Intestinal helminth communities of the red fox (Vulpes vulpes L.) in the Italian Alps Anna R. Di Cerbo 1 , Maria T. Manfredi 1* , Karin Trevisiol 2 , Marco Bregoli 3 , Nicola Ferrari 1 , Francesca Pirinesi 1 and Selvaggia Bazzoli 1 1 Department of Animal Pathology Hygiene and Public Health (DIPAV), Section of General Pathology and Parasitology, Faculty of Veterinary Medicine of Milan, University of Milan, Via Celoria 10, I-20133 Milan; 2 Istituto Zooprofilattico Sperimentale Section of Bolzano, Via Bivio 59, 39100 Bolzano; 3 Istituto Zooprofilattico Sperimentale, Section of Trento, Via Lavisotto 129, Trento; Italy Abstract The study analyses the intestinal helminth communities found in 645 red foxes of alpine populations from five areas of Northern Italy. In particular, the aim was to evaluate the contribution of both environmental variability and the structure of fox popula- tion (extrinsic and intrinsic factors, respectively) in shaping the structure and composition of the intestinal helminth commu- nities. To identify the influence of the characteristics of the host (i.e. age and sex) together with extrinsic factors (year, site, season, and altitude of collection) on number of species, total load per fox, prevalence and abundance of each parasite species, general linear models were performed. Fifteen helminth species have been detected in the intestine of 545 infected animals (84.5%) with a total of 17,144 parasites collected. The analysis of factors influencing both prevalence and abundance of infec- tion of the parasite species revealed the preponderance of extrinsic factors on intrinsic ones. In particular, geographical areas influenced prevalence and abundance of every parasite species, showing high spatial variability. The lower influence of host factors may suggest that, in this case, host dynamics play a trivial role with respect to spatial variability in determining para- site abundance. These results pointed out high prevalence of intestinal helminth infections in the Italian Alpine fox, confirmed the typical composition of parasite fauna within fox populations, underlined local differences in the structure and composition of the helminth communities. Moreover, this study highlights the major role of extrinsic factors vs intrinsic ones. Keywords Red fox, helminth community, Toxocara canis, Echinococcus multilocularis, Alps, Northern Italy * Corresponding author: [email protected] Introduction The study concerning the structure of helminth communities, the relationships among parasite species, and the intercours- es between parasites and the environment has recently been applied to ecological studies on the parasite fauna of different species among wild animals (Poulin 2001). A parasite infec- tion reflects the exposition and susceptibility of their host; thus, investigations on the composition and structure of their communities can be highly informative on the factors influ- encing the dynamics of parasite transmission. Conceptually, a parasite community can be defined as all parasite individ- uals of all species within the host population (Bush et al. 1997). Nevertheless, it can be approached at different hierar- chical levels; in fact, interactions between infrapopulations in an individual host, i.e. all members of a given parasite species within a single host, may determine how many parasite species can coexist within a single host (the infracommunity structure), and consequently in a parasite component commu- nity (Holmes and Price 1986, see Poulin 2001 for a review). These classifications demonstrate to be very helpful for inves- tigation as the effect of different factors may be identified at different levels. While the effect of host factors, i.e. sex and age, are more prone to act at infracommunity level, environ- mental factors can shape difference between component com- munities. Ecological studies also involve the evaluation of richness and species diversity within the helminth community. Several indices have been proposed to this aim (e.g., see review by Poulin 2001). For example, a very simple index to evaluate the species richness is offered by the number of the species involved. However, it cannot explain the distribution of the DOI: 10.2478/s11686-008-0042-7 © 2008 W. Stefañski Institute of Parasitology, PAS Acta Parasitologica, 2008, 53(3), 302–311; ISSN 1230-2821

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Intestinal helminth communities of the red fox(Vulpes vulpes L.) in the Italian Alps
Anna R. Di Cerbo1, Maria T. Manfredi1*, Karin Trevisiol2, Marco Bregoli3, Nicola Ferrari1, Francesca Pirinesi1 and Selvaggia Bazzoli1
1Department of Animal Pathology Hygiene and Public Health (DIPAV), Section of General Pathology and Parasitology, Faculty of Veterinary Medicine of Milan, University of Milan, Via Celoria 10, I-20133 Milan;
2Istituto Zooprofilattico Sperimentale Section of Bolzano, Via Bivio 59, 39100 Bolzano;3Istituto Zooprofilattico Sperimentale, Section of Trento, Via Lavisotto 129, Trento; Italy
AbstractThe study analyses the intestinal helminth communities found in 645 red foxes of alpine populations from five areas of NorthernItaly. In particular, the aim was to evaluate the contribution of both environmental variability and the structure of fox popula-tion (extrinsic and intrinsic factors, respectively) in shaping the structure and composition of the intestinal helminth commu-nities. To identify the influence of the characteristics of the host (i.e. age and sex) together with extrinsic factors (year, site,season, and altitude of collection) on number of species, total load per fox, prevalence and abundance of each parasite species,general linear models were performed. Fifteen helminth species have been detected in the intestine of 545 infected animals(84.5%) with a total of 17,144 parasites collected. The analysis of factors influencing both prevalence and abundance of infec-tion of the parasite species revealed the preponderance of extrinsic factors on intrinsic ones. In particular, geographical areasinfluenced prevalence and abundance of every parasite species, showing high spatial variability. The lower influence of hostfactors may suggest that, in this case, host dynamics play a trivial role with respect to spatial variability in determining para-site abundance. These results pointed out high prevalence of intestinal helminth infections in the Italian Alpine fox, confirmedthe typical composition of parasite fauna within fox populations, underlined local differences in the structure and compositionof the helminth communities. Moreover, this study highlights the major role of extrinsic factors vs intrinsic ones.
KeywordsRed fox, helminth community, Toxocara canis, Echinococcus multilocularis, Alps, Northern Italy
*Corresponding author: [email protected]
Introduction
The study concerning the structure of helminth communities,the relationships among parasite species, and the intercours-es between parasites and the environment has recently beenapplied to ecological studies on the parasite fauna of differentspecies among wild animals (Poulin 2001). A parasite infec-tion reflects the exposition and susceptibility of their host;thus, investigations on the composition and structure of theircommunities can be highly informative on the factors influ-encing the dynamics of parasite transmission. Conceptually,a parasite community can be defined as �all parasite individ-uals of all species within the host population� (Bush et al.1997). Nevertheless, it can be approached at different hierar-chical levels; in fact, interactions between infrapopulations inan individual host, i.e. all members of a given parasite species
within a single host, may determine how many parasitespecies can coexist within a single host (the infracommunitystructure), and consequently in a parasite component commu-nity (Holmes and Price 1986, see Poulin 2001 for a review).These classifications demonstrate to be very helpful for inves-tigation as the effect of different factors may be identified atdifferent levels. While the effect of host factors, i.e. sex andage, are more prone to act at infracommunity level, environ-mental factors can shape difference between component com-munities.
Ecological studies also involve the evaluation of richnessand species diversity within the helminth community. Severalindices have been proposed to this aim (e.g., see review byPoulin 2001). For example, a very simple index to evaluatethe species richness is offered by the number of the speciesinvolved. However, it cannot explain the distribution of the
DOI: 10.2478/s11686-008-0042-7© 2008 W. Stefañski Institute of Parasitology, PASActa Parasitologica, 2008, 53(3), 302–311; ISSN 1230-2821

Helminth communities of V. vulpes in Northern Italy
species within a given area, as it gives equal weight to all taxa(Oindo 2001). On the other hand, diversity indices are morereliable, as they consider both the number of species collect-ed and the parasite abundance in each area (Magurran 1988).
Many studies on the parasite ecology of wild mammalshave been reported so far (e.g., Pence and Windberg 1984,Pence 1990, Zaffaroni et al. 1996, Roberts et al. 2002, Man-fredi et al. 2003b, Matthee et al. 2004, Segovia et al. 2004,Eira et al. 2006a, b). For instance, studies on helminth com-munities of coyotes (Canis latrans) in North America indicateboth extrinsic (i.e., season) and intrinsic factors operating in-dependently or in combination on the structure of the parasitecommunity and on the distribution of certain species (e.g.,Pence and Windberg 1984). A similar approach could be usedalso to assess infracommunity and community structure of hel-minths in foxes, and to provide epidemiological comparisonsas these canids are ecological equivalents.
Vulpes vulpes (L., 1758), the red fox, is one of the mostcommon wild carnivores nearly all over Italy. This speciesshows great ecological plasticity and a variable diet partiallyincluding human food sources such as cultivated fruits or rub-bish, and even domestic animals (Cagnacci et al. 2003, Ma-rinelli et al. 2003). Their high adaptability allows red foxes tocolonize all the Alps and any environment within this area.They colonize both rural and urban areas, and small or medi-um villages located in mountain areas (Lucherini et al. 1995,Richards et al. 1995, Hofer et al. 2000, Deplazes et al. 2004).In some Alpine localities fox populations are influenced by thegrowing pressure of tourism, which implies both direct andindirect interactions between them and humans (Meriggi andRosa 1991, Guglielmi et al. 2003), and can mean hazardoussituations when foxes harbour potentially zoonotic parasites(Guberti and Poglayen 1991, Pozio 1991, Manfredi et al. 2002).
There are few reports concerning the structure of helminthcommunities of the fox (e.g., Richards et al. 1995, GortBzaret al. 1998, Segovia et al. 2004); moreover, most of those con-cerning the European fox population have been aimed at de-termining the presence of parasites that represent a risk forpublic health, in particular Echinococcus multilocularis (e.g.,Deplazes et al. 2004, Manfredi et al. 2004). In Italy there havebeen sporadic reports on the parasite populations in foxes, allof them carried out on a limited number of animals comingfrom a defined area (Soldati et al. 1976, Rossi et al. 1983,Poglayen et al. 1985, Iori et al. 1990, Stancampiano et al.1998). All these studies show low attention to the factors shap-ing the structure and composition of the parasite communities;as a result, we do not know whether parasite species occur dueto environmental differences or to differences in the structureof fox population. An important implication can derive froma different approach which can also represent a first step inmanaging zoonotic parasite infections.
The aim of the present study was to evaluate the contribu-tion of both extrinsic and intrinsic factors in shaping the struc-ture and composition of the intestinal helminth communitiesof red foxes from five areas of the Italian Alps. In particular,we investigated whether a helminth community is shaped by
environmental variability, or by a difference in the structure offox population.
Materials and methods
Sample collection
From 1998 to 2006, we examined a total of 645 foxes (285males, 211 females, and 149 specimens of undetermined gen-der) from the following regions: Trentino Alto Adige (pro-vinces of Trento, n = 181, and Bolzano, n = 299), Veneto (Bel-luno province, n = 79), Valle d�Aosta (Aosta province, n = 38),Lombardia (Bergamo, Brescia, Como, Sondrio and Varese prov-inces, n = 48) (Fig. 1). These animals were found either nat-urally dead or hunted according to national law no. 157/92 andregional regulations in areas situated between 100 and 2,200m a s.l.
Their carcasses were brought to the provincial offices ofthe Veterinary Public Health Services (Zooprophylactic Insti-tutes), where the small intestine was taken from each fox. Allsamples were sent to the Faculty of Veterinary Medicine ofMilan (DIPAV). The samples were frozen (at least 2 weeks at�80°C and then at �20°C) and were left overnight at roomtemperature before being examined to reduce any risk ofinfection for laboratory staff. Parasites were isolated using thesedimentation and counting technique (SCT) on intestinalcontents, as described by Hofer et al. (2000). The parasiteswere preserved in 70% ethanol before being clarified orstained and identified using a light microscope (Zeiss Axio-scop).
Data on individual foxes were collected (finding area, alti-tude and date, sex, age, cause of death); the samples weregrouped according to their province of origin, but the samplesfrom Lombardy, whose data were pooled due to the low num-ber of foxes examined per province.
Out of 330 foxes of which age was determined, 40 wereclassified as very young (cubs), 77 as young and 213 as adultconsidering their individual dentition and teeth wear togeth-er with their body size.
As to the finding dates (n = 528), data were subdividedinto 4 seasonal periods: winter (December-February) n = 131;spring (March-May) n = 69; summer (June-August) n = 159;autumn (September-November) n = 169. While consideringthe finding altitude (n = 433), data were grouped into fourclasses: 1, 0�500 m (n = 40); 2, >500�1,000 m (n = 143); 3,>1,000�1,500 m (n = 195) and 4, >1,500 m (n = 55).
Analytical methods
Epidemiological indices (mean abundance, mean intensity,and prevalence) were calculated for each taxon of helminth(Bush et al. 1997). The degree of parasite aggregation is ex-pressed by the k parameter estimated through maximum like-lihood function (Wilson et al. 2002). Diversity indices wereused to evaluate species richness and diversity in the helminthcommunities. For each fox group the following diversity in-dices were calculated (Magurran 1988):
303
�l¹ski

Anna R. Di Cerbo et al.
Dominance measure using Simpson�s index: D = Σ (n1 (n1 � 1)) (ni is the worm burdens of species i and
N is the total worm burden) corresponds to the proportion ofindividuals in the species i. This index calculates the proba-bility that 2 randomly selected individuals in the communitybelong to the same category and ranges from 0 and 1.0.
Diversity index of Shannon: H′ = �Σ pi log2 pi (where p =ni/N). This assumes that individuals are randomly sampled.
Evenness of the abundance of species for Shannon�s index:EH′ = H′/ln S. This parameter measures the equitability of dis-tribution for individuals of the same species and ranges from0 and 1.
Richness, using Menhinick�s richness index: R = S/√N,where S is the number of species recorded within a province,while N is the total worm burden in the same area.
Moreover, we used Thul Importance index (Thul et al.1985) to classify helminth communities hierarchically. TheImportance value (It) was calculated for each of the helminthtaxon by the formula: Itj= (Mj) Aj Bj/Σ Aj Bj
*100, where Mj= maturity factor equals 1.0 if mature specimens were foundor 0 if they were absent, Aj = number of individual parasitesin species j, Bj = number of hosts infected by parasites speciesj. This index weighting simultaneously by prevalence andabundance classify the species as either: dominant (It>1), spe-cies strongly characteristic of the community; codominant(0.01<It<1), species contributing significantly to the commu-nity, though to a lesser degree; subordinate (0<It<0.01), spe-cies occurring infrequently that may develop and reproduce
but do not contribute significantly to the community; unsuc-cessful pioneer (It = 0), species characteristic of another hostthat gain access to the host but do not mature or reproduce andcontribute little to the community.
Statistical analysis
To identify the influence of the characteristics of the host suchas sex and age, together with extrinsic factors, e.g., year, site,season, and altitude, on (i) number of species, (ii) total loadper fox, (iii) prevalence and abundance of each parasite spe-cies, general linear models (GLMs) were performed. Maxi-mal model containing up to all second order interactions wasfitted and the minimal adequate model, i.e. the model includ-ing factors and interactions that best contribute to explain datavariation, was obtained through deletion test (McCullagh andNelder 1983, Crawley 2002). According to the distribution ofresponse variable the following error structure and link func-tion were used: for the aggregate distribution of the parasitespecies, abundance was analyzed by modified negative bino-mial GLM with log-ratio link function (McCullagh and Nel-der 1983). Prevalence was analysed by classifying each foxeither positive or negative to the infection of parasites, sologistic regression with binomial error structure and logit linkfunction was used. Finally, the number of species was ana-lysed fitting a Poisson error distribution with a log link func-tion. The distribution of parasite intensity was tested if signif-icantly differing from negative binomial through nb.fit func-tion under S-PLUS 6.1.2 (developed by Darren Shaw). The
304
N (N � 1)
Fig. 1. Map of study areas with regions (marked differently) and provinces of origin of examined foxes

Helminth communities of V. vulpes in Northern Italy
parasite aggregation is expressed by the parameter k comput-ed by the same function through maximum likelihood meth-ods. The 95% confidence intervals for prevalence was calcu-lated according to the method proposed by Wilson (1927), andrevised by Newcombe (1998).
All statistical analyses were performed with S-PLUS 6.1.2software packages.
Results
Composition of the intestinal helminth fauna
Out of 645 red foxes examined, 100 showed no intestinalhelminths, while 545 animals (84.5%; 95% CI: 81.5�87.1%)resulted infected by one or more parasite species with a totalof 17,144 parasites collected (Table I). We recorded 15 speciesbelonging to Cestoda (Mesocestoides lineatus, Taenia crassi-ceps, T. hydatigena, T. taeniaeformis, Echinococcus multilo-cularis), Nematoda (Uncinaria stenocephala, Toxocara canis,Pterygodermatites affinis, Molineus legerae, Trichuris vulpis,Capillaria sp., Oxynema crassispiculum) and Trematoda (Pha-ryngostomum cordatum, Metorchis vulpis, Plagiorchis ele-gans).
At individual host level, the evaluation of infracommuni-ty structure gave the following results: 7.9% (95% CI: 6.1�10.3%) of the examined foxes harboured only cestodes while35.2% (95% CI: 31.6�38.9%) had only nematodes; 40.0%(95% CI: 36.3�43.8%) showed both cestodes and nematodes,and respectively 1.7% (95% CI: 1.0�3.0%) had cestodes and1 nematode species, 29.8% (95% CI: 26.4�33.4%) cestodesand two nematode species, 7.3% (95% CI: 5.5�9.6%) ces-todes and 3 nematode species, 1.1% (95% CI: 0.5�2.2%) ces-todes and 4 nematode species, 0.2% (95% CI: 0.03�0.9%)cestodes and 5 nematode species. Finally, 1.4% of the samples(95% CI: 0.7�2.6%) had cestodes, nematodes, and trema-todes.
Helminth community patterns in the five study areas
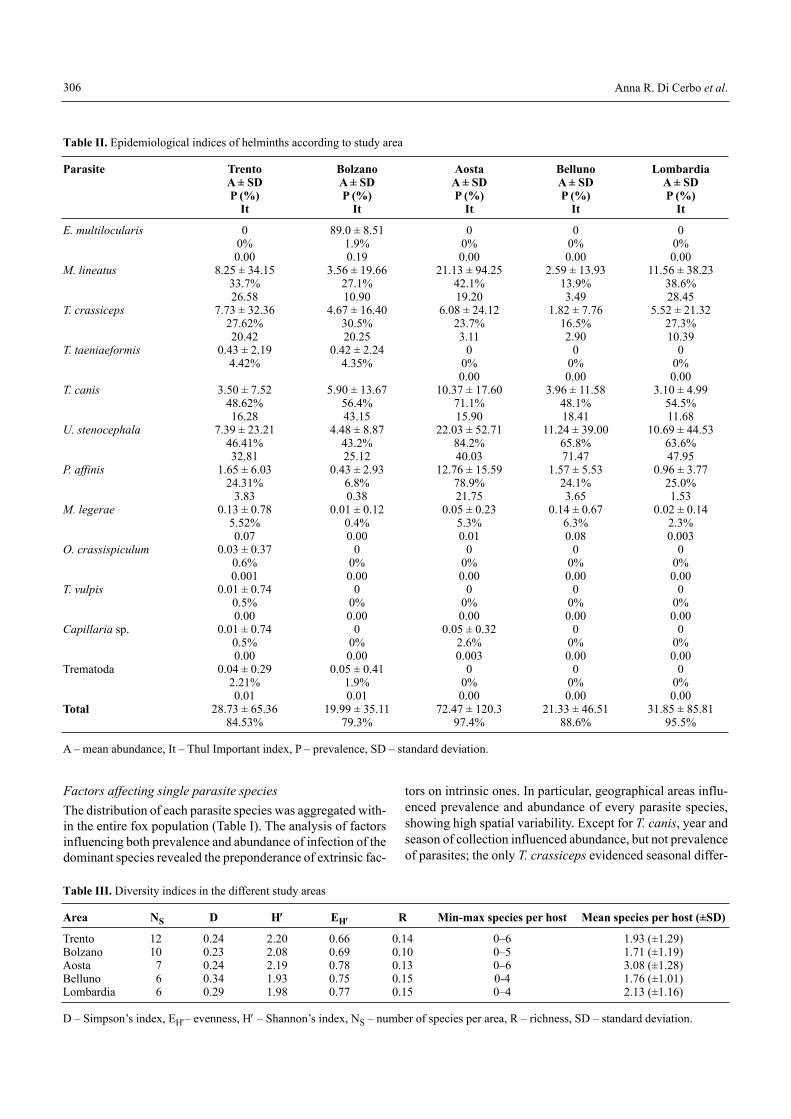
The total prevalence calculated in the different provincesranged from 79.3% (Bolzano province) to the maximum97.4% (Aosta province) and 95.5% (Lombardy region) (TableII). On the basis of hierarchical categories derived from valuesof Thul Importance index, several differences inside the par-asite communities� structure were noted. As dominant species(It>1) resulted M. lineatus, T. crassiceps, T. canis, U. steno-cephala, P. affinis (except for Bolzano where the last specieswas codominant, 0.01<It<1). Wherever E. multilocularis waspresent (Bolzano province), it gave a significant contributionto the parasite community (codominant species). The otherhelminth populations did not contribute significantly to thecommunities (subordinate species and unsuccessful pioneer).
In Table III the values calculated according to differentindices are presented, reflecting various aspects of diversity atthe infracommunity level and at the component communitylevel (parasites within the study areas).
The highest species richness was recorded in Lombardyand in the province of Belluno (both: R = 0.15), even thoughthey did not have the highest number of parasite species col-lected (6 vs 12 in Trento province); these same areas regis-tered the highest values of dominance (D = 0.29 and D = 0.34,respectively). The highest species diversity was recorded inTrento and Aosta provinces (H′ = 2.20 and H′ = 2.19, respec-tively).
At the infracommunity level, the number of parasite taxacollected per host showed significant differences among yearsand areas, with the highest values registered in Aosta region,and the lowest ones in Bolzano province. Moreover, the areasshowed a different variation between them with respect to theyears. The age was the only significant host factor (Table IV).
Similarly, area and year of sampling showed a significantinfluence even on total parasite load, with the highest valuesin Aosta region and the lowest in Bolzano; no effect by hostfactors was observed (Table V).
305
Roborzyñski rosb��æv fjad kadsææ¿æ
Table I. Population parameters of helminths in red foxes collected from five study areas in Northern Italy
Parasite Range A ± SD I ± SD P (%; 95% CI) k
E. multilocularis 0�100 0.41 ± 5.80 53.00 ± 44.07 0.78 (0.33�1.81) 0.001M. lineatus 0�581 6.39 ± 34.15 23.28 ± 62.23 27.44 (24.14�31.01) 0.068T. crassiceps 0�300 5.33 ± 22.24 18.37 ± 38.35 28.99 (25.62�32.61) 0.081T. hydatigena 0�15 0.028 ± 0.61 9.00 ± 8.48 0.32 (0.09�1.16) 0.001T. taeniaeformis 0�22 0.32 ± 1.95 9.71 ± 4.87 3.36 (2.14�5.18) 0.028T. canis 0�162 5.05 ± 11.89 9.27 ± 14.85 54.42 (50.56�58.23) 0.253U. stenocephala 0�333 7.62 ± 26.36 14.85 ± 35.33 51.32 (47.47�55.16) 0.188P. affinis 0�65 1.68 ± 6.39 8.66 ± 12.30 19.38 (16.51�22.61) 0.066M. legerae 0�9 0.06 ± 0.49 2.11 ± 2.00 2.95 (1.90�4.56) 0.023O. crassispiculum 0�5 0.01 ± 0.20 5.00 0.16 (0.03�0.88) 0.001T. vulpis 0�1 0.00 ± 0.04 1.00 0.16 (0.03�0.88) �Capillaria sp. 0�2 0.00 ± 0.09 1.50 ± 0.71 0.31 (0.09�1.12) 0.004Trematoda 0�5 0.03 ± 0.32 2.33 ± 1.41 1.40 (0.74�2.64) 0.009Total 0�615 26.58 ± 59.55 31.46 ± 63.59 84.50 (81.50�87.09) �
A � mean abundance, CI � confidence interval, I � mean intensity, k � parasite aggregation parameter, P � prevalence, SD � standard devi-ation.
Anna R. Di Cerbo et al.
Factors affecting single parasite species
The distribution of each parasite species was aggregated with-in the entire fox population (Table I). The analysis of factorsinfluencing both prevalence and abundance of infection of thedominant species revealed the preponderance of extrinsic fac-
tors on intrinsic ones. In particular, geographical areas influ-enced prevalence and abundance of every parasite species,showing high spatial variability. Except for T. canis, year andseason of collection influenced abundance, but not prevalenceof parasites; the only T. crassiceps evidenced seasonal differ-
306
Table II. Epidemiological indices of helminths according to study area
Parasite Trento Bolzano Aosta Belluno LombardiaA ± SD A ± SD A ± SD A ± SD A ± SDP (%) P (%) P (%) P (%) P (%)
It It It It It
E. multilocularis 0 89.0 ± 8.51 0 0 00% 1.9% 0% 0% 0%0.00 0.19 0.00 0.00 0.00
M. lineatus 8.25 ± 34.15 3.56 ± 19.66 21.13 ± 94.25 2.59 ± 13.93 11.56 ± 38.2333.7% 27.1% 42.1% 13.9% 38.6%26.58 10.90 19.20 3.49 28.45
T. crassiceps 7.73 ± 32.36 4.67 ± 16.40 6.08 ± 24.12 1.82 ± 7.76 5.52 ± 21.3227.62% 30.5% 23.7% 16.5% 27.3%20.42 20.25 3.11 2.90 10.39
T. taeniaeformis 0.43 ± 2.19 0.42 ± 2.24 0 0 04.42% 4.35% 0% 0% 0%
0.00 0.00 0.00T. canis 3.50 ± 7.52 5.90 ± 13.67 10.37 ± 17.60 3.96 ± 11.58 3.10 ± 4.99
48.62% 56.4% 71.1% 48.1% 54.5%16.28 43.15 15.90 18.41 11.68
U. stenocephala 7.39 ± 23.21 4.48 ± 8.87 22.03 ± 52.71 11.24 ± 39.00 10.69 ± 44.5346.41% 43.2% 84.2% 65.8% 63.6%32.81 25.12 40.03 71.47 47.95
P. affinis 1.65 ± 6.03 0.43 ± 2.93 12.76 ± 15.59 1.57 ± 5.53 0.96 ± 3.7724.31% 6.8% 78.9% 24.1% 25.0%
3.83 0.38 21.75 3.65 1.53M. legerae 0.13 ± 0.78 0.01 ± 0.12 0.05 ± 0.23 0.14 ± 0.67 0.02 ± 0.14
5.52% 0.4% 5.3% 6.3% 2.3%0.07 0.00 0.01 0.08 0.003
O. crassispiculum 0.03 ± 0.37 0 0 0 00.6% 0% 0% 0% 0%0.001 0.00 0.00 0.00 0.00
T. vulpis 0.01 ± 0.74 0 0 0 00.5% 0% 0% 0% 0%0.00 0.00 0.00 0.00 0.00
Capillaria sp. 0.01 ± 0.74 0 0.05 ± 0.32 0 00.5% 0% 2.6% 0% 0%0.00 0.00 0.003 0.00 0.00
Trematoda 0.04 ± 0.29 0.05 ± 0.41 0 0 02.21% 1.9% 0% 0% 0%0.01 0.01 0.00 0.00 0.00
Total 28.73 ± 65.36 19.99 ± 35.11 72.47 ± 120.3 21.33 ± 46.51 31.85 ± 85.8184.53% 79.3% 97.4% 88.6% 95.5%
A � mean abundance, It � Thul Important index, P � prevalence, SD � standard deviation.
Table III. Diversity indices in the different study areas
Area NS D H′ EH′′ R Min-max species per host Mean species per host (±SD)
Trento 12 0.24 2.20 0.66 0.14 0�6 1.93 (±1.29)Bolzano 10 0.23 2.08 0.69 0.10 0�5 1.71 (±1.19)Aosta 7 0.24 2.19 0.78 0.13 0�6 3.08 (±1.28)Belluno 6 0.34 1.93 0.75 0.15 0-4 1.76 (±1.01)Lombardia 6 0.29 1.98 0.77 0.15 0�4 2.13 (±1.16)
D � Simpson�s index, EH′� evenness, H′ � Shannon�s index, NS � number of species per area, R � richness, SD � standard deviation.
Helminth communities of V. vulpes in Northern Italy
ence as to prevalence. Further, abundance of all species exceptU. stenocephala was influenced by the altitude of collectionwhich linked to a lower degree of urbanization.
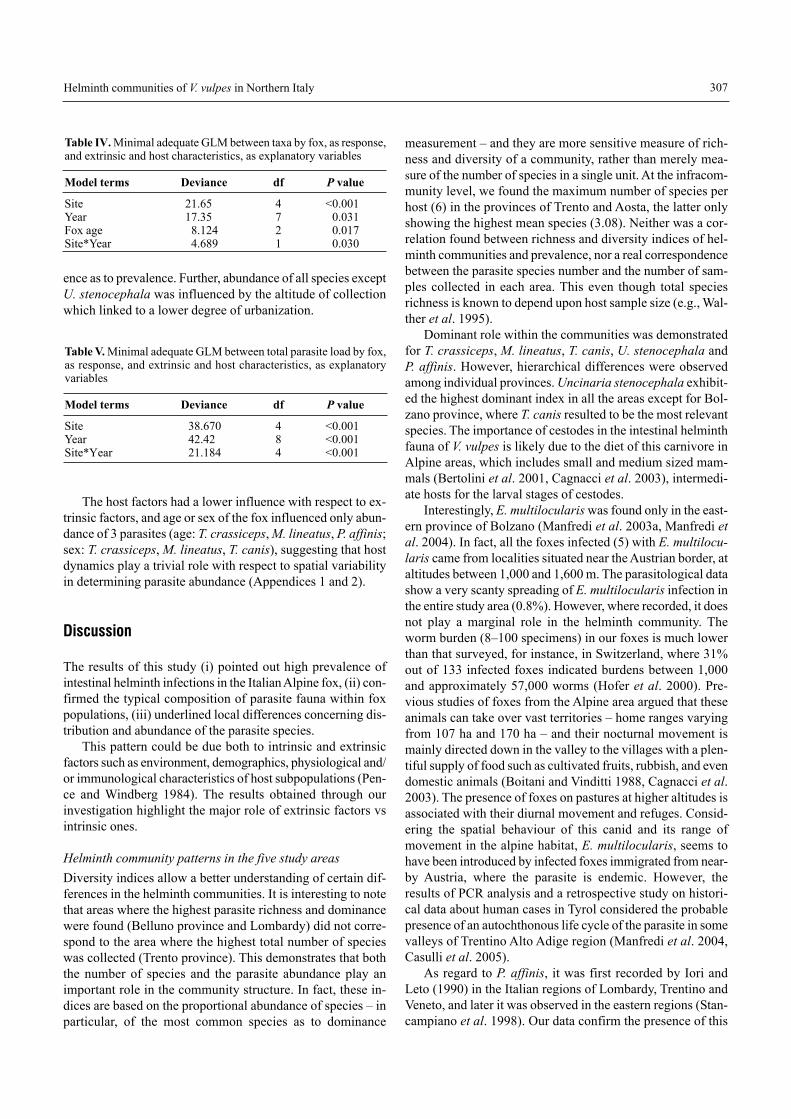
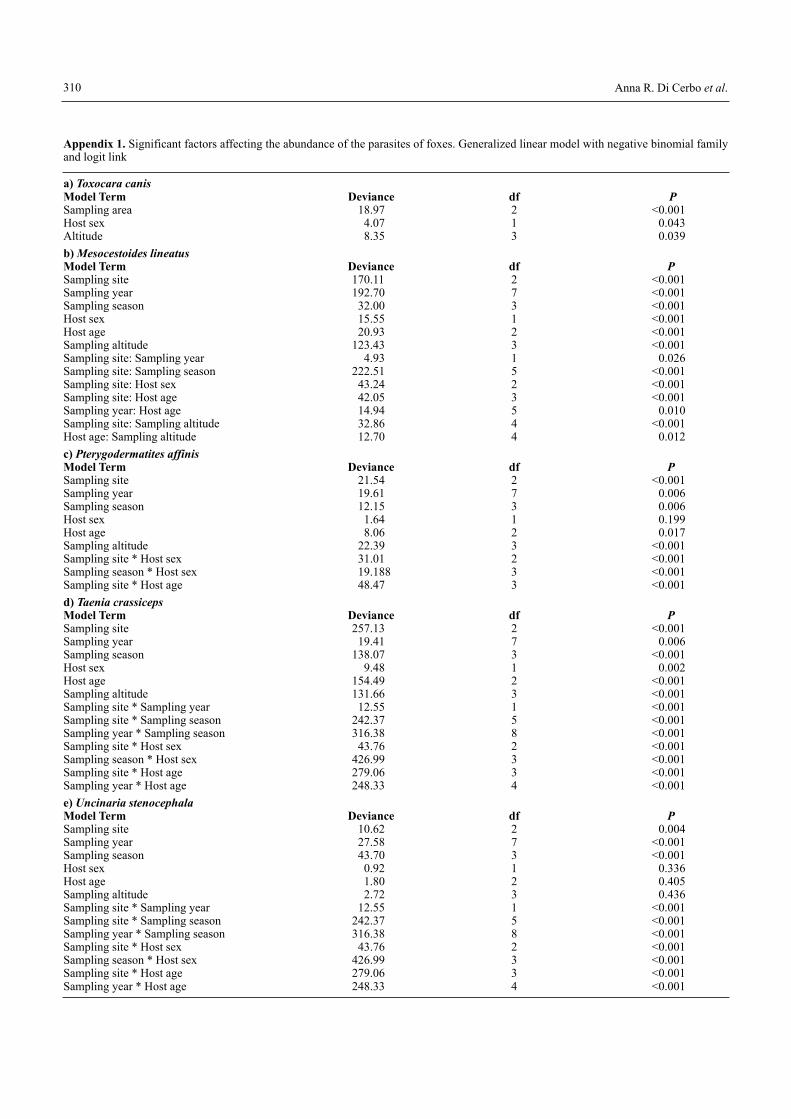
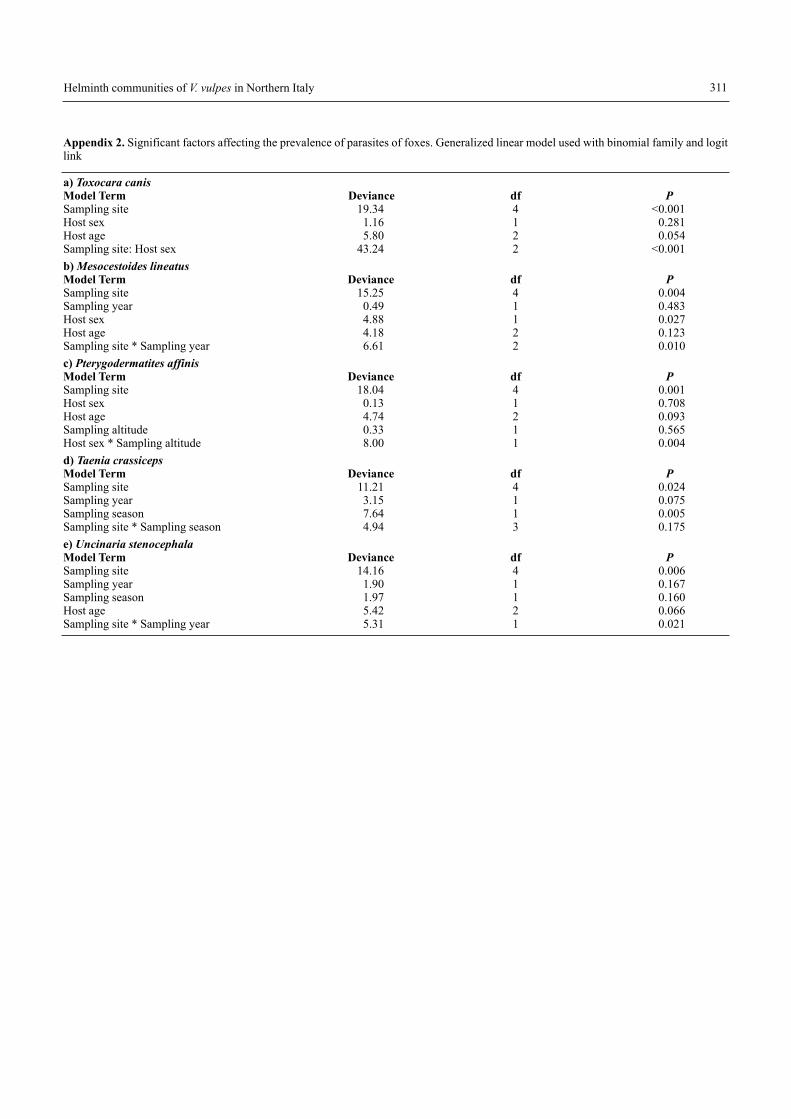
The host factors had a lower influence with respect to ex-trinsic factors, and age or sex of the fox influenced only abun-dance of 3 parasites (age: T. crassiceps, M. lineatus, P. affinis;sex: T. crassiceps, M. lineatus, T. canis), suggesting that hostdynamics play a trivial role with respect to spatial variabilityin determining parasite abundance (Appendices 1 and 2).
Discussion
The results of this study (i) pointed out high prevalence ofintestinal helminth infections in the Italian Alpine fox, (ii) con-firmed the typical composition of parasite fauna within foxpopulations, (iii) underlined local differences concerning dis-tribution and abundance of the parasite species.
This pattern could be due both to intrinsic and extrinsicfactors such as environment, demographics, physiological and/or immunological characteristics of host subpopulations (Pen-ce and Windberg 1984). The results obtained through ourinvestigation highlight the major role of extrinsic factors vsintrinsic ones.
Helminth community patterns in the five study areas
Diversity indices allow a better understanding of certain dif-ferences in the helminth communities. It is interesting to notethat areas where the highest parasite richness and dominancewere found (Belluno province and Lombardy) did not corre-spond to the area where the highest total number of specieswas collected (Trento province). This demonstrates that boththe number of species and the parasite abundance play animportant role in the community structure. In fact, these in-dices are based on the proportional abundance of species � inparticular, of the most common species as to dominance
measurement � and they are more sensitive measure of rich-ness and diversity of a community, rather than merely mea-sure of the number of species in a single unit. At the infracom-munity level, we found the maximum number of species perhost (6) in the provinces of Trento and Aosta, the latter onlyshowing the highest mean species (3.08). Neither was a cor-relation found between richness and diversity indices of hel-minth communities and prevalence, nor a real correspondencebetween the parasite species number and the number of sam-ples collected in each area. This even though total speciesrichness is known to depend upon host sample size (e.g., Wal-ther et al. 1995).
Dominant role within the communities was demonstratedfor T. crassiceps, M. lineatus, T. canis, U. stenocephala andP. affinis. However, hierarchical differences were observedamong individual provinces. Uncinaria stenocephala exhibit-ed the highest dominant index in all the areas except for Bol-zano province, where T. canis resulted to be the most relevantspecies. The importance of cestodes in the intestinal helminthfauna of V. vulpes is likely due to the diet of this carnivore inAlpine areas, which includes small and medium sized mam-mals (Bertolini et al. 2001, Cagnacci et al. 2003), intermedi-ate hosts for the larval stages of cestodes.
Interestingly, E. multilocularis was found only in the east-ern province of Bolzano (Manfredi et al. 2003a, Manfredi etal. 2004). In fact, all the foxes infected (5) with E. multilocu-laris came from localities situated near the Austrian border, ataltitudes between 1,000 and 1,600 m. The parasitological datashow a very scanty spreading of E. multilocularis infection inthe entire study area (0.8%). However, where recorded, it doesnot play a marginal role in the helminth community. Theworm burden (8�100 specimens) in our foxes is much lowerthan that surveyed, for instance, in Switzerland, where 31%out of 133 infected foxes indicated burdens between 1,000and approximately 57,000 worms (Hofer et al. 2000). Pre-vious studies of foxes from the Alpine area argued that theseanimals can take over vast territories � home ranges varyingfrom 107 ha and 170 ha � and their nocturnal movement ismainly directed down in the valley to the villages with a plen-tiful supply of food such as cultivated fruits, rubbish, and evendomestic animals (Boitani and Vinditti 1988, Cagnacci et al.2003). The presence of foxes on pastures at higher altitudes isassociated with their diurnal movement and refuges. Consid-ering the spatial behaviour of this canid and its range of movement in the alpine habitat, E. multilocularis, seems tohave been introduced by infected foxes immigrated from near-by Austria, where the parasite is endemic. However, theresults of PCR analysis and a retrospective study on histori-cal data about human cases in Tyrol considered the probablepresence of an autochthonous life cycle of the parasite in somevalleys of Trentino Alto Adige region (Manfredi et al. 2004,Casulli et al. 2005).
As regard to P. affinis, it was first recorded by Iori andLeto (1990) in the Italian regions of Lombardy, Trentino andVeneto, and later it was observed in the eastern regions (Stan-campiano et al. 1998). Our data confirm the presence of this
307
Table IV. Minimal adequate GLM between taxa by fox, as response,and extrinsic and host characteristics, as explanatory variables
Model terms Deviance df P value
Site 21.65 4 <0.001Year 17.35 7 0.031Fox age 8.124 2 0.017Site*Year 4.689 1 0.030
Table V. Minimal adequate GLM between total parasite load by fox,as response, and extrinsic and host characteristics, as explanatoryvariables
Model terms Deviance df P value
Site 38.670 4 <0.001Year 42.42 8 <0.001Site*Year 21.184 4 <0.001

Anna R. Di Cerbo et al.
parasite in Lombardy, Trentino and Veneto, and also its geo-graphical distribution to include Aosta Valley region. The lat-ter, compared with other Italian Alpine regions, shows a ratherdry climate since its high mountains create a barrier to thewetter weather from the west. In particular, its central part in-cluded between the village of Saint-Vincent and the regionalcapital, Aosta, undergoes an average rainfall less than 600 mmper yr. Just here we recorded the highest values of prevalenceand abundance of the parasite. In agreement with our obser-vations, it can be reminded that P. affinis seems to be partic-ularly associated with a semiarid habitat (GortBzar et al.1998), which can at least partially explain the data recorded.
Factors affecting single parasite species
General linear models revealed some environmental factorsinfluencing mean abundance and prevalence of the parasites.First, the composition of helminth communities can vary ac-cording to geographical areas. Second, the altitude seems toaffect cestodes and T. canis and P. affinis. Third, a seasonaldynamic of some parasites is likely. In fact, U. stenocephalaand P. affinis were more abundant in summer. U. stenocepha-la has a direct life cycle and the infection occurs via oral-ingestion of free-living larvae (L3). Temperature represents alimiting factor. This parasite developing time, from its hatch-ing to its infective stage, is inversely proportional to the tem-perature (Gibbs and Gibbs 1959). For instance, its abundanceincreases between July and September in open areas (Urqu-hart et al. 1996).
Mesocestoides lineatus and T. crassiceps were more fre-quently observed in summer and in autumn respectively, assmall mammals, i.e. paratenic or intermediate hosts, are morecommon in the diet of Alpine fox population in these periods(Bertolini et al. 2001, Cantini 1991).
Differences were, however, recorded in the age of infect-ed foxes, as young individuals generally harboured more par-asites than the adults. Particularly, differences emerged for U.stenocephala, perhaps due to the inefficient immunity ofyoung foxes and to their greater exposure to the various stagesof infections (Richards et al. 1995, Suchentrunck and Sat-tmann 1994).
Generally speaking, the intestinal helminth fauna of foxesof the Italian Alps appears to be abundant and characterized byspecies already considered typical. Nonetheless, in our pres-ent study we were able to highlight the significant prevalenceof P. affinis in the more western fields of the Alps and torecord for the first time the presence of the nematode O. cras-sispiculum and trematodes M. vulpis and P. elegans in Italy. Inaddition, we signalled low prevalence as to E. multilocularisand only in foxes from the Bolzano province.
Summing up, on the basis of our epidemiological data V.vulpes may constitute a reservoir of selected parasites of vet-erinary and public health importance in Northern Italy; werefer, for example, to T. canis, responsible for visceral andocular larva migrans. This parasite was found to be geo-graphically widespread and abundant in Italian fox popula-
tions, in addition to E. multilocularis which causes a muchmore serious zoonosis, namely alveolar echinococcosis.
Acknowledgments. All animals used in this research were collect-ed according to Italian national law (L. 157/92) and to Regional reg-ulations. The research was funded by a grant from MIUR, PRIN2003 Prot. 2003070410_005. The authors are grateful to two anony-mous referees for valuable comments on the manuscript, and toGigliola Canepa for her review of our original English manuscript.
References
Bertolini M., Pallaveri A., Pedrini P. 2001. Spettro alimentare dellavolpe (Vulpes vulpes) in Trentino (NE Italia) mediante l�anali-si dei contenuti gastrici. Studi Trentini di Scienze Naturali �Acta Biologica, 78, 71�76.
Boitani L., Vinditti R.M. 1988. La volpe rossa. Edagricole, Bologna,241 pp.
Bush A.O., Lafferty K.D., Lotz M.J., Shostak A.W. 1997. Parasi-tology meets ecology on its own terms: Margolis et al. revis-ited. Journal of Parasitology, 83, 575�583. DOI: 10.2307/3284227.
Cagnacci F., Lovari S., Meriggi A. 2003. Carrion dependence andfood habitus of the red fox in an Alpin area. Italian Journalof Zoology, 70, 31�38.
Cantini M. 1991. The diet of the fox (Vulpes vulpes) in woodlandsof Orobie Alps (Lombardy region, Northern Italy. Hystrix,Italian Journal of Mammology, 3, 83�89.
Casulli A., La Rosa G., Manfredi M.T., Di Cerbo A.R., Dinkel A.,Romig T., Deplazes P., Genchi C., Pozio E. 2005. Echino-coccus multilocularis in red foxes (Vulpes vulpes) of theItalian Alpine region: is there a focus of autochthonous trans-mission? International Journal for Parasitology, 35, 1079�1083. DOI: DOI: 10.1016/j.ijpara.2005.04.005.
Crawley M.J. 2002. Statistical Computing: An Introduction to dataanalysis using S-plus. John Wiley & Sons, Ltd, Chichester,772 pp.
Deplazes P., Hegglin D., Gloor S., Romig T. 2004. Wilderness in thecity: The urbanization of Echinococcus multilocularis. Trendsin Parasitology, 20, 77�84. DOI: 10.1016/j.pt.2003.11.011.
Eira C., Torres J., Viganda J., Miquel J. 2006a. Ecological aspectsinfluencing the helminth community of the wood mouse Apo-demus sylvaticus in Dunas de Mira, Portugal. Acta Parasito-logica, 51, 300�308. DOI: 10.2478/s11686-006-0046-0.
Eira C., Viganda J., Torres J., Miquel J. 2006b. The helminth com-munity of the red fox, Vulpes vulpes, in Dunas de Mira (Por-tugal) and its effect on host condition. Wildlife Biology inPractice, 2, 26�36. DOI: 10.2461/wbp.2006.2.5.
Gibbs H.C., Gibbs K.E. 1959. The effect of temperature on the devel-opment of the free-living stages of the Dochmoides steno-cephala (Railliet, 1884) (Ancylostomidae: Nematoda). Cana-dian Journal of Zoology, 37, 247�257. DOI: 10.1139/z59-030.
GortBzar C., Villafuerte R., Lucientes J., FernBndez-de-Luco D.1998. Habitat related differences in helminth parasites of redfoxes in the Ebro Valley. Veterinary Parasitology, 80, 75�81.DOI: 10.1016/S0304-4017(98)00192-7.
Guberti V., Poglayen G. 1991. Zoonosi parassitarie: indagini in volpi(Vulpes vulpes) dell�Appennino Settentrionale. Hystrix, Ital-ian Journal of Mammology, 3, 167�173.
Guglielmi R., Nappi A., Giannotti M. 2003. Presenza di volpe Vulpesvulpes in contesto fortemente urbanizzato di periferia metro-politana: il caso del quartiere ScampXa a Napoli. Hystrix, Ital-ian Journal of Mammology, 14, 55�56.
Hofer S., Gloor S., Muller U., Mathis A., Hegglin D., Deplazes P.2000. High prevalence of Echinococcus multilocularis in ur-
308

Helminth communities of V. vulpes in Northern Italy
ban red foxes (Vulpes vulpes) and voles (Arvicola terrestris)in the city of Zurich, Switzerland. Parasitology, 120, 135�142. DOI: 10.1017/S0031182099005351.
Holmes J.C., Price P.W. 1986. Communities of parasites. In: (Eds J.Kikkawa and D.J. Anderson) Community Ecology: Patternand Process. Blackwell Scientific Publications, Oxford, 187�213.
Iori A., Costantini R., Cancrini G. 1990. Parassiti in volpi provenientida alcune regioni italiane. Parassitologia, 32, 153�154.
Iori A., Leto A. 1990. Prima segnalazione di Pterygodermatites (Mul-tipectines) affinis (Jagerskiold, 1904; Quentin, 1969) nellavolpe in Italia. Parassitologia, 32, 359�362.
Lucherini M., Lovari S., Crema G. 1995. Habitat use and rangingbehaviour of the red foxes in a mediterranean rural area: isshelter availability a key factor? Journal of Zoology, 237,577�591.
Magurran A.E. 1988. Ecological diversity and its measurement.Great Britain University Press, Cambridge, 192 pp.
Manfredi M.T., Di Cerbo A.R., Trevisiol K. 2004. An update of theepidemiological situation of Echinococcus multilocularis inTrentino Alto-Adige (Northern Italy). Parassitologia, 46, 431�433.
Manfredi M.T., Di Cerbo A.R., Trevisiol K., Lombardo D., BregoliM., Ferro N. 2003a. La diffusione di Echinococcus multilo-cularis nel nord Italia (Cestoda; Taenidae). Giornale Italianodi Malattie Infettive, 9, 200�204.
Manfredi M.T., Genchi C., Deplazes P., Trevisiol K., Fraquelli C.2002. Echinococcus multilocularis infection in red foxes inItaly. Veterinary Record, 150, 757�758.
Manfredi M.T., Giacometti A., Fraquelli C., Piccolo G. 2003b. Studiodella popolazione elmintica in volpi (Vulpes vulpes) del Tren-tino Alto Adige. Atti II Convegno Nazionale di Ecopatologiadella Fauna, Bormio 8�10 October 1998. Journal of Moun-tain Ecology, 7, 261�263.
Marinelli A., Sorino R., Bux M., Scillitani G. 2003. L�ecologia trofi-ca della volpe Vulpes vulpes e della faina Martes foina in unhabitat mediterraneo. Hystrix, Italian Journal of Mammology,14, 59.
Matthee S., Krecek R.C., McGeoch M.A. 2004. A comparison of theintestinal helminth communities of Equidae in Southern Af-rica. Journal of Parasitology, 90, 1263�1273. DOI: 10.1645/GE-3353.
McCullagh P., Nelder J.A. 1983. Generalized linear models. Chap-man & Hall, London, 261 pp.
Meriggi A., Rosa P. 1991. Fattori influenzanti la distribuzione delletane di volpe (Vulpes vulpes) in Italia Settentrionale. Hystrix,Italian Journal of Mammology, 3, 31�39.
Newcombe R.G. 1998. Two-sided confidence intervals for the singleproportion: comparison of seven methods. Statistics in Medi-cine, 17, 857�872. DOI: 10.1002/(SICI)1097-0258(19980430)17:8<857::AID-SIM777>3.0.CO;2-E.
Oindo B.O. 2001. Body size and abundance relationship: an indexof diversity for herbivores. Biodiversity and Conservation,10, 1923�1931. DOI: 10.1023/A:1013191419831.
Pence D.B. 1990. Helminth community of mammalian hosts: con-cepts at the infracommunity, component and compound com-munity levels. In: (Eds. G.W. Esch, A.O. Bush and J.M. Aho)Parasite Communities: Patterns and Processes. Chapman andHall, London, 233�260.
Pence D.B., Windberg L.A. 1984. Population dynamics across selec-ted habitat variables of the helminth community in coyotes, Canis latrans, from south Texas. Journal of Parasitology, 70,735�746. DOI: 10.2307/3281756.
(Accepted March 18, 2008)
Poglayen G., Guberti V., Leoni B. 1985. Parassiti presenti in volpi(Vulpes vulpes) nella provincia di ForlX. Parassitologia, 27,303�311.
Poulin R. 2001. Interactions between species and structure of hel-minth communities. Parasitology, 122, S3�S11. DOI: 10.1017/S0031182000016991.
Pozio E. 1991. La volpe (Vulpes vulpes L.) principale serbatoio dellatrichinellosi in Italia. Hystrix, Italian Journal of Mammology,3, 175�186.
Richards D.T., Harris S., Lewis J.W. 1995. Epidemiological studieson intestinal helminth parasites of rural and urban red foxes(Vulpes vulpes) in the United Kingdom. Veterinary Parasi-tology, 59, 39�51. DOI: 10.1016/0304-4017(94)00736-V.
Roberts M.G., Dobson A.P., Arneberg P., de Leo G.A., Krecek R.C.,Manfredi M.T., Lanfranchi P., Zaffaroni E. 2002. Parasitecommunity ecology and biodiversity. In: (Eds. P.J. Hudson,A.P. Rizzoli, B.T. Grenfell, H. Heesterbeek and A.P. Dobson)The Ecology of Wildlife Diseases. Oxford University Press,Oxford, 63�82.
Rossi L., Iori A., Cancrini G. 1983. Osservazioni sulla fauna paras-sitaria della popolazione di volpi presente nel parco regionale�La Mandria�. Parassitologia, 25, 340�343.
Segovia J.M., Torres J., Miquel J. 2004. Helminth parasites of the redfox (Vulpes vulpes L., 1758) in the Iberian Peninsula: An eco-logical study. Acta Parasitologica, 49, 67�79.
Soldati G., Pavesi M., Canestri Trotti G., Cocchi M.G., Gaiardi S.,Morganti L., Prosperi S., Sanguinetti V., Stanzani F. 1976.Research on infectious and parasitic agents in foxes on theModenese Apennines. Rivista di Parassitologia, 37, 229�233.
Stancampiano L., Capelli G., Schiavon E., Mutinelli F., Bozzolan G.1998. Trichinellosis, sarcoptic mange, filariosis and intesti-nal helminths stability in a fox population (Vulpes vulpes).Parassitologia, 40, 171.
Suchentrunk F., Sattmann H. 1994. Prevalence of intestinal hel-minths in Austrian red foxes (Vulpes vulpes). Annalen desNaturhistorischen Museums Wien, 96B, 29�38.
Thul J.E., Forrester D.J., Abercombie C.L. 1985. Ecology of parasitichelminths of wood duck, Air sponsa, in the Atlantic flyway.Proceedings of the Helminthological Society of Washington,52, 297�310.
Urquhart G.M., Armour J., Duncan J.L., Dunn A.M., Jennings F.W.1996. Veterinary Parasitology. Blackwell Science Publica-tions, Iowa State, 307 pp.
Walther B.A., Cotgreave P., Gregory R.D., Price R.D., Clayton D.H.1995. Sampling effort and parasite species richness. Para-sitology Today, 11, 306�310. DOI: 10.1016/0169-4758(95)80047-6.
Wilson E.B. 1927. Probable inference, the law of succession, and sta-tistic inference. Journal of American Statistical Association,22, 209�212. DOI: 10.2307/2276774.
Wilson K., Birrnstad O.N., Dobson A.P., Merler S., Poglayen G.,Randolph S.E., Read A.F., Skorping A. 2002. Heterogeneitiesin macroparasite infection: patterns and processes. In: (Eds.P.J. Hudson, A.P. Rizzoli, B.T. Grenfell, H. Heesterbeek andA.P. Dobson) The Ecology of Wildlife Diseases. Oxford Uni-versity Press, Oxford, 6�44.
Zaffaroni E., Fraquelli C., Manfredi M.T., Siboni A., Lanfranchi P.,Sartori E., Partel P. 1996. Abomasal helminth communities ineastern Alpine sympatric roe deer (Capreolus capreolus) andchamois (Rupicapra rupicapra) populations. SupplementoRicerche Biologia della Selvaggina, 24, 53�68.
309
Anna R. Di Cerbo et al.310
Appendix 1. Significant factors affecting the abundance of the parasites of foxes. Generalized linear model with negative binomial familyand logit link
a) Toxocara canis Model Term Deviance df PSampling area 18.97 2 <0.001Host sex 4.07 1 0.043Altitude 8.35 3 0.039b) Mesocestoides lineatusModel Term Deviance df PSampling site 170.11 2 <0.001Sampling year 192.70 7 <0.001Sampling season 32.00 3 <0.001Host sex 15.55 1 <0.001Host age 20.93 2 <0.001Sampling altitude 123.43 3 <0.001Sampling site: Sampling year 4.93 1 0.026Sampling site: Sampling season 222.51 5 <0.001Sampling site: Host sex 43.24 2 <0.001Sampling site: Host age 42.05 3 <0.001Sampling year: Host age 14.94 5 0.010Sampling site: Sampling altitude 32.86 4 <0.001Host age: Sampling altitude 12.70 4 0.012c) Pterygodermatites affinis Model Term Deviance df PSampling site 21.54 2 <0.001Sampling year 19.61 7 0.006Sampling season 12.15 3 0.006Host sex 1.64 1 0.199Host age 8.06 2 0.017Sampling altitude 22.39 3 <0.001Sampling site * Host sex 31.01 2 <0.001Sampling season * Host sex 19.188 3 <0.001Sampling site * Host age 48.47 3 <0.001d) Taenia crassicepsModel Term Deviance df PSampling site 257.13 2 <0.001Sampling year 19.41 7 0.006Sampling season 138.07 3 <0.001Host sex 9.48 1 0.002Host age 154.49 2 <0.001Sampling altitude 131.66 3 <0.001Sampling site * Sampling year 12.55 1 <0.001Sampling site * Sampling season 242.37 5 <0.001Sampling year * Sampling season 316.38 8 <0.001Sampling site * Host sex 43.76 2 <0.001Sampling season * Host sex 426.99 3 <0.001Sampling site * Host age 279.06 3 <0.001Sampling year * Host age 248.33 4 <0.001e) Uncinaria stenocephalaModel Term Deviance df PSampling site 10.62 2 0.004Sampling year 27.58 7 <0.001Sampling season 43.70 3 <0.001Host sex 0.92 1 0.336Host age 1.80 2 0.405Sampling altitude 2.72 3 0.436Sampling site * Sampling year 12.55 1 <0.001Sampling site * Sampling season 242.37 5 <0.001Sampling year * Sampling season 316.38 8 <0.001Sampling site * Host sex 43.76 2 <0.001Sampling season * Host sex 426.99 3 <0.001Sampling site * Host age 279.06 3 <0.001Sampling year * Host age 248.33 4 <0.001
Helminth communities of V. vulpes in Northern Italy 311
Appendix 2. Significant factors affecting the prevalence of parasites of foxes. Generalized linear model used with binomial family and logitlink
a) Toxocara canis Model Term Deviance df PSampling site 19.34 4 <0.001Host sex 1.16 1 0.281Host age 5.80 2 0.054Sampling site: Host sex 43.24 2 <0.001b) Mesocestoides lineatusModel Term Deviance df PSampling site 15.25 4 0.004Sampling year 0.49 1 0.483Host sex 4.88 1 0.027Host age 4.18 2 0.123Sampling site * Sampling year 6.61 2 0.010c) Pterygodermatites affinisModel Term Deviance df PSampling site 18.04 4 0.001Host sex 0.13 1 0.708Host age 4.74 2 0.093Sampling altitude 0.33 1 0.565Host sex * Sampling altitude 8.00 1 0.004d) Taenia crassiceps Model Term Deviance df PSampling site 11.21 4 0.024Sampling year 3.15 1 0.075Sampling season 7.64 1 0.005Sampling site * Sampling season 4.94 3 0.175e) Uncinaria stenocephalaModel Term Deviance df PSampling site 14.16 4 0.006Sampling year 1.90 1 0.167Sampling season 1.97 1 0.160Host age 5.42 2 0.066Sampling site * Sampling year 5.31 1 0.021
Related Documents











