Insecticide resistance in Culex pipiens quinquefasciatus and Aedes albopictus mosquitoes from La Réunion Island Michaël Luciano Tantely a, 1 , Pablo Tortosa b , Haoues Alout a , Claire Berticat a , Arnaud Berthomieu a , Abdoul Rutee c , Jean-Sébastien Dehecq c , Patrick Makoundou a , Pierrick Labbé a , Nicole Pasteur a , Mylène Weill a, * a Université Montpellier 2, UMR CNRS 5554, Institut des Sciences de l'Evolution, Génétique de l'Adaptation, C.C. 065, Place Eugène Bataillon, 34095 Montpellier 05, France b Centre de Recherche et de Veille sur les Maladies Emergentes dans l'Océan Indien, 2 rue Maxime Rivière, 97410 Ste Clotilde, Ile de La Réunion, France c Direction Régionale des Affaires Sanitaires et Sociales, 2 bis avenue Georges Brassens, BP50 97408 Saint-Denis, Ile de La Réunion, France article info Article history: Received 1 December 2009 Received in revised form 16 February 2010 Accepted 18 February 2010 Keywords: Insecticide resistance Culex pipiens quinquefasciatus Aedes albopictus Vector control Dieldrin resistance abstract Resistance to insecticides was monitored on Culex pipiens quinquefasciatus mosquitoes collected in twelve localities of La Réunion, a geographically isolated island of the Indian Ocean. This mosquito is of medical concern in the region as a known vector for lariasis and a potential vector for West Nile and Rift Valley Fever viruses. Our bioassays indicated the presence of resistance to all tested insecticides, i.e. organo- chlorides, organophosphates and pyrethroids. A molecular investigation revealed a higher frequency of resistance genes in the coastal areas compared to elevated rural sites, probably reecting the different nature of insecticide pressures together with the genetic cost of resistance alleles. A simple molecular test was developed to detect Rdl R allele, encoding a g-aminobutyric acid (GABA) receptor resistant to dieldrin. Unexpectedly high Rdl R frequencies were recorded over the whole island, despite this insecticide having been banned for over 15 years. This resistant allele was also detected for the rst time in two samples of Aedes albopictus, a species recently involved in severe Chikungunya epidemics on the island. Rdl R selection in these two mosquito species discloses current insecticide pressures in urban areas, from unknown origins, that should be taken into account to develop vector control strategies. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Resistance to insecticides that impairs nervous transmission has been extensively studied as it is critical for a proper management of insect control. It is also of particular interest for evolutionary biologists: it represents an almost unique example for monitoring the genetic consequences of a well characterized alteration of the environment. In mosquitoes, insecticide resistance is essentially achieved through two mechanisms: increased detoxication and target insensitivity (review in Nauen, 2007; Whalon et al., 2008). The former results from an increased activity of detoxifying enzymes: esterases, cytochrome P450 oxidases or glutathione- S-transferases. The latter is associated with target modications that lower their afnity for the considered insecticide. There are few insecticide targets: the synaptic acetylcholinesterase (AChE1) encoded by the ace-1 gene, the g-aminobutyric acid (GABA) receptor gene encoded by Rdl and the voltage-dependent sodium channel encoded by kdr . Each of these genes has a limited number of possible mutable positions. As far as mosquitoes are concerned, three substitutions conferring resistance have been recorded in the synaptic AChE1 (G119S, F290V and F331W), two mutations (A302S and A302G) in the GABA receptor, and two substitutions in the sodium channel (L1014F and L1014S) in Culex and Anopheles, while additional mutations were recorded on the same target in Aedes aegypti (ffrench-Constant et al., 2000; Brengues et al., 2003; Weill et al., 2003; Du et al., 2005; Alout et al., 2007a, 2007b; Pinto et al., 2007; Saavedra-Rodriguez et al., 2007). Among all the species controlled with insecticides, Culex pipiens is of particular interest because it can be considered as an indicator to monitor the nature and persistence of insecticides in the envi- ronment: most of its natural breeding sites are indeed localized in various types of drainages that collect lthy waters over large areas. A full set of biochemical and molecular tools has been developed to identify resistance genes in this species. Moreover, its resistance alleles evolve rapidly and can be easily tracked. Besides, over the last 30e40 years, substantial physiological and molecular data have been documented on the evolutionary constraints of insecticide * Corresponding author. Fax: þ33 4 67 14 36 22. E-mail address: [email protected] (M. Weill). 1 Present address: Laboratoire d'Entomologie Médicale, Institut Pasteur de Madagascar, B.P. 1274, Ambatofotsikely, Antananarivo 101, Madagascar. Contents lists available at ScienceDirect Insect Biochemistry and Molecular Biology journal homepage: www.elsevier.com/locate/ibmb 0965-1748/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.ibmb.2010.02.005 Insect Biochemistry and Molecular Biology 40 (2010) 317e324

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Insecticide resistance in Culex pipiens quinquefasciatus and Aedes albopictusmosquitoes from La Réunion Island
Michaël Luciano Tantely a,1, Pablo Tortosa b, Haoues Alout a, Claire Berticat a, Arnaud Berthomieu a,Abdoul Rutee c, Jean-Sébastien Dehecq c, Patrick Makoundou a, Pierrick Labbé a,Nicole Pasteur a, Mylène Weill a,*aUniversité Montpellier 2, UMR CNRS 5554, Institut des Sciences de l'Evolution, Génétique de l'Adaptation, C.C. 065, Place Eugène Bataillon, 34095 Montpellier 05, FrancebCentre de Recherche et de Veille sur les Maladies Emergentes dans l'Océan Indien, 2 rue Maxime Rivière, 97410 Ste Clotilde, Ile de La Réunion, FrancecDirection Régionale des Affaires Sanitaires et Sociales, 2 bis avenue Georges Brassens, BP50 97408 Saint-Denis, Ile de La Réunion, France
a r t i c l e i n f o
Article history:Received 1 December 2009Received in revised form16 February 2010Accepted 18 February 2010
Keywords:Insecticide resistanceCulex pipiens quinquefasciatusAedes albopictusVector controlDieldrin resistance
a b s t r a c t
Resistance to insecticides was monitored on Culex pipiens quinquefasciatusmosquitoes collected in twelvelocalities of La Réunion, a geographically isolated island of the Indian Ocean. This mosquito is of medicalconcern in the region as a known vector for !lariasis and a potential vector for West Nile and Rift ValleyFever viruses. Our bioassays indicated the presence of resistance to all tested insecticides, i.e. organo-chlorides, organophosphates and pyrethroids. A molecular investigation revealed a higher frequency ofresistance genes in the coastal areas compared to elevated rural sites, probably re"ecting the differentnature of insecticide pressures together with the genetic cost of resistance alleles. A simple molecular testwas developed to detect RdlR allele, encoding a g-aminobutyric acid (GABA) receptor resistant to dieldrin.Unexpectedly high RdlR frequencies were recorded over the whole island, despite this insecticide havingbeen banned for over 15 years. This resistant allele was also detected for the !rst time in two samples ofAedes albopictus, a species recently involved in severe Chikungunya epidemics on the island. RdlR selectionin these two mosquito species discloses current insecticide pressures in urban areas, from unknownorigins, that should be taken into account to develop vector control strategies.
! 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Resistance to insecticides that impairs nervous transmission hasbeen extensively studied as it is critical for a proper management ofinsect control. It is also of particular interest for evolutionarybiologists: it represents an almost unique example for monitoringthe genetic consequences of a well characterized alteration of theenvironment. In mosquitoes, insecticide resistance is essentiallyachieved through two mechanisms: increased detoxi!cation andtarget insensitivity (review in Nauen, 2007; Whalon et al., 2008).The former results from an increased activity of detoxifyingenzymes: esterases, cytochrome P450 oxidases or glutathione-S-transferases. The latter is associated with target modi!cationsthat lower their af!nity for the considered insecticide. There arefew insecticide targets: the synaptic acetylcholinesterase (AChE1)encoded by the ace-1 gene, the g-aminobutyric acid (GABA)
receptor gene encoded by Rdl and the voltage-dependent sodiumchannel encoded by kdr. Each of these genes has a limited numberof possible mutable positions. As far as mosquitoes are concerned,three substitutions conferring resistance have been recorded in thesynaptic AChE1 (G119S, F290V and F331W), two mutations (A302Sand A302G) in the GABA receptor, and two substitutions in thesodium channel (L1014F and L1014S) in Culex and Anopheles, whileadditional mutations were recorded on the same target in Aedesaegypti (ffrench-Constant et al., 2000; Brengues et al., 2003; Weillet al., 2003; Du et al., 2005; Alout et al., 2007a, 2007b; Pintoet al., 2007; Saavedra-Rodriguez et al., 2007).
Among all the species controlled with insecticides, Culex pipiensis of particular interest because it can be considered as an indicatorto monitor the nature and persistence of insecticides in the envi-ronment: most of its natural breeding sites are indeed localized invarious types of drainages that collect !lthy waters over large areas.A full set of biochemical and molecular tools has been developed toidentify resistance genes in this species. Moreover, its resistancealleles evolve rapidly and can be easily tracked. Besides, over the last30e40 years, substantial physiological and molecular data havebeen documented on the evolutionary constraints of insecticide
* Corresponding author. Fax: !33 4 67 14 36 22.E-mail address: [email protected] (M. Weill).
1 Present address: Laboratoire d'Entomologie Médicale, Institut Pasteur deMadagascar, B.P. 1274, Ambatofotsikely, Antananarivo 101, Madagascar.
Contents lists available at ScienceDirect
Insect Biochemistry and Molecular Biology
journal homepage: www.elsevier .com/locate/ ibmb
0965-1748/$ e see front matter ! 2010 Elsevier Ltd. All rights reserved.doi:10.1016/j.ibmb.2010.02.005
Insect Biochemistry and Molecular Biology 40 (2010) 317e324

resistance in Cx. pipiens (Raymond et al., 2001; Weill et al., 2003,2004, 2005). In particular, it was clearly shown that some resis-tance genes can be associated with an important !tness cost(Lenormand et al., 1999; Gazave et al., 2001; Berticat et al., 2002,2008; Bourguet et al., 2004; Duron et al., 2006). As a result, thefrequency of resistance alleles generally decreases with the end ofinsecticide treatments (Silvestrini et al., 1998) or in non-treatedareas (Lenormand et al., 1999).
In this study, we investigated the levels of resistance ofCx. pipiens quinquefasciatus against three insecticide families,organophosphates (OPs), pyrethroids (PYRs) and organochlorides(OCs), and the frequencies of the associated resistance genesthrough different ecotopes of La Réunion Island. Located in theSouth West of Indian Ocean, the geographic isolation of thissubtropical island makes natural migrations rare, and mainlyassociated with hurricane episodes or unintentional transport byhumans. Therefore, resistance levels and resistance gene frequen-cies are expected to re"ect the insecticide pressure that is locallyapplied. The island is mountainous: the center, rural and high, isless submitted to insecticide treatments than the coastal areaswhich are more populated and warmer. The coast is of disparatenature, both urban or rural and dedicated to sugar cane or truckfarming agriculture. La Réunion is therefore an interesting islandmodel for investigating various insecticide selective pressures(agriculture and/or public health). Finally, the island was severelyaffected by Chikungunya epidemics in the year 2006, which led theauthorities to implement a large mosquito control program tar-geting larval and adult stages in all populated coastal areas.Temephos and fenithrothion (OP), targeting larval and adult stages,respectively, were implemented until mid February 2006. Then,temephos was replaced by Bti (Bacillus thuringiensis var. israelensistoxins) and fenithrothion by deltamethrin (PYR) until 2007.
Our study shows that resistance to all tested insecticide classesis found in Cx. pipiens natural populations. The distribution ofresistance alleles turned to be rather different on the coast versusthe island center, probably re"ecting the distinct insecticide pres-sures exerted on mosquitoes in the different sites and the !tnesscost associated with resistance genes. Interestingly, a very highlevel of dieldrin resistance (OC), the use of which was of!ciallyprohibited in 1994, was recorded. Dieldrin resistant allelefrequencies ranged from less than 10% in the high altitude areas, toclose !xation in the North-West quarter of the island, including thecapital St Denis. Interestingly, RdlR resistant allele was also detectedin Ae. albopictus although with lower frequencies in the twoinvestigated populations. A possible explanation for this unex-pected resistance is proposed.
2. Materials and methods
2.1. Mosquito samples and strains
Cx. pipiens was collected as larvae or adults in 12 localities of LaRéunion (Table 1, Fig. 1) in 2008. Larvae were reared to adults underlaboratoryconditions, and stored in liquid nitrogen. Two collections,Ste Suzanne and St Leu, were allowed to breed in the laboratory forestablishing two strains (Suz and Leu) which were maintainedduring the course of this study. Reference strains included Slabsusceptible line (Georghiou et al., 1966), SR and Homoval lineswhich are homozygous for the G119S or F290V resistant AChE1alleles, respectively (Alout et al., 2009; Berticat et al., 2002), andvarious lines homozygous for different over-produced esterases[SA1, SA2, SA4, Berticat et al., 2002]. Ae. albopictus specimens werecollected as adults in St Denis and as larvae in natural breeding sitesof Ravine à Jacques (RaJ, Fig. 1).
2.2. Bioassays
Bioassays were performed as described previously (Raymondet al., 1986) using ethanol solutions of permethrin (PYR, 99% [AI],Riedel de Haen, Seelze, Germany), deltamethrin (PYR, 98.8% [AI],Bayer AG, Leverkusen), temephos (OP, 95% [AI], AmericanCyanamid,Princeton, NJ), ethyl-chlorpyrifos (OP, 99% [AI], Dow Chemicals,Midland, MI), malathion (OP, 96% [AI], Procida, Marseille, France),and dieldrin (OC, 95% [AI], Cil Cluzeau, Courbevoie, France). Mostbioassays included 5 doses and up to 3 replicates per dose, whenenough larvae were available. They were conducted on sets of 20early 4th instars in a !nal volume of 100ml of water containing 1mlof ethanol solution of the tested insecticide. The effect of Pb(piperonyl butoxide, Fluka AG, Buchs, Switzerland)was investigatedby exposing larvae to a standard sublethal dose (5 mg/L) beforeaddition of the tested insecticide.
Mortality was recorded after 24 h exposure and analyzed usingthe Priprobit software (Sakuma, 1998) based on Finney (1971). Foreach strain, this software analyzes the doseemortality response in(a) testing its linearity and computing the slope with its standard
Table 1Cx. pipiens quinquefasciatus collected in 12 localities of La Réunion. Exact locations ofsites are given Fig. 1.
Sample # Sample name Nature of collections
1 St Joseph Larvae (domestic breeding sites in a cemetery)2 St Pierre Adults (2 ovitraps in the “Ravine d'Abord”)3 St Leu Larvae (water treating plant)4 La Possession Larvae (domestic breeding sites in a cemetery)5 St Bernard Adults (7 ovitraps in a cemetery)6 St Denis Adults (10 ovitraps in a public park)7 Ste Suzanne Larvae (broken sewage pipe)8 St André Adults (7 ovitraps in a cemetery)9 Bras Panon Adults (6 ovitraps in a cemetery)10 St Benoît Adults (11 ovitraps in a cemetery)11 Ste Rose Larvae (domestic breeding sites in a cemetery)12 Plaine des Palmistes Adults (8 ovitraps in a cemetery)
Fig. 1. Localities of sampling in the island of La Réunion. Grey areas indicate regions ofhuman habitat and the “communes” of La Réunion are delineated. Locations of Culexpipiens sampling are shown as black dots and numbered along the coast from West ofthe active volcano Piton de la Fournaise. Locations of Aedes albopictus sampling arepointed by red arrows. One corresponds to the same location as the Cx. pipiens samplenamed St Denis, the other to the Ravine à Jacques (RaJ). (For interpretation of thereferences to colour in this !gure legend, the reader is referred to the web version ofthis article.)
M.L. Tantely et al. / Insect Biochemistry and Molecular Biology 40 (2010) 317e324318
deviation (SD), and (b) estimating the lethal concentration givinga chosen mortality (here 50%, named LC50) together with its 95%con!dence limits (CL). It also compares two doseemortalityresponses, tests their parallelism and computes the resistance ratio(RR) or the synergism ratio (SR) with its 95% CL at LC50. Twosamples displaying RR or SR with overlapping CL are consideredidentical within the 5% error margin.
2.3. Enzymatic phenotyping of the ace-1 and Ester genes
The phenotypes of AChE1 (acetylcholinesterase-1), encoded bythe ace-1 gene, were studied using the previously described TDPtest (Alout et al., 2009). Over-produced esterases (Ester locus) wereinvestigated after separating the proteins of thorax homogenatesby starch gel electrophoresis in Tris-Maleate buffer-EDTA (pH 7.4).Esterase activity was revealed using a- and b-naphthyl acetates andFast Garnett (Pasteur et al., 1987). The esterases were identi!ed bycomparing their electrophoretic mobility to that of the laboratoryreference strains.
2.4. Characterization of kdr and Rdl mutations conferring resistance
Total DNA of single mosquitoes was extracted using a CTABprotocol (Rogers and Bendich, 1988). The Leu to Phe kdr substitu-tion was identi!ed with the PASA method (Martinez-Torres et al.,1998). A PCR-RFLP test was developed to detect the Ala to Sersubstitution in Rdl of Cx. pipiens and Ae. albopictus mosquitoes.A 232 bp fragment of the GABA receptor encoding gene wasampli!ed by PCRwithmqGABAdir (50-TGTACGTTCGATGGGTTAT-30)and mqGABArev (50-CATGACGAAGCATGTGCCTA-30) primers. PCRwas run for 30 cycles (94 "C for 30 s, 52 "C for 30 s, and 72 "C for1 min), followed by 5 min at 72 "C. The PCR product was thendigested during 2 h at 60 "C with BstAPI restriction endonuclease(New England Biolabs) which selectively cleaves the susceptibleallele. The Rdl PCR-RFLP test was validated by sequencing. PCRproducts were puri!ed with the QIAquick gel extraction kit (QIA-GEN, Valencia, CA) and directly sequenced on an ABI Prism 310sequencer using the Big Dye terminator kit (Applied Biosystems,Foster City, CA).
3. Results
3.1. Analysis of resistance levels
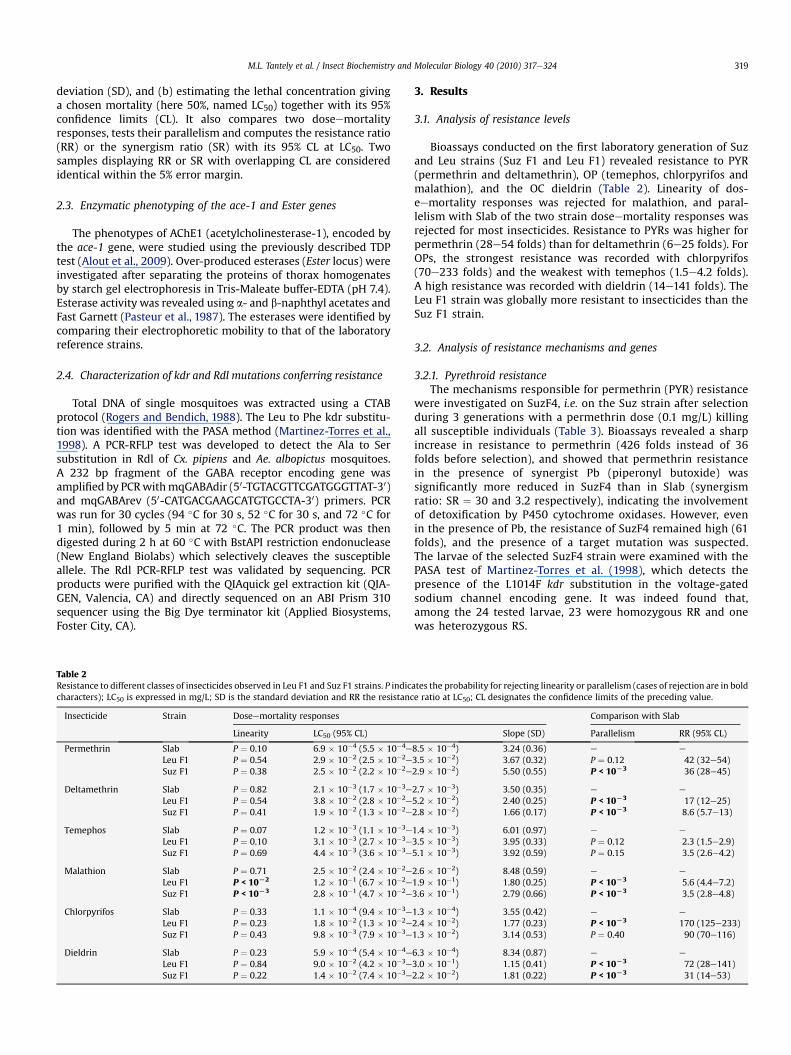
Bioassays conducted on the !rst laboratory generation of Suzand Leu strains (Suz F1 and Leu F1) revealed resistance to PYR(permethrin and deltamethrin), OP (temephos, chlorpyrifos andmalathion), and the OC dieldrin (Table 2). Linearity of dos-eemortality responses was rejected for malathion, and paral-lelism with Slab of the two strain doseemortality responses wasrejected for most insecticides. Resistance to PYRs was higher forpermethrin (28e54 folds) than for deltamethrin (6e25 folds). ForOPs, the strongest resistance was recorded with chlorpyrifos(70e233 folds) and the weakest with temephos (1.5e4.2 folds).A high resistance was recorded with dieldrin (14e141 folds). TheLeu F1 strain was globally more resistant to insecticides than theSuz F1 strain.
3.2. Analysis of resistance mechanisms and genes
3.2.1. Pyrethroid resistanceThe mechanisms responsible for permethrin (PYR) resistance
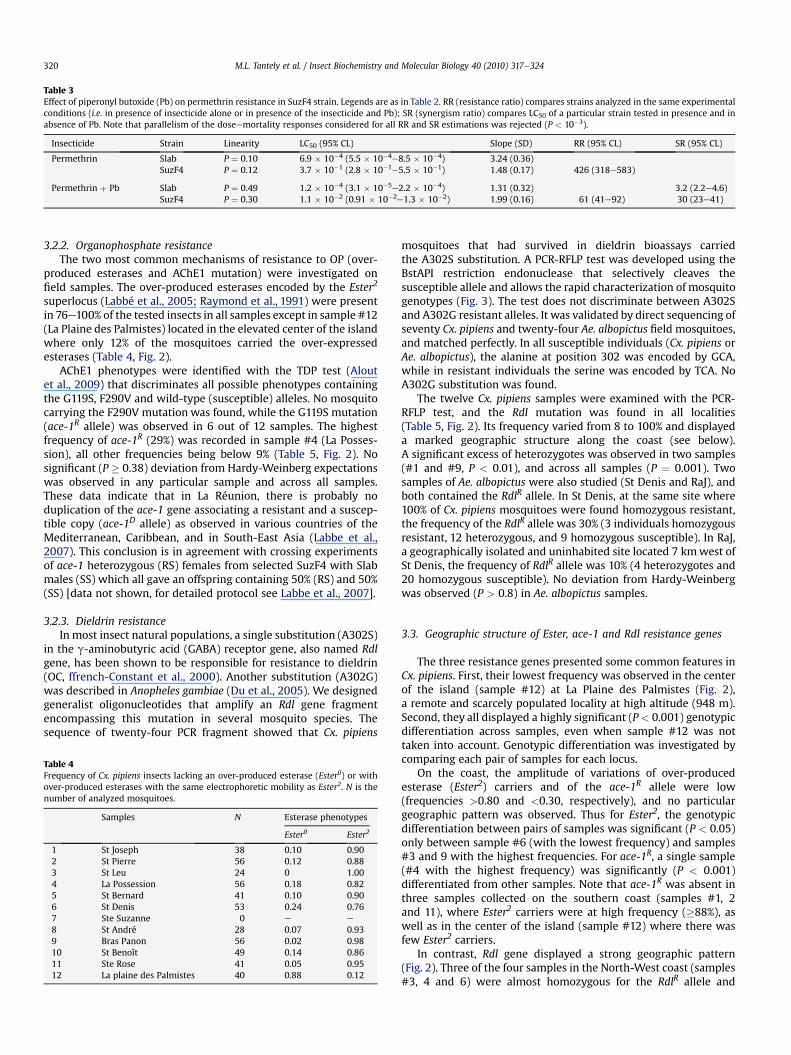
were investigated on SuzF4, i.e. on the Suz strain after selectionduring 3 generations with a permethrin dose (0.1 mg/L) killingall susceptible individuals (Table 3). Bioassays revealed a sharpincrease in resistance to permethrin (426 folds instead of 36folds before selection), and showed that permethrin resistancein the presence of synergist Pb (piperonyl butoxide) wassigni!cantly more reduced in SuzF4 than in Slab (synergismratio: SR # 30 and 3.2 respectively), indicating the involvementof detoxi!cation by P450 cytochrome oxidases. However, evenin the presence of Pb, the resistance of SuzF4 remained high (61folds), and the presence of a target mutation was suspected.The larvae of the selected SuzF4 strain were examined with thePASA test of Martinez-Torres et al. (1998), which detects thepresence of the L1014F kdr substitution in the voltage-gatedsodium channel encoding gene. It was indeed found that,among the 24 tested larvae, 23 were homozygous RR and onewas heterozygous RS.
Table 2Resistance to different classes of insecticides observed in Leu F1 and Suz F1 strains. P indicates the probability for rejecting linearity or parallelism (cases of rejection are in boldcharacters); LC50 is expressed in mg/L; SD is the standard deviation and RR the resistance ratio at LC50; CL designates the con!dence limits of the preceding value.
Insecticide Strain Doseemortality responses Comparison with Slab
Linearity LC50 (95% CL) Slope (SD) Parallelism RR (95% CL)
Permethrin Slab P # 0.10 6.9 $ 10%4 (5.5 $ 10%4e8.5 $ 10%4) 3.24 (0.36) e e
Leu F1 P # 0.54 2.9 $ 10%2 (2.5 $ 10%2e3.5 $ 10%2) 3.67 (0.32) P # 0.12 42 (32e54)Suz F1 P # 0.38 2.5 $ 10%2 (2.2 $ 10%2e2.9 $ 10%2) 5.50 (0.55) P < 10L3 36 (28e45)
Deltamethrin Slab P # 0.82 2.1 $ 10%3 (1.7 $ 10%3e2.7 $ 10%3) 3.50 (0.35) e e
Leu F1 P # 0.54 3.8 $ 10%2 (2.8 $ 10%2e5.2 $ 10%2) 2.40 (0.25) P < 10L3 17 (12e25)Suz F1 P # 0.41 1.9 $ 10%2 (1.3 $ 10%2e2.8 $ 10%2) 1.66 (0.17) P < 10L3 8.6 (5.7e13)
Temephos Slab P # 0.07 1.2 $ 10%3 (1.1 $ 10%3e1.4 $ 10%3) 6.01 (0.97) e e
Leu F1 P # 0.10 3.1 $ 10%3 (2.7 $ 10%3e3.5 $ 10%3) 3.95 (0.33) P # 0.12 2.3 (1.5e2.9)Suz F1 P # 0.69 4.4 $ 10%3 (3.6 $ 10%3e5.1 $ 10%3) 3.92 (0.59) P # 0.15 3.5 (2.6e4.2)
Malathion Slab P # 0.71 2.5 $ 10%2 (2.4 $ 10%2e2.6 $ 10%2) 8.48 (0.59) e e
Leu F1 P < 10L2 1.2 $ 10%1 (6.7 $ 10%2e1.9 $ 10%1) 1.80 (0.25) P < 10L3 5.6 (4.4e7.2)Suz F1 P < 10L3 2.8 $ 10%1 (4.7 $ 10%2e3.6 $ 10%1) 2.79 (0.66) P < 10L3 3.5 (2.8e4.8)
Chlorpyrifos Slab P # 0.33 1.1 $ 10%4 (9.4 $ 10%3e1.3 $ 10%4) 3.55 (0.42) e e
Leu F1 P # 0.23 1.8 $ 10%2 (1.3 $ 10%2e2.4 $ 10%2) 1.77 (0.23) P < 10L3 170 (125e233)Suz F1 P # 0.43 9.8 $ 10%3 (7.9 $ 10%3e1.3 $ 10%2) 3.14 (0.53) P # 0.40 90 (70e116)
Dieldrin Slab P # 0.23 5.9 $ 10%4 (5.4 $ 10%4e6.3 $ 10%4) 8.34 (0.87) e e
Leu F1 P # 0.84 9.0 $ 10%2 (4.2 $ 10%3e3.0 $ 10%1) 1.15 (0.41) P < 10L3 72 (28e141)Suz F1 P # 0.22 1.4 $ 10%2 (7.4 $ 10%3e2.2 $ 10%2) 1.81 (0.22) P < 10L3 31 (14e53)
M.L. Tantely et al. / Insect Biochemistry and Molecular Biology 40 (2010) 317e324 319
3.2.2. Organophosphate resistanceThe two most common mechanisms of resistance to OP (over-
produced esterases and AChE1 mutation) were investigated on!eld samples. The over-produced esterases encoded by the Ester2
superlocus (Labbé et al., 2005; Raymond et al., 1991) were presentin 76e100% of the tested insects in all samples except in sample #12(La Plaine des Palmistes) located in the elevated center of the islandwhere only 12% of the mosquitoes carried the over-expressedesterases (Table 4, Fig. 2).
AChE1 phenotypes were identi!ed with the TDP test (Aloutet al., 2009) that discriminates all possible phenotypes containingthe G119S, F290V and wild-type (susceptible) alleles. No mosquitocarrying the F290V mutation was found, while the G119S mutation(ace-1R allele) was observed in 6 out of 12 samples. The highestfrequency of ace-1R (29%) was recorded in sample #4 (La Posses-sion), all other frequencies being below 9% (Table 5, Fig. 2). Nosigni!cant (P & 0.38) deviation from Hardy-Weinberg expectationswas observed in any particular sample and across all samples.These data indicate that in La Réunion, there is probably noduplication of the ace-1 gene associating a resistant and a suscep-tible copy (ace-1D allele) as observed in various countries of theMediterranean, Caribbean, and in South-East Asia (Labbe et al.,2007). This conclusion is in agreement with crossing experimentsof ace-1 heterozygous (RS) females from selected SuzF4 with Slabmales (SS) which all gave an offspring containing 50% (RS) and 50%(SS) [data not shown, for detailed protocol see Labbe et al., 2007].
3.2.3. Dieldrin resistanceIn most insect natural populations, a single substitution (A302S)
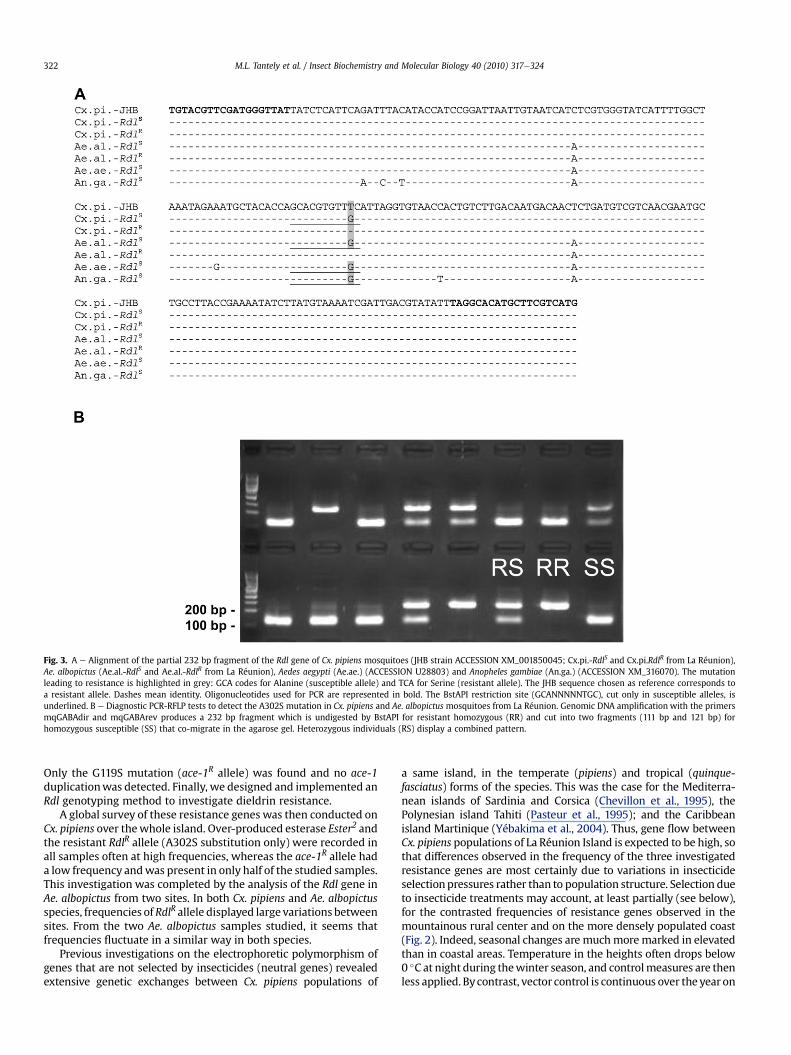
in the g-aminobutyric acid (GABA) receptor gene, also named Rdlgene, has been shown to be responsible for resistance to dieldrin(OC, ffrench-Constant et al., 2000). Another substitution (A302G)was described in Anopheles gambiae (Du et al., 2005). We designedgeneralist oligonucleotides that amplify an Rdl gene fragmentencompassing this mutation in several mosquito species. Thesequence of twenty-four PCR fragment showed that Cx. pipiens
mosquitoes that had survived in dieldrin bioassays carriedthe A302S substitution. A PCR-RFLP test was developed using theBstAPI restriction endonuclease that selectively cleaves thesusceptible allele and allows the rapid characterization of mosquitogenotypes (Fig. 3). The test does not discriminate between A302Sand A302G resistant alleles. It was validated by direct sequencing ofseventy Cx. pipiens and twenty-four Ae. albopictus !eld mosquitoes,and matched perfectly. In all susceptible individuals (Cx. pipiens orAe. albopictus), the alanine at position 302 was encoded by GCA,while in resistant individuals the serine was encoded by TCA. NoA302G substitution was found.
The twelve Cx. pipiens samples were examined with the PCR-RFLP test, and the Rdl mutation was found in all localities(Table 5, Fig. 2). Its frequency varied from 8 to 100% and displayeda marked geographic structure along the coast (see below).A signi!cant excess of heterozygotes was observed in two samples(#1 and #9, P < 0.01), and across all samples (P # 0.001). Twosamples of Ae. albopictus were also studied (St Denis and RaJ), andboth contained the RdlR allele. In St Denis, at the same site where100% of Cx. pipiens mosquitoes were found homozygous resistant,the frequency of the RdlR allele was 30% (3 individuals homozygousresistant, 12 heterozygous, and 9 homozygous susceptible). In RaJ,a geographically isolated and uninhabited site located 7 kmwest ofSt Denis, the frequency of RdlR allele was 10% (4 heterozygotes and20 homozygous susceptible). No deviation from Hardy-Weinbergwas observed (P > 0.8) in Ae. albopictus samples.
3.3. Geographic structure of Ester, ace-1 and Rdl resistance genes
The three resistance genes presented some common features inCx. pipiens. First, their lowest frequency was observed in the centerof the island (sample #12) at La Plaine des Palmistes (Fig. 2),a remote and scarcely populated locality at high altitude (948 m).Second, they all displayed a highly signi!cant (P< 0.001) genotypicdifferentiation across samples, even when sample #12 was nottaken into account. Genotypic differentiation was investigated bycomparing each pair of samples for each locus.
On the coast, the amplitude of variations of over-producedesterase (Ester2) carriers and of the ace-1R allele were low(frequencies >0.80 and <0.30, respectively), and no particulargeographic pattern was observed. Thus for Ester2, the genotypicdifferentiation between pairs of samples was signi!cant (P < 0.05)only between sample #6 (with the lowest frequency) and samples#3 and 9 with the highest frequencies. For ace-1R, a single sample(#4 with the highest frequency) was signi!cantly (P < 0.001)differentiated from other samples. Note that ace-1R was absent inthree samples collected on the southern coast (samples #1, 2and 11), where Ester2 carriers were at high frequency (&88%), aswell as in the center of the island (sample #12) where there wasfew Ester2 carriers.
In contrast, Rdl gene displayed a strong geographic pattern(Fig. 2). Three of the four samples in the North-West coast (samples#3, 4 and 6) were almost homozygous for the RdlR allele and
Table 3Effect of piperonyl butoxide (Pb) on permethrin resistance in SuzF4 strain. Legends are as in Table 2. RR (resistance ratio) compares strains analyzed in the same experimentalconditions (i.e. in presence of insecticide alone or in presence of the insecticide and Pb); SR (synergism ratio) compares LC50 of a particular strain tested in presence and inabsence of Pb. Note that parallelism of the doseemortality responses considered for all RR and SR estimations was rejected (P < 10%3).
Insecticide Strain Linearity LC50 (95% CL) Slope (SD) RR (95% CL) SR (95% CL)
Permethrin Slab P # 0.10 6.9 $ 10%4 (5.5 $ 10%4e8.5 $ 10%4) 3.24 (0.36)SuzF4 P # 0.12 3.7 $ 10%1 (2.8 $ 10%1e5.5 $ 10%1) 1.48 (0.17) 426 (318e583)
Permethrin ! Pb Slab P # 0.49 1.2 $ 10%4 (3.1 $ 10%5e2.2 $ 10%4) 1.31 (0.32) 3.2 (2.2e4.6)SuzF4 P # 0.30 1.1 $ 10%2 (0.91 $ 10%2e1.3 $ 10%2) 1.99 (0.16) 61 (41e92) 30 (23e41)
Table 4Frequency of Cx. pipiens insects lacking an over-produced esterase (Ester0) or withover-produced esterases with the same electrophoretic mobility as Ester2. N is thenumber of analyzed mosquitoes.
Samples N Esterase phenotypes
Ester0 Ester2
1 St Joseph 38 0.10 0.902 St Pierre 56 0.12 0.883 St Leu 24 0 1.004 La Possession 56 0.18 0.825 St Bernard 41 0.10 0.906 St Denis 53 0.24 0.767 Ste Suzanne 0 e e
8 St André 28 0.07 0.939 Bras Panon 56 0.02 0.9810 St Benoît 49 0.14 0.8611 Ste Rose 41 0.05 0.9512 La plaine des Palmistes 40 0.88 0.12
M.L. Tantely et al. / Insect Biochemistry and Molecular Biology 40 (2010) 317e324320

showed no genotypic differentiation (P > 0.49) between them.However, they were signi!cantly different (P < 0.001) from allother samples, including sample #5 collected in this sector but at analtitude of 450 m, where human habitat is at low density. South ofsample #3 (St Leu) and East of sample #6 (St Denis), RdlR allelefrequency decreased signi!cantly (P < 0.001). There was nosigni!cant (P> 0.05) genotypic differentiation between samples #1and #2 in the South-East, and between samples #7, 8, 9 and 10 inthe North-East when taking into account multiple tests (Bonferronisequential test, Holm, 1979). In the South-East, RdlR frequencycontinued to decrease (18% in sample #11). Finally in the center ofthe island, RdlR frequency was 8% (sample #12), and there was nosigni!cant (P# 0.30) genotypic differentiation between this sampleand sample #11 from the South-East coast.
From the two Ae. albopictus samples studied, it appears that Rdlgene also displays a signi!cant (P # 0.03) genotypic differentiationin this species. It can be noted that in Ae. albopictus as well as inCx. pipiens, the samples captured in the same site of St Denis(old botanical park) have a signi!cantly higher frequency of RdlR
allele than those captured west of this site in areas (RaJ andSt Bernard) where human habitat is scarce or non existent.
4. Discussion
A survey of insecticide resistance in Cx. pipiens mosquitoescollected in 2008 on the island of La Réunion was conducted withbioassays performed on two samples (St Leu and Ste Suzanne). Itrevealed a marked resistance (&10 folds) to the pyrethroids (PYR)permethrin and deltamethrin, to chlorpyrifos (OP) and to theorganochloride (OC) dieldrin. Permethrin resistance was due inpart to an increased detoxi!cation (by P450 oxidases), and in part tothe presence of the L1014F kdr mutation in the strain isolated fromSt Suzanne. The simultaneous presence of these two PYR resistancemechanisms is in agreement with the wide use of this insecticideclass in local agriculture as well as by vector control units, whichhave been particularly active since the 2006 Chikungunyaepidemics. As for OP resistance, both over-produced esterases andmutation on the ace-1 gene encoding OPs target were observed.
Fig. 2. Comparison of the resistance gene frequencies in the various samples from LaRéunion. Samples are numbered as in Fig. 1. For each locus, the frequency of theresistant allele is represented in black.
Table 5Frequency of the ace-1R and RdlR alleles in Cx. pipiens samples from la Réunion. Fisindicates excess (Fis < 0) or de!cit (Fis > 0) of heterozygotes in each sample andacross all samples; P is the probability that observations deviate from expectations(bold when signi!cant). All estimates were computed with GENEPOP (Raymond andRousset, 1995).
Samples ace-1 gene Rdl gene
N ace-1R Fis P N RdlR Fis P
1 St Joseph 38 0 e e 17 0.56 %0.778 0.0022 St Pierre 56 0 e e 23 0.70 %0.005 0.693 St Leu 23 0.04 %0.023 0.98 23 1.00 e e
4 La Possession 56 0.29 0.021 0.56 24 0.96 %0.022 0.985 St Bernard 41 0 e e 27 0.39 %0.150 0.366 St Denis 56 0.08 %0.078 0.71 21 1.00 e e
7 Ste Suzanne 24 0.02 0.000 e 24 0.48 %0.232 0.238 St André 28 0.07 %0.059 0.89 26 0.39 0.206 0.269 Bras Panon 56 0 e e 23 0.57 %0.760 0.000310 St Benoît 56 0.05 %0.048 0.87 23 0.41 %0.147 0.4011 Ste Rose 41 0 e e 20 0.18 %0.191 0.5312 Plaine des
Palmistes40 0 e e 24 0.08 %0.071 0.87
All samples 515 %0.147 0.38 275 %0.225 0.0009
M.L. Tantely et al. / Insect Biochemistry and Molecular Biology 40 (2010) 317e324 321
Only the G119S mutation (ace-1R allele) was found and no ace-1duplicationwas detected. Finally, we designed and implemented anRdl genotyping method to investigate dieldrin resistance.
A global survey of these resistance genes was then conducted onCx. pipiens over thewhole island. Over-produced esterase Ester2 andthe resistant RdlR allele (A302S substitution only) were recorded inall samples often at high frequencies, whereas the ace-1R allele hada low frequency andwas present in only half of the studied samples.This investigation was completed by the analysis of the Rdl gene inAe. albopictus from two sites. In both Cx. pipiens and Ae. albopictusspecies, frequencies of RdlR allele displayed large variations betweensites. From the two Ae. albopictus samples studied, it seems thatfrequencies "uctuate in a similar way in both species.
Previous investigations on the electrophoretic polymorphism ofgenes that are not selected by insecticides (neutral genes) revealedextensive genetic exchanges between Cx. pipiens populations of
a same island, in the temperate (pipiens) and tropical (quinque-fasciatus) forms of the species. This was the case for the Mediterra-nean islands of Sardinia and Corsica (Chevillon et al., 1995), thePolynesian island Tahiti (Pasteur et al., 1995); and the Caribbeanisland Martinique (Yébakima et al., 2004). Thus, gene "ow betweenCx. pipiens populations of La Réunion Island is expected to be high, sothat differences observed in the frequency of the three investigatedresistance genes are most certainly due to variations in insecticideselectionpressures rather than to population structure. Selection dueto insecticide treatments may account, at least partially (see below),for the contrasted frequencies of resistance genes observed in themountainous rural center and on the more densely populated coast(Fig. 2). Indeed, seasonal changes are muchmoremarked in elevatedthan in coastal areas. Temperature in the heights often drops below0 "C at night during thewinter season, and controlmeasures are thenless applied. Bycontrast, vector control is continuous over the year on
Fig. 3. A e Alignment of the partial 232 bp fragment of the Rdl gene of Cx. pipiens mosquitoes (JHB strain ACCESSION XM_001850045; Cx.pi.-RdlS and Cx.pi.RdlR from La Réunion),Ae. albopictus (Ae.al.-RdlS and Ae.al.-RdlR from La Réunion), Aedes aegypti (Ae.ae.) (ACCESSION U28803) and Anopheles gambiae (An.ga.) (ACCESSION XM_316070). The mutationleading to resistance is highlighted in grey: GCA codes for Alanine (susceptible allele) and TCA for Serine (resistant allele). The JHB sequence chosen as reference corresponds toa resistant allele. Dashes mean identity. Oligonucleotides used for PCR are represented in bold. The BstAPI restriction site (GCANNNNNTGC), cut only in susceptible alleles, isunderlined. B e Diagnostic PCR-RFLP tests to detect the A302S mutation in Cx. pipiens and Ae. albopictus mosquitoes from La Réunion. Genomic DNA ampli!cation with the primersmqGABAdir and mqGABArev produces a 232 bp fragment which is undigested by BstAPI for resistant homozygous (RR) and cut into two fragments (111 bp and 121 bp) forhomozygous susceptible (SS) that co-migrate in the agarose gel. Heterozygous individuals (RS) display a combined pattern.
M.L. Tantely et al. / Insect Biochemistry and Molecular Biology 40 (2010) 317e324322

the coast. In addition, the low frequency of resistance allelesmeasured in the elevated areas could be reinforced by their !tnesscost, which has been well documented for over-produced esterasesand for the ace-1R allele (Silvestrini et al., 1998; Lenormand et al.,1999; Gazave et al., 2001; Berticat et al., 2002; Bourguet et al.,2004; Duron et al., 2006). Along the coast, the three resistancegenes displayed different patterns. Ester2 frequency was high in alllocalities (>76%)while ace-1R had an overall lower frequency (<30%)and was absent from the south of the island. Ester2 is among the !rstOP resistance genes identi!ed in Cx. pipiens, andwas recorded in EastAfrica in 1972 (Curtis and Pasteur,1981). This resistant allele is highlyinvasive (Raymond et al., 1991; Yébakima et al., 2004), and waspresumably introduced in La Réunion several decades ago. Its highfrequency on the coast is not surprising since OPs have been inten-sively used in agriculture against fruit "ies (Diptera: Tephritidae,Ceratitis capitata, Ceratitis rosa, Bactrocera zonata) and for mosquitocontrol before they were banned in February 2006 by the mosquitocontrol agency. The contrast between Ester2 and ace-1R frequenciesmost probably indicates that ace-1R is a relatively recent allele in LaRéunion, since both mechanisms confer resistance to OPs. Further-more we can infer that ace-1R appeared by mutation or migration inthe north of the island (La Possession, sample #4, is close to the onlyinternational harbor of the island). This view is in agreementwith theabsence of ace-1R in the South despite the high frequencies of Ester2.From an evolutionary perspective, it would be interesting tomonitorthe ace-1 resistance evolutionary dynamics in La Réunion, as theisland geographic structure may in"uence its spreading, and as newalleles, including ace-1 duplication, would probably occur throughlocal mutation rather than migration.
Of the three resistance genes investigated in Cx. pipiens, Rdldisplayed the largest variability, being almost !xed inmost samplesfrom the North-West coast of the island (from St Leu to St Denis).The excess of heterozygotes observed in all samples (signi!cant insamples #1 and #9, and across all samples) may indicate a dupli-cation of the gene, similar to that found in the aphidMyzus persicae(Anthony et al., 1998), and this aspect is worth investigating in thefuture. The high frequency of RdlR allele in Cx. pipiens and itspresence in Ae. albopictus are surprising as none of the pesticides towhich this allele confers resistance are used or have been used formosquito control in La Réunion. These pesticides include dieldrinand particularly endosulfan used in arboriculture (OCs, ffrench-Constant et al., 2000) which have been banned for all usages inFrance (La Réunion is an overseas department of France) since 1994(décret 92-1074 1992) and 2007 (Journal Of!ciel 2006),respectively. This high RdlR frequency may thus indicate a persis-tence of dieldrin and/or endosulfan in the soil or, more worrisome,an illegal use of these insecticides, despite banning measures. Itmay also be due to !pronil (Kristensen et al., 2005; Le Goff et al.,2005), which is currently used to control termites on the island,or to lindane (Lees and Calder, 1996) and/or antiparasites of theavermectin family (Buckingham et al., 2005), which are also widelyused by veterinarians. Noteworthy, dieldrin, !pronil and lindanehave been detected in coastal waters (ARVAM, Agence pour laRecherche et la VAlorisation Marines, personal communication).Thus, the present data on Cx. pipiens indicate that at least onepesticide targeting the GABA receptor exhibits important concen-tration variations, as RdlR allele is close to !xation over the North-West quarter of La Réunion coast, in contrast with a frequency ofonly 18% in the South-East (Fig. 2).
The high frequencies of the different resistance genes (despitetheir sometimes high !tness cost) undoubtedly attest the applicationin La Réunion of contemporary or very recent strong insecticidepressures. From this work, we consider Cx. pipiens as a promisingsentinel species to survey insecticide pressures due to vectorcontrol, agricultural or any other usages of pesticides. Indeed, direct
measurement of the presence of pesticides in water (liquid chro-matography) detects doses much higher than those toxicto mosquitoes and other non-target species (Eritja and Chevillon,1999). The surprisingly high RdlR frequency detected in Cx. pipiensprompted us to extend this study to Ae. albopictus and allowed toidentify the !rst insecticide resistance gene present in populations ofthis species on La Réunion.
These data highlight the fact that many xenobiotics fromdistinct and not clearly characterized origins exert selection pres-sures on mosquito vectors that transmit serious pathogens.Cx. pipiens is of medical interest in the region as a known vector forlymphatic !lariasis in Comoros (Sabatinelli et al., 1994) andMadagascar (Champetier De Ribes et al., 2000) and might be ofconcern for the Rift Valley Fever virus ongoing epidemics andendemic West Nile Virus (Zeller, 1998). Ae. albopictus is the mainvector for Chikungunya and Dengue viruses on La Réunion (Delatteet al., 2008). The spread of resistance in these species could gravelyimpede the strategies of disease control, and facilitate thehappening of future outbreaks. Altogether, our data strengthen theneed for taking into account pest management measures imple-mented by all economic actors of a region in order to developef!cient vector control strategies.
Acknowledgements
We are very grateful to S. Unal for technical assistance, SamuelHuet for actively participating in Cx. pipiens collections, LaurenceDijoux and Jean Turquet for stimulating discussions on xenobi-otics implementations on La Réunion. This work was !nanced inpart by ANR CHIKVENDOM Santé-Environnement (Ministèredélégué à la Recherche) and by the Conseil Régional de LaRéunion. All sequence data were obtained on the EnvironmentalGenomic Platform of the IFR Montpellier-Environnement-Biodiversité. Contribution 2010.016 of the Institut des Sciences del'Evolution de Montpellier (UMR CNRS 5554).
References
Alout, H., Berthomieu, A., Cui, F., Tan, Y., Berticat, C., Qiao, C., Weill, M., 2007a.Different amino-acid substitutions confer insecticide resistance throughacetylcholinesterase 1 insensitivity in Culex vishnui and Culex tritaeniorhynchus(Diptera: Culicidae) from China. J. Med. Entomol. 44, 463e469.
Alout, H., Berthomieu, A., Hadjivassilis, A., Weill, M., 2007b. A new amino-acidsubstitution in acetylcholinesterase 1 confers insecticide resistance to Culexpipiens mosquitoes from Cyprus. Insect Biochem. Mol. Biol. 37, 41e47.
Alout, H., Labbe, P., Berthomieu, A., Pasteur, N., Weill, M., 2009. Multiple duplica-tions of the rare ace-1 mutation F290V in Culex pipiens natural populations.Insect Biochem. Mol. Biol. 39, 884e891.
Anthony, N., Unruh, T., Ganser, D., ffrench-Constant, R., 1998. Duplication of the RdlGABA receptor subunit gene in an insecticide-resistant aphid, Myzus persicae.Mol. Gen. Genet. 260, 165e175.
Berticat, C., Bonnet, J., Duchon, S., Agnew, P., Weill, M., Corbel, V., 2008. Costs andbene!ts of multiple resistance to insecticides for Culex quinquefasciatusmosquitoes. BMC Evol. Biol. 8, 104.
Berticat, C., Boquien, G., Raymond, M., Chevillon, C., 2002. Insecticide resistancegenes induce a mating competition cost in Culex pipiensmosquitoes. Genet. Res.79, 41e47.
Bourguet, D., Guillemaud, T., Chevillon, C., Raymond, M., 2004. Fitness costs ofinsecticide resistance in natural breeding sites of the mosquito Culex pipiens.Evolution 58, 128e135.
Brengues, C., Hawkes, N.J., Chandre, F., McCarroll, L., Duchon, S., Guillet, P.,Manguin, S., Morgan, J.C., Hemingway, J., 2003. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 17, 87e94.
Buckingham, S.D., Biggin, P.C., Sattelle, B.M., Brown, L.A., Sattelle, D.B., 2005. InsectGABA receptors: splicing, editing, and targeting by antiparasitics and insecti-cides. Mol. Pharmacol. 68, 942e951.
Champetier De Ribes, G., Ranaivoson, G., Lamina Rakotonjanabelo, A.,Radoerimanana, R., Rabeson, D., 2000. Bancroftian !lariasis in Madagascar:persistent endemicity. Med. Trop. (Mars) 60, 141e145.
Chevillon, C., Addis, G., Raymond, M., Marchi, A., 1995. Population structure inMediterranean islands and risk of genetic invasion in Culex pipiens L. (Diptera:Culicidae). Biol. J. Linnean Soc. 55, 329e343.
M.L. Tantely et al. / Insect Biochemistry and Molecular Biology 40 (2010) 317e324 323

Curtis, C.F., Pasteur, N., 1981. Organophosphate resistance in vector populations ofthe complex of Culex pipiens L. (Diptera: Culicidae). Bull. Entomol. Res. 71,153e161.
Delatte, H., Paupy, C., Dehecq, J.S., Thiria, J., Failloux, A.B., Fontenille, D., 2008. Aedesalbopictus, vector of Chikungunya and dengue viruses in Reunion Island:biology and control. Parasite 15, 3e13.
Du, W., Awolola, T.S., Howell, P., Koekemoer, L.L., Brooke, B.D., Benedict, M.Q.,Coetzee, M., Zheng, L., 2005. Independent mutations in the Rdl locus conferdieldrin resistance to Anopheles gambiae and An. arabiensis. Insect Mol. Biol. 14,179e183.
Duron, O., Labbe, P., Berticat, C., Rousset, F., Guillot, S., Raymond, M., Weill, M., 2006.High Wolbachia density correlates with cost of infection for insecticide resistantCulex pipiens mosquitoes. Evolution 60, 303e314.
Eritja, R., Chevillon, C., 1999. Interruption of chemical mosquito control andevolution of insecticide resistance genes in Culex pipiens (Diptera: Culicidae).J. Med. Entomol. 36, 41e49.
ffrench-Constant, R.H., Anthony, N., Aronstein, K., Rocheleau, T., Stilwell, G., 2000.Cyclodiene insecticide resistance: from molecular to population genetics. Annu.Rev. Entomol. 45, 449e466.
Finney, D.J., 1971. Probit Analysis. Cambridge University Press, Cambridge, England.Gazave, E., Chevillon, C., Lenormand, T., Marquine, M., Raymond, M., 2001. Dis-
secting the cost of insecticide resistance genes during the overwintering periodof the mosquito Culex pipiens. Heredity 87, 441e448.
Georghiou, G.P., Metcalf, R.L., Gidden, F.E., 1966. Carbamate-resistance in mosquitos.Selection of Culex pipiens fatigans Wiedemann (#C. quinquefasciatus Say) forresistance to Baygon. Bull. World Health Organ. 35, 691e708.
Holm, S., 1979. A simple sequentially rejective multiple test procedure. Scand. J. Stat.6, 65e70.
Kristensen, M., Hansen, K.K., Jensen, K.M., 2005. Cross-resistance between dieldrinand !pronil in German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol.98, 1305e1310.
Labbe, P., Berthomieu, A., Berticat, C., Alout, H., Raymond, M., Lenormand, T.,Weill, M., 2007. Independent duplications of the acetylcholinesterase geneconferring insecticide resistance in the mosquito Culex pipiens. Mol. Biol. Evol.24, 1056e1067.
Labbé, P., Lenormand, T., Raymond, M., 2005. On the worldwide spread of aninsecticide resistance gene: a role for local selection. J. Evol. Biol. 18, 1471e1484.
Le Goff, G., Hamon, A., Berge, J.B., Amichot, M., 2005. Resistance to !pronil inDrosophila simulans: in"uence of two point mutations in the RDL GABA receptorsubunit. J. Neurochem. 92, 1295e1305.
Lees, G., Calder, J., 1996. Interaction of lindane isomers with chloride currents ininsect membranes: steric requirements for channel modulation and block.Pestic. Biochem. Physiol. 55, 40e48.
Lenormand, T., Bourguet, D., Guillemaud, T., Raymond, M., 1999. Tracking theevolution of insecticide resistance in the mosquito Culex pipiens. Nature 400,861e864.
Martinez-Torres, D., Chandre, F., Williamson, M.S., Darriet, F., Berge, J.B.,Devonshire, A.L., Guillet, P., Pasteur, N., Pauron, D., 1998. Molecular character-ization of pyrethroid knockdown resistance (kdr) in the major malaria vectorAnopheles gambiae s.s. Insect Mol. Biol. 7, 179e184.
Nauen, R., 2007. Insecticide resistance in disease vectors of public health impor-tance. Pest Manage. Sci. 63, 628e633.
Pasteur, N., Marquine, M., Rousset, F., Failloux, A.-B., Chevillon, C., Raymond, M.,1995. The role of passive migration in the dispersal of resistance genes in Culexpipiens quinquefasciatus within French Polynesia. Genet. Res. 66, 139e146.
Pasteur, N., Pasteur, G., Bonhomme, F., Catalan, J., Britton-Davidian, J., 1987. Manueltechnique de génétique par électrophorèse des protéines. Technique et Docu-mentation. Lavoisier, Paris.
Pinto, J., Lynd, A., Vicente, J.L., Santolamazza, F., Randle, N.P., Gentile, G.,Moreno, M., Simard, F., Charlwood, J.D., do Rosario, V.E., Caccone, A., DellaTorre, A., Donnelly, M.J., 2007. Multiple origins of knockdown resistancemutations in the Afrotropical mosquito vector Anopheles gambiae. PLoSOne 2, e1243.
Raymond, M., Berticat, C., Weill, M., Pasteur, N., Chevillon, C., 2001. Insecticideresistance in the mosquito Culex pipiens: what have we learned about adap-tation? Genetica 112e113, 287e296.
Raymond, M., Callaghan, A., Fort, P., Pasteur, N., 1991. Worldwide migration ofampli!ed insecticide resistance genes in mosquitoes. Nature 350,151e153.
Raymond, M., Fournier, D., Bride, J.M., Cuany, A., Berge, J., Magnin, M., Pasteur, N.,1986. Identi!cation of resistance mechanisms in Culex pipiens (Diptera: Culi-cidae) from southern France: insensitive acetylcholinesterase and detoxifyingoxidases. J. Econ. Entomol. 79, 1452e1458.
Raymond, M., Rousset, F., 1995. GENEPOP (Version-1.2) e population geneticssoftware for exact tests and ecumenicism. J. Hered. 86, 248e249.
Rogers, S.O., Bendich, A.J., 1988. Extraction of DNA from plant tissues. In:Gelvin, S.B., Schilperoort, R.A. (Eds.), Plant Molecular Biology Manuel. KluwerAcademic Publisher, Boston.
Saavedra-Rodriguez, K., Urdaneta-Marquez, L., Rajatileka, S., Moulton, M.,Flores, A.E., Fernandez-Salas, I., Bisset, J., Rodriguez, M., McCall, P.J.,Donnelly, M.J., Ranson, H., Hemingway, J., Black, WCt, 2007. A mutation in thevoltage-gated sodium channel gene associated with pyrethroid resistance inLatin American Aedes aegypti. Insect Mol. Biol. 16, 785e798.
Sabatinelli, G., Ranieri, E., Gianzi, F.P., Papakay, M., Cancrini, G., 1994. Role of Culexquinquefasciatus in the transmission of bancroftian !lariasis in the FederalIslamic Republic of Comoros (Indian Ocean). Parasite 1, 71e76.
Sakuma, M., 1998. Probit analysis of preference data. Appl. Entomol. Zool. 33,339e347.
Silvestrini, F., Severini, C., Di Pardo, V., Romi, R., De Matthaeis, E., Raymond, M., 1998.Population structure and dynamics of insecticide resistance genes in Culexpipiens populations from Italy. Heredity 81, 342e348.
Weill, M., Berthomieu, A., Berticat, C., Lutfalla, G., Negre, V., Pasteur, N., Philips, A.,Leonetti, J.-P., Fort, P., Raymond, M., 2004. Insecticide resistance: a silent baseprediction. Curr. Biol. 14, R552eR553.
Weill, M., Labbé, P., Duron, O., Pasteur, N., Fort, P., Raymond, M., 2005. Insecticideresistance in the mosquito Culex pipiens: towards an understanding of theevolution of ace genes. In: Fellowes, M.D.E., Holloway, G.J., Rolff, J. (Eds.), InsectEvolutionary Ecology. Cabin, Oxon.
Weill, M., Lutfalla, G., Mogensen, K., Chandre, F., Berthomieu, A., Berticat, C.,Pasteur, N., Philips, A., Fort, P., Raymond, M., 2003. Comparative genomics:insecticide resistance in mosquito vectors. Nature 423, 136e137.
Whalon, M.E., Mota-Sanchez, D., Hollingworth, R.M., 2008. Analysis of globalpesticide resistance in arthropods. In: Whalon, M.E., Mota-Sanchez, D.,Hollingworth, R.M. (Eds.), Global Pesticide Resistance in Arthropods. CABInternational, Cambridge, MA, p. 192.
Yébakima, A., Marquine, M., Rosine, J., Yp-Tcha, M.M., Pasteur, N., 2004. Evolution ofresistance under insecticide selection pressure in Culex pipiens quinquefasciatus(Diptera, Culicidae) from Martinique. J. Med. Entomol. 41, 718e725.
Zeller, H.G., 1998. Dengue, arbovirus and migrations in the Indian Ocean. Bull. Soc.Pathol. Exot. 91, 56e60.
M.L. Tantely et al. / Insect Biochemistry and Molecular Biology 40 (2010) 317e324324
Related Documents











