Neuron Article Intra-axonal Patterning: Intrinsic Compartmentalization of the Axonal Membrane in Drosophila Neurons Takeo Katsuki, 1,2 Deepak Ailani, 1,2 Masaki Hiramoto, 1,3 and Yasushi Hiromi 1,2, * 1 Department of Developmental Genetics, National Institute of Genetics, 1111 Yata, Mishima, Shizuoka 411-8540, Japan 2 Department of Genetics, SOKENDAI, 1111 Yata, Mishima, Shizuoka 411-8540, Japan 3 Present address: ICND 216, The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA *Correspondence: [email protected] DOI 10.1016/j.neuron.2009.08.019 SUMMARY In the developing nervous system, distribution of membrane molecules, particularly axon guidance receptors, is often restricted to specific segments of axons. Such localization of membrane molecules can be important for the formation and function of neural networks; however, how this patterning within axons is achieved remains elusive. Here we show that Drosophila neurons in culture establish intra-axonal patterns in a cell-autonomous manner; several membrane molecules localize to either proximal or distal axon segments without cell-cell contacts. This distinct patterning of membrane proteins is not explained by a simple temporal control of expression, and likely involves spatially controlled vesicular targeting or retrieval. Mobility of transmembrane molecules is restricted at the boundary of intra-axonal segments, indicating that the axonal membrane is compartmentalized by a barrier mechanism. We propose that this intra- axonal compartmentalization is an intrinsic property of Drosophila neurons that provides a basis for the structural and functional development of the nervous system. INTRODUCTION The process by which different parts of the body acquire distinct properties such as morphology, function, and molecular distri- bution is generally called patterning. Likewise, patterning events that take place within a single cell may be called intracellular patterning. In the nervous system of vertebrates and inverte- brates, different regions of axons are often characterized by differential expression of membrane molecules (Bastiani et al., 1987; Brittis et al., 2002; Callahan et al., 1995; Dodd et al., 1988; Kidd et al., 1998; Patel et al., 1987; Rajagopalan et al., 2000a, 2000b; Simpson et al., 2000a, 2000b), suggesting that axons are ‘‘patterned’’ into intra-axonal segments. For example, in the ventral nerve cord of Drosophila embryos, axon guidance receptor Roundabout (ROBO), and other members of this family, ROBO2 and ROBO3, accumulate on longitudinal axon tracts, but are excluded from commissures (Kidd et al., 1998; Rajago- palan et al., 2000a, 2000b; Simpson et al., 2000a, 2000b) (Figures 1A and 1C). This specific distribution pattern is due to the localization of ROBO proteins to distal axon segments of individual commissural neurons (Kidd et al., 1998; Simpson et al., 2000a)(Figure 1D). The distribution of another guidance receptor, Derailed (DRL), which, by contrast, is enriched on the anterior commissures, provides a second example of this regulated distribution (Bonkowsky et al., 1999; Callahan et al., 1995)(Figures 1B–1D). In the spinal cord of mouse embryos, commissural axons express elevated levels of Robo1 and Robo2 proteins on the axon segments that have crossed the floor plate, while an isoform of Rig-1/Robo3 is mostly present on the precrossing axon segment (Chen et al., 2008; Sabatier et al., 2004). Thus, the spatial regulation of axon guidance receptors within an axon appears to be conserved across species. Despite the generality of the intra-axonal localization of membrane molecules, little is known about how such elabo- rate patterns emerge. These patterns may result from the influ- ence of extrinsic cues, an intrinsic ability of cells, or both. It is also elusive how polarized distribution of these membrane proteins along axons can be established and maintained without diffusing into a uniform distribution. Here, using a low-density primary cell culture prepared from Drosophila embryos, we show that neurons possess an ability to generate intra-axonal patterns of membrane molecules cell-autono- mously. ROBO3 and ROBO2 are localized to distal axon segments, while DRL is localized to proximal axon segments in a complementary manner. These localization patterns are not explained by a simple temporal control of receptor expres- sion, and likely involve spatially controlled vesicular targeting or retrieval pathways. The temperature-sensitive dynamin mutant reveals that DRL requires Dynamin-dependent endocy- tosis for its localization to proximal axon segments, whereas ROBO3 localization is relatively insensitive to blocking Dyna- min function. We also show that the exchange of membrane proteins between the distal and proximal axon segments is restricted at a medial point of the axon, which may maintain the compartmentalized distribution of membrane proteins along axons. 188 Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neuron
Article
Intra-axonal Patterning: IntrinsicCompartmentalization of the Axonal Membranein Drosophila NeuronsTakeo Katsuki,1,2 Deepak Ailani,1,2 Masaki Hiramoto,1,3 and Yasushi Hiromi1,2,*1Department of Developmental Genetics, National Institute of Genetics, 1111 Yata, Mishima, Shizuoka 411-8540, Japan2Department of Genetics, SOKENDAI, 1111 Yata, Mishima, Shizuoka 411-8540, Japan3Present address: ICND 216, The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA*Correspondence: [email protected]
DOI 10.1016/j.neuron.2009.08.019
SUMMARY
In the developing nervous system, distribution ofmembrane molecules, particularly axon guidancereceptors, is often restricted to specific segmentsof axons. Such localization of membrane moleculescan be important for the formation and functionof neural networks; however, how this patterningwithin axons is achieved remains elusive. Here weshow that Drosophila neurons in culture establishintra-axonal patterns in a cell-autonomous manner;several membrane molecules localize to eitherproximal or distal axon segments without cell-cellcontacts. This distinct patterning of membraneproteins is not explained by a simple temporalcontrol of expression, and likely involves spatiallycontrolled vesicular targeting or retrieval. Mobilityof transmembrane molecules is restricted at theboundary of intra-axonal segments, indicating thatthe axonal membrane is compartmentalized bya barrier mechanism. We propose that this intra-axonal compartmentalization is an intrinsic propertyof Drosophila neurons that provides a basis for thestructural and functional development of the nervoussystem.
INTRODUCTION
The process by which different parts of the body acquire distinct
properties such as morphology, function, and molecular distri-
bution is generally called patterning. Likewise, patterning events
that take place within a single cell may be called intracellular
patterning. In the nervous system of vertebrates and inverte-
brates, different regions of axons are often characterized by
differential expression of membrane molecules (Bastiani et al.,
1987; Brittis et al., 2002; Callahan et al., 1995; Dodd et al., 1988;
Kidd et al., 1998; Patel et al., 1987; Rajagopalan et al., 2000a,
2000b; Simpson et al., 2000a, 2000b), suggesting that axons
are ‘‘patterned’’ into intra-axonal segments. For example, in
the ventral nerve cord of Drosophila embryos, axon guidance
188 Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc.
receptor Roundabout (ROBO), and other members of this family,
ROBO2 and ROBO3, accumulate on longitudinal axon tracts,
but are excluded from commissures (Kidd et al., 1998; Rajago-
palan et al., 2000a, 2000b; Simpson et al., 2000a, 2000b)
(Figures 1A and 1C). This specific distribution pattern is due to
the localization of ROBO proteins to distal axon segments of
individual commissural neurons (Kidd et al., 1998; Simpson
et al., 2000a) (Figure 1D). The distribution of another guidance
receptor, Derailed (DRL), which, by contrast, is enriched on
the anterior commissures, provides a second example of this
regulated distribution (Bonkowsky et al., 1999; Callahan et al.,
1995) (Figures 1B–1D). In the spinal cord of mouse embryos,
commissural axons express elevated levels of Robo1 and
Robo2 proteins on the axon segments that have crossed the
floor plate, while an isoform of Rig-1/Robo3 is mostly present
on the precrossing axon segment (Chen et al., 2008; Sabatier
et al., 2004). Thus, the spatial regulation of axon guidance
receptors within an axon appears to be conserved across
species.
Despite the generality of the intra-axonal localization of
membrane molecules, little is known about how such elabo-
rate patterns emerge. These patterns may result from the influ-
ence of extrinsic cues, an intrinsic ability of cells, or both. It is
also elusive how polarized distribution of these membrane
proteins along axons can be established and maintained
without diffusing into a uniform distribution. Here, using
a low-density primary cell culture prepared from Drosophila
embryos, we show that neurons possess an ability to generate
intra-axonal patterns of membrane molecules cell-autono-
mously. ROBO3 and ROBO2 are localized to distal axon
segments, while DRL is localized to proximal axon segments
in a complementary manner. These localization patterns are
not explained by a simple temporal control of receptor expres-
sion, and likely involve spatially controlled vesicular targeting
or retrieval pathways. The temperature-sensitive dynamin
mutant reveals that DRL requires Dynamin-dependent endocy-
tosis for its localization to proximal axon segments, whereas
ROBO3 localization is relatively insensitive to blocking Dyna-
min function. We also show that the exchange of membrane
proteins between the distal and proximal axon segments is
restricted at a medial point of the axon, which may maintain
the compartmentalized distribution of membrane proteins
along axons.
Neuron
Intra-axonal Patterning in Drosophila Neurons
RESULTS
Intra-axonal Localization of Axon Guidance ReceptorsCan Be Generated Cell AutonomouslyWe sought to examine whether Drosophila neurons possess any
intrinsic ability to generate intra-axonal distribution patterns of
membrane molecules, and if so, by what mechanism. To explore
the cell-autonomous properties of Drosophila neurons, we used
a low-density primary cell culture system, in which neurons
extend their axons in the absence of cell-cell contacts. We tested
whether or not neurons placed in culture exhibit a localized distri-
bution of ROBO receptors. Immunostaining revealed that ROBO
A
E
F
G
H
I
J
K
L
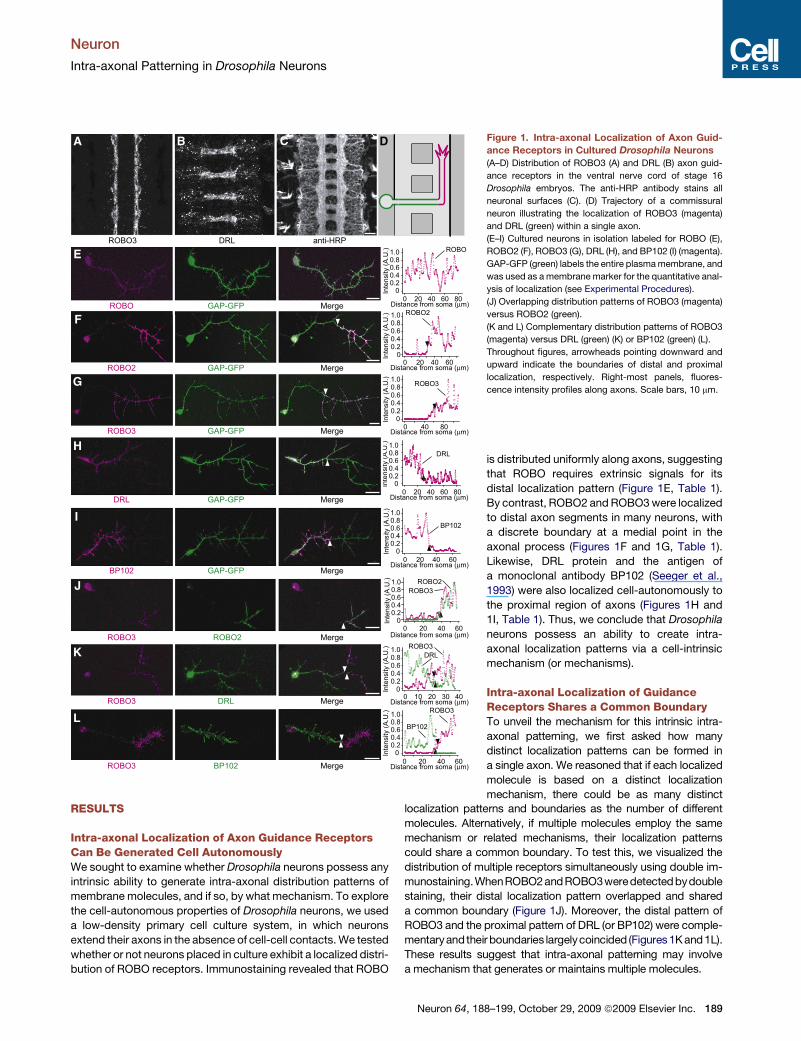
B C D Figure 1. Intra-axonal Localization of Axon Guid-
ance Receptors in Cultured Drosophila Neurons
(A–D) Distribution of ROBO3 (A) and DRL (B) axon guid-
ance receptors in the ventral nerve cord of stage 16
Drosophila embryos. The anti-HRP antibody stains all
neuronal surfaces (C). (D) Trajectory of a commissural
neuron illustrating the localization of ROBO3 (magenta)
and DRL (green) within a single axon.
(E–I) Cultured neurons in isolation labeled for ROBO (E),
ROBO2 (F), ROBO3 (G), DRL (H), and BP102 (I) (magenta).
GAP-GFP (green) labels the entire plasma membrane, and
was used as a membrane marker for the quantitative anal-
ysis of localization (see Experimental Procedures).
(J) Overlapping distribution patterns of ROBO3 (magenta)
versus ROBO2 (green).
(K and L) Complementary distribution patterns of ROBO3
(magenta) versus DRL (green) (K) or BP102 (green) (L).
Throughout figures, arrowheads pointing downward and
upward indicate the boundaries of distal and proximal
localization, respectively. Right-most panels, fluores-
cence intensity profiles along axons. Scale bars, 10 mm.
is distributed uniformly along axons, suggesting
that ROBO requires extrinsic signals for its
distal localization pattern (Figure 1E, Table 1).
By contrast, ROBO2 and ROBO3 were localized
to distal axon segments in many neurons, with
a discrete boundary at a medial point in the
axonal process (Figures 1F and 1G, Table 1).
Likewise, DRL protein and the antigen of
a monoclonal antibody BP102 (Seeger et al.,
1993) were also localized cell-autonomously to
the proximal region of axons (Figures 1H and
1I, Table 1). Thus, we conclude that Drosophila
neurons possess an ability to create intra-
axonal localization patterns via a cell-intrinsic
mechanism (or mechanisms).
Intra-axonal Localization of GuidanceReceptors Shares a Common BoundaryTo unveil the mechanism for this intrinsic intra-
axonal patterning, we first asked how many
distinct localization patterns can be formed in
a single axon. We reasoned that if each localized
molecule is based on a distinct localization
mechanism, there could be as many distinct
localization patterns and boundaries as the number of different
molecules. Alternatively, if multiple molecules employ the same
mechanism or related mechanisms, their localization patterns
could share a common boundary. To test this, we visualized the
distribution of multiple receptors simultaneously using double im-
munostaining.WhenROBO2andROBO3weredetectedbydouble
staining, their distal localization pattern overlapped and shared
a common boundary (Figure 1J). Moreover, the distal pattern of
ROBO3 and the proximal pattern of DRL (or BP102) were comple-
mentaryandtheir boundaries largelycoincided (Figures 1K and1L).
These results suggest that intra-axonal patterning may involve
a mechanism that generates or maintains multiple molecules.
Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc. 189
Neuron
Intra-axonal Patterning in Drosophila Neurons
Localization Patterns of Receptors Are Not Determinedby Their Temporal Expression ProfilesA simple mechanism for intra-axonal localization of guidance
receptors would be to control distribution by the temporal
expression profile of the receptors at the growth cone during
axonal elongation (Dodd et al., 1988; Keleman et al., 2002). For
example, the complementary distribution pattern of DRL and
ROBO3 could be generated if the expression of DRL and
ROBO3 switch at a certain time point in axonal elongation;
DRL may be expressed during the earlier stages of axonal elon-
gation, whereas ROBO3 may be expressed only after the
expression of DRL has been turned off. This hypothesis implies
that the spatial pattern along the proximal-distal axis of the
axon is generated from information on the time axis.
To test this possibility, we manipulated the timing of receptor
expression using GAL4/UAS-based systems. When EGFP-
tagged receptors (hereafter referred to as ROBO-EGFP, etc.)
were expressed throughout axonal elongation under the control
of elav-GAL4, EGFP-tagged receptors showed localization
patterns similar to those of endogenous proteins; ROBO-EGFP
was uniformly distributed, ROBO2-EGFP and ROBO3-EGFP
were localized to the distal axon segment, and DRL-EGFP was
localized to the proximal axon segment (Figures 2A–2D, Table 2).
Even when ROBO3-mRFP and DRL-EGFP were simultaneously
expressed in the same cell, they showed complementary locali-
zation patterns with similar or identical boundary positions (Fig-
ures 2E–2G). Complementary localization of tagged ROBO3 and
DRL molecules was also verified in vivo, by coexpression in
a subset of identified neurons under the control of a GAL4 driver
(Figure S1 available online). These results indicate that localiza-
tion is not determined by the temporal profile of receptor expres-
sion at the transcriptional level.
Because the above result does not rule out the possibility that
localization is determined by temporal control at the posttran-
scriptional level, we next used an expression system that can
switch on the expression of transgenic receptors by a brief
heat pulse (see Experimental Procedures for details). This
system enables us to delay expression, including at the posttran-
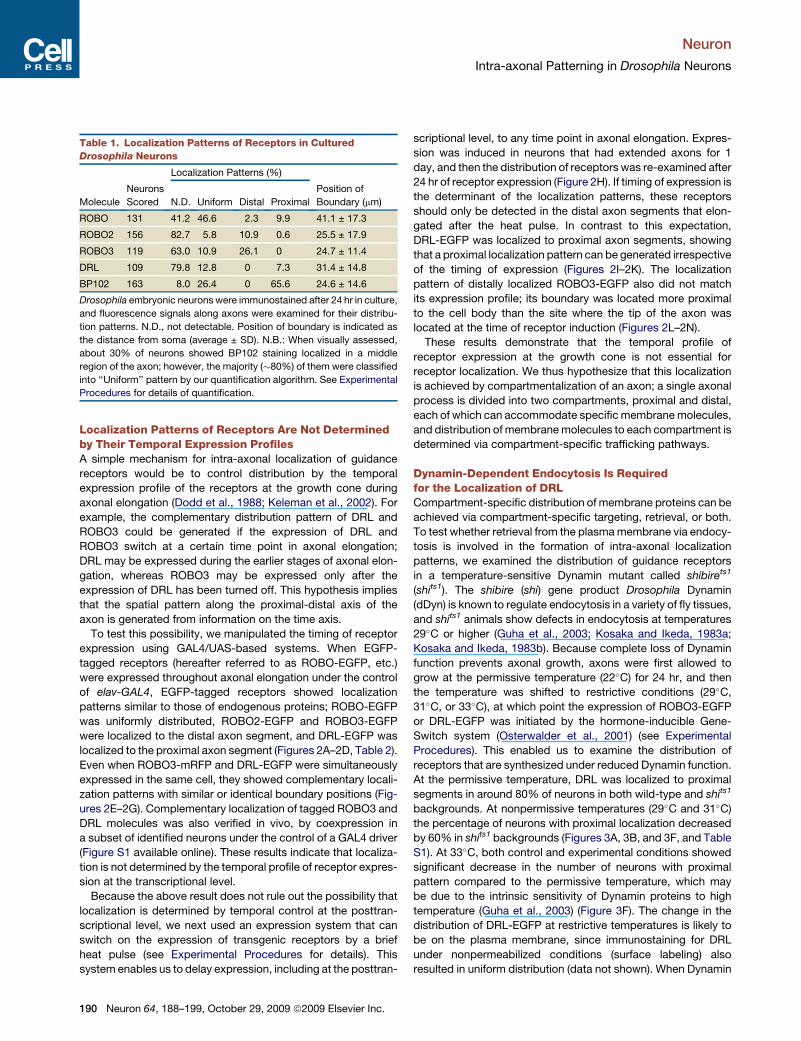
Table 1. Localization Patterns of Receptors in Cultured
Drosophila Neurons
Localization Patterns (%)
Molecule
Neurons
Scored N.D. Uniform Distal Proximal
Position of
Boundary (mm)
ROBO 131 41.2 46.6 2.3 9.9 41.1 ± 17.3
ROBO2 156 82.7 5.8 10.9 0.6 25.5 ± 17.9
ROBO3 119 63.0 10.9 26.1 0 24.7 ± 11.4
DRL 109 79.8 12.8 0 7.3 31.4 ± 14.8
BP102 163 8.0 26.4 0 65.6 24.6 ± 14.6
Drosophila embryonic neurons were immunostained after 24 hr in culture,
and fluorescence signals along axons were examined for their distribu-
tion patterns. N.D., not detectable. Position of boundary is indicated as
the distance from soma (average ± SD). N.B.: When visually assessed,
about 30% of neurons showed BP102 staining localized in a middle
region of the axon; however, the majority (�80%) of them were classified
into ‘‘Uniform’’ pattern by our quantification algorithm. See Experimental
Procedures for details of quantification.
190 Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc.
scriptional level, to any time point in axonal elongation. Expres-
sion was induced in neurons that had extended axons for 1
day, and then the distribution of receptors was re-examined after
24 hr of receptor expression (Figure 2H). If timing of expression is
the determinant of the localization patterns, these receptors
should only be detected in the distal axon segments that elon-
gated after the heat pulse. In contrast to this expectation,
DRL-EGFP was localized to proximal axon segments, showing
that a proximal localization pattern can be generated irrespective
of the timing of expression (Figures 2I–2K). The localization
pattern of distally localized ROBO3-EGFP also did not match
its expression profile; its boundary was located more proximal
to the cell body than the site where the tip of the axon was
located at the time of receptor induction (Figures 2L–2N).
These results demonstrate that the temporal profile of
receptor expression at the growth cone is not essential for
receptor localization. We thus hypothesize that this localization
is achieved by compartmentalization of an axon; a single axonal
process is divided into two compartments, proximal and distal,
each of which can accommodate specific membrane molecules,
and distribution of membrane molecules to each compartment is
determined via compartment-specific trafficking pathways.
Dynamin-Dependent Endocytosis Is Requiredfor the Localization of DRLCompartment-specific distribution of membrane proteins can be
achieved via compartment-specific targeting, retrieval, or both.
To test whether retrieval from the plasma membrane via endocy-
tosis is involved in the formation of intra-axonal localization
patterns, we examined the distribution of guidance receptors
in a temperature-sensitive Dynamin mutant called shibirets1
(shits1). The shibire (shi) gene product Drosophila Dynamin
(dDyn) is known to regulate endocytosis in a variety of fly tissues,
and shits1 animals show defects in endocytosis at temperatures
29�C or higher (Guha et al., 2003; Kosaka and Ikeda, 1983a;
Kosaka and Ikeda, 1983b). Because complete loss of Dynamin
function prevents axonal growth, axons were first allowed to
grow at the permissive temperature (22�C) for 24 hr, and then
the temperature was shifted to restrictive conditions (29�C,
31�C, or 33�C), at which point the expression of ROBO3-EGFP
or DRL-EGFP was initiated by the hormone-inducible Gene-
Switch system (Osterwalder et al., 2001) (see Experimental
Procedures). This enabled us to examine the distribution of
receptors that are synthesized under reduced Dynamin function.
At the permissive temperature, DRL was localized to proximal
segments in around 80% of neurons in both wild-type and shits1
backgrounds. At nonpermissive temperatures (29�C and 31�C)
the percentage of neurons with proximal localization decreased
by 60% in shits1 backgrounds (Figures 3A, 3B, and 3F, and Table
S1). At 33�C, both control and experimental conditions showed
significant decrease in the number of neurons with proximal
pattern compared to the permissive temperature, which may
be due to the intrinsic sensitivity of Dynamin proteins to high
temperature (Guha et al., 2003) (Figure 3F). The change in the
distribution of DRL-EGFP at restrictive temperatures is likely to
be on the plasma membrane, since immunostaining for DRL
under nonpermeabilized conditions (surface labeling) also
resulted in uniform distribution (data not shown). When Dynamin
Neuron
Intra-axonal Patterning in Drosophila Neurons
function was restored by down-shifting the temperature to
a permissive condition (a 5 hr chase), the proximal localization
pattern increased by about 30% (data not shown). These results
suggest that the proximal localization of DRL on the axonal
membrane requires Dynamin-dependent endocytosis.
In contrast to DRL, the distal localization of ROBO3 was only
mildly affected by the shits1 mutation: the number of neurons
A
E
F G
H
I J K
NML
B
C
D
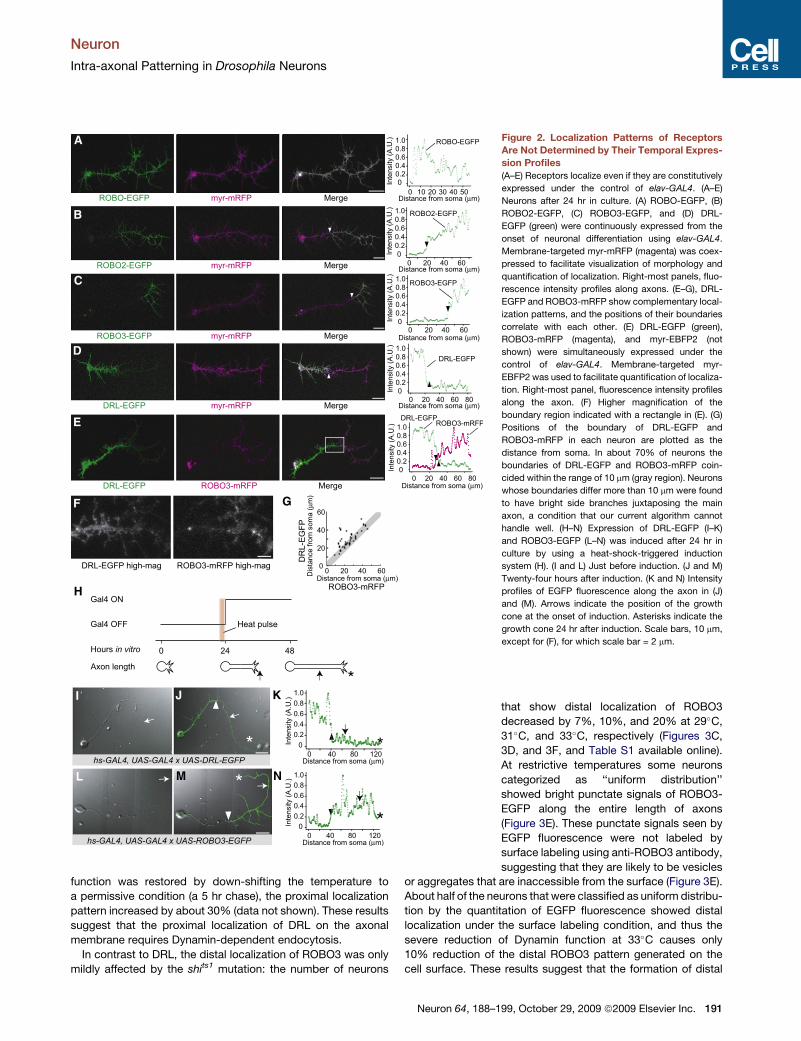
Figure 2. Localization Patterns of Receptors
Are Not Determined by Their Temporal Expres-
sion Profiles
(A–E) Receptors localize even if they are constitutively
expressed under the control of elav-GAL4. (A–E)
Neurons after 24 hr in culture. (A) ROBO-EGFP, (B)
ROBO2-EGFP, (C) ROBO3-EGFP, and (D) DRL-
EGFP (green) were continuously expressed from the
onset of neuronal differentiation using elav-GAL4.
Membrane-targeted myr-mRFP (magenta) was coex-
pressed to facilitate visualization of morphology and
quantification of localization. Right-most panels, fluo-
rescence intensity profiles along axons. (E–G), DRL-
EGFP and ROBO3-mRFP show complementary local-
ization patterns, and the positions of their boundaries
correlate with each other. (E) DRL-EGFP (green),
ROBO3-mRFP (magenta), and myr-EBFP2 (not
shown) were simultaneously expressed under the
control of elav-GAL4. Membrane-targeted myr-
EBFP2 was used to facilitate quantification of localiza-
tion. Right-most panel, fluorescence intensity profiles
along the axon. (F) Higher magnification of the
boundary region indicated with a rectangle in (E). (G)
Positions of the boundary of DRL-EGFP and
ROBO3-mRFP in each neuron are plotted as the
distance from soma. In about 70% of neurons the
boundaries of DRL-EGFP and ROBO3-mRFP coin-
cided within the range of 10 mm (gray region). Neurons
whose boundaries differ more than 10 mm were found
to have bright side branches juxtaposing the main
axon, a condition that our current algorithm cannot
handle well. (H–N) Expression of DRL-EGFP (I–K)
and ROBO3-EGFP (L–N) was induced after 24 hr in
culture by using a heat-shock-triggered induction
system (H). (I and L) Just before induction. (J and M)
Twenty-four hours after induction. (K and N) Intensity
profiles of EGFP fluorescence along the axon in (J)
and (M). Arrows indicate the position of the growth
cone at the onset of induction. Asterisks indicate the
growth cone 24 hr after induction. Scale bars, 10 mm,
except for (F), for which scale bar = 2 mm.
that show distal localization of ROBO3
decreased by 7%, 10%, and 20% at 29�C,
31�C, and 33�C, respectively (Figures 3C,
3D, and 3F, and Table S1 available online).
At restrictive temperatures some neurons
categorized as ‘‘uniform distribution’’
showed bright punctate signals of ROBO3-
EGFP along the entire length of axons
(Figure 3E). These punctate signals seen by
EGFP fluorescence were not labeled by
surface labeling using anti-ROBO3 antibody,
suggesting that they are likely to be vesicles
or aggregates that are inaccessible from the surface (Figure 3E).
About half of the neurons that were classified as uniform distribu-
tion by the quantitation of EGFP fluorescence showed distal
localization under the surface labeling condition, and thus the
severe reduction of Dynamin function at 33�C causes only
10% reduction of the distal ROBO3 pattern generated on the
cell surface. These results suggest that the formation of distal
Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc. 191

Neuron
Intra-axonal Patterning in Drosophila Neurons
localization of ROBO3 is largely independent of Dynamin
function.
We next addressed the possibility of selective targeting of DRL
to the membrane. The dependence of DRL on shi function for
endocytosis enabled us to design an experiment to distinguish
between directed and uniform targeting. We reasoned that if
DRL is preferentially targeted to the proximal segment, proteins
newly appearing on the cell surface should exhibit proximal
localization even when endocytosis is blocked. To test this, the
distribution of DRL-EGFP in shits1 mutant was examined at
2 hr, 4 hr, and 6 hr after the temperature was shifted to a restric-
tive condition (29�C). To assay only newly synthesized proteins
appearing on the cell surface, the expression of DRL-EGFP
was initiated by the GeneSwitch system simultaneously with
shift up to the restrictive temperature, and DRL-EGFP on the
axonal surface was detected by surface labeling. At 2 hr from
DRL-EGFP induction, a large fraction (64%) of neurons exhibited
proximal localization pattern, even in shits1 mutant (Figure 3G).
This suggests that newly synthesized molecules appear prefer-
entially on the proximal axonal membrane. The severe effect of
compromised Dynamin function on DRL localization manifested
only after longer chase periods; at 4 hr from DRL-EGFP induc-
tion, shits1 cells showed a significant reduction in the number
of proximal localizations, with further reduction at 6 hr (Fig-
ure 3G). This likely reflects a specific targeting property of
DRL, because similar experiments using ROBO3 revealed that
ROBO3-EGFP was distally localized at all time points (Fig-
ure 3G). These results suggest that there is a preferential target-
ing of DRL to the proximal axon compartment, although this
does not exclude the possibility that DRL is also selectively
retrieved from the distal axon compartment.
Mobility of Membrane Molecules Is Restrictedat the Compartment Boundary by a Diffusion BarrierMechanismWhile compartment-specific retrieval and targeting pathways
likely contribute to the establishment and/or maintenance of
the compartmentalized distribution of receptors, an additional
effective mechanism for pattern maintenance over time is the
mobility restriction of proteins on the fluid plasma membrane,
either within the compartment or at the compartment bound-
Table 2. Localization Patterns of EGFP-Tagged Receptors in
Cultured Drosophila Neurons
Localization Patterns (%)
Molecule
Neurons
Scored Uniform Distal Proximal
Position of
Boundary (mm)
ROBO-EGFP 108 67.6 4.6 27.8 25.8 ± 11.2
ROBO2-EGFP 98 2.0 98.0 0 23.8 ± 9.8
ROBO3-EGFP 120 2.5 97.5 0 22.4 ± 8.7
DRL-EGFP 99 16.2 0 83.8 31.8 ± 14.8
Drosophila embryonic neurons 24–36 hr in culture expressing EGFP-
tagged receptors and myr-mRFP under the control of a panneuronal
driver elav-GAL4 were imaged live, and fluorescence signals along axons
were examined for their distribution patterns. Position of boundary is indi-
cated as the distance from soma (average ± SD). See Experimental
Procedures for details of quantification.
192 Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc.
aries. Compartmentalized membrane proteins may become
immobile by being anchored to the cytoskeleton (Garrido et al.,
2003; Pan et al., 2006), or their mobility could be restricted at
certain locations on the membrane by a barrier mechanism
(Kobayashi et al., 1992; Nakada et al., 2003; Takizawa et al.,
2000; Winckler et al., 1999). To test whether or not localized
molecules are mobile, we measured the mobility of ROBO3-
EGFP and DRL-EGFP, which are localized to distal and proximal
axon segments, respectively, using fluorescence recovery after
photobleaching (FRAP) (Lippincott-Schwartz et al., 1999).
When ROBO3-EGFP fluorescence was bleached in a part of
the distal axon segment, the fluorescence recovered to the
bleached area from both ends (Figures 4A–4D). Similar results
were obtained with DRL-EGFP in proximal axon segments
(Figures 4E–4H). These results suggest that these molecules
are laterally mobile within an axonal compartment.
We next asked whether or not a barrier exists that could retain
membrane molecules within one axon compartment. To this
end, we examined whether CD8-GFP, a model transmembrane
protein that labels the entire plasma membrane of Drosophila
neurons (Lee and Luo, 1999), can move from one axon segment
to another. CD8-GFP and ROBO3-mRFP were coexpressed in
neurons in culture, and photobleaching of CD8-GFP was per-
formed in regions neighboring the boundary of ROBO3-mRFP
(Figure 4I–4K). The recovery at the side abutting the boundary
was significantly slower than that at the side away from the
boundary, indicating that the movement of CD8-GFP across
the compartment boundary is restricted (Figure 4K–4M). Similar
mobility restriction was observed with ROBO-EGFP and a GPI-
linked GFP (GFPgpi, Greco et al., 2001), but not with GAP-
GFP, which is targeted to the inner membrane (Figure 4N–4P).
These data suggest that the exchange of transmembrane
proteins between axon segments is generally limited at the
boundary region.
To gain further insight into the nature of this constraint, we per-
formed spot-size (�1 mm2) FRAP experiments on CD8-GFP
along the axon (Figures 5A and 5B). In contrast to the quick
recovery at locations away from the boundary of ROBO3-
mRFP, the region in proximity of the boundary exhibited dimin-
ished levels of recovery, with both a larger immobile fraction
and a longer half-recovery time (Figures 5C and 5D). Such
regions of reduced mobility were confined to within 10 mm of
axon length around the boundary (Figures 5E and 5F). These
data suggest that a diffusion barrier, which is thought to hinder
the mobility of molecules through the anchoring and friction-
like effects of membrane proteins (Nakada et al., 2003), exists
at the boundary region of intra-axonal compartments. This
barrier may contribute to the maintenance of intra-axonal
patterns of guidance receptors by limiting the exchange of
membrane proteins between distinct axon segments, and may
also serve as a positional cue for the specification of intra-axonal
compartments.
Presynaptic Proteins Accumulate to the Distal AxonCompartmentThe formation of the functional neuronal circuit relies on correct
axonal wiring and the subsequent establishment of synaptic
connections. In the Drosophila ventral nerve cord, synaptic
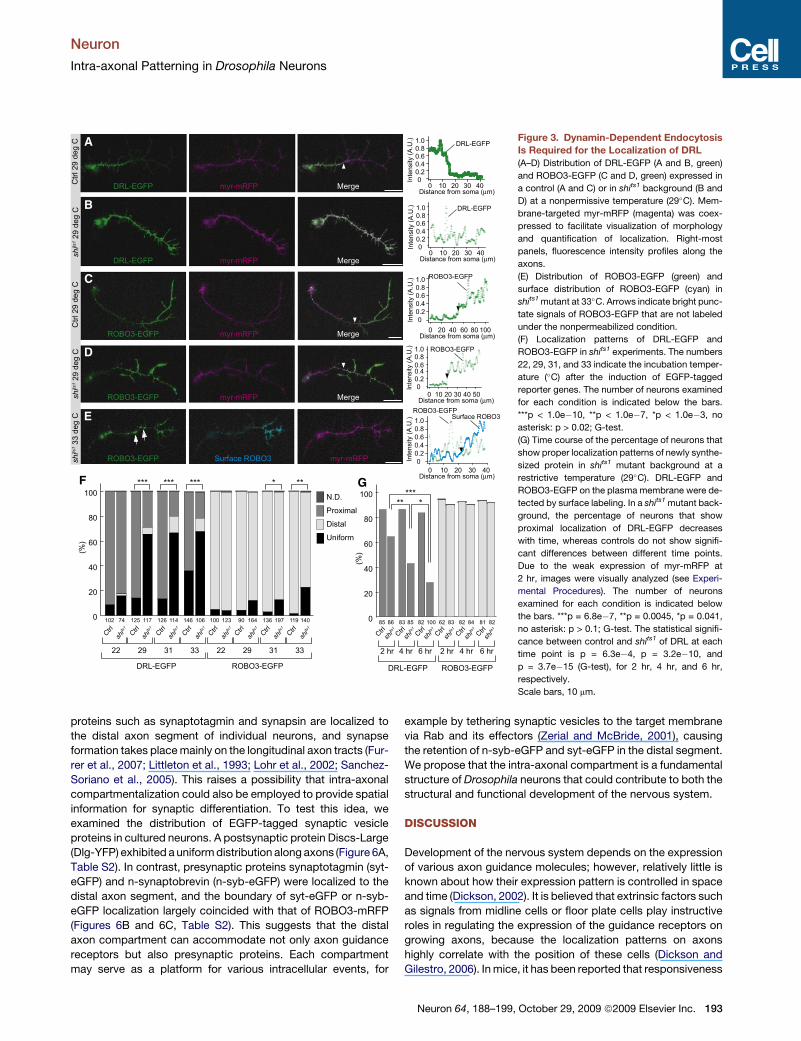
Neuron
Intra-axonal Patterning in Drosophila Neurons
A
B
C
D
E
F G
Figure 3. Dynamin-Dependent Endocytosis
Is Required for the Localization of DRL
(A–D) Distribution of DRL-EGFP (A and B, green)
and ROBO3-EGFP (C and D, green) expressed in
a control (A and C) or in shits1 background (B and
D) at a nonpermissive temperature (29�C). Mem-
brane-targeted myr-mRFP (magenta) was coex-
pressed to facilitate visualization of morphology
and quantification of localization. Right-most
panels, fluorescence intensity profiles along the
axons.
(E) Distribution of ROBO3-EGFP (green) and
surface distribution of ROBO3-EGFP (cyan) in
shits1 mutant at 33�C. Arrows indicate bright punc-
tate signals of ROBO3-EGFP that are not labeled
under the nonpermeabilized condition.
(F) Localization patterns of DRL-EGFP and
ROBO3-EGFP in shits1 experiments. The numbers
22, 29, 31, and 33 indicate the incubation temper-
ature (�C) after the induction of EGFP-tagged
reporter genes. The number of neurons examined
for each condition is indicated below the bars.
***p < 1.0e�10, **p < 1.0e�7, *p < 1.0e�3, no
asterisk: p > 0.02; G-test.
(G) Time course of the percentage of neurons that
show proper localization patterns of newly synthe-
sized protein in shits1 mutant background at a
restrictive temperature (29�C). DRL-EGFP and
ROBO3-EGFP on the plasma membrane were de-
tected by surface labeling. In a shits1 mutant back-
ground, the percentage of neurons that show
proximal localization of DRL-EGFP decreases
with time, whereas controls do not show signifi-
cant differences between different time points.
Due to the weak expression of myr-mRFP at
2 hr, images were visually analyzed (see Experi-
mental Procedures). The number of neurons
examined for each condition is indicated below
the bars. ***p = 6.8e�7, **p = 0.0045, *p = 0.041,
no asterisk: p > 0.1; G-test. The statistical signifi-
cance between control and shits1 of DRL at each
time point is p = 6.3e�4, p = 3.2e�10, and
p = 3.7e�15 (G-test), for 2 hr, 4 hr, and 6 hr,
respectively.
Scale bars, 10 mm.
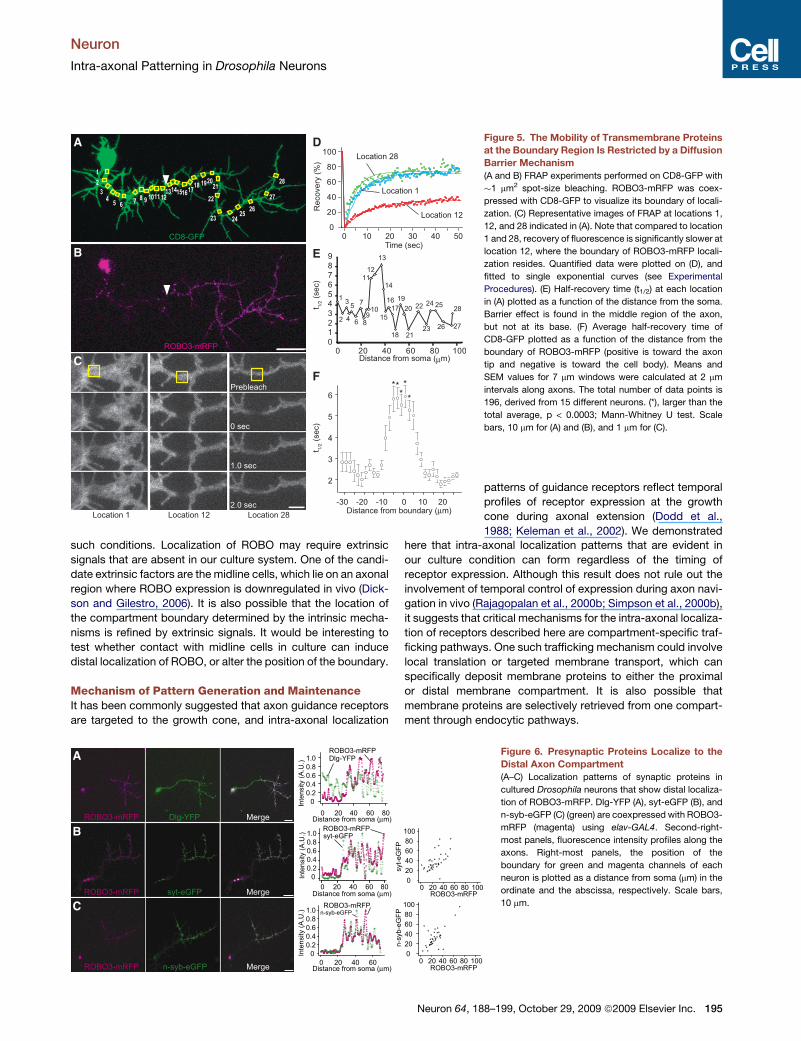
proteins such as synaptotagmin and synapsin are localized to
the distal axon segment of individual neurons, and synapse
formation takes place mainly on the longitudinal axon tracts (Fur-
rer et al., 2007; Littleton et al., 1993; Lohr et al., 2002; Sanchez-
Soriano et al., 2005). This raises a possibility that intra-axonal
compartmentalization could also be employed to provide spatial
information for synaptic differentiation. To test this idea, we
examined the distribution of EGFP-tagged synaptic vesicle
proteins in cultured neurons. A postsynaptic protein Discs-Large
(Dlg-YFP) exhibited a uniform distribution along axons (Figure 6A,
Table S2). In contrast, presynaptic proteins synaptotagmin (syt-
eGFP) and n-synaptobrevin (n-syb-eGFP) were localized to the
distal axon segment, and the boundary of syt-eGFP or n-syb-
eGFP localization largely coincided with that of ROBO3-mRFP
(Figures 6B and 6C, Table S2). This suggests that the distal
axon compartment can accommodate not only axon guidance
receptors but also presynaptic proteins. Each compartment
may serve as a platform for various intracellular events, for
example by tethering synaptic vesicles to the target membrane
via Rab and its effectors (Zerial and McBride, 2001), causing
the retention of n-syb-eGFP and syt-eGFP in the distal segment.
We propose that the intra-axonal compartment is a fundamental
structure of Drosophila neurons that could contribute to both the
structural and functional development of the nervous system.
DISCUSSION
Development of the nervous system depends on the expression
of various axon guidance molecules; however, relatively little is
known about how their expression pattern is controlled in space
and time (Dickson, 2002). It is believed that extrinsic factors such
as signals from midline cells or floor plate cells play instructive
roles in regulating the expression of the guidance receptors on
growing axons, because the localization patterns on axons
highly correlate with the position of these cells (Dickson and
Gilestro, 2006). In mice, it has been reported that responsiveness
Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc. 193
Neuron
Intra-axonal Patterning in Drosophila Neurons
of axons to guidance cues changes in the presence of the floor
plate (Shirasaki et al., 1998), and a ligand expressed at the floor
plate is required for the restricted expression of guidance recep-
tors (Long et al., 2004). On the other hand, not much work has
been devoted to unveiling the roles of cell-intrinsic programs in
regulating the intra-axonal localization of guidance receptors.
Cell-Intrinsic and -Extrinsic Pattern Generationwithin the AxonWe have described here a patterning phenomenon that takes
place within single axonal processes as a cell-intrinsic event.
This patterning involves compartmentalization of the axonal
membrane with a diffusion barrier located at a medial point of
the axon. Our data suggest that this patterning ability is a funda-
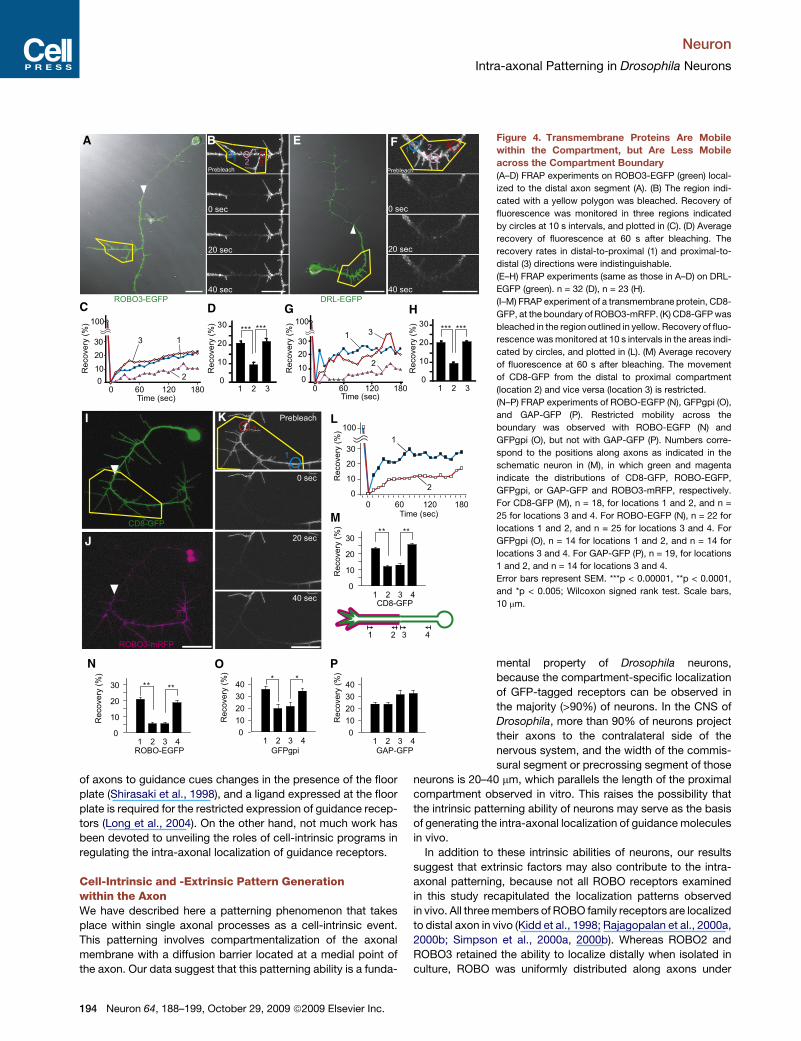
A
C D G H
LKI
J
M
PON
B E F Figure 4. Transmembrane Proteins Are Mobile
within the Compartment, but Are Less Mobile
across the Compartment Boundary
(A–D) FRAP experiments on ROBO3-EGFP (green) local-
ized to the distal axon segment (A). (B) The region indi-
cated with a yellow polygon was bleached. Recovery of
fluorescence was monitored in three regions indicated
by circles at 10 s intervals, and plotted in (C). (D) Average
recovery of fluorescence at 60 s after bleaching. The
recovery rates in distal-to-proximal (1) and proximal-to-
distal (3) directions were indistinguishable.
(E–H) FRAP experiments (same as those in A–D) on DRL-
EGFP (green). n = 32 (D), n = 23 (H).
(I–M) FRAP experiment of a transmembrane protein, CD8-
GFP, at the boundary of ROBO3-mRFP. (K) CD8-GFP was
bleached in the region outlined in yellow. Recovery of fluo-
rescence was monitored at 10 s intervals in the areas indi-
cated by circles, and plotted in (L). (M) Average recovery
of fluorescence at 60 s after bleaching. The movement
of CD8-GFP from the distal to proximal compartment
(location 2) and vice versa (location 3) is restricted.
(N–P) FRAP experiments of ROBO-EGFP (N), GFPgpi (O),
and GAP-GFP (P). Restricted mobility across the
boundary was observed with ROBO-EGFP (N) and
GFPgpi (O), but not with GAP-GFP (P). Numbers corre-
spond to the positions along axons as indicated in the
schematic neuron in (M), in which green and magenta
indicate the distributions of CD8-GFP, ROBO-EGFP,
GFPgpi, or GAP-GFP and ROBO3-mRFP, respectively.
For CD8-GFP (M), n = 18, for locations 1 and 2, and n =
25 for locations 3 and 4. For ROBO-EGFP (N), n = 22 for
locations 1 and 2, and n = 25 for locations 3 and 4. For
GFPgpi (O), n = 14 for locations 1 and 2, and n = 14 for
locations 3 and 4. For GAP-GFP (P), n = 19, for locations
1 and 2, and n = 14 for locations 3 and 4.
Error bars represent SEM. ***p < 0.00001, **p < 0.0001,
and *p < 0.005; Wilcoxon signed rank test. Scale bars,
10 mm.
mental property of Drosophila neurons,
because the compartment-specific localization
of GFP-tagged receptors can be observed in
the majority (>90%) of neurons. In the CNS of
Drosophila, more than 90% of neurons project
their axons to the contralateral side of the
nervous system, and the width of the commis-
sural segment or precrossing segment of those
neurons is 20–40 mm, which parallels the length of the proximal
compartment observed in vitro. This raises the possibility that
the intrinsic patterning ability of neurons may serve as the basis
of generating the intra-axonal localization of guidance molecules
in vivo.
In addition to these intrinsic abilities of neurons, our results
suggest that extrinsic factors may also contribute to the intra-
axonal patterning, because not all ROBO receptors examined
in this study recapitulated the localization patterns observed
in vivo. All three members of ROBO family receptors are localized
to distal axon in vivo (Kidd et al., 1998; Rajagopalan et al., 2000a,
2000b; Simpson et al., 2000a, 2000b). Whereas ROBO2 and
ROBO3 retained the ability to localize distally when isolated in
culture, ROBO was uniformly distributed along axons under
194 Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc.
Neuron
Intra-axonal Patterning in Drosophila Neurons
such conditions. Localization of ROBO may require extrinsic
signals that are absent in our culture system. One of the candi-
date extrinsic factors are the midline cells, which lie on an axonal
region where ROBO expression is downregulated in vivo (Dick-
son and Gilestro, 2006). It is also possible that the location of
the compartment boundary determined by the intrinsic mecha-
nisms is refined by extrinsic signals. It would be interesting to
test whether contact with midline cells in culture can induce
distal localization of ROBO, or alter the position of the boundary.
Mechanism of Pattern Generation and MaintenanceIt has been commonly suggested that axon guidance receptors
are targeted to the growth cone, and intra-axonal localization
A D
E
F
B
C
Figure 5. The Mobility of Transmembrane Proteins
at the Boundary Region Is Restricted by a Diffusion
Barrier Mechanism
(A and B) FRAP experiments performed on CD8-GFP with
�1 mm2 spot-size bleaching. ROBO3-mRFP was coex-
pressed with CD8-GFP to visualize its boundary of locali-
zation. (C) Representative images of FRAP at locations 1,
12, and 28 indicated in (A). Note that compared to location
1 and 28, recovery of fluorescence is significantly slower at
location 12, where the boundary of ROBO3-mRFP locali-
zation resides. Quantified data were plotted on (D), and
fitted to single exponential curves (see Experimental
Procedures). (E) Half-recovery time (t1/2) at each location
in (A) plotted as a function of the distance from the soma.
Barrier effect is found in the middle region of the axon,
but not at its base. (F) Average half-recovery time of
CD8-GFP plotted as a function of the distance from the
boundary of ROBO3-mRFP (positive is toward the axon
tip and negative is toward the cell body). Means and
SEM values for 7 mm windows were calculated at 2 mm
intervals along axons. The total number of data points is
196, derived from 15 different neurons. (*), larger than the
total average, p < 0.0003; Mann-Whitney U test. Scale
bars, 10 mm for (A) and (B), and 1 mm for (C).
A
B
C
Figure 6. Presynaptic Proteins Localize to the
Distal Axon Compartment
(A–C) Localization patterns of synaptic proteins in
cultured Drosophila neurons that show distal localiza-
tion of ROBO3-mRFP. Dlg-YFP (A), syt-eGFP (B), and
n-syb-eGFP (C) (green) are coexpressed with ROBO3-
mRFP (magenta) using elav-GAL4. Second-right-
most panels, fluorescence intensity profiles along the
axons. Right-most panels, the position of the
boundary for green and magenta channels of each
neuron is plotted as a distance from soma (mm) in the
ordinate and the abscissa, respectively. Scale bars,
10 mm.
patterns of guidance receptors reflect temporal
profiles of receptor expression at the growth
cone during axonal extension (Dodd et al.,
1988; Keleman et al., 2002). We demonstrated
here that intra-axonal localization patterns that are evident in
our culture condition can form regardless of the timing of
receptor expression. Although this result does not rule out the
involvement of temporal control of expression during axon navi-
gation in vivo (Rajagopalan et al., 2000b; Simpson et al., 2000b),
it suggests that critical mechanisms for the intra-axonal localiza-
tion of receptors described here are compartment-specific traf-
ficking pathways. One such trafficking mechanism could involve
local translation or targeted membrane transport, which can
specifically deposit membrane proteins to either the proximal
or distal membrane compartment. It is also possible that
membrane proteins are selectively retrieved from one compart-
ment through endocytic pathways.
Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc. 195

Neuron
Intra-axonal Patterning in Drosophila Neurons
Our time course experiment in shits1 mutant backgrounds
suggests that DRL is preferentially targeted to the proximal
compartment. We have also shown that the correct intra-axonal
localization of DRL requires Dynamin-dependent endocytosis;
however, at present we cannot distinguish whether or not the
endocytosis of DRL is compartment specific. Because our
FRAP experiments on CD8-GFP suggest that the barrier
between the proximal and distal axon compartments does not
completely block the movement of membrane proteins between
the compartments, it is possible that the Dynamin-dependent
endocytosis is required to remove DRL that leaks into the distal
compartment, serving to maintain the pattern generated by tar-
geting. Alternatively, endocytosis itself may be compartment
specific, contributing to the establishment of the pattern.
In contrast to DRL, ROBO3 does not appear to require shibire
function for its localization, demonstrating the presence of differ-
ential trafficking mechanisms for DRL and ROBO3. Due to this
shi-independence of ROBO3, it is not possible to conclusively
demonstrate the presence of preferential targeting of ROBO3
by performing a time course experiment. Even if there is prefer-
ential targeting, it is likely that ROBO3 also needs to be removed
from the incorrect compartment, because ROBO3 shows a level
of lateral mobility on the axon similar to that of DRL. Since
ROBO3 localization is largely independent of Dynamin function,
such a retrieval pathway must be based on Dynamin-indepen-
dent mechanisms. While the complementary localization pat-
terns of DRL and ROBO3 suggests that intra-axonal com-
partments are fundamental units for localization of multiple
molecules, molecular mechanisms for generating or maintaining
their compartmental localization could be diverse.
Another critical issue raised in the previous studies in vivo is
how the intra-axonal localization of guidance receptors is main-
tained over time (Keleman et al., 2002). If the guidance receptors
are freely diffusible on the axonal membrane, they may spread
along the axon, leading to a uniform distribution. Our FRAP
experiments in cultured neurons revealed that localized recep-
tors (ROBO3-EGFP and DRL-EGFP) are indeed mobile within
the intra-axonal compartment. Although the mobility of these
localized receptors across the compartment boundary was not
directly measurable, the mobility of several transmembrane
proteins (ROBO-EGFP and CD8-GFP) and lipid-anchored pro-
tein (GFPgpi) that distribute along the entire axon length was
significantly restricted at the compartment boundary. This
restriction is likely due to the diffusion barrier that spans over
a 10 mm axon length around the boundary. We propose that
this barrier is a part of the mechanisms that maintain the pattern
of compartment-specific membrane proteins, as shown in
different subcellular regions such as the tight junction of epithe-
lial cells (Shin et al., 2006), the posterior ring of sperm (James
et al., 2004), the cleavage furrow of dividing yeast and mamma-
lian cells (Schmidt and Nichols, 2004; Takizawa et al., 2000), and
the initial segment of mammalian neurons (Kobayashi et al.,
1992; Nakada et al., 2003; Winckler et al., 1999). We did not
detect significant barrier effect on GAP-GFP, which resides in
the inner leaflet of the plasma membrane. We have also
observed that vesicles containing membrane proteins pass
through the barrier region (data not shown). Thus, we favor
a model in which the barrier becomes effective only after the
196 Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc.
molecules are inserted into the axonal membrane. It would be
important to test whether or not a diffusion barrier exists
in vivo, and whether or not it plays a role in the development of
the nervous system.
Potential Roles of Intra-axonal PatterningAn important but yet poorly explored question is the role of the
guidance receptors localized on axon shafts. A straightforward
explanation can be offered based on non-cell-autonomous func-
tions of guidance receptors or membrane proteins in general;
they may ‘‘label’’ axon pathways through specific adhesion
(fasciculation), or through presenting their ligands, thereby
providing instructive spatial cues for the navigation of other
axons (Goodman et al., 1984; Hiramoto et al., 2000; Lin et al.,
1994; Raper et al., 1984). For example, Fasciclin cell-adhesion
molecules have been suggested to provide pathway labels for
guiding other growth cones (Goodman et al., 1984; Lin et al.,
1994; Raper et al., 1984). Drosophila Netrin receptor Frazzled/
DCC relocates its ligand Netrin to strategic positions in the
nervous system, thereby generating guidance information for
a longitudinal pioneer neuron (Hiramoto et al., 2000). Other
studies reported that guidance receptors can also play non-
cell-autonomous roles in cell migration and synaptogenesis
(Ango et al., 2004; Kraut and Zinn, 2004). Thus, spatial patterns
of molecules on axon shafts likely have direct roles in neuronal
circuit formation.
Lastly, we propose that the compartmentalization of the
axonal membrane could be a common basis for the structure
and function of the nervous system. In the Drosophila ventral
nerve cord, formation of the longitudinal axon tracts depends
on the expression of ROBO receptors. On the other hand, longi-
tudinal axon tracts are considered as the site for synapse forma-
tion, because synaptic proteins such as synaptotagmin and syn-
apsin accumulate on the longitudinal tracts (Littleton et al., 1993;
Lohr et al., 2002). We found that in cultured neurons both ROBO
receptors and synaptic proteins localize to the distal axon
compartment. This may suggest that the spatial distribution of
guidance molecules and synaptic proteins can be collectively
governed by the compartmentalization of the axonal membrane.
Future work to identify the molecular basis of the compartmen-
talization, and to establish the link between cellular identity and
this intracellular pattern, will aid in determining how intra-axonal
patterning contributes to tissue organization.
EXPERIMENTAL PROCEDURES
Drosophila Strains
The following stocks were used: y w, shits1(Grigliatti et al., 1973), elav-
GAL4(c155) (Lin and Goodman, 1994), UAS-GAP-GFP (generated by Akira
Chiba), UAS-myr-mRFP (generated by Henry Chang), UAS-mCD8-GFP (Lee
and Luo, 1999), hs-hid Y (generated by Mark Van Doren), UAS-syt-eGFP
(Zhang et al., 2002), UAS-n-syb-eGFP (Zhang et al., 2002) (from Bloomington
Stock Center), hs-GAL4 (Halfon et al., 1997), UAS-GAL4 (Hassan et al., 2000),
UAS-Nod-lacZ (Clark et al., 1997), UAS-Kin-lacZ (Clark et al., 1997), NP5280
(Hiyashi et al., 2002) (from DGRC, Kyoto), UAS-GFPgpi (Greco et al., 2001)
(kindly provided by Suzanne Eaton), UAS-Dlg-YFP (Kohsaka et al., 2007)
(kindly provided by Akinao Nose), elav-GeneSwitch (Osterwalder et al.,
2001) (kindly provided by Thomas Osterwalder), and UAS-HA-ROBO3
(Rajagopalan et al., 2000a) (kindly provided by Barry Dickson). hs-hid Y was
used for virgin collection of elav-GAL4. Targeted gene expression in transgenic

Neuron
Intra-axonal Patterning in Drosophila Neurons
Drosophila was achieved using the binary GAL4/UAS system (Brand and
Perrimon, 1993). Transgenes UAS-ROBO-EGFP, UAS-ROBO2-EGFP, UAS-
ROBO3-EGFP, UAS-ROBO3-mRFP, UAS-DRL-EGFP, and UAS-myr-EBFP2
were created using standard PCR-based methods from cDNA clones of
ROBO, ROBO2, and ROBO3 (Simpson et al., 2000a) (kindly provided by Julie
Simpson); DRL (Callahan et al., 1995) (kindly provided by Shingo Yoshikawa);
and EBFP2 (Ai et al., 2007) (Addgene). EGFP was fused to the C termini of
receptors, removing their 30-UTRs. Myristoylation signal for UAS-myr-EBFP2
was cloned from the UAS-myr-mRFP transgenic line.
Drosophila Primary Cell Cultures
Primary cell cultures of Drosophila embryonic neurons were prepared as
described previously (Patel et al., 1987), with slight modifications. Postmitotic
neurons before axonogenesis were obtained by homogenizing embryos at
stages 9–11. The cells obtained were suspended in Schneider’s Drosophila
medium (GIBCO) supplemented with 10% FBS, and cultured on glass bottom
dishes (MatTek) coated with 1 mg/ml poly-DL-ornithine (Sigma). Cell density
was adjusted to minimize cell-cell contacts. Cultures were kept at 25�C for 1
to 2 days without additional CO2. Axons and dendrites were defined by the
localization of Kin-bgal and Nod-bgal, respectively (Clark et al., 1997). Kin-
bgal accumulates to the tip of the longest neurite, while Nod-bgal localizes
to the cell body or to shorter neurites directly extending from the cell body
(Figure S1). The longest neurite is therefore referred to as an axon in this report.
A temporal shift in induction was achieved through heat-shock-triggered
induction, using a combination of hs-GAL4 (Halfon et al., 1997) and UAS-
GAL4 (Hassan et al., 2000) strains. A brief heat-shock induction of hs-GAL4
at 37�C for 30 min maintains GAL4 expression through the UAS-GAL4 autor-
egulatory loop. Neurons were imaged just before induction; the same neurons
were identified 24 hr after induction with the help of a grid-coverslip (MatTek),
and their morphology and fluorescence signals were compared.
Temperature shift experiments for the shits1 mutant were performed as
follows. Cells were cultured at 22�C for 24 hr before the temperature was
shifted to restrictive conditions. The expression of receptors under the control
of elav-GeneSwitch was initiated by adding 20 nM RU486 (Sigma) to the
culture medium immediately before shifting the temperature to the permissive
or nonpermissive conditions. Temperature shift was achieved by keeping
glass bottom culture dishes on a water bath placed in an incubator set at
a desired temperature. Fixation of cells was also done on the water bath in
the incubators after 6 hr (for 29�C, 31�C, and 33�C conditions) or 12 hr (for
22�C condition) from the temperature shift.
For the pulse chase experiments described in Figure 3G, fixation was done
after 2 hr, 4 hr, and 6 hr from the temperature shift (shifted to 29�C). Detection
of newly synthesized proteins was done by surface labeling (see Immunostain-
ing section below). We have confirmed that the labeling and detection condi-
tion we used for this experiment does not produce any signal in the absence of
RU486. Reversibility of the effects of temperature up-shift on DRL-EGFP local-
ization was verified as follows. After 24 hr of culture at 22�C, cells were incu-
bated for 6 hr at a restrictive temperature (29�C), and then transferred to
22�C for 5 hr before fixation.
Immunostaining
The following primary antibodies were used: rabbit anti-ROBO2 (Simpson
et al., 2000a) (1:1000 dilution), rat anti-DRL (Callahan et al., 1995) (1:1000 dilu-
tion), anti-HRP (Jackson Laboratories), rat anti-GFP (nacalai tesque), mouse
anti-HA (nacalai tesque), and rat anti-bgal. We also used the following anti-
bodies obtained from the Developmental Studies Hybridoma Bank, developed
under the auspices of the NICHD and maintained by The University of Iowa,
Department of Biological Sciences, Iowa City, IA 52242: anti-ROBO (Kidd
et al., 1998) (mAb 13C9) and anti-ROBO3 (Simpson et al., 2000a) (mAb
14C9 and 15H2), BP102 anti-CNS axons [1:10–1:20, developed by Corey
S. Goodman], and anti-tubulin (mAb 12G10) [1:20, developed by Joseph
Frankel and E. Marlo Nelsen]. Immunostaining of whole embryos was per-
formed as described previously (Patel, 1994). For staining cultured cells, cells
were fixed with 4% paraformaldehyde/PBS for 20 min at room temperature
and then permeabilized with 0.1% Tween-20/PBS. For surface labeling exper-
iments, cells were fixed for 10 min with 4% paraformaldehyde/PBS, and PBS
was used for all the following steps. Blocking was carried out using Block Ace
(Dainihon Pharmaceutical) for 30 min. Cells were incubated overnight with
primary antibodies in 10% Block Ace, 0.1% Tween-20/PBS at 4�C and then
stained with secondary antibodies for 1 hr at room temperature. The following
secondary antibodies were used: Alexa 405 conjugated goat anti-mouse IgG;
Alexa 488 conjugated goat anti-rabbit IgG (Invitrogen, used at 1:1000 dilution);
and DyLight488-, Cy2-, Cy3-, or Cy5-conjugated donkey anti-mouse, anti-
rabbit, or anti-rat IgG (Jackson Laboratories, used at 1:1000 dilution). Zenon
Alexa Labeling Kit (Invitrogen) was used for the double staining with mAb
BP102 and mAb 14C9. All double staining experiments were done by sequen-
tial labeling with two antibodies.
Quantitative Analysis of Localization
To avoid subjective differences between observers and to improve reproduc-
ibility between analyses, localization patterns and the positions of boundaries
were determined by quantitative criteria, except for Figure 3G (see below). All
the images were collected at an 8-bit depth, and axonal trajectories were
traced by using the ImageJ plug-in NeuronJ (Imagescience). Either fluores-
cence signals of membrane-targeted fluorescent protein (GAP-GFP or myr-
mRFP) or color-inverted bright field images (phase contrast or DIC) were
used for tracing. Pixel intensity along the trace was analyzed by the following
algorithm, using a custom program written in R software (The R Foundation).
Images were processed with a 5 3 5 median filter, and background subtrac-
tion was applied. If the maximum pixel intensity along the trace was less
than 20% of the maximum value of the dynamic range, the neuron was sorted
as ‘‘not detectable’’ (N.D.). To obtain intensity per membrane area, pixel inten-
sity of the molecule was divided by the amount of a membrane marker, GAP-
GFP or myr-mRFP, where possible. Intensity was normalized so that 1 and
0 represented the maximum and minimum intensity, respectively. Localization
was determined by a statistical significance test where the average normalized
intensities for the proximal and distal halves of an axon were compared. If a p
value was larger than 1.0e�13 (Wilcoxon signed rank test), the neuron was
classified as having a uniform distribution of markers. The position of a
boundary was defined as the point where normalized intensity becomes
continuously higher than 0.2 over a 4 mm length. When there was no point
matching the above criteria of a boundary, the neuron was again classified
as having a uniform distribution of markers. Thus ‘‘uniform’’ distribution here
should be considered as having no clear localization with a boundary.
Table 1 and Table 2 list a classification of neurons using the algorithm
described above. The percentages of neurons judged to have localized pattern
(e.g., distal for ROBO2 and ROBO3, proximal for DRL) differ between the
endogenous proteins (Table 1) and EGFP-tagged transgenes (Table 2). There
are several factors that contribute to this difference. While the transgenes are
expressed in all neurons using elav-GAL4 driver (Lin and Goodman, 1994),
endogenous genes are driven by their own promoter and are expressed only
in a subset of neurons. Thus any potential cell-type-specific differences in
the intra-axonal patterning process could cause a difference in the frequency
of neurons with localized pattern. The classification is also influenced by the
signal intensity and the signal-to-noise ratio, which in turn depends on the
method of observation (antibody versus GFP fluorescence, the quality of anti-
body, expression levels). Thus the percentages of neurons with localized
patterns are not directly comparable, especially between endogenous
proteins (Table 1) and GFP-tagged reporters (Table 2).
In Figure 3G, the fluorescence signal of the membrane marker myr-mRFP
was barely detectable at 2 hr after temperature shift, and thus we analyzed
all the data for Figure 3G visually.
Photobleaching Experiments
All photobleaching experiments were done by expressing fluorescent reporter
genes using elav-GAL4 driver. A Zeiss LSM 510 META confocal microscope
was used for all imaging, using a 100 X 1.45 N.A. objective lens at 25�C.
Bleaching was performed at the maximum laser intensity of a 488 nm argon
laser. Image acquisition was typically performed at 0.3%–0.8% intensity.
Image analysis was performed using ImageJ software (NIH) or a custom
program written in R software (The R Foundation). Recovery was calculated
according to the following formula: Recovery (%) = 100[It� I0]/[Ipre� I0], where
I = (Ia � Ib), Ia is the average fluorescence intensity of the ROI, Ib is the average
fluorescence intensity of the background region of equal size, It is the intensity
Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc. 197

Neuron
Intra-axonal Patterning in Drosophila Neurons
at each time point, I0 is the postbleach intensity, and Ipre is prebleach intensity.
The mobile fraction and the half-recovery time, which is the time required to
reach half of the intensity of the mobile fraction, were calculated by fitting
the curves obtained by a least-squares regression against the following equa-
tion: Recovery (%) = A[1� exp(�t/t)], where A is the mobile fraction and t is the
time constant. Statistical analysis was performed using R software.
All experiments presented in this study were replicated at least two times
with independent experiments and their results were reproducible.
SUPPLEMENTAL DATA
Supplemental data for this article include two Supplemental Figures and two
Supplemental Tables and can be found at http://www.cell.com/neuron/
supplemental/S0896-6273(09)00631-X.
ACKNOWLEDGMENTS
We thank J. Simpson, S. Yoshikawa, S. Eaton, A. Nose, B. Dickson, the Bloo-
mington Drosophila Stock Center, DGRC Kyoto, and DSHB for providing
materials. We thank N. Patel for advice on culture experiments; K. Fukatsu
for advice on FRAP experiments; T. Takano-Shimizu for advice on statistical
analyses; T. Yoshimori for advice on shi experiments; T. Hirata and T. De Falco
for comments on the manuscript; members of the Suzuki, Emoto, Hirose, and
Hiromi laboratories for discussions; and M. Aono, C. Asaka, E. Suzuki, Y. Ike-
tani, H. Hatanaka, and Y. Noguchi for technical assistance. This work was sup-
ported by the grants from the JSPS (T.K.), the PREST program of JST (M.H.),
the CREST program of JST, and the Takeda Science Foundation (Y.H.). The
authors declare no competing financial interests.
Accepted: August 20, 2009
Published: October 28, 2009
REFERENCES
Ai, H.W., Shaner, N.C., Cheng, Z., Tsien, R.Y., and Campbell, R.E. (2007).
Exploration of new chromophore structures leads to the identification of
improved blue fluorescent proteins. Biochemistry 46, 5904–5910.
Ango, F., di Cristo, G., Higashiyama, H., Bennett, V., Wu, P., and Huang, Z.J.
(2004). Ankyrin-based subcellular gradient of neurofascin, an immunoglobulin
family protein, directs GABAergic innervation at purkinje axon initial segment.
Cell 119, 257–272.
Bastiani, M.J., Harrelson, A.L., Snow, P.M., and Goodman, C.S. (1987).
Expression of fasciclin I and II glycoproteins on subsets of axon pathways
during neuronal development in the grasshopper. Cell 48, 745–755.
Bonkowsky, J.L., Yoshikawa, S., O’Keefe, D.D., Scully, A.L., and Thomas, J.B.
(1999). Axon routing across the midline controlled by the Drosophila Derailed
receptor. Nature 402, 540–544.
Brand, A.H., and Perrimon, N. (1993). Targeted gene expression as a means of
altering cell fates and generating dominant phenotypes. Development 118,
401–415.
Brittis, P.A., Lu, Q., and Flanagan, J.G. (2002). Axonal protein synthesis
provides a mechanism for localized regulation at an intermediate target. Cell
110, 223–235.
Callahan, C.A., Muralidhar, M.G., Lundgren, S.E., Scully, A.L., and Thomas,
J.B. (1995). Control of neuronal pathway selection by a Drosophila receptor
protein-tyrosine kinase family member. Nature 376, 171–174.
Chen, Z., Gore, B.B., Long, H., Ma, L., and Tessier-Lavigne, M. (2008). Alter-
native splicing of the Robo3 axon guidance receptor governs the midline
switch from attraction to repulsion. Neuron 58, 325–332.
Clark, I.E., Jan, L.Y., and Jan, Y.N. (1997). Reciprocal localization of Nod and
kinesin fusion proteins indicates microtubule polarity in the Drosophila oocyte,
epithelium, neuron and muscle. Development 124, 461–470.
Dickson, B.J. (2002). Molecular mechanisms of axon guidance. Science 298,
1959–1964.
198 Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc.
Dickson, B.J., and Gilestro, G.F. (2006). Regulation of commissural axon path-
finding by slit and its Robo receptors. Annu. Rev. Cell Dev. Biol. 22, 651–675.
Dodd, J., Morton, S.B., Karagogeos, D., Yamamoto, M., and Jessell, T.M.
(1988). Spatial regulation of axonal glycoprotein expression on subsets of
embryonic spinal neurons. Neuron 1, 105–116.
Furrer, M.P., Vasenkova, I., Kamiyama, D., Rosado, Y., and Chiba, A. (2007).
Slit and Robo control the development of dendrites in Drosophila CNS. Devel-
opment 134, 3795–3804.
Garrido, J.J., Giraud, P., Carlier, E., Fernandes, F., Moussif, A., Fache, M.P.,
Debanne, D., and Dargent, B. (2003). A targeting motif involved in sodium
channel clustering at the axonal initial segment. Science 300, 2091–2094.
Goodman, C.S., Bastiani, M.J., Doe, C.Q., du Lac, S., Helfand, S.L., Kuwada,
J.Y., and Thomas, J.B. (1984). Cell recognition during neuronal development.
Science 225, 1271–1279.
Greco, V., Hannus, M., and Eaton, S. (2001). Argosomes: a potential vehicle for
the spread of morphogens through epithelia. Cell 106, 633–645.
Grigliatti, T.A., Hall, L., Rosenbluth, R., and Suzuki, D.T. (1973). Temperature-
sensitive mutations in Drosophila melanogaster. XIV. A selection of immobile
adults. Mol. Gen. Genet. 120, 107–114.
Guha, A., Sriram, V., Krishnan, K.S., and Mayor, S. (2003). Shibire mutations
reveal distinct dynamin-independent and -dependent endocytic pathways in
primary cultures of Drosophila hemocytes. J. Cell Sci. 116, 3373–3386.
Halfon, M.S., Kose, H., Chiba, A., and Keshishian, H. (1997). Targeted gene
expression without a tissue-specific promoter: creating mosaic embryos using
laser-induced single-cell heat shock. Proc. Natl. Acad. Sci. USA 94, 6255–
6260.
Hassan, B.A., Bermingham, N.A., He, Y., Sun, Y., Jan, Y.N., Zoghbi, H.Y., and
Bellen, H.J. (2000). atonal regulates neurite arborization but does not act as
a proneural gene in the Drosophila brain. Neuron 25, 549–561.
Hayashi, S., Ito, K., Sado, Y., Taniguchi, M., Akimoto, A., Takeuchi, H., Aigaki,
T., Matsuzaki, F., Nakagoshi, H., Tanimura, T., et al. (2002). GETDB, a database
compiling expression patterns and molecular locations of a collection of Gal4
enhancer traps. Genesis 34, 58–61.
Hiramoto, M., Hiromi, Y., Giniger, E., and Hotta, Y. (2000). The Drosophila
Netrin receptor Frazzled guides axons by controlling Netrin distribution. Nature
406, 886–889.
James, P.S., Hennessy, C., Berge, T., and Jones, R. (2004). Compartmentali-
sation of the sperm plasma membrane: a FRAP, FLIP and SPFI analysis of
putative diffusion barriers on the sperm head. J. Cell Sci. 117, 6485–6495.
Keleman, K., Rajagopalan, S., Cleppien, D., Teis, D., Paiha, K., Huber, L.A.,
Technau, G.M., and Dickson, B.J. (2002). Comm sorts robo to control axon
guidance at the Drosophila midline. Cell 110, 415–427.
Kidd, T., Brose, K., Mitchell, K.J., Fetter, R.D., Tessier-Lavigne, M., Goodman,
C.S., and Tear, G. (1998). Roundabout controls axon crossing of the CNS
midline and defines a novel subfamily of evolutionarily conserved guidance
receptors. Cell 92, 205–215.
Kobayashi, T., Storrie, B., Simons, K., and Dotti, C.G. (1992). A functional
barrier to movement of lipids in polarized neurons. Nature 359, 647–650.
Kohsaka, H., Takasu, E., and Nose, A. (2007). In vivo induction of postsynaptic
molecular assembly by the cell adhesion molecule Fasciclin2. J. Cell Biol. 179,
1289–1300.
Kosaka, T., and Ikeda, K. (1983a). Possible temperature-dependent blockage
of synaptic vesicle recycling induced by a single gene mutation in Drosophila.
J. Neurobiol. 14, 207–225.
Kosaka, T., and Ikeda, K. (1983b). Reversible blockage of membrane retrieval
and endocytosis in the garland cell of the temperature-sensitive mutant of
Drosophila melanogaster, shibirets1. J. Cell Biol. 97, 499–507.
Kraut, R., and Zinn, K. (2004). Roundabout 2 regulates migration of sensory
neurons by signaling in trans. Curr. Biol. 14, 1319–1329.
Lee, T., and Luo, L. (1999). Mosaic analysis with a repressible cell marker for
studies of gene function in neuronal morphogenesis. Neuron 22, 451–461.

Neuron
Intra-axonal Patterning in Drosophila Neurons
Lin, D.M., and Goodman, C.S. (1994). Ectopic and increased expression of
Fasciclin II alters motoneuron growth cone guidance. Neuron 13, 507–523.
Lin, D.M., Fetter, R.D., Kopczynski, C., Grenningloh, G., and Goodman, C.S.
(1994). Genetic analysis of Fasciclin II in Drosophila: defasciculation, refasci-
culation, and altered fasciculation. Neuron 13, 1055–1069.
Lippincott-Schwartz, J., Presley, J.F., Zaal, K.J., Hirschberg, K., Miller, C.D.,
and Ellenberg, J. (1999). Monitoring the dynamics and mobility of membrane
proteins tagged with green fluorescent protein. Methods Cell Biol. 58,
261–281.
Littleton, J.T., Bellen, H.J., and Perin, M.S. (1993). Expression of synaptotag-
min in Drosophila reveals transport and localization of synaptic vesicles to the
synapse. Development 118, 1077–1088.
Lohr, R., Godenschwege, T., Buchner, E., and Prokop, A. (2002). Compart-
mentalization of central neurons in Drosophila: a new strategy of mosaic anal-
ysis reveals localization of presynaptic sites to specific segments of neurites.
J. Neurosci. 22, 10357–10367.
Long, H., Sabatier, C., Ma, L., Plump, A., Yuan, W., Ornitz, D.M., Tamada, A.,
Murakami, F., Goodman, C.S., and Tessier-Lavigne, M. (2004). Conserved
roles for Slit and Robo proteins in midline commissural axon guidance. Neuron
42, 213–223.
Nakada, C., Ritchie, K., Oba, Y., Nakamura, M., Hotta, Y., Iino, R., Kasai, R.S.,
Yamaguchi, K., Fujiwara, T., and Kusumi, A. (2003). Accumulation of anchored
proteins forms membrane diffusion barriers during neuronal polarization. Nat.
Cell Biol. 5, 626–632.
Osterwalder, T., Yoon, K.S., White, B.H., and Keshishian, H. (2001). A condi-
tional tissue-specific transgene expression system using inducible GAL4.
Proc. Natl. Acad. Sci. USA 98, 12596–12601.
Pan, Z., Kao, T., Horvath, Z., Lemos, J., Sul, J.Y., Cranstoun, S.D., Bennett, V.,
Scherer, S.S., and Cooper, E.C. (2006). A common ankyrin-G-based mecha-
nism retains KCNQ and NaV channels at electrically active domains of the
axon. J. Neurosci. 26, 2599–2613.
Patel, N.H. (1994). Imaging neuronal subsets and other cell types in whole-
mount Drosophila embryos and larvae using antibody probes. Methods Cell
Biol. 44, 445–487.
Patel, N.H., Snow, P.M., and Goodman, C.S. (1987). Characterization and
cloning of fasciclin III: a glycoprotein expressed on a subset of neurons and
axon pathways in Drosophila. Cell 48, 975–988.
Rajagopalan, S., Nicolas, E., Vivancos, V., Berger, J., and Dickson, B.J.
(2000a). Crossing the midline: roles and regulation of Robo receptors. Neuron
28, 767–777.
Rajagopalan, S., Vivancos, V., Nicolas, E., and Dickson, B.J. (2000b). Select-
ing a longitudinal pathway: Robo receptors specify the lateral position of axons
in the Drosophila CNS. Cell 103, 1033–1045.
Raper, J.A., Bastiani, M.J., and Goodman, C.S. (1984). Pathfinding by
neuronal growth cones in grasshopper embryos. IV. The effects of ablating
the A and P axons upon the behavior of the G growth cone. J. Neurosci. 4,
2329–2345.
Sabatier, C., Plump, A.S., Le, M., Brose, K., Tamada, A., Murakami, F., Lee,
E.Y., and Tessier-Lavigne, M. (2004). The divergent Robo family protein
rig-1/Robo3 is a negative regulator of slit responsiveness required for midline
crossing by commissural axons. Cell 117, 157–169.
Sanchez-Soriano, N., Bottenberg, W., Fiala, A., Haessler, U., Kerassoviti, A.,
Knust, E., Lohr, R., and Prokop, A. (2005). Are dendrites in Drosophila
homologous to vertebrate dendrites? Dev. Biol. 288, 126–138.
Schmidt, K., and Nichols, B.J. (2004). A barrier to lateral diffusion in the
cleavage furrow of dividing mammalian cells. Curr. Biol. 14, 1002–1006.
Seeger, M., Tear, G., Ferres-Marco, D., and Goodman, C.S. (1993). Mutations
affecting growth cone guidance in Drosophila: genes necessary for guidance
toward or away from the midline. Neuron 10, 409–426.
Shin, K., Fogg, V.C., and Margolis, B. (2006). Tight junctions and cell polarity.
Annu. Rev. Cell Dev. Biol. 22, 207–235.
Shirasaki, R., Katsumata, R., and Murakami, F. (1998). Change in chemoat-
tractant responsiveness of developing axons at an intermediate target.
Science 279, 105–107.
Simpson, J.H., Bland, K.S., Fetter, R.D., and Goodman, C.S. (2000a). Short-
range and long-range guidance by Slit and its Robo receptors: a combinatorial
code of Robo receptors controls lateral position. Cell 103, 1019–1032.
Simpson, J.H., Kidd, T., Bland, K.S., and Goodman, C.S. (2000b). Short-range
and long-range guidance by slit and its Robo receptors. Robo and Robo2 play
distinct roles in midline guidance. Neuron 28, 753–766.
Takizawa, P.A., DeRisi, J.L., Wilhelm, J.E., and Vale, R.D. (2000). Plasma
membrane compartmentalization in yeast by messenger RNA transport and
a septin diffusion barrier. Science 290, 341–344.
Winckler, B., Forscher, P., and Mellman, I. (1999). A diffusion barrier maintains
distribution of membrane proteins in polarized neurons. Nature 397, 698–701.
Zerial, M., and McBride, H. (2001). Rab proteins as membrane organizers. Nat.
Rev. Mol. Cell Biol. 2, 107–117.
Zhang, Y.Q., Rodesch, C.K., and Broadie, K. (2002). Living synaptic vesicle
marker: synaptotagmin-GFP. Genesis 34, 142–145.
Neuron 64, 188–199, October 29, 2009 ª2009 Elsevier Inc. 199
Related Documents