Drug Resistance Updates 15 (2012) 237–247 Contents lists available at SciVerse ScienceDirect Drug Resistance Updates journa l h omepage: www.elsevier.com/locate/drup Insights into the global molecular epidemiology of carbapenem non-susceptible clones of Acinetobacter baumannii Nabil Karah a,b , Arnfinn Sundsfjord a,b , Kevin Towner c , Ørjan Samuelsen a,∗ a Reference Centre for Detection of Antimicrobial Resistance, Department of Microbiology and Infection Control, University Hospital of North Norway, Tromsø, Norway b Research Group for Host-Microbe Interactions, Department of Medical Biology, Faculty of Health Sciences, University of Tromsø, Tromsø, Norway c Department of Clinical Microbiology, University Hospital Queen’s Medical Centre, Nottingham, UK a r t i c l e i n f o Article history: Received 18 April 2012 Accepted 29 June 2012 Keywords: A. baumannii Carbapenemases MLST International clones Multidrug resistance Resistance islands a b s t r a c t The global emergence of multidrug resistance (MDR) among Gram-negative bacteria has dramatically limited the therapeutic options. During the last two decades, Acinetobacter baumannii has become a pathogen of increased clinical importance due to its remarkable ability to cause outbreaks of infections and to acquire resistance to almost all currently used antibiotics, including the carbapenems. This review considers the literature on A. baumannii and data from multilocus sequence typing studies to explore the global population structure of A. baumannii and detect the occurrence of clonality, with the focus on the presence of specific resistance mechanisms such as the OXA-carbapenemases. The worldwide dissemination of MDR and carbapenem non-susceptible A. baumannii is associated with diverse genetic backgrounds, but predominated by a number of extensively distributed clones, such as CC92 B /CC2 P and CC109 B /CC1 P , which have frequently been supplemented by acquired OXA-type carbapenemase genes. © 2012 Elsevier Ltd. All rights reserved. 1. Background 1.1. Acinetobacter baumannii – a pathogen of increasing importance A. baumannii is a non-fermentative, oxidase-negative, non- motile, strictly aerobic, Gram-negative coccobacillary organism (Perez et al., 2007). Although cases of community-acquired infec- tions have been reported, the primary pathogenic activity of A. baumannii is undoubtedly linked to hospital-acquired infections, mostly among immunocompromised patients in intensive care units (Dijkshoorn et al., 2007). A. baumannii is mainly implicated in ventilator-associated pneumonia, catheter-related bloodstream and urinary tract infections, and wound infections. The risk fac- tors predisposing individuals to infection by A. baumannii include: (i) major trauma, particularly burns, and surgery, (ii) previous antimicrobial therapy, (iii) prolonged hospital and ICU stay, and (iv) utilization of mechanical ventilators, drainage tubes, and indwelling catheters. However, careful clinical judgment is always required to separate between cases of infection and nosocomial colonization (Perez et al., 2007). ∗ Corresponding author at: Reference Centre for Detection of Antimicrobial Resis- tance, Department of Microbiology and Infection Control, University Hospital of North Norway, Postbox 56, N-9038 Tromsø, Norway. Tel.: +47 77627043; fax: +47 77627015. E-mail address: [email protected] (Ø. Samuelsen). A. baumannii is characterized by remarkable capabilities for the acquisition of antibiotic resistance determinants, intra- and inter-hospital outbreak spread, and national and international clonal dissemination (Dijkshoorn et al., 2007). The occurrence and dissemination of A. baumannii poses major medical and organiza- tional burdens to healthcare facilities since infections caused by A. baumannii are difficult to treat, and because eradication of this pathogen from the hospital environment and prevention of further spreading might require solid targeted measures, such as the isola- tion of patients and temporary closure of wards (Dijkshoorn et al., 2007; Perez et al., 2007). Comparative typing of outbreak strains of A. baumannii from geographically scattered European hospitals demonstrated the occurrence of three successful clones, originally named “European clones I–III” (Dijkshoorn et al., 1996; van Dessel et al., 2004). These three European clones have subsequently been identified worldwide and accordingly re-named as “international clones I–III” (Diancourt et al., 2010). In addition to these major clones, a wide geographic distribution of some other clones has been reported (Giannouli et al., 2009; Towner et al., 2008). The emergence of such highly successful clones seems to play a leading role in the increasing global occurrence of A. baumannii infections (Diancourt et al., 2010). 1.2. Antimicrobial resistance – the emergence of carbapenem non-susceptible strains Antibiotics of choice for the treatment of infections caused by A. baumannii include the aminoglycosides, fluoroquinolones, and 1368-7646/$ – see front matter © 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.drup.2012.06.001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ic
Na
b
c
a
ARA
KACMIMR
1
1i
m(tbmuiat(a(irc
tNf
1h
Drug Resistance Updates 15 (2012) 237– 247
Contents lists available at SciVerse ScienceDirect
Drug Resistance Updates
journa l h omepage: www.elsev ier .com/ locate /drup
nsights into the global molecular epidemiology of carbapenem non-susceptiblelones of Acinetobacter baumannii
abil Karaha,b, Arnfinn Sundsfjorda,b, Kevin Townerc, Ørjan Samuelsena,∗
Reference Centre for Detection of Antimicrobial Resistance, Department of Microbiology and Infection Control, University Hospital of North Norway, Tromsø, NorwayResearch Group for Host-Microbe Interactions, Department of Medical Biology, Faculty of Health Sciences, University of Tromsø, Tromsø, NorwayDepartment of Clinical Microbiology, University Hospital Queen’s Medical Centre, Nottingham, UK
r t i c l e i n f o
rticle history:eceived 18 April 2012ccepted 29 June 2012
eywords:
a b s t r a c t
The global emergence of multidrug resistance (MDR) among Gram-negative bacteria has dramaticallylimited the therapeutic options. During the last two decades, Acinetobacter baumannii has become apathogen of increased clinical importance due to its remarkable ability to cause outbreaks of infectionsand to acquire resistance to almost all currently used antibiotics, including the carbapenems. This review
. baumanniiarbapenemasesLST
nternational clonesultidrug resistance
considers the literature on A. baumannii and data from multilocus sequence typing studies to explorethe global population structure of A. baumannii and detect the occurrence of clonality, with the focuson the presence of specific resistance mechanisms such as the OXA-carbapenemases. The worldwidedissemination of MDR and carbapenem non-susceptible A. baumannii is associated with diverse geneticbackgrounds, but predominated by a number of extensively distributed clones, such as CC92B/CC2P and
freq
esistance islands CC109B/CC1P, which have. Background
.1. Acinetobacter baumannii – a pathogen of increasingmportance
A. baumannii is a non-fermentative, oxidase-negative, non-otile, strictly aerobic, Gram-negative coccobacillary organism
Perez et al., 2007). Although cases of community-acquired infec-ions have been reported, the primary pathogenic activity of A.aumannii is undoubtedly linked to hospital-acquired infections,ostly among immunocompromised patients in intensive care
nits (Dijkshoorn et al., 2007). A. baumannii is mainly implicatedn ventilator-associated pneumonia, catheter-related bloodstreamnd urinary tract infections, and wound infections. The risk fac-ors predisposing individuals to infection by A. baumannii include:i) major trauma, particularly burns, and surgery, (ii) previousntimicrobial therapy, (iii) prolonged hospital and ICU stay, andiv) utilization of mechanical ventilators, drainage tubes, andndwelling catheters. However, careful clinical judgment is always
equired to separate between cases of infection and nosocomialolonization (Perez et al., 2007).∗ Corresponding author at: Reference Centre for Detection of Antimicrobial Resis-ance, Department of Microbiology and Infection Control, University Hospital oforth Norway, Postbox 56, N-9038 Tromsø, Norway. Tel.: +47 77627043;
ax: +47 77627015.E-mail address: [email protected] (Ø. Samuelsen).
368-7646/$ – see front matter © 2012 Elsevier Ltd. All rights reserved.ttp://dx.doi.org/10.1016/j.drup.2012.06.001
uently been supplemented by acquired OXA-type carbapenemase genes.© 2012 Elsevier Ltd. All rights reserved.
A. baumannii is characterized by remarkable capabilities forthe acquisition of antibiotic resistance determinants, intra- andinter-hospital outbreak spread, and national and internationalclonal dissemination (Dijkshoorn et al., 2007). The occurrence anddissemination of A. baumannii poses major medical and organiza-tional burdens to healthcare facilities since infections caused byA. baumannii are difficult to treat, and because eradication of thispathogen from the hospital environment and prevention of furtherspreading might require solid targeted measures, such as the isola-tion of patients and temporary closure of wards (Dijkshoorn et al.,2007; Perez et al., 2007). Comparative typing of outbreak strainsof A. baumannii from geographically scattered European hospitalsdemonstrated the occurrence of three successful clones, originallynamed “European clones I–III” (Dijkshoorn et al., 1996; van Desselet al., 2004). These three European clones have subsequently beenidentified worldwide and accordingly re-named as “internationalclones I–III” (Diancourt et al., 2010). In addition to these majorclones, a wide geographic distribution of some other clones hasbeen reported (Giannouli et al., 2009; Towner et al., 2008). Theemergence of such highly successful clones seems to play a leadingrole in the increasing global occurrence of A. baumannii infections(Diancourt et al., 2010).
1.2. Antimicrobial resistance – the emergence of carbapenem
non-susceptible strainsAntibiotics of choice for the treatment of infections caused byA. baumannii include the aminoglycosides, fluoroquinolones, and

2 ance U
ctcfgm(
crdebrbqrems2
a(baaa22dabsbatiAubf
t2l(bbe2P2cA
tbsPbost(I
38 N. Karah et al. / Drug Resist
arbapenems (Dijkshoorn et al., 2007). A. baumannii is, in clinicalerms, intrinsically resistant to ampicillin, amoxicillin-clavulanate,efazolin, cefotaxime, ceftriaxone, ertapenem, trimethoprim, andosfomycin (http://www.eucast.org/expert rules/). However, thereatest problem concerns the large number of acquired resistanceechanisms that have increasingly been emerging in A. baumannii
Dijkshoorn et al., 2007; Peleg et al., 2008; Perez et al., 2007).Resistance to quinolones in A. baumannii is primarily caused by
hromosomal mutations in the quinolone resistance determiningegion (QRDR) of the gyrA and parC genes, and the subsequent pro-uction of modified bacterial DNA gyrase and topoisomerase IVnzymes (Peleg et al., 2008). A double mutation in the QRDRs ofoth GyrA and ParC is necessary to obtain high levels of quinoloneesistance in A. baumannii, with Ser-83 in GyrA and Ser-80 in ParCeing the most frequently mutated amino acid residues in theuinolone-resistant isolates (Vila et al., 1997). However, quinoloneesistance can also result from decreased influx and/or increasedfflux of quinolones (Peleg et al., 2008). Of note, the plasmid-ediated quinolone resistance aac(6′)-Ib-cr and qnr genes have,
o far, not been identified in A. baumannii (Adams-Haduch et al.,008; Peleg et al., 2008).
Resistance to aminoglycosides in A. baumannii is mainly medi-ted by the production of aminoglycoside-modifying enzymesAMEs) (Peleg et al., 2008). Genes encoding different AMEs haveeen detected in A. baumannii, including: the aphA1, aphA6, andphA15 phosphotransferase, the aacC1, aacC2, aacA4, aac(6′)-Iad,ac(6′)-Im, and aac(6′)-II acetylttransferase, and the aadB, aadA1,nd aadA4 nucleotidyltransferase genes (Adams-Haduch et al.,008; Boo et al., 2009; Cho et al., 2009; Han et al., 2008; Nemec et al.,004; Perez et al., 2007). The co-presence of more than one AMEeterminant and the occurrence of some of these genes (such asphA6, aacC1, aacA4, aadA1, and aadB) within class 1 integrons haveeen reported (Boo et al., 2009; Nemec et al., 2004). A. baumanniitrains producing the 16S rRNA methylase ArmA enzyme have alsoeen identified (Cho et al., 2009; Doi et al., 2007). ArmA mediates
high level of pan-aminoglycoside resistance, based on protectinghe antimicrobial target by post-transcriptional methylation lead-ng to loss of affinity of 16S rRNA for aminoglycosides (Doi andrakawa, 2007). The armA gene in A. baumannii was surroundedpstream by tnpU, a putative transposase gene, and downstreamy tnpD, another putative transposase gene, and was repeatedlyound to be plasmid-mediated (Cho et al., 2009; Doi et al., 2007).
Carbapenem resistance in A. baumannii is mainly mediated byhe production of carbapenem-hydrolyzing enzymes (Peleg et al.,008). Genes encoding a variety of carbapenem-hydrolyzing �-
actamases, belonging to class A (blaGES-14 and blaKPC), class BblaIMP, blaVIM, blaSIM-1, and blaNDM), and class D (blaOXA-23-like,laOXA-24-like, blaOXA-51-like, blaOXA-58-like, blaOXA-104, blaOXA-143,laOXA-164, and blaOXA-182), have been detected in A. baumannii (Chut al., 2001; Di Popolo et al., 2011; Kaase et al., 2011; Lee et al.,005; Lim et al., 2007; Lin et al., 2010; Moubareck et al., 2009;oirel and Nordmann, 2006a; Poirel et al., 2010; Robledo et al.,010). Associated with a notable diversity, the class D OXA-typearbapenemases are by far the most prevalent carbapenemases in. baumannii (Table 1) (Poirel and Nordmann, 2006a).
While blaGES-14, blaIMP, blaVIM, and blaSIM-1 are commonly foundo be located on class 1 integrons, blaNDM and the OXA-type car-apenemase genes have generally been associated with insertionequence (IS) elements (Lee et al., 2005; Moubareck et al., 2009;oirel and Nordmann, 2006a; Poirel et al., 2010). ISAba1 is proba-ly responsible for the mobility of the blaOXA-23-like genes by meansf either two surrounding copies forming a composite transpo-
on defined as Tn2006, or a single copy located at one side ofhe gene and forming a one-ended transposon defined as Tn2008Adams-Haduch et al., 2008; Corvec et al., 2007). A single copy ofSAba4 located upstream of blaOXA-23-like can also be responsiblepdates 15 (2012) 237– 247
for its mobility, by forming another one-ended transposon definedas Tn2007 (Corvec et al., 2007). Similarly, the blaOXA-58-like geneshave been linked with different IS elements, such as ISAba1, ISAba2,ISAba3, IS18, and ISAba825 (Poirel and Nordmann, 2006b; Ravasiet al., 2011).
The class B �-lactamases confer high levels of carbapenem resis-tance, as well as resistance to all other ß-lactams except aztreonam(Poirel and Nordmann, 2006a). In contrast, the substrate profileof the carbapenem-hydrolyzing class A, except for KPC, and classD �-lactamases is generally diverse, with most of them showinglow to moderate hydrolytic activities against carbapenems, partic-ularly meropenem (Moubareck et al., 2009; Poirel and Nordmann,2006a; Poirel et al., 2010; Robledo et al., 2010). Interestingly, resis-tance to carbapenems mediated by the coexistence of blaNDM-1,blaOXA-23, and blaIMP has been identified in several pan-drug resis-tant A. baumannii isolates from China (Chen et al., 2011a). Similarly,the coexistence of armA, blaOXA-23, and blaNDM-1 has recently beenreported in three clinical isolates from India (Karthikeyan et al.,2010).
Importantly, a number of resistance islands have recently beenidentified among MDR isolates of A. baumannii, mainly belonging toeither international clone I or II (Adams et al., 2008; Fournier et al.,2006; Nigro and Hall, 2012). The majority of these islands, desig-nated as AbaR islands, are characterized by being transposed into aspecific position in the chromosomally located comM gene, creatinga duplication of five nucleotides “ACCGC” (Post et al., 2010). Over-all, the AbaR islands detected among isolates from internationalclone II are different in their genetic structure from those iden-tified among isolates belonging to international clone I (Nigro andHall, 2012). However, the increasingly detected AbaR4-like islands,characterized by carrying a blaOXA-23-like enzyme within Tn2006,has been described among isolates from both international clones Iand II (Adams et al., 2008; Hamidian and Hall, 2011; Hornsey et al.,2011; Turton et al., 2011). All of these islands have most likely beenderived from Tn6021, a backbone transposon found in the suscep-tible A. baumannii strain ATCC 17978 (Post et al., 2010; Smith et al.,2007).
1.3. Molecular methods for strain typing
Several molecular methods with different resolution have beenused to type A. baumannii strains (Dijkshoorn et al., 2007). A typ-ing scheme based on two multiplex polymerase chain reactions(PCRs) targeting three genes under selective pressure (ompA, csuE,and blaOXA-51-like) has been a convenient method for rapid assign-ment of A. baumannii isolates into three major PCR-based groups(Gs) corresponding to international clones I (G2), II (G1), and III (G3)(Turton et al., 2007). Using this scheme, additional groups (G4–G7and putative G8–G14) have been defined according to new combi-nations of the PCR amplicons (Table 2) (Bonin et al., 2011; Giannouliet al., 2010; Grosso et al., 2011a; Karah et al., 2011a,b; Towner et al.,2008). Of note, the labels proposed for the PCR-based Gs do not cor-respond to those proposed by another typing approach based oncomparative sequence analysis of the same three genes (Giannouliet al., 2009, 2010; Turton et al., 2007).
Sequence analysis of the blaOXA-51-like gene has also been pro-posed as a useful typing method for A. baumannii isolates, giventhat evident correlation has been detected between particularblaOXA-51-like enzymes and particular epidemic lineages (Evanset al., 2008; Hamouda et al., 2010; Turton et al., 2007). In this regard,a study conducted on sixty A. baumannii isolates collected world-
wide demonstrated that isolates belonging to international cloneII encoded blaOXA-51-like enzymes belonging to the OXA-66 clusterwhile those belonging to international clone I encoded enzymesfrom the OXA-69 cluster (Evans et al., 2008; Hamouda et al., 2010).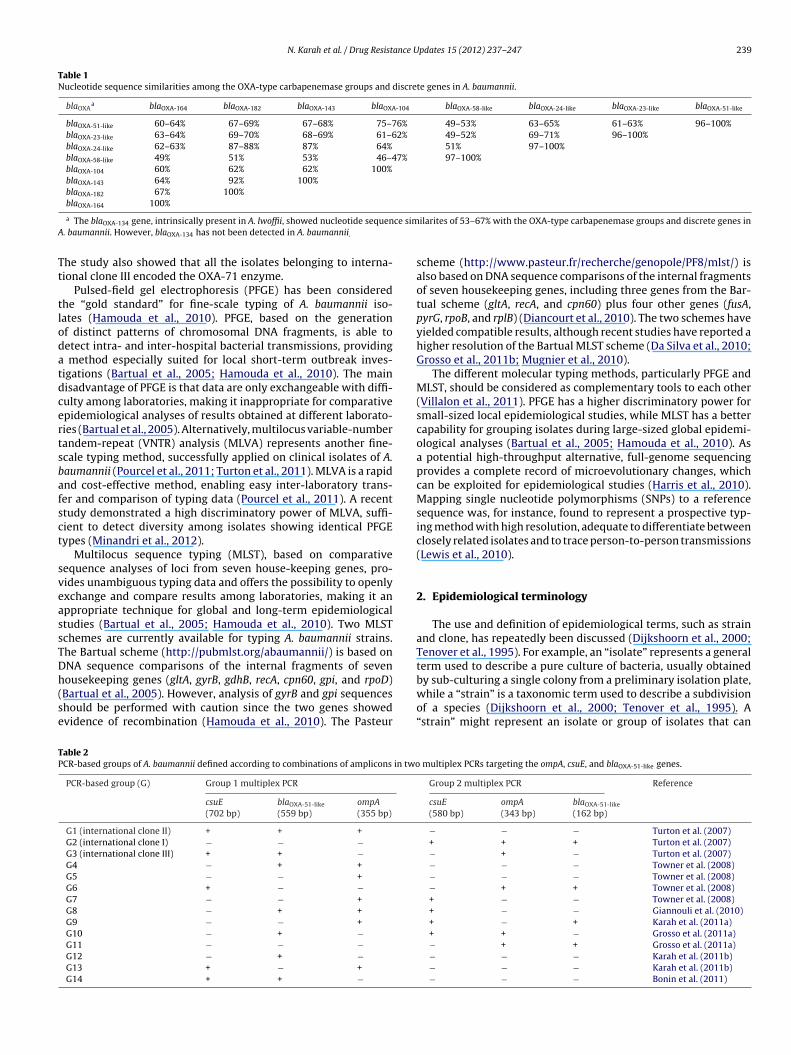
N. Karah et al. / Drug Resistance Updates 15 (2012) 237– 247 239
Table 1Nucleotide sequence similarities among the OXA-type carbapenemase groups and discrete genes in A. baumannii.
blaOXAa blaOXA-164 blaOXA-182 blaOXA-143 blaOXA-104 blaOXA-58-like blaOXA-24-like blaOXA-23-like blaOXA-51-like
blaOXA-51-like 60–64% 67–69% 67–68% 75–76% 49–53% 63–65% 61–63% 96–100%blaOXA-23-like 63–64% 69–70% 68–69% 61–62% 49–52% 69–71% 96–100%blaOXA-24-like 62–63% 87–88% 87% 64% 51% 97–100%blaOXA-58-like 49% 51% 53% 46–47% 97–100%blaOXA-104 60% 62% 62% 100%blaOXA-143 64% 92% 100%blaOXA-182 67% 100%bla 100%
ce simA
Tt
tlodatdcertsbafsct
sveassTDh(se
TP
OXA-164
a The blaOXA-134 gene, intrinsically present in A. lwoffii, showed nucleotide sequen. baumannii. However, blaOXA-134 has not been detected in A. baumannii.
he study also showed that all the isolates belonging to interna-ional clone III encoded the OXA-71 enzyme.
Pulsed-field gel electrophoresis (PFGE) has been consideredhe “gold standard” for fine-scale typing of A. baumannii iso-ates (Hamouda et al., 2010). PFGE, based on the generationf distinct patterns of chromosomal DNA fragments, is able toetect intra- and inter-hospital bacterial transmissions, providing
method especially suited for local short-term outbreak inves-igations (Bartual et al., 2005; Hamouda et al., 2010). The mainisadvantage of PFGE is that data are only exchangeable with diffi-ulty among laboratories, making it inappropriate for comparativepidemiological analyses of results obtained at different laborato-ies (Bartual et al., 2005). Alternatively, multilocus variable-numberandem-repeat (VNTR) analysis (MLVA) represents another fine-cale typing method, successfully applied on clinical isolates of A.aumannii (Pourcel et al., 2011; Turton et al., 2011). MLVA is a rapidnd cost-effective method, enabling easy inter-laboratory trans-er and comparison of typing data (Pourcel et al., 2011). A recenttudy demonstrated a high discriminatory power of MLVA, suffi-ient to detect diversity among isolates showing identical PFGEypes (Minandri et al., 2012).
Multilocus sequence typing (MLST), based on comparativeequence analyses of loci from seven house-keeping genes, pro-ides unambiguous typing data and offers the possibility to openlyxchange and compare results among laboratories, making it anppropriate technique for global and long-term epidemiologicaltudies (Bartual et al., 2005; Hamouda et al., 2010). Two MLSTchemes are currently available for typing A. baumannii strains.he Bartual scheme (http://pubmlst.org/abaumannii/) is based onNA sequence comparisons of the internal fragments of seven
ousekeeping genes (gltA, gyrB, gdhB, recA, cpn60, gpi, and rpoD)Bartual et al., 2005). However, analysis of gyrB and gpi sequenceshould be performed with caution since the two genes showedvidence of recombination (Hamouda et al., 2010). The Pasteurable 2CR-based groups of A. baumannii defined according to combinations of amplicons in two
PCR-based group (G) Group 1 multiplex PCR
csuE(702 bp)
blaOXA-51-like
(559 bp)ompA(355 bp)
G1 (international clone II) + + +
G2 (international clone I) − − −
G3 (international clone III) + + −
G4 − + +
G5 − − +
G6 + − −
G7 − − +
G8 − + +
G9 − − +
G10 − + −
G11 − − −
G12 − + −
G13 + − +
G14 + + −
ilarites of 53–67% with the OXA-type carbapenemase groups and discrete genes in
scheme (http://www.pasteur.fr/recherche/genopole/PF8/mlst/) isalso based on DNA sequence comparisons of the internal fragmentsof seven housekeeping genes, including three genes from the Bar-tual scheme (gltA, recA, and cpn60) plus four other genes (fusA,pyrG, rpoB, and rplB) (Diancourt et al., 2010). The two schemes haveyielded compatible results, although recent studies have reported ahigher resolution of the Bartual MLST scheme (Da Silva et al., 2010;Grosso et al., 2011b; Mugnier et al., 2010).
The different molecular typing methods, particularly PFGE andMLST, should be considered as complementary tools to each other(Villalon et al., 2011). PFGE has a higher discriminatory power forsmall-sized local epidemiological studies, while MLST has a bettercapability for grouping isolates during large-sized global epidemi-ological analyses (Bartual et al., 2005; Hamouda et al., 2010). Asa potential high-throughput alternative, full-genome sequencingprovides a complete record of microevolutionary changes, whichcan be exploited for epidemiological studies (Harris et al., 2010).Mapping single nucleotide polymorphisms (SNPs) to a referencesequence was, for instance, found to represent a prospective typ-ing method with high resolution, adequate to differentiate betweenclosely related isolates and to trace person-to-person transmissions(Lewis et al., 2010).
2. Epidemiological terminology
The use and definition of epidemiological terms, such as strainand clone, has repeatedly been discussed (Dijkshoorn et al., 2000;Tenover et al., 1995). For example, an “isolate” represents a generalterm used to describe a pure culture of bacteria, usually obtained
by sub-culturing a single colony from a preliminary isolation plate,while a “strain” is a taxonomic term used to describe a subdivisionof a species (Dijkshoorn et al., 2000; Tenover et al., 1995). A“strain” might represent an isolate or group of isolates that canmultiplex PCRs targeting the ompA, csuE, and blaOXA-51-like genes.
Group 2 multiplex PCR Reference
csuE(580 bp)
ompA(343 bp)
blaOXA-51-like
(162 bp)
− − − Turton et al. (2007)+ + + Turton et al. (2007)− + − Turton et al. (2007)− − − Towner et al. (2008)− − − Towner et al. (2008)− + + Towner et al. (2008)+ − − Towner et al. (2008)+ − − Giannouli et al. (2010)+ − + Karah et al. (2011a)+ + − Grosso et al. (2011a)− + + Grosso et al. (2011a)− − − Karah et al. (2011b)− − − Karah et al. (2011b)− − − Bonin et al. (2011)

2 ance U
bneit1ooic
goaeebbl“cAicmdmho
3
stpABee2eKeeHe22(ocwawaTvt(ssAi
40 N. Karah et al. / Drug Resist
e distinguished from other isolates of the same species by aumber of phenotypic and/or genotypic characteristics (Tenovert al., 1995). An outbreak of infections is an increased incidence ofnfections in a specific location during a given period that is abovehe baseline rate for that location and time frame (Tenover et al.,995). An outbreak strain (a strain responsible for the occurrencef an outbreak of infections) will accordingly represent a groupf phenotypically, genotypically, and epidemiologically relatedsolates which are assumed to be a progeny of a recent singleommon precursor isolate (Tenover et al., 1995).
A “clonal complex” (CC) is an MLST-based term representing aroup of related sequence types (STs) sharing alleles at 5/7 or 6/7f the loci (http://eburst.mlst.net). On the other hand, a “clone” is
general term that should probably be used to describe a group ofpidemiologically unrelated (independently obtained from differ-nt sources, in different locations, and perhaps at different times)ut genotypically and phenotypically related isolates which areelieved to be a progeny of an old single common precursor iso-
ate (Orskov and Orskov, 1983). The terms “clonal complex” andclone” are not interchangeable; a clonal complex represents alone only when it includes epidemiologically unrelated isolates.
clone will subjectively be defined in this review as a group ofsolates belonging to one ST/CC but obtained from more than oneountry. Occurrence in at least two countries is chosen to mini-ize the possibility of a recent inter-hospital spread, either by a
irect patient transfer or by an intermediate circulation in the com-unity (Dijkshoorn et al., 1996). This subjective definition might,
owever, be biased since it does not take into consideration the sizef individual countries and the occurrence of import.
. MLST-based global population structure
A subset of 496 A. baumannii isolates, identified by previoustudies and/or retrieved from the MLST databases, were selectedo perform an analysis of the current structure of the globalopulation of A. baumannii (Acosta et al., 2011; Adams et al., 2008;dams-Haduch et al., 2011; Ansaldi et al., 2011; Bartual et al., 2005;onin et al., 2011; Carretto et al., 2011; Da Silva et al., 2010; Deplanot al., 2011; Diancourt et al., 2010; Di Nocera et al., 2011; Di Popolot al., 2011; Fu et al., 2010; Giannouli et al., 2010; Gogou et al.,011; Grosso et al., 2011a,b; Hamidian and Hall, 2011; Hamoudat al., 2010; He et al., 2011; Ho et al., 2010; Kaase et al., 2011;arah et al., 2011a,b; Ko et al., 2010; Krizova and Nemec, 2010; Leet al., 2011; Mammina et al., 2011; Martins et al., 2012; Mezzatestat al., 2012; Mugnier et al., 2010; Nigro et al., 2011; Nigro andall, 2012; Park et al., 2009b, 2010; Post and Hall, 2009; Runnegart al., 2010; Sahl et al., 2011; Tian et al., 2010; Villalon et al.,011; Wisplinghoff et al., 2008; Yamamoto et al., 2011; Zhou et al.,011) (http://www.pasteur.fr/recherche/genopole/PF8/mlst/)http://pubmlst.org/abaumannii/). The proposed analysis includednly one isolate per country per ST (geographically and genotypi-ally divergent isolates); additional isolates from the same countryith the same ST were considered as replicates of an outbreak or
n endemic strain and were excluded regardless of whether theyere divergent by date of isolation or different by their phenotypic
nd genotypic resistance characteristics. The Minimum Spanningree (MST) algorithm implemented in the BioNumerics software.6.01 (Applied Maths, Sint-Martens-Latem, Belgium) was usedo group related STs into given CCs under stringent parameters6/7 shared allels). In order to differentiate between the two MLST
chemes, STs and CCs were designated as STB/CCB for the Bartualcheme and STP/CCP for the Pasteur scheme (Karah et al., 2011a).n ST or CC was subjectively considered to represent a clone onlyf it included isolates from more than one country.
pdates 15 (2012) 237– 247
Based on these criteria, the analysis included 308 isolatestyped using the Bartual scheme, 150 isolates typed usingthe Pasteur scheme, and 38 isolates typed using both of theschemes (Tables S1–S3, respectively). The isolates belongedto 272 Bartual STs (ST1-88B, ST90-187B, ST189-197B, ST200B,ST202-259B, ST265B, and ST345-361B) and 97 Pasteur STs(ST1-32P, ST34-59P, ST77-86P, ST94-106P, ST109-111P, ST117P,ST118P, ST120P, ST123P, ST124P, ST126P, ST127P, and ST129-134P) (http://www.pasteur.fr/recherche/genopole/PF8/mlst/)(http://pubmlst.org/abaumannii/). Overall, the analysis identified26 A. baumannii clones (i.e., STs and CCs geographically distributedin more than one country), accounting for 283/496 of the isolates.Eighteen clones were distributed in more than one continentand were described as “international clones”, while eight clonesappeared to be, so far, restricted to Europe (n = 6) and Asia (n = 2)(Table 3) (Figs. 1 and 2). On the other hand, 213/496 of the isolatesbelonged to non-clonal STs and CCs that were geographicallyrestricted to only one country.
3.1. International clone CC92B/CC2P
CC92B/CC2P was by far the largest and most widely distributedA. baumannii clone, accounting for 104/496 of the isolates andshowing a very broad international distribution among 34 coun-tries (Italy, Spain, Germany, UK, Greece, Netherlands, Denmark,Czech Republic, France, Poland, Turkey, Norway, Sweden, Portugal,Ireland, Belgium, Romania, Japan, China, Korea, Thailand, India,Malaysia, Philippines, Taiwan, Singapore, Vietnam, Lebanon, Brazil,South Africa, Reunion, Tahiti, New Caledonia, USA, and Australia)(Figs. 1 and 2). CC92B/CC2P extensively included outbreak and mul-tidrug resistant (MDR) isolates. Various carbapenemases (blaVIM-1,blaOXA-23-like, blaOXA-24-like, and/or blaOXA-58-like) were detectedamong the CC92B/CC2P isolates. In addition, the clone includedseveral isolates producing the 16S rRNA methylase ArmA enzyme,as well as one isolate resistant to colistin (Park et al., 2010). Anequine isolate was also found to belong to this clone (Diancourtet al., 2010). The oldest two isolates from this clone were col-lected in 1982 from Netherlands (RUH 134, representing a MDRoutbreak strain) and 1984 from Denmark (RUH 3422, an antibiotic-susceptible sporadic isolate) (Diancourt et al., 2010). Worringly,the occurrence of ST92B isolates resistant to all available antimi-crobial agents, including colistin, polymyxin B, and tigecycline, hasrecently been reported in Korea (Park et al., 2009a). In addition, thisclone included isolates carrying the AbaR2, AbaR4-like, AbaR22,and AbaR22-like resistance islands (Adams-Haduch et al., 2011;Chen et al., 2011b; Hornsey et al., 2011; Iacono et al., 2008; Nigroand Hall, 2012; Park et al., 2011; Zhou et al., 2011).
The intra-clonal diversity of phenotypic and genotypic resis-tance characteristics among isolates of CC92B/CC2P could be causedby the scattered occurrence of this clone throughout the world,resulting in access to a wide range of varied pools of transmissi-ble resistance elements (Nemec et al., 2004). The current analysisconfirmed that CC92B/CC2P was closely linked with internationalclone II and the OXA-66 cluster (the blaOXA-66, blaOXA-82, blaOXA-83,and blaOXA-109 variants) of the intrinsic blaOXA-51-like gene (Evanset al., 2008; Hamouda et al., 2010). The isolate from ST47P (single-ton) also belonged to international clone II. However, ST47P wasfound to be a DLV of CC2P (Diancourt et al., 2010).
3.2. International clone CC109B/CC1P
CC109B/CC1P was the second major A. baumannii clone, account-
ing for 59/496 of the isolates, with a broad international distributionamong 31 countries (Germany, Italy, Norway, Bulgaria, UK, Poland,Slovenia, Croatia, Czech Republic, Ireland, Belgium, Switzerland,Spain, France, Greece, Netherlands, Romania, Turkey, India, China,
N. Karah et al. / Drug Resistance Updates 15 (2012) 237– 247 241
Table 3The geographic distribution of MLST-designated A. baumannii clones.
A. baumannii clonea Geographic distributionb
CC92B/CC2P International clone with 104 isolates recovered from 34 countries in Europe, Asia, Africa, Australia, USA, and South AmericaCC109B/CC1P International clone with 59 isolates recovered from 31 countries in Europe, Asia, Africa, The Middle East, Australia, USA, and South AmericaCC131B South and North American clone with 20 isolates recovered from Argentina, Brazil, and USACC187B/CC3P International clone with 13 isolates recovered from 6 European countries, USA, and LebanonCC104B/CC15P International clone with 13 isolates obtained from 8 countries in Europe and South America. One isolate was exported from PakistanCC20B International clone with 11 isolates recovered from 7 countries in Europe, Asia, and AfricaST25P International clone with 7 isolates recovered from Europe, Asia, and USACC10P International clone with 6 isolates recovered from 4 European countries and AustraliaCC110B International clone with 5 isolates recovered from Korea, Argentina, and USACC32P European clone with 5 isolates recovered from Sweden, Denmark, Spain, and PortugalCC119B International clone with 4 isolates recovered from Korea, Thailand, and ArgentinaCC69B International clone with 4 isolates recovered from Korea and AustraliaCC222/228/229B International clone with 3 isolates recovered from Brazil and JapanST17B International clone with 3 isolates recovered from Germany, China, and KoreaCC105P International clone with 3 isolates recovered from Czech Republic and ChinaCC34/120/130P International clone with 3 isolates recovered from Japan, Taiwan, and SwedenCC54/99p European clone with 2 isolates from Greece and Turkey as well as the reference isolate ATCC 17904ST19B European clone with 2 isolates recovered from Germany and UKST73B International clone with 2 isolates recovered from Australia and KoreaST184B Asian clone with 2 isolates recovered from China and KoreaCC80/360B Asian clone with 2 isolates recovered from Japan and KoreaCC87/253B International clone with 2 isolates recovered from Japan and UAEST5P European clone with 2 isolates recovered from Poland and Swedenc
CC6/85P European clone with 2 isolates recovered from UK and GreeceCC83/109P European clone with 2 isolates recovered from Turkey and Swedend
ST49P International clone with 2 isolates recovered from Netherlands and USA
a CC110B and CC222/228/229B were DLVs of each other; CC69B and ST19B were DLVs of CC92B; ST49P was a DLV of ST3P.rab Em
isolatwas e
JU(rCbdbtbg6smHSg2ceflfaaaaAHe
3C
i
b USA: the United States of America; UK: the United Kingdom; UAE: the United Ac Could probably represent a Polish outbreak-related or endemic strain since thed Could probably represent an international clone since the isolate from Sweden
apan, Korea, South Africa, Singapore, Libya, Algeria, Lebanon, thenited Arab Emirates, Bahrain, Argentina, USA, and Australia)
Figs. 1 and 2). Recently, a CC1P isolate from Iraq has also beeneported (Huang et al., 2012). Similar to the CC92B/CC2P clone,C109B/CC1P included many outbreak and MDR isolates. ThelaVIM-4, blaOXA-23-like, and blaOXA-58-like carbapenemase genes wereetected. None of the CC109B/CC1P isolates were found to producelaOXA-24-like enzymes. The clone was closely linked with interna-ional clone I and the OXA-69 cluster (the blaOXA-69, blaOXA-107,laOXA-110 and blaOXA-112 variants) of the intrinsic blaOXA-51-likeene (Evans et al., 2008; Hamouda et al., 2010). One isolate (K55-1) was assigned into ST94P (a member of CC1P) by the Pasteurcheme but represented a singleton (ST194B), rather than being aember of CC109B, by the Bartual scheme (Karah et al., 2011a).owever, the analysis showed that ST194B was a DLV of severalTs from CC109B. Interestingly, this isolate contained the blaOXA-69ene, but did not belong to international clone I (Karah et al.,011a). Similarly, one isolate of ST248B (singleton) was found toarry the blaOXA-92 gene, belonging to the OXA-69 cluster (Evanst al., 2008; Hamouda et al., 2010). However, ST248B was alsoound to be a DLV of several STs from CC109B. The oldest iso-ate (HK302) known to belong to this clone was collected in 1977rom Switzerland (Krizova and Nemec, 2010). HK302 represented
MDR but imipenem-susceptible outbreak-related strain (Krizovand Nemec, 2010). Isolates carrying the AbaR1, AbaR3-AbaR21,nd AbaR3-like resistance islands belonged to this clone (GenBankccession numbers JF262178 and HM590877) (Adams et al., 2008;dams-Haduch et al., 2011; Fournier et al., 2006; Hamidian andall, 2011; Krizova et al., 2011; Krizova and Nemec, 2010; Nigrot al., 2011; Post et al., 2010; Post and Hall, 2009).
.3. International clones CC131B, CC187B/CC3P, and
C104B/CC15PCC131B has so far been discribed only in South and North Amer-ca. This clone included 20 isolates obtained from Argentina, Brazil,
erates.e from Sweden was exported from Poland.xported from Thailand.
and USA (Fig. 1). Several isolates from this clone were obtainedfrom hospital environmental sites (personnel hands, cushion, bed-rail, soap dispenser etc.) (http://pubmlst.org/abaumannii/). CC131B
included isolates producing the blaOXA-23-like and blaOXA-24-like car-bapenemases. A probable linkage could exist between CC131B andthe singleton ST79P from the Pasteur scheme (Villalon et al., 2011).Interestingly, two of the CC131B isolates belonged to internationalclone II and carried the blaOXA-66 and blaOXA-65 genes from to theOXA-66 cluster (Hamouda et al., 2010; Villalon et al., 2011). ST79P
and ST121B, a DLV of several STs from CC131B, included isolatesrecovered from Spain, indicating that the distribution of this clonemay not be restricted to South and North America (Villalon et al.,2011).
International clone CC187B/CC3P included 13 isolates obtainedfrom European countries (France, Germany, Spain, Netherlands,Italy, and Belgium), USA and Lebanon (Figs. 1 and 2). In addi-tion, a CC3P isolate from Iraq has recently been reported (Huanget al., 2012). CC187B/CC3P was closely linked with internationalclone III and the blaOXA-71 gene (Evans et al., 2008; Hamouda et al.,2010). Many of the CC187B/CC3P isolates were MDR, harbored ablaOXA-58-like gene, and were associated with outbreaks of infec-tions. A canine isolate was assigned to ST13P, which was found to bea DLV of ST3P (Diancourt et al., 2010). The limited intra-clonal diver-sity observed within CC187B/CC3P, compared with CC92B/CC2P,may indicate a relatively short-term existence (Nemec et al., 2004).
CC104B/CC15P, another international clone, included 13 isolatesobtained from 9 countries in Europe and South America (Norway,Portugal, Czech Republic, Netherlands, Turkey, Spain, Greece,Argentina, and Brazil) (Figs. 1 and 2). The Norwegian isolate wasassociated with an import event from Pakistan (Karah et al., 2011a).This clone also included outbreak, MDR, carbapenemase-encoding(blaOXA-23-like and blaOXA-58-like), and armA-encoding isolates. The
analysis detected a linkage between CC104B/CC15P and PCR-basedgroup 4 (Grosso et al., 2011a,b; Karah et al., 2011a). Isolatesfrom this clone carried the intrinsic blaOXA-51 and blaOXA-132 vari-ants. Two isolates from ST249B and ST250B, DLVs of CC104B,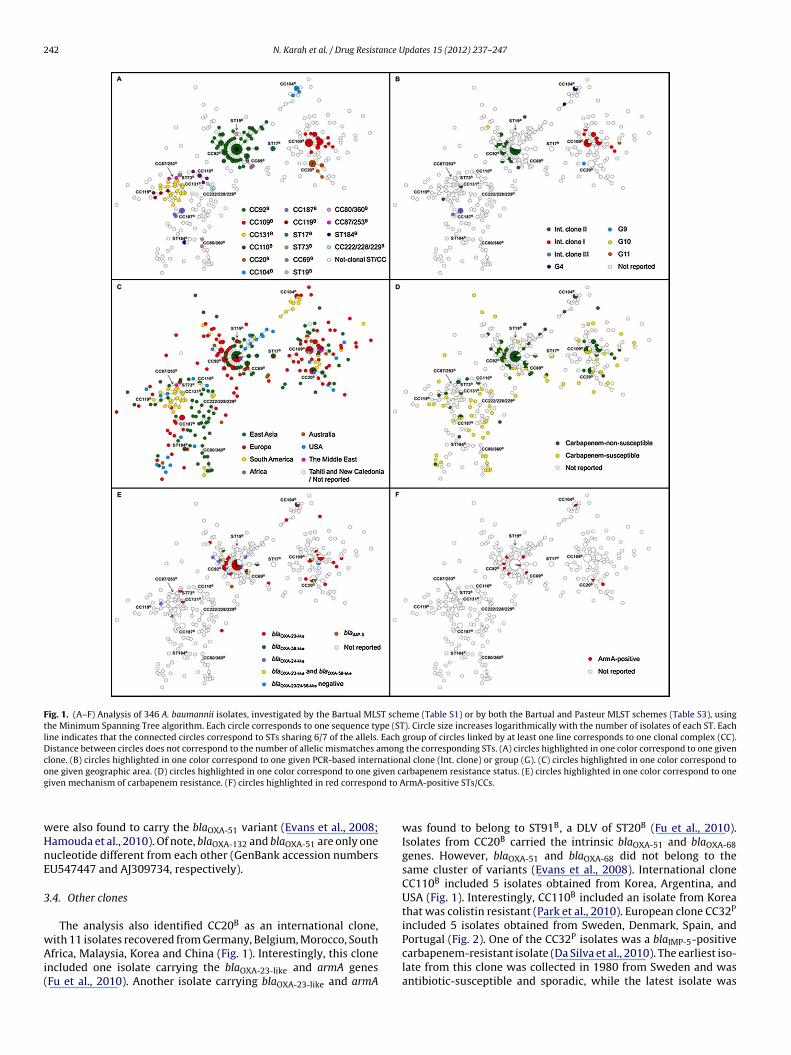
242 N. Karah et al. / Drug Resistance Updates 15 (2012) 237– 247
Fig. 1. (A–F) Analysis of 346 A. baumannii isolates, investigated by the Bartual MLST scheme (Table S1) or by both the Bartual and Pasteur MLST schemes (Table S3), usingthe Minimum Spanning Tree algorithm. Each circle corresponds to one sequence type (ST). Circle size increases logarithmically with the number of isolates of each ST. Eachline indicates that the connected circles correspond to STs sharing 6/7 of the allels. Each group of circles linked by at least one line corresponds to one clonal complex (CC).Distance between circles does not correspond to the number of allelic mismatches among the corresponding STs. (A) circles highlighted in one color correspond to one givenclone. (B) circles highlighted in one color correspond to one given PCR-based international clone (Int. clone) or group (G). (C) circles highlighted in one color correspond too ven cag d to A
wHnE
3
wAi(
ne given geographic area. (D) circles highlighted in one color correspond to one giiven mechanism of carbapenem resistance. (F) circles highlighted in red correspon
ere also found to carry the blaOXA-51 variant (Evans et al., 2008;amouda et al., 2010). Of note, blaOXA-132 and blaOXA-51 are only oneucleotide different from each other (GenBank accession numbersU547447 and AJ309734, respectively).
.4. Other clones
The analysis also identified CC20B as an international clone,
ith 11 isolates recovered from Germany, Belgium, Morocco, Southfrica, Malaysia, Korea and China (Fig. 1). Interestingly, this clonencluded one isolate carrying the blaOXA-23-like and armA genesFu et al., 2010). Another isolate carrying blaOXA-23-like and armA
rbapenem resistance status. (E) circles highlighted in one color correspond to onermA-positive STs/CCs.
was found to belong to ST91B, a DLV of ST20B (Fu et al., 2010).Isolates from CC20B carried the intrinsic blaOXA-51 and blaOXA-68genes. However, blaOXA-51 and blaOXA-68 did not belong to thesame cluster of variants (Evans et al., 2008). International cloneCC110B included 5 isolates obtained from Korea, Argentina, andUSA (Fig. 1). Interestingly, CC110B included an isolate from Koreathat was colistin resistant (Park et al., 2010). European clone CC32P
included 5 isolates obtained from Sweden, Denmark, Spain, and
Portugal (Fig. 2). One of the CC32P isolates was a blaIMP-5-positivecarbapenem-resistant isolate (Da Silva et al., 2010). The earliest iso-late from this clone was collected in 1980 from Sweden and wasantibiotic-susceptible and sporadic, while the latest isolate was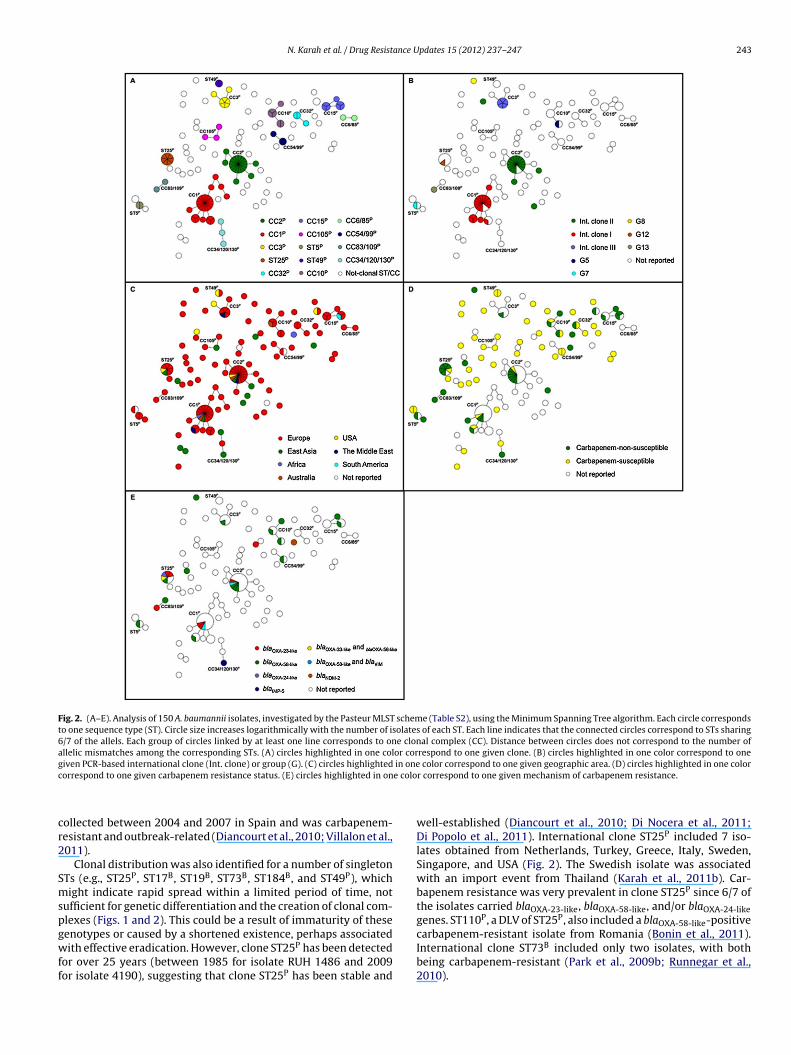
N. Karah et al. / Drug Resistance Updates 15 (2012) 237– 247 243
Fig. 2. (A–E). Analysis of 150 A. baumannii isolates, investigated by the Pasteur MLST scheme (Table S2), using the Minimum Spanning Tree algorithm. Each circle correspondsto one sequence type (ST). Circle size increases logarithmically with the number of isolates of each ST. Each line indicates that the connected circles correspond to STs sharing6/7 of the allels. Each group of circles linked by at least one line corresponds to one clonal complex (CC). Distance between circles does not correspond to the number ofallelic mismatches among the corresponding STs. (A) circles highlighted in one color correspond to one given clone. (B) circles highlighted in one color correspond to oneg in onec e colo
cr2
Smspgwff
iven PCR-based international clone (Int. clone) or group (G). (C) circles highlightedorrespond to one given carbapenem resistance status. (E) circles highlighted in on
ollected between 2004 and 2007 in Spain and was carbapenem-esistant and outbreak-related (Diancourt et al., 2010; Villalon et al.,011).
Clonal distribution was also identified for a number of singletonTs (e.g., ST25P, ST17B, ST19B, ST73B, ST184B, and ST49P), whichight indicate rapid spread within a limited period of time, not
ufficient for genetic differentiation and the creation of clonal com-lexes (Figs. 1 and 2). This could be a result of immaturity of these
enotypes or caused by a shortened existence, perhaps associatedith effective eradication. However, clone ST25P has been detectedor over 25 years (between 1985 for isolate RUH 1486 and 2009or isolate 4190), suggesting that clone ST25P has been stable and
color correspond to one given geographic area. (D) circles highlighted in one colorr correspond to one given mechanism of carbapenem resistance.
well-established (Diancourt et al., 2010; Di Nocera et al., 2011;Di Popolo et al., 2011). International clone ST25P included 7 iso-lates obtained from Netherlands, Turkey, Greece, Italy, Sweden,Singapore, and USA (Fig. 2). The Swedish isolate was associatedwith an import event from Thailand (Karah et al., 2011b). Car-bapenem resistance was very prevalent in clone ST25P since 6/7 ofthe isolates carried blaOXA-23-like, blaOXA-58-like, and/or blaOXA-24-likegenes. ST110P, a DLV of ST25P, also included a blaOXA-58-like-positive
carbapenem-resistant isolate from Romania (Bonin et al., 2011).International clone ST73B included only two isolates, with bothbeing carbapenem-resistant (Park et al., 2009b; Runnegar et al.,2010).
244 N. Karah et al. / Drug Resistance Updates 15 (2012) 237– 247
Fig. 3. Predominance of CC92B/CC2P. Favorable genetic backgrounds and the acquisition of virulence and antimicrobial resistance genes are probably main factors for theoccurrence of specific strains with increased epidemicity. International travel and/or independent but parallel emergence of smaller clusters might explain the extensivew
3
wrc2SarfStIHiteSrSS(nuS(otefbecne5
orldwide distribution of specific clones.
.5. Non-clonal STs and CCs
A large proportion of the isolates in the present study (∼43%)ere assigned to non-clonal STs and CCs. These CCs did not rep-
esent clones since they were geographically restricted to oneountry. For instance, CC164B could basically represent a Korean008 outbreak strain although it included 5 isolates from 5 differentTs (Lee et al., 2011). Similarly, CC32/33/46/47B and CC85/86B prob-bly represented two endemic strains from Germany and Japan,espectively. As expected, many of the non-clonal STs and CCs wereound to include sporadic antibiotic-susceptible isolates. However,T96B and ST78P, for instance, represented two clinically impor-ant outbreak strains showing extensive spread in Hong Kong andtaly, respectively (Di Nocera et al., 2011; Giannouli et al., 2010;o et al., 2010). The selection of the Hong Kong ST96B and Ital-
an ST78P strains might have been favored by the acquisition ofhe blaOXA-23-like and blaOXA-58-like genes, respectively (Giannoulit al., 2010; Ho et al., 2010). Other non-clonal STs (such as ST115B,T125B, and ST134B) also included MDR and/or carbapenem-esistant isolates carrying the blaOXA-23-like genes. Furthermore,T117P and ST118P included blaCTX-M-15-positive isolates, whileT103P included a blaNDM-2-positive carbapenem-resistant isolateKaase et al., 2011). The occurrence of isolates belonging to inter-ational clone II within the non-clonal ST252B and ST59P wasnanticipated. In contrast to what has previously been reported,T59P diverged from CC2P by at least four of the seven allelesDiancourt et al., 2010). ST252B also diverged from CC92B by sixf the seven alleles. Furthermore, the blaOXA-108 gene carried byhe ST252B isolate did not belong to the OXA-66 cluster (Evanst al., 2008). This could be explained by either a distant descentrom a single ancestor, lack in the discriminatory power of the PCR-ased clonal lineage typing method, or by occasional inaccuracy inxperimental results. In addition, the non-clonal ST2B and ST134B
uriously included isolates carrying blaOXA-69, representive of inter-ational clone I (Grosso et al., 2011a; Hamouda et al., 2010; Evanst al., 2008). The isolates SDF, LUH 5691, RUH 1317, LUH 5684, LUH685 and LUH 5687, recovered from a human body louse, mink, cat,
horse, and dogs, respectively, were all assigned to different non-clonal STs (Diancourt et al., 2010). Nonetheless, ST13P, ST42P, andST43P, containing isolates recovered from animals, were DLVs ofother STs (ST3P and ST5P, ST106P, and ST99P and ST21P, respec-tively) containing isolates recovered from humans. The referencestrains DSM 30007 and ATCC 17978 obtained in 1911 and 1951,respectively, were also assigned to non-clonal STs (Diancourt et al.,2010).
3.6. Predominance of CC92B/CC2P
Several factors can be proposed to explain the apparent consid-erable capability of CC92B/CC2P for epidemic clonal spread (Fig. 3).The human immune system and antibiotic treatment are mainstress conditions and crucial evolutionary bottlenecks for clinicallyrelevant bacteria (Martinez and Baquero, 2002). These conditionsgenerally decrease the biodiversity and support the emergence ofclonality within a bacterial population. Accordingly, virulence andantibiotic resistance are vital adaptive mechanisms of selectionused by particular bacterial strains and clones, such as the A. bau-mannii CC92B/CC2P clone, to survive under stress conditions, sincecompetition with isolates from other strains and clones is avoided(Martinez and Baquero, 2002.
A recent study based on comparative full-genome sequenceanalysis reported that specific genomic islands (G6, G11, G37, andG32) involved in different metabolic and ABC-type multidrug trans-port processes could be specific for, but not exclusive to ST2P
isolates (Di Nocera et al., 2011). Another study demonstrated thatACICU, an epidemic CC92B/CC2P strain, had a higher capability forbiofilm formation compared with AYE, an epidemic CC109B/CC1P
strain, and compared with ATCC 17978, a non-epidemic ST77P
strain (Antunes et al., 2011). Isolates from CC92B/CC2P have repeat-edly expressed high resistance rates to all antimicrobial agents
(Adams-Haduch et al., 2011; Diancourt et al., 2010). The fre-quent acquisition of diverse antimicrobial resistance determinantsis probably a major factor in the ability of CC92B/CC2P isolatesto survive in the hospital setting, thereby providing them with
ance U
a(glveft(
cwosroctlattoeiT1tobpirsb(
4
ncCt3csfdatwMenh
A
Ntap
N. Karah et al. / Drug Resist
dditional time for dissemination and further clonal expansionPark et al., 2010; Sahl et al., 2011). It is notable that the emer-ence of drug-resistant isolates belonging to CC92B/CC2P has mostikely resulted from independent events involving acquisition ofarious resistance determinants associated with various mobilelements (Mugnier et al., 2010). Nonetheless, specific determinantsor transmission, colonization, and/or invasion, which may explainhe epidemic behavior of CC92B/CC2P, have not yet been identifiedPerez et al., 2007; Sahl et al., 2011).
The successful occurrence of CC92B/CC2P might primarily beaused by extensive clonal spread of a single recent ancestorhich, to a certain degree of accuracy, indicates the occurrence
f a sustained and prolonged international outbreak. The theory isupported by a study from Korea showing that all the carbapenem-esistant ST92B isolates obtained from five cities belonged tone outbreak pulsotype (Fu et al., 2010). Likewise, eight of ninearbapenem-resistant isolates of ST92B obtained from three con-inents showed related PFGE patterns (Mugnier et al., 2010). Theocal, national, and international circulation of isolates is prob-bly facilitated by the increase in international travel, includingransfer of patients between different hospitals and different coun-ries (Peleg et al., 2008). Alternatively, the extensive distributionf CC92B/CC2P could be related to the independent but parallelmergence of smaller clusters expanding from several progen-tors possibly sharing a common but ancient ancestor (Fig. 3).he carbapenem-resistant ST92B isolates (n = 72) obtained from6 cities in China were distributed among at least three dis-inct outbreak pulsotypes (Ho et al., 2010). Similarly, a numberf carbapenem-resistant ST92B isolates from China were found toelong to sporadic pulsotypes (Fu et al., 2010). In addition, discreteulsotypes were assigned to isolates belonging to CC92B collected
n Norway (Karah et al., 2011a). In this regard, the similarity inesistance traits and determinants among isolates of different pul-otypes (proposed to be independently emerging) could be causedy comparable selection pressures in different parts of the worldNemec et al., 2004).
. Conclusion
The global population structure of MDR and carbapenemon-susceptible A. baumannii isolates is diverse, although spe-ific clones/CCs seem to have predominated. CC92B/CC2P andC109B/CC1P, representing international clones II and I, respec-ively, have shown extensive global dissemination in more than0 countries, supplemented by a significant assemblage of antimi-robial resistance determinants. The reason(s) for the successfulpread of these clones remain to be defined, but their capabilityor acquiring resistance is most likely a selective factor. The broadistribution of CC131B, CC187B/CC3P, CC104B/CC15P, and CC20B islso considerable, suggesting that they are major components inhe A. baumannii global population. The use of strain typing systemshich are amenably interchangeable between laboratories, such asLST, is important for conducting continued global surveillance of
merging clinically significant microorganisims such as A. bauman-ii. Nonetheless, full-genome sequencing is probably an unrivaledigh-throughput approch for prospective epidemiological studies.
cknowledgments
Part of this work was supported by a grant from the Northernorway Regional Health Autority. We also acknowledge the Geno-
yping of Pathogens and Public Health (Institut Pasteur) platformnd the PubMLST database curators for coding the MLST alleles androfiles.
pdates 15 (2012) 237– 247 245
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.drup.2012.06.001.
References
Acosta, J., Merino, M., Viedma, E., Poza, M., Sanz, F., Otero, J.R., Chaves, F., Bou, G.,2011. Multidrug-resistant Acinetobacter baumannii harboring OXA-24 carbapen-emase, Spain. Emerging Infectious Diseases 17, 1064–1067.
Adams, M.D., Goglin, K., Molyneaux, N., Hujer, K.M., Lavender, H., Jamison, J.J., Mac-Donald, I.J., Martin, K.M., Russo, T., Campagnari, A.A., Hujer, A.M., Bonomo, R.A.,Gill, S.R., 2008. Comparative genome sequence analysis of multidrug-resistantAcinetobacter baumannii. Journal of Bacteriology 190, 8053–8064.
Adams-Haduch, J.M., Paterson, D.L., Sidjabat, H.E., Pasculle, A.W., Potoski, B.A., Muto,C.A., Harrison, L.H., Doi, Y., 2008. Genetic basis of multidrug resistance in Acine-tobacter baumannii clinical isolates at a tertiary medical center in Pennsylvania.Antimicrobial Agents and Chemotherapy 52, 3837–3843.
Adams-Haduch, J.M., Onuoha, E.O., Bogdanovich, T., Tian, G.B., Marschall, J., Urban,C.M., Spellberg, B.J., Rhee, D., Halstead, D.C., Pasculle, A.W., Doi, Y., 2011. Molec-ular epidemiology of carbapenem-non-susceptible Acinetobacter baumannii inthe United States. Journal of Clinical Microbiology 49, 3849–3854.
Ansaldi, F., Canepa, P., Bassetti, M., Zancolli, M., Molinari, M.P., Talamini, A., Ginoc-chio, F., Durando, P., Mussap, M., Orengo, G., Viscoli, C., Icardi, G., 2011. Sequentialoutbreaks of multidrug-resistant Acinetobacter baumannii in intensive care unitsof a tertiary referral hospital in Italy: combined molecular approach for epidemi-ological investigation. Journal of Hospital Infection 79, 134–140.
Antunes, L.C.S., Imperi, F., Carattoli, A., Visca, P., 2011. Deciphering the multifactorialnature of Acinetobacter baumannii pathogenicity. PLoS One 6, e22674.
Bartual, S.G., Seifert, H., Hippler, C., Luzon, M.A., Wisplinghoff, H., Rodriguez-Valera,F., 2005. Development of a multilocus sequence typing scheme for charac-terization of clinical isolates of Acinetobacter baumannii. Journal of ClinicalMicrobiology 43, 4382–4390.
Bonin, R.A., Poirel, L., Licker, M., Nordmann, P., 2011. Genetic diversity of carbapenemhydrolysing �-lactamases in Acinetobacter baumannii from Romanian hospitals.Clinical Microbiology and Infection 17, 1524–1528.
Boo, T.W., Walsh, F., Crowley, B., 2009. Molecular characterization of carbapenem-resistant Acinetobacter species in an Irish university hospital: predominance ofAcinetobacter genomic species 3. Journal of Medical Microbiology 58, 209–216.
Carretto, E., Barbarini, D., Dijkshoorn, L., van der Reijden, T.J.K., Brisse, S., Passet,V., Farina, C., 2011. Widespread carbapenem resistant Acinetobacter baumanniiclones in Italian hospitals revealed by a multicenter study. Infection, Geneticsand Evolution 11, 1319–1326.
Chen, Z., Qlu, S., Wang, Y., Liu, S., Wang, Z., Du, X., Wang, L., Guo, J., Liu, N., Yuan, J.,Song, H., Huang, L., 2011a. Coexistence of blaNDM-1 with the prevalent blaOXA23
and blaIMP in pan-drug resistant Acinetobacter baumannii isolates in China. Clin-ical Infectious Diseases 52, 692–693.
Chen, C.C., Lin, Y.C., Sheng, W.H., Chen, Y.C., Chang, S.C., Hsia, K.C., Liao, M.H., Li,S.Y., 2011b. Genome sequence of a dominant multidrug-resistant Acinetobacterbaumannii strain TCDC-AB0715. Journal of Bacteriology 193, 2361–2362.
Cho, Y.J., Moon, D.C., Jin, J.S., Choi, C.H., Lee, Y.C., Lee, J.C., 2009. Genetic basisof resistance to aminoglycosides in Acinetobacter spp. and spread of armAin Acinetobacter baumannii sequence group 1 in Korean hospitals. DiagnosticMicrobiology and Infectious Disease 64, 185–190.
Chu, Y.W., Afzal-Shah, M., Houang, E.T., Palepou, M.I., Lyon, D.J., Woodford, N.,Livermore, D.M., 2001. IMP-4, a novel metallo-ß-lactamase from nosocomialAcinetobacter spp. collected in Hong Kong between 1994 and 1998. AntimicrobialAgents and Chemotherapy 45, 710–714.
Corvec, S., Poirel, L., Naas, T., Drugeon, H., Nordmann, P., 2007. Genetics and expres-sion of the carbapenem-hydrolyzing oxacillinase gene blaOXA-23 in Acinetobacterbaumannii. Antimicrobial Agents and Chemotherapy 51, 1530–1533.
Da Silva, G.J., Mendonca, N., Batista, G., Duarte, A., 2010. Sequence types of Por-tuguese carbapenem-resistant Acinetobacter baumannii isolates collected over10 years. Journal of Antimicrobial Chemotherapy 65, 2254–2256.
Deplano, A., Denis, O., Rodriguez-Villalobos, H., De Ryck, R., Struelens, M.J., Hallin, M.,2011. Controlled performance evaluation of the DiversiLab repetitive-sequence-based genotyping system for typing multidrug-resistant health care-associatedbacterial pathogens. Journal of Clinical Microbiology 49, 3616–3620.
Diancourt, L., Passet, V., Nemec, A., Dijkshoorn, L., Brisse, S., 2010. The populationstructure of Acinetobacter baumannii: expanding multiresistant clones from anancestral susceptible genetic pool. PLoS One 5, e10034.
Dijkshoorn, L., Aucken, H., Gerner-Smidt, P., Janssen, P., Kaufmann, M.E., Garaizar, J.,Ursing, J., Pitt, T.L., 1996. Comparison of outbreak and nonoutbreak Acinetobac-ter baumannii strains by genotypic and phenotypic methods. Journal of ClinicalMicrobiology 34, 1519–1525.
Dijkshoorn, L., Ursing, B.M., Ursing, J.B., 2000. Strain, clone and species: comments
on three basic concepts of bacteriology. Journal of Medical Microbiology 49,397–401.Dijkshoorn, L., Nemec, A., Seifert, H., 2007. An increasing threat in hospitals:multidrug-resistant Acinetobacter baumannii. Nature Reviews Microbiology 5,939–951.

2 ance U
D
D
D
D
E
F
F
G
G
G
G
G
H
H
H
H
H
H
H
H
I
K
K
46 N. Karah et al. / Drug Resist
i Nocera, P.P., Rocco, F., Giannouli, M., Triassi, M., Zarrilli, R., 2011. Genome orga-nization of epidemic Acinetobacter baumannii strains. BMC Microbiology 11,224.
i Popolo, A., Giannouli, M., Triassi, M., Brisse, S., Zarrilli, R., 2011. Molecular epidemi-ological investigation of multidrug-resistant Acinetobacter baumannii strains infour Mediterranean countries with a multilocus sequence typing scheme. Clin-ical Microbiology and Infection 17, 197–201.
oi, Y., Adams, J.M., Yamane, K., Paterson, D.L., 2007. Identification of 16S rRNAmethylase-producing Acinetobacter baumannii clinical strains in North America.Antimicrobial Agents and Chemotherapy 51, 4209–4210.
oi, Y., Arakawa, Y., 2007. 16S ribosomal RNA methylation: emerging resistancemechanism against aminoglycosides. Clinical Infectious Diseases 45, 88–94.
vans, B.A., Hamouda, A., Towner, K.J., Amyes, S.G., 2008. OXA-51-like �-lactamasesand their association with particular epidemic lineages of Acinetobacter bau-mannii. Clinical Microbiology and Infection 14, 268–275.
ournier, P.E., Vallenet, D., Barbe, V., Audic, S., Ogata, H., Poirel, L., Richet, H., Robert,C., Mangenot, S., Abergel, C., Nordmann, P., Weissenbach, J., Raoult, D., Claverie,J.M., 2006. Comparative genomics of multidrug resistance in Acinetobacter bau-mannii. PLoS Genetics 2, e7.
u, Y., Zhou, J., Zhou, H., Yang, Q., Wei, Z., Yu, Y., Li, L., 2010. Wide dissemination ofOXA-23-producing carbapenem-resistant Acinetobacter baumannii clonal com-plex 22 in multiple cities of China. Journal of Antimicrobial Chemotherapy 65,644–650.
iannouli, M., Tomasone, F., Agodi, A., Vahaboglu, H., Daoud, Z., Triassi, M., Tsakris, A.,Zarrilli, R., 2009. Molecular epidemiology of carbapenem-resistant Acinetobacterbaumannii strains in intensive care units of multiple Mediterranean hospitals.Journal of Antimicrobial Chemotherapy 63, 828–830.
iannouli, M., Cuccurullo, S., Crivaro, V., Di Popolo, A., Bernardo, M., Tomasone, F.,Amato, G., Brisse, S., Triassi, M., Utili, R., Zarrilli, R., 2010. Molecular epidemiol-ogy of multidrug-resistant Acinetobacter baumannii in a tertiary care hospital inNaples, Italy, shows the emergence of a novel epidemic clone. Journal of ClinicalMicrobiology 48, 1223–1230.
ogou, V., Pournaras, S., Giannouli, M., Voulgari, E., Piperaki, E.T., Zarrilli, R.,Tsakris, A., 2011. Evolution of multidrug-resistant Acinetobacter baumanniiclonal lineages: a 10 year study in Greece (2000–2009). Journal of AntimicrobialChemotherapy 66, 2767–2772.
rosso, F., Quinteira, S., Peixe, L., 2011a. Understanding the dynamics ofimipenem-resistant Acinetobacter baumannii lineages within Portugal. ClinicalMicrobiology and Infection 17, 1275–1279.
rosso, F., Carvalho, K.R., Quinteira, S., Ramos, A., Carvalho-Assef, A.P., Asensi, M.D.,Peixe, L., 2011b. OXA-23-producing Acinetobacter baumannii: a new hotspot ofdiversity in Rio de Janeiro? Journal of Antimicrobial Chemotherapy 66, 62–65.
amidian, M., Hall, R.M., 2011. AbaR4 replaces AbaR3 in a carbapenem-resistantAcinetobacter baumannii isolate belonging to global clone 1 from an Australianhospital. Journal of Antimicrobial Chemotherapy 66, 2484–2491.
amouda, A., Evans, B.A., Towner, K.J., Amyes, S.G., 2010. Characterization of epi-demiologically unrelated Acinetobacter baumannii isolates from four continentsby use of multilocus sequence typing, pulsed-field gel electrophoresis, andsequence-based typing of blaOXA-51-like genes. Journal of Clinical Microbiology48, 2476–2483.
an, H.L., Jang, S.J., Park, G., Kook, J.K., Shin, J.H., Shin, S.H., Kim, D.M., Cheon, J.S.,Moon, D.S., Park, Y.J., 2008. Identification of an atypical integron carrying anIS26-disrupted aadA1 gene cassette in Acinetobacter baumannii. InternationalJournal of Antimicrobial Agents 32, 165–169.
arris, S.R., Feil, E.J., Holden, M.T.G., Quail, M.A., Nickerson, E.K., Chantratita, N.,Gardete, S., Tavares, A., Day, N., Lindsay, J.A., Edgeworth, J.D., de Lencastre, H.,Parkhill, J., Peacock, S.J., Bentleyet, S.D., 2010. Evolution of MRSA during hospitaltransmission and intercontinental spread. Science 327, 469–474.
e, C., Xie, Y., Fan, H., Kang, M., Tao, C., Zhang, R., Hu, Y., Chen, Z., Wang, L., 2011.Spread of imipenem-resistant Acinetobacter baumannii of European clone II inWestern China. International Journal of Antimicrobial Agents 38, 257–260.
o, P.L., Ho, A.Y., Chow, K.H., Lai, E.L., Ching, P., Seto, W.H., 2010. Epidemiologyand clonality of multidrug-resistant Acinetobacter baumannii from a healthcareregion in Hong Kong. Journal of Hospital Infection 74, 358–364.
ornsey, M., Loman, N., Wareham, D.W., Ellington, M.J., Pallen, M.J., Turton, J.F.,Underwood, A., Gaulton, T., Thomas, C.P., Doumith, M., Livermore, D.M., Wood-ford, N., 2011. Whole-genome comparison of two Acinetobacter baumanniiisolates from a single patient, where resistance developed during tigecyclinetherapy. Journal of Antimicrobial Chemotherapy 66, 1499–1503.
uang, X.-Z., Chahine, M.A., Frye, J.G., Cash, D.M., Lesho, E.P., Craft, L.E., Lindler,D.W., Nikolich, M.P., 2012. Molecular analysis of imipenem-resistant Acinetobac-ter baumannii isolated from US service members wounded in Iraq, 2003–2008.Epidemiology and Infection 25, 1–6.
acono, M., Villa, L., Fortini, D., Bordoni, R., Imperi, F., Bonnal, R.J., Sicheritz-Ponten,T., De Bellis, G., Visca, P., Cassone, A., Carattoli, A., 2008. Whole-genome pyrose-quencing of an epidemic multidrug-resistant Acinetobacter baumannii strainbelonging to the European clone II group. Antimicrobial Agents and Chemother-apy 52, 2616–2625.
aase, M., Nordmann, P., Wichelhaus, T.A., Gatermann, S.G., Bonnin, R.A., Poirel, L.,2011. NDM-2 carbapenemase in Acinetobacter baumannii from Egypt. Journal ofAntimicrobial Chemotherapy 66, 1260–1262.
arah, N., Haldorsen, B., Hermansen, N.O., Tveten, Y., Ragnhildstveit, E., Skut-laberg, D.H., Tofteland, S., Sundsfjord, A., Samuelsen, O., 2011a. Emergence ofOXA-carbapenemase- and 16S rRNA methylase-producing international clonesof Acinetobacter baumannii in Norway. Journal of Medical Microbiology 60,515–521.
pdates 15 (2012) 237– 247
Karah, N., Giske, C.G., Sundsfjord, A., Samuelsen, O., 2011b. A diversity ofOXA-carbapenemases and class 1 integrons among carbapenem-resistantAcinetobacter baumannii clinical isolates from Sweden belonging to differentinternational clonal lineages. Microbial Drug Resistance 17, 545–549.
Karthikeyan, K., Thirunarayan, M.A., Krishnan, P., 2010. Coexistence of blaOXA-23 withblaNDM-1 and armA in clinical isolates of Acinetobacter baumannii from India.Journal of Antimicrobial Chemotherapy 65, 2253–2254.
Ko, K.S., Kim, H.W., Choi, J.Y., Song, J.H., 2010. Dissemination of limited imipenem-resistant Acinetobacter baumannii clones in Asian countries. 20th EuropeanCongress of Clinical Microbiology and Infectious Diseases, P1722.
Krizova, L., Nemec, A., 2010. A 63 kb genomic resistance island found in a multidrug-resistant Acinetobacter baumannii isolate of European clone I from 1977. Journalof Antimicrobial Chemotherapy 65, 1915–1918.
Krizova, L., Dijkshoorn, L., Nemec, A., 2011. Diversity and evolution of AbaR genomicresistance islands in Acinetobacter baumannii strains of European clone I. Antimi-crobial Agents and Chemotherapy 55, 3201–3206.
Lee, K., Yum, J.H., Yong, D., Lee, H.M., Kim, H.D., Docquier, J.D., Rossolini, G.M., Chong,Y., 2005. Novel acquired metallo-ß-lactamase gene, blaSIM-1, in a class 1 integronfrom Acinetobacter baumannii clinical isolates from Korea. Antimicrobial Agentsand Chemotherapy 49, 4485–4491.
Lee, Y., Lee, J., Jeong, S.H., Bae, I.K., Lee, K., 2011. Carbapenem-non-susceptible Acine-tobacter baumannii of sequence type 92 or its single-locus variants with a G428Tsubstitution in zone 2 of the rpoB gene. Journal of Antimicrobial Chemotherapy66, 66–72.
Lim, Y.M., Shin, K.S., Kim, J., 2007. Distinct antimicrobial resistance patterns andantimicrobial resistance-harboring genes according to genomic species of Acine-tobacter isolates. Journal of Clinical Microbiology 45, 902–905.
Lin, Y.C., Sheng, W.H., Chen, Y.C., Chang, S.C., Hsia, K.C., Li, S.Y., 2010. Differencesin carbapenem resistance genes among Acinetobacter baumannii, Acinetobac-ter genospecies 3 and Acinetobacter genospecies 13TU in Taiwan. InternationalJournal of Antimicrobial Agents 35, 439–443.
Lewis, T., Loman, N.J., Bingle, L., Jumaa, P., Weinstock, G.M., Mortiboy, D., Pallen, M.J.,2010. High-throughput whole-genome sequencing to dissect the epidemiologyof Acinetobacter baumannii isolates from a hospital outbreak. Journal of HospitalInfection 75, 37–41.
Mammina, C., Bonura, C., Aleo, A., Calà, C., Caputo, G., Cataldo, M.C., Benedetto, A.D.,Distefano, S., Fasciana, T., Labisi, M., Sodano, C., Palma, D.M., Giammanco, A.,2011. Characterization of Acinetobacter baumannii from intensive care units andhome care patients in Palermo, Italy. Clinical Microbiology and Infection 17,E12–E15.
Martinez, J.L., Baquero, F., 2002. Interactions among strategies associated with bac-terial infection: pathogenicity, epidemicity, and antibiotic resistance. ClinicalMicrobiology Reviews 15, 647–679.
Martins, N., Martins, I.S., de Freitas, W.V., de Matos, J.A., Magalhaes, A.C.G.,Girao, V.B.C., Dias, R.C.S., de Souza, T.C., Pellegrino, F.L., Costa, L.D., Boasque-visque, C.H.R., Nouer, S.A., Riley, L.W., Santoro-Lopes, G., Moreira, B.M., 2012.Severe infection in a lung transplant recipient caused by donor-transmittedcarbapenem-resistant Acinetobacter baumannii. Transplant Infectious Disease14, 316–320.
Mezzatesta, M.L., D’Andrea, M.M., Migliavacca, R., Giani, T., Gona, F., Nucleo, E.,Fugazza, G., Pagani, L., Rossolini, G.M., Stefani, S., 2012. Epidemiological char-acterization and distribution of carbapenem-resistant Acinetobacter baumanniiclinical isolates in Italy. Clinical Microbiology and Infection 18, 160–166.
Minandri, F., D’Arezzo, S., Antunes, L.C.S., Pourcel, C., Principe, L., Petrosillo, N., Visca,P., 2012. Evidence of diversity among epidemiologically related carbapenemase-producing Acinetobacter baumannii strains belonging to international clonallineage II. Journal of Clinical Microbiology 50, 590–597.
Moubareck, C., Bremont, S., Conroy, M.C., Courvalin, P., Lambert, T., 2009. GES-11, anovel integron-associated GES variant in Acinetobacter baumannii. AntimicrobialAgents and Chemotherapy 53, 3579–3581.
Mugnier, P.D., Poirel, L., Naas, T., Nordmann, P., 2010. Worldwide dissemination ofthe blaOXA-23 carbapenemase gene of Acinetobacter baumannii. Emerging Infec-tious Diseases 16, 35–40.
Nemec, A., Dolzani, L., Brisse, S., van den Broek, P., Dijkshoorn, L., 2004. Diversityof aminoglycoside-resistance genes and their association with class 1 integronsamong strains of pan-European Acinetobacter baumannii clones. Journal of Med-ical Microbiology 53, 1233–1240.
Nigro, S.J., Post, V., Hall, R.M., 2011. The multiresistant Acinetobacter baumannii Euro-pean clone I type strain RUH875 (A297) carries a genomic antibiotic resistanceisland AbaR21, plasmid pRAY and a cluster containing ISAba1-sul2-CR2-strB-strA. Journal of Antimicrobial Chemotherapy 66, 1928–1930.
Nigro, S.J., Hall, R.M., 2012. Antibiotic resistance islands in A320 (RUH134), the refer-ence strain for Acinetobacter baumannii global clone 2. Journal of AntimicrobialChemotherapy 67, 335–338.
Orskov, F., Orskov, I., 1983. From the national institutes of health. Summary of aworkshop on the clone concept in the epidemiology, taxonomy, and evolutionof the Enterobacteriaceae and other bacteria. Journal of Infectious Diseases 148,346–357.
Park, Y.K., Peck, K.R., Cheong, H.S., Chung, D.R., Song, J.H., Ko, K.S., 2009a. Extremedrug resistance in Acinetobacter baumannii infections in intensive care units,South Korea. Emerging Infectious Diseases 15, 1325–1327.
Park, Y.K., Choi, J.Y., Jung, S.I., Park, K.H., Lee, H., Jung, D.S., Heo, S.T., Kim, S.W.,Chang, H.H., Cheong, H.S., Chung, D.R., Peck, K.R., Song, J.H., Ko, K.S., 2009b.Two distinct clones of carbapenem-resistant Acinetobacter baumannii isolatesfrom Korean hospitals. Diagnostic Microbiology and Infectious Disease 64,389–395.

ance U
P
P
P
P
P
P
P
P
P
P
R
R
R
S
N. Karah et al. / Drug Resist
ark, Y.K., Lee, G.H., Baek, J.Y., Chung, D.R., Peck, K.R., Song, J.H., Ko, K.S., 2010. A singleclone of Acinetobacter baumannii, ST22, is responsible for high antimicrobialresistance rates of Acinetobacter spp. isolates that cause bacteremia and urinarytract infections in Korea. Microbial Drug Resistance 16, 143–149.
ark, J.Y., Kim, S., Kim, S.M., Cha, S.H., Lim, S.K., Kim, J., 2011. Complete genomesequence of multidrug-resistant Acinetobacter baumannii strain 1656-2, whichforms sturdy biofilm. Journal of Bacteriology 193, 6393–6394.
eleg, A.Y., Seifert, H., Paterson, D.L., 2008. Acinetobacter baumannii: emergence of asuccessful pathogen. Clinical Microbiology Reviews 21, 538–582.
erez, F., Hujer, A.M., Hujer, K.M., Decker, B.K., Rather, P.N., Bonomo, R.A., 2007.Global challenge of multidrug-resistant Acinetobacter baumannii. AntimicrobialAgents and Chemotherapy 51, 3471–3484.
oirel, L., Nordmann, P., 2006a. Carbapenem resistance in Acinetobacter bauman-nii: mechanisms and epidemiology. Clinical Microbiology and Infection 12,826–836.
oirel, L., Nordmann, P., 2006b. Genetic structures at the origin of acquisitionand expression of the carbapenem-hydrolyzing oxacillinase gene blaOXA-58 in Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy 50,1442–1448.
oirel, L., Naas, T., Nordmann, P., 2010. Diversity, epidemiology, and genetics of classd ß-lactamases. Antimicrobial Agents and Chemotherapy 54, 24–38.
ost, V., Hall, R.M., 2009. AbaR5, a large multiple-antibiotic resistance regionfound in Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy 53,2667–2671.
ost, V., White, P.A., Hall, R.M., 2010. Evolution of AbaR-type genomic resis-tance islands in multiply antibiotic-resistant Acinetobacter baumannii. Journalof Antimicrobial Chemotherapy 65, 1162–1170.
ourcel, C., Minandri, F., Hauck, Y., D’Arezzo, S., Imperi, F., Vergnaud, G., Visca,P., 2011. Identification of variable-number tandem-repeat (VNTR) sequencesin Acinetobacter baumannii and interlaboratory validation of an optimizedmultiple-locus VNTR analysis typing scheme. Journal of Clinical Microbiology49, 539–548.
avasi, P., Limansky, A.S., Rodriguez, R.E., Viale, A.M., Mussi, M.A., 2011. ISAba825,a functional insertion sequence modulating genomic plasticity and blaOXA-58
expression in Acinetobacter baumannii. Antimicrobial Agents and Chemotherapy55, 917–920.
obledo, I.E., Aquino, E.E., Sante, M.I., Santana, J.L., Otero, D.M., Leon, C.F., Vazquez,G.J., 2010. Detection of KPC in Acinetobacter spp. in Puerto Rico. AntimicrobialAgents and Chemotherapy 54, 1354–1357.
unnegar, N., Sidjabat, H., Goh, H.M., Nimmo, G.R., Schembri, M.A., Paterson, D.L.,
2010. Molecular epidemiology of multidrug-resistant Acinetobacter baumanniiin a single institution over a 10-year period. Journal of Clinical Microbiology 48,4051–4056.ahl, J.W., Johnson, J.K., Harris, A.D., Phillippy, A.M., Hsiao, W.W., Thom, K.A.,Rasko, D.A., 2011. Genomic comparison of multi-drug resistant invasive and
pdates 15 (2012) 237– 247 247
colonizing Acinetobacter baumannii isolated from diverse human body sitesreveals genomic plasticity. BMC Genomics 12, 291.
Smith, M.G., Gianoulis, T.A., Pukatzki, S., Mekalanos, J.J., Ornston, L.N., Gerstein,M., Snyder, M., 2007. New insights into Acinetobacter baumannii pathogenesisrevealed by high-density pyrosequencing and transposon mutagenesis. Genesand Development 21, 601–614.
Tenover, F.C., Arbeit, R.D., Goering, R.V., Mickelsen, P.A., Murray, B.E., Persing, D.H.,Swaminathan, B., 1995. Interpreting chromosomal DNA restriction patterns pro-duced by pulsed-field gel electrophoresis: criteria for bacterial strain typing.Journal of Clinical Microbiology 33, 2233–2239.
Tian, G.B., Adams-Haduch, J.M., Bogdanovich, T., Pasculle, A.W., Quinn, J.P., Wang,H.N., Doi, Y., 2010. Identification of diverse OXA-40-group carbapenemasesincluding a novel variant, OXA-160, from Acinetobacter baumannii in Pennsyl-vania. Antimicrobial Agents and Chemotherapy 55, 429–432.
Towner, K.J., Levi, K., Vlassiadi, M., 2008. Genetic diversity of carbapenem-resistantisolates of Acinetobacter baumannii in Europe. Clinical Microbiology and Infec-tion 14, 161–167.
Turton, J.F., Gabriel, S.N., Valderrey, C., Kaufmann, M.E., Pitt, T.L., 2007. Use ofsequence-based typing and multiplex PCR to identify clonal lineages of out-break strains of Acinetobacter baumannii. Clinical Microbiology and Infection 13,807–815.
Turton, J.F., Baddal, B., Perry, C., 2011. Use of the accessory genome for characteriza-tion and typing of Acinetobacter baumannii. Journal of Clinical Microbiology 49,1260–1266.
van Dessel, H., Dijkshoorn, L., van der Reijden, T., Bakker, N., Paauw, A., van den Broek,P., Verhoef, J., Brisse, S., 2004. Identification of a new geographically widespreadmultiresistant Acinetobacter baumannii clone from European hospitals. Researchin Microbiology 155, 105–112.
Vila, J., Ruiz, J., Goni, P., Jimenez de Anta, T., 1997. Quinolone-resistance mutations inthe topoisomerase IV parC gene of Acinetobacter baumannii. Journal of Antimi-crobial Chemotherapy 39, 757–762.
Villalon, P., Valdezate, S., Medina-Pascual, M.J., Rubio, V., Vindel, A., Saez-Nieto, J.A.,2011. Clonal diversity of nosocomial epidemic Acinetobacter baumannii strainsisolated in Spain. Journal of Clinical Microbiology 49, 875–882.
Wisplinghoff, H., Hippler, C., Bartual, S.G., Haefs, C., Stefanik, D., Higgins, P.G., Seifert,H., 2008. Molecular epidemiology of clinical Acinetobacter baumannii and Acine-tobacter genomic species 13TU isolates using a multilocus sequencing typingscheme. Clinical Microbiology and Infection 14, 708–715.
Yamamoto, M., Nagao, M., Matsumura, Y., Matsushima, A., Ito, Y., Takakura, S.,Ichiyama, S., 2011. Interspecies dissemination of a novel class 1 integron car-
rying blaIMP-19 among Acinetobacter species in Japan. Journal of AntimicrobialChemotherapy 66, 2480–2483.Zhou, H., Zhang, T., Yu, D., Pi, B., Yang, Q., Zhou, J., Hu, S., Yu, Y., 2011. Genomicanalysis of the multi-drug resistant Acinetobacter baumannii strain MDR-ZJ06widely spread in China. Antimicrobial Agents and Chemotherapy 55, 4506–4512.
Related Documents











