Molecular Cell, Vol. 18, 319–329, April 29, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2005.03.028 Interaction of Era with the 30S Ribosomal Subunit: Implications for 30S Subunit Assembly Manjuli R. Sharma, 1,7 Chandana Barat, 1,7 Introduction Daniel N. Wilson, 2,7 Timothy M. Booth, 1 The ribosome, a complex macromolecular machine Masahito Kawazoe, 3 Chie Hori-Takemoto, 3 composed of several rRNAs and a large number of pro- Mikako Shirouzu, 3 Shigeyuki Yokoyama, 3,4,5 tein molecules, carries out the translation of an organ- Paola Fucini, 2 and Rajendra K. Agrawal 1,6, * ism’s genetic information into proteins (reviewed by Ra- 1 Division of Molecular Medicine makrishnan, 2002). The biogenesis of the ribosome is a Wadsworth Center highly complex process that includes the cotranscrip- New York State Department of Health tional processing of the precursor rRNAs and the ribo- Empire State Plaza somal protein-assisted folding and maturation of the Albany, New York 12201 rRNAs into the fully assembled ribosomal subunits (see 2 Max-Planck Institute for Molecular Genetics Culver, 2003). The bacterial, small ribosomal subunit Ihnestrasse 63-73 (30S) is composed of a 16S rRNA and 21 ribosomal D-14195 Berlin proteins (designated S1–S21) (Wittmann-Liebold, 1986) Germany and plays a direct role in the decoding of the genetic 3 RIKEN Genomic Sciences Center message (see Ogle et al., 2003). Although in vitro as- 1-7-22 Suehiro-cho sembly of the 30S subunit using only its RNA and pro- Tsurumi, Yokohama, 230-0045 tein components has been achieved (Traub and No- Japan mura, 1968; Culver and Noller, 2000), efficient assembly 4 RIKEN Harima Institute at SPring-8 in vivo of fully active ribosomes requires accessory Mikazuki-cho factors, such as RNA helicases and other molecular Sayo, Hyogo, 679-5148 chaperones (Williamson, 2003). Japan Era, a multifunctional, essential GTPase is found in 5 Department of Biophysics and Biochemistry all bacteria with the exception of some obligate intra- Graduate School of Science cellular eubacteria (Caldon and March, 2003; Leipe et The University of Tokyo al., 2002), and it couples cell growth with cytokinesis 7-3-1 Hongo (Gollop and March, 1991; Britton et al., 1998). Era has Bunkyo-ku, Tokyo, 113-0033 also been implicated in cell division and energy metab- Japan olism (Britton et al., 1998; Inoue et al., 2002). It is pre- 6 Department of Biomedical Sciences sent in amounts that are typical for regulatory proteins, State University of New York at Albany and its synthesis increases as the cell’s growth rate in- Albany, New York 12201 creases (Bardwell et al., 1989; Britton et al., 1998; Mat- sunaga et al., 1996). Homologs of Era have been iden- tified in eukaryotes; depletion of these proteins leads to cell-cycle arrest at G1 phase and apoptosis (Gohda et al., 2003). One of the characteristic properties of Era Summary is its RNA binding ability, and its role in cell-cycle regu- lation is dependent on this feature (Johnstone et al., Era (E. coli Ras-like protein) is a highly conserved and 1999; Gohda et al., 2003). In bacteria, depletion of Era essential GTPase in bacteria. It binds to the 16S ribo- leads to an increase in intracellular levels of unassoci- somal RNA (rRNA) of the small (30S) ribosomal sub- ated 30S subunits and 50S large subunits, relative to unit, and its depletion leads to accumulation of an 70S ribosomes (Sayed et al., 1999), and a concomitant unprocessed precursor of the 16S rRNA. We have ob- accumulation of 17S rRNA, the unprocessed precursor tained a three-dimensional cryo-electron microscopic of the 16S rRNA (Inoue et al., 2003). Era can functionally map of the Thermus thermophilus 30S-Era complex. compensate for deletion of the gene encoding the cold- Era binds in the cleft between the head and platform shock adaptation protein RbfA (Inoue et al., 2003), of the 30S subunit and locks the subunit in a confor- which is required for efficient processing of the 16S mation that is not favorable for association with the rRNA (Dammel and Noller, 1993). Furthermore, Era- large (50S) ribosomal subunit. The RNA binding KH depleted cell extracts are incapable of translation, and motif present within the C-terminal domain of Era in- addition of Era does not restore translational ability teracts with the conserved nucleotides in the 3 re- (Sayed et al., 1999). These observations suggest that gion of the 16S rRNA. Furthermore, Era makes con- Era is not directly involved in the process of translation tact with several assembly elements of the 30S per se but is instead involved in the processing and subunit. These observations suggest a direct involve- maturation of the 30S subunit to a conformational state ment of Era in the assembly and maturation of the suitable for association with the 50S subunit. An X-ray crystallographic study has shown that Era 30S subunit. has a two-domain structure (Chen et al., 1999) with a GTPase-containing N-terminal domain (NTD) and an RNA binding C-terminal domain (CTD). The NTD com- *Correspondence: [email protected] 7 These authors contributed equally to this work. prises six β sheet strands flanked by five α helices,

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Cell, Vol. 18, 319–329, April 29, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2005.03.028
Interaction of Era with the 30S Ribosomal Subunit:Implications for 30S Subunit Assembly
Manjuli R. Sharma,1,7 Chandana Barat,1,7
Daniel N. Wilson,2,7 Timothy M. Booth,1
Masahito Kawazoe,3 Chie Hori-Takemoto,3
Mikako Shirouzu,3 Shigeyuki Yokoyama,3,4,5
Paola Fucini,2 and Rajendra K. Agrawal1,6,*1Division of Molecular MedicineWadsworth CenterNew York State Department of HealthEmpire State PlazaAlbany, New York 122012Max-Planck Institute for Molecular GeneticsIhnestrasse 63-73D-14195 BerlinGermany3RIKEN Genomic Sciences Center1-7-22 Suehiro-choTsurumi, Yokohama, 230-0045Japan4RIKEN Harima Institute at SPring-8Mikazuki-choSayo, Hyogo, 679-5148Japan5Department of Biophysics and BiochemistryGraduate School of ScienceThe University of Tokyo7-3-1 HongoBunkyo-ku, Tokyo, 113-0033Japan6Department of Biomedical SciencesState University of New York at AlbanyAlbany, New York 12201
Summary
Era (E. coli Ras-like protein) is a highly conserved andessential GTPase in bacteria. It binds to the 16S ribo-somal RNA (rRNA) of the small (30S) ribosomal sub-unit, and its depletion leads to accumulation of anunprocessed precursor of the 16S rRNA. We have ob-tained a three-dimensional cryo-electron microscopicmap of the Thermus thermophilus 30S-Era complex.Era binds in the cleft between the head and platformof the 30S subunit and locks the subunit in a confor-mation that is not favorable for association with thelarge (50S) ribosomal subunit. The RNA binding KHmotif present within the C-terminal domain of Era in-teracts with the conserved nucleotides in the 3� re-gion of the 16S rRNA. Furthermore, Era makes con-tact with several assembly elements of the 30Ssubunit. These observations suggest a direct involve-ment of Era in the assembly and maturation of the30S subunit.
*Correspondence: [email protected]
7 These authors contributed equally to this work.Introduction
The ribosome, a complex macromolecular machinecomposed of several rRNAs and a large number of pro-tein molecules, carries out the translation of an organ-ism’s genetic information into proteins (reviewed by Ra-makrishnan, 2002). The biogenesis of the ribosome is ahighly complex process that includes the cotranscrip-tional processing of the precursor rRNAs and the ribo-somal protein-assisted folding and maturation of therRNAs into the fully assembled ribosomal subunits (seeCulver, 2003). The bacterial, small ribosomal subunit(30S) is composed of a 16S rRNA and 21 ribosomalproteins (designated S1–S21) (Wittmann-Liebold, 1986)and plays a direct role in the decoding of the geneticmessage (see Ogle et al., 2003). Although in vitro as-sembly of the 30S subunit using only its RNA and pro-tein components has been achieved (Traub and No-mura, 1968; Culver and Noller, 2000), efficient assemblyin vivo of fully active ribosomes requires accessoryfactors, such as RNA helicases and other molecularchaperones (Williamson, 2003).
Era, a multifunctional, essential GTPase is found inall bacteria with the exception of some obligate intra-cellular eubacteria (Caldon and March, 2003; Leipe etal., 2002), and it couples cell growth with cytokinesis(Gollop and March, 1991; Britton et al., 1998). Era hasalso been implicated in cell division and energy metab-olism (Britton et al., 1998; Inoue et al., 2002). It is pre-sent in amounts that are typical for regulatory proteins,and its synthesis increases as the cell’s growth rate in-creases (Bardwell et al., 1989; Britton et al., 1998; Mat-sunaga et al., 1996). Homologs of Era have been iden-tified in eukaryotes; depletion of these proteins leadsto cell-cycle arrest at G1 phase and apoptosis (Gohdaet al., 2003). One of the characteristic properties of Erais its RNA binding ability, and its role in cell-cycle regu-lation is dependent on this feature (Johnstone et al.,1999; Gohda et al., 2003). In bacteria, depletion of Eraleads to an increase in intracellular levels of unassoci-ated 30S subunits and 50S large subunits, relative to70S ribosomes (Sayed et al., 1999), and a concomitantaccumulation of 17S rRNA, the unprocessed precursorof the 16S rRNA (Inoue et al., 2003). Era can functionallycompensate for deletion of the gene encoding the cold-shock adaptation protein RbfA (Inoue et al., 2003),which is required for efficient processing of the 16SrRNA (Dammel and Noller, 1993). Furthermore, Era-depleted cell extracts are incapable of translation, andaddition of Era does not restore translational ability(Sayed et al., 1999). These observations suggest thatEra is not directly involved in the process of translationper se but is instead involved in the processing andmaturation of the 30S subunit to a conformational statesuitable for association with the 50S subunit.
An X-ray crystallographic study has shown that Erahas a two-domain structure (Chen et al., 1999) with aGTPase-containing N-terminal domain (NTD) and anRNA binding C-terminal domain (CTD). The NTD com-prises six β sheet strands flanked by five α helices,

Molecular Cell320
whereas the CTD is a tightly folded structure of three βsheet strands on one side and three α helices on theother. The CTD possesses an RNA binding KH-signa-ture domain with a βααβ motif and a conserved se-quence, VIGxxGxxI, mapping within the helix-turn-helixmotif. The KH domain is known to be critical for RNArecognition and binding (Grishin, 2001). Deletion of theβ2, α2, α1, or β1 within the KH domain abolishes theability of these truncated Era genes to complement anE. coli mutant strain defective in Era expression (Hanget al., 2001), thus illustrating the importance of the KHdomain for 16S rRNA binding activity and bacterialgrowth. Although no functional coordination betweenthe NTD and CTD has yet been established, both do-mains are important for Era function (Lerner et al., 1995;Pillutla et al., 1995). Mutations in the CTD that interferewith Era’s RNA binding ability also interfere with its abil-ity to participate in the process of 30S-subunit matura-tion (Inoue et al., 2003; also see Johnstone et al., 1999).While there is no evidence for direct involvement ofEra’s GTPase activity in its function, earlier studieshave shown that mutations of conserved residueswithin the GTPase portion of the NTD that significantlyalter Era’s GTPase activity have no effect on its abilityto bind RNA (Johnstone et al. 1999). However, someother NTD mutations, which do not involve its GTPaseportion, were shown to affect Era’s RNA binding abilityand functionality (Johnstone et al. 1999), suggesting afunctional interaction between NTD and CTD. Other
Fstudies have shown that an Era mutant, in which the(effector (G2) region within the NTD has been deleted, issan efficient suppressor of defective ribosome assemblyu
and accumulation of 17S rRNA in a RbfA deletion mu- Ctant (Inoue et al., 2003). a
Here, we determine the binding position of Era on an tmS1-depleted Thermus thermophilus 30S subunit, usinglthe techniques of cryo-electron microscopy (cryo-EM)aand three-dimensional (3D) image reconstruction (Frank(
et al., 2000). Sucrose-density gradient and PAGE analy- (ses of the 30S-Era complex suggest a high affinity of 3Era for the 30S subunit. Comparison of the cryo-EM s
amap with X-ray crystallographic structures of Era (Chenhet al., 1999) and the 30S subunit (Wimberly et al., 2000)
reveals that Era binds to the 3# minor domain of the16S rRNA, through the highly conserved KH domain of sits CTD. Analysis of molecular contacts of Era with the avarious components of the 30S subunit provides a bet- iter understanding of the role of this highly conserved (protein during assembly and maturation of the 30S tsubunit. d
tmResultsmnBinding of Era to the 30S Subunit and Cryo-EM
Structure of the 30S-Era Complex faEra binds to the S1-depleted 30S subunit with high af-
finity: it remains bound to the 30S subunit after passagecthrough a 10%–30% sucrose gradient and pelleting of
the 30S subunits by overnight ultracentrifugation (Fig- fture 1A). The stoichiometry of the complex formed in the
presence of a 4-fold excess of Era was determined to btbe w80%, and higher excesses of Era did not signifi-
cantly improve the occupancy (data not shown). Con- b
igure 1. Binding of Era to the 30S Ribosomal Subunit
A) SDS-PAGE analysis of Era binding to the 30S subunit. The 30Subunit fraction, obtained by sucrose density gradient ultracentrif-gation of the 30S-Era complex, was analyzed by SDS PAGE andoomassie blue staining. Lane 1 shows the 30S-subunit proteins,nd lane 4 shows the Era protein. Lanes 2 and 3 were loaded withhe 30S-Era complex obtained in the presence of 2- and 4-foldolar excess of Era, respectively. Lane 5 shows a molecular weight
adder. The Era band (molecular mass w34 kDa) is marked by anrrow at the left.
B and C) 3D cryo-EM map showing the binding position of Erared) on the 30S subunit (yellow). The Era density isolated from the0S-Era complex is superimposed on the map of the control 30Subunit. The 30S subunit is shown in (B), an interface view, and (C),platform-side view. Landmarks and abbreviations are as follows:
, head; b, body; p, platform; n, neck; and sp, spur.
istent with the high occupancy seen in the PAGEnalysis, an extra mass of density corresponding to Era
s clearly resolved in the cryo-EM map of the complexFigures 1B and 1C), when compared with the map ofhe S1-depleted 30S subunit (control). This bilobed Eraensity feature spans the entire cleft region betweenhe head and platform of the 30S subunit, such that itakes contacts with three of the four structural do-ains of the subunit’s 16S rRNA (Wimberly et al., 2000),
amely, the central domain located in the subunit plat-orm, the 3# major domain located in the subunit head,nd the 3# minor domain.Superposition of cryo-EM maps of the Era bound and
ontrol 30S subunits reveals a significantly altered con-ormation of the subunit in its Era bound state. In par-icular, the 30S subunit head bends toward the platformy w4°, while the platform itself arcs downward, ex-ending away from the head by w3° (Figure 2A). Thus,oth head and platform show significant movements
Interaction of Era with the 30S Ribosomal Subunit321
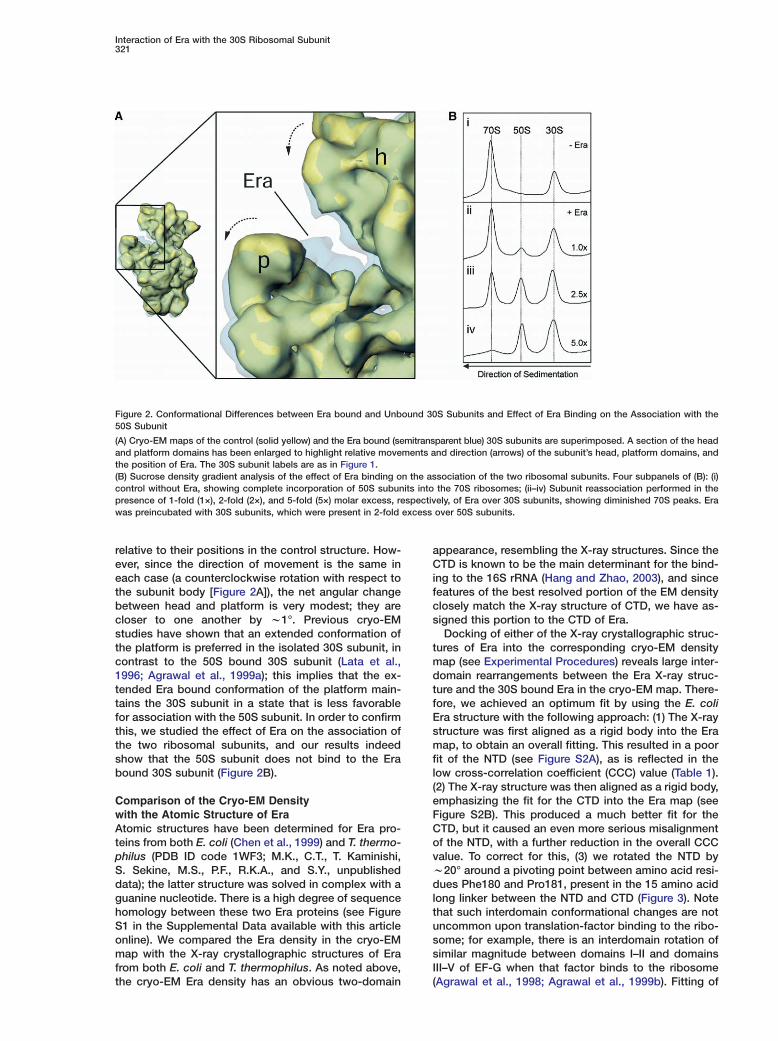
Figure 2. Conformational Differences between Era bound and Unbound 30S Subunits and Effect of Era Binding on the Association with the50S Subunit
(A) Cryo-EM maps of the control (solid yellow) and the Era bound (semitransparent blue) 30S subunits are superimposed. A section of the headand platform domains has been enlarged to highlight relative movements and direction (arrows) of the subunit’s head, platform domains, andthe position of Era. The 30S subunit labels are as in Figure 1.(B) Sucrose density gradient analysis of the effect of Era binding on the association of the two ribosomal subunits. Four subpanels of (B): (i)control without Era, showing complete incorporation of 50S subunits into the 70S ribosomes; (ii–iv) Subunit reassociation performed in thepresence of 1-fold (1×), 2-fold (2×), and 5-fold (5×) molar excess, respectively, of Era over 30S subunits, showing diminished 70S peaks. Erawas preincubated with 30S subunits, which were present in 2-fold excess over 50S subunits.
relative to their positions in the control structure. How-ever, since the direction of movement is the same ineach case (a counterclockwise rotation with respect tothe subunit body [Figure 2A]), the net angular changebetween head and platform is very modest; they arecloser to one another by w1°. Previous cryo-EMstudies have shown that an extended conformation ofthe platform is preferred in the isolated 30S subunit, incontrast to the 50S bound 30S subunit (Lata et al.,1996; Agrawal et al., 1999a); this implies that the ex-tended Era bound conformation of the platform main-tains the 30S subunit in a state that is less favorablefor association with the 50S subunit. In order to confirmthis, we studied the effect of Era on the association ofthe two ribosomal subunits, and our results indeedshow that the 50S subunit does not bind to the Erabound 30S subunit (Figure 2B).
Comparison of the Cryo-EM Densitywith the Atomic Structure of EraAtomic structures have been determined for Era pro-teins from both E. coli (Chen et al., 1999) and T. thermo-philus (PDB ID code 1WF3; M.K., C.T., T. Kaminishi,S. Sekine, M.S., P.F., R.K.A., and S.Y., unpublisheddata); the latter structure was solved in complex with aguanine nucleotide. There is a high degree of sequencehomology between these two Era proteins (see FigureS1 in the Supplemental Data available with this articleonline). We compared the Era density in the cryo-EMmap with the X-ray crystallographic structures of Erafrom both E. coli and T. thermophilus. As noted above,the cryo-EM Era density has an obvious two-domain
appearance, resembling the X-ray structures. Since theCTD is known to be the main determinant for the bind-ing to the 16S rRNA (Hang and Zhao, 2003), and sincefeatures of the best resolved portion of the EM densityclosely match the X-ray structure of CTD, we have as-signed this portion to the CTD of Era.
Docking of either of the X-ray crystallographic struc-tures of Era into the corresponding cryo-EM densitymap (see Experimental Procedures) reveals large inter-domain rearrangements between the Era X-ray struc-ture and the 30S bound Era in the cryo-EM map. There-fore, we achieved an optimum fit by using the E. coliEra structure with the following approach: (1) The X-raystructure was first aligned as a rigid body into the Eramap, to obtain an overall fitting. This resulted in a poorfit of the NTD (see Figure S2A), as is reflected in thelow cross-correlation coefficient (CCC) value (Table 1).(2) The X-ray structure was then aligned as a rigid body,emphasizing the fit for the CTD into the Era map (seeFigure S2B). This produced a much better fit for theCTD, but it caused an even more serious misalignmentof the NTD, with a further reduction in the overall CCCvalue. To correct for this, (3) we rotated the NTD byw20° around a pivoting point between amino acid resi-dues Phe180 and Pro181, present in the 15 amino acidlong linker between the NTD and CTD (Figure 3). Notethat such interdomain conformational changes are notuncommon upon translation-factor binding to the ribo-some; for example, there is an interdomain rotation ofsimilar magnitude between domains I–II and domainsIII–V of EF-G when that factor binds to the ribosome(Agrawal et al., 1998; Agrawal et al., 1999b). Fitting of

Molecular Cell322
Table 1. Crosscorrelation Values for Three Schemes of Fitting between the Cryo-EM Density and X-Ray Coordinates of Era Protein
Organism from Which EraStructure Was Derived Two Domains Fitted Rigid-Body Fit in Alternate(PDB ID code) Rigid-Body Fit Separately Orientationb
E. coli (1EGA) 0.69 0.79 0.59T. thermophilus (1WF3)a 0.72 0.69 0.64
a M.K., C.T., T. Kaminishi, S. Sekine, M.S., P.F., R.K.A., and S.Y., unpublished data.b Orientations of CTD and NTD were flipped to fit them into putative NTD and CTD, respectively, of the cryo-EM map.
entation (see Table 1). density (see Figure S2B). The homologous α helix in the
Figure 3. Comparison of the X-Ray Crystallographic Structure of Era with the Cryo-EM Density Map
(A) Stereo representation of the fitted X-ray structure (Chen et al., 1999) of E. coli Era (CTD in green and NTD in blue) into the cryo-EM densitymap (semitransparent red). The X-ray structure of Era is embedded within the EM density, recognizable by altered hues for the two Eradomains (light green for CTD and magenta for NTD), while portions of the X-ray structure that protrude from the cryo-EM density are visiblein their original (green or blue) colors. The Era EM density is shown at a threshold value lower than the value used in Figure 2.(B) The geometric pivot points (F180 and G284) that we used to obtain the best fit are highlighted. The axes (solid red lines) and planes(dashed lines) of the rotations are indicated. Ribbons in gray show the unfitted positions of the NTD and the C-terminal α helix of the CTD.The orientation of the 30S-Era complex is depicted to the lower right. The 30S subunit labels are as in Figure 1. C and N, respectively, indicatethe carboxy and amino termini of Era.
the domains independently produced a CCC of 0.793, alvalue significantly higher than the values obtained from
rigid-body fittings or the values obtained when the NTD CEand CTD were fitted independently in the opposite ori-
However, in the fitted Era structure, a short α helix,ocated at the C terminus and linked to the rest of theTD through a 5 amino acid long loop in the E. colira structure, protrudes completely out of the cryo-EM

Interaction of Era with the 30S Ribosomal Subunit323
T. thermophilus Era structure is oriented toward the restof the CTD and assumes a more compact structure,indicating that this α helix is flexible and capable ofadapting alternate conformations. Therefore, to accom-modate the C-terminal α helix (CTH) within the cryo-EMdensity, we allowed the G284–L295 segment to rotateby w8° around position G284, enabling a better fit ofthe CTH. In order to eliminate the possibility of abnor-mal bond geometry or unfavorable steric interactionsintroduced during independent fittings of three struc-tural components of Era (NTD, CTD, and separateCTH), we subjected the resultant Era structure to en-ergy minimization. The CCC value between the energy-minimized Era and cryo-EM map was 0.80 (Figure 3A).Even after these steps, portions of helix α2 of the NTDstill protruded from the Era density. A comparison ofthe Era structures from E. coli and from the guaninenucleotide bound T. thermophilus revealed a large dif-ference in orientation of this helix with respect to therest of the NTD (see Figure S3). This helix is part of theswitch II region of the Era GTPase and undergoes largeconformational changes upon GTP hydrolysis (Sprang,1997).
Fitting of the T. thermophilus Era structure into thecryo-EM map, using the same approach, resulted insignificantly poorer CCC values (Table 1), which couldbe due to either or both of the following factors: (1) The30S-Era complex analyzed here was obtained in theabsence of any nucleotide, whereas the T. thermophilusEra structure represents a guanine nucleotide boundform, and (2) it is likely that the E. coli Era structure inthe crystal represents a conformation that is moreclosely related to the 30S bound form than to the nucle-otide bound T. thermophilus Era structure. Ultimately,since the model with the three separately fitted compo-nents of the E. coli Era structure gives the highest CCCvalue (0.80), we used this model to determine all of thecontacts of Era with the 30S subunit.
Contact Sites of Era on the 30S SubunitInteraction between the KH Domain of Eraand the 3# End of the 16S rRNAIn order to determine the interaction sites of Era on the30S subunit, we separately fitted the four structural do-mains of the 30S subunit (Wimberly et al., 2000),namely, the head, platform, main body, and the 3# minordomain of its 16S rRNA, into our cryo-EM map. Withthe assumption that there are no major intradomain re-arrangements in Era’s 30S bound form, fittings of theX-ray structures reveal that the amino acid stretchV246–I254 (246VIGNKGAKI254), located at the turn of thehelix-turn-helix motif of the CTD, makes direct andextensive contacts with the 3# minor domain nucleo-tides 1530–1534 of the 16S rRNA (Figure 4). This findingis consistent with results from earlier studies (Chen etal., 1999; Johnstone et al., 1999), which predicted theinvolvement of the highly conserved signature se-quence VIGxxGxxI of the KH domain in binding of Erato the 16S rRNA. Our result is, to our knowledge, thefirst direct localization of the binding site of the KH do-main of Era to the 3# end of the 16S rRNA.
Interactions between Era and AssemblyElements of the 30S SubunitEra has been implicated in the assembly of the 30Ssubunit (Sayed et al., 1999; Inoue et al., 2003). The pro-cess of assembly of ribosomal subunits is known to behighly cooperative and hierarchical in nature (see Cul-ver, 2003). Based on the order in which the 30S subunitproteins interact with the 16S rRNA during 30S assem-bly, these proteins have been classified as primary, se-condary, or tertiary binding proteins. In addition to itsinteraction with the 3# end of the 16S rRNA, the CTDof Era interacts with ribosomal protein S7, a primaryassembly protein, and with the 16S rRNA helix 28 (h28)(Figure 5), which is involved in the latter phase of as-sembly of the 30S subunit (Holmes and Culver, 2004).The amino acid sequence S223–G227 (SNERG), whichlies in the loop between βA and βB of Era’s CTD (Chenet al., 1999), makes contact with Y151–W156 (YAHYRW)of protein S7. The amino acid residues W285–E289(WADDE) of the αD chain of the Era CTD interact withnucleotides G928–C930, and amino acid residues R293and S294 interact with nucleotide U1381 of h28. Thus,the contact sites of Era with S7 and h28 flank the βααβmotif housing the KH domain, which is believed to bethe main ribosomal binding site of Era.
The NTD and the NTD-CTD linker region (amino acidresidues 172–188) are involved in several interactionswith two components of the 30S subunit platform (Fig-ure 5): protein S18, a secondary-assembly protein (Heldet al., 1974), and h26 of the 16S rRNA, also involved inthe same stage of assembly (Agalarov and Williamson,2000). The linker residues E182 and D183 contact theP16–A20 stretch (PSRKA) of protein S18, whereas theamino acid residues G30–K32 (GQK) of NTD lie in closeproximity (within 4–8 Å) to G832 and U833 of h26. Also,I33 of the NTD might interact with nucleotide U723 ofh23. Some of the ribosomal components that are notknown to be primary- or secondary-assembly elementsof the 30S subunit, such as protein S2 and rRNA h37of the 30S-subunit head, may also interact with Era(Figure 5). Amino acid residues A133 and L136 of Era’sNTD are within w5–8 Å of the amino acid residues G99and R96, respectively, of protein S2, and the E131–D134 stretch (EKAD) of Era’s NTD makes contact withnucleotides G1099 and G1100 of rRNA h37. Amino acidresidues A271 and P272 of Era’s CTD interact with E92of the platform protein S11.
Discussion
The 16S rRNA nucleotides 1530–1534 (GAUCA), whichmake direct contact with the KH domain of Era, are po-sitioned immediately upstream of the anti-Shine-Dal-garno (anti-SD) sequence and are highly conservedamong eubacteria, archaea, and eukaryotes (Raué etal., 1988; Noller, 1993). These nucleotides lie in closeproximity to the ribosome bound messenger RNA(mRNA), as the rRNA nucleotide G1530 crosslinks to anmRNA at a site between its SD region and the AUGstart codon (Rinke-Appel et al., 1994). The conservednature of this rRNA region and the region’s close prox-imity to the anti-SD region suggest that this region ofthe 30S subunit is functionally crucial. Indeed, 30S sub-

Molecular Cell324
Figure 4. Interactions between the Helix-Turn-Helix Motif of the KH Domain of Era’s CTD and the 3# Segment of the 16S rRNA
The amino acid residues of the KH domain (green) that make direct contact with the rRNA (purple) are shown with side chains (blue). TherRNA residues either making direct contact with or in close proximity to Era are highlighted as beads (orange). Left panel depicts the overallorientation of the 30S subunit, with the 16S rRNA 3#-minor domain (purple) highlighted. The 30S subunit labels are the same as in Figure 1.
units bearing 16S rRNA mutations of the invariant pu- svrines G1530/A1531–A1530/G1531 are unable to form
70S ribosomes in vitro, in the absence of any factors dl(Firpo et al., 1996). These mutated 30S subunits are
also incapable of binding fMet-tRNAfMet and hence are a
aunable to form the preinitiation complex. Conversely,although 70S ribosomes containing mutant 30S sub- t
1units, produced in vivo, had a high affinity for IF3, theability of IF3 to dissociate these ribosomes into sub- h
1units was lost (Firpo et al., 1996). These results high-light the importance of the 16S rRNA nucleotides i
bG1530–A1534 in the formation of the translation initia-tion complex. Specific binding of Era to this rRNA re- c
ogion could, to a certain extent, functionally mimic theeffects of the above-described mutations, by inhibiting a
cthe formation of an initiation complex on an incom-pletely assembled 30S subunit. Furthermore, we find a
2that the binding site of Era significantly overlaps that ofprotein S1 (Figure 6), a protein known to directly influ- t
fence SD/anti-SD interactions (Komarova et al., 2002).Our finding suggests that Era and S1 cannot coexist on i
hthe 30S subunit and that the binding of Era would pre-vent the recruitment of mRNA to an incompletely as- s
ssembled 30S subunit.The CTD of Era makes direct contact with protein S7 l
(Figure 5A). Protein S7 is located at the corner of the30S subunit head that encompasses two multiple rRNA l
tstem junctions, one involving 16S rRNA helices h28,h29, and h43, and the other involving helices h29, h30, t
ch41, and h42 (Brodersen et al., 2002). Footprinting datasuggest that the binding of S7 stabilizes the packing s
sof h43 with h29 and h41 and induces packing of RNAelements outside the primary interaction sites (Powers c
hand Noller, 1995). The other 30S head element that in-teracts with the Era CTD is h28 (Figure 5B), the confor- S
omation of which is known to be important for ribosomefunction (Ericson et al., 1995). Earlier studies have t
hown that the 30S subunit undergoes reversible inacti-ation in vitro at low concentrations of monovalent andivalent cations (Zamir et al., 1974). The 30S subunit
oses its tRNA binding capacity and is unable to associ-te with the 50S subunit. The inactivation process isccompanied by large changes in chemical reactivity inhe 920–927 (h28) and 1391–1401 (h44) regions in the6S rRNA, indicating that h28 and part of the adjacent44 are disrupted in the inactive state (Moazed et al.,986). Mutational studies also indicate that h28 may be
mportant in determining the conformation of the tRNAinding site, either by organizing rRNA structure (in-luding G926 and G1401) or by controlling the correctrientation of the head of the 30S subunit (Ericson etl., 1995). Protein S7 and h28 have been implicated inrucial transitions that occur late in both the in vitrond in vivo assembly processes (Holmes and Culver,004). Since S7 also participates in long-range interac-ions among three of the structural domains (head, plat-orm, and body) of the 30S subunit, and since h28 isnvolved in determining the correct orientation of theead domain, direct interactions of Era with these ribo-omal elements during final stages of 30S subunit as-embly could be key in defining proper interdomain re-
ationships.As mentioned above, the NTD and the NTD-CTD
inker region of Era interact mainly with the platform ofhe 30S subunit. Protein S18 and h26 of the 16S rRNA,he two platform components with which Era makesontact, are both elements involved in the secondarytage of 30S-subunit assembly. S18 is important for as-embly of the platform domain because it stabilizeslose packing of the secondary rRNA elements (helices22, h23a, and h26) (Agalarov and Williamson, 2000).18 is proposed to be involved in overall stabilizationf the 30S subunit (Held and Nomura, 1975). In addi-ion, S18 interacts with proteins S11 and S6. Like S18,

Interaction of Era with the 30S Ribosomal Subunit325
Figure 5. Stereo Representations of Interactions of Era with 30S-Subunit Proteins and rRNA Segments
(A) Interactions of Era (red) with proteins S2 and S7 of the head (purple) and proteins S11 and S18 of the platform (green); and (B) interactionswith the 16S rRNA helices h28 and h37 of the head (blue) and helices h23 and h26 of the platform (brown). The corresponding orientationsof the 30S subunit are depicted to the left of each panel. The 30S subunit labels are the same as in Figure 1.
main. The only component of the head domain of the It is likely that the interactions of Era with S18, S11,
Figure 6. Comparison of the Binding Posi-tions of Era and S1 on the 30S Subunit
Binding positions of (A) Era (red) and (B) pro-tein S1 (blue), shown side by side, in an inter-face view of the 30S subunit (yellow). The S1density was isolated by comparing the pro-tein-only map computed for the 30S-subunitportion of the E. coli 70S ribosome contain-ing the S1 protein with the protein-only mapof the S1-depleted T. thermophilus 30S sub-unit (also see Sengupta et al., 2001). The 30Ssubunit labels are as in Figure 1.
S11 is important for stabilization of the platform struc-ture (Brodersen et al., 2002), and it also interacts withEra’s CTD. Interaction of S11 with S18, together withtheir contacts with the 690 (h23b) and 790 (h24) loopsof the 16S rRNA of the platform, brings this domain inclose proximity to h45 of the 16S rRNA 3# minor do-
30S subunit that contacts the NTD is the S2 protein. S2lies in the hinge region between the head and platform,on the solvent side of the 30S subunit, and joins thesubunit during the final stages of assembly. Era couldbe also involved in the recruitment of S2 to the 30Ssubunit.

Molecular Cell326
and h26 cause the 30S-subunit platform to extend out- eward. As stated earlier, an extended platform represents aa 30S-subunit conformation that is unfavorable for as- esociation with the 50S subunit (see Lata et al., 1996; dAgrawal et al., 1999a). Since the platform is directly in- tvolved in the formation of several bridges between the mtwo subunits (Gabashvili et al., 2000; Yusupov et al., g2001), Era binding-associated conformational changes tin the 30S platform (Figure 2A) might maintain the 30S hsubunit in an association-incompetent conformation t(Figure 2B) until the process of its assembly is com- tplete. This observation does not necessarily conflict awith the fact that Era depletion in vivo leads to a greater 1proportion of unassociated 30S and 50S subunits, rela- Etive to 70S ribosomes (Sayed et al., 1999), since ab- ssence of Era leads to the presence of precursor 17S rrRNA (Inoue et al., 2003), which can still form into a 30S fsubunit. However, such a 30S subunit is inactive, due Eto the presence of flanking 3# and 5# regions that form csecondary structure with one another; these regions amust be removed for activation of the 30S subunit(Wireman and Sypherd, 1974a; Wireman and Sypherd, w1974b; also see, Culver, 2003). Their removal is proba- fbly also necessary to bring the 30S subunit into an as- psociation-competent conformation. It is possible that dthe binding of Era precedes the final processing steps Fof maturation of the 3# end of the 16S rRNA precursor ion the ribosome. Because mature 16S rRNA is present cin our 30S subunits, and because Era has not been re- mleased from these S1-depleted 30S subunits, it appears tthat we have captured Era in a specific physiological tconformation during the latter stages of the 30S assem- fbly and after the rRNA has been processed. Further- tmore, because the primary binding site of Era (via the oKH domain) lies at the 3# end of the 16S rRNA, the over- Eall binding position of Era is likely to remain largely un- caltered during this late assembly process.
Although there is no direct evidence that Era’sEGTPase activity plays a functional role, the presence of
both the GTP binding NTD and CTD, and the properP
interaction between the NTD and the CTD, are impor- atant for Era’s functioning (Lerner et al., 1995; Pillutla et Tal., 1995). Era binds to the 30S subunit both in presence o
iand absence of GTP. However, its RNA binding abilityNis reduced upon GTP binding (Sayed et al., 1999). This,ttaken with the significant conformational differencesuobserved between the NTDs of GTP bound (M.K., C.T., s
T. Kaminishi, S. Sekine, M.S., P.F., R.K.A., and S.Y., un- spublished data) and nucleotide-free (Chen et al., 1999) rEra (also see Figure S3), may suggest that GTP hydroly- b
vsis (induced by an as-yet-unknown mechanism) is re-squired for the eventual release of Era from the fully as-Hsembled 30S subunit.s
It has been demonstrated that the body, head, and (platform of the 30S subunit can assemble separately ain vitro (Agalarov et al., 1998; Agalarov et al., 1999; Sa- �
maha et al., 1994). Based on the pattern of interaction (uof the primary-, secondary-, and tertiary-assembly pro-pteins with the 16S rRNA, it appears that assembly ofothe 30S subunit proceeds in a 5# to 3# direction alongb
the 16S rRNA (Powers et al., 1993). These observations bsuggest that interdomain interactions occur late in the tsubunit assembly process. However, based on the c
gatomic structure of the mature 30S subunit (Wimberly
t al., 2000), we cannot rule out that whereas primary-ssembly proteins may help to nucleate assembly ofach of the subunit’s large domains, concomitant inter-omain interactions are also necessary to determinehe subunit’s final structure. Our study reveals that Eraakes several contacts with the head and platform re-ions of the 30S subunit. Previously described interac-ions of Era with protein S2, and with 16S RNA helices37 and h26, could be critical in establishing the rela-ive orientations of the head and platform. Furthermore,he contact of Era with protein S11 could influence thebility of S11 to bring the subunit head and h45 of the6S rRNA 3# minor domain into close proximity. Thus,ra could play a direct role in bringing together all fourtructural domains (body, head, platform, and the 16SRNA 3# minor domain) of the 30S subunit during theinal stages of the assembly process. Alternatively,ra’s role could be indirect, in merely influencing theonformational changes of these components associ-ted with the assembly process.Thus, in addition to its role as an rRNA chaperone,hereby it stabilizes the secondary structures needed
or correct processing of the 3# end of the 16S rRNArecursor (Inoue et al., 2003), Era appears to play a roleuring the final stages of the 30S subunit assembly.urthermore, by maintaining the 30S-subunit platform
n an extended conformation, and by binding to theonserved pre anti-SD nucleotides, Era inhibits the for-ation of the translation initiation complex on a prema-
urely assembled 30S subunit; complete assembly ishe key to ensuring the high fidelity and speed neededor mRNA decoding on the 30S subunit. The fact thathe binding site of Era overlaps significantly with thatf ribosomal protein S1 suggests that dissociation ofra and incorporation of S1 are the steps that mark theompletion of the assembly process.
xperimental Procedures
reparation of T. thermophilus 30S Ribosomal Subunitsnd the 30S-Era Complex. thermophilus 30S ribosomal subunits were isolated by passagef a crude ribosome preparation over a 10%–30% sucrose gradient
n buffer A (10 mM Hepes-KOH [pH 7.8], 10 mM MgOAc, 60 mMH4Cl, and 6 mM β-mercaptoethanol). The fraction corresponding
o the 30S peak was pooled, and the ribosomes were pelleted byltracentrifugation at 80,000 × g for 24 hr at 4°C. SDS-PAGE analy-is of the 30S subunits revealed that the amounts of S1 are sub-toichiometric, relative to the other ribosomal proteins (see 60 kDaegion of Figure 1A). The gene for T. thermophilus Era was amplifiedy PCR from HB8 genomic DNA and was cloned in an expressionector pET11b (Novagen). It was overexpressed in the BL21(DE3)train, and was purified by successive column chromatography oniPrep Butyl FF and MONO Q (Amersham Bioscience) from theoluble fraction, after a heat treatment at 70°C, as described earlierKato et al., 2003). 30S subunits (2.4 �M) were incubated eitherlone or with a 2- to 4-fold molar excess of purified Era protein (9.6M) at 60°C for 15 min in buffer A. After incubation, a small aliquot
w20 �l) was snap-frozen in liquid nitrogen and stored at −80°C,ntil needed. For the preparation of cryo-EM grids, these com-lexes were diluted in buffer A, containing a 10-fold molar excessf Era. The remainder of the reaction was used to determine theinding efficiency of Era to the 30S subunit. This was accomplishedy separating unbound Era protein in two different ways: (1) Reac-
ions were applied to 10%–30% sucrose gradients (in buffer A) andentrifuged in an SW40 rotor at 41,000 × g for 18 hr at 4°C. Theradients were monitored at 260 nm, and the 30S peak fractions

Interaction of Era with the 30S Ribosomal Subunit327
were pooled and centrifuged in a TLA100.3 rotor at 80,000 × g for24 hr at 4°C. (2) Reactions were applied to a 10% sucrose cushionin buffer A and were centrifuged at 215,000 × g for 30 min. In bothcases, the pellets were resuspended in 20 �l of buffer A, the con-centration was determined using the absorbance at 260 nm, and10 pmol of complex was analyzed by SDS-PAGE. The occupancyof the complex was determined by densitometric analysis of theSDS-PAGE gels, comparing the Era band within the Era-30S com-plexes with known amounts of Era protein, and then normalizingthese values against the constant amount of 30S subunit loadedper sample (10 pmol).
Reassociation of Ribosomal Subunits in the Presence of EraThe reassociation protocol was modified from that described byBlaha and coworkers (2000). E. coli 30S subunits (1 �M) were prein-cubated with varying amounts of T. thermophilus Era protein (1–5�M) at 37°C for 20 min in buffer B (20 mM Hepes-KOH [pH 7.8],20 mM MgOAc, 30 mM KCl, and 4 mM β-mercaptoethanol) in thepresence of 100 �M GDPNP. Subsequently, 50S subunits (0.5 �M)were added, and the reaction was incubated for a further 30 min at37°C, before being loaded onto 10%–30% sucrose gradients inbuffer B and centrifuged in an SW40 rotor at 61,000 × g for 18 hrat 4°C. Control experiments, performed under the same conditionsbut with Era protein replaced by equivalent amounts of bovine se-rum albumin showed no inhibition of subunit association (data notshown), demonstrating that the Era inhibition does not result fromnonspecific interactions.
Cryo-Electron Microscopy and 3D ReconstructionCryo-EM grids were prepared according to standard procedures(Wagenknecht et al., 1988). Data were collected on a Philips FEI(Eindhoven, The Netherlands) Tecnai F20 FEG microscope equip-ped with low-dose kit and an Oxford cryotransfer holder, at a mag-nification of 50,000×. Micrographs were scanned on a Zeiss flatbedscanner, with a step size of 14 �m, corresponding to 2.82 Å onthe object scale. For the 30S-Era complex, 104 micrographs werescanned. Initially, 107,244 selected particle images were sorted into21 defocus groups (ranging from 1.18–3.94 �m under focus). Aftermanual screening, removal of images from over-representedgroups within 83 equispaced views of the ribosome, and elimina-tion of images based on ranking of CCC values, a total of 56,333particles were retained for the final 3D reconstruction. The pro-jection-matching procedure (Penczek et al., 1994) in conjunctionwith the SPIDER software, was used to obtain the CTF-corrected3D map (Frank et al., 2000). Similarly, a cryo-EM map of the uncom-plexed T. thermophilus 30S subunit was obtained as control. Forthis, 44,390 particle images from 37 micrographs were initiallysorted into 14 defocus groups (ranging from 1.87–4.04 �m). A totalof 26,216 images were included in the final 3D reconstruction. Theresolutions, estimated using the Fourier-shell-correlation (FSC) witha cutoff value of 0.5, were 13.5 Å (or 8.5 Å by the 3σ criterion;Orlova et al., 1997) for the 30S-Era complex and 15.5 Å (or 9.7 Å by3σ) for the 30S-subunit control.
Fitting of Atomic Structures into the Cryo-EM MapsTo obtain an optimum fitting of the X-ray crystallographic structureof the T. thermophilus 30S subunit (Wimberly et al., 2000; PDB IDcode 1FJF), we divided the X-ray coordinates into four structuraldomains (head, body, platform, and 16S rRNA 3# minor domain),and we then fitted each domain individually into the cryo-EM mapsof the control 30S and the 30S-Era complex. Fitted coordinateswere converted into electron density maps using a pixel size of2.82 Å, with filtration to the resolution of the individual cryo-EMmaps. The density assigned to Era was isolated through compari-son of the cryo-EM map of the 30S-Era complex and the densitymap obtained from the fitting of the 30S X-ray coordinates into thecryo-EM maps of the 30S-Era complex. The atomic structures ofEra were fitted initially with Era considered as a single rigid body;next, with the atomic structure considered as two separate rigidbodies, corresponding to NTD and CTD; and finally, with the C-ter-minal helix within the CTD considered as the third rigid body so asto optimize the fit into the isolated Era density. The CCC valuesbetween the fitted X-ray coordinates of Era and the corresponding
cryo-EM density were determined after conversion of the fitted co-ordinates into the density map, through computation of averageddensities within volume elements scale-matched to those of thecryo-EM map. The accuracy of fittings of atomic structures intocryo-EM maps has been estimated to be in the range of 1/6 to1/10 of the resolution of the cryo-EM map (see, e.g., Rossmann,2000), which would yield w1.8 Å in the case of our results with Era.
Docking of the X-ray structures into the cryo-EM map was donewith O (Jones et al., 1991), and visualization of the fitted atomicstructures and the cryo-EM density maps was done with Ribbons(Carson, 1991) and IRIS EXPLORER (Numerical Algorithms group,Inc., Downers Grove, IL), respectively. Insight II (Accelrys Inc.,USA), with the molecular mechanics/dynamics program DIS-COVER, was used to perform energy minimization. All of the num-bering for Era refers to the E. coli protein sequence; for all ribo-somal components, rRNA and ribosomal proteins, T. thermophilusnumbering is used.
Supplemental DataSupplemental Data include three additional figures and can befound with this article online at http://www.molecule.org/cgi/content/full/18/3/319/DC1/.
Acknowledgments
The authors thank Adriana Verschoor for critical reading of themanuscript. This work was supported in part by grants from theNational Institutes of Health, GM61576 (to R.K.A.) and from the Hu-man Frontier Science Program, RGY232003 (to R.K.A.). Supportfrom National Science Foundation grant DBI9871347, for the Wads-worth Center’s EM infrastructure, is gratefully acknowledged.
Received: December 16, 2004Revised: March 16, 2005Accepted: March 24, 2005Published: April 28, 2005
References
Agalarov, S.C., and Williamson, J.R. (2000). A hierarchy of RNA sub-domains in assembly of the central domain of the 30S ribosomalsubunit. RNA 6, 402–408.
Agalarov, S.C., Zheleznyakova, E.N., Selivanova, O.M., Zheleznaya,L.A., Matvienko, N.I., Vasiliev, V.D., and Spirin, A.S. (1998). In vitroassembly of a ribonucleoprotein particle corresponding to the plat-form domain of the 30S ribosomal subunit. Proc. Natl. Acad. Sci.USA 95, 999–1003.
Agalarov, S.C., Selivanova, O.G., Zheleznyakova, E.N., Zheleznaya,L.A., Matvienko, N.I., and Spirin, A.S. (1999). Independent in vitroassembly of all three major morphological parts of the 30S ribo-somal subunit of Thermus thermophilus. Eur. J. Biochem. 266,533–537.
Agrawal, R.K., Penczek, P., Grassucci, R.A., and Frank, J. (1998).Visualization of elongation factor G on the Escherichia coli 70S ri-bosome: the mechanism of translocation. Proc. Natl. Acad. Sci.USA 95, 6134–6138.
Agrawal, R.K., Lata, R.K., and Frank, J. (1999a). Conformationalvariability in Escherichia coli 70S ribosome as revealed by 3D cryo-electron microscopy. Int. J. Biochem. Cell Biol. 31, 243–254.
Agrawal, R.K., Heagle, A.B., Penczek, P., Grassucci, R.A., andFrank, J. (1999b). EF-G-dependent GTP hydrolysis induces translo-cation accompanied by large conformational changes in the 70Sribosome. Nat. Struct. Biol. 6, 643–647.
Bardwell, J.C.A., Regnier, P., Chen, S.-M., Nakamura, Y., Grunberg-Manago, M., and Court, D.L. (1989). Autoregulation of RNase IIIoperon by mRNA processing. EMBO J. 27, 3401–3407.
Blaha, G., Stelzl, U., Spahn, C.M., Agrawal, R.K., Frank, J., andNierhaus, K.H. (2000). Preparation of functional ribosomal com-

Molecular Cell328
plexes and effect of buffer conditions on tRNA positions observed Ioby cryoelectron microscopy. Methods Enzymol. 317, 292–309.eBritton, R.A., Powell, B.S., Dasgupta, S., Sun, Q., Margolin, W., Lup-Mski, J.R., and Court, D.L. (1998). Cell cycle arrest in Era GTPase
mutants: a potential growth rate-regulated checkpoint in Esche- Jrichia coli. Mol. Microbiol. 27, 739–750. R
TBrodersen, D.E., Clemons, W.M., Carter, A.P., Wimberly, B.T., andmRamakrishnan, V. (2002). Crystal structure of the 30S ribosomal1subunit from Thermus thermophilus: structure of the proteins and
their interactions with 16S RNA. J. Mol. Biol. 316, 725–768. JpCaldon, C.E., and March, P.E. (2003). Function of the universallymconserved bacterial GTPases. Curr. Opin. Microbiol. 6, 135–139.ACarson, M. (1991). Ribbons 2.0 J. Acta. Crystallogr. D Biol. Crystal-Klogr. 24, 103–106.SChen, X., Court, D.L., and Ji, X. (1999). Crystal structure of Era: aoGTPase-dependent cell cycle regulator containing an RNA bindingBmotif. Proc. Natl. Acad. Sci. USA 96, 8396–8401.KCulver, G.M. (2003). Assembly of the 30S ribosomal subunit. Bio-Ppolymers 68, 234–249.D
Culver, G.M., and Noller, H.F. (2000). In vitro reconstitution of 30SLribosomal subunits using complete set of recombinant proteins.FMethods Enzymol. 318, 446–460.r
Dammel, C.S., and Noller, H.F. (1993). A cold-sensitive mutation inL16S rRNA provides evidence for helical switching in ribosome as-csembly. Genes Dev. 7, 660–670.M
Ericson, G., Minchew, P., and Wollenzien, P. (1995). StructuralLchanges in base-paired region 28 in 16S rRNA close to the decod-ding region of the 30S ribosomal subunit are correlated to changeslin tRNA binding. J. Mol. Biol. 250, 407–419.F
Firpo, M.A., Connelly, M.B., Goss, D.J., and Dahlberg, A.E. (1996).MMutations at two invariant nucleotides in the 3#-minor domain ofEEscherichia coli 16S rRNA affecting translational initiation and initi-ration factor 3 function. J. Biol. Chem. 271, 4693–4698.MFrank, J., Penczek, P., Agrawal, R.K., Grassucci, R.A., and Heagle,IA.B. (2000). Three-dimensional cryoelectron microscopy of ribo-asomes. Methods Enzymol. 317, 276–291.o
Gabashvili, I.S., Agrawal, R.K., Spahn, C.M., Grassucci, R.A., Sver-Ngun, D.I., Frank, J., and Penczek, P. (2000). Solution structure of themE. coli 70S ribosome at 11.5 Å resolution. Cell 100, 537–549.JGohda, J., Nomura, Y., Suzuki, H., Arai, H., and Inoue, J. (2003).1Elimination of the vertebrate Escherichia coli Ras-like protein ho-Omologue leads to cell cycle arrest at G1 phase and apoptosis. On-tcogene 22, 1340–1348.BGollop, N., and March, P.E. (1991). A GTP-binding protein (Era) hasOan essential role in growth rate and cell cycle control in EscherichiaJcoli. J. Bacteriol. 173, 2265–2270.tGrishin, N.V. (2001). KH domain: one motif, two folds. Nucleic AcidsaRes. 29, 638–643.PHang, J.Q., and Zhao, G. (2003). Characterization of the 16S rRNA-iand membrane-binding domains of Streptococcus pneumoniaerEra GTPase: structural and functional implications. Eur. J. Bio-Uchem. 270, 4164–4172.PHang, J.Q., Meier, T.I., and Zhao, G. (2001). Analysis of the interac-Ction of 16S rRNA and cytoplasmic membrane with the C-terminalmpart of the Streptococcus pneumoniae Era GTPase. Eur. J. Bio-achem. 268, 5570–5577.2
Held, W.A., and Nomura, M. (1975). Escherichia coli 30 S ribosomalPproteins uniquely required for assembly. J. Biol. Chem. 250, 3179–r3184.PHeld, W.A., Ballou, B., Mizushima, S., and Nomura, M. (1974). As-isembly mapping of 30 S ribosomal proteins from Escherichia coli.BFurther studies. J. Biol. Chem. 249, 3103–3111.RHolmes, K., and Culver, G.M. (2004). Mapping structural differencesobetween 30S ribosomal subunit assembly intermediates. Nat.RStruct. Mol. Biol. 11, 179–186.sInoue, K., Chen, J., Kato, I., and Inouye, M. (2002). Specific growthsinhibition by acetate of an Escherichia coli strain expressing Era-
dE, a dominant negative Era mutant. J. Mol. Microbiol. Biotechnol. Rs4, 379–388.
noue, K., Alsina, J., Chen, J., and Inouye, M. (2003). Suppressionf defective ribosome assembly in a rbfA deletion mutant by over-xpression of Era, an essential GTPase in Escherichia coli. Mol.icrobiol. 48, 1005–1016.
ohnstone, B.H., Handler, A.A., Chao, D.K., Nguyen, V., Smith, M.,yu, S.Y., Simons, E.L., Anderson, P.E., and Simons, R.W. (1999).he widely conserved Era G-protein contains an RNA-binding do-ain required for Era function in vivo. Mol. Microbiol. 33, 1118–
131.
ones, T.A., Zou, J.Y., Cowan, S.W., and Kjeldgaard, M. (1991). Im-roved methods for building protein models in electron densityaps and the location of errors in these models. Acta Crystallogr.47, 110–119.
ato, M., Shirouzu, M., Terada, T., Yamaguchi, H., Murayama, K.,akai, H., Kuramitsu, S., and Yokoyama, S. (2003). Crystal structuref the 2#-5# RNA ligase from Thermus thermophilus HB8. J. Mol.iol. 329, 903–911.
omarova, A.V., Tchufistova, L.S., Supina, E.L., and Boni, I.V. (2002).rotein S1 counteracts the inhibitory effect of the extended Shine-algarno sequence on translation. RNA 8, 1137–1147.
ata, K.R., Agrawal, R.K., Penczek, P., Grassucci, R., Zhu, J., andrank, J. (1996). Three-dimensional reconstruction of the Esche-ichia coli 30 S ribosomal subunit in ice. J. Mol. Biol. 262, 43–52.
eipe, D.D., Wolf, Y.I., Koonin, E.V., and Aravind, L. (2002). Classifi-ation and evolution of P-loop GTPases and related ATPases. J.ol. Biol. 317, 41–72.
erner, C.G., Gulati, P.S., and Inouye, M. (1995). Cold-sensitive con-itional mutations in Era, an essential Escherichia coli GTPase, iso-
ated by localized random polymerase chain reaction mutagenesis.EMS Microbiol. Lett. 126, 291–298.
atsunaga, J., Dyer, M., Simons, E.L., and Simons, R.W. (1996).xpression and regulation of the rnc and pdxJ operons of Esche-ichia coli. Mol. Microbiol. 27, 977–989.
oazed, D., Van Stolk, B.J., Douthwaite, S., and Noller, H.F. (1986).nterconversion of active and inactive 30 S ribosomal subunits isccompanied by a conformational change in the decoding regionf 16 S rRNA. J. Mol. Biol. 191, 483–493.
oller, H.F. (1993). On the origin of ribosome: coevolution of subdo-ains of tRNA and rRNA. In The RNA World, R.F. Gesteland and
.F. Atkins, eds. (Cold Spring Harbor, NY: CSHL Press), pp. 137–56.
gle, J.M., Carter, A.P., and Ramakrishnan, V. (2003). Insights intohe decoding mechanism from recent ribosome structures. Trendsiochem. Sci. 28, 259–266.
rlova, E.V., Dube, P., Harris, J.R., Beckman, E., Zemlin, F., Markl,., and van Heel, M. (1997). Structure of keyhole limpet hemocyaninype 1 (KLH1) at 15 Å resolution by electron cryomicroscopy andngular reconstitution. J. Mol. Biol. 271, 417–437.
enczek, P., Grassucci, R.A., and Frank, J. (1994). The ribosome atmproved resolution: new techniques for merging and orientationefinement in 3D cryo-electron microscopy of biological particles.ltramicroscopy 53, 251–270.
illutla, R.C., Sharer, J.D., Gulati, P.S., Wu, E., Yamashita, Y., Lerner,.G., Inouye, M., and March, P.E. (1995). Cross-species comple-entation of the indispensable Escherichia coli era gene highlights
mino acid regions essential for activity. J. Bacteriol. 177, 2194–196.
owers, T., and Noller, H.F. (1995). Hydroxyl radical footprinting ofibosomal proteins on 16S rRNA. RNA 1, 194–209.
owers, T., Daubresse, G., and Noller, H.F. (1993). Dynamics ofn vitro assembly of 16 S rRNA into 30 S ribosomal subunits. J. Mol.iol. 232, 362–374.
amakrishnan, V. (2002). Ribosome structure and the mechanismf translation. Cell 108, 557–572.
aué, H.A., Klootwijk, J., and Musters, W. (1988). Evolutionary con-ervation of structure and function of high molecular weight ribo-omal RNA. Prog. Biophys. Mol. Biol. 51, 77–129.
inke-Appel, J., Jünke, N., Brimacombe, R., Lavrik, I., Dokudov-kaya, S., Dontsova, O., and Bogdanov, A. (1994). Contacts be-

Interaction of Era with the 30S Ribosomal Subunit329
tween 16S ribosomal RNA and mRNA, within the spacer regionseparating the AUG initiator codon and the Shine-Dalgarno se-quence; a site-directed cross-linking study. Nucleic Acids Res. 22,3018–3025.
Rossmann, M.G. (2000). Fitting atomic models into electron-microscopy maps. Acta. Crystallogr. D Biol. Crystallogr. 56, 1341–1349.
Samaha, R.R., O’Brien, B., O’Brien, T.W., and Noller, H.F. (1994).Independent in vitro assembly of a ribonucleoprotein particle con-taining the 3# domain of 16S rRNA. Proc. Natl. Acad. Sci. USA 91,7884–7888.
Sayed, A., Matsuyama, S., and Inouye, M. (1999). Era, an essentialEscherichia coli small G-protein, binds to the 30S ribosomal sub-unit. Biochem. Biophys. Res. Commun. 264, 51–54.
Sengupta, J., Agrawal, R.K., and Frank, J. (2001). Visualization ofprotein S1 within the 30S ribosomal subunit and its interaction withmessenger RNA. Proc. Natl. Acad. Sci. USA 98, 11991–11996.
Sprang, S.R. (1997). G protein mechanisms: insights from structuralanalysis. Annu. Rev. Biochem. 66, 639–678.
Traub, P., and Nomura, M. (1968). Structure and function of E. coliribosomes. V. Reconstitution of functionally active 30S ribosomalparticles from RNA and proteins. Proc. Natl. Acad. Sci. USA 59,777–784.
Wagenknecht, T., Grassucci, R., and Frank, J. (1988). Electron mi-croscopy and computer image averaging of ice-embedded largeribosomal subunits from Escherichia coli. J. Mol. Biol. 199, 137–145.
Williamson, J.R. (2003). After the ribosome structures: how are thesubunits assembled? RNA 9, 165–167.
Wimberly, B.T., Brodersen, D.E., Clemons, W.M., Jr., Morgan-War-ren, R.J., Carter, A.P., Vonrhein, C., Hartsch, T., and Ramakrishnan,V. (2000). Structure of the 30S ribosomal subunit. Nature 407, 327–339.
Wireman, J.W., and Sypherd, P.S. (1974a). In vitro assembly of 30Sribosomal particles from precursor 16S RNA of Escherichia coli.Nature 247, 552–554.
Wireman, J.W., and Sypherd, P.S. (1974b). Properties of 30S ribo-somal particles reconstituted from precursor 16S ribonucleic acid.Biochemistry 13, 1215–1221.
Wittmann-Liebold, B. (1986). Ribosomal proteins: their structureand evolution. In Structure, function and genetics of ribosomes, B.Hardesty and G. Kramer, eds. (NY: Springer), pp. 326–361.
Yusupov, M.M., Yusupova, G.Z., Baucom, A., Lieberman, K., Ear-nest, T.N., Cate, J.H., and Noller, H.F. (2001). Crystal structure ofthe ribosome at 5.5 Å resolution. Science 292, 883–896.
Zamir, A., Miskin, R., Vogel, Z., and Elson, D. (1974). The inactiva-tion and reactivation of Escherichia coli ribosomes. Methods Enzy-mol. 30, 406–426.
Accession Numbers
Relevant coordinates of the fitted X-ray crystallographic structuresof Era and the 30S subunit components into the cryo-EM map ofthe 30S-Era complex have been deposited in the Protein DataBank, http://www.rcsb.org/pdb/, as two separate files (PDB IDcodes 1X18 and 1X1L)
Related Documents