Downloaded from www.microbiologyresearch.org by IP: 54.144.197.59 On: Thu, 21 Apr 2016 12:21:25 Internalization of a thiazole-modified peptide in Sinorhizobium meliloti occurs by BacA-dependent and -independent mechanisms Silvia Wehmeier, 1 3 Markus F. F. Arnold, 1 3 Victoria L. Marlow, 1 Mustapha Aouida, 2 Kamila K. Myka, 1 Vivien Fletcher, 1 Monica Benincasa, 3 Marco Scocchi, 3 Dindial Ramotar 2 and Gail P. Ferguson 1 Correspondence Gail P. Ferguson [email protected] Received 10 March 2010 Revised 17 May 2010 Accepted 24 May 2010 1 School of Medicine and Dentistry, Institute of Medical Sciences, University of Aberdeen, Foresterhill, Aberdeen AB25 2ZD, UK 2 Maisonneuve-Rosemont Hospital Research Centre, University of Montreal, 5415, Boul. de l’Assomption, Montreal, QC H1T 2M4, Canada 3 Department of Life Sciences, University of Trieste, Via Giorgieri 1, 34127 Trieste, Italy BacA proteins play key roles in the chronic intracellular infections of Sinorhizobium meliloti, Brucella abortus and Mycobacterium tuberculosis within their respective hosts. S. meliloti, B. abortus and M. tuberculosis BacA-deficient mutants have increased resistance to the thiazole- modified peptide bleomycin. BacA has been previously hypothesized, but not experimentally verified, to be involved in bleomycin uptake. In this paper, we show that a BacA-dependent mechanism is the major route of bleomycin internalization in S. meliloti. We also determined that the B. abortus and S. meliloti BacA proteins are functional homologues and that the B. abortus BacA protein is involved in the uptake of both bleomycin and proline-rich peptides. Our findings also provide evidence that there is a second, BacA-independent minor mechanism for bleomycin internalization in S. meliloti. We determined that the BacA-dependent and -independent mechanisms of bleomycin uptake are energy-dependent, consistent with both mechanisms of bleomycin uptake involving transport systems. INTRODUCTION Sinorhizobium meliloti can be found either free-living in the soil or in a symbiotic relationship with leguminous plants such as alfalfa (for recent reviews refer to Gibson et al., 2008; Jones et al., 2007). After entry into alfalfa cells, S. meliloti differentiates into a nitrogen-fixing bacteroid, which persists for extensive periods. Despite being bene- ficial for the host, this interaction can be viewed as a chronic intracellular infection (Gibson et al., 2008). The BacA protein is essential for the chronic intracellular infection of S. meliloti within alfalfa (Glazebrook et al., 1993), as in the absence of BacA, an S. meliloti mutant can enter into the plant cell but lyses and dies shortly after entry. The chronic mammalian pathogen Brucella abortus also has a BacA protein, which shares 68 % identity with the S. meliloti BacA protein and is essential for chronic infections of BALB/c mice (LeVier et al., 2000). Recently, the Mycobacterium tuberculosis BacA protein (Rv1819c), which is 39 % similar to the B. abortus BacA protein, was also determined to be involved in the maintenance of chronic B6D2/F1 mice infections (Domenech et al., 2009). Since the mechanism of chronic bacterial infections is poorly understood, elucidation of the function of different BacA proteins could reveal important insights into this process. To understand more about the function of BacA proteins, S. meliloti, B. abortus and M. tuberculosis bacA mutants have also been characterized in their free-living states relative to their respective parent strains. The lipid A of S. meliloti and B. abortus is unusually modified with very- long-chain fatty acids (VLCFAs), and it was discovered that S. meliloti and B. abortus bacA null mutants have an approximately 50 % reduction in their lipid A VLCFA content (Ferguson et al., 2004). Lipid A is a component of the lipopolysaccharide that forms the outer leaflet of the outer membrane of Gram-negative bacteria (Raetz & Whitfield, 2002). However, by constructing and character- izing S. meliloti acpXL and lpxXL mutants that are deficient in the biosynthesis of the lipid A VLCFA modification, it Abbreviations: BY, BODIPY FL-N-(2-aminorthyl)maleimide; DAPI, 49,69- diamidino-2-phenylindole; DNP, dinitrophenol; fluoro-BLM-A5, fluores- cently labelled bleomycin A5; MMS, methyl methane sulfonate; NCR peptides, nodule-specific cysteine-rich peptides; TMD, transmembrane domain; VLCFA, very-long-chain fatty acid. 3These authors contributed equally to this work. Microbiology (2010), 156, 2702–2713 DOI 10.1099/mic.0.039909-0 2702 039909 G 2010 SGM Printed in Great Britain

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
Internalization of a thiazole-modified peptide inSinorhizobium meliloti occurs by BacA-dependentand -independent mechanisms
Silvia Wehmeier,13 Markus F. F. Arnold,13 Victoria L. Marlow,1
Mustapha Aouida,2 Kamila K. Myka,1 Vivien Fletcher,1 Monica Benincasa,3
Marco Scocchi,3 Dindial Ramotar2 and Gail P. Ferguson1
Correspondence
Gail P. Ferguson
Received 10 March 2010
Revised 17 May 2010
Accepted 24 May 2010
1School of Medicine and Dentistry, Institute of Medical Sciences, University of Aberdeen,Foresterhill, Aberdeen AB25 2ZD, UK
2Maisonneuve-Rosemont Hospital Research Centre, University of Montreal, 5415, Boul. del’Assomption, Montreal, QC H1T 2M4, Canada
3Department of Life Sciences, University of Trieste, Via Giorgieri 1, 34127 Trieste, Italy
BacA proteins play key roles in the chronic intracellular infections of Sinorhizobium meliloti,
Brucella abortus and Mycobacterium tuberculosis within their respective hosts. S. meliloti, B.
abortus and M. tuberculosis BacA-deficient mutants have increased resistance to the thiazole-
modified peptide bleomycin. BacA has been previously hypothesized, but not experimentally
verified, to be involved in bleomycin uptake. In this paper, we show that a BacA-dependent
mechanism is the major route of bleomycin internalization in S. meliloti. We also determined that
the B. abortus and S. meliloti BacA proteins are functional homologues and that the B. abortus
BacA protein is involved in the uptake of both bleomycin and proline-rich peptides. Our findings
also provide evidence that there is a second, BacA-independent minor mechanism for bleomycin
internalization in S. meliloti. We determined that the BacA-dependent and -independent
mechanisms of bleomycin uptake are energy-dependent, consistent with both mechanisms of
bleomycin uptake involving transport systems.
INTRODUCTION
Sinorhizobium meliloti can be found either free-living in thesoil or in a symbiotic relationship with leguminous plantssuch as alfalfa (for recent reviews refer to Gibson et al.,2008; Jones et al., 2007). After entry into alfalfa cells, S.meliloti differentiates into a nitrogen-fixing bacteroid,which persists for extensive periods. Despite being bene-ficial for the host, this interaction can be viewed as achronic intracellular infection (Gibson et al., 2008). TheBacA protein is essential for the chronic intracellularinfection of S. meliloti within alfalfa (Glazebrook et al.,1993), as in the absence of BacA, an S. meliloti mutant canenter into the plant cell but lyses and dies shortly afterentry. The chronic mammalian pathogen Brucella abortusalso has a BacA protein, which shares 68 % identity withthe S. meliloti BacA protein and is essential for chronic
infections of BALB/c mice (LeVier et al., 2000). Recently,the Mycobacterium tuberculosis BacA protein (Rv1819c),which is 39 % similar to the B. abortus BacA protein, wasalso determined to be involved in the maintenance ofchronic B6D2/F1 mice infections (Domenech et al., 2009).Since the mechanism of chronic bacterial infections ispoorly understood, elucidation of the function of differentBacA proteins could reveal important insights into thisprocess.
To understand more about the function of BacA proteins,S. meliloti, B. abortus and M. tuberculosis bacA mutantshave also been characterized in their free-living statesrelative to their respective parent strains. The lipid A of S.meliloti and B. abortus is unusually modified with very-long-chain fatty acids (VLCFAs), and it was discovered thatS. meliloti and B. abortus bacA null mutants have anapproximately 50 % reduction in their lipid A VLCFAcontent (Ferguson et al., 2004). Lipid A is a component ofthe lipopolysaccharide that forms the outer leaflet of theouter membrane of Gram-negative bacteria (Raetz &Whitfield, 2002). However, by constructing and character-izing S. meliloti acpXL and lpxXL mutants that are deficientin the biosynthesis of the lipid A VLCFA modification, it
Abbreviations: BY, BODIPY FL-N-(2-aminorthyl)maleimide; DAPI, 49,69-diamidino-2-phenylindole; DNP, dinitrophenol; fluoro-BLM-A5, fluores-cently labelled bleomycin A5; MMS, methyl methane sulfonate; NCRpeptides, nodule-specific cysteine-rich peptides; TMD, transmembranedomain; VLCFA, very-long-chain fatty acid.
3These authors contributed equally to this work.
Microbiology (2010), 156, 2702–2713 DOI 10.1099/mic.0.039909-0
2702 039909 G 2010 SGM Printed in Great Britain

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
was found that the unusual modification of the lipid A isimportant but not essential for the chronic infection of S.meliloti in alfalfa (Ferguson et al., 2005; Haag et al., 2009;Sharypova et al., 2003). Additionally, these studies demon-strated that the effect of BacA on the lipid A VLCFAmodification is unlikely solely to account for its essentialrole in chronic infections. Consistent with this, the BacAprotein of M. tuberculosis had no effect on the fatty acidcontent of membrane lipids (Domenech et al., 2009).Combined, these studies provide support for a modelwhereby BacA proteins also have another function or effecton bacterial cells that is important for their essential role inchronic bacterial infections.
The S. meliloti BacA protein also shares 64 % identity withthe Escherichia coli SbmA protein (Glazebrook et al., 1993;Ichige & Walker, 1997). The sbmA gene was first identifiedby its ability to confer sensitivity of E. coli to theantimicrobial peptide microcin B17 (Lavina et al., 1986;Yorgey et al., 1994). Microcin B17 is a glycine-rich,thiazole-modified peptide produced by E. coli strainscarrying a naturally occurring plasmid and is an inhibitorof DNA gyrase (Yorgey et al., 1994). Since mutations insbmA only protected E. coli against exogenous microcinB17 and conferred no growth advantage to the microcinB17-producing strain, this provided indirect evidence thatSbmA is involved in peptide uptake (Lavina et al., 1986). Itwas subsequently shown, using fluorescently labelledpeptides, that the SbmA protein is involved in the uptakeof truncated Bac7 peptides into E. coli (Mattiuzzo et al.,2007). Full-length Bac7 is produced by bovine leukocytesand is a linear proline/arginine-rich peptide (Frank et al.,1990). At present, it is not known whether SbmA plays adirect or indirect role in peptide uptake in E. coli(Mattiuzzo et al., 2007). However, it has been proposedthat SbmA and BacA proteins form the transmembranedomains of an ABC transporter and the M. tuberculosisBacA protein has a fused ATPase domain (Domenech et al.,2009; LeVier & Walker, 2001), suggesting that they couldbe directly involved in peptide uptake.
S. meliloti is highly resistant to killing by microcin B17(Ichige & Walker, 1997). However, S. meliloti is sensitive totruncated Bac7 peptides, and the BacA protein has beenshown to play an essential role in their uptake (Marlowet al., 2009). In contrast, there appear to have been noprevious studies investigating the role of the B. abortus andM. tuberculosis BacA proteins in peptide uptake. Neverthe-less, every sbmA/bacA mutant characterized to date showsincreased resistance to the glycopeptide antibiotic bleomy-cin (Domenech et al., 2009; Ichige & Walker, 1997; LeVieret al., 2000). In S. meliloti, it was shown that reductions inthe lipid A VLCFA do not confer resistance to bleomycin,providing evidence that the increased resistance of the S.meliloti bacA null mutant to bleomycin is independent ofits lipid A alteration (Ferguson et al., 2006). Bleomycin issynthesized by the soil bacterium Streptomyces verticillus(Umezawa et al., 1966) and has been shown previouslyto damage DNA through a Fe(II)-mediated oxidative
mechanism (Kane & Hecht, 1994). It had been proposedpreviously that BacA-mediated sensitivity to bleomycincould be due to BacA being involved in iron uptake (Ichige& Walker, 1997). However, an M. tuberculosis BacA-deficient mutant was unaffected in iron uptake, suggestingthat this was not the case (Domenech et al., 2009).Interestingly, although the peptide sequences of bleomycinand microcin B17 differ, both peptides are modified withthiazole rings (Yorgey et al., 1994). Based on this findingand the increased resistance of bacA/sbmA mutants tobleomycin it was hypothesized that SbmA/BacA proteinscould also be involved in the uptake of bleomycin intobacterial cells (Ichige & Walker, 1997; Yorgey et al., 1994).However, this hypothesis had never been verified experi-mentally. Bleomycin is also known to damage the cell walland membrane of yeast cells (Lim et al., 1995). Therefore,we could not exclude the possibility that BacA/SbmA weresensitizing bacterial cells to bleomycin through anothermechanism rather than uptake.
Since bleomycin is produced by a soil bacterium and plantsproduce thiazole-containing compounds (Umezawa et al.,1966; Yorgey et al., 1994), the effect of bleomycin on S.meliloti could have relevance to both its free-living andsymbiotic states. Additionally, bleomycin is an antibiotic(Umezawa et al., 1966; Yorgey et al., 1994). Therefore,investigating the factors involved in sensitizing B. abortusto this peptide could provide information relevant to thetreatment of chronic bacterial infections. In this study, weinvestigated the hypothesis that BacA proteins are involvedin bleomycin internalization. We found that both the S.meliloti and B. abortus BacA proteins play important rolesin bleomycin internalization in S. meliloti. However, wealso discovered a second, BacA-independent, energy-dependent mechanism for bleomycin internalization in S.meliloti.
METHODS
Bacterial strains and growth conditions. All bacterial strains andplasmids used in this work are described in Table 1. The S. melilotistrains used are all derivatives of the Rm1021 sequenced strain(Galibert et al., 2001). For all experiments, S. meliloti strains weregrown in either Luria–Bertani broth (LB) (Sambrook et al., 1982)prepared with 10 g NaCl l21or LB supplemented with 2.5 mM CaCl2and 2.5 mM MgSO4 (LB/MC) for 48 h at 30 uC. Unless indicatedotherwise, antibiotics were used at the following concentrations (mgml21): gentamicin (Gm), 50; streptomycin (Sm), 500; spectinomycin(Spc), 100; neomycin (Nm) 200; and tetracycline (Tc), 5.
Filter disc and liquid culture viability assays. For the filter discassays, late-exponential-phase cultures (OD600 ~3.0) were washed inLB, resuspended to OD600 ~0.2 and then the assays were performedexactly as described previously (Ferguson et al., 2002). The agar plateswere incubated at 30 uC (72 h) and then the diameters of growthinhibition were recorded. The growth inhibition zone was measuredfrom at least three plates for each strain and condition and the resultswere averaged. The liquid culture viability assays, unless statedotherwise, were also performed with late-exponential-phase cultures,which were harvested, washed and diluted to OD600 ~0.1 in LBmedium. After addition of the appropriate form of bleomycin, the
Internalization of bleomycin in S. meliloti
http://mic.sgmjournals.org 2703

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
cultures were incubated at 30 uC. At defined times, samples were
removed, serially diluted in LB medium, then 10 ml aliquots were
plated in triplicate on LB agar plates. C.f.u. were determined after
72 h at 30 uC. The mean c.f.u. ml21 in the three 10 ml aliquots was
plotted. All experiments were repeated at least twice and the data
shown are a representative dataset.
For the 2,4-dinitrophenol (DNP) treatment, cultures were grown to
mid-exponential phase and then resuspended to OD600 ~0.8 in LB
medium supplemented or not with the defined amount of DNP. After
50 min with shaking (200 r.p.m.) at 30 uC, the cultures were washed
twice with LB and the viability determined as described above. To
determine whether DNP treatment affected sensitivity to bleomycin,
the cultures were diluted to OD600 ~0.1 in LB medium and the
bleomycin sensitivity assay performed as described above.
Genomic DNA degradation assay. Cultures were grown to mid-
exponential phase (OD600 ~0.9) in LB/MC, washed three times with
LB and then recovered by centrifugation at 2264 g (10 min). The cell
pellet was resuspended in 2 ml LB to OD600 ~2, split equally between
two glass test tubes and then one tube was treated with either
bleomycin A5 (20 mg ml21) or the defined amount (1 or 0.2 % v/v) of
methyl methane sulfonate (MMS) (Sigma). The second tube served as
an untreated control. All tubes were incubated with shaking at 30 uCfor 2 h (bleomycin treated) or the defined time (30 min to 2 h) for
MMS; the cells were then washed once, recovered by centrifugation at
2264 g (10 min) and the cell pellets were stored at 280 uC until
required. The genomic DNA was extracted as described previously
(Wilson, 1987) with the following modifications: in step 4 chloroform
only was used, in step 5 phenol/chloroform (1 : 1) was used instead of
phenol/chloroform/isoamyl alcohol (25 : 24 : 1) and in step 7 the
genomic DNA was dissolved in 25 ml H2O. The genomic DNA was
quantified using UV spectroscopy and 2 mg of the DNA was analysed
on a 1 % (w/v) agarose gel.
4§,6§-Diamidino-2-phenylindole (DAPI) fluorescence. Cultures
were treated with and without bleomycin A5 exactly as for the
genomic DNA preparations except that following the centrifugation
the cells were resuspended in 100 ml 1 % (v/v) toluene, vortexed and
stored at 4 uC. When required, the cells were diluted to OD600 ~0.08
in dilution buffer (10 mM NaCl, 6.6 mM Na2SO4, 5 mM HEPES
pH 7.0). The DAPI assay was performed following a previously
described procedure (Johnson, 1994) except that DAPI was added tothe cells at a final concentration of 0.1 mg ml21 in a final volume of100 ml and the cells were incubated with DAPI overnight at 4 uC. The
fluorescence intensity was measured using a FLUOstar optima platereader (BMG Labtech) spectrophotometer with excitation andemission at 350 and 450 nm, respectively.
Cloning of the B. abortus and S. meliloti bacA genes. The B.abortus bacA gene, including 150 bp upstream, was amplified by PCRusing the primers BabacA+150-F (59-GCTTGGCTGCAGCCTAA-CACCCTATG GGCCGT-39) and BabacA-R (59-ATGATGGATCCT-CAGCCCGCCCCTG-39). The fragment was then digested with
BamHI and PstI, and ligated into pRF771 (Wells & Long, 2002),under control of the trp promoter. The S. meliloti bacA gene wasamplified by PCR using the primers Smeliloti_bacA_NsiI_RBS_F (59-GCTAGAATGCATGAAACGAGAGTGCCGTCCCCCTTGTTCCAA-TCCTTCTTCCCC-39) and Smeliloti_bacA_XbaI_R (59-TTATCGT-
CTAGAATGCCCGCCGTTACAGA-39). The PCR fragment was thendigested with NsiI and XbaI and ligated into pRF771 under control ofptrp. In both cases, the ligated plasmids were transformed into E. coliDH5a and transformants were selected on LB Tc agar. The insertionswere then confirmed by amplification of the inserts by PCR using the
plasmid-specific primers pkX-US-F (59-CCGGCTCGTATGTTGTGTGG-39) and pkX-DS-R (59-CGAAAGGGGGATGTGCTGC-39), fol-lowed by sequencing. The correct clones, pBabacA and pSmbacA,were then conjugated into the S. meliloti Rm1021 bacA null mutant(Ferguson et al., 2002) using E. coli MT616 with the helper plasmid
pRK600 (Finan et al., 1986), and selected on LB Sm Tc agar.
S. meliloti–alfalfa interaction experiments. To determine theability of S. meliloti to form a successful symbiosis with alfalfa, 3-day-
old seedlings were inoculated with 1 ml S. meliloti culture,resuspended to OD600 ~0.05 in sterile water, on Jensen’s agar asdescribed previously (Leigh et al., 1985). The plates were incubated at25 uC, and plant growth and nodule morphology were determinedafter 4 weeks.
Bac7 sensitivity and Bac71–16-BY uptake assays. The N-terminalfragments 1–16 and 1–35 of Bac7 were synthesized and prepared asdescribed previously (Benincasa et al., 2004). The Bac7 sensitivityassays were performed using mid-exponential-phase cultures exactlyas described previously (Marlow et al., 2009). A fluorescently labelled
Table 1. Bacterial strains and plasmids used
Strain or plasmid Relevant characteristics Source or reference
S. meliloti
Rm1021 Smr derivative of SU47 Meade et al. (1982)
SmGF1 Rm1021, DbacA654 : : Spcr Ferguson et al. (2002)
E. coli
DH5a supE44 DlacU169 (w80lacZDM15) hsdR17 recA1 endA1 gyrA96 thi-1 relA1 BRL
MT616 MM294A recA56 (pRK600) Cmr Finan et al. (1986)
Plasmids
pRK404 Broad-host-range control plasmid, Tcr Ditta et al. (1985)
pJG51A pRK404 carrying the S. meliloti wild-type bacA gene with upstream region, Tcr Glazebrook et al. (1993)
pRF771 RK2 derivative Ptrp expression vector, Tcr Wells & Long (2002)
pBabacA pRF771 carrying the entire B. abortus bacA gene with 150 bp upstream region, Tcr This study
pSmbacA pRF771 carrying the entire S. meliloti bacA gene with 19 bp upstream region, Tcr This study
Bacteriophage
wM12 Generalized transducing phage for S. meliloti Rm1021 T. Finan*
*Center for Environmental Genomics, Department of Biology, McMaster University, Hamilton, Ontario, Canada.
S. Wehmeier and others
2704 Microbiology 156

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
version of Bac71–16, Bac71–16-BY, was prepared by linkage of the
thiol-reactive dye BODIPY-FL N-(2-aminorthyl)maleimide (BY)(Invitrogen) to an additional C-terminal cysteine residue as reported
previously (Scocchi et al., 2008). The uptake of Bac71–16-BY wasperformed as described previously (Marlow et al., 2009), with
modifications. Mid-exponential-phase cultures of OD600 ~1.0 were
harvested, washed and resuspended in fresh LB medium. After theaddition of 0.5 mM Bac71–16-BY, the cultures were incubated at 30 uCfor 1 h. To account for extracellular binding of Bac71–16-BY, the
cultures were then treated with the extracellular quencher offluorescence, Trypan Blue (TB; 1 mg ml21) for 10 min at room
temperature. The fluorescence of cells was determined on poly-L-
lysine-coated slides using a Carl Zeiss Axioskop microscope (1006objective magnification with UV light and FITC filter). Images were
taken with a Canon Powershot camera and using the AxioVisionversion 2.0 software.
Statistical analysis. Where shown, the significance of differences
among bacterial strains was assessed by the Student’s unpaired t-testusing Microsoft Excel.
RESULTS
BacA sensitizes S. meliloti to different forms ofbleomycin
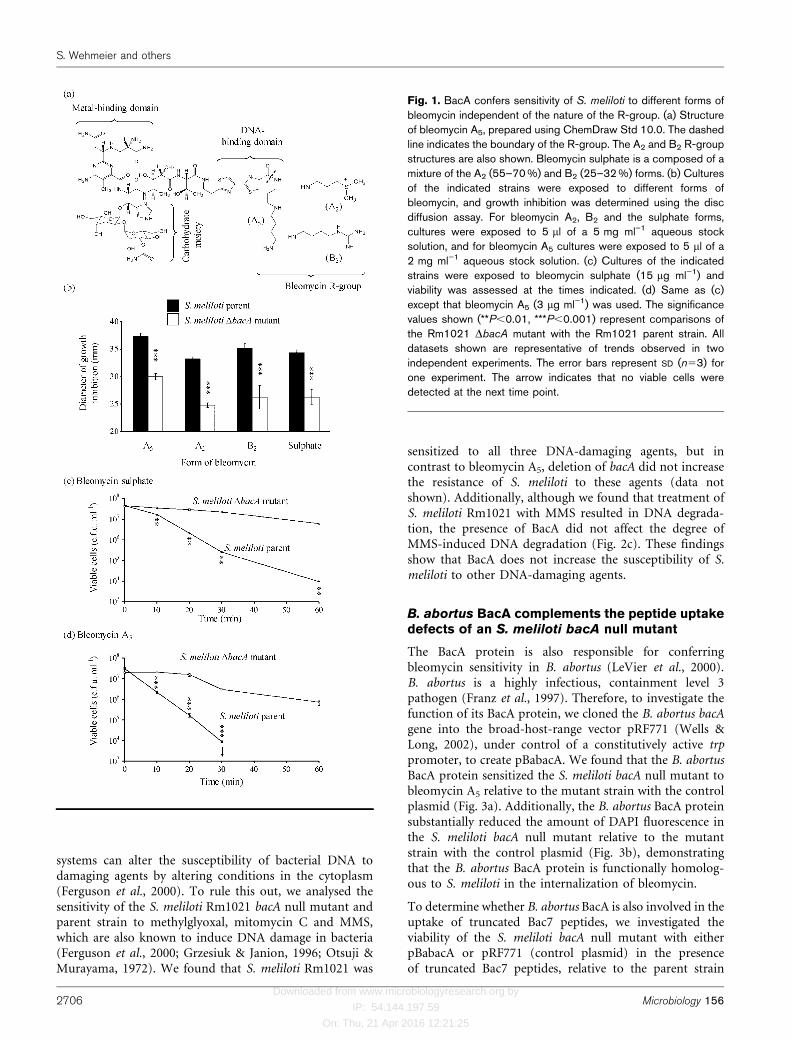
The R-group of bleomycin differs depending upon theform (Fig. 1a). Previous studies investigating the bleomy-cin-resistance phenotypes of S. meliloti Rm1021 bacAmutants have used the A2 form (Ferguson et al., 2006;Ichige & Walker, 1997; LeVier & Walker, 2001). Toinvestigate whether the nature of the R-group affectedBacA-mediated sensitivity of S. meliloti to bleomycin, weperformed disc diffusion assays using different forms ofbleomycin and compared the sensitivity of an S. melilotiRm1021 bacA null mutant and the parent strain (Fig. 1b).We found that the S. meliloti parent strain showed anincreased sensitivity to all the forms of bleomycin testedrelative to the S. meliloti bacA null mutant. We alsodetermined that bleomycin sulphate and bleomycin A5
reduced the viability of S. meliloti Rm1021 in liquid cultureand that deletion of bacA conferred protection against bothforms (Fig. 1c, d). These findings show that BacA sensitizesS. meliloti to different forms of bleomycin, irrespective ofthe nature of the R-group.
BacA is involved in the internalization ofbleomycin in S. meliloti
It has been shown previously by transmission electronmicroscopy (TEM) that bleomycin, in addition to damag-ing the DNA, also damages the cell wall and membrane ofyeast cells (Lim et al., 1995). However, using TEM, we didnot detect any bleomycin-induced lesions in the cellenvelope components of S. meliloti Rm1021 or the bacAnull mutant (data not shown). Additionally, unlikepolymyxin B, which damages both the inner and outermembranes (Benincasa et al., 2009), bleomycin A5 treat-ment did not increase the amount of propidium iodidefluorescence of either the S. meliloti mutant lacking BacA
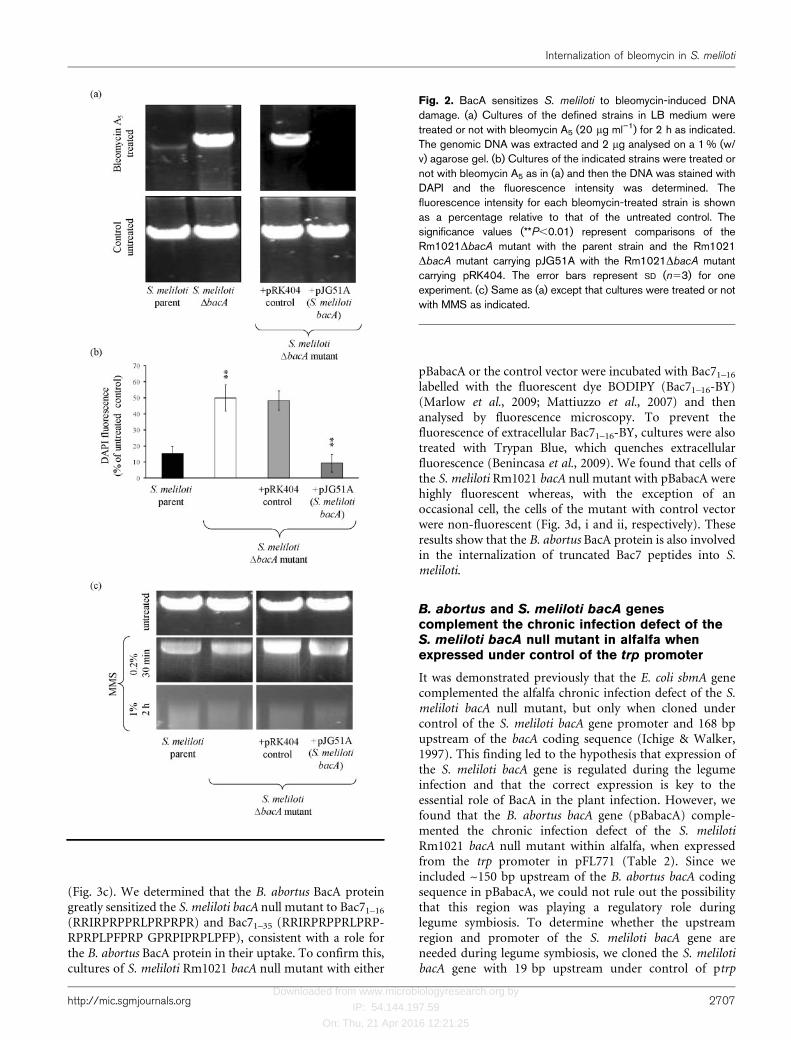
or the parent strain (data not shown). To investigate apotential role for BacA in bleomycin uptake, we preparedfluorescently labelled bleomycin A5 (fluoro-BLM-A5) byconjugating it with FITC (Aouida et al., 2004). The result-ing fluoro-BLM-A5 contained a 1 : 1 ratio of FITC andbleomycin A5 (data not shown). However, unlike withyeast cells (Aouida et al., 2004), we found that theconjugation of FITC to bleomycin A5 substantially reducedits toxicity and prevented uptake into S. meliloti (data notshown). For this reason, we used bleomycin-induced DNAdegradation in the absence of membrane-damaging effectsas an assay to monitor bleomycin A5 internalization in S.meliloti. We found that treatment of an S. meliloti Rm1021culture with bleomycin A5 induced substantial degradationof the genomic DNA relative to the DNA from theuntreated control culture (Fig. 2a), confirming the intracel-lular presence of bleomycin in S. meliloti. In contrast, wedid not observe any significant loss of the genomic DNAafter bleomycin A5 treatment of the S. meliloti Rm1021bacA null mutant relative to the DNA from the untreatedcontrol culture (Fig. 2a). Additionally, the presence ofpJG51A (encoding the S. meliloti wild-type bacA gene inpRK404) but not the control plasmid (pRK404) in the S.meliloti Rm1021 bacA null mutant strain increased theamount of bleomycin-induced DNA degradation relativeto untreated control cultures (Fig. 2a). Taken together,these findings are consistent with a role for BacA in theinternalization of bleomycin, which then leads to DNAdamage and the subsequent degradation of the genomicDNA.
We also quantified the effect of bleomycin A5 treatmenton the S. meliloti parent strain and bacA null mutant insitu by permeabilizing cells and measuring fluorescence ofDAPI, a fluorescent dye that binds DNA (Johnson, 1994).We found a substantial decrease in the amount of DAPIfluorescence after treatment of either S. meliloti Rm1021or the S. meliloti Rm1021 bacA null mutant with pJG51Awith bleomycin A5 relative to the untreated controlcultures (Fig. 2b). In contrast, treatment of either the S.meliloti Rm1021 bacA null mutant with or withoutpRK404 (control vector) with bleomycin A5 resulted ina much smaller decrease in the amount of DAPIfluorescence relative to their untreated control cultures(Fig. 2b). These data confirm that BacA-dependent uptakeis the major route of bleomycin internalization in S.meliloti. However, the fact that we observed a reduction inDAPI fluorescence after bleomycin treatment of the S.meliloti bacA null mutant shows that there is also a BacA-independent mechanism for bleomycin internalization inS. meliloti.
BacA does not affect the sensitivity of S. melilotito other DNA-damaging agents
Although BacA is predicted to be in the inner membrane(Glazebrook et al., 1993), it has been demonstratedpreviously that the activation of inner-membrane transport
Internalization of bleomycin in S. meliloti
http://mic.sgmjournals.org 2705
Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
systems can alter the susceptibility of bacterial DNA todamaging agents by altering conditions in the cytoplasm(Ferguson et al., 2000). To rule this out, we analysed thesensitivity of the S. meliloti Rm1021 bacA null mutant andparent strain to methylglyoxal, mitomycin C and MMS,which are also known to induce DNA damage in bacteria(Ferguson et al., 2000; Grzesiuk & Janion, 1996; Otsuji &Murayama, 1972). We found that S. meliloti Rm1021 was
sensitized to all three DNA-damaging agents, but incontrast to bleomycin A5, deletion of bacA did not increasethe resistance of S. meliloti to these agents (data notshown). Additionally, although we found that treatment ofS. meliloti Rm1021 with MMS resulted in DNA degrada-tion, the presence of BacA did not affect the degree ofMMS-induced DNA degradation (Fig. 2c). These findingsshow that BacA does not increase the susceptibility of S.meliloti to other DNA-damaging agents.
B. abortus BacA complements the peptide uptakedefects of an S. meliloti bacA null mutant
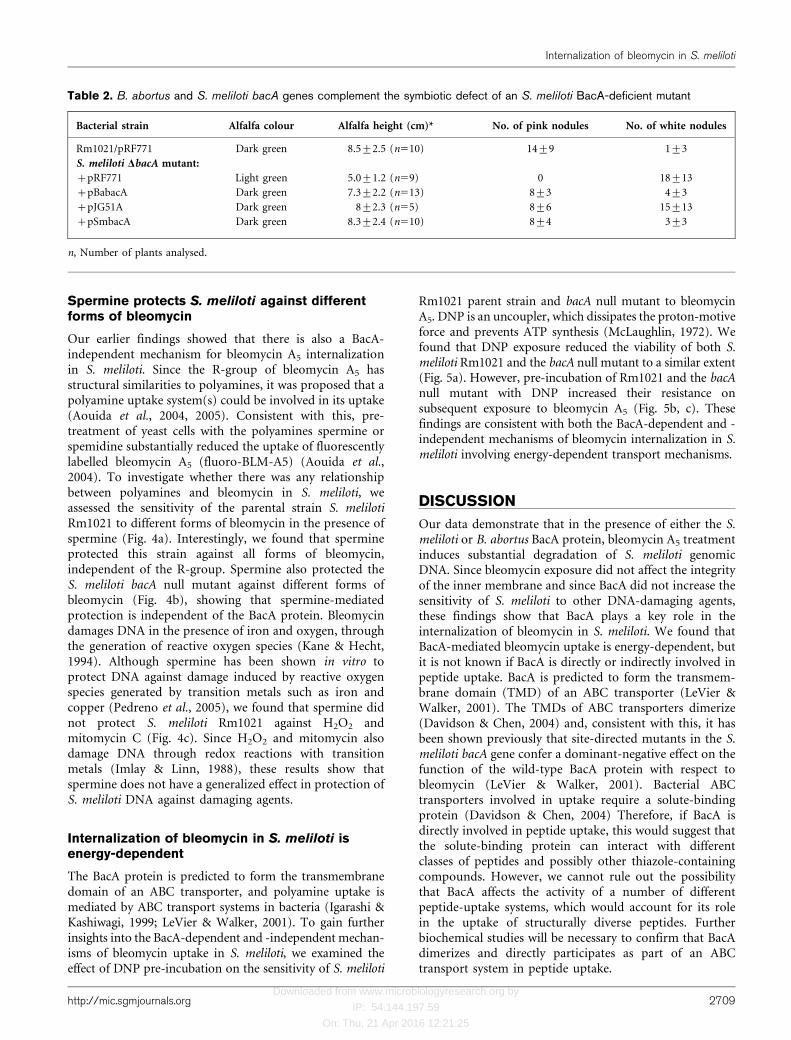
The BacA protein is also responsible for conferringbleomycin sensitivity in B. abortus (LeVier et al., 2000).B. abortus is a highly infectious, containment level 3pathogen (Franz et al., 1997). Therefore, to investigate thefunction of its BacA protein, we cloned the B. abortus bacAgene into the broad-host-range vector pRF771 (Wells &Long, 2002), under control of a constitutively active trppromoter, to create pBabacA. We found that the B. abortusBacA protein sensitized the S. meliloti bacA null mutant tobleomycin A5 relative to the mutant strain with the controlplasmid (Fig. 3a). Additionally, the B. abortus BacA proteinsubstantially reduced the amount of DAPI fluorescence inthe S. meliloti bacA null mutant relative to the mutantstrain with the control plasmid (Fig. 3b), demonstratingthat the B. abortus BacA protein is functionally homolog-ous to S. meliloti in the internalization of bleomycin.
To determine whether B. abortus BacA is also involved in theuptake of truncated Bac7 peptides, we investigated theviability of the S. meliloti bacA null mutant with eitherpBabacA or pRF771 (control plasmid) in the presenceof truncated Bac7 peptides, relative to the parent strain
Fig. 1. BacA confers sensitivity of S. meliloti to different forms ofbleomycin independent of the nature of the R-group. (a) Structureof bleomycin A5, prepared using ChemDraw Std 10.0. The dashedline indicates the boundary of the R-group. The A2 and B2 R-groupstructures are also shown. Bleomycin sulphate is a composed of amixture of the A2 (55–70 %) and B2 (25–32 %) forms. (b) Culturesof the indicated strains were exposed to different forms ofbleomycin, and growth inhibition was determined using the discdiffusion assay. For bleomycin A2, B2 and the sulphate forms,cultures were exposed to 5 ml of a 5 mg ml–1 aqueous stocksolution, and for bleomycin A5 cultures were exposed to 5 ml of a2 mg ml”1 aqueous stock solution. (c) Cultures of the indicatedstrains were exposed to bleomycin sulphate (15 mg ml”1) andviability was assessed at the times indicated. (d) Same as (c)except that bleomycin A5 (3 mg ml”1) was used. The significancevalues shown (**P,0.01, ***P,0.001) represent comparisons ofthe Rm1021 DbacA mutant with the Rm1021 parent strain. Alldatasets shown are representative of trends observed in twoindependent experiments. The error bars represent SD (n53) forone experiment. The arrow indicates that no viable cells weredetected at the next time point.
S. Wehmeier and others
2706 Microbiology 156
Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
(Fig. 3c). We determined that the B. abortus BacA proteingreatly sensitized the S. meliloti bacA null mutant to Bac71–16
(RRIRPRPPRLPRPRPR) and Bac71–35 (RRIRPRPPRLPRP-RPRPLPFPRP GPRPIPRPLPFP), consistent with a role forthe B. abortus BacA protein in their uptake. To confirm this,cultures of S. meliloti Rm1021 bacA null mutant with either
pBabacA or the control vector were incubated with Bac71–16
labelled with the fluorescent dye BODIPY (Bac71–16-BY)(Marlow et al., 2009; Mattiuzzo et al., 2007) and thenanalysed by fluorescence microscopy. To prevent thefluorescence of extracellular Bac71–16-BY, cultures were alsotreated with Trypan Blue, which quenches extracellularfluorescence (Benincasa et al., 2009). We found that cells ofthe S. meliloti Rm1021 bacA null mutant with pBabacA werehighly fluorescent whereas, with the exception of anoccasional cell, the cells of the mutant with control vectorwere non-fluorescent (Fig. 3d, i and ii, respectively). Theseresults show that the B. abortus BacA protein is also involvedin the internalization of truncated Bac7 peptides into S.meliloti.
B. abortus and S. meliloti bacA genescomplement the chronic infection defect of theS. meliloti bacA null mutant in alfalfa whenexpressed under control of the trp promoter
It was demonstrated previously that the E. coli sbmA genecomplemented the alfalfa chronic infection defect of the S.meliloti bacA null mutant, but only when cloned undercontrol of the S. meliloti bacA gene promoter and 168 bpupstream of the bacA coding sequence (Ichige & Walker,1997). This finding led to the hypothesis that expression ofthe S. meliloti bacA gene is regulated during the legumeinfection and that the correct expression is key to theessential role of BacA in the plant infection. However, wefound that the B. abortus bacA gene (pBabacA) comple-mented the chronic infection defect of the S. melilotiRm1021 bacA null mutant within alfalfa, when expressedfrom the trp promoter in pFL771 (Table 2). Since weincluded ~150 bp upstream of the B. abortus bacA codingsequence in pBabacA, we could not rule out the possibilitythat this region was playing a regulatory role duringlegume symbiosis. To determine whether the upstreamregion and promoter of the S. meliloti bacA gene areneeded during legume symbiosis, we cloned the S. melilotibacA gene with 19 bp upstream under control of ptrp
Fig. 2. BacA sensitizes S. meliloti to bleomycin-induced DNAdamage. (a) Cultures of the defined strains in LB medium weretreated or not with bleomycin A5 (20 mg ml”1) for 2 h as indicated.The genomic DNA was extracted and 2 mg analysed on a 1 % (w/v) agarose gel. (b) Cultures of the indicated strains were treated ornot with bleomycin A5 as in (a) and then the DNA was stained withDAPI and the fluorescence intensity was determined. Thefluorescence intensity for each bleomycin-treated strain is shownas a percentage relative to that of the untreated control. Thesignificance values (**P,0.01) represent comparisons of theRm1021DbacA mutant with the parent strain and the Rm1021DbacA mutant carrying pJG51A with the Rm1021DbacA mutantcarrying pRK404. The error bars represent SD (n53) for oneexperiment. (c) Same as (a) except that cultures were treated or notwith MMS as indicated.
Internalization of bleomycin in S. meliloti
http://mic.sgmjournals.org 2707

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
(pSmbacA). We found that the S. meliloti bacA gene clonedunder control of ptrp was able to complement thesymbiotic defect of the S. meliloti bacA null mutant to asimilar extent to S. meliloti bacA cloned under control of its
own promoter with an extended upstream region(pJG51A) (Table 2). These findings show that the regulatedexpression of bacA is not essential for S. meliloti to formthe legume symbiosis.
Fig. 3. BacA of B. abortus is also involved in peptide uptake. (a) Cultures, containing approximately 1�109 c.f.u. ml”1, weretreated with bleomycin A5 (20 mg ml”1) for 2 h and then the viability was determined. (b) Same as (a) except that cultures weretreated or not with bleomycin and then the DNA was quantified by DAPI fluorescence. The fluorescence intensity for eachbleomycin-treated strain is shown as a percentage relative to that of the untreated control. (c) Cultures of the indicated strainswere incubated with and without Bac71–16 and Bac71–35 (1 mM) in LB medium for 1 h and then the viability was assessed. Thearrows indicate that no viable cells were detected. The datasets shown are representative of the trends observed in twoindependent experiments and the error bars represent SD (n53) for one experiment. The significance values (***P,0.001)represent comparison of the Rm1021 DbacA mutant carrying pRF771 (control) compared with pBabacA. (d) Cultures of theindicated strains were incubated with Bac71–16 labelled with the fluorescent dye BODIPY (Bac71–16-BY), treated with TrypanBlue and then analysed by microscopy as indicated. The images shown are representative fields of view from at least twoindependent experiments (bars, 20 mm).
S. Wehmeier and others
2708 Microbiology 156
Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
Spermine protects S. meliloti against differentforms of bleomycin
Our earlier findings showed that there is also a BacA-independent mechanism for bleomycin A5 internalizationin S. meliloti. Since the R-group of bleomycin A5 hasstructural similarities to polyamines, it was proposed that apolyamine uptake system(s) could be involved in its uptake(Aouida et al., 2004, 2005). Consistent with this, pre-treatment of yeast cells with the polyamines spermine orspemidine substantially reduced the uptake of fluorescentlylabelled bleomycin A5 (fluoro-BLM-A5) (Aouida et al.,2004). To investigate whether there was any relationshipbetween polyamines and bleomycin in S. meliloti, weassessed the sensitivity of the parental strain S. melilotiRm1021 to different forms of bleomycin in the presence ofspermine (Fig. 4a). Interestingly, we found that spermineprotected this strain against all forms of bleomycin,independent of the R-group. Spermine also protected theS. meliloti bacA null mutant against different forms ofbleomycin (Fig. 4b), showing that spermine-mediatedprotection is independent of the BacA protein. Bleomycindamages DNA in the presence of iron and oxygen, throughthe generation of reactive oxygen species (Kane & Hecht,1994). Although spermine has been shown in vitro toprotect DNA against damage induced by reactive oxygenspecies generated by transition metals such as iron andcopper (Pedreno et al., 2005), we found that spermine didnot protect S. meliloti Rm1021 against H2O2 andmitomycin C (Fig. 4c). Since H2O2 and mitomycin alsodamage DNA through redox reactions with transitionmetals (Imlay & Linn, 1988), these results show thatspermine does not have a generalized effect in protection ofS. meliloti DNA against damaging agents.
Internalization of bleomycin in S. meliloti isenergy-dependent
The BacA protein is predicted to form the transmembranedomain of an ABC transporter, and polyamine uptake ismediated by ABC transport systems in bacteria (Igarashi &Kashiwagi, 1999; LeVier & Walker, 2001). To gain furtherinsights into the BacA-dependent and -independent mechan-isms of bleomycin uptake in S. meliloti, we examined theeffect of DNP pre-incubation on the sensitivity of S. meliloti
Rm1021 parent strain and bacA null mutant to bleomycinA5. DNP is an uncoupler, which dissipates the proton-motiveforce and prevents ATP synthesis (McLaughlin, 1972). Wefound that DNP exposure reduced the viability of both S.meliloti Rm1021 and the bacA null mutant to a similar extent(Fig. 5a). However, pre-incubation of Rm1021 and the bacAnull mutant with DNP increased their resistance onsubsequent exposure to bleomycin A5 (Fig. 5b, c). Thesefindings are consistent with both the BacA-dependent and -independent mechanisms of bleomycin internalization in S.meliloti involving energy-dependent transport mechanisms.
DISCUSSION
Our data demonstrate that in the presence of either the S.meliloti or B. abortus BacA protein, bleomycin A5 treatmentinduces substantial degradation of S. meliloti genomicDNA. Since bleomycin exposure did not affect the integrityof the inner membrane and since BacA did not increase thesensitivity of S. meliloti to other DNA-damaging agents,these findings show that BacA plays a key role in theinternalization of bleomycin in S. meliloti. We found thatBacA-mediated bleomycin uptake is energy-dependent, butit is not known if BacA is directly or indirectly involved inpeptide uptake. BacA is predicted to form the transmem-brane domain (TMD) of an ABC transporter (LeVier &Walker, 2001). The TMDs of ABC transporters dimerize(Davidson & Chen, 2004) and, consistent with this, it hasbeen shown previously that site-directed mutants in the S.meliloti bacA gene confer a dominant-negative effect on thefunction of the wild-type BacA protein with respect tobleomycin (LeVier & Walker, 2001). Bacterial ABCtransporters involved in uptake require a solute-bindingprotein (Davidson & Chen, 2004) Therefore, if BacA isdirectly involved in peptide uptake, this would suggest thatthe solute-binding protein can interact with differentclasses of peptides and possibly other thiazole-containingcompounds. However, we cannot rule out the possibilitythat BacA affects the activity of a number of differentpeptide-uptake systems, which would account for its rolein the uptake of structurally diverse peptides. Furtherbiochemical studies will be necessary to confirm that BacAdimerizes and directly participates as part of an ABCtransport system in peptide uptake.
Table 2. B. abortus and S. meliloti bacA genes complement the symbiotic defect of an S. meliloti BacA-deficient mutant
Bacterial strain Alfalfa colour Alfalfa height (cm)* No. of pink nodules No. of white nodules
Rm1021/pRF771 Dark green 8.5±2.5 (n510) 14±9 1±3
S. meliloti DbacA mutant:
+pRF771 Light green 5.0±1.2 (n59) 0 18±13
+pBabacA Dark green 7.3±2.2 (n513) 8±3 4±3
+pJG51A Dark green 8±2.3 (n55) 8±6 15±13
+pSmbacA Dark green 8.3±2.4 (n510) 8±4 3±3
n, Number of plants analysed.
Internalization of bleomycin in S. meliloti
http://mic.sgmjournals.org 2709

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
Combining these results with those of our previous study(Marlow et al., 2009), we found that the S. meliloti BacAprotein is involved in the uptake of both proline-rich and
Fig. 4. Spermine protects S. meliloti against bleomycin but notH2O2 or mitomycin C. (a , b) Cultures of the indicated strains wereexposed to different forms of bleomycin, at the concentrationsdescribed for Fig. 1(b), and growth inhibition was determined bythe disc diffusion assay in the presence (black bars) or absence(white bars) of 1 mM spermine. (c) Cultures of S. meliloti Rm1021were exposed to either 5 ml H2O2 (30 %, w/w) or mitomycin C(0.5 mg ml”1), and growth inhibition was determined by the discdiffusion assay in the presence (black bars) or absence (whitebars) of 1 mM spermine. All datasets shown are a representativeset from at least two independent experiments. The error barsrepresent SD (n53) for one experiment. The significance valuesshown (***P,0.001) represent comparisons of the sensitivities ofboth strains in LB alone and with spermine added.
Fig. 5. Uptake of bleomycin is energy-dependent. (a) Cultures ofthe indicated strains were treated with (black bars) and without(white bars) 5 mM DNP for 50 min in LB medium and then theviability was determined. (b, c) Cultures of the indicated strainswere treated (black bars) or not (white bars) with DNP asdescribed in (a), diluted to OD600 ~0.1 in LB, exposed to 3 mgbleomycin A5 ml”1 for the indicated time, and then the viabilitywas determined. The results are expressed as a percentage ofthe viable cells at time zero. The datasets shown arerepresentative of at least two independent experiments and theerror bars represent SD (n53) for one experiment. The signifi-cance values shown (**P,0.01, ***P,0.001) represent com-parisons of the sensitivities of both strains with and without DNPpretreatment.
S. Wehmeier and others
2710 Microbiology 156

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
thiazole-modified peptides. Therefore, within the soil andduring legume symbiosis, BacA-mediated uptake ofdifferent classes of peptides and/or thiazole-containingcompounds into S. meliloti could be important for envi-ronmental survival and/or bacteroid development. Inter-estingly, plants produce thiazole-containing compounds inresponse to invading micro-organisms (Yorgey et al.,1994). Therefore, the BacA-mediated uptake of one ormore of these compounds could be important during thelegume symbiosis. Additionally, hundreds of nodule-specific cysteine-rich (NCR) peptides have been identifiedin the legume host of S. meliloti (Alunni et al., 2007;Mergaert et al., 2003, 2006). The NCR peptides haverecently been shown to play a key role in S. melilotibacteroid differentiation (Van de Velde et al., 2010). Atpresent, S. meliloti factors involved in the NCR-mediatedresponse have not been identified. However, since it hasbeen shown that NCR peptides can enter S. meliloti (Vande Velde et al., 2010) and since we found that BacA isinvolved in the uptake of structurally diverse peptides,BacA may also be playing a key role in the uptake of one ormore of these NCR peptides.
We found by functional complementation experimentsthat the B. abortus BacA protein is also involved in uptakeof bleomycin and truncated Bac7 peptides in S. meliloti.These results suggest that BacA-mediated peptide uptakecould also be important for B. abortus during hostinfections. B. abortus causes abortions in cattle, and inhumans it results in brucellosis, a severe debilitating disease(Franz et al., 1997). During mammalian infections, B.abortus is expected to encounter a number of differentclasses of peptides such as the proline-rich Bac7 peptidesused in this study, as they were derived from a full-lengthpeptide produced by bovine leukocytes (Frank et al., 1990).Other mammalian hosts are also known to produceproline-rich peptides (Gennaro et al., 2002), and threemajor families of antimicrobial peptides (defensins,cathelicidins and histatins) are produced as part of thehuman innate immune response (for a recent review referto Rivas-Santiago et al., 2009). However, further studieswill be necessary to understand the effect of these differentclasses of peptides on B. abortus and the role of BacA in thisresponse.
Our findings also demonstrate that there is a BacA-independent mechanism of bleomycin uptake in S. meliloti.Based on work with bleomycin in yeast (Aouida et al.,2004, 2005), we hypothesize that the BacA-independentmechanism of bleomycin uptake in S. meliloti occurs viaone or more polyamine uptake system(s). Consistent withthis, we showed that spermine protected S. meliloti againstbleomycin and that the BacA-independent mechanism ofbleomycin uptake is energy-dependent. Since our fluores-cently labelled bleomycin was unable to enter S. meliloti, wehave not been able to demonstrate conclusively thatspermine reduces bleomycin uptake in S. meliloti. How-ever, since spermine did not protect S. meliloti againstother agents, which damage DNA via a similar mechanism
to bleomycin (Imlay & Linn, 1988), our findings suggestthat spermine is not mediating its protective effect againstbleomycin at the level of the DNA. Polyamine uptake inbacteria has been most extensively investigated in E. coli(Igarashi & Kashiwagi, 1999), in which spermine uptakeoccurs via the ATP-binding cassette spermidine-preferen-tial uptake system encoded by the four genes potA–D.Based on sequence similarity to the E. coli potA–D genes,we have identified five different probable ABC transporters(SMc01963–66, SMb20380–83, SMc01652–55, SMb20281–84 and PotF–I) encoded in the S. meliloti Rm1021 genome(Galibert et al., 2001), which are potential polyamine-uptake systems. Like the E. coli spermine uptake system,each of the S. meliloti probable ABC transporters consistsof four proteins, which have between 22% and 53 %identity to the corresponding proteins in the E. coli PotA–D system (data not shown). Further studies will benecessary to construct single and combination mutants inthese different systems, to determine how they affectpolyamine and bleomycin uptake.
In summary, our findings show the importance of BacAproteins in the uptake of structurally diverse peptides.Since the BacA protein also sensitizes M. tuberculosis tobleomycin (Domenech et al., 2009), it will be interesting todetermine whether this protein is also involved in peptideuptake in this pathogen, and whether BacA-mediatedpeptide uptake is important during the host infection.
ACKNOWLEDGEMENTS
An MRC New Investigator (G0501107) grant and a BBSRC (BB/D000564/1) grant awarded to G. P. F. supported this work. S. W. wasfunded by the BBSRC and V. F. is funded by the MRC grant. Thestudentship of V. L. M. was funded by the BBSRC and M. A. and K. M.are SULSA-funded PhD students. The Canadian Institute of Health(MPO-67196) funds D. R. Thanks to Graham Walker and AndreasHaag for helpful discussion and Marty Roop II for provision of the B.abortus DNA. Thanks also to Hazel Phillips for preparing thebleomycin structure figure and Xiaming Yang for technical assistance.
REFERENCES
Alunni, B., Kevei, Z., Redondo-Nieto, M., Kondorosi, A., Mergaert, P.& Kondorosi, E. (2007). Genomic organisation and evolutionaryinsights on GRP and NCR genes, two large nodule-specific genefamilies in Medicago truncatula. Mol Plant Microbe Interact 20, 1138–1148.
Aouida, M., Leduc, A., Wang, H. & Ramotar, D. (2004).Characterization of a transport and detoxification pathway for theantitumour drug bleomycin in Saccharomyces cerevisiae. Biochem J384, 47–58.
Aouida, M., Leduc, A., Poulin, R. & Ramotar, D. (2005). AGP2 encodesthe major permease for high affinity polyamine import inSaccharomyces cerevisiae. J Biol Chem 280, 24267–24276.
Benincasa, M., Scocchi, M., Podda, E., Skerlavaj, B., Dolzani, L. &Gennaro, R. (2004). Antimicrobial activity of Bac7 fragments againstdrug-resistant clinical isolates. Peptides 25, 2055–2061.
Benincasa, M., Pacor, S., Gennaro, R. & Scocchi, M. (2009). Rapidand reliable detection of antimicrobial peptide penetration into
Internalization of bleomycin in S. meliloti
http://mic.sgmjournals.org 2711

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
Gram-negative bacteria based on fluorescence quenching. Antimicrob
Agents Chemother 53, 3501–3504.
Davidson, A. L. & Chen, J. (2004). ATP-binding cassette transporters
in bacteria. Annu Rev Biochem 73, 241–268.
Ditta, G., Schmidhauser, T., Yakobson, E., Lu, P., Liang, X.-W., Finlay,D. R., Guiney, D. & Helinski, D. R. (1985). Plasmids related to the
broad host range vector, pRK290, useful for gene cloning and formonitoring gene expression. Plasmid 13, 149–153.
Domenech, P., Kobayashi, H., Levier, K., Walker, G. C. & Barry, C. E.,III (2009). BacA: an ABC transporter involved in maintenance of
chronic murine infections with Mycobacterium tuberculosis. J Bacteriol
191, 477–485.
Ferguson, G. P., Battista, J. R., Lee, A. T. & Booth, I. R. (2000).Protection of the DNA during the exposure of Escherichia coli cells to
a toxic metabolite: the role of the KefB and KefC potassium channels.Mol Microbiol 35, 113–122.
Ferguson, G. P., Roop, R. M., II & Walker, G. C. (2002). Deficiency ofSinorhizobium meliloti bacA mutant in alfalfa symbiosis correlates
with alteration of cell envelope. J Bacteriol 184, 5625–5632.
Ferguson, G. P., Datta, A., Baumgartner, J., Roop, R. M., II, Carlson,R. W. & Walker, G. C. (2004). Similarity to peroxisomal-membrane
protein family reveals that Sinorhizobium and Brucella BacA affect
lipid-A fatty acids. Proc Natl Acad Sci U S A 101, 5012–5017.
Ferguson, G. P., Datta, A., Carlson, R. W. & Walker, G. C. (2005).Importance of unusually modified lipid A in Sinorhizobium stress
resistance and legume symbiosis. Mol Microbiol 56, 68–80.
Ferguson, G. P., Jansen, A., Marlow, V. L. & Walker, G. C. (2006).BacA-mediated bleomycin sensitivity in Sinorhizobium meliloti isindependent of the unusual lipid A modification. J Bacteriol 188,
3143–3148.
Finan, T. M., Kunkel, B., de Vos, G. F. & Signer, E. R. (1986). Secondsymbiotic megaplasmid in Rhizobium meliloti carrying exopolysac-
charide and thiamine synthesis genes. J Bacteriol 167, 66–72.
Frank, R. W., Gennaro, R., Schneider, K., Przybylski, M. & Romeo, D.(1990). Amino acid sequences of two proline-rich bactenecins.
Antimicrobial peptides of bovine neutrophils. J Biol Chem 265,18871–18874.
Franz, D. R., Jahrling, P. B., Friedlander, A. M., McClain, D. J., Hoover,D. L., Bryne, W. R., Pavlin, J. A., Christopher, G. W. & Eitzen, E. M., Jr(1997). Clinical recognition and management of patients exposed to
biological warfare agents. JAMA 278, 399–411.
Galibert, F., Finan, T. M., Long, S. R., Puhler, A., Abola, P., Ampe, F.,Barloy-Hubler, F., Barnett, M. J., Becker, A. & other authors (2001).The composite genome of the legume symbiont Sinorhizobiummeliloti. Science 293, 668–672.
Gennaro, R., Zanetti, M., Benincasa, M., Podda, E. & Miani, M. (2002).Pro-rich antimicrobial peptides from animals: structure, biologi-cal functions and mechanism of action. Curr Pharm Des 8, 763–
778.
Gibson, K. E., Kobayashi, H. & Walker, G. C. (2008). Molecular
determinants of a symbiotic chronic infection. Annu Rev Genet 42,
413–441.
Glazebrook, J., Ichige, A. & Walker, G. C. (1993). A Rhizobium
meliloti homolog of the Escherichia coli peptide-antibiotic transport
protein SbmA is essential for bacteroid development. Genes Dev 7,1485–1497.
Grzesiuk, E. & Janion, C. (1996). MMS-induced mutagenesis andDNA repair in Escherichia coli dnaQ49: contribution of UmuD9 to
DNA repair. Mutat Res 362, 147–154.
Haag, A. F., Wehmeier, S., Beck, S., Marlow, V. L., Fletcher, V.,James, E. K. & Ferguson, G. P. (2009). The Sinorhizobium meliloti
LpxXL and AcpXL proteins play important roles in bacteroid
development within alfalfa. J Bacteriol 191, 4681–4686.
Ichige, A. & Walker, G. C. (1997). Genetic analysis of the Rhizobium
meliloti bacA gene: functional interchangeability with the Escherichiacoli sbmA gene and phenotypes of mutants. J Bacteriol 179, 209–
216.
Igarashi, K. & Kashiwagi, K. (1999). Polyamine transport in bacteriaand yeast. Biochem J 344, 633–642.
Imlay, J. A. & Linn, S. (1988). DNA damage and oxygen radicaltoxicity. Science 240, 1302–1309.
Johnson, J. L. (1994). Similarity analysis of DNAs. In Methods for
General and Molecular Bacteriology, pp. 655–682. Edited byP. Gerhardt, R. G. E. Murray, W. A. Wood & N. R. Kreig.
Washington, DC: American Society of Microbiology.
Jones, K. M., Kobayashi, H., Davies, B. W., Taga, M. E. & Walker, G. C.(2007). How rhizobial symbionts invade plants: the Sinorhizobium–
Medicago model. Nat Rev Microbiol 5, 619–633.
Kane, S. A. & Hecht, S. M. (1994). Polynucleotide recognition and
degradation by bleomycin. Prog Nucleic Acid Res Mol Biol 49, 313–
352.
Lavina, M., Pugsley, A. P. & Moreno, F. (1986). Identification,
mapping, cloning and characterization of a gene (sbmA) required formicrocin B17 action on Escherichia coli K12. J Gen Microbiol 132,
1685–1693.
Leigh, J. A., Signer, E. R. & Walker, G. C. (1985). Exopolysaccharide-deficient mutants of Rhizobium meliloti that form ineffective nodules.
Proc Natl Acad Sci U S A 82, 6231–6235.
LeVier, K. & Walker, G. C. (2001). Genetic analysis of the
Sinorhizobium meliloti BacA protein: differential effects of mutations
on phenotypes. J Bacteriol 183, 6444–6453.
LeVier, K., Phillips, R. W., Grippe, V. K., Roop, R. M., II & Walker, G. C.(2000). Similar requirements of a plant symbiont and a mammalian
pathogen for prolonged intracellular survival. Science 287, 2492–2493.
Lim, S. T., Jue, C. K., Moore, C. W. & Lipke, P. N. (1995). Oxidative cell
wall damage mediated by bleomycin-Fe(II) in Saccharomycescerevisiae. J Bacteriol 177, 3534–3539.
Marlow, V. L., Haag, A. F., Kobayashi, H., Fletcher, V., Scocchi, M.,Walker, G. C. & Ferguson, G. P. (2009). Essential role for the BacA
protein in the uptake of a truncated eukaryotic peptide in
Sinorhizobium meliloti. J Bacteriol 191, 1519–1527.
Mattiuzzo, M., Bandiera, A., Gennaro, R., Benincasa, M., Pacor, S.,Antcheva, N. & Scocchi, M. (2007). Role of the Escherichia coli SbmA
in the antimicrobial activity of proline-rich peptides. Mol Microbiol66, 151–163.
McLaughlin, S. (1972). The mechanism of action of DNP onphospholipid bilayer membranes. J Membr Biol 9, 361–372.
Meade, H. M., Long, S. R., Ruvkun, G. B., Brown, S. E. & Ausubel, F. M.(1982). Physical and genetic characterization of symbiotic andauxotrophic mutants of Rhizobium meliloti induced by transposon
Tn5 mutagenesis. J Bacteriol 149, 114–122.
Mergaert, P., Nikovics, K., Keleman, Z., Maunoury, N., Vaubert, D.,Kondorosi, A. & Kondorosi, E. (2003). A novel family of Medicago
truncatula consisting of more than 300 nodule-specific genes codingfor small, secreted polypeptides with conserved cysteine motifs. Plant
Physiol 132, 161–173.
Mergaert, P., Uchiumi, T., Alunni, B., Evanno, G., Cheron, A., Catrice, O.,Mausett, A. E., Barloy-Hubler, F., Galibert, F. & other authors (2006).Eukaryotic control on bacterial cell cycle and differentiation in the
Rhizobium–legume symbiosis. Proc Natl Acad Sci U S A 103, 5230–5235.
S. Wehmeier and others
2712 Microbiology 156

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 12:21:25
Otsuji, N. & Murayama, I. (1972). Deoxyribonucleic acid damageby monofunctional mitomycins and its repair in Escherichia coli.J Bacteriol 109, 475–483.
Pedreno, E., Lopez-Contreras, A. J., Cremades, A. & Penafiel, R.(2005). Protecting or promoting effects of spermine on DNA strandbreakage induced by iron or copper ions as a function of metalconcentration. J Inorg Biochem 99, 2074–2080.
Raetz, C. R. & Whitfield, C. (2002). Lipopolysaccharide endotoxins.Annu Rev Biochem 71, 635–700.
Rivas-Santiago, B., Serrano, C. J. & Enciso-Moreno, J. A. (2009).Susceptibility to infectious diseases based on antimicrobial peptideproduction. Infect Immun 77, 4690–4695.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1982). Molecular Cloning:a Laboratory Manual, 2nd edn. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory.
Scocchi, M., Mattiuzzo, M., Benincasa, M., Antcheva, N., Tossi, A. &Gennaro, R. (2008). Investigating the mode of action of proline-richantimicrobial peptides using a genetic approach: a tool to identifynew bacterial targets amenable to the design of novel antibiotics.Methods Mol Biol 494, 161–176.
Sharypova, L. A., Niehaus, K., Scheidle, H., Holst, O. & Becker, A.(2003). Sinorhizobium meliloti acpXL mutant lacks the C28 hydro-
xylated fatty acid moiety of lipid A and does not express a slow
migrating form of lipopolysaccharide. J Biol Chem 278, 12946–12954.
Umezawa, H., Maeda, K., Takeuchi, T. & Okami, Y. (1966). New
antibiotics, bleomycin A and B. J Antibiot (Tokyo) 19, 200–209.
Van de Velde, W., Zehirov, G., Szatmari, A., Debreczeny, M.,Ishihara, H., Kevei, Z., Farkas, A., Mikulass, K., Nagy, A. & otherauthors (2010). Plant peptides govern terminal differentiation of
bacteria in symbiosis. Science 327, 1122–1126.
Wells, D. H. & Long, S. R. (2002). The Sinorhizobium meliloti stringent
response affects multiple aspects of symbiosis. Mol Microbiol 43,
1115–1127.
Wilson, K. (1987). Preparation of genomic DNA from bacteria. In
Current Protocols in Molecular Biology, pp. 2.4.1–2.4.5. Edited by F.
M. Ausubel, R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J.
A. Smith & K. Struhl. New York: Green Publishing & Wiley-
Interscience.
Yorgey, P., Lee, J., Kordel, J., Vivas, E., Warner, P., Jebaratnam, D. &Kolter, R. (1994). Posttranslational modifications in microcin B17
define an additional class of DNA gyrase inhibitor. Proc Natl Acad Sci
U S A 91, 4519–4523.
Edited by: M. F. Hynes
Internalization of bleomycin in S. meliloti
http://mic.sgmjournals.org 2713
Related Documents











