Neiv Phytol. (1995), 133, 113-1A2 Inter- and intraspecific variation within the morphologically-similar arbuscular mycorrhizal fungi Glomus mosseae and Glomus coronatum BY J. C. DODDS S, ROSENDAHL^ M, GIOVANNETTP, A, BROOME*, L, LANFRANCC^ AND C. WALKER* ^ International Institute of Biotechnology/Department of Biosciences, University of Kent, Canterbury, Kent CT2 7YW, UK ^Department of Mycology, University of Copenhagen, 0ster Farimagsgade 2D, DK-1353 Copenhagen K, Denmark ^ Instituto di Microbiologia Agraria, Centro di Studio per la Microbiologia del Suolo, Via del Borghetto 80, 56124 Pisa, Italy "^Forestry Commission, The Forestry Authority, Northern Research Station, Roslin, Midlothian EH25 9SY, UK ^ Dipartimento di Biologia Vegetale & CSMT-CNR, Universita di Torino, Viale Mattioli 25, 10125 Torino, Italy {Received 13 November 1995) This collaborative investigation was aimed at using morphological and molecular characters to study inter- and intraspecific variation within isolates of Glomus mosseae and Glomus coronatum from different parts of the world, A secondary aim was to assess whether any divergence found was genetically based and/or correlated with the biogeographic origin of isolates. Morphological evaluations of various possible taxonomic characters including spore colour, size, sporocarp architecture and hyphal attachment morphology, showed that only spore colour couid discriminate the two groups, Isozyme analysis of malate dehydrogenase and esterase loci clearly confirmed this grouping of the two species complexes, SDS profiles and cluster analysis showed the same separation on a selection of isolates from the two groups, A comparison of a representative isolate from the G, coronatum group (BEG 49) wrth other arbuscular mycorrhizal fungi using G, mo^^eae-specific primers also provided evidence for separation of the two species complexes. The data and methodology employed provide a blueprint for future multimodal and niultidisciplinary approaches to the unravelling of taxonomic problems within this ancient group of symbiotic fungi. Key words: Arbuscular mycorrhizal fungi, molecular markers, Glomus mosseae, Glomus coronatum, morphological diversity, ,,,.„ molecular data obtained from nucleotide sequences INTRODUCTION of the 18S rRNA genes (Berbee & Taylor, 1993; Arbuscular mycorrhizal (AM) fungi {Glomales, Simon eta/,, 1993), The taxonomy of tbese obiigately iygomycotina) are one of the tnost widespread biotrophic fungi bas principally relied on the mor- =;roups of fungi on eartb. The symbiotic relationship phological cbaracteristics of tbe resting spores and vhich tbey establish with plant roots, mycovthiza, is spore wall cbaracteristics, leading to tbe currentlj' ound in most biomes and witb most plant species, described six genera and at least 150 species (Walker fbere is evidence from fossil plants that AM fungi & Trappe, 1993), In many cases, however, species colonized the roots of tbe first land plants have been described wbich are clearly identical with Pyrozynski&Dalpe, 1989; Taylor eJ a/,, 1995), and previously named species (Rosendabl, Dodd & hat both the plants and tbe fungal partner have Walker, 1994), Little is known of the range of probably co-evolved during the last 400 million diversity of tbese fungi because of tbe difficulty in ears. This is strongly supported by analysis of identifying them in the field (Giovannetti & Gianin-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Neiv Phytol. (1995), 133, 113-1A2
Inter- and intraspecific variation within themorphologically-similar arbuscularmycorrhizal fungi Glomus mosseae andGlomus coronatum
BY J. C. D O D D S S, ROSENDAHL^ M, GIOVANNETTP, A, BROOME*,L, LANFRANCC^ AND C. WALKER*
^ International Institute of Biotechnology/Department of Biosciences, University of Kent,Canterbury, Kent CT2 7YW, UK^Department of Mycology, University of Copenhagen, 0ster Farimagsgade 2D, DK-1353Copenhagen K, Denmark^ Instituto di Microbiologia Agraria, Centro di Studio per la Microbiologia del Suolo, Viadel Borghetto 80, 56124 Pisa, Italy"^Forestry Commission, The Forestry Authority, Northern Research Station, Roslin,Midlothian EH25 9SY, UK^ Dipartimento di Biologia Vegetale & CSMT-CNR, Universita di Torino, VialeMattioli 25, 10125 Torino, Italy
{Received 13 November 1995)
This collaborative investigation was aimed at using morphological and molecular characters to study inter- andintraspecific variation within isolates of Glomus mosseae and Glomus coronatum from different parts of the world,A secondary aim was to assess whether any divergence found was genetically based and/or correlated with thebiogeographic origin of isolates. Morphological evaluations of various possible taxonomic characters includingspore colour, size, sporocarp architecture and hyphal attachment morphology, showed that only spore colour couiddiscriminate the two groups, Isozyme analysis of malate dehydrogenase and esterase loci clearly confirmed thisgrouping of the two species complexes, SDS profiles and cluster analysis showed the same separation on a selectionof isolates from the two groups, A comparison of a representative isolate from the G, coronatum group (BEG 49)wrth other arbuscular mycorrhizal fungi using G, mo^^eae-specific primers also provided evidence for separationof the two species complexes. The data and methodology employed provide a blueprint for future multimodal andniultidisciplinary approaches to the unravelling of taxonomic problems within this ancient group of symbioticfungi.
Key words: Arbuscular mycorrhizal fungi, molecular markers, Glomus mosseae, Glomus coronatum, morphologicaldiversity,
,,,.„ molecular data obtained from nucleotide sequencesINTRODUCTION
of the 18S rRNA genes (Berbee & Taylor, 1993;Arbuscular mycorrhizal (AM) fungi {Glomales, Simon eta/,, 1993), The taxonomy of tbese obiigatelyiygomycotina) are one of the tnost widespread biotrophic fungi bas principally relied on the mor-=;roups of fungi on eartb. The symbiotic relationship phological cbaracteristics of tbe resting spores andvhich tbey establish with plant roots, mycovthiza, is spore wall cbaracteristics, leading to tbe currentlj'ound in most biomes and witb most plant species, described six genera and at least 150 species (Walkerfbere is evidence from fossil plants that AM fungi & Trappe, 1993), In many cases, however, speciescolonized the roots of tbe first land plants have been described wbich are clearly identical withPyrozynski&Dalpe, 1989; Taylor eJ a/,, 1995), and previously named species (Rosendabl, Dodd &hat both the plants and tbe fungal partner have Walker, 1994), Little is known of the range ofprobably co-evolved during the last 400 million diversity of tbese fungi because of tbe difficulty inears. This is strongly supported by analysis of identifying them in the field (Giovannetti & Gianin-

114 y.C. Dodd and others
azzi-Pearson, 1994). The problem is exacerbatedby the diffictjlty in obtaining AM fungi in axenicculture. There is, however, a relatively low level ofmorphological diversity which might, in part, be dueto the criteria used in selecting the importanttaxonomic characters (Walker, 1992; Bentivenga &Morton, 19946; Rosendahl et aL, 1994).
The morphological similarity might concealphysiological diversity in these fungi. Some studieshave suggested that these difTerences could begenetically based, using the analysis of isozymevariation within spore-cluster-forming Glomus spp.(Rosendahl, 1989) and Glomus mosseae (Hepper etaL, 1988), although in the latter study the resultshave subsequently been questioned, as the variationmight have been interspecific rather than intra-specific (Rosendahl et aL, 1994). Ecologicaldifferences between species could be used to supporttheir taxonomic separation, but it would be im-portant to determine whether the habitat-correlatedvariation is induced by environmental conditions oris genetically based.
Another taxonomic problem with certain speciessuch as Glomus mosseae, is documentation of therange of morphological variation which can occurbased on colour, size, wall structure and hyphalattachment. This species has become associated withthe yellow-vacuolate (YV) isolate described hyMosse & Bowen (1968) which had a funnel-shapedattachment, but the species description was based ona different isolate (Gerdemann & Trappe, 1974). It isone of the most frequently reported species instudies done around the globe and consequentlywidely used in experimentation. Another type whichbas been described as being more dark brown witha more prominent funnel-shaped attachment hasbeen regularly described in ecological surveys inMediterranean zones around the world, e.g. Israel(Dodd & Krikun, 1984), Italy (Giovannetti &Nicolson, 1983), Li.hya (El-Giahmi, Nicolson &Daft, 1976), Portugal, Spain and Australia (Sward,Hallam & Holland, 1978) and possibly more recentlyin Northern Europe (Blaszkowski, 1994), althoughthe latter isolates appear not to have the typicalfunnel-shaped attachment. Giovannetti, Avio &Salutini (1991) subsequently decided that there weresufficient differences, based on morphological, cyto-chemical and ontogenetic characteristics of thespores and sporocarps, to erect the new species calledGlomus coronatum. Both species form uniquesporocarps with a hyphal peridium (Meier &Charvat, 1992), which also delimits them from otherspecies of Glomus.
The objective of the present study was to usemolecular characters to compare YV isolates of G.mosseae from different parts of the world with G.coronatum and morphologically-similar dark brownfunnel-shaped AM fungi.
MATERI.'iLS ."iND METHODS
Source and maintenance of the fungi
Isolates were either maintained in pot culture atKent, Edinburgh, Copenhagen or Pisa, or samples ofpot cultures were obtained from other sources as air-dried inoculum (see Table 1). The fungi weremaintained in different substrates including theattapulgite clay product Terragreen® (Turfpro Ltd,UK), a coarse durite sand (a calcined flint of pH 8-5in water) and a variety of other sands or sandy soils.The culturing conditions and isolation history of thepot cultures varied; most were open-pot culturesmade from either single-spore or multi-spore cultureattempts. The isolate of Glomus dimorphicum. and theVenezuelan isolate of G. mosseae (BEG 61) wereoriginally multi-spore cultures, but were re-isolatedfrom a single spore in the UK. The isolate Glomussp. (AD-1) was included as it appeared to have anocclusion of the attachment similar to that found inthe G. mosseaejG. coronatum grouping but wasoccasionally observed to have an innermost mem-branous wall not yet observed in G. coronatum (BEG28).
Morphological characterization
Table 1 lists the isolates used within the study. Pot-cuiture material from each isolate was carefullypassed through a 710/(m sieve and collected on a45 /im sieve before backwashing into Petri dishes.The pot cultures were carefully checked for purityand, with one exception, were found to contain onlythe required species. The exception was the fungusG. mosseae (DDA3). This culture was from a multi-spore isolation and two species, G. mosseae andGlomus etunicatum were found. These species,however, are readily separated, and spores of theformer were carefully selected under a dissectingmicroscope. Spore colours were determined using acolour chart (Anon., 1969) under a stereomicroscopewith a split fibre-optic light source with a colourtemperature of 3200 °K, at magnifications of up to50. The presence or absence of sporocarps wasassessed for each isolate, together with sporocarpperidium colour and texture, and the number ofspores per sporocarp. The colour and Melzer'sreaction (data not presented) of endocarpic (excisedfrom the sporocarps) and ectocarpic spores were alsoinvestigated. More detailed microscopic examinationof the spores was made from specimens mounted inpolyvinyl-alcohol lacto-glycerol (Omar, Bolland &Heather, 1979) with or without the addition ofMelzer's reagent. Mounted specimens wereexamined under a compound microscope atmagnifications of up to x 2000, with brightfield anddifferential interference contrast (Nomarski) optics.Prepared slides and spores preserved in 0-025%sodium azide were kept as voucher specimens and

Variation in two Glomus species 115
Table 1. List of isolates included in the morphological j isozyme study
AM fungus
Yellow-vacuoiate typesG. mosseaeG. mosseaeG. mosseaeG. MosseaeG. mosseaeG. mosseaeG. mosseaeG. mosseaeG. mosseaeG. mosseaeG. mosseaeG. mosseaeG. dimorphicum
Brown funnel-shapedG. coronatumGlontus sp.Glomus sp.Glomus sp.
Brown inner wallGlomus sp.
Out-groupsG. caledoniumG. geosporum
BEGcode*
—BEG 12BEG 24BEG 25BEG 46BEG 54BEG 55BEG 61BEG 29BEG 57——BEG 59
BEG 28BEG 49BEG 22—
—
BEG 20BEG 11
Localcode
DK 154LPA 5KentSussexECPC 1107PALU-3PHIL-IDECPC 1132V 57V 139DD.A.3MAID-5A 65-1
—.4.LM-1LPA 13Al l 1-1
.4D-1
—Kent
Voucherno.
W 1854W 1857W 1858W 1859W 1860W 1863W 1862W 1865W 1804W 1805W 2354W 1903W 1805
W 1856W 1855W 1867W 1861
W 1868
W 1936W 1869
Source
S. RosendahlV. Gianinazzi-PearsonJ. C. DoddJ. C. DoddG. BethlenfalvayJ. C, Dodd/Y. SetiadiJ. C, DoddI. CuencaM. VestbergM. VestbergC. Walker/MehrotraJ. C. Dodd/C. WalkerD. McArthur
M. GiovannettiJ. C. Dodd/P. JeffriesV. Gianinazzi-PearsonL. Abbott
J .C. Dodd
RothamstedJ .C. Dodd
Country oforigin
DenmarkUKUKUKU S AIndonesiaThe PhilippinesVenezuelaFinlandFinlandIndiaNigeriaCanada
ItalySpainAustraliaAustralia
Abu-Dhabi
UKUK
* BEG is La Banque Europeenne des Glomales (European Bank of Glomales, Dodd et al., 1994).
given accession numbers (e.g. W1854) in the per-sonal herbarium of one of us (C.W.).
Native polyacrylamide gel electrophoresis
Spores in Petri dishes were collected on clean filterpaper (Whatman No. 1) and subsequently trans-ferred to Eppendorf tubes containing sucrose-tritonextraction-buffer (STEB): 20 mM trisma base;10 mM NaHCO.,, 10 mM MgCl^. 6Hi,O; 0-1 mMNa.jEDTA.2H2O; 10 mM mercaptoethanol;100 g 1-1 Triton® X-100. The spores collected in thetubes were crushed over ice for 2 tnin using a glasspestle formed from a flamed Pasteur pipette andcentrifuged for 20 min at 4 °C (200005-). Thesupernatants were transferred to another Eppendorfand stored at — 80 °C until required. A HoeferMighty Small (Pharmacia) vertical gel electro-phoresis system was used. The electrode buffercontained 25 mM Tris and 192 mM glycine at pH 8-3.The proteins were separated at pH 8'8 in a 375 mMTris-HCl buffer and stacked at pH 6-8 in a 125 mMTris-HCl buffer. The non-denaturing resolving gelcontained 7-5 % acrylamide, and the stacking gel 5 %acrylamide. The thickness of the gels was 0-75 mm.The gels were stained for esterase (EST,EC 3.1.1.1) or malate dehydrogenase (MDH,EC 1.1.1.37). Gel runs were made on three separateoccasions and gels stained for both enzymes in twoseparate laboratories (Denmark & Kent, UK). Otherenzyme stains were used but did not providediscriminatory data.
Sodium dodecyl sulphate (SDS)~polyacrylamide gelelectrophoresis
G. coronatum (BEG 28), and three dark-sporedGlomus sp. isolates (BEG 49, AD-1 and BEG 22) andfour YV isolates of G. mosseae (BEG 54, BEG 55,DK 154 and BEG 12) were compared using G.caledonium (BEG 20) and G. geosporum (BEG 11) asout-groups. Spores were ground at 4 °C and solubleproteins extracted in a buffer containing 75 mMTris/HCI (pH 8-5), 2 mM phenylmethylsulphonylfluoride (PMSE), 290 mM mercaptoethanol, 20%glycerol, 2 % sodium dodecyl sulphate (SDS), 1 '5 /(Maprotinin and 50 /iM leupeptin. Samples were boiledat 100 °C for 10 min, centrifuged at 14000^ for20 min at 4 °C and were then either stored at — 80 °Cor loaded immediately onto gels after adding 0-002 %bromophenol blue. Aiiquots of the proteins extractedwere precipitated with acetone atid the proteincontent was assessed by the Bio-Rad® assay pro-cedure using bovine serine albumin as standard(Bradford, 1976). One-dimensional SDS-PAGE wascarried out using a discontinuous buffer system(Laemmli, 1970). Extracts (2 fig of protein) wereloaded onto linear 10% polyacrylamide gels(10 cm X 8 cm x 0-75 mm) and run in a Bio-Rad MiniProtean II slab cell systena at 25 mA and 200 V atroom tenaperature. Gels were stained in BrilliantBlue R-250 (Sigtna) and then silver-stained,photographed and air-dried in cellulose sheets.
The number of bands detected was registered and

116 J. C. Dodd and others
the similarity between the isolates was estimatedusing the Kulczynski index:
where A is the number of shared bands, B is thenumber of bands found only in isolate 1 and C is thenumber of bands found only in isolate 2. Thesimilarity matrix was clustered by UPGMA, singleand complete linked clustering using the NTSYS(Numerical taxonomy and multivariate analysissystem, Exeter Software, ver. 1.80; Rohlf, 1994).Only the result of the UPGMA is presented as aphenogram. The cophenetic correlation wascalculated from comparisons between the originalsimilarity matrix and the matrix of cophenetic valuescalculated from the UPGMA phenogram.
DNA analyses
The brown-spored Glomus sp. (BEG 49) was selectedfrom the putative G. coronatum. group to checkagainst other isolates of Glomus using three DNAtechniques: (i) ITSl -4 primers, (ii) G. mosseae-specific primers designed by Lanfranco et al. (1995)and (iii) RAPD-PCR primers. The other isolates ofAM fungi used in this brief comparative study weremaintained separately in Torino and had voucherspecimen numbers from the Herbarium Crypto-gamicum (HC) of the Dipartintiento di BiologiaVegetale deH'Universita di Torino. These were G.mosseae (ROTH), which has the same origin as BEG12, Glomus versiforme (USA-Voucher specimenHC/F-EOl), Gigaspora margarita (BEG 34 - HC/E-ElO) and Glomus fiavisporum (Walker).
Extraction of DN.'\ and RAPD experiments wereperformed as described in Wyss & Bonfante (1993).Pooled spores (n = 5—10) were cleaned by sonication(4 X 15 s) and by four rinses with sterile bi-distilledwater, then crushed with sterile forceps in 100/tlwater. To them 50 /il of 20 % Chelex* (Bio-Rad)suspension were added immediately. After a 15 spulse sonication and freezing/thawing in liquidnitrogen four times, the preparation was heated for1 5 min at 95 °C. The supernatants were stored at- 2 0 °C.
ITS region analysis. The primers used to analyse theITS region (ITS1-ITS4) were described by Whiteet al. (1990). The reaction of PCR amplification wascarried out in SO/<1 of final volume containing:10 mM Tris-HCl (pH 9-0), 50 mM KCl, 0-1 % TritonX-100, 1-5 mM MgCla, 200/(M of dNTP, 1/IM ofeach primer, 0'5 units of SuperTaq®—Polymerase(Stehelin, Basel, Switzerland) and 10 /«1 of undilutedDNA extracted from spores. The reaction mixtureswere overlaid with 40 /tl of naineral oil (Sigma) toprevent evaporation. ITS1-ITS4 primers wereobtained from Primm s.r.l. (Milan, Italy). Fortycycles of amplifieation were carried out in a DNAthermal cycler (Perkin-Elmer-Cetus) using a step
programme; 94 °C 1 min, 50 °C 1 min 30 s, 72 °C1 min 30 s, followed by a 5 min final extension at72 °C. Next 10 /j.1 of the PCR reaction were digestedwith 5 units of Hint I (2 h at 37 °C) and loaded on a2 % agarose gel.
Glomus mosseae-specific P0-M3 primers. The PCRconditions for the P0-M3 reactions are as describedin Lanfranco et al (1995).
RAPD analysis. Amplification of DNA was carriedout in one-pritner reactions. The primers OPA-7,OP A-11 and OPA-18 were obtained from OperonTechnologies, Inc. (Alameda, CA, USA). Thereaction mix consisted of 10 mM Tris-HCl (pH 9-0),50 mM KCl, 0-1% Triton X-100, 2 mM MgCl^,200 fiM of each dNTP, 15 ng of primer, 0-4 units ofSuperTaq*-Polynierase (Stehelin, Basel, Switzer-land) and 6 ml of the template diluted 5-10-fold in afinal volume of 15 /A. Mineral oi! (15 [iX) was addedon the top of the reaction mixture. The amplificationreaction was performed in a Perkin Elmer thermalcycler programmed as follows: 1 x 2-5 min at 94 °C,45 X 30 s at 94 °C, 1 min at 36 °C, 2 min at 72 °C,1 x 5 min at 72 °C. Control experiments were per-formed without adding the template DNA. Theamplification products were separated electro-phoretically on 1-4 % agarose gels and stained withethidiuna bromide (Williams et al., 1990).
RESULTS
Morphological comparisons
Sporocarpic spore colour, and ectocarpic sporecolour where present, clearly distinguished (Table 2)the YV G. mosseae group from the brown funnel-shaped G. coronatum group and Glomus sp. (AD-1).There was some overlap in the ranges of sporecolours between the two but generally G. mosseaeisolates (YV) were yellow and the G. coronatumgroup were brown (Sienna on the colour chart). Allisolates produced sporocarps except the out-groupfungi, Glomus sp. (AD-1), G. caledonium (BEG 20)and G. geosporum (BEG 11). None was observed inGlomus sp. (BEG 22) but they have been previously(Gianinazzi-Pearson, unpublished) and sporocarpswere subsequently observed in the G. mosseae(MAID-5) isolate. There were no easily definabledifTerences in peridium texture nor in the ranges ofspore numbers per sporocarp (Table 2). Eurtherstudy of the developmental stages of the differentisolates would be needed, with replication withinisolates, if this taxonomic character is to be used withany degree of accuracy. Similarly Melzer's reactionsof the hyaline outer wall were variable (data notpresented) which correlated directly with the degreeof persistence of this layer on the spores investigated.As mentioned above this reaction may be useful
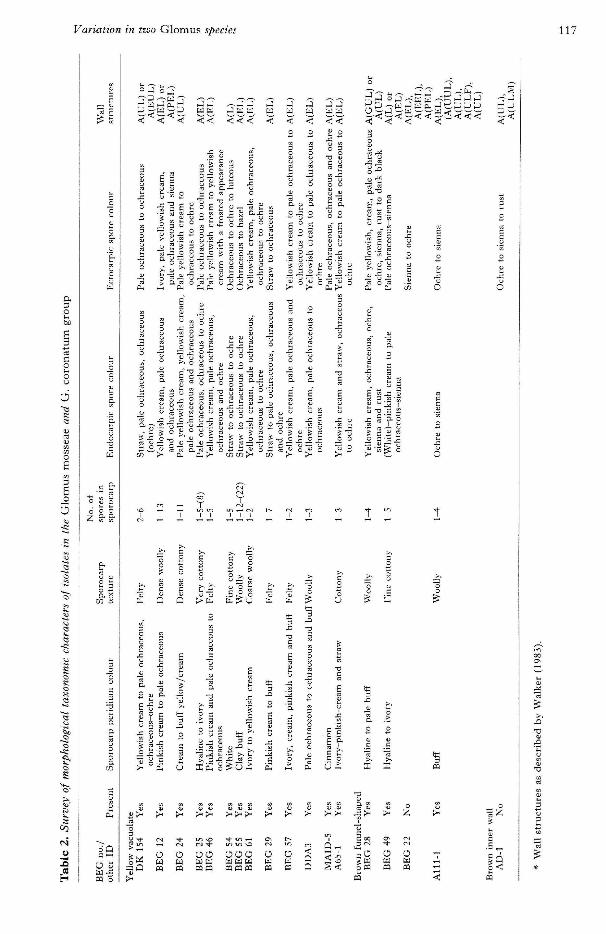
Variation in two Glomus species 117
(Si
3O)
o
1 I
°9
cq 5
.--^ t ^ ...- . p^
M C C
S S
•3 S-c 2 i x :
> a s o o o
E s,
tuJS
OJI
wit
h a
aiH
u
aj
X NU KO X0
3•OJ
brae
a
Beo
us t
ois
h er
eabr
ae
OO
low
Yel
1 oc
h)
23O
Xo
CO
ceou
:o o
chra
Str
a;
r-H 0Cra
su
Xu:
low
Yel
w
0i;
XO
P.
o
•B
ish
crea
low
Yel
s, o
chr
aceo
uoc Pal
a0
£
UX1/;
low
;:h
re
1°
c
crea
X
SV
1)
Pal
, ru
stsi
enna
,hr
e,
0
s-si
e
§rakH
Xo
Pal
? X o -
D m F ^ OO 3 « S ?̂u o u ™ i:
UJ Li f- "^^ "ra ^ ; = u S
r
ra ;H y rt «•c^ £ a au X
E ° o E E"ra O -3 ra
S I 2
Q Or c o
JJ O
a ii
£ • £ £C u ""̂
I ? S2 TO Cj
| i |S 2 «
£ it
2 o .tl
• ^ d , k^
c o
s
^ !
..Q•3
>. :i: :r
? m ^
= QO O 00
enOOOu u uCQ P3 CQ
o os <

118 J.C. Dodd and others
1 2 3 4 5 6 7
(a) , 1 . .
1 2 3 4 5 6 7
Figure 1. Native-PAGE gels of spore extracts of different isolates of the G. mosseae and G. coronatum groupsstained for (a) MDH, {b) esterase: lane 1, G. mosseae (BEG 61); lane 2, G. mosseae (BEG 55); lane 3, G.mosseae (BEG 54); lane 4, G. coronatum. (BEG 28); lane 5, Glomus sp. (BEG 22); lane 6, Glomus sp. ( A l l l -1); lane 7, Glotrtus sp. (BEG 49) and lane 8, G. mosseae (DK154).
1 2 3 4 5 6 7 8 9 10 n 12 13 14 15 16 17 18 19(al
Figure 2. Diagrams of the Native-PAGE gels run showing the (a) MDH and (6) esterase patterns of all isolatesrun on three different occasions and in two different laboratories: lane 1, Glomus sp. (BEG 49); lane 2, Glomussp. (BEG 22); lane 3, Glomus sp. (Al l l -1 ) ; lane 4, G, coronatum (BEG 28); lane 5, Glomus sp. (AD-1); lane6, G. mosseae (BEG 54); lane 7, G. mosseae (BEG 55); lane 8, G. mosseae (BEG 61); lane 9, G. mosseae (DK154); kne 10, G. mosseae (BEG 24); lane 11, G. mosseae (BEG 25); lane 12, G. mosseae (BEG 25); lane 13, G.mosseae (BEG 46); lane 14, G. dimorphicum (BEG 59); lane 15, G. mosieae (BEG 29); kne 16, G. mosseae (BEG57); lane 17, G. mosseae (DDA-3); lane 18, G. geosporum (BEG 11) and lane 19, G. caledonium (BEG 20).

Variation in two Glomus species 119
10 11 12 13 14
29
Figure 3. SDS-PAGE of total proteins extracted from spores of diflerent isolates of the G. mosseae and G.coronatum groups: lane 1, molecular weight markers (/?-galactosidase 116 kDa; albumen, bovine 66 kDa;albumen, egg 45 kDa; carbonic anhydrase 29 kDa (Sigma); lane 2, G. caledonium (BEG 20); lane 3, G.geosporum (BEG 11); lane 4, G. mmseae (BEG 54); lane 5, G. mosseae (BEG 55); lane 6, G. mosseae (BEG 25);lane 7, G. mosseae (DK154); lane 8, molecuiar weight markers; lane 9, Glomus sp. (AD-l); lane 10, G. coronatum(BEG 28); lane 11, Glomus sp. (BEG 49); lane 12, Glomus sp. (All 1-1); lane 13, G. geosporum (BEG 11) andlane 14, G. caledonium (BEG 20).
when spores are tested at earlier stages of maturationwhen the hyaline wall is present on most spores.
Isozyme comparisons
The results of the isozyme profiles, from MDH andesterase, showed that the isolates of G. mosseaeformed a well-defined group including G.dimorphicum (BEG 59) (Figs 1, 2). G. coronatum(BEG 28) and the brown funnel-shaped isolatesformed their own group (Figs 1, 2). G. caledonium(BEG 20) and G. geosporum (BEG 11) formed adistinct out-group (Figs 1, 2). Only a single MDHlocus was found in the (YV) isolates whose Rf valuesvaried little. The isolates with brown funnel-shapedspores all had two putative loci, a slow-movingmonomorphic locus and a faster-moving polymor-phic locus. This included the non-funnel-shapedspores oi Glomus sp. (AD-1) (Figs 1,2). The esterasepattern showed far greater variation between YV G.mosseae isolates (Figs 1, 2) but the locus found in thedark brown isolates showed no variation. Theyellow-spored isolates showed an esterase patternwith several closely-migrating bands. The patternwas interpreted as one locus, as the number of bandsvaried between runs.
SDS protein profiles
Figure 3 shows the soluble protein profiles resultingfrom SDS-PAGE on the selected isolates from bothmorphological groups. Only minor differences werefound between the phenograms generated by theanalysis using different clustering methods. Thephenogram based on the UPGMA clustering (Fig.4) shows the YV G. mosseae isolates falling into onecluster with the two European isolates BEG 25 andDKl 54 the two most closely linked. The dark brown'.solates including G. coronatum (BEG 28) alsoformed a unique cluster. Within this cluster theisolates Glomus sp. (AD-1) and the Italian ex-type of
Similarity
0-2 0-4 0-6 0-8 1-0
G. caledonium (BEG 20)
G. geosporum (BEG 11)
Glomus sp. (Alll-IiGlomus sp. {BEG 49)
G. coronatum (BEG 28)
Glomus sp. (AD-1)
G. mosseae (BEG 54)
G. mosseae (BEG 55)
G. mosseae {BBG 25)
G. mosseae (DKl 54)
Fig. 4. UPGMA cluster analysis of selected G. mosseae andG. coronatum isolates. The cophenetic correlation was0-864.
G. coronatum (BEG 28) had the greatest similarity.The out-group G. geosporum and G. caledonium forma cluster distinct from either of the two abovegroups. The cophenetic correlation between thematrix used for the UPGMA clustering and thematrix of cophenetic values was 0-864, and wasinterpreted as a good fit.
DNA analyses
Three approaches were tested to analyse DNApolymorphisms, firstly with PCR with ITSribosomal primers; secondly PCR with specificprimers P0-M3 (Lanfranco et al., 1995) and lastlyPCR using primers of random sequences (RAPDs).Figure 5 shows that the ITS ribosomal regionamplified with ITS] and ITS4 primers had a similarsize {c. 600 bp) in the Glomus sp. (BEG 49), G.mosseae (ROTH-BEG 12) and G. versiforme (USA),whereas a lower molecular weight band was observedfor G. fiavisporum (Walker). On closer inspection,slight differences were observed in fragment sizebetween G. mosseae (ROTH-BEG 12) and Glomussp. (BEG 49) generated by the primers ITSl and

120 y. C. Dodd and others
0 . 5 6
M 1 2 3 4 5 1 2 3 4 5
Fig. 5. Undigested or Hini I-digested ITS region from thefollowing species: lane 1, Glomus sp. (BEG 49); lane 2,G, tnosseae (ROTH-BEG 12); lane 3, G. versiforme (USA);lane 4, G. fiavisporum (Walker); lane 5, Gigasporamargarita (BEG 34); lane M, lambda DNA digested withHind III—i?foR I, Some of the low molecular bands in theHini I digestions are PCR-aspecific products and/orprimer artefacts since they are present sometimes also inthe undigested samples.
2 3 5 6
kbp
0-56 —
Fig. 6. PCR amplification products using PO—M3 primers(Lanfranco et ai., 1995): lane 1, Glomus sp, (BEG 49);lane 2, G, mosseae (ROTH-BEG 12); lane 3, G, versiforme(USA); lane 4, G. fiavisporum (Walker); lane 5, Gigasporamargarita (BEG 34); lane 6, no sample; lane M, lambdaDNA digested with Hind III-BcoR L
M 1kbp
Fig. 7. RAPD amplification products using OPA-7, OPA-11 and OPA-18 primers on templates from: lane 1,Glomus sp, (BEG 49); lane 2, G. mosseae (ROTH-BEG12); lane 3, Gigaspora margarita (BEG 34); lane M,lambda DNA digested with Hind III-iicoR I. Each re-action IS repeated twice,
ITS4 before and after digestion by Hini 1 (Fig. 5).By contrast, the fragtnents generated for G.versiforme (USA) after Hin{ I digestion were of acompletely different length (Fig, 5). Using theprimers P0-M3 (Lanfranco et aL, 1995) gave theexpected specific amplification of about 550 bp for
the G. mosseae (ROTH-BEG 12) isolate (Lanfrancoet aL, 1995) whilst no signal was detected for Glomussp. (BEG 49) or any of the other templates (Fig, 6),The R.4,PD amplification profiles (Fig. 7) obtainedby using OPA-7, OPA-11 and OPA-18 randomprimers differed clearly between G. mosseae (ROTH-BEG 12) and Glomus sp. (BEG 49) and Gi. margarita(BEG 34),
DISCUSSION
The use of MDH and esterase isozyme profilesclearly allows characterization of YV or brownfunnel-shaped isolates frona different biogeo-graphical origins into either the G. mosseae or G.coronatum groups and so supports the division of thetwo groups at the species level. Other investigationswith different fungal groups have prodtjced similarresults using isozymes to deflne species previouslyonly explored using morphological traits(Oudemans, Forster & Coffey, 1994), although theability to culture their organisms allowed moreenzytnes to be tested. The successful use of only twoisozymes to compare the organisms in this studyshould not detract from the consistently useful dataobtained in similar studies using these two enzymeswith other well-defined isolates of AM fungi(Rosendahl et aL, 1994), The morphological dataindicate that spore colour (in a broad sense) was theonly character discriminating the G, mosseae isolatesfrom those of the darker brown G. coronatum isolates,Isozyme analysis of the MDH and esterase loci,however, clearly distinguished the two groups, G.dimorphicum fell distinctly within the YV G. mosseaegroup; consideration should be given to re-classi-fying it as an isolate of G, mosseae, based on ourmorphological and isozyme data. The G, coronatumgrouping also included Glomus sp, (AD-1), whichwas not initially considered to belong to either groupbecause it had a different wall structure, a straighterhyphal attachment and a lack of sporocarps incultures at Kent and the Northern Research Station,Edinburgh (UK), although it did have a G.mosseaejG. coronatum-\ike septum in the attachment.It also had the same spore colour profile as the G.coronatum group. SDS profiles and cluster analysisof the data indicated a close sitniiarity between G.coronatum (BEG 28) and Glomus sp. (AD-1) withinthe dark brown funnel-shaped Glomus spp. Thismay imply that a broader definition of whatencompasses the species definition of G. coronatum isneeded or that more isolates of this group need tobe studied to see if Glomus sp. (AD-1) is an anotnaly.We originally hypothesized that these dark brownfunnel-shaped Glomus spp, occurred only in semi-arid or arid calcareous soils and were ecologicallyseparated from the YV G. mosseae isolates whichwere never found in the same soil types. More recentobservations have noted the occurrence of YV G,

Variation in two Glomus species 121
•mosseae isolates in the same biogeographic zones andthere are preliminary indications that they mightoccur in the same soils (Dodd, Jeffries & Walker,unpublished). Ali the brown funnel-shaped Glomussp. isolates and Glomus sp. (AD-1) used in our studycame from soils in semi-arid or arid zones and itwould therefore be interesting to compare theisolates from Poland and Germany reported byBlaszkowski (1994) to see if they too fall within theclusters produced by isozyme analysis and SDSprotein profiles. Bentivenga & Morton (19946)considered colour and ornamentation to be tertiarycharacters not useful at the higher taxonomiccategories but capable of delimiting glomaleanspecies. They suggested a future well-definedweighting of characters based on developmentalpatterns.
This problem of selecting appropriate morpho-logical characters for taxonomic differentiation hasbeen highlighted in the use of sporocarp formation todelimit certain species. Our morphological studyindicates that sporocarpic formation might only beuseful to delimit the G. mosseae/G. coronatumcomplex (Meier & Charvat, 1992) and under moreenvironmental control depending on culture con-ditions. The ability to form sporocarps has beenknown to disappear occasionally in some isolates(Walker & Giovannetti, unpublished). A speciescalled Glomus monosporum (Gerdemann & Trappe,1974) is described which also forms sporocarpsreportedly containing only one spore, one of theoriginal reasons for separating it fro'm G. mosseae.Our data suggest that spore number in sporocarps isa variable characteristic and its use as a taxonomiccharacter is doubtful. The isolate of G. mosseae(BEG 25) was considered to be an isolate of G.monosporum in previous publications (Dodd et al.,1987) but has now been placed within the G. mosseaecomplex on the basis of this and earlier studies(Hepper et al., 1988). The presence of the outerhyaline wall on YV G. mosseae isolates was not aconsistent feature on mature spores nor was it usefulas a diagnostic character for G. coronatum, whereonly in ex-type material was it observed to expand inacidic mountants (Giovannetti et al., 1991), and notin all spores in this study. The degree of variation inspore size (data not presented) and biogeographicorigin within both G. mosseae isolates and thosegrouping with G. coronatum was refiected in theisozyme analysis. The esterase locus revealed isolatevariation for the former group but the MDH locishowed most variation for the latter group.
DNA fingerprints obtained with different PCRapproaches demonstrated that the isolate Glomus sp.BEG 49) is different from the other Glomus species.The P0-M3 primers which were designed for G.•nosseae (Lanfranco et al., 1995) confirmed theiripecificity, failing to amplify DNA from Glomus sp.BEG 49). These results strongly suggest that the
two Glomus isolates can be considered as belongingto two genetically different groups, Lloyd-MacGilpet al. (1996) also compared Glomus sp. (BEG 49)with different isolates of G. mosseae (including BEG12) and found that the ITS] and 2 sequences fromthe former were clearly distinct (11-16% diver-gence) from the other isolates of G. mosseae.Interestingly the pattern after Hini I digestion of theG. mosseae (ROTH-BEG 12) ITS regioncorresponds to that shown by Sanders et al. (1995)who also used this isolate.
In conclusion, therefore, much of the confusion indistinguishing species in the Glomales, and par-ticularly in the genus Glomus, can be attributed toinadequate taxonomic characterization that hasresulted in possible synonymy. The species in theGlomus mosseae complex (G. mosseae, G. monosporum,G. dimorphicum, G. fragilistratum and possibly G.fecundisporum) are all morphologically very similar,and the new species descriptions have not alwaysinvolved a review of type material, but rather haverelied on the original species descriptions(protologues). Evidence presented here indicatesthat there is wide morphological and molecularvariation among isolates of fungi that will fit thedescription of G. mosseae/G. coronatum. Thisunderlines and reinforces the need for a co-ordinatedeffort to examine species of similar morphologieswith a unified view of their taxonomic characters.Only by clarifying the species concept can we beginto discover more about the true biodiversity of thesefungi and the possible ecological roles played bypopulations of AM fungi. A range of molecularcharacters has been used to study AM fungiincluding: PCR-RELP of ribosomal DNA (Sanderset al., 1995), RAPD-PCR (Lanfranco et al., 1995),ITS sequences (Lloyd-MacGilp et aL, 1996),isozymes (Rosendahl, 1989), total soluble proteins(this study), glycoproteins (Wright, Morton &SviJorobuk, 1987; Thingstrup et al., 1995) and fattyacids (Jabaji-Hare, 1988; Bentivenga & Morton,1994 a); the users of each have addressed the meritsor drawbacks of their approaches. The present study,however, is one of the first to address the criticismsof Morton & Bentivenga (1994) in being careful indefining correct inter- and intraspecific comparisonsby prior characterization of the organisms involved.The use of isozymes and soluble protein profiles hasproduced data to establish two clear sub-groups ofthe two species of AM fungi investigated in thisstudy and to support the morphological inter-pretation. The use of G. mosseae-specific primersand DNA fingerprinting has provided further e '̂i-dence to support this taxonomic separation. Thisstudy, using the combined expertise of morpho-logical taxonomy and of the developing molecularapproaches, provides a blueprint for future investi-gations of the current taxononmic and systematicproblems of this important ancient fungal group.

122 J.C. Dodd and others
ACKNOWLEDGEMENTS
The authors J.C.D., C.W. and M.G. would like to thankthe President (Dr Silvio Gianinazzi) and managementcommittee of COST Action 8.21 for providing travelgrants to allow the initiation of this project. M.G. wouldlike to thank Dr L. Avio for his help in preparing theSDS-PAGE. The authors would also like to thank DrVestberg, Dr Mehrotra, Dr Cuenca, Dr D. McArthur andM. Verinumbe for providing some original soil samples orcultures. J .C.D. would also like to acknowledge two ECprojects CI1*-CT91-0904 and EV5V-O488 which havehelped generate cultures used in this study. L.L. wishes tothank the National Research Council of Italy for fundingthe molecular vi-ork and the EC-IMPACT project B102-CT93-OO53.
REFERENCES
Anonymous. 1969. Royal Botanic Garden Edinburgh. Flora ojBritish fungi. Colour identification chart. Edinburgh, UK: HerMajesty's Stationery Office.
Bentivenga SP, Morton JB. 1994a. Stability and heritability offatty acid methyl ester profiles of glomalean endomycorrhizalfungi. Mycological Research 98: 1419-1426.
Bentivenga SP, Morton JB. 19946. Systematics of glomaleanendomycorrhizal fungi: current views and future directions.In: Pfieger FL, Linderman RG, eds, Mycorrhizae and PlantHealth. St Paul, MK, USA: American PhytopathologicalSociety Press, 283-308.
Berbee ML, Taylor JW. 1993. Dating the evolutionary radiationsof the true fungi. Canadian Journal oJ Botany 71: 1114-1127.
Blaszkowski J. 1994. First record and notes on Glomus coronatumin Poland and Germany. Mycologia 86: 630-634,
Bradford MM. 1976. A rapid and sensitive method for thequantification of microgram quantities of protein utilising theprinciple of protein-dye binding. Analytical Biochemistrv 72:248-254.
Dodd JC, Krikun J. 1984. Obser'i'ations on endogonaceous sporesin the Negev de:sert (Israel). Transaction.^ of the BritishMycological Society 82: 536-540.
Dodd JC, Burton C, Burns RG, Jeffries P. 1987. Soilphosphatase activity around roots of three host plants infectedwith VAM fungi. New Phytologist 107: 163-172.
Dodd JC, Gianinazzi-Pearson V, Rosendahl S, Walker C.1994. European Bank of Glomales — an essential tool forefficient international and interdisciplinary collaboration, ln:Gianinazzi S, Schuepp H, eds. Impact of ArbuscutarMycorrhizas on Sustainable Agriculture and Natural Ecosystems.Basel, Switzerland: Birkhauser Press, 41-46.
El-Giahmi AA, Nicolson T, Daft M. 1976. Endomycorrhizalfungi from Libyan soils. Transactions of the British MycologicalSociety 67: 164-169.
Gerdemann J, Trappe JM. 1974. The Endogonaceae of thePacific Northwest. Mycological Memoir No. 5. MycologicalSociety of America, New York Botanical Garden, New York,
Giovannetti M,, AvSo L, Salutini L. 1991. Morphological,cytochemica] and ontogenetic characteristics of a new species ofvesiciilar-arhuscular mycorrhizal fungus, Canadian Journal ofBotany 69: 161-167.
Giovannetd M, Giaeinazzi-Pearson V. 1994. Biodiversity inarbuscular mycorrhiza] fungi. Mycological Research 98:703-715,
Giovaneetti M, Nicolson TH. 1983. Vesicular-arbuscularmycorrhizas in Italian sand dunes. Transactions of the BritishMycolagical Society 80: 552-557,
Hepper CM, Sen R, Azeon-Agiiilar C, Grace C. 1988.Variation in certain isozymes amongst different geographicalisolates of the vesicular-arbuscular mycorrhizal fungi Glomusclarmn, Glomus monosporum and Glomus mosseae. Soil Biologyand Biochemistry 20: 51-59.
Jabaji-Hare S. 1988. Lipid and fatty acid profiles of somevesicular-arbuscular mycorrhizal fungi: contribution to tax-onomy, Mycoiogia 80: 622-629,
Laemmli UK. 1970. Cleavage of structural proteins during theassembly of the head of bactetiophage T4, Nature 227:680-685,
Lanfranco L, Wyss P, Marzachi C, Bonfante P. 1995.Generation of RAPD-PCR primers for the identification ofisolates of Glomits mosseae, an arhuscular mycorrhizal fungus.Molecular Ecology 4: 61-68.
Lloyd-MacGilp SA, Chambers SM, Dodd JC, Fitter AH,Walker €,, Young JPW. 1996. Diversity of the internaltranscribed spacers wdthin and among isolates of Glomus mosseaeand related arbuscular mycorrhizal fungi. Neti' Phytologist, thisvolume, OO'O-OOO,
Meier R, Charvat I. 1992. Peridial development in Glomusmosseae (Glomaceae), A^nerican Journal of Botany 79: 928-936.
Morton JB, Bentivenga SP. 1994. Levels of diversity inendomycorrhizal fungi (Glomales, Zygomycetes) and their rolein defining taxonomic and non-taxonomic groups. Plant andSoil 159: 47-59,
Mosse B, Bowen GD. 1968. A key to the recognition of someEndogone spore types. Transactions of the British MycologicalSociety 51: 469-4'83.
Omar MB, Bolland L, Heather WA. 1979. A permanentmounting medium for fungi. Bulletin of the British MycologicalSociety 13: 31-32.
Oudemans P, Forster H, Coffey MD. 1994. Evidence fordistinct isozyme subgroups within Phytophthora citricola andclose relationships with P. capsici and P. cilrophthora. Mv-cological Research 9S: lS9-\99.
Pyrozynski KA, Dalpe Y. 1989. Geological history of theGlomaceae with particular reference to mycorrhizal symbiosis.Symbiosis 7: 1-36,
Rohlf FJ. 1994. NTSYS, Numerical taxonomy and multivariateanalysis system. Exeter Software, ver, 1,80.
Rosendahl S. 1989. Comparisons of spore-cluster foriningGlomus species (Endogonaceae) based on morphologicalcharacteristics and isoenzyme banding patterns. Opera Botanica100:215-223,
Rosendahl S, Dodd JC, Walker C. 1994. Taxonomy andphylogeny of the Glomales. In : Gianinazzi S, Schuepp H, eds.Impact of Arhuscular Mycorrhizas on Sustainable Agricultureand Natural Ecosvstems. Basel, Sxvitzerland: Birkhauser Press,1-12.
Sanders IR, Alt M, Groppe K, Boiler T, Wiemken A. 1995.Identification of rihosomal DNA polymorphisms among andwithin spores of the Glomales: application to studies on thegenetic diversity of arbuscular mycorrhiza] fungal communities,Neic Phytologist 130: 419^27.
Simon L, Bousquet J, Levesque RC, Lalonde M. 1993. Originand diversification of endomycorrhizal fungi and coincidencewith vascular plants. Nature 363: 67-69.
Sward RJ, Hallam ND, Holland AA. 1978. Endogone spores ina heathland area of South-Fastern Australia, Australian Journalof Botany 26: 2 9 ^ 3 .
Taylor TN, Remy W, Hass H, Kerp H. 1995. Fossil arbuscularmycorrhizae from the Farly Devonian. Mycologia 87 : 560-573,
Thingstrup I, Rozycka M, Jeffries P, Rosendahl S, Dodd JC.1995. Detection of the arbuscular mycorrhizal fungusScutellospora heterogama within roots using polydonal aotiseia.Mycological Research 99: 1225-1232,
Walker C. 1983. Taxonomic concepts in the Endogonaceae:spore wall concepts in species descriptions. Mycotaxon 18:443-455.
Walker C. 1992. Systematics and taxonomy of the arhuscularendomycorrhizal fungi (Glomales) - a possible way forward,Agronomie 12: 887-897.
WalkerC, Trappe JM. 1993. Names and epithets in the Glomalesand Fndogonales. Myeological Research 97: 339-344.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification anddirect sequencing of fungal ribosomal RNA genes forphylogenetics. In: Innis MA, Gelfaud DH, Sninsky JJ, WhiteTJ, eds, PCR Protocols. A Guide to Methods and Applications.San Diego: .Academic Press, 315-322,
Williams JGK, Kubelik AR, Livak KJ, Rafalski JA, TingeySV. 1990, DNA polymorphisms amplified by arbitrary primersare useful as genetic markers. Nucleic Acid Research 18:6531-6535.
Wright SF, Morton JB, Sworobuk JE. 1987. Identification of avesicular-arbuscular mycorrhizal fungus hy using monoclonalantibodies in an enzyme-Hnked immunosorbent assay. Appliedand Environmental Microbiology 53: 2222—2225.
Wyss P, Bonfante P. 1993. Amplification of genomic DNA ofarbuscular-mycorrhizal (AM) fungi by PCR using shortarbitrary primers. Mycological Research 97: 1351-1357.

Phytol. {1996), 133, 123-134
The genetic diversity of arbuscularmycorrhizal fungi in natural ecosystems - akey to understanding the ecology andfunctioning of the mycorrhizal symbiosis
BY IAN R. SANDERS\ JUSTIN P. CLAPP- AND ANDRES WIEMKEN^
^Botanical Institute, University of Basle, CH-4056 Basle., Switzerland^Department of Biology, University of York, PO Box No. 373, York YOl 5YW, UK
(Received 13 November 1995)
SUMMARY
The mi'corrhizal symbiosis formed between plant roots and the arbuscular mycorrhizal (AM) fungi or Glomalesis of great interest to ecologists because of its potential influence on ecosystem processes, its role in determiningplant diversity m natural communities and the ability of the fungi to induce a wide variety of growth responsesin coexisting plant species. Little attention, however, has been paid to the ecological role of diversity of AM fungi.Difficulties in identification, the inability to grow the fungi in pure culture, problems of taxonomic classification,and a lack of basic information on the life histories of AM fungi hinder studies of the ecological significance ofdiversity of .AM fungi. Nucleic acid based techniques have the potential to fill this gap in our knowledge by offeringbetter means of identification and the opportunity to study links between the genetic diversity of AM fungi andfunctional and morphological diversity. The application of genus-specific molecular markers has shown thatdifferent genera of AM fungi coexist in plant roots and that this is a common occurrence. Molecular techniqueshave alsO' shown that natural AM fungai populations exhibit unexpectedly high genetic diversity, despite theassumption that diversity in these seemingly asexual fungi should be low. The high diversity occurs in multicopytibosomal genes and their internal transcribed spacers, which are normally well conserved and homogeneouswithin an individual organLsm. The results show that sequence heterogeneity of the ribosomal genes can occur evenin single spores of AM fungi, and we discuss how genetic diversity m.ay be promoted and maintained. Contrastingresults, indicating that genetic diversity among replicate spores from pot-cultured material is low (even thoughthey contain within spore sequence heterogeneity), suggest that there are mechanisms which promote high geneticdiversity of AM fungi in natural ecosystems.
We propose that AM fungi could be heterokaryotic as a result of the exchange of nuclei following hyphal fusionwith other individuals but that other mechanisms, such as gene turnover and molecular drive, might also explainthe generation of high genetic diversity without any exchange of genetic material among individuals. The highdiversity in ribosomal gene sequences in AM fungi might cause problems in their use as molecular markers in fieldstudies. \ better understanding of the levels of genetic diversity of ribosomal genes within spores, among sporesof the same morphology, and among spores of differing morphology is essential to the development of soundmolecular markers for field studies and to the development of a phylogenetic classification.
We concltide that an understanding of tbe mechanisms which promote and maintain genetic diversity in the AMiungi is crucial, not only to further advances in ecological and evolutionary studies but also to studies of theTiolecular basis of the regulation of the symbiosis. Moreover, we predict that while observational investigationsm ."VM fungal ecology and diversity using molecular techniques are of high value they will not give aninderstanding of the role of AM fungi in natural ecosystems and that further studies should also aim to fill the;aps in current knowledge of links between genetic diversity and distribution of AM fungi in natural ecosystems,;nd their functional diversity,
vey words: Ribosomal genes, internal transcribed spacer, molecular drive, gene turnover, Glomales.
without using any specialized or sophisticated tech-N T R O D U C T I O N . T i !• • i j l •/= •
niques. In additton, the taxonomj/ and classificationThe distribution, abundance and life history of of many plant and animal groups is well establishednany plants and animals can easily be observed to the species level. This information forms a sound

124 /. R. Sanders, J. P. Clapp and A. Wiemken
basis from which to pursue experimental studies oftheir ecology, evolution and role within naturalecosystetns. In the case of some organisms, par-ticularly those below ground, the difficulty of thetask of describing their taxonomy, distribution,abundance and life history hinders meaningfulexperimental investigations of their role in naturalecosystems. One such group of organisms is theZygomycete fungi of the order Glomales, which arealso commonly known as arbuscular mycorrhizal(AM) fungi. The AM fungi form the arbuscularmycorrhizal symbiosis with the roots of plants, andin nnany cases this association is beneficial for bothpartners (Harley & Smith, 1983). Despite the lack ofbasic ecological and biological information about theAM fungi, they are of particular interest to ecologists(Fitter, 1990).
The lack of basic ecological information about theAM fungi stems from a few fundamental difficultiesencountered in their identification, culture andtaxonotny, coupled with the great difficulty inmanipulating AM fungi in natural ecosystems with-out greatly modifying their environment in otherways. Some of the problems of identification, andpossibly of taxonomy, can be overcome by usingmolecular techniques, and these techniques couldprovide us with an insight into the structure ofnatural communities of AM fungi. In this review weoutline briefly why the understanding of AM fungalecology is important to the understanding of thefunctioning of ecosystems, why the study of AMfungal diversity warrants attention, precisely whatthe gaps in our knowledge are, what moleculartechniques are available to fill in the gaps and howthe application of molecular techniques has alreadycontributing to our understanding of AM fungaldiversity and of the role of .4M fungi in ecology. Thetechniques available have only recently beenmodified for specific use with AM fungi,, and westress that the results that we present (includingresults from our own complementary researchprogrammes in York and Basle) are preliminary, buthave already yielded interesting and, indeed, sur-prising information. We also discuss their implic-ations in the understanding of the molecularmechanisms which promote and maintain diversityof AM fungi in natural ecosystems.
THE ECOLOG1C.1L SIGNIFICANCE OF AM
FUNGAL DIVERSITY
The potential impact of AM fungi on the functioningof ecosystems is not insignificant. Arbuscular mycor-rhiza are ubiquitous, with c. 90 % of terrestrial plantspecies forming the AM symbiosis. The AM fungican induce large growth benefits in the host plant. Inmany cases, the growth benefit has been attributed toimproved plant phosphorus nutrition resulting froman increased efficiency ir> phosphorus acquisition
from the soil by the extraradicle AM fungal hyphae(Koide, 1991). In return, the plants donate carbo-hydrates to the fungal partner (Smith & Smith.1990) and this carbohydrate can move from oneplant to another through hyphae (Grime et aL,1987). Thus, at the ecosystem level the AM fungihave the potential to influence both the carbon andphosphorus cycles.
Recentl}', great emphasis has been placed onunderstanding how ecosystems function and, inparticular, the role that diversity (or biodiversity)plays in ecosystem; the impetus being the concernover both global and local losses in biodiversity andthe effects that these might have on the environment.AM fungi have been shown to play a significant rolein the fioristic diversity and structure of annual andperennial plant communities (Grime et aL, 1987;Gange, Brown & Farmer, 1990; Sanders & Koide,1994). However, in most of these ecological in-vestigations, little or no attention has been paid to thediversity of AM fungi themselves. This is becauseexperiments conducted in pots have indicated thatAM fungi show little host specificity. Consequently,it was thought that few selection pressures thatwould favour extensive divergence arise in mutu-alistic symbioses (Law, 1985). In support of this, themorphological diversity of AM fungi is, indeed, low(c. 152 species in six genera; Walker & Trappe,1993). However, Jakobsen, Abbott & Robson (1992)have shown that different isolates of AM fungi canresult in diiTerent effects on plant growth. Un-fortunately, few of the experiments demonstratingdifferent efTects of AM fungal isolates on plantgrowth have been conducted in an ecological context;the fungi used have been isolated from different soilsand environmental conditions but have then beencompared on the basis of their functionality withplants with which they might never naturally grow.
Studies conducted in Basle on the Swiss Bio-diversity Programme have used a collection of fungiwhich were all isolated from the same plant com-munity (a species-rich calcareous grassland calledNenzlinger Weide). Their functionality with severalplant species from the grassland has been assessed;functionality is defined as an effect on the overallgrowth, growth habit, vegetative and reproductivefitness of the plant and of the fungus. The resultshave shown that isolates indeed differ in their effectson plant growth but that these effects differ accordingto plant species. An extreme case is seen in the twodominant coexisting grasses at Nenzlinger Weide,Bromus erectus and Festuca ovina. One fungal isolatefrom the field site induced faster growth in Festucaovina and a reduced growth rate (similar to that ofthe non-mycorrhizal situation) of Bromus erectus;another isolate from the same site had exactly theopposite effect (M. van der Heijden, pers. comm.).Other experiments on the functionality of thesefungal isolates with Prunella vulgaris and P. grandi-

Genetic diversity of AM fungi 125
ora have indicated that AM fungal diversity islikely to affect the plant's reproductive strategy and,ul particular, whether reproduction is likely to besexual or asexual (clonal) (Streitwolf-Engel et al.,unpublished). These studies outline the potentialimportance of diversity of AM fungi in naturalcommunities.
Understanding the real, rather than the potential,significance of AM fungal diversity in naturalcommunities has posed a greater problem. For agiven plant community we need to know how diversethe AM fungal community is, which plant roots arecolonized by which AM fungi, whether there isseasonality in the patterns of colonization, whetherany specificity' between plants and AM fungi occursand what the effect of those AM fungi will be onplants and the ecosystem. However, the difficulty ofidentifying AM fungi in the roots of plants hasalways been an obstacle to their study in naturalcommunities. Although identification of AM fungiin roots based on morphological observations hasbeen successful in pot experiments (Abbott, 1982; J.Merryweather, pers. comm.) the usefulness of thistechnique for field investigations must be limitedbecause the hyphal morphology of one .\M fungalisolate is likely to vary with host species (Gailaud,1905; Gerdemann, 1965; Lackie et aL, 1987).Molecular or nucleic acid techniques have been usedsuccessfully for in planta identification of ecto-mycorrhizai fungi (Gardes et al., 1991; Bruns &Gardes, 1993; Gardes & Bruns, 1993) and endo-phytic fungi such as Epichloe sp. (Groppe et al.,1995) and the techniques available for identificationof AM fungi in plant roots are described in'Molecular Techniques Available for Studying AMFungal Fcology' (below).
TAXONOMY AND DIVERSIFICATION OF AM
FUNGI
The arbuscular mycorrhizal symbiosis is ancient;the fossil record suggests that it occurred in the firstland plants (Pyrozynski & Dalpe, 1989; Remy et al.,1994). Morphological diversification is low, withonly 152 recognized species in six genera (Walker &Trappe, 1993). Law (1985) pointed out that con-sidering the age of the symbiosis this diversifications low, and he suggests that this is typical ofnutualistic symbioses, as there are few external•ressures which would favour the selection of newtaits. However, the low divergence of AM fungieems at odds with their functional diversity,nformation regarding the genetic diversity of AM™gi can also be obtained using molecular tech-iques and the results of such investigations can, andave, helped to construct a phylogenetic classi-cation of AM fungi (Simon, Levesque & Lalonde,993).Until recently,, methods for studying AM fungal
diversity and the ecology of the symbiosis have beenwholly reliant on the morphology of the spore phase.Spores of AM fungi are relatively large, easy toextract from soil, and have several morphologicalcharacters that allow species identity to be de-termined by experienced personnel. However, theuse of spore data alone for the assessment ofecosystem diversity and ecology has long beenrecognized as unsatisfactory (Walker, Mize &McNabb, 1982; Klironomos et al., 1993; Rolden-Fajardo, 1994). Situations can be envisaged whichcould give rise to misleading conclusions, such as thepresence of non-sporulating fungi, spore types thatare isomorphic but are in fact distinct, or thepresence of spores in soil in differing numbers beingassumed to reflect the situation within the root. Inaddition, data derived from observation of sporenumbers reflect past events in the symbiosis; howthis information relates to the contemporary situ-ation is at best uncertain. The study of spores alone,therefore, is a suspect method for the study of theecology of the symbiosis and is especially likely tolead to a low estimation of diversity.
The taxonomy of AM fungi has also been based onthe morphology of spores and, therefore, ecologicalstudies have also been reliant on the taxonomicclassification. The ecological worth of constructingdata on AM fungal community structure which arebased on morphological characteristics is question-able, since the relationship between the morpho-logical diversity of AM fungi and their genetic andfunctional diversity has not been established. Ac-cording to Walker (1992) many described taxa are' workable' in that they are recognizable and arefound repeatedly in different parts of the world,,albeit in diverse environments. For example, severalspecies of AM fungi which were reported in a semi-arid site in Australia (McGee, 1989) also occurred ina species-rich meadow in the north of England(Sanders, 1993), both ecosystems comprising com-pletely different soils, vegetation and climate. Thisindicates that AM fungi are very plastic in theirenvironmental adaptability. There are many otherexamples in the literature of the enormous plasticityof AM fungal species which is reflected by theirdistribution in extremely diverse environments. Thelack of host specificity in the mycorrhizal symbiosisalso indicates that the .A.M fungi are very plastic withrespect to host range. How this enormous plasticitycomes about in AM fungi is not yet explained.
Molecular techniques can give information aboutthe genetic diversity of AM fungal species whichmight help to explain these phenomena but the lattermust only be adequately resolved if individual clonesof AM fungi can be identified and recognized so thatgenetic characteristics can be used to identify themorphological, functional and ecological differencesbetween AM fungi (Walker, 1992). However,, it isuncertain whether AM fungal clones exist. The

126 I. R. Sanders, J. P. Clapp and A. Wiemken
fungi are coenocytic organisms where single sporescontain thousands of nuclei (Burggraaf & Beringer,1989; Bianciotto & Bonfante, 1992), Although AMfungi are thought to be asexual, it is not certainwhether their spores are homokaryotic or hetero-kary-otic (Walker, 1992), The likelihood of theexistence of heterokaryote AM fungal spores isdiscussed in ' Inriplications for the Biology of the AMFungi', below.
MOLECULAR TECHNIQUES AVAILABLE FOR
STUDYING AM FUNGAL ECOLOGY
Techniques for studying genetic diversity
It is not our aim to describe all the moleculartechniques available for ecological studies but tooutline those which have been used, or modifiedspecifically, for studying AM fungi, and those whichare likely to be useful for future studies. For adescription of the use of molecular techniques inecology we refer the reader to Bachmann (1994), Formany organisms, a DNA fingerprint for ident-ification or for the estimation of genetic diversity canbe achieved by using restriction fragment lengthpolymorphism (RFLP) analysis. The technique canonly be used where a large amount of DNA can beobtained from individual organisms. Because AMfungi cannot be maintained in pure culture, obtain-ing sufficient DNA for RFLP is difficult. Conse-quently, all the nucleic acid techniques employed foridentifying AM fungi or for studying their geneticdiversity use the polymerase chain reaction (PCR),which amplifies relatively small amounts of DNA,
A universal molecular phylogeny has been basedlargely on sequences of the small subunit ribosomalRN.A gene (18S in eukaryotes and 16S in pro-karyotes) and other regions of ribosomal DNA(Winker & Woese, 1991), For elucidating thephylogenetic relationships of fungi, the 18S and 5,8SrRNA genes, along with flanking regions known asinternal transcribed spacers (ITS), can be amplifiedand sequenced (White et al., 1990), The 18S and5,8S genes evolve relatively slowly and are useful forstudies of distantly-related organisms. The ITSregion evolves faster than the 18S and 5,8S genesand can differ between species. The advantage ofusing these regions of DNA is that multiple copies ofthe genes are present in all organisms. It is nosurprise, therefore, that the first report of DNAamplified from AM fungi using the polymerasechain reaction (PCR) and sequenced was from the18S rRNA gene (Simon, Lalonde & Bruns, 1992),The DNA was amplified from small numbers ofspores using so-called universal primers; shortlength oligonucleotides which are complementary tohighly conserved sequences and which thereforecause the amplification of DNA from a wide varietyof difTerent organisms, Simon found that the sequen-
ces of the 18S rRNA gene differed between speciesof AM fungi and, following the design of a Glomale?specific primer (VANSl) was able to amplify andsequence the 18S rRNA gene from several differentspecies of AM fungi which represented all the generaof the Glomales (Simon et al., 1993a),
The studies of Simon et al. (1992) demonstratethat if sequences of common genes differ betweenspecies or isolates of AMF it is possible to use thesefor identification purposes and for studies of geneticdiversity. The techniques have been applied to AMfungal spores, and the ITS region (which is thoughtto be less conserved than the 18S) has been amplifiedfrom single spores of several isolates (Sanders et al.,1995), Amplification of DNA from single sporespermits studies of AM fungal diversity in naturalpopulations, and Sanders et al. (1995) used it for thispurpose, ITS regions from single spores of differentknown .AM fungal isolates were amplified using theuniversal primers ITSl and ITS4, By cutting theITS fragment with restriction enzymes, differentfingerprints could be obtained for the differentisolates. The technique, known as PCR-RFLP, wasused for the rapid identification of genetically diversespores. The method is, however, severely limitedunless the ITS regions are actually sequenced, Adifference in banding pattern between two amplifiedproducts from two different spores confirms that thelatter are genetically different. However, if nodifferences are found between the two fingerprintsthe result does not mean that the two ITS regions areidentical, but might mean that the correct restrictionenzyme for finding those differences was not used.Sequencing of the ITS regions is an obvious way toovercome this. However, in field investigations, thegenetic diversity of AM fungal spores was so greatthat PCR-RFLP was sensitive enough to detectgenetic diversity in the community without the needto sequence the amplified products (Sanders et al.,1995),
Another technique, known as random amplifiedpolymorphic DNA (RAPD-PCR) is also availablefor studies of genetic diversity. The technique ispotentially more sensitive in detecting genetic difTer-ences between individuals, and by screening largenumbers of arbitrary primers, the desired sensitivitycan be found for detecting genus, species orindividual genotype differences, Wyss & Bonfante(1993) successfully used RAPD-PCR to identifyspores at the level of species and to differentiatebetween isolates of the same species. So far thetechnique has not been used for studying thediversity of natural AM fungal communities.
Techniques for identifying AM fungi in roots
If the sequence of a known region of DNA differsbetween species or isolates it is possible to developspecific primers for their identification and quanti-

Genetic diversity of AM fungi 127
ication. Once primers are developed they can bejsed to identify AM fungi from spore, hyphal orcolonized root material. As many ecological studiesof AM fungi are aimed at determining the re-lationship hetween the host and colonist the mostdesirable site from which to obtain valid informationIS clearly the root itself.
Initial uses of molecular methods involved iso-zymes. Hepper, Sen & Maskall (1986) found that itwas possible to detect the presence of single andmixed species of Glomus in pot cultures of maize byobserving allozyme patterns. This work was ex-tended into the first assessment of AM fungi in asemi-natural grassland by Rosendahl, Rosendahl &S0chting (1990) and showed that several distinctfungi could be isolated from a single host plant,indicating that not only could more than one genetict3'pe exist in a single plant but also that the fungaldiversity might be greater than was expected frominformation derived from spore abundance. Imnnu-nological methods have also been developed fordetection of AM fungi in roots but have not beenused for field investigations (Wright & Morton,1989; Sanders et al., 1992; Hahn, Horn & Hock,1995). Currently, however the most favoured tech-niques available use the extremely high definitiongiven by DNA sequence information for the identi-fication of AM fungal spores and endophytichyphae. All these methods use the polymerase chainreaction (PCR) or its derivatives.
Simon et al. (1992) used universal fungal primersto sequence the 18S ribosomal RNA gene fromspores of Glomus intraradices and Gigasporamargarita. The sequence information obtained wasused to design a primer that was specific to theGlomales, VANSl. They showed that this primer,when used in conjunction with the universal primerNS21, was able to amplify AM fungal DNA directlyfrom leek roots infected with Glomus vesiculiferumand this has been repeated in four other plants (DiBonito, Elliott & Des Jardin, 1995). Following thedevelopment of family-specific primers (Simon,Levesque & Lalonde, 1993 b) this work was extendedto identify different genera of AM fungi in roots.Simon et al. (199341) also exploited single strandconformational polymorphism (SSCP) rapidly toscreen PCR-amplified DNA sequences for variation.SSCP has been reported to be able to detect single3ase substitutions between homologous DNA se-juences (Ainsworth, Surh & Coulter-Mackie, 1991;Vlakino et al., 1992). These differences are detected>y electrophoresing the samples on a high-resolution>oK'acrylamide gel under conditions where the twotrands of the amplified product run as two in-'ependent molecules, their rate of progress beingiirectly affected by their conformation within theiiatrix of the gel. Simon et al. (1993^i) showed thatmplified fragments differing by only a few basesould be detected and selected for sequencing. This
technique could prove to be a useful screeningmethod before resorting to lengthy sequencingprojects for the assessment of AM fungal diversity.
There have, as yet, been few studies in which theuse of nucleic acid information has been extendedinto a non-laboratory environment. Clapp et al.(1995) found difficulty with the VANSl primerwhen used for the PCR amplification of AM fungalDNA from field-collected bluebell roots (Hya-cinthoides non-scripta). This apparently stemmedfrom interference by the host plant DNA. Whendilution of the template DNA was found to have noeffect, the host-plant DNA was removed by sub-tractive hybridization, the overall process beingnamed SEAD (selective enrichment of amplifiedDNA), whereafter amplifications using VANSl weresuccessful. Parallel control pot experiments, withonion roots experimentally infected with Glomusmosseae, were found to be easy to amplify withoutresorting to SEAD.
A different approach to the development ofspecies-specific primers was taken by Lanfranco etal. (1995) who compared banding profiles generatedby RAPD-PCR to identify unique bands in differentAM fungi. These bands were cloned and sequenced,and the information used to design species-specificprimers. There is difficulty, however, in directlyapplying RAPD-PCR based techniques to the endo-phytic stage as the primers will almost certainlyproduce PCR products derived from the host plantDNA. It might, however, be possible to utilize aSEAD-based procedure to allow the identification ofsingle infections. The identification of multiplecolonizations by RAPD-PCR might prove moredifficult.
Nucleic acid based technology can potentially beused for the quantification of AM fungal colonizationand is essential for making estimations of the relativeabundance of different AM fungi and allowing anassessment to be made of co-existence of AM fungiin roots. Simon, Levesque & Lalonde (1992) havedeveloped a quantitative PCR technique for theestimation of AM funga! colonization. The PCRamplification of a fragment from an AM fungus andthe deletion of a small number of bases allows theproduction of an amplification standard that whencloned and introduced into the early stages of a DNAextraction will subsequently co-amplify with theroot-derived AM fungal template. A comparison ofthe ratio of brightness of the two bands produced bystaining with ethidium bromide on an agarose gel(with a small correction for the lower size of thestandard) reflects the initial concentration of thetemplate with respect to the known concentration ofthe standard, provided that the conditions of thePCR are identical. This technique is being usedextensi\'ely at the University of York with theprimers PO and M3 which were found to be morereliable than VANSl for the quantification of

128 J. R. Sanders, J. P. Clapp and A. Wiemken
colonization of Glomus mosseae under experimentalconditions (S. E. Edwards, pers. comm.). Problemswith the formation of hybrid molecules between thefungal product and the standard, represented as athird band on the gel, have been largely overcome bythe use of totally unrelated template DNA asten:iplate for the amplification. This is isolated byPCR amplification of DNA from an unrelatedorganism using the sanae primers at low annealingtemperature (Uberla et aL, 1991), resulting in amolecule of similar size to, but with no sequencehomology with, the AM fungal template sequence.
Population genetics
The methods described are of limited use for studiesof population genetics of .A-M fungi. A traditionalapproach in population genetics is to use isozymeanalysis but this can be expensive and can requirelarge amounts of material. In the case of non-culturable organisms such as AM fungi, where thesmallest individual unit is a spore, sufficient materialmight not always be readily available. An alternativeis to use microsatellite markers. Microsatellite DNA(also known as short tandem repeats or simplesequence length polymorphisms) are stretches oftandem mono-, di-, tri- and tetranucleotide repeatsof various lengths. The loci containing micro-satellites act as heritable, but selectively neutralmarkers (they are usually non-coding regions ofDNA), which often differ in length between indi-viduals. Their use for studies of genetic diversity,gene How in populations, maternity and paternityanalysis and relatedness, and genetic distancebetween individuals, species and populations, isreviewed by Bachmann (1994). .Although, to ourknowledge, microsatellite markers have not yet beenfound for the Glomales there is no reason to supposethat they do not exist. Regions of microsatelliteDNA have been found in the Epichloe endophyte(Groppe et aL, 1995) and the length of the regiondiffers between genetically different individuals ofthe fungus. Development of specific primers toamplify a region of Epichloe' DN.4 containing themicrosatellite region, for use in studies of diversity inpopulations of these fungi, has been successful. Byreviewing the GenBank and EMBL sequence data-bases, Groppe et al. (1995) have also shown thatmicrosatellite DNA is common in naany fungi andtherefore there is no reason to expect that it does notoccur in the Glomales.
Linking genetic diversity and functionality of AMfungi in natural ecosystems
The methods described above are all for theidentification of AMF or the detection of geneticdiversity among .-̂ M fungi. In each case, the targetedregions of DNA which are used for characterizationof AM fungi are either genes which bear little
relation to the symbiosis-specific functions of AMfungi, e.g, the ribosomal genes or, in the case ofRAPD-PCR and microsatellite DNA, are frequentlynon-coding regions. Thus, obtaining markers forstudying the functional ecology of AM fungi isunlikely to advance with the use of these techniques.It is possible that certain genes are symbiosis-specific and that their expression indicates particularfunctions occurring in the symbiosis. These could bevaluable for field studies if their expression could bedetected and quantified.
A possible method for the detection of sytnbiosis-specific genes would be the technique of differentiallyexpressed mRNA display (Liang, Averboukh &Pardee, 1993). The procedure works by extractingmRNA from two individuals, one of which hasundergone a specific treatment; in the case of studiesof AM fungi this might be from both colonized anduncolonized roots. A reverse transcription PCRreaction (RT-PCR) converts mRNA into cDNAwhich is then amplified (Bauer et al., 1993). Thedifferential display of mRNA, which is a represent-ation of differential gene expression between thetwo treatments, can then easily be observed on a gel.The result is that bands which are specific to themycorrhizal symbiosis can be identified,, clonedand sequenced. Finding out whether sequencedregions are of fungal or plant origin would requiresubsequent verification b\' Southern hybridizationanalysis. The technique would allow an experimentalapproach to understanding how AM fungi interactwith environmental variables by manipulating en-vironmental conditions, and by observing differentialgene expression in the symbiosis. Such an approachcould also lead to the development of genetic markersfor symbiosis-specific functions for use in fieldstudies. This would give an insight into the geneticregulation of the AM symbiosis under manipulatedenvironmental conditions. In addition, carrying outdifferential display techniques on plants colonizedby different AM fungal isolates would allow theidentification of genes which may be specific to aparticular plant/AM fungal isolate interaction, thefirst step' to finding a link between genetic andfunctional diversity among AM fungi. To ourknowledge, the use of differential display for thepurposes of understanding AM fungal interactionswith the environment has not yet been performed.
A second approach to isolating specifically ex-pressed mRNAs involved in the interaction betweenthe AM fungal colonist and its host plant could beachieved by subtraction methodology. Such pro-cedures combine cD NA products resulting from RT-PCR of, for example, AM fungal colonized and un-colonized plants in such a way that cDNA fragmentscommon to both, and therefore not relevant to theinteraction, would be lost. The only cDNAs that areretained are those that were induced in the host plantby the presence of the fungus. As in the case of

Genetic diversity of AM fungi 129
iifferentially expressed mRNA, the plant or fungaljrigin of resultant products would have to be verifiedby Southern hybridization, but this approach has theadvantage that far fewer cDNA products will beinvolved in the final screening, allowing easierinterpretation. This technique has been successfullyused to isolate differentially expressed genes inducedby salt stress (Guhck & Dvorak, 1990) and low-temperatures (Aguan et al., 1991). There are manymore examples of its use for the isolation ofdifferentially expressed mRNAs and there is noreason why it cannot be used to isolate genes inducedby AM fungal colonization.
MOLECUL.'iR EVIDENCE FOR THE DIVERSITY OF
AM FUNGI
Diversity of AM fungi in laboratory-culturedmaterial
On the basis of the 18S ribosomal RNA genesequences, Simon et al. (1993 a) and Simon (1996)have produced a phylogeny for the Glomales whichthey compared to the major periods of divergence ofterrestrial plants. The numbers of base differencesamong species and genera of the Glomales were usedto estimate the likely periods of divergence of thedifferent genera. The data of Simon et al. (1993 a)corroborate the fossil and taxonomic e\'idence of thedivergence of the Glomales, based on spore mor-phology, and confirm that AME are ancient and thatthere has been little diversification (Pyrozynski &Dalpe, 1989; Remy et al., 1994).
Evidence from other molecular studies suggeststhat the genetic diversity of mycorrhizal fungi is notquite so simple. Using RAPD-PCR, Wyss &Bonfante (1993) developed fingerprints from dif-ferent isolates of Glomus mosseae. Some of the isolatesoriginated from the same culture but were thenmaintained in different laboratories for several years.Slightly different fingerprints were found betweenthose isolates, which could indicate that geneticdifferences in the Glomales can arise over relativelyshort periods of time (c. 12 yr).
Information from the sequencing of the ITSregion has revealed surprising information about thepossible genetic diversity of the Glomales. Sandersst al. (1995) were able to obtain reproducible specificingerprints from ITS regions of AMF using PCR-RFLP. The DNA came from single spores orig-aating from single spore isolates. Some fingerprints;ave multiple banding patterns which, whenummed, did not add up to the original size of theTS region. This suggested that different sequencesf the ITS region were amplified from single spores'Ut in a reproducible manner (other spores from theame culture yielding the same pattern). Cloning andequencing of the amplified ITS region from singlepores of Glomus mosseae revealed that different
sequences of the ITS region are, undoubtedly,present in single spores. Two sequences obtainedfrom one spore were sufficiently similar to suggestthat they are closely related. Comparison withanother, similar, ITS sequence obtained followingphage cloning, confirmed that the sequences werefrom Glomus mosseae (Franken & Gianinazzi-Pearson, 1996). The implications of these findings forthe diversity of the arbuscular mycorrhizal symbiosisare discussed in ' Innplications for the Bioiogy of theAM Fungi', below.
The diversity of AM fungi in natural ecosystems
There is very little information on the diversity ofAM fungi in natural ecosystems, although evidenceis emerging that the diversity is greater than has beeninferred from morphological studies of spores.Typical estimates of species diversity in naturalecosystems on the basis of spore counts rangebetween approx. 5—20 different species in a com-munity. Using the PCR-RFLP method to analysethe genetic diversity' in natural AM fungal popu-lations, Sanders et al. (1995) found that the ITSregion of ten morphologically identical spores of thegenus Glomus were genetically different. This resultindicates an unexpectedly high heterogeneity in theITS region in natural populations of AM fungi.
Investigations of the diversity of natural Scutel-lospora populations in an oak woodland in the northof England highlight the diversity that can occuramong and within spores in natural populations ofthe Glomales. In order to confirm that the VANSlprimer site was conserved in the Scutellospora speciesat the study site, DNA was extracted from single,field-collected spores and PCR-amplified using SS38(universal primer) and VAGIGA (family-specificprimer) primers (designed by Simon et al. 19936).These primers amplify a region of DNA whichshould contain the VANSl primer site. The PCRproducts were cloned and several clones weresequenced from each spore. More than one 18Sribosomal RNA gene sequence occurred in a singlespore, confirming the evidence from the ITSsequences of the genetic diversity of single spores.More surprisingly the sequences contained differentvariants of the sequence corresponding to theVANSl primers, usually varying by three bases overits 21 bp length, indicating that the VANSl site isnot conserved (Clapp et al., unpuhlished). It isconceivable that these sequence differences couldreduce the efficiency of annealing of the VANSlprimer and explain the difficulties that have pre-viously been encountered using this primer toamplify from field samples (Clapp et al., 1995). Theresults raise questions regarding the validity of usingVANSl sequences for the phylogenetic classificationof the Glonriales (see ' Implications for the Biology ofthe AM Fungi' below).

130 /, R. Sanders, jf. P. Clapp and A. Wiemken
Despite the confusing and overwhelming geneticdiversity seen in natural populations and communi-ties of the Glomales, studies using ribosomal genesequences have given the first indications of dis-tribution of AM fungi in the roots of a natural plantcommunity. Studies at the University of York haveconcentrated on the Hyadnthoides non-scriptaljAM.fungi symbiosis which has been described in relationto the life history and phosphorus nutrition of H.non-scripta (Merryweather & Fitter, 1995 a, ft). Tothis end, amplified DNA from both spores andendophytic hyphae have been sequenced to associatethe rhizosphere species with the endophytic stagesand to characterize the distribution of AM fungi intime and space in the roots of these plants. The workhas concentrated on the 5' end of the IBS rRNA geneand the results obtained so far appear to imply thatthe situation is more complex than at first envisaged.Initial studies, using SEAD (Clapp et al., 1995),found that more than one fungal genus could berepresented in a single root of bluebell and that thisseems to be the normal situation rather than anexception. Up to three different genera of AM fungi{Acaulospora, Glomus and Scutellospora) are repre-sented in the roots of the bluebell population. Thepresence of Acaulospora and Scutellospora fungi inthe roots was expected on the basis of spore countsmade from the soil at the field site, although Glomussequences within the roots were not expected, asspores from this genus had been found onlyinfrequently in the soil. This observation indicatesthat it is very likely that many more species arepresent in roots than can be found by solelysurveying spores in the rhizosphere.
Direct sequencing of PCR products derived fromthe family-specific primers (Simon et at., 1993i)and VANSl in these roots, however, was notsuccessful as more than one sequence was present.To overcome this difficulty, the DNAs were clonedand two colonies were sequenced from each family-specific amplification. In every case the sequencesfrom each genus were found to be different (Clappet al., 1995), This apparent diversity is undergoingfurther investigation which concentrates on thegenus Scutellospora from the same field site. Similarstudies on field-collected roots have also yieldedseveral sequences, but whether these can be assignedto different species or to multiple ribosomal genesequences within single species, is as yet impossibleto say (Clapp et al., unpublished).
To date, it appears that molecular informationobtained from colonized roots in natural ecosystemsis likely to identify the presence of many species orgenotypes of AM fungi that are as yet undetected. Itmay be possible tO' counter the difficulties inobtaining fungal sequence data from natural eco-systems (possibly due to the variation within theVANSl site) by the use of degenerate primers whichmight then allow direct amplification of sequences
from roots without the need to resort to morecomplicated procedures such as SEAD,
IMPLICATIONS FOR THE BIOLOGY OF THE AM
FUNGI
Implications for the maintenance of genetic variationin AM fungi
Molecular studies of sequences of ribosomal genes inthe Glomales reveal unexpectedly wide geneticdiversity, especially in organisms which are thoughtto be asexual. We are not aware of other organisms inwhich different ribosomal genes occur within asingle individual, A major question now is whetherthe variation is a result of the presence of distinctgenotypes in different nuclei in the same spore(heterokaryotic) or gene heterogeneity within singlenuclei. With our current lack of knowledge of biologyof AM fungi either or both scenarios are possible.
The heterokaryote hypothesis proposes that dif-ferent sequences of ribosomal genes occur in difTerentnuclei in a single spore. This is highly pertinent,because the presence of different genotypes of nucleioccurring in single spores has strong implications forthe biology, ecology and evolution of the mycorrhizalsymbiosis, Heterokaryote spores could not be con-sidered as genetic individuals but as populations.This raises questions of how the heterokaryote hasarisen and whether there is any transfer of nucleibetween hyphae of different AM fungi, Hetero-karyotic mycelia occur frequently in higher fungi(Alexopoulos & Mims, 1979) and exchange of nucleifollowing anastomosis occurs in many filamentousfungi, A similar mechanism has been proposed forthe diversification of Epichloe endophytes in Festucarubra following the discovery of several differentsequences of ^-tubuiin {tub2) gene copies in thefungus which can best be explained by the hybrid-ization of vegetative hyphae (Tsai et al., 1994),
The high genetic diversity that would be main-tained in this way may help to explain how an asexualorganism can exhibit such high plasticity both inenvironment and host range and why the Glomalesseem to give remarkably inconsistent results in manyscientific experiments. It is also tempting to believethe heterokaryote hypothesis when such high geneticdiversity has been observed in both the York andBasle studies among and within morphologicallyidentical spores in natural populations. In contrast,results from pot-cultured material from single sporeswith PCR-RFLP suggest genetic consistency amongspores from the same culture (even though they aregenetically diverse in a single spore). This indicatesthat some mechanism must be operative in naturalpopulations of AM fungi which promotes andmaintains genetic diversity. The presence of genetic-ally diverse nuclei (heterokaryotic) does notnecessarily demonstrate that the exchange of genetic

Genetic diversity of AM fungi 131
material occurs between individual fungi. Variationin ribosomal genes among or within nuclei of AMfungi could occur by other mechanisms, and those ofgene turnover and molecular drive are addressedbelow.
Knowledge of gene turnover and molecular drive(Hoelzel & Dover, 1991) may explain the occurrenceof highly variable ribosomal gene sequences. Themultiple copies of ribosomal genes are normallyhomogeneous within a single genome. It is becauseof this homogeneity that ribosomal gene and ITSsequences are useful for studies of phylogeny. Insexual organisms, incomplete alignment and sub-sequent unequal crossing over can occur duringrecombination. These events are responsible for thegeneration of multicopy genes (Smith, 1976) and formaintaining homogeneity of the sequences (Dover,1986 ; Hoelzel & Dover, 1991). In asexual organisms,mitotic recombination occurs very infrequently(Darnell, Lodish & Baltimore, 1990). As a conse-quence, the opportunity for unequal crossing overwould also be infrequent. Therefore, perhaps acertain amount of heterogeneity among the copiescould be expected, owing to mutations which are notfrequently corrected by unequal crossing over. Thiscould explain the high variability in the ribosomalgene sequences seen within single spores of theGlomales if the Glomales are asexual. Sexualreproduction events have not been observed in theAM fungi, although this does not rule out thepossibility of their occurrence. If sexual repro-duction were a regular occurrence in the .^M fungi,then different sequences of ribosomal genes wouldbe unlikely to occur within a single nucleus orbetween different nuclei (assuming that recombin-ation were a regular event) because recombinationwould maintain homogeneity of the gene copies.Therefore, our results point strongly to AM fungibeing asexual. If the lack of frequent recombinationaccounts for the genetic variability of ribosomalgenes in single spores of the Glomales then thedifferent sequences could, theoretically, occur bothwithin and among nuclei in a spore. It would seemthat the distribution of different ribosomal genesequences in the spore can best be elucidated if singlenuclei can be isolated by microdissection, thenamplified by PCR. However, this might also notprovide the conclusive evidence of the mechanisms-ontroliing genetic variation in the Glomales.
Unequal crossing over is not the only mechanismvhich helps to maintain homogeneity among geneopies. Gene conversion can also be responsible forhis homogeneity. The process of gene conversionlormally occurs during recombination but can also'ccur in mitotic cells, although it is a much rarervent. Some gene conversion in mitotic cells prob-bly occurs during DNA replication, wheremismatch repair mechanisms are in operationKourilsky, 1986). This would allow some repair of
mutated ribosomal gene copies. Whether the mech-anism of gene conversion in the absence ofrecombination can also be responsible for the in-corporation of new gene sequences throughout thegene family is unclear. In the case of the Glomales,the question is whether the rate of gene conversionproceeds at a rate fast enough to repair the ribosomalgenes as the mutations occur. This would causehomogeneity of ribosomal genes within a nucleus,but heterogeneity of sequences among nuclei in thesame spore. The other possibility is that mutationscould occur at a much faster rate than geneconversion, therefore resulting in some hetero-geneity of gene sequences within and among nuclei.
Implications for a phylogeny of AM fungi
The existence of rRN.\ sequence variation withinsingle spores of AM fungi puts into question thevalidity of the use of these sequences for under-standing the phylogeny of this fungal group. In view'of the probable lack of recombination in the AMFand the unchecked mutations which could occur, thevariation seen in the ITS region in spores is notsurprising, because mutations are much more likelyto persist in non-coding regions than in codingregions. This is because selection reduces thenumber of deleterious mutations persisting in apopulation, whereas mutation in non-coding regionswould frequently be selectively neutral (Wilson,Ochman & Prager, 1987). However, the variation inthe VANSl region observed in a natural populationof Scutellospora indicates that the suggested phy-logeny based on 18S ribosomal gene sequenceregions (using the VANSl primer), might notindicate the phylogeny of the AM fungi themselvesbut rather the phylogeny of those 18S gene sequenceswhich were selected from spores because theycontained the conserved VANSl region itself. Ahigh level of diversity' in the 18S gene would then bemissed. A similar problem has previously beenaddressed in relation to the phylogeny of bacteriallineages based on the 16S ribosomal gene (Woese,1987,1992); the question being whether the bacterialphylogenetic tree is really a phylogeny of the bacteriaor of the 16S ribosomal gene. In the case of thebacteria, this question arises because genes can betransferred horizontally, i.e. bacteria contain anunknown number of promiscuous genes and, there-fore, the molecular phylogeny based on some genesmight have little relation to the true phylogeny of theorganism. Establishing how much the ribosomalgene sequences can vary within single spores,between spores of identical morphology and betweenmorphologically different spores will resolve theproblem of relating ribosomal gene sequences tophylogeny in AM fungi and, therefore, whetherribosomal gene sequences can be used for con-structing a phylogeny of .^M fungi. In the case of the

132 /. R. Sanders, J. P. Clapp and A. Wiemken
ITS regions, only a small number of base differenceswere found between two sequences within a singleGlomus mosseae spore and many more base dif-ferences in one ITS sequence obtained from Glomusgeosporum, indicating that the differences betweenspores of different morphology are much greaterthan the differences within a single spore. It must beretnembered, however, that the fungi came fromsingle spore isolates and, therefore, replicate sporesfrom the same isolate would be genetically similar.An experiment has been carried out at the Universityof York to compare the interspecific and intraspecificvariation of ITS regions of several species of Glomus(Lloyd-MacGilp et al., 1996). The intraspecificvariation has been found to be as great as thatbetween some species. In addition, intra-sporesequence variation has also been observed. ThisrRNA gene diversity supports the variation de-scribed by Sanders et al. (1995), Clapp et al. (1995)and Clapp et al. (unpuhlished).
However, the fact that phylogeny based on the 18Sgene sequence containing the VANSl primer site(Simon et al., 1993) fits well with conventionalstudies of Glomales phylogeny, based on sporemorphology, cannot be ignored.
Implications of distribution of AM fungi in the rootsof natural plant communities
The knowledge that can be obtained regarding thedistribution of AM fungi in the roots of natural plantcommunities is still limited by the lack of goodspecies-specific molecular markers for AM fungithat can be used successfully in field investigations.One clear problem here is the lack of knowledge ofgenetics of AM fungi. If there is little homogeneityamong gene sequences in a population then findingreliable markers is difficult. Despite these problems,the work of Clapp et al. (1995) with the family-specific markers clearly shows the coexistence ofdifferent AM fungi in the roots of an individual plantand, therefore, the potential for competitive inter-actions to occur among AM fungi. This has strongcomplications for mycorrhizal functioning since it isknown that different isolates of AM fungi can havedifferent effects on the nutrition and growth of plants(Jakobsen et al., 1992). McGonigle (1988) foundlittle evidence for a relationship between colonizationby AM fungi and stimulation of growth under fieldconditions. It has been suggested by Fitter (1990)and Sanders & Fitter (1992) that coexisting AMfungi might be responsible for the lack of thisrelationship because the growth stimulation by onefungus might be masked by the presence, anddifferent stimulation of growth, by another fungus.The development of further markers for AM fungiwill allow the possibility of investigating this in thefuture.
Competitive interactions among coexisting AM
fungi within the root have not been studied in thefield. However, there is evidence from pot ex-periments that competition between AM fungi occursin plant roots (Pearson, Abbott & Jasper, 1993) andthat it is mediated by the host. The presence ofseveral genera, and possibly species, within singleroots raises questions regarding the mechanisms ofcoexistence of AM fungi and whether the fungicould be temporally and spatially separated in theroot to avoid direct competition. Furthermore, thefunctional role of coexisting AM fungi needs to beexamined. Such investigations may require the useof fiuorescent oligonucleotide probes to identify thevarious AM fungi within the root.
FUTURE DIRECTIONS IN RESEARCH ON AM
FUNGI
Understanding the genetics of AM fungi
We now have interesting data regarding the geneticsof AM fungi but much more work is required toelucidate the mechanisms controlling the geneticdiversity' in these fungi. The task clearly is of highpriority, not only for ecological studies of AM fungibut in order to understand the relationships betweenecology, functionality and morphology in this group.If the genetic diversity in AM fungi is as great as theresults from the York and Basle projects suggest,then it could also have important implications forinvestigations into the underlying molecular pro-cesses governing the development of the symbiosis.There is good reason to suggest that if the processesleading to the genetic diversity of AM fungi are notunderstood they could lead to inconsistent, irre-producible and uninterpretable results in these typesof investigations. If we cannot find conservation inribosomal gene sequences than what chances do wehave of finding universal or conserved symbiosis-related genes from the fungal side of the arbuscularmycorrhizal symbiosis ?
New molecular markers for field studies of AM fungi
The continuation of investigations into ecology ofAM fungi in natural populations requires thedevelopment of reliable molecular markers. Tbe useof ribosomal genes might turn out to be inap-propriate for this purpose and the development ofother likely candidates will probably have to await abetter knowledge of the genetics of AM fungi. Untilthen we do not know whether other genes with alower copy number can be conserved in the genomesof Glomales or whether these genes are suitable foruse as markers in field studies. However, what wemay find is that taxa of AM fungi have particular setsof sequences associated with them which allow aspecies to be identified by a sequence fingerprint. It

Related Documents











