J Cancer Res Clin Oncol (2012) 138:405–414 DOI 10.1007/s00432-011-1111-0 123 ORIGINAL PAPER Inhibition of Aurora kinases enhances chemosensitivity to temozolomide and causes radiosensitization in glioblastoma cells Kleiton Silva Borges · Angel Maurício Castro-Gamero · Daniel Antunes Moreno · Vanessa da Silva Silveira · Maria Sol Brassesco · Rosane Gomes de Paula Queiroz · Harley Francisco de Oliveira · Carlos Gilberto Carlotti Jr. · Carlos Alberto Scrideli · Luiz Gonzaga Tone Received: 27 September 2011 / Accepted: 28 November 2011 / Published online: 9 December 2011 © Springer-Verlag 2011 Abstract Background Glioblastoma remains one of the most devastating human malignancies, and despite therapeutic advances, there are no drugs that signiWcantly improve the patient survival. Altered expression of the Aurora kinases was found in diVerent malignancies, and their inhibition has been studied in cancer therapy. In this study, we analyzed the expression of Aurora A and Aurora B in glioblastoma samples and also analyzed whether the eVects of Aurora kinase inhibition were associated with temozolomide or not on cell lines and primary cultures of glioblastoma. Materials and methods RT-PCR assays were used to determine the mRNA expression in glioblastoma tumor samples and in the cell lines. Cell proliferation was mea- sured by XTT assay, and apoptosis was determined by Xow cytometry. Drug combination analyses were made based in Chou-Talalay method. Gamma radiation for clonogenic survival used the doses of 2, 4 and 6 Gy. Changes in Aurora B level were assessed by Western blot analysis. Results Aurora A and B were expressed in glioblastoma samples as well as in the glioblastoma cell lines (n = 6). Moreover, ZM447439, a selective Aurora kinase inhibitor, decreased the proliferation separately and synergistically with temozolomide in primary cultures and cell lines of glioblastoma. ZM also enhanced the eVects of radiation on the two cell lines studied (U343 and U251), mainly when associated with TMZ in U343 cells. Treatment with ZM induced apoptotic cell death and diminished Aurora B pro- tein level. Conclusions These data suggest that Aurora kinase inhi- bition may be a target for glioblastoma treatment and could be used as adjuvant to chemo- and radiotherapy. Keywords Glioblastoma · Aurora A · Aurora B · ZM447439 · Temozolomide · Radiation Introduction Glioblastoma (GBM) is the most common type of primary malignant brain tumor in adults and is associated with a high mortality rate. Despite access to multimodality treat- ment (surgery, radiotherapy and chemotherapy with the alkylating agent temozolomide), patients diagnosed with GBM have a low survival rate (Khasraw and Lassman 2010; Ohgaki and Kleihues 2009). The Aurora kinase fam- ily is a collection of highly related and conserved serine– threonine kinases consisting of three members (A, B and C). These proteins are involved in the mitotic (M) phase of the cell cycle, playing essential roles in ensuring correct K. S. Borges (&) · A. M. Castro-Gamero · D. A. Moreno · L. G. Tone Department of Genetics, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Avenida Bandeirantes 3900, Ribeirão Preto, SP 14048-900, Brazil e-mail: [email protected] V. da Silva Silveira · M. S. Brassesco · R. G. de Paula Queiroz · C. A. Scrideli · L. G. Tone Department of Pediatrics, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Ribeirão Preto, Brazil H. F. de Oliveira Department of Clinical Medicine, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Ribeirão Preto, Brazil C. G. Carlotti Jr. Department of Surgery and Anatomy, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Ribeirão Preto, Brazil

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

J Cancer Res Clin Oncol (2012) 138:405–414
DOI 10.1007/s00432-011-1111-0ORIGINAL PAPER
Inhibition of Aurora kinases enhances chemosensitivity to temozolomide and causes radiosensitization in glioblastoma cells
Kleiton Silva Borges · Angel Maurício Castro-Gamero · Daniel Antunes Moreno · Vanessa da Silva Silveira · Maria Sol Brassesco · Rosane Gomes de Paula Queiroz · Harley Francisco de Oliveira · Carlos Gilberto Carlotti Jr. · Carlos Alberto Scrideli · Luiz Gonzaga Tone
Received: 27 September 2011 / Accepted: 28 November 2011 / Published online: 9 December 2011© Springer-Verlag 2011
AbstractBackground Glioblastoma remains one of the mostdevastating human malignancies, and despite therapeuticadvances, there are no drugs that signiWcantly improve thepatient survival. Altered expression of the Aurora kinaseswas found in diVerent malignancies, and their inhibition hasbeen studied in cancer therapy. In this study, we analyzedthe expression of Aurora A and Aurora B in glioblastomasamples and also analyzed whether the eVects of Aurorakinase inhibition were associated with temozolomide or noton cell lines and primary cultures of glioblastoma.Materials and methods RT-PCR assays were used todetermine the mRNA expression in glioblastoma tumorsamples and in the cell lines. Cell proliferation was mea-sured by XTT assay, and apoptosis was determined by Xow
cytometry. Drug combination analyses were made based inChou-Talalay method. Gamma radiation for clonogenicsurvival used the doses of 2, 4 and 6 Gy. Changes in AuroraB level were assessed by Western blot analysis.Results Aurora A and B were expressed in glioblastomasamples as well as in the glioblastoma cell lines (n = 6).Moreover, ZM447439, a selective Aurora kinase inhibitor,decreased the proliferation separately and synergisticallywith temozolomide in primary cultures and cell lines ofglioblastoma. ZM also enhanced the eVects of radiation onthe two cell lines studied (U343 and U251), mainly whenassociated with TMZ in U343 cells. Treatment with ZMinduced apoptotic cell death and diminished Aurora B pro-tein level.Conclusions These data suggest that Aurora kinase inhi-bition may be a target for glioblastoma treatment and couldbe used as adjuvant to chemo- and radiotherapy.
Keywords Glioblastoma · Aurora A · Aurora B · ZM447439 · Temozolomide · Radiation
Introduction
Glioblastoma (GBM) is the most common type of primarymalignant brain tumor in adults and is associated with ahigh mortality rate. Despite access to multimodality treat-ment (surgery, radiotherapy and chemotherapy with thealkylating agent temozolomide), patients diagnosed withGBM have a low survival rate (Khasraw and Lassman2010; Ohgaki and Kleihues 2009). The Aurora kinase fam-ily is a collection of highly related and conserved serine–threonine kinases consisting of three members (A, B andC). These proteins are involved in the mitotic (M) phase ofthe cell cycle, playing essential roles in ensuring correct
K. S. Borges (&) · A. M. Castro-Gamero · D. A. Moreno · L. G. ToneDepartment of Genetics, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Avenida Bandeirantes 3900, Ribeirão Preto, SP 14048-900, Brazile-mail: [email protected]
V. da Silva Silveira · M. S. Brassesco · R. G. de Paula Queiroz · C. A. Scrideli · L. G. ToneDepartment of Pediatrics, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Ribeirão Preto, Brazil
H. F. de OliveiraDepartment of Clinical Medicine, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Ribeirão Preto, Brazil
C. G. Carlotti Jr.Department of Surgery and Anatomy, School of Medicine of Ribeirão Preto, University of São Paulo (USP), Ribeirão Preto, Brazil
123

406 J Cancer Res Clin Oncol (2012) 138:405–414
bipolar spindle formation, alignment of centrosomes on themitotic spindle, centrosome separation, cytokinesis andmonitoring of the mitotic checkpoint. Alterations in theexpression of these genes cause disorders at diVerent stagesof the mitotic phase and consequently the development ofaneuploid cells with genomic instability and defects in themitotic apparatus (Boss et al. 2009; Katayama and Sen2010; Green et al. 2011). Overexpression of Aurora Acaused the transformation of rodent Wbroblasts, furthersupporting the role of Aurora A as a potential oncogene(BischoV et al. 1998). In colorectal cell lines and in murineembryo Wbroblasts raised Aurora B protein was associatedwith aneuploidy and increased levels of phosphorylated H3(Ser10). Moreover, these transformed Wbroblasts formedinvasive tumors in nude mice (Ota et al. 2002). Severalreports have shown the overexpression of Aurora A andAurora B in a variety of cancers such as non-small-cell lungcarcinoma (Zhang et al. 2008), breast cancer (Nadler et al.2008; Tanaka et al. 1999) and head and neck carcinoma(Reiter et al. 2006), medulloblastoma (Neben et al. 2004)and neuroblastoma (Morozova et al. 2010). Taken together,these data support the importance of these proteins in thetumorigenesis process and indicate that inhibiting thesekinases might be a powerful antitumor strategy. A numberof small-molecule inhibitors of one or more Aurora kinasesmembers have been developed to target these proteins andto investigate the antineoplastic eVects of Aurora kinaseinhibition (Boss et al. 2009). ZM447439, AZD1152, VX-680 and hesperadin are examples of Aurora kinaseinhibitors that reduce histone H3 phosphorylation, endore-duplication and apoptosis (Green et al. 2011). ZM447439 isan ATP selective inhibitor of Aurora kinases A and B(DitchWeld et al. 2003). Previous studies have reported theantineoplastic eVects of ZM447439 in several cell linesfrom diVerent tumors (Georgieva et al. 2010; Long et al.2008; Walsby et al. 2008). The present study shows theexpression of Aurora A and Aurora B genes in the samplesof patients diagnosed with glioblastoma as well as in apanel of glioblastoma cell lines. Furthermore, exposure ofthe cell lines and primary cultures of glioblastoma toZM447439 induced growth inhibition and synergisticeVects when combined with TMZ. ZM also enhanced radia-tion eVects, alone or associated with TMZ, and causedapoptosis in glioblastoma cell lines.
Materials and methods
Patients analyzed
For this study, 20 fresh-frozen microdissected tumor sam-ples were obtained from gross total surgical resection ofglioblastomas according to the WHO classiWcation (Louis
et al. 2007), from subjects admitted for diagnosis and treat-ment to the University Hospital of the School of Medicineof Ribeirão Preto, SP, Brazil. Five samples of microdis-sected non-neoplastic brain tissue (white matter) wereobtained from patients who had undergone surgery for thetreatment of epilepsy. The average age of the patients was44.3 years, and eleven subjects were men and nine werewomen. The study was approved by the Research EthicsCommittee of the University Hospital, School of Medicineof Ribeirão Preto, University of São Paulo (protocol num-ber 8273/2008).
Cell lines and primary culture
The human adult glioblastoma cell lines U251, U343,T98G and U87 were purchased from the American TypeCulture Collection, and cell line LN319 was kindly pro-vided by Dr. Frank Furnari (Ludwig Institute for CancerResearch, La Jolla, CA). Pediatric glioblastoma cell lineSF188 was kindly provided by Dr Michael S. Bobola(Department of Neurological Surgery, University of Wash-ington, Seattle, WA). Cells were cultured in HAM F10(Gibco BRL, Life Technologies®, Carlsbad, CA, USA)supplemented with 10% fetal bovine serum, penicillin (100U/mL) and streptomycin (100 �g/mL) (Sigma® ChemicalCo., St. Louis, MO, USA) at 37°C in a humidiWed 5% CO2
incubator. The primary culture was derived from the biopsyspecimens of glioblastoma patients under a protocolapproved by the institutional review board at the UniversityHospital of the School of Medicine of Ribeirão Preto, SP,Brazil. Cultures were obtained according to Brassesco et al.(2009) and maintained at low passage numbers (p2–p5)under standard culture conditions.
Drugs
ZM447439 (ZM) was obtained from Tocris Cookson Inc.(Ellisville, MO). The compound was diluted to 10 mmol/Lusing 99% DMSO (Mallinckrodt. Chemical Works, St.Louis, Mo) and stored at ¡20°C. Temozolomide wasobtained from Sigma (Sigma® Chemical Co., St. Louis,MO, USA) and Schering-Plough Brazil. The former wasdiluted to 200 mM using DMSO and stored at ¡20°C, andthe latter was prepared as previously described (Borgeset al. 2011) and used only for the radiation assay.
RNA isolation and reverse transcription-PCR
Total RNA was extracted from each cell line using the Trizol®
reagent (Gibco BRL, Life technologies®, Carlsbad, CA,USA). Complementary DNA (cDNA) was obtained withthe High Capacity® kit (Applied Biosystems®, Foster City,CA, USA) according to the manufacturers’ instructions. cDNA
123

J Cancer Res Clin Oncol (2012) 138:405–414 407
levels of genes Aurora A and Aurora B were measuredusing an ABI 7500 Real Time PCR System (PE AppliedBiosystems). AmpliWcations were obtained using on-demandTaqMan® probes: AURKA (Hs00269212_m1), AURKB(Hs00177782_m1), TBP (4326322E0811008), HPRT (4310809E0502006) and GAPDH (4326317-E 0905031) (AppliedBiosystems). Blank and standard controls (calibrators) wererun in parallel to verify the ampliWcation eYciency withineach experiment. The level of internal control genes, humanGAPDH, TBP and HPRT, were used to normalize the cDNAlevels of the Aurora genes. The material was prepared andstored according to the manufacturer’s instructions, and theWnal volume of each reaction was optimized to 12 �L (6.0 �Lof TaqMan PCR Master Mix, 0.6 �L of TaqMan probe and5.4 �L of cDNA—diluted 1:20). Real-time PCR wasperformed in duplicate, and a standard deviation (SD) of <0·7between duplicates was accepted. Expression levels arereported as a ratio between either Aurora A or Aurora B andthe reference genes GAPDH, for cell lines, and TBP andHPRT for glioblastoma samples. The calibrators of the rela-tive target gene expression were the T98G and U343 cells forthe cell lines and for the glioblastoma samples, respectively.The expression levels of the Aurora kinase genes andreference genes were calculated using the 2¡��CT method(Livak and Schmittgen 2001).
Proliferation assay
To investigate the growth-inhibiting eVects of ZM alone orassociated with TMZ in glioblastoma cells, proliferationassays were conducted. Cells were seeded in 96-well platesat 3 £ 103 per well and maintained in culture. After 24 h,cells were treated with drugs and controls. At diVerent timepoints, the culture medium was removed and replaced withculture medium containing 10 �L of XTT dye (3 mg/mL)(XTT II; Roche Molecular Biochemicals, Indianapolis, IN)in each well. The plates were incubated for 2 h at 37°C, andthe formazan product was measured at 450 nm using aniMark microplate reader (Bio-Rad Laboratories). Valuesrepresent the means § standard deviation (SD) of at leastthree separate experiments performed in triplicate for cul-tured cell lines or at least one experiment depending uponthe cell availability for primary samples. Drug-inducedcytotoxic synergy was analyzed using the CalcuSyn soft-ware (Biosoft, Cambridge, UK). The combination index isa method for quantifying drug cytotoxic synergism, basedon the mass-action law approach and the median eVectprinciple derived from enzyme kinetic models developedby Chou and Talalay (1984) that have been widely used toevaluate the interactions between antineoplastic drugs. Acombination index (CI) of 1 indicates an additive druginteraction, whereas a CI of >1 is antagonistic and a scorelower than 1 is synergistic.
Radiation clonogenic survival assay
A clonogenic assay was performed to study the eVects ofradiation as described previously (Tao et al. 2007). U343and U251 cells were plated into six-well plates with 300cells per dish. After 24 h, cells were treated with ZM(400 nM), TMZ (1 �M) or ZM + TMZ for 48 h. Cells werethen irradiated with 2, 4 and 6 Gy of X-ray radiation (irradi-ation rate of 3.20 Gy/min) using a Gammatron S-80 deviceequipped with a Cobalt-60 source (Siemens Medical Sys-tems, Inc., South Iselin, USA). The irradiated cells werethen cultured in a 5% CO2 incubator at 37°C for an addi-tional 10–14 days. Individual colonies (>50 cells per col-ony) were Wxed with methanol, stained with crystal violetand subsequently counted. Plating eYciency (PE) repre-sents the percentage of cells seeded that grow into coloniesunder a speciWc culture condition of a given cell line. Thesurvival fraction, expressed as a function of irradiation, wascalculated as follows: survival fraction (SF) at 2 Gy(SF) = colonies counted of 2 Gy/(cells seeded of 2 Gy*PE/100). In the clonogenic survival curve, we normalized thediVerent conditions according to the control. The radiationdose enhancement ratio (DER) of the diVerent treatmentswas calculated using the following formula: DER = (SF atan indicated dose of radiation alone)/(SF at an indicateddose of radiation + treatment). Radiosensitization is theterm used when the treatment increases the sensitivity ofcells to radiation. This is calculated with the formula listedabove and represented in the form of DER. Thus, DER isdeWned as the ratio of surviving cells with radiation alonecompared with a combination of radiation and diVerentdrug exposures. A dose enhancement ratio = 1 suggests anadditive radiation eVect and DER >1, a supra-additiveeVect as opposed to a sub-additive eVect in the case ofDER <1.
Western blotting
Anti-AURKB and anti-GAPDH antibodies were purchasedfrom Santa Cruz Biotechnology (Santa Cruz, CA). Equalamounts of protein were size-fractionated by 12.0% SDS-PAGE, blotted onto a nitrocellulose membrane (AmershamHybond™ ECL™, GE Healthcare) and incubated in Tris-buVered saline—0.1% Tween-20 (TBST) containing 5%(w/v) dried non-fat milk for 1 h at room temperature. Afterblocking and washing in PBS with 0.1% Tween 20 for30 min, each membrane was incubated with appropriatelydiluted primary antibodies at 4°C for 2 h. Following mem-brane incubation, the membrane was washed three times inPBS with 0.1% Tween 20 and bound to biotin-labeledhorseradish peroxidase–conjugated species-speciWc sec-ondary antibody (AbCam, CA, USA). The complexes werevisualized using an enhanced chemiluminescence reagent
123
408 J Cancer Res Clin Oncol (2012) 138:405–414
(ECL; Amersham, Uppsala, Sweden). The Wlms wereexposed for 30–60 s.
Apoptosis assessment by annexin V/PI staining
Apoptotic cell death was determined by labeling withAnnexin V—Xuorescein isothiocyanate (FITC) (BD Bio-sciences Pharmigen, USA). BrieXy, after drug treatment,150,000 cells were trypsinized and centrifuged at 1,000 rpmfor 5 min at 4°C, washed with ice-cold PBS and then resus-pended in 300 �L of 1X Annexin V binding buVer (BDBiosciences Pharmigen, USA). Cells were stained with5 �L of annexin V-FITC and 50 �L of a solution 50 �Mpropidium iodide (PI) and incubated at room temperature inthe dark. The samples were analyzed using a BD FACSCal-ibur™ Xow cytometer (BD Biosciences, San Jose, CA,USA).
Statistical analysis
Statistical analysis was performed using one-way or two-way ANOVA followed by Bonferroni’s test, as appropriate.P < 0.05 was considered to be statistically signiWcant. TheeVect of ZM, ZM + TMZ and of dose irradiation on the per-centage of cells leading to colony formation was tested bylogistic regression. Data analysis was carried out using theSPSS17.0 statistical software package. Results were pre-sented as means § SDs.
Results
Aurora A and Aurora B are expressed in glioblastoma samples and cell lines
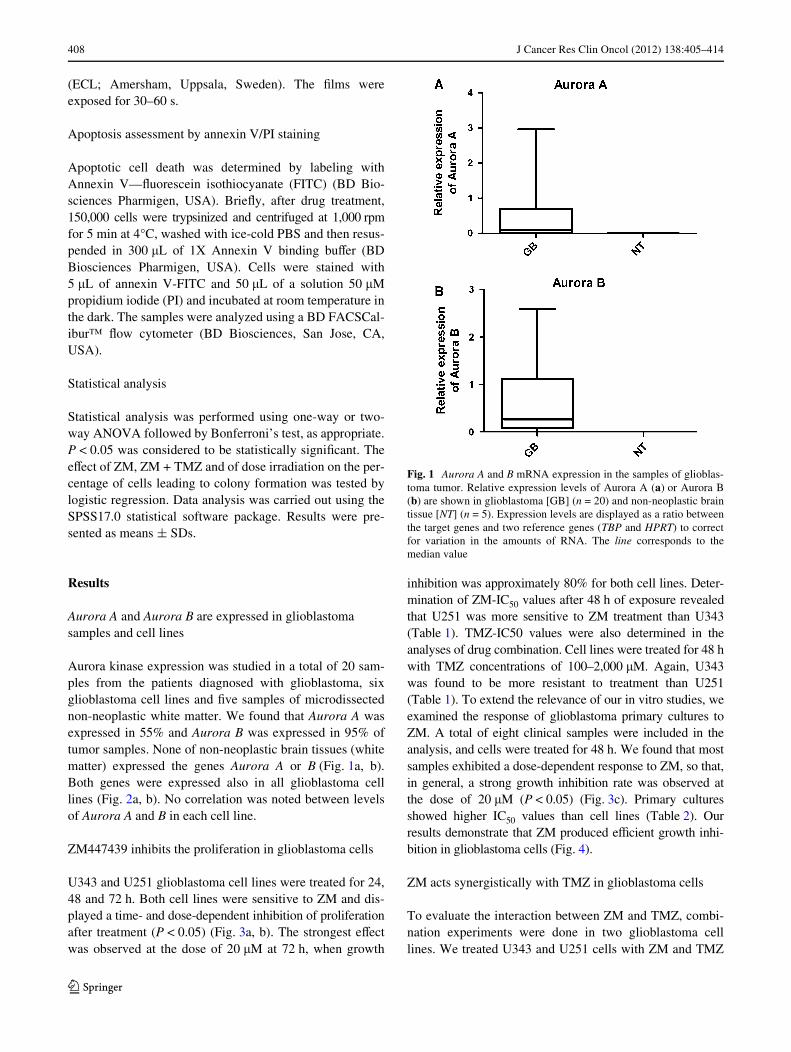
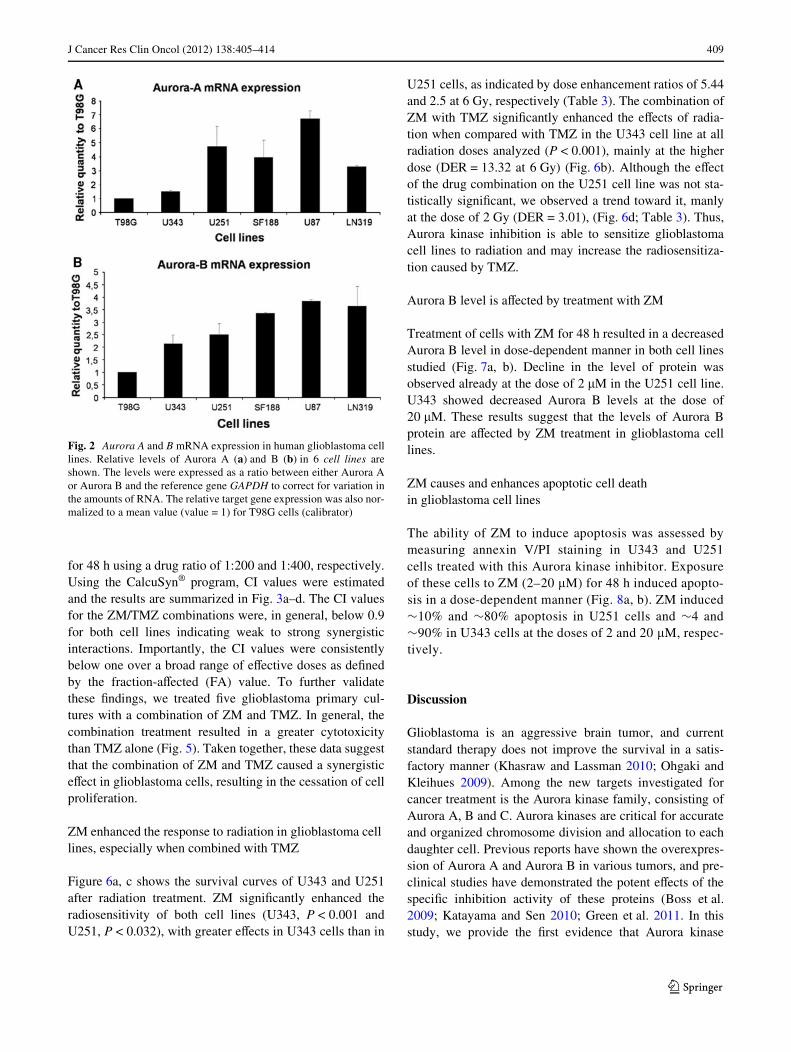
Aurora kinase expression was studied in a total of 20 sam-ples from the patients diagnosed with glioblastoma, sixglioblastoma cell lines and Wve samples of microdissectednon-neoplastic white matter. We found that Aurora A wasexpressed in 55% and Aurora B was expressed in 95% oftumor samples. None of non-neoplastic brain tissues (whitematter) expressed the genes Aurora A or B (Fig. 1a, b).Both genes were expressed also in all glioblastoma celllines (Fig. 2a, b). No correlation was noted between levelsof Aurora A and B in each cell line.
ZM447439 inhibits the proliferation in glioblastoma cells
U343 and U251 glioblastoma cell lines were treated for 24,48 and 72 h. Both cell lines were sensitive to ZM and dis-played a time- and dose-dependent inhibition of proliferationafter treatment (P < 0.05) (Fig. 3a, b). The strongest eVectwas observed at the dose of 20 �M at 72 h, when growth
inhibition was approximately 80% for both cell lines. Deter-mination of ZM-IC50 values after 48 h of exposure revealedthat U251 was more sensitive to ZM treatment than U343(Table 1). TMZ-IC50 values were also determined in theanalyses of drug combination. Cell lines were treated for 48 hwith TMZ concentrations of 100–2,000 �M. Again, U343was found to be more resistant to treatment than U251(Table 1). To extend the relevance of our in vitro studies, weexamined the response of glioblastoma primary cultures toZM. A total of eight clinical samples were included in theanalysis, and cells were treated for 48 h. We found that mostsamples exhibited a dose-dependent response to ZM, so that,in general, a strong growth inhibition rate was observed atthe dose of 20 �M (P < 0.05) (Fig. 3c). Primary culturesshowed higher IC50 values than cell lines (Table 2). Ourresults demonstrate that ZM produced eYcient growth inhi-bition in glioblastoma cells (Fig. 4).
ZM acts synergistically with TMZ in glioblastoma cells
To evaluate the interaction between ZM and TMZ, combi-nation experiments were done in two glioblastoma celllines. We treated U343 and U251 cells with ZM and TMZ
Fig. 1 Aurora A and B mRNA expression in the samples of glioblas-toma tumor. Relative expression levels of Aurora A (a) or Aurora B(b) are shown in glioblastoma [GB] (n = 20) and non-neoplastic braintissue [NT] (n = 5). Expression levels are displayed as a ratio betweenthe target genes and two reference genes (TBP and HPRT) to correctfor variation in the amounts of RNA. The line corresponds to themedian value
123
J Cancer Res Clin Oncol (2012) 138:405–414 409
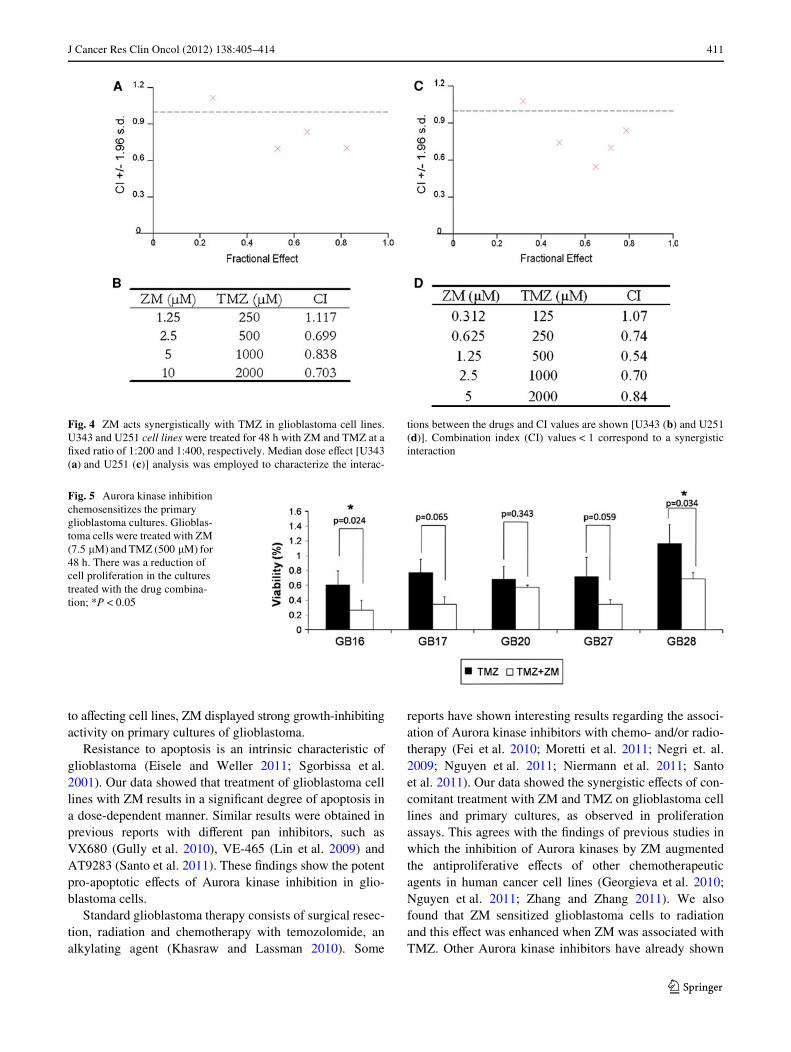
for 48 h using a drug ratio of 1:200 and 1:400, respectively.Using the CalcuSyn® program, CI values were estimatedand the results are summarized in Fig. 3a–d. The CI valuesfor the ZM/TMZ combinations were, in general, below 0.9for both cell lines indicating weak to strong synergisticinteractions. Importantly, the CI values were consistentlybelow one over a broad range of eVective doses as deWnedby the fraction-aVected (FA) value. To further validatethese Wndings, we treated Wve glioblastoma primary cul-tures with a combination of ZM and TMZ. In general, thecombination treatment resulted in a greater cytotoxicitythan TMZ alone (Fig. 5). Taken together, these data suggestthat the combination of ZM and TMZ caused a synergisticeVect in glioblastoma cells, resulting in the cessation of cellproliferation.
ZM enhanced the response to radiation in glioblastoma cell lines, especially when combined with TMZ
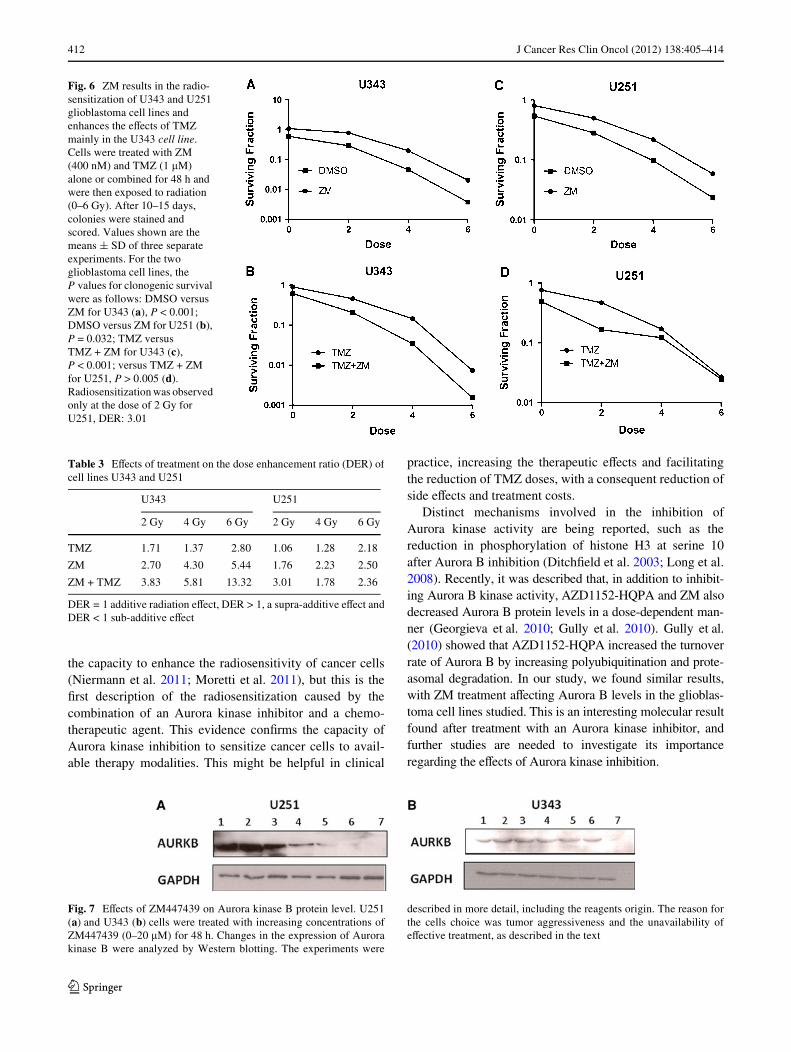
Figure 6a, c shows the survival curves of U343 and U251after radiation treatment. ZM signiWcantly enhanced theradiosensitivity of both cell lines (U343, P < 0.001 andU251, P < 0.032), with greater eVects in U343 cells than in
U251 cells, as indicated by dose enhancement ratios of 5.44and 2.5 at 6 Gy, respectively (Table 3). The combination ofZM with TMZ signiWcantly enhanced the eVects of radia-tion when compared with TMZ in the U343 cell line at allradiation doses analyzed (P < 0.001), mainly at the higherdose (DER = 13.32 at 6 Gy) (Fig. 6b). Although the eVectof the drug combination on the U251 cell line was not sta-tistically signiWcant, we observed a trend toward it, manlyat the dose of 2 Gy (DER = 3.01), (Fig. 6d; Table 3). Thus,Aurora kinase inhibition is able to sensitize glioblastomacell lines to radiation and may increase the radiosensitiza-tion caused by TMZ.
Aurora B level is aVected by treatment with ZM
Treatment of cells with ZM for 48 h resulted in a decreasedAurora B level in dose-dependent manner in both cell linesstudied (Fig. 7a, b). Decline in the level of protein wasobserved already at the dose of 2 �M in the U251 cell line.U343 showed decreased Aurora B levels at the dose of20 �M. These results suggest that the levels of Aurora Bprotein are aVected by ZM treatment in glioblastoma celllines.
ZM causes and enhances apoptotic cell death in glioblastoma cell lines
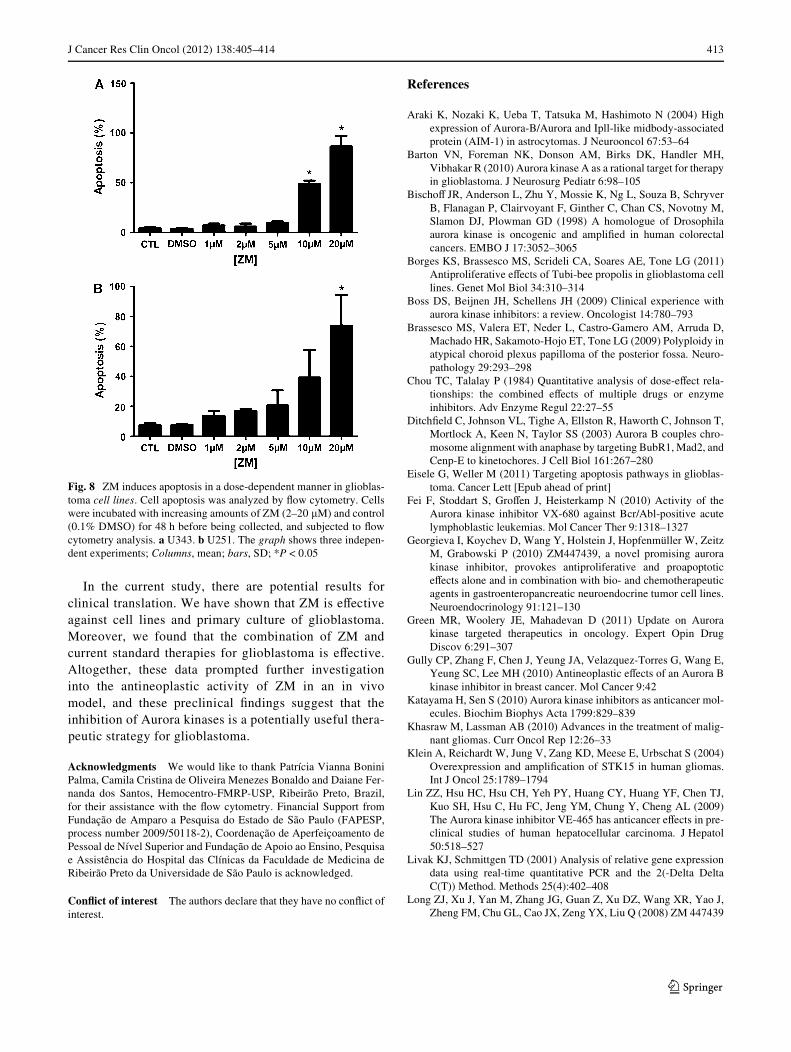
The ability of ZM to induce apoptosis was assessed bymeasuring annexin V/PI staining in U343 and U251cells treated with this Aurora kinase inhibitor. Exposureof these cells to ZM (2–20 �M) for 48 h induced apopto-sis in a dose-dependent manner (Fig. 8a, b). ZM induced»10% and »80% apoptosis in U251 cells and »4 and»90% in U343 cells at the doses of 2 and 20 �M, respec-tively.
Discussion
Glioblastoma is an aggressive brain tumor, and currentstandard therapy does not improve the survival in a satis-factory manner (Khasraw and Lassman 2010; Ohgaki andKleihues 2009). Among the new targets investigated forcancer treatment is the Aurora kinase family, consisting ofAurora A, B and C. Aurora kinases are critical for accurateand organized chromosome division and allocation to eachdaughter cell. Previous reports have shown the overexpres-sion of Aurora A and Aurora B in various tumors, and pre-clinical studies have demonstrated the potent eVects of thespeciWc inhibition activity of these proteins (Boss et al.2009; Katayama and Sen 2010; Green et al. 2011. In thisstudy, we provide the Wrst evidence that Aurora kinase
Fig. 2 Aurora A and B mRNA expression in human glioblastoma celllines. Relative levels of Aurora A (a) and B (b) in 6 cell lines areshown. The levels were expressed as a ratio between either Aurora Aor Aurora B and the reference gene GAPDH to correct for variation inthe amounts of RNA. The relative target gene expression was also nor-malized to a mean value (value = 1) for T98G cells (calibrator)
123

410 J Cancer Res Clin Oncol (2012) 138:405–414
inhibition may be a promising anticancer agent for glioblas-toma monotherapy as well as in combination with temozol-omide and/or radiation.
The proposal of Aurora A and Aurora B as potential tar-gets for glioblastoma treatment arose from studies showingthe aberrant expression of these genes in this tumor (Arakiet al. 2004; Barton et al. 2010; Klein et al. 2004; Samaraset al. 2009; Scrideli et al. 2008; Zeng et al. 2007). Zeng et al.(2007) found that Aurora B expression levels were mark-edly correlated with a shortened survival, and Barton et al.(2010) showed that Aurora A was associated with a poorprognosis. In this study, we also showed Aurora A andAurora B mRNA expression in most glioblastoma samples.Besides, all glioblastoma cell lines expressed both genes.All these data provide enough evidence to evaluate thesetargets as potential alternatives for glioblastoma treatment.
Several small molecules have been studied to inhibitAurora kinase activity; among them, ZM447439 (ZM) is anovel and speciWc Aurora A and Aurora B inhibitor withpotent antineoplastic eVects (DitchWeld et al. 2003; Georgievaet al. 2010; Green et al. 2011; Long et al. 2008). Weshowed that ZM eVectively inhibited the proliferation ofglioblastoma cell lines and was also able to inhibit thegrowth of primary culture specimens. Primary tumor cul-tures are believed to be more representative of in vivotumors than established cancer cell lines and generally dis-play weaker responses to chemotherapeutics than do celllines (Qian et al. 2006). Thus, it is notable that, in addition
Fig. 3 ZM447439 inhibited the proliferation of glioblastoma cells.U343 (a) and U251 (b) glioblastoma cell lines and primary cultures ofglioblastoma tumors (c) were cultured in the presence of various concen-
trations of ZM447439 (2–20 �M) for 24, 48 and 72 h for cell lines and48 h for the primary cultures. Proliferation was measured with the XTTkit. *P < 0.05
Table 1 IC50 values for ZM447439 (ZM) and temozolomide (TMZ)in the U343 and U251 cell lines
Cell line IC50 (�M)
ZM TMZ
U343 12.2 § 4.4 825 § 90
U251 7.7 § 0.68 315.7 § 52.7
Table 2 IC50 values for ZM447439 in glioblastoma primary cultures
Tumor samples IC50 (�M)
GB03 10.82
GB12 20.80
GB15 9.97
GB17 22.37
GB20 15.55
GB22 13.56
GB27 28.21
GB28 n.p*
GB41 21.89* It was not possible to calculate IC50
123
J Cancer Res Clin Oncol (2012) 138:405–414 411
to aVecting cell lines, ZM displayed strong growth-inhibitingactivity on primary cultures of glioblastoma.
Resistance to apoptosis is an intrinsic characteristic ofglioblastoma (Eisele and Weller 2011; Sgorbissa et al.2001). Our data showed that treatment of glioblastoma celllines with ZM results in a signiWcant degree of apoptosis ina dose-dependent manner. Similar results were obtained inprevious reports with diVerent pan inhibitors, such asVX680 (Gully et al. 2010), VE-465 (Lin et al. 2009) andAT9283 (Santo et al. 2011). These Wndings show the potentpro-apoptotic eVects of Aurora kinase inhibition in glio-blastoma cells.
Standard glioblastoma therapy consists of surgical resec-tion, radiation and chemotherapy with temozolomide, analkylating agent (Khasraw and Lassman 2010). Some
reports have shown interesting results regarding the associ-ation of Aurora kinase inhibitors with chemo- and/or radio-therapy (Fei et al. 2010; Moretti et al. 2011; Negri et. al.2009; Nguyen et al. 2011; Niermann et al. 2011; Santoet al. 2011). Our data showed the synergistic eVects of con-comitant treatment with ZM and TMZ on glioblastoma celllines and primary cultures, as observed in proliferationassays. This agrees with the Wndings of previous studies inwhich the inhibition of Aurora kinases by ZM augmentedthe antiproliferative eVects of other chemotherapeuticagents in human cancer cell lines (Georgieva et al. 2010;Nguyen et al. 2011; Zhang and Zhang 2011). We alsofound that ZM sensitized glioblastoma cells to radiationand this eVect was enhanced when ZM was associated withTMZ. Other Aurora kinase inhibitors have already shown
Fig. 4 ZM acts synergistically with TMZ in glioblastoma cell lines.U343 and U251 cell lines were treated for 48 h with ZM and TMZ at aWxed ratio of 1:200 and 1:400, respectively. Median dose eVect [U343(a) and U251 (c)] analysis was employed to characterize the interac-
tions between the drugs and CI values are shown [U343 (b) and U251(d)]. Combination index (CI) values < 1 correspond to a synergisticinteraction
Fig. 5 Aurora kinase inhibition chemosensitizes the primary glioblastoma cultures. Glioblas-toma cells were treated with ZM (7.5 �M) and TMZ (500 �M) for 48 h. There was a reduction of cell proliferation in the cultures treated with the drug combina-tion; *P < 0.05
123
412 J Cancer Res Clin Oncol (2012) 138:405–414
the capacity to enhance the radiosensitivity of cancer cells(Niermann et al. 2011; Moretti et al. 2011), but this is theWrst description of the radiosensitization caused by thecombination of an Aurora kinase inhibitor and a chemo-therapeutic agent. This evidence conWrms the capacity ofAurora kinase inhibition to sensitize cancer cells to avail-able therapy modalities. This might be helpful in clinical
practice, increasing the therapeutic eVects and facilitatingthe reduction of TMZ doses, with a consequent reduction ofside eVects and treatment costs.
Distinct mechanisms involved in the inhibition ofAurora kinase activity are being reported, such as thereduction in phosphorylation of histone H3 at serine 10after Aurora B inhibition (DitchWeld et al. 2003; Long et al.2008). Recently, it was described that, in addition to inhibit-ing Aurora B kinase activity, AZD1152-HQPA and ZM alsodecreased Aurora B protein levels in a dose-dependent man-ner (Georgieva et al. 2010; Gully et al. 2010). Gully et al.(2010) showed that AZD1152-HQPA increased the turnoverrate of Aurora B by increasing polyubiquitination and prote-asomal degradation. In our study, we found similar results,with ZM treatment aVecting Aurora B levels in the glioblas-toma cell lines studied. This is an interesting molecular resultfound after treatment with an Aurora kinase inhibitor, andfurther studies are needed to investigate its importanceregarding the eVects of Aurora kinase inhibition.
Fig. 6 ZM results in the radio-sensitization of U343 and U251 glioblastoma cell lines and enhances the eVects of TMZ mainly in the U343 cell line. Cells were treated with ZM (400 nM) and TMZ (1 �M) alone or combined for 48 h and were then exposed to radiation (0–6 Gy). After 10–15 days, colonies were stained and scored. Values shown are the means § SD of three separate experiments. For the two glioblastoma cell lines, the P values for clonogenic survival were as follows: DMSO versus ZM for U343 (a), P < 0.001; DMSO versus ZM for U251 (b), P = 0.032; TMZ versus TMZ + ZM for U343 (c), P < 0.001; versus TMZ + ZM for U251, P > 0.005 (d). Radiosensitization was observed only at the dose of 2 Gy for U251, DER: 3.01
Table 3 EVects of treatment on the dose enhancement ratio (DER) ofcell lines U343 and U251
DER = 1 additive radiation eVect, DER > 1, a supra-additive eVect andDER < 1 sub-additive eVect
U343 U251
2 Gy 4 Gy 6 Gy 2 Gy 4 Gy 6 Gy
TMZ 1.71 1.37 2.80 1.06 1.28 2.18
ZM 2.70 4.30 5.44 1.76 2.23 2.50
ZM + TMZ 3.83 5.81 13.32 3.01 1.78 2.36
Fig. 7 EVects of ZM447439 on Aurora kinase B protein level. U251(a) and U343 (b) cells were treated with increasing concentrations ofZM447439 (0–20 �M) for 48 h. Changes in the expression of Aurorakinase B were analyzed by Western blotting. The experiments were
described in more detail, including the reagents origin. The reason forthe cells choice was tumor aggressiveness and the unavailability ofeVective treatment, as described in the text
123
J Cancer Res Clin Oncol (2012) 138:405–414 413
In the current study, there are potential results forclinical translation. We have shown that ZM is eVectiveagainst cell lines and primary culture of glioblastoma.Moreover, we found that the combination of ZM andcurrent standard therapies for glioblastoma is eVective.Altogether, these data prompted further investigationinto the antineoplastic activity of ZM in an in vivomodel, and these preclinical Wndings suggest that theinhibition of Aurora kinases is a potentially useful thera-peutic strategy for glioblastoma.
Acknowledgments We would like to thank Patrícia Vianna BoniniPalma, Camila Cristina de Oliveira Menezes Bonaldo and Daiane Fer-nanda dos Santos, Hemocentro-FMRP-USP, Ribeirão Preto, Brazil,for their assistance with the Xow cytometry. Financial Support fromFundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP,process number 2009/50118-2), Coordenação de Aperfeiçoamento dePessoal de Nível Superior and Fundação de Apoio ao Ensino, Pesquisae Assistência do Hospital das Clínicas da Faculdade de Medicina deRibeirão Preto da Universidade de São Paulo is acknowledged.
ConXict of interest The authors declare that they have no conXict ofinterest.
References
Araki K, Nozaki K, Ueba T, Tatsuka M, Hashimoto N (2004) Highexpression of Aurora-B/Aurora and Ipll-like midbody-associatedprotein (AIM-1) in astrocytomas. J Neurooncol 67:53–64
Barton VN, Foreman NK, Donson AM, Birks DK, Handler MH,Vibhakar R (2010) Aurora kinase A as a rational target for therapyin glioblastoma. J Neurosurg Pediatr 6:98–105
BischoV JR, Anderson L, Zhu Y, Mossie K, Ng L, Souza B, SchryverB, Flanagan P, Clairvoyant F, Ginther C, Chan CS, Novotny M,Slamon DJ, Plowman GD (1998) A homologue of Drosophilaaurora kinase is oncogenic and ampliWed in human colorectalcancers. EMBO J 17:3052–3065
Borges KS, Brassesco MS, Scrideli CA, Soares AE, Tone LG (2011)Antiproliferative eVects of Tubi-bee propolis in glioblastoma celllines. Genet Mol Biol 34:310–314
Boss DS, Beijnen JH, Schellens JH (2009) Clinical experience withaurora kinase inhibitors: a review. Oncologist 14:780–793
Brassesco MS, Valera ET, Neder L, Castro-Gamero AM, Arruda D,Machado HR, Sakamoto-Hojo ET, Tone LG (2009) Polyploidy inatypical choroid plexus papilloma of the posterior fossa. Neuro-pathology 29:293–298
Chou TC, Talalay P (1984) Quantitative analysis of dose-eVect rela-tionships: the combined eVects of multiple drugs or enzymeinhibitors. Adv Enzyme Regul 22:27–55
DitchWeld C, Johnson VL, Tighe A, Ellston R, Haworth C, Johnson T,Mortlock A, Keen N, Taylor SS (2003) Aurora B couples chro-mosome alignment with anaphase by targeting BubR1, Mad2, andCenp-E to kinetochores. J Cell Biol 161:267–280
Eisele G, Weller M (2011) Targeting apoptosis pathways in glioblas-toma. Cancer Lett [Epub ahead of print]
Fei F, Stoddart S, GroVen J, Heisterkamp N (2010) Activity of theAurora kinase inhibitor VX-680 against Bcr/Abl-positive acutelymphoblastic leukemias. Mol Cancer Ther 9:1318–1327
Georgieva I, Koychev D, Wang Y, Holstein J, Hopfenmüller W, ZeitzM, Grabowski P (2010) ZM447439, a novel promising aurorakinase inhibitor, provokes antiproliferative and proapoptoticeVects alone and in combination with bio- and chemotherapeuticagents in gastroenteropancreatic neuroendocrine tumor cell lines.Neuroendocrinology 91:121–130
Green MR, Woolery JE, Mahadevan D (2011) Update on Aurorakinase targeted therapeutics in oncology. Expert Opin DrugDiscov 6:291–307
Gully CP, Zhang F, Chen J, Yeung JA, Velazquez-Torres G, Wang E,Yeung SC, Lee MH (2010) Antineoplastic eVects of an Aurora Bkinase inhibitor in breast cancer. Mol Cancer 9:42
Katayama H, Sen S (2010) Aurora kinase inhibitors as anticancer mol-ecules. Biochim Biophys Acta 1799:829–839
Khasraw M, Lassman AB (2010) Advances in the treatment of malig-nant gliomas. Curr Oncol Rep 12:26–33
Klein A, Reichardt W, Jung V, Zang KD, Meese E, Urbschat S (2004)Overexpression and ampliWcation of STK15 in human gliomas.Int J Oncol 25:1789–1794
Lin ZZ, Hsu HC, Hsu CH, Yeh PY, Huang CY, Huang YF, Chen TJ,Kuo SH, Hsu C, Hu FC, Jeng YM, Chung Y, Cheng AL (2009)The Aurora kinase inhibitor VE-465 has anticancer eVects in pre-clinical studies of human hepatocellular carcinoma. J Hepatol50:518–527
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta DeltaC(T)) Method. Methods 25(4):402–408
Long ZJ, Xu J, Yan M, Zhang JG, Guan Z, Xu DZ, Wang XR, Yao J,Zheng FM, Chu GL, Cao JX, Zeng YX, Liu Q (2008) ZM 447439
Fig. 8 ZM induces apoptosis in a dose-dependent manner in glioblas-toma cell lines. Cell apoptosis was analyzed by Xow cytometry. Cellswere incubated with increasing amounts of ZM (2–20 �M) and control(0.1% DMSO) for 48 h before being collected, and subjected to Xowcytometry analysis. a U343. b U251. The graph shows three indepen-dent experiments; Columns, mean; bars, SD; *P < 0.05
123

414 J Cancer Res Clin Oncol (2012) 138:405–414
inhibition of aurora kinase induces Hep2 cancer cell apoptosis inthree-dimensional culture. Cell Cycle 7:1473–1479
Louis DN, Ohgaki H, Wiestler OD (2007) The 2007 WHO classiWca-tion of tumours of the central nervous system. Acta Neuropathol114:97–109
Moretti L, Niermann K, Schleicher S, Giacalone NJ, Varki V, KimKW, Kopsombut P, Jung DK, Lu B (2011) LN8054, a small mol-ecule inhibitor of aurora kinase a, sensitizes androgen-resistantprostate cancer to radiation. Int J Radiat Oncol Biol Phys80:1189–1197
Morozova O, Vojvodic M, Grinshtein N, Hansford LM, Blakely KM,Maslova A, Hirst M, Cezard T, Morin RD, Moore R, Smith KM,Miller F, Taylor P, Thiessen N, Varhol R, Zhao Y, Jones S, MoVatJ, Kislinger T, Moran MF, Kaplan DR, Marra MA (2010) System-level analysis of neuroblastoma tumor-initiating cells implicatesAURKB as a novel drug target for neuroblastoma. Clin CancerRes 16:4572–4582
Nadler Y, Camp RL, Schwartz C, Rimm DL, Kluger HM, Kluger Y(2008) Expression of Aurora A (but not Aurora B) is predictive ofsurvival in breast cancer. Clin Cancer Res 15:4455–4462
Neben K, Korshunov A, Benner A, Wrobel G, Hahn M, Kokocinski F,Golanov A, Joos S, Lichter P (2004) Microarray-based screeningfor molecular markers in medulloblastoma revealed STK15 asindependent predictor for survival. Cancer Res 64:3103–3111
Negri JM, McMillin DW, Delmore J, Mitsiades N, Hayden P, KlippelS, Hideshima T, Chauhan D, Munshi NC, Buser CA, Pollard J,Richardson PG, Anderson KC, Mitsiades CS (2009) In vitro anti-myeloma activity of the Aurora kinase inhibitor VE-465. Br JHaematol 147:672–676
Nguyen T, Dai Y, Attkisson E, Kramer L, Jordan N, Nguyen N, KolluriN, Muschen M, Grant S (2011) HDAC inhibitors potentiate theactivity of the BCR/ABL kinase inhibitor KW-2449 in imatinib-sensitive or -resistant BCR/ABL+ leukemia cells in vitro and invivo. Clin Cancer Res 17(10):3219–3232
Niermann KJ, Moretti L, Giacalone NJ, Sun Y, Schleicher SM,Kopsombut P, Mitchell LR, Kim KW, Lu B (2011) Enhancedradiosensitivity of androgen-resistant prostate cancer: AZD1152-mediated Aurora kinase B inhibition. Radiat Res 175:444–451
Ohgaki H, Kleihues P (2009) Genetic alterations and signaling path-ways in the evolution of gliomas. Cancer Sci 100:2235–2241
Ota T, Suto S, Katayama H, Han ZB, Suzuki F, Maeda M, Tanino M,Terada Y, Tatsuka M (2002) Increased mitotic phosphorylation ofhistone H3 attributable to AIM-1/Aurora-B overexpression contrib-utes to chromosome number instability. Cancer Res 62:5168–5177
Qian X, LaRochelle WJ, Ara G, Wu F, Petersen KD, Thougaard A,Sehested M, Lichenstein HS, JeVers M (2006) Activity ofPXD101, a histone deacetylase inhibitor, in preclinical ovariancancer studies. Mol Cancer Ther 5:2086–2095
Reiter R, Gais P, Jütting U, Steuer-Vogt MK, Pickhard A, Bink K,Rauser S, Lassmann S, HöXer H, Werner M, Walch A (2006)
Aurora kinase A messenger RNA overexpression is correlatedwith tumor progression and shortened survival in head and necksquamous cell carcinoma. Clin Cancer Res 1:5136–5141
Samaras V, Stamatelli A, Samaras E, Arnaoutoglou C, ArnaoutoglouM, Stergiou I, Konstantopoulou P, Varsos V, Karameris A,Barbatis C (2009) Comparative immunohistochemical analysis ofaurora-A and aurora-B expression in human glioblastomas.Associations with proliferative activity and clinicopathologicalfeatures. Pathol Res Pract 205:765–773
Santo L, Hideshima T, Cirstea D, Bandi M, Nelson EA, Gorgun G,Rodig S, Vallet S, Pozzi S, Patel K, Unitt C, Squires M, Hu Y,Chauhan D, Mahindra A, Munshi NC, Anderson KC, Raje N(2011) Antimyeloma activity of a multitargeted kinase inhibitor,AT9283, via potent Aurora kinase and STAT3 inhibition eitheralone or in combination with lenalidomide. Clin Cancer Res17:3259–3271
Scrideli CA, Carlotti CG Jr, Okamoto OK, Andrade VS, Cortez MA,Motta FJ, Lucio-Eterovic AK, Neder L, Rosemberg S, Oba-ShinjoSM, Marie SK, Tone LG (2008) Gene expression proWle analysisof primary glioblastomas and non-neoplastic brain tissue: identi-Wcation of potential target genes by oligonucleotide microarrayand real-time quantitative PCR. J Neurooncol 88:281–291
Sgorbissa A, Tomasella A, Potu H, Manini I, Brancolini C (2001) TypeI IFNs signaling and apoptosis resistance in glioblastoma cells.Apoptosis 21 Aug 2011. [Epub ahead of print]
Tanaka T, Kimura M, Matsunaga K, Fukada D, Mori H, Okano Y(1999) Centrosomal kinase AIK1 is overexpressed in invasiveductal carcinoma of the breast. Cancer Res 1:2041–2044
Tao Y, Zhang P, Frascogna V, Lecluse Y, Auperin A, Bourhis J,Deutsch E (2007) Enhancement of radiation response by inhibi-tion of Aurora-A kinase using siRNA or a selective Aurora kinaseinhibitor PHA680632 in p53-deWcient cancer cells. Br J Cancer97:1664–1672
Walsby E, Walsh V, Pepper C, Burnett A, Mills K (2008) EVects of theaurora kinase inhibitors AZD1152-HQPA and ZM447439 ongrowth arrest and polyploidy in acute myeloid leukemia cell linesand primary blasts. Haematologica 93:662–669
Zeng WF, Navaratne K, Prayson RA, Weil RJ (2007) Aurora B expres-sion correlates with aggressive behaviour in glioblastoma multi-forme. J Clin Pathol 60:218–221
Zhang L, Zhang S (2011) ZM447439, the Aurora kinase B inhibitor,suppresses the growth of cervical cancer SiHa cells, enhances thechemosensitivity to cisplatin. J Obstet Gynaecol Res 37:591–600
Zhang XH, Rao M, Loprieato JA, Hong JA, Zhao M, Chen GZ,Humphries AE, Nguyen DM, Trepel JB, Yu X, Schrump DS(2008) Aurora A, Aurora B and survivin are novel targets oftranscriptional regulation by histone deacetylase inhibitors innon-small cell lung cancer. Cancer Biol Ther 7:1388–1397
123
Related Documents











