RESEARCH ARTICLE Cathy Craig Gert-Jan Pepping Madeleine Grealy Intercepting beats in predesignated target zones Received: 27 April 2004 / Accepted: 12 February 2005 / Published online: 24 May 2005 Ó Springer-Verlag 2005 Abstract Moving to a rhythm necessitates precise timing between the movement of the chosen limb and the tim- ing imposed by the beats. However, the temporal information specifying the moment when a beat will sound (the moment onto which one must synchronise one’s movement) is not continuously provided by the acoustic array. Because of this informational void, the actors need some form of prospective information that will allow them to act sufficiently ahead of time in order to get their hand in the right place at the right time. In this acoustic interception study, where participants were asked to move between two targets in such a way that they arrived and stopped in the target zone at the same time as a beat sounded, we tested a model derived from tau-coupling theory (Lee DN (1998) Ecol Psychol 10:221–250). This model attempts to explain the form of a potential timing guide that specifies the duration of the inter-beat intervals and also describes how this infor- mational guide can be used in the timing and guidance of movements. The results of our first experiment show that, for inter-beat intervals of less than 3 s, a large proportion of the movement (over 70%) can be ex- plained by the proposed model. However, a second experiment, which augments the time between beats so that it surpasses 3 s, shows a marked decline in the percentage of information/movement coupling. A close analysis of the movement kinematics indicates a lack of control and anticipation in the participants’ movements. The implications of these findings, in light of other re- search studies, are discussed. Keywords Interception Acoustic beats Tau-coupling Movement Control Dancing to our favourite song demands that the move- ments of our body parts are spatially controlled relative to the temporal structure being imposed by the music. This task, not unlike catching a moving object, has both spatial and temporal constraints in that the unfolding of the action is guided by the rhythm of the music. In the case of rhythms, however, there is no continuous online information that constantly informs participants about the time remaining until the next beat sounds. This informational void has to be filled by internal timing mechanisms that allow participants to prospectively guide their movements. A rich body of research exists on the intrinsic temporal mechanisms underlying rhythmi- cally-paced movements (for example, Treisman et al 1990; Wing and Kristofferson 1973). Commonly, these studies measure the degree of temporal synchrony be- tween some movement (usually a finger tap) and some temporal framework (a beat imposed by a metronome), and are characterised by assessing temporal variability. Whilst this ‘‘tapping paradigm’’ is extensively used to provide evidence for the existence of internal timing mechanisms, the models proposed thus far do not ex- plain how the finger is controlled in order to make the tap in the first place. In particular, the existing models fall short of addressing the spatial–temporal character- istics required to control such movements. C. Craig (&) UMR ‘Mouvement et Perception’, Faculte´ des Sciences du Sport, Universite´ de la Me´diterrane´e, 163 Avenue de Luminy, 13288 Marseille, France E-mail: [email protected] Tel.: +33-4-91172275 Fax: +33-4-91172252 G.-J. Pepping Department of Physical Education, Sport and Leisure Studies, School of Education, University of Edinburgh, St. Leonard’s Land, Holyrood Road, Edinburgh, EH8 8AQ, Scotland M. Grealy Department of Psychology, University of Strathclyde, Graham Hills Building, 40 George Street, Glasgow, G1 1QE, Scotland Exp Brain Res (2005) 165: 490–504 DOI 10.1007/s00221-005-2322-x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH ARTICLE
Cathy Craig Æ Gert-Jan Pepping Æ Madeleine Grealy
Intercepting beats in predesignated target zones
Received: 27 April 2004 / Accepted: 12 February 2005 / Published online: 24 May 2005� Springer-Verlag 2005
Abstract Moving to a rhythm necessitates precise timingbetween the movement of the chosen limb and the tim-ing imposed by the beats. However, the temporalinformation specifying the moment when a beat willsound (the moment onto which one must synchroniseone’s movement) is not continuously provided by theacoustic array. Because of this informational void, theactors need some form of prospective information thatwill allow them to act sufficiently ahead of time in orderto get their hand in the right place at the right time. Inthis acoustic interception study, where participants wereasked to move between two targets in such a way thatthey arrived and stopped in the target zone at the sametime as a beat sounded, we tested a model derived fromtau-coupling theory (Lee DN (1998) Ecol Psychol10:221–250). This model attempts to explain the form ofa potential timing guide that specifies the duration of theinter-beat intervals and also describes how this infor-mational guide can be used in the timing and guidance
of movements. The results of our first experiment showthat, for inter-beat intervals of less than 3 s, a largeproportion of the movement (over 70%) can be ex-plained by the proposed model. However, a secondexperiment, which augments the time between beats sothat it surpasses 3 s, shows a marked decline in thepercentage of information/movement coupling. A closeanalysis of the movement kinematics indicates a lack ofcontrol and anticipation in the participants’ movements.The implications of these findings, in light of other re-search studies, are discussed.
Keywords Interception Æ Acoustic beats ÆTau-coupling Æ Movement Æ Control
Dancing to our favourite song demands that the move-ments of our body parts are spatially controlled relativeto the temporal structure being imposed by the music.This task, not unlike catching a moving object, has bothspatial and temporal constraints in that the unfolding ofthe action is guided by the rhythm of the music. In thecase of rhythms, however, there is no continuous onlineinformation that constantly informs participants aboutthe time remaining until the next beat sounds. Thisinformational void has to be filled by internal timingmechanisms that allow participants to prospectivelyguide their movements. A rich body of research exists onthe intrinsic temporal mechanisms underlying rhythmi-cally-paced movements (for example, Treisman et al1990; Wing and Kristofferson 1973). Commonly, thesestudies measure the degree of temporal synchrony be-tween some movement (usually a finger tap) and sometemporal framework (a beat imposed by a metronome),and are characterised by assessing temporal variability.Whilst this ‘‘tapping paradigm’’ is extensively used toprovide evidence for the existence of internal timingmechanisms, the models proposed thus far do not ex-plain how the finger is controlled in order to make thetap in the first place. In particular, the existing modelsfall short of addressing the spatial–temporal character-istics required to control such movements.
C. Craig (&)UMR ‘Mouvement et Perception’,Faculte des Sciences du Sport,Universite de la Mediterranee,163 Avenue de Luminy, 13288 Marseille,FranceE-mail: [email protected].: +33-4-91172275Fax: +33-4-91172252
G.-J. PeppingDepartment of Physical Education,Sport and Leisure Studies,School of Education,University of Edinburgh,St. Leonard’s Land,Holyrood Road, Edinburgh, EH8 8AQ,Scotland
M. GrealyDepartment of Psychology,University of Strathclyde,Graham Hills Building, 40 George Street,Glasgow, G1 1QE,Scotland
Exp Brain Res (2005) 165: 490–504DOI 10.1007/s00221-005-2322-x

The current lack of focus on the temporal evolutionof action within an interval frame possibly stems fromthe work of Wing and Kristofferson (1973). Their two-level model partitions variability in timing into a centraltimekeeper and motor implementation, with Wing(1980) clearly showing that the central timer can beexperimentally manipulated while the motor componentremains constant. These components have also beenshown to be dissociable in single interval productiontasks (Rosenbaum and Patashnik 1980a, 1980b). Thevariance being discussed here, though, refers to the delayin motor implementation that a centrally-generatedinterval is subject to, whereas, when dancing or playingmusic, it is clear that there is synchronisation betweenthe unfolding of the movements of the different bodyparts over time and the temporal structure being im-posed by the music. In other words, body and limbmovements are patterned relative to the music that isplayed or danced to. This poses a genuine problem in theapplication of the current models to the overall temporalorganisation of the control of goal-directed action. Toaddress this problem, we propose a very different ap-proach to conceptualising the organisation of temporalcoordination. We suggest that it is more appropriate tothink of time’s passage (or a time interval) as beingrepresented by an evolving event, rather than an internalclock which demarcates the passage of time in a constantmanner.
If we take into account Mach’s (1883) reasoning that‘‘...time is an abstraction, at which we arrive by means ofthe change of things’’ and Gibson’s (1975) who arguedthat ‘‘events are perceivable, but time is not’’, it might bemore useful to think of time perception as an ever-changing representation of an event. In the physicalworld it is known that the way in which somethingchanges over time, for instance the closure of a spatialgap between a ball and a target, provides robust tem-poral information that can be used in the control of anaction (Lee et al 2001). This evolution of change,otherwise known as ‘‘tau’’, is specified as the time-to-closure of a motion-gap (such as ball to target) at itscurrent closure-rate. Theoretically the tau of a motion-gap can be sensed directly as its value is specified by tau-variables in optic, acoustic, haptic and other sensoryflow fields (Lee 1998). Tau is powerful information forguiding movement because it is a universal (temporal)measure applicable to any motion-gap, whether it bedistance, angle or force. Moreover, tau is sufficientinformation to guide closure of a motion-gap; in otherwords, more complex information about gap size,velocity and acceleration of closure and so on is notneeded. Data from many behavioural experimentsindicate that humans and other animals use tau infor-mation when guiding movement (Craig et al 2000; Leeet al 1999, 2001). Thus, tau is simple, general and robustinformation that delineates the guiding of movements atboth behavioural and neural levels. Furthermore, it isprospective; that is to say, it provides temporal infor-mation about the time remaining until a motion gap is
closed (gap closure is denoted where tau=0). The appealof a prospective informational variable in the context oftiming is that it allows for the preparation and initiationof an action ahead of time.
Although the example of catching a ball differs inmany ways from tapping along to acoustic beats, thereare important similarities with respect to their co-ordi-nation. When catching a ball one has to place one’s handin a particular spatial location at a particular moment intime. Hence precise temporal information is required soas to be able to co-ordinate one’s actions with themovement of the ball ahead of time. It is hypothesisedthat this prospective temporal information, the timeremaining until the ball arrives, is visually specified inthe unfolding of the event, or more specifically by thetau of the gap between the ball and the target (Lee et al2001). Imagine now that an observer wishes to interceptthis ball in a target zone. According to the tau-couplinghypothesis, the observer would directly pick up temporalinformation, visually specified, through the way thespatial gap between the ball and the target is closing, andget their hand to the target at the same time as the ballarrives by adjusting the closure of the hand/target gap insuch a way that the tau of this gap, or the time to closureof this gap, is kept directly proportional to the closure ortau of the ball/target gap. By doing this they both willarrive at the interception point at the same time.
In the case of tapping to discrete acoustic beats thereare likewise very stringent spatial and temporal con-straints in that one has to place one’s finger in the rightplace (the contact zone) at the right time (when the beatsounds). In other words, the closure of two gaps has tobe synchronised—the spatial gap between the finger andthe target zone (the movement gap) and the temporalgap between two beats (the information gap). Whentapping, however, the temporal information specifyingthe time remaining until the next beat arrives cannot bedirectly perceived from the environment in the unfoldingof the event (unlike the case of a ball moving to thetarget). Instead, this informational void, so to speak, hasto be filled by some kind of internal temporal informa-tion, generated within the CNS, that is accessible to theactor and which reliably specifies the time remaininguntil the next beat sounds.
This intrinsic temporal information, which bridgesthe external information gap, may well take the form ofan endogenous oscillatory process or networks of neu-rons providing interval-based representations. However,an alternative view to clock/timer like mechanisms is theidea that temporal information is embedded within thestructure of a changing event. By suggesting that time isrepresented by the means in which an event evolves overtime, one might hypothesise that a gap between twobeats is represented in the CNS by the way in which thefiring of neural structures changes across a given inter-beat interval. Here we present a model that proposeshow the closure of the temporal gap between two beats,the inter-beat interval, could be temporally specified in atau-like form.
491

The temporal guide (sg) information has beenmathematically described by Lee (1998) as
sg ¼ 1
2t � T 2
t
� �ð1Þ
where T is the inter-beat interval and t is the evolvingtime series within the inter-beat interval. In order to timea movement so that it coincides with the arrival of abeat, one would simply synchronise the way the move-ment changes over time (the time-to-closure of themovement gap, sm) onto this temporal information (thetime-to-sounding of the next beat, sg) in a constant ratiok, whose different values represent alternative kinematicforms of movement that will ensure the closure of thesame spatial gap. In effect, the temporal information (sg)generated within the nervous system guides the move-ment so that
sm ¼ ksg ð2Þ
The model predicts that an actor has incessant accessto this ever-changing temporal information provided bythe tau-guide, and that they control the closure of thegap (that is, the remaining distance, angle, pressure, andso on, to the desired goal, see also Lee 1998) by linkingtheir movement onto this continuously available dy-namic temporal information (see Craig et al 2000; Craigand Lee 1999; Lee et al 1999). This process, called tau-coupling, is a parsimonious solution to the problem ofsynchronising gap closure, where the tau of the move-ment gap (sm—the movement gap divided by its closurerate) is kept in constant proportion to the tau-guide (sg)so that sm=ksg.
In this study, participants were asked to synchronisetheir arrival in a target zone with the sounding of arhythmic beat. In other words, to perform the taskcorrectly, participants have to ‘‘know’’ how much time isleft before the next beat sounds so they can start movingahead of time in order to temporally synchronise theirarrival in the pre-designated target zone with the playingof a beat. That is, prior to moving, the participant isaware of how much time remains and adjusts the starttime of their movement accordingly. We propose thatthis information, which has to inform about the currentfuture, takes the form of a tau-guide and specifies thetime to closure of the temporal gap between beats. It ishypothesised that this tau-guide provides a temporalframework which participants use to control the closureof the spatial gap between where they are at a givenmoment in time and where they want to be in the nearfuture (the target zone). In other words, participants usethis tau-guide to control the tau of their movement,which specifies the time to closure of the spatial gapbetween the effector and the designated target. Thetheory hypothesises that by keeping the tau of themovement linearly coupled onto the tau-guide, bothgaps will be closed in a synchronous manner; or insimpler terms, both taus will reach zero simultaneously,indicating temporal concordance. If this were the case
we would expect a linear relationship, as indicated byhigh r2 values between the two taus.
Experiment 1
This experiment is different from other continuation andsynchronisation tapping paradigms. In tapping tasks,participants produce a tap by moving their finger in sucha way that it rebounds off a surface and produces asound that corresponds to the sounding of the next beatin a sequence of beats, either with the sound of the beatpresent (synchronisation) or absent (continuation para-digm), where participants reproduce the inter-beatintervals from memory alone. In this study, the task wasdesigned to resemble an interception task, where aclearly-defined spatial gap had to be closed in such a wayso as to arrive in the right place (a predefined targetzone) at the right time (when the beep or bop sounded).The aim of this task was therefore twofold: to syn-chronise the closure of a spatial gap (that is visuallyspecified) with an acoustic gap (that is specified by someinternal timing mechanism). The task therefore de-manded a high degree of both spatial and temporalprecision. Although tapping tasks likewise demand ahigh degree of temporal precision, the spatial location isless well defined, with several critics (for instance, Kolersand Brewster 1985) claiming that the participants maybe spatially synchronising another part of the movementto the beat, and that this may not correspond to the partof the movement that makes a sound when the fingerrebounds off the chosen surface. This lack of clarity isfurther emphasised in the temporal errors produced. Alarge number of studies (for example, Mates et al 1994;McAnally 2002) have reported asymmetric distributions(anticipation tendency) in temporal errors made whentapping, in other words an undershoot with respect tothe sounding of the beat. This discrepancy may simplybe due to a difference between what is being physicallydefined as being temporally synchronous (time of con-tact with a surface and the time of a beat) and what thesubject perceives as being temporally synchronous(the duration of their tapping sound and the duration ofthe beat) (Aschersleben and Prinz 1995).
Method
Participants
Eight right-handed non-musicians, average age 25(SD=3.2), participated in the study. Ethics approvalwas granted by the local ethics committee.
Apparatus
Two targets (1.5-cm wide, 1.8-cm long) 10 cm apartwere drawn on a piece of paper and placed under theplastic covering of a graphics tablet (Wacom UD 1218,
492
spatial resolution better than 0.25 mm). The width ofthe target zones and the distance between them werecalculated so as to correspond to an Index of Difficultyof 3 (a comfortable movement according to Fitts’ Law).Movements across the graphics tablet were made with agraphics’ tablet pen. The positional changes made by thetip of the pen were recorded at 100 Hz using a Macin-tosh G3.
The sounds (a beep and a bop—two tones of slightlydifferent pitch), which the participants had to interceptin the target zones, lasted 50 ms and were also producedby a Macintosh G3. The time that these beeps and bopswere sounded was recorded in parallel with the pointingdata. There were essentially two categories of sounds:those that were presented in a symmetrical manner (thesame time interval between a beep–bop and a bop–beep)and those presented in an asymmetrical manner (differ-ent time intervals between a beep–bop and a bop–beep).The sounds were presented cyclically (beep–bop–beep–bop), with the time interval being fixed at either 1.5/1.5or 2/2 s for the symmetrical sound presentation, and1.05/1.95 or 1.4/2.6 s for the asymmetrical sound pre-sentation. A 35:65 ratio between beats for the asym-metrical condition was chosen so as to preventparticipants from spontaneously producing naturalrhythms. Instead we wanted them to attend to the inter-beat intervals and really try and synchronise theirmovements to the beats provided.
These different presentation stimuli gave rise to fourdifferent experimental conditions. It should be notedthat, although the durations of the inter-beat intervalswere different, a complete beep–beep or bop–bop cyclewas either 3 s (symmetrical=1.5 + 1.5 s; asymmetri-cal=1.05 + 1.95 s) or 4 s (symmetrical=2 + 2 s;asymmetrical=1.4 + 2.6 s).
Procedure
Participants were told to move the pen back and forthbetween the targets on the graphics tablet in such a waythat the arrival and stopping of the pen in the targetzone was synchronised with the playing of the sound (seeFig. 1A). At the start of each trial, participants weregiven a period of familiarisation (15 pointing move-ments) during which they could adjust their movementso that it coincided with the sounding of the beeps andbops. After this period recording was started.
A total of 20 beep interceptions in the target on oneside and 20 bop interceptions in the target on the otherside (40 pointing movements) were recorded for allparticipants in each of the four sound conditions (twosymmetrical and two asymmetrical). The presentation ofthe experimental conditions (the time intervals) wascounter-balanced between participants. The recordedmovement data from the graphics tablet were subse-quently filtered using a Gaussian filter with a cut-off of
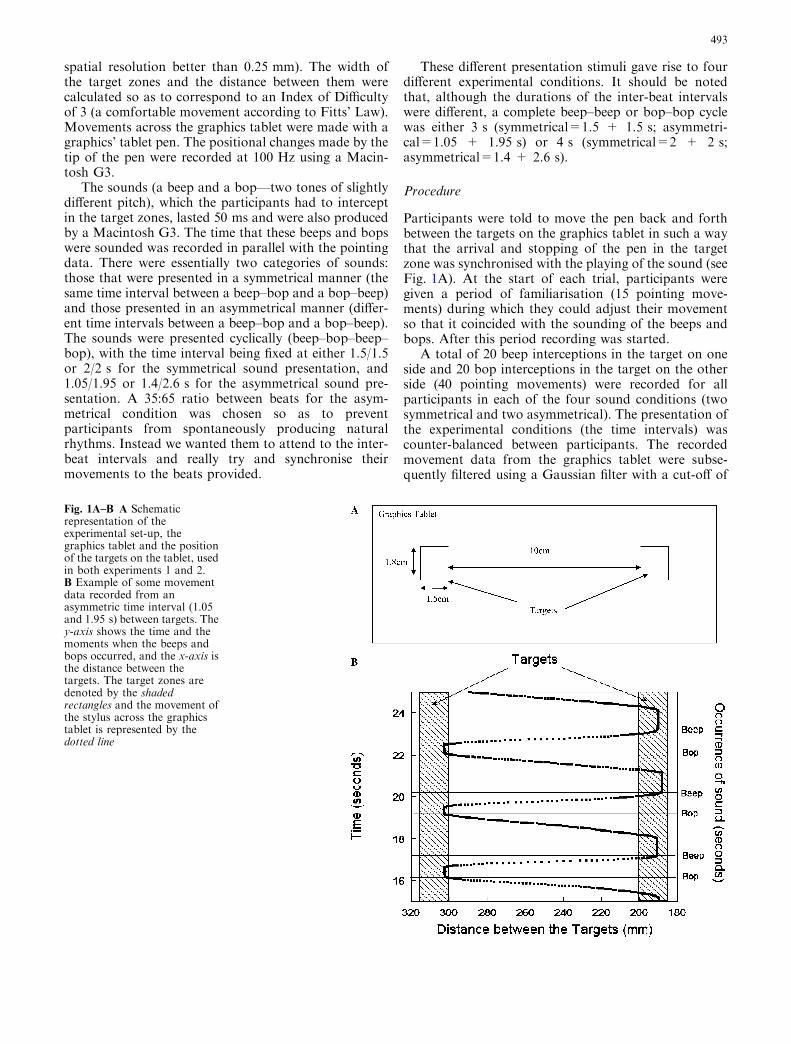
Fig. 1A–B A Schematicrepresentation of theexperimental set-up, thegraphics tablet and the positionof the targets on the tablet, usedin both experiments 1 and 2.B Example of some movementdata recorded from anasymmetric time interval (1.05and 1.95 s) between targets. They-axis shows the time and themoments when the beeps andbops occurred, and the x-axis isthe distance between thetargets. The target zones aredenoted by the shadedrectangles and the movement ofthe stylus across the graphicstablet is represented by thedotted line
493
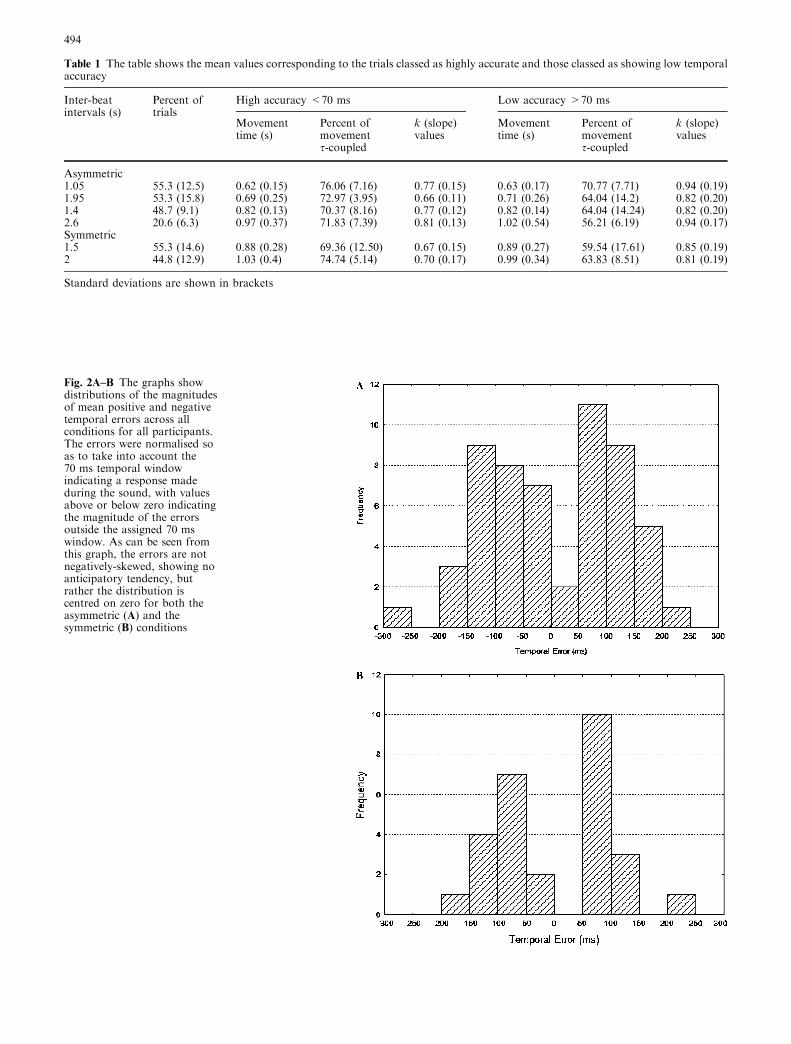
Fig. 2A–B The graphs showdistributions of the magnitudesof mean positive and negativetemporal errors across allconditions for all participants.The errors were normalised soas to take into account the70 ms temporal windowindicating a response madeduring the sound, with valuesabove or below zero indicatingthe magnitude of the errorsoutside the assigned 70 mswindow. As can be seen fromthis graph, the errors are notnegatively-skewed, showing noanticipatory tendency, butrather the distribution iscentred on zero for both theasymmetric (A) and thesymmetric (B) conditions
Table 1 The table shows the mean values corresponding to the trials classed as highly accurate and those classed as showing low temporalaccuracy
Inter-beatintervals (s)
Percent oftrials
High accuracy <70 ms Low accuracy >70 ms
Movementtime (s)
Percent ofmovements-coupled
k (slope)values
Movementtime (s)
Percent ofmovements-coupled
k (slope)values
Asymmetric1.05 55.3 (12.5) 0.62 (0.15) 76.06 (7.16) 0.77 (0.15) 0.63 (0.17) 70.77 (7.71) 0.94 (0.19)1.95 53.3 (15.8) 0.69 (0.25) 72.97 (3.95) 0.66 (0.11) 0.71 (0.26) 64.04 (14.2) 0.82 (0.20)1.4 48.7 (9.1) 0.82 (0.13) 70.37 (8.16) 0.77 (0.12) 0.82 (0.14) 64.04 (14.24) 0.82 (0.20)2.6 20.6 (6.3) 0.97 (0.37) 71.83 (7.39) 0.81 (0.13) 1.02 (0.54) 56.21 (6.19) 0.94 (0.17)Symmetric1.5 55.3 (14.6) 0.88 (0.28) 69.36 (12.50) 0.67 (0.15) 0.89 (0.27) 59.54 (17.61) 0.85 (0.19)2 44.8 (12.9) 1.03 (0.4) 74.74 (5.14) 0.70 (0.17) 0.99 (0.34) 63.83 (8.51) 0.81 (0.19)
Standard deviations are shown in brackets
494
9 Hz, and the velocity of the movement calculated usingfinite differences.
Results
It should be emphasised at this point that this experi-ment was deliberately intended to break from traditionaltapping (both continuation and synchronisation)experimental protocols. The reasons for this werethreefold: (1) traditional tapping studies often do notspecify a specific target location for the effector when thebeat sounds, which makes synchronisation errors diffi-cult to measure (Kolers and Brewster 1985), (2) syn-chronisation is taken to be the moment of beat onseteven though a beat may have a duration of 50 ms(Aschersleben and Prinz 1995) and (3) movementamplitudes in tapping studies are often very small or notrecorded. To examine in detail the link between move-ment coordination and beat onset, we asked participantsto synchronise their pointing movements to clearly de-fined spatial targets (10 cm apart) with the sounding ofan acoustic beat. The aim of this task was, therefore, tosynchronise the closure of a spatial gap specified visu-ally, with an acoustic gap, specified intrinsically by atiming mechanism.
Spatial–temporal accuracy
To be spatially accurate, participants had to have thepen positioned in the target zone when the beep orbop sounded. Participants’ performance satisfied thesespatial task requirements extremely well; all partici-pants in all conditions arrived in the required targetzone (see Fig. 1B for example). Given the low index ofdifficulty of the task (according to Fitts’ Law; largetarget width with respect to the inter-target distanceand large temporal window in which the movementhas to take place), this was expected.
Spatio-temporal accuracy was defined with respect tothe velocity of the pen movement. For each pointingmovement, start and end times were defined as the timesat which the velocity of the pen rose above 5% of peakvelocity and when it dropped below 5% of peak velocity,respectively, for a given movement. The temporal win-dow, within which there could have been coincidencebetween the arrival of the pen and the beat, was 70 ms(the duration of the sound, 50 ms, plus 10 ms samplingerrors for the apparatus—before and after the beat).Participants satisfied these stringent spatio-temporaltask requirements in about 50% of the trials (seeTable 1). A closer look at the data shows that the 2.6 scondition produced a significantly lower percentage ofcorrect responses (mean=20.6%). An analysis of vari-ance yielded a significant effect for the time intervalcondition [F(5,36)=4.57, p<0.003] with Tukey HSD posthoc test revealing a significant difference between the2.6 s condition and all the other time interval conditions
(p<0.05). While participants were able to achieve a highlevel of synchronisation between the spatial/temporalcontrol of their movements and the rhythm being im-posed by the beats, the 2.6 s interval appears to be muchmore difficult in comparison to the other time intervalconditions.
An analysis of temporal errors was undertaken tosee if there was a tendency to make anticipatory re-sponses in line with previous findings (Mates et al1994; McAnally 2002). All errors outside the 70-mswindow were normalised (temporal error ±70) so thatthose falling just outside the window corresponded toerrors just above or below zero. This normalisationmeant that any errors less than zero corresponded to apremature arrival in the target zone, and any valuesgreater than zero corresponded to a late arrival in thetarget zone. From Fig. 2, it can be clearly seen that themean positive and negative errors across conditions forall participants did not yield the same negative asym-metry or anticipatory tendency as previously observedin other tapping studies (Mates et al 1994; McAnally2002). In fact, when comparing the number of timesthe participants were early or late, they were late asignificantly greater number of times than they wereearly [t(82)=3.49, p<0.001]. This again suggests thatthere are some fundamental differences in the way thatthis task is executed and in the way a traditionaltapping study is carried out.
Movement organisation
This part of the analysis deals only with the trials thatwere considered to be highly accurate. The results of asimilar analysis for the less accurate trials will be pre-sented at the end of the ‘‘Results’’ section.
Using the start and the end times for each pointingmovement (calculated according to the criteria outlined
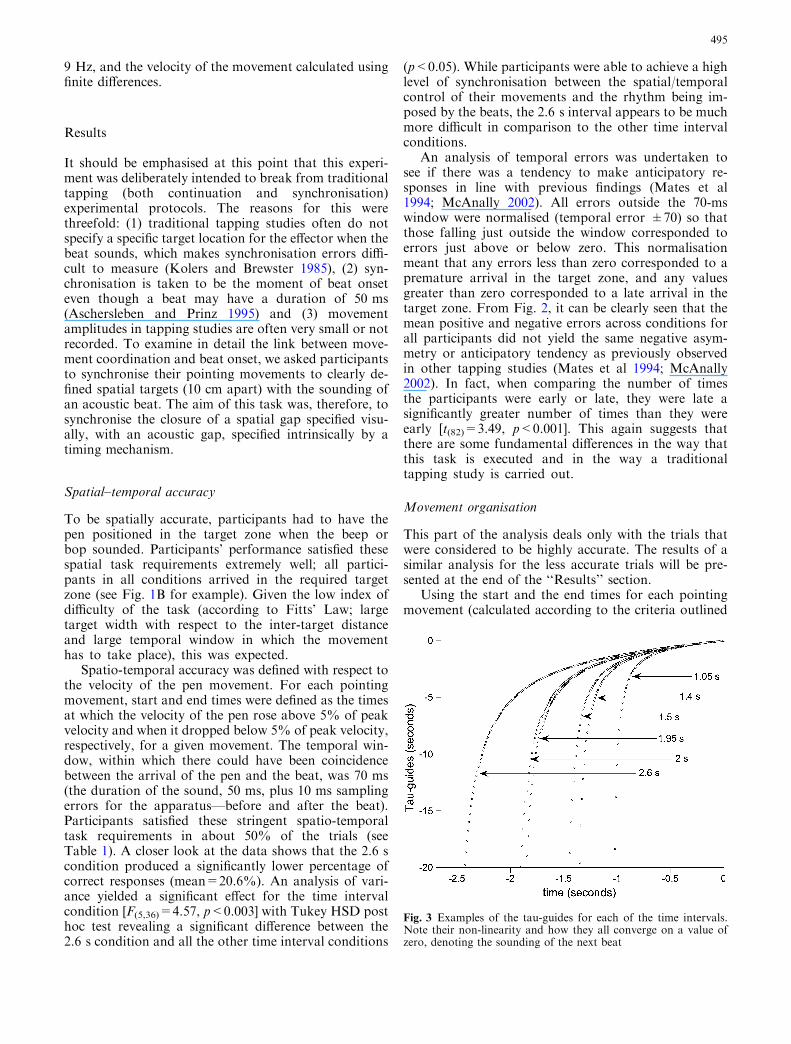
Fig. 3 Examples of the tau-guides for each of the time intervals.Note their non-linearity and how they all converge on a value ofzero, denoting the sounding of the next beat
495
above), waiting times (time between the beat beingsounded and movement initiation), and movement times(time between movement initiation and movement end)were calculated. Given that we asked participants to usea metronome to guide the timing of their movements,one might think that the simplest way of organisingone’s movement would be to move in a cyclical mannerfor the total duration of the time between the beeps andthe bops. However, looking more closely at the organi-sation of the movements during the specified timeintervals (the time between a beep and a bop and viceversa), it appears that the time needed to cover the dis-tance between the two targets was smaller than the timeinterval allotted between the two acoustic signals. Par-ticipants typically waited in the target zone for a variableamount of time before initiating their movement (seeFig. 1B). A regression analysis carried out on the wait-ing times and inter-beat intervals revealed that thewaiting time increased linearly across the different timeintervals (r2=0.91).
To test whether participants were simply keepingmovement times constant, a one-way analysis of vari-ance was performed. A significant main effect of move-ment times was found for the different inter-beatintervals [F(5,30)=3.81, p<0.01] with a Tukey HSD posthoc test revealing significant differences between the1.05 s (mean movement time=0.62) and the 2 s (meanmovement time=1.03) and 2.6 s (mean movementtime=0.97) time intervals (p<0.05).
The finding that participants only move for a certainpart of the inter-beat interval goes against the idea of
controlling the movement by simply setting the move-ment time so that it directly corresponds to the length ofthe inter-beat interval. On the other hand, the observedbehaviour may be explained by the tau-coupling theory,since it proposes that participants are being continu-ously informed by a tau-guide, a dynamic temporalrepresentation of the inter-beat interval, of the timeremaining until the next beat sounds. This type ofinformation would allow participants to determine howlong they should wait in the target zone. The actor cansimply use this information as a temporal guide in orderto control the unfolding of their movement so that theystop in the target zone when the next beat sounds.
Timing information
To test the extent to which participants followed theprinciples of tau guidance, sm=ksg, tau-guides werecalculated (dynamically representing the time intervalbetween a beep and a bop) for each of the different timeintervals (see Fig. 3). In addition, for the closure of eachmovement gap (distance between the targets) a move-ment tau was calculated by dividing current gap size (themovement amplitude at any given moment) by currentvelocity of gap closure (the velocity of the movement atthat time) or x= _x where x is the current distance to thenext target and _x its derivative.
To assess the degree of coupling between the two tau-variables, the tau of the movement gap was linearly re-gressed onto the tau-guide (see Fig. 4) for each trial. We
k = 0.38
R2 = 0.9581% Movement
-7
-6
-5
-4
-3
-2
-1
0-3 -2 -1 0
Tau guide (s)
Tau M
ovement (s)
-120
-100
-80
-60
-40
-20
0
0 0.2 0.4 0.6 0.8
Time (s)
Pos
ition
(m
m)
0
50
100
150
200
250
Velocity (m
m/s)
-120
-100
-80
-60
-40
-20
0
0 0.1 0.2 0.3 0.4 0.5
Time (s)
Po
sitio
n (m
m)
0
50
100
150
200
250
300
350
400
Velocity (m
m/s)
k = 0.41
R2 = 0.9638% Movement
-4.5
-4
-3.5
-3
-2.5
-2
-1.5
-1
-0.5
0-0.8 -0.6 -0.4 -0.2 0
Tau guide (s)T
au Movem
ent (s)A BFig. 4A–B A Examples of the
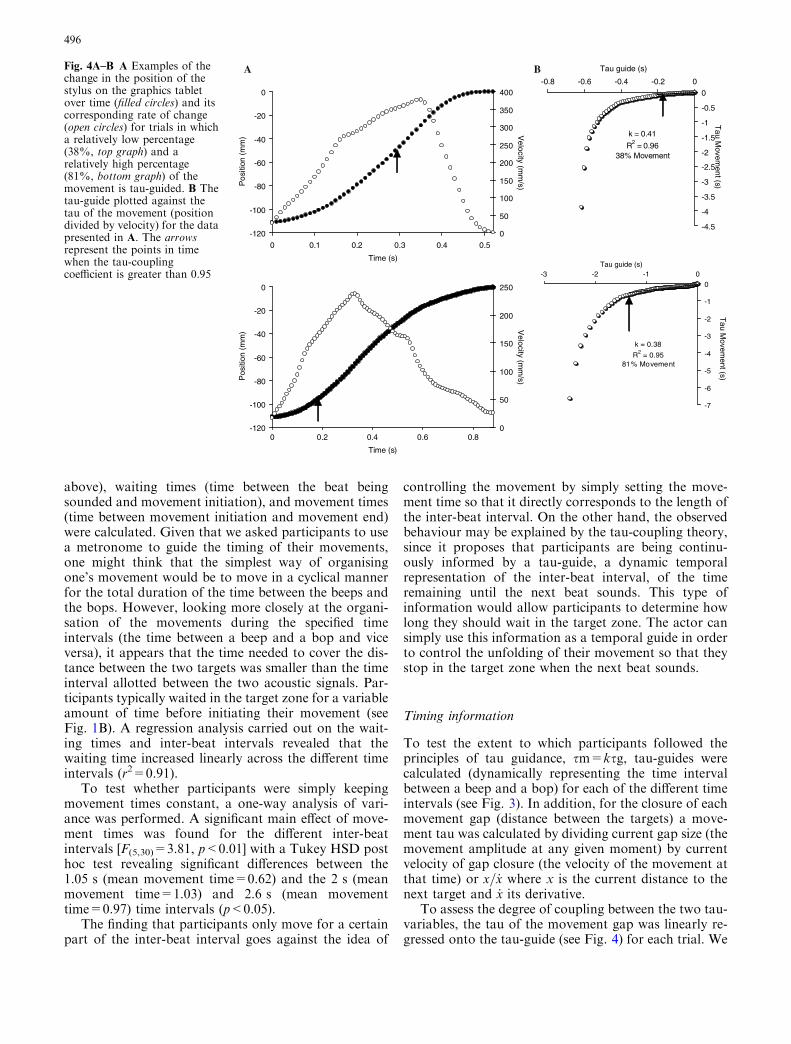
change in the position of thestylus on the graphics tabletover time (filled circles) and itscorresponding rate of change(open circles) for trials in whicha relatively low percentage(38%, top graph) and arelatively high percentage(81%, bottom graph) of themovement is tau-guided. B Thetau-guide plotted against thetau of the movement (positiondivided by velocity) for the datapresented in A. The arrowsrepresent the points in timewhen the tau-couplingcoefficient is greater than 0.95
496
then determined for how much of the movement the twotau-variables were linearly related such that the tau-coupling model would explain more than 95% of thevariability. The degree of coupling between two taus wasmeasured by running a recursive linear regression algo-rithm. This successively computed the r2 when 0,1,2...data points were removed from the beginning of the dataseries. The algorithm stopped when r2 first exceeded0.95, and the current value of r2 and the current per-centage of data points in the regression were both re-corded. Although the value 0.95 is rather arbitrary,Table 1 shows a summary of the mean percentage ofmovement times where the two taus are coupledaccording to these criteria. As can be seen from Table 1,for more than 70% of the movement there is a stronglinear relationship or coupling between the tau-guide(the intrinsic informational variable) and the taumovement (the movement variable).
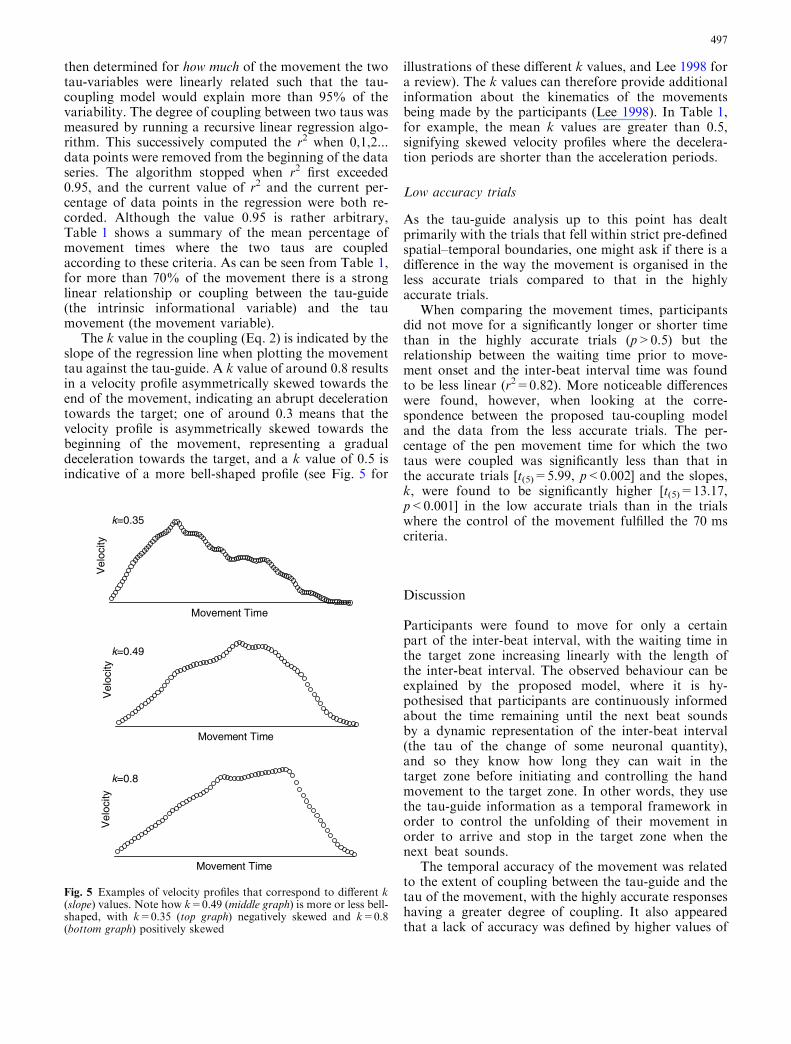
The k value in the coupling (Eq. 2) is indicated by theslope of the regression line when plotting the movementtau against the tau-guide. A k value of around 0.8 resultsin a velocity profile asymmetrically skewed towards theend of the movement, indicating an abrupt decelerationtowards the target; one of around 0.3 means that thevelocity profile is asymmetrically skewed towards thebeginning of the movement, representing a gradualdeceleration towards the target, and a k value of 0.5 isindicative of a more bell-shaped profile (see Fig. 5 for
illustrations of these different k values, and Lee 1998 fora review). The k values can therefore provide additionalinformation about the kinematics of the movementsbeing made by the participants (Lee 1998). In Table 1,for example, the mean k values are greater than 0.5,signifying skewed velocity profiles where the decelera-tion periods are shorter than the acceleration periods.
Low accuracy trials
As the tau-guide analysis up to this point has dealtprimarily with the trials that fell within strict pre-definedspatial–temporal boundaries, one might ask if there is adifference in the way the movement is organised in theless accurate trials compared to that in the highlyaccurate trials.
When comparing the movement times, participantsdid not move for a significantly longer or shorter timethan in the highly accurate trials (p>0.5) but therelationship between the waiting time prior to move-ment onset and the inter-beat interval time was foundto be less linear (r2=0.82). More noticeable differenceswere found, however, when looking at the corre-spondence between the proposed tau-coupling modeland the data from the less accurate trials. The per-centage of the pen movement time for which the twotaus were coupled was significantly less than that inthe accurate trials [t(5)=5.99, p<0.002] and the slopes,k, were found to be significantly higher [t(5)=13.17,p<0.001] in the low accurate trials than in the trialswhere the control of the movement fulfilled the 70 mscriteria.
Discussion
Participants were found to move for only a certainpart of the inter-beat interval, with the waiting time inthe target zone increasing linearly with the length ofthe inter-beat interval. The observed behaviour can beexplained by the proposed model, where it is hy-pothesised that participants are continuously informedabout the time remaining until the next beat soundsby a dynamic representation of the inter-beat interval(the tau of the change of some neuronal quantity),and so they know how long they can wait in thetarget zone before initiating and controlling the handmovement to the target zone. In other words, they usethe tau-guide information as a temporal framework inorder to control the unfolding of their movement inorder to arrive and stop in the target zone when thenext beat sounds.
The temporal accuracy of the movement was relatedto the extent of coupling between the tau-guide and thetau of the movement, with the highly accurate responseshaving a greater degree of coupling. It also appearedthat a lack of accuracy was defined by higher values of
Movement Time
Vel
ocity
k=0.35
k=0.49
k=0.8
Movement Time
Vel
ocity
Movement Time
Vel
ocity
Fig. 5 Examples of velocity profiles that correspond to different k(slope) values. Note how k=0.49 (middle graph) is more or less bell-shaped, with k=0.35 (top graph) negatively skewed and k=0.8(bottom graph) positively skewed
497

the coupling constant k, suggesting that abrupt brakingstrategies could be a reflection of, or responsible for,increased error.
Experiment 2
The previous experiment showed that participants werequite adept at coordinating their movements so thattheir arrival in a spatially-defined target zone coincidedwith the sounding of an acoustic beat. The analysis did,however, indicate that overall accuracy fell off for the2.6-s inter-beat interval. Other studies that have lookedat the temporal asynchrony between the production of atap and the sounding of a beat have also shown that, atintervals shorter than 2.5 s, participants can relativelyaccurately anticipate the arrival of a beat. However, atlarger inter-beat intervals (>2.5 s), temporal variabilityin tap production increases dramatically, with partici-pants reacting after the arrival of the acoustic stimulusrather than anticipating it (Szelag et al 2002). Thisfinding may be explained by the idea that internal timingmechanisms have an upper inter-beat time interval limit,beyond which it becomes very difficult to anticipate thearrival of the next beat (Peters 1989; Mates et al 1994). Itshould be noted that the amplitudes of the movementsmade in many of the previous studies were relativelysmall (around 1 cm). Furthermore, no analysis of thekinematics of the movements was carried out. The aimof this second experiment is to further explore this no-tion of limitations in intrinsic timing mechanisms byexamining effects of increased inter-beat intervals onmovement kinematics. Moreover, we wanted to investi-gate whether the tau-coupling model proposed can ex-plain any possible effects.
Method
Participants
Seven non-musicians (average age 24, SD=2.6) who didnot participate in Experiment 1 participated in Experi-ment 2.
Apparatus
The same pointing apparatus that was used in the pre-vious experiment was again implemented here. In orderto see if the lower percentage of tau-coupling observedfor the 2.6 s time interval was due to the asymmetricnature of the acoustic stimuli presentation, the inter-beattime intervals in this experiment were all symmetrical.Intervals of 1.5, 3 or 4.5 s were chosen, with the threedifferent time interval conditions being presented onthree separate occasions. The inter-beat interval thatparticipants began with was randomly chosen.
Procedure
As before, the aim of the task was to move the penbetween the targets in such a way so as to arrive andstop in one of the target zones at the same time as thebeat sounded. Recording began after participants hadcompleted a familiarisation period of 20 pointingmovements and was stopped when participants hadcompleted a further 60 movements back and forth be-tween the targets. Positional changes of the graphics penacross the tablet were smoothed using a Gaussian filterwith a cut-off of 9 Hz, as in Experiment 1, and thevelocity of the movement was calculated using finitedifferences.
Results
Spatial–temporal accuracy
To assess the spatial–temporal accuracy of the partici-pants’ movements, we had to ascertain whether theparticipants had come to a stop in the target zone whenthe beat sounded. First, positional changes of thegraphics tablet pen were aligned with the time series dataindicating the moment the beat started to sound. Thetemporal window in which the movement was classifiedas being synchronised with the beat was again taken tobe 70 ms (50 ms beat duration ± 10 ms sampling error).Next, the velocity of the graphics tablet pen was exam-ined. The movement of the graphics tablet pen wasjudged to have stopped if the velocity was less than 5%of the value of peak velocity at some moment during the70 ms temporal window. All the pointing movementsthat fulfilled these criteria (movement velocity at lessthan 5% of peak velocity within the 70 ms beat temporalwindow) were classified as being accurate.
Although Table 2 shows that the average percentageof highly accurate responses for all the participants was,in general, not very high, a marked effect of the in-creased time interval between beats on accuracy wasnoted. A repeated measures ANOVA on these percent-ages for the three different inter-beat intervals confirmsthis observation [F(2,18)=11.17, p £ 0.001]. A Tukey
Table 2 The mean percentage of correct movements, the move-ment time, the percentage of movement s-coupled and the coeffi-cient k for all seven participants in each of the different inter-beatintervals
Timeinterval(s)
Percent oftrials within70 ms criteria
Movementdurationtime (s)
Percent ofmovement times-coupled
k (slope)values
1.5 40 (12) 1.01 (0.29) 63.2 (14.8) 0.64 (0.16)3 23 (16) 1.89 (0.87) 47.6 (23.8) 0.70 (0.13)4.5 11 (4) 2.97 (1.41) 30.4 (24.5) 0.82 (0.10)
The standard deviations of the mean values are indicated inbrackets
498

HSD post hoc test shows a significant difference betweenthe 1.5 and 4.5 s intervals (p<0.001) and the 1.5 and 3 sintervals (p<0.005).
Movement kinematics
To understand why such a marked decrease in thenumber of highly accurate trials was observed, welooked more closely at the kinematics of the pointingmovements. Inspection of the average movement timesfor each subject in each condition indicated that par-ticipants took longer to move between the targets whenthe time interval increases, even though the distancebetween the targets did not change. A repeated measuresANOVA showed that the effect of the length of the in-ter-beat interval on the movement times was significant[F(2,18)=7.12, p<0.005]. More specifically, a post hoctest (Tukey HSD test) showed that there were significantdifferences in movement times between the 1.5 and the4.5 s inter-beat time intervals (p<0.05). Not surpris-ingly, this increase in movement time entrains a sig-nificant decrease in the peak velocity as the inter-beatintervals increased [F =4.29, p<0.05], with differencesagain being found between the 1.5 and 4.5 s intervals(Tukey HSD, p<0.05).
These kinematic differences between the movementsfor the different inter-beat intervals become veryapparent when one looks at the phase planes and Hookeportraits (see Fig. 6). A phase plane shows the rela-tionship between the movement velocity and positionalchanges where a perfect sinusoidal movement is char-acterised by a circle. The phase planes for unconstrainedmovements that are prospectively guided and have asmooth single peak velocity profile usually have a cir-cular-type form (with half a circle corresponding to asingle reaching movement). Note that this is not to saythat such a movement is simply harmonic; there is noevidence that natural movements can be characterised assuch. However, the phase plane representation can beused to get more insight into the extent to which amovement is prospectively guided and controlled. Ascan be seen from Fig. 6, the phase plane for the 1.5 scondition has a rounded appearance. However, as theinter-beat interval increases, the forms of the velocity-position plots progressively flatten, indicating a drop inthe velocity. This effect is even more pronounced for the4.5 s interval, where the phase planes appear more likeflattened ellipses than circles.
These kinematic differences are also mirrored in theHooke portraits, which show change in acceleration as afunction of positional change, and are sloped straightlines when the movement is sinusoidal. As noted earlier,in response to the specific instructions given to partici-pants (to arrive and stop in the target zone at the sametime as the beat sounds), participants spent a certainpercentage of the inter-beat interval waiting prior tomovement onset. This behaviour is evidenced in theHooke portraits for the 1.5 s condition, which clearly
illustrate a deceleration period when the subject enters atarget zone and an acceleration period when the subjectleaves a target zone. As the time interval increased, therewas much less of a distinction between the behaviouroutside and inside the target zones. The slope of the linecontinually decreases until it becomes very flat in the 4.5-s inter-beat interval condition.
Tau-coupling
If these differences were due to limitations in theintrinsic timing mechanisms we would expect to seedifferences in the parameters of the tau-coupling modelproposed to account for this behaviour.
As previously proposed, the hypothesised tau-guide(sg in Eq. 1) specifies the temporal gap of an inter-beatinterval and continually informs the subject of the timeremaining until the next beat sounds. It is hypothesisedthat in order to successfully control the movement sothat it spatially and temporally coincides with thesounding of the next beat, participants should couple thetau of their movements (sm—the time to target gapclosure at its current closure rate) onto the intrinsictemporal tau-guide (sg).
The model was tested by calculating the percentage ofmovement time for which the movement was coupledonto the intrinsic tau-guide, as in Experiment 1. Only thepointing movements that were considered to be accuratewere analysed so that a direct comparison between thetwo experiments could be made. As can be seen fromTable 2, there is a marked decrease in the percentage ofthe movement time that is coupled onto the guide whenthe inter-beat interval increases. Likewise, the couplingcoefficient, k, is affected by the increased time intervalsbetween beats (see Table 2). This increase in the value ofk signifies an abrupt deceleration at the end of themovement (Lee 1998).
Discussion
The results of this experiment clearly show that the abil-ities of participants to accurately guide and synchronisetheir movements to inter-beat intervals varies as theduration of the interval increases. Of the three experi-mental conditions (1.5, 3 and 4.5 s inter-beat intervals),participants could best synchronise their movements tothe 1.5-s interval. These findings are in accordance withprevious experiments, which have shown that inter-beatintervals above 2.5 s give rise to a much greater temporalasynchrony between the tap produced by the subject andthe sounding of the beat (Mates et al 1994; Peters 1989;Szelag et al 2002). Experiment 1,where participants had tomove between target zones to intercept acoustic beats,also shows that synchronising movements to the 2.6 sinter-beat interval is much more difficult than the otherfive inter-beat intervals used in the experiment (1.05, 1.4,1.5, 1.95 and 2 s).
499
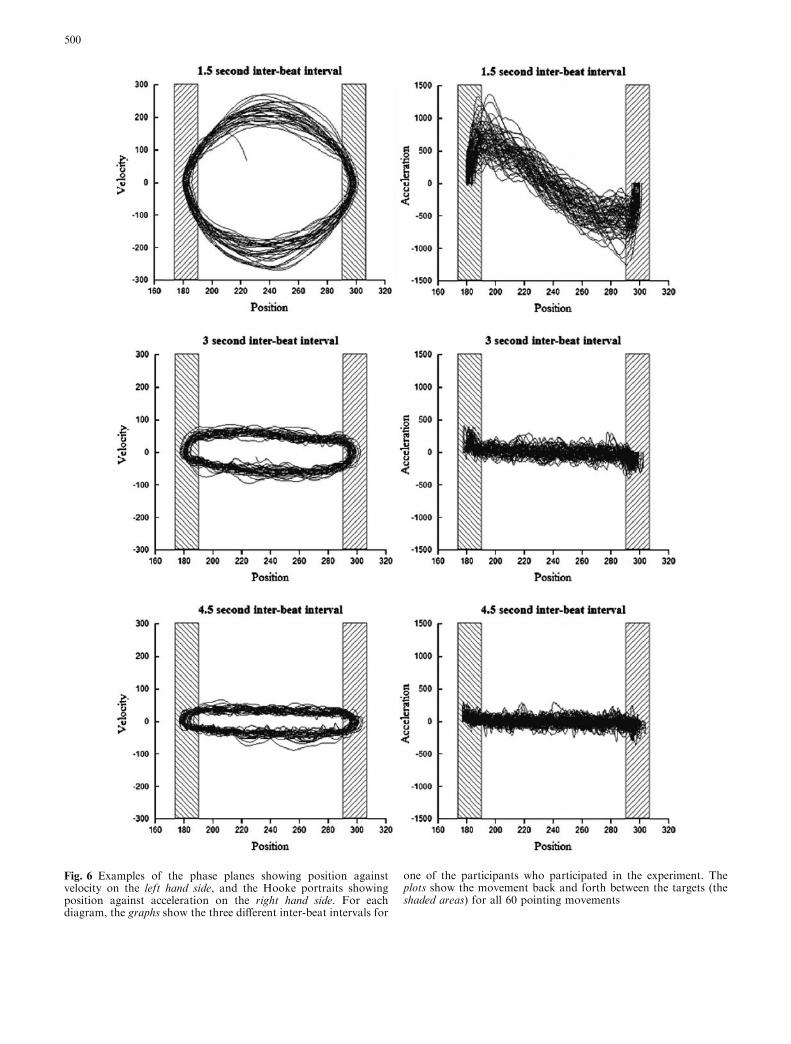
Fig. 6 Examples of the phase planes showing position againstvelocity on the left hand side, and the Hooke portraits showingposition against acceleration on the right hand side. For eachdiagram, the graphs show the three different inter-beat intervals for
one of the participants who participated in the experiment. Theplots show the movement back and forth between the targets (theshaded areas) for all 60 pointing movements
500

Another curious finding in this study concerns theorganisation of the movement. Even though the distancebetween the targets did not change, participants tooklonger to move between the targets when the timeinterval increased. It would appear that participantswere unsure of when the beat was going to arrive andhence moved more slowly, estimating, rather than pro-spectively anticipating, when to arrive in the target zonewhen the beats sounded.
These results support the idea of an internal timingmechanism with a limited capacity. If, as the tau-guidehypothesis proposes, the duration of an inter-beatinterval is represented by the way in which some neuralquantity discharges over time, then the upper limit mustbe linked to the limitations in the neural structuresrepresenting the temporal interval. Intervals that surpassthe 2.5/3-s barrier can no longer be represented accu-rately and hence the unfolding of the movement overtime cannot be coupled onto the temporal guide speci-fying the time remaining until the next beat. Thebreakdown in coupling between the two means partici-pants can no longer synchronise their movements to thebeats.
General discussion
To successfully intercept an acoustic beat in a giventarget zone, one has to begin one’s actions sufficientlyahead of time. The spatial gap between the current po-sition of the hand and the target zone has to be closed insuch a way that the hand stops in the target zone at thesame time as the beat is sounded. How do participantsknow when the next beat is going to arrive? The modelpresented in this study proposes that the temporal gapbetween two successive acoustic beats is represented inthe form of a ‘‘tau-guide’’, a dynamic neural represen-tation that prospectively informs about the timeremaining until the next beat. By linearly coupling thetau of the movement (the rate of closure of the hand/target gap at its current closure rate) onto the tau-guide,both will reach zero simultaneously and will ensure thatthe participant’s hand arrives in the right place (thetarget zone) at the right time (when the next beat issounded) (see Fig. 7). The high percentage of the vari-ance that is explained by the linear regression betweenthe tau of the hand movement and the tau-guide, alongwith the high percentage of the movement duration thatseems to be tau-coupled, lend support to this model.
Additional support is provided by the way in whichthe participants organise their hand movements: theyonly move for a certain part of the inter-beat interval.This would go against the idea of controlling themovement by simply coupling together two oscillators,one for the inter-beat interval and one for the handmovement. In line with the proposed model, participantswould know how long to wait in the target zone as thedynamic representation of the inter-beat interval, the tauof the change of some neuronal quantity, would reliably
provide the necessary prospective information about thetime remaining until the next beat arrives. The subjectcould simply use this information as a temporal guide soas to control the unfolding of their movement so thatthey stop in the target zone when the next beat sounds.
Cariani (1999) proposes that when participants per-form a rhythm continuation task (continuing to tap outan inter-beat interval that is no longer presented to theparticipants) they construct a neural temporal dischargethat becomes a reverberating memory of the temporalpattern. He suggests that participants use this memorytrace to trigger motor systems, which control the timingof the tap. Although he does not elaborate on how thereverberating memory trace is coded and how themovement is then controlled or initiated, he does suggestthat there are some common underlying invariantsconcerning the temporal structure in the perceptual andmotor domains. Cariani (1999) proposes that thesecommon time structures for both sensory coding andmotor control may help explain the integration of per-ception and action. Georgopoulos (2002) also arguesthat the control of action requires the merging of sepa-rate processes that share common general principles forextracting information in space and executing tasks intime. He suggests that tau is a good candidate for ageneral timing principle.
Evidence supporting this idea of common neuralstructures for perception and action has also been pro-vided by other studies (Ivry 1996). Recent fMRI exper-iments have shown that there is a common cortical andsub-cortical basis for time perception and motor timing,with the premotor cortex being involved in sequencingand timing activities that are independent of the execu-tion of movement (Schubotz et al 2000). To have tem-poral coincidence between tasks it is essential to have alink between motor timing and time perception. Theproblems that Parkinson’s patients have in tapping outtemporal intervals in continuation paradigms (O’Boyleet al 1996) and estimating the duration of differentintervals (Pastor et al 1992) suggests that the basalganglia may have a role to play in the representation oftemporal information.
In trying to understand the link between music andmovements, Cooper and Meyer (1960) talked about abeat sense that ‘‘tends to be continued in the mind andmusculature of the listener’’ even after the rhythm hasstopped. This description suggests that the beat formssome sort of dynamic temporal imprint in both the mindand the muscles, which likewise infers some commontiming mechanism. Recent research on music therapy hasshown that patient populations that normally have dif-ficulty controlling or coordinating self-paced movements(movements made in the absence of any extrinsically-provided perceptual information) improve the overallspatio-temporal characteristics of their movement whenthey try to move in time to an acoustic stimulus (Thautet al 1999; McIntosh et al 1997). In this case it wouldappear that this so-called ‘‘sensory cueing’’ provided bythe extrinsic aid, namely the acoustic beats of the met-
501
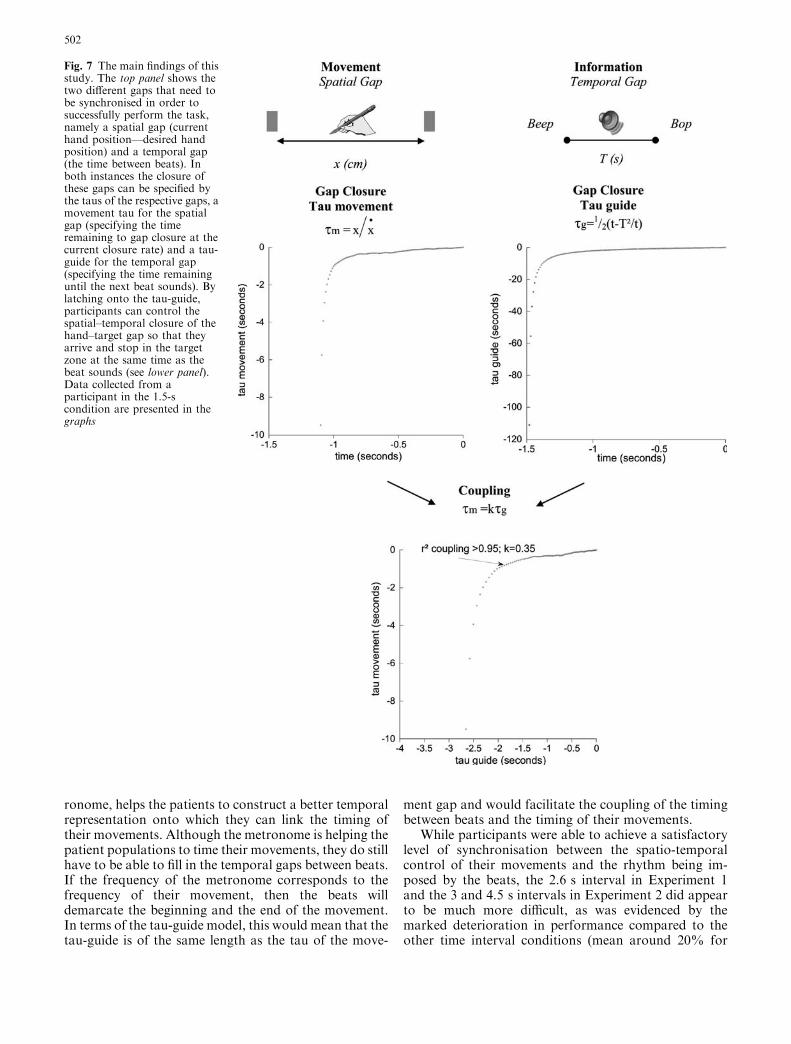
ronome, helps the patients to construct a better temporalrepresentation onto which they can link the timing oftheir movements. Although the metronome is helping thepatient populations to time their movements, they do stillhave to be able to fill in the temporal gaps between beats.If the frequency of the metronome corresponds to thefrequency of their movement, then the beats willdemarcate the beginning and the end of the movement.In terms of the tau-guide model, this would mean that thetau-guide is of the same length as the tau of the move-
ment gap and would facilitate the coupling of the timingbetween beats and the timing of their movements.
While participants were able to achieve a satisfactorylevel of synchronisation between the spatio-temporalcontrol of their movements and the rhythm being im-posed by the beats, the 2.6 s interval in Experiment 1and the 3 and 4.5 s intervals in Experiment 2 did appearto be much more difficult, as was evidenced by themarked deterioration in performance compared to theother time interval conditions (mean around 20% for
Fig. 7 The main findings of thisstudy. The top panel shows thetwo different gaps that need tobe synchronised in order tosuccessfully perform the task,namely a spatial gap (currenthand position—desired handposition) and a temporal gap(the time between beats). Inboth instances the closure ofthese gaps can be specified bythe taus of the respective gaps, amovement tau for the spatialgap (specifying the timeremaining to gap closure at thecurrent closure rate) and a tau-guide for the temporal gap(specifying the time remaininguntil the next beat sounds). Bylatching onto the tau-guide,participants can control thespatial–temporal closure of thehand–target gap so that theyarrive and stop in the targetzone at the same time as thebeat sounds (see lower panel).Data collected from aparticipant in the 1.5-scondition are presented in thegraphs
502

the 2.6 and 3 s intervals and 11% for the 4.5 s interval,as opposed to 50% for all the other conditions). Thisfinding is interesting and in accordance with severalstudies that have shown that the ability to accuratelysynchronise taps to inter-beat intervals is restricted totime intervals of less than 2.5 s (Mates et al 1994; Peters1989). These studies, along with our results, suggest thatthe way in which an inter-beat interval is intrinsicallyrepresented is limited by the way the neural structurescan physically determine this time period. If the repre-sentation of the time interval was indeed determined bythe way in which a neural quantity changed over time,this would suggest that the tau-guide has a maximumsize in terms of duration, determined by the physicalnature of the neural structures involved.
These apparent limits in time perception have beenaddressed before (Mates et al 1994; Peters 1989; Szelaget al 2002). Fraisse (1984) made an important distinctionbetween time perception and the estimation of time.Time perception, according to Fraisse, is related toJames’ (1950) description of the ‘‘specious present’’ (or‘‘feeling of nowness’’) and corresponds to time intervalsof 100 ms to a few seconds. The limits of this so-called‘‘feeling of nowness’’ are subjective. According to Fra-isse, time intervals beyond this boundary can no longerbe perceived and must be estimated, as shown by amarked decrease in temporal synchronicity betweenstimuli and participants’ anticipatory responses.
A possible solution to this inter-beat durationboundary may be found in music (Thaut et al 1997).Filling the informational void between beats with amelody has been shown to provide additional informa-tion onto which participants can couple their move-ments and hence reduce the magnitude of the temporalerror between the production of the tap (the movement)and the sounding of the beat.
In the present study, the surpassing of the 2.5/3 sboundary resulted in a significant decrease in the par-ticipants’ performance. This was evidenced by a break-down in the coupling between the information (thedynamic neural representation modelled by the tau-guide) and the movement of the hand between the tar-gets. It is suggested that this de-coupling, so to speak,could be attributed to the limitations in the neuralstructures implicated in the construction of or the use ofthe tau-guide.
To conclude, it would appear that, when moving tobeats, the timing for the movement is inextricably boundup in the time imposed by the inter-beat duration.Synchronisation of movements to beats requires pro-spective information about the time remaining until thenext beat sounds. Given that the beats mark the end ofthe movement, some intrinsic timing mechanism has tofill in the temporal gap. Instead of thinking of time inconstant units, as is the case for man-made time passagemarkers such as clocks, one should maybe think of therepresentation of time’s passage as the way that someneural quantity changes within that time interval. Thedynamics of that change could then denote the time
remaining until the next beat sounds and provide atemporal guide for the spatio-temporal unfolding of themovement.
References
Aschersleben G, Prinz W (1995) Synchronizing actions with events:the role of sensory information. Percept Psychophys 57(3):305–317
Cariani P (1999) Closing the loop: common temporal structure inneural substrates for perception and action. Paper presented atthe 9th Annual Meeting of the New England Sequencing andTiming (NEST) Workshop, Hunter Laboratory, Brown Uni-versity, Rhode Island
Cooper G, Meyer LB (1960) The rhythmic structure of music.University of Chicago Press, Chicago, IL
Craig CM, Lee DN (1999) Neonatal control of nutritive suckingpressure: evidence for an intrinsic tau-guide. Exp Brain Res124(3):371–382
Craig CM, Delay D, Grealy MA, Lee DN (2000) Guiding theswing in golf putting. Nature 405(6784):295–296
Fraisse P (1984) Perception and estimation of time. Annu RevPsychol 35:1–36
Georgopoulos AP (2002) Cognitive motor control: spatial andtemporal aspects. Curr Opin Neurobiol 12:678–683
Gibson JJ (1975) Events are perceivable but time is not. In: FraserJT, Lawrence N (eds) The study of time, 2. Springer, BerlinHeidelberg New York
Ivry RB (1996) The representation of temporal informationin perception and motor control. Curr Opin Neurobiol 6:851–857
James W (1950) The principles of psychology, vol 1. Dover Pub-lications (original work published 1890), New York
Kolers PA, Brewster JM (1985) Rhythms and responses. J ExpPsychol Hum Percept Perform 11(2):150–167
Lee DN (1998) Guiding movement by coupling taus. Ecol Psychol10:221–250
Lee DN, Craig CM, Grealy MA (1999) Sensory and intrinsiccoordination of movement. Proc R Soc Lond B Biol Sci266(1432):2029–2035
Lee DN, Georgopoulos AP, Clark MJO, Craig CM, Port NL(2001) Guiding contact by coupling the taus of gaps. Exp BrainRes 139(2):151–159
Mach E (1883) The science of mechanics: a critical and historicalexposition of its principles. Chicago University Press, Chicago,IL
Mates J, Muller U, Radil T, Poppel E (1994) Temporal integrationin sensorimotor synchronization. J Cogn Neurosci 6:332–340
McAnally KI (2002) Timing of finger tapping to frequency mod-ulated acoustic stimuli. Acta Psychol 109(3):331–338
McIntosh GC, Brown SH, Rice RR, Thaut MH (1997) Rhyth-mic auditory-motor facilitation of gait patterns in patientswith Parkinson’s disease. J Neurol Neurosurg Psychiatry62:22–26
O’Boyle DJ, Freeman JS, Cody FWJ (1996) The accuracy andprecision of timing of self-paced, repetitive movements in sub-jects with Parkinson’s disease. Brain 119:51–70
Pastor MA, Artieda J, Jahanshahi M, Obeso JA (1992) Timeestimation and reproduction is abnormal in Parkinson’s dis-ease. Brain 115:211–225
Peters M (1989) The relationship between variability of intertapintervals and interval duration. Psychol Res 51:38–42
Rosenbaum DA, Patashnik O (1980a) A mental clock settingprocess revealed by reaction times. In: Stelmach GE, Requin J(eds) Tutorials in motor behavior. North-Holland, Amsterdam,pp 487–499
Rosenbaum DA, Patashnik O (1980b) Time to time in the humanmotor system. In: Nickerson RS (ed) Attention and perfor-mance, vol 8. Erlbaum, Hillsdale, NJ
503

Schubotz RI, Friederici AD, von Cramon DY (2000) Time per-ception and motor timing: a common cortical and subcorticalbasis revealed by fMRI. Neuroimage 11(1):1–12
Szelag E, Kowalska J, Rymarczyk K, Poppel E (2002) Durationprocessing in children as determined by time reproduction:implications of a few seconds time window. Acta Psychol110:1–119
Thaut MH, McIntosh GC, Rice RR (1997) Rhythmic facilitationof gait training in hemiparetic stroke rehabilitation. J NeurolSci 151(2):207–212
Thaut MH, Kenyon GP, Schauer ML, McIntosh GC (1999) Theconnection between rhythmicity and brain function. IEEE EngMed Biol 18:101–108
TreismanM, Faulkner A,Naish PLN, BroganD (1990) The internalclock: evidence for a temporal oscillator underlying time per-ception with some estimates of its characteristic frequency. Per-ception 19:705–743
Wing AM (1980) The long and short of timing in response se-quences. In: Stelmach GE, Requin J (eds) Tutorials in motorbehavior. North-Holland, Amsterdam
Wing AM, Kristofferson AB (1973) Response delays and the tim-ing of discrete motor responses. Percept Psychophys 14:5–12
504
Related Documents









![Intercepting Licensing Rights: Why College Athletes Need a … · 2016. 5. 6. · 2012] INTERCEPTING LICENSING RIGHTS 87 Seventeen athletes are named among the “Forbes 100 Most](https://static.cupdf.com/doc/110x72/60b573235092ec449e54290b/intercepting-licensing-rights-why-college-athletes-need-a-2016-5-6-2012-intercepting.jpg)

