Interval-timing deficits in individuals at high risk for schizophrenia q Trevor B. Penney a, * , Warren H. Meck b , Simone A. Roberts c , John Gibbon d , L. Erlenmeyer-Kimling e a Department of Psychology, Chinese University of Hong Kong, Shatin, N.T., Hong Kong, Hong Kong b Department of Psychological and Brain Sciences, Duke University, United States c Department of Medical Genetics, New York State Psychiatric Institute, United States d Department of Psychology, Columbia University and New York State Psychiatric Institute, United States e Department of Medical Genetics, Columbia University and New York State Psychiatric Institute, United States Accepted 16 September 2004 Available online 2 December 2004 Abstract A duration-bisection procedure was used to study the effects of signal modality and divided attention on duration classification in participants at high genetic risk for schizophrenia (HrSz), major affective disorder (HrAff), and normal controls (NC). Participants learned short and long target durations during training and classified probe durations during test. All groups classified visual signals as shorter than equivalent duration auditory signals. However, the difference between auditory and visual signal classification was significantly larger for the HrSz group than for the NC group. We posit a model in which there is a clock rate difference between auditory and visual signals due to an attentional effect at the level of a mode switch that gates pulses into an accumulator. This attentionally mediated clock rate difference was larger for the HrSz participants than for the NC participants, resulting in a larger auditory/visual difference for the HrSz group. Ó 2004 Elsevier Inc. All rights reserved. 1. Introduction Overestimation of duration (e.g., a 10-s stimulus elic- its a verbal label larger than 10 s) in the seconds range has been reported for patients with schizophrenia (Sz) when they are asked to verbally estimate the duration of a recently presented stimulus (Clausen, 1950; Densen, 1977; Johnson & Petzel, 1971; Lhamon & Goldstone, 1956; Wahl & Sieg, 1980; Weinstein, Goldstone, & Boardman, 1958). In addition, underproduction in the seconds range (e.g., the participant responds before 10 s has elapsed when required to respond at 10 s) has been reported when participants with Sz are presented with either a standard duration stimulus (reproduction task) or a verbal label for that standard duration (pro- duction task) and are required respond when the com- parison stimulus has been present for the appropriate amount of time (Clausen, 1950; Johnson & Petzel, 1971; Tysk, 1983, 1990; Wahl & Sieg, 1980). The overes- timation and underproduction results obtained in these tasks suggest that the representation of time is systemat- ically distorted in individuals with Sz. Explanations of the timing dysfunction in Sz are considered in greater detail below, but, in brief, the distortion could be due to a ‘‘fast’’ clock, meaning subjective time accumulates faster than objective time, or, alternatively, a systematic shortening of duration in memory, meaning a duration held in memory appears shorter when compared with 0278-2626/$ - see front matter Ó 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.bandc.2004.09.012 q This research was supported in part by National Institute of Mental Health Grant MH19560 from the US Public Health Service (L. Erlenmeyer-Kimling) and Grant MH20906 from the National Institute of Mental Health (Clinical Research Center, New York State Psychiatric Institute), Rockville, MD. We thank Donald Rock, Ph.D., for his statistical advice, Stephen Fairhurst for his program- ming efforts, and Courtney Strand for her help with data entry and analysis. * Corresponding author. Fax: +1 852 2603 5019. E-mail address: [email protected] (T.B. Penney). www.elsevier.com/locate/b&c Brain and Cognition 58 (2005) 109–118

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Interval-timing deficits in individuals at high risk for schizophreniaq
Trevor B. Penneya,*, Warren H. Meckb, Simone A. Robertsc,John Gibbond, L. Erlenmeyer-Kimlinge
a Department of Psychology, Chinese University of Hong Kong, Shatin, N.T., Hong Kong, Hong Kongb Department of Psychological and Brain Sciences, Duke University, United States
c Department of Medical Genetics, New York State Psychiatric Institute, United Statesd Department of Psychology, Columbia University and New York State Psychiatric Institute, United States
e Department of Medical Genetics, Columbia University and New York State Psychiatric Institute, United States
Accepted 16 September 2004Available online 2 December 2004
Abstract
A duration-bisection procedure was used to study the e!ects of signal modality and divided attention on duration classification inparticipants at high genetic risk for schizophrenia (HrSz), major a!ective disorder (HrA!), and normal controls (NC). Participantslearned short and long target durations during training and classified probe durations during test. All groups classified visual signalsas shorter than equivalent duration auditory signals. However, the di!erence between auditory and visual signal classification wassignificantly larger for the HrSz group than for the NC group. We posit a model in which there is a clock rate di!erence betweenauditory and visual signals due to an attentional e!ect at the level of a mode switch that gates pulses into an accumulator. Thisattentionally mediated clock rate di!erence was larger for the HrSz participants than for the NC participants, resulting in a largerauditory/visual di!erence for the HrSz group.! 2004 Elsevier Inc. All rights reserved.
1. Introduction
Overestimation of duration (e.g., a 10-s stimulus elic-its a verbal label larger than 10 s) in the seconds rangehas been reported for patients with schizophrenia (Sz)when they are asked to verbally estimate the durationof a recently presented stimulus (Clausen, 1950; Densen,1977; Johnson & Petzel, 1971; Lhamon & Goldstone,1956; Wahl & Sieg, 1980; Weinstein, Goldstone, &
Boardman, 1958). In addition, underproduction in theseconds range (e.g., the participant responds before10 s has elapsed when required to respond at 10 s) hasbeen reported when participants with Sz are presentedwith either a standard duration stimulus (reproductiontask) or a verbal label for that standard duration (pro-duction task) and are required respond when the com-parison stimulus has been present for the appropriateamount of time (Clausen, 1950; Johnson & Petzel,1971; Tysk, 1983, 1990; Wahl & Sieg, 1980). The overes-timation and underproduction results obtained in thesetasks suggest that the representation of time is systemat-ically distorted in individuals with Sz. Explanations ofthe timing dysfunction in Sz are considered in greaterdetail below, but, in brief, the distortion could be dueto a ‘‘fast’’ clock, meaning subjective time accumulatesfaster than objective time, or, alternatively, a systematicshortening of duration in memory, meaning a durationheld in memory appears shorter when compared with
0278-2626/$ - see front matter ! 2004 Elsevier Inc. All rights reserved.doi:10.1016/j.bandc.2004.09.012
q This research was supported in part by National Institute ofMental Health Grant MH19560 from the US Public Health Service (L.Erlenmeyer-Kimling) and Grant MH20906 from the National Instituteof Mental Health (Clinical Research Center, New York StatePsychiatric Institute), Rockville, MD. We thank Donald Rock,Ph.D., for his statistical advice, Stephen Fairhurst for his program-ming e!orts, and Courtney Strand for her help with data entry andanalysis.
* Corresponding author. Fax: +1 852 2603 5019.E-mail address: [email protected] (T.B. Penney).
www.elsevier.com/locate/b&c
Brain and Cognition 58 (2005) 109–118

a currently elapsing duration of equivalent magnitude(Meck, 1996).
Sz is also of interest for investigations of interval tim-ing because it has been linked to dysfunctions of thedopamine (DA) system and a relationship between braindopaminergic systems and interval timing has beenestablished. For example, the clinical potencies of e!ec-tive antipsychotic drugs are positively correlated withtheir ability to block specific DA receptor subtypes(e.g., D2 and D4—Seeman, 1992, 1995) and, impor-tantly, a correlation between binding a"nity for theD2 receptor and the neuroleptic dose required to achievea 10–15% reduction in the magnitude of perceived dura-tion in an interval timing task has been observed in rats,and has been related to frontal–striatal function in hu-mans and other animals (Lustig & Meck, 2005; Meck,1986; Meck & Benson, 2002).
The importance of normal DA function for temporalprocessing is also suggested by data from individualswith Parkinson!s Disease (PD). For example, PD pa-tients evaluated o! LL-dopa medication underestimatedduration in a verbal time estimation task, overproducedduration in a temporal reproduction task, and, com-pared to controls, required a larger minimum time inter-val between paired stimuli in order to perceive the twostimuli as separated in time and administration of LL-dopa significantly improved the accuracy of patientson all three tasks (Artieda, Pastor, Lacruz, & Obeso,1992; Pastor, Artieda, Jahanshahi, & Obeso, 1992).More recently, Malapani and colleagues have demon-strated dissociable deficits of temporal memory encod-ing and retrieval in patients with PD by manipulatingwhether PD patients were on or o! LL-dopa medicationat study and test in a reproduction task (the peak-inter-val procedure). They found that, depending on the spe-cific combination of drug manipulation with study-testcharacteristics, dopamine deficiency could result in dis-tortions when storing multiple time intervals that ledto overproduction of all intervals at test, or retrieval dis-tortions such that short intervals were distorted longerand long intervals were distorted shorter (Malapaniet al., 1998, Malapani, Deweer, & Gibbon, 2002).
Given the known timing deficits manifested in Sz andthe experimental evidence implicating the DA system ininterval timing, further examination of interval timing inSz populations appears warranted. One di"culty withstudying cognitive processing in a psychiatric popula-tion, however, is that performance deficits may be a con-sequence of a lack of motivation or task comprehension.In addition, psychiatric patients are often medicated andmost psychoactive drugs have varied e!ects on cognitiveprocessing. Studying individuals at genetic risk for thepsychiatric disorder in question, however, may circum-vent some of these di"culties. Given that individualsat high risk for developing schizophrenia (i.e., first de-gree relatives of diagnosed patients) sometimes exhibit
structural and functional abnormalities of the brain,specifically frontal–striatal circuitry, that are similar tothose exhibited by patients with schizophrenia (seeCannon, 1996; for review) they form an interestingpopulation for further studies of timing and timeperception.
Although the first-degree relatives of diagnosed Szpatients are at substantially greater risk of developingSz than are members of the general population, the pat-tern of empiric genetic risks for schizophrenia in rela-tives of schizophrenic probands is not Mendelian, butsuggests instead the action of multiple genes (Gottes-man, McGu"n, & Farmer, 1987). Moreover, given thatthe concordance rate for the disorder is only about 45%in monozygotic twins (Gottesman, Shields, & Hanson,1982; Kendler, 1988), it is clear that the clinical illnessis not expressed in all individuals who carry the full ge-netic liability. Based on cumulative data collectedacross a number of studies, the risk to o!spring ofone schizophrenic parent, the relatives most commonlystudied in high-risk research, has been estimated to be12% (Gottesman et al., 1987; Holzman & Matthysse,1990). Some clinically una!ected individuals, however,may show cognitive or other neurobehavioral deficitsassociated with schizophrenia. Thus, the rate of suchdeficits in high-risk individuals may be greater thanthe expected rate of the illness itself (e.g., Erlenmeyer-Kimling, 1996).
Here, o!spring of patients diagnosed with Sz werestudied to determine whether individuals at relativelyhigh risk for developing Sz (HrSz) show interval-timingdeficits as compared to normal controls (NC). In addi-tion, a group of participants at relatively high risk fora!ective disorders (HrA!) was included to ensure thatany deficits were specific to the risk for Sz and not a gen-eral risk for psychiatric disorders. Using high-risk par-ticipants allowed us to avoid some of the problemsassociated with studies of psychiatric populations de-scribed above.
We used a duration-bisection procedure to examineinterval timing. The participants were initially trainedto discriminate between a short and a long duration sig-nal, the anchor durations. In a subsequent test phase,the participants classified probe signals as short or longrelative to the anchor durations learned in training.Some of these probe signals were the same as the anchordurations, but most were of intermediate duration.Moreover, in the present experiment, participants wererequired to simultaneously time an auditory and a visualprobe signal on some trials and time a single probe sig-nal (auditory or visual) on other trials. Duration-bisec-tion designs including two signal modalities and trialsrequiring one or two signals to be timed have been usedpreviously to examine attention and memory e!ects ininterval timing (e.g., Lustig & Meck, 2001; Meck,2003; Penney, 2003; Penney, Gibbon, & Meck, 2000).
110 T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118

2. Method
2.1. Participants
The participants in the present study were from theNew York High-Risk Project (NYHRP), a longitudinalstudy of Sz, which has been described in detail by Erlen-meyer-Kimling and Cornblatt (1987, 1992). In brief, par-ticipants were enrolled in the NYHRP in 1977–1979(Sample B). They were 7–12 years old at the time, andwere free of mental retardation or serious psychiatric dis-turbances. Participants in the group at high risk forschizophrenia (HrSz) had one parent diagnosed asschizophrenic. Participants in the group at high risk formajor a!ective disorder (HrA!) had one parent diag-nosed with major a!ective disorder. The parents of theparticipants had been diagnosed at several large state psy-chiatric hospitals in lower New York State. Diagnosedparents were white, English speaking, and in an intactmarriage with the child!s other biological parent at thetime of child recruitment. The normal control group(NC) included only the o!spring of parents not diagnosedwith any psychiatric disorder. They came from familiesliving in neighborhoods similar to those of the at-riskfamilies, and had been matched on the basis of age, sex,and socioeconomic status. The participants in the exper-iment described here formed a sub-sample of Sample B.
The HrSz group comprised 5 females and 12 males,with an average age of 25.5 ± 2.1 (SD) and 13.2 ± 1.7years of education at the time of study. There weretwo control groups, a high-risk major a!ective disordercontrol group, and a normal control group. The HrA!group comprised 8 females and 8 males, with an averageage of 27.1 ± 1.7 and 12.9 ± 1.4 years of education atthe time of study. The NC group comprised 14 femalesand 20 males, with an average age of 25.9 ± 2.2 and14.9 ± 1.8 years of education at the time of study.
2.2. Stimuli and apparatus
The participants were seated approximately 60 cm infront of a Macintosh computer, which was used to pres-ent all experimental stimuli and to record responses. Inthe training phase, signals were presented for either ashort or a long anchor duration. In the testing phase, se-ven signal durations were presented, the two anchorsand five geometrically spaced intermediate probe dura-tions (Penney et al., 2000). Auditory and visual signalswere presented. The auditory signals were tones(880 Hz) produced by the computer tone generatorand played over the computer speaker. The visual sig-nals were black squares measuring approximately 5 cmby 5 cm on the computer monitor. The training phaseanchor durations were 3 and 6 s. In the testing phase,the signal durations were 3.00, 3.67, 3.78, 4.24, 4.76,5.34, and 6.00 s.
2.3. Design and procedure
At the beginning of the session, participants were in-structed that they would have to classify auditory and/or visual signals, presented by computer, as either shortor long in duration. A training phase provided partici-pants with examples of the short and long signal dura-tions. Participants were told not to count or sub-dividethe intervals in any way.
After signal o!set, participants verbally indicatedtheir duration decision (short or long) and the experi-menter entered the decision via the computer keyboard.On screen feedback indicated whether the choice wascorrect or not. Following correct responses, a circle ap-peared around the selected short or long choice box andthe words ‘‘correct—it was short’’ or ‘‘correct—it waslong’’ appeared above the choice box. When the partic-ipant chose incorrectly, a large · appeared on the se-lected choice box and the words ‘‘incorrect—it wasshort’’ or ‘‘incorrect it was long’’ appeared above thechoice box. Feedback was displayed for 3 s. The inter-trial interval varied randomly from 0 to 2 s.
Participants were told that although in the test phaseshort and long judgments were required, they couldthink of the judgment as closer to short or closer to longbecause most of the durations presented would be be-tween the short and the long durations learned in train-ing. They were also told there were no correct orincorrect classifications of these intermediate durations.As a consequence, in the test phase participants receivedfeedback only for those signals that were the same dura-tion (short or long) as those learned in training. Thefeedback was the same as that given in the trainingsession.
Participants were presented with 12 trials in the train-ing phase. Trial type and signal duration were selected inthe following manner. Before each trial the computergenerated a random number between 1 and 100. If thenumber was from 1 to 50 an auditory signal was pre-sented, and if from 51 to 100 a visual signal was pre-sented. Next, a second random number was generatedto determine signal duration. If the number was from1 to 50 a 3-s signal (short) was presented and if from51 to 100 a 6-s signal (long) was presented.
The 150 trials in the test phase were randomly se-lected from single auditory, single visual, and simulta-neous auditory and visual trials in a similar manner,with the exception that random numbers from 1 to 33led to auditory trials, 34 to 66 to visual trials, and 67to 100 to simultaneous trials. In simultaneous trials,both a visual signal and an auditory signal were pre-sented together but with asynchronous onset and vari-able overlap. A delay to signal start was selected foreach signal in two steps. A delay that was uniformly ran-dom between zero and one second was selected for thefirst signal, and summed with a second delay that was
T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118 111

uniformly random between 0 and 1.5 s selected for thesecond signal. The modality order (auditory first or vi-sual first) was random. Participants classified each signalas short or long upon signal termination. This means theparticipant had to attend to and time both signals simul-taneously during the period of overlap. For example, ifthe auditory signal began first the participant com-menced timing the auditory signal, and when the visualsignal began the participant initiated timing that signalas well. When one of the signals terminated the partici-pant stopped timing that signal, but continued timingthe other signal until it also terminated.
Signal duration was determined in two steps. A com-puter generated random number determined whetherone of the two anchor durations or an intermediateprobe duration was presented (numbers from 1 to 30 re-sulted in an anchor duration, 31 to 100 in an intermedi-ate probe). A second random number determined whichanchor (numbers 1–50 resulted in the short anchor and51–100 in the long anchor) or which probe (a 20 numberrange between 1 and 100 for each of the five intermedi-ate probe durations) was presented.
2.4. Data and analysis
The data were analyzed and interpreted within theframework of Scalar Timing Theory, a mathematicaldescription of interval timing that allows the patternsof variability in temporally controlled behavior to beattributed to specific stages of information processing(e.g., Church, 2003; Gibbon, 1977, 1991; Gibbon &Church, 1984; Gibbon, Church, & Meck, 1984). Theinformation-processing model of interval timing de-scribed by Gibbon et al. (1984) consists of three stages:clock, memory, and decision. In the presence of a timingsignal, pulses from a pacemaker are gated, via an atten-tion-modulated switch, into an accumulator. When thesignal terminates, the value of the record in the accumu-lator is compared to the representation of the target timeheld in memory (see Fig. 1). The mathematical, behav-ioral, and physiological underpinnings of this modelare described in detail elsewhere (e.g., Gibbon et al.,1984; Meck, 1996, 2003) as is its application to the dura-tion-bisection procedure used here (Allan & Gibbon,1991; Gibbon, 1981, 1986; Penney, Allan, Meck, & Gib-bon, 1998; Penney, Gibbon, & Meck, 2000).
The classification data obtained in the duration-bisec-tion procedure may be quantified as the proportion oflong responses the participant makes for each signalduration. These data can usually be well described bya sigmoidal function.
Three main measures of bisection performance thatcan be calculated from a fitted sigmoidal function arethe point of subjective equality (PSE or T1/2), the di!er-ence limen (DL), and the Weber fraction (WF). The PSEis the signal duration at which a participant is equally
likely to classify the signal as short or long. It representsthe subjective midpoint between the short and long an-chor values the participant learned in training. TheDL is a measure of the slope of the participant!s re-sponse function. It is calculated by subtracting the dura-tion the participant classifies as long 25% of the timefrom the duration the participant classifies as long 75%of the time and dividing by two. It can be interpretedas a measure of a participant!s temporal precision be-cause steep slopes are indicative of precise temporal pro-cessing whereas shallow slopes indicate greatervariability in the interval-timing system. In a few casesin the present experiment, the fitted sigmoidal functiondid not reach 75% long before the value of the long an-chor duration. When this occurred, a regression equa-tion of area under the response function against the75% long value was determined using the response func-tions from the same condition provided by the otherparticipants. The resulting regression equation was usedto extrapolate a replacement for the missing 75% value(Gibbon & Fairhurst, 1994) for 6 out of the 268 re-sponse functions reported here.
The WF is calculated by dividing the DL by the PSE.It is a measure of timing variability that takes into ac-count the duration being timed. According to ScalarTiming Theory, one of the hallmarks of interval timingis that the coe"cient of variation, here the WF, is con-stant across durations (Gibbon, 1977). That is to say,although the DL increases with increasing GM of theanchor durations the WF value is expected to remainthe same.
In addition, we applied a quantitative model of ScalarTiming Theory developed for duration-bisection dataobtained when the auditory and visual signals are pre-sented in the same test session (Penney et al., 2000). Inthis Sample Known Exactly-Mixed Memories (SKE-
Fig. 1. An information-processing model of interval timing based onscalar timing theory. (Adapted from Meck (1996)).
112 T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118
MM) version of the model there are four parameters. Asensitivity parameter, c, is the coe"cient of variation ofremembered time. It reflects the degree of noise in thememory representation of the short and long signaldurations and is comparable to, although not identicalto, the empirically based Weber ratio. The locationparameter, b, may be thought of as a bias parameterfor the report of long. For the SKE-MM model it is as-sumed that there is no variability in b, or in xT, the per-cept of current time. Relative Rate (RR) is the ratio ofthe visual clock speed to the auditory clock speed. RRequals one when the auditory and visual clock speedsare equivalent and is less than one when the auditoryclock speed is faster than the visual clock speed. Pa isthe proportion of the memory mixture representing theanchor durations contributed by the auditory modalitysignals. Pa can range from zero, meaning no auditorycontribution to the memory, to one, meaning completeauditory dominance of the memory. The mathematicalmodel is developed in full elsewhere (Penney et al.,2000).
The SKE-MM model was fit to individual partici-pant!s response functions using a simplex fitting algo-rithm (Press, Teukolsky, Vettering, & Flannery, 1994).The parameters for the best fit were recorded and sub-mitted to an ANOVA.
3. Results
PSE, DL, and WF values, calculated for eachparticipant!s four psychophysical functions, were testedstatistically by separate three-way, Group (3 levels) ·Modality (2 levels) · Trial Type (2 levels), repeatedmeasures ANOVAs. Group was a between-subjectsfactor and Modality and Trial Type were within-subjects factors.
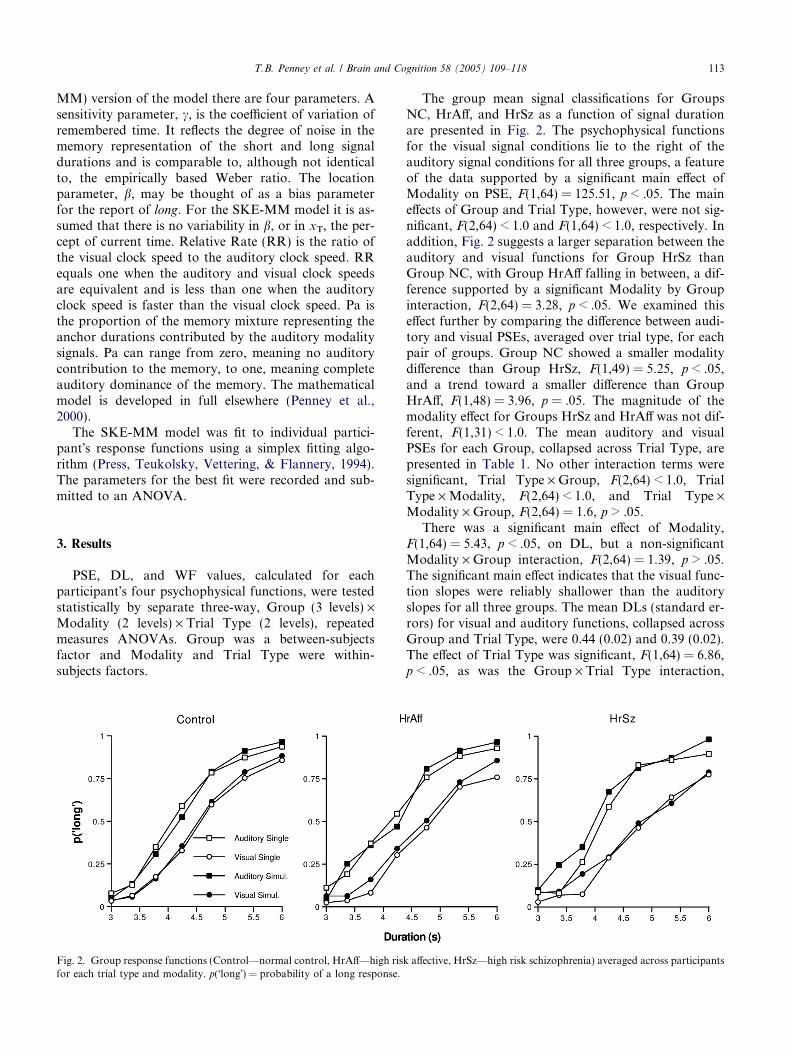
The group mean signal classifications for GroupsNC, HrA!, and HrSz as a function of signal durationare presented in Fig. 2. The psychophysical functionsfor the visual signal conditions lie to the right of theauditory signal conditions for all three groups, a featureof the data supported by a significant main e!ect ofModality on PSE, F(1,64) = 125.51, p < .05. The maine!ects of Group and Trial Type, however, were not sig-nificant, F(2,64) < 1.0 and F(1,64) < 1.0, respectively. Inaddition, Fig. 2 suggests a larger separation between theauditory and visual functions for Group HrSz thanGroup NC, with Group HrA! falling in between, a dif-ference supported by a significant Modality by Groupinteraction, F(2,64) = 3.28, p < .05. We examined thise!ect further by comparing the di!erence between audi-tory and visual PSEs, averaged over trial type, for eachpair of groups. Group NC showed a smaller modalitydi!erence than Group HrSz, F(1,49) = 5.25, p < .05,and a trend toward a smaller di!erence than GroupHrA!, F(1,48) = 3.96, p = .05. The magnitude of themodality e!ect for Groups HrSz and HrA! was not dif-ferent, F(1,31) < 1.0. The mean auditory and visualPSEs for each Group, collapsed across Trial Type, arepresented in Table 1. No other interaction terms weresignificant, Trial Type · Group, F(2,64) < 1.0, TrialType · Modality, F(2,64) < 1.0, and Trial Type ·Modality · Group, F(2,64) = 1.6, p > .05.
There was a significant main e!ect of Modality,F(1,64) = 5.43, p < .05, on DL, but a non-significantModality · Group interaction, F(2,64) = 1.39, p > .05.The significant main e!ect indicates that the visual func-tion slopes were reliably shallower than the auditoryslopes for all three groups. The mean DLs (standard er-rors) for visual and auditory functions, collapsed acrossGroup and Trial Type, were 0.44 (0.02) and 0.39 (0.02).The e!ect of Trial Type was significant, F(1,64) = 6.86,p < .05, as was the Group · Trial Type interaction,
Fig. 2. Group response functions (Control—normal control, HrA!—high risk a!ective, HrSz—high risk schizophrenia) averaged across participantsfor each trial type and modality. p("long!) = probability of a long response.
T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118 113
F(2,64) = 4.28, p < .05. We examined the Trial Type ·Group interaction in greater detail by comparing thesingle and simultaneous trial DLs, averaged over modal-ity, separately for each group. Groups NC and HrA!failed to show a significant e!ect of Trial Type on DL,F(1,33) < 1.0, and F(1,15) = 2.44, p > .05, respectively.However, the e!ect of Trial Type was significant forGroup HrSz, F(1,16) = 8.43, p < 0.05. The main e!ectof Group and the Trial Type · Modality and Group ·Trial Type · Modality interactions were not significant,F(2,64) = 2.64, p > .05, F(1,64) < 1.0, and F(2,64) < 1.0,respectively. Group mean DL values for eachTrial Type, collapsed across modality, are presented inTable 1.
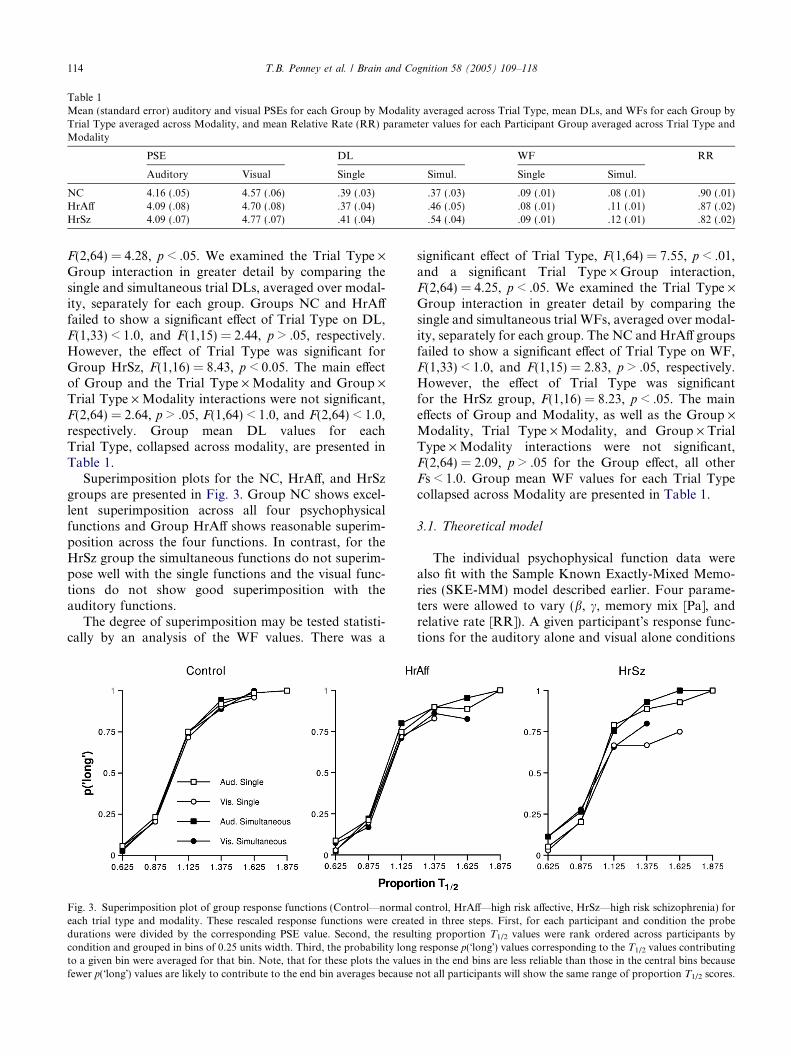
Superimposition plots for the NC, HrA!, and HrSzgroups are presented in Fig. 3. Group NC shows excel-lent superimposition across all four psychophysicalfunctions and Group HrA! shows reasonable superim-position across the four functions. In contrast, for theHrSz group the simultaneous functions do not superim-pose well with the single functions and the visual func-tions do not show good superimposition with theauditory functions.
The degree of superimposition may be tested statisti-cally by an analysis of the WF values. There was a
significant e!ect of Trial Type, F(1,64) = 7.55, p < .01,and a significant Trial Type · Group interaction,F(2,64) = 4.25, p < .05. We examined the Trial Type ·Group interaction in greater detail by comparing thesingle and simultaneous trial WFs, averaged over modal-ity, separately for each group. The NC and HrA! groupsfailed to show a significant e!ect of Trial Type on WF,F(1,33) < 1.0, and F(1,15) = 2.83, p > .05, respectively.However, the e!ect of Trial Type was significantfor the HrSz group, F(1,16) = 8.23, p < .05. The maine!ects of Group and Modality, as well as the Group ·Modality, Trial Type · Modality, and Group · TrialType · Modality interactions were not significant,F(2,64) = 2.09, p > .05 for the Group e!ect, all otherFs < 1.0. Group mean WF values for each Trial Typecollapsed across Modality are presented in Table 1.
3.1. Theoretical model
The individual psychophysical function data werealso fit with the Sample Known Exactly-Mixed Memo-ries (SKE-MM) model described earlier. Four parame-ters were allowed to vary (b, c, memory mix [Pa], andrelative rate [RR]). A given participant!s response func-tions for the auditory alone and visual alone conditions
Table 1Mean (standard error) auditory and visual PSEs for each Group by Modality averaged across Trial Type, mean DLs, and WFs for each Group byTrial Type averaged across Modality, and mean Relative Rate (RR) parameter values for each Participant Group averaged across Trial Type andModality
PSE DL WF RR
Auditory Visual Single Simul. Single Simul.
NC 4.16 (.05) 4.57 (.06) .39 (.03) .37 (.03) .09 (.01) .08 (.01) .90 (.01)HrA! 4.09 (.08) 4.70 (.08) .37 (.04) .46 (.05) .08 (.01) .11 (.01) .87 (.02)HrSz 4.09 (.07) 4.77 (.07) .41 (.04) .54 (.04) .09 (.01) .12 (.01) .82 (.02)
Fig. 3. Superimposition plot of group response functions (Control—normal control, HrA!—high risk a!ective, HrSz—high risk schizophrenia) foreach trial type and modality. These rescaled response functions were created in three steps. First, for each participant and condition the probedurations were divided by the corresponding PSE value. Second, the resulting proportion T1/2 values were rank ordered across participants bycondition and grouped in bins of 0.25 units width. Third, the probability long response p("long!) values corresponding to the T1/2 values contributingto a given bin were averaged for that bin. Note, that for these plots the values in the end bins are less reliable than those in the central bins becausefewer p("long!) values are likely to contribute to the end bin averages because not all participants will show the same range of proportion T1/2 scores.
114 T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118

were fit concurrently (Fit-type AV-Single). This meansthat a single value for b, c, Pa, and RR was determinedfor both the auditory single and the visual single re-sponse functions of that participant. The auditorysimultaneous and visual simultaneous functions (Fit-type AV-Simultaneous) were also fit concurrently witheach other, but separately from the single functions.As a consequence it was possible for b, c, Pa, and RRto be di!erent for the single and simultaneous condi-tions. However, concurrent fits of this model precludefinding between-modality di!erences in b and c.
The obtained parameter values for b, c, RR, and Pawere submitted to separate 3 · 2 repeated measuresANOVAs with Group as the between-subjects factor(3 levels) and Fit-Type (2 levels) as the within-subjectsfactor.
There was a significant e!ect of Group on RelativeRate (RR), F(2,64) = 8.11, p < .05. The main e!ect ofFit Type, F(1,64) < 1.0, and the Group · Fit Type inter-action, F(2,64) = 1.15, p > .05, were not significant. AFisher!s Protected LSD post-hoc analysis of the Groupe!ect indicated that Group HrSz was significantly di!er-ent from both the NC group and the HrA! group atp < .05. As RR is a measure of the clock speed di!eren-tial between the auditory and visual modalities, thismeans there was a larger di!erence between auditoryand visual clock speed for the HrSz group than therewas for the NC or the HrA! groups. The group meanRR values collapsed over Fit Type are presented inTable 1.
There was a significant e!ect of Fit Type on b,F(1,64) = 5.73, p < .05. The main e!ect of Group andthe Fit Type · Group interaction were not significant,both Fs(2,64) < 1.0. The average b value collapsedacross Groups was higher for the AV-Simultaneous fits(M = 1.0, SE = .02) than the AV-Single fits (M = .93,SE = .03). Participants were less biased (1.0 equals nobias) toward short reports on simultaneous trials thanthey were on single trials.
There were no significant e!ects of any factor orinteraction of factors for c, Group, F(2,64) = 1.70,p > .05, Fit Type F(1,64) < 1.0, Fit Type · GroupF(2,64) = 1.51, p > .05, or Pa, Group F(2,64) < 1.0, FitType F(1,64) < 1.0, Fit Type · Group F(2,64) = 2.53,p > .05. This means the degree of memory variabilityand the degree of memory mixing was e!ectivelyequivalent for the di!erent Participant Groups and FitTypes.
4. Discussion
A di!erence between auditory and visual signal clas-sification is a robust finding in duration bisection whentwo signal modalities are presented in the same testsession (Lustig & Meck, 2001; Meck, 1991, 2003; Penney
et al., 2000). We have previously argued (Penney, 2003;Penney et al., 2000) that visual signals are classified asshorter than equivalent duration auditory signals be-cause auditory signals are associated with a faster clockspeed than are visual signals, and both auditory and vi-sual temporal accumulations contribute to non-modali-ty-specific memory representations of the short and longdurations (auditory and visual memory mixing). Bothslow-clock visual accumulations and fast-clock auditoryaccumulations are compared to these common memoryrepresentations with the result that visual signals seemshorter than equivalent duration auditory signals. Note,however, that if a fast-clock auditory accumulation iscompared to a fast-clock auditory memory representa-tion and a slow-clock visual accumulation is comparedwith a slow-clock visual memory representation, thenthe auditory and visual response functions, at least interms of PSE, do not di!er.
Such a clock rate di!erence between auditory and vi-sual signals may be a function of a mode-switch e!ect inthat the amount of attention allocated to the timing sig-nal may determine the rate of pulse accumulation. Un-der full attention, the mode switch governing temporalaccumulation closes and remains closed for the entireduration of the signal (see Fig. 1). Under conditions ofless than full attention, however, the mode switch mayoscillate or flicker between a closed and opened stateresulting in a slower rate of pulse accumulation. If lessattention is allocated to mode-switch closure during vi-sual signals relative to auditory signals, the clock ratefor visual signals will be e!ectively smaller than thatfor auditory signals (Penney, 2003; Penney et al., 2000).
The larger di!erence between auditory and visualPSE!s for the HrSz group than for the NC and HrA!groups suggests there was a greater auditory/visualclock speed di!erence for the HrSz participants thanfor the NC or HrA! participants. The HrSz participantsmay have a slower visual temporal accumulation thannormal because they are less able to maintain the modeswitch in a closed state, indicating an attentional dys-function. Shallower visual than auditory slopes for eachParticipant Group also suggest a clock speed di!erence.Everything else being equal, a temporal accumulationgenerated with a slow clock has less precision and great-er variability than one generated with a fast clock. In theduration-bisection procedure, increased variability is re-vealed by the slope of the psychophysical function and,within Scalar Timing Theory, shallower slopes are fre-quently interpreted as attentional in origin Meck (1991).
If a flickering mode switch is the mechanism mediat-ing the modality and group e!ects, one might expect tosee these di!erences exacerbated on simultaneous trialsbecause the participant has to keep two mode switchesclosed while maintaining two pulse accumulations. In-deed, earlier work with normal participants has demon-strated that timing accuracy can be a!ected by cognitive
T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118 115

variables such as manipulations of attention (Fortin,2003). For example, in dual-task paradigms that useda non-timing task as the secondary task, perceived dura-tion was shortened by reductions in the amount of atten-tional resources allocated to the timing task (Brown,1985; Macar, Grondin, & Casini, 1994; Sawyer, Meyers,& Huser, 1994). Of particular relevance here is a studyby Brown and West (1990) in which participants timedone or more signals simultaneously. Temporal repro-duction accuracy decreased as a function of signal num-ber, but the e!ect was non-directional as there were bothover and underestimations of duration.
The absence in the present study of a main e!ect orinteraction involving Trial Type on the PSE suggeststhat all participants were equally proficient on both sin-gle and simultaneous trials. However, the significantinteraction between Group and Trial Type for the DLmeasure, due to larger simultaneous than single DLsfor the HrSz group, but not for the NC group, indicatesthat the HrSz participants had some di"culty processingtwo signals simultaneously. It is likely that this di"cultywas a direct consequence of the need to maintain twomode switches in a closed state rather than a problemin monitoring for the onset of a second signal given thatparticipants were essentially performing a divided atten-tion task even on single-signal trials. This interpretationis justifiable because single and simultaneous trial typeswere randomly mixed within a test session, e!ectivelyrequiring participants to monitor for the presentationof a second signal even when no such signal was pre-sented (see Fortin, 2003).
The superimposition of psychophysical functions isalso diagnostic of the nature of the modality e!ect fortemporal processing shown by all groups as well as thetrial type e!ect shown by the participants in the HrSzgroup. The NC and HrA! groups showed superimposi-tion of all psychophysical functions. This means that thedi!erence between the auditory and visual psychophysi-cal functions is multiplicative (i.e., the di!erence is aconstant proportion of the probe duration.). If the dif-ference were additive (i.e., the di!erence is an absoluteamount for each probe duration) the functions wouldshow a systematic failure of superimposition. The multi-plicative nature of the di!erence is consistent with amode switch that flickers at a constant rate throughoutsignal presentation. Longer signals lose more time as afunction of the switch being in the open state more fre-quently. Graphically, superimposition appeared to failfor the HrSz group both between-trial types and be-tween modalities (Fig. 3). The WF analysis supports thisconclusion for trial type, but not modality. Moreover,the absence of between-trial type superimposition indi-cates that the trial-type di!erence for these participantshas an additional feature. For the participants in theHrSz group, monitoring two signals a!ects initial switchclosure when signal onset occurs, in addition to a!ecting
maintenance of the switch in the closed state during sig-nal presentation. The di"culty these participants have ininitiating timing of simultaneous signals means an addi-tive di!erence between single and simultaneous signals iscombined with the multiplicative di!erence, resulting inpoor superimposition for the single and simultaneous re-sponse functions. The di"culty in initial switch closureis similar to delays in switch closure that we have foundusing the prior-entry method (Meck, 1984; Penney,Holder, & Meck, 1996).
The application of a theoretical account of intervaltiming (e.g., Scalar Timing Theory—Gibbon et al.,1984) allows for a more rigorous interpretation of di!er-ences in psychophysical functions than does the analysisof simple descriptive measures such as PSE, DL, andWF. A RR parameter value less than 1.0 indicates thatvisual clock speed is less than auditory clock speed. Allgroups showed this clock speed di!erence, but to vary-ing extents. The smaller RR parameter values for theHrSz group suggests that these participants had a largerdiscrepancy between auditory and visual clock ratesthan participants in the NC and HrA! groups.Although the theory indicates that the overall modalitye!ect is due to visual clock speed being slower than audi-tory clock speed, it does not specify whether the largerdiscrepancy exhibited by participants in the HrSz groupis due to a slower than normal visual clock speed or afaster than normal auditory clock speed. It is parsimoni-ous to assign the e!ect to a slower than normal visualclock speed, as that interpretation is consistent withthe PSE and DL results.
As described above, patients with Sz are usually ob-served to overestimate event durations when performingverbal estimation tasks and to underproduce event dura-tions when performing production or reproductiontasks. These studies have been interpreted as indicativeof increased clock rate in Sz participants, an interpreta-tion that has been assumed consistent with animal stud-ies of the e!ects of DA agonists on timing (Rammsayer,1990). However, within the framework provided bypacemaker-accumulator models of interval timing, suchas that illustrated in Fig. 1, a faster than normal clockspeed results in a shift in an estimate of duration onlywhen the experience of time on the test trial is measuredwith a faster clock than that used to measure the refer-ence duration (Meck, 1996). For example, in a repro-duction task, if the reference (target) duration, e.g.10 s, is measured with a relatively slower clock speedand the value stored, and on the test trial a faster clockis used, then the target temporal accumulation obtainsin less than 10 s and elicits an early response (underpro-duction). Given the same contingencies in a verbal esti-mation task, the 10 s signal seems longer than it really isand it is overestimated. Hence the findings for patientswith Sz might be interpreted as due to di!erential clockspeeds for reference and comparison durations. Note,
116 T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118

however, that if the fast clock speed is used both whenthe reference duration is encoded and when the compar-ison duration is experienced, then signal classificationsare accurate (i.e., neither under nor over estimationsoccur).
An alternative possibility is that the overestimationsand underproductions are a consequence of systematicmemory distortions. In this case, if the reference (target)duration is systematically distorted short when it isstored in memory, then the comparison duration accu-mulated during the test trial is larger than the referencememory of what was, in reality, exactly the same objec-tive duration. Therefore, durations are overestimated onverbal estimation tasks because the current accumula-tion is larger than some comparison duration andunderproduced on reproduction tasks because the sub-jective target duration, erroneously distorted short inmemory, is accumulated earlier than the objective targetduration. Moreover, the participant will not be able tocorrect the error even when given feedback becausethe new, ‘‘correct’’, accumulation will also be distortedshort and the same overestimation or underproductionwill occur on subsequent trials. Therefore, it is possiblethat any timing dysfunctions observed in diagnosed Szpatients may be due to disruption of frontal lobestructures involved in memory storage and retrieval.Interestingly, Volz and colleagues recently reported hyp-oactivity of the prefrontal cortex in participants with Sz,as compared to normal controls, when completing a tim-ing task (Volz et al., 2001).
Moreover, this memory distortion account also re-ceives some support from work by Malapani and col-leagues, who, as noted above, demonstrated dissociablememory storage and retrieval distortions in a durationreproduction task by manipulating dopamine levels inPD patients (Malapani et al., 1998, 2002). Perhaps defi-cits in dopamine balance in patients with Sz, caused eitherby the disease state itself or its treatment with anti-psy-chotic drugs, lead to temporal memory dysfunction.
In the present study with HrSz participants, however,we did not obtain systematic overestimations of dura-tion that require a memory dysfunction explanation.Although the HrSz visual functions were shifted fartherto the right than those for the NC and HrA! partici-pants, the HrSz auditory function PSEs were equal tothose of the NC and HrA! participants. Moreover,our interpretation of the observed interval-timing defi-cits for HrSz participants does not require a faster over-all clock rate for the HrSz participants—although alarger relative di!erence between auditory and visualclock rates is prescribed. Given that a number of studieshave found attentional deficits in both Sz patients andHrSZ individuals (e.g., Erlenmeyer-Kimling & Cornbl-att, 1992; Erlenmeyer-Kimling, Cornblatt, & Fleiss,1979; Mirsky, Ingraham, & Kugelmass, 1995; Nuechter-lein, 1977), the more parsimonious explanation for our
results may be to interpret the increased auditory/visualdi!erence in clock speed for HrSz participants as result-ing from an attentional e!ect operating at the level ofthe mode switch which gates pacemaker pulses into anaccumulator.
References
Allan, L. G., & Gibbon, J. (1991). Human bisection at the geometricmean. Learning and Motivation, 22, 39–58.
Artieda, J., Pastor, M. A., Lacruz, F., & Obeso, J. A. (1992). Temporaldiscrimination is abnormal in Parkinson!s disease. Brain, 115,199–210.
Brown, S. W. (1985). Time perception and attention: The e!ects ofprospective versus retrospective paradigms and task demands onperceived duration. Perception & Psychophysics, 38, 115–124.
Brown, S. W., & West, A. N. (1990). Multiple timing and theallocation of attention. Acta Psychologica, 75, 102–121.
Cannon, T. D. (1996). Abnormalities of brain structure and function inschizophrenia: Implications for aetiology and pathophysiology.Annals of Medicine, 28, 533–539.
Church, R. M. (2003). A concise introduction to scalar timing theory.In W. H. Meck (Ed.), Functional and neural mechanisms of intervaltiming (pp. 3–22). Boca Raton, FL: CRC Press..
Clausen, J. (1950). An evaluation of experimental methods of timejudgment. Journal of Experimental Psychology, 40, 76–761.
Densen, M. E. (1977). Time perception and schizophrenia. Perceptualand Motor Skills, 44, 436–438.
Erlenmeyer-Kimling, L. (1996). A look at the evolution of develop-mental models of schizophrenia. In S. Matthysse, D. Levy, J.Kagan, & F. M. Benes (Eds.), Psychopathology: The evolvingscience of mental disorder (pp. 229–252). Cambridge, UK: Cam-bridge University Press.
Erlenmeyer-Kimling, L., & Cornblatt, B. (1987). The New York High-Risk Project: A follow-up report. Schizophrenia Bulletin, 13,451–463.
Erlenmeyer-Kimling, L., & Cornblatt, B. A. (1992). A summary ofattentional findings in the New York High-Risk Project. Journal ofPsychiatry Research, 26, 405–426.
Erlenmeyer-Kimling, L., Cornblatt, B., & Fleiss, J. (1979). High-riskresearch in schizophrenia. Psychiatric Annals, 9, 79–111.
Fortin, C. (2003). Attentional time-sharing in interval timing. In W. H.Meck (Ed.), Functional and neural mechanisms of interval timing(pp. 235–260). Boca Raton, FL: CRC Press.
Gibbon, J. (1977). Scalar expectancy theory and Weber!s Law inanimal timing. Psychological Review, 84, 279–325.
Gibbon, J. (1981). On the form and location of the psychometricbisection function for time. Journal of Mathematical Psychology,24, 58–87.
Gibbon, J. (1986). The structure of subjective time: How time flies. InG. Bower (Ed.), The psychology of learning and motivation(pp. 105–135). New York: Academic Press.
Gibbon, J. (1991). Origins of scalar timing. Learning and Motivation,22, 3–38.
Gibbon, J., & Church, R. M. (1984). Sources of variance in aninformation processing theory of timing. In H. L. Roitblat, T. G.Bever, & H. S. Terrace (Eds.), Animal cognition (pp. 465–488).Hillsdale, NJ: Lawrence Erlbaum Associates.
Gibbon, J., Church, R. M., & Meck, W. H. (1984). Scalar timing inmemory. In J. Gibbon & L. Allan (Eds.), Timing and timeperception (pp. 58–87). New York: Academy of Sciences.
Gibbon, J., & Fairhurst, S. (1994). Ratio versus di!erence comparatorsin choice. Journal of the Experimental Analysis of Behavior, 62,409–434.
T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118 117

Gottesman, I. I., McGu"n, P., & Farmer, A. E. (1987). Clinicalgenetics as clues to the real genetics of schizophrenia. A decade ofmodest gains while playing for time. Schizophrenia Bulletin, 13,23–47.
Gottesman, I. I., Shields, J., & Hanson, D. R. (1982). Schizophrenia:The epigenetic puzzle. Cambridge, England: Cambridge UniversityPress.
Holzman, P. S., & Matthysse, S. (1990). The Genetics of schizophre-nia: A review. Psychological Science, 1, 279–286.
Johnson, J. E., & Petzel, T. P. (1971). Temporal orientation and timeestimation in chronic schizophrenics. Journal of Clinical Psychol-ogy, 27, 194–196.
Kendler, K. S. (1988). The genetics of schizophrenia and relateddisorders. In D. L. Dunner, E. S. Gershon, & J. E. Barrett (Eds.),Relatives at risk for mental disorder. New York: Raven Press.
Lhamon, L. T., & Goldstone, S. (1956). The time sense: Estimation ofone second durations by schizophrenic patients. Archives ofNeurology & Psychiatry, 76, 625–629.
Lustig, C., & Meck, W. H. (2001). Paying attention to time as one getsolder. Psychological Science, 12, 478–484.
Lustig, C., & Meck, W. H. (2005). Chronic treatment with haloperidolinduces deficits in feedback e!ects of interval timing. Brain andCognition, in press.
Macar, F., Grondin, S., & Casini, L. (1994). Controlled attentionsharing influences time estimation. Memory & Cognition, 22,673–686.
Malapani, C., Rakitin, B., Levy, R., Meck, W. H., Deweer, B.,Dubois, B., et al. (1998). Coupled temporal memories in Parkin-son!s disease: A dopamine related dysfunction. Journal of CognitiveNeuroscience, 10, 316–331.
Malapani, C., Deweer, B., & Gibbon, J. (2002). Separating storagefrom retrieval dysfunction of temporal memory in Parkinson!sdisease. Journal of Cognitive Neuroscience, 14, 311–322.
Meck, W. H. (1984). Attentional bias between modalities: E!ects onthe internal clock, memory, and decision stages used in animal timediscrimination. In J. Gibbon & L. Allan (Eds.), Timing and timeperception (pp. 528–541). New York: New York Academy ofSciences.
Meck, W. H. (1986). A"nity for the dopamine D2 receptor predictsneuroleptic potency in decreasing the speed of an internalclock. Pharmacology, Biochemistry & Behavior, 25, 1185–1189.
Meck, W. H. (1991). Modality-specific circadian rhythmicities influ-ence mechanisms of attention and memory for interval timing.Learning and Motivation, 22, 153–179.
Meck, W. H. (1996). Neuropharmacology of timing and timeperception. Cognitive Brain Research, 3, 227–242.
Meck, W. H., 2003. Introduction: Persistence of time. In W. H. Meck,(Ed.), Functional and neural mechanisms of interval timing (pp. xvii–xli). Boca Raton, FL: CRC Press.
Meck, W. H., & Benson, A. M. (2002). Dissecting the brain!s internalclock: How frontal-striatal circuitry keeps time and shifts attention.Brain and Cognition, 48, 195–211.
Mirsky, A. F., Ingraham, L. J., & Kugelmass, S. (1995). Neuropsy-chological assessment of attention and its pathology in the Israelicohort. Schizophrenia Bulletin, 21, 193–204.
Nuechterlein, K. H. (1977). Refocusing on attentional dysfunctions inschizophrenia. Schizophrenia Bulletin, 3, 457–468.
Pastor, M. A., Artieda, J., Jahanshahi, M., & Obeso, J. A. (1992).Time estimation and reproduction is abnormal in Parkinson!sdisease. Brain, 115, 211–225.
Penney, T. B. (2003). Modality di!erences in interval timing: Atten-tion, clock speed, and memory. In W. H. Meck (Ed.), Functionaland neural mechanisms of interval timing (pp. 209–233). BocaRaton, FL: CRC Press.
Penney, T. B., Allan, L., Meck, W. H., & Gibbon, J. (1998). Memorymixing in duration bisection. In D. A. Rosenbaum & C. E. Collyer(Eds.), Timing of behavior: Neural, computational, and psychologicalperspectives (pp. 165–193). Cambridge, MA: MIT Press.
Penney, T. B., Gibbon, J., & Meck, W. H. (2000). Di!erential e!ects ofauditory and visual signals on clock speed and temporal memory.Journal of Experimental Psychology: Human Perception and Per-formance, 26, 1770–1787.
Penney, T. B., Holder, M. D., & Meck, W. H. (1996). Clonidine-induced antagonism of norepinephrine modulates the attentionalprocesses involved in peak-interval timing. Experimental andClinical Psychopharmacology, 4, 82–92.
Press, W. H., Teukolsky, S. A., Vettering, W. T., & Flannery, B. P.(1994). Numerical Recipes in C: The art of scientific computing (2nded.). Cambridge: Cambridge University Press.
Rammsayer, T. (1990). Temporal discrimination in schizophrenic anda!ective disorders: Evidence for a dopamine-dependent internalclock. International Journal of Neuroscience, 53, 111–120.
Sawyer, T. F., Meyers, P. J., & Huser, S. J. (1994). Contrasting taskdemands alter the perceived duration of brief time intervals.Perception & Psychophysics, 56, 649–657.
Seeman, P. (1992). Dopamine receptor sequences: Therapeutic levels ofneuroleptics occupy D2 receptors, clozapine occupies D4. Neuro-psychopharmacology, 7, 261–284.
Seeman, P. (1995). Dopamine receptors: Clinical correlates. In F. E.Bloom&D. J.Kupfer et al. (Eds.),Psychopharmacology: The fourthgeneration of progress: An o!cial publication of the american collegeof psychopharmacology (pp. 295–302). New York: Raven Press.
Tysk, L. (1983). Estimation of time and the subclassification ofschizophrenic disorders. Perceptual and Motor Skills, 57, 911–918.
Tysk, L. (1990). Estimation of time by patients with positive andnegative schizophrenia. Perceptual and Motor Skills, 71, 826.
Volz, H. P., Nenadic, I., Gaser, C., Rammsayer, T., Hager, F., &Sauer, H. (2001). Time estimation in schizophrenia: An fMRIstudy at adjusted levels of di"culty. NeuroReport, 12, 313–316.
Wahl, O. F., & Sieg, D. (1980). Time estimation among schizophrenics.Perceptual and Motor Skills, 50, 535–541.
Weinstein, A. D., Goldstone, S., & Boardman, W. R. (1958). The e!ectof recent and remote frames of reference on temporal judgments ofschizophrenic patients. Journal of Abnormal and Social Psychology,57, 241–244.
118 T.B. Penney et al. / Brain and Cognition 58 (2005) 109–118
Related Documents











