INTRASPECIFIC DIVERSITY AND DISTRIBUTION OF THE COSMOPOLITAN SPECIES PSEUDO-NITZSCHIA PUNGENS (BACILLARIOPHYCEAE): MORPHOLOGY, GENETICS, AND ECOPHYSIOLOGY OF THE THREE CLADES 1 Jin Ho Kim, Bum Soo Park, Joo Hwan Kim, Pengbin Wang Department of Life Science, College of Natural Sciences, Hanyang University, Seoul 133-791, South Korea and Myung Soo Han 2 Department of Life Science, College of Natural Sciences, Hanyang University, Seoul 133-791, South Korea Research Institute for Natural Sciences, Hanyang University, Seoul 133-791, South Korea Three clades of Pseudo-nitzschia pungens, determined by the internal transcribed space (ITS) region, are distributed throughout the world. We studied 15 P. pungens clones from various geographical locations and confirmed the existence of the three clades within P. pungens, based on ITS sequencing and described the three subgroups (IIIaa, IIIab, and IIIb) of clade III. Clade III (clade IIIaa) populations were reported for the first time in Korean coastal waters and the East China Sea. In morphometric analysis, we found the ultrastructural differences in the number of fibulae, striae, and poroids that separate the three clades. We carried out physiological tests on nine clones belonging to the three clades growing under various culture conditions. In temperature tests, only clade III clones could not grow at lower temperatures (10°C and 15°C), although clade I and II clones grew well. The estimated optimal growth range of clade I clones was wider than that of clades II and III. Clade II clones were considered to be adapted to lower temperatures and clade III to higher temperatures. In salinity tests, clade II and III clones did not grow well at a salinity of 40. Clade I clones were regarded as euryhaline and clade II and III clones were stenohaline. This supports the hypothesis that P. pungens clades have different ecophysiological characteristics based on their habitats. Our data show that physiological and morphological features are correlated with genetic intraspecific differentiation in P. pungens. Key index words: distribution; genotype; intraspecific diversity; physiology; pseudo-nitzschia pungens Abbreviations : ANOVA, analysis of variance; C max , maximal growth rate; L:D, light:dark; LM, light microscope; MAST-4, marine stramenopile group 4; ML, Maximum-likelihood; MP, maximum parsi- mony; NJ, neighbor-joining; NOAA, National Oceanic and Atmospheric Administration; NODC, National Oceanographic Data Center; PCR, poly- merase chain reaction; rDNA, ribosomal DNA; R op , optimal growth range; SEM, scanning electron microscope; l max , maximum growth rate Recently, species of the pennate diatom genus Pseudo-nitzschia pungens, some of which produce the neurotoxin domoic acid (Rhodes et al. 1996, Bates et al. 1998, Trainer et al. 1998), have garnered sig- nificant attention. They are distributed throughout the world’s oceans and coasts and are regarded as a common component of marine phytoplankton (Hasle 2002, Lelong et al. 2012, Trainer et al. 2012). Among the Pseudo-nitzschia species, P. pun- gens is the most widely distributed, from 50°S to 70°N latitude, including the Atlantic and Pacific Oceans, and blooms of P. pungens have been recorded from a wide variety of marine ecosystems (Hasle 2002). The genetic intraspecific diversity of P. pungens is broad and includes three clades that can be distinguished from one another by internal transcribed spacer (ITS) analysis sequences (Caste- leyn et al. 2008, Lim et al. 2014) and ribulose 1,5- bisphosphate carboxylase sequences (Casteleyn et al. 2010). Each clade shows distinct biogeograph- ic distribution patterns and has slight but consistent morphological differences (Casteleyn et al. 2008, Churro et al. 2009, Lim et al. 2014). As reviewed by Casteleyn et al. (2009), clade I (P. pungens var. pun- gens) has a cosmopolitan distribution in temperate waters of the Atlantic and Pacific Oceans. Clade II (P. pungens var. cingulata) has only been found in the northeastern Pacific, where it co-occurs with clade I. Clade III (P. pungens var. aveirensis) occurs in the tropical to warm-temperate waters of the Atlantic and Pacific Oceans. However, research on the physiology of these clades has never been reported. Innumerable biological and ecological parame- ters affect intraspecific diversity (Medlin et al. 2000, Montresor et al. 2003). Population diver- gence is driven by natural selection, which 1 Received 10 February 2014. Accepted 29 October 2014. 2 Author for correspondence: e-mail: [email protected]. Editorial Responsibility: M. Cock (Associate Editor) J. Phycol. 51, 159–172 (2015) © 2014 Phycological Society of America DOI: 10.1111/jpy.12263 159

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

INTRASPECIFIC DIVERSITY AND DISTRIBUTION OF THE COSMOPOLITAN SPECIESPSEUDO-NITZSCHIA PUNGENS (BACILLARIOPHYCEAE): MORPHOLOGY, GENETICS, AND
ECOPHYSIOLOGY OF THE THREE CLADES1
Jin Ho Kim, Bum Soo Park, Joo Hwan Kim, Pengbin Wang
Department of Life Science, College of Natural Sciences, Hanyang University, Seoul 133-791, South Korea
and Myung Soo Han2
Department of Life Science, College of Natural Sciences, Hanyang University, Seoul 133-791, South Korea
Research Institute for Natural Sciences, Hanyang University, Seoul 133-791, South Korea
Three clades of Pseudo-nitzschia pungens,determined by the internal transcribed space (ITS)region, are distributed throughout the world. Westudied 15 P. pungens clones from variousgeographical locations and confirmed the existenceof the three clades within P. pungens, based on ITSsequencing and described the three subgroups(IIIaa, IIIab, and IIIb) of clade III. Clade III (cladeIIIaa) populations were reported for the first timein Korean coastal waters and the East China Sea. Inmorphometric analysis, we found the ultrastructuraldifferences in the number of fibulae, striae, andporoids that separate the three clades. We carriedout physiological tests on nine clones belonging tothe three clades growing under various cultureconditions. In temperature tests, only clade IIIclones could not grow at lower temperatures (10°Cand 15°C), although clade I and II clones grew well.The estimated optimal growth range of clade Iclones was wider than that of clades II and III.Clade II clones were considered to be adapted tolower temperatures and clade III to highertemperatures. In salinity tests, clade II and IIIclones did not grow well at a salinity of 40. Clade Iclones were regarded as euryhaline and clade II andIII clones were stenohaline. This supports thehypothesis that P. pungens clades have differentecophysiological characteristics based on theirhabitats. Our data show that physiological andmorphological features are correlated with geneticintraspecific differentiation in P. pungens.
Key index words: distribution; genotype; intraspecificdiversity; physiology; pseudo-nitzschia pungens
Abbreviations: ANOVA, analysis of variance; Cmax,maximal growth rate; L:D, light:dark; LM, lightmicroscope; MAST-4, marine stramenopile group 4;ML, Maximum-likelihood; MP, maximum parsi-mony; NJ, neighbor-joining; NOAA, NationalOceanic and Atmospheric Administration; NODC,
National Oceanographic Data Center; PCR, poly-merase chain reaction; rDNA, ribosomal DNA; Rop,optimal growth range; SEM, scanning electronmicroscope; lmax, maximum growth rate
Recently, species of the pennate diatom genusPseudo-nitzschia pungens, some of which produce theneurotoxin domoic acid (Rhodes et al. 1996, Bateset al. 1998, Trainer et al. 1998), have garnered sig-nificant attention. They are distributed throughoutthe world’s oceans and coasts and are regarded asa common component of marine phytoplankton(Hasle 2002, Lelong et al. 2012, Trainer et al.2012). Among the Pseudo-nitzschia species, P. pun-gens is the most widely distributed, from 50°S to70°N latitude, including the Atlantic and PacificOceans, and blooms of P. pungens have beenrecorded from a wide variety of marine ecosystems(Hasle 2002). The genetic intraspecific diversity ofP. pungens is broad and includes three clades thatcan be distinguished from one another by internaltranscribed spacer (ITS) analysis sequences (Caste-leyn et al. 2008, Lim et al. 2014) and ribulose 1,5-bisphosphate carboxylase sequences (Casteleynet al. 2010). Each clade shows distinct biogeograph-ic distribution patterns and has slight but consistentmorphological differences (Casteleyn et al. 2008,Churro et al. 2009, Lim et al. 2014). As reviewed byCasteleyn et al. (2009), clade I (P. pungens var. pun-gens) has a cosmopolitan distribution in temperatewaters of the Atlantic and Pacific Oceans. Clade II(P. pungens var. cingulata) has only been found inthe northeastern Pacific, where it co-occurs withclade I. Clade III (P. pungens var. aveirensis) occursin the tropical to warm-temperate waters of theAtlantic and Pacific Oceans. However, research onthe physiology of these clades has never beenreported.Innumerable biological and ecological parame-
ters affect intraspecific diversity (Medlin et al.2000, Montresor et al. 2003). Population diver-gence is driven by natural selection, which
1Received 10 February 2014. Accepted 29 October 2014.2Author for correspondence: e-mail: [email protected] Responsibility: M. Cock (Associate Editor)
J. Phycol. 51, 159–172 (2015)© 2014 Phycological Society of AmericaDOI: 10.1111/jpy.12263
159

operates when a species interacts with its environ-ment, leading to adaptations (Schluter 2001). Also,ecological niches and the distribution of bioticcommunities are closely related (Grinnell 1917).Three important factors determine species distribu-tion (Guisan and Thuiller 2005, Soberon 2005). Thefirst is the species’ ownmovement, however, the plank-tonic diatom P. pungens has been moving passivelywith the currents. The second factor includes interac-tions among species, i.e., competition, predation, andinfection. Lastly are the environmental conditionsthat influence the establishment, survival, andreproduction of the species. Here, we focusedon the latter, to determine ecophysiological character-istics that will help to understand the intraspecificdifferentiation and distribution of P. pungensclades.
The fitness of a species can now be linked togenes or other DNA sequences (Ellegren and Shel-don 2008). An increasing amount of eco-genomicinformation has been applied to understand thesuccess of diatoms in aquatic environments, e.g.,Thalassiosira pseudonana and Phaeodactylum tricornutum(Armbrust et al. 2004, Bowler et al. 2008). T. pseudo-nana clones isolated from neritic and oceanic loca-tions show differences in the electrophoretic mobilityof isozymes and in reproduction rates. T. pseudonanastrains display genetic variability and differentiationin different temperature niches (Brand et al. 1981).Skeletonema costatum has a great ecotypic diversity andlarge variation in LSU ribosomal DNA (rDNA)sequences, which have been influenced by their glo-bal distribution (Gallagher 1982, Kooistra et al.2008). According to Gallagher et al. (1984), eachS. costatum clone has particular photoadaptive strate-gies under different light conditions in culture, con-tributing to its great ecological success. Informationabout intraspecific genetic diversity and ecotypes pro-vides the key information to explain a species’ suc-cess. Knowledge of how environmental factors affectgrowth can be useful for understanding the abun-dance and distribution of a species (Kibler et al.2012).
We hypothesized that genetic diversity reflectsphysiological and morphological characteristics inP. pungens, and those are related to the environ-mental conditions of a clone’s geographical origin.Our goal was to understand the ecophysiologicalcharacteristics of each P. pungens clone, isolatedfrom different regions of the world, by estimatingits optimal growth range (Rop) under various cul-ture conditions (i.e., temperature, salinity, lightintensity and nutrients) and investigate morphologi-cal differences among the clades. We then lookedto relate this information to understanding the dis-tinct distribution pattern of each P. pungens cladeworldwide and survived coastal areas to understandwhether emergences of each clade reflect on theirphysiological features.
MATERIALS AND METHODS
Sampling and cell cultures. P. pungens clones for this studyoriginated from diverse regions (Fig. 1; Table 1). Clonal cul-tures of P. pungens clade I were established from Koreancoastal waters. Clade II clones were obtained from theNational Oceanic and Atmospheric Administration (NOAA),USA. Clones of clade III originated from the South Sea ofKorea, the East China Sea and Philippine coastal waters.
For Pseudo-nitzschia isolation, seawater was collected with a4.2-L Van Dorn sampler (Wildlife Supply Company, Yulee,FL, USA) from the surface layer (at 50 cm depth). When celldensity was low (<100 cells � mL�1), samples were concen-trated using a 20-lm mesh. Single cells or single chains wereisolated by Pasteur pipette (Hilgenberg, Germany) under aninverted microscope (IX71; Olympus, Tokyo, Japan) at 2009magnification. Isolated cells were washed three times withfresh seawater, filtered through a 47-mm diameter glass-fiberfilter (GF/F, Whatman, Florham Park, NJ, USA) and trans-ferred into individual wells of a 96-well plate that were filledwith 200 lL of sterilized f/2 medium (Guillard and Ryther1962). To establish clonal cultures, single cells of each isolatewere transferred into a culture flask (cat. no. 70025; SPL,Daegu, Korea) containing 25 mL of sterile f/2 medium. Cul-tures were maintained at 20°C, with a 12:12 light:dark (L:D)photoperiod and 30–50 lmol photons � m�2 � s�1 providedby cool-white fluorescent lamps.
DNA extraction, amplification and sequencing. Based on ITSregions, three clades are found in P. pungens (Casteleyn et al.2008). The rDNA-ITS region was analyzed to categorize ourclones by clade. Cells of each clone were harvested into 1.5-mL micro-tubes (Axygen, Union City, CA, USA) and pelletedby centrifugation at 10,000g (4°C, 5 min). DNA extractionwas carried out using a DNeasy Plant Mini Kit (Qiagen,Valencia, CA, USA) following the manufacturer’s protocol.Extracted DNA was stored at �20°C before amplification.
ITS rDNA was amplified using two primers sets: universalprimers, designed to amplify the ITS1 and 5.8S, UniITSF(GAA GTC GTA ACA AGG TTT CCG) and UniITSR (TTACGT ATC GCA TTT CGC TG), and Pseudo-nitzschia-specificprimers designed to amplify 5.8S and ITS2 region PnITSF(ACT TTC AGC GGT GGA TGT CTA) and PnITSR (CTTGAT CTG AGA TCC GGA ATT). The reaction mixture ran ina 19 Ex Taq Buffer that consisted of <0.1 lg template DNA,0.3 lM of each primer, 0.25 lM of deoxynucleoside triphos-phate and 1 unit of TaKaRa Ex Taq, with the final volume ofthe mixture set to 20 lL.
Polymerase chain reactions (PCR) were conducted in thefollowing steps: initial denaturation at 94°C for 5 min; 36cycles of denaturation for 1 min at 94°C, annealing for1 min at 57°C, and extension for 2 min at 72°C; and finalextension at 72°C for 10 min. PCR-amplified products wereanalyzed by electrophoresis using 1.3% agarose and werepurified using the Qiaquick PCR purification kit (Qiagen,Hilden, Germany). Finally, products were directly sequencedwith the aid of a Big DyeTM Terminator Cycle SequencingReady reaction Kit (Applied Biosystems, Foster City, CA,USA).
Phylogenetic analysis. Both strands of the PCR product weresequenced and the reverse compliment matched, aligned andedited using Bioedit version 7.1.3.0 software (North CarolinaState University). Edited partially, ITS1, 5.8S and ITS2sequences were aligned with published P. pungens and P. mul-tiseries sequences selected from the NCBI GenBank (http://www.ncbi.nlm.nih.gov). All analyzed sequence information isshown in Table 1.
A maximum parsimony (MP) tree was constructed inMEGA version 5.10 software (Tamura et al. 2011). The MP
160 JIN HO KIM ET AL.

tree was obtained using the tree-bisection-regrafting algo-rithm (Nei and Kumar 2000) with search level 1, in whichthe initial trees were obtained by the random addition ofsequences (10 replicates). The tree is drawn to scale, withbranch lengths calculated using the average pathway method(Nei and Kumar 2000). All positions containing gaps andmissing data were eliminated. The analysis involved 39 nucle-otide sequences. All positions containing gaps and missingdata were eliminated. There were 637 positions in the finaldataset. Maximum-likelihood (ML) and neighbor-joining(NJ) analyses based on the Hasegawa-Kishino-Yano model,and the best sequence-evolutional fitting model determinedby jModelTest version 2.13 software (https://code.go-ogle.com/p/jmodeltest2/), were carried out using PAUP ver-sion 4.0b.8 software. ML analyses were conducted throughheuristic searches with the tree bisection reconnectionbranch-swapping algorithm with random-addition-sequencereplication (10 replicates). A bootstrap test was performedfor all of the phylogenetic trees (1,000 replicates; Felsenstein1985).
Microscopy. Light microscope (LM; Axioplan, Zeiss, Jena,Germany) was used to measure the length and width of thevalve, using Image-Pro Plus version 6.0 software. Ten cells ofeach clone were measured at 4009 magnification. Scanningelectron microscope (SEM) was used to observe theultrastructure of P. pungens and to compare morphometric
characteristics among the clones of each clade. Stationary-phase cells of each clone were subsampled and acid-treated(Hendey 1974) to remove organic material, then rinsed threetimes with distilled water. Cleaned samples were placed ontoglass cover slips that were coated with Poly-L-Lysine (Sigma,Saint Louis, MO, USA) in water (0.1% w/v), and dried. Speci-mens were coated with Pt for 300 s and observed under SEM(JSM-6701F; JEOL, Tokyo, Japan). Statistical analyses of themorphometric data were carried out using SPSS version 8.0software (SPSS Inc., Chicago, IL, USA). One-way analysis ofvariance (ANOVA) was performed to compare morphometricdata between clones. Tukey’s test was carried out for signifi-cance verification.
Measurement of in vivo fluorescence. In vivo fluorescence wasused to measure P. pungens growth rate (Guillard 1973). Todetermine the initial inoculation concentration and confirmthe relationship between in vivo fluorescence and cell abun-dance, triplicate cultures (four clones of clade I, two of cladeII and three of clade III) were pre-cultured in 25 9 150-mmtest-tubes (Pyrex, Corning, NY, USA) filled with 25 mL of f/2medium, under the above conditions. Each test tube reachedat stationary phase, was placed in a fluorometer (model 10-AU; Turner Designs, Sunnyvale, CA, USA) to measure in vivofluorescence. Measurements were made after the cultureswere diluted with fresh filtered seawater to 1/2, 1/4, 1/8,and 1/16 of the origin culture. Aliquots (1 mL) of each
FIG. 1. Sampling sites ▲ with station numbers. Stations 1–11 are the origins of Pseudo-nitzschia pungens clones. Station 12 (Masan Bay)is the location for samples used for the single cell/chain PCR to survey the proportion of clade III cells.
INTRASPECIFIC DIVERSITY OF THREE CLADES OF PSEUDO-NITZSCHIA PUNGENS 161

TABLE1.
Inform
ationontheoriginofclones
usedin
thisstudyan
daccessionnumbersfrom
Gen
Ban
k.Station(St.)numbersreferto
those
shownin
Figure
1.
Species
Clonenam
eor
AccessionNo.(cladeNo.)
Geo
grap
hic
origin
Analysis
Datesampled
Referen
cePhyloge
netic
Morphological
Physiological
P.pu
ngens
HYM
R11
04PP34
B1(I)
Korea,
Port
Maryang(St.1)
oo
o20
11/04
Thisstudy
HYS
S110
5PP35
H10
(I)
Korea,
Port
Susan(St.2)
o20
11/05
Thisstudy
HYS
C11
05PP35
G6(I)
Korea,
Port
Samch
uk(St.3)
o20
11/05
Thisstudy
HYC
S110
5PP35
F4(I)
Korea,
Port
Chuksan
(St.4)
o20
11/05
Thisstudy
HYG
C11
05PP35
D3(I)
Korea,
Port
Gilch
un(St.5)
oo
o20
11/05
Thisstudy
HYN
G10
10PP29
F5(IIIaa)
Korea,
Nam
hae
Island(St.6)
o20
10/10
Thisstudy
HYN
H10
10PP29
B9(IIIaa)
Korea,
Betwee
nNaro-Geu
mo
Islands(St.7)
o20
10/10
Thisstudy
HYJJ120
2PP38
B2(I)
Korea,
Port
Jeju
(St.8)
oo
2011
/08
Thisstudy
HYE
C11
08PP36
B6(I)
EastChina,
nearIeoIsland(St.9)
oo
o20
11/08
Thisstudy
HYE
C11
08PP36
C7(IIIaa)
EastChina,
nearIeoIsland(St.9)
oo
o20
11/08
Thisstudy
HYE
C11
08PP36
E10
(IIIaa)
EastChina,
nearIeoIsland(St.9)
oo
o20
11/08
Thisstudy
HYP
P11
10PP37
F8(IIIab
)Philippines,Pagbilao
Port
(St.10
)o
oo
2011
/10
Thisstudy
HYP
P11
10PP37
F9(IIIab
)Philippines,Pagbilao
Port
(St.10
)o
2011
/10
Thisstudy
NWFSC
356(II)
USA
,Clam
Bay,WashingtonState(St.11
)o
oo
2011
/10
Thisstudy
NWFSC
357(II)
USA
,Clam
Bay,WashingtonState(St.11
)o
oo
2012
/04
Thisstudy
AM77
8777
(I)
Spain,Bay
ofVigo
oCasteleyn
etal.20
08AM77
8785
(I)
Spain,Bay
ofVigo
oCasteleyn
etal.20
08AM77
8803
(I)
USA
,Se
quim
Bay
StatePark,
WashingtonState
oCasteleyn
etal.20
08
AM77
8815
(I)
New
Zealand,Stee
lsRee
fo
Casteleyn
etal.20
08AM77
8817
(I)
New
ZealandTaylor’sMistake
oCasteleyn
etal.20
08AM77
8786
(I)
Can
ada,
Cardigan
River,Prince
EdwardIsland
oCasteleyn
etal.20
08
AM77
8787
(I)
Can
ada,
Oak
Point,Miram
ichiBay,
New
Brunswick
oCasteleyn
etal.20
08
AM77
8788
(I)
Can
ada,
Deadman
sHbr.,Bay
ofFundy,
New
Brunswick
oCasteleyn
etal.20
08
AM77
8769
(I)
Netherlands,Bresken
so
Casteleyn
etal.20
08AM77
8775
(I)
Netherlands,Bresken
so
Casteleyn
etal.20
08AM77
8812
(I)
Japan
,OfunatoBay,IwatePrefecture
oCasteleyn
etal.20
08FN82
3051
(I)
Den
mark,
Lim
fjord,Thisted
oCasteleyn
etal.20
10AM77
8795
(II)
USA
,LaPush,WashingtonState
oCasteleyn
etal.20
08AM77
8796
(II)
USA
,Eld
Inlet,MudBay,WashingtonState
oCasteleyn
etal.20
08AM77
8804
(II)
USA
ECOHABI(coastalWashingtonState)
oCasteleyn
etal.20
08AM77
8811
(II)
USA
ECOHABI(coastalWashingtonState)
oCasteleyn
etal.20
08DQ16
6533
(IIIaa)
Vietnam
,Hai
Phong
oUnpublished
DQ06
2665
(IIIab
)Vietnam
,Khan
HoaBay
oUnpublished
AY2
5784
6(IIIb)
Mex
ico,NearTuxp
anm
oLundholm
etal.20
03AY2
5784
5(IIIb)
Portugal,CostaNova
oLundholm
etal.20
03EU68
4234
(IIIb)
Portugal,Aveiroco
astallago
on
oChurroet
al.20
09EU68
4236
(IIIb)
Portugal,Aveiroco
astallago
on
oChurroet
al.20
09P.multiseries
DQ44
5651
USA
,ChoptankRiver,Chesap
eake
Bay
oThessenet
al.20
09AY2
5784
4USA
,MontereyBay,California
oLundholm
etal.20
03
162 JIN HO KIM ET AL.

diluted culture were then placed into 1.5-mL micro-tubes andimmediately preserved with 0.3% Lugol’s solution (Saraceniand Ruggiu 1974) for cell counts. Counting was carried outusing a Sedgwick-Rafter chamber with a LM (Axioplan, Zeiss)at 2009 magnification.
Physiological tests: temperature, salinity, irradiance, and nutri-ents. Duplicate cultures of nine P. pungens clones (Table 1)were used for measurement of growth rates. Cultures wereinoculated into test tubes filled with 25 mL of f/2 medium,some modified to change the nitrate and silicate concentra-tion (see below). In vivo fluorescence was quantified at 2-d(until day 16) and 3-d (until the day 31) intervals. Initial celldensity was ~2 9 103 cells � mL�1, determined by a derivedlinear equation from the relationship between in vivo fluores-cence and cell abundance of each clone. Growth rates weredetermined by the following equation (Guillard 1973): l = ln(F2/F1)/T2 � T1, where l is the growth rate (d�1), F1 and F2are relative in vivo fluorescence units of averaged duplicatecultures, and T1 and T2 are respective experimental timepoints (days). Each clade’s maximum growth rate (lmax) wasdetermined as the mean of each culture’s highest l value.
In the temperature experiment, cultures were grown in f/2medium at 10, 15, 20 and 25°C; at a salinity of 30 and anirradiance of 80 lmol photons � m�2 � s�1. Before inocula-tion, all clonal cultures were acclimated at each temperaturefor 4 d. For the 10°C test, cultures were first grown at 15°Cfor 2 d before being moved to 10°C, where they remainedfor another 2 d.
In the salinity experiment, filtered seawater was diluted toa salinity of 10 and NaCl was added to bring salinity levels upto 20, 35, and 40. Seawater from these prepared salinity levels(10, 20, 35, and 40) was used to make the f/2 medium.Before the experiment, the salinity of each culture wasgradually adjusted by replacing half of the medium with freshf/2 medium of the desired salinity (10, 20, 35, and 40) everyday; this treatment was continued for 4 d. Cultures weregrown at 20°C and with 80 lmol photons � m�2 � s�1 ofirradiance.
In the irradiance experiment, different light intensities(25, 50, 100, and 300 lmol photons � m�2 � s�1) were estab-lished by regulating the distance between cultures and thefluorescent lamps. Before inoculation, all clonal cultures wereacclimated at each irradiance level for 4 d. For the 300 lmolphotons � m�2 � s�1 test, cultures were first grown at100 lmol photons � m�2 � s�1 for 2 d before being moved to300 lmol photons � m�2 � s�1, where they were grown foranother 2 d. Cultures were grown at 20°C and a salinity of 30.
In the nutrient experiment, modified f/2 media was pre-pared with the silicate and nitrate concentrations shownbelow. Nitrate was added aseptically after autoclaving toachieve solutions with N:P ratios of 6, 10, 16, 24, and 100,with phosphorus at 36.2 lmol � L�1, and silicate at107 lmol � L�1 (f/2 concentrations). Silicate was added toachieve N:Si ratios of 4, 9, 18, and 35, with nitrate at217.2 lmol � L�1 and phosphate at 36.2 lmol � L�1. Prior toinoculation, all clonal cultures were acclimated at each nutri-ent condition for 4 d. Cultures were grown at 20°C, a salinityof 30 and at 80 lmol photons � m�2 � s�1 of irradiance. Allexperiments were carried out under a 12:12 L:D cycle.
ANOVA and Tukey’s test were performed to compare thelmax among the clades. Second-order polynomial curve fit-ting was used to estimate the Rop and conditions for maximalgrowth (Cmax), using the following equation:y = ax2 + bx + c, where y is the averaged lmax of each clade,x is the value of the given culture condition (temperature,salinity, and irradiance), and a, b, and c are constants, calcu-lated using Microsoft Excel version 14 software. Cmax isdefined by the x value corresponding to the maximal y value.
Rop is the x range corresponding with y values higher thanCmax � 0.9 in each experiment.
Nested PCR of single cell/chain of P. pungens. To confirmthe occurrence of a clade III population according environ-mental changing, we carried out single-cell PCR using P. pun-gens cells collected from Masan Bay, within Jinhae Bay (Fig. 1,St. 12), which is known to contain Pseudo-nitzschia spp. (Choet al. 2001a,b). Samples were taken four times during seawa-ter temperature decreasing season that July 28, August 19,September 7, and September 30 (2011). Water samples forP. pungens cell counts were preserved using 3% Lugol’s solu-tion. Counts were performed using a Sedgwick-Rafter cham-ber with a LM (Axioplan, Zeiss) at 2009 magnification.
Individual cells or short chains of P. pungens, totaling 123(48, 43, 32, and 0 each sampling day), were isolated into 0.2-mL microtubes to extract DNA, using the direct extractionmethod (Ki et al. 2004). Nested PCR amplifications targetingthe ITS2 region were carried out using two primer sets. First,PCR amplification was performed using an external primerset, UniITSF (used in this study, located at the end portionof 18S) and PsITSR2 (50-ACA TCC TGA GCT AGT CCC G-30,sequences located at the start of 28S). PCR were run in thefollowing steps: initial denaturation at 9°C for 10 min; 30cycles of denaturation for 1 min at 94°C, annealing for1 min at 58°C and extension for 2 min at 72°C; and finalextension at 72°C for 10 min. One lL of PCR product wasprovided as a template for the next round of PCR. A secondPCR amplification was performed using a PnITSF andPnITSR primer set (used in this study). PCR were run in thefollowing steps: initial denaturation at 94°C for 5 min; 35cycles of denaturation for 1 min at 94°C, annealing for1 min at 57°C and extension for 1 min at 72°C; and finalextension at 72°C for 5 min. The components of both PCRmixtures, the first and the second, were the same as thosementioned above. Finally, products were directly sequencedwith the aid of a Big DyeTM Terminator Cycle SequencingReady Reaction Kit (Applied Biosystems). Clade affinity ofthe isolates was determined by comparison of loci of ITS2with other sequence data in this phylogenetic study. NJ andMP trees were constructed using MEGA 5.10 version software(1,000 replicates).
RESULTS
Genetic diversity and distribution of P. pungens. Atotal of 37 P. pungens sequences containing 680bases of partial ITS rDNA (ITS1 = 252, 5.8S = 171and ITS2 = 257 bases) were aligned with two P. mul-tiseries sequences from GenBank for outgroups. Atotal of 645 bases were conserved and 35 were vari-able. Out of 35 variable loci, 14 were located inITS1, 20 in ITS2, and 1 in the 5.8S region. Themost parsimonious tree (length = 129) is shown inFigure 2. Three main clades could be distinguished(I, II, and III), which correspond to the cladesdetermined by Casteleyn et al. (2008). Within cladeIII, three subgroups (IIIaa, IIIab, and IIIb) could bediscerned. The clades I, II, and the three subgroupsof clade III (IIIaa, IIIab, and IIIb) were supportedby high bootstrap values (>72%).The genetic distance calculated between clade I
and clade II was 1.20%; between I and III 2.91%;and between II and III 3.20%. The genetic distanceswithin clades I and II were <0.01%, but those forclade III were relatively high (0.91%). The inter-
INTRASPECIFIC DIVERSITY OF THREE CLADES OF PSEUDO-NITZSCHIA PUNGENS 163

subclade genetic difference in clade III between II-Iaa and IIIab was 0.97%; between IIIaa and IIIb1.22%; and between IIIab and IIIb 1.54%.
The geographical distributions of the clonesshown in the phylogenetic tree are shown inFigure 3. Clade I clones were isolated from Korean
FIG. 2. Maximum parsimony tree of Pseudo-nitzschia based on internal transcribed space sequences. Bootstrap values are shown aboveeach branch (MP/NJ/ML). ♦: clones used in this study.
FIG. 3. Worldwide distribution of Pseudo-nitzschia pungens based on internal transcribed spacer information. Circles indicate locations ofclones isolated for this study. Squares indicate clones from published studies. Black = clade I; gray = clade II; white = clade III (IIIaa);white marked X = clade III (IIIab); white dotted = clade III (IIIb); half-filled circle = co-occurrence of clades I and III.
164 JIN HO KIM ET AL.

coastal waters, including eastern, western, andsouthern portions. Clade II clones were only foundin Puget Sound, USA, and clade III clones werefound in the Philippines, East China Sea, and Kor-ean south coast waters. These findings are inagreement with Casteleyn et al. (2008, 2009, 2010).However, this is the first record of the appearanceof clade III (IIIaa) populations in Korean coastalwaters and the East China Sea.Identification and morphometric data of P. pun-
gens. Isolated cells were successfully identified asP. pungens using the ultrastructure characteristicsfrom SEM images and the sequenced ITS regions(compared with information from GenBanksequences, with a similarity of >97%).
The shape of P. pungens cells was spindle-likeand linear in valve view (Fig. 4, a, d, g). Gener-ally, one stria consisted of two rows of poroids(Fig. 4, b, e, h). Frequently, additional poroidswere observed the between two rows of poroids,near the raphe sternum (Fig. 4f, indicated byarrows). The length of the transapical and apicalaxes, as well as the number of fibulae, striae, andporoids in 10 lm, is shown in Table 2. Clonesfrom all clades had valvocopulae consisting of onerow of circular to square, or oval, poroids (Fig. 4,k and l). In clade II cells, in particular, we occa-sionally found distinguishable valvocopulae thathad one row of square structures that consisted offour poroids (Fig. 4m). This is consistent with it
TABLE 2. Morphometric data (averages of ten cells � SD) of each clone and averages in each clade. Values in bold are sig-nificant (P � 0.001).
CladeClone name (sub-groups
of clade III)Valve
length (lm)Valve
width (lm)Fibulae
(in 10 lm)Striae
(in 10 lm)Poroids
(in 1 lm)
Additionalporoids
(in 10 striae)
I HYMR1104PP34B1 102.60 � 1.32 4.03 � 0.32 10.70 � 0.82 10.40 � 0.52 2.60 � 0.24 0.45 � 0.59HYGC1105PP35D3 106.46 � 1.64 4.00 � 0.24 10.30 � 0.95 10.10 � 0.74 2.47 � 0.16 1.08 � 1.75HYEC1108PP36B6 85.13 � 1.91 3.84 � 0.15 10.50 � 0.53 10.30 � 0.48 2.69 � 0.23 0.20 � 0.33
II HYNWFSC356 84.02 � 1.85 3.94 � 0.32 12.00 � 0.49 11.90 � 0.69 3.31 � 0.25 7.32 � 5.20HYNWFSC357 109.31 � 1.71 3.81 � 0.34 11.90 � 0.38 12.20 � 0.95 3.56 � 0.25 6.71 � 4.64
III HYEC1108PP36E10 (IIIaa) 108.03 � 0.99 3.63 � 0.23 11.70 � 0.67 12.20 � 0.79 3.28 � 0.24 1.16 � 2.52HYPP1110PP36C7 (IIIaa) 105.20 � 0.80 3.85 � 0.28 13.60 � 0.70 13.60 � 0.70 3.26 � 0.41 1.12 � 2.10HYPP1110PP37F8 (IIIab) 98.84 � 1.60 3.88 � 0.28 12.00 � 1.05 12.10 � 0.88 3.27 � 0.26 0.62 � 1.05
Clade I average 98.07 � 9.57 3.95 � 0.25 10.50 � 0.78A 10.27 � 0.58A 2.59 � 0.23A 0.57 � 1.11A
Clade II average 96.67 � 13.08 3.93 � 0.33 11.95 � 0.76B 12.10 � 0.94B 3.44 � 0.27B 7.02 � 4.80B
Clade III average 104.02 � 4.07 3.76 � 0.26 12.43 � 1.17B 12.63 � 1.03B 3.27 � 0.30C 0.96 � 1.94A
In each content, A, B, and C are significantly different (P = 0.01, Turkey’s test) subgroups among clade.
FIG. 4. SEM micrographs of Pseudo-nitzschia pungens. (a–c, k): Clade I (a, k; HYGC1105PP35D3, b; HYEC1108PP36B6, c;HYMR1104PP34B1;). (d–f, j, l): Clade II (d, e, j; NWFSC357, f, l; NWFSC356). (g–I, m): Clade III (g, m; HYEC1110PP36C7, h;HYPP1110PP37F8, I; HYEC1108PP36E10). (a, d, g): Whole-valve view. (b, e, h): Internal-valve view. (c, f, i): External-valve view. (j): Exter-nal-girdle view. (k–m): Valvocopulae with poroids. Note the additional poroids, indicated with an arrow in (f). Scales: 10 lm (b, d, g);1 lm (b, c, e, f, h, i, j, k, l, m).
INTRASPECIFIC DIVERSITY OF THREE CLADES OF PSEUDO-NITZSCHIA PUNGENS 165
being considered as P. pungens var. cingulata (Vil-lac and Fryxell 1998).
The ANOVA test showed significant differencesamong clades with respect to ultrastructure charac-teristics. From our morphometric data, each cladewas subdivided into two or three subgroups(Table 2). Clade I was separated from the otherclades, based on the number of fibulae and striae(F2,77 = 59.58 and 33.82, P < 0.001). Clade I cloneshad significantly fewer fibulae and striae in 10 lmthan did clade II and III (10.50 � 0.78 and10.27 � 0.78 in 10 lm, respectively). Clade II wasdistinguishable from the other clades based on thenumber of additional poroids in 10 striae(F2,77 = 38.85, P < 0.001). As well, each clade wasdistinguished based on the density of poroids in1 lm (F2,77 = 20.34, P < 0.001). Clade II clones hadlarger number of poroids. However, there is no sig-nificant difference between two subgroups (IIIaaand IIIab) of clade III (data not shown). The cellshapes and morphometric data presented here aresimilar to those of previous studies (Casteleyn et al.2008, Churro et al. 2009).Growth rates of P. pungens under various treat-
ments. High correlation coefficients (R2 > 0.97, atleast) were found between in vivo fluorescence andcell concentration of nine P. pungens clones used inphysiological experiments (Fig. S1 in the Support-ing Information). Measurement of in vivo fluores-cence therefore provides a credible estimate ofcell concentration and growth rate for each P. pun-
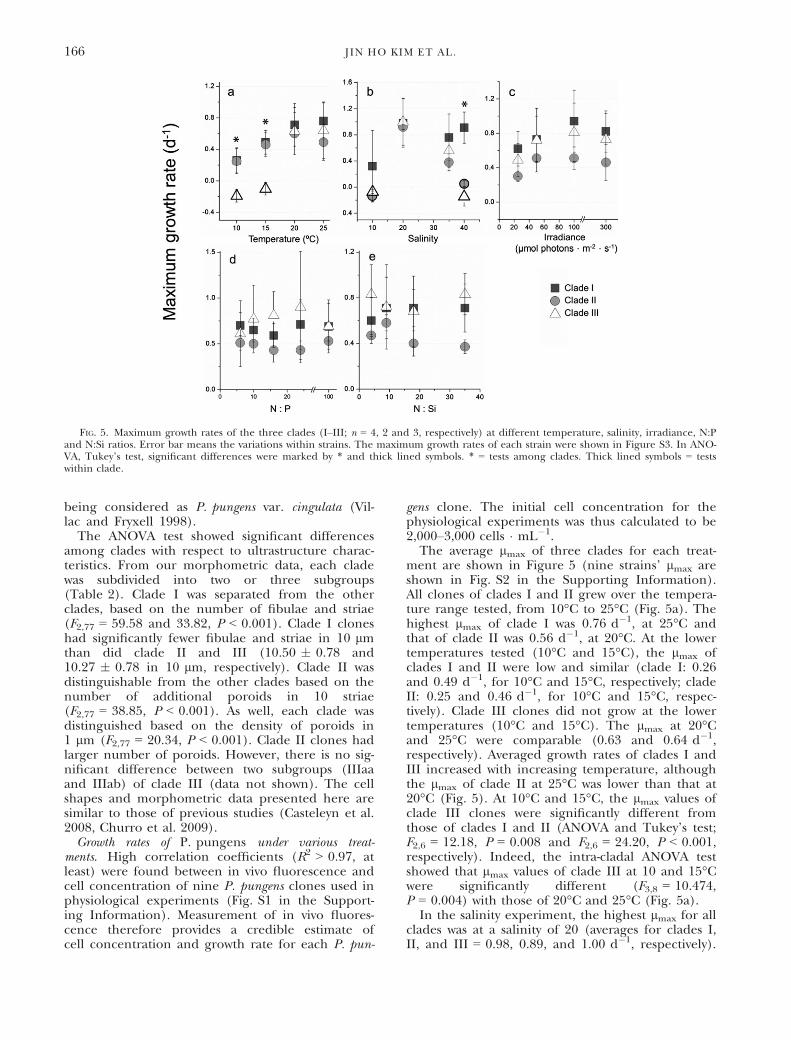
gens clone. The initial cell concentration for thephysiological experiments was thus calculated to be2,000–3,000 cells � mL�1.The average lmax of three clades for each treat-
ment are shown in Figure 5 (nine strains’ lmax areshown in Fig. S2 in the Supporting Information).All clones of clades I and II grew over the tempera-ture range tested, from 10°C to 25°C (Fig. 5a). Thehighest lmax of clade I was 0.76 d�1, at 25°C andthat of clade II was 0.56 d�1, at 20°C. At the lowertemperatures tested (10°C and 15°C), the lmax ofclades I and II were low and similar (clade I: 0.26and 0.49 d�1, for 10°C and 15°C, respectively; cladeII: 0.25 and 0.46 d�1, for 10°C and 15°C, respec-tively). Clade III clones did not grow at the lowertemperatures (10°C and 15°C). The lmax at 20°Cand 25°C were comparable (0.63 and 0.64 d�1,respectively). Averaged growth rates of clades I andIII increased with increasing temperature, althoughthe lmax of clade II at 25°C was lower than that at20°C (Fig. 5). At 10°C and 15°C, the lmax values ofclade III clones were significantly different fromthose of clades I and II (ANOVA and Tukey’s test;F2,6 = 12.18, P = 0.008 and F2,6 = 24.20, P < 0.001,respectively). Indeed, the intra-cladal ANOVA testshowed that lmax values of clade III at 10 and 15°Cwere significantly different (F3,8 = 10.474,P = 0.004) with those of 20°C and 25°C (Fig. 5a).In the salinity experiment, the highest lmax for all
clades was at a salinity of 20 (averages for clades I,II, and III = 0.98, 0.89, and 1.00 d�1, respectively).
FIG. 5. Maximum growth rates of the three clades (I–III; n = 4, 2 and 3, respectively) at different temperature, salinity, irradiance, N:Pand N:Si ratios. Error bar means the variations within strains. The maximum growth rates of each strain were shown in Figure S3. In ANO-VA, Tukey’s test, significant differences were marked by * and thick lined symbols. * = tests among clades. Thick lined symbols = testswithin clade.
166 JIN HO KIM ET AL.

At a salinity of 10, only clones HYMR1104PP34B1and HYEC1108PP36B6 (clade I) grew (lmax = 1.09and 0.39 d�1, respectively). At a salinity of 40, theaverage lmax of all clones in clades II and III wasvery low (0.05 and �0.15 d�1, respectively). Other-wise, clade I clones grew well (0.91 d�1) at a salinityof 40, and lmax values were significantly differentfrom those of clades II and III (ANOVA andTukey’s test; F2,6 = 28.40, P < 0.001). In intra-cladalANOVA test, the lmax values of clade II and III atsalinities of 10 and 40 were significantly different(clade II, F3,4 = 64.908, P < 0.001 and clade III,F3,8 = 15.929, P < 0.001, respectively; Fig. 5b).
In the irradiance test, the highest lmax values forclades I, II and III were at 100 lmol pho-tons � m�2 � s�1 (0.94, 0.51 and 0.81 d�1, respec-tively). At 300 lmol photons � m�2 � s�1, lmax
values of all clades were lower (0.82, 0.46 and0.73 d�1, respectively). However, there were no sig-nificant differences in lmax values at any irradiancelevel among the clades.
In the nutrient test, there were no significant dif-ferences among lmax values for any of the clades atall N:P and N:Si ratios (Fig. 5, d and e).
To estimate Cmax and Rop (Table 3), we used sec-ond-order fitting curves. In the nutrient tests (N:Pand N:Si), this fitting analysis was not carried outbecause its lmax values were not fitted significantly.
In the temperature test, maximum values for cladeIII Rop and Cmax were not detected in our testedrange because a decreasing area of the curve wasnot detected by the fitting equation. However, itseems that clade III clones were better adapted tothe higher temperature than the other clades, as itsRop range was at least >23.7°C. In the salinity test,clade I clones had the widest salinity tolerancerange among the three clades; those of clades IIand III were similar and narrower compared toclade I. In the irradiance test, the Rop range ofclade II was lower relative to the other clades,although there were no significant differencesamong lmax values.Nested PCR of single cell/chain of P. pungens. In
the direct PCR results, 80 of 123 extracted DNAsamples were amplified successfully (384 base pairsof partial 5.8S and ITS2 regions) and successratios were 36/48, 28/43, 18/32 and 0/0 (couldnot find the P. pungens cell) at each date sam-pled. Sequences of PCR products determined theclade categorization by comparison with thesequences of our tested clones. They were thendivided into two clades (I and III) by the NJ andMP phylogenetic trees (Fig. 6). Separatedsequences were the same as the other sequencesin the same group (100%). Clade II sequenceswere not detected.
TABLE 3. Optimal growth range (Rop) and conditions for maximal growth (Cmax) for each clade, estimated by second-orderpolynomial equations, for temperature (°C), salinity and irradiance (lmol photons � m�2 � s�1). R2 values of each clade’sfitting curves are 0.992, 0.991, and 0.847; temperature test, 0.758, 0.958, and 1.000; salinity test 0.997, 0.662, and 0.878;irradiance test, for clade I, clade II, and clade III, respectively.
Clade
Temperature Salinity Irradiance
Rop Cmax Rop Cmax Rop Cmax
I 20.1–32.9 26.5 21.1–35.8 28.4 107–261 184II 15.1–22.9 19.0 20.5–29.5 25.0 102–243 173III >23.7 n.d. 20.3–29.0 24.7 108–257 183
n.d.: not determined in this test.
FIG. 6. Maximum parsimony tree of Pseudo-nitzschia pungens cells isolated from Masan Bay revealed by partial 5.8S and ITS2 sequencesinformation. Bootstrap values are shown above each branch (MP/NJ).
INTRASPECIFIC DIVERSITY OF THREE CLADES OF PSEUDO-NITZSCHIA PUNGENS 167
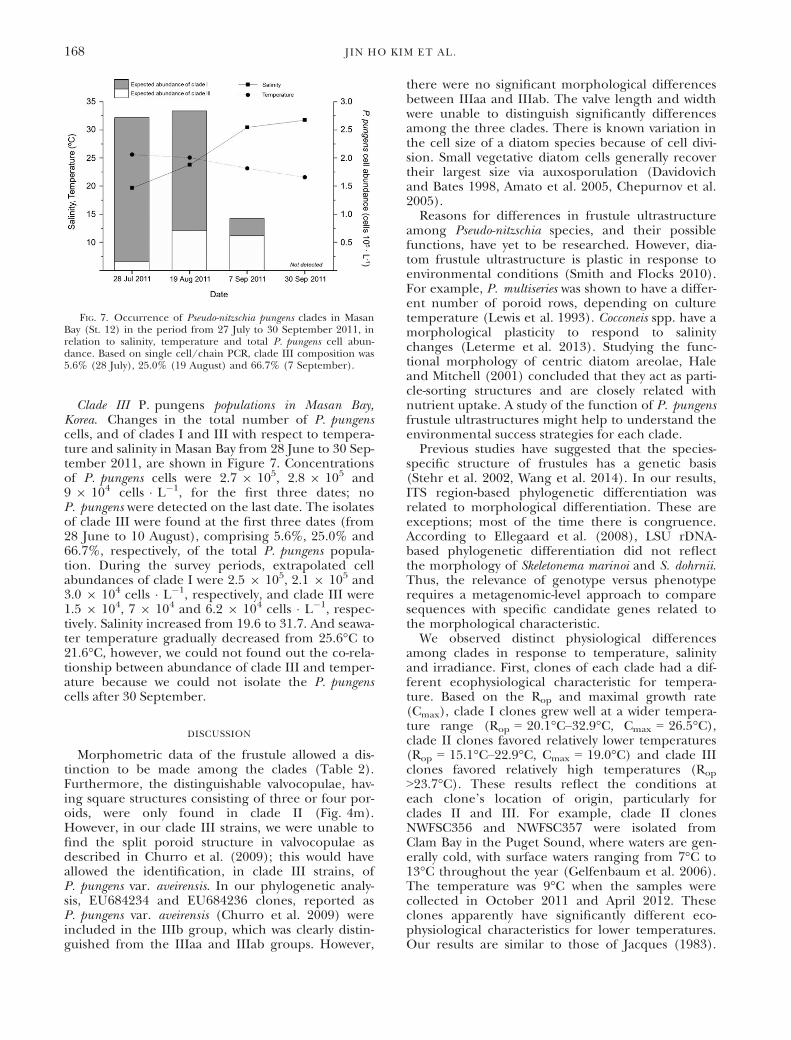
Clade III P. pungens populations in Masan Bay,Korea. Changes in the total number of P. pungenscells, and of clades I and III with respect to tempera-ture and salinity in Masan Bay from 28 June to 30 Sep-tember 2011, are shown in Figure 7. Concentrationsof P. pungens cells were 2.7 9 105, 2.8 9 105 and9 9 104 cells � L�1, for the first three dates; noP. pungens were detected on the last date. The isolatesof clade III were found at the first three dates (from28 June to 10 August), comprising 5.6%, 25.0% and66.7%, respectively, of the total P. pungens popula-tion. During the survey periods, extrapolated cellabundances of clade I were 2.5 9 105, 2.1 9 105 and3.0 9 104 cells � L�1, respectively, and clade III were1.5 9 104, 7 9 104 and 6.2 9 104 cells � L�1, respec-tively. Salinity increased from 19.6 to 31.7. And seawa-ter temperature gradually decreased from 25.6°C to21.6°C, however, we could not found out the co-rela-tionship between abundance of clade III and temper-ature because we could not isolate the P. pungenscells after 30 September.
DISCUSSION
Morphometric data of the frustule allowed a dis-tinction to be made among the clades (Table 2).Furthermore, the distinguishable valvocopulae, hav-ing square structures consisting of three or four por-oids, were only found in clade II (Fig. 4m).However, in our clade III strains, we were unable tofind the split poroid structure in valvocopulae asdescribed in Churro et al. (2009); this would haveallowed the identification, in clade III strains, ofP. pungens var. aveirensis. In our phylogenetic analy-sis, EU684234 and EU684236 clones, reported asP. pungens var. aveirensis (Churro et al. 2009) wereincluded in the IIIb group, which was clearly distin-guished from the IIIaa and IIIab groups. However,
there were no significant morphological differencesbetween IIIaa and IIIab. The valve length and widthwere unable to distinguish significantly differencesamong the three clades. There is known variation inthe cell size of a diatom species because of cell divi-sion. Small vegetative diatom cells generally recovertheir largest size via auxosporulation (Davidovichand Bates 1998, Amato et al. 2005, Chepurnov et al.2005).Reasons for differences in frustule ultrastructure
among Pseudo-nitzschia species, and their possiblefunctions, have yet to be researched. However, dia-tom frustule ultrastructure is plastic in response toenvironmental conditions (Smith and Flocks 2010).For example, P. multiseries was shown to have a differ-ent number of poroid rows, depending on culturetemperature (Lewis et al. 1993). Cocconeis spp. have amorphological plasticity to respond to salinitychanges (Leterme et al. 2013). Studying the func-tional morphology of centric diatom areolae, Haleand Mitchell (2001) concluded that they act as parti-cle-sorting structures and are closely related withnutrient uptake. A study of the function of P. pungensfrustule ultrastructures might help to understand theenvironmental success strategies for each clade.Previous studies have suggested that the species-
specific structure of frustules has a genetic basis(Stehr et al. 2002, Wang et al. 2014). In our results,ITS region-based phylogenetic differentiation wasrelated to morphological differentiation. These areexceptions; most of the time there is congruence.According to Ellegaard et al. (2008), LSU rDNA-based phylogenetic differentiation did not reflectthe morphology of Skeletonema marinoi and S. dohrnii.Thus, the relevance of genotype versus phenotyperequires a metagenomic-level approach to comparesequences with specific candidate genes related tothe morphological characteristic.We observed distinct physiological differences
among clades in response to temperature, salinityand irradiance. First, clones of each clade had a dif-ferent ecophysiological characteristic for tempera-ture. Based on the Rop and maximal growth rate(Cmax), clade I clones grew well at a wider tempera-ture range (Rop = 20.1°C–32.9°C, Cmax = 26.5°C),clade II clones favored relatively lower temperatures(Rop = 15.1°C–22.9°C, Cmax = 19.0°C) and clade IIIclones favored relatively high temperatures (Rop
>23.7°C). These results reflect the conditions ateach clone’s location of origin, particularly forclades II and III. For example, clade II clonesNWFSC356 and NWFSC357 were isolated fromClam Bay in the Puget Sound, where waters are gen-erally cold, with surface waters ranging from 7°C to13°C throughout the year (Gelfenbaum et al. 2006).The temperature was 9°C when the samples werecollected in October 2011 and April 2012. Theseclones apparently have significantly different eco-physiological characteristics for lower temperatures.Our results are similar to those of Jacques (1983).
FIG. 7. Occurrence of Pseudo-nitzschia pungens clades in MasanBay (St. 12) in the period from 27 July to 30 September 2011, inrelation to salinity, temperature and total P. pungens cell abun-dance. Based on single cell/chain PCR, clade III composition was5.6% (28 July), 25.0% (19 August) and 66.7% (7 September).
168 JIN HO KIM ET AL.

Three diatoms, isolated from the Indian sector ofthe Antarctic Ocean, likewise grew well at low tem-peratures. When the temperature was increasedfrom 5°C to 10°C in culture, the cells’ growth ratedecreased to zero (Jacques 1983). Clade III clonesHYPP1110PP37F8 and HYPP1110PP37F9 were iso-lated from Pagbilao Bay (Philippines), 14°C fromthe equator. The average surface water temperatureof Pagbilao Bay is 28.5°C (National OceanographicData Center [NODC] data for 2005 and 2009;http://www.nodc.noaa.gov) and temperature rangeis 23.8°C to 30.4°C (Pinto 1987). Our results indi-cate that the Rop for clade III clones was over23.7°C, at least; thus, it is assumed they have signifi-cant differences in ecophysiological characteristicsfor higher temperatures. Unfortunately, we couldnot detect the end values of the Rop because themethod used (second-order polynomial equations)relies on detecting the decreasing point of the curvecalculated by the equation; in our tested tempera-ture range, it could not be shown. A wider tempera-ture range for the growth experiment would benecessary to capture the entire range. We attributethe different temperature adaptations of P. pungensto its original growth environment. Our results arecompatible with the discussion on the marine stra-menopile group 4 (MAST-4), an uncultured groupof picoeukaryotes (Rodr�ıguez-Mart�ınez et al. 2013).The major factor leading to the worldwide distribu-tion patterns of MAST-4 populations was tempera-ture. Each population was separated into five cladesby a phylogenetic tree constructed by the ends of 18S rDNA and complete ITS1 sequences.
From the salinity test, we concluded that clade Iclones were euryhaline (Rop = salinity 21.1 to 35.8,Cmax = salinity of 28.4), while clades II and III arestenohaline (Rop = salinity 20.5 to 29.5 and 20.3 to29.0, Cmax = salinity of 25.0 and 24.7, respectively).Salinity is an important abiotic factor that affectsphytoplankton growth (Brand 1984, Prestegardet al. 2014). Species of the Pseudo-nitzschia genus arewidespread in coastal waters with various salinities(Hasle 2002, Thessen et al. 2005, Lelong et al.2012). In several studies, P. pungens did not growwell at low salinity, but, did so at a salinity of >30(Jackson et al. 1992, Thessen et al. 2005). Ourresults corroborate these studies, with the exceptionof one clone (HYMR1104PP34B1), which grew wellat a salinity of 10 (lmax, 1.09). Clade I clones stillgrew well at a high salinity (40), but clades II andIII did not. In the case of P. multiseries, the nearestneighbor species, some clones even grew well at >40(Jackson et al. 1992, Thessen et al. 2005). To under-stand what factors may have led to clade II and IIIclones failing to grow at higher salinities, we mayconsider the regions from which they were isolated.The surface waters of Puget Sound have an annualaverage salinity of 27. There is a large influx offreshwater from rivers and limited mixing with thehigher-salinity open ocean (Gelfenbaum et al.
2006). Pagbilao Bay (Philippines), the origin ofclade III clones, is located near the equator. Gener-ally, surface waters in equatorial regions have alower average salinity because of the long rainy sea-son. The seasonal variation in salinity ranges from11.3 to 33.7 in Pagbilao Bay (Pinto 1987). However,the salinity remains under 30 for most of the year,with the exception of periods during the dry season(Pinto 1987). Confirmation that these regions arecharacterized by lower salinities is provided byNODC (http://www.nodc.noaa.gov). We concludethat high salinity is one of the crucial factors limit-ing the growth of clade II and III clones.In marine environments, water depth and latitude
are two major factors that affect the amount of inso-lation that reaches phytoplankton habitats. Speciesinhabit their respective ecological niches in thesame water column depending on their differentlight intensity preferences. The cyanobacterium Pro-chlorococcus has different light-adaptation capabili-ties, depending on the water depth of their habitat,and two distinct ecotypes, with distinguishablegenetic variation, coexist (Moore et al. 1998). Thereis, however, no comparable research about latitudi-nal adaptation in relation to light intensity in phyto-plankton. The amount of insolation decreasesgradually with increasing latitude. Puget Sound islocated at 47° to 48°N latitude, where there is lowinsolation. From the results of our irradiance test,clade II clones have relatively lower lmax values ateach irradiance level, and lower Rop and Cmax val-ues, than the other clades. We therefore hypothe-size that the lower Rop and Cmax values of clade IIclones is consistent with the habitat conditions atthe latitude where the P. pungens was isolated. Weassumed that clade II clones would have been out-competed by clade I and III clones everywhere withregard to efficiency of capturing light. Our basicphysiological data provides some insight into therelationship between the latitudinal difference in aspecies’ origin and intraspecific differences in physi-ological response. However, lmax values from irradi-ance experiments were not significantly different inthe ANOVA test. Proof of this hypothesis thereforerequires more clonal isolates from various latitudes.The N:P ratio could be an important factor that
influences blooms of non-siliceous phytoplanktongroups in natural environments (Hodgkiss and Ho1997). Competition between diatoms and other phy-toplankton that do not require Si is normally influ-enced by silicate availability (Hecky and Kilham1988, Sommer 1994). In the nutrient test (N:P andN:Si), no statistical differences were observed in thelmax among clades I, II and III (Fig. 5, d and e).However, we determined the general physiologicalfeatures of P. pungens via the maximum cell abun-dance estimated from Figure S1. The highest N:Pratio condition (N:P = 100; N sufficient) showed alower maximum cell abundance than the N:P ratioof 16 (by 34.8%, Fig. S3a in the Supporting Infor-
INTRASPECIFIC DIVERSITY OF THREE CLADES OF PSEUDO-NITZSCHIA PUNGENS 169

mation). In the N:P tests, the amount of silicate wassufficient (the standard concentration in f/2 med-ium), although the nitrate concentration was highand potentially a stress. According to Hu et al.(2008), the lmax of P. pungens occurred at an N:Pratio of 10:1, and P. pungens required lower nitrateand phosphate levels compared to other species. Inthe N:Si ratio test, the estimated maximum cellabundance was diminished by only 15.8% in thecondition that lacked added Si (N:Si = 35) com-pared with the Si-sufficient condition (N:Si = 4);there were no significant differences in cell abun-dances among the N:Si ratios (Fig. S3b). It has beenobserved that blooms of P. pungens were onlyslightly affected by Si limitation. For example,P. pungens could be dominant at relatively low Si:Nratios compared with other diatoms (Sommer1994). Our results suggest that the growth of P. pun-gens is less affected by the low-nutrient conditions(N and Si), which could provide it with a competi-tive advantage.
In previous studies, clade III populations havebeen detected only from Vietnam, China, Mexicoand Portugal (Casteleyn et al. 2008, 2010).Although sequences from the strains from Mexicoand Portugal (the IIIb group) designated them asbeing clade III, the genetic difference with IIIaa was1.22% and 1.54% for IIIab. These gaps are greaterthan those found between clades I and II (1.20%).Geographically, the origins of clones IIIaa and IIIabare close, but the IIIb subgroup is far. For the firsttime, we report clade III P. pungens, belonging tothe IIIaa subgroup, in Korean coastal watersand the East China Sea. Previously, clones of theIIIa and IIIab subgroups were found only in SWPacific regions (Fig. 3), not farther northwest.These results are both ecologically and evolutionallyimportant. First ecologically, Korean coastal watersare located in a temperate zone and have a largeseasonal temperature range. Our results show thatclones of clade III did not grow well at lower tem-peratures (<15°C). From composite images of sea-surface temperature estimated by NOAA satellites,winter (January and February) Korean coastal watersrange from 5°C to 15°C, whereas summer andautumn (July to October) waters are stronglyaffected by warm currents (Isobe 1999, Takikawaand Yoon 2005), resulting in temperatures rangingfrom 20°C to 27°C. Clade III clonesHYNG1010PP29F5 and HYNH1010PP29B9 were iso-lated from Korean southern coastal waters in Octo-ber 2010. In addition, sequence data for clade IIIwere obtained from Masan Bay in late summer,2011, via direct PCR. The question thus remains asto whether clade III P. pungens was recently intro-duced into Korean coastal waters via warm currentsor ballast water, or has it always been present.
Because water temperature is increasing due toglobal warming, and populations are transported viacurrents or ballast water, inhabited environments
are continuously changing. In particular, the mostaccelerated sea surface temperature change in theNW Pacific, including Korean coastal waters (Han-sen et al. 2006). Different environmental prefer-ences might help ecological succession ordominance. Our results show that each P. pungensclade has slightly different adaptive strategies fortemperature (Table 3). In Masan survey, we couldnot find out the occurrence of clade III related withseawater temperature. However, clades I and IIIpopulations were coexisted in Korean coastal waterswhere the annual temperature range is 5°C to 27°C.It is difficult to speculate which P. pungens clade willdominate during different seasons and laterexpected warming. Development of new markersspecific to each clade and use of the quantitativereal-time PCR assay could help understand thedynamics of P. pungens clades. Actually, in case ofCochlodinium polykrikoides, their different ribotypeswere dominant during different seasons in Koreacoastal waters (Park et al. 2014).From an evolutionary viewpoint, it would be
important to know if clades I and III from Koreanwaters and the East China Sea can interbreed suc-cessfully. Churro et al. (2009) found that Ukrai-nian/USA clade I strains and Portuguese clade IIIstrains did not interbreed. However, these strainsoriginated from distant locations, and Portugueseclade III strains belonged to a different subgroup(IIIb). On the other hand, natural hybrids were pro-duced between clades I and II, which appeartogether in USA west-coast waters (Casteleyn et al.2009). Our sampling locations, where we observedboth clades I and III, are therefore potentiallyimportant “natural laboratories” for studying thegenetic flow and evolution of P. pungens.In conclusion, we observed differences in physio-
logical and morphological characteristics that corre-spond to clades. Our results show that the Rop fortemperature and salinity appear to be closely relatedto the environmental conditions of the habitatsfrom which the clones were originally isolated. Thisstudy provides the valuable information for under-standing the distribution of P. pungens, especiallyfor clades II and III.
This research was supported by a grant from the Marine Bio-technology Program, funded by the Ministry of Oceans andFisheries, Korea. Thanks to Vera Trainer and Brian D. Bill(NOAA) for providing cultures, Sunju Kim (ChonnamNational University) sampling assistance in the Philippines,and Stephen S. Bates (Fisheries and Oceans Canada) forcommenting on our manuscript and discussing our research.
Amato, A., Orsini, L., D’Alelio, D. & Montresor, M. 2005. Lifecycle, size reduction patterns, and ultrastructure of the pen-nate planktonic diatom Pseudo-nitzschia delicatissima (Bacillari-ophyceae). J. Phycol. 41:542–56.
Armbrust, E. V., Berges, J. A., Bowler, C., Green, B. R., Martinez,D., Putnam, N. H., Zhou, S., Allen, A. E., Apt, K. E. & Bech-ner, M. 2004. The genome of the diatom Thalassiosira pseudo-nana: ecology, evolution, and metabolism. Science 306:79–86.
170 JIN HO KIM ET AL.

Bates, S. S., Garrison, D. L. & Horner, R. A. 1998. Bloom dynam-ics and physiology of domoic-acid-producing Pseudo-nitzschiaspecies. In Anderson, D. M., Cembella, A. D. & Hallegraeff,G. M. [Eds.] Physiological Ecology of Harmful Algal Blooms.Springer-Verlag, Heidelberg, Germany, pp. 267–92.
Bowler, C., Allen, A. E., Badger, J. H., Grimwood, J., Jabbari, K.,Kuo, A., Maheswari, U., Martens, C., Maumus, F. & Otillar,R. P. 2008. The Phaeodactylum genome reveals the evolution-ary history of diatom genomes. Nature 456:239–44.
Brand, L. E. 1984. The salinity tolerance of forty-six marine phy-toplankton isolates. Est. Coast. Shelf Sci. 18:543–56.
Brand, L. E., Murphy, L. S., Guillard, R. R. L., Lee, H. T. 1981.Genetic variability and differentiation in the temperatureniche component of the diatom Thalassiosira pseudonana.Mar. Biol. 62:103–10.
Casteleyn, G., Adams, N. G., Vanormelingen, P., Debeer, A. E.,Sabbe, K. & Vyverman, W. 2009. Natural hybrids in the mar-ine diatom Pseudo-nitzschia pungens (Bacillariophyceae):genetic and morphological evidence. Protist 160:343–54.
Casteleyn, G., Chepurnov, V. A., Leliaert, F., Mann, D. G., Bates,S. S., Lundholm, N., Rhodes, L., Sabbe, K. & Vyverman, W.2008. Pseudo-nitzschia pungens (Bacillariophyceae): a cosmo-politan diatom species? Harmful Algae 7:241–57.
Casteleyn, G., Leliaert, F., Backeljau, T., Debeer, A. E., Kotaki, Y.,Rhodes, L., Lundholm, N., Sabbe, K. & Vyverman, W. 2010.Limits to gene flow in a cosmopolitan marine planktonic dia-tom. Proc. Natl. Acad. Sci. USA 107:12952–7.
Chepurnov, V. A., Mann, D. G., Sabbe, K., Vannerum, K., Casteleyn,G., Verleyen, E., Peperzak, L. & Vyverman, W. 2005. Sexualreproduction, mating system, chloroplast dynamics andabrupt cell size reduction in Pseudo-nitzschia pungens from theNorth Sea (Bacillariophyta). Eur. J. Phycol. 40:379–95.
Cho, E. S., Kotaki, Y. & Park, J. G. 2001a. The comparisonbetween toxic Pseudo-nitzschia multiseries (Hasle) Hasle andnon-toxic P. pungens (Grunow) Hasle isolated from JinhaeBay. Korea. Algae 16:275–85.
Cho, E. S., Park, J. G., Oh, B. C. & Cho, Y. C. 2001b. The applica-tion of species specific DNA-targeted probes and fluorescentlytagged lectin to differentiate several species of Pseudo-nitzschia(Bacillophyceae) in Chinhae Bay, Korea. Sci. Mar. 65:207–14.
Churro, C. I., Carreira, C. C., Rodrigues, F. J., Craveiro, S. C., Ca-lado, A. J., Casteleyn, G. & Lundholm, N. 2009. Diversity andabundance of potentially toxic Pseudo-nitzschia Peragallo inAveiro coastal lagoon, Portugal and description of a new vari-ety, P. pungens var. aveirensis var. nov. Diatom Res. 24:35–62.
Davidovich, N. A. & Bates, S. S. 1998. Sexual reproduction in thepennate diatoms Pseudo-nitzschia multiseries and P. pseudodelica-tissima (Bacillariophyceae). J. Phycol. 34:126–37.
Ellegaard, M., Godhe, A., H€arnstr€om, K. & McQuoid, M. 2008.The species concept in a marine diatom: LSU rDNA-basedphylogenetic differentiation in Skeletonema marinoi/dohrnii(Bacillariophyceae) is not reflected in morphology. Phycologia47:156–67.
Ellegren, H. & Sheldon, B. C. 2008. Genetic basis of fitness differ-ences in natural populations. Nature 452:169–75.
Felsenstein, J. 1985. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39:783–91.
Gallagher, J. C. 1982. Physiological variation and electrophoreticbanding patterns of genetically different seasonal popula-tions of Skeletonema costatum (Bacillariophycee). J. Phycol.18:148–62.
Gallagher, J. C., Wood, A. M. & Alberte, R. S. 1984. Ecotypic dif-ferentiation in the marine diatom Skeletonema costatum: influ-ence of light intensity on the photosynthetic apparatus. Mar.Biol. 82:121–34.
Gelfenbaum, G., Mumford, T., Brennan, J., Case, H., Dethier, M.,Fresh, K., Goetz, F., van Heeswijk, M., Leschine, T. M. &Logsdon, M. 2006. Coastal habitats in Puget Sound: aresearch plan in support of the Puget Sound Nearshore Part-nership. Partnership Report No. 2006-1. U.S. Geological Sur-vey, Seattle, Washington. Available at http://pugetsoundnearshore.org.
Grinnell, J. 1917. The niche-relationships of the CaliforniaThrasher. Auk 34:427–33.
Guillard, R. R. L. 1973. Division rates. Handbook of PhycologicalMethods 1:289–312.
Guillard, R. R. L. & Ryther, J. H. 1962. Studies of marine plank-tonic diatoms. I. Cyclotella nana Hustedt and Detonula conferv-acea Cleve. Can. J. Microbiol. 8:229–39.
Guisan, A. & Thuiller, W. 2005. Predicting species distribution:offering more than simple habitat models. Ecol. Let. 8:993–1009.
Hale, M. S. & Mitchell, J. G. 2001. Functional morphology of dia-tom frustule microstructures: hydrodynamic control ofBrownian particle diffusion and advection. Aquat. Microb.Ecol. 24:287–95.
Hansen, J., Sato, M., Ruedy, R., Lo, K., Lea, D. W. & Medina-Eli-zade, M. 2006. Global temperature change. Proc. Natl. Acad.Sci. USA 103:14288–93.
Hasle, G. R. 2002. Are most of the domoic acid-producing speciesof the diatom genus Pseudo-nitzschia cosmopolites? HarmfulAlgae 1:137–46.
Hecky, R. E. & Kilham, P. 1988. Nutrient limitation of phyto-plankton in freshwater and marine environments: a review ofrecent evidence on the effects of enrichment. Limnol. Ocea-nogr. 33:796–822.
Hendey, N. 1974. The permanganate method for cleaning freshlygathered diatoms. Microscopy 32:423–6.
Hodgkiss, I. J. & Ho, K. C. 1997. Are changes in N: P ratios incoastal waters the key to increased red tide blooms? InWong, Y. S. & Tam, F. Y. [Ed.] Asia-Pacific Conference on Sci-ence and Management of Coastal Environment. Springer, Hydro-biologia, pp. 141–7.
Hu, Z. X., Xu, N., Li, A. F. & Duan, S. S. 2008. Effects of differ-ent N: P ratios on the growth of Pseudo-nitzschia pungens,Prorocentrum donghaiense and Phaeocystis globosa. Acta Hydrobiol.Sin. 4:482–7.
Isobe, A. 1999. On the origin of the Tsushima Warm Currentand its seasonality. Cont. Shelf Res. 19:117–34.
Jackson, A. E., Ayer, S. W. & Laycock, M. V. 1992. The effect ofsalinity on growth and amino acid composition in the mar-ine diatom Nitzschia pungens. Can. J. Bot. 70:2198–201.
Jacques, G. 1983. Some ecophysiological aspects of the Antarcticphytoplankton. Polar Biol. 2:27–33.
Ki, J. S., Jang, G. Y. & Han, M. S. 2004. Integrated method forsingle-cell DNA extraction, PCR amplification, and sequenc-ing of ribosomal DNA from harmful dinoflagellates Cochlodi-nium polykrikoides and Alexandrium catenella. Mar. Biotechnol.6:587–93.
Kibler, S. R., Litaker, R. W., Holland, W. C., Vandersea, M. W. &Tester, P. A. 2012. Growth of eight Gambierdiscus (Dinophy-ceae) species: effects of temperature, salinity and irradiance.Harmful Algae 19:1–14.
Kooistra, W. H. C. F., Diana, S., Sergio, B., Gu, H., Anderson, R.A. & Zingone, A. 2008. Global diversity and biogeography ofSkeletonema species (Bacillariophyta). Protist 159:177–93.
Lelong, A., H�egaret, H., Soudant, P. & Bates, S. S. 2012. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amne-sic shellfish poisoning: revisiting previous paradigms. Phycolo-gia 51:168–216.
Leterme, S. C., Prime, E., Mitchell, J., Brown, M. H. & Ellis, A. V.2013. Diatom adaptability to environmental change: a casestudy of two Cocconeis species from high-salinity areas. DiatomRes. 28:29–35.
Lewis, N. I., Bates, S. S., McLachlan, J. L. & Smith, J. C. 1993.Temperature effects on growth, domoic acid production,and morphology of the diatom Nitzschia pungens f. multiseries.In Smayda, T. J. & Shimizu, Y. [Eds.] Toxic PhytoplanktonBlooms in the Sea. Elsevier Sci. Publ. B.V., Amsterdam, TheNetherlands, pp. 601–6.
Lim, H. C., Lim, P. T., Teng, S. T., Bates, S. S. & Leaw, C. P.2014. Genetic structure of Pseudo-nitzschia pungens (Bacillario-phyceae) populations: implications of a global diversificationof the diatom. Harmful Algae 37:142–52.
INTRASPECIFIC DIVERSITY OF THREE CLADES OF PSEUDO-NITZSCHIA PUNGENS 171

Lundholm, N., Moestrup, Ø., Hasle, G. R. & Hoef-Emden, K.2003. A study of the Pseudo-nitzschia pseucodelicatissima/cuspi-data complex (Bacillariophycae): what is P. pseudodelicatis-sima? J. Phycol. 39:797–813.
Medlin, L. K., Lange, M. & Nothig, E. 2000. Genetic diversity inthe marine phytoplankton: a review and a consideration ofAntarctic phytoplankton. Antarctic Sci. 12:325–33.
Montresor, M., Sgrosso, S., Procaccini, G. & Kooistra, W. H. C. F.2003. Intraspecific diversity in Scrippsiella trochoidea (Dinopby-ceae): evidence for cryptic species. Phycologia 42:56–70.
Moore, L. R., Rocap, G. & Chisholm, S. W. 1998. Physiology andmolecular phylogeny of coexisting Prochlorococcus ecotypes.Nature 393:464–7.
Nei, M. & Kumar, S. 2000. Molecular Evolution and Phylogenetics.Oxford University Press, New York, 333 pp.
Park, B. S., Wang, P., Kim, J. H., Kim, J. H., Gobler, C. J. & Han,M. S. 2014. Resolving the intra-specific succession withinCochlodinium polykrikoides populations in southern Koreancoastal waters via use of quantitative PCR assays. HarmfulAlgae. 37:133–41.
Pinto, L. 1987. Environmental factors influencing the occurrenceof juvenile fish in the mangroves of Pagbilao, Philippines.Hydrobiologia 150:283–301.
Prestegard, S. K., Knutsen, G. & Herfindal, L. 2014. Adenosinecontent and growth in the diatom Phaeodactylum tricornutum(Bacillariophyceae): effect of salinity, light, temperature andnitrate. Diatom Res. 28:361–9.
Rhodes, L., White, D., Syhre, M. & Atkinson, M. 1996. Pseudo-nitzs-chia species isolated from New Zealand coastal waters: do-moic acid production in vitro and links with shellfishtoxicity. In Yasumoto, T., Oshima, Y. & Fukuyo, Y. [Eds.]Harmful and Toxic Algal Blooms. Intergov. Oceanogr. Comm.,UNESCO, Paris, France, pp. 155–8.
Rodr�ıguez-Mart�ınez, R., Rocap, G., Salazar, G. & Massana, R.2013. Biogeography of the uncultured marine picoeukaryoteMAST-4: temperature-driven distribution patterns. ISME J.7:1531–43.
Saraceni, C. & Ruggiu, D. 1974. Techniques for sampling waterand phytoplankton. A manual on methods for measuringprimary production in aquatic environments. IBP Handbook12:5–7.
Schluter, D. 2001. Ecology and the origin of species. Trends Ecol.Evol. 16:372–80.
Smith, K. E. & Flocks, J. G. 2010. Environmental investigationsusing diatom microfossils. US Geological Survey Fact Sheet2010–3115:1.
Soberon, J. 2005. Interpretation of models of fundamental eco-logical niches and species’ distributional areas. Biodivers. In-format. 2:1–10.
Sommer, U. 1994. Are marine diatoms favoured by high Si: Nratios? Mar. Ecol. Prog. Ser. 115:309–15.
Stehr, C. M., Connell, L., Baugh, K. A., Bill, B. D., Adams, N. G.& Trainer, V. L. 2002. Morphological, toxicological, andgenetic differences among Pseudo-nitzschia (Bacillariophy-ceae) species in inland embayments and outer coastal watersof Washington State, USA. J. Phycol. 38:55–65.
Takikawa, T. & Yoon, J. H. 2005. Volume transport through theTsushima Straits estimated from sea level difference. J. Ocea-nogr. 61:699–708.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. &Kumar, S. 2011. MEGA5: molecular evolutionary geneticsanalysis using maximum likelihood, evolutionary distance,and maximum parsimony methods. Molec. Biol. Evol.28:2731–9.
Thessen, A. E., Bowers, H. A. & Stoecker, D. K. 2009. Intra- andinterspecies differences in growth and toxicity of Pseudo-nitzschia while using different nitrogen sources. HarmfulAlgae. 8:792–810.
Thessen, A. E., Dortch, Q., Parsons, M. L. & Morrison, W. 2005.Effect of salinity on Pseudo-nitzschia species (Bacillariophy-ceae) growth and distribution1. J. Phycol. 41:21–9.
Trainer, V. L., Bates, S. S., Lundholm, N., Thessen, A. E., Coch-lan, W. P., Adams, N. G. & Trick, C. G. 2012. Pseudo-nitzschiaphysiological ecology, phylogeny, toxicity, monitoring andimpacts on ecosystem health. Harmful Algae 14:271–300.
Trainer, V. L., Wekell, J. C., Horner, R. A., Hatfield, C. L. &Stein, J. E. 1998. Domoic acid production by Pseudo-nitzschiapungens. In Reguera, B., Blanco, J., Fern�andez, M. L. &Wyatt, T. [Eds.] Harmful Algae. Xunta de Galicia and theInternational Oceanographic Commission of UNESCO,Paris, pp. 337–40.
Villac, M. C. & Fryxell, G. A. 1998. Pseudo-nitzschia pungens var.cingulata var. nov. (Bacillariophyceae) based on field and cul-ture observations. Phycologia 37:269–74.
Wang, P., Park, B. S., Kim, J. H., Kim, J. H., Lee, H. O. & Han,M.-S. 2014. Phylogenetic position of eight Amphora sensu lato(Bacillariophyceae) species and comparative analysis of mor-phological characteristics. Algae 29:57–73.
Supporting Information
Additional Supporting Information may befound in the online version of this article at thepublisher’s web site:
Figure S1. Relationship between the cell con-centration of nine clones of Pseudo-nitzschia pun-gens and in vivo fluorescence, and the resultingcoefficient of determination (R2) values are indi-cated at each graph.
Figure S2. Maximum growth rate of the ninestrains (I-III; n = 4, 2 and 3, respectively) at differ-ent temperature, salinity, irradiance, N:P and N:Siratios. Black = clade I; gray = clade II;white = clade III.
Figure S3. Relative estimated maximum cellabundance of nine Pseudo-nitzschia pungens cul-tures in the N:P (a) and N:Si (b) experiments.
172 JIN HO KIM ET AL.
Related Documents











