between trnL and tmF varied from 373-398 bp, and that between atpB and rbcL varied from 705-732bp (Table2). The lengths after multiple alignment of the sequences were 743, 431 and 744 bp, respectively.
The total numbers of polymorphic characters in the three spacers were 141 nucleotide substitutions and 30 indels among all accessions, and 33 nucleotide substitu- tions and 12 indels within P. chamissonis (l 'able 2 and Fig. 3). P. verticillate, P. respinata subsp, oppositifolia, P. schistostegia, P. iwatensis, and P. yezoensis were distin- guished from Haplotype A of P. chamissonis by 55 nu- cleotide substitutions and 11 indels, 41 site changes and 12 indels, 38 nucleotide substitutions and 11 indels, 57 site changes and 13 indels, and 48 nucleotide substitutions and 10 indels, respectively.
We determined the proportion of observed mutational events among all accessions, including the outgroups (Table 2). The total values among all accessions ranged from 0.001-0.046, and the intraspecific values of P. chamissonis ranged from 0.001-0.017. We compared the values among the three intergenic spacers. Though

202 N. Fujii et ah
Fig. 4. SSCP band patterns of P. chamissonis in the populations of Mrs. Daisetsu, Mts. Poroshiri and Mts. lide. Sample number of each lane corresponds to that of Table 1. The first and second lanes represent the PCR products of the intergenic spacer between atpB and rbcL digested by EcoRI; the other lanes represent the PCR products of the intergenic spacer between/TnT (UGU) and/TnL (UAA) 5'exon digested by Haelll.
each region indicated similar values, those of the inter- genic spacer between trnL and trnF were slightly higher.
Genealogical analysis We first constructed MP, NJ and ML trees based solely
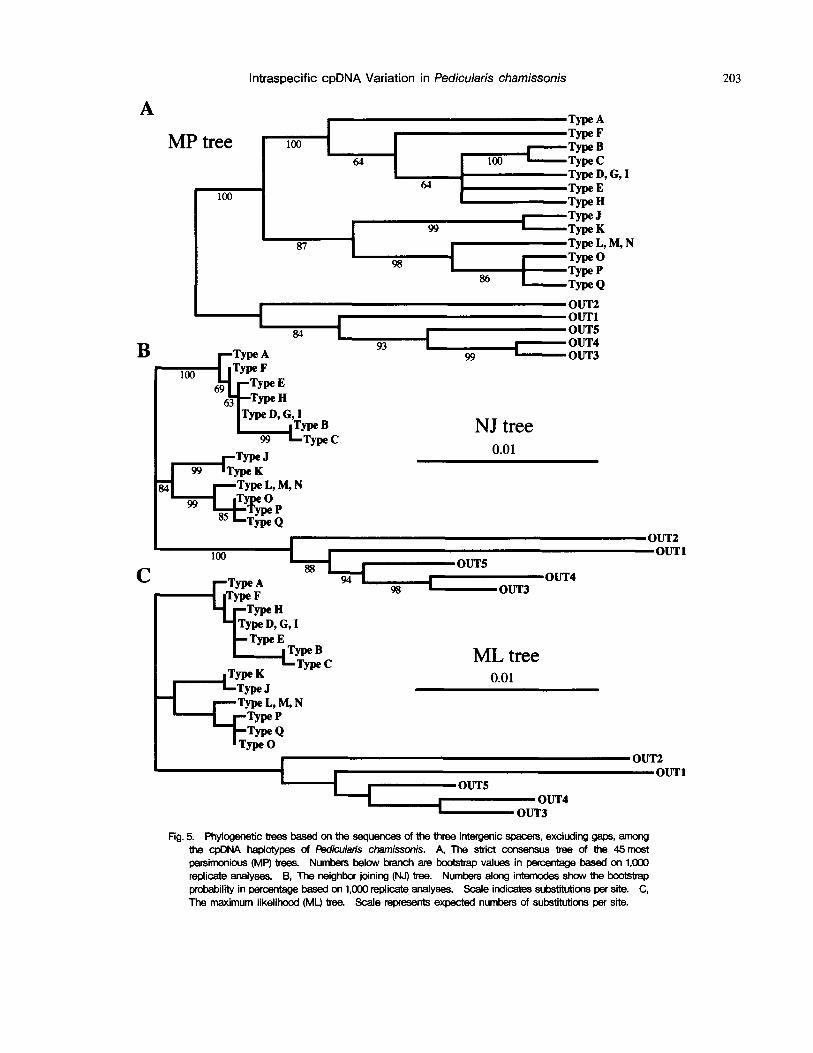
on site-change data for the outgroup species and 17 cpDNA haplotypes of P. chamissonis (Fig. 5). In MP analysis, 45 MP trees were obtained, which required 156 steps; Consistency Index including uninformative charac- ters (CI)=0.936, Retention Index (RI)=0.940. The strict consensus tree of the 45 is presented in Fig. 5A. NJ analysis was performed using the data shown in Table 3. The range of genetic distance among all accessions was 0.001-0.039, and the intraspecific distance of P. chamis- sonis ranged from 0.001-0.014. The NJ tree based on these data is depicted in Fig. 5B. The ML tree is shown in Fig. 5C. Though the topologies of the three trees were partially different from one another, all the trees revealed the same fundamental topology.
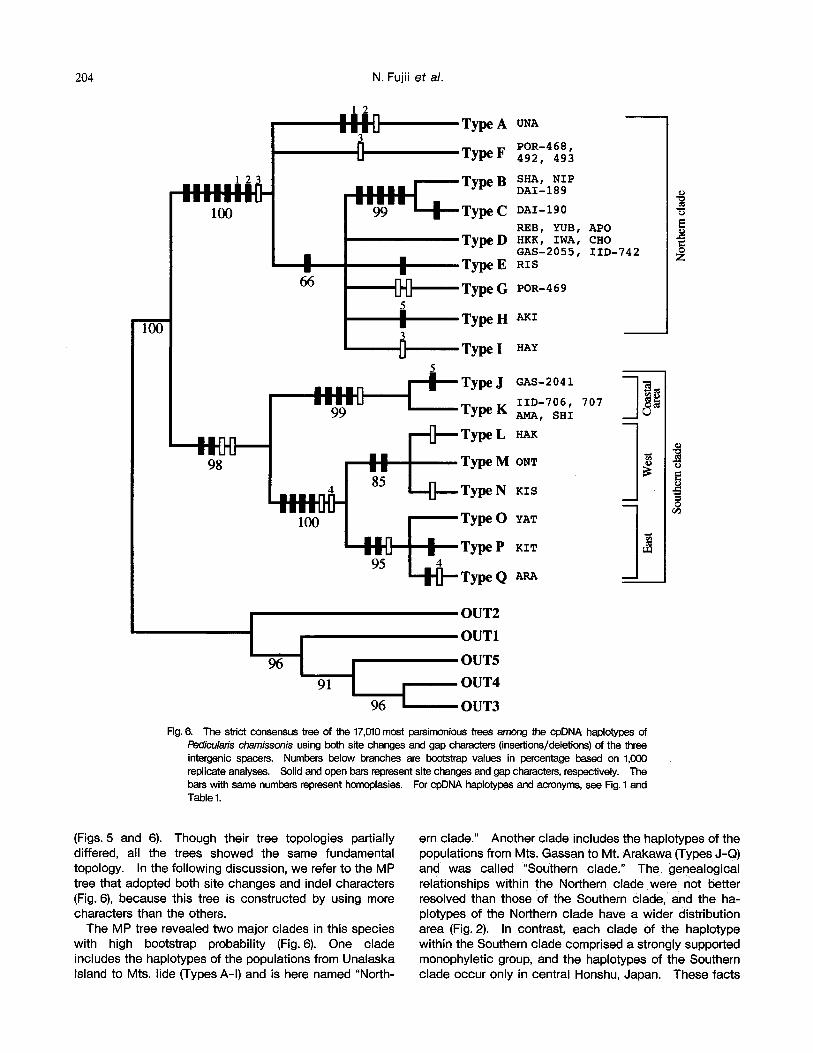
In MP analysis adopting both site changes and indel characters, 17,010MP trees were obtained using the branch and bound search option. The trees required 203 steps, with CI of 0.926 and RI of 0.939. The strict consensus tree of the 17,010 is presented in Fig. 6. The tree shows that the topology of haplotypes from Type J-Q was the same as that of the above trees (Fig. 5), however, the clade of haplotypes from Type A-I had unresolved trichotomies.
The MP tree shows the following genealogical relation- ships of cpDNA haplotypes (Fig. 6). (1) The haplotypes from the populations of P. chamissonis were monophyletic with a bootstrap value of 100% against the outgroup species. (2)The haplotypes of the populations from
northern Honshu in Japan to Unalaska Island in the Aleutians (Types A-I) made a clade supported by a 100% confidence value. The haplotypes of the populations of Hokkaido and northern Honshu (Types B-I excluding Type F) constituted a single clade with 66% bootstrap probabil- ity. The haplotypes of the populations of eastern Hok- kaido (Types B and C) comprised a strongly supported monophyletic group (99%). (3)The haplotypes of the populations from central Honshu, Japan (Types J-Q) constituted a single clade with a bootstrap value of 98%. Within this clade, the haplotypes of the populations from Mt. Gassan to Mt. Shirouma (Types J and K) made a clade with 99% probability, and those from Mts. Hakusan to Mt. Arakawa (Types L-Q) were supported in all bootstrap replicates. Furthermore, the haplotypes of the popula tions from Mts. Hakusan to Mt. Kisokomagatake (Types L- N) constituted a single clade (85%), and the haplotypes of the populations from Mts. Yatsugatake to Mt. Arakawa (Types O-Q) made another single clade supported by a 95% confidence value. (4)P. verticillata is a sister species to the other four species with a bootstrap value of 96%. P. respinata subsp, oppositifolia, P. yezoensis and P. schistostegia constituted a single clade with 91% prob- ability. P. respinata subsp, oppositifolia and P. yezoensis comprised a monophyletic group (96%).
Discussion
Molecular phylogeny and evolution of P. chamissonis The extensive cpDNA diversity in P. chamissonis
affords an opportunity for phylogeny reconstruction at the population level. In this study, we constructed MP, NJ and ML trees among haplotypes of the present species
In t raspec i f i c c p D N A Var ia t ion in Pedicular is chamisson is 203
A
B
C
I
p
M P tree
100
100 I i I I 100
99
86
I 99
98
100
99
99
I
i I 84
6963~T ~ypeType A
Type F ype E ypeH
D, G, I nType B
99 L-Type C
~Type Type J K
[----Type L, M, N ~
~ e %~'E'rype Q
100 [ I 88 I
~ T TyType A ypeF Type H
D, G, I ype 17
Type B "--Type C
ypeJ
93
NJ tree 0.01
i OUT5 94 ! OUT4
9~ ; ou'r3
M L tree 0.01
Type A Type F Type B Type C Type D, G, I ' l 'y~ E Type H TypeJ Type K Type L, M, N Type O Type P Type Q
OUT2 OUTI OUTS OUT4 OUT3
r ==- Type L, M, N
l ~ T Type P Type Q ype O
u OUT5 OUT4
= ~ OUT3
OUT2 OUTI
OUT2 OUTI
Fig. 5. Phylogenetic bees based on the sequences of the ~ree intergenic spacers, excluding gaps, among the cpDNA haplotypes of Pedicularis chamissonis. A, The sbict consensus tree of the 45 most parsimonious (MP) trees. Numbers below bcanch are 10ootstrap values in percentage based on 1,000 replicate analyses. B, The neighbor joining (NJ) tree. Numbem along intemock~ show the I:x:x~tstrap probability in 13ercentage based on 1,000 replicate analyses. Scale indicates substitutions per site. C, The maximum likelihood (ML) tree. Scale represents expected numbers of s u b s ~ per site.
204 N. Fujii et al.
_J 100
98
1 2 l l ,m~ Type
D Type
|
66
••. Type
Type
Type
�9 Type
Type G 5 I Type H 3
Type I 5
A UNA
POR-468, F 492, 493
B SHA, NIP DAI-189
C DAI-190
REB, YUB, D HKK, IWA,
GAS-2055, E RIS
POR-469
AKI
HAY
•••--• Type J
Type K
.__-.fL.m Type L �9 �9 [ u �9 �9 Type M ONT
85 ~-- Type N KIS
I00 I ___ [ TypeO YA~
=44
GAS-2041
IID-706, AMA, SHI
HAK
APO CHO
707
IID-742
1, 1,
96 91
OUT2
OUT1
OUT5
OUT4
OUT3 96 Fig. 6. The strict consensus tree of the 17,010 most parsimonious trees among the cpDNA haplotypes of
Pedicularis chamissonis using both site changes and gap characters (insertions/deletions) of the three intergenic spacers. Numbers below branches are bootstrap values in percentage based on 1,(X)0 replicate analyses. Solid and open bars represent site changes and gap characters, respectively. The bars with same numbers represent homoplasies. For cpDNA haplotypes and acronyms, see Fig. 1 and Table 1.
:
"5 E
o Z
c.)
E
O
(Figs. 5 and 6). Though their tree topologies partially differed, all the trees showed the same fundamental topology. In the following discussion, we refer to the MP tree that adopted both site changes and indel characters (Fig. 6), because this tree is constructed by using more characters than the others.
The MP tree revealed two major clades in this species with high bootstrap probability (Fig. 6). One clade includes the haplotypes of the populations from Unalaska Island to Mts. lide (Types A-I) and is here named "North-
ern clade." Another clade includes the haplotypes of the populations from Mts. Gassan to Mt. Arakawa (Types J-Q) and was called "Southern clade." The genealogical relationships within the Northern clade were not better resolved than those of the Southern clade, and t h e ha- plotypes of the Northern clade have a wider distribution area (Fig. 2). In contrast, each clade of the haplotype within the Southern clade comprised a strongly supported monophyletic group, and the haplotypes of the Southern clade occur only in central Honshu, Japan. These facts

Intraspecific cpDNA Variation in Pedicularis chamissonis 205
suggest that the haplotypes of these two major lineages were a result of different evolutionary processes, and perhaps the Southern clade is older than the Northern clade (see Fig. 5B, C).
SSCP and sequencing analyses revealed intra- populational variations in the populations of Mt. Gassan and Mts. lide (Figs. 2 and 4). Two cpDNA haplotypes (2041, Type J; and 2055, Type D) could be recognized in the population of Mt. Gassan, and two haplotypes (742, Type D; and 706 and 707, Type K) also could be observed in the population of Mts. lide. Therefore, the haplotypes of both Northern and Southern clades were recognized in the Northern Honshu, Japan. The Mt. Gassan population is the southernmost population in the Northern clade, and the Mts. lide population is the northernmost population in the Southern clade.
The Northern clade was a strongly supported mono- phyletic group (Fig. 6). This result indicates that the haplotypes of the Northern clade diverged from a common ancestral genome. The haplotypes of the Northern clade, excluding Haplotypes A and F, were monophyletic, although the confidence value was somewhat low (66%). These haplotypes from the populations of Hokkaido and northern Honshu, Japan might have diverged from a common ancestor. The haplotypes of the populations of Mts. Daisetsu, Mt. Nipesotsu and Mt. Shari (Types B and C) possess many more autapomorphies than the other haplotypes of the Northern clade (Fig. 6). This result might mean that the haplotypes of eastern Hokkaido have a unique evolutionary history in the Northern clade. Although the haplotypes of the Northern clade have wider distribution (Fig. 2), the genetic distances among the haplotypes of this clade were significantly smaller than those of the Southern clade (Mann & Whitney's U-test, P<0.001). The genetic distances among the haplotypes of the Northern clade ranged from 0.001 to 0.005, with an average of 0.00225 (Table3). Those of the Southern clade ranged from 0.001 to 0.008, with an average of 0.00536. If the haplotypes of both clades have the same molecular evolutionary rates, these results suggest that the haplotypes of the Northern clade were recently differ- entiated.
The Southern clade also constituted a strongly support- ed monophyletic group (Fig. 6). This fact suggests that the haplotypes of the Southern clade diverged from a common ancestral genome. The Southern clade was highly structured in topology, i.e., each branch of the clade was supported by high confidence value and had at least two synapomorphies. This may indicate that geo- graphical isolation helped diversify effectively the ha- plotypes of the Southern clade. Three major sub-clades were recognized clearly in the Southern clade. We here refer to these subclades as the "Coastal area subclade" (Types J and K), "Western subclade" (Types L-N) and "Eastern subclade" (Types O-Q). The Western and East- ern subclades comprised a strongly supported mono- phyletic group (100%), and were sisters to the Coastal area subclade. This suggests that the haplotypes of the
Western and Eastern subclades diverged from a common ancestral genome.
Soltis et al. (1992b) indicated that the patterns of geo- graphic distribution of cpDNA haplotypes were shared across taxa in the member of Saxifragaceae. They investigated the geographic pattern of cpDNA variation in Tiarella trifoliata, Tellima grandiflora (Soltis et al. 1991), and Tolmiea menziesii (Soltis et al. 1989), all of which share similar geographic distributions and life histories. These three species all exhibited "northern" and "southern" cpDNA clades in spite of the presence of a continuous geographic distribution from northern California to south- eastern Alaska. The authors suggested that Pleistocene glaciation may have played a major role in forming this similar genetic structure.
An interesting finding of the present survey is the concordance in the general distribution pattern of the cpDNA haplotypes of Pedicularis chamissonis to that of a previous report for the Primula cuneifolia (Fujii et al. 1995). Pr. cuneifolia is an alpine meadow plant in Japan, and shows a similar geographic distribution to that of Pe. chamissonis. In the parsimony analysis of Pr. cuneifolia, the haplotypes of the populations from Mts. lide to Mts. Hakusan in central Honshu, Japan were monophyletic with 64% probability (Fig. 1). In the present study, the haplotypes of the populations south of Mt. Gassan formed a single clade with a bootstrap value of 98% (the Southern clade). Kita et al. (1995) clarified that Japanese diploid Aconitum (Ranunculaceae) in Hokkaido and Mts. lide constituted a single clade, which is supported by a 99% bootstrap value. These species occur in subalpine or alpine regions. In Pe. chamissonis, a monophyletic group (the Northern clade) was recognized in the haplotypes of the populations of Unalaska Island, Hokkaido and north- ern Honshu (Fig. 6). Consequently, these results indicate that a common historical factor might have played a major role in the evolutionary processes of these species.
We can propose three hypotheses with respect to the evolutionary patterns of the two major clades of Pedicular- is chamissonis. In general, monophyletic groups distin- guished by large phylogenetic gaps usually arise from long-term extrinsic barriers to gene flow (Avise 1994). In the case of P. chamissonis, the border between the clades is located at northern Honshu. This may imply that the barrier to gene flow exists in the northern Honshu region. However, it is nearly impossible to consider any special geographical barriers sufficient to prevent gene flow over a long period in that region. A second hypothesis is that two major lineages (the Northern and Southern clades) entered the Japanese Archipelago over different land bridges. In the Pleistocene, it is known that northern Japan was connected to the Sakhalin and/or the Kuriles, and that western Japan was connected to the Korean peninsula (Minato and Ijiri 1976). However, this hypothe- sis seems less likely, because P. chamissonis are not distributed at all in western Japan, the Korean peninsula, or China (Fig. 2). The third hypothesis is that two major lineages had traveled southwards to the Japanese Archi-

206 N. Fujii et al.
pelago during a different glacial period. It has been suggested that glacial advances and retreats occurred at least four times in the Northern hemisphere during the Pleistocene (Minato and Ijiri 1976). During climatic coo- ling,-the ancestral genomes of the Southern clade traveled southwards to central Honshu, Japan, and remained alive only on high mountains due to climatic warming. When it was cool again, the ancestral genomes of the Northern clade moved southwards to northern Honshu, The ancestral genomes of the Southern clade that had been distributed in the northern Honshu and northward have been replaced by the ancestral genomes of the Northern clade through the competitive exclusion of local popula- tions. This hypothesis is supported by the result that the haplotypes of the Northern clade have a wider distribution, and by the fact that their genetic diversifications were lower than those of the Southern clade (Figs. 5 and 6). We therefore regard this last hypothesis as the most rational.
We have preliminary data of cpDNA in other species: Campanula lasiocarpa (Campanulaceae), Orchis aristata (Orchidaceae), and Fritillaria camtschatcensis (Liliaceae). These species occur in subalpine or alpine regions in Japan, and have distribution areas similar to that of P. chamissonis. However, no intraspecifir taxa, such as those of P. chamissonis, have been recognized for these species, except a triploid taxon in F. camtschatcensis. Previously, we sequenced the intergenic spacer between trnL and trnF, which was the same region used in the present study, and found little molecular polymorphisms in the three species. In C. lasiocarpa, a single nucleotide substitution was detected among six populations from Mts. Hakusan of central Honshu to Unalaska Island of the Aleutians. There were no molecular polymorphisms in O. aristata among 15 populations from Mt. Kitadake of central Honshu to Unalaska Island. In F. camtschatcensis, a single gap was detected among six populations from Mt. Kitadake to Unalaska Island. If they have the same molecular evolutionary rates as P. chamissonis, the chlor- oplast genomes of these species may have come down to the Japanese Archipelago in a recent glacial period.
Comparisons of the former taxonomic systems and cpDNA data in P. chamissonis
Pedicularis chamissonis is so variable in external mor- phology that several intraspecific classifications for the species have been proposed. Several taxonomists have accepted two varieties: vars. chamissonis and japonica (Miq.) Maxim. (Hulten 1930, Hara 1948, Kitamura et al. 1958, Ohwi 1978). The two varieties are distinguished by the length of beak of corolla. The former is distributed in the Aleutians, Kamchatka and northern Kuriles, and the latter is distributed in the southern Kuriles, Sakhalin, and Hokkaido and Honshu in Japan. Shimizu (1982) recog- nized three varieties, chamissonis, hokkaidoensis T. Shim- izu, and japonica. The second variety is distinguished from the others by a large inflorescence with 7-12 whorls, and he designated the distribution area as Hokkaido,
Japan. Yamazaki (1987) described the population of Rebun Island off the northernmost Hokkaido as P. chamis- sonis var. rebunensis Yamazaki. This variety has a large plant body (70-100 cm tall) and large inflorescence with 20-30whorls. Furthermore, Yamazaki (1982) described the plants that are characterized by long beak of corolla (5-7 mm) as P. chamissonis var. Iongirostrata Yamazaki. This variety is distributed mainly in the southern part of central Honshu. In short, he accepted four varieties in this species: chamissonis, rebunensis, japonica, and Ion- girostrata (Yamazaki 1993), but he did not accept var. hokkaidoensis.
The obtained gene trees were considerably concordant with our classification for present materials based on their external morphology and traditional classification. We identified the present materials from Unalaska Island as P. chamissonis var. chamissonis, the populations from Hok- kaido (excluding Rebun Island) and northern Honshu as var. hokkaidoensis, those of Mt. Gassan (2041), Mts. lide (706 and 707), Mt. Shirouma, Mts. Hakusan, Mt. Ontake and Mt. Kisokomagatake as var. japonica, and those from Mt. Kitadake as var. Iongirostrata (Table 1). In the cpDNA analyses, the haplotype from the population of Unalaska Island was unique (Type A), and the haplotypes from the populations of Hokkaido and northern Honshu, except Haplotype F, constituted a monophyletic group (Fig. 6). The haplotypes from the populations of central Honshu made a single clade, and within this clade, the haplotype from the population of Mt. Kitadake was unique (Type P). In the populations of Mt. Gassan and Mts. lide, we could recognize two varieties (var. hokkaidoensis, 2055 and 742; and var. japonica, 2041, 706 and 707) according to their external morphology (Table 1). In fact, our identification was supported by the present molecular data.
We classified the present materials from Rebun Island as P. chamissonis var. rebunensis (Table1). In the cpDNA data, however, the haplotype from the population of Rebun Island (Type D) was identical to those from the populations of Mt. Yubari, Mt. Apoi, Mts. Hakkoda, Mt. Iwate, Mt. Chokai, Mt. Gassan (2055), and Mts. lide (742) (Fig. 2). This might mean that the giant plants of Rebun Island are merely an extreme type of var. hokkaidoensis.
We express our sincere thanks to Dr. T. Terachi for providing information on primer sequences for the inter- genic spacer between atpB and rbcL, and to Drs. H. Tachida and M. Maki for their valuable suggestions in the statistical analyses. Our hearty thanks are extended to Dr. S. Okitsu and Messrs. Y. Watanabe, N. Shirai and T. Nakajima for provision of plant materials. The present study was supported in part by a Grant-in-Aid for Encour- agement of the JSPS Fellowships for Japanese Junior Scientists (to N. F.), and by the Joint Research Utilizing Science and Technology Potential in Region (to T. S.) from the Science and Technology Agency, Japan.

Intraspecific cpDNA Variation in Pedicularis chamissonis 20?
References
Adachi, J. and Hasegawa, M. 1996. MOLPHY: Programs for Molecular Phylogenetics, version 2.3b3. Institute of Statistical Mathematics, Tokyo.
Avise, J.C. 1994. Molecular markers, natural history and evolution. Chapman and Hall, New York.
Avise, J.C., Arnold, J., Ball, R.M., Bermingham, E., Lamb, T., Neigel, J.E., Reeb, C.A. and Saunders, N.C. 1987. Intraspecific phytogeography: The mitochondrial DNA bridge between population genetics and system- atics. Ann. Rev. Ecol. Syst. 18: 489-522.
Doyle, J.J. and Dickson, E.E. 1987. Preservation of plant samples for DNA restriction endonuclease analysis. Taxon 36: 715-722.
Fujii, N., Ueda, K., Watano, Y. and Shimizu, T. 1995. Intraspecific sequence variation in chloroplast DNA of Primula cuneifolia Ledeb. J. Phytogeogr. & Taxon 43: 15-24.
Hara, H. 1948. A bibliographic enumeration of flowering plants indigenous to or long cultivated in Japan and its adjacent islands. Iwanami shoten, Tokyo.
Hasegawa, M., Kishino, H. and Yano, T. 1985. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. MoL Evol. 22: 160-174.
Hult6n, E. 1930. Pedicularis chamissonis. In E. Hulten, Flora of Kamchatka and the Adjacent Islands 4, Almqvist and Wiksells, Stockholm, pp 109-111.
Kawano, S. 1971. Studies on the alpine flora of Hok- kaido, Japan I. phytogeography. J. Coll. Lib. Arts, Toyama Univ. Nat. Sci. 4: 13-96.
Kimura, M. 1980. A simple method for estimating evolu- tionary rates of base substitutions through compara- tive studies of nucleotide sequences. J. Mol. Evol. 18: 111-120.
Kita, Y., Ueda, K. and Kadota, Y. 1995. Molecular phylogeny and evolution of the Asian Aconitum sub- genus Aconitum (Ranunculaceae). J. Plant Res. 108: 429-442.
Kitamura, S,, Murata, G. and Hori, M. 1958. Colored Illustrations of Herbaceous Plants of Japan vol. 1 (Sympetalae), rev. ed. Hoikusha, Osaka. (in Japanese)
Koidzumi, G. 1919. Genetic and floristic phytogeography of the alpine flora of Japan. Bot. Mag. Tokyo 33: 193-222. (in Japanese)
Minato, M. and Ijiri, S. 1976. The Japanese Archipelago. 3rd ed., Iwanamishoten, Tokyo. (in Japanese)
O'Donnell, K. 1992. Ribosomal DNA internal transcribed spacers are highly divergent in the phytopathogenic ascomycete Fusarium sambucinum (Gibberella pulicaris). Curr. Genet. 22: 213-220.
Ohwi, J. 1978. Flora of Japan, rev. and enlarg, ed. Shibundo, Tokyo. (in Japanese)
Orita, M., Iwahara, H., Kanazawa, H., Hayashi, K. and Sekiya, T. 1989a. Detection of polymorphisms of
human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc. Natl. Acad. Sci. USA 86: 2771-2774.
Orita, M., Suzuki, Y., Sekiya, T. and Hayashi, K. 1989b. Rapid and sensitive detection of point mutations and DNA polymorphisms using the polymerase chain reaction. Genomics 5: 874-879.
Shimizu, T. 1982. The new alpine flora of Japan in color vol. 1. Hoikusha, Osaka. (in Japanese)
Soltis, D.E., Mayer, M.S., Soltis, P.S. and Edgerton, M. 1991. Chloroplast DNA variation in Tellima grandiflor- a (Saxifragaceae). Amer. J. Bot. 78: 1379-1390.
Soltis, D.E. and Soltis, P.S. and Milligan, B.G. 1992a. Intraspecific chloroplast DNA variation: Systematics and phylogenetic implication. In P.S. Soltis, D.E. Soltis and J.J. Doyle, eds., Molecular Systematics of Plants, Chapman and Hall, New York, pp. 117-150.
Soltis, D.E., SolUs, P.S., Kuzoff, R.K. and Tucker, T.L. 1992b. Geographic structuring of chloroplast DNA genotypes in Tiarella trifoliata (Saxifragaceae). PI. Syst. Evol. 181: 203-216.
Soltis, D.E., Soltis, P.S., Ranker, T.A. and Ness, B.D. 1989. Chloroplast DNA variation in a wild plant, Tolmiea menziesii. Genetics 121: 819-826.
Swofford, D.L. 1993. PAUP: Phylogenetic Analysis Using Parsimony, version 3.1.1. The Illinois Natural History Survey, Champaign.
Taberlet, P., Gielly, L., Pautou, G. and Bouvet, J. 1991. Universal primers for amplification of three non-cod- ing regions of chloroplast DNA. PI. Mol. Bio. 17: 1105-1109.
Terachi, T. 1993. Structural alterations of chloroplast genome and their significance to the higher plant evolution. Bull. Inst. Natl. Land Util. Developm. Kyoto Sangyo Univ. 14: 138-148.
Thompson, J.D., Higgins, D.G. and Gibson, T.J. 1996. Clustal W version 1.6. EMBL, Heidelberg.
Toyokuni, H. 1981. A preliminary note on the floristic phytogeography of the alpine flora of Japan. J. Fac. Lib. Arts, Shinshu Univ. Nat. Sci. 15: 81-96.
Yamazaki, T. 1982. Nomenclatural changes and new taxa of some Scrophulariaceae plants in Japan. J. Jap. Bot. 57: 212-215. (in Japanese with Latin description)
Yamazaki, T. 1987. A new variety of Pedicularis chamis- sonis Steven. J. Jap. Bot. 62: 54. (in Japanese with Latin description)
Yamazaki, T. 1993. Pedicularis. in K. Iwatsuki, T. Yam- azaki, D.E. Boufford and H. Ohba, eds., Flora of Japan, vol. 3a. Kodansha, Tokyo. pp. 364-371.
Watano, Y., Imazu, M. and Shimizu, T. 1995. Chloroplast DNA typing by PCR-SSCP in the Pinus pumila-P. parviflora var. pentaphylla complex (Pinaceae). J. Plant Res. 108: 493-499.
(Received November 15, 1996: Accepted March 8, 1997)
Related Documents










